© 2018 anil sigdel - ufdcimages.uflib.ufl.edu
TRANSCRIPT

GENETIC ANALYSIS OF HEAT TOLERANCE FOR PRODUCTION, REPRODUCTION AND HEALTH TRAITS IN US HOLSTEIN COWS
By
ANIL SIGDEL
A THESIS PRESENTED TO THE GRADUATE SCHOOL OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT
OF THE REQUIREMENTS FOR THE DEGREE OF MASTER OF SCIENCE
UNIVERSITY OF FLORIDA
2018

© 2018 Anil Sigdel

To dairy farmers’ around the globe

ACKNOWLEDGMENTS
First and foremost, I would like to express my deepest gratitude to my advisor Dr.
Francisco Peñagaricano for his intellect support, critical analysis of data and for offering
valuable ideas and suggestions during my master study. Without his valuable
suggestions, meticulous supervision, and constructive criticism, this manuscript would
not come up to its present quality. Many thanks also go to members of my committee;
Dr. Peter J. Hansen and Dr. Raluca Mateescu for their advice and suggestions.
I am very thankful to Dr. Rostam Abdollahi-Arpanahi for his guidance and
expertise that he shared with me during my research. I wish to thank the past and
current members of Quantitative Genetics and Genomics group at University of Florida
for providing friendly and enthusiastic environment. Special thanks go to Yi Han, Rocio
Amorin, Fernanda Rezende, Juan Pablo Nani, Leeher for their friendship and
assistance. I would also like to thank Animal Science Graduate Students’ Association
here in the Department of Animal Sciences, UF for enriching my personal and
professional experiences.
I appreciate the support from South East Milk Check Off and Department of
Animal Sciences, UF for funding my study. I would also like to thank Council on Dairy
Cattle Breeding and North Florida Holsteins for providing necessary data for this study. I
wish to thank my parents Mr. Bishnu Prasad Sigdel and Mrs. Bhagirathi Sigdel, and my
sisters and brother for their love and encouragement. Finally, I will like to thank my
brother Narendra and friend Bikalpa for their advices and suggestions on my scientific
work, future professional career, and personal life. I remain indebted to them.

5
TABLE OF CONTENTS page
ACKNOWLEDGMENTS .................................................................................................. 4
LIST OF TABLES ............................................................................................................ 7
LIST OF FIGURES .......................................................................................................... 8
LIST OF ABBREVIATIONS ............................................................................................. 9
ABSTRACT ................................................................................................................... 10
CHAPTER
1 INTRODUCTION .................................................................................................... 12
2 LITERATURE REVIEW .......................................................................................... 14
Thermoneutral Zone ............................................................................................... 15
Heat Stress ............................................................................................................. 16 Responses to Heat Stress ...................................................................................... 17 Methods of Heat Transfer ....................................................................................... 17
Conduction and Convection ............................................................................. 18 Radiation .......................................................................................................... 19
Evaporative Heat Loss ..................................................................................... 19 Physiological Responses of Cattle to Heat Stress .................................................. 20
Reduced Feed Intake ....................................................................................... 20 Decreased Milk Production ............................................................................... 21
Decreased Reproduction .................................................................................. 22 Increased Water Intake .................................................................................... 23 Increased Respiration Rate .............................................................................. 23
Changes in Blood Hormones Concentration .................................................... 23 Methods for Assessment of Heat Stress ................................................................. 24
Rectal Temperature .......................................................................................... 24
Milk Temperature ............................................................................................. 25 Respiration Rate ............................................................................................... 25 Heat Shock Proteins (HSPs) ............................................................................ 26
Strategies for Reduction of Effects of Heat Stress .................................................. 27 Physical Modifications of the Environment ....................................................... 27
Nutritional Strategies for Managing Heat Stress in Dairy Cows ........................ 28 Genetic Development of Heat Resistant Animals ............................................. 28
Genetic Studies on Heat Stress .............................................................................. 30
3 GENETIC ANALYSIS OF HEAT TOLERANCE FOR PRODUCTION TRAITS IN US HOLSTEIN COWS ............................................................................................ 33
Background ............................................................................................................. 33

6
Materials and Methods............................................................................................ 35
Phenotypic and Genotypic Data ....................................................................... 35
Statistical Model ............................................................................................... 36 Statistical Analysis ............................................................................................ 37 Genome Wide Association Mapping using ssGBLUP ...................................... 38 Gene Set Analysis ............................................................................................ 39
Assignment of SNPs to genes ................................................................... 39
Assignment of genes to pathways ............................................................. 40 Pathway-based association analysis ......................................................... 40
Results and Discussion........................................................................................... 41 Genetic Dissection of Heat Stress for MY, FY and PY ..................................... 41 Whole-Genome Mapping for Thermotolerance Genes affecting MY ................ 42
Pathway-Based Association Analysis for MY ................................................... 45
Summary ................................................................................................................ 48
4 GENETIC ANALYSIS OF HEAT TOLERANCE FOR REPRODUCTION AND UDDER HEALTH IN US HOLSTEIN COWS .......................................................... 55
Background ............................................................................................................. 55 Materials and Methods............................................................................................ 58
Phenotypic and Genotypic Data ....................................................................... 58
Statistical Model ............................................................................................... 59 Statistical Analysis ............................................................................................ 60
Genome-wide Association Mapping using ssGBLUP ....................................... 61 Gene Set Analysis ............................................................................................ 62
Assignment of SNPs to genes ................................................................... 62
Assignment of genes to pathways ............................................................. 62
Pathway-based association analysis ......................................................... 62 Results and Discussion........................................................................................... 63
Genetic Dissection of Heat Stress for Conception and Udder Health ............... 63
Variance Component Estimates for CI ............................................................. 63 Variance Component Estimates for SCS.......................................................... 64 Genomic Scans for Thermotolerance Genes Affecting CI and SCS ................. 64
Whole Genome association analysis for CI................................................ 64 Whole Genome association analysis for SCS ............................................ 66
Pathway-Based Analysis .................................................................................. 68 Gene-set analysis for CI ............................................................................ 69 Gene-set analysis for SCS ......................................................................... 70
Summary ................................................................................................................ 72
5 CONCLUSIONS ..................................................................................................... 79
LIST OF REFERENCES ............................................................................................... 81
BIOGRAPHICAL SKETCH ............................................................................................ 94

7
LIST OF TABLES
Table page 2-1 General (σ2
a) and heat tolerance (100σ2v) additive variances, genetic
correlations and estimates of heritability h2f(10) at THI of 78 (10 units above
THI threshold of 68) for MY(Kg)2 ........................................................................ 50
2-2 General (σ2a) and heat tolerance (100σ2
v) additive variances, genetic correlations and estimates of heritability h2
f(10) at THI of 78 (10 units above THI threshold of 68) for FY(Kg x 100)2 ............................................................... 51
2-3 General (σ2a) and heat tolerance (100σ2
v) additive variances, genetic correlations and estimates of heritability h2
f(10) at THI of 78 (10 units above THI threshold of 68) for PY(Kg x 100)2 ............................................................... 52
2-4 GO terms significantly enriched with genes associated with MY ....................... 53
2-5 MeSH terms significantly enriched with genes associated with MY ................... 53
3-1 General (σ2a) and heat tolerance (100σ2
v) additive variances, genetic correlations and estimates of heritability h2
f(10) at THI of 78 (10 units above THI threshold) for CI ........................................................................................... 73
3-2 General (σ2a) and heat tolerance (100σ2
v) additive variances, genetic correlation and estimates of heritability h2
f(10) at THI of 78 (10 units above THI threshold) for SCS .............................................................................................. 74
3-3 GO terms significantly enriched with genes associated with CI ......................... 75
3-4 MeSH terms significantly enriched with genes associated with CI .................... 75
3-5 GO terms significantly enriched with genes associated with SCS ..................... 76
3-6 MeSH terms significantly enriched with genes associated with SCS ................. 76

8
LIST OF FIGURES
Figure page 2-1 Manhattan plot showing the results of genome-wide association mapping for
MY. ..................................................................................................................... 54
3-1 Manhattan plot showing the results of genome-wide association mapping for CI. ....................................................................................................................... 77
3-2 Manhattan plot showing the results of genome-wide association mapping for SCS .................................................................................................................... 78

9
LIST OF ABBREVIATIONS
BTA Bos taurus autosome
CI Conception per insemination
CM Clinical mastitis
DEG Differentially expressed genes
DIM Days in milk
DNA Deoxyribonucleic acid
FY Fat yield
MY Milk yield
PY Protein yield
RR Respiration rate
SCS Somatic cell score
THI Temperature humidity index
TNZ Thermoneutral zone

10
Abstract of Thesis Presented to the Graduate School of the University of Florida in Partial Fulfillment of the
Requirements for the Master of Science
GENETIC ANALYSIS OF HEAT TOLERANCE FOR PRODUCTION, REPRODUCTION AND HEALTH TRAITS IN US HOLSTEIN COWS
By
AniI Sigdel
May 2018
Chair: Francisco Peñagaricano Major: Animal Sciences
Heat stress is an important economic issue in dairy farming, especially in the
southern states of the US where the climate is sub-tropical and subject to prolonged
periods of high ambient temperature and humidity. Genetic selection for heat tolerance
is an attractive alternative for reducing the effects of heat stress on animal performance.
Our first goal was to estimate variance components of Milk yield (MY), Fat yield (FY),
Protein yield (PY), Conception per insemination (CI) and Somatic cell score (SCS)
across lactations considering heat stress. Our second goal was to reveal genes and
pathways responsible for thermotolerance. Multi-trait repeatability test-day models with
random regressions on THI values were used to estimate variance components. The
models included herd-test-day and DIM classes as fixed effects, and regular and heat
tolerance additive and permanent environmental as random effects. Interestingly,
genetic variance for all traits under study increased across parities, suggesting that
cows become more sensitive to heat stress as they age. In addition, our study revealed
negative genetic correlations of MY, FY, PY and CI and positive genetic correlation of
SCS between general merit and thermotolerance. Whole-genome scans and gene-set
analyses were carried out to identify genomic regions, individual genes and pathways

11
responsible for thermotolerance. Interestingly, some significant regions harbor strong
candidate genes for thermoregulation, such as HSF1. The gene-set analysis revealed
several functional categories, such as cellular response to heat, response to DNA
damage stimulus, protein refolding that are involved in biological processes closely
related with thermotolerance. Overall, this study contributes to a better understanding of
the genetics underlying heat stress and points out novel opportunities for improving
thermotolerance in dairy cattle.

12
CHAPTER 1 INTRODUCTION
Heat stress alters a variety of physiological functions affecting production,
reproduction and health traits in dairy cattle thereby causing huge economic losses to
the dairy industry. Heat stress is likely to become more prevalent in years to come as
global climate change is expected to increase the frequency and duration of heat-stress
related events. Dairy cows are more susceptible to heat stress because of higher
metabolic heat production. Heat stress has been a huge issue in dairy farming
especially in the southern states of the US where climate is sub-tropical and subject to
prolonged periods of high ambient temperature and humidity. As a result, heat load in
the cow increases to the point that the body temperature rises, intake declines, milk
yield reduces, and health problem increases (Kadzere et al., 2002). In 2010, heat stress
estimates the loss of annual milk production for average US dairy by about $39,000
accounting for total $1.2 billion loss in milk production for the US dairy sector (Key and
Sneeringer, 2014). Hence, adopting strategies to alleviate heat stress and restore cow’s
health, production and reproduction efficiency is necessary for improving profitability of
the dairy farm. Over several years, heat stress has been defined as the function of
temperature -humidity index. As a result, different approaches like physical
modifications of the environment and improved nutritional and management practices
have been used to alleviate the effects of heat stress in dairy cows. However, these
practices increase production costs, and in general, they cannot eliminate heat stress
completely. One complementary strategy for reducing the effects of heat stress on dairy
cattle performance is the identification and subsequent selection of animals that are
genetically more heat tolerant. Identification of such heat tolerant animal can be made

13
based on the measurement of their immediate response like rectal temperature,
respiration rate among others. However, it is not feasible to use these records in
national evaluation system because collection of such records at a national level would
be time consuming and labor intensive. Alternatively, decline in performance due to
heat stress can be used as an indicator trait of heat tolerance (Ravagnolo et al., 2000).
The animal with minimum decline of performance per degree increase of temperature is
identified as heat tolerant. Dry bulb temperatures combined with humidity in an index
are used to assess the impacts of heat stress on dairy cattle performance. These
climatic variables are available from public weather stations. Meteorological data from
nearest weather stations can describe environmental conditions of farms (Freitas et al.,
2006). Additionally, since this methodology does not require any additional
measurements (e.g., body temperature), it can be applied to datasets as large as those
utilized for the national evaluation.
The main objective of this research proposal was to perform a comprehensive
genomic analysis of heat tolerance in dairy cattle. Production, reproduction and health
records, climatic data, and genome-wide dense single nucleotide polymorphism
markers were jointly analyzed using test-day models that include a random regression
on a heat stress function based on THI values. This strategy allowed us to identify and
characterize genomic regions, and individual genes and pathways, responsible for heat
tolerance in dairy cattle. The relevant information thus generated in this study will help
us better understand genetics underlying heat stress and subsequent use of marker-
assisted selection in commercial breeding schemes.

14
CHAPTER 2 LITERATURE REVIEW
Cattle, being homeothermic animal, try to maintain relatively constant core body
temperature regardless of changes in environmental temperature. The normal body
temperature of cattle is around 38.6°C. It is important to note that the core body
temperature of cattle is generally higher than the peripheral because of extensive
metabolic heat production by internal organs. In cows, body temperature also vary with
the stage of lactation, amount of milk production, nutritional and health status, stage of
the estrous cycle and genetic diversity within a population (Wrenn et al., 1961, Shearer
and Beede, 1990). Therefore, to maintain constant core body temperature, cows
typically utilize different behavioral, physiological and immunological adaptations.
Several physiological variables such as rectal temperature, respiratory rate, and
hormone concentrations have been used to predict body temperature in dairy cattle. For
instance, when rectal temperature of the cow is greater than 39°C, it is very likely that
the cow undergoes several physiological adaptations that compromises both
performance and well-being. Curtis (1983) reported that rectal temperature is not an
accurate indicator of body temperature in dairy cattle as there is a time lag between
rectal temperature and body temperature. Respiration rate (RR) has also been used as
an indicator of body temperature in dairy cows. However, RR varies according to the
animal conditions, prior exposure to hot conditions, ambient environment conditions and
cooling strategies. Similarly, there is also a time lag of two to four hours between body
temperature and RR (Gaughan et al., 2000). High environmental temperature also
reduces the plasma concentration of thyroid hormones in the cattle. However, it should
be considered that increase in ambient temperature is accompanied with decrease in

15
feed intake and reduction in hormone concentrations. Therefore, decrease in feed
intake should be considered as a confounding factor to correlate high temperature with
low hormone concentration (Saber et al., 2009).
Body temperature can also be assessed by measuring meteorological variables
such as temperature and humidity. Temperature Humidity Index (THI) formed by
combination of temperature and humidity is used to predict potential heat stress
conditions. These climatic variables are available from nearest public weather stations
which can accurately describe environmental conditions of farms (Freitas et al., 2006).
Additionally, this methodology has been used for genetic evaluation of heat tolerance in
dairy cattle (Ravagnolo et al., 2000, Aguilar et al., 2009, Sanchez et al., 2009). Since
this methodology does not require any additional measurements (e.g., body
temperature), it can be applied to datasets as large as those utilized for national
evaluations.
Thermoneutral Zone
Thermoneutral zone (TNZ) is a range of environmental temperature in which
change in ambient temperature does not lead to change in animal heat production. In
TNZ, animal is experiencing basal metabolic rate. As a result, animals can regulate
temperature by non- evaporative physical process alone. Generally, temperature range
of TNZ varies with age, species, feed intake, physiological status, behavioral and
immunological conditions of the animal (Yousef, 1985). TNZ is bounded by lower critical
temperature (LCT) at the lower end, and upper critical temperature (UCT) at the upper
end. LCT is defined as the environmental temperature below which homeotherms must
increase metabolic heat production to maintain constant core body temperature. As air
temperature drops below the lower critical temperature, animals increase their

16
metabolic heat production by increasing feed intake, shivering and activating the brown
adipose tissue (Carstens, 1994).
The upper end of the thermoneutral zone is the upper critical temperature (UCT)
which is defined as the ambient temperature above which animals must decrease heat
production to maintain thermal balance. The estimates of UCT for cows producing 30 kg
milk per day range from 12 to 24°C (Berman and Meltzer, 1973). When air temperature
exceeds the UCT, the heat gradient reduces, as a result, the potential for non-
evaporative heat loss is reduced and animals depend on evaporative cooling such as
sweating and panting to dissipate any excess heat received from the environment or
generated by metabolism (McArthur and Clark, 1988). Above UCT, an increase in body
temperature negatively influences animal performance and well-being as the cow enters
heat stress conditions.
Heat Stress
Stress is the sum total of the forces outside the bodily system that acts to
displace the system from the ground state (Yousef, 1985). Heat stress is a combination
of air temperature, humidity, wind speed, solar radiation and other environment
variables that makes it difficult for the cow to lose bodily heat to environment thereby
increasing body temperature. Strain is an internal displacement of the bodily system
(Yousef, 1985). Strain from heat stress is in fact, a change in body temperature in the
animal that can be easily measured. Heat stress is an important economic issue in dairy
farming that negatively impacts the performance and health of livestock. In dairy cows,
heat stress decreases milk yield, reduces milk quality, and depresses fertility. Economic
losses due to heat stress are estimated to be between $897 and $1500 million per year
for the US dairy industry (St-Pierre et al., 2003). Since heat stress is an important

17
economic issue in dairy farming, it has been extensively reviewed by several authors
(Kadzere et al., 2002, St-Pierre et al., 2003, West, 2003, Garcia-Ispierto et al., 2006).
Responses to Heat Stress
Adaptation is the animal response to reduce the magnitude of physiological strain
caused by heat stress. Most of the adaptations responding to heat stress involve
increasing heat transfer to the environment and reducing the production of metabolic
heat (Kadzere et al., 2002). However, adaptive response themselves reduce
physiological functions thereby compromising performance. For instance, during heat
stress, milk production decreases because of the adaptive responses to prevent rise in
body temperature. One of the physiological adaptive responses to regulate body
temperature during heat stress include reduction in feed intake. When cows eat less,
less of the energy will be available for milk production thereby causing reduce in milk
production (West, 2003). Another reason why performance suffers during heat stress is
failure to successfully adapt to high temperature during heat stress thereby disrupting
physiological functions. One example could be reduced fertility during heat stress owing
to the effects of elevated temperature on oocyte and embryo development (Hansen,
2009). Thus, maintenance of homeothermy is critical for both performance and well-
being of the cow.
Methods of Heat Transfer
Homeotherms attempt to balance heat production and heat loss. Heat stress
occurs when sum of heat produced from metabolic heat and heat received from the
environment exceeds heat lost to the environment. Heat is exchanged between the cow
and the environment via two types of mechanisms, viz., sensible heat loss mechanism
and latent heat loss mechanism. Sensible heat loss mechanisms, which includes

18
conduction, convection, and radiation, depend on temperature gradient between animal
surface and environment on which animal is exchanging heat. Sensible heat loss
reduces as the temperature gradient between environment and cow’s surface
temperature decreases (Maia et al., 2005). After sensible heat loss mechanisms
become ineffective, the latent heat loss becomes effective. Latent heat loss mechanism,
which includes evaporation and condensation, occurs along the vapor gradient between
the animal surface and the environmental vapor pressure. When the environmental
humidity is high, little heat is lost by evaporative cooling. Therefore, when cows are
exposed to high air temperature coupled with high humidity, it becomes increasing
difficult for the cow to lose heat to the environment thereby building increasing heat load
in the body.
Conduction and Convection
Conduction refers to physical transfer of heat from warm object to cool object
without any object moving. Examples are from cow feet to floor. Cows undergo
physiological adaptations to regulate conductive heat loss. During heat stress, cows
increase blood flow from core to the periphery, thereby increasing surface temperature.
This is possible because cows have special adaptations in blood vessels that help
increase blood flow to the skin. During periods of heat stress conditions, when body
temperature rises, anastomosis between arterioles and venules open thereby allowing
rapid flow of blood from core to skin (Walloe, 2016). If exists a temperature gradient
between skin surface and environment, then heat lost occurs to the environment.
Convective heat exchange refers to transfer of heat from warm substance to cool
substance where the substances are moving past each other. Examples are from cow
to wind. Like conductive heat loss, cows can also regulate convective heat loss through

19
behavioral modification, posture, orientation and by increasing the convective coefficient
of wind around animal body (Kadzere et al., 2002).
Radiation
The radiant energy from the sun is solar radiation. The heat exchange with solar
radiation refers to production and absorption of electromagnetic radiation by animal
surface when exposed to sunlight. The amount of heat absorbed by an animal exposed
to direct sunlight depends on the absorptivity of the surface. The absorptivity of the
white-coat cow is 0.50 while the black coat-cow is 0.90 (Shearer and Beede, 1990)
which means net transfer of heat via solar radiation is 1.8 times higher for black coat
cows as compared to white coat cows when exposed to sunlight. Radiation is another
important medium of heat loss especially during night when the animal radiates heat to
the cooler sky. However, high humidity and clouds reduce heat loss by radiation
(Fuquay, 1981). When there is no cloud in the sky during night, the temperature
gradient between outer sky and the animal surface is greater, allowing large flow of heat
from animal surface to outer environment. However, in a cloudy night, a cow outside at
night will lose less heat because clouds have a warmer temperature than objects in the
outer space.
Evaporative Heat Loss
Evaporative heat loss is an important avenue for heat exchange particularly
when the temperature gradient between cows’ surface and air temperature falls.
Evaporative heat loss occurs when dew point temperature of the air surrounding animal
is lower than evaporative surface temperature of the animal (West, 2003). Increased air
speed and low humidity facilitates evaporative heat loss. When air temperature is about

20
40 °C, approximately 84% of the total heat loss is through evaporative heat loss, such
as sweating (Yousef, 1985).
Physiological Responses of Cattle to Heat Stress
Cows have TNZ within which no additional energy above maintenance is needed
to regulate the body temperature. Hyperthermia results from a negative energy balance
between heat produced and absorbed by the animal with respect to heat lost to the
environment. During heat stress, animals undergo several behavioral and physiological
changes from sub-cellular to the whole animal level. All these behavioral and
physiological responses involve the expenditure of energy to remove or reduce the
impacts of heat stress. Sweating, high respiration rate, vasodilation with increased
blood flow to skin surface, reduced metabolic rate, decreased dry matter intake, and
altered water metabolism are the physiological responses animals typically undergo
during heat stress. Most of these adaptations have in general negative impacts on
animal performance, health, and well-being (Shearer and Beede, 1990).
Reduced Feed Intake
Feed intake starts to decline at ambient temperature of 25-26 °C and drops more
rapidly above 36 °C in dairy cows. Reduced feed intake has been identified as one of
the main causes of reduced milk production in dairy cows during heat stress conditions.
Heat stress stimulates the medial satiety center of hypothalamus which, in turn, inhibits
the lateral appetite center thereby declining both reduced dietary intake and milk
production (Albright and Alliston, 1971). Maust et al. (1972) reported a negative genetic
correlation between rectal temperature and feed intake which suggests that elevated
body temperature reduces feed intake. Similarly, Bouraoui et al. (2002) reported a

21
9.6% decrease in DMI when THI increases from 69 to 78. According to Baumgard et al.
(2013), about 50% decrease in MY is due to reduced feed intake.
Decreased Milk Production
Heat stress negatively impacts milk production. Several studies have reported
decline in milk production due to high environmental temperature (Ravagnolo and
Misztal, 2000, Aguilar et al., 2009, Sanchez et al., 2009). This may be because heat
stress has negative effects on energy balance and secretory functions of the udder
(Silanikove, 1992). For instance, Ravagnolo et al. (2000) reported loss of milk
production per cow per year around 0.2 kg per unit of THI greater than 72. West et al.
(2003) found a negative relationship between heat stress and milk production. However,
it is important to note that there is a lag effect between the onset of high temperature
and humidity on milk production. The lag could be due to reduction in feed intake, a
delay between intake and utilization of nutrients, and hormonal changes (West, 2003).
The stage of lactation at which cows’ experience heat stress also affects the
effect of heat stress on milk yield. Cows in mid lactation are most adversely affected,
cows in early lactation are least affected, while those in late lactation are intermediately
affected. Under mild conditions of heat stress, Abeni et al. (2007) reported that heat
stress reduces milk yield in early, mid and late stages of lactation by 13%, 24% and
16.5%, respectively. This could be because cows in early lactation produce highest MY
and decrease in feed intake due to heat stress is compensated with the catabolism of
body fat to replace deficits in energy intake. However, the effects of heat stress are
more severe in cows of mid lactation as there are the highest alterations of blood
parameters related with energy balance and enzyme activities. High-milk-producing
cows are more affected by heat stress compared to low-milk producing cows (Kadzere

22
et al., 2002). Multiple-parity cows are known to be more susceptible to heat stress than
first-parity cows (Aguilar et al., 2009). Similarly, heat stress during the dry period affects
milk production in the subsequent lactation, as it was reported by several authors (West,
2003, Avendano-Reyes et al., 2006, Tao et al., 2011).
Decreased Reproduction
Heat stress negatively impacts most aspects of the reproductive functions in
mammals. Heat stress depresses estrous behavior, delays oocyte maturation, reduces
conception rate, and retards early embryonic development and growth. Badinga et al.
(1993) reported that heat stress on the day of ovulation reduces the size of dominant
follicle during estrous cycle. Hansen and Arechiga (1999) reported that heat stress
reduces expression of estrous, resulting in less number of cows eligible for embryo
transfer during summer season. Ravagnolo and Misztal (2002) reported a negative
genetic correlation of -0.35 between non-return rate at 90 days (NR90) and THI,
suggesting an unfavorable relationship between heat tolerance and reproduction.
The deleterious effects of heat stress on reproductive functions could be due to
the results of hyperthermia affecting the functions or physiological adjustments made by
the heat-stressed animal to regulate body temperature (Hansen, 2009) Several studies
have reported that heat stress decreases oocyte competence for fertilization. Even if
high temperature does not affect oocyte competence for fertilization, the resultant
embryo develops very slowly and abnormally (Hansen, 2013). Many of the effects of
heat stress on oocyte and gametes are closely related with the production of reactive
oxygen species which affect the quality of the oocytes and embryos, fertilization and
pregnancy success.

23
Increased Water Intake
Heat stress conditions alter water intake in lactating dairy cows (Murphy et al.,
1983). Heat stressed cows increase water intake above the minimal need for
maintenance and metabolism. This is because cows must compensate for losses of
water from evaporative heat loss through sweating and panting during the periods of
heat stress. Also, cows lose large amount of potassium through sweating. So, mineral
salts containing potassium are recommended to be included in the feed of heat stressed
cows (Shearer and Beede, 1990).
Increased Respiration Rate
Cows undergo physiological adjustments during high ambient temperature,
including increased respiration rate (Coppock et al., 1982). In a study involving high-
producing dairy cows in heat-stress conditions, Berman et al. (1985) found that
respiratory rate is 50-60 breaths per minute higher than in a thermo-neutral
environment. Increase in respiration rate is associated with increase in evaporative heat
loss (e.g. panting) which helps regulate animal body temperature.
Changes in Blood Hormones Concentration
Secretion of hormones associated with metabolism and water balance are
altered during heat stress. The hormones associated with adaptation to heat stress
include prolactin (PRL), growth hormone (GH), thyroid hormones, glucocorticoids,
mineralocorticoids, acetacholamines and antidiuretic hormone (Farooq et al., 2010).
Giesecke (1985) reported an increase in the concentration of plasma PRL during
thermal stress in dairy cows. This is because PRL is involved in regulating water and
electrolyte demands of heat stressed cows. Growth hormone, a calorigenic hormone
released from anterior pituitary, is also altered during heat stress. The decreased

24
concentration of GH leads to less heat production, which helps regulate body
temperature during heat stress. The thyroid hormones triiodothyronine (T3) and
thyroxine (T4) are primary hormones involved in basal metabolic rate and their
concentrations are found to be reduced during heat stress (Purwanto et al., 1991). The
decline in thyroid hormones along with decline in GH has synergic effect to reduce heat
production and regulate thermal temperature in dairy cattle. There is an increase in the
concentration of glucocorticoid hormone due to activation of adrenocorticotropic
hormone releasing factor in the hypothalamus during heat stress (Farooq et al., 2010).
The glucocorticoids work as vasodilators, thus increasing blood flow from core to
periphery helping regulate body temperature. There is increased concentration of
catecholamine during heat stress (Farooq et al., 2010). Catecholamine is involved in
regulating sweat gland activity. Similarly, there is increased concentration of antidiuretic
hormone (ADH) during heat stress conditions in dairy cattle. Increased water loss
through evaporative cooling in the respiratory tract and skin invokes increases secretion
of ADH, which in turn reduces water loss and increases water intake.
Methods for Assessment of Heat Stress
Rectal Temperature
Rectal temperature is commonly used as indicator trait to predict core body
temperature in dairy cattle. During heat stress condition, rectal temperature rises above
a normal body temperature of 39°C (Bianca, 1965, Igono and Johnson, 1990).
However, there is a limitation in the use of rectal temperature for quantification of heat
stress in dairy cattle because rectal temperature rises slowly in response to rise in
temperature (Curtis, 1983). Moreover, the process of measuring rectal temperature
through thermometer is labor intensive, time consuming, and prone to errors such as

25
variability probe, and variation in animal handlers (West et al., 2003). Additionally,
restraining animals to collect rectal temperature may subject animals to further stress
that can alter temperature, therefore rectal temperature might not provide a very
accurate measure of body temperature in dairy cattle (Hahn et al., 1990, Prendiville et
al., 2003).
Milk Temperature
Milk temperature has also been used as an indicator of body temperature in dairy
cattle. Several studies have used milk temperature to monitor body temperature and
have reported correlations between milk temperature and body temperature between
0.78 and 0.99 (Fordham et al., 1984, Igono et al., 1987, West et al., 1990). However,
West (2003) reported that milk temperature is very sensitive to climatic variables at the
time of milk collection. Indeed, West (2003) found that the climatic variable having the
greatest influence on cow morning milk temperature is the current day minimum air
temperature, while evening milk temperature is most influenced by current day mean air
temperature. Moreover, it seems milk temperature is also sensitive to other variables.
For instance, West et al. (1990) reported that milk temperature was greater for cows
administered bST compared with controls, and Igono et al. (1990) observed higher milk
temperature in high-producing cows compared to low-producing cows
Respiration Rate
Respiration Rate (RR) has also be used as an indicator of heat stress in cattle.
RR can be measured by counting flank movement. Cows exposed to hot weather
conditions show increased respiration rate, a physiological adjustment to high ambient
temperature (Coppock et al., 1982). Berman et al. (1985) reported that respiratory
frequency increases above 50-60 breaths/minute at ambient temperature higher than 25

26
°C in dairy cows. Increase in RR during hot humid conditions is mainly due to increase
in relative humidity which reduces respiratory and surface evaporation thereby
increasing body temperature. It should be noted that RR can vary according to animal
conditions, prior exposures to hot conditions, any cooling strategies applied and other
environmental factors. Since there is a time lag between RR and body temperature, it is
recommended that RR measurements are made two to three hours prior to the hottest
time of the day (Gaughan et al., 2000).
Heat Shock Proteins (HSPs)
Heat shock proteins (HSPs) have been implicated in thermotolerance in cells. In
fact, HSPs are molecular chaperones that protect cells from thermal damage, prevent
protein denaturation and block apoptosis (Kampinga, 2006).Based on molecular weight,
HSPs are classified into five families viz., Hsp100, Hsp90, Hsp70, Hsp60 and the small
HSPs. These proteins are found to be produced in increased amount in response to
heat shock and are involved in folding, unfolding and refolding of proteins (Sorensen et
al., 2003). In livestock, the Hsp70 family has been extensively studied in relation to its
involvement in thermo-tolerance. The cellular response to heat stress is regulated at
the transcription level and controlled by inducible expression of Hsp70 genes. However,
the biosynthesis and chaperonin activity of Hsp70 requires a critical threshold level of
temperature above which Hsp70 gene will be induced (Somero, 2002). Hsp70 genes
prevent aggregation of misfolded proteins and facilitate folding of misfolded protein to
the native state (Mayer and Bukau, 2005).

27
Strategies for Reduction of Effects of Heat Stress
Several strategies have been commonly used to reduce the effects of heat stress
in dairy cattle, including physical modifications of the environment, nutritional strategies
and genetic development of heat resistant animals.
Physical Modifications of the Environment
Physical modification of the environment is a commonly used strategy to reduce
the effects of heat stress in dairy cows. This strategy aims to either modify the
environment to reduce the exposure of heat stress on cows (e.g. shading) or to
enhance heat exchange between cows and surrounding (e.g. fans, sprinkles). However,
the degree of abatement of heat stress varies with the climate and production level of
the cows. In a hot and dry climate, shading is a cost-effective solution for minimizing
heat stress in dairy cattle. Indeed, shading reduces direct solar radiation and helps
reduce heat load on the animals. Several studies have indicated that cows with access
to shade had lower increase in rectal temperature than those without access to shade
in hot climate (Fisher et al., 2008, Collier and Gebremedhin, 2015, Fournel et al., 2017).
Shaded environment also impacts production and reproduction performance of
dairy cows. Cows in shade generally produce 0.7 kg more milk than cows which are not
in shade and the conception rate generally increases from 25.3% in an unshaded cows
to 44.4% in a shaded cows (Romanponce et al., 1977).
The use of sprinklers combined with fans is another effective method to promote
heat loss during high temperature. Sprinkles produce droplets that wet the cow’s
surface. Fans, when used with sprinkles, cause evaporative cooling to take place on the
surface. The impact of sprinkles combined with fans was assessed on dairy cows in hot
conditions. In a study conducted by Strickland (1989) in Florida, sprinkles and fans

28
reduced the RR from 95 to 57, increased DMI from 17.8 to 19.1 kg, and increased milk
yield from 18.1 to 20.2 kg.
Nutritional Strategies for Managing Heat Stress in Dairy Cows
Nutritional management is another commonly used strategy to reduce the effects
of heat stress in dairy cows. Nutritional management includes changes in feeding
schedule, and ration composition. High quality forages with less fibers produce less
heat than high fiber forages during digestion (West, 1999). Therefore, it is necessary to
provide good quality forages during heat stress as cows during hot weather will reduce
feed intake. Additionally, it is also necessary to increase concentrate of certain minerals
supplements to compensate for the loss from the body during heat stress. For instance,
it is recommended that cows should be supplemented with 1.5-1.6% DM of potassium
and 0.5-0.6% DM of sodium to improve milk yield when cows are under heat stress
conditions. Feeding 150–200 g/cow/day of sodium bicarbonate during hot conditions
helps buffer rumen and improves the appetite of the dairy cows. Additionally, to
minimize heat generation, it is a good strategy that the greater part of the cows’ ration
should be fed during cooler period of the day (e.g., between 4.00 and 6.00 am, and 9.00
and 11.00 pm). Another strategy could be, instead of feeding large bulk of feed, provide
smaller amount of feed and increase the number of feedings per day (Beede and
Collier, 1986).
Genetic Development of Heat Resistant Animals
Improving milk production has been a major breeding goal in dairy cattle over
several decades. As a result, over the last fifty years, average milk production in US
dairy cattle increased by 5,997 kg with 56% of this increase due to genetic selection
(VanRaden, 2004). However, several studies have reported that intense selection for

29
production traits has led to decline in thermo-regulatory physiology of cows (Ravagnolo
et al 2002; Kadzere et al., 2002; Dikmen et al. 2012). High producing cows are
characterized by larger body size, consumption of more feed and production of more
metabolic heat. Moreover, thermoneutral zone of high producing cows has shifted to
lower temperature thereby making high producing cows more susceptible to heat stress.
Several studies have reported that heat tolerance is a heritable trait (Ravagnolo
and Misztal, 2000, Dikmen et al., 2012, Garner et al., 2017). So, genetic selection can
be utilized to increase heat tolerance in dairy cattle. Some studies have estimated the
heritability of indicator traits of heat tolerance like body temperature and respiration rate.
For instance, Dikmen et al. (2013) estimated the heritability of rectal temperature for US
Holsteins around 0.13 to 0.17, which allows selection for lower rectal temperature under
heat stress. Body temperature heritability estimates range from 0.11 to 0.68 (Mackinnon
et al., 1991, Burrow, 2001, Howard et al., 2014). Heritability estimates of respiration rate
range from 0.76 to 0.84 (Seath and Miller, 1947). The estimates of these indicator traits
for heat stress show that there is enough genetic variability to allow selection for heat
tolerance. There are also studies indicating genes underlying heat tolerance in dairy
cattle. For instance, Olson et al. (2003) reported slick hair gene in the bovine genome.
This gene can maintain body temperature at lower rates. Several studies have reported
the presence of heat shock genes as candidate genes for thermotolerance in dairy
cattle (Lee et al., 2006, Page et al., 2006, Xiong et al., 2013). These are molecular
chaperones that protects cells from thermal damage, prevents protein denaturation and
blocks apoptosis (Kampinga, 2006).

30
Using random regression models, Ravagnolo et al. (2000) estimated genetic
components of heat tolerance in dairy cattle at different values of THI. At THI ≥72, the
study estimated the drop of 0.2 kg/unit of THI for milk. The study also reported a
negative correlation (r = -0.35) between milk production and thermotolerance which
means selection for milk yield will make cows more susceptible to heat stress. The
study also estimated the breeding value (BV) under no heat stress condition and
another BV for performance under heat stress conditions. The application of this model
helps calculate simultaneously the genetic merit for performance and the genetic merit
for heat tolerance, and therefore help predict rankings of animals in various
environments with different climatic conditions.
Genetic Studies on Heat Stress
Different approaches including cooling, shading and nutrition are commonly used
to mitigate the effects of heat stress. However, these practices increase production
costs and in general, they cannot eliminate heat stress completely. One of the
complementary strategies of reducing the effects of heat stress is the selection of heat
tolerant sires. Several studies have reported the existence of additive genetic variability
for heat stress in dairy cattle (Ravagnolo et al., 2000, Aguilar et al., 2009, Sanchez et
al., 2009, Nguyen et al., 2016, Macciotta et al., 2017). Ngyuen et al.(2017) estimated
breeding value for heat tolerance in Australian dairy cattle which provides opportunity to
breed cows that are more tolerant to heat with less impact on milk production. Note that
selective breeding of dairy cattle for thermotolerance is permanent, cumulative, and
hence, the most cost-effective approach for mitigating the effects of heat stress in dairy
cattle.

31
Test-day models have been used for genetic evaluation of thermotolerance in
dairy cattle. Nguyen et al. (2017) used random regression test day models to estimate
genomic estimated breeding values (GEBV) for heat tolerance for milk, fat and protein
yield in Australian dairy cattle. The use of test-day models provide an opportunity to
accommodate different recording schemes, utilize incomplete lactation records and
account for time-dependent effects for each test-day (Swalve, 2000). Many studies in
dairy cattle have used repeatability test-day models in which the shape of the lactation
curve has been modeled either using fixed regression with fixed effects (Ptak and
Schaeffer, 1993) or with function of days in milk.
The actual measurements of heat load on an animal could be performed by
measuring rectal temperature, milk temperature or respiratory rate, but this information
is expensive and/or difficult to measure in large populations. On the other hand, a linear
regression of regularly recorded traits such as milk production, health traits and
reproductive traits on climatic data can be fitted to predict the relationship between
performance and weather conditions. Ravagnolo et al. (2000) proposed a model to
study genetic components of heat stress in dairy cattle by using performance data
augmented with public weather information. This model assumes that cow performance
is unaffected until a certain level of THI, and above that level the performance declines
linearly with increasing THI. Once THI for heat stress is estimated, a heat load function
can be calculated as HL = max (0, THI-THIT), where THI is the observed THI value for a
given day, THIT is a threshold of THI value, and HL is the heat load value. Once the
heat load is defined, it can be incorporated in reaction norm models for evaluating
genetic tolerance to heat stress. In reaction norm analysis, the weather data (THI) is

32
treated as continuous, and performance data are regressed on this continuous scale.
However, the decline in performance as indicated by the slope is different for different
cows. If the variation in the slope contains sizeable additive genetic components, then
selection for heat tolerant animals is possible.

33
CHAPTER 3 GENETIC ANALYSIS OF HEAT TOLERANCE FOR PRODUCTION TRAITS IN US
HOLSTEIN COWS
Background
Improving animal’s ability to cope with adverse environmental condition has been
a challenge in animal industry. Adverse environmental conditions, especially heat
stress, are important economic issues in dairy farming as they negatively impact the
production of dairy cows (Aguilar et al., 2010b, Nardone et al., 2010, Biffani et al.,
2016). During heat stress, dairy cows undergo several behavioral and physiological
changes from sub-cellular to whole animal level, thereby affecting production traits
(West, 2003, Nardone et al., 2010). Annual economic losses due to heat stress are
estimated between $897 and $1,500 million for the US dairy industry
(St-Pierre et al., 2003). Since heat stress is an important economic issue in dairy
farming, different approaches including cooling, shading and nutrition are commonly
used to mitigate the effects of heat stress. However, these practices increase
production costs and in general, they cannot eliminate heat stress completely. One of
the complementary strategies of reducing the effects of heat stress is the selection of
heat tolerant animals. Some indicator traits of response to heat stress such as rectal
temperature and respiration rate exhibit a sizable genetic component. For instance,
Dikmen et al. (2013) estimated heritability of rectal temperature in US Holsteins around
0.13 to 0.17. Similarly, heritability estimates of RR range from 0.76 to 0.84 (Seath and
Miller, 1947). Heritability estimates of these indicator traits suggest that genetic
selection for thermotolerance is feasible. However, the inclusion of these indicator traits
in a national genetic evaluation is both expensive and time consuming.

34
An alternative methodology to evaluate heat tolerance is to examine changes in
performance over environmental variables (Ravagnolo et al., 2000). In genetic
evaluation of heat stress, the environmental variable is often expressed as
Temperature-Humidity Index (THI), a combination of temperature and humidity,
obtained from nearest public weather stations. In this model, a linear regression of
regularly recorded traits such as MY, FY and PY on climatic data is fitted to predict the
relationship between production and weather conditions. This model assumes that
production is unaffected until a certain level of THI, and above that level the production
declines linearly with increasing THI. For genetic evaluation of heat stress, several
studies have assumed that all animals have a common threshold above which
production declines linearly (Ravagnolo and Misztal, 2000, Aguilar et al., 2009,
Macciotta et al., 2017). Sanchez et al. (2009) estimated however, a separate threshold
and slope for each animal and reported a strong genetic correlation of 0.95 between
these two components, indicating that animals with higher threshold of heat stress
would also have lower rate of performance decline and vice-versa. Therefore, a simple
model assuming constant threshold and different slopes would suffice for genetic
evaluation of heat stress in dairy cattle.
The identification of genomic regions, and preferably individual genes and
pathways, responsible for genetic variation of thermotolerance will enhance
understanding of biological mechanisms and point out novel opportunities for improving
thermotolerance through selective breeding. However, there is a limited information on
genes and genomic regions associated with thermotolerance. Dikmen et al. (2013)
reported a genomic region on BTA24 harbors potential candidate genes for regulating

35
rectal temperature in dairy cows. Macciotta et al. (2017) reported a genomic region on
BTA26 that harbors a potential candidate gene BTRC known to be associated with
thermotolerance for MY.
The first objective of this study was to estimate variance components of MY, FY
and PY across lactations using multi trait repeatability test-day models considering heat
stress. Test-day records of MY, FY and PY were merged with daily THI values, and
were jointly analyzed using test-day models that include random regressions on a
function of THI values. The second objective of this study was to identify and
characterize genomic regions, and preferably individual genes and pathways,
underlying thermotolerance. For this, test-day records of MY, weather data, and
genome-wide SNP markers were jointly analyzed. As a result, this study will contribute
to better understanding of the genetics underlying heat stress and subsequent use of
marker-assisted selection in breeding program.
Materials and Methods
Phenotypic and Genotypic Data
Data comprised 254,215 MY, FY and PY test-day records on 17,522 Holstein
cows calved from 2006 through 2016 on two dairy farms viz., North Florida Holsteins
and Dairy Research Unit of University of Florida, USA. Lactation records were required
to have at least 8 test-days and be between 5 and 305 DIM. Pedigree file was created
by tracing the pedigree of cows back to five generations available at the Council on
Dairy Cattle Breeding website. The pedigree file included 35,006 animals.
Genotype data for 60,671 single nucleotide polymorphism (SNP) markers were
available for 4,776 cows with MY test-day records and 1,592 sires in the pedigree. The
SNP data were provided by the Council on Dairy Cattle Breeding and the Cooperative

36
Dairy DNA Repository. Those SNP markers that mapped to sex chromosomes or were
monomorphic or had minor allele frequency less than 1% were removed from the
dataset. After data editing, a total of 57,954 single nucleotide polymorphisms (SNPs)
were retained for subsequent genomic analyses.
Weather data were obtained from Florida Automated Weather Network for
Alachua county and hourly THI was calculated as proposed by Ravagnolo and Misztal
(2000) as:
THI = (1.8 × temp + 32) −(0.55−0.55 × rh ) x (1.8 × temp − 26) (2-1)
where temp is the temperature in degree Celsius and rh is the relative humidity in
percentage. Mean daily THI of 3 days prior the test day was assigned to each test-day
record as suggested by Bohmanova et al.(2007).
A function of THI (f(THI)) was created to estimate reduction in MY due to heat
stress. Therefore,
f(THI) = {if THI ≤ THIthr. 0if THI > THIthr. THI − THIthr.
(2-2)
where the value of THIthr. was set to 68 and thus f(THI) was equal to
max (0, THI - THIthr.)
Statistical Model
The following multi-trait repeatability test-day model was used to estimate the
variance components of production traits, considering multiple lactations as different
traits:
yklmn = HTDkl + DIMm + anl + pnl + vnl[f(THI)] + qnl [f(THI)] + eklmn
(2-3)

37
where yklmn is the record for MY, FY, and PY, HTDkl is herd-test-day k within parity l ( l =
1, 2,3), DIMm is the mth DIM class with classes defined every 20 days, anl is the general
random additive genetic effect of animal n in parity l , f(THI) is the heat stress function
for herd test day k , vnl is the random additive genetic effect of heat tolerance for the
animal n in parity l, pnl is the random permanent environmental effect of cow n in parity l,
𝑞𝑛𝑙 is the random permanent environmental effect of heat tolerance of the cow n in
parity l and eklmn is the random residual effect.
Let a= [𝑎′𝑛𝑙 𝑣′𝑛𝑙] be vector of random additive genetic effects and p = [𝑝′𝑛𝑙 𝑞′𝑛𝑙] be a
vector of random permanent environment effects for parities n = 1 to 3.
The variance- covariance structure was
V [𝑎𝑝𝑒
] = ⌈𝐴 ⊗ 𝐺 0 0
0 𝐼 ⊗ 𝑃 00 0 𝐼 ⊗ 𝑅
⌉ (2-4)
where A is the numerator relationship matrix, and G and P are 6x6 matrices of
(co)variances for additive and permanent environment effects respectively. R is a 3x3
diagonal matrix of residual variances corresponding to each trait.
Statistical Analysis
Initial estimates for multiple trait analyses were obtained from Aguilar et al.
(2009). Variance component for MY, FY and PY using multi-trait repeatability test-day
models were estimated in Bayesian framework using GIBBS2F90 (Misztal et al., 2002).
Genomic data were not included for variance components estimation. Of a total of
500,000 samples, first 100,000 were discarded as burn-in, and every 100th sample was
retained to calculate variance components and posterior standard deviations.
Heritability for the generic merit of MY was estimated as

38
h2 = σa
2
σtotal2
(2-5)
Where
σtotal2 = σa
2 + σp2 + σe
2
(2-6)
And heritability for MY at heat stress level f(i) was calculated as
h2 = σa
2 + f(i)2σv2 + 2f(i)σav
σa2 + f(i)2σv
2 + 2f(i)σav + σp2 + f(i)2σq
2 + 2f(i)σpq + σe2
(2-7)
The genetic correlation between the generic and heat tolerance additive effects
was calculated as
corr [a,f(i)v] = f(i)σav
√σ𝑎2 ∗ 𝑓(𝑖)2𝜎𝑣
2
(2-8)
Genome Wide Association Mapping using ssGBLUP
The whole-genome association mapping was performed using single-step
genomic BLUP methodology (ssGBLUP). The ssGBLUP is indeed a classical BLUP but
it replaces the inverse of the pedigree relationship matrix (A-1) with the inverse of the
realized relationship matrix (H-1) that combines both pedigree and genomic information
(Aguilar et al., 2010a). The combined pedigree genomic relationship matrix H-1 was
calculated as follows
H-1 = A-1 + [0 00 𝐺−1 − 𝐴22
−1] (2-9)
where 𝐺−1 is the inverse of the genomic relationship matrix, and 𝐴22−1 is the inverse of
the numerator relationship matrix for genotyped animals. Here, 𝐺−1 has the dimension
of 6,368 x 6,368 which includes 4,776 cows with test-day service records and 1,592

39
sires in the pedigree. Similarly, A matrix has a dimension of 35,006 X 35,006 which is
based on a-five generation pedigree.
Candidate genomic regions associated with general merit and thermotolerance
additive merit for MY were identified based on the amount of genetic variance explained
by 2.0 Mb moving windows of adjacent SNPs. Given the genomic estimated breeding
values (GEBVs), the SNP effects can be estimated as ŝ = DZ′[ZDZ′]-1 âg, where ŝ is the
vector of SNP marker effects, D is a diagonal matrix of weights of SNPs, and âg is the
vector of GEBVs (Wang et al., 2012). The percentage of genetic variance explained by
a given 2.0 Mb of moving window of adjacent SNPs was then calculated as
𝑉𝑎𝑟𝑢𝑖
𝜎𝑢2 x 100 =
𝑣𝑎𝑟 ∑ 𝑍𝑗𝑆𝑗𝐵𝑗=1
𝜎𝑢2 x 100
(2-10)
where ui is the genetic value of the ith genomic region under consideration, B is the total
number of adjacent SNPs within 2.0 Mb region, and Sj is the marker effect of the jth SNP
within the ith region. All the ssGBLUP calculations were performed using BLUPF90
(Misztal et al., 2002).
Gene Set Analysis
The gene set analysis includes basically three different steps: i) assignment of
SNPs to genes ii): assignment of genes to pathways iii) Pathway-based association
analysis
Assignment of SNPs to genes
The UMD 3.1 bovine genome sequence assembly was used for SNP assignment
using the Bio Conductor R package biomaRt (Zimin et al., 2009). SNPs were assigned
to genes if they were located within the genomic sequence of the gene or at most 15 kb
either upstream or downstream the gene. If a SNP was found to be located within the

40
gene or at most 15kb either upstream or downstream the gene, that gene will be
included for further analysis. Finally, a gene was considered significant gene for MY, if
the gene is flagged by a top 1% SNP that explained the highest genetic variances for
MY.
Assignment of genes to pathways
Gene Ontology (GO) (Ashburner et al., 2000) and Medical Subject Headings
(MeSH) (Nelson et al., 2004) databases were used to define functional sets of genes.
Genes assigned to the same functional category are considered more closely related
rather than random sets of genes.
Pathway-based association analysis
The association of a give pathway with MY was analyzed using Fisher’s exact
test. This test, based on cumulative hypergeometric distribution, enables to search for
an overrepresentation of significant genes among all the genes in the pathway. The P-
value of observing 𝑘 significant genes in the pathway was calculated by
P-value = 1 - ∑(S
i )(N−Sm−i)
(Nm)
k−1i=0
(2-11)
where S is the total number of genes that were significantly associated with MY, N is the
total number of genes that were analyzed in the study, and m is the total number of
genes in the pathway (Abdalla et al., 2016). The GO gene set enrichment analysis was
performed using the R-package goseq (Young et al., 2010) whereas MeSH enrichment
analysis was carried out using the R-package meshr (Tsuyuzaki et al., 2015).

41
Results and Discussion
Genetic Dissection of Heat Stress for MY, FY and PY
Variance components of general production level (intercept) and animal ability to
respond to heat stress (slope) for MY, FY and PY using multi-trait repeatability test day
models (MREP) were estimated respectively (Table 2-1, Table 2-2, Table 2-3).
Similarly, additive genetic variances for thermotolerance and relevant genetic
parameters, namely heritability and genetic correlation at heat stress level ф (THI) = 78
(i.e. 10 units above THI threshold of 68) were also estimated.
Estimates of variance components for both generic and heat stress additive
effects were comparable to those estimated by Aguilar et al. (2009) for first three
parities in US Holsteins, however, the estimates reported in this study are higher than
those reported by Ravagnolo and Misztal (2000). Both generic and heat tolerance
additive genetic variances for MY, FY and PY increased across parity. Genetic
variances for MY under-heat stress increased by 8.31% from parity one to parity two
and 5.18 % from parity two to parity three suggesting that cows become more sensitive
to heat stress as they age. Additive genetic variances for thermotolerance increased
from first to second parity in both FY and PY. Increase from first to second parity were
more than 100% for FY and more than 16% for PY.
This is in agreement with the findings of Bernabucci et al. (2014) who also
reported that multiple-parity cows are more susceptible to heat stress than first parity
cows. Genetic correlations between generic and heat tolerance effects for MY in all
three lactations were negative, ranging from -0.30 to -0.55. Genetic correlations
between generic and heat tolerance effects for FY and PY in all three lactations were
also negative, ranging from -0.18 to -0.68. This is in agreement with the findings of

42
Aguilar et al. (2009) and Ravagnolo and Misztal (2000) in US Holsteins. Our findings
suggest that there is unfavorable genetic correlation between MY, FY and PY and cow
thermotolerance, and that continued selection for MY, FY and PY without consideration
for thermotolerance will result in greater susceptibility to heat stress.
Heritability estimates for MY at THI 78 were between 0.17 to 0.32 across
lactations, which is comparable with the heritability estimates reported by Ravagnolo et
al. (2000). Similarly, heritability estimates for FY for first three lactations were between
13 to 20 % whereas heritability estimates for PY across first three lactations were
between 18 to 26%. The estimates of heritability decreased across the parity. This could
be because phenotypic variances increase across the parity as cows become more
sensitive to heat stress in later lactations. Genetic correlations between parities for
general additive effect for MY were positive and high (≥0.82) whereas genetic
correlations between parities for additive thermotolerance were also positive but slightly
lower ranging from 0.61 to 0.78. For FY and PY, genetic correlations between parities
for general additive effect were also positive and high (≥0.76) whereas genetic
correlations between parities for heat tolerance additive effects were positive but slightly
lower, ranging between 0.34 to 0.78. The lower genetic correlation between parities for
heat tolerance additive effects could be due to interaction between parity and THI.
However, it is important to note that variances and genetic parameters estimated in this
study are in lower bounds as MREP ignore the dynamics of heat stress with respect to
function of DIM.
Whole-Genome Mapping for Thermotolerance Genes affecting MY
Single-step genomic BLUP methodology was utilized to perform GWAS of
thermotolerance for MY. The ssGBLUP methodology allows to identify genomic regions

43
that explain genetic variance for a trait of interest under consideration. The advantage of
ssGBLUP is it captures less noise and yields well-defined peaks across the entire
genome, however, with ssGBLUP, there is no statistical test to test the significance of a
genetic marker for a trait of interest.
The association analysis identified several regions in the bovine genome strongly
associated with general production level and animal ability to respond to heat stress.
Figure 2-1 displays Manhattan plots for MY across the three lactations under study; the
left plots show the results for general merit (intercept) while the right plots show the
results for thermotolerance (slope). The results are presented in terms of the proportion
of the genetic variance explained by 2.0 Mb SNP windows. For MY, as expected, two
different regions on BTA14 (14:1379063-3371507 and 14:3513577-5494654) harbor
genes with known relevant biological functions such as DGAT1, CPSF1, FABP4 across
all three parities. These two QTL regions on BTA14 explained together about 8.28, 5.28
and 4.47% of genetic variances across the first three parities, respectively. Gene
DGAT1 is a strong candidate gene for MY that encodes a key enzyme involved in the
synthesis of milk triglycerides (Cases et al., 1998). Knockdown of DGAT1 gene in
mouse reduces MY (Smith et al., 2000). Cochran et al. (2013) reported SNP located
within CPSF1 to be significantly associated with MY which is involved in 3’ end-
processing of pre-messenger RNAs into messenger RNAs. FABP4 regulates both
medium and long chain fatty acids in milk and is upregulated during lactation (Zhou et
al., 2015). Another 2.0 Mb SNP window on BTA20 (20:31051302-33048635) explained
substantial number of genetic variances for MY (1.07%). Interestingly, this region
harbors GHR, a growth hormone receptor known to have a major effect on milk yield

44
and milk composition (Blott et al., 2003, Viitala et al., 2006). GHR is implicated in lipid
and carbohydrate metabolism and plays a pivotal role in the growth hormone axis by
initiating and maintaining lactation (Parmentier et al., 1999).
The association study also identified several regions associated with
thermotolerance. Indeed, one region on BTA15 (15:75595790-77595636) was strongly
associated with thermotolerance across all three parities. This region harbors PEX16,
MAPK8IP1, CREB3L1, CRY2; all genes implicated in cellular response to heat stress.
MAPK8IP1 is involved in controlling cellular response to external stimuli such as heat
shock, which in turn increases transcription activity of many heat-stress target genes
including cell survival, cell proliferation, apoptosis and morphogenesis (Thompson et al.,
2001). Therefore, MAPK8IP1 helps repair DNA replication error and damage occurred
at high temperature. PEX16 is involved in cell membrane biosynthesis (Farr et al.,
2016). PEX16 thus plays an important role in protection against heat shock. CREB3L1
is implicated in endoplasmic reticulum (ER) stress response caused due to the
accumulation of misfolded protein and promotes cell survival during heat stress (Liu and
Chang, 2008, Greenwood et al., 2015). CRY2 is involved in thermal tolerance and
knockdown of CRY2 increased sensitivity to temperature (Sanchez-Bermejo et al.,
2015).
The QTL region on BTA22 (22:16637720-18628331) identified in the slope of first
lactation harbors potential candidate genes for heat stress, such as FANCD2. Gene
FANCD2 is involved in Hsp90-mediated regulation of DNA damage response (Oda et
al., 2007).

45
Another SNP window on BTA14 (14:1651311-3649589) harbors genes HSF1,
EEF1D, VPS28 and TONSL which are involved in heat stress response. HSF1 is
involved in regulating HSP gene expression during heat stress. Upon heat stimulus,
HSF1 binds promoters containing heat shock elements (HSE) and activates heat stress
target gene transcription (Calderwood et al., 2010). EEF1D directs DNA binding at heat
shock promoter elements (HSE) and regulates heat shock responsive genes through
association with heat shock transcription factors (Cui et al., 2016). VPS28 is a vacuolar
protein sorting gene involved in heat shock resistance and any mutation in VPS28 will
lead to heat-shock sensitivity (Jarolim et al., 2013). TONSL is involved in DNA repair
and maintenance of genome stability in the presence of DNA damaging stimulus such
as heat shock (Saredi et al., 2016).
Another QTL region on BTA15 (15:82479789-84450812) harbors potential
candidate genes for heat stress, such as GLYAT. Gene GLYAT activates HSE signaling
pathway and is known to involve in detoxification of xenobiotic acyl-CoA’s in mammalian
cells (Zhang et al., 2007).
Pathway-Based Association Analysis for MY
In this study, we complemented the whole-genome scan with a gene set
enrichment analysis to detect potential functional categories and molecular mechanisms
associated with heat tolerance. The first step of the pathway-based analysis was to
assign SNPs to genes. Of the 58,046 SNP markers evaluated in the analysis, a total of
27,488 SNPs was located within annotated genes or within 15kb upstream or
downstream from annotated genes. This set of SNPs defined a total of 17,238 genes
annotated in the UMD 3.1 bovine genome sequence assembly, which in turn were
evaluated for pathway analysis. Additionally, a subset of 344 of these 17,238 genes had

46
been flagged by at least one SNP whose effect was in the top 1% distribution and were
significantly associated with heat tolerance for MY. The second step in the analysis
was to assign 17,238 genes into pathways. We tested GO and MeSH categories using
a hypergeometric test (Fisher’s exact test).
Several GO terms showed significant overrepresentation of genes statistically
associated with thermotolerance for MY. Noticeably, some of these terms are directly
associated with heat tolerance, such as cellular response to heat (GO:00346), response
to temperature stimulus (GO:0009266), and cellular response to stress (GO:0080135).
These three categories are highly related in the GO hierarchy and had two significant
genes in common, HSF1 and MAPT. There is a well-known connection between heat
tolerance and HSF1. Upon heat stimulus, HSF1 activates heat stress target gene
transcription including molecular chaperones essential for recovery from cellular
damage. Molecular chaperones are involved in protein folding and protect cell against
heat stress (Luft et al., 2001). MAPT is involved in regulating and coordinating diverse
cellular responses to stress. Heat stress leads to protein damage thereby giving rise to
misfolded protein and cell death. MAPT initiates mechanisms to repress translation and
ameliorate misfolded protein accumulation thereby promoting cell survival under
stressful conditions (Parker et al., 2014). Furthermore, GO:0080135, cellular response
to stress, also includes gene GAB1, a gene involved in angiogenesis through mediating
angiogenic and survival signaling (Zhao et al., 2011). Development of peripheral blood
vessels will enhance blood flow from core to periphery thereby removing heat from core
and exchanging with the environment (Charkoudian, 2010).Two significant GO terms
are related to DNA repair mechanism during heat stress. These GO terms showed a

47
very close relationship in the GO hierarchy: response to DNA damage stimulus
(GO:2001020 and double strand break repair (GO:0006302). These highly related GO
terms had two genes in common, HSF1 and BABAM2. BABAM2 is anti-apoptotic and
positive regulator of DNA repair which promotes cell survival during heat stress.
Furthermore, many significant GO terms were associated with signal transduction, ion
transport and homeostasis, including regulation of calcium-mediated signaling
(GO:0050848), ion channel regulator activity (GO:0099106), glutamine family amino
acid catabolic process (GO:0009065) and alpha-amino acid catabolic process
(GO:1901606). The regulation of calcium and heat tolerance is well-documented.
Calcium can reverse the effects of heat implying protection against heat damage. The
term GO:0050848 is significantly enriched with genes FKBP1B and RCAN2. Gene
FKBP1B plays a role in protein folding thereby acting as a molecular chaperone and
preventing protein damage during heat stress (Calderwood, 2018). RCAN2 acts as a
molecular chaperone and regulates protein folding. Ion channel regulator activity
(GO:0099106) is significantly enriched with at least two genes PRKG1 and PRKCB.
These genes are key mediators of NO signaling pathway. NO is implicated in
maintaining cellular homeostasis during heat stress by acting as an antioxidant,
protecting membrane from damage and decreasing ROS levels (Parankusam et al.,
2017).
The term GO:0009065, Glutamine family amino acid catabolic process, is
significantly enriched with a gene called GLUD1 which plays a key role in cellular
protection during heat stress through enhancement of heat shock protein 70 (HSP70)
(Hamiel et al., 2009). Alpha-amino acid catabolic process (GO:1901606) is enriched

48
with a significant gene GCAT which has a role in production of glutathione synthesis.
Glutathione protects cells from death by removing free oxygen radicals produced during
heat shock (Yao et al., 2011).
Table 2-5 shows a panel of MeSH terms significantly enriched with genes
associated with heat tolerance of MY. Many of these terms are closely related with heat
stress such as HSP20 Heat-Shock Proteins (D050886), Heat-Shock Response
(D018869), Glutathione Transferase (D005982), Nitroso Compounds (D009603).
Additionally, functional categories including Inositol 1,4,5-Trisphosphate Receptors
(D053496), Calcium-Calmodulin-Dependent Protein Kinase Type (D054732) were also
detected as significant in the MeSH-informed enrichment analysis.
Summary
The results of this study reinforce the idea that there is a negative relationship
between production traits and thermotolerance. Therefore, continued selection for MY,
FY and PY without consideration of thermotolerance will result in greater susceptibility
to heat stress. In this study, we also performed GWAS and pathway-association
analysis with the purpose of understanding the genetic architecture underlying
thermotolerance. Genomic regions in BTA14, BTA15 and BTA22 were associated with
thermotolerance. Some of these genomic regions harbor genes that have been
implicated in cellular response to heat stress, such as HSF1, GLYAT, FANCD2.
Moreover, gene set analysis revealed several functional significant terms such as
response to heat conditions, response to DNA damage stimulus, and calcium-mediated
signaling. Most of these terms are directly implicated in biological process related with
thermotolerance. Overall, this study contributes to better understanding of the genetics

49
underlying heat stress and points out novel opportunities for improving thermotolerance
in dairy cattle.
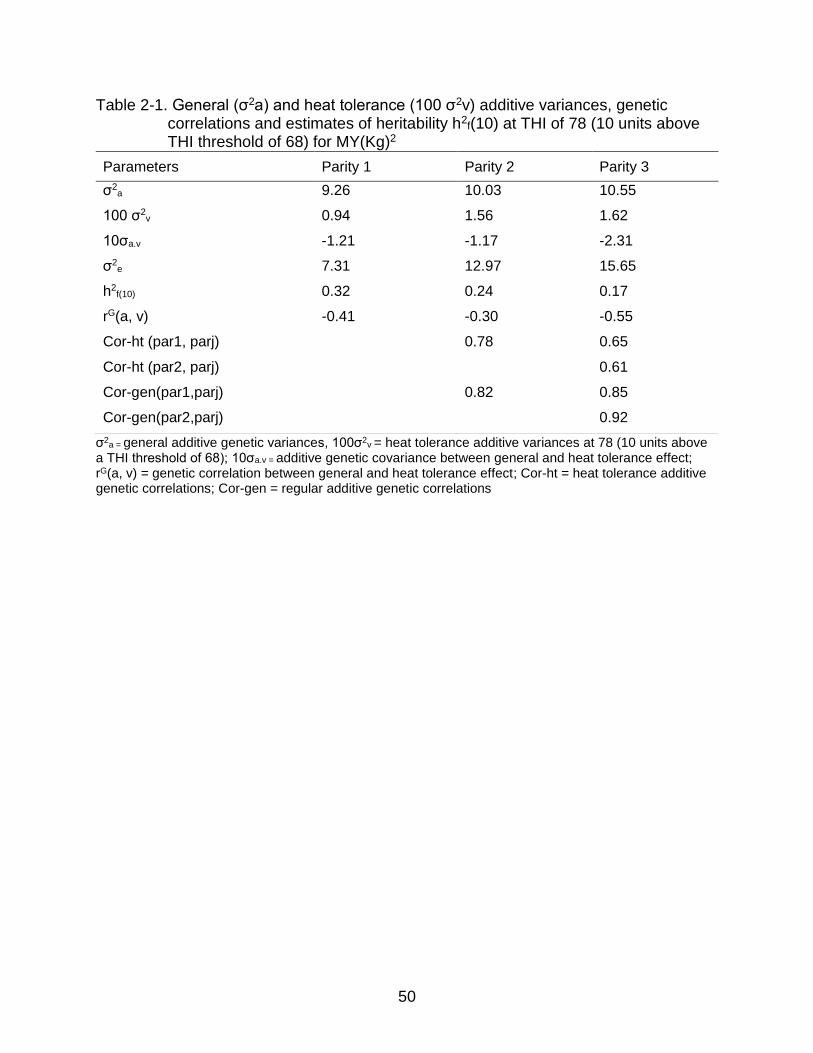
50
Table 2-1. General (σ2a) and heat tolerance (100 σ2v) additive variances, genetic correlations and estimates of heritability h2
f(10) at THI of 78 (10 units above THI threshold of 68) for MY(Kg)2
σ2a = general additive genetic variances, 100σ2
v = heat tolerance additive variances at 78 (10 units above a THI threshold of 68); 10σa.v = additive genetic covariance between general and heat tolerance effect; rG(a, v) = genetic correlation between general and heat tolerance effect; Cor-ht = heat tolerance additive genetic correlations; Cor-gen = regular additive genetic correlations
Parameters Parity 1 Parity 2 Parity 3
σ2a 9.26 10.03 10.55
100 σ2v 0.94 1.56 1.62
10σa.v -1.21 -1.17 -2.31
σ2e 7.31 12.97 15.65
h2f(10) 0.32 0.24 0.17
rG(a, v) -0.41 -0.30 -0.55
Cor-ht (par1, parj) 0.78 0.65
Cor-ht (par2, parj) 0.61
Cor-gen(par1,parj) 0.82 0.85
Cor-gen(par2,parj) 0.92

51
Table 2-2. General (σ2a) and heat tolerance (100σ2
v) additive variances, genetic correlations and estimates of heritability h2
f(10) at THI of 78 (10 units above THI threshold of 68) for FY (Kg x 100)2
σ2a = General additive genetic variances, 100σ2
v = heat tolerance additive variances at 78 (10 units above a THI threshold of 68); 10σa.v = additive genetic covariance between general and heat tolerance effect; rG (a, v) = genetic correlation between general and heat tolerance effect; Cor- ht = heat tolerance additive genetic correlations; Cor-gen = regular additive genetic correlations
Parameters Parity 1 Parity 2 Parity 3
σ2a 119.76 205.77 252.33
100 σ2v 18.189 48.614 33.78
10σa.v -11.44 -37.90 -63.03
σ2e 351.15 666.04 840.74
h2f(10) 0.20 0.17 0.13
rG(a, v) -0.25 -0.38 -0.68
Cor-ht (par1, parj) 0.46 0.34
Cor-ht (par2, parj) 0.38
Cor-gen (par1, parj) 0.91 0.95
Cor-gen (par2, parj) 0.95
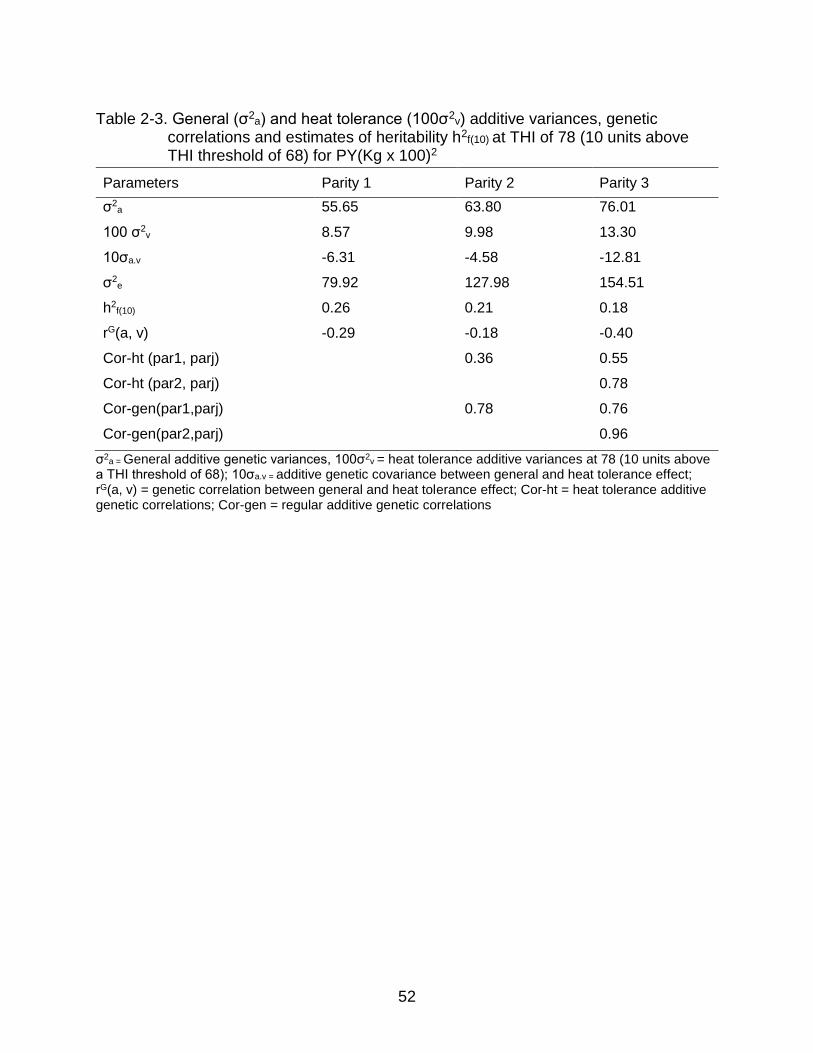
52
Table 2-3. General (σ2a) and heat tolerance (100σ2
v) additive variances, genetic correlations and estimates of heritability h2
f(10) at THI of 78 (10 units above THI threshold of 68) for PY(Kg x 100)2
σ2a = General additive genetic variances, 100σ2
v = heat tolerance additive variances at 78 (10 units above a THI threshold of 68); 10σa.v = additive genetic covariance between general and heat tolerance effect; rG(a, v) = genetic correlation between general and heat tolerance effect; Cor-ht = heat tolerance additive genetic correlations; Cor-gen = regular additive genetic correlations
Parameters Parity 1 Parity 2 Parity 3
σ2a 55.65 63.80 76.01
100 σ2v 8.57 9.98 13.30
10σa.v -6.31 -4.58 -12.81
σ2e 79.92 127.98 154.51
h2f(10) 0.26 0.21 0.18
rG(a, v) -0.29 -0.18 -0.40
Cor-ht (par1, parj) 0.36 0.55
Cor-ht (par2, parj) 0.78
Cor-gen(par1,parj) 0.78 0.76
Cor-gen(par2,parj) 0.96

53
Table 2-4. GO terms significantly enriched with genes associated with MY
Table 2-5. MeSH terms significantly enriched with genes associated with MY
GO ID Term No. genes
No. sig. genes P-value
0034605 Cellular response to heat 15 3 0.003
0009266 Response to temperature stimulus 33 3 0.027
0080135 Cellular response to stress 96 5 0.043
2001020 Response to DNA damage stimulus 35 3 0.032
0050848 Calcium-mediated signaling 15 3 0.003
0009065 Glutamine family amino acid catabolic
process 9 2 0.013
MeSH ID Term No. genes
No. sig. genes P-value
18869 Heat-Shock Response 9 2 0.015
50886 HSP20 Heat-Shock Proteins 2 1 0.004
05982 Glutathione Transferase 50 4 0.002
09603 Nitroso Compounds 2 1 0.004
53496 Inositol 1,4,5-Trisphosphate Receptors 17 2 0.040
54732 Calcium-Calmodulin-Dependent Protein
Kinase Type 2
13 2 0.002
51116 Receptors, Scavenger 2 1 0.004

54
Figure 2-1. Manhattan plot showing the results of genome-wide association mapping for Milk yield (MY): the left plots show the result of general merit and the right plots show the results of thermotolerance across lactations numbered vertically as lac1, lac2 and lac3.

55
CHAPTER 4 GENETIC ANALYSIS OF HEAT TOLERANCE FOR REPRODUCTION AND UDDER
HEALTH IN US HOLSTEIN COWS
Background
Heat stress depresses fertility and increases incidence of health disorders in
dairy cows. The effects of heat stress on fertility are mainly because of the failure of the
inseminated cows to establish and maintain pregnancy. Several studies have reported
that there is a marked decrease in conception per insemination during summer (Hansen
and Arechiga, 1999, de Vries and Risco, 2005, Huang et al., 2009). Indeed, de Vries
(2004) reported that the reduction in conception rate is around 53% during summer in
Florida. Reduction in conception rate is mainly because heat stress damages oocyte
and early embryo (Hansen, 2013). Embryo loss is 3.7 times more likely during summer
season as compared to winter season (Thatcher et al., 2001). Heat stress also has
negative effects on udder health, particularly the count of somatic cells is elevated in
response to high temperature. Also, high level of circulating stress hormones interferes
with immune system thereby increasing susceptibility of cows to clinical mastitis (Hagiya
et al., 2017). It is not surprise that in Florida, where more than 257 days have
Temperature-Humidity Index (THI) values greater than 68, heat stress has a huge toll in
both reproduction performance and health traits of dairy cattle (Ferreira et al., 2016).
Conception per insemination (CI) can be found as low as 10% to 20 % during summer
season (Hansen and Arechiga, 1999). Even with heat abatement systems, CI is found
to be lower during heat stress conditions (Hansen, 2009). Embryo transfer has been
considered as one of the most viable tools for improving fertility during heat stress.
However, farmers need to spend additional cost of $60 per cow for timed embryo
transfer as compared to $20 for conventional AI during summer months (de Vries,

56
2004). Moreover, it is known that during heat stress, the immune system is
compromised, which in turn makes cows more susceptible to infection, increasing
markedly the number of cases of clinical mastitis during summer (Kadzere et al., 2002).
Economic loss estimates about $179 per case of clinical mastitis. Since heat stress is a
huge economic problem in the dairy industry, especially in the southeast, various
strategies such as cooling, shading and nutrition have been used to alleviate effects of
heat stress and improve performance and well-being in dairy cows. Nonetheless, there
is a clear decline in fertility and udder health in dairy cows in hot and tropical climates.
In this context, genetic selection of heat tolerance for CI and SCS is an attractive
alternative for reducing effects of heat stress and subsequently improving fertility and
udder health in dairy cattle.
A growing body of literature suggests that there exists an additive genetic
components of heat tolerance for reproduction and udder health traits in dairy cattle
(Ravagnolo and Misztal, 2002, Aguilar et al., 2009). Indicator traits of heat stress like
respiration rate, body temperature have moderate to strong heritability. Estimates of
heritability for rectal temperature during heat stress is 0.17 which suggests genetic
improvement of heat tolerance through genetic selection is possible (Dikmen et al.,
2012). Ravagnolo and Misztal (2002) estimated heritability for non-return rate of estrous
during heat stress condition (THI = 70) and reported heritability estimate increased
considerably from 0.06% at 45 days to 5.3% at 90 days. However, there exists a
negative genetic correlation between heat tolerance and fertility trait. In the same study,
Ravagnolo and Misztal (2002) reported a negative genetic correlation of
-0.25, -0.77 and -0.95 between general merit and heat tolerance estimated at 45, 60

57
and 90 days respectively after insemination. Regarding udder health, Hagiya et al.
(2017) reported heritability estimates for SCS under heat stress conditions in the range
of 0.18 to 0.20 across lactations. Ravagnolo and Misztal (2002) presented a
methodology for genetic evaluation of heat tolerance by combining test day records with
public weather station records. Freitas et al. (2006) reported that information from
weather stations can be as accurate as that of recorded in farm, and hence, it can be
used to dissect the genetics underlying thermotolerance. In this methodology, test day
records and daily THI values are jointly analyzed using test-day models that include
random regressions on THI values.
Although traditional selection methods could be used to identify and select
thermotolerant cows, genetic gains for fertility and health traits could be accelerated if
the genes and pathways responsible for thermotolerance are identified and then used in
marker-assisted selection in commercial breeding schemes. As such, the first objective
of this study was to estimate variance components for CI and SCS across lactations
using multi trait repeatability test-day models considering heat stress. The second
objective of the study was to identify and characterize genomic regions, individual
genes and biological processes responsible for the variation in thermotolerance
underlying fertility and udder health. Pregnancy and SCS test-day records, weather
data, and genome-wide SNP markers were jointly analyzed. This relevant information
can contribute to a better understanding of the genetics underlying heat stress and
provide novel opportunities for use of marker-assisted selection in dairy cattle breeding
schemes.

58
Materials and Methods
Phenotypic and Genotypic Data
Conception per Insemination (CI) is one of the important fertility traits in dairy
cattle, which is defined as the ability of cow to establish and maintain pregnancy
provided cow is inseminated at the time of ovulation (Averill et al., 2004). Somatic cell
score (SCS) is an indicator trait for clinical mastitis, and somatic cell counts increase
with the severity of udder infection incurring huge economic loss to the dairy industry
(Sahana et al., 2013). Our data comprised 74,221 CI records on 13,704 Holstein cows.
All available service records maintained between 5 and 400 DIM were used. For SCS,
data comprised 355,546 test-day records on 18,975 cows obtained between 5 and 305
DIM. Cows calved from 2006 through 2016 on two dairy farms in Florida, USA. A
complete pedigree file was created by tracing the pedigree of cows back to five
generations using information available at the Council on Dairy Cattle Breeding website
Genotype data for 60,671 single nucleotide polymorphism (SNP) markers were
available for 4,700 cows with phenotype records, and 1,592 sires in the pedigree. The
SNP data were provided by the Council on Dairy Cattle Breeding and the Cooperative
Dairy DNA Repository. Those SNP markers that mapped to sex chromosomes or were
monomorphic or had minor allele frequency less than 1% were removed from the
dataset. After data editing, a total of 57,954 single nucleotide polymorphisms (SNPs)
were retained for subsequent genomic analyses.
Weather data were obtained from Florida Automated Weather Network for
Alachua county and hourly THI values were calculated as proposed by Ravagnolo and
Misztal (2002) as

59
THI = (1.8 × temp +32) −(0.55−0.55 × rh ) x (1.8 × temp −26) (3-1)
where temp is the temperature in degree Celsius and rh is the relative humidity in
percentage.
A function of THI (f(THI)) was created to estimate effects of heat stress on CI and
SCS. Therefore,
f(THI) = {if THI ≤ THIthr. 0if THI > THIthr. THI − THIthr.
(3-2)
where the value of THIthr. was set to 68 and thus f(THI) was equal to max (0, THI –
THIthr).
Statistical Model
The following multi-trait repeatability test day model was used to estimate the
variance components of CI and SCS for heat tolerance, considering multiple lactations
as different traits.
yklmn = HTDkl + DIMm + anl + pnl + vnl[f(THI)] + qnl [f(THI)] + eklmn (3-3)
where yklmn is the record for CI and SCS, HTDkl is herd-test-day k within parity l ( l = 1,
2,3), DIMm is the mth DIM class ( m = 1 to 20 for CI and m = 1 to 16 for SCS), with
classes defined every 20 days, anl is the general random additive genetic effect of
animal n in parity l , f(THI) is the heat stress function for herd test day k , vnl is the
random additive genetic effect of heat tolerance for the animal n in parity l, pnl is the
random permanent environmental effect of cow n in parity l, qnl is the random
permanent environmental effect of heat tolerance of the cow n in parity l and 𝑒𝑘lmn is the
random residual effect.
Let a= [𝑎′𝑛𝑙 𝑣′𝑛𝑙] be vector of random additive genetic effects and p = [𝑝′𝑛𝑙 𝑞′𝑛𝑙]
be a vector of random permanent environment effects for parities n = 1 to 3

60
The variance- covariance structure was
V [𝑎𝑝𝑒
] = ⌈𝐴 ⊗ 𝐺 0 0
0 𝐼 ⊗ 𝑃 00 0 𝐼 ⊗ 𝑅
⌉ (3-4)
where A is the numerator relationship matrix and G and P are 6x6 matrices of
(co)variances for additive and permanent environment effects respectively. R is a 3x3
diagonal matrix of residual variances corresponding to each trait.
Statistical Analysis
Variance components were estimated in a Bayesian framework using
GIBBS2F90 (Misztal et al., 2002). Genomic data were not included for variance
component estimation. Of a total of 500,000 samples, first 100,000 were discarded as
burn-in, and every 100th sample was retained to calculate variance components and
posterior standard deviations of relevant genetic parameters.
Heritability was estimated as
h2 = σa
2
σtotal2
(3-5)
Where
σtotal2 = σa
2 + σp2 + σe
2 (3-6)
and heritability at heat stress level f(i) was calculated as
ℎ2 = σa
2 + f(i)2σv2 + 2f(i)σav
σa2 + f(i)2σv
2 + 2f(i)σav + σp2 + f(i)2σq
2 + 2f(i)σpq + σe2
(3-7)
The genetic correlation between the generic and heat tolerance additive effects was
calculated as

61
corr [𝑎,𝑓(𝑖)𝑣] = f(i)σav
√σa2∗ f(i)2σv
2
(3-8)
Genome-wide Association Mapping using ssGBLUP
The whole-genome association mapping was performed using single-step
genomic BLUP methodology (ssGBLUP). ssGBLUP is indeed a classical BLUP but here
pedigree relationship matrix (A-1) is replaced with the inverse of the realized relationship
matrix (H-1) that combines both pedigree and genomic information (Aguilar et al.,
2010a). The combined pedigree genomic relationship matrix H-1 is calculated as follows,
H-1 = A-1 + [0 00 𝐺−1 − 𝐴22
−1] (3-9)
where G−1 is the inverse of the genomic relationship matrix, and 𝐴22−1 is the inverse of
the numerator relationship matrix for genotyped animals. Here, G−1 has the dimension
of 6,362 x 6,362 which includes 4,700 cows with phenotypic records and 1,592 sires in
the pedigree. Similarly, A matrix has a dimension of 28,620 X 28,620 for CI and 31,412
X 31,412, based on a-five generation pedigree.
Candidate genomic regions associated with general merit and heat tolerance
additive merit for CI and SCS were identified based on the amount of genetic variance
explained by 2.0 Mb windows of SNPs. Given the genomic estimated breeding values
(GEBVs), the SNP effects can be estimated as ŝ = DZ′[ZDZ′]-1 âg, where ŝ is the vector
of SNP marker effects, D is a diagonal matrix of weights of SNPs, and âg is the vector
of GEBVs (Wang et al., 2012). The percentage of genetic variance explained by a given
2.0 Mb genomic region was then calculated as
𝑣𝑎𝑟𝑢𝑖
𝜎𝑢2 x 100 =
𝑣𝑎𝑟 ∑ 𝑍𝑗𝑆𝑗𝐵𝑗=1
𝜎𝑢2 x 100
(3-10)

62
where ui is the genetic value of the ith genomic region under consideration, B is the total
number of adjacent SNPs within the 2.0 Mb region, and Sj is the marker effect of the jth
SNP within the ith region. All the ssGBLUP calculations were performed using
BLUPF90 family of programs (Misztal et al., 2002)
Gene Set Analysis
The gene set analysis includes basically three different steps: i) assignment of
SNPs to genes ii): assignment of genes to pathways iii) pathway-based association
analysis
Assignment of SNPs to genes
The UMD 3.1 bovine genome sequence assembly was used for SNP assignment
using the Bio Conductor R package biomaRt (Zimin et al., 2009). SNPs were assigned
to genes if they were located within the genomic sequence of the gene or at most 15 kb
either upstream or downstream the gene. If a SNP was found to be located within the
gene or at most 15kb either upstream or downstream the gene, that gene will be
included for further analysis. Finally, a gene was considered significantly associated
with a trait, if the gene is flagged by a SNP in top 1% distribution.
Assignment of genes to pathways
Gene Ontology (GO) (Ashburner et al., 2000)and Medical Subject Headings
(MeSH) (Nelson et al., 2004) databases were used to define functional sets of genes.
Genes assigned to the same functional category are considered more closely related in
terms of biology rather than random sets of genes.
Pathway-based association analysis
The association of a give pathway with either CI or SCS was analyzed using
Fisher’s exact test which is based on the cumulative hypergeometric distribution. This

63
test enables to search for an overrepresentation of significantly associated genes. The
P-value of observing k significant genes in the pathway was calculated by
P-value = 1 - ∑(S
i )(N−Sm−i)
(Nm)
k−1i=0
(3-11)
where S is the total number of genes that were significantly associated with MY, N is the
total number of genes that were analyzed in the study, and m is the total number of
genes in the pathway (Abdalla et al., 2016). The GO gene set enrichment analysis was
performed using the R package goseq (Young et al., 2010) whereas MeSH enrichment
analysis was carried out using the R package meshr (Tsuyuzaki et al., 2015).
Results and Discussion
Genetic Dissection of Heat Stress for Conception and Udder Health
Variance components of general performance level (intercept) and animal ability
to respond under heat stress conditions (slope) for CI and SCS were estimated using
multi-trait repeatability test day models (MREP) (Table 3-1, 3-2). Similarly, heritability
and genetic correlations at heat stress level ф (THI) = 78 (i.e. 10 units above THI
threshold of 68) for the first three lactations were also estimated.
Variance Component Estimates for CI
Both generic and heat tolerance additive genetic variances for CI increased
across parity. Generic additive genetic variances ranged from 0.2% to 0.4% in first three
lactations. Genetic variances for CI under-heat stress increased between 0.5% to
0.15% in consecutive parities suggesting that heat stress has greater effects on
conception rates in later lactations. This finding agrees with the results of Schueller et
al. (2014) who reported that multiple-parity cows are more affected by heat stress than
monoparous cows. Genetic correlations between generic and heat tolerance effects for

64
CI were negative in all three lactations, ranging from -0.35 to -0.82. This agrees with the
findings of Dikmen et al. (2012) who also reported a negative genetic correlation
between rectal temperature and fertility. These results suggest that there is unfavorable
genetic correlation between CI and heat tolerance, and that continued selection for
successful pregnancy in thermoneutral conditions will depress fertility under heat stress
condition. Moreover, heritability estimates for CI at THI 78 were between 2 to 3%, which
is comparable with the heritability estimates reported by Dikmen et al. (2012). Genetic
correlations between parities for general additive effect for CI were positive and high
(≥0.58), whereas genetic correlations between parities for additive heat tolerances were
also positive but slightly lower ranging from 0.17 to 0.49.
Variance Component Estimates for SCS
Both general and thermotolerance additive genetic variances increased with
parity. This is in agreement with the previous study (Hammami et al., 2015). Heritability
estimates at THI 78 ranged from 0.10 to 0.15 across three parities, which are in the
range of heritability estimates reported by Santana et al. (2017) in Brazilian Holsteins.
Interestingly, genetic correlations between generic and thermotolerance were positive in
all three lactations, ranging from +0.10 to +0.43. This suggests that continued selection
for lower SCS, will also yield a positive selection response in the cow’s ability to
respond under heat stress conditions.
Genomic Scans for Thermotolerance Genes Affecting CI and SCS
Whole Genome association analysis for CI
The association analysis identified several regions in the bovine genome strongly
associated with CI under heat stress. Figure 3-1 displays Manhattan plots across three
lactations under study; the left plots show the results for general merit (intercept), while

65
the right plots show the results for CI under heat conditions (slope). The results are
presented in terms of the proportion of the genetic variance explained by non-
overlapping 2.0 Mb SNP windows.
Two different regions on BTA23 (23:3626219-5618724 and 23:10620999-
12579560) were found to be strongly associated with general merit for CI across all
three parities. These regions together explained almost 3% of the additive genetic
variances. Interestingly, these strongly associated regions harbor several putative
genes closely related to reproduction, including PADI6 and HCRTR2. PADI6 is a
maternal-effect gene that regulates mitochondrial activity in oocyte. Mitochondrial
activity in oocyte influences the competence of oocyte to fertilize and early embryonic
development. Mouse oocyte lacking PADI6 triggers intrinsic apoptotic pathway resulting
in early embryonic death following ovulation and fertilization (Fernandes et al., 2012).
Gene HCRTR2 binds the hypothalamic neuropeptides orexin A and orexin B which are
implicated in normal pregnancy. This orexin system is expressed in uterus, conceptus
and trophoblast and is involved in maternal recognition of pregnancy, implantation and
proliferation of endometrial epithelial cells (Smolinska et al., 2017). Two genomic
regions on BTA2 (2:134837645-136697236) and BTA12(12:14654029-16641931) were
also found associated with CI, particularly for first lactation cows. The region on BTA2
harbors gene RPA2 which is involved in DNA damage response in oocyte and embryos.
DNA damage leads to cell cycle arrest, which in turn compromises the chances of
embryo survival. RPA2 is involved in DNA replication, recombination and repair and
maintain cellular and long-term embryo viability through effects on genomic integrity
(Zheng et al., 2005). The region on BTA12, which explained more than 1% of the

66
genetic variance for CI, harbors several putative genes, including TPT1 which is
expressed in placenta and is implicated in calcium binding and homeostasis of
trophoblast cells. It is known that placental transfer of calcium is important for growth
and development of fetus (Arcuri et al., 2005). The association study also identified
several regions associated with thermotolerance. Indeed, one region on BTA10 (10:
80756063-82748913) explained 1.00% of genetic variance for CI under heat stress in
first lactation cows. This region harbors gene RGS6 which is involved in cellular
response to heat stress. Gene RGS6 is implicated in subcellular trafficking of proteins to
nucleoli in response to heat stress and prevents further damage of proteins from heat
stress (Chatterjee and Fisher, 2003). Another region on BTA11(11:27185742 -
29174959) explained 1.40% of the genetic variance for CI under heat stress in second
lactation cows. This region harbors PRKCE which is also involved in heat shock
response. The heat shock response through the activation of HSFs protects proteins by
stabilizing and refolding protein-folding intermediates (Morimoto et al., 1997).
Another region on BTA21 (21:15031948- 17028796) explained 1% of the genetic
variance for thermotolerance associated with CI. This region harbors KLHL25 which is
involved in protein ubiquitination (Zhang et al., 2016).
Whole Genome association analysis for SCS
The whole-genome scan identified several regions strongly associated with SCS
in both thermoneutral conditions and under heat stress. Figure 3-2 displays Manhattan
plots for SCS across the three lactations under study; the left plots show the results for
general merit (intercept), while the right plots show the results for SCS thermotolerance
(slope). The results are presented in terms of the proportion of the genetic variance
explained by 2.0 Mb SNP windows.

67
Two different regions on BTA29 (29:2294521-4291391 and 29:41755825-
43726898) were found to be strongly associated with SCS across all three parities.
These regions harbor FAT3 and OTUB1. Gene FAT3 is a calcium ion binding gene that
reduces calcium in blood. Reduction in blood calcium level impairs immunity, reduces
smooth muscle function in teat sphincter, leading to partial or incomplete closure of teat
canals. Lack of teat sphincter closure leads to entry of pathogens into teat canal and
hence increases risk of CM (Mulligan et al., 2006). Gene OTUB1 serves as a marker of
tumor and therefore could play a role in the development of tumors in mammary gland
(Liu et al., 2014). A 2.0 Mb SNP window on BTA6 (6:92648735-94627787) explained
0.54, 0.73 and 0.66% of the genetic variance for SCS for first, second, and third
lactation, respectively. This region harbors several genes, including vitamin D-binding
protein precursor (GC) and neuropeptide FF receptor 2 (NPFFR2), which are strong
candidates for udder health. The GC gene encodes Gc-globulin which transports
vitamin D to target cells such as monocytes. In presence of microbes in the mammary
gland, vitamin D stimulates monocyte antibacterial activity by increasing production of
antimicrobial peptides and enhances mechanism associated with autophagy (Hewison,
2011). Neuropeptide FF receptor 2 is a membrane protein which suppresses the
production of nitric oxide in the inflammation process and exhibit anti-inflammatory
activity (Sun et al., 2013). Besides, it plays a role in the proliferation of T cells and
modulation of immune responses (Minault et al., 1995).
Similarly, one region on BTA24 (24:60394194-62332550) explained 0.83, 0.64
and 0.69% of the genetic variances for SCS for parity one, two, and three, respectively.
This region harbors several genes including BCL2 which is an apoptotic gene

68
significantly upregulated in the mammary tissues challenged with Staphylococcus
aureus (Long et al., 2001). Another QTL region on BTA4 (4:60094523-62089794)
explained 0.52 and 0.54% of the genetic variances for parity two and three,
respectively. This region harbors a potential candidate gene, namely AOAH. This gene
is expressed in the neutrophils, and its activity decreases when dairy cows are
challenged with endotoxin and E. coli, resulting in the appearances of lipo-
polysaccharides in the blood which causes inflammation in mammary tissues (Mehrzad
et al., 2007).
The association study also identified regions associated with SCS under heat
stress conditions. One genomic region on BTA2 (2:24639094-26617396) was strongly
associated with thermotolerance in first two lactations. This region harbors two
candidate genes, DLX1 and DLX2 which downregulate cytokine signaling pathway and
prevent inflammation (Dulken et al., 2017). Another QTL region on BTA19
(19:42020856-44001678) explained 0.61% of thermotolerance genetic variances for
SCS in first lactation cows. This region harbors genes STAT5B and STAT5A which are
directly implicated in the heat stress response. Indeed, STAT5B and STAT5A are
involved in activating immune responses during heat stress. Knockout of Stat5a−/−
and Stat5b−/− in mouse exhibit decreased immunity thereby suggesting their role in
development of immune functions (Lin et al., 2012).
Pathway-Based Analysis
We complemented the whole-genome scans with alternative gene set
enrichment analyses to detect potential functional categories and molecular mechanism
associated with thermotolerance for conception and udder health. Of the 58,046 SNP
markers evaluated in the analysis, a total of 27,488 SNPs was located within annotated

69
genes or within 15kb upstream or downstream from annotated genes. This set of SNPs
defined a total of 17,235 genes annotated in the UMD 3.1 bovine genome sequence
assembly. A subset of 345 and 361 of these 17,235 genes were significantly associated
with CI and SCS, respectively. A gene was declared as significant if it had at least one
SNP in the top 1% of the SNP effect distribution.
Gene-set analysis for CI
Several GO terms showed significant overrepresentation of genes statistically
associated with CI under heat stress. Noticeably, some of these terms are directly
associated with heat response such as protein refolding (GO:0042026), inter-strand
cross link repair (GO:0036297), and regulation of intra-cellular protein transport
(GO:0033157). There is well-known connection between heat tolerance and protein
refolding. Interestingly, protein refolding (GO:0042026) is significantly enriched with a
gene HSPA1A which acts as a molecular chaperone and prevents unfolding and
degradation of protein during heat stress (Calderwood, 2018).Inter-strand cross link
repair is implicated in DNA replication, maintenance and repair which promotes cell-
cycle, growth and survival during heat stress (McMahon et al., 2016). Regulation of
intra-cellular protein transport is implicated in protein trafficking under cellular stress. In
response to heat stress, the nucleolus responds to cellular stress by sequestering and
releasing a variety of proteins that affect cell cycle and DNA repair (Nalabothula et al.,
2010).
Two significant GO terms are directly related with reproduction process: steroid
hormone mediated signaling pathway (GO:0043401) and lipid modification
(GO:0030258). Lipid modification greatly occurs during pregnancy which is important for
fetal growth and development. Two important lipid modifications viz., maternal

70
accumulation of fat and hyperlipidemia greatly benefits fetal growth particularly under
conditions of dietary intake (Herrera and Ortega-Senovilla, 2010).
Two relevant GO terms, namely germ cell development (GO:0007281) and post-
embryonic development (GO:0009791) also showed significant overrepresentation of
genes associated with conception under heat stress. Interestingly, post-embryonic
development includes the gene GATA3 which is selectively expressed in trophectoderm
of peri-implantation embryo and regulates morula to blastocyst transformation.
Knockdown of GATA3 in pre-implantation mouse embryo inhibits embryonic
development (Home et al., 2009).
Gene-set analysis for SCS
Several GO terms showed significant enrichment of genes statistically associated
with SCS under heat stress. Some of these terms are directly associated with heat
response such as chaperone mediated protein folding (GO:0061077). This term had
three significant genes, FKBP10, FKBP11 and TCP1. Genes FKBP10 and FKBP11 are
chaperones involved in protein folding and prevent the aggregation of unfolded protein
in ER at high temperature (Jeffries et al., 2014). TCP1 is another chaperone gene which
interacts with Hsp70 to assist protein folding and prevents aggregation of misfolded
protein during heat stress (Young et al., 2004). Another significant GO term for SCS is
DNA replication (GO:0006260). During heat stress conditions, nucleolin normally found
in the nucleolus is translocated to nucleoplasm and interacts with recombinant
replication protein, a key DNA replication factor. It is the contribution of these regulatory
process that helps in DNA replication during heat stress (Wang et al., 2001)
Interestingly, four significant GO terms are related to mastitis resistance, namely
positive regulation of lymphocyte proliferation (GO:0050671), phagocytosis

71
(GO:0006909), regulation of inflammatory response (GO:0050727) and defense
response to bacterium (GO:0046903). The relationship between lymphocyte
proliferation and mastitis resistance is well documented. Lymphocytes are able to
secrete certain cytokines which based on the kind of cytokines facilitate either a cell-
mediated or humoral response and produce antibodies against invading pathogens
thereby developing resistance against mastitis (Sordillo and Streicher, 2002). In
addition, heat stress is known to induce death signals that lead to apoptosis of
mammary cells. During inflammatory reactions, phagocytosis of apoptotic cells is to be
recognized and engulfed by macrophages to prevent inflammation of mammary tissues
(Kitamura et al., 2005). It should be note that the cellular stress response, including heat
shock response and DNA damage response, and their cross talk with inflammatory
pathways regulate host defense against bacterial infections in the udder (Muralidharan
and Mandrekar, 2013).
Intercalated disc (GO:0014704) is implicated in placental function and
development. Several biological processes are related with this term including diffusion
of molecules such as cAMP, cGMP, inositol triphosphate (IP3), Ca++ between mother
and fetus thereby allowing growth and development of fetus (Malassine and Cronier,
2005).
Cellular response to tumor necrosis factor (GO:0071356) showed significant
overrepresentation of genes associated with successful pregnancy. TNF-α is also
essential for early event in pregnancy such as implantation. Besides, TNF-α is also
involved in secreting placental hormone such as chorionic gonadotrophin which is
crucial for gestation and successful pregnancy outcome (Haider and Knoefler, 2009).

72
Summary
The results of this study suggest that there is a negative genetic correlation for
conception between thermoneutral and heat stress conditions. Therefore, the continued
selection for greater fertility ignoring heat stress conditions will result in increasing even
more the susceptibility to heat stress. On the other hand, we identified a positive genetic
correlation for SCS between neutral and heat stress, and hence continued selection for
lower SCS will also yield a favorable selection response for SCS under heat stress.
Whole-genome scan identified significant regions on BTA10, BTA11 and BTA21
associated with CI under heat stress. Some of the putative genes are RGS6, PRKCE,
KLHL25. Moreover, gene set analyses revealed several functional significant terms
such as protein refolding, inter-strand cross link repair, regulation of intra-cellular protein
transport, and intercalated disc. For SCS, genomic regions on BTA2, BTA3, BTA19 and
BTA23 were associated with thermotolerance for SCS. These genomic regions harbor
genes such as DLX1, STAT5B, NR2F6 CNPY3 and LCP1 which are potential candidate
genes for response of SCS to heat stress. Additionally, gene set analysis revealed
several functional significant terms such as chaperone mediated protein folding, DNA
replication, positive regulation of lymphocyte proliferation, phagocytosis, regulation of
inflammatory response and defense response to bacterium are associated with
thermotolerance. Overall, this study contributes to better understanding of the genetics
underlying heat stress and points out novel opportunities for improving thermotolerance
in dairy cattle.

73
Table 3-1. General (σ2a) and heat tolerance (100σ2
v) additive variances, genetic correlations and estimates of heritability h2
f(10) at THI of 78 (10 units above THI threshold) for CI
σ2a = General additive genetic variances, 100 σ2
v = heat tolerance additive variances at 78 (10 units above a THI threshold of 68); 10σa.v = additive genetic covariance between general and heat tolerance effect; rG(a, v) = genetic correlation between general and heat tolerance effect; Cor-ht = heat tolerance additive genetic correlations; Cor-gen = regular additive genetic correlations.
Parameters Parity 1 Parity 2 Parity 3
σ2a 0.002 0.004 0.008
100 σ2v 0.005 0.009 0.015
10σa.v -0.001 -0.003 -0.008
σ2e 0.20 0.19 0.18
h2f(10) 0.024 0.032 0.024
rG(a, v) -0.35 -0.58 -0.82
Cor-ht (par1, parj) 0.17 0.49
Cor-ht (par2, parj) 0.32
Cor- gen(par1, parj) 0.58 0.83
Cor- gen(par2, parj) 0.88

74
Table 3-2. General (σ2a) and heat tolerance (100 σ2
v) additive variances, genetic correlations and estimates of heritability h2
f(10) at THI of 78 (10 units above THI threshold) for SCS
σ2a = General additive genetic variances, 100 σ2
v = heat tolerance additive variances at 78 (10 units above a THI threshold of 68); 10σa.v = additive genetic covariance between general and heat tolerance effect; rG(a, v) = genetic correlation between general and heat tolerance effect; Cor-ht = heat tolerance additive genetic correlations; Cor-gen = regular additive genetic correlations
Parameters Parity 1 Parity 2 Parity 3
σ2a 0.27 0.28 0.38
100 σ2v 0.042 0.049 0.091
10σa.v 0.026 0.011 0.081
h2f(10) 0.11 0.10 0.15
σ2e 1.34 1.35 1.55
rG(a, v) 0.24 0.10 0.43
Cor- ht (par1, parj) 0.18 0.68
Cor- ht (par2, parj) 0.33
Cor- gen(par1, parj) 0.77 0.79
Cor- gen(par2, parj) 0.90

75
Table 3-3. GO terms significantly enriched with genes associated with CI
GO ID Term No. genes
No. sig. genes
P – value
0042026 Protein folding 5 2 0.002
0036297 Inter-strand cross link repair 7 2 0.005
0003157 Regulation of intra cellular protein
transport
66 5 0.010
0043401 Steroid hormone mediated signaling
pathway
34 3 0.030
0030258 Lipid modification 48 4 0.015
0007281 Germ cell development 40 3 0.045
0009791 Post embryonic development 14 2 0.031
Table 3-4. MeSH terms significantly enriched with genes associated with CI
MeSH ID Term No. genes No. sig. genes P - value
D050884 HSP72 Heat-Shock Proteins 2 1 0.032
D018840 HSP70 Heat-Shock Proteins 26 3 0.008
D050883 HSC70 Heat-Shock Proteins 10 2 0.011
D005982 Glutathione Transferase 50 3 0.048
D017871 Calcium-Calmodulin-Dependent
Protein Kinases
16 2 0.028
D052250 Nitric Oxide Synthase Type III 73 4 0.043
D005306 Fertilization 32 3 0.029

76
Table 3-5. GO terms significantly enriched with genes associated with SCS
GO ID Term No. genes
No. sig. genes
P -value
0061077 Chaperone mediated protein folding 26 3 0.016
0006260 DNA replication 57 4 0.031
0050671 Positive regulation of lymphocyte proliferation 27 4 0.002
0006909 Phagocytosis 31 4 0.003
0050727 Regulation of inflammatory response 56 4 0.029
0042742 Defense response to bacterium 41 4 0.007
Table 3-6. MeSH terms significantly enriched with genes associated with SCS
MeSH ID Term No. genes No. sig. genes
P- value
D0061077 Major Histocompatibility complex 14 2 0.036
D0006260 Immunity Innate 35 4 0.004
D0050671 Interleukin-1 29 3 0.012
D0015850 Interleukin-6 32 3 0.016
D0054732 Calcium-Calmodulin-Dependent
Protein Kinase Type 2
13 2 0.019
D0053496 Inositol 1,4,5-Triphosphate
Receptors
17 2 0.032

77
Figure 3-1. Manhattan plot showing the results of genome-wide association mapping for CI: the left plots show the result of general merit and the right plots show the results of thermotolerance across lactations numbered vertically as lac1, lac2 and lac3.

78
Figure 3-2. Manhattan plot showing the results of genome-wide association mapping for SCS: the left plots show the result of general merit and the right plots show the results of thermotolerance across lactations numbered vertically as lac1, lac2 and lac3

79
CHAPTER 4 CONCLUSIONS
The results from this study indicate that test-day records combined with
meteorological data from public weather stations can be used for genetic-genomic
evaluations of heat tolerances. These studies also indicate that heat tolerances additive
genetic variances for production, reproduction and health traits increased with parity
suggesting that cows become more sensitive to heat stress in later lactations. The
genetic correlations between regular and heat stress effects for production and
reproduction traits were negative and ranged between -0.30 and -0.55 for MY traits and
between -0.35 and -0.82 for reproduction trait. Continued selection for production and
reproduction trait without heat stress can result in animals with lower heat stress
tolerance and will also deteriorate genetic merit for these traits under heat stress
condition. However, the study estimated positive genetic correlations between regular
and heat stress effects for SCS ranging between +0.10 to +0.43. Continued selection
for lower somatic cell score will also yield positive selection response for heat
tolerances in cows.
These studies also have identified genomic regions, and a list of candidate genes
and pathways that show significant association with responses of production,
reproduction and health traits to heat stress. Our findings reveal that thermotolerance is
a quantitative trait influenced by many regions of the genome. Several potential
candidate genes and pathways with known roles in thermal regulation, immune
response, autophagy, anti-apoptosis among others were identified in these genomic
regions. These findings contribute to better understanding of the genetics underlying
heat stress and in addition these results can be useful for future fine mapping studies,

80
identification of causative mutations, and functional studies. Finally, the information
provided in this study provides novel opportunities for improving thermotolerance in
dairy cattle by means of selective breeding.
Heat stress in our study explained only a small portion of genotype by
environment interaction. Moreover, the model in our study identifies only acute
response to heat stress and do not consider chronic response to heat stress. It also fails
to consider the differences between test days. Therefore, the model could be improved
by modeling test days as random effects and by modeling the genotype and
environment interactions, but this will increase the computational cost and may raise the
issues of convergence of parameters.

81
LIST OF REFERENCES
Abdalla, E. A., F. Penagaricano, T. M. Byrem, K. A. Weigel, and G. J. M. Rosa. 2016. Genome-wide association mapping and pathway analysis of leukosis incidence in a US Holstein cattle population. Animal Genetics 47(4):395-407.
Abeni, F., L. Calamari, and L. Stefanini. 2007. Metabolic conditions of lactating Friesian cows during the hot season in the Po valley. 1. Blood indicators of heat stress. International Journal of Biometeorology 52(2):87-96.
Aguilar, I., I. Misztal, D. L. Johnson, A. Legarra, S. Tsuruta, and T. J. Lawlor. 2010a. Hot topic: A unified approach to utilize phenotypic, full pedigree, and genomic information for genetic evaluation of Holstein final score. J. Dairy Sci.93(2):743-752.
Aguilar, I., I. Misztal, and S. Tsuruta. 2009. Genetic components of heat stress for dairy cattle with multiple lactations. J. Dairy Sci.92(11):5702-5711.
Aguilar, I., I. Misztal, and S. Tsuruta. 2010b. Short communication: Genetic trends of milk yield under heat stress for US Holsteins. J. Dairy Sci.93(4):1754-1758.
Albright, J. L. and C. W. Alliston. 1971. Effects of varying the environment upon the performance of dairy cattle. J. Anim. Sci.32(3):66-77.
Arcuri, F., S. Papa, A. Meini, A. Carducci, R. Romagnoli, L. Bianchi, M. G. Riparbelli, J. C. Sanchez, M. Palmi, P. Tosi, and M. Cintorino. 2005. The translationally controlled tumor protein is a novel calcium binding protein of the human placenta and regulates calcium handling in trophoblast cells. Biology of Reproduction 73(4):745-751.
Ashburner, M., C. A. Ball, J. A. Blake, D. Botstein, H. Butler, J. M. Cherry, A. P. Davis, K. Dolinski, S. S. Dwight, J. T. Eppig, M. A. Harris, D. P. Hill, L. Issel-Tarver, A. Kasarskis, S. Lewis, J. C. Matese, J. E. Richardson, M. Ringwald, G. M. Rubin, G. Sherlock, and C. Gene Ontology. 2000. Gene Ontology: tool for the unification of biology. Nature Genetics 25(1):25-29.
Avendano-Reyes, L., F. D. Alvarez-Valenzuela, A. Correa-Calderon, J. S. Saucedo-Quintero, P. H. Robinson, and J. G. Fadel. 2006. Effect of cooling Holstein cows during the dry period on postpartum performance under heat stress conditions. Livestock Science 105(1-3):198-206.
Averill, T. A., R. Rekaya, and K. Weigel. 2004. Genetic analysis of male and female fertility using longitudinal binary data. J. Dairy Sci.87(11):3947-3952.
Badinga, L., W. W. Thatcher, T. Diaz, M. Drost, and D. Wolfenson. 1993. Effect of environmental heat-stress on follicular development and steroidogenesis in lactating holstein cows. Theriogenology 39(4):797-810.

82
Baumgard, L. H. and R. P. Rhoads, Jr. 2013. Effects of heat stress on postabsorptive metabolism and energetics. Annual Review of Animal Biosciences, 1(1):311-337.
Beede, D. K. and R. J. Collier. 1986. Potential nutritional strategies for intensively managed cattle during thermal-stress. J. Anim. Sci.62(2):543-554.
Berman, A., Y. Folman, M. Kaim, M. Mamen, Z. Herz, D. Wolfenson, A. Arieli, and Y. Graber. 1985. Upper critical-temperatures and forced ventilation effects for high-yielding dairy-cows in a sub-tropical climate. J. Dairy Sci.68(6):1488-1495.
Berman, A. and A. Meltzer. 1973. Critical temperatures in lactating dairy cattle a new approach to an old problem. International Journal of Biometeorology 17(2):167-176.
Bernabucci, U., S. Biffani, L. Buggiotti, A. Vitali, N. Lacetera, and A. Nardone. 2014. The effects of heat stress in Italian Holstein dairy cattle. J. Dairy Sci.97(1):471-486.
Bianca, W. 1965. Reviews of progress of dairy science .a. physiology cattle in a hot environment. Journal of Dairy Research 32(3):291
Biffani, S., U. Bernabucci, A. Vitali, N. Lacetera, and A. Nardone. 2016. Short communication: Effect of heat stress on nonreturn rate of Italian Holstein cows. J. Dairy Sci.99(7):5837-5843.
Blott, S., J. J. Kim, S. Moisio, A. Schmidt-Kuntzel, A. Cornet, P. Berzi, N. Cambisano, C. Ford, B. Grisart, D. Johnson, L. Karim, P. Simon, R. Snell, R. Spelman, J. Wong, J. Vilkki, M. Georges, F. Farnir, and W. Coppieters. 2003. Molecular dissection of a quantitative trait locus: A phenylalanine-to-tyrosine substitution in the transmembrane domain of the bovine growth hormone receptor is associated with a major effect on milk yield and composition. Genetics 163(1):253-266.
Bohmanova, J., I. Misztal, and J. B. Cole. 2007. Temperature-humidity indices as indicators of milk production losses due to heat stress. J. Dairy Sci.90(4):1947-1956.
Bouraoui, R., M. Lahmar, A. Majdoub, M. Djemali, and R. Belyea. 2002. The relationship of temperature-humidity index with milk production of dairy cows in a Mediterranean climate. Animal Research 51(6):479-491.
Burrow, H. M. 2001. Variances and covariances between productive and adaptive traits and temperament in a composite breed of tropical beef cattle. Livestock Production Science 70(3):213-233.
Calderwood, S. K. 2018. Heat shock proteins and cancer: intracellular chaperones or extracellular signalling ligands? Philosophical transactions of the Royal Society of London. Series B, Biological sciences 373(1738).

83
Calderwood, S. K., Y. Xie, X. Wang, M. A. Khaleque, S. D. Chou, A. Murshid, T. Prince, and Y. Zhang. 2010. Signal transduction pathways leading to heat shock transcription. Signal transduction insights 2:13-24.
Carstens, G. E. 1994. Cold thermoregulation in the newborn calf. Veterinary Clinics of North America-Food Animal Practice 10(1):69-106.
Cases, S., S. J. Smith, Y. W. Zheng, H. M. Myers, S. R. Lear, E. Sande, S. Novak, C. Collins, C. B. Welch, A. J. Lusis, S. K. Erickson, and R. V. Farese. 1998. Identification of a gene encoding an acyl CoA : diacylglycerol acyltransferase, a key enzyme in triacylglycerol synthesis. Proceedings of the National Academy of Sciences of the United States of America 95(22):13018-13023.
Charkoudian, N. 2010. Mechanisms and modifiers of reflex induced cutaneous vasodilation and vasoconstriction in humans. Journal of Applied Physiology 109(4):1221-1228.
Chatterjee, T. K. and R. A. Fisher. 2003. Mild heat and proteotoxic stress promote unique subcellular trafficking and nucleolar accumulation of RGS6 and other RGS proteins - Role of the RGS domain in stress-induced trafficking of RGS proteins. Journal of Biological Chemistry 278(32):30272-30282.
Cochran, S. D., J. B. Cole, D. J. Null, and P. J. Hansen. 2013. Discovery of single nucleotide polymorphisms in candidate genes associated with fertility and production traits in Holstein cattle. Bmc Genetics 14:49.
Collier, R. J. and K. G. Gebremedhin. 2015. Thermal Biology of Domestic Animals. Annual Review of Animal Biosciences, 3 (3):513-532.
Coppock, C. E., P. A. Grant, S. J. Portzer, D. A. Charles, and A. Escobosa. 1982. Lactating dairy-cow responses to dietary-sodium, chloride, and bicarbonate during hot weather. J. Dairy Sci.65(4):566-576.
Cui, Y., Y. Hao, J. Li, W. Bao, G. Li, Y. Gao, and X. Gu. 2016. Chronic heat stress induces immune response, oxidative stress response, and apoptosis of finishing pig liver: a proteomic approach. International journal of molecular sciences 17(5).
Curtis, S. E. 1983. Environmental management in animal agriculture. Iowa State University Press, Ames, Iowa.
de Vries, A. 2004. Economics of delayed replacement when cow performance is seasonal. J. Dairy Sci.87(9):2947-2958.
de Vries, A. and C. A. Risco. 2005. Trends and seasonality of reproductive performance in Florida and Georgia dairy herds from 1976 to 2002. J. Dairy Sci.88(9):3155-3165.

84
Dikmen, S., J. B. Cole, D. J. Null, and P. J. Hansen. 2012. Heritability of rectal temperature and genetic correlations with production and reproduction traits in dairy cattle. J. Dairy Sci.95(6):3401-3405.
Dikmen, S., J. B. Cole, D. J. Null, and P. J. Hansen. 2013. Genome-wide association mapping for identification of quantitative trait loci for rectal temperature during heat stress in holstein cattle. Plos One 8(7).
Dulken, B. W., D. S. Leeman, S. C. Boutet, K. Hebestreit, and A. Brunet. 2017. Single-cell transcriptomic analysis defines heterogeneity and transcriptional dynamics in the adult neural stem cell lineage. Cell Reports 18(3):777-790.
Farooq, U., H. A. Samad, F. Shehzad, and A. Qayyum. 2010. Physiological responses of cattle to heat stress. World Applied Sciences Journal 8(Special Issue):38-43.
Farr, R. L., C. Lismont, S. R. Terlecky, and M. Fransen. 2016. Peroxisome biogenesis in mammalian cells: The impact of genes and environment. Biochimica Et Biophysica Acta-Molecular Cell Research 1863(5):1049-1060.
Fernandes, R., C. Tsuda, A. L. Perumalsamy, T. Naranian, J. Chong, B. M. Acton, Z.-B. Tong, L. M. Nelson, and A. Jurisicova. 2012. NLRP5 mediates mitochondrial function in mouse oocytes and embryos. Biology of Reproduction 86(5).
Ferreira, F. C., R. S. Gennari, G. E. Dahl, and A. De Vries. 2016. Economic feasibility of cooling dry cows across the United States. J. Dairy Sci.99(12):9931-9941.
Fisher, A. D., N. Roberts, S. J. Bluett, G. A. Verkerk, and L. R. Matthews. 2008. Effects of shade provision on the behaviour, body temperature and milk production of grazing dairy cows during a New Zealand summer. New Zealand Journal of Agricultural Research 51(2):99-105.
Fordham, D. P., T. T. McCarthy, and P. Rowlinson. 1984. Oestrus detection in dairy cows by milk temperature measurement. British Society of Animal Production. Winter Meeting. 26-28 March 1984, Grand Hotel, Scarborough.135 (2) .
Fournel, S., V. Ouellet, and E. Charbonneau. 2017. Practices for alleviating heat stress of dairy cows in humid continental climates: A Literature Review. Animals : an open access journal from MDPI 7(5).
Freitas, M. S., I. Misztal, J. Bohmanova, and J. West. 2006. Utility of on- and off-farm weather records for studies in genetics of heat tolerance. Livestock Science 105(1-3):223-228.
Fuquay, J. W. 1981. Heat-stress as it affects animal production. J. Anim. Sci.52(1):164-174.

85
Garcia-Ispierto, I., F. Lopez-Gatius, P. Santolaria, J. L. Yaniz, C. Nogareda, M. Lopez-Bejar, and F. De Rensis. 2006. Relationship between heat stress during the peri-implantation period and early fetal loss in dairy cattle. Theriogenology 65(4):799-807.
Garner, J. B., M. L. Douglas, S. R. O. Williams, W. J. Wales, L. C. Marett, T. T. T. Nguyen, C. M. Reich, and B. J. Hayes. 2017. Genomic selection improves heat tolerance in dairy cattle. Scientific Reports 7.
Gaughan, J. B., S. M. Holt, G. L. Hahn, T. L. Mader, and R. Eigenberg. 2000. Respiration rate - Is it a good measure of heat stress in cattle? Asian-Australasian Journal of Animal Sciences 13:329-332.
Giesecke, W. H. 1985. The effect of stress on udder health of dairy-cows. Onderstepoort Journal of Veterinary Research 52(3):175-193.
Greenwood, M., M. P. Greenwood, J. F. R. Paton, and D. Murphy. 2015. Transcription factor CREB3L1 regulates endoplasmic reticulum stress response genes in the osmotically challenged rat hypothalamus. Plos One 10(4).
Hagiya, K., K. Hayasaka, T. Yamazaki, T. Shirai, T. Osawa, Y. Terawaki, Y. Nagamine, Y. Masuda, and M. Suzuki. 2017. Effects of heat stress on production, somatic cell score and conception rate in Holsteins. Animal Science Journal 88(1):3-10.
Hahn, G. L., R. A. Eigenberg, J. A. Nienaber, E. T. Littledike, and R. L. Hruska. 1990. Measuring physiological-responses of animals to environmental stressors using a microcomputer-based portable datalogger. J. Anim. Sci.68(9):2658-2665.
Haider, S. and M. Knoefler. 2009. Human Tumour Necrosis Factor: Physiological and pathological roles in placenta and endometrium. Placenta 30(2):111-123.
Hamiel, C. R., S. Pinto, A. Hau, and P. E. Wischmeyer. 2009. Glutamine enhances heat shock protein 70 expression via increased hexosamine biosynthetic pathway activity. American Journal of Physiology-Cell Physiology 297(6):C1509-C1519.
Hammami, H., J. Vandenplas, M. L. Vanrobays, B. Rekik, C. Bastin, and N. Gengler. 2015. Genetic analysis of heat stress effects on yield traits, udder health, and fatty acids of Walloon Holstein cows. J. Dairy Sci.98(7):4956-4968.
Hansen, P. J. 2009. Effects of heat stress on mammalian reproduction. Philosophical Transactions of the Royal Society B-Biological Sciences 364(1534):3341-3350.
Hansen, P. J. 2013. Cellular and molecular basis of therapies to ameliorate effects of heat stress on embryonic development in cattle. Animal Reproduction 10(3):322-333.
Hansen, P. J. and C. F. Arechiga. 1999. Strategies for managing reproduction in the heat-stressed dairy cow. J. Anim. Sci.77:36-50.

86
Herrera, E. and H. Ortega-Senovilla. 2010. Maternal lipid metabolism during normal pregnancy and its implications to fetal development. Clinical Lipidology 5(6):899-911.
Hewison, M. 2011. Antibacterial effects of vitamin D. Nature Reviews Endocrinology 7(6):336-344.
Home, P., S. Ray, D. Dutta, I. Bronshteyn, M. Larson, and S. Paul. 2009. GATA3 is selectively expressed in the trophectoderm of peri-implantation embryo and directly regulates Cdx2 gene expression. Journal of Biological Chemistry 284(42):28729-28737.
Howard, J. T., S. D. Kachman, W. M. Snelling, E. J. Pollak, D. C. Ciobanu, L. A. Kuehn, and M. L. Spangler. 2014. Beef cattle body temperature during climatic stress: a genome-wide association study. International Journal of Biometeorology 58(7):1665-1672.
Huang, C., S. Tsuruta, J. K. Bertrand, I. Misztal, T. J. Lawlor, and J. S. Clay. 2009. Trends for conception rate of Holsteins over time in the southeastern United States. J. Dairy Sci.92(9):4641-4647.
Igono, M. O. and H. D. Johnson. 1990. Physiological stress index of lactating dairy-cows based on diurnal pattern of rectal temperature. Journal of Interdisciplinary Cycle Research 21(4):303-320.
Igono, M. O., H. D. Johnson, B. J. Steevens, G. F. Krause, and M. D. Shanklin. 1987. Physiological, productive, and economic-benefits of shade, spray, and fan system versus shade for holstein cows during summer heat. J. Dairy Sci.70(5):1069-1079.
Jarolim, S., A. Ayer, B. Pillay, A. C. Gee, A. Phrakaysone, G. G. Perrone, M. Breitenbach, and I. W. Dawes. 2013. Saccharomyces cerevisiae genes involved in survival of heat shock. G3-Genes Genomes Genetics 3(12):2321-2333.
Jeffries, K. M., S. G. Hinch, T. Sierocinski, P. Pavlidis, and K. M. Miller. 2014. Transcriptomic responses to high water temperature in two species of Pacific salmon. Evolutionary Applications 7(2):286-300.
Kadzere, C. T., M. R. Murphy, N. Silanikove, and E. Maltz. 2002. Heat stress in lactating dairy cows: a review. Livestock Production Science 77(1):59-91.
Kampinga, H. H. 2006. Chaperones in preventing protein denaturation in living cells and protecting against cellular stress. Handbook of experimental pharmacology (172):1-42.
Key, N. and S. Sneeringer. 2014. Potential effects of climate change on the productivity of U.S. dairies. American Journal of Agricultural Economics 96(4):1136-1156.

87
Kitamura, C., T. Nishihara, Y. Ueno, M. Nagayoshi, S. Kasugai, and M. Terashita. 2005. Thermotolerance of pulp cells and phagocytosis of apoptotic pulp cells by surviving pulp cells following heat stress. Journal of Cellular Biochemistry 94(4):826-834.
Lee, W. C., H. C. Wen, C. P. Chang, M. Y. Chen, and M. T. Lin. 2006. Heat shock protein 72 overexpression protects against hyperthermia, circulatory shock, and cerebral ischemia during heatstroke. Journal of Applied Physiology 100(6):2073-2082.
Lin, J.-X., P. Li, D. Liu, H. T. Jin, J. He, M. A. U. Rasheed, Y. Rochman, L. Wang, K. Cui, C. Liu, B. L. Kelsall, R. Ahmed, and W. J. Leonard. 2012. Critical role of STAT5 transcription factor tetramerization for cytokine responses and normal immune function. Immunity 36(4):586-599.
Liu, Y. and A. Chang. 2008. Heat shock response relieves ER stress. Embo Journal 27(7):1049-1059.
Liu, Z. B., J. M. Wang, F. X. Yang, L. Yang, Y. F. Yue, J. B. Xiang, M. Gao, F. J. Xiong, D. Lv, X. J. Wu, N. Liu, X. Zhang, X. F. Li, and Y. Yang. 2014. A novel membrane-bound E3 ubiquitin ligase enhances the thermal resistance in plants. Plant Biotechnology Journal 12(1):93-104.
Long, E., A. V. Capuco, D. L. Wood, T. Sonstegard, G. Tomita, M. J. Paape, and X. Zhao. 2001. Escherichia coli induces apoptosis and proliferation of mammary cells. Cell Death and Differentiation 8(8):808-816.
Luft, J. C., I. J. Benjamin, R. Mestril, and D. J. Dix. 2001. Heat shock factor 1-mediated thermotolerance prevents cell death and results in G(2)/M cell cycle arrest. Cell Stress & Chaperones 6(4):326-336.
Macciotta, N. P. P., S. Biffani, U. Bernabucci, N. Lacetera, A. Vitali, P. Ajmone-Marsan, and A. Nardone. 2017. Derivation and genome-wide association study of a principal component-based measure of heat tolerance in dairy cattle. J. Dairy Sci.100(6):4683-4697.
Mackinnon, M. J., K. Meyer, and D. J. S. Hetzel. 1991. Genetic-variation and covariation for growth, parasite resistance and heat tolerance in tropical cattle. Livestock Production Science 27(2-3):105-122.
Maia, A. S. C., R. G. daSilva, and C. M. B. Loureiro. 2005. Sensible and latent heat loss from the body surface of Holstein cows in a tropical environment. International Journal of Biometeorology 50(1):17-22.
Malassine, A. and L. Cronier. 2005. Involvement of gap junctions in placental functions and development. Biochimica Et Biophysica Acta-Biomembranes 1719(1-2):117-124.

88
Maust, L. E., R. E. McDowell, and N. W. Hooven. 1972. Effect of summer weather on performance of holstein cows in 3 stages of lactation. J. Dairy Sci.55(8):1133-&.
Mayer, M. P. and B. Bukau. 2005. Hsp70 chaperones: Cellular functions and molecular mechanism. Cellular and Molecular Life Sciences 62(6):670-684.
McArthur, A. J. and J. A. Clark. 1988. Body-temperature of homeotherms and the conservation of energy and water. Journal of Thermal Biology 13(1):9-13.
McMahon, S. J., J. Schuemann, H. Paganetti, and K. M. Prise. 2016. Mechanistic modelling of DNA repair and cellular survival following radiation-induced DNA damage. Scientific Reports 6.
Mehrzad, J., H. Dosogne, B. De Spiegeleer, L. Duchateau, and C. Burvenich. 2007. Bovine blood neutrophil Acyloxyacyl Hydrolase (AOAH) activity during endotoxin and coliform mastitis. Veterinary Research 38(5):655-668.
Minault, M., J. C. Lecron, S. Labrouche, G. Simonnet, and J. Gombert. 1995. Characterization of binding-sites for neuropeptide FF on T-lymphocytes of the jurkat cell-line. Peptides 16(1):105-111.
Misztal, I., S. Tsuruta, T. Strabel, B. Auvray, T. Druet, and D. H. Lee. 2002. BLUPF90 and related programs (BGF90). Proceedings of the 7th World Congress on Genetics Applied to Livestock Production, Montpellier, France, August, 2002. Session 28:1-2.
Morimoto, R. I., M. P. Kline, D. N. Bimston, and J. J. Cotto. 1997. The heat-shock response: regulation and function of heat-shock proteins and molecular chaperones. Essays in Biochemistry,32(32):17-29.
Mulligan, F., L. O'Grady, D. Rice, and M. Doherty. 2006. Production diseases of the transition cow: Milk fever and subclinical hypocalcaemia. Irish Veterinary Journal 59(12):697-702.
Muralidharan, S. and P. Mandrekar. 2013. Cellular stress response and innate immune signaling: integrating pathways in host defense and inflammation. Journal of Leukocyte Biology 94(6):1167-1184.
Murphy, M. R., C. L. Davis, and G. C. McCoy. 1983. Factors affecting water-consumption by Holstein cows in early lactation. J. Dairy Sci.66(1):35-38.
Nalabothula, N., F. E. Indig, and F. Carrier. 2010. The nucleolus takes control of protein trafficking under cellular stress. Molecular and cellular pharmacology 2(5):203-212.
Nardone, A., B. Ronchi, N. Lacetera, M. S. Ranieri, and U. Bernabucci. 2010. Effects of climate changes on animal production and sustainability of livestock systems. Livestock Science 130(1-3):57-69.

89
Nelson, S. J., M. Schopen, A. G. Savage, J.-L. Schulman, and N. Arluk. 2004. The MeSH translation maintenance system: structure, interface design, and implementation. Studies in health technology and informatics 107(Pt 1):67-69.
Nguyen, T. T. T., P. J. Bowman, M. Haile-Mariam, G. J. Nieuwhof, B. J. Hayes, and J. E. Pryce. 2017. Short communication: Implementation of a breeding value for heat tolerance in Australian dairy cattle. J. Dairy Sci.100(9):7362-7367.
Nguyen, T. T. T., P. J. Bowman, M. Haile-Mariam, J. E. Pryce, and B. J. Hayes. 2016. Genomic selection for tolerance to heat stress in Australian dairy cattle. J. Dairy Sci.99(4):2849-2862.
Oda, T., T. Hayano, H. Miyaso, N. Takahashi, and T. Yamashita. 2007. Hsp90 regulates the Fanconi anemia DNA damage response pathway. Blood 109(11):5016-5026.
Olson, T. A., C. Lucena, C. C. Chase, and A. C. Hammond. 2003. Evidence of a major gene influencing hair length and heat tolerance in Bos taurus cattle. J. Anim. Sci.81(1):80-90.
Page, T. J., D. Sikder, L. Yang, L. Pluta, R. D. Wolfinger, T. Kodadek, and R. S. Thomas. 2006. Genome-wide analysis of human HSF1 signaling reveals a transcriptional program linked to cellular adaptation and survival. Molecular Biosystems 2(12):627-639.
Parankusam, S., S. S. Adimulam, P. Bhatnagar-Mathur, and K. K. Sharma. 2017. Nitric oxide (NO) in plant heat stress tolerance: current knowledge and perspectives. Frontiers in Plant Science 8.
Parker, A. L., M. Kavallaris, and J. A. McCarroll. 2014. Microtubules and their role in cellular stress in cancer. Frontiers in oncology 4:153-153.
Parmentier, I., D. Portetelle, N. Gengler, A. Prandi, C. Bertozzi, L. Vleurick, R. Gilson, and R. Renaville. 1999. Candidate gene markers associated with somatotropic axis and milk selection. Domestic Animal Endocrinology 17(2-3):139-148.
Prendiville, D. J., J. Lowe, B. Earley, C. Spahr, and P. Kettlewell. 2003. Radiotelemetry systems for measuring body temperature in cattle. Farm & Food 13(1):15-18.
Ptak, E. and L. R. Schaeffer. 1993. Use of test day yields for genetic evaluation of dairy sires and cows. Livestock Production Science 34(1-2):23-34.
Purwanto, B. P., M. Fujita, M. Nishibori, and S. Yamamoto. 1991. Effect of environmental temperature and feed intake on plasma concentration of thyroid hormones in dairy heifers. Asian-Australasian Journal of Animal Sciences 4(3):293-298.
Ravagnolo, O. and I. Misztal. 2000. Genetic component of heat stress in dairy cattle, parameter estimation. J. Dairy Sci.83(9):2126-2130.

90
Ravagnolo, O. and I. Misztal. 2002. Effect of heat stress on nonreturn rate in Holstein cows: Genetic analyses. J. Dairy Sci.85(11):3092-3100.
Ravagnolo, O., I. Misztal, and G. Hoogenboom. 2000. Genetic component of heat stress in dairy cattle, development of heat index function. J. Dairy Sci.83(9):2120-2125.
Romanponce, H., W. W. Thatcher, D. E. Buffington, C. J. Wilcox, and H. H. Vanhorn. 1977. Physiological and production responses of dairy-cattle to a shade structure in a subtropical environment. J. Dairy Sci.60(3):424-430.
Saber, A. P. R., M. T. Jalali, D. Mohjeri, A. A. Akhoole, H. Z. N. Teymourluei, M. Nouri, and S. Garachorlo. 2009. The effect of ambient temperature on thyroid hormones concentration and histopathological changes of thyroid gland in cattle in tabriz, iran. Asian Journal of Animal and Veterinary Advances 4(1):28-33.
Sahana, G., B. Guldbrandtsen, B. Thomsen, and M. S. Lund. 2013. Confirmation and fine-mapping of clinical mastitis and somatic cell score QTL in Nordic Holstein cattle. Animal Genetics 44(6):620-626.
Sanchez, J. P., I. Misztal, I. Aguilar, B. Zumbach, and R. Rekaya. 2009. Genetic determination of the onset of heat stress on daily milk production in the US Holstein cattle. J. Dairy Sci.92(8):4035-4045.
Sanchez-Bermejo, E., W. Zhu, C. Tasset, H. Eimer, S. Sureshkumar, R. Singh, V. Sundaramoorthi, L. Colling, and S. Balasubramanian. 2015. Genetic architecture of natural variation in thermal responses of arabidopsis. Plant Physiology 169(1):647-659.
Saredi, G., H. Huang, C. M. Hammond, C. Alabert, S. Bekker-Jensen, I. Forne, N. Reveron-Gomez, B. M. Foster, L. Mlejnkova, T. Bartke, P. Cejka, N. Mailand, A. Imhof, D. J. Patel, and A. Groth. 2016. H4K20me0 marks post-replicative chromatin and recruits the TONSL-MMS22L DNA repair complex. Nature 534(7609):714-718.
Schueller, L. K., O. Burfeind, and W. Heuwieser. 2014. Impact of heat stress on conception rate of dairy cows in the moderate climate considering different temperature-humidity index thresholds, periods relative to breeding, and heat load indices. Theriogenology 81(8):1050-1057.
Seath, D. M. and G. D. Miller. 1947. Repeatability of heat tolerance observations. J. Dairy Sci.30(12):957-962.
Shearer, J. K. and D. K. Beede. 1990. Heat-stress .1. thermoregulation and physiological-responses of dairy-cattle in hot weather. Agri-Practice 11(4):5
Silanikove, N. 1992. Effects of water scarcity and hot environment on appetite and digestion in ruminants - a review. Livestock Production Science 30(3):175-194.

91
Smith, S. J., S. Cases, D. R. Jensen, H. C. Chen, E. Sande, B. Tow, D. A. Sanan, J. Raber, R. H. Eckel, and R. V. Farese. 2000. Obesity resistance and multiple mechanisms of triglyceride synthesis in mice lacking Dgat. Nature Genetics 25(1):87-90.
Smolinska, N., M. Kiezun, K. Dobrzyn, K. Szeszko, A. Maleszka, and T. Kaminski. 2017. Adiponectin, orexin A and orexin B concentrations in the serum and uterine luminal fluid during early pregnancy of pigs. Animal reproduction science 178:1-8.
Somero, G. N. 2002. Thermal physiology and vertical zonation of intertidal animals: Optima, limits, and costs of living. Integrative and Comparative Biology 42(4):780-789.
Sordillo, L. M. and K. L. Streicher. 2002. Mammary gland immunity and mastitis susceptibility. Journal of Mammary Gland Biology and Neoplasia 7(2):135-146.
Sorensen, J. G., T. N. Kristensen, and V. Loeschcke. 2003. The evolutionary and ecological role of heat shock proteins. Ecology Letters 6(11):1025-1037.
St-Pierre, N. R., B. Cobanov, and G. Schnitkey. 2003. Economic Losses from Heat Stress by US Livestock Industries. J. Dairy Sci.86:E52-E77.
Strickland, J. T., R. A. Bucklin, R. A. Nordstedt, D. K. Beede, and D. R. Bray. 1989. Sprinkler and fan cooling system for dairy cows in hot, humid climates. Applied Engineering in Agriculture 5(2):231-236.
Sun, Y.-L., X.-Y. Zhang, T. Sun, N. He, J.-Y. Li, Y. Zhuang, Q. Zeng, J. Yu, Q. Fang, and R. Wang. 2013. The anti-inflammatory potential of neuropeptide FF in vitro and in vivo. Peptides 47:124-132.
Swalve, H. H. 2000. Theoretical basis and computational methods for different test-day genetic evaluation methods. J. Dairy Sci.83(5):1115-1124.
Tao, S., J. W. Bubolz, B. C. do Amaral, I. M. Thompson, M. J. Hayen, S. E. Johnson, and G. E. Dahl. 2011. Effect of heat stress during the dry period on mammary gland development. J. Dairy Sci.94(12):5976-5986.
Thatcher, W. W., A. Guzeloglu, R. Mattos, M. Binelli, T. R. Hansen, and J. K. Pru. 2001. Uterine-conceptus interactions and reproductive failure in cattle. Theriogenology 56(9):1435-1450.
Thompson, N. A., J. A. Haefliger, A. Senn, T. Tawadros, F. Magara, B. Ledermann, P. Nicod, and G. Waeber. 2001. Islet-brain1/JNK-interacting protein-1 is required for early embryogenesis in mice. Journal of Biological Chemistry 276(30):27745-27748.

92
Tsuyuzaki, K., G. Morota, M. Ishii, T. Nakazato, S. Miyazaki, and I. Nikaido. 2015. MeSH ORA framework: R/Bioconductor packages to support MeSH over-representation analysis. Bmc Bioinformatics 16.
VanRaden, P. M. 2004. Invited review: Selection on net merit to improve lifetime profit. J. Dairy Sci.87(10):3125-3131.
Viitala, S., J. Szyda, S. Blott, N. Schulman, M. Lidauer, A. Maki-Tanila, M. George, and J. Vilkki. 2006. The role of the bovine growth hormone receptor and prolactin receptor genes in milk, fat and protein production in Finnish Ayrshire dairy cattle. Genetics 173(4):2151-2164.
Walloe, L. 2016. Arterio-venous anastomoses in the human skin and their role in temperature control. Temperature (Austin, Tex.) 3(1):92-103.
Wang, H., I. Misztal, I. Aguilar, A. Legarra, and W. M. Muir. 2012. Genome-wide association mapping including phenotypes from relatives without genotypes. Genetics Research 94(2):73-83.
Wang, Y. Z., J. Guan, H. Y. Wang, Y. Wang, D. Leeper, and G. Iliakis. 2001. Regulation of DNA replication after heat shock by replication protein A-nucleolin interactions. Journal of Biological Chemistry 276(23):20579-20588.
West, J. W. 1999. Nutritional strategies for managing the heat-stressed dairy cow. J. Anim. Sci.77:21-35.
West, J. W. 2003. Effects of heat-stress on production in dairy cattle. J. Dairy Sci.86(6):2131-2144.
West, J. W., B. G. Mullinix, J. C. Johnson, K. A. Ash, and V. N. Taylor. 1990. Effects of bovine somatotropin on dry-matter intake, milk-yield, and body-temperature in holstein and jersey cows during heat-stress. J. Dairy Sci.73(10):2896-2906.
Wrenn, T. R., J. F. Sykes, and J. Bitman. 1961. Diurnal patterns of bovine body temperature. J. Dairy Sci.44:2077-2080.
Xiong, Q., J. Chai, H. Xiong, W. Li, T. Huang, Y. Liu, X. Suo, N. Zhang, X. Li, S. Jiang, and M. Chen. 2013. Association analysis of HSP70A1A haplotypes with heat tolerance in Chinese Holstein cattle. Cell Stress & Chaperones 18(6):711-718.
Yao, J., K. Liao, Q.-L. Li, C.-F. Wang, H.-M. Wang, S.-J. Lai, and J.-F. Zhong. 2011. mRNA expression of glutathione s-transferase pi (gstp1) under heat stress and association of genotypes with heat tolerance ability in holstein. Journal of Animal and Veterinary Advances 10(6):757-763.
Young, J. C., V. R. Agashe, K. Siegers, and F. U. Hartl. 2004. Pathways of chaperone-mediated protein folding in the cytosol. Nature Reviews Molecular Cell Biology 5(10):781-791.

93
Young, M. D., M. J. Wakefield, G. K. Smyth, and A. Oshlack. 2010. Gene ontology analysis for RNA-seq: accounting for selection bias. Genome Biology 11(2).
Yousef, M. K. 1985. Stress physiology in livestock. Volume I. Basic principles. Stress physiology in livestock. Volume I. Basic principles.217.
Zhang, C., J. Liu, G. Huang, Y. Zhao, X. Yue, H. Wu, J. Li, J. Zhu, Z. Shen, B. G. Haffty, W. Hu, and Z. Feng. 2016. Cullin3-klhl25 ubiquitin ligase targets acly for degradation to inhibit lipid synthesis and tumor progression. Genes & Development 30(17):1956-1970.
Zhang, H., Q. Lang, J. Li, Z. Zhong, F. Xie, G. Ye, B. Wan, and L. Yu. 2007. Molecular cloning and characterization of a novel human glycine-n-acyltransferase gene GLYATL1, which activates transcriptional activity of hse pathway. International Journal of Molecular Sciences 8(5):433-444.
Zhao, J., W. Wang, C. H. Ha, J. Y. Kim, C. Wong, E. M. Redmond, A. Hamik, M. K. Jain, G.-S. Feng, and Z. G. Jin. 2011. Endothelial GRB2-associated binder 1 is crucial for postnatal angiogenesis. Arteriosclerosis, Thrombosis and Vascular Biology 31(5):1016-1147.
Zheng, P., R. D. Schramm, and K. E. Latham. 2005. Developmental regulation and in vitro culture effects on expression of DNA repair and cell cycle checkpoint control genes in rhesus monkey oocytes and embryos. Biology of Reproduction 72(6):1359-1369.
Zhou, H., L. Cheng, W. Azimu, S. Hodge, G. R. Edwards, and J. G. H. Hickford. 2015. Variation in the bovine FABP4 gene affects milk yield and milk protein content in dairy cows. Scientific Reports 5.
Zimin, A. V., A. L. Delcher, L. Florea, D. R. Kelley, M. C. Schatz, D. Puiu, F. Hanrahan, G. Pertea, C. P. Van Tassell, T. S. Sonstegard, G. Marcais, M. Roberts, P. Subramanian, J. A. Yorke, and S. L. Salzberg. 2009. A whole-genome assembly of the domestic cow, Bos taurus. Genome Biology 10(4).

94
BIOGRAPHICAL SKETCH
Anil was born and raised in Kathmandu, Nepal. After completing his high school,
Anil joined Tribhuvan University where he completed his undergraduate degree of
veterinary science and animal husbandry (BVSc & AH). In spring of 2016, he came to
the United States to pursue his master’s degree in animal sciences and joined
University of Florida (UF). He conducted research under the supervision of Dr.
Francisco Peñagaricano in dairy cattle genetics and genomics. At UF, he has been
conducting his master research to understand genetic aspects of thermotolerance in
dairy cattle. His research contributes to a better understanding of the genes and
biological mechanisms underlying heat stress and points out novel genomic tools for
improving thermotolerance in dairy cattle. His research interests include development
and application of statistical and computational models for the analysis of phenotypic
and molecular data in dairy cattle.