1, aïcha daher2, kaitlin j. soye2,3, lisa b. frankel2,3...
TRANSCRIPT

siRNAs against TRBP inhibit HIV 1
siRNAs against the TAR RNA Binding Protein, TRBP, a Dicer cofactor,1
inhibit HIV-1 long terminal repeat expression and viral production.2
3
Helen S. Christensen1, Aïcha Daher2, Kait lin J. Soye2,3, Lisa B. Frankel2,3,‡, Marina R.4
Alexander1, Sébast ien Lainé2,3, Sylvie Bannwarth2,3,#, Chi L. Ong1, Sean W.L. Chung1,5
Shahan M. Campbell1, Damian F. J. Purcell1,*, Anne Gat ignol2,3,4,*6
1Department of Microbiology and Immunology, University of Melbourne, Parkville,7
Aust ralia; 2Virus-Cell Interact ions Laboratory, Lady Davis Inst itute for Medical8
Research; Department of 3Microbiology and Immunology, and 4Experimental Medicine,9
McGill University, Mont réal, Québec, Canada.10
11
*Equal work from AG and DP laboratories12
Corresponding authors:13
Dr. Anne Gat ignol14
Virus-Cell Interact ions Laboratory, Lady Davis Inst itute for Medical Research,15
3755 Côte Ste Catherine, Mont réal, QC, H3T 1E2, Canada.16
Tel: 1 514 340-8260 ext : 5284; Fax: 1 514 340 757617
E mail: anne.gat [email protected]
19
Dr. Damian F. J. Purcell20
Department of Microbiology and Immunology21
The University of Melbourne, Parkville 3010, Victoria, Aust ralia.22
Tel: 61 3 8344 6753; Fax: 61 3 9347 154023
E. mail: df j [email protected]
25
Present addresses: ‡ Biotech Research and Innovat ion Center, University of26
Copenhagen, Copenhagen, Denmark. # Laboratoire de Génét ique Moléculaire, Hôpital27
de l'Archet 2, Nice, France.28
Word count for the abst ract : 245; Word count for the text : 515729
ACCEPTED
Copyright © 2007, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.J. Virol. doi:10.1128/JVI.01511-06 JVI Accepts, published online ahead of print on 14 March 2007
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 2
ABSTRACT1
RNA interference (RNAi) is now widely used for gene silencing in mammalian2
cells. The mechanism uses the RNA-induced silencing complex, in which Dicer, Ago23
and the HIV-1 TAR RNA binding protein, TRBP are the main components. TRBP is a4
protein that increases HIV-1 expression and replicat ion by inhibit ion of the interferon-5
induced protein kinase PKR and by increasing t ranslat ion of viral mRNA. Af ter HIV6
infect ion, TRBP could rest rict t he viral RNA through it s act ivit y in RNAi or could7
cont ribut e more t o t he enhancement of viral repl icat ion. To det ermine which8
funct ion wil l be predominant in t he virological context , we analyzed whether t he9
inhibit ion of it s expression could enhance or decrease HIV replicat ion. We have10
generated small interfering (si)RNAs against TRBP and found that they decrease HIV-111
long terminal repeat (LTR) basal expression two fold, and LTR Tat t rans-act ivated12
level up to tenfold. In the context of HIV replicat ion, siRNAs against TRBP decrease13
the expression of viral genes and inhibit viral product ion up to f ivefold. The moderate14
increase of PKR expression and act ivat ion indicates that it cont ributes part ially to viral15
gene inhibit ion. The moderate decrease of miRNA biogenesis by TRBP siRNAs suggests16
t hat in t he cont ext of HIV repl icat ion, TRBP funct ions ot her t han RNAi are17
predominant . In addit ion, siRNAs against Dicer decrease viral product ion twofold and18
impedes miRNA biogenesis. These result s suggest t hat , in t he cont ext of HIV19
replicat ion, TRBP cont ributes mainly to the enhancement of virus product ion and that20
Dicer does not mediate HIV rest rict ion by RNAi.21
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 3
INTRODUCTION1
RNA interference (RNAi) is a natural mechanism used by eukaryotes for gene2
silencing (27, 38, 80). While invertebrates use 23bp double-st randed (ds) RNA, called3
smal l int erfering (si) RNAs, wit h perfect complement arit y t o degrade t arget ed4
messenger RNA (mRNA) in t he RNA-induced silencing complex (RISC), vertebrates5
predominant ly use imperfect ly paired 23bp micro (mi) RNAs t o cont rol mRNA6
t ranslat ion using RISC. In the RISC complex the Dicer protein has RNAse III domains7
and generates the siRNAs, whereas Argonaute, an enzyme with RNAse H-like domains8
mediates gene silencing. The abil it y of 21-23 nt siRNAs, short hairpin (sh) RNAs and9
miRNAs to inact ivate gene expression, while avoiding the dsRNA-act ivated protein10
kinase (PKR) act ivat ion (5, 25), has allowed the use of RNAi as a tool for specif ically11
decreasing t he expression of bot h cel lular (22, 51) and viral (17, 37) genes in12
mammalian cells.13
While inact ivat ing Human Immunodef iciency Virus (HIV) genes wit h siRNA,14
shRNA and miRNAs decreases viral repl icat ion (9, 11, 16, 41, 50, 73), t he high15
mutat ion rate in the HIV genome allows the virus to readily escape this sequence16
specif ic mechanism (8, 77), highlight ing the advantages of t arget ing cellular genes17
necessary for viral replicat ion. Cellular factors required for HIV replicat ion cycle that18
have been downregulated by RNAi include cell surface receptors CD4, CCR5, CXCR419
(55, 58, 65), expression factors NF B, Cyclin T1, CDK9, SPT5, PARP1 (15, 43, 62, 74)20
and proteins involved in int racellular t raff icking and viral packaging including Staufen,21
tRNA synthetase, Arp2/ 3, Cyclophilin A and Rab9 GTPase (13, 35, 45, 53, 57). In each22
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 4
case, HIV replicat ion was signif icant ly decreased with minimal cell death indicat ing1
that it is a very promising approach.2
Human t rans-act ivat ion response (TAR) RNA binding proteins (TRBP)1 and TRBP23
were init ially ident if ied as proteins that bind the HIV-1 TAR RNA and act ivate the long4
terminal repeat (LTR) expression in the absence and in the presence of the viral t rans-5
act ivator Tat (24, 29, 30). The two proteins dif fer by 21 addit ional amino acids in the6
N-terminal end of TRBP2 (4). TRBPs have two double-st randed RNA binding domains7
(dsRBD), the second one containing a KR-helix mot if that mediates dsRNA binding (21,8
26, 28, 44). A third basic domain in the C-terminal end of TRBP mediates protein-9
protein interact ions (36, 47). TRBPs have a physiological role in spermatogenesis and10
growth cont rol during development (49, 81). They also bind the interferon (IFN)-11
induced dsRNA-act ivat ed prot ein kinase PKR (19). TRBPs are oncogenic upon12
overexpression, l ikely due to their associat ion with PKR (7), with the PKR act ivator13
PACT [(60);G. Laraki, A. Daher and A. Gat ignol, unpublished data] and with the tumor14
suppressor Merlin (47, 48).15
In the context of HIV-1 replicat ion, TRBP1 and TRBP2 increase viral expression16
similarly by blocking the inhibit ory ef fect of PKR on viral t ranslat ion. TRBPs also17
restore the t ranslat ion of TAR-containing RNAs by a PKR-independent pathway (2, 7,18
19, 23, 24). In t he gl ioblastoma/ ast rocytoma cell l ine U251MG, an enhanced PKR19
response blocks the t ranslat ion of HIV st ructural proteins and inhibits viral product ion20
(59). Increasing levels of TRBP rescued the expression of HIV-1 proteins and virion21
product ion. This abil it y can be explained by the low endogenous TRBP1 and TRBP222
expression in primary ast rocytes and U251MG cells, which are unable to modulate PKR23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 5
act ivat ion (4, 59). The specif ic low expression of TRBP1 in ast rocytes is due, at least1
in part , to a lack of the NF-Y t ranscript ion factor in these cells (3). All available data2
indicate t hat TRBP proteins cont ribute t o t he high level of HIV-1 expression and3
repl icat ion in permissive cel ls and suggest t hat reducing TRBP expression could4
decrease HIV replicat ion (2).5
Recent data in the elucidat ion of the RNAi mechanism in mammalian cells have6
shown t hat t he Dicer protein is associated both wit h Ago2, a protein f rom the7
Argonaute family, and with TRBP (14, 36, 52, 75). The use of siRNAs directed against8
TRBP in funct ional assays has shown that TRBP is involved in the RNAi mechanism as a9
Dicer partner (14, 36, 67). This act ivit y may cont ribute to the role of TRBP during10
development . This recent discovery that TRBP is involved in both the RNAi mechanism11
and HIV repl icat ion raises t he quest ion of it s role during t he early st eps of HIV12
infect ion (31). In this study, we invest igated if a decrease in TRBP or Dicer expression13
could decrease HIV-1 product ion. We targeted TRBP and Dicer mRNAs by RNAi using14
siRNAs and show t hat inhibit ing t heir expression induces a decrease in HIV-115
expression and product ion in permissive cells to dif ferent extents.16
17
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 6
MATERIALS AND METHODS1
siRNA synthesis2
Target sequences for TRBP and EGFP were chosen using the Ambion software3
pr ogr am and t hei r spec i f i c i t y ver i f i ed by BLAST sear ch4
(www.ncbi.nlm.nih.gov/ BLAST). The non-silencing (NS) siRNA sequence was designed5
by Qiagen t o have no homology t o any mammalian gene. The siRNA sequences6
target ing HIV-1 Tat (SF2) protein and TAR RNA were from (16) and (41) respect ively.7
siRNA sequences are indicated in Table 1. All siRNAs t ransfected alongside TRBP8
siRNAs were synthesized in vi t ro using the Si lencerTM siRNA Const ruct ion Kit (Ambion9
Inc). Their concent rat ion and integrit y were verif ied by OD and gel analysis. The NS10
and Dicer siRNAs used to assess Dicer decrease were purchased from Qiagen.11
12
Semi-quantitative RT-PCR13
Total cel l RNA was ext racted using TRIzol® reagent 48 hr af ter t ransfect ion14
(Invit rogen). cDNA was made from 3 µg of total RNA in a 20 µL react ion containing 315
µM of annealed random hexamer, 100 mM DTT, 8 unit s AMV-RT (Promega), 1.25 mM16
dNTP and 40 units RNasin (Promega) at 42ºC for 1 hr. 1 µL of the cDNA template was17
used for PCR amplif icat ion in a 20 µL react ion containing 0.2 µL phusion polymerase18
(Finnzymes), 200 µM dNTP, 0.5 µM each of forward and reverse primers and a MgCl219
concent rat ion of eit her 1.5 mM for Dicer or 2.5 mM for GAPDH ampl if icat ion20
r espec t i ve l y . Pr i mer s used t o amp l i f y GAPDH w er e21
5’ TGAAGGTCGGAGTCAACGGATTTGGT3’ and 5’ CATGTGGGCCATGAGGTCCACCAC3’ ,22
whi l e pr i mer s f or Di cer wer e 5’ CTGAGCTTAGGAGATCTGAG3’ and23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 7
5’ GGAACCTGAGGTTGATTAGC3’ . Condit ions for amplif icat ion were 98ºC for 30 sec;1
either 23 cycles or 27 cycles of 98ºC 10 sec, 50ºC 20 sec, and 72ºC 30 sec for GAPDH2
or Dicer respect ively. The product s were resolved on a 1.5% agarose gel and3
quant if ied using Fuj i Film Image Gauge software.4
5
Plasmids6
pLTR-Luc, pCMV1-Tat (f rom HIV-1 SF2 st rain) (19) and pEGFP-C1-TRBP2 (59)7
expression plasmids were previously described. EGFP pre-miRNA sequence was8
derived by incorporat ion of t he EGFP siRNA sequence into the stem of the miR-309
miRNA as described (9). To generate pEGFP pre-miRNA vector, a casset te expressing10
EGFP pre-miRNA f rom t he U6+27 promot er (32) was made by t wo st ep PCR as11
described (12) and l igated into the pCRII -TOPO cloning vector (Invit rogen). The12
pNL4-3 (1) and pAD8 (76) proviral plasmids were obtained from M. Mart in (Nat ional13
Inst it ut e of Al lergy and Infect ious Diseases, Nat ional Inst it ut es of Healt h, USA).14
Proviral plasmids pELI-1, pMAL-2 (61) and pROD-10 (68) were obtained from K. Peden15
(Cent re for Biological Evaluat ion and Research, FDA, USA). Proviral plasmid p89.616
(18) was obt ained t hrough t he AIDS Research and Reference Reagent Program,17
Division of AIDS, NIAID, NIH, from R. J. Collman.18
19
Cells and transfections20
HeLa cel ls (ATCC) were maintained in Dulbecco’ s modif ied Eagle’ s medium21
(DMEM; Invit rogen) supplemented with 10% Fetal Bovine Serum (FBS; Hyclone), 2 mM22
L-Glutamine and 1% penicil l in-st reptomycin (Invit rogen). For t ransfect ions followed23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 8
by Fluorescence Act ivated Cell Sorter (FACS) analysis, 7.5 x 104 HeLa cel ls were1
seeded in 24-well plates 16 hr prior t o co-t ransfect ion with pEGFP-C1-TRBP2 and2
siRNA using Lipofect amineTM 2000 (Invit rogen). For t ransfect ions fol lowed by3
luciferase expression assay or immunoblot , 1.6 x 105 HeLa cells were plated in 6-well4
plates 16 hr prior t o t ransfect ions with siRNAs and the LTR-Luc plasmid or siRNAs5
alone using FuGENE 6 Reagent (Roche) at a 1:3 DNA/ RNA:FuGENE rat io. Luciferase6
expression was measured 48 hr post -t ransfect ion and normalized to the same amount7
of protein as previously described (19). Transfect ions of cel ls wit h HIV proviral8
const ruct s were eit her in T25 f lasks seeded with 7.5 x 105 cel ls or 6 well plates9
seeded wit h 3.0 x 105 cel ls 24 hr prior t o co-t ransfect ion wit h siRNA using10
LipofectamineTM 2000 (Invit rogen). To cont rol for t ransfect ion ef f iciency, eit her11
pEGFP-N1 (Clontech) was co-t ransfected as a t ransfect ion ef f iciency reporter and12
assessed by FACS, or experiments were performed at least three t imes to account for13
variat ions. Large variat ions in siRNA concent rat ions among the experiments are due14
to dif ferent experimental condit ions when using FuGENE 6 TM or lipofectamine TM 2000.15
Transfect ion ef f iciency was verif ied by PCR using 4 µl of cell lysate and 250 ng of16
luciferase primers t o ampl i f y a 456 nt DNA. The Luc sense primer was 5’ -17
CTATCCTCTAGAGGATGGAACC-3’ a n d t h e an t i sense w as 5 ’ -18
CGTCTACATCGACTGAAATCCC-3’ . Amplif icat ion was performed at 94oC 2 min, (94oC19
45 sec; 55oC 45 sec; 72oC 2 min) 30 cycles, 72oC 2 min. 10% of t he react ion was20
loaded on an agarose gel. Only t ransfect ions in which the ef f iciencies vary by less21
than 5% were considered for the average value calculat ion.22
23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 9
FACS analysis1
Cells were analyzed for EGFP expression on a FACsort (Becton-Dickinson), using2
the Cellquest cont rol sof tware (Becton-Dickinson). Transfected cells were gated by3
green f luorescence greater than cells in a mock t ransfect ion. Relat ive f luorescence4
values were calculated as the product of the percentage of EGFP f luorescent cells5
gated and the mean f luorescence of cells gated posit ive.6
For cell viabilit y, 2 x 105 HeLa cells were seeded in 6 well plates 24 hr prior to7
t ransfect ion wit h 14 nM of siRNA or 1.8 µg of poly(I) • poly(C) (Sigma) using 1 µl8
LipofectamineTM 2000 (Invit rogen). 48 hr later, cells were harvested with t rypsin, and9
st ained wit h 7-amino-act inomycin D (7-AAD) solut ion (Pharmingen) fol lowing10
manufacturer's protocol. The non-f luorescent populat ion represent ing viable cells11
was gated and calculated as a percentage of the total populat ion.12
13
Fluorescence14
HeLa cells were plated in 12-well plates on coverslips (Fischer Scient if ic) and15
were 70% conf luent at t he t ime of t ransfect ion. 100 nM of siRNAs were co-16
t ransfect ed wit h 0.5 µg pEGFP-C1-TRBP2 using FuGENE (Roche). 48 hr post -17
t ransfect ion, the cells were washed twice in phosphate-buffered saline (PBS). The18
cells were f ixed in a 4% paraformaldehyde solut ion for 10 min at room temperature19
followed by two washes in PBS. Fixed cells were mounted in Airvol (Air Products and20
Chemicals, Allentown, PA) and the f luorescence was detected on an Olympus BX-5121
microscope.22
23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 10
Measurement of mRNA stability1
Cells were grown in serum-free medium in the presence of 5 µg/ ml of either2
t he t ranscript ional inhibit or Act inomycin D (ActD; Sigma) or t he same volume of3
ethanol (cont rol). Af ter various t imes of ActD t reatment , cells were harvested and4
t otal RNA was isolated using the TRIzol® isolat ion t reatment (Invit rogen). RT-PCR5
react ion was performed as previously described (4). cDNA was synthesized from 5 µg6
of total RNA using 5 pmol of TRBP ant isense-(5’ -CTCAATGAAACGCTCCAC-3’ ) or c-myc7
ant isense primer (5’ -GGGGCTGGTGCATTTTCGGTTGTTGC-3’ ). PCR amplif icat ions8
were performed in a 100 µl react ion mixture containing 250 ng of each TRBP (5’ -9
CGGGTCACCGTTGGCGAC-3’ ) or c-myc primer (5’ -GCTCCTGGCAAAAGGTCAGAGTCTGG-10
3’ ). Ant isense primers were as described above for reverse react ion. To respect the11
PCR exponent ial phase, the PCR amplif icat ions for TRBP mRNA were performed with12
23 cycles. The products were resolved on a 1.5% agarose gel.13
14
HIV Reverse Transcriptase (RT) assay15
The reverse t ranscriptase assay was performed as previously described (39).16
Each react ion contained 6 µl viral supernatant in a 30 µl RT cocktail (60 mM Tris-HCl17
(pH7.8), 75 mM KCl, 5 mM MgCl2, 0.1% (w/ v) Nonidet P-40 (NP40, Fisons), 1 mM EDTA,18
5 µg/ ml poly-A, 162.5 ng/ ml oligo(dT), 4 µM dithiothreitol (DTT), 1 µCi/ ml ( -32P)19
dTTP (PerkinElmer) and was incubated at 37°C for 2 hr. 6 µl of each react ion was20
spot t ed ont o diet hyl aminoet hyl cel lulose (DEAE) f i l t er paper (What man21
Internat ional), which was subsequent ly washed four t imes in 2X SSC (3 M NaCl, 0.3 M22
t risodium cit rate (pH7.0)) and twice in 100% ethanol (ten min per wash) before being23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 11
air-dried and exposed to a Fuj if ilm BAS-MS-IP 2340 imaging plate, and reading on a1
phosphorimager (FLA3000 Fuj i Phot of i lm Co. ). Quant i t at ion of samples was2
performed using Fuj i Film Image Gauge software.3
4
Immunoblotting5
48 hr post -t ransfect ion, cells were washed twice with PBS and lysed in cold6
lysis buffer (50 mM Tris-HCl (pH 7.4), 150 mM NaCl, 5 mM EDTA (pH 8), 10% Glycerol,7
1% NP-40) with the protease inhibitor cocktail (Roche) and with phosphatase inhibitors8
(30 mM sodium f luoride, 10 mM p-nit rophenyphosphate, 40 mM ß-glycerophosphate9
and 1 mM sodium orthovanadate) when a phospho-specif ic ant ibody was used. The10
lysates were chilled on ice and cent rifuged for 15 min. Equivalent amounts of whole11
cel l ext ract , measured by Bradford assay (Bio-Rad), were separated by sodium12
dodecyl sulfate (SDS) polyacrylamide gel elect rophoresis (PAGE). The proteins were13
t ransferred to a Hybond ECL nit rocellulose membrane (Amersham) as described (6).14
The membrane was blocked for 1 hr in 5% nonfat milk and Tris-buffered saline-0.1%15
Tween 20 (TBST) (69) or 5% BSA and 0.1% TBST for ant i PKR and ant i-P-PKR, or in 5%16
milk/ PBST for HIV serum. The membranes were incubated overnight at 4˚ C with ant i-17
TRBP672 (21) at a 1/ 500 dilut ion, with ant i Dicer 349 (36) at a 1/ 1000 dilut ion, or18
with serum from a HIV-1 subtype B pat ient at a 1/ 5000 dilut ion in the corresponding19
buffers. They were incubated for 1 hr at room temperature with a monoclonal ant i-20
act in (Chemicon) at a 1/ 10000 dilut ion. For probing PKR and phosphorylated PKR,21
t hey were incubated overnight at 4˚ C with monoclonal ant i-PKR 71/ 10 (46, 56) or22
polyclonal ant i-P-PKR (Biosource) at a 1/ 1000 dilut ion in 3% BSA/ TBST. Af t er 523
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 12
washes in TBST or PBST, membranes were incubated with peroxidase-conj ugated1
secondary goat ant i-rabbit ant ibody (Amersham) for TRBP, P-PKR, Dicer, goat ant i-2
mouse (Amersham) f or PKR and act in, and rabbi t ant i -human ant ibody3
(DacoCytomat ion) for HIV-1 at a 1/ 10000 dilut ion. The bands were visualized as4
described (6).5
6
Northern blot analysis.7
Total RNA was harvested using TRIzol® reagent (Invit rogen), of which 10 µg was8
resolved in a 15% denaturing polyacrylamide 7M urea gel and then t ransferred by9
elect ro-blot t ing onto a GeneScreen Plus nylon membrane (NEN Life Sciences). An10
EGFP (5’ -GGGCATCGACTTCAAGGAG-3’ ) radiolabeled ol igo probe was hybridized in11
buffer (0.5 M sodium phosphate (pH 7.2), 7% (w/ v) SDS, 1 mM EDTA) to the membrane12
overnight at 42°C. Membranes were washed twice in wash buffer 1 (1X SSC, 1% (w/ v)13
SDS) for 20 min and twice in wash buffer 2 (0.5X SSC, 0.1% (w/ v) SDS) for 40 min at14
50°C and then exposed to imaging plates for 2 days. Blots were quant if ied using Fuj i15
Film Image Gauge software.16
17
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 13
RESULTS1
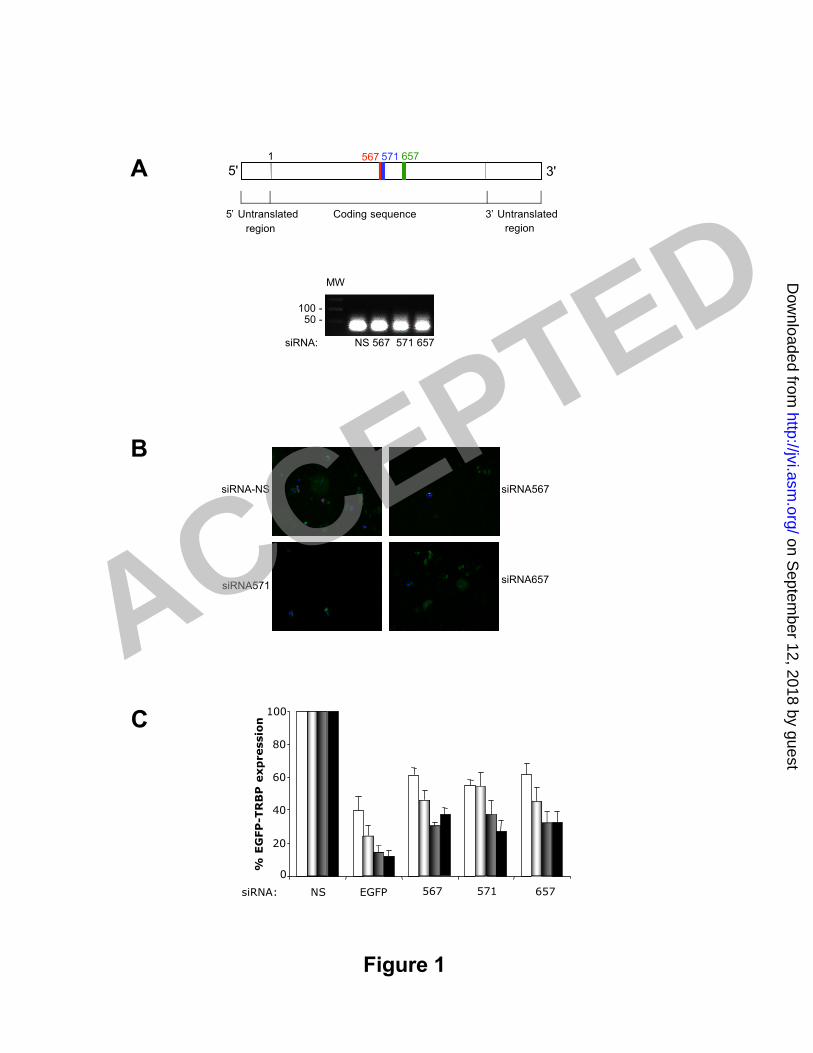
siRNAs against TRBP decrease exogenous TRBPs2
To analyze t he ef fect of decreasing TRBP protein on HIV-1 expression and3
replicat ion, six potent ial siRNAs against TRBP were synthesized and t ransfected in4
HeLa cells. The total RNA was ext racted and assayed by semi-quant itat ive RT-PCR for5
the amount of TRBP mRNA (data not shown). Three sequences that decreased TRBP6
mRNA signif icant ly were chosen (Fig 1A) and numbered according to the nucleot ide7
sequence start ing at the init iat ing AUG of TRBP2. All t hree sequences target both8
TRBP1 and TRBP2.9
To assess the act ivit y of t he selected siRNAs in decreasing the expression of10
TRBP, HeLa cells were t ransfected with a reporter plasmid expressing EGFP-TRBP211
fusion protein and the dif ferent siRNAs (Fig 1B). Compared to the NS siRNA cont rol all12
TRBP siRNAs decreased the expression of t ransfected EGFP-TRBP2 with increasing13
act ivit ies f rom siRNA 657 to 567 and 571. To quant ify these result s and assess the14
act ivit y of the siRNAs at dif ferent concent rat ions, reporter expression was measured15
by FACS analysis (Fig 1C). All three TRBP siRNAs decreased the expression of EGFP-16
TRBP from two to fourfold when compared to the siRNA-NS cont rol. siRNA571 had the17
most potent silencing act ivit ies at the highest concent rat ion.18
19
siRNAs against TRBP decrease endogenous TRBPs20
Because newly synthesized mRNA from t ransfected plasmid may have dif ferent21
accessibil it y and stabil it y compared with the endogenous mRNA, we next assessed22
TRBP mRNA stabil it y and the act ivit y of t he siRNAs to decrease endogenous TRBPs23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 14
(Fig. 2). We f irst determined TRBP mRNA half-life in the Jurkat lymphocyt ic cell l ine,1
which supports HIV replicat ion (Fig. 2A). The TRBP mRNA stabilit y was measured after2
t reat ing the cells with the t ranscript ional inhibitor Act inomycin D (ActD) between 13
and 10 hr. The amount of mRNA was then analyzed by RT-PCR and compared to4
cont rol cells without ActD (Fig. 2A, top); c-myc mRNA, which is known to be very5
unstable in lymphoblastoid as well as in HeLa cells (20, 66), was used as a reference.6
The decay of TRBP and c-myc mRNAs was plot ted as the percentage of the original7
amount of mRNA at t ime 0 (Fig. 2A, bot tom). The half-life of TRBP mRNA was about 38
hr, whereas the half-l ife of c-myc mRNA was less than 1 hr indicat ing that TRBP mRNA9
is about four t imes more stable than c-myc mRNA.10
The act ivit y of siRNAs to decrease endogenous TRBP1 and TRBP2 proteins was11
t hen evaluated in HeLa cells (Fig 2B). We used an ant ibody which recognizes both12
proteins as shown previously (3). The siRNA-NS did not decrease the concent rat ion of13
the endogenous proteins, but all three TRBP siRNAs did. The siRNA567 and siRNA65714
caused a part ial decrease in endogenous TRBP, whereas siRNA571 induced an almost15
complete inhibit ion of the protein. Because some siRNAs can act ivate the interferon16
pathway (10, 72), we verif ied if t he TRBP decrease by siRNA571 could be due to a17
t ranslat ional shut down by act ivated PKR. PKR phosphorylat ion remained at the same18
weak endogenous level whet her t he cel ls were t ransfect ed by NS or siRNA57119
indicat ing that the low TRBP amount is not due to PKR-induced t ranslat ion shut -down20
(Fig 2C). Previous reports with dif ferent siRNAs target ing TRBP showed an incomplete21
loss of TRBP protein (14, 36). Therefore, siRNA571 gives rise to a higher level of22
expression inhibit ion.23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 15
1
siRNAs against TRBP decrease the expression of the HIV-1 LTR.2
TRBP1 and TRBP2 were previously shown to inf luence HIV-1 gene expression and3
t o act in concert with Tat although at dif ferent levels (2). Therefore, we expected4
that a decrease in TRBPs would decrease HIV-1 basal LTR expression and Tat -mediated5
t rans-act ivat ion. The siRNAs directed against TRBPs were assayed on HIV-1 LTR6
expression in the absence and in the presence of Tat (Fig. 3). To bet ter evaluate their7
act ivit y, they were compared to the previously described siRNAs directed against TAR8
or Tat RNAs (16, 41). Whereas siRNA657 had lit t le act ivit y, siRNA-TAR, siRNA567 and9
siRNA571 showed a twofold inhibit ion of LTR basal expression (Fig. 3A). These results10
indicate that TRBP or TAR inact ivat ion has the same effect on LTR basal expression.11
This is consist ent wit h siRNAs-TAR that have a maximum of 50% reduct ion in a12
comparable luciferase reporter gene assay due to the t ight TAR RNA st ructure (79). In13
the context of Tat t rans-act ivat ion, siRNA657 had the same act ivit y as siRNA-TAR with14
an approximately twofold reduct ion in LTR expression, whereas siRNA567 showed a15
threefold reduct ion (Fig. 3B). The act ivit y of siRNA571 was close to that of siRNA-Tat16
wit h a t enfold reduct ion over t he NS cont rol . Overal l , t he decrease in TRBP17
expression by siRNAs result s in a 50 to 90% reduct ion of HIV-1 LTR expression in the18
presence of the Tat t ransact ivator.19
20
siRNAs against TRBP decrease HIV-1 production in transfected cells.21
The act ivit y of siRNAs against TRBP was next evaluated for t heir act ivit y t o22
decrease HIV-1 product ion. HeLa cel ls were t ransfected wit h t he previous TRBP23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 16
siRNAs, or another siRNA (Tat1c) target ing a region of Tat present in the HIV molecular1
clone pNL4-3. Virus product ion was monitored by RT assay and the expression of2
int racellular viral proteins evaluated by western blot (Fig. 4). In agreement with3
previous result s (16) the Tat siRNA was very ef fect ive at inhibit ing HIV-1 product ion4
with a twenty-fold reduct ion of RT act ivity (Fig. 4A). siRNAs against TRBP also showed5
high act ivit y, with siRNA567 and siRNA571 inhibit ing RT act ivit y by four to f ive-fold6
(lanes 4 and 5) and siRNA657 by twofold (lane 6). Int racel lular viral proteins,7
part icularly t he viral capsid p24, were mildly decreased with siRNA657 but highly8
decreased with siRNA567 and siRNA571 compared to the negat ive cont rol (Fig 4B).9
Unsurprisingly, target ing virtually all viral spliced mRNAs, except Nef1, one of f ive Nef10
mRNAs (63), the Tat1c siRNA abolished all viral protein product ion.11
To determine if the reduced expression of viral proteins and virion product ion12
could be ascribed to an increased PKR act ivat ion, we measured PKR expression and13
PKR act ivat ion on the same ext racts (Fig 4C). In assays with siRNA Tat , siRNA567 and14
571, PKR expression was increased twofold compared to the siRNA-NS suggest ing a15
part ial act ivat ion of the interferon pathway likely due to the concomitant presence of16
viral RNA and act ive siRNAs. In paral lel, phosphorylated PKR increased in similar17
proport ion suggest ing t hat act ivated PKR is only a moderate component of t he18
reduced HIV product ion. These resul t s demonst rat e t hat decreased levels of19
endogenous TRBP induce a signi f icant reduct ion in vi ral prot ein synt hesis.20
Subsequent ly, viral product ion is inhibited to levels comparable with direct ly reducing21
Tat expression.22
23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 17
siRNAs against TRBP decrease HIV production of various strains.1
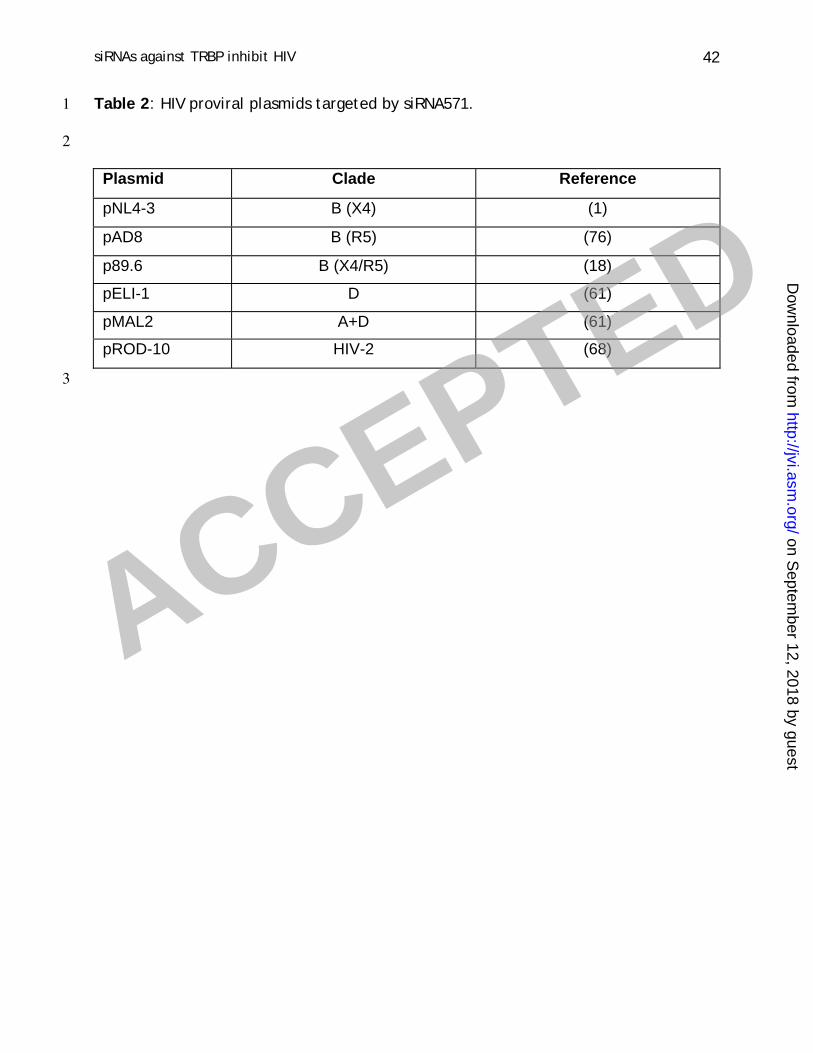
One advantage to t arget cel lular factors required for HIV replicat ion is t he2
possibil i t y t o t arget many viral st rains wit h t he same sequence. We t herefore3
assessed if decreasing TRBP concent rat ion wil l also af fect t he product ion of other4
lymphot ropic (X4) or macrophage-t ropic (R5) HIV-1 st rains as well as an HIV-2 st rain.5
The HIV st rains that were assayed are summarized in Table 2. All HIV st rains showed a6
decreased virus product ion in the presence of siRNA571 compared to siRNA-NS (Fig. 5).7
The HIV-2 pROD-10 st rain showed a 40% decrease indicat ing that TRBP is also required8
to some extent for this virus. The pELI-1 (HIV-1 D clade) was the most affected with a9
90% reduct ion in virus product ion indicat ing a st rong TRBP requirement . Overall, HIV-10
1 virion product ion was reduced 60-90% by a decrease in TRBP.11
12
siRNAs against Dicer decrease HIV-1 production in transfected cells.13
To determine if t he act ivit y of siRNAs against TRBP could be ascribed to it s14
act ivit y with HIV RNA or to it s funct ion in RISC, we targeted another member of the15
RISC. It has previously been shown that reduced Dicer expression diminishes the16
act ivit y of RISC (40). We evaluated the act ivit y of siRNA against Dicer by RT-PCR and17
western blot (Fig. 6A) and found that t ransfect ion of cells with 14 nM of siRNA Dicer18
using Lipofectamine ef fect ively reduced both Dicer mRNA and protein levels. To19
assess the ef fect s t his decreased Dicer expression had on viral expression we co-20
t ransfected the HIV molecular clone pNL4-3 with siRNADicer (Fig. 6B). Surprisingly,21
HIV product ion was reduced up t o 40% suggest ing t hat t he act ivit y of t he RISC22
cont ributes to, rather than inhibits, viral product ion.23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 18
1
siRNAs against TRBP or Dicer do not affect cell viability.2
To determine if some or al l act ivit y of siRNAs against TRBP or Dicer on HIV3
expression and product ion could be at t ributed to a non-specif ic cytotoxic ef fect we4
measured cell viabil it y of t ransfected cells. Using 7-AAD, a f luorescent dye taken up5
by non-viable cel ls, we were able t o discriminate between viable and non-viable6
t ransfected cell populat ions based on non-f luorescence (71). The result s show that7
the siRNAs do not reduce cell viabil it y at the concent rat ions used in this study, while8
the poly(I) • poly(C) cont rol signif icant ly reduced cell viabilit y (Fig. 7A).9
10
siRNAs against TRBP or Dicer decrease the processing of miRNAs against GFP.11
Because siRNA-TRBP and siRNA-Dicer affect HIV-1 product ion, this act ivity could12
be due to their funct ion in RNAi or to another funct ion. To discriminate between13
t hese, we assessed t he ext ent t o which t he respect ive siRNAs af fect t he RNAi14
pathway. Using a system reliant on the processing of a vector delivered EGFP pre-15
miRNA into mature miRNA we direct ly assessed the effects various siRNAs had on the16
product ion of EGFP miRNA species by nort hern blot . SiRNA567 and siRNA57117
decreased the miRNA processing by 23 and 41% respect ively, whereas siRNA-Dicer18
decreased it by 73% compared to the siRNA-NS. Therefore, TRBP siRNAs disrupt the19
RNAi pathway part ially while Dicer siRNA largely impedes it . 20
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 19
DISCUSSION1
RNAi has been widely used within the last f ive years to inhibit gene expression2
in mammalian cells. Applicat ions range from fundamental knowledge in genomics to3
t herapeut ics in disease-associat ed genes (51). In paral lel , t he int ricat e RNAi4
mechanism is being elucidated to understand how si- or miRNAs funct ion in the cells5
and recent progress has been made t o elucidate t he dif ferences between lower6
eukaryotes and mammalian cells (14, 34, 36, 52, 54). It has become clear within the7
last year that Dicer, Ago2 and TRBP are the main components of the mammalian RISC8
that t rigger either RNA cleavage or t ranslat ion inhibit ion, although other proteins have9
been ident if ied in this complex (14, 36, 52). Both TRBP1 and TRBP2 bind to Dicer in10
several assays and the two isoforms may play a similar role in the RISC (36).11
Because of the high eff iciency of siRNAs, they have been used to target viruses12
and combat viral diseases. Viruses whose repl icat ion has been silenced include13
Respiratory Syncyt ial , Inf luenza, Pol io, Herpes, Hepat it is B, Hepat it is C, Human14
Papil loma, Human Immunodef iciency viruses and a growing l ist of other viruses (17,15
37). In the choice of targets against a virus, the best one seems to be the virus it self16
because a specif ic target will less likely damage the cell. However, one concern with17
HIV is it s high mutat ion rate that induces drug resistance. Indeed, a single mismatch18
in the sequence can decrease or inhibit the act ivit y of siRNAs (41, 64). In support of19
these concerns, HIV was shown to escape RNAi by mutat ing the targeted sequence (8,20
77). The choice of a highly conserved sequence or the use of mult iple targets are21
ways to overcome this inconvenience but may also have some limitat ions. Target ing22
cel lular fact ors required for HIV repl icat ion is anot her way t o circumvent t his23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 20
problem. This st rategy has already been used against HIV by inhibit ing cel lular1
receptors, expression factors and factors required for viral t raf f icking and packaging.2
In each case, the inhibit ion of the targeted cellular factor decreased HIV product ion3
and did not damage the cell.4
TRBP is a cellular factor that enhances HIV replicat ion by it s act ivit y on HIV-15
gene expression and more specif ically on t ranslat ion (7, 19, 23, 24, 29). We recent ly6
showed that ast rocytes that have a low level of TRBP expression have a concomitant7
low level of HIV mRNA t ranslat ion and poor HIV replicat ion (3, 4, 33, 59). Ast rocytes8
and live Tarbp2-/ - mice (81) indicate that cells can live with low or no TRBP protein.9
Therefore, it seemed l ikely t hat inact ivat ing TRBP should be an ef fect ive way to10
decrease HIV replicat ion. The recent elucidat ion of components of RISC required for11
RNAi in mammalian cel ls showed t hat TRBP is a Dicer part ner and a necessary12
component of the RNAi mechanism (14, 36, 67). Because RNAi may be part of the cell13
react ion against viruses in mammalian cells, it became less obvious that decreasing14
TRBP may result in HIV reduced replicat ion (31, 70, 78).15
The results shown in this paper indicate that t ransfected and endogenous TRBPs16
can be effect ively decreased by RNAi, using dif ferent t ransfect ion protocols (Fig. 1-2).17
In previous cellular set t ings (14, 36), the inhibit ion was good but not complete, l ikely18
because TRBP is part of the RISC and because the mRNA and the protein are quite19
stable. Here, siRNA571 gave a high decrease in protein concent rat ion, indicat ing that20
ef fect ive inhibit ion can be achieved (Fig. 2). Because TRBP overexpression can21
increase HIV-1 LTR basal expression (19, 24, 29), we expected that it s inhibit ion may22
lower this level. The results show that inhibit ing TRBP mRNA by siRNAs decreases HIV23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 21
LTR basal expression (Fig. 3A) to a mild extent similar to that induced by siRNA-TAR in1
t he best case. This twofold ef fect of siRNA571 may be mainly t he result of an2
unrepressed t ranslat ion cont rol mediated by TAR (23), rather than by an increased3
PKR phosphorylat ion as siRNA571 does not act ivate PKR (Fig. 2C). In cont rast , in the4
presence of Tat , the act ivit y of siRNA571 on HIV-1 LTR t rans-act ivated expression was5
close to the act ivit y of siRNA-Tat and the weakest siRNA657 had an act ivit y similar to6
siRNA-TAR (Fig. 3B). These result s suggest t hat TRBP cont ributes t o t he t rans-7
act ivated level of HIV-1 expression to a higher extent than to the basal level, which is8
compat ible with the observed synergist ic ef fect between TRBP and Tat (29). The9
comparison between the ef f iciency of siRNA-TAR and siRNA571 on the basal and the10
t ransact ivated level suggest s t hat siRNA-TAR has a consist ent twofold decrease.11
Indeed various TAR siRNAs have been extensively studied and were shown to have a12
maximum eff iciency of 50% in a comparable luciferase gene reporter assay due to the13
highly st ructured TAR RNA prevent ing access to the siRNA-TAR (79). Because TRBP14
act s mainly t o increase t ranslat ion of mRNAs, i t is l ikely t hat by reducing it s15
concent rat ion in cells, the remaining amount is almost suff icient to t ranslate the LTR16
basal level, but in l imit ing amount to t ranslate the large amount of mRNA present17
after t rans-act ivat ion (compare Fig. 3A lane 5 with 3B lane 6).18
These result s suggest that TRBP cont ributes largely to HIV expression and we19
assayed TRBP siRNAs on HIV-1 product ion. In this context , the decreased expression20
of TRBP reduced expression of HIV-1 proteins and format ion of viral part icles (Fig. 4).21
This rest rict ion was also observed with dif ferent HIV-1 st rains and HIV-2, indicat ing22
that TRBP is a protein required for all st rains (Fig. 5). Together with studies that show23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 22
overexpression of TRBP can overcome the t ranslat ion and replicat ion block caused by1
act ivated PKR in lymphocytes (7) and ast rocytes (59), our result s show TRBP is2
important to HIV replicat ion. This requirement is l ikely due to all TRBP act ivit ies on3
PKR inhibit ion (7, 19), increased t ranslat ion of TAR containing RNAs (23), PACT4
inhibit ion (G. Laraki, A. Daher and A. Gat ignol, unpublished data) and possibly it s5
funct ion in RNAi (14, 36). Results in Figure 4C show a moderate increase in PKR and6
act ivated PKR with both siRNA Tat and siRNA TRBP, demonst rat ing that increased PKR7
act ivat ion only part ly cont ributes t o reduced HIV product ion. Furt hermore, t he8
act ivit y of TRBP siRNAs cannot be ascribed to a loss of cell viabilit y (Fig. 7A) and their9
modest impediment of t he miRNA biogenesis pathway (Fig. 7B) cannot explain the10
ent ire act ivit y. The results in the context of Tat t rans-act ivat ion and HIV product ion11
show comparable act ivity (Fig. 3B and 4A). Therefore, the overall results suggest that12
the act ivit y of TRBP siRNAs on HIV expression and product ion can be mainly ascribed13
t o t he TRBP funct ions other t han RNAi. This study st rongly suggests t hat , in t he14
context of HIV replicat ion, TRBP is more important to help HIV replicat ion than to15
rest rict viral RNA by RNAi as suggested by other experiments (31, 59). To determine16
more precisely if RNAi funct ion cont ributes to HIV replicat ion, we also decreased17
Dicer. Surprisingly, we did not obtain an increase in viral product ion that would have18
supported a role for RNAi in HIV rest rict ion. Instead, l ike with the TRBP siRNAs, we19
obtained a decrease in HIV-1 product ion, although to a lesser extent (Fig. 6). Because20
Dicer siRNA largely impedes miRNA processing (Fig. 7), we conclude that , in t his21
cellular and viral context , RNAi does not rest rict HIV replicat ion (31, 70, 78), rather it22
cont ributes moderately to virus product ion, suggest ing that similar to HCV, HIV uses23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 23
RNAi for it s own benef it rather t han being rest ricted by RNA cleavage (42). The1
mechanism by which this funct ion is accomplished, and if it can also be observed in2
lymphocytes and macrophages, which are the natural targets of HIV, remains to be3
determined.4
5
ACKNOWLEDGMENTS6
We would l ike to thank E. Meurs and W. Fil ipowicz for the ant i-PKR and the7
ant i-Dicer ant ibodies. We also thank M. Mart in, K. Peden and R. Collman for HIV8
proviral plasmids. We are grat eful t o A. Mouland and R. Ramsay for helpful9
discussions and comments. This work was supported by the Canadian Inst it ute for10
Health Research (CIHR) grant HOP38112 to AG and by Nat ional Health and Medical11
Research Council (NHMRC) of Aust ralia proj ect grant 400302 to DP and by the Early12
Career Researchers Grant Scheme to SMC. SB was supported by a CIHR postdoctoral13
fellowship. AG is the recipient of a Hugh and Helen McPherson memorial award.14
The authors declare that they have no compet ing f inancial interests.15 ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 24
REFERENCES1
1. Adachi, A., H. E. Gendelman, S. Koenig, T. Folks, R. L. Willey, A. Rabson,2
and M. A. Martin. 1986. Product ion of acquired immunodef iciency syndrome-3
associat ed ret rovirus in human and nonhuman cel ls t ransfect ed wit h an4
infect ious molecular clone. J. Virol. 59:284-291.5
2. Bannwarth, S., and A. Gatignol. 2005. HIV-1 TAR RNA: the target of molecular6
interact ions between the virus and its host . Curr. HIV Res. 3:61-71.7
3. Bannwarth, S., S. Lainé, A. Daher, N. Grandvaux, G. Clerzius, A. C. LeBlanc,8
J. Hiscott, and A. Gatignol. 2006. Cell-specif ic regulat ion of TRBP1 promoter9
by NF-Y t ranscript ion fact or in lymphocyt es and ast rocyt es. J. Mol. Biol .10
355:898-910.11
4. Bannwarth, S. , L. Talakoub, F. Letourneur, M. Duarte, D. F. Purcell, J.12
Hiscott, and A. Gatignol. 2001. Organizat ion of the human tarbp2 gene reveals13
two promoters t hat are repressed in an ast rocyt ic cel l l ine. J. Biol. Chem.14
276:48803-48813.15
5. Bass, B. L. 2001. RNA interference. The short answer. Nature 411:428-429.16
6. Battisti, P.-L. , A. Daher, S. Bannwarth, J. Voortman, K. W. C. Peden, J.17
Hiscott, A. J. Mouland, R. Benarous, and A. Gatignol. 2003. Addit ive Act ivit y18
between the Trans-Act ivat ion Response RNA-Binding Protein, TRBP2, and Cyclin19
T1 on HIV Type 1 Expression and Viral Product ion in Murine Cells. AIDS Res.20
Hum. Ret roviruses. 19:767- 778.21
7. Benkirane, M. , C. Neuveut, R. F. Chun, S. M. Smith, C. E. Samuel, A.22
Gatignol, and K.-T. Jeang. 1997. Oncogenic potent ial of TAR RNA-binding23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 25
protein TRBP and it s regulatory interact ion with Protein Kinase PKR. EMBO J.1
16:611-624.2
8. Boden, D., O. Pusch, F. Lee, L. Tucker, and B. Ramratnam. 2003. Human3
immunodef iciency virus t ype 1 escape f rom RNA int erference. J. Virol .4
77:11531-11535.5
9. Boden, D., O. Pusch, R. Silbermann, F. Lee, L. Tucker, and B. Ramratnam.6
2004. Enhanced gene silencing of HIV-1 specif ic siRNA using microRNA designed7
hairpins. Nuc. Acids Res. 32:1154-1158.8
10. Bridge, A. J. , S. Pebernard, A. Ducraux, A. L. Nicoulaz, and R. Iggo. 2003.9
Induct ion of an interferon response by RNAi vectors in mammalian cells. Nat .10
Genet . 34:263-264.11
11. Capodici, J. , K. Kariko, and D. Weissman. 2002. Inhibit ion of HIV-1 infect ion12
by small interfering RNA-mediated RNA interference. J. Immunol. 169:5196-13
5201.14
12. Castanotto, D., H. Li, and J. J. Rossi. 2002. Funct ional siRNA expression from15
t ransfected PCR products. RNA 8:1454-1460.16
13. Chatel-Chaix, L. , J. F. Clement, C. Martel, V. Beriault, A. Gatignol, L.17
DesGroseillers, and A. J. Mouland. 2004. Ident if icat ion of Staufen in t he18
human immunodef iciency virus t ype 1 Gag ribonucleoprotein complex and a19
role in generat ing infect ious viral part icles. Mol. Cell. Biol. 24:2637-2648.20
14. Chendrimada, T. P., R. I. Gregory, E. Kumaraswamy, J. Norman, N. Cooch,21
K. Nishikura, and R. Shiekhattar. 2005. TRBP recruit s the Dicer complex to22
Ago2 for microRNA processing and gene silencing. Nature 436:740-744.23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 26
15. Chiu, Y. L. , H. Cao, J. M. Jacque, M. Stevenson, and T. M. Rana. 2004.1
Inhibi t ion of human immunodef iciency virus t ype 1 repl icat ion by RNA2
interference directed against human t ranscript ion elongat ion factor P-TEFb3
(CDK9/ CyclinT1). J. Virol. 78:2517-2529.4
16. Coburn, G. A., and B. R. Cullen. 2002. Potent and specif ic inhibit ion of human5
immunodef iciency virus t ype 1 repl icat ion by RNA int erference. J. Virol .6
76:9225-9231.7
17. Colbere-Garapin, F. , B. Blondel, A. Saulnier, I. Pelletier, and K. Labadie.8
2005. Silencing viruses by RNA interference. Microbes Infect . 7:767-775.9
18. Collman, R. , J. W. Balliet, S. A. Gregory, H. Friedman, D. L. Kolson, N.10
Nathanson, and A. Srinivasan. 1992. An infect ious molecular clone of an11
unusual macrophage-t ropic and highly cyt opat hic st rain of human12
immunodeficiency virus type 1. J. Virol. 66:7517-7521.13
19. Daher, A. , M. Longuet, D. Dorin, F. Bois, E. Segeral, S. Bannwarth, P.-L.14
Battisti, D. Purcell, R. Benarous, C. Vaquero, E. F. Meurs, and A. Gatignol.15
2001. Two dimerizat ion domains in the Trans-act ivat ion response RNA-binding16
Protein (TRBP) individually reverse the Protein Kinase R inhibit ion of HIV-1 Long17
Terminal Repeat expression. J. Biol. Chem. 276:33899-33905.18
20. Dani, C., J. M. Blanchard, M. Piechaczyk, S. El Sabouty, L. Marty, and P.19
Jeanteur. 1984. Ext reme instabil it y of myc mRNA in normal and t ransformed20
human cells. Proc. Nat l. Acad. Sci. USA 81:7046-7050.21
21. Daviet, L., M. Erard, D. Dorin, M. Duarte, C. Vaquero, and A. Gatignol. 2000.22
The analysis of a binding dif ference between the two dsRNA binding domains in23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 27
TRBP reveals t he modular funct ion of a KR-hel ix mot if . Eur. J. Biochem.1
267:2419-2431.2
22. Dillon, C. P., P. Sandy, A. Nencioni, S. Kissler, D. A. Rubinson, and L. Van3
Parij s. 2005. RNAi as an experimental and t herapeut ic t ool t o st udy and4
regulate physiological and disease processes. Annu. Rev. Physiol. 67:147-173.5
23. Dorin, D. , M. C. Bonnet, S. Bannwarth, A. Gatignol, E. F. Meurs, and C.6
Vaquero. 2003. The TAR RNA-binding protein, TRBP, st imulates the expression7
of TAR-containing RNAs in vi t ro and in vivo independent ly of it s abil it y t o8
inhibit the dsRNA-dependent kinase PKR. J. Biol. Chem. 278:4440-4448.9
24. Duarte, M., K. Graham, A. Daher, P.-L. Battisti, S. Bannwarth, E. Segeral, K.-10
T. Jeang, and A. Gatignol. 2000. Characterizat ion of TRBP1 and TRBP2: stable11
stem-loop st ructure at the 5' end of TRBP2 mRNA resembles HIV-1 TAR and is12
not found in it s processed pseudogene. J. Biomed. Sci. 7:494-506.13
25. Elbashir, S. M., J. Harborth, W. Lendeckel, A. Yalcin, K. Weber, and T.14
Tuschl. 2001. Duplexes of 21-nucleot ide RNAs mediate RNA interference in15
cultured mammalian cells. Nature 411:494-498.16
26. Erard, M., D. G. Barker, F. Amalric, K.-T. Jeang, and A. Gatignol. 1998. An17
Arg/ Lys-rich core pept ide mimics TRBP binding to the HIV-1 TAR RNA upper-18
stem/ loop. J. Mol. Biol. 279:1085-1099.19
27. Filipowicz, W. 2005. RNAi: the nuts and bolts of the RISC machine. Cell 122:17-20
20.21
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 28
28. Gatignol, A. , C. Buckler, and K.-T. Jeang. 1993. Relatedness of an RNA1
binding mot if in HIV-1 TAR RNA binding protein TRBP to human P1/ dsI kinase2
and Drosophila Staufen. Mol. Cell. Biol. 13:2193-2202.3
29. Gatignol, A. , A. Buckler-White, B. Berkhout, and K. -T. Jeang. 1991.4
Characterizat ion of a human TAR RNA-binding protein that act ivates the HIV-15
LTR. Science 251:1597-1600.6
30. Gatignol, A. , M. Duarte, L. Davie t, Y. -N. Chang, and K.-T. Jeang. 1996.7
Sequent ial steps in Tat t rans-act ivat ion of HIV-1 mediated through cellular DNA,8
RNA, and protein binding factors. Gene Expr. 5:217-228.9
31. Gatignol, A. , S. Lainé, and G. Clerzius. 2005. Dual role of TRBP in HIV10
repl icat ion and RNA interference: viral diversion of a cel lular pathway or11
evasion from ant iviral immunity? Ret rovirology 2:65.12
32. Good, P. D. , A. J. Krikos, S. X. Li, E. Bertrand, N. S. Lee, L. Giver, A.13
Ellington, J. A. Zaia, J. J. Rossi, and D. R. Engelke. 1997. Expression of small,14
therapeut ic RNAs in human cell nuclei. Gene Ther. 4:45-54.15
33. Gorry, P. R., J. L. Howard, M. J. Churchill, J. L. Anderson, A. Cunningham,16
D. Adrian, D. A. McPhee, and D. F. Purcell. 1999. Diminished product ion of17
human immunodef iciency virus t ype 1 in ast rocytes result s f rom inef f icient18
t ranslat ion of gag, env, and nef mRNAs despite eff icient expression of Tat and19
Rev. J. Virol. 73:352-361.20
34. Gregory, R. I. , T. P. Chendrimada, N. Cooch, and R. Shiekhattar. 2005.21
Human RISC couples microRNA biogenesis and post t ranscript ional gene22
silencing. Cell 123:631-640.23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 29
35. Guo, F. , S. Cen, M. Niu, H. Javanbakht, and L. Kleiman. 2003. Specif ic1
inhibit ion of the synthesis of human lysyl-tRNA synthetase results in decreases2
in t RNA(Lys) incorporat ion, tRNA(3)(Lys) anneal ing t o viral RNA, and viral3
infect ivity in human immunodeficiency virus type 1. J. Virol. 77:9817-9822.4
36. Haase, A. D., L. Jaskiewicz, H. Zhang, S. Lainé, R. Sack, A. Gatignol, and W.5
Filipowicz. 2005. TRBP, a regulator of cellular PKR and HIV-1 virus expression,6
interacts with Dicer and funct ions in RNA silencing. EMBO Rep. 6:961-967.7
37. Haasnoot, J. , and B. Berkhout. 2006. RNA interference: it s use as ant iviral8
therapy. Handb. Exp. Pharmacol. 173:117-150.9
38. Hammond, S. M. 2005. Dicing and sl icing: t he core machinery of t he RNA10
interference pathway. FEBS Let t . 579:5822-5829.11
39. Huang, L. M., A. Joshi, R. Willey, J. Orenstein, and K.-T. Jeang. 1994. Human12
immunodef iciency viruses regulated by alt ernat ive t rans-act ivators: genet ic13
evidence for a novel non-t ranscript ional funct ion of Tat in virion infect ivit y.14
EMBO J. 13:2886-2896.15
40. Hutvagner, G., J. McLachlan, A. E. Pasquinelli, E. Balint, T. Tuschl, and P. D.16
Zamore. 2001. A cellular funct ion for the RNA-interference enzyme Dicer in the17
maturat ion of the let -7 small temporal RNA. Science 293:834-838.18
41. Jacque, J. M. , K. Triques, and M. Stevenson. 2002. Modulat ion of HIV-119
replicat ion by RNA interference. Nature 418:435-438.20
42. Jopling, C. L. , M. Yi, A. M. Lancaster, S. M. Lemon, and P. Sarnow. 2005.21
Modulat ion of hepat it is C virus RNA abundance by a l iver-specif ic MicroRNA.22
Science 309:1577-1581.23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 30
43. Kameoka, M., S. Nukuzuma, A. Itaya, Y. Tanaka, K. Ota, K. Ikuta, and K.1
Yoshihar a. 2004. RNA int erference direct ed against Poly(ADP-Ribose)2
polymerase 1 ef f icient ly suppresses human immunodef iciency virus t ype 13
replicat ion in human cells. J. Virol. 78:8931-8934.4
44. Kharrat, A. , M. J. Macias, T. J. Gibson, M. Nilges, and A. Pastore. 1995.5
St ructure of the dsRNA binding domain of E.coli RNase III. EMBO J. 14:3572-6
3584.7
45. Komano, J., K. Miyauchi, Z. Matsuda, and N. Yamamoto. 2004. Inhibit ing the8
Arp2/ 3 complex limits infect ion of both int racellular mature vaccinia virus and9
primate lent iviruses. Mol. Biol. Cell. 15:5197-5207.10
46. Laurent, A. G., B. Krust, J. Galabru, J. Svab, and A. G. Hovanessian. 1985.11
Monoclonal ant ibodies to an interferon-induced Mr 68,000 protein and their use12
for the detect ion of double-st randed RNA-dependent protein kinase in human13
cells. Proc. Nat l. Acad. Sci. USA 82:4341-4345.14
47. Lee, J. Y., H. Kim, C. H. Ryu, J. Y. Kim, B. H. Choi, Y. Lim, P. W. Huh, Y. H.15
Kim, K. H. Lee, T. Y. Jun, H. K. Rha, J. K. Kang, and C. R. Choi. 2004. Merlin,16
a t umor suppressor, int eract s wit h t ransact ivat ion-responsive RNA-binding17
protein and inhibits it s oncogenic act ivity. J. Biol. Chem. 279:30265-30273.18
48. Lee, J. Y., H. J. Moon, W. K. Lee, H. J. Chun, C. W. Han, Y. W. Jeon, Y. Lim,19
Y. H. Kim, T. P. Yao, K. H. Lee, T. Y. Jun, H. K. Rha, and J. K. Kang. 2006.20
Merlin facil it ates ubiquit inat ion and degradat ion of t ransact ivat ion-responsive21
RNA-binding protein. Oncogene 25:1143-1152.22
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 31
49. Lee, K. , M. A. Fajardo, and R. E. Braun. 1996. A t est is cytoplasmic RNA-1
binding protein that has the propert ies of a t ranslat ional repressor. Mol. Cell.2
Biol. 16:3023-3034.3
50. Lee, N. S., T. Dohjima, G. Bauer, H. Li, M. J. Li, A. Ehsani, P. Salvaterra, and4
J. Rossi. 2002. Expression of small interfering RNAs targeted against HIV-1 rev5
t ranscripts in human cells. Nat . Biotechnol. 20:500-505.6
51. Leung, R. K. , and P. A. Whittaker. 2005. RNA int erference: f rom gene7
silencing to gene-specif ic therapeut ics. Pharmacol. Ther. 107:222-239.8
52. Liu, J., M. A. Carmell, F. V. Rivas, C. G. Marsden, J. M. Thomson, J. J. Song,9
S. M. Hammond, L. Joshua-Tor, and G. J. Hannon. 2004. Argonaute2 is the10
catalyt ic engine of mammalian RNAi. Science 305:1437-1441.11
53. Liu, S. , M. Asparuhova, V. Brondani, I. Ziekau, T. Klimkait , and D.12
Schumperli. 2004. Inhibit ion of HIV-1 mult iplicat ion by ant isense U7 snRNAs and13
siRNAs target ing cyclophilin A. Nuc. Acids Res. 32:3752-3759.14
54. Maniataki, E. , and Z. Mourelatos. 2005. A human, ATP-independent , RISC15
assembly machine fueled by pre-miRNA. Genes Dev. 19:2979-2990.16
55. Martinez, M. A. , A. Gutierrez, M. Armand-Ugon, J. Blanco, M. Parera, J.17
Gomez, B. Clotet, and J. A. Este. 2002. Suppression of chemokine receptor18
expression by RNA interference allows for inhibit ion of HIV-1 replicat ion. AIDS19
16:2385-2390.20
56. Meurs, E. F., Y. Watanabe, S. Kadereit, G. N. Barber, M. G. Katze, K. Chong,21
B. R. Williams, and A. G. Hovanessian. 1992. Const itut ive expression of human22
double-st randed RNA-act ivat ed p68 kinase in murine cel ls mediat es23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 32
phosphorylat ion of eukaryot ic init iat ion factor 2 and part ial resist ance t o1
encephalomyocardit is virus growth. J. Virol. 66:5805-5814.2
57. Murray, J. L., M. Mavrakis, N. J. McDonald, M. Yilla, J. Sheng, W. J. Bellini,3
L. Zhao, J. M. Le Doux, M. W. Shaw, C. C. Luo, J. Lippincott-Schwartz, A.4
Sanchez, D. H. Rubin, and T. W. Hodge. 2005. Rab9 GTPase is required for5
replicat ion of human immunodef iciency virus t ype 1, f iloviruses, and measles6
virus. J. Virol. 79:11742-11751.7
58. Novina, C. D., M. F. Murray, D. M. Dykxhoorn, P. J. Beresford, J. Riess, S. K.8
Lee, R. G. Collman, J. Lieberman, P. Shankar, and P. A. Sharp. 2002. siRNA-9
directed inhibit ion of HIV-1 infect ion. Nat . Med. 8:681-686.10
59. Ong, C. L. , J. C. Thorpe, P. R. Gorry, S. Bannwarth, A. Jaworowski, J. L.11
Howard, S. Chung, S. Campbell, H. S. Christensen, G. Clerzius, A. J.12
Mouland, A. Gatignol, and D. F. Purcell. 2005. Low TRBP levels support an13
innat e human immunodef iciency virus t ype 1 resist ance in ast rocyt es by14
enhancing the PKR ant iviral response. J. Virol. 79:12763-12772.15
60. Patel, R. C., and G. C. Sen. 1998. PACT, a protein act ivator of the interferon-16
induced protein kinase, PKR. EMBO J. 17:4379-4390.17
61. Peden, K. , M. Emerman, and L. Montagnier. 1991. Changes in growt h18
propert ies on passage in t issue cult ure of viruses derived f rom infect ious19
molecular clones of HIV-1LAI, HIV-1MAL, and HIV-1ELI. Virology 185:661-672.20
62. Ping, Y. H., C. Y. Chu, H. Cao, J. M. Jacque, M. Stevenson, and T. M. Rana.21
2004. Modulat ing HIV-1 replicat ion by RNA interference directed against human22
t ranscript ion elongat ion factor SPT5. Ret rovirology 1:46.23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 33
63. Purcell, D. F. , and M. A. Mart in. 1993. Al t ernat ive spl icing of human1
immunodef iciency virus t ype 1 mRNA modulat es viral prot ein expression,2
replicat ion, and infect ivity. J. Virol. 67:6365-6378.3
64. Pusch, O., D. Boden, R. Silbermann, F. Lee, L. Tucker, and B. Ramratnam.4
2003. Nucleot ide sequence homology requirements of HIV-1-specif ic short5
hairpin RNA. Nuc. Acids Res. 31:6444-6449.6
65. Qin, X. F. , D. S. An, I. S. Chen, and D. Baltimore. 2003. Inhibit ing HIV-17
infect ion in human T cells by lent iviral-mediated delivery of small interfering8
RNA against CCR5. Proc. Nat l. Acad. Sci. USA 100:183-188.9
66. Rabbitts, P. H. , A. Forster, M. A. Stinson, and T. H. Rabbitts. 1985.10
Truncat ion of exon 1 f rom the c-myc gene result s in prolonged c-myc mRNA11
stabilit y. EMBO J. 4:3727-3733.12
67. Rossi, J. J. 2005. Mammalian Dicer f inds a partner. EMBO Rep. 6:927-929.13
68. Ryan-Graham, M. A., and K. W. Peden. 1995. Both virus and host components14
are important for the manifestat ion of a Nef- phenotype in HIV-1 and HIV-2.15
Virology 213:158-168.16
69. Sambrook, J. , E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: a17
laboratory manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor,18
New York.19
70. Saumet, A., and C. H. Lecellier. 2006. Ant i-viral RNA silencing: do we look like20
plants? Ret rovirology 3:3.21
71. Schmid, I. , W. J. Krall, C. H. Uittenbogaart, J. Braun, and J. V. Giorgi. 1992.22
Dead cell discriminat ion with 7-amino-act inomycin D in combinat ion with dual23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 34
color immunof luorescence in single laser f low cytomet ry. Cytomet ry 13:204-1
208.2
72. Sledz, C. A., M. Holko, M. J. de Veer, R. H. Silverman, and B. R. Williams.3
2003. Act ivat ion of the interferon system by short -interfering RNAs. Nat . Cell.4
Biol. 5:834-839.5
73. Song, E., S. K. Lee, D. M. Dykxhoorn, C. Novina, D. Zhang, K. Crawford, J.6
Cerny, P. A. Sharp, J. Lieberman, N. Manjunath, and P. Shankar. 2003.7
Sustained small interfering RNA-mediated human immunodeficiency virus type 18
inhibit ion in primary macrophages. J. Virol. 77:7174-7181.9
74. Surabhi, R. M., and R. B. Gaynor. 2002. RNA interference directed against viral10
and cellular targets inhibit s human immunodeficiency Virus Type 1 replicat ion.11
J. Virol. 76:12963-12973.12
75. Tahbaz, N., F. A. Kolb, H. Zhang, K. Jaronczyk, W. Filipowicz, and T. C.13
Hobman. 2004. Characterizat ion of the interact ions between mammalian PAZ14
PIWI domain proteins and Dicer. EMBO Rep. 5:189-194.15
76. Theodore, T. S., G. Englund, A. Buckler-White, C. E. Buckler, M. A. Martin,16
and K. W. Peden. 1996. Const ruct ion and characterizat ion of a stable ful l-17
lengt h macrophage- t ropic HIV t ype 1 molecular clone t hat direct s t he18
product ion of high t it ers of progeny virions. AIDS Res. Hum. Ret roviruses.19
12:191-194.20
77. Westerhout, E. M., M. Ooms, M. Vink, A. T. Das, and B. Berkhout. 2005. HIV-121
can escape f rom RNA interference by evolving an alternat ive st ructure in it s22
RNA genome. Nuc. Acids Res. 33:796-804.23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 35
78. Yeung, M. L. , Y. Bennasser, S. Y. Le, and K. T. Jeang. 2005. siRNA, miRNA1
and HIV: promises and challenges. Cell. Res. 15:935-946.2
79. Yoshinari, K., M. Miyagishi, and K. Taira. 2004. Ef fects on RNAi of the t ight3
st ruct ure, sequence and posit ion of t he t arget ed region. Nuc. Acids Res.4
32:691-699.5
80. Zamore, P. D., and B. Haley. 2005. Ribo-gnome: the big world of small RNAs.6
Science 309:1519-1524.7
81. Zhong, J., A. H. Peters, K. Lee, and R. E. Braun. 1999. A double-st randed RNA8
binding protein required for act ivat ion of repressed messages in mammalian9
germ cells. Nat . Genet . 22:171-174.10
11
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 36
FIGURE LEGEND1
2
Figure 1: siRNAs against TRBP decrease exogenous TRBP expression. A) Schematic3
representation of TRBP2 mRNA and location of the siRNAs. One µg of each siRNA4
was run on a 2% agarose gel and visual ized by ethidium bromide staining. MW5
indicat es DNA molecular weight markers. B) siRNAs against TRBP reduce the6
fluorescence of EGFP-TRBP. HeLa cells were co-t ransfected with 0.5 µg pEGFP-C1-7
TRBP2 and with 100 nM of siRNA NS, 567, 571 and 657 as indicated using FuGENE. 48h8
post -t ransfect ion cel ls were f ixed, mount ed and assayed for GFP expression by9
f luorescence. C) Titration of EGFP-TRBP by FACS after co-transfection with siRNAs.10
HeLa cells were co-t ransfected with 0.5 µg pEGFP-C1-TRBP2 and with 0 (black bar), 311
(white bars), 7 (grey bars), 14 (st riped bars) or 18 (dot ted black bars) nM of siRNA NS,12
GFP, 567, 571 and 657 as indicated using Lipofectamine. Reporter expression is13
calculated as the percentage of EGFP-TRBP2 expression in the absence of siRNA. This14
result is the average of six independent experiments ± SEM.15
16
Figure 2. siRNAs against TRBP decrease endogenous protein expression. A)17
Determination of TRBP mRNA stability. Jurkat cells were incubated with ActD (518
µg/ ml) for 1, 2, 4, 6, 8, 10 hrs. 5 µg of t otal RNA were reverse t ranscribed and19
subj ected t o PCR amplif icat ion wit h specif ic primers for TRBP and c-myc. PCR20
products were quant if ied by densitomet ric scanning of t he gel (Typhoon scanner).21
TRBP (open squares) and c-myc (sol id circles) mRNA levels were expressed as22
percentage of init ial value and plot ted against t ime after ActD t reatment . The results23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 37
are t he means of t wo separat e experiment s. B) siRNAs against TRBP decrease1
endogenous TRBPs. HeLa cells were t ransfected with 0 (lane 4), 20 (lanes 1,5), 402
(lanes 2,6) or 80 (lanes 3,7) nM of siRNA-NS, siRNA567, siRNA571 or siRNA657 as3
indicated using FuGENE. Cells were t ransfected with 0.5 µg pCDNA3-TRBP1 (lane 8) or4
pCDNA3-TRBP2 (lane 9). 200 (lanes 1-7) or 20 (lanes 8,9) µg of cell ext ract were5
resolved by SDS-PAGE, analyzed by immunoblot t ing with an ant ibody against TRBP and6
exposed for 1hr or against act in and exposed for 1 min. TRBP1 start codon is included7
within TRBP2 reading frame. This is a representat ive experiment of three that gave8
similar result s. C) siRNAs do not activate PKR. HeLa cells were t ransfected with 09
(lane 1) or 80 nM of siRNA-NS (lane 2) or siRNA571 (lane 3) using FuGENE. 200 µg of10
cell ext ract were resolved by SDS-PAGE, analyzed by immunoblot t ing with ant ibodies11
against phosphorylated PKR (top panel), PKR (middle panel) and act in (lower panel)12
and exposed for 1 min. This is a representat ive experiment of three that gave similar13
results.14
15
Figure 3: siRNAs against TRBP decrease the expression of the HIV-1 LTR. A) siRNAs16
against TAR and TRBP reduce HIV-1 LTR basal expression. HeLa cells were mock17
t ransfected (lane 1) or cot ransfected with 0.05 µg of LTR-Luc, and 80 nM of t he18
indicated siRNAs (lanes 2-6) using FuGENE. B) siRNAs against Tat, TAR and TRBP19
reduce HIV-1 LTR trans-activated expression. HeLa cells were mock t ransfected20
(lane 1) or cot ransfected with 0.05 µg of LTR-Luc (lanes 2-8), 0.01 µg of pCMV1-Tat21
and 80 nM of the indicated siRNAs (lanes 3-8) using FuGENE. Luciferase act ivit y is the22
rat io between the luciferase level in the presence of the siRNA versus NS normalized23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 38
to 100% for cel ls t ransfected wit h NS. Each value represent s t he average of 51
independent experiments ± SEM. A representat ive quant itat ive PCR on the luciferase2
gene is shown at the bot tom of each graph as a t ransfect ion cont rol.3
4
Figure 4: siRNAs against TRBP decrease HIV-1 production in transfected HeLa cells.5
A) HIV-1 RT activity in cell supernatants. HeLa cel ls were mock t ransfected or6
cot ransfected with 0.5 µg of pNL4-3 and 14 nM of siRNAs NS, Tat1c, 567, 571 or 657 as7
indicated using Lipofectamine in T25 f lask format . RT act ivit y was calculated by8
densit omet ry using the sof tware referred in methods. Each value represents t he9
average of 4 independent experiments normalized as a percentage of the siRNA-NS RT10
value ± SEM. B) HIV Protein expression in cell lysates. HeLa cells were t ransfected11
as above. 120 µg of cel l lysat es were resolved by SDS-PAGE and analyzed by12
immunoblot t ing with an ant ibody against HIV-1 and exposed for 15 min. C) Cellular13
protein expression in cell lysates. 200 µg of the above cell lysates were resolved by14
SDS-PAGE and analyzed by immunoblot t ing with ant ibodies against P-PKR, PKR, TRBP15
and act in successively. The blots were exposed for 10 min for P-PKR, 1 min for PKR16
and TRBP, and 10 sec for act in. The blots shown in B and C are representat ive data17
among 4 independent experiments.18
19
Figure 5: siRNAs against TRBP decrease production of various HIV strains in20
transfected HeLa cells. HeLa cells were cot ransfected with 0.5 µg of pNL4-3, and 1 µg21
each of pAD8, p89.6, pELI-1, pMAL-2 or pROD-10 and 14 nM of siRNAs NS or TRBP 57122
as indicated using Lipofectamine in a 6 well format . RT act ivit y was calculated as in23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 39
Fig. 4. Each value represents the average of 3 independent experiments normalized1
as a percentage of the NS siRNA RT value ± SEM.2
3
Figure 6: siRNAs against Dicer decrease HIV-1 production in transfected HeLa cells.4
A) siRNAs against Dicer decrease mRNA and protein levels. HeLa cells were mock5
t ransfected or t ransfected with 14 nM of siRNAs NS or 4, 8 or 14 nM siRNA Dicer as6
indicated using Lipofectamine. (Top) Semi-quant it at ive RT-PCR with primers t hat7
amplify Dicer or GAPDH mRNA as indicated. (Bottom) 200 µg of cell lysates were8
resolved by SDS-PAGE and analyzed by immunoblot t ing with ant ibodies against Dicer9
or act in as indicated. The blots were exposed for 10 min for Dicer and 10 sec for10
act in. B) HIV-1 RT activity in cell supernatants. HeLa cells were mock t ransfected or11
cot ransfected with 0.5 µg of pNL4-3 and 14 nM of siRNAs NS, Tat1c or 4, 8 or 14 nM12
siRNA Dicer as indicated using Lipofectamine in a 6 well format . RT act ivit y was13
calculat ed as in Fig. 4 and correct ed for t ransfect ion ef f iciency. Each value14
represents the average of 4 independent experiments normalized as a percentage of15
the NS siRNA RT value ± SEM.16
17
Figure 7: Effects of siRNAs against TRBP or Dicer on cell viability and RNAi. A)18
siRNAs do not affect cell viability. HeLa cells were t ransfected with no siRNA (lane19
1), 1.8 µg poly(I) • poly(C) (lane 2) or 14 nM of either siRNA NS (lanes 3,4), siRNA EGFP20
(lane 5) siRNA-Tat (lane 6), siRNA567 (lane 7), siRNA571 (lane 8) siRNA657 (lane 9), or21
siRNA-Dicer (lane 10). All siRNAs were generated from the Ambion siRNA const ruct ion22
kit , except NS2 and siRNA-Dicer, which were purchased from Qiagen. Cell viabil it y23
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 40
was assessed by FACS analysis of 7-AAD stained cells. The viable cell populat ion of the1
mock t ransfected cel ls was set at 100%. Each value represents t he average of 42
independent experiments ± SEM. B) miRNA biogenesis is impaired in HeLa cells3
t ransfected with siRNA567, siRNA571 or siRNA-Dicer . (Top) Schemat ic4
representat ion of the vector delivered EGFP pre-miRNA and the predicted st ructure of5
t he fully processed EGFP miRNA. The EGFP sense sequence is highlighted. (Bot tom)6
HeLa cells were cot ransfected with 0 (lane 1) or 2 µg EGFP pre-miRNA vector (lanes 2-7
5) and 14 nM of siRNA NS (lane 2) siRNA567 (lane 3), siRNA571 (lane 4) or siRNA-Dicer8
(lane 5) in 6 well plates. Expression of EGFP miRNA was determined by Northern blot9
and quant i f ied by densit omet ry. The resul t s represent t he average of t hree10
independent experiments ± SEM.11
12
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

siRNAs against TRBP inhibit HIV 41
Table 1 : Sequence of the siRNAs used in this study.1
2
Target : Sequence: Reference:
Non silencing (NS) 5' AAUUCUCCGAACGUGUCACGU 3' Qiagen
EGFP 5’ AAGGGCAUCGACUUCAAGGAG 3’ This manuscript
Tat (SF2) 5’ AACUGCUUGUAACAAUUGCUA 3’ (16)
Tat1c (NL4-3) 5’ AAGAGCUCAUCAGAACAGUCA 3’ This manuscript
TAR 5’ AGACCAGAUCUGAGCCUGGUU 3' (41)
TRBP567 5’ AAAGAAUUCACCAUGACCUGU 3’ This manuscript
TRBP571 5’ AAUUCACCAUGACCUGUCGAG 3' This manuscript
TRBP657 5’ AAAUGCUGCUUCGAGUGCACA 3’ This manuscript
Dicer 5’ ACTGCTTGAAGCAGCTCTGGA 3’ Qiagen; (40)
3
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
siRNAs against TRBP inhibit HIV 42
Table 2 : HIV proviral plasmids targeted by siRNA571.1
2
Plasmid Clade Reference
pNL4-3 B (X4) (1)
pAD8 B (R5) (76)
p89.6 B (X4/R5) (18)
pELI-1 D (61)
pMAL2 A+D (61)
pROD-10 HIV-2 (68)
3
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
A
BsiRNA-NS siRNA567
siRNA571 siRNA657
1 567 6575' 3'
Coding sequence 3’ Untranslatedregion
5’ Untranslatedregion
571
Figure 1
C
siRNA: NS 567 571 657EGFP0
20
40
60
80
100
% E
GFP-
TRBP
exp
ress
ion
siRNA: NS 567 571 657
MW
50 -100 -
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

A 0 1 2 4 6 8 10 1 2 4 6 8 10Act. D EtOH
c-myc
TRBP
Time (h)
0
20
40
60
80
100
0 1 2 4 6 8 10Time (h) after Act. D treatment
c-mycTRBP
Perce
nt of
mRNA
rema
ining
C - NS 571P-PKR
Actin
PKR
siRNA
1 2 3
Figure 2
BsiRNA-NS TR
BP1TR
BP2siRNA567
TRBP2TRBP1
1 2 3 4 5 6 7 8 9
-
Actin
TRBP2TRBP1
siRNA571 siRNA657 TRBP1
TRBP2
1 2 3 4 5 6 7 8 9
-
Actin
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

A
0
20
40
60
80
100
Lucif
eras
e ac
tivity
(%)
- NS TAR 567 571 657siRNA:LTR-Luc: - + + + + +
1 2 3 4 5 6
Figure 3
B
0
20
406080
100
- NS Tat 567 571 657 TARsiRNA:
Lucif
eras
e ac
tivity
(%)
LTR-Luc:Tat:
- + + + + + +- - + + + + +
1 2 3 4 5 6 7 8++
NS
456 nt
456 nt
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
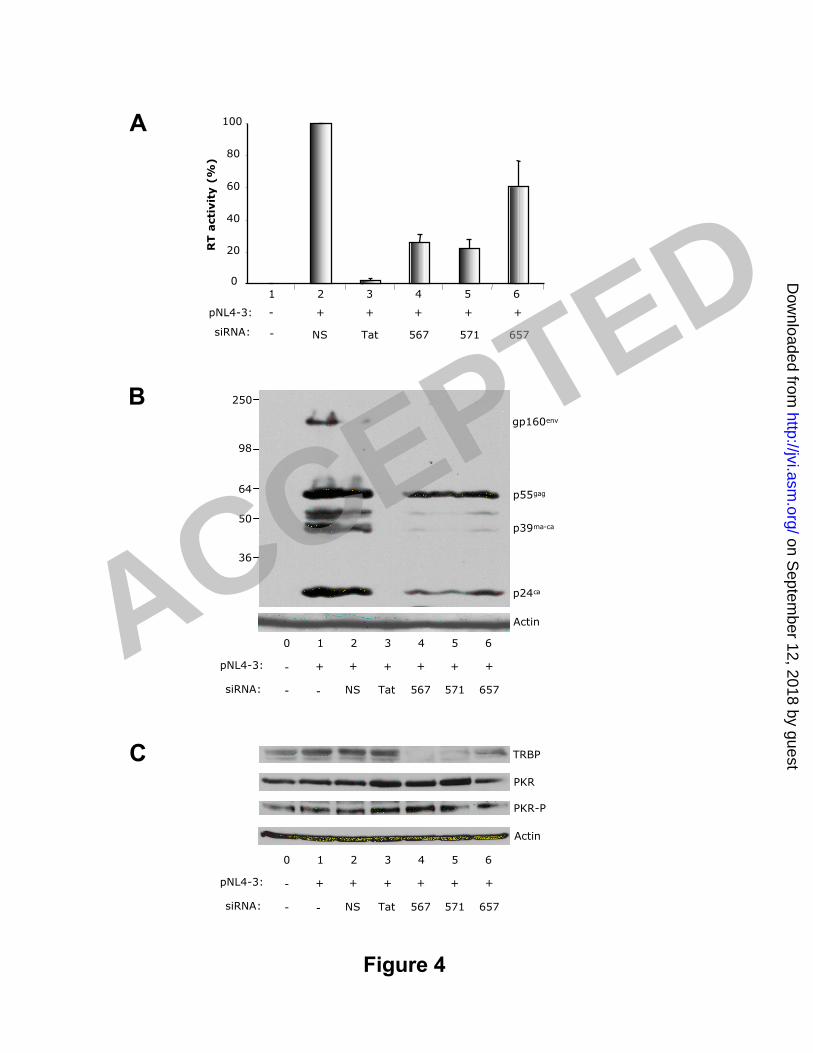
Figure 4
A
RT a
ctiv
ity (%
)
- NS Tat 567 571 657siRNA:pNL4-3: - + + + + +
1 2 3 4 5 6 0
20
40
60
80
100
siRNA:pNL4-3: - + + ++ +
-+- NS Tat 567 571 657
0 1 2 3 4 5 6
PKR
PKR-P
Actin
TRBPC
Bgp160env
p55gag
p39ma-ca
p24ca
250
98
64
50
36
Actin
siRNA:pNL4-3: - + + ++ +
-+- NS Tat 567 571 657
0 1 2 3 4 5 6 ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

Figure 5
0
20
40
60
80
100
pNL4-3 pAD8 p89.6 pELI-1 pMAL-2 pROD-10HIV-1 HIV-2
RT a
ctiv
ity (%
)
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
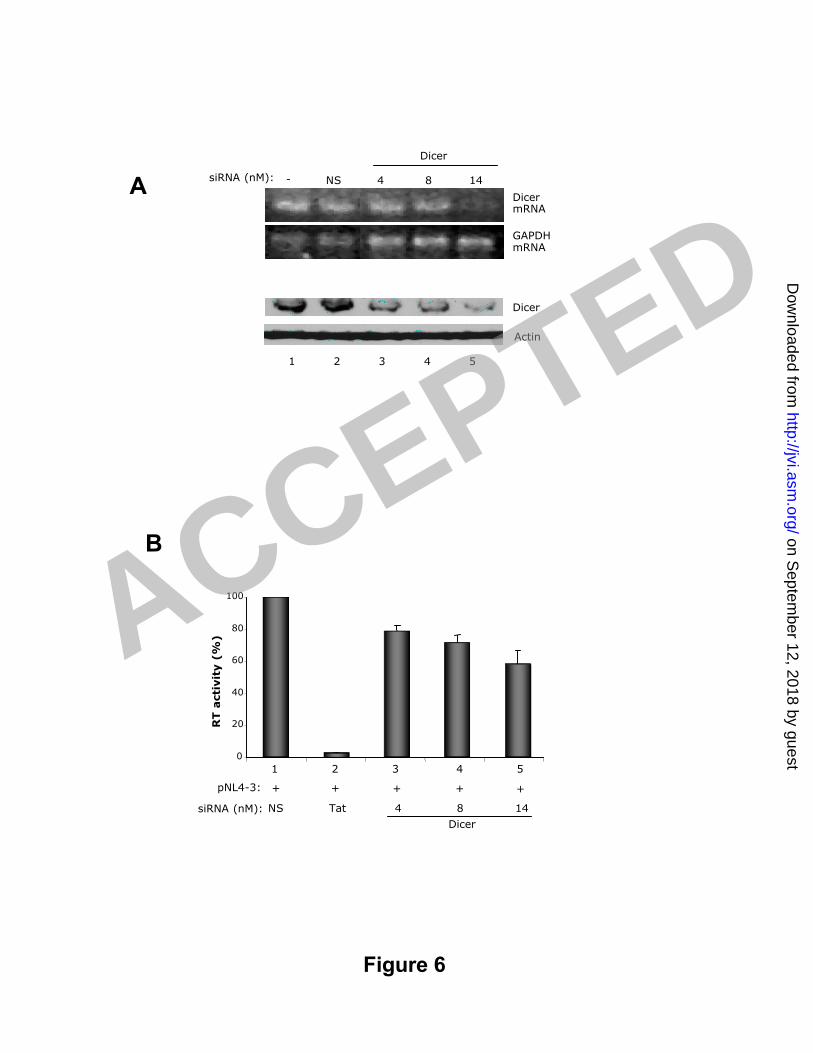
Figure 6
B
0
20
40
60
80
100
siRNA (nM):
RT a
ctiv
ity (%
)
NS Tat 4 8 14Dicer
pNL4-3: + + + + +1 2 3 4 5
A - NS 4 8 14siRNA (nM):Dicer
Actin
Dicer
1 2 3 4 5
DicermRNA
GAPDHmRNA
ACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

Figure 7
B
0
20
40
60
80
100
mock NS 567 571 DicersiRNA:
EGFP
miR
NA e
xpre
ssio
n (%
NS)
1 2 3 4 5
0 pI.pC NS NS2 EGFP Tat 567 571 657 DicerRNA:1 2 3 4 5 6 7 8 9 10
0
20
40
60
80
100
cell
viab
ility
(%)
siRNA
A
--| A ----- A GCG GGGCAUCGACUUCAAGGAGCU GUG A CGU CCCGUAGCUGAAGUUCCUCGG CAC GUU C GUAGA C
EGFPpre-miRNA
GGGCAUCGACUUCAAGGAGCUG UCCCCGUAGCUGAAGUUCCUCG
EGFPmiRNAACCEPTED
on Septem
ber 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from