1 cisplatin hypersensitivity of testicular germ cell - cancer research
TRANSCRIPT

1
Cisplatin hypersensitivity of testicular germ cell tumors is determined by high
constitutive Noxa levels mediated by Oct4
Matthias Gutekunst1, Thomas Mueller2, Andrea Weilbacher1, Michael A. Dengler1, Jens
Bedke3, Stephan Kruck3, Moshe Oren4, Walter E. Aulitzky5, Heiko van der Kuip1*
1 Dr. Margarete Fischer-Bosch Institute of Clinical Pharmacology and University of
Tuebingen, Stuttgart, Germany 2 Department of Internal Medicine IV, Oncology and Hematology, Martin-Luther-University
of Halle-Wittenberg, Halle, Germany 3 Department of Urology, Eberhard Karls University of Tuebingen, Tuebingen, Germany 4 Department of Molecular Cell Biology, Weizmann Institute of Science, Rehovot, Israel 5 2nd Department of Internal Medicine, Oncology and Hematology, Robert Bosch Hospital,
Stuttgart, Germany
*correspondence
Dr. Margarete Fischer-Bosch Institute of Clinical Pharmacology
Auerbachstr. 112
70376 Stuttgart
Germany
Telephone: #49-711-8101-3730; Fax: #49-711-859295
Email: [email protected]
Running title: Oct-4-dependent constitutive Noxa levels determine Cisplatin sensitivity
Key words: Oct-4, Noxa, Testicular Germ Cell Tumors (TGCT), Cisplatin hypersensitivity
Disclosure of Potential Conflicts of Interest: The authors declare no potential conflicts of
interest.
Grant Support: The research leading to these results has received support from the Robert
Bosch Foundation (project 11.5.8000.0094.0 and O2A to MG, AW, MAD, MO, WEA, HK)
and the Federal State of Saxony Anhalt (FKZ 3646A/0907 to TM).
Word counts: abstract: 250 manuscript text: 3912
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876

2
Abstract:
Testicular germ cell tumors (TGCT) are considered a paradigm of chemosensitive tumors.
Embryonal carcinoma (EC) cells represent the pluripotent entity of TGCT and are
characterized by expression of Oct-4, a key regulator of pluripotency and a determinant of
their inherent hypersensitivity to Cisplatin. However, the mechanisms underlying this Oct-4-
mediated sensitivity are poorly understood. We previously demonstrated that p53 is a major
player in Cisplatin hypersensitivity and therefore investigated whether Oct-4 may directly
affect p53 activity. Despite a significant decrease in sensitivity, depletion of Oct-4 did neither
alter Cisplatin-induced transactivation of p53 target genes nor its subcellular localization.
These data indicate that, rather than directly modulating p53 activity, Oct-4 provides a cellular
context that augments the proapoptotic activity of p53. Since mitochondrial priming by the
Bcl-2 family is a known determinant of chemosensitivity we compared the constitutive levels
of these proteins in Oct-4-positive and Oct-4-depleted cells. We identified Noxa as the only
Bcl-2 family protein to be highly correlated with Oct-4 status and Cisplatin sensitivity.
Compared to differentiated cells, constitutive Noxa levels were significantly higher in Oct-4-
positive cell lines and cancer patient samples. Furthermore, RNAi-mediated knockdown of
Oct-4 resulted in reduced Noxa transcript, in an almost complete loss of constitutive Noxa
protein and decreased Cisplatin hypersensitivity to a similar extent as did Noxa depletion.
In conclusion, our study indicates that Noxa is a central determinant of hypersensitivity to
Cisplatin. Oct-4-dependent high constitutive levels of this BH3-only protein prime EC cells to
undergo rapid and massive apoptosis in response to p53 activation.
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876

3
Introduction:
TGCT are the most frequent carcinomas in young male adults with a still increasing incidence
worldwide. Cisplatin-based chemotherapy cures the majority of patients even in advanced
stages due to the extraordinary sensitivity of TGCT to Cisplatin. Thus, TGCT are considered
as the paradigm of a chemosensitive tumor (1). Several mechanisms have been considered to
underlie this hypersensitivity, including the high apoptotic propensity of TGCT cells (2, 3).
The transcription factor Oct-4, a key regulator of pluripotency, is exclusively expressed in
cells of non-malignant pluripotent nature, i.e. embryonic stem cells (ESC) and germ cells, as
well as their malignant counterparts EC and seminoma, where it serves as a specific
malignancy marker (4, 5). Loss of Oct-4 causes a significant reduction of Cisplatin
hypersensitivity and was proposed to account for acquired Cisplatin resistance in refractory
tumors (6, 7). We and others have shown that Cisplatin hypersensitivity is highly dependent
on a functional p53 protein (8–10). The central role of this tumor suppressor was also
demonstrated by the finding that these cells are sensitive not only to Cisplatin but also to
DNA damage-independent p53 activators such as Nutlin-3 (10–12). Together, these results
indicate that hypersensitivity to p53 activation requires an Oct-4-mediated cellular context.
We now report that the low apoptotic threshold of Oct-4-positive TGCT cells is dictated by
the constitutive presence of high Noxa protein levels.
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876

4
Materials & Methods:
Cell culture: Cell lines from non-seminomatous germ cell tumors were analyzed: H12.1 (EC),
H12.5 (EC), 2102EP (EC), 1777NRpmet (differentiated state), 1411HP (differentiated state),
NTERA-2D1 (EC), H12.1ODM (in vitro differentiated derivative of H12.1). Origin and
cultivation procedures of these cell lines were described previously (6, 10). The cell lines
833K (EC), kindly provided by Prof. Andrews (University of Sheffield, UK), and GCT-72
(differentiated state), kindly provided by Prof. Pera (University of Southern California, USA),
were cultivated under same conditions. The cell line H12.1RA (in vitro differentiated
derivative of H12.1) was established by treatment of H12.1 with all-trans-retinoid acid during
cultivation in GCT-72-conditioned medium followed by cultivation under same condition as
the other cell lines (6, 10).
H460 (lung) and A2780 (ovary) cells were obtained from the NCI-60 cell line panel and
cultivated in RPMI-1640 with 10 % FCS and glutamine. PBMCs and Fibroblasts were
isolated and maintained as described previously (13, 14).
Reagents: Cisplatin and Nutlin-3 (Sigma) were used at a concentration of 10 µM, Etoposide
was used at 5 µM and Doxorubicin at 100 ng/ml. MG132 was used at a concentration of
1 µM.
Protein expression: Cells were lysed to obtain cellular protein according to standard
protocols. Western blot was performed using a SDS-PAGE Gel Electrophoresis system.
mRNA Expression: Total RNA was extracted and transcribed to cDNA according to standard
protocols. Expression was analyzed using BioMark HD System (Fluidigm) according to
manufacturer’s instructions. TaqMan assays were obtained from Applied Biosystems
(Supplementary Table 1). The relative amount of Noxa mRNA and pre-mRNA was
determined by a SYBR Green-based qPCR assay (7900HT Fast Real-Time PCR System,
Applied Biosystems). Contamination of RNA samples with genomic DNA was excluded by
DNase digestion followed by reverse transcription to cDNA. An intron-spanning primer pair
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876

5
was designed to detect Noxa mRNA levels whereas Noxa pre-mRNA was detected using
primers that span an exon-intron junction (for primer sequences see Supplementary Table 2).
Primers for detection of actin expression for normalization were kindly provided by Claudia
Kalla (Institute of Clinical Pharmacology, Stuttgart, Germany).
SRB microculture colorimetric assay: IC50 of Cisplatin and Nutlin-3 were determined as
described (15).
Subcellular fractionation: Cytoplasm and nuclei were separated using Mitochondria Isolation
Kit for Cultured Cells (Pierce) according to manufacturer’s instructions.
Apoptosis assay: Apoptosis was assessed by FITC-conjugated Annexin-V staining (16).
siRNA experiments: To silence Oct-4, Noxa, Puma and p53 we used siGenome SMARTpool
siRNA (Dharmacon, for sequences see Supplementary Table 3). Bim was silenced using Bim
siRNA from Santa Cruz. As a control we used siGenome Non-Targeting siRNA #1
(Dharmacon). Cells were transfected using DharmaFECT#3 (Dharmacon). 48 h after
transfection cells were treated according to requirements. To evaluate the efficacy of siRNA
silencing, protein lysates were prepared and analyzed by Western Blot.
Tissue samples: Tissue samples from TGCT patients were provided by the Department of
Urology, Eberhard Karls University of Tuebingen, Tuebingen, Germany. The local ethics
committee approved the collection of patient samples (315/2012BO2 and 396/2005V) and
informed consent was obtained from the patients. Specimen were frozen in liquid nitrogen
immediately after surgery and stored at -80°C for further use. To obtain protein lysates for
Western Blot, tissue was lysed using FastPrep-24 (MP Biomedicals) and sonication.
Statistics: Data are expressed as standard deviation of the mean (SD). Correlations are
expressed as Spearman rank correlation coefficient.
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876

6
Results:
Oct-4 depletion does not compromise the p53 response to Cisplatin
It has previously been demonstrated that Oct-4 protein levels are highly correlated with
Cisplatin hypersensitivity in TGCT cells (6). Therefore, we first tested whether Oct-4 is also
required for the more general sensitivity of TGCT to DNA damage-independent p53
activators (10). This was addressed by assessing the impact of Nutlin-3 on NTERA-2D1 cells
subjected to RNAi-mediated Oct-4 knockdown. As shown in Figure 1A, Oct-4 depletion
reduced Nutlin-3-induced apoptosis to a degree comparable to that observed in cells treated
with Cisplatin. These results suggest that efficient induction of apoptosis by p53 requires the
pluripotent, Oct-4-positive context of EC cells.
Consequently, we investigated possible differences in the accumulation and activation of p53
upon Cisplatin in the presence or absence of Oct-4. In response to cellular stress p53
activation leads to induction of a wide variety of target genes. Hence, we screened a panel of
46 bona fide p53 transcriptional targets ((17, 18); Supplementary Table 1) by qPCR in Oct-4-
positive and Oct-4-deprived NTERA-2D1 cells exposed to Cisplatin. A similar analysis was
also performed with the EC cell line 2102EP that retains a nullipotent phenotype, allowing us
to discriminate between differentiation-dependent and direct effects of Oct-4. Surprisingly,
despite its pronounced impact on apoptosis, Oct-4 depletion did not compromise the p53-
dependent transcriptional response to Cisplatin in both cell lines (Figure 1B). Hence,
induction of a p53-mediated transcriptional program is not the primary determinant of Oct-4-
mediated hypersensitivity.
Beyond its function as a transcription factor, p53 is capable of facilitating apoptosis in the
cytoplasm. Thus, we monitored the accumulation of p53 upon Cisplatin exposure in both
nucleus and cytoplasm. Notably, upon Cisplatin treatment, p53 accumulated to a similar
extent in both compartments (Figure 1C). Nevertheless, neither p53 accumulation nor its
localization was significantly influenced by knockdown of Oct-4 (Figure 1C), which further
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876

7
argues against a direct influence of Oct-4 on p53 activity. Together, these results indicate that
rather than directly altering p53 activity, Oct-4 provides an apoptosis-prone cellular context
that augments the proapoptotic efficacy of p53.
Oct-4 determines Cisplatin sensitivity by modulating the Bcl-2 family composition
Recently, it was demonstrated that the response to cytotoxic chemotherapy is dependent on
the relative composition of Bcl-2 family proteins which determines the apoptotic threshold
(19). We therefore investigated a possible correlation of individual Bcl-2 family proteins with
Oct-4 and Cisplatin sensitivity in NTERA-2D1 and 2102EP cells. To that end, we silenced
Oct-4 by RNAi-mediated knockdown and examined the protein levels of the proapoptotic
Bcl-2 family members Bax, Bak, Noxa, Puma, Bim and Bid, as well as the anti-apoptotic
members Bcl-2, Bcl-xL, Bcl-w and Mcl-1. Notably, Noxa was the only Bcl-2 family protein
which could be detected selectively in the presence of Oct-4 (Figure 2A). The fact that this
was observed in both pluripotent NTERA-2D1 and nullipotent 2102EP cells (Figure 2A)
suggests that the reduction of Noxa protein is not just a secondary effect of differentiation but
a direct consequence of Oct-4 depletion. Mcl-1, the major anti-apoptotic binding partner of
Noxa, was also reduced upon Oct-4 knockdown (Figure 2A). Since Noxa levels became
practically undetectable under such conditions (Figure 2A), the Noxa/Mcl-1 ratio was shifted
considerably in favor of Mcl-1, probably establishing a higher apoptotic threshold.
Knockdown of Oct-4 also reduced the levels of the BH3-only proteins Puma and Bim,
especially in 2102EP cells, although not as strongly as observed for Noxa (Figure 2A).
Notably, a role of Puma in hypersensitivity of pluripotent EC cells was previously proposed
(10). The protein levels of all other Bcl-2 family members analyzed did not exhibit
concomitant changes in these cell lines upon Oct-4 depletion (Figure 2A).
Next, we investigated the impact of the Oct-4-dependent Bcl-2 family proteins Noxa, Puma
and Bim on Cisplatin hypersensitivity. Therefore, we transfected NTERA-2D1 and 2102EP
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876

8
cells with the corresponding siRNAs and obtained a similar knockdown efficiency for each
siRNA (Figure 2B, right panel). Silencing of Noxa led to a marked decrease in sensitivity to
Cisplatin in both cell lines, comparable to that observed upon Oct-4 knockdown (Figure 2B,
left panel). A similar trend was also seen with Puma, although in NTERA-2D1 cells, silencing
of Puma did not reduce apoptosis as efficiently as did Noxa knockdown. In contrast, Bim
knockdown had only minor effects on Cisplatin hypersensitivity (Figure 2B, left panel). To
investigate possible cross-regulation between these pro-apoptotic Bcl-2 family members,
NTERA-2D1 cells were depleted of each protein and levels of the respective others were
quantified. We found no significant cross-regulation between Noxa, Puma and Bim
(Supplementary Figure 1).
It is of note that knockdown of either Oct-4 or Noxa reduced sensitivity of TGCT not only to
Cisplatin but also to other genotoxic agents such as Etoposide and Doxorubicin as well as to
the DNA damage-independent p53 activator Nutlin-3 (Supplementary Figure 2). These data
suggest that Oct-4 and Noxa are central mediators of the general hypersensitivity of TGCT.
Constitutive Noxa protein levels are closely correlated with Oct-4 and Cisplatin
hypersensitivity in a panel of TGCT cell lines and patient samples
Noxa and Puma are induced in response to several cellular stress stimuli. However, Noxa
protein was also found to be highly expressed in unstressed, undifferentiated EC cells but not
in cell lines derived from other tumor entities (10). We therefore investigated if constitutive
Noxa and Puma levels are maintained via transcriptional mechanisms in EC cells. In contrast
to Puma, Noxa transcript was significantly higher expressed in Oct-4-positive NTERA-2D1
and 2102EP cells when compared to Oct-4-negative lymphocytes, fibroblasts or cancer cell
lines (Figure 3A, left panels). A direct effect of Oct-4 on Noxa mRNA expression was
confirmed by Oct-4 knockdown experiments. Oct-4 depletion reduced Noxa transcript in
NTERA-2D1 and 2102EP cells to a level comparable to that observed in differentiated cells
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876

9
(Figure 3A, left panels). To investigate the possibility that Oct-4 may regulate mRNA stability
by post-transcriptional mechanisms, expression of Noxa pre-mRNA was examined and found
to be similarly dependent on Oct-4 as was mature Noxa mRNA (Supplementary Figure 3).
This result indicates that Oct-4 regulates Noxa gene transcription. Puma, on the other hand,
was only slightly reduced upon Oct-4 depletion (Figure 3A, left panels). Together, these data
suggest an Oct-4-dependent transcriptional mechanism resulting in high constitutive Noxa
protein levels.
The effect of Oct-4 on constitutive expression of Noxa in Oct-4-positive and Oct-4-depleted
EC cells was even more pronounced at the protein level (Figure 3A, right panel).
Furthermore, the absolute levels of Noxa protein in Oct-4-positive cells were 9.2fold
(NTERA-2D1 vs. PBMC) and 11.2fold (2102EP vs. PBMC) higher as compared to those of
stimulated hematopoietic cells (Figure 3A, right panel) which have been shown to exhibit
constitutive Noxa protein (20), whereas the difference in transcript levels was only 2.3fold
(NTERA-2D1 vs. PBMC) and 1.3fold (2102EP vs. PBMC) (Figure 3A, left panel). It is
therefore likely that Oct-4 also modulates Noxa protein translation and/or stability by a yet
unknown mechanism. In contrast to Noxa, Puma levels were found to be higher only in one of
the two Oct-4-positive cell lines when compared to differentiated cells (Figure 3A, right
panel).
To further investigate the correlation of Oct-4 and Cisplatin sensitivity with Noxa and Puma,
we analyzed constitutive protein levels of both BH3-only proteins (Figure 3B, upper left
panel) together with Cisplatin IC50 values (Figure 3B, lower left panel) in an extended TGCT
cell line panel consisting of 5 Oct-4-negative and 5 Oct-4-positive cell lines. We confirmed a
correlation of Cisplatin sensitivity with Oct-4 (Spearman r=-0.842, p=0.004; Figure 3B, lower
right panels). Importantly, Noxa was significantly correlated with Oct-4 expression
(Spearman r=0.891, p=0.001; Figure 3B, upper right panels) as well as with Cisplatin
sensitivity (Spearman r=-0.806, p=0.007; Figure 3B, lower right panels). On the other hand,
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876

10
while Puma protein levels were also positively correlated with Oct-4 status, this correlation
was weaker than for Noxa (Spearman r=0.685, p=0.035; Figure 3B, upper right panel), and no
significant correlation was seen with Cisplatin sensitivity (Spearman r=-0.527, p>0.05; Figure
3B, lower right panel). Notably, similar results were observed with Nutlin-3 (Supplementary
Figure 4). These data further support the conjecture that Oct-4 lowers the apoptotic threshold
in TGCT primarily by maintaining high constitutive Noxa protein levels.
In addition, we sought to extend our in vitro findings to cancer patient samples. Indeed, we
could confirm a close correlation between Oct-4 and Noxa protein levels in primary tissue
samples derived from 5 ECs and 8 seminomas (Spearman r=0.832; p<0.0001; Figure 3C).
Accumulation of both Noxa and p53 is required to induce cell death in Oct-4-depleted
TGCT cells
We have previously shown that accumulation of p53 is a central determinant in TGCT
hypersensitivity (10). In the present study, we established that Oct-4-positive TGCT cells are
characterized by high constitutive Noxa protein levels, which seems to be a requirement for
their p53-mediated hypersensitivity. Therefore, in Oct-4-depleted cells, pharmacological
induction of both Noxa and p53 protein should result in cell death. In order to test this
hypothesis, NTERA-2D1 and 2102EP cells, respectively, were depleted of Oct-4 by RNAi-
mediated gene silencing (Figure 4, upper panels). In contrast to Cisplatin or Nutlin-3 (Figure
1A), treatment with the proteasome inhibitor MG132 resulted in efficient induction of
apoptosis in both Oct-4-depleted cell lines (Figure 4, lower panels). Importantly, MG132
treatment induced simultaneous accumulation of Noxa and p53 protein (Figure 4, middle
panels). Prevention of accumulation of either protein by RNAi-mediated knockdown led to
increased survival upon MG132 treatment in NTERA-2D1 and 2102EP cells (Figure 4, lower
panels). These data further demonstrate that both Noxa and p53 protein are required to induce
apoptosis in TGCT cells.
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876

11
Discussion:
Cisplatin-based chemotherapy has been a successful cure for TGCT for decades. A variety of
molecular determinants of these neoplasms were proposed to account for their exquisite
sensitivity to Cisplatin such as a reduced DNA repair capacity, a deregulated G1/S transition
or marginal p21 expression (2; 3; 21). Much of the research done in this field has focused on
p53 which is almost exclusively expressed in its wild type conformation in TGCT (22). We
recently demonstrated a central role for p53 in TGCT hypersensitivity to Cisplatin and
suggested an intrinsic sensitivity of these tumors to activation of p53 which leads to robust
induction of apoptosis in this specific cellular context (10). In the present study we found that
this sensitivity is dependent on the presence of Oct-4, although p53 activation is not directly
influenced by this mediator of pluripotency. Rather, by maintaining high constitutive Noxa
protein levels, Oct-4 provides a cellular context that primes TGCT cells to p53-dependent
apoptosis.
TGCT derive from intrinsically apoptosis-prone tissue. Therefore, their unique responsiveness
to chemotherapy might be due to inherent biological characteristics (23). An intrinsic
molecular determinant of the pluripotent compartment of TGCT is the expression of
regulators of pluripotency such as Oct-4. Recent work related loss of Oct-4 to an increase in
Cisplatin resistance (6). In the present study we show that Oct-4 depletion not only protected
cells from Cisplatin-induced cell death but also led to a reduction of the apoptotic response to
other genotoxic and non-genotoxic p53 inducers (Figure 1A and Supplementary Figure 2).
These data imply that Oct-4 mediates the general intrinsic hypersensitivity to p53 activators in
pluripotent EC cells. Several studies have attributed the increased susceptibility to apoptosis
to overexpression of p53 (24; 25). However, we and others could show that high p53 protein
levels are not a characteristic of TGCT (10; 26). Here we demonstrate that Oct-4 had no
impact on constitutive p53 levels or on its accumulation upon Cisplatin treatment (Figure 1C).
Furthermore, Oct-4 depletion did not influence the Cisplatin-induced transactivation of 46
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876

12
representative bona fide p53 target genes (Figure 1B). These data indicate that the
transcriptional p53 response is not substantially altered in Oct-4-positive hypersensitive
TGCT cells. In addition to its capability to launch the apoptotic program through
transactivation of proapoptotic target genes, p53 can directly induce apoptosis in the
cytoplasm by interacting with Bcl-2 family proteins. Subcellular fractionation revealed that
besides an accumulation of p53 in the nucleus, Cisplatin treatment resulted in a similar
accumulation in the cytoplasm (Figure 1C), suggesting that p53 could function independent of
its transcriptional activity to launch the apoptotic program in TGCT; this was also proposed as
a proapoptotic mechanism in ESC (27). However, Oct-4 depletion did not significantly
influence subcellular distribution of p53 (Figure 1C). Together, these data demonstrate that
neither transcriptional nor cytoplasmic functions of p53 are dependent on the presence of Oct-
4 in TGCT cells. Since Oct-4 is central for Cisplatin sensitivity, it must provide a cellular
context allowing an effective proapoptotic p53 response.
Recently, Letai and co-workers established a model to explain and predict chemotherapeutic
response by mitochondrial priming (19). According to this hypothesis, occupation of anti-
apoptotic Bcl-2 family proteins by their proapoptotic counterparts determines the proximity of
a cell to the threshold of apoptosis (28). Our data demonstrate that the presence of Oct-4 in
pluripotent EC cells dictates a Bcl-2 family profile dominated by high constitutive Noxa
protein levels which augments the cell’s commitment to apoptosis. We found that Noxa
protein levels are tightly correlated with both Oct-4 status and Cisplatin sensitivity in a panel
of cell lines and also in samples from primary TGCT.
The relative levels of other Bcl-2 proteins have also previously been proposed to determine
hypersensitivity in TGCT. Several studies suggested that the enhanced sensitivity to
genotoxic agents is dependent on a high Bax/Bcl-2 ratio (29) and low levels of Bcl-2 and Bcl-
xL (30; 31). A chemoprotective role of Bcl-xL in these tumors has also been proposed (32).
However, we found no correlation between the protein levels of Bcl-2, Bax and Bak and Oct-
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876

13
4 status (Figure 2A). Indeed, recent data demonstrated that Bcl-2, Bcl-xL, Bax or Bak levels
were similar among TGCT cell lines with differential Cisplatin sensitivity (15; 33). Together,
these data indicate that Noxa is the only Bcl-2 family protein that is tightly correlated to both
Oct-4 status and Cisplatin sensitivity, suggesting that constitutive Noxa protein is a key
determinant of the exclusive sensitivity of Oct-4 positive EC cells to Cisplatin. Moreover, we
show that the general sensitivity of TGCT to genotoxic and non-genotoxic agents is also
dependent on Noxa (Supplementary Figure 2). The prominent role of Noxa for TGCT
hypersensitivity is further supported by the finding that Noxa protein was found to be
correlated with good clinical prognosis in patients with EC (34).
Noxa was shown to preferentially bind to Mcl-1 and A1 (35). The apoptotic potential of this
BH3-only protein when overexpressed was therefore suggested to be quite variable due to its
inability to antagonize Bcl-2 and Bcl-xL (36). However, as mentioned above, TGCT exhibit
low expression of Bcl-2 and Bcl-xL (30; 31), potentially providing an ideal premise for an
increased apoptotic potential of Noxa overexpression. Moreover, our results demonstrate a
considerable shift of the Noxa/Mcl-1 ratio in favor of Mcl-1 upon Oct-4 depletion (Figure
2A), enforcing a higher apoptotic threshold. A similar dependence on the Noxa/Mcl-1 balance
was shown for HeLa and pancreatic cancer cells treated with camptothecin (37; 38). It is of
note that Noxa was recently shown to be capable of binding Bcl-xL upon DNA damage and
thus facilitate apoptosis induction (39), a conceivable mechanism that broadens Noxa’s
proapoptotic capacity. In addition, it seems most likely that while Noxa inhibits the anti-
apoptotic functions of Mcl-1, a robust induction of other BH3-only proteins by p53 sequesters
Bcl-xL and activates Bax and Bak. This hypothesis is supported by the fact that Puma was
found to be as important as was Noxa for the hypersensitivity to Cisplatin (Figure 2B).
However, constitutive Puma protein levels could not to be linked to Cisplatin sensitivity
(Figure 3B). These results indicate that Puma exerts its prominent proapoptotic function in
TGCT cells only upon induction by p53 as previously described for other cell types (40).
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876

14
Due to the established function of Oct-4 as a transcription factor it is tempting to speculate
that transcription of the Noxa gene is directly promoted by Oct-4. Interestingly, in a recent
study, an Oct-4 binding site was mapped within 8 kb of the Noxa promoter (41). Moreover,
Noxa mRNA was shown to be upregulated in ESCs (42) and its expression was found to be
correlated with Oct-4 in this cell type (43; 44). The finding that Noxa pre-mRNA was
downregulated upon Oct-4 depletion to a similar extent as the corresponding mature mRNA
(Supplementary Figure 3) suggests that Oct-4 may indeed directly regulate the transcription of
the Noxa gene. However, since Oct-4 is involved in the transactivation of many genes,
including other transcription factors (44), it remains possible that this regulation might as well
include indirect effects.
Noxa was commonly seen as a transcriptionally activated stress-responsive gene (36) rather
than a protein present at high levels in unstressed cells. However, upregulated Noxa protein
levels were reported to restrain lymphocyte expansion and can be triggered by glucose
deprivation (20). Together with the high constitutive protein levels reported in the present
work (Figure 3A), these data suggest an additional regulation at the protein level, e.g. through
the ubiquitin-proteasome system (45; 46). Since Noxa protein regulation by Oct-4 was
demonstrated to be more pronounced when compared to regulation at the mRNA level (Figure
3A), altered Noxa protein stability cannot be excluded as a possible additional explanation for
high constitutive Noxa protein levels.
We established that loss of high constitutive Noxa protein levels in Oct-4-depleted cells
causes resistance to Cisplatin and Nutlin-3. However, these cells were still sensitive to
proteasomal inhibition by MG132 (Figure 4). Importantly, MG132 resulted in stabilization of
Noxa protein reaching levels comparable to that observed in Oct-4-positive cells.
Concomitantly, p53 was stabilized upon proteasomal inhibition. RNAi-mediated knockdown
of Noxa or p53, respectively, rescued Oct-4-depleted cells from cell death upon MG132
treatment (Figure 4). These data further indicate that hypersensitivity of TGCT is dependent
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876

15
on both high levels of Noxa and induction of p53.
We propose that, under basal non-stressed conditions, EC cells may buffer the high apoptotic
pressure driven by high constitutive Noxa levels by a parallel increase in its anti-apoptotic
binding partner Mcl-1. Upon cellular stress, p53-mediated induction of additional BH3-only
proteins such as PUMA (10) and concomitant Mcl-1 degradation (47) may then shift the
balance and lead to a rapid and robust induction of apoptosis.
In conclusion, our present study demonstrates that Oct-4-dependent high constitutive Noxa
levels determine Cisplatin hypersensitivity in TGCT. Despite the predominant role of p53 in
TGCT hypersensitivity, Oct-4 does not directly influence p53 activation but rather provides
an extremely low apoptotic threshold by maintaining high constitutive Noxa protein levels.
Therefore, in response to cellular stress, accumulation of p53 triggers a rapid and massive
induction of apoptosis.
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876

16
Acknowledgements:
We are grateful to Prof. Hans-Joachim Schmoll (Martin-Luther-University of Halle-
Wittenberg, Germany) for the initiation of the collaboration between Stuttgart and Halle as
well as Tabea Lieberich (Stuttgart) and Franziska Reipsch (Halle) for technical assistance.
Grant Support:
This work was financially supported by the Robert Bosch Foundation (project
11.5.8000.0094.0 and O2A) and the Federal State of Saxony Anhalt (FKZ 3646A/0907).
References:
1. Di Pietro A, Vries EGE de, Gietema JA, Spierings DCJ, De Jong S. Testicular germ cell
tumours: the paradigm of chemo-sensitive solid tumours. Int. J. Biochem. Cell Biol.
2005;37(12):2437–2456.
2. Masters JRW, Köberle B. Curing metastatic cancer: lessons from testicular germ-cell
tumours. Nat. Rev. Cancer. 2003;3(7):517–525.
3. Spierings DCJ, De Vries EGE, Vellenga E, De Jong S. The attractive Achilles heel of
germ cell tumours: an inherent sensitivity to apoptosis-inducing stimuli. J. Pathol.
2003;200(2):137–148.
4. De Jong J, Looijenga LHJ. Stem cell marker OCT3/4 in tumor biology and germ cell
tumor diagnostics: history and future. Crit Rev Oncog. 2006;12(3-4):171–203.
5. Mueller T, Luetzkendorf J, Nerger K, Schmoll H-J, Mueller LP. Analysis of OCT4
expression in an extended panel of human tumor cell lines from multiple entities and in
human mesenchymal stem cells. Cell. Mol. Life Sci. 2009;66(3):495–503.
6. Mueller T, Mueller LP, Luetzkendorf J, Voigt W, Simon H, Schmoll H-J. Loss of Oct-3/4
expression in embryonal carcinoma cells is associated with induction of cisplatin
resistance. Tumour Biol. 2006;27(2):71–83.
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876

17
7. Koster R, Di Pietro A, Timmer-Bosscha H, Gibcus JH, Van den Berg A, Suurmeijer AJ,
et al. Cytoplasmic p21 expression levels determine cisplatin resistance in human
testicular cancer. J. Clin. Invest. 2010;120(10):3594–3605.
8. Lutzker SG, Mathew R, Taller DR. A p53 dose-response relationship for sensitivity to
DNA damage in isogenic teratocarcinoma cells. Oncogene. 2001;20(23):2982–2986.
9. Kerley-Hamilton JS, Pike AM, Li N, DiRenzo J, Spinella MJ. A p53-dominant
transcriptional response to cisplatin in testicular germ cell tumor-derived human
embryonal carcinoma. Oncogene. 2005;24(40):6090–6100.
10. Gutekunst M, Oren M, Weilbacher A, Dengler MA, Markwardt C, Thomale J, et al. p53
hypersensitivity is the predominant mechanism of the unique responsiveness of testicular
germ cell tumor (TGCT) cells to cisplatin. PLoS ONE. 2011;6(4):e19198.
11. Li B, Cheng Q, Li Z, Chen J. p53 inactivation by MDM2 and MDMX negative feedback
loops in testicular germ cell tumors. Cell Cycle. 2010;9(7):1411–1420.
12. Bauer S, Mühlenberg T, Leahy M, Hoiczyk M, Gauler T, Schuler M, et al. Therapeutic
potential of Mdm2 inhibition in malignant germ cell tumours. Eur. Urol. 2010;57(4):679–
687.
13. Fanta S, Sonnenberg M, Skorta I, Duyster J, Miething C, Aulitzky WE, et al.
Pharmacological inhibition of c-Abl compromises genetic stability and DNA repair in
Bcr-Abl-negative cells. Oncogene. 2008;27(31):4380–4384.
14. Haubeiss S, Schmid JO, Mürdter TE, Sonnenberg M, Friedel G, Van der Kuip H, et al.
Dasatinib reverses cancer-associated fibroblasts (CAFs) from primary lung carcinomas to
a phenotype comparable to that of normal fibroblasts. Mol. Cancer. 2010;9168.
15. Mueller T, Voigt W, Simon H, Fruehauf A, Bulankin A, Grothey A, et al. Failure of
activation of caspase-9 induces a higher threshold for apoptosis and cisplatin resistance in
testicular cancer. Cancer Res. 2003;63(2):513–521.
16. Skorta I, Oren M, Markwardt C, Gutekunst M, Aulitzky WE, Van der Kuip H. Imatinib
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876

18
mesylate induces cisplatin hypersensitivity in Bcr-Abl+ cells by differential modulation
of p53 transcriptional and proapoptotic activity. Cancer Res. 2009;69(24):9337–9345.
17. Wei C-L, Wu Q, Vega VB, Chiu KP, Ng P, Zhang T, et al. A global map of p53
transcription-factor binding sites in the human genome. Cell. 2006;124(1):207–219.
18. Riley T, Sontag E, Chen P, Levine A. Transcriptional control of human p53-regulated
genes. Nat. Rev. Mol. Cell Biol. 2008;9(5):402–412.
19. Ni Chonghaile T, Sarosiek KA, Vo T-T, Ryan JA, Tammareddi A, Moore VDG, et al.
Pretreatment mitochondrial priming correlates with clinical response to cytotoxic
chemotherapy. Science. 2011;334(6059):1129–1133.
20. Alves NL, Derks IAM, Berk E, Spijker R, Van Lier RAW, Eldering E. The Noxa/Mcl-1
axis regulates susceptibility to apoptosis under glucose limitation in dividing T cells.
Immunity. 2006;24(6):703–716.
21. Bartkova J, Lukas C, Sørensen CS, Meyts ER-D, Skakkebaek NE, Lukas J, et al.
Deregulation of the RB pathway in human testicular germ cell tumours. J. Pathol.
2003;200(2):149–156.
22. Peng HQ, Hogg D, Malkin D, Bailey D, Gallie BL, Bulbul M, et al. Mutations of the p53
gene do not occur in testis cancer. Cancer Res. 1993;53(15):3574–3578.
23. Blagosklonny MV. NCI’s provocative questions on cancer: some answers to ignite
discussion. Oncotarget. 2011;2(12):1352–1367.
24. Riou G, Barrois M, Prost S, Terrier MJ, Theodore C, Levine AJ. The p53 and mdm-2
genes in human testicular germ-cell tumors. Mol. Carcinog. 1995;12(3):124–131.
25. Guillou L, Estreicher A, Chaubert P, Hurlimann J, Kurt AM, Metthez G, et al. Germ cell
tumors of the testis overexpress wild-type p53. Am. J. Pathol. 1996;149(4):1221–1228.
26. Kersemaekers A-MF, Mayer F, Molier M, Van Weeren PC, Oosterhuis JW, Bokemeyer
C, et al. Role of P53 and MDM2 in treatment response of human germ cell tumors. J.
Clin. Oncol. 2002;20(6):1551–1561.
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876

19
27. Qin H, Yu T, Qing T, Liu Y, Zhao Y, Cai J, et al. Regulation of apoptosis and
differentiation by p53 in human embryonic stem cells. J. Biol. Chem. 2007;282(8):5842–
5852.
28. Davids MS, Letai A. Targeting the B-Cell Lymphoma/Leukemia 2 Family in Cancer.
Journal of Clinical Oncology: J Clin Oncol. 2012 Sep 1;30(25):3127-35.
29. Chresta CM, Masters JR, Hickman JA. Hypersensitivity of human testicular tumors to
etoposide-induced apoptosis is associated with functional p53 and a high Bax:Bcl-2 ratio.
Cancer Res. 1996;56(8):1834–1841.
30. Mayer F, Stoop H, Scheffer GL, Scheper R, Oosterhuis JW, Looijenga LHJ, et al.
Molecular determinants of treatment response in human germ cell tumors. Clin. Cancer
Res. 2003;9(2):767–773.
31. Soini Y, Paakko P. Extent of apoptosis in relation to p53 and bcl-2 expression in germ
cell tumors. Hum. Pathol. 1996;27(11):1221–1226.
32. Arriola EL, Rodriguez-Lopez AM, Hickman JA, Chresta CM. Bcl-2 overexpression
results in reciprocal downregulation of Bcl-X(L) and sensitizes human testicular germ
cell tumours to chemotherapy-induced apoptosis. Oncogene. 1999;18(7):1457–1464.
33. Burger H, Nooter K, Boersma AW, Kortland CJ, Stoter G. Lack of correlation between
cisplatin-induced apoptosis, p53 status and expression of Bcl-2 family proteins in
testicular germ cell tumour cell lines. Int. J. Cancer. 1997;73(4):592–599.
34. Grande L, Bretones G, Rosa-Garrido M, Garrido-Martin EM, Hernandez T, Fraile S, et
al. Transcription factors sp1 and p73 control the expression of the proapoptotic protein
noxa in the response of testicular embryonal carcinoma cells to cisplatin. J Biol Chem.
2012 Aug 3;287(32):26495-505.
35. Chen L, Willis SN, Wei A, Smith BJ, Fletcher JI, Hinds MG, et al. Differential targeting
of prosurvival Bcl-2 proteins by their BH3-only ligands allows complementary apoptotic
function. Mol. Cell. 2005;17(3):393–403.
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876

20
36. Ploner C, Kofler R, Villunger A. Noxa: at the tip of the balance between life and death.
Oncogene. 2008;27 Suppl 1S84–92.
37. Mei Y, Xie C, Xie W, Tian X, Li M, Wu M. Noxa/Mcl-1 balance regulates susceptibility
of cells to camptothecin-induced apoptosis. Neoplasia. 2007;9(10):871–881.
38. Naumann K, Schmich K, Jaeger C, Kratz F, Merfort I. Noxa/Mcl-1 balance influences the
effect of the proteasome inhibitor MG-132 in combination with anticancer agents in
pancreatic cancer cell lines. Anticancer Drugs. 2012 Jul;23(6):614-26.
39. Lopez H, Zhang L, George NM, Liu X, Pang X, Evans JJD, et al. Perturbation of the Bcl-
2 network and an induced Noxa/Bcl-xL interaction trigger mitochondrial dysfunction
after DNA damage. J. Biol. Chem. 2010;285(20):15016–15026.
40. Yu J, Yue W, Wu B, Zhang L. PUMA sensitizes lung cancer cells to chemotherapeutic
agents and irradiation. Clin. Cancer Res. 2006;12(9):2928–2936.
41. Boyer LA, Lee TI, Cole MF, Johnstone SE, Levine SS, Zucker JP, et al. Core
transcriptional regulatory circuitry in human embryonic stem cells. Cell.
2005;122(6):947–956.
42. Madden DT, Davila-Kruger D, Melov S, Bredesen DE. Human embryonic stem cells
express elevated levels of multiple pro-apoptotic BCL-2 family members. PLoS ONE.
2011;6(12):e28530.
43. Cai J, Chen J, Liu Y, Miura T, Luo Y, Loring JF, et al. Assessing self-renewal and
differentiation in human embryonic stem cell lines. Stem Cells. 2006;24(3):516–530.
44. Babaie Y, Herwig R, Greber B, Brink TC, Wruck W, Groth D, et al. Analysis of Oct4-
dependent transcriptional networks regulating self-renewal and pluripotency in human
embryonic stem cells. Stem Cells. 2007;25(2):500–510.
45. Ploner C, Rainer J, Lobenwein S, Geley S, Kofler R. Repression of the BH3-only
molecule PMAIP1/Noxa impairs glucocorticoid sensitivity of acute lymphoblastic
leukemia cells. Apoptosis. 2009;14(6):821–828.
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876

21
46. Jia L, Yang J, Hao X, Zheng M, He H, Xiong X, et al. Validation of SAG/RBX2/ROC2
E3 ubiquitin ligase as an anticancer and radiosensitizing target. Clin. Cancer Res.
2010;16(3):814–824.
47. Gomez-Bougie P, Ménoret E, Juin P, Dousset C, Pellat-Deceunynck C, Amiot M. Noxa
controls Mule-dependent Mcl-1 ubiquitination through the regulation of the Mcl-
1/USP9X interaction. Biochem. Biophys. Res. Commun. 2011;413(3):460–464.
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876

22
Figure legends:
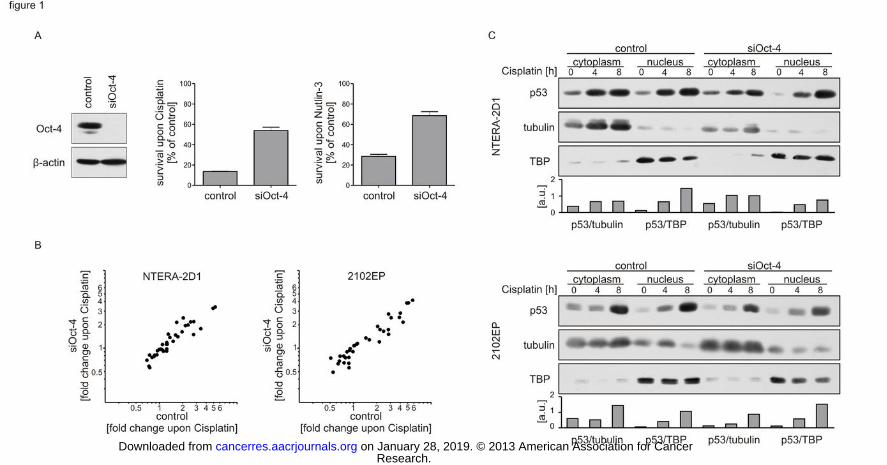
Figure 1: Oct-4 depletion does not compromise the p53 activity in response to Cisplatin.
Cells were transfected with indicated siRNA and incubated for 48 h prior to Cisplatin or
Nutlin-3 treatment. Controls reflect cells treated with a non-targeting siRNA. (A) p53
hypersensitivity is dependent on Oct-4 in NTERA-2D1 cells. Left panel: verification of Oct-4
knockdown by Western Blot. Right panels: quantification of survival upon Cisplatin by flow
cytometry. Cells were stained with Annexin-V/FITC and PI upon treatment with Cisplatin
(16 h) or Nutlin-3 (24 h). Graph reflects means ±SD of survival upon Cisplatin or Nutlin-3
from 3 independent experiments. (B) Oct-4 depletion does not influence the transcriptional
response of p53 to Cisplatin. Cells were incubated with or without Cisplatin for 6 h. To
evaluate p53 transactivation activity upon Cisplatin treatment in dependency of Oct-4,
transcripts of 46 bona fide p53 target genes were quantified by qPCR. For this, RNA was
isolated, transcribed to cDNA and applied to qPCR analysis using the primers listed in
Supplementary Table 1. Graph shows means of fold changes upon Cisplatin of detected
transcripts from 3 independent experiments, dots depict fold change values of each transcript
in cells pre-treated with control (X axis) or Oct-4 siRNA (Y axis). (C) Oct-4 depletion does
not affect p53 accumulation and localization. Cells were treated with Cisplatin for indicated
times and subjected to subcellular fractionation. p53 protein was analyzed by Western Blot
along with tubulin, representing the cytoplasmic compartment and TBP representing the
nuclear compartment. Graphs reflect densitometric quantification of p53 protein levels
normalized to tubulin or TBP, respectively.
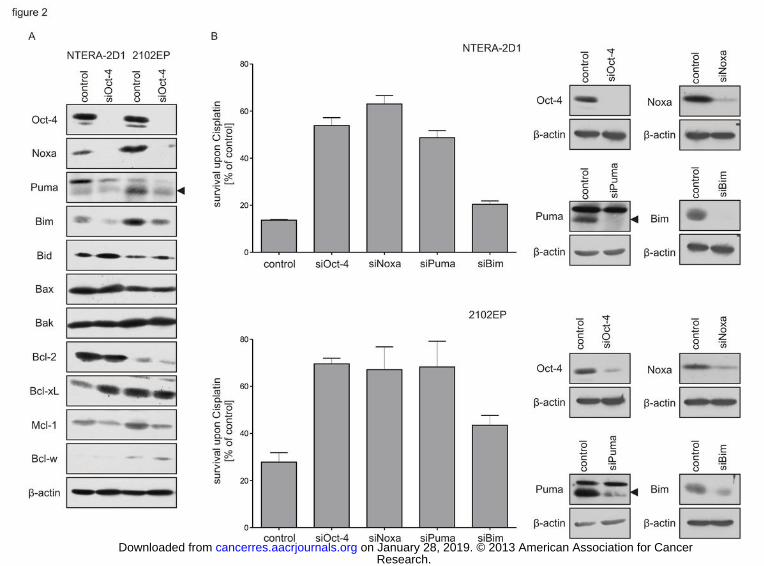
Figure 2: Composition of Bcl-2 family proteins is dependent on Oct-4 and determines
Cisplatin hypersensitivity. Cells were transfected with indicated siRNA and incubated for
48 h. Controls reflect cells treated with a non-targeting siRNA. (A) Noxa, Puma and Bim
expression is dependent on Oct-4. Transfected cells were lysed and subjected to Western Blot
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876

23
analysis to determine protein levels of Bcl-2 family members. (B) Silencing of Noxa and
Puma but not Bim leads to a marked reduction in Cisplatin sensitivity comparable to that
observed upon Oct-4 knockdown. Left panel: quantification of survival upon Cisplatin by
flow cytometry. Transfected cells were treated with Cisplatin for 16 h (NTERA-2D1) or 12 h
(2102EP) and stained with Annexin-V/FITC and PI. Graph reflects means ±SD of survival
upon Cisplatin from 3 independent experiments. Right panel: verification of indicated
knockdowns by Western Blot
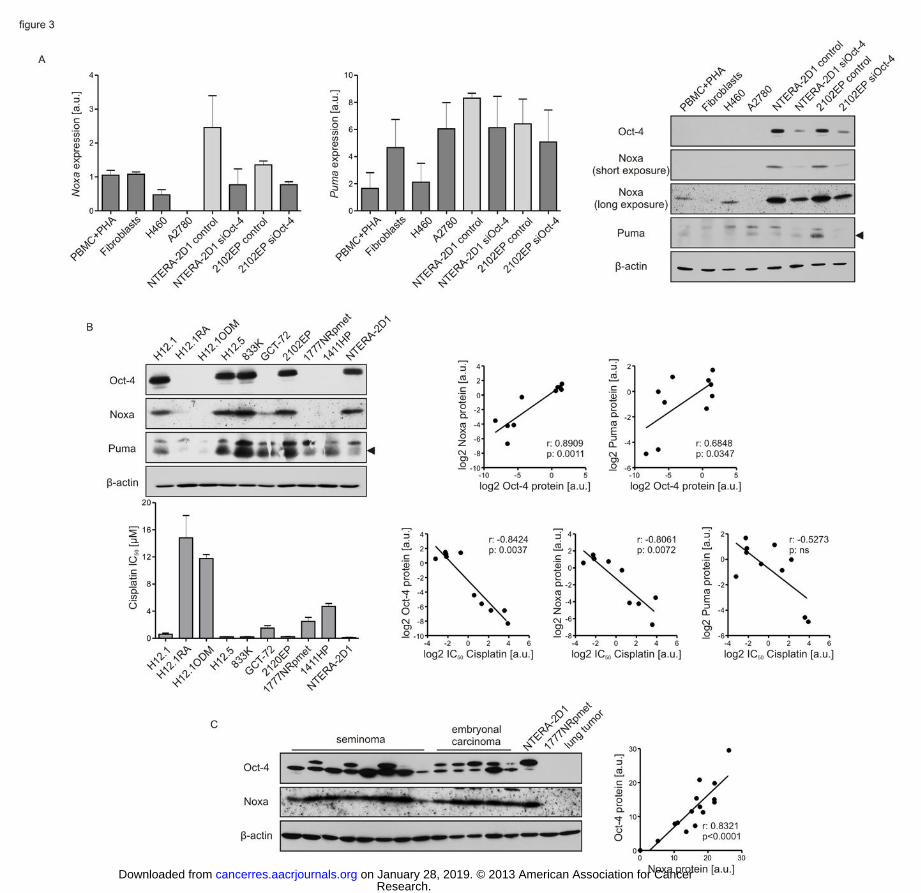
Figure 3: Noxa protein levels are tightly correlated with Oct-4 status and Cisplatin
sensitivity in TGCT cell lines and patient samples. (A) Oct-4-positive EC cells express
high constitutive Noxa mRNA and protein levels. NTERA-2D1 and 2102EP cells were
transfected with indicated siRNA and incubated for 48 h prior to analysis. Controls reflect cell
treated with a non-targeting siRNA. The other cells were left untreated and used as a control.
Left panels: RNA was isolated, transcribed to cDNA and applied to qPCR analysis. Graph
reflects means ±SD of Noxa and Puma transcript from 3 independent experiments. Right
panel: cells were harvested and lysed to analyze Oct-4, Noxa and Puma protein levels by
Western Blot. (B) Constitutive Noxa protein levels are highly correlated with Oct-4 and
Cisplatin hypersensitivity in TGCT cell lines. Upper left panel: cells were harvested and lysed
to analyze Oct-4, Noxa and Puma protein levels by Western Blot. Lower left panel: IC50 of
Cisplatin were determined by SRB microculture colorimetric assay. Graph reflects means
±SD from 3 independent experiments. Upper right panels: dot blots reflect correlation of Oct-
4 with Noxa or Puma protein as determined by densitometric analysis. Lower right panels: dot
blots reflect correlation of Oct-4, Noxa and Puma protein levels with IC50 of Cisplatin. (C)
Noxa and Oct-4 protein levels are highly correlated in cancer patient samples. Whole protein
lysates were obtained from frozen patient samples of 5 EC, 8 seminoma and a lung tumor and
applied to Western Blot analysis to determine Oct-4 and Noxa protein levels. Lysates from
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876

24
NTERA-2D1 and 1777NRpmet cell lines served as positive or negative control, respectively.
Graph reflects correlation of Oct-4 and Noxa protein levels.
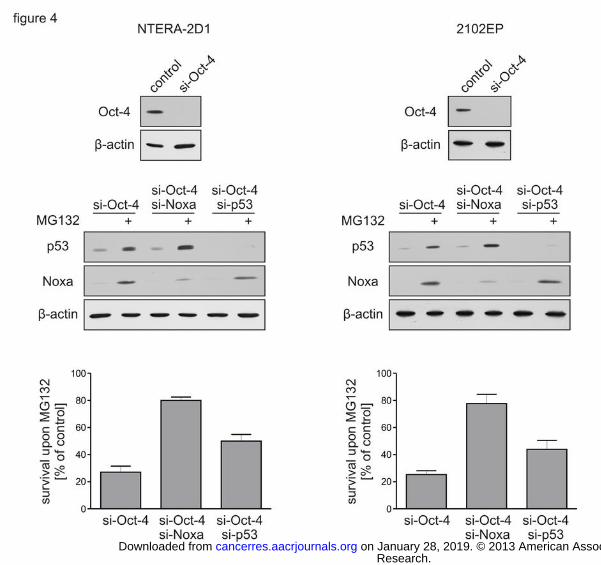
Figure 4: Accumulation of both Noxa and p53 is required to induce cell death in Oct-4-
depleted TGCT cells
Cells were transfected with indicated siRNAs and incubated for 48 h before MG132
treatment. Controls reflect cells treated with a non-targeting siRNA. Efficient gene silencing
was verified by Western Blot analysis (upper and middle panels). Transfected cells were
treated with MG132 for 20 h (NTERA-2D1) or 16 h (2102EP), protein lysates were prepared
and subjected to Western Blot analysis to determine protein levels of p53 and Noxa (middle
panels) and stained with Annexin-V/FITC and PI for cell death analysis by flow cytometry
(lower panels). Graph reflects means ±SD of survival upon MG132 from 3 independent
experiments.
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876

Published OnlineFirst January 9, 2013.Cancer Res Matthias Gutekunst, Thomas Mueller, Andrea Weilbacher, et al. determined by high constitutive Noxa levels mediated by Oct4Cisplatin hypersensitivity of testicular germ cell tumors is
Updated version
10.1158/0008-5472.CAN-12-2876doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2013/01/09/0008-5472.CAN-12-2876.DC1
Access the most recent supplemental material at:
Manuscript
Authoredited. Author manuscripts have been peer reviewed and accepted for publication but have not yet been
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/early/2013/01/08/0008-5472.CAN-12-2876To request permission to re-use all or part of this article, use this link
Research. on January 28, 2019. © 2013 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on January 9, 2013; DOI: 10.1158/0008-5472.CAN-12-2876