10 11 12 14 - kofstmycobio.kofst.or.kr/upload/pds/epapyrus/mycobiology/14-0148/14... · 23...
TRANSCRIPT

1
The Mushrooms: an ethnomycological, molecular and nutritional analysis from the perspective of ethnic 1
communities 2
Ethnomycobiology and mushrooms 3
Madhusmita Borthakur, Polashree Khaund , S.R. Joshi* 4
Microbiology Laboratory, Department of Biotechnology & Bioinformatics, 5
North-Eastern Hill University, Shillong 793022, Meghalaya, India 6
Corresponding address: [email protected] 7
8
Abstract: 9
Micro-organisms are considered as priceless sources for various important biomolecules having enormous 10
commercial applications in medicine, food industry and bio-anthropology. Biodiversity hotspot regions are 11
considered to be genetic paragon of plant, animal and microbial resources. This mega-biodiversity zone encompasses 12
areas such as the sacred groves which are considered as the abode of deities by ethnic people. These groves are rich 13
in flora and fauna besides the microbial and macrofungal components. Mushrooms are considered as highly 14
nutritious which carry added medicinal value. However, their bioprospection and commercial exploitation is very 15
scarce as compared to other life forms. This may be due to the limited information available on their nutritional and 16
genetic constituents. Proteins and polysaccharide comprise the main component of their dry matter with glycogen, 17
chitin, trehalose, mannitol as the main carbohydrate while n-3 fatty acids are negligible. High proportion of fibre 18
content, less expensive and easy availability has attracted both the interest of researchers and consumers to the 19
science of mushrooms. The ethnomycological, nutritional and molecular aspects of mushrooms has been reviewed in 20
this paper. 21
Key words: Ethnic community, Bioanthropology, Ethnomycology, Mushrooms 22

2
INTRODUCTION 23
Microorganisms are considered to be the richest repository in natural environment [1]. Microbial diversities are 24
found pervasive in every corner of earth. There are as many as 25 biodiversity rich regions of the world [2] and Asia 25
is considered as one of the mega-diversity hotspots which hold 60-70% of the world’s biodiversity [3] including 26
microorganisms. Among various microorganisms, fungi play a vital role in maintaining the sustainability of our 27
mother Earth. The diversity of the fungi was debated for more than 200 years but in last few decades, the DNA 28
sequence has provided a myriad of new characteristics for the analysis of 100 phylum from the kingdom fungi [4, 5]. 29
Fungi are found from the equatorial regions to the poles, from marine ecosystems to fresh water environment [6] 30
surpassed by only bacterial community in their inability to withstand water activity, extreme temperature conditions 31
and carbon sources [5]. Fungal community are considered to be existing in two diverse characteristics - sociological 32
which includes their geographical distribution, community structure, their diversity in various community, habitat 33
specificity and their second aspect of characteristics is their behavioral characteristics which relates to their function 34
in an ecosystem [7]. There are estimates of more than 1.5 million fungi [8] in the world out of which only 98,998 35
species have been described [9]. Mycotechnology has advanced in last five decades for their potential in various 36
organic acids, extracellular enzymes, antibiotics, antioxidant activity, in the field of bioremediation, biome-seed, etc 37
[10, 11]. 38
Malnutrition in terms of deficiency in protein is one of the foremost factors responsible for the morbidity and 39
mortality in under-developed as well as other developing countries in the world [12]. With population explosion, the 40
protein deficiency is expected to become more acute in the coming years and recommending animal protein is 41
beyond the limit for some developing countries. Non-traditional food resources can partly meet the dietary deficit. 42
With the advent of modern molecular biology tools, fungus has been renamed as a ‘cell factory’ for various 43
heterologous proteins [13, 10] which also include human proteins. Mushrooms produce many bioactive proteins and 44
peptides, primarily including lectins, laccasses, fungal immunomodulatory proteins (FIP), antimicrobial/antifungal 45
proteins, ribosome inactivating proteins (RIP), ribonucleases and many of these bioactive compounds have entered 46
into phase I, II and III clinical trials and are successful in treating cancers in world [14]. 47
This review emphasizes mostly on the filamentous fungi, their nutritional perspectives and cytogenetics, since bulk 48
of research has been done in this group and is considered as a alternative food source for sustainable development of 49
a country’s economy. 50

3
MUSHROOMS - THE MACROSCOPIC FUNGI 51
Mushrooms are considered as well-balanced food stuffs and good source of nutrition in the era where whole world 52
suffers from scarcity of food. Geologically, mushrooms existed on earth even before the appearance of human, as 53
evidenced from the fossil records of the lower cretaceous period [15]. The term mushroom was first defined ‘as 54
macrofungi with a defined fruiting body that can be either epigeous or hypogeous’ which can be easily visible to 55
naked eye [16]. Mushrooms belong to basidiomycetes family with stalk and umbrella shape fruiting body bearing 56
either gills or pores although from taxonomical view, some species in ascomycetes family also belong to mushroom. 57
It first appears as a white tiny balls with a cap, also known as pileus and a short stem also known as stipe. The 58
membrane or veil, also known as velum covers the envelope which gets torn off and cap become visible with gills or 59
lamellae radiating down beneath the cap [12]. Several species of mushrooms are known because of its characteristics 60
color, texture and pleasant odour [16](Fig I). It has been estimated that there are more than 140000 mushrooms with 61
only 10% being known [16, 17]. Being from basidiomycetes family, they have compatible haploids commonly the 62
mycelia being composed of filamentous hyphae. Through plasmogamy, haploid compatible mycelia fuse with each 63
other and nuclei migrate into each other’s mycelia and get paired with the resident nuclei. Since karyogamy is 64
delayed, compatible nuclei remain in dikaryotic stage for some time. Often it is seen that mycelium in dikaryotic 65
stage is more vigorous than the monokaryotic stage (the haploid mycelia) and can take up the substrate more easily 66
in which they are growing. Basidiomycota is characterized with the formation of clamp connection. The long lasting 67
dikaryotic period produce basidia, in which a congruent nuclei fuse (karyogamy) to form a diploid cell. Meiosis is 68
followed; by the production of 4 haploid nuclei that transfer into the external part forming basidiospores. However, 69
variation are always observed ; Craterellus spp. often have 6-spored basidia while Sistotrema spp. have 2,4, 6 or 8 70
basidiospores and the common button mushroom, Agaricus bisporous has 1, 2, 3 or 4 spored basidia under some 71
circumstances. Basidiospores are discharged to start up a new haploid mycelia also known as monokaryons. The 72
karyotype of mushrooms has not yet been very clear because of contingency of endomitosis and small size of 73
mushroom chromosomes [2]. The cultivation of mushroom is termed as “non green revolution”[18]. The genome 74
size and chromosome numbers vary in mushrooms when analyzed through various analytic techniques which include 75
Restriction Fragment Length Polymorphism (RFLP) and Randomly Amplified Polymorphic DNA (RAPD). 76
Mushrooms are being consumed over 10 kg yearly in few European countries with a statistical mean of 5.6 kg of 77
cultivated fresh mushrooms per household/year [19]. World- wide mushroom cultivation is dominated by Agaricus 78

4
bisporus [20] which is not only edible but also has been valued as a source of traditional medication by humankind 79
[17]. Considering their nutraceutical and ethnocultural relevance, there is a need of conservation, preservation and 80
characterization of the wild mushrooms from the pristine forests owing to the vigorous destruction of the wild habitat 81
by various anthropogenic activities. The economic prospects of such wild mushrooms which are yet to be identified 82
for their activities including medicinal and antioxidant can assist in their conservation and economic subsistence. 83
SOURCES AND ETHNOMYCOLOGY OF MUSHROOMS 84
In the recent era, the hub for research on mushrooms has turned to undisturbed forests and pristine forests like sacred 85
groves. Sacred groves with a concept of “presiding deity” have been protected by the tribal community. The section 86
contains geographical features along with vegetations and other forms of flora and fauna, demarcated by the 87
indigenous people to keep them in a protected and undisturbed state which have formed an intricate relationship 88
between human and nature Earth [21]. These virgin forests are rich in biodiversity with a huge amount of endemic 89
and endangered species. These forests in recent days have attracted attention for numerous issues including reservoir 90
of threatened species, repositories for various medicinal plants, macrofungi (mushrooms), giant trees, gene bank for 91
economic species, recreation center for urbanites and a huge untapped paradox for researchers and environmentalist 92
[22, 23]. Due to numerous environmental transformations such as climatic variations, human interference such as 93
shifting cultivation and other anthropogenic activities, these forests have dwindled its size leading to the loss of 94
pristine habitats. 95
Sacred groves are protected and largely untrapped with a limited knowledge of the indigenous flora, fauna and 96
microbes. Traditional knowledge on conservation and preservation of its wild habitat are seen among the native tribal 97
communities. The knowledge emerged over the generation for conservation of these groves is as much a resource as 98
a biodiversity. Wild mushrooms (macrofungi) are highly dominant in such groves and only few reports on wild 99
mushrooms are available [24] while their medicinal prospects, antimicrobial, anti-oxidative, trace element prospects 100
are yet to be documented. Tribal Communities are known to collect the wild mushrooms based on their 101
ethnomycological knowledge. Wild poisonous mushrooms are distinguished from their edible counterparts in having 102
a pointed cap, bright colored fruiting body, unpleasant odour, colour of the fruiting body often turns black upon 103
breakage, absence of worms and insects [25]. 104
Ethnomycology is considered as the study of fungi for the development of cultures, religion and mythology. In this 105
respect mushroom received attention for their potential as a hallucinogen, their edibility for the presence of abundant 106

5
bioactive and nutritional compounds and for their toxicity. China have a long tradition of using mushroom since last 107
5000 years due to its therapeutic and consumable purpose but its therapeutic value has also been recognized by many 108
early civilizations including Greeks, Aryan, Romans, Egyptians, Romans and Mexicans. Mushrooms were 109
considered as a “food of God” by Romanians, “strength food” by Greek warriors, “elixir of life” by Chinese, as 110
“Somrus” a decoction of mushroom by Indians [26]. The brown rot fungi, Formitopsis officinalis are recognised as a 111
“Bread of Ghosts” by the indigenous people of Pacific Northwest and use it to mark the grave of shamen. Shamen of 112
Pacific Northwest use the sporophores of Formitopsis officinalis for various rituals and to treat illnesses caused by 113
supernatural powers. They believe the shape of the mushroom sporophore gave mushroom its spirit catching abilities. 114
Apart from Formitopsis, an obligate plant pathogen Exobasidium vaccinii infect stem, leaves and flowers of 115
Ericaceous plant and subsequently form vegetative deformities thus forming a basidia covered gills which are 116
consumed by the indigenous coastal groups of Pacific Northwest and considered them to be berries. The berries are 117
referred to as “Ears of Ghost” and believe in a myth that they are associated with corpse-stealing, child- kidnapping 118
creature [26]. Burk (1983) reported the spiritual and religious use of puffballs mushrooms by the people of North 119
America and they believed that they have the ability to ward off ghosts [27]. Puffballs also received attention in 120
British Columbian Indian spiritual world as they are regarded as “ghost’s make up” or “ground ghost” or “corpse” as 121
reviewed by Nicholson (2009)[26]. In Asia, the mycophilic societies are associated among the indigenous people of 122
North Eastern India, Western ghats and North Western India, China. The local inhabitants collect mushrooms from 123
their neighboring localities, meadows and forests for consumption and sell them to earn revenue for their family 124
during monsoon season when other forest nonwood products are unavailable in the market. Among various species 125
of mushrooms which are commonly consumed by the people are Termitophilous which include T. microcarpus, T. 126
aurantiacus, T. eurhizus, T. clypeatus, T. Tyleranus [28] Few indigenous tribes of North-Eastern corridor of the India 127
such as Khasis of Meghalaya use a traditional technique ‘narsuh’ for cooking mushrooms where they heat the tip of a 128
small iron rod and place it in the middle of the bowl containing cooked mushroom. They believe that heat release 129
from the tip is responsible to destroy or absorb the poisonous harmful substance from the mushrooms [25].Some use 130
other traditional methods. The fruiting bodies are washed and boiled with a few cloves of garlic. If the cloves of 131
garlic turn black in color, they believe that the mushrooms are poisonous in nature. However, if there is no color 132
change then they are safe to eat. 133
CYTOGENETICS OF MUSHROOM 134

6
Basidiomycetes contain two types of mycelia, monokaryon (asexual) and dikaryon (sexual) which allow easy 135
exchange of nuclear partners in monokaryotic - dikaryotic and dikaryotic- dikaryotic mycelial fusion [29]. For most 136
cultivable mushroom in the world, Agaricus bisporus researchers have not been able to provide comprehensive 137
information on various expect such as the genetic, physiology, cytology and breeding [30]. Since fungal 138
chromosomes are minute, pulse field gel electrophoresis enables the karyotyping of the various mushroom species 139
[31] showing molecular size and numbers of chromosomes. A study by Sagawa and Nagata (1992) reported the size 140
of six chromosome ranging from 2.1 to 5.7Mbp (million base pair) per chromosome and genomic size to be 20.8Mbp 141
of Pleurotus ostreatus[32]. The size of mushroom chromosomes DNAs range from 2Mb to 30Mbp. Fruiting bodies 142
normally develop on dikaryon. Karyotypic analysis of the mushroom fruiting bodies can be revealed by spreading 143
the chromosome at meiotic prophase stage. Karyogamic in basidiomycetes occur directly before meiosis where intra-144
tetrad fertilization occur, that is the nuclei of one tetrad unite in pairs but do not fuse [33] which normally takes place 145
in the gills of the fruiting bodies which is followed by meiosis (maturation of basidia, an autonomous endotrophic 146
process) and by production of meiotic basidiospores. A report by Larraya et al (1999) suggest that chromosomal 147
length polymorphism are most common in basidiomycetes and are analyzed by electrophoretic karyotyping, however 148
two approaches have been proposed for polymorphism to occur, by increasing the copy number of ribosomal DNA 149
segment or subtelomeric region of DNA and by mitotic and meiotic recombination process [34]. From various 150
studies it has been reported that both monokaryon and dikaryon can form multicellular resting bodies or sclerotia and 151
various mitotic spores, the tiny uninucleate aerial oidia and large thick walled chlamydospores within submerged 152
mycelium [35, 36, 37]. In a study by Summerbell et al (1989) it has been reported that one spore receives two non-153
sister nuclei, i.e. the progeny is formed of two different nuclei in first mitotic division [38]. However, in 154
basidiomycetes, mating is supported by population genetics evidence of recombination and also from intravarietal 155
hybrids from the environment [39]. Spore production can be examined by analyzing molecular markers of progeny 156
that are derived from a single basidium. A report suggest that in Cryptococcus neoformans basidium, there occurs a 157
single meiosis and other multiple rounds of mitosis to generate nuclei in the basidium [40]. Spores remain in 158
aneuploid state and do not germinate rather remain dormant. Chains are formed from heterogenous population of 159
spores of different genotypes due to nuclear movements which occurs in basidium [41]. Nuclei can remain fixed in 160
the basidia from which the spores bud or can move after every mitosis which can get influenced by environmental 161
factors including temperature, light, humidity and nutrients. Light conditions determine the formation of hyphal 162

7
knots or oidiophores with oidia and also determine whether hyphal knots will mature into sclerotia or fruiting body. It 163
induces hymenium differentiation and finally the karyogamy. Among genes specifically expressed within fruiting 164
body for hyphal aggregation are β-galactoside binding lectins [29]. Variation in temperature also determines the 165
formation of fruiting body. Lower temperature favors the formation of fruiting body initial and lower temperature 166
favors the formation of oidia [36, 42, 43, 44, 45, and 46]. Fruiting body needs high humidity whereas sclerotia and 167
chlamydospores will appear at low humidity. In later half of the development after 48 h, light is required for the 168
induction of karyogamy during DNA synthesis prior to the formation of nuclear fusion [43, 47, 48, 49, 50, 51, 52, 169
53] while stipe elongation and maturation of cap takes place parallelly. For the formation of fruiting bodies, around 170
20 to 37 0C temperature is necessary but prior to the meiotic DNA replication, a two hours dark period at 37
0C is 171
utmost important. The ratio of carbon and nitrogen also determine the initiation of fruiting body formation and 172
sclerotium maturation, whereas fruiting body initiation is inhibited by glucose and glucose analogues [54, 55, 56, 57, 173
and 58]. 174
TAXONOMICAL STUDIES TO ANALYZE VARIOUS STRAINS 175
The study of mushrooms to distinguish among different strains is perhaps inadequate based on morphological 176
features. For the genetic analysis, it is essential to express both the alleles in heterozygous which is possible by 177
allozyme analysis (recognition of codominant phenotype) which involves movement and separation of enzymes in 178
gel. Electrophoretic mobility of the allozyme is directly correlated with the genotypic variations among different 179
strains within a single species [16]. Among the species identified by multilocus enzyme electrophoresis are wild 180
mushroom including Agaricus bisporus, Agaricus campestris, Volvariella volvacea, Lentinula edodes, Pleurotus spp. 181
and Morchella spp. 182
Arbitrarily primed polymerase chain reaction, a marker based upon DNA probe technique is also advantageous 183
process to distinguish between different strains of mushrooms over allozyme analysis. The amplicons are revealed by 184
electrophoresis and are characterized by direct sequencing. 185
Edible and poisonous mushrooms are characterized by mycologists based on certain palynological, sporological and 186
morphological trait, but very few reports are available on their molecular aspects. In the genus Psilocybe, P. 187
semilanceata and P. cubensis produce hallucinogenic effect while P. merdaria and P. montana donot produce such 188
effect [59]. Few toxic mushrooms belonging to the genus Lepiota are identical to that of the edible mushroom of 189
genus Macrolepiota. Similarly, Inocybe asteropora can be misidentified as Armillaris spp.[60]. Rapid molecular tools 190

8
including molecular markers, PCR analysis would support the identification of poisonous nature of mushrooms. 191
Various toxins including amanitoxins, phallotoxins, monomethylhydrazine, orellanine, muscarine, ibotenic acid, 192
muscimol, coprine, psilocybin, psilocin are confined to various wild macrofungi [61]. An interesting poisonous 193
mushroom is Gomphus floccosus. Consumption of this mushroom in the northern United States and in Canada has 194
been reported to cause severe gastrointestinal disorders [62]. The consumption of this poisonous mushroom by the 195
ethnic population of Meghalaya without any apparent harm may be hypothesized to reduced toxicity of the specific 196
local strain available in the region, unique indigenous cooking practices and methodology or even to genetic 197
resistance to the mushroom toxin developed among the local population [25]. Poisonous and edible ones are grouped 198
and named as toadstool by English speakers and “jhasmuka” or “lu” among the Mayans [63]. Epis et al (2010) 199
developed species specific molecular marker to identify the poisonous nature of four different mushrooms including 200
Amanita phalloides, Lepiota cristata, Lepiota brunneoincarnata, Inocybe asterospora based on the ITS1-5S rDNA-201
ITS2 sequences which are highly conserved among the fungi [60]. Lee et al (2000) developed primers 202
complementary to ITS1 locus to differentiate between hallucinogen mushroom Panaeolus and Psilocybe based on 203
the length of the PCR product [64]. 204
Identification of novel molecular markers and rapid PCR protocol can be an approachable strategy to differentiate the 205
hallucinogen from edible counterpart. 206
In a study on Amanita mushrooms it was found that amatoxins, the lethal constituents of these poisonous mushrooms, 207
are bicyclic octapeptides [65]. Two genes in Amanita bisporigera, AMA1 and PHA1, directly encode α-amanitin, an 208
amatoxin, and the related bicyclic heptapeptide phallacidin, a phallotoxin, indicating that these compounds are 209
synthesized on ribosomes and not by nonribosomal peptide synthetases. AMA1 and PHA1 are present in other toxic 210
species of Amanita section Phalloidae but are absent from nontoxic species in other sections. The genomes of 211
Amanita bisporigera and Amanita phalloides contain multiple sequences related to AMA1 and PHA1. In another such 212
study by Qing et al (2012), to establish a standard DNA barcode for poisonous species of Amanita in China, three 213
candidate markers, the large subunit nuclear ribosomal RNA (nLSU), the internal transcribed spacer (ITS), and the 214
translation elongation factor 1 alpha (tef1α) were tested using the eukaryotic general primers for their feasibility as 215
barcodes to identify seven species of lethal fungi and two species of edible ones which can easily be confused with 216
the lethal ones known from China [66]. This study showed that the tef1α and nLSU can be proposed as 217
supplementary barcodes for the genus Amanita, while ITS can be used as a primary barcode marker considering that 218

9
the ITS region may become a universal barcode marker for the fungal kingdom. 219
In the study on the wild mushrooms which are consumed by the ethnic tribal population of India, it was found that 220
the differentiation between the poisonous and edible varieties made strictly based on the morphology and molecular 221
characterization often do not bestow apparent results. Molecular characterization being confined to one conserved 222
locus, ITS1 and ITS4 only identify various macrofungi rather than discriminating among edible and closely 223
associated poisonous mushroom (Fig II, Fig III). An alternative DNA barcode based on mitochondrial COI gene 224
allows correct identification of only 20-70% of fungal species [67, 68]. In a study on wild edible mushrooms, 225
morphological features and ITS markers have been proposed as barcode for mushroom identification [69]. So, a hunt 226
for a novel primer for rapid differentiation among the poisonous and edible one are in progress to discriminate the 227
two varieties to a greater extent. 228
MUSHROOMS AS DIETARY SUPPLEMENTS 229
Mushrooms have been used as a food item since time immemorial [70]. Mushrooms are considered as the world’s 230
largest untapped resources of dietary supplements. Their consumption increased due to the presence of high amount 231
of protein content and trace mineral [71, 72]. In comparison with other eatables including fruit, vegetables and 232
legumes, mushroom or basidiomycetes can prove better as a rich source of certain dietary factors that can be 233
beneficial to human health and growth [73]. They are considered as a healthy food which contains essential fatty 234
acids, low in the amount of calories and high in the concentration of the protein, fats and minerals [5]. As wild 235
mushrooms are considered to have more amount of protein content than commercially available mushroom, several 236
reports reveal that they are used as a diet to combat various diseases [15]. For a developing country like India, they 237
can serve as a good substitute for protein malnutrition [20]. Mushrooms with fleshy fruiting bodies can be used as a 238
good source of biopharmaceutical product as they contain polysaccharides and triterpenoids [70]. Various studies 239
have shown that after cooking, dried mushrooms show highest level of nutritional value due to insufficient 240
rehydration [74]. In general mushrooms contain 90% water and 10% dry wight [75]. In a study by Orgundana and 241
Fagade (1981), mushrooms are reported to contain about 16.5% dry matter, the approximate composition of crude 242
protein is 14.6%, crude fibre is 7.4%, and composition of fat and oil is around 4.48% [76]. Though mushrooms are 243
commercially used as a source of nutritional supplement, they have an adverse effect in the health of human as they 244
are also considered as a source of human poisoning and are associated with various carcinogenesis while 245
experimenting in animal [73]. According to a study by Chang (1990), only 2,000 mushrooms are considered edible of 246

10
which 20 are commercially cultivated and 4 to 5 are cultivated industrially [77]. Among various higher species of 247
basidiomycetes, Agaricaceae are widely consumed worldwide. Many polysaccharide and protein based 248
polysaccharide isolated from Agaricaceae have anti-tumour activity and specific or non specific immune response 249
activity [36, 37, 42, and 74]. The dietary fibre of mushroom is due to the polysaccharides and triterpenoids contained 250
in them. The cell wall of mushroom contains a mixture of fibrillar and matrix components which comprises chitin, 251
polymer of β 1- 4 linkages N-acetyl-glucosamine and polysaccharides of 1-3 linkage of β-D-glucans and mannans 252
respectively. Mushrooms bear mainly the water insoluble dietary fibers which include both the chitin and β-glucans. 253
Under unfavorable environmental conditions, mushrooms regenerate a special structure known to be a sclerotia 254
which are compact mycelium structure bearing chitin and β-glucans with β 1-3 backbone and 1- 6 linked side 255
branches and due to this they are considered to be a novel source of dietary component as human enzyme cannot 256
digest them [70]. 257
PROTEIN COMPOSITION 258
Mushrooms being rich in the protein component comprise mostly of the essential amino acid component lysin, 259
histidine, arginine, threonine and tryptophan that are commonly not present in cereals. The component of lysin has 260
been reported as a most abundant essential amino acid and methionine and tryptophan being the least abundant 261
amino acid. The content of free amino acid is low which consist of only 1% of the dry weight of mushroom, thus 262
limiting the nutritional level but they participate in the taste of mushroom [19]. The protein component of mushroom 263
depends on its growth substratum, the level of nitrogen and its location, the size of the pileus, varies among species 264
and its harvesting time [78]. Immature primordia contain more amount of protein than mature primordia as reported 265
in Agaricus bisporus and Pleurotus spp. [79]. The composition does not change during oven drying of mushroom in 266
40 degree centigrade but significant reduction in its composition can be visualized upon boiling of fresh mushroom 267
(19). Protein production efficiency for mushroom is about twice that of cabbage and asparagus, four times as of 268
orange, 12 times as of apples [77]. On the basis of its dry weight it contains 65% protein as compared to 25.2% in 269
milk, 20% for pork meat and 4% for beef , 39.1% in soybean, 7.3% in rice, 13.2% in wheat [77, 80]. An early study 270
by Rose (1937) had compared the presence of all essential amino acid for the growth of mammalian cells, which 271
revealed that the composition of amino acid in mushroom is adequate except in phenylalanine and methionine[81]. 272
The palatable taste of the mushroom is enhanced by the presence of aspartic and glutamic acid which are 273
monosodium glutamate-like (MSG-like) components [82]. It has been reported by Bauer-Petrovska (2001) that mean 274

11
level of protein fraction such as albumins, prolamines, globulins, glutenins, glutenin-like material and prolamine-like 275
material are likely to be 24.8, 5.7, 11.5, 11.5, 7.4 and 5.3% respectively [83]. Proteins and peptides produced by 276
mushrooms possess interesting biological activities such as lectin, antimicrobial protein, laccases, ribonucleases, 277
ribosome inactivating protein and fungal immunomodulatory protein [14]. 278
A wide range of bioactive compounds have been isolated from mushrooms which include protein- polysaccharide 279
complexes, proteins, polysaccharides, etc [84, 85]. Among those the most specific are lectin protein, which are 280
glycoproteins that can bind to cell surface carbohydrates with their ability for cell agglutination. They exhibit 281
antiproliferative activity towards tumor cell lines specifically to human leukemic T cells, breast cancer MCF7 cell, 282
hepatoma Hep G2 cells and can recognise human blood group A determinant carbohydrates [14]. The sacred 283
mushroom Reishi (Ganoderma spp.) contain a polysaccharide, β-glucan which stimulate or modulate the level of 284
immune system by activating T cells and macrophages as well as activating immunoglobin levels which can combat 285
and respond against foreign cells [86]. Crystal structure of lectin protein was first determined by Cioci et al (2006) 286
with a regular seven-bladed beta-propeller fold with N-terminal region being stuck into the central cavity around 287
pseudo 7-fold axis [87]. Ribosome inactivating protein enzymes which eliminate one or more adenosine residue from 288
rRNA inactivates ribosomes, inhibit proliferation of HIV- 1 reverse transcriptase activity [88]. 289
Another isolated bioactive compound from mushroom is laccase, which is a multicopper oxidase with its 290
implication in inhibiting HIV-1 reverse transcriptase, proliferation of hepatoma HepG2 cells and MCF 7 tumour cell 291
[89]. Fungal immunomodulating proteins was isolated from various mushrooms which include Ganoderma sp., 292
Russula paludosa, Pleurotus citrinopileatus, Grifola frondosa, Antrodia camphorata [90, 91, 92, 93, 94, 95, 96, 97] 293
with various immunomodulating, antitumour and antiviral activity. A novel Se-containing protein Se-GL-P (36 kDa) 294
isolated from Ganoderma lucidum using ammonium sulphate precipitation, indicated the incorporation of selenium 295
in the protein in the form of selenocysteine and selenomethionine having a positive response towards inhibiting the 296
multiplication of tumour cells [91]. Various other proteins have been isolated which include trichogin protein from 297
Tricholoma giganteum with their antifungal activity against Mycospaerella arachidicola, Fusarium oxysporum and 298
Psylla piricola [98]. Chen et al (2009) isolated a glycoprotein PCP-3A from the fresh fruiting body of oyster 299
mushroom which showed a positive antitumour activity [94]. A report by Chang et al (2007) assessed the utilization 300
of two proteins, Agaricus bisporus lectin (ABL) from Agaricus bisporus and immunomodulating Agaricus polytricha 301
protein (APP) from Auricularia polytricha which showed good stability after thermal, freezing, acid, alkali and 302

12
dehydration treatments, thus indicating the proteins to be stable immune-stimulant for food, health and 303
pharmaceutical applications [99]. 14-3-3 protein with molecular mass of 28.8kDa was isolated from Sparassia crispa 304
and Hericium erinaceum which can be a potential candidate protein for phylogenetic relationship analysis among 305
mushroom species because of its molecular biological feature such as similar pI, molecular mass and slightly 306
differing amino acid sequence [100]. 307
LIPID COMPOSITION 308
Fatty acids are considered to play a major role in immune system functioning and balancing the hormonal level [101]. 309
In basidiomycetes, lipid content ranges from 0.6 to 18.4% (w/w) [102]. The fatty acid profile of mushroom is 310
relatively higher in basidiomycetes which ranges from saturated palmitic acid (16: 0) to oleic acid (18:1, Δ9c) and 311
linoleic acid (18:2 Δ9c, 12c) that are present in the membrane of basidiomycetes [102, 103]. Mushrooms are mostly 312
dominated by the presence of unsaturated fatty acids. However, low proportion of oleic acid is characterized for the 313
mushroom species Agaricus spp. and Cantharellus cibarius [19]. Boletus species are known for their higher 314
concentration of monoenic acid. The concentration of oleic acid are higher in B. edulis, B. piperatus, B. subglabripes, 315
B. erythropus, B. subtomentosus and B. variipes as reported by Hanus et al (2008)[101]. From the above mentioned 316
Boletus species, the polar lipids, including phospholipids and betaine, showed 317
diacylglyceryltrimethylhomoserine(DGTS) and phosphotidylcholine (PC) as a major polar lipid of varying 318
concentration of 72-93% from the total amount of polar lipids. Neutral lipid content ranges from 0.7 to 9.4% and the 319
range of polar lipid content in case of Agaricus sp. ranges from 2.4 to 11.8% [102]. The quantity of polar lipids 320
account for more than 50% of total lipids. Unsaturated fatty acids account for an average of 74.4 % of the total fatty 321
acid. The ratios of unsaturated to saturated fatty acid (U:S) ranges from 1.61 for Lycoperdon pyriforme to 5.36 for 322
Boletus edulis. However, few unusual fatty acids, cyclopropane FA, 11,12 methyleneoctadecanoic acids are found in 323
nonpolar lipid fraction of many basidiomycetes [103]. Davidoff and Kom (1963) reported the presence of isomer cis-324
11 of heptadecenoic acid in the basidiomycetes(104). Elaidic acid (18:1 D9t), a common fatty acid in milk fat and in 325
the tissues of ruminant animals and occasionally in seed oils are found in few basidiomycetes as reported by Pfeuffer 326
and Schrezenmeir (2006)[105]. From human nutritional point, long chain of polyunsaturated linoleic acid and α-327
linolenic acid are important for basal metabolism in human and mushrooms are known to contain all these additional 328
value. Overall, the low value of lipid content in basidiomycetes is due to low proportion of desirable n-3 fatty acid. 329
CARBOHYDRATES AND FIBER CONTENT 330

13
The polysaccharide of mushroom is glycogen unlike that of starch as in plants. Mannitol, glucose and α, α- 331
trehalose(alphaD-glucopyranosyl-(1-1)alpha-Glucopyranoside) are main representatives of monosaccharides. 332
Carbohydrates usually account for the prevailing component of fruiting bodies. The content of carbohydrate on dry 333
weight basis ranges from 47.38 to 63.40g/100g in various basidiomycetes including Agaricus bisporus, Lentinus 334
edodes, Pleurotus ostreatus, Pleurotus pulmonarius [106]. However, the contents of trehalose and glucose in the 335
fruiting body of mushrooms are low, in order of 100 per gram of dry matter. In Agaricus bisporus, the synthesis of 336
mannitol is mediated by NADPH-dependent mannitol dehydrogenase using fructose as its substrate. It functions as 337
an osmolyte, which get accumulated at higher concentration in the fruiting body while after sporulation its amount 338
decreases drastically. Trehalose also serves as reserve carbohydrate in Agaricus bisporus which get synthesized in 339
mycelium and later translocated to fruiting body [107]. Mannitol concentration also varies widely among 340
basidiomycetes. The concentration is found to be of 1.0, 6.5 and 13.7% for dry matter of Tricholoma portentosum, 341
Agaricus arvensis and Lactarius deliciosus respectively [108] and 0.8, 0.2, 11.7 and 13.9% of dry matter of Lepista 342
Nuda, Lycoperdon perlatum, Ramaria botrytis and Cantharellus cibarius respectively [19]. The amount of trehalose 343
and mannitol get reduced considerably while boiling the fresh mushrooms, while freezing and drying result in only 344
limited losses [108]. In Cantharallus cibarius, trehalose and arginine are the most important compounds for carbon 345
assimilation [108]. Chitin is a water-insoluble structural polysaccharide that accounts for up to 80–90% of dry matter 346
in cell walls of mushroom. Contents of dry matter as reported from eight samples of Boletus spp. ranged from 6.8–347
10.2% [74]. Chitin being indigestible for humans apparently decreases the availability of other mushroom 348
components. 349
Information on dietary fibre content in wild growing mushrooms has been very limited. Boletus spp. is reported to 350
contain 4.2–9.2% and 22.4–31.2% of dry matter for soluble and insoluble fibres respectively for oyster mushroom 351
and Agariocus bisporus [109, 70]. Cheung (1997) determined hemicelluloses and pectic substances in 352
basidiomycetes [110]. Information on chitin and fibre changes during different preservation and cooking remedy has 353
been lacking. Great attention has recently been given to fiber content of the mushroom, the β-glucans which act as a 354
health-promoting factor [86]. A recent study reported that two mushroom carbohydrates inhibit breast cancer cell 355
growth by enhancing the immune function [111]. 356
MINERAL COMPOSITION 357
Wild mushrooms are assumed to accumulate a large amount of both macro and micro-minerals. Mineral content 358

14
varies among different species of basidiomycetes. A research on 11 different mushroom species showed the level of 359
potassium concentration in Tricholoma saponaceum to be around 39.8 mg/g, which is lower in case of Laccaria 360
laccata and Candida rugosa being 30.2 mg/g and 28.9 mg/g [112]. Selenium content in the tubes and gills of 361
mushrooms are quite high which act as an antioxidant. It is also required for the biosynthesis of selenoenzymes 362
which include glutathione peroxidase, thioredoxin reductases, Selenoprotein W and Selenoprotein P, iodothyronine 363
5’-deiodases. Selenium content in mushrooms ranges from approximately 5μg/g in Lycoperdon spp. to about 200μg/g 364
in Albatrellus pes-caprae. King Bolete(Boletus edulis) is considered to contain an average concentration of 365
approximately 20μg/g of selenium whereas Pinewood King Bolete(Boletus pinicola) contain selenium on an average 366
of 40μg/g based on its dry weight. The fruiting body of Lurid Bolete(Boletus luridus), Lepista luscina and Parasol 367
Mushroom (Macrolepiota procera) contained, respectively, 49, 91 and 47 of selenium in crude mushroom[113]. 368
Potassium is not distributed evenly within fruiting bodies, the concentration seems to be higher in cap followed by 369
stipe, spore forming part and finally to spores. It seems to be higher by 20 to 40 folds in the fruiting bodies than the 370
remaining part [114]. Followed by potassium, the second most abundant major element in mushroom fruiting bodies 371
is phosphorus which ranges from 5 to 10 g per kg of dry matter [19]. A report shows high amount of calcium 372
concentration of 1.600 mg/g in Craterellus tubaeformis followed by 10.50 mg/g in Laccaria Laccatta [112]. Sodium 373
concentration is relatively less in mushroom species due to which they have been considered good for patient with 374
hypertension which ranges from 100 to 400 mg per kg of dry matter [19]. Few mushrooms including Formitopsis 375
officinalis offer chlorine in the form of coumarine which include 6-chloro-4-phenyl-2H-chromen-2-one and ethyl 6-376
chloro-2-oxo-4-phenyl-2H-chromen-3-carboxylate [115]. As per the report of Rudawska and Leski (2005), total 377
sulphur content per kg of dry matter in Amanita rubescens and Xerocomus chrysenteron is found to be in between 378
900 and 4400 mg per kg dry matter respectively [116]. Mushrooms are considered to contain high amount of trace 379
elements including cadmium, mercury, lead, copper, antimony in their fruiting bodies [117]. Agaricus spp. 380
accumulates a high level of mercury and cadmium upto a range of 130 mg of cadmium per kg of dry weight of 381
fruiting body [118]. Other mercury accumulators include Lycoperdon perlatum, Lepista spp. and Macrolepiota spp. 382
[118]. Chromium and nickel concentration is higher in Lactarius deliciosus with around 4.51 mg per kg of dry 383
weight and 9.9 mg per kg of dry weight respectively in Tricholoma terreum [119]. Mushrooms are also known as 384
zinc accumulator. Lactarius spp. is known for its higher concentration of zinc accumulation which ranges from 385
55.7mg/kg to 158mg/kg on the basis of its dry weight. The gills of Clitocybe alexandri are considered to contain high 386

15
amount of copper which ranges from 26.3 to 95.9mg/kg on its dry weight basis. The concentration of iron seems to 387
be slightly higher in Volvariella speciosa which ranges from 220 to 7162 mg/kg [120]. 388
During Chernobyl accident, mushrooms fruiting bodies (radiotrophic fungi) were seen to accumulate huge amount of 389
toxic heavy metal [121]. It has been reported that natural radionuclide 40
K accumulates highly in mushroom and 390
accumulation factor ranges from 20 to 40 [114,122]. However, Boletus group are considered to contain high amount 391
of selenium content [19]. Heavy metal radiocaesium are also found to a greater extent in mushrooms of coniferous 392
forest then deciduous forest [114, 123]. 393
CONCLUSION 394
Various wild macrofungi are traditionally used as a delicacy and source of medicine since time immemorial. 395
However, large sections of their population are still unknown due to fragmentary and poor information about their 396
bioavailability and toxicity. Furthermore, the use of morphological and molecular identification based on ITS 397
sequences can to some extent be used in terms of their ecological perspective. 398
n-3 fatty acid concentration is low in mushroom which contributes negligibly in its nutritional profile but high 399
proportion of insoluble fibre encompassing chitin, β-glucan and other polysaccharides seems to be highly valuable 400
both in nutritional and pharmacological aspects. With the high concentration of unsaturated fatty acid, they prove to 401
be beneficial for basal metabolism in human. The concentration of potassium and selenium is high in the tubes and 402
gills of mushrooms for which they can be used as a source of antioxidant. Few fruiting bodies can accumulate quite a 403
large concentration of trace elements and radioactive components which make them vulnerable to consume. However, 404
a large section of poisonous mushrooms resemble their non-toxic counterparts. So, a need of a barcode to 405
differentiate their toxic nature will further develop their biological and therapeutic application and open vistas for 406
their bio-prospection and use for human benefits. 407
LITERATURE CITED 408
1. Bhattacharya PN, Baruah C, Jha DK, Sharma DK. Potential Application of Microbial Resources in North East 409
India: Future Prospects and Challenges. NeBIO 2011; 2(3):12-18. 410
2. Myers N, Mittermeier RA, Mittermerier CG. Biodiversity hotspots for conservation priorities. Nature 2000; 403: 411
853–858. 412
3. Smith D. Culture collections over the World. Int Microbiol 2003; 6: 95-100. 413
4. Khoshoo TN. Census of India's biodiversity: Tasks ahead. Curr Sci 1995; 69: 14-17. 414

16
5. Blackwell M. The Fungi: 1, 2, 3 … 5.1 Million Species? Am J Bot 2011; 98(3): 426–438. 415
6. Watling R. Pulling the threads together: habitat diversity. Biodivers Conserv1997; 6: 753-763. 416
7. Christensen M. A view of fungal ecology. Mycologia 1989; 81(1): 1-19. 417
8. Hawksworth DL.The magnitude of fungal diversity: the 1.5 million species estimate revisited. Mycol Res 2001; 418
105 (12): 1422- 1432. 419
9. Kirk P, Cannon PF, Minter DW, Stalpers JA. Ainsworth & Bisby’s Dictionary of the Fungi. 10th edn. CAB 420
International,Wallingford, UK; 2008. 421
10. Raghukumar C. Marine fungal biotechnology: an ecological perspective. Fungal Divers 2008; 31: 19-35. 422
11. Ahmed M, Abdel-Azeem. The history, fungal biodiversity, conservation, and future perspectives for mycology in 423
Egypt. IMA Fungus 2010; 1(2):123-142. 424
12. Aneja KR. Experiments in Microbiology, Plant pathology and Biotechnology. 4th Edn., New Age International 425
Pvt. Ltd., India; 2005. 426
13. Punt PJ, Biezen NV, Conesa A, Albers A, Mangnus J, Hondel C. Filamentous fungi as cell factories for 427
heterologous protein production. Trends Biotechnol 2002; 20: 200-206. 428
14. Xu X, Yan H, Chen J, Zhang X. Bioactive proteins from mushrooms. Biotechnol Adv 2011; 29: 667–674. 429
15. Wani B.A, Bodha RH, Wani AH. Nutritional and medicinal importance of mushrooms. J Med Plants Res 2010; 430
4(24):2598-2604. 431
16. Chang ST, Miles PG. Mushrooms: cultivation, nutritional value, medicinal effect and environmental impact. CRC 432
Press; 1989. p. 480. 433
17. Oyetayo OV. Medicinal uses of mushrooms in Nigeria: towards full and sustainable exploitation. Afr J. Tradit. 434
Complement. Altern. Med 2011; 8(3): 267-274. 435
18. Fan L, Pan H, Soccol AT, Pandey A, Soccol CR. Advances in Mushroom research in the last decade. Food 436
Technol. Biotechno 2006; l 44(3):303-311. 437
19. Kalac P. Chemical composition and nutritional value of European species of wild growing mushrooms: A review. 438
Food Chem 2009; 113: 9-16. 439
20. Kredics L, Jimenez LG, Naeimi S, Czifra D, Urban P, Manczinger L, Vagvolgyi C, Hatvani L. A challenge to 440
mushroom growers: the green mould disease of cultivated champignons. Current research, technology and education 441
topics in applied microbiology and microbial biotechnology. A Mendez- Vilas (Ed.); 2010. p. 295-305. 442

17
21. Jeeva S, Mishra BP, Venugopal N, Laloo RC. Sacred forests: Traditional ecological heritage in Meghalaya. J. 443
Scott Research Forum 1 2005; 93-97. 444
22. Somashekar BS. Treasure house in trouble. Amruth 1998; 2: 3-7. 445
23. Ramanujam MP, Kadamban D. Plant biodiversity of two tropical dry evergreen forests in the Pondicherry region 446
of south India and the role of belief systems in their conservation. Biodiversity Conserv 2001; 10: 1203-1217. 447
24. Agrahar-Murugkar D, Pal PP. Intake of nutrients and food sources of nutrients among the Khasi tribal women of 448
India. Nutrition 2004; 20(3): 268-273. 449
25. Khaund P, Joshi SR. Wild edible macrofungal species consumed by the Khasi tribe of Meghalaya, India. IJNPR. 450
2013; 4(2): 197-204. 451
26. Nicholson MS. Some spiritualistic uses of Mushrooms. Fungi 2009; 2(2): 26-27. 452
27. Burk W. Puffball usages among North American Indians. J. Ethnobiol 1983; 3(1):55–62. 453
28. Kumari B, Atri NS, Upadhyay RC. Culinary status and sociobiology of Termitophilous and Lepiotoid 454
mushrooms of North West India. World J Agricult Sci 2012; 8(4): 415-420. 455
29. Kues U. Life history and development processes in the basidiomycetes Coprinus cinereus. Microbiol Mol Biol R 456
2000; 64(2): 316-353. 457
30. Chang ST, Hayes W. The Biology and Cultivation of Edible Mushrooms. Academic Press. New York; 1978.p. 458
230. 459
31. Lai E, Birren BW, Clark SM, Simon MI, Hood L. Pulsed field gel electrophoresis. BioTechniques 1989; 7: 34-42. 460
32. Sagawa I, Nagata Y. Analysis of chromosomal DNA of mushrooms in genus Pleurotus by pulsed field gel 461
electrophoresis. J. Gen. Appl. Microbiol 1992; 38:47–52. 462
33. Zakharov IA. Intratetrad mating and its genetic and evolutionary consequences. Russ J Genet+ 2005; 41(4): 402-463
411. 464
34. Larraya L, Penas MM, Pe rez G, Santos C, Ritter E, Pisabarro AG, Ramı´rez. L. Identification of incompatibility 465
alleles and characterisation of molecular markers genetically linked to the A incompatibility locus in the white rot 466
fungus Pleurotus ostreatus. Curr Genet 1999; 34:486–493. 467
35. Waters H, Moore D, Butler RD. Morphogenesis of aerial sclerotia of Coprinus lagopus. New Phyto 1975; 74: 468
207–213. 469

18
36. Moore D. Developmental genetics of Coprinus cinereus: genetic evidence that carpophores and sclerotia share a 470
common pathway of initiation. Curr Genet 1981; 3:145–150. 471
37. Kues U, Granado J D, Hermann R, Boulianne RP, Kertesz-Chaloupkova K, Aebi M. The A mating type and blue 472
light regulate all known differentiation processes in the basidiomycete Coprinus cinereus. Mol Gen Genet 1998; 260: 473
81–91. 474
38. Summerbell RC, Castle AJ, Horgen PA, Anderson JB. Inheritance of restriction fragment length polymorphisms 475
in Agaricus brunnescens. Genetics1989; 123: 293-300. 476
39. Idnurm A. A tertrad analysis of the basidiomycetes fungus Cryptococcus neoformans. Genetics 2010; 185: 153-477
163. 478
40. Kwon-Chung KJ, Edman JC, Wickes BL. Genetic association of mating types and virulence in Cryptococcus 479
neoformans. Infect Immun 1992; 60: 602–605. 480
41. Kwon-Chung KJ. Nuclear genotypes of spore chains in Filobasidiella neoformans (Cryptococcus neoformans). 481
Mycologia 1980; 72: 418 – 422. 482
42. Madelin MF. The influence of light and temperature on fruiting of Coprinus lagopus Fr. in pure culture. Ann Bot 483
1956; 20:467–480. 484
43. Lu BC. Meiosis in Coprinus V. The role of light on basidiocarp initiation, mitosis and hymenium differentiation 485
in Coprinus lagopus. Can J Bot 1974; 52: 299–305. 486
44. Morimoto N, Suda S, Sagara N. Effect of ammonia on fruitbody induction of Coprinus cinereus in darkness. 487
Plant Cell Physiol 1981; 22: 247–254. 488
45. Chaloupkova K, Walser KPJ, Granado JD, Aebi M, Kues U. Blue light overrides repression of asexual 489
sporulation by mating type genes in the basidiomycete Coprinus cinereus. Fungal Genet Biol 1998; 23: 95–109. 490
46. Kues U, Granado JD, Kertesz-Chaloupkova K, Walser PJ, Hollenstein M, Polak E, Liu Y, Boulianne RP, Bottoli 491
APF, Aebi M.Mating types and light are major regulators of development in Coprinus cinereus. In L. J. L. D. Van 492
Griensven and J. Visser (ed.), Proceedings of the Fourth Meeting on the Genetics and Cellular Biology of 493
Basidiomycetes. Mushroom Experimental Station, Horst, The Netherlands; 1998. p. 113–118. 494
47. Tsusue YM. Experimental control of fruitbody formation in Coprinus macrorhizus. Dev Growth Differ 1969; 495
11:164–178. 496

19
48. Morimoto N, Oda Y. Effects of light on fruitbody formation in a basidiomycete, Coprinus macrorhizus. Plant Cell 497
Physiol 1973; 14: 217–225. 498
49. Morimoto N, Oda V. Photo-induced karyogamy in a basidiomycete, Coprinus macrorhizus. Plant Cell Physiol 499
1974; 15:183–186. 500
50. Uno I, Yamaguchi M, Ishikawa T. The effect of light on fruiting body formation and adenosine 3’, 5’ cyclic 501
monophosphate metabolism in Coprinus macrorhizus. Proc Natl Acad Sci 1974; 71: 479–483. 502
51. Lu BC, Jeng DY. Meiosis in Coprinus. VII. The pre - karyogamy S-phase and the post karyogamy DNA 503
replication in C. lagopus. J Cell Sci 1975; 17: 461– 470. 504
52. Kamada T, Kurita R, Takemaru T. Effects of light on basidiocarp maturation in Coprinus macrorhizus. Plant Cell 505
Physiol 1978; 19: 263–275. 506
53. Ballou LR, Holton RW. Synchronous initiation and sporulation of fruitbodies by Coprinus cinereus on a defined 507
medium. Mycologia 1985; 77:103–108. 508
54. Matthews TR, Niederpruem DJ. Differentiation in Coprinus lagopus. I. Control of fruiting and cytology of initial 509
events. Arch Mikrobiol 1972; 87: 257–268. 510
55. Uno I, Ishikawa T. Effect of glucose on the fruiting body formation and adenosine 39, 59-cyclic monophosphate 511
levels in Coprinus macrorhizus. J Bacteriol 1974; 120: 96–100. 512
56. Uno I, Ishikawa T. Subcellular localization of adenylate cyclase during the fruiting body formation in Coprinus 513
macrorhizus. J Gen Appl Microbiol 1975; 21: 335–343. 514
57. Nyunoya H, Ishikawa T. Effect of 2-deoxy-D-glucose on the induction of tyrosinase in Coprinus marorhizus. J 515
Gen Microbiol 1980; 26: 229– 238. 516
58. Uno I, Nyunoya H, Ishikawa T. Effects of 2-deoxy-D-glucose and quinidine on the fruiting body formation in 517
Coprinus macrorhizus. J Gen Appl Microbiol 1981; 27: 219–228. 518
59. Zuber A, Kowalczyk M, Sekula A, Mleczko P, Kupiec T. Methods used in species identification of hallucinogenic 519
and other poisonous mushrooms in forensic investigations. Probl Forensic Sci 2011; 86: 151–161. 520
60. Epis S, Matinato C, Gentili G, Varotto F, Bandi C, Sassera D. Molecular detection of poisonous mushrooms in 521
different matrices. Mycologia 2010; 102(3): 747-754. 522
61. Botany.hawaii.edu. http://www. botany. Hawaii.edu. 523

20
62. Ammirati JF, Traquair JA, Horgen PA. Poisonous Mushrooms of the Northern United States and Canada, 524
University of Minnesota Press, Minneapolis; 1985. 525
63. Orijel RG, Terrazo AR, Velazquez MO. Women care about local knowledge, experiences from ethnomycology. J 526
Ethnobiol Ethnomed 2012; 8(25): 1-12. 527
64. Lee JC, Cole M, Linacre A. Identification of members of genera Panaeolus and Psilocybe by a DNA test. A 528
preliminary test for hallucinogenic fungi. Forensic Sci Int 2000; 112: 123–133. 529
65. Hallen HE, Luo H, Scott-Craig, JS, Walton JD. Gene family encoding the major toxins of lethal Amanita 530
mushrooms. Proc Natl Acad Sci 2007; 104(48): 19097-19101. 531
66. Qing CA, Li-Ping T, Zhu-Liang, Y. DNA Barcoding of Economically Important Mushrooms: A Case Study on 532
Lethal Amanitas from China. PDR 2012; 34(6): 614-622. 533
67. Frezel L, Leblois R. Four years of DNA barcoding: Current advances and prospects. Infect Genet Evol 2008; 534
8(5): 727-736. 535
68. Zuber A, Kowalczyk M, Sekula A, Mleczko P, Kupiec T. Methods used in species identification of hallucinogenic 536
and other poisonous mushrooms in forensic investigations. Probl Forensic Sci 2011; 86: 151–161. 537
69. Khaund P, Joshi SR. DNA barcoding of wild edible mushrooms consumed by the ethnic tribes of India.Gene 538
2014; 550(1): 123-130. 539
70. Cheung PCK. Mini-review on edible mushrooms as source of dietary fiber: Preparation and health benefits. Food 540
Sci Hum Wellness 2013; 22: 1-5. 541
71. Breene WM. Nutritional and medicinal value of specialty mushrooms. J Food Protect 1990; 53: 883- 894. 542
72. Ng TB. A review of research on the protein bound polysaccharide (Polysaccharopeptide, PSP) from the 543
mushroom Coriolus versicolor (Basidiomycetes: Polyporaceae). Gen Pharmac 1998; 30(1): 1-4. 544
73. Ribeiro LR, Salvadori DMF. Dietary components may prevent mutation-related diseases in humans. Mutat Res 545
2003; 544: 195-201. 546
74. Manzi P, Marconi S, Aguzzi A, Pizzoferrato L. Commercial mushrooms: nutritional quality and effect of cooking. 547
Food Chem 2004; 84: 201-206. 548
75. Crisan EW, Sand. A Nutritional value. In: Chang ST and Hayes WA (Eds.). The Biology and Cultivation of 549
Edible Mushrooms, Academic press, New York; 1978. p. 172-189. 550

21
76. Orgundana SK, Fagade O. The nutritive value of some Nigerian edible mushrooms. In: Mushroom Science XI, 551
Proceedings of the Eleventh International Scientific Congress on the Cultivation of Edible Fungi, Australia; 1981.p. 552
123-131. 553
77. Chang ST. Future trends in cultivation of alternative mushrooms. Mushroom J 1990; 215:422- 423. 554
78. Kakon AJ, Choudhury Md BK, Saha S. Mushroom is an ideal food supplement. J Dhaka National Med Cell Hos 555
2012; 18(1): 58-62. 556
79. Garch HS, Khanna PK, Soni GL. Nutritional importance of mushrooms. In Mushroom Biology and Mushroom 557
Products, Proceedings of the First International Congress, eds Chang ST, Buswell JA, Chiu SW. The Chinese 558
University Press, Hong Kong; 1993. p. 227-236. 559
80. Atri NS, Sharma SK, Joshi R, Gulati A, Gulati A. Amino acid composition of five wild Pleurotus species chosen 560
from North West India. Eur J Biol Sci 2012; 4 (1): 31-34. 561
81. Rose WC. The nutritive significance of the amino acids and certain related compounds. Science 1937; 86: 298-562
300. 563
82. Wang XM, Zhang J, Wu LH, Zhao YL, Li T, Li JQ, Wang YZ, Liu HG. A mini- review of chemical composition 564
and nutritional value of edible wild-grown mushroom from China. Food Chem 2014; 151:279-285. 565
83. Bauer Petrovska B. Protein fraction in edible Macedonian mushrooms. Eur Food Res Technol 2001; 212: 469-566
472. 567
84. Ferreira ICFR, Barros L, Abreu RM. Antioxidants in wild mushrooms. Curr Med Chem 2009; 16: 1543–1560. 568
85. Quang DN, Hashimoto T, Asakawa Y. Inedible mushrooms: A good source of biologically active substances. 569
Chem Rec 2005; 6: 79-99. 570
86. Wasser SP. Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl 571
Microbiol Biot 2002; 60: 258–274. 572
87. Cioci G, Mitchell EP, Chazalet V, Debray H, Oscarson S, Lahmann M, Gautier C, Breton C, Perez S, Imberty A 573
Beta-propeller crystal structure of Psathyrella velutina lectin: an integrin-like fungal protein interacting with 574
monosaccharides and calcium. J Mol Biol 2006; 357(5):1575-1591. 575
88. Wong JH, Wang HX, Ng TB. Marmorin, a new ribosome inactivating protein with antiproliferative and HIV-1 576
reverse transcriptase inhibitory activities from the mushroom Hypsizigus marmoreus. Appl Microbiol Biotechnol 577
2008; 81(4): 669–674. 578

22
89. Zhang GQ, Wang YF, Zhang XQ, Ng TB, Wang HX. Purification and characterization of a novel laccase from the 579
edible mushroom Clitocybe maxima. Process Biochem 2010b; 45(5): 627–633. 580
90. Wang HX, Ng TB. Ganodermin, an antifungal protein from fruiting bodies of the medicinal mushroom 581
Ganoderma lucidum. Peptides 2006b; 27(1): 27–30. 582
91. Du M, Zhao L, Li CR, Zhao GH, Hu XS. Purification and characterization of a novel fungi Se-containing protein 583
from Se-enriched Ganoderma Lucidum mushroom and its Se dependent radical scavenging activity. Eur Food Res 584
Technol 2007; 224(5): 659–665. 585
92. Jeurink PV, Noguera CL, Savelkoul HFJ, Wichers HJ. Immunomodulatory capacity of fungal proteins on the 586
cytokine production of human peripheral blood mononuclear cells. Int Immunopharmacol 2008; 8(8):1124–1133. 587
93. Maiti S, Bhutia SK, Mallick SK, Kumar A, Khadgi N, Maiti TK. Antiproliferative and immunostimulatory 588
protein fraction from edible mushrooms. Environ Toxicol Pharmacol 2008; 26(2):187–191 589
94. Chen JN, Wang YT, Wu JSB. A glycoprotein extracted from golden oyster mushroom Pleurotus citrinopileatus 590
exhibiting growth inhibitory effect against U937 leukemia cells. J Agricult Food Chem 2009; 57:6706–6711. 591
95. Lin YL, Liang YC, Tseng YS, Huang HY, Chou SY, Hseu RS, et al. An immunomodulatory protein, Ling Zhi-8, 592
induced activation and maturation of human monocytederived dendritic cells by the NF-kappa B and MAPK 593
pathways. J Leukoc Biol 2009; 86(4):877–89. 594
96. Sheu F, Chien PJ, Hsieh KY, Chin KL, Huang WT, Tsao CY, et al. Purification, cloning, and functional 595
characterization of a novel immunomodulatory protein from Antrodia camphorata (Bitter Mushroom) that exhibits 596
TLR2-dependent NF-kappa B activation and M1 polarization within murine macrophages. J Agric Food Chem 2009; 597
57(10): 4130–4141. 598
97. Kodama N, Mizuno S, Nanba H, Saito N. Potential antitumor activity of a low-molecular weight protein fraction 599
from Grifola frondosa through enhancement of cytokine production. J Med Food 2010; 13(1):20 –30. 600
98. Guo YX, Wang HX, Ng TB. Isolation of trichogin, an antifungal protein from fresh fruiting bodies of the edible 601
mushroom Tricholoma giganteum. Peptides2005; 26(4):575–580. 602
99. Chang HH, Chien PJ, Tong MH, Sheu F. Mushroom immunomodulatory proteins possess potential 603
thermal/freezing resistance, acid/alkali tolerance and dehydration stability. Food Chem 2007; 105(2):597–605. 604

23
100. Horie K, Rakwal R, Hirano M, Shibato J, Nam HW, Kim YS, KouzumaY, Agarwal GN, Masuo Y, Yonekura M. 605
Proteomics of two cultivated mushrooms Sparassia crispa and Hericium erinaceum provides insights into their 606
numerous functional protein components and diversity. J Proteome Res 2008; 7: 1819-1835. 607
101. Hanus LM, Shkrob I, Dembitsky VM. Lipids and fatty acids of wild edible mushrooms of the genus Boletus. J 608
Food Lipids 2008; 15: 370-383. 609
102. Pedneault K, Angers P, Gosselin A, Tweddell RJ. Fatty acid composition of lipids from mushrooms belonging to 610
the family Boletaceae. Mycol Res2006; 110: 1179–1183. 611
103. Weete JD. Fungal lipid biochemistry: Distribution and Metabolism. Monographs in Lipid Research, v. 612
1.Kritchevshy D. (Series Ed.), New York: Plenum Press; 1974. 613
104. Davidoff F, Kom ED. Fatty acid and phospholipid composition of the cellular slime mold, Dictyostelium 614
discoideum: the occurrence of previously undescribed fatty acids. J Biol Chem 1963; 238: 3199-3209. 615
105. Pfeuffer M, Schrezenmeir J. Impact of trans fatty acid of ruminant origin compared with those from partially 616
hydrogenated vegetable oils on CHD risk. Int Dairy J 2006; 16 (11): 1383-1388. 617
106. Stamets P. Mycelium Running: How mushroom can save the World. Ten Speed Press, Crown Publishing Group, 618
Random House Digital, Inc., New York, USA; 2005. 619
107. Patyshakuliyeva A, Jurak E, Kohler A, Baker A, Battaglia E, de Bruijn W, Burton KS et al. Carbohydrate 620
utilization and metabolism is highly differentiated in Agaricus bisporus. BMC Genomics 2013; 14: 663-676. 621
108. Barros L, Morais JS, Ferreira ICFR. Effects of conservation treatment and cooking on the chemical composition 622
and antioxidant activity of Portuguese wild edible mushrooms. J Agricult Food Chem 2007b; 55: 4781–4788. 623
109. Rangel-Castro JI, Danell E, Pfeffer PE. A 13
C-NMR study of exudation and storage of carbohydrates and amino 624
acids in the ectomycorrhizal edible mushroom Cantharellus cibarius. Mycologia 2002; 94(2): 190-199. 625
110. Cheung PCK. Dietary fibre content and composition of some edible fungi determined by two methods of 626
analysis. J Sci Food and Agr 1997; 72: 255–260. 627
111. Jeong SC, Koyyalamudi SR, Jeong YT, Song CH, Pang G. Macrophage immunomodulating and antitumor 628
activities of polysaccharides isolated from Agaricus bisporus white button mushrooms. J Med Food 2012; 15: 58-65. 629
112. Ayaz FA, Torun H, Colak A, Sesli E, Millson M, Glew RH. Macro and microelement contents of fruiting bodies 630
of wild- edible mushrooms growing in the East Black Sea region of Turkey. Food Nutr Sci 2011; 2: 53-59. 631
113. Falandysz J. Selenium in edible mushrooms. J Environ Sci Health C 2008; 26: 256-299. 632

24
114. Seeger R.Content of potassium in higher fungi. Zeitschrift fur Lebensmittel Untersuchung und Forschung 1978; 633
167: 23–31 [in German]. 634
115. Hwang CH, Jaki BU, Klein LL, Lankin DC, McAlpine JB, Napolitano JG, Fryling NA, Franzblau SG, Cho SH, 635
Stamets PE, Wang Y, Pauli GF. Chlorinate coumarins from the Polypore Mushroom Fomitopsis officinalis and their 636
activity against Mycobacterium tuberculosis. J Nat Prod 2013; 76 (10): 1916-1922. 637
116. Rudawska M, Leski T. Macro- and microelement contents in fruiting bodies of wild mushrooms from Notecka 638
forest in west-central Poland. Food Chem 2005; 92 : 499–506. 639
117. Vetter J.“Arsenic Content of Some Edible Mushroom Species”. Eur Food Res Technol 2004; 219: 71-74. 640
118. Lodenius M, Herranen M. Influence of a chloralkali plant on the mercury contents of fungi. Chemosphere 1981; 641
10: 313–318. 642
119. Vetter J, Berta E. Mercury content of some wild edible mushrooms. Food Res Techn 1997; 205: 316-320. 643
120. Vetter J. Chromium and nickel content of some common edible mushroom species Acta Alimentaria 1997; 26: 644
163-170. 645
121. Isiloglu M, Yilmaz F, Merdivan M. Concentration of trace elements in wild edible mushrooms. Food Chem 646
2009; 73: 169-175. 647
122. Khaund P, Joshi SR. Micromorphological characterization of wild edible mushroom spores using Scanning 648
Electron Microscopy. Natl Acad Sci Lett 2014; 37(6):521-527. 649
123. Nikkarinen M, Mertanen E. Impact of geological origin on trace element composition of edible mushrooms. J 650
Food Comp Anal 2004; 17: 301-310. 651
124. Mycoprotein. org, http://www. mycoprotein.org. 652
LEGENDS TO FIGURES 653
Fig I Wild macrofungi : (MS1)Formitopsis spp., (MS2)Russula spp., (MS3) Mycena spp., (MS4)Hypholoma spp., 654
(MS5)Trichaptum spp., (MS6)Panus spp., (MS7)Galerina spp., (MS8)Lactarius spp. 655
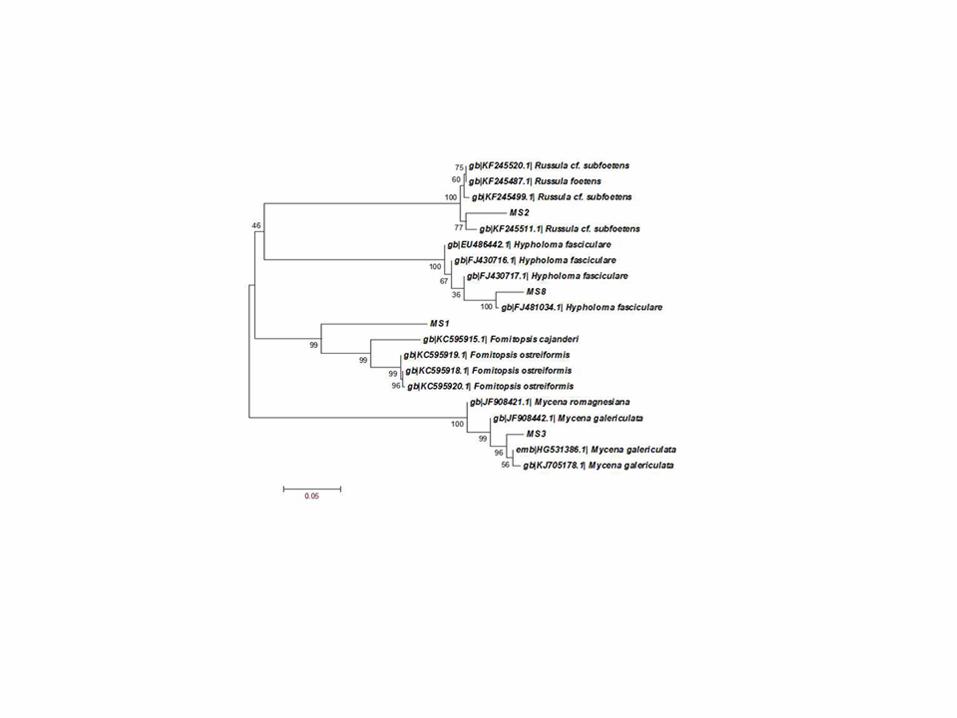
Fig II Phylogenetic tree generated for edible macrofungi 656
Fig III Phylogenetic tree generated of inedible macrofungi 657

25

TABLE I. Essential amino acid per 100 grams of proteins of mushroom [82, 83, 124]
Amino acid Mycoprotein (g) Egg (g) Milk (g) Wheat (g)
Arginine....................... 6.7 6.4 - -
Histidine...................... 0.35- 2.1 2.1 0.09 0.32
Lysine.......................... 0.83- 5.0 7.2 0.26 0.30
Tryptophan.................. 0.16- 0.9 1.5 0.05 0.18
Phenylalanine.............. 0.49- 2.0 6.3 0.16 0.68
Methionine.................. 0.21- 1.26 4.1 0.08 0.22
Threonine.................... 0.55- 4.2 4.9 0.15 0.37
Leucine....................... 0.86- 4.4 9.2 0.32 0.93
Isoleucine................... 0.52- 5.8 8.0 0.20 0.53
Valine.......................... 0.62- 4.65 7.3 0.22 0.59
TABLE II. Nutritional composition in mushroom fruiting body g/100g (% of dry matter)
Species Crude protein Lipid Carbohydrates Fibre Reference
Agaricus bisporus 26.3- 56.3 1.8-2.7 37.5-59.96 - [50, 51]
Agaricus arvensis 56.3 2.7 37.5 - [50]
Amanita rubescens 26-31.9 7.2 - 27.5 30.6-62.2 - [50, 84, 85]
Armillariella mellea 21.9 1.8-6.08 70 - [50, 84]
Auricularia polytrica 7.7 0.8 87.6 - [51]
B. Aereus 26.9 2.1 34 17.0 [86]
Boletus edulis 26.5-28.7 2.8-4.1 30.6- 65.4 15.3 [50, 86]
B. speciosus 28.1 2.9 28.6 21.0 [86]
Cantharellus cibarius 34.17- 53.7 1.40- 2.9 31.9-57 - [50, 84]
C. aureus 14.1 4.0 61.5 5.2 [86]
C. Cornucopioides 50.10 5.89 34 - [84]

Flaninnilbia velutipes 17.6 1.9 73.1 - [51]
H. repandum 34.14 8.80 55 - [84]
Lactarius deliciosus 20.2- 29.8 2.2-4.0 5.1-25 36.8 [50, 51]
Lactarius hatsudake 15.3 1.0 38.2 31.8 [51]
Lactarius volemus 17.6- 87.57 3.98-6.7 15- 64 40.0 [84, 51]
Lentinula edodes 17.1 1.9 30.2 39.4 [51]
Lentinula modes 17.5 8.0 67.5 - [86]
Lepista nuda 19.8-59.4 1.8-9 20.3-71.0 - [50]
Lycoperdon perlatum 17.2- 44.93 0.4 -10.58 42- 50.4 - [50, 84]
L. Crocipodium 29.3 1.0 12.8 37.9 [86]
Macrolepiota procera 23.9 2.3 68.4 - [50]
Pleurotus ostreatus 10.5 1.6 81.8 - [51]
Ramaria botrytis 39.0 1.4 50.8 - [50]
R. Flava 35.55 5.20 65 - [84]
R. virescens 28.3 1.5 13.4 32.8 [86]
S. Imbricatus 27.45 8.85 57 - [84]
Suillus granulatus 16.5 4.0 74.3 - [50]
S. aspratus 12.0 2.8 64.6 5.1 [86]
Tricholoma flavovirens 18.1 2.0 37.0 - [50]
Tricholoma portentosum 19.6-30.5 5.5-5.8 34.6 30.1 [87]
Tricholoma terreum 20.1 6.6 31.1 30.1 [87]
Trcinclla fuciforinis 4.6 0.2 94.8 - [51]
T. Matsutake 14.3 5.0 36.7 29.1 [86]
Volvariella volvacea 29.3 5.7 60.0 - [51]
TABLE III. Accumulation of various trace elements by the mushroom fruiting bodies [78, 79, 88, 89, 90]
Element Content Accumulators
Antimony 0.05-0.15 Suillus sp., Laccaria amethystina, Amanita rubescens
Arsenic < 1 Laccaria amethystina, L. Laccata, Agaricus sp., Lepista nuda
Lycoperdon perlatum
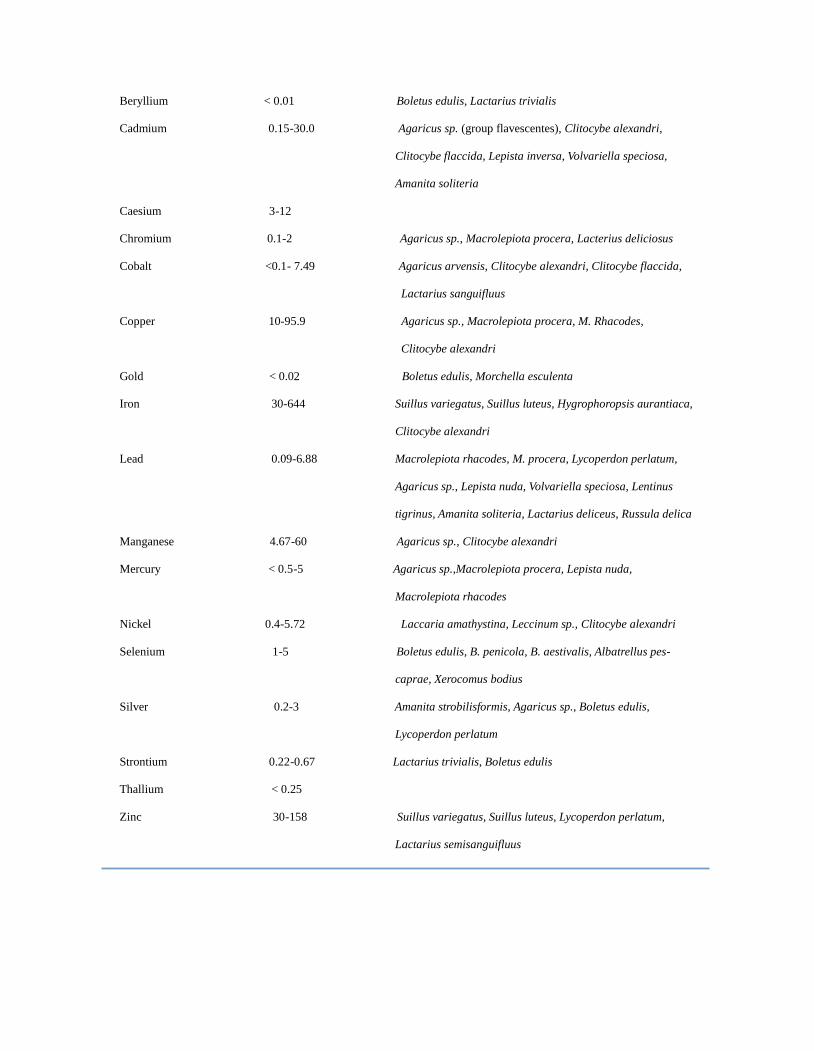
Beryllium < 0.01 Boletus edulis, Lactarius trivialis
Cadmium 0.15-30.0 Agaricus sp. (group flavescentes), Clitocybe alexandri,
Clitocybe flaccida, Lepista inversa, Volvariella speciosa,
Amanita soliteria
Caesium 3-12
Chromium 0.1-2 Agaricus sp., Macrolepiota procera, Lacterius deliciosus
Cobalt <0.1- 7.49 Agaricus arvensis, Clitocybe alexandri, Clitocybe flaccida,
Lactarius sanguifluus
Copper 10-95.9 Agaricus sp., Macrolepiota procera, M. Rhacodes,
Clitocybe alexandri
Gold < 0.02 Boletus edulis, Morchella esculenta
Iron 30-644 Suillus variegatus, Suillus luteus, Hygrophoropsis aurantiaca,
Clitocybe alexandri
Lead 0.09-6.88 Macrolepiota rhacodes, M. procera, Lycoperdon perlatum,
Agaricus sp., Lepista nuda, Volvariella speciosa, Lentinus
tigrinus, Amanita soliteria, Lactarius deliceus, Russula delica
Manganese 4.67-60 Agaricus sp., Clitocybe alexandri
Mercury < 0.5-5 Agaricus sp.,Macrolepiota procera, Lepista nuda,
Macrolepiota rhacodes
Nickel 0.4-5.72 Laccaria amathystina, Leccinum sp., Clitocybe alexandri
Selenium 1-5 Boletus edulis, B. penicola, B. aestivalis, Albatrellus pes-
caprae, Xerocomus bodius
Silver 0.2-3 Amanita strobilisformis, Agaricus sp., Boletus edulis,
Lycoperdon perlatum
Strontium 0.22-0.67 Lactarius trivialis, Boletus edulis
Thallium < 0.25
Zinc 30-158 Suillus variegatus, Suillus luteus, Lycoperdon perlatum,
Lactarius semisanguifluus