1071155923398 lehane histamine scombroid report
TRANSCRIPT

Histamine (Scombroid) FishPoisoning
a review in a risk-assessment framework
Leigh Lehane and June Olley
National Office of Animal and Plant HealthCanberra 1999

Histamine Fish Poisoning
ii
© Commonwealth of Australia 1999
This work is copyright. It may be reproduced in whole or in part subject to the inclusion of anacknowledgment of the source and no commercial usage or sale. Reproduction, for purposes other thanthose indicated above, requires prior written permission from the Commonwealth available fromAusInfo. Requests and inquiries concerning reproduction and rights should be addressed to theManager, Legislative Services, AusInfo, GPO Box 1920, Canberra, ACT 2601.
Note:
To ensure timely publication, this report has not been subjected to either formal external peer review orprofessional editing. It was prepared for publication by the Animal Health Science and EmergencyManagement Branch, of the National Office of Animal and Plant Health, Department of Agriculture,Fisheries and Forestry, Australia.
Revised 4/5/2000
Published by:
Animal Health Science and Emergency Management BranchNational Office of Animal and Plant HealthAgriculture, Fisheries and Forestry — AustraliaGPO Box 858Canberra ACT 2601
Preferred way to cite this publication:
Lehane, L. and Olley, J* (1999) Histamine (Scombroid) Fish Poisoning: a review in a risk-assessmentframework. National Office of Animal and Plant Health, Canberra.
* Dr June Olley is an Honorary Research Associate in the School of Agricultural Science at theUniversity of Tasmania.
Acronyms used frequently in this report:
DAO, diamine oxidase or histaminaseDSP, diarrhetic shellfish poisonsHACCP, Hazard Analysis and Critical Control PointHAL, L-histidine ammonia lysase, or histidaseHD, histidine decarboxylaseHDB, histidine decarboxylating bacteriaHFP, histamine fish poisoningHMT, histamine methyl transferaseMAO, monoamine oxidasePSP, paralytic shellfish poisons

Histamine Fish Poisoning
iii
CONTENTS
SUMMARY 1Introduction 1Hazard identification 1Dose−response assessment 3Exposure assessment 4Risk characterisation 5
1. INTRODUCTION 7
2. HAZARD IDENTIFICATION 82.1 Definition of hazard identification 8 2.2 Production of histamine, other biogenic amines and imidazole compounds in spoiling fish 9 2.3 Metabolism of histidine and histamine in mammals 11 2.4 Possible mechanisms of toxicity 13 2.4.1 Toxicity of histamine and other biogenic amines 13 2.4.2 Inhibition of histamine detoxification by histamine potentiators 15 2.4.2.1 Background 15 2.4.2.2 In vitro studies 16 2.4.2.3 In vivo studies 16 2.4.3 Barrier disruption hypothesis 18 2.4.4 Release of endogenous (mast cell) histamine by scombroid toxin(s) 19 2.4.5 Are paralytic and diarrhetic shellfish poisons involved? 22 2.4.6 Absorption of histamine from mouth and throat 22 2.5 Clinical characteristics and treatment 23 2.5.1 Clinical signs and symptoms 23 2.5.2 Diagnosis 24 2.5.3 Treatment 25 2.5.4 Clinical complications 25 2.6 Detection of histamine and other biogenic amines in fish 25 2.6.1 Detection of histamine-producing bacteria 25 2.6.2 Analysis of histamine 26 2.6.3 Analysis of other biogenic amines, and chemical quality index 28 2.6.4 Analysis of urocanic acid 30
3. DOSE−RESPONSE ASSESSMENT 31 3.1 Definition of dose−response assessment 31 3.2 Incidence of histamine fish poisoning 31 3.2.1 Background 31 3.2.2 Africa 32 3.2.3 Asia 32 3.2.4 Australia 32 3.2.4.1 Outbreaks described by Smart (1992) 33 3.2.4.2 Cases described by Brown (1993) 34 3.2.5 Canada 34 3.2.6 Europe 34 3.2.7 New Zealand 36 3.2.8 United States 36 (Dose–Response Assessment continued over page)

Histamine Fish Poisoning
iv
3.3 Fish characteristics that affect the clinical response 37 3.3.1 Species 37 3.3.2 Intrinsic potential for histamine and cadaverine accumulation 38 3.3.3 Seasonal and other variability 39 3.3.4 Parts of fish consumed 40 3.3.5 Nature and amount of bacterial contamination in spoiling fish 40 3.3.5.1 The common histamine-producing bacteria 40 3.3.5.2 Time–temperature relationships and histamine production 41 3.3.5.3 Studies on variability in species/strain composition of microflora 43 3.3.5.4 Species/strain variability in decarboxylase activity 44 3.3.5.5 Bacterial destruction of histamine 46 3.3.6 Histamine levels and toxic dose 46 3.4 Human factors that affect the clinical response 48 3.4.1 Variation in individual susceptibility 48 3.4.2 Influence of diet 49 3.4.3 Influence of medication 49 3.4.4 Disease states and age 50 3.5 Morbidity and mortality rates 50
4. EXPOSURE ASSESSMENT 51 4.1 Definition of exposure assessment 51 4.2 Factors affecting the probability of histamine fish poisoning occurring 51 4.2.1 Post-catching contamination 51 4.2.2 Temperature abuse on fishing vessels 52 4.2.3 Inadequate chill-storage procedures 53 4.2.4 Inadequate freezing and thawing procedures 54 4.2.5 Temperature abuse in the preparation of dried and/or smoked products 55 4.2.6 Poor canning procedures 57 4.2.7 Low-quality fermented products 57 4.2.8 Temperature abuse of raw tuna for the sashimi market 574.3 Histamine levels in fish products in Australia 584.3.1 Survey of histamine in canned tuna 584.3.2 Survey of biogenic amines in fish products in the ACT 58 4.3.3 Monitoring by the Australian Government Analytical Laboratories 59 4.3.4 Monitoring by industry 594.4 Amounts and types of fish consumed, and at-risk population groups 594.5 Future exposure trends 60
5. RISK CHARACTERISATION 615.1 Definition of risk characterisation 615.2 Nature and magnitude of risk 625.2.1 Impact on human health 625.2.2 Impact on fishing industries 625.3 Uncertainties and problem areas in risk characterisation 625.3.1 Defining histamine fish poisoning and elucidating its pathogenesis 625.3.2 Investigating and managing post-harvesting contamination 63
REFERENCES 66
Appendix 1: Acknowledgments 80

Histamine Fish Poisoning
1
SUMMARY
Introduction
Histamine fish poisoning (HFP) is a foodborne chemical intoxication caused by theconsumption of spoiled, or bacterially contaminated, fish. Fish species associated withHFP are harmless when caught. They may still have a normal appearance and odourafter they have become toxic. Spoiled ’fresh’ fish, and frozen and smoked fish andcanned fish products have all caused the disease.
HFP occurs worldwide. Recent reports in the literature have suggested that it is asignificant public health and safety concern. Its true incidence has probably beenunderestimated, because of under-reporting and misdiagnosis owing to confusion withsymptoms of other illnesses, particularly food allergy.
Histamine, a physiological amine involved in allergic reactions, is the main toxininvolved in HFP, but HFP is not uncomplicated histamine poisoning. Although thedisease is generally associated with high levels of histamine (��� mg/100g) in spoiledfish, its pathogenesis has not been elucidated.
Scientific information on HFP is reviewed here in a risk-assessment framework inorder to arrive at an informed characterisation of risk. An attempt has been made toaddress, as accurately as possible, the questions: ’What causes outbreaks of HFP?’’What are the underlying factors contributing to outbreaks?’ and ’What are theconsequences of outbreaks?’
Hazard identification
The involvement of histamine as the main hazard in HFP is supported by: symptomsidentical to those of intravenous histamine administration or allergic reaction; theefficacy of antihistamine therapy; and the presence of increased levels of histamine inspoiled fish that cause the syndrome.
Histamine production in fish is related to the histidine content of the fish, the presenceof bacterial histidine decarboxylase (HD), and environmental conditions. Bacterialdecarboxylase enzymes acting on free histidine and other amino acids in the fishmuscle form histamine and other biogenic amines. Fish of the family Scombridae,notably tuna and mackerel, contain abundant amounts of histidine and are mostcommonly implicated. However, many species, both scombroid and non-scombroid(e.g. mahi-mahi, bluefish and sardines), have caused HFP so the term scombroid fishpoisoning is a misnomer.
The main bacteria responsible for histidine decarboxylation and HFP are members ofthe family Enterobacteriaceae. Specific bacteria present in the marine environment orintroduced during food handling produce HD, which converts histidine to histamine,particularly when fish are not kept chilled or frozen. Other bacteria in spoiling fishmuscle, again often members of the Enterobacteriaceae family, produce otherbreakdown products such as biogenic amines putrescine from ornithine andcadaverine from lysine.

Histamine Fish Poisoning
2
Histamine consumed in spoiled fish is more toxic than an equal amount of histaminetaken orally in an aqueous solution. It has been proposed that other biogenic aminesproduced in spoiled fish may potentiate the action of histamine by inhibiting theenzymes diamine oxidase (DAO, or histaminase) and histamine methyl transferase(HMT). These two enzymes normally detoxify histamine in the intestinal tract andprevent the absorption of unmetabolised histamine into the circulation. Putrescine andcadaverine have been identified as histamine potentiators. However, results ofexperiments to support the potentiator hypothesis are not totally convincing. Anotherpossible mechanism of potentiation of histamine by other biogenic amines is the’barrier disruption hypothesis’, which suggests that potentiators might interfere withthe protective actions of intestinal mucin, which binds histamine.
Still more hypotheses have been proposed in an attempt to explain the pathogenesis ofHFP. Some scientists have suggested that a scombroid toxin (or toxins) inscombrotoxic fish causes the release of endogenous histamine from mast cells, whichaugments the exogenous histamine consumed in spoiled fish in causing symptoms oftoxicity. Others have proposed that paralytic and/or diarrhetic shellfish poisons areresponsible. Yet another hypothesis, long discarded, is that histamine in a fishsubstrate may be better absorbed from the mouth or throat than pure histamine insolution.
Thus, the pathogenesis of HFP remains obscure. After sifting the evidence, the presentauthors believe that endogenous release of histamine from mast cells probably doesoccur in HFP, at least in some cases. We suggest that urocanic acid may be themissing factor (’scombroid toxin’) in HFP for the following reasons. In spoiling fish,histidine may be metabolised to histamine, or to urocanic acid and glutamate by analternative metabolic pathway. The first step in the alternative pathway is the loss ofammonia from histidine by the action of L-histidine ammonia lysase (HAL, orhistidase). HAL has a wide distribution among bacteria and, unlike HD, is also foundnaturally in fish muscle. Bacteria also produce urocanase, but HAL activity is greaterthan that of urocanase and urocanic acid accumulates with histamine in spoiling fish.Most importantly, urocanic acid has recently been recognised as a mast celldegranulator.
Histamine is catabolised by several routes in the human body. The main catabolicpathways involve the enzymes DAO and HMT. HMT activity in particular iswidespread in many body tissues and imidazole histamine metabolites are excreted inthe urine.
Histamine exerts its toxicity by interacting with histamine receptors (H1, H2 and H3)on cellular membranes. The most common symptoms of histamine poisoning arecardiovascular —flushing, urticaria, hypotension and headache. Other symptoms aregastrointestinal — abdominal cramps, diarrhoea and vomiting — and neurological —pain and itching associated with urticarial lesions. However, HFP is usually a milddisease of quick onset (several minutes) and short duration (about 8 h). It respondswell to antihistamine treatment.

Histamine Fish Poisoning
3
Numerous tests are available for detecting histidine decarboxylating bacteria (HDB)and histamine. Problems associated with histamine detection in fish as a control ordiagnostic measure for HFP include: the lack of uniform distribution of histamine intoxic fish; the large numbers of methods available and the lack of standardisationbetween countries; and the fact that histamine is not the only factor involved in thepathogenesis of HFP. High-performance liquid chromatography (HPLC) and capillaryelectrophoresis are often used, and rapid enzyme-linked immunosorbent assay(ELISA) test kits are also available. Methods including HPLC are used for detectingother biogenic amines and urocanic acid.
Dose−response assessment Since 1970, most reports of HFP have come from Japan, the US and Great Britain.Outbreaks have been reported less frequently in various other countries, includingAustralia and New Zealand. There are only two reports of HFP in Australia in thescientific literature. Juvenile Western Australian salmon caught in South Australianwaters were responsible for two outbreaks, affecting a total of seven people; and twopeople were affected by eating a tuna meal at a restaurant in Brisbane. The trueincidence of HFP in Australia is unknown.
Scombroid fish that cause HFP include mackerel (Scomber spp.), tuna (Thunnus spp.),saury (Cololabis saira) and bonito (Sarda app.) (scombroid fish). Non-scombroid fishthat cause HFP include mahi-mahi or dolphin fish (Coryphaena spp.), sardines(Sardinella spp.), pilchards (Sardina pilchardus), anchovies (Engraulis spp.), herring(Clupea spp.), marlin (Makaira spp.) and tailor or bluefish (Pomatomus spp.). Othernon-scombroid species, Western Australian salmon (Arripis truttaceus), sockeyesalmon (Oncorhynchus nerka) and Cape yellowtail (Seriola lalandii), have also beenimplicated.
Since free histidine in fish muscle is the substrate for microbial decarboxylation toproduce histamine, species difference in free histidine content has a large effect on thepotential hazards of poor handling and storage practices. Levels of ornithine (toproduce putrescine) and lysine (to produce cadaverine) may also be important.Histamine concentrations in fish tend to be greater adjacent to the gills or intestines.
Spoilage and ammonia and biogenic amine production are enhanced at elevatedtemperatures, with histamine production by bacteria such as Morganella morganii,Klebsiella pneumoniae and Hafnia alvei being optimal at 30oC. Once a large bacterialpopulation has been established, residual enzyme activity continues slowly atrefrigeration temperatures (0–5oC), although bacterial growth ceases. Histamine isalso produced, but to a lesser extent, by bacteria that can grow at refrigerationtemperatures (e.g Vibrio spp., Photobacterium spp.).

Histamine Fish Poisoning
4
It is not possible to predict accurately the effect of temperature abuse on histamineformation. Different bacterial species have different temperatures for maximumgrowth, and the histamine concentration in any part of a fish represents that producedby a population of bacteria with capacities for histamine production and/or destructionranging from high to zero. However, as spoilage progresses, the proportion of HDB inthe microbial population increases. In addition, quantification of bacterialcontamination is not meaningful. Subsequent heat processing can destroy bacterialcontaminants and even HD activity, but has little or no effect on histamine levels.
Although histamine is not solely responsible for HFP, levels of histamine in suspectfish serve as an important indicator of bacterial contamination, and many countrieshave set guidelines for maximum permitted levels. The current level for histamine infish in the Australian Food Standards Code is 100 mg/kg. However, concentrations ofhistamine within a fish are extremely variable, as is the threshold toxic dose.
If we regard 10 mg histamine/100 g of fish as the highest level that can be consumedsafely by most people, this must be related to the amount of fish eaten and the weightof the person to calculate a likely safe dose. If a 60-kg person eats, say, 300 g (wetweight) of this fish, this dose would be 0.5 mg/kg bodyweight. Such a calculation is oflimited value, however, considering the variable nature of HFP and the lack ofunderstanding of its pathogenesis.
The severity of the clinical response depends on the amount of toxin(s) ingested andthe variation in individual susceptibility. In some outbreaks the morbidity rate mayreach 100%. There is large variation in individual susceptibility. Certain dietarycomponents and medications such as isoniazid, aminoguanidine and someantihistaminic drugs increase susceptibility. Disease states, such as allergies andmastocytosis, may also affect the clinical manifestation of HFP.
Exposure assessment
Post-catching contamination with HDB may occur aboard the fishing vessel, at theprocessing plant, in the distribution system (fresh and frozen fish), and at the level ofthe user, for example in a restaurant. The key to the reduction of histamine productionis the rapid cooling of the fish after catching. As most histamine-producing bacteria require temperatures >15oC for growth, low-temperature storage (<10°C) effectively controls their growth. However, somebacteria can produce smaller amounts of histamine in fish stored at temperaturesbetween 0 and 10°C. Regardless of the species involved, bacteria must grow to a largeenough population for significant production of histamine to occur. If fish are subject to elevated temperatures, even for short periods, a large microbialpopulation is established. During subsequent refrigeration, although bacterial growthceases, residual enzyme activity continues slowly and histamine (and putrescine andcadaverine) levels continue to increase. If the fish are then hot smoked or canned, theheat will destroy the residual microflora and HD, but not histamine.

Histamine Fish Poisoning
5
Indications that HFP is not common in Australia are supported by results of limitedsurveys and routine monitoring. Of 104 cans of tuna screened in Melbourne retailmarkets in 1987 (45 from Australia and 59 from Asian countries), 101/104 cans had<5 mg/100 g histamine, and 3/104 had 5–10 mg/100 g. In a recent ACT Governmentsurvey of biogenic amines in seafood and seafood products, only 1/64 samples, driedfish from Asia, failed to comply with the Australian Food Standards Code level of100 mg/kg histamine. However, two dried fish samples contained >100 mg/kgputrescine, and two contained >100 mg/kg cadaverine. Ongoing random monitoring of imports and domestic product by the AustralianGovernment Analytical Laboratories (AGAL) has revealed only a small percentage ofsamples with >100 mg/kg histamine. The only remaining tuna cannery in Australia,Port Lincoln Tuna Processors Pty Ltd, South Australia, monitors histamine levels inall batches of fish entering the cannery, as well as in finished product, to ensure thatstrict food safety standards are maintained.
In developed countries where HFP still occurs quite frequently, such as the UnitedStates and Japan, most outbreaks are the result of consumption of fish caught byrecreational fishers, with insufficient knowledge of the problem and without properchilling facilities on fishing boats.
Although not reported to any extent, HFP is probably still common in developingcountries, where fish preserved by traditional methods is an important part of the diet.The problem would be expected to be greater in tropical countries, where highambient temperatures promote the growth of the most active HDB.
Great progress has been made in ensuring the quality of fish products despite the hugeexpansion in trade in recent years. This is the result of the introduction of internationalstandards in food hygiene and the application of risk analysis and Hazard Analysis andCritical Control Point (HACCP) principles. Although incidents of HFP caused by highlevels of histamine in canned tuna have occurred all over the world, improvements inhandling and processing associated with the establishment of quality controlprocedures are now widespread and taking effect.
Risk characterisation
HFP does not have a major impact on human health in Australia, partly because fishdoes not form a large part of the diet of most Australians. However, the disease isimportant from the food safety aspect and it is possible that toxic products,particularly imports, will escape the random monitoring safety net from time to time.Consumers are becoming more demanding and litigation following food poisoningincidents is becoming more common. Producers, distributors and restaurants willincreasingly be held liable for the quality of the products they handle and sell.
If a major outbreak of HFP were to occur in Australia, as has happened in Japan andthe United States, resulting media attention would affect fish consumption and have anegative impact on the marketing of seafood. An outbreak in another country causedby Australian exports would seriously affect trade. Although such events arebecoming increasingly less likely because of the widespread adoption of HACCP

Histamine Fish Poisoning
6
analysis and quality assurance, constant monitoring is necessary to allow for factorssuch as equipment failure, human error or negligence.
Other activities could also place Australia in a better position to characterise the riskof HFP and prevent or rapidly manage an outbreak should it occur. The pathogenesisof HFP requires elucidation. More research is needed to determine the threshold oftoxic dose for histamine and the role that potentiators or other toxins play in causingtoxicity. The possible role of urocanic acid in HFP, as a potential mast celldegranulator, should be investigated, initially in laboratory animals. There is also aneed to better assess the actual incidence of fish allergy and to estimate whatpercentage of cases diagnosed as fish allergy represent misdiagnosis of HFP.
There is a need for global standardisation of histamine detection methods, andlaboratory accreditation and proficiency testing, if histamine is to remain the mainindicator of microbial spoilage in histidine-containing fish. From an environmentalhealth perspective, a rapid and cheap assay for detecting histamine in fish would be ofvalue if made available to the public and, in particular, to recreational fishers.
However, monitoring fish histamine levels alone may not always ensure protectionfrom HFP. There may be advantage in the simultaneous detection of other biogenicamines, such as cadaverine and putrescine. The detection of histamine, cadaverine andputrescine can be achieved satisfactorily by the use of two-dimensional TLC orHPLC, but this is neither rapid nor cheap. Urocanic acid may be a useful alternative tohistamine as a spoilage index in scombroid and other fish that are rich in endogenoushistidine, and may be linked to the potential of fish to cause HFP.
Reporting of suspected cases of HFP to local food authorities should lead to removalof contaminated fish from the marketplace and prevention of additional cases.Mechanisms should be put in place, where these are not present already, to allowefficient and complete traceback of incriminated fish to point of origin, in order torectify problems leading to spoilage. In addition, there needs to be education ofrecreational fishers and the public about the need for good refrigeration and hygiene tominimise the possible hazards of consuming fish.

Histamine Fish Poisoning
7
1. INTRODUCTION
Scombroid fish poisoning, or more correctly histamine fish poisoning (HFP), is afoodborne chemical intoxication caused by eating spoiled, or bacteriallycontaminated, fish. Fish species associated with HFP are harmless when fresh, andafter they have become toxic they may still have a normal appearance and odour(Sapin-Jaloustre and Sapin-Jaloustre 1957). No available method of preparation,including freezing, canning and smoking, will destroy the causative toxin(s) (Etkind etal 1987).
Histamine — which is the main toxin involved in HFP — is also a naturally occurringsubstance in mammalian physiology. It is contained in mast cells and basophils, andits powerful biological effects are usually seen only when it is released in largeamounts in the course of allergic and other reactions. Histamine exerts its effects bybinding to receptors on cellular membranes in the respiratory, cardiovascular,gastrointestinal and haematological/immunological systems and the skin (Cavanahand Casale 1993). The pathogenesis of HFP has not been elucidated. The disease isnot uncomplicated histamine poisoning, but is generally associated with high levels ofhistamine (��� mg/100g) in spoiled fish.
Although it is a common form of fish poisoning, many incidents of HFP gounreported because of the mildness of the disease, lack of required reporting, andmisdiagnosis. Symptoms may be confused with those of other illnesses, particularly'Salmonella�LQIHFWLRQ��5XVVHOO�DQG�0DUHWLü�������DQG�IRRG�DOOHUJ\��7D\ORU������Taylor et al 1989; Institute of Medicine 1991; Sánchez-Guerrero et al 1997; Kerr andParke 1998). The syndrome is that of histamine toxicity, but there is individualvariation in susceptibility, and clinical signs are more severe in people takingmedications such as isoniazid, which inhibit enzymes responsible for normalhistamine-detoxification reactions in the intestine (Stratton et al 1991).
HFP is a significant public health and safety concern (Sanchez-Guerrero et al 1997;Wu et al 1997; Fletcher et al 1998) and a trade issue (Anon. 1998a). The earliestrecord of the disease was in 1828 (Henderson 1830). Since then, it has been describedin many countries, including Australia and New Zealand (Taylor 1985), and is nowthe most prevalent form of seafood-borne disease in the United States (Lipp and Rose1997). The worldwide network for harvesting, processing and distributing fishproducts has made HFP a global problem. Because the toxic condition is aconsequence of improper handling or storage of fish and there are effective testingmethods to identify likely toxic fish, control and prevention are possible (Institute ofMedicine 1991).
Scientific information on HFP is reviewed here in a risk-assessment framework(Kindred 1996; Buchanan et al 1998) in order to arrive at an informed characterisationof risk. An attempt has been made to address, as accurately as possible, the questions:'What causes outbreaks of HFP?', 'What are the underlying factors contributing tooutbreaks?' and 'What are the consequences of outbreaks?'
Risk assessment forms the scientific foundation for risk analysis, which comprisesrisk assessment, risk management and risk communication. The purpose of risk

Histamine Fish Poisoning
8
analysis is to foster informed management decision making. The risk assessmentpresented here is intended for use in a subsequent study to weigh policy alternativesand, if required, to select appropriate control options, including risk communication.
2. HAZARD IDENTIFICATION
2.1 Definition of hazard identification Kindred (1996) defined hazard identification as ’The determination of whether aparticular chemical is or is not causally linked to particular health effects’. Topicsdiscussed under hazard identification may include:
• sources of the hazard;• the frequency with which it may occur in food, and in what amounts;• the usual food vehicle(s);• the mechanism of toxicity;• the pathogenesis, clinical signs, pathology and diagnosis of the disease the hazard
causes; and• methods of detection of the hazard.
The involvement of histamine as the main hazard in scombroid poisoning or HFP issupported by:
• symptoms identical to those of intravenous histamine administration or allergicreaction;
• the efficacy of antihistamine therapy; and• the presence of increased levels of histamine in spoiled fish that cause the
syndrome.
In 1991, Morrow et al claimed that there was definitive evidence that histamine is thetoxic agent in HFP after analysing the urine of poisoned subjects. They found thaturinary levels of histamine in patients with HFP were higher than those of subjectswho had been injected intravenously with histamine to produce toxic symptoms. However, HFP is not simply histamine poisoning. Consuming spoiled fish containinghistamine is more likely to cause toxic effects than taking the same amount of purehistamine by mouth. Pure histamine taken orally is substantially metabolised incrossing the intestinal wall or in the liver, and produces only mild symptoms at highdoses (Taylor et al 1984). Thus, while elevated blood histamine levels areundoubtedly characteristic of HFP, there is uncertainty about how these occur. Thepathogenesis of the disease is complicated and there is speculation about the identityof other possible ’scombroid toxins’ acting in concert with histamine. Taylor (1986) said that HFP appears to be caused by exogenous histamine (i.e. fromspoiled fish), potentiated by other biogenic amines and possibly other substances.Clifford et al (1991) and Ijomah et al (1991), on the other hand, postulated that atoxin or toxins other than histamine in the spoiled fish cause release of endogenoushistamine from mast cells in the human body. The indication that there may be

Histamine Fish Poisoning
9
multiple toxins, perhaps acting synergistically, may account for the considerablevariability of clinical signs and symptoms encountered.
2.2 Production of histamine, other biogenic amines and imidazole compoundsin spoiling fish
Histamine production in fish is related to the histidine content of the fish, the presenceof bacterial histidine decarboxylase (HD), and environmental conditions (Ienistea1973). During spoilage, certain bacteria produce decarboxylase enzymes, which act onfree histidine and other amino acids in the fish muscle to form histamine and otherbiogenic amines. Chemically, histamine (from histidine), putrescine (from ornithine),cadaverine (from lysine), and spermidine and spermine (from arginine), which areproduced post-mortem in fish muscle, are low-molecular-weight, aliphatic, alicyclicor heterocyclic organic bases (Eitenmiller and De Souza 1984; Rawles et al 1996). Fish species belonging to the families Scombridae (e.g. tuna and mackerel) andScomberesocidae (e.g. saury) are most commonly associated with HFP, but non-scombroid fish, such as mahi-mahi, sardines, pilchards, anchovies, herring, marlin andbluefish can also be involved (Taylor 1986). These fish species are characterised byhaving relatively high levels of histidine in their flesh (Taylor 1986; Institute ofMedicine 1991). Histidine levels vary from 1 g/kg in herring to as much as 15 g/kg intuna (Ijomah et al 1992). Frank et al (1981) found that fresh (skipjack) tuna(Katsuwonus pelamis) contained negligible quantities of histamine, usually<0.1 mg/100 g. Histamine can be produced rapidly by bacterial decarboxylases in scombroid andother fish that have relatively high free histidine levels in their muscles when alive(Love 1980), before post-mortem proteolysis liberates additional histidine frommuscle protein. This explains why toxic levels of histamine are generally limited tothe ’red meat of free-swimming species’ (Arnold and Brown 1978) and why histaminecan reach high concentrations without the formation of organoleptic (sensory)spoilage indicators (Sapin-Jaloustre and Sapin-Jaloustre 1957). After investigatingHD production by Morganella morganii in mackerel, Eitenmiller et al (1982)concluded that the ready availability of free histidine in the muscle to act as both aninducer and substrate makes scombroid fish muscle an ideal environment forhistamine formation. The main bacteria responsible for histidine decarboxylation and HFP are members ofthe family Enterobacteriaceae (Frank et al 1985; Taylor and Sumner 1986).Endogenous production of decarboxylase enzymes is insignificant when comparedwith the exogenous (bacterial) pathway (Rawles et al 1996). Spoilage, ammoniaproduction and biogenic amine production are enhanced at elevated storagetemperatures, with histamine production being optimal at around 30oC (Arnold et al1980). Once a large bacterial population has been established, residual enzymeactivity continues slowly at refrigeration temperatures (0–5oC), although bacterialgrowth ceases (Institute of Medicine 1991). Histamine is also produced, but to a lesserextent, by bacteria that can grow at refrigeration temperatures (Okuzumi et al 1981).

Histamine Fish Poisoning
10
Only free histidine can be decarboxylated (Arnold and Brown 1978). Geiger (1944b)observed that the ability of Escherichia coli to decarboxylate histidine was highlyspecific, since histamine formation was inhibited by acylation of the amino group ofhistidine. He later studied histamine formation with a Clostridium perfringens enzymepreparation, a strain of E. coli and a strain of a marine bacterium. All were able toform histamine from free L-histidine, but none were able to form histamine fromaspartyl-histidine or histidyl-histidine. Geiger (1944b) concluded that peptide linkagesto either the -COOH or -NH2 groups of histidine prevented bacterial decarboxylation. The decarboxylation of histidine to form histamine is only one of two routes ofhistidine metabolism, and the occurrence of this pathway in fish spoilage is quitelimited. According to Baranowski (1985), the pathway favoured by most bacteria is acatabolic one in which glutamate is the ultimate product formed. The first step in thispathway is the loss of ammonia from histidine by the action of L-histidine ammonialysase (HAL), or histidase, resulting in the formation of urocanic acid (Fig. 1 below).
Figure 1. Degradation of histidine by HAL to form urocanic acid and ammonia. (FromWhite et al 1973.) HAL has a wide distribution among bacteria (Shibatani et al 1974; Baranowski 1985)and, unlike HD, is also found as an endogenous component of fish muscle (Kawai andSukaguchi 1968; Mackie and Fernández-Salgéuro 1977). Some bacteria also possessurocanase, which catabolises urocanic acid, but Shibatani et al (1974) found that nostrain of 106 tested had urocanase activity equal to its HAL activity. Urocanic acidwas found at a much higher concentration than histamine (4.74 versus0.19 mg/100 mL) in mackerel after 18 days' storage at 0oC (Mackie and Fernández-Salgéuro 1977; Fernández-Salgéuro and Mackie 1979), although in this experiment itwas attributed to endogenous rather than bacterial HAL. Shewan (1955) investigated the distribution of free bases and amino acids in musclefrom fresh and spoiled mackerel by fractionation by displacement chromatography onion-exchange resins of aqueous–alcoholic extracts. Following spoilage, he found largeamounts of histamine and moderate amounts of an unidentified (probably imidazole)component (compound 8) not found in fresh muscle. Later, Hughes (1959)

Histamine Fish Poisoning
11
demonstrated a major imidazole spoilage product (compound X) from spoiled herringby paper chromatography, which gave cherry red colour with Pauly’s reagent,suggesting the imidazole portion of the molecule was still intact. However, his effortsto identify compound X as urocanic acid (imidazolylacrylic acid),imidazolylcarboxylic acid or imidazolylacetic acid by comparison with ’authenticsamples’ (of unspecified origin) were unsuccessful.
2.3 Metabolism of histidine and histamine in mammals Humans metabolise histidine to urocanic acid through the activity of HAL, to formglutamate and then α-ketoglutarate, which enters the citric acid cycle (White et al1973; Furuta et al 1996), or to histamine through the activity of HD (Stryer 1981). Histamine can be catabolised by several routes (Fig. 2). It can be oxidativelydeaminated by diamine oxidase (DAO, or histaminase) to imidazole acetaldehyde andimidazoleacetic acid, methylated by histamine methyl transferase (HMT) to formmethylhistamine, or its side chain can be methylated or acetylated (Rice et al 1976;Taylor 1986).
Figure 2. Histamine catabolic routes. (From Taylor 1986.)

Histamine Fish Poisoning
12
The fact that oral histamine alone is not toxic to adults except at doses ≥100 mgappears to be due mainly to the presence of DAO and HMT in the intestinal tract,which detoxify histamine (references cited by Taylor 1986). Histamine is alsoconverted to inactive acetylhistamine in the intestine, presumably by bacterial enzyme(references cited by Taylor 1986). All these enzymes decrease the amount ofunmetabolised histamine available for absorption into the circulation (Granerus 1968;Taylor and Lieber 1979; Lyons et al 1983; Hui and Taylor 1985; Taylor 1986). The relative importance of the DAO and HMT pathways varies among species. Theoxidative deamination pathway predominates in rats and guinea pigs, and themethylation pathway is of prime importance in humans, mice, cats, pigs and hamsters(Taylor 1986). When [14C]-histamine was administered orally to humans, 68–80% of the radioactivedose was recovered in the urine (Sjaastad and Sjaastad 1974). Some histamineremained unchanged in the faeces, and additional amounts were catabolised byintestinal bacteria and radioactivity was exhaled as 14CO2 from the lungs. Hesterberget al (1984) demonstrated that HMT activity was widespread in human tissues, withthe order of activity being liver >> colon >spleen > lung > small intestine > stomach.By contrast, DAO was mainly localised in the intestine (Hesterberg et al 1984).Helander et al (1965) had shown earlier that the human kidney possessed considerablecapacity for removing histamine from the blood. When they infused healthyindividuals with histamine intravenously, a large proportion was methylated by thekidney and excreted in the urine, while a smaller proportion was excreted unchangedin the urine. HMT is very selective for histamine, and requires S-adenosylmethionine as a methyldonor. HMT is inhibited by analogues of methylmethionine, such as adenosyl-homocysteine, antimalarial drugs and numerous agonists and antagonists of histaminereceptors. HMT is also subject to substrate inhibition by high concentrations ofhistamine. The type of inhibition (competitive, uncompetitive, non-competitive)varies depending on the inhibitor. (References cited by Taylor 1986.) DAO oxidises other diamines, such as putrescine, as well as histamine. Manyinhibitors of DAO have been identified, such as aminoguanidine and someantihistaminic drugs. DAO is also subject to substrate inhibition when certaindiamines including histamine are used as substrates. A number of foodbornesubstances are inhibitors of DAO, including carnosine, thiamine, cadaverine andtyramine. Monoamine oxidase (MAO) is also important in histamine metabolism.(References cited by Taylor 1986.)

Histamine Fish Poisoning
13
2.4 Possible mechanisms of toxicity 2.4.1 Toxicity of histamine and other biogenic amines In the body, histamine exerts its toxicity by interacting with receptors on cellularmembranes. There are three types of histamine receptors, H1, H2 and H3 (Cavanah andCasale 1993). The most common symptoms of histamine poisoning result from theaction of histamine on the cardiovascular system, which involves H1 and H2 receptors.Histamine causes dilatation of peripheral blood vessels, capillaries and arteries,causing urticaria, hypotension, flushing and headache. Histamine-induced contractionof intestinal smooth muscle, mediated by H1 receptors, causes abdominal cramps,diarrhoea and vomiting. Pain and itching associated with the urticarial lesions may bedue to sensory and motor neuron stimulation, which is also mediated by H1 receptors(Taylor 1986). There is compelling evidence to implicate histamine as the causative agent in HFP —high levels of histamine have been found in food samples implicated in outbreaks, thesymptoms noted in the patients are consistent with histamine poisoning, andantihistamines are effective in counteracting the symptoms. However, there is not astraightforward dose–response relationship. Spoiled fish containing histamine tends tobe more toxic than the equivalent amount of pure histamine dosed orally. Studies onHFP have been confused by interacting variables affecting both fish samples and theconsumer and, according to Ijomah et al (1992), most investigations have been limitedin their ability to disentangle these. Potentially confusing factors are listed in the tablebelow. Potentially confusing factors in HFP
Samples Consumers Variable histamine levels in sample Microbial toxins Other toxins/contaminants/ metabolites Wrong sample analysed
Wrong diagnosis Bodyweight differences Individual variation Sex differences in metabolism Concomitant medication Idiosyncratic intolerance True allergy
Weiss et al (1932) found that 180 mg histamine base (given as 500 mg histaminephosphate) administered orally was without noticeable effect in humans, while 7 µgadministered intravenously caused vasodilatation and increased heart rate. Granerus(1968) gave humans up to 67.5 mg histamine orally without toxic effect. Motil andScrimshaw (1979) found that administration of 100–180 mg histamine orally mixed ingrapefruit juice (containing citric acid) or in 100 g high-quality tuna causedcharacteristic symptoms of mild histamine poisoning (mild-to-severe headache andobvious flushing) in some people (1/4 and 4/8 respectively). These doses represented1.6–3 mg/kg bodyweight (bw), assuming an average weight of 60 kg. Subsequently,Clifford et al (1989) gave volunteers 50 g of fresh mackerel to which 300 mghistamine had been added (a dose of about 5 mg/kg bw), and recorded only mildsymptoms of histamine poisoning (oral tingling, headache and flushing in some

Histamine Fish Poisoning
14
subjects). However, 50 g of spoiled mackerel with 300 mg added histamine was nostronger in its effect. Theories have been put forward to help explain why fish that cause histaminepoisoning often do not contain levels of histamine considered high enough to be toxicorally. Some scientists maintain that, in HFP, factors (potentiators) act to makeexogenous histamine more toxic, possibly by increasing its absorption or decreasingits detoxification in the intestinal tract or elsewhere in the body, such as the liver(Taylor and Lieber 1979; Lyons et al 1983; Hui and Taylor 1985). Others believe thatanother toxin or toxins might act to liberate endogenous histamine, which contributesto histamine toxicity (Clifford et al 1991; Ijomah et al 1991). A survey of scombrotoxic fish poisoning in Britain by Bartholomew et al (1987)suggests that most cases may be uncomplicated histamine poisoning, and that othertoxins may be involved when the suspect fish contains little histamine, or whensymptoms are not typical. Between 1976 and 1986, Bartholomew et al (1987)followed up 258 incidents of suspected scombrotoxic fish poisoning. Of 240 fishsamples from these incidents, 101 contained >5 mg histamine/100 g fish. Thesymptoms most consistently reported were rash, diarrhoea, flushing and headache. Inany one incident, the symptoms of all patients were similar, although each patient didnot experience all symptoms. Of fish with >20 mg histamine/100 g, 94% were fromincidents in which scombrotoxic symptoms were characteristic. Where fish containedonly 5–20 mg histamine/100 g fish, only 38% of incidents were 'clinically distinctive'.The authors regarded rash, flushing and burning of the mouth as distinguishingfeatures of scombrotoxic fish poisoning. Although gastrointestinal symptomsespecially diarrhoea were frequently experienced, in contrast with some previousreports (Arnold and Brown 1978), gastrointestinal symptoms alone were not regardedas indicative of scombrotoxic fish poisoning. Surprisingly, 36/93 fish samples (39%)with <5 mg/100 g histamine also gave rise to characteristic scombrotoxic (or HFP)symptoms. Analyses were not done for other biogenic amines, which may have beentoxic in their own right, or histamine potentiators. Til et al (1997) examined the acute and subacute toxicity of five biogenic amines —tyramine, spermidine, spermine, putrescine and cadaverine — in Wistar rats. For theacute studies, the amines were administered orally as a 25% aqueous solution atvarious dose levels to 8–13/week-old rats (2/sex/group). The rats were observed for14 days and clinical signs and deaths were recorded. The rats that died and thesurvivors killed after 14 days were examined pathologically. For the subacute studiesthe rats were fed diets containing 0, 200, 2000 or 10 000 ppm (or mg/kg food)cadaverine or tyramine; 0, 200, 2000 or 5000 ppm spermine or putrescine; or 0, 20,200 or 500 ppm spermidine for 5–6 weeks. The major findings are summarised in thetable below.

Histamine Fish Poisoning
15
Summary of major findings obtained in acute and subacute studies with five biogenic amines
Observations Tyr Spd Spm Put Cad Approx. LD50 (mg/kg) NOEL (ppm) Mortality Paralysis Bodyweight Anaemia Haemoconcentration Impaired renal function Histopathology – Liver –Kidneys –Heart
>2000 2000
0 0 – 0 0 0 0 0 0
600 1000
0 0 – 0 0 0 0 0 0
600 200 ++ +
– – – + 0
+ +
+ + + +
2000 2000
0 0 – 0 0 0 0 0 0
5000 2000
0 0
– – 0 + 0 0 0 0
Tyr = tyramine; Spd = spermidine; Spm = spermine; Put = putrescine; Cad = cadaverine LD50 = single (oral) dose required to cause death in 50% of animals dosed NOEL = no observable effect level 0 = not affected; + = slightly increased or slightly affected; – = slightly decreased; – – = moderatelydecreased; – – – = markedly decreased. The doses of biogenic amines tested and tolerated in rats are many times higher thanwould be expected in humans following the ingestion of a meal of spoiled fish.However, the amines were tested individually, not in combination with one another,or with histamine. Spermine was the most toxic of the five amines tested, causingliver changes, elevated activity of plasma enzymes associated with the liver,nephrotoxicity and myocardial degeneration. It is somewhat reassuring that Mietz andKarmas (1977) found that levels of spermine and spermidine, the most toxic amines inthis series, tended to fall and sometimes reached zero during the decompositionprocess in tuna. 2.4.2 Inhibition of histamine detoxification by histamine potentiators 2.4.2.1 Background Among the scientists who have postulated that histamine is potentiated by some othercomponent(s) in the toxic fish are Bjeldanes et al (1978), Taylor and Lieber (1979),Chu and Bjeldanes (1981), Lyons et al (1983), Taylor (1986, 1988), and Stratton et al(1991). By definition, such a potentiator(s) would act to decrease the threshold dose ofhistamine needed to provoke an adverse reaction in humans challenged orally. Certaindrugs have definitely been implicated as contributing factors in cases of histaminepoisoning (Chin et al 1989; Stratton et al 1991). Taylor (1986) stated that doses of pure histamine required to produce mild reactionswere ’several times higher than the doses producing more severe symptoms whenconsumed with spoiled fish’. In the same paper he stated that ’the variability inhistamine levels in spoiled fish makes estimates of the toxic threshold difficult toobtain’ and quoted an estimated threshold dose of histamine in fish at about60 mg/100 g flesh. Even after allowing for variability in human susceptibility andvariable histamine content in different parts of fish (Lerke et al 1978), it seems that

Histamine Fish Poisoning
16
there is indeed some difference between the relative lack of toxicity of pure histamineand the (often) apparent toxicity of histamine in spoiled fish. In support of the histamine-potentiator hypothesis, several in vivo and in vitro studieshave suggested that the absorption, metabolism, and/or potency of one biogenic aminemight be modified in the presence of a second amine (Bjeldanes et al 1978; Taylorand Lieber 1979; Lyons et al 1983). The biogenic amines putrescine and cadaverineoccur in appreciable quantities in toxic fish (Arnold and Brown 1978) and at lowlevels in non-toxic fish (Mietz and Karmas 1977). When given in higher ratiosrelative to histamine than those that usually seem to occur in toxic fish, these aminespotentiate the biological activity of histamine in laboratory animals. However, according to Hui and Taylor (1985), uptake of unmetabolised histaminealone would not be sufficient to elicit some of the symptoms observed in HFP. Toexert its full toxic effects, histamine must reach the peripheral tissues. According tothese authors, the detoxification of histamine in extra-intestinal tissues must also beinhibited to achieve the full effects. 2.4.2.2 In vitro studies Mongar (1957) observed that cadaverine and putrescine competitively inhibited DAOand potentiated histamine-induced contractions in guinea pig ileum. Taylor and Lieber(1979) tested the effects of 38 chemicals (mainly nitrogen-containing bases) likely tobe consumed with tuna on the in vitro activity of rat jejunal mucosal HMT and DAO.The most potent inhibitors (at 10 mM concentrations) were monoamines, diaminesand guanidines, including tyramine, β-phenylethylamine and tryptamine, butcorrelations between chemical structure and inhibitory activity were difficult todefine. The scientists reported that many of the identified chemicals are found inspoiled tuna, along with histamine, and their ability to inhibit intestinal histaminecatabolism could magnify the oral toxicity of histamine and explain its apparentlygreater toxicity when consumed with spoiled tuna. Cadaverine and aminoguanidine were both strong inhibitors of rat intestinal DAO andHMT in vitro (cadaverine, 87% and 35%; and aminoguanidine, 100% and 81% forDAO and HMT respectively). Putrescine was a weak inhibitor (11%) of HMT only(Taylor and Lieber 1979). Lyons et al (1983) subsequently showed that cadaverineand aminoguanidine enhanced the absorption of unmetabolised histamine in perfusedrat intestinal segments by inhibiting the conversion of histamine to less-toxicmetabolites. Aminoguanidine is not known to occur in spoiled fish (Lyons et al 1983). 2.4.2.3 In vivo studies Parrot and Nicot (1966) found that the oral toxicity of histamine in the guinea pig wasincreased 10 times when it was administered 40 min after oral administration ofputrescine. And Bjeldanes et al (1978) found that the oral toxicity of histamine in theguinea pig was potentiated by simultaneous administration of cadaverine. Otherbiogenic amines that may act as potentiators of histamine toxicity include tyramine (apressor amine that inhibits MAO), tryptamine (which inhibits DAO), and β-

Histamine Fish Poisoning
17
phenylethylamine (a DAO and HMT inhibitor) (references cited by Stratton et al1991). However, Bjeldanes et al (1978), in an article on the aetiology of HFP, put thequestion of a synergist for histamine in proper perspective. They pointed out that, toexhibit synergism, as measured by LD50 values in the guinea pig, Parrot and Nicot(1966) used an unrealistically high ratio of putrescine to histamine (5:1, comparedwith 1:100 in toxic fish). In their own LD50 experiments on guinea pigs, Bjeldanes etal (1978) found that cadaverine was synergistic only at a ratio to histamine of 1:5 orgreater, whereas in toxic fish (tuna) the ratio is 1:10 (Mietz and Karmas 1977; Kim1978). They also demonstrated that while optimal cadaverine synergism was obtainedby simultaneous administration with histamine to guinea pigs, putrescine required atime lag before histamine was administered. As it stands, the above study suggests that it is unlikely that putrescine in spoiled fishacts as a histamine potentiator, and that cadaverine may only possibly be involved.However, Klausen and Lund (1986) found, when comparing mackerel and herring,that levels of cadaverine in mackerel may exceed those of histamine by 2–5 times,which may well explain why this species commonly causes HFP. Hui and Taylor (1985) determined the effects of enzyme inhibitors of histamine invivo and their possible role in potentiating histamine toxicity by studying the effects ofknown inhibitors of DAO, HMT and MAO, both foodborne and pharmacological, onthe urinary excretion pattern of histamine and its metabolites in rats. When theyadministered [14C]-histamine orally to rats (female Sprague-Dawley, 3–5/group), anaverage of 80% of the administered radioactivity was recovered in the urine after 24 h.About 10% of the total dose was excreted via the faeces. Analysis of 4-h urinesamples showed imidazoleacetic acid was the predominant metabolite (60.6%), andN-methyl-imidazoleacetic acid (8.6%), N-methylhistamine (7.3%) and N-acetylhistamine (4.5%) were minor metabolites. Histamine metabolism was inhibited by simultaneous oral administration of the drugsaminoguanidine, isoniazid and quinacrine, and the biogenic amines cadaverine,putrescine, tyramine and β-phenylethylamine. The administration of inhibitorsresulted in an increased amount of unmetabolised histamine and a decreased amountof metabolites reaching the urine. However, the total rate of excretion of histamineand its metabolites in the urine was similar in the presence and absence ofpotentiators, indicating that the total amount of absorption and excretion of histamineand its metabolites was equivalent, with or without potentiators. Hui and Taylor (1985) found that pharmacological inhibitors were more potent andhad a longer duration of action in rats than foodborne ones. Cadaverinedihydrochloride (≥2 mmol/kg, equivalent to ≥204 mg cadaverine/kg), putrescinedihydrochloride (2.5 mmol/kg), tyramine hydrochloride (2.5 mmol/kg) and β-phenylethylamine hydrochloride (2.5 mmol/kg) were weak inhibitors, effective atdoses 4–5 times higher than that of simultaneously administered histaminedihydrochloride (0.5 mmol/kg, equivalent to 55 mg histamine/kg).

Histamine Fish Poisoning
18
Inhibitors of DAO (aminoguanidine, isoniazid, cadaverine, β-phenylethylamine andtyramine) had a more profound effect on excretion of unmetabolised histamine in ratsthan inhibitors of only HMT. To test the possibility that more than one potentiatormay be involved in foods implicated in histamine poisoning incidents, Hui and Taylor(1985) tested a mixture of histamine (0.5 mmol/kg) and the foodborne inhibitorscadaverine, putrescine, tyramine, β-phenylethylamine and tryptamine (each dosed at0.5 mmol/kg). The mixture increased the amount of unmetabolised histamine in 4-hurine samples 100% over that present when histamine was dosed alone. These results indicate that cadaverine and these other amines could indeed potentiatehistamine toxicity in spoiled fish if present alone (in the case of cadaverine) orcollectively at levels ≥4 times greater than that of histamine. Furthermore, it is likelythat all the possible histamine potentiators have not been identified. 2.4.3 Barrier disruption hypothesis Another possible mechanism of potentiation of histamine by other biogenic amines isthe ’barrier disruption hypothesis’. The hypothesis was first proposed by Parrot andNicot (1966), who suggested that potentiators might interfere with the protectiveactions of intestinal mucin. Intestinal mucin is known to bind histamine, and it hasbeen suggested that this binding is essential to prevent the intestinal absorption ofhistamine. Potentiation would occur by disruption of the bonding and resultingenhancement of absorption and an increase in histamine toxicity (Jung and Bjeldanes1979; Chu and Bjeldanes 1981). Jung and Bjeldanes (1979) found that cadaverine exhibited a marked influence on therate of transport of 14C-labelled histamine and metabolites across the gut wall inisolated gut sections of the guinea pig. However, cadaverine had a minor effect onhistamine metabolism: the percentage of free histamine appearing outside the gut sacincreased from 30.7% when histamine alone was present, to 35.2% and 37.2% whencadaverine was added in ratios to histamine of 1:2 and 1:1 respectively. Whenhistamine alone was placed inside the gut, 41.2% of radioactivity was transportedacross the gut wall as histamine and metabolites. The percentage increasedsignificantly to 48% by adding cadaverine in the ratio of cadaverine:histamine of 1:10,to 52.8% at the ratio 1:2, and to 53.6% at the ratio 1:1. Jung and Bjeldanes (1979) suggested that cadaverine potentiation of histaminetoxicity may result from induction of increased rates of absorption of histamine andmetabolites, and that the established antihistaminase activity of cadaverine does notappear to play a significant role. They believed their results were consistent with aproposed role of substances such as mucin in maintaining a barrier to histaminetransport. According to Taylor (1986) and Mitchell (1993), experiments based on this theoryhave not so far provided a convincing rationale for the apparent toxicity of histaminein fish.

Histamine Fish Poisoning
19
2.4.4 Release of endogenous (mast cell) histamine by scombroid toxin(s) In 1957, Mongar had reported that diamines in the series NH2(CH2)nNH2 releasehistamines from isolated tissues. His results were obtained using minced guinea piglung and isolated rat diaphragm to measure histamine-releasing activity of diamines ofchain length C5–C15. The activity increased steadily with chain length from less than5% release with cadaverine to >90% release with pentadecamethylene-diamine duringa test period of 15 min. The effect of all diamines increased steeply withconcentration. In a concentration of 1 mM, cadaverine was inactive both on mincedguinea-pig lung and rat diaphragm, but, at 100 mM, it released about 80% of thehistamine content of the tissue. As mentioned above, Mongar (1957) also reported on the antihistamine activity ofdiamines, which produced a depression of the histamine contractions of guinea-pigileum. Histamine-releasing activity increased steadily with chain length, but theactivity of diamines as inhibitors of histaminase decreased with chain length. Mongar(1957) noted that the parallelism between histamine-releasing activity and toxicityextended to monoamines in the series CH3(CH2)n-1NH2. Histamine-releasing activityof both series increased with pH and was attributed mainly to the non-ionised base.However, since the compounds were inactive at pH 7 and their activity rose sharplywith pH to reach a high value at pH 8.5, they would be ionised at physiological pHand not active in producing endogenous histamine in the human body. However, the idea that release of endogenous histamine may be involved in HFPpersisted. Attention began to focus on mast cells, the granules of which contain ahistamine–heparin complex (Riley 1959). In 1972, Olley postulated that 'some otherbasic substance produced during spoilage [of fish] may release histamine from thehistamine–heparin complex of mast cells'. Arnold and Brown (1978) also pointed outthat release of histamine from mast cell granules could account for some, if not all, ofthe symptoms associated with scombroid toxicity. Subsequently, Clifford et al (1991) and Ijomah et al (1991) of the UK Ministry ofAgriculture, Fisheries and Food postulated that the scombroid toxin(s) is a mast celldegranulator, and the antihistamine therapy is effective because it eliminates the effectof endogenous histamine. Ijomah et al (1991, 1992) fed mackerel deliberately exposed to poor storageconditions to volunteers. They found that the volunteers fell into susceptible and non-susceptible subgroups, and that the level of histamine in fish did not correlate withtoxicity exhibited. However, when subjects were dosed with either placebo or H1
antagonist (chlorpheniramine 4 mg), the antihistamine convincingly abolishedvomiting and diarrhoea associated with the ingestion of scombrotoxic fish. Thescientists concluded that dietary histamine played a minor role in the toxicity and thatthe major contributor was a postulated agent that released histamine by degranulationof mast cells in the gastrointestinal tract. Clifford et al (1991) went further, to analyse mackerel fillets that had been associatedwith an outbreak of 'scombrotoxicosis' for their contents of cadaverine, histamine,putrescine, spermidine, spermine and tyramine. The same fillets were fed to healthy

Histamine Fish Poisoning
20
volunteers. Susceptibility to the disease was quite variable. Of the 86 fillets examined,30 rapidly induced nausea/vomiting and/or diarrhoea when 50-g portions wereconsumed. The remaining fillets failed to provoke such symptoms, even thoughvolunteers proven to be susceptible to scombrotoxicosis tested 17 of them. Statisticalanalysis failed to detect differences in amine content between fillets shown to bescombrotoxic and those failing to induce symptoms, and failed to establish significantrelationships between the amine doses and volunteer responses, even aftermanipulations to simulate additive or synergistic interactions. The minimum dosesassociated with scombrotoxicosis and the maximum doses giving negative responsesin susceptible volunteers were: histamine, 0.23 and 3.33; cadaverine, <0.02 and 0.38;putrescine, 0.0008 and 0.01; tyramine, 0.009 and 0.75 mg/kg bodyweight (bw)respectively. The scientists concluded that the contents of such amines in mackerelhave little or no role in the aetiology of scombrotoxicosis, even when acting additivelyor synergistically, and that the primary scombrotoxin, which remained unknown, wasresponsible for mast cell degranulation. Other clinical reports of HFP, for examples those caused by the consumption ofsalmon, support the suggestion that exogenous histamine is not always responsible forintoxication, and that other toxins and/or endogenous histamine may be involved.Bartholomew et al (1987) reported six incidents of scombrotoxic fish poisoning incanned salmon in Britain in 1983. In five of the incidents, the incriminated fishcontained <1 mg/100 g histamine, despite the presence of characteristic symptoms andincubation period of histamine poisoning. Salmon from the sixth episode containedonly 17 mg/100 g histamine. In addition, Gessner et al (1996) described a typical and severe scombrotoxic-likeillness following the ingestion of smoked sockeye salmon that demonstrated lowhistamine levels and high toxicity on mouse bioassay. Symptoms included flushing,pruritus, nausea, sweating, vomiting, diarrhoea, dizziness etc. The implicated salmonhad histamine levels 25-fold less than the US Food and Drug Administration (FDA)toxicity level for tuna of 50 mg/100 g (FDA 1998). The patient ate an estimated0.0006 mg of histamine/kg bw — far less than the estimated 1 mg of histamine/kg bwreported to cause human illness (Taylor 1986). Estimated levels for putrescine andcadaverine were 0.67 mg/100 g and 0.19 mg/100 g. (The analyses were performed onfish caught and treated at the same time and in the same way as the incriminated fish.) Some imidazole compounds are known to release histamine from mast cells by a non-immunological mechanism; for example, imidazole fungicides (Gietzen et al 1996). Itis interesting to note that urocanic acid, an imidazole compound and a histidinemetabolite of spoiling fish (Kawai and Sakaguchi 1968; Mackie and Fernández-Salguero 1977; Baranowski 1985), has recently been described as an inducer ofhistamine in vivo in mice (Hart et al 1997) and as a mast cell degranulator in humanskin organ cultures (Wille and Kydonieus 1995; Wille et al 1999). Urocanic acid maybe one of the 'toxic imidazole compounds' in spoiling fish mentioned by Olley in 1972and the 'scombroid toxin' that researchers have been seeking for decades. Some experimental evidence seems to reject the mast cell degranulation theory, butthis may reflect the large number of variables involved in the pathogenesis of HFP.Mast cell degranulation may not occur in all cases of the disease, but may be an

Histamine Fish Poisoning
21
important feature in other cases. When Van Gelderen et al (1992) gave eight healthyvolunteers either herring paste containing 90 mg histamine formed from inducedspoilage with photobacteria or fresh fish to which 90 mg histamine had been added,the maximum histamine concentration in plasma did not differ significantly betweenthe two dose regimes. However, herring may not have been the most suitable fish touse in this experiment. Klausen and Lund (1986) found that herring contained 4–5times less free histidine and free lysine (cadaverine precursor) than fresh mackerel. Inaddition, the photobacteria used to induce spoilage may not have possessed HAL,which is required to produce urocanic acid from histidine. Morrow et al (1991) measured the urinary excretion of histamine and its metabolite,N-methylhistamine, in three people who had HFP after eating marlin that containedhigh levels of histamine (842–2503 µmol/100 g, equivalent to 93–278 mg/100 g).They also measured 9α,11β-dihydroxy-15-oxo-2,3,18,19-tetranorprost-5-ene-1,20-dioic acid (PGD-M), the principal metabolite of prostaglandin D2, a mast cellsecretory product, to assess whether mast cells had been activated to releasehistamine. Flushing and headache began in all three people 10–30 min after ingestionof the fish. One person also had severe diarrhoea of sudden onset. The levels ofhistamine and N-methylhistamine were 9–20 and 15–20 times respectively the normalmeans in urine samples collected 1–4 h after the meal. During the subsequent 24 h,the levels fell to 4–15 and 4–11 times the normal values, and 14 days later they hadreturned to normal. PGD-M excretion was not increased at any time. Two of the threepatients were treated with an antihistamine (diphenhydramine), which was rapidlyeffective. In the study of Morrow et al (1991), HFP was associated with urinary excretion ofhistamine in quantities far exceeding those required to produce toxicity. The highhistamine levels in the incriminated fish, together with the failure to find increasedendogenous release of prostaglandin D2, indicated that the histamine was derived fromthe fish. Analyses were not done for other biogenic amines, which could have beenhistamine potentiators. However, increases in the urinary excretion of both histamineand N-methylhistamine of similar magnitude in the poisoned people suggested that thespoiled fish did not contain a substance or substances that potentiated histaminetoxicity by inhibiting its inactivation by HMT. Like Morrow et al (1991), Sánchez-Guerrero et al (1997) rejected the proposal thatendogenous histamine is involved in HFP. The latter authors used immunoassay tomeasure tryptase, another indicator of mast cell degranulation, in seven patientsintoxicated after eating tuna. The quantification of tryptase release in vivo allows aprecise valuation of mast cell activation, as the enzyme is located in secretory granulesof mast cells and released at the same time as histamine during degranulation. Thelow levels obtained caused the authors to reject an anaphylactic-type reaction andfavour poisoning by an exogenous source of histamine (fish). The role of endogenous histamine release, if any, in histamine/scombroid fishpoisoning remains uncertain and unproved. The poisoning syndrome described byIjomah et al (1991, 1992) and Clifford et al (1991) was not what would be consideredtypical of HFP. Many toxins produce nausea, vomiting and diarrhoea. Mitchell (1993)wrote that a possible factor in explaining the apparent contradiction of this work with

Histamine Fish Poisoning
22
previous observations on HFP is the definition of poisoning that was applied. Ijomahet al (1991) considered that vomiting and/or diarrhoea were essential factors inpoisoning. Flushing and tingling in the mouth were considered ’minor’ and possiblycaused by dietary histamine. The syndrome described by Ijomah, Clifford andcolleagues may well have been complicated by a toxin or toxins additional to thosecommonly associated with HFP. The complexity of fish poisoning makes it difficultto do clinical trials on HFP. Unless there is consensus of opinion on the clinical signsand symptoms of HFP, such trials are of little value. 2.4.5 Are paralytic and diarrhetic shellfish poisons involved? Clifford et al (1993) found low levels of paralytic shellfish poisons (saxitoxins) intoxic mackerel (0.02–1.30 µg saxitoxin equivalents/kg) by enzyme-linkedimmunosorbent assay (ELISA) and suggested that, possibly in combination withdiarrhetic shellfish poisons, they may be responsible for scombroid fish poisoning.These concentrations of saxitoxin equivalents corresponded to doses in the range0.04–1.0 ng/kg bw and were orders of magnitude less than the intraperitoneal and oralLD50 values in mice (10 and 263 µg/kg respectively). Calculations made following an outbreak of mussel poisoning in England suggestedthat doses as low as 9 µg saxitoxin equivalents/kg bw may cause illness in humans,but doses as high as 86 µg/kg bw are not necessarily fatal (references cited by Cliffordet al 1993). Nine of 23 mackerel samples associated with scombrotoxic incidentsapparently contained less saxitoxin than the negative control sample, and dose(saxitoxin equivalents) could not be correlated with the volunteer response. Dosesassociated with nausea/vomiting and/or diarrhoea ranged from 0.11 to 1.0 ng/kg bw,whereas doses not producing these symptoms ranged up to 0.5 ng/kg bw. Clifford etal (1993) concluded that paralytic shellfish poisons are at least partly responsible for'scombrotoxicosis'. However, the observation during volunteer testing of nausea andvomiting at doses orders of magnitude less than previously reported for saxitoxin,coupled with the presence of diarrhoea, which is not characteristic of paralyticshellfish poisoning, suggest that this is unlikely. It would appear that a clear definition of histamine/scombroid fish poisoning isrequired, based on the clinical signs/symptoms described by Taylor (1985) and Wu etal (1997) below. Many instances of fish poisoning are probably the result of a numberof unrelated toxins acting in concert. As such, they are not typical ofhistamine/scombroid fish poisoning, and should not be given that classification. Muchresearch is needed to sort out the many toxins in spoiling fish and the syndromes theycause. 2.4.6 Absorption of histamine from mouth and throat An unusual hypothesis (cited by Lange 1988) is that when affected fish are eaten thehistamine may be absorbed through the mucous membranes of the mouth and throat,thus bypassing the digestive process that destroys it. It is difficult to propose amechanism whereby a fish substrate would enhance the absorption of histaminethrough mucous membranes.

Histamine Fish Poisoning
23
2.5 Clinical characteristics and treatment 2.5.1 Clinical signs and symptoms Signs and symptoms of HFP occur from several minutes to several hours afteringestion of the toxic fish. The illness typically lasts a few hours, but may continue forseveral days. The primary signs/symptoms are those of histamine poisoning —cutaneous (rash, urticaria, oedema and localised inflammation), gastrointestinal(nausea, vomiting, diarrhoea), haemodynamic (hypotension) and neurological(headache, palpitations, tingling, burning, itching) (Taylor 1985; Wu et al 1997). Insevere cases there may be bronchospasm and respiratory distress (Shalaby 1996).
The most consistent clinical sign reported by Arnold and Brown (1978) was flushingof the skin of the face and neck, which caused a feeling of intense heat and generaldiscomfort. Because this striking erythema predominantly involves exposed areas, itresembles sunburn (Kim 1979). Gastrointestinal symptoms were experienced by fewerthan 25% of victims described by Arnold and Brown (1978). When present, theygenerally took the form of nausea and diarrhoea. However, when Gilbert et al (1980)reported on 150 patients affected in 30 separate outbreaks of 'scombrotoxic fishpoisoning' in Britain, in each outbreak the symptoms were similar, and diarrhoea waspredominant. Symptoms were recorded for each incident rather than individually, anda breakdown of patient numbers was not provided. The following table lists thenumber of outbreaks in which particular symptoms were noted. Signs and symptoms associated with scombrotoxic fish poisoning in 30 outbreaks (Gilbert et al 1980)
Sign or symptom No. of outbreaks Diarrhoea Hot flush/sweating Erythematous rash Nausea Headache Abdominal pain Palpitations Burning in mouth Vomiting Feverish Giddy or dizzy Tight chest Respiratory distress Facial swelling
24 15 14 13 13 6 6 5 4 3 2 1 1 1
Diarrhoea was also a prominent clinical sign in 77% of patients (second in frequencyonly to skin rash, which occurred in 82%) in ten incidents of HFP involving 22patients in South Africa (Müller et al 1992). Cape yellowtail (Seriola lalandii), a non-scombroid fish, was involved in all cases. Other signs were palpitations, headache,nausea and abdominal cramps, paraesthesia, an unusual taste sensation and breathingdifficulties. The patients responded well to antihistamine therapy and most hadrecovered within 12–24 h.

Histamine Fish Poisoning
24
Wu et al (1997) described two outbreaks of HFP that occurred in Taiwan in 1996 andwere confirmed by toxin analysis. In the first, caused by a non-scombroid fish, themain symptoms were facial flush, dizziness, headache, conjunctival hyperaemia andhypotension. In the second outbreak, in which a scombroid fish was involved,dizziness, diarrhoea, flushing and headache were the most common symptoms. Levelsof histamine and cadaverine in fish samples taken from the two outbreaks were 84 and8.5 mg/100 g (1st outbreak) and 272 and 23 mg/100 g (2nd outbreak) respectively. $Q[LHW\�LV�RIWHQ�PHQWLRQHG�DV�D�SURPLQHQW�V\PSWRP�RI�+)3��H�J��5XVVHOO�DQG�0DUHWLü1986; Specht 1998; Sabroe and Kobza Black 1998). A case of transient loss of visionin association with atrial tachycardia with a 2:1 atrioventricular block was alsoreported. Other signs and symptoms in this patient were typical of HFP, and responseto antihistamine therapy, including return of vision, was rapid (McInerney et al 1996;Clark 1997). In 1997, Ascione et al reported two cases of ’scombroid syndrome’ in Palermo, Italy,following ingestion of cooked tuna. Apart from symptoms and clinical signs typical ofHFP, both patients exhibited cardiovascular shock. This was associated withsubendocardial myocardial infarction in one case and acute pulmonary oedema withmyocardial ischaemia in the other. There appears to be no information on the effect of HFP in a woman on anembryo/foetus or breastfed infant. 2.5.2 Diagnosis A diagnosis of HFP depends on a history of fish eaten by the patient immediatelybefore onset of the illness. If the symptoms are typical of histamine poisoning, theonset time is short, and the patient has eaten a type of fish that has previously beenimplicated in cases of histamine poisoning, then a tentative diagnosis of HFP can bemade. The diagnosis can be confirmed by detecting high levels of histamine in thecontaminated food, meal remnants or in a similar fish obtained from the same source(Taylor 1986). Apart from being somewhat inconsistent, the symptoms of HFP are not definitive.Many can occur with other illnesses, both foodborne and non-foodborne. Whendiarrhoea is the predominant symptom, histamine may not be the main toxin involved.In addition, histamine poisoning is often confused diagnostically with food allergies.Identical symptoms occur, and antihistamines are equally effective in treating bothillnesses. However, histamine poisoning can easily be distinguished from food allergyon the basis of: the lack of a previous history of allergic reactions to the incriminatedfood; the high attack rate in group outbreaks; and the detection of high levels ofhistamine in contaminated food. With the fish that are commonly associated withHFP, histamine poisoning would be encountered much more frequently than allergicreactions. IgE-mediated allergic reactions could be detected by using skin prick testswith extracts of similar fish with known low histamine content (Taylor 1986).

Histamine Fish Poisoning
25
2.5.3 Treatment Symptoms of HFP rapidly subside after antihistamine treatment. H1 antagonists, suchas diphenhydramine or chlorpheniramine, are usually selected, but H2 antagonists suchas cimetidine may also be effective (Guss 1998). Induced emesis has also beenrecommended as a treatment (Downs 1997). Since the disease is self-limited, runninga short course, pharmacological intervention is not always necessary (Taylor 1986). 2.5.4 Clinical complications Serious complications are rarely encountered in cases of HFP. However, on rareoccasions, serious cardiac and respiratory complications occur in individuals with pre-existing cardiac and respiratory conditions (Taylor et al 1989). For example, RussellDQG�0DUHWLü��������GHVFULEHG�WKH�FDVH�RI�D�\RXQJ�FKLOG�ZLWK�D�KLVWRU\�RI�EURQFKLDOasthma, who suffered respiratory collapse after consuming fish contaminated withexcessive amounts of histamine. To determine whether kidney disease could exacerbate the symptoms of HFP,Helander et al (1965) investigated the renal removal of histamine in a person withseverely impaired renal function caused by chronic pyelonephritis. The percent renalextraction of injected [14C]-histamine from whole blood was reduced (to 51%)compared with the values of 74–86% obtained in healthy subjects.
2.6 Detection of histamine and other biogenic amines in fish 2.6.1 Detection of histamine-producing bacteria Many methods have been developed to detect histamine-producing bacteria in foodssuch as fish. Most of these detection systems are based on specific media that areselective and/or differential for histamine-producing bacteria (e.g. Niven et al 1981).Some methods require the preliminary isolation of bacteria so that individual isolatescan be assessed for histamine formation. One system utilises a histidine-fortifiedtrypticase soy broth that is optimised for bacterial histamine production (Taylor andWoychik 1982). Another utilises a tuna fish infusion broth (Omura et al 1978), butthis is a poorly defined medium and dependent on the availability of high quality rawtuna (Taylor and Woychik 1982). In these procedures, the histamine is extracted andmeasured by fluorometric assay. Another system for the detection of histamine-forming bacteria in fish is a differentialmedium containing bromocresol purple. Histamine formation by a colony causes acolour change due to a shift in pH. MRS broth fortified with histidine can be used inthe identification of histamine-producing lactobacilli. The histamine can be extractedand measured fluorometrically, or a procedure based on diamine oxidase andleucocrystal violet can be applied directly to the growth medium (references cited byStratton and Taylor 1991). However, there have been problems with identification of histamine-producingbacteria. Taylor et al (1978a) listed bacteria previously identified as histamineproducers, but found that evidence firmly linking some of the bacterial species on that

Histamine Fish Poisoning
26
list to histamine production was lacking. They considered that discrepancies could bedue to differences between strains within a particular species, growth media, oranalytical methodology used in the detection of histamine or HD. For example,Niven’s medium (Niven et al 1981) has been associated with false positive results.Garland (1985) attributed this to the non-specific alkaline products generated throughthe metabolism of tryptone and yeast extract by bacteria. McMeekin (1986) suggestedthat the incorporation of glucose at 0.1% in the medium could balance non-specificalkaline products. However, this medium still gave problems with M. morganii thatsuggested that the amount of acid formed from glucose produced by differentorganisms was different, depending on the bacterial glucose fermentation pathway.When 0.1% maltose was incorporated by Anggawati (1986), most false positiveresults were eliminated (Leung 1987). According to Taylor (1986), many early studies of bacterial histamine production werenot comparative, making it difficult to determine whether histamine production bybacteria was prolific or inconsequential. A reason for measuring histamine productionby bacteria rather than HD activity is that several bacterial species also producehistaminase, which may limit histamine accumulation by these species in foods(Ienistea 1971). Klausen and Huss (1987a) developed a method based on automated conductancemeasurements in a histidine-containing medium incubated at 25oC. The method israpid, with histamine-producing bacteria being detected within 24 h. Such a methodcould be used to screen large numbers of fish samples that may be contaminated withsignificant amounts of histamine-producing bacteria, and thus could be part of aquality assurance program (Klausen and Huss 1987a). However, the method has thedisadvantage of requiring expensive instrumentation (Stratton and Taylor 1991). An improved test for screening for histamine-producing bacteria is required, ideallyencompassing and distinguishing cadaverine- and putrescine-producing bacteria(James and Olley 1985). 2.6.2 Analysis of histamine Researchers in the field, as well as industry representatives, unanimously agree thatcanned tuna with high levels of histamine imparts a ’peppery’ feel to the mouth whenchewed, but tasting on a routine basis is not a feasible means of quality assurance(Etkind et al 1987). There is considerable evidence associating several othermeasurements of fish deterioration with histamine levels. For example, Cantoni et al(1976) compared histamine production with that of volatile acids and volatile bases intuna stored at 18–20oC. Toxic levels of histamine (>100 mg histamine/100 g fish)were noted after 4 days. During the same 4-day period, high levels of volatile acidsand bases were also noted. Hungerford et al (1997) developed a simple field test fordecomposition of seafood based on volatile bases. However, no objectivemeasurement, short of determining histamine concentration, has emerged as aneffective indicator of histamine levels. The classical method for determination of histamine was based on the fact that it willcause contraction of guinea pig ileum (Arnold and Brown 1978). The first application

Histamine Fish Poisoning
27
of this method to the analysis of histamine in fish was reported by Geiger (1944a). Heand fellow workers had earlier identified a biologically active substance in marine fishas histamine (Geiger et al 1944). Geiger (1944a) detailed how the bioassay could beapplied to fish and reported findings on histamine levels in raw and canned sardinesand mackerel. He pointed out that canning did not interfere with subsequent analysisfor histamine, and demonstrated the practical application of histamine determinationin canned tuna. Chemical testing is an effective means of detecting the presence in fish muscle.However, the validity of such testing depends on the design of the sampling plan. Forthis reason, chemical testing alone will not normally provide assurance that the hazardhas been controlled. Because histamine is generally not uniformly distributed in adecomposed fish, a guidance level of 50 ppm has been set (FDA 1998). If 50 ppm isfound in one section, there is the possibility that other sections may exceed thetoxicity level of 500 ppm (FDA 1998). Additionally, recent studies suggest that, if HDlevels are high, histamine formation can continue even in frozen storage (Price 1999). Two other problems remain. One is the lack of standardisation of histamine detectionmethodology, with numerous different methods being used around the world; and thesecond is that the presence of histamine alone is not necessarily a reliable indicator offish likely to cause HFP. Since the guinea pig ileum method of Geiger (1944a) for histamine in unprocessedand canned fish, many diverse analytical procedures have been published. Some of themore important include tests for histamine in:
• fish products: thin layer chromatographic method (Shultz et al 1976)• canned tuna: fluorometric method (Lerke and Bell 1976)• fish: fluorometric method (Taylor et al 1978b)• fish: enzyme-based screening test (Lerke et al 1983)• seafood: flow-injection method (Hungerford et al 1990)• canned fish: HPLC (Yen and Hsieh 1991)• fish: capillary electrophoresis (Mopper and Sciacchitano 1994)• tuna: copper chelation method (Bateman et al 1994)• fish: oxygen-sensor-based method (Ohashi et al 1994)• seafood: biological method (Association of Official Analytical Chemists, AOAC,
1995a)• seafood: chemical method (AOAC 1995b)• seafood: fluorometric method (AOAC 1995c)• seafood: modification of AOAC method (Rogers and Staruszkiewicz 1997). Historically, determination of histamine has been difficult due to the lack of rapidtesting methods. However, rapid testing methods are now available (Hungerford et al1997; Miller et al 1997; Price 1999). For example, commercial competitive ELISAkits are available for histamine detection, such as ALERTTM (sensitivity 5–50 ppm,qualitative, 2 h) and VeratoxTM (sensitivity <5 ppm, quantitative from 0–50 ppm, 1 h)from Neogen Corporation, USA (and Elisa Systems, Australia). There are also severalenzyme immunoassay test kits, including Histamarine Test Kit (sensitivity 0.5 ppm,

Histamine Fish Poisoning
28
quantitative from 1–500 ppm, 1 h) from Immunotech, France, approved by theAssociation of Official Analytical Chemists (AOAC) (Price 1999). In the ELISA kits, histamine is extracted from a sample in a quick water-extractionprocess. The extract is filtered, then diluted into a buffer solution prior to running onthe ELISA. These kits provide controls for rapid distinct visual colour results andeliminate the need for complicated chemical extraction of samples. They provide goodreproducibility and 'low' false negative and false positive rates when tested against theofficial AOAC fluorometric method for histamine (Anon. 1999; Rob Sherlock, ElisaSystems, pers. comm. 1999). However, at more than $10/test, they are still tooexpensive for routine testing of numerous samples on a quality-assurance orpreventive-testing basis. They are, however, suitable for disease outbreakinvestigatory studies. 2.6.3 Analysis of other biogenic amines and chemical quality index The assessment of the biogenic amine content in foods has received much attentionfor many years due to the possibility of using amine concentrations as an index offood quality, and has included various HPLC methods besides those for histamine(Mietz and Karmas 1977; Edwards et al 1987). Because histamine alone is not always useful as an indicator of fish quality, Mietz andKarmas (1977) established a chemical quality index of canned tuna for estimating theextent of decomposition in fresh tuna prior to canning. They used qualitative andquantitative HPLC to examine the relationship of dansyl derivatives of five amines(histamine, putrescine, cadaverine, spermine and spermidine) extracted from groundcanned fish. They prepared standards of pure putrescine, cadaverine, spermine andspermidine at concentrations of 1 mg/100 mL, and the level of each of the fivecompounds in canned fish samples was calculated on a ppm basis to generate an indexof tuna decomposition. Mietz and Karmas (1977) prepared and canned 48 cans of tuna ('authentic pack') asgood, borderline and decomposed; received 6 cans from the US FDA as authenticpack 'good', 'borderline' and 'decomposed'; and purchased 87 cans of commercialcanned tuna. The tuna included skipjack, yellowfin and albacore. All 141 cans wereevaluated organoleptically after opening and scored from 1 to 10, with the highestvalues representing the most decomposition. Subsequently the scores were translatedto good, borderline, and decomposed. The classification of borderline indicated initialstages of decomposition. Significant differences were seen between the good, borderline and decomposed tuna.Low levels of putrescine, cadaverine and histamine were typical of good tuna.Relatively high levels of spermine and spermidine were likewise found in good tuna.The borderline tuna showed a significant rise in the levels of putrescine, cadaverineand histamine, and generally decreased levels of spermine and spermidine. Indecomposed tuna, the putrescine, cadaverine and especially histamine levelscontinued to increase significantly, while the spermine and spermidine levels fell, withmany samples showing the absence of these two amines (thus necessitating a factor of1 in the denominator of the index below). Average values for histamine, putrescine,

Histamine Fish Poisoning
29
cadaverine, spermidine and spermine in ’authentic pack’ decomposed tuna were 253,2.5, 19, 0.7 and 1.9 ppm (or µg/g) respectively. Each of the five compounds per fish sample in ppm was submitted to the formula: Index = ppm histamine + ppm putrescine + ppm cadaverine 1 + ppm spermine + ppm spermidine Nominal cut-off values were established for the index scores to establish Class 1,Class 2 or Class 3 fish. The decomposition index followed an exponential value. Thevalues were Class 1, 0–1; Class 2, 1–10; and Class 3, >10. These nominal cut-offvalues were found to be effective in discriminating between the classes of fish, andappeared not to be dependent on species of fish or packing media. The resulting chemical index scores (see table below) compared favourably to'authentic pack' and organoleptic value scores. Organoleptic scores versus index scores for all samples
Index scores (141) Organoleptic scores(141) Class 1 (109) Class 2 (15) Class 3 (17)
Good (85) Borderline (36) Decomposed (20)
73a
25 11b
11 4a
0
1b
7 9a
% agreement (a) = 61% % disagreement (b) = 8.5% (Borderline samples classified either good or decomposed by either score were not included in the %agreement or disagreement.) Since it measures several different compounds resulting from several differentprocesses of decomposition, this method could be used as a chemical indicator fordecomposition of tuna. However, it is too complex to be used for routine screening. Wagener (1984) prepared dansyl derivatives of extracted amines from fresh andspoiled fish using the method of Hui and Taylor (1983) and examined them by HPLC.He confirmed the earlier findings, that levels of histamine, putrescine and cadaverinerise rapidly during the initial stages of fish spoilage. He also identified tryptamine intuna, hake and mackerel and tyramine in mackerel, but found that levels of these didnot change much during storage. Interestingly, there were unidentified peaks on thechromatograms, some increasing and some decreasing during spoilage. An unknownpeak marked 'X' showed an increase in both tuna and hake, and was very prominent incanned mackerel. Wagener (1984) speculated that the peak could be the substance'saurine' noted by Japanese workers in spoiling saury (Kawabata et al 1955).(However, Foo identified saurine as the phosphate salt of histamine in 1976.) Theunidentified compound was apparently not tested by Pauly's reagent, and may be thesame as Shewan's compound 8 (Shewan 1955) or Hughes' compound X (Hughes1959). It is interesting to note that Mietz and Karmas (1977) also found a dansylatedpeak (shoulder) between cadaverine and histamine in decomposed canned tuna, whichremained unidentified.

Histamine Fish Poisoning
30
In 1997, the US FDA published details of a gas chromatographic (GC) method forputrescine and cadaverine in seafood. The extracted diamines are converted tofluorinated derivatives, which are passed through solid-phase extraction columns andthe derivatives quantified by electron-capture GC after separation. Fourteenlaboratories analysed 14 samples of canned tuna and raw mahi-mahi (including blindduplicates and a spike) containing 0.2–2.6 ppm putrescine and 0.6–9.1 ppmcadaverine using the method. Recoveries ranged from 71 to 102% for putrescine andfrom 77 to 112% for cadaverine. AOAC International has adopted the GC method fordetermination of putrescine in canned tuna and cadaverine in canned tuna and mahi-mahi (Rogers and Staruszkiewicz 1997). More recently, an Italian group has developed an improved method for thesimultaneous determination of underivatised biogenic amines, cadaverine, putrescine,spermidine, histamine, tyramine and some amino acid precursors, histidine andtyrosine, in food products. The method is based on ion-exchange chromatography (IC)with integrated pulsed amperometric detection (IPAD). The method was usedsuccessfully for the analysis of biogenic amines and amino acids in food both ofvegetable and animal origin (pilchards) and in fermented foods, such as cheese andsalami. The main advantages of this method over a previous HPLC method coupledwith IPAD (Draisci et al 1993) are its application to a larger number of analytes andmatrices, a simpler extraction and clean-up procedure, and an improvedchromatographic separation and a lower detection limit. The IC-IPAD method issuitable for the detection of biogenic amines in a large number of samples and isparticularly useful for routine checks in repetitive analyses (Draisci et al 1998). 2.6.4 Analysis of urocanic acid Although he did not postulate that urocanic may be a candidate scombroid toxin,Baranowski (1985) suggested that urocanic acid may be a useful alternative tohistamine as a spoilage index in scombroid and other fish rich in endogenoushistidine. He used thin-layer chromatography to analyse for urocanic acid usingstandards in 95% alcohol. Morrison et al (1980) and Caron et al (1982) have usedHPLC methods.

Histamine Fish Poisoning
31
3. DOSE−RESPONSE ASSESSMENT
3.1 Definition of dose−response assessment
Kindred (1996) defined dose−response assessment as ’The determination of therelationship between the magnitude of exposure and the probability of occurrence ofthe health effects in question.’ Dose−response assessment is often termed hazardcharacterisation (Codex Alimentarius Commission 1996). The kind of topics that may be discussed under dose–response assessment include:
• the quantity of hazard in relation to the frequency and magnitude of adverseeffects;
• susceptibility of the general population and sub-populations;• food composition characteristics that affect host–hazard interaction;• special characteristics of the agent that affect host–pathogen interaction;• rates of intoxication;• morbidity and mortality rates (severity of the disease, sequelae);• human volunteer feeding studies;• epidemiological data;• animal data; and• clinical and laboratory studies. The data required in this area of risk assessment are largely quantitative rather thanqualitative, to complement the descriptive elements of HFP discussed under HazardIdentification. Because of the highly variable nature of the epidemiology and clinicalfeatures of the disease, this section is difficult to address. It has been approached bylooking at the incidence of HFP; fish characteristics that affect the clinical response;amount of bacterial contamination in relation to histamine; bacterial species involved;concentration of histamine and other biogenic amines in fish and the toxic dose;human factors that affect the clinical response; and morbidity and mortality rates.
3.2 Incidence of histamine fish poisoning 3.2.1 Background HFP occurs throughout the world and is perhaps the most common form of toxicitycaused by the ingestion of fish (Mines et al 1997). However, statistics on its incidencedo not exist. Many countries do not have adequate systems for reporting foodbornediseases. In those countries that do, cases are missed because of the often mild natureof the illness, which causes patients not to seek medical attention, or ignorance bymedical personnel, who may misdiagnose it as a food allergy. Moreover, scombroidpoisoning is not a reportable disease, even in those countries that keep records (Taylor1986). Under reporting of this illness is a worldwide problem (FDA 1992). Since 1970, the countries with the most reported incidents of HFP are Japan, the USand Great Britain, although this most likely represents better reporting by thesecountries. Less frequent outbreaks have been reported in various other countries

Histamine Fish Poisoning
32
including Canada, New Zealand, France, Germany, Norway, Sweden,Czechoslovakia, the Netherlands, Australia, Sri Lanka, Indonesia, South Africa andEgypt. Since only a few of these countries keep official records on incidents of fishpoisoning, it would be safe to assume that many incidents in these countries are notreported (Taylor 1986). 3.2.2 Africa Although no information was found on the incidence of HFP in Africa, Ababouch etal (1986) discussed histamine levels in commercially processed fish in Morocco,indicating that the problem has existed there. Morocco has a large fisheries industrybased in the Atlantic Ocean, with sardines the predominant fish landed. Fishmeal isthe main product, followed by canned sardines, tuna and mackerel and fresh or frozenfish. In 1986, worldwide sales of Moroccan fish products were falling, owing tocomplaints about the quality of canned products and several outbreaks of foodpoisoning in Europe implicating Moroccan products. In a survey of 248 samples ofcommercially processed fish, histamine concentrations ranged from <0.01 to694 mg/100 g. Ten samples (4%) had toxicologically significant levels of>50 mg/100 g. Tuna showed the highest percentage of samples (7%) with histaminelevels >50 mg/100g. Morocco has since developed a progressive strategy to meet European standards forfish products, including the use of mandatory in-plant Hazard Analysis and CriticalControl Point (HACCP)-based quality control systems (Ababouch 1997). 3.2.3 Asia HFP was recognised as a major cause of foodborne disease in Japan in the early1950s, and remains a major foodborne disease in that country. The first reportedincident involved 700 people, which remains one of the largest outbreaks on record.Forty-two outbreaks involving 4122 cases were reported by the Ministry of Health andWelfare, Japan, in 1970–80. Incriminated fish included mackerel, tuna, anchovies,sardines and marlin. The largest outbreak yet recorded in the world, involving 2656cases, occurred in Japan in 1973 from the consumption of dried horse mackerel(Trachurus japonicus). Given the Japanese preference for raw fish, it is surprising thatcooked fish have been involved in more incidents than raw fish. The utilisation ofonly the highest quality fish in the raw fish market of Japan is probably the reason forthis (Taylor 1986). HFP probably occurs frequently in Asia. High levels of histamine in fish on sale inAsian countries have been reported in various FAO Fisheries reports (references citedby Mitchell 1993). There are also reports of extremely high levels of histamine insome salted, and dried or salted, fermented products in Asia. Histamine in cannedproducts from Asia has also been a problem (James and Olley 1985). 3.2.4 Australia There is little in the scientific literature on outbreaks of HFP in Australia, whichindicates that the disease is not common in this country. Incidents have been reported

Histamine Fish Poisoning
33
associated with the consumption of tailor (Pomatomus saltatrix) (Taylor 1985),juvenile Australian salmon (Arripis truttaceus) (Smart 1992) and tuna of unknownsource (Brown 1993) (see below). The present authors are unaware of any recentmedia items on HFP. Information on surveys and monitoring presented here under’Histamine levels in fish products in Australia’ is reassuring, even if histamine is notthe only toxin involved, and indicates that HFP is unlikely to be prevalent inAustralia, at least in commercial fish products. 3.2.4.1 Outbreaks described by Smart (1992) Smart (1992) reported two separate outbreaks of HFP caused by consumption ofjuvenile Australian salmon (Arripis truttaceus), or ’salmon trout’, caught in SouthAustralian waters. The fish had been purchased from two different local Adelaide fishsuppliers. In the first outbreak, in March 1990, a family consisting of mother, child and infantwas affected. The fish was frozen within 1 h of purchase. Two weeks later, it waswashed and baked after partial defrosting at room temperature for 2–3 h and then in amicrowave oven. The mother, who noted that the fish had a peppery taste, requiredparenteral promethazine (an antihistamine drug) and overnight hospitalisation. The second outbreak, in March 1991, involved a family of three adults and one child.The fish was frozen within half an hour of purchase. Before baking 1 week later, itwas defrosted in a refrigerator for 12 h. A 35-year-old man and a 69-year-old womanrequired parenteral antihistamines, but were not hospitalised. Another woman and an8-year-old boy did not require treatment. In all cases, onset of symptoms occurred within half an hour of fish ingestion. Theyincluded erythema and urticaria of the skin, facial flushing and sweating, palpitations,hot flushes, headache, nausea, vomiting and dizziness. The patients in each outbreakhad minimal gastrointestinal symptoms. The infant, who consumed only oneteaspoonful of fish, was least affected of the seven patients, exhibiting only a transienterythematous rash. The following table summarises the clinical signs and symptoms in the seven patientsinvolved in the two outbreaks described by Smart (1992). Signs and symptoms associated with HFP in seven patients (Smart 1992)
Sign or symptom No. of patients Erythema and urticaria Facial flushing/sweating Palpitations Hot flush of body Headache Nausea Vomiting Dizziness Peppery taste
7 5 5 3 3 2 1 1 1

Histamine Fish Poisoning
34
The diagnosis of scombroid poisoning was made both clinically and on thedemonstration of high histamine levels in the cooked fish (80 mg/100 g and254 mg/100 g in the first and second outbreaks respectively). This indicates grosstemperature abuse. 3.2.4.2 Cases described by Brown (1993) Brown (1993) reported two cases of HFP in patients attending a suburban generalpractice in Brisbane on one afternoon. The first patient was a 35-year-old nurse, whopresented with a florid flush of her entire skin surface. She complained of a poundingheart, tightness of the chest and a feeling of doom. She had eaten tuna at a restaurant2 hours before. Her symptoms had begun about 75 min before presentation andsteadily increased in intensity, in association with an extreme sense of alarm. She hadno itch, nausea or respiratory compromise. The woman was treated with 50 mgpromethazine by intramuscular injection. After 15 min, the erythema began to wane,disappearing over the next 10 min. The patient said she felt much better. After afurther 10 min, loose bowel motions began, and continued intermittently for the next45 min. The second patient reported by Brown (1993) was a 31-year-old man, who had eatenthe same tuna dish. His symptoms were a hot sensation, nausea, diarrhoea, shakes andheadache. A third person who had eaten the dish was reported as having similar, butless intense, symptoms. 3.2.5 Canada In Canada, HFP was first confirmed in 1975, although there were probably incidentsbefore then. In the 1970s and 1980s, canned tuna was the most likely vehicle ofcontamination, but between 1990 and 1995 it was fresh tuna, mahi-mahi or marlin.The three largest outbreaks were caused by the ingestion of smoked mackerel(October 1987: 14 cases), fresh marlin (July 1991: 12 cases) and fresh tuna (August1994: 12 cases). Histamine levels of 28–710 mg/100 g were implicated in sevenseparate incidents involving tuna imported from Sri Lanka. Most incidents in Canadahave been associated with imported fish (Todd et al 1992; Todd 1997). 3.2.6 Europe An early outbreak of HFP in Great Britain occurred among a group of British sailorsaboard the Triton of Leith in 1828 (Henderson 1830). Five crew members became illwith symptoms of severe headache, dilated blood vessels, oedema, facial swelling andflushing, swelling of the entire body and shivering following the consumption ofbonito. Bonito is a scombroid fish and a likely vehicle for HFP. After this earlyincident, reports of HFP in Great Britain were scarce or non-existent until 1976. From1976, numerous outbreaks have been reported in England, Wales and Scotland,associated with mackerel, tuna, sardines, herring and pilchards (e.g. Gilbert et al 1980;Bartholomew et al 1987). Scoging (1991) reported on incidents of scombrotoxic fish poisoning in the UnitedKingdom between 1976 and 1990. There were 441 suspected incidents involving 962

Histamine Fish Poisoning
35
cases. Fish associated with 167 incidents were confirmed as having histamine levelsof >5 mg/100 g. Between 1987 and 1996, ’fresh’ tuna and canned tuna became increasingly associatedwith incidents of HFP. Scoging (1998) investigated 405 incidents during this period:243 sporadic incidents (60%), 105 general outbreaks (26%) and 56 family outbreaks(14%). In 148/405 incidents (37%), food remnants contained histamine at levels>5 mg/100 g, and 117/148 (79%) had histamine levels >100 mg/100 g. (Histaminewas detected using HPLC.) Of the 257 incidents not confirmed by elevated histamine levels, 61% (156) wereassociated with atypical symptoms or with fish not commonly associated with thissyndrome (e.g. shellfish, crustaceans, trout, haddock, skate, shark), or directlyimplicated remnants were unavailable for analysis. In the remaining 101 (39%),symptoms and onset times were typical, but the levels of histamine found in remnantswere within acceptable limits. The latter would suggest uneven distribution ofhistamine in contaminated fish or that unidentified agents or toxins may contribute tothe syndrome (Scoging 1998). Tuna (fresh/frozen and canned) and mackerel were most commonly implicated in bothsuspected and confirmed (elevated histamine) scombrotoxic incidents. Salmon wasinvolved in 30 suspected incidents, but remnants from only one incident containedslightly elevated histamine levels (5.1 mg/100 g). Salmon is therefore of particularinterest as a vehicle for other possible causative toxins. Bacterial contaminationoccurred both during harvesting and processing and during food retail and preparation(e.g. after canned tuna was opened) (Scoging 1998). Sockett (1991) reported on food poisoning outbreaks associated with manufacturedfoods in England and Wales between 1980 and 1989. Thirty-five outbreaks associatedwith processed fish and shellfish were reported during this period, including twooutbreaks caused by imported products. Scombrotoxin poisoning was the mostcommon cause of illness, and gave rise to 20 outbreaks affecting 59 people. Eachoutbreak affected two or three people. All outbreaks were associated with mackerelproducts, including smoked mackerel (18), mackerel paté (1) and filleted mackerel(1). The symptoms were those of histamine poisoning. Molinari et al (1989) reviewed the hygiene and health importance of histaminetoxicity in Italy and reported that the problem is widely underestimated in thatcountry. Tuna and mackerel were the fish most commonly associated with toxicity.An outbreak of HFP occurred in Palermo in 1979 that affected 'nearly 250' people. In 25 years (1966–91), 76 poisoning incidents after intake of tuna were reported to theSwiss Toxicological Information Centre (Maire et al 1992). Twenty-seven reportscame from physicians and, of these, 18 fulfilled the criteria of scombroid fishpoisoning.

Histamine Fish Poisoning
36
3.2.7 New Zealand From 1973 to 1975, several incidents of histamine poisoning occurred in NewZealand. The outbreaks were associated with consumption of canned mackerel,smoked kahawai, kingfish and trumpeter fish. In addition, New Zealand authoritiesassisted in the investigation of an outbreak in 1974 involving canned skipjack tuna inthe Solomon Islands (Taylor 1985). In a report on HFP prepared in 1993 for the New Zealand Ministry of Health (Mitchell1993), survey data from 1992–93 on histamine levels in (mostly smoked) fish werepresented, together with a summary of recent (1990–93) HFP outbreaks in NewZealand. Nineteen outbreaks were reported between March 1990 and June 1993, allresulting from the consumption of smoked fish, including kahawai and mackerel.Histamine levels in the incriminated fish ranged from 20 to 500 mg/100 g. Mitchell's(1993) report concluded that, because of the number of poisoning incidents and thenumber of samples exceeding the acceptable level of histamine in retail smoked fish,further monitoring and investigations of the causes of high levels of histamine in fishwere warranted. 3.2.8 United States It is difficult to determine the true incidence of HFP in the US, because of incompleterecord keeping. Local health departments recommend, but do not require, that casesbe reported. Statistics on the disease are also inaccurate because of misdiagnosis(Lange 1988). Data compiled by the CDC in the mid-1970s indicated no particularseason for HFP and suggested Hawaii and California as the states with largest numberof outbreaks (Hughes et al 1977). In 1980, histamine poisoning was one of the mostprevalent forms of foodborne disease of chemical aetiology in the United States,ranking only behind ciguatera fish poisoning (Sours and Smith 1980). By 1997,scombroid toxicity or HFP was the most prevalent form of seafood-borne disease inthe United States — Lipp and Rose (1997) reported outbreaks of seafood-bornedisease associated with fish, 1983–92, and listed the aetiological agents as scombroid(57% of outbreaks), ciguatoxin (19%), bacteria (14%), unknown (9%) and chemicals(1%). In contrast to the Japanese situation, most of the US outbreaks have involved asmall number of people, typically fewer than five individuals. Between 1970 and 1974, 'scombroid fish poisoning' was responsible for 29 (43%) ofthe 68 outbreaks caused by fish and shellfish toxins. In 45% of outbreaks, laboratorytests demonstrated elevated histamine levels in the incriminated fish. Tuna and mahi-mahi were most frequently implicated. The place where fish were mishandled, leadingto the outbreak, was identified in 10 outbreaks. Errors leading to contamination of thefood were reported in 18 outbreaks, and improper storage or holding temperature wasidentified in 16 (89%) of these outbreaks (Hughes et al 1977). In 1970, some 40 children in a school lunch program became ill after eating importedcanned tuna. A larger outbreak of HFP occurred in 1973, involving commerciallycanned tuna (Merson et al 1974). There were 254 clinical cases in eight states. Thesymptoms included immediate oral burning and blistering, followed in 30–45 min byheadache, abdominal cramps, diarrhoea and flushing. No cases required

Histamine Fish Poisoning
37
hospitalisation, and the duration of illness was generally 8 h or less. Nine assays forhistamine produced values ranging from 68 to 280 mg/100 g, with controls at3 mg/100 g. FDA recalled the incriminated lots, which showed evidence ofhoneycombing, indicating advanced decomposition (Merson et al 1974; Arnold andBrown 1978). This outbreak is probably still the largest on record in the United States,and it signalled an increasing awareness of histamine poisoning in that country(Taylor 1986). In 1979–80, more than 200 people became ill after consumingimported frozen mahi-mahi. Other incidents of intoxication in the United States have resulted from theconsumption of canned abalone-like products, canned anchovies, and fresh and frozenamberjack, bluefish (tailor), sole and scallops. In particular, shipments of unfrozenfish packed in refrigerated containers have posed a significant problem because ofinadequate temperature control (FDA 1999). 3.3 Fish characteristics that affect the clinical response 3.3.1 Species Fish that cause HFP include mackerel (Scomber spp.), tuna (Thunnus spp.), saury(Cololabis saira) and bonito (Sarda app.) (scombroid fish); and mahi-mahi(Coryphaena spp.), sardines (Sardinella spp.), pilchards (Sardina pilchardus),anchovies (Engraulis spp.), herring (Clupea spp.), marlin (Makaira spp.) and bluefish(Pomatomus spp.) (non-scombroid fish) (Taylor 1986; Institute of Medicine 1991).Non-scombroid species Australian salmon (Arripis truttaceus), sockeye salmon(Oncorhynchus nerka) and Cape yellowtail (Seriola lalandii) have also beenimplicated in HFP (Smart 1992; Müller et al 1992; Gessner et al 1996). As non-scombroid fish can cause HFP, the term scombroid fish poisoning is a misnomer(Taylor 1988). In the USA, of 73 outbreaks of HFP reported to the Centers for Disease Control from1978 to 1982, 42% implicated mahi-mahi as the vehicle. An epidemic in Alabama andTennessee in 1986 involved amberjack (Seriola spp.), a non-scombroid species.Bluefish have been responsible for many cases of HFP along the eastern seaboard ofthe US (Karolus et al 1985; Etkind et al 1987; Lange 1988). In New Zealand, incidents involving smoked kahawai, kingfish and canned mackerelin the early 1970s were reported and discussed by Foo (1975a,b, 1976, 1977). Otherreports involved kingfish and smoked kahawai (Mitchell 1984) and smoked mackerel(Mitchell and O'Brien 1992). Mitchell (1993) gave a summary of recent casesinvolving smoked kahawai, smoked marlin, smoked mackerel and smoked trevally. Australian cases were caused by improperly stored tailor or bluefish (Pomatomussaltatrix) (Taylor 1985); juvenile Australian salmon (Arripis truttaceus) (Smart 1992);and tuna (Brown 1993).

Histamine Fish Poisoning
38
3.3.2 Intrinsic potential for histamine and cadaverine accumulation Since free histidine in fish muscle is the substrate for microbial decarboxylation toproduce histamine, species difference in histidine content has a large effect on thepotential hazards of poor handling practices (Pan 1985b). Although the muscle offatty, red-meat, active and migratory species contains much more free histidine thanthat of white-meat, slower species, the latter still contain combined histidine as acomponent of muscle protein. In the active fish, the free amino acid may act as abuffer, protecting the tissues from the effects of sudden increases in lactic acid.Histidine deaminase [HAL] and urocanase, which mediate the first two steps of thedegradative process, regulate the level of histidine in the live fish (Love 1980). Arnold and Brown (1978) cited a series of papers dealing with the freshness of fishand the amount of histamine present. They discussed factors such as the effect of pHand temperature on the amount of histamine produced in ’red-meat fish’ such asmackerel and ’white-meat fish’ such as rockfish. The terms ’red-meat fish’ and ’white-meat fish’ are based on the general surface appearance of the fish. The terms ’redmuscle’ and ’white muscle’ refer to muscle types within individual fish, red musclesbeing those containing predominantly red fibre types and white muscles being thosecontaining predominantly white fibre types. All scombroid fish contain both muscletypes in varying amounts, depending on species. In the commercial processing of tuna,only white muscle is canned for human consumption, the red muscle being usedprimarily for pet food (Arnold and Brown 1978). Takagi et al (1969) examined the amounts of histidine and histamine in 21 species ofaquatic animals during spoilage and found that more histamine was produced in red-meat fish such as mackerel than in white-meat fish such as rockfish. Suyama andYoshizawa (1973) found relatively high free histidine contents in 13 species ofmigratory red-meat fish (286–1460 mg/100 g), whereas white-meat fish were low infree histidine (0–38.2 mg/100 g) and did not produce histamine during spoilage.However, within the same species of fish, more histidine was found in white than inred muscle, and the resulting histamine formation followed this pattern with regard tomuscle type; i.e. more histamine in white muscle (Takagi et al 1969; Orejana et al1983; Pan 1985b). The following table illustrates the results of random sampling offish from a fish market for histamine content of red and white muscle. Histamine content in the white and red muscle of scombroid fish (after Pan 1985b)
Histamine (ppm) Common name Scientific name White muscle Red muscle
Spotted mackerel Frigate mackerel Oceanic bonito
Scomber australasicus Auxis thazard Euthynnus affinis
25 45 80
3 6 35
Klausen and Lund (1986) set up an experiment to demonstrate the differences in theamine formation between a scombroid and a non-scombroid fish. They chosemackerel (Scomber scombrus) and herring (Clupea harengus), because in Denmarkthe methods of catching, handling and manufacturing of the two species are verymuch alike, but only mackerel causes HFP. The fish were stored in refrigerated tanks

Histamine Fish Poisoning
39
or ice until they arrived at the laboratory and were eviscerated, vacuum packed andstored. When stored in vacuum packs at 2 or 10oC, mackerel and herring accumulatedsimilar amounts of histamine (herring, 4.2 and 10.9 mg/100 g; mackerel, 4.3 and11.4 mg/100 g respectively) at the time of rejection, which was assessed by sensorymeans by a panel of six. With cadaverine, the situation was different. In herring, only small amounts wereformed (0.7 mg/100 g at 2oC and 4.9 mg/100 g at 10oC). In mackerel, high contents ofcadaverine rapidly accumulated 20 mg/100 g at 2oC and 22.6 mg/100 g at 10oC(before rejection). These values for mackerel correspond to about 2–5 times thehistamine contents. Thus the levels of cadaverine were high enough in relation tohistamine to potentiate the oral toxicity of histamine as described in guinea pigs byBjeldanes et al (1978), and may explain why mackerel are much more frequentlyinvolved than herring in HFP incidents. In addition, according to Klausen and Lund (1986) the contents of free histidine andfree lysine are 4–5 times higher in fresh mackerel compared with fresh herring. Whenexposed to high temperatures (20–30oC) and contamination with histidine-decarboxylating bacteria, mackerel therefore has the capacity for accumulation ofmuch higher levels of histamine and cadaverine than herring. Gilbert et al (1980)examined 37 samples (almost all of them mackerel) involved in HFP and found>100 mg/100 g of histamine in 22/37. In herring, on the other hand, only moderatelytoxic levels of histamine (50 mg/100 g) could be formed from the free histidinepresent in the fresh fish (Klausen and Lund 1986). Cadaverine is more frequently found in spoiled fish than is histamine. The amountvaries, but levels of 10–60 mg/100 g have often been found. Putrescine levels inspoiled fish are usually much lower than those of cadaverine, usually <10 mg/100 g,probably because of the limited quantities of ornithine in fish tissues (Taylor andSumner 1986). While the significance of cadaverine and putrescine as potentiators of histamineintoxication has not been fully established, it could be expected that if they are presentat ratios to histamine higher than those normally found in spoiled fish, they mayinhibit the detoxification of histamine. They may also act in an additive or synergisticfashion with other, as yet undiscovered, potentiators. 3.3.3 Seasonal and other variability Whereas data compiled by the CDC in the mid-1970s indicated no particular seasonfor HFP (cited by Lange 1988), there seems to be a definite connection betweenseason, temperature, histidine content and histamine formation in fish products (Pan1985b). Free histidine content in herring varied with seasons from 260 to 1600 mg/kg,being highest in summer (Hughes 1959). Less histamine was found in sardines andbonito during winter than in other seasons under the same handling conditions(Simidu and Hibiki 1954). Skovgaard and Elleman (1978) reported that confirmedhistamine poisoning outbreaks caused by smoked mackerel occurred during latesummer, when both fish and water have the highest temperature, and the mackerel areat their fattest.

Histamine Fish Poisoning
40
Clifford et al (1989) noted that mackerel show quite marked fish-to-fish and seasonalvariations in chemical composition and susceptibility to spoilage by variousmicrobiological, enzymic and auto-oxidative processes. The authors concluded that itis possible that only certain fish in an apparently homogenous batch will develop allfactors necessary to induce HFP. 3.3.4 Parts of fish consumed Concentrations of histamine in fish tend to vary widely from one part of the fish toanother. Histamine in raw fish is usually present at higher levels in tissue adjacent tothe gills or the intestines, the main reservoirs of the histidine decarboxylating bacteria(HDB) (Lerke et al 1978; Taylor 1986). Frank et al (1981) observed a histaminegradient in spoiled skipjack tuna, with highest concentrations near the gut cavity in theanterior section, and progressively lower concentrations moving posteriorly. Orejanaet al (1983) found that during 15-day iced storage of skipjack tuna, the tail sectionshowed the lowest histamine content throughout the period. Hardy and Smith (1976)showed that the histamine content of ungutted mackerel was 10 times more than thatof gutted fish after storage at ambient temperature for 140 h. 3.3.5 Nature and amount of bacterial contamination in spoiling fish 3.3.5.1 The common histamine-producing bacteria Although only about 1% of the surface microflora of live fish represents histamineproducers (Kimata 1961), HDB form a greater proportion of the microbial populationas a fish spoils. Omura et al (1978) reported that 31% of isolates from spoiledskipjack tuna and jack mackerel growing at warm temperatures were histamineproducers, and Yoshinaga and Frank (1982) found 13.4% in decomposing skipjacktuna. Frank and Baranowski (1984) found with mahi-mahi that 7% of isolates growingat warm temperatures and 9% growing at refrigeration temperatures were histamineproducers. Taylor et al (1978a) identified 112 species of bacteria that are known to possess HD.Most were members of the family Enterobacteriaceae, or the genera Clostridium andLactobacillus. Enteric bacteria, specifically Morganella morganii, certain strains ofKlebsiella pneumoniae and a few strains of Hafnia alvei are the most prolifichistamine producers in fish when they are maintained at temperatures greater than 4oC(Stratton and Taylor 1991) and are the most commonly associated with scombrotoxicfish (Taylor 1983). Certain strains of Lactobacillus that are also prolific histamineproducers are probably of importance only in fermented fish (Taylor 1986). Niven et al (1981) identified Vibrio spp. as histamine producers, and histamine-producing isolates of V. alginolyticus have been found in decomposing skipjack tunaand mahi-mahi (Frank et al 1985). V. harvei, V. fisheri and Photobacterium leiognathiare capable of producing histamine at warm temperatures (Ramesh and Venugopalan1986). Other Photobacterium spp. and Vibrio spp. may be primarily responsible forhistamine production in scombroid fish at lower temperatures (Morii et al 1986; VanSpreekens 1987; Leung 1987).
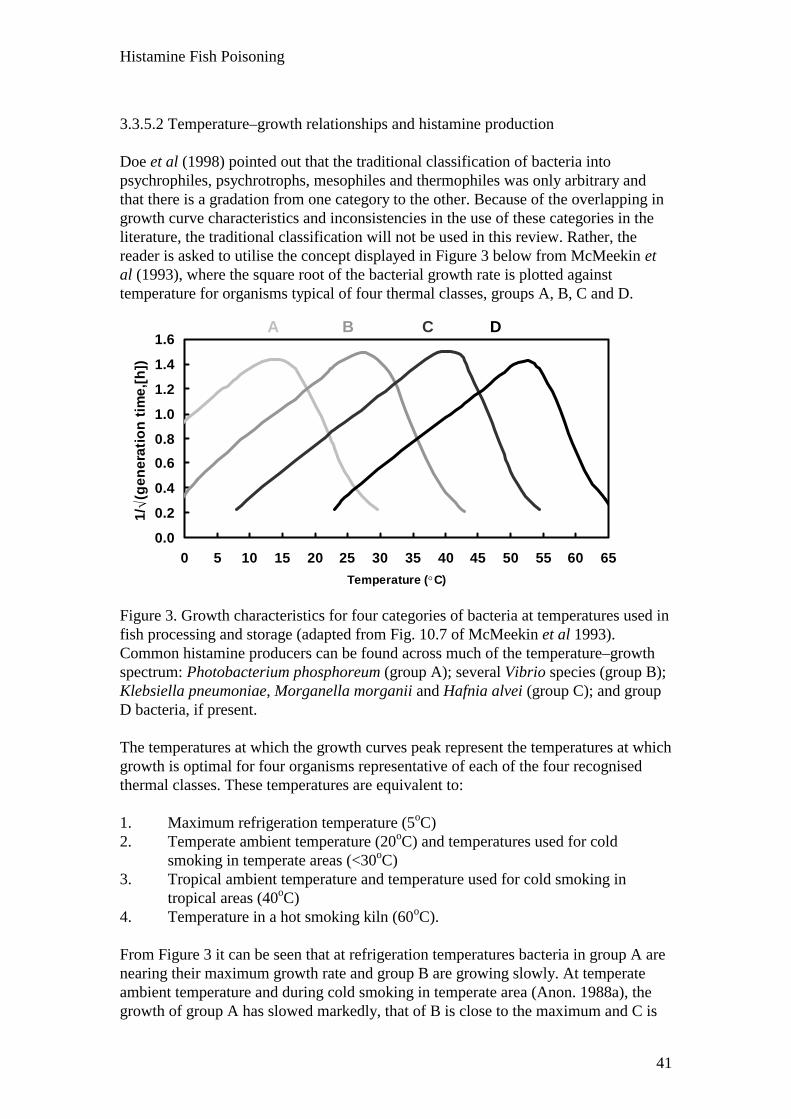
Histamine Fish Poisoning
41
3.3.5.2 Temperature–growth relationships and histamine production Doe et al (1998) pointed out that the traditional classification of bacteria intopsychrophiles, psychrotrophs, mesophiles and thermophiles was only arbitrary andthat there is a gradation from one category to the other. Because of the overlapping ingrowth curve characteristics and inconsistencies in the use of these categories in theliterature, the traditional classification will not be used in this review. Rather, thereader is asked to utilise the concept displayed in Figure 3 below from McMeekin etal (1993), where the square root of the bacterial growth rate is plotted againsttemperature for organisms typical of four thermal classes, groups A, B, C and D.
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
0 5 10 15 20 25 30 35 40 45 50 55 60 65
Temperature (�C)
1/»
(gen
erat
ion
tim
e,[h
])
A B C D
Figure 3. Growth characteristics for four categories of bacteria at temperatures used infish processing and storage (adapted from Fig. 10.7 of McMeekin et al 1993).Common histamine producers can be found across much of the temperature–growthspectrum: Photobacterium phosphoreum (group A); several Vibrio species (group B);Klebsiella pneumoniae, Morganella morganii and Hafnia alvei (group C); and groupD bacteria, if present. The temperatures at which the growth curves peak represent the temperatures at whichgrowth is optimal for four organisms representative of each of the four recognisedthermal classes. These temperatures are equivalent to: 1. Maximum refrigeration temperature (5oC) 2. Temperate ambient temperature (20oC) and temperatures used for cold
smoking in temperate areas (<30oC) 3. Tropical ambient temperature and temperature used for cold smoking in
tropical areas (40oC) 4. Temperature in a hot smoking kiln (60oC). From Figure 3 it can be seen that at refrigeration temperatures bacteria in group A arenearing their maximum growth rate and group B are growing slowly. At temperateambient temperature and during cold smoking in temperate area (Anon. 1988a), thegrowth of group A has slowed markedly, that of B is close to the maximum and C is

Histamine Fish Poisoning
42
growing slowly. At tropical ambient temperature and temperatures used for coldsmoking in tropical areas (Chng and Kuang 1987), group C is almost at its optimumgrowth rate, B is almost at its upper limit of temperature, and D is beginning to growwell. In temperatures in the hot smoking kiln (Anon. 1988b), bacteria in groups A, Band C have been eliminated, but 60oC has not destroyed group-D bacteria,representing the thermophile group, if there are any present. Each bacterial strain has a notional temperature (Tmin) at which growth is zero on thetemperature axis; a temperature at which growth is most rapid (Topt); and atemperature at which thermal death reduces the growth rate to zero (Tmax). Cardinaltemperatures predicted from data for a wide range of organisms were listed byMcMeekin et al (1993). From these data, it is evident that there are no clear-cutdistinctions between the temperature groups, but rather a continuum of Tmin values. Itis interesting to note the lower limits for production of toxicologically significantlevels of histamine in tuna fish infusion broth for some common HDB were 7oC for K.pneumoniae, 15oC for two M. morganii strains, and 30oC for H. alvei, Citrobacterfreundii and E. coli (Behling and Taylor 1982). Because of the importance of temperature in the production of histamine, severalattempts have been made to predict histamine formation in spoiling fish at differenttemperatures. For example, Frank (1985) constructed normographs to predicthistamine production in skipjack tuna, and Pan (1985a) used Arrhenius plots for suchestimations in mackerel and bonito. However, the methods involved too manyassumptions and were inaccurate in their predictions. A further development,temperature function integrators, which take temperature fluctuations into account,were also considered unsuitable for predicting the complicated process of histamineproduction in spoiling fish (Olley and McMeekin 1985). It is clearly not possible to relate the amount of bacterial contamination to histamineproduction by such methods. Numerous factors introduce bias. First, a reasonableestimate of the initial numbers of significant bacteria needs to be made. Thenhistamine concentration represents the amount produced by potentially diversepopulation of bacteria with different capacities for histamine production and/ordestruction in association with bacteria that are not histamine producers or destroyers.Even when one group or strain of HDB predominates, histamine production isunlikely to follow a growth curve fitting that group/strain, not least becausemetabolism of histamine by histaminase may be taking place simultaneously. Further,the amount of histamine production varies from site to site within a fish, and substratechanges such as proteolysis at high temperatures affect the amount of histidineavailable for histamine formation. Subsequent cooking or processing of a spoiled fish will further alter the relationshipbetween bacterial numbers and histamine production by reducing or removing themicrobial population without affecting the histamine content significantly. Forexample, Fletcher et al (1995) reported that, when whole fresh kahawai (Arripistrutta) was stored under a variety of temperature regimes, elevated levels of histamine(defined as ≥50 mg/kg) occurred only when aerobic plate counts at 20 or 35oCexceeded 106 colony forming units (CFU)/g. There was only one sample with elevatedhistamine where the aerobic plate count was <107 CFU/g. When such fish were

Histamine Fish Poisoning
43
smoked, bacterial numbers, but not histamine concentration and toxicity, wereconsiderably reduced (Fletcher et al 1998). 3.3.5.3 Studies on variability in species/strain composition of microflora Scientists have found that factors affecting the dominating species/strains of HDB inthe microflora of a particular spoiling fish are many and varied. The diversity can beattributed to differences in the species of fish, handling procedures, holding times andtemperatures. In addition, the character of the microflora can be influenced by thefish’s feeding habits, geographical location, the season, ocean temperature etc.(Kimata 1961; Shewan 1977; Yoshinaga and Frank 1982). It is not surprising thatresults have been variable in many studies on the effect of storage temperature ofhistamine formation in fish (Behling and Taylor 1982).
HDB that grow at refrigeration temperatures usually produce histamine in smallerquantities than those produced by species that grow at warmer temperatures, and toxiclevels of histamine may not be reached. Okuzumi et al (1984a) looked at theoccurrence of various histamine-forming bacteria on/in fresh fish purchased from fishmarkets in Japan. For the summer samples, M. morganii was found in the greatestnumbers, followed by the ’N-group’ bacteria, later identified as Photobacteriumphosphoreum by Fujii et al (1997). For the winter samples, only N-group bacteriawere detected. This fits in with the observation of Simidu and Hibiki (1954), whofound less histamine in sardines and bonito during winter than in other seasons underthe same handling conditions. In a subsequent study, Okuzumi et al (1984b) studied microbial populations at variousstorage temperatures in mackerel bought at fish markets, and correlated these withhistamine production. In fish samples stored at 5oC and 10oC, the main histamineformers were N-group bacteria, which reached levels of 107–108/g after 7 days. Insamples stored at 15oC, the formation of histamine was still attributed mainly to N-group bacteria, particularly in the first 3 days, although numbers of M. morganiiincreased throughout. In samples stored at 20oC, a great part of histamine at the earlypart of storage (0–4 days) was attributed to N-group bacteria, and at a later stage (6–8 days) to M. morganii. However, at the final stage (11–14 days), histamine contentdecreased from >700 mg/100 g, and remained constant at about 500 mg/100 g. Insamples stored at 25oC and 30oC, M. morganii was the main histamine producer. Ryser et al (1984) obtained 60 isolates of indigenous bacteria from raw tuna fish, andidentified them as Pseudomonas spp. Fewer than half (35%) the isolates producedhistamine during incubation at 21oC for 48 h, and the maximum amount produced bya single isolate was 3.2 mg/100 mL, far less than the 50 mg/100 g believed necessaryto induce symptoms of HFP. This would suggest that bacteria that grow at lowertemperatures are not significant toxicologically. However, Okuzumi et al (1981)reported significant histamine production by halophilic (salt-loving) N-group bacteriagrowing at 5oC (300–400 mg histamine/100 mL of mackerel homogenate). Husain (1996) estimated the growth rates of different HDB such as Vibrio spp.,Bacillus spp. and Morganella spp. incubated at 28oC in seawater broth (75% sterile

Histamine Fish Poisoning
44
seawater, 0.5% peptone and 0.5% yeast extract) at different salt concentrations. Atsodium chloride concentrations ranging from 1% to 10%, a visible growth was seen inall cultures. Maximum growth occurred at 4%, 3% and 7% sodium chloride for Vibriospp., Bacillus spp. and Morganella spp. respectively. Husain (1996) also found that traditional Indian preservatives garlic, turmeric, gingerand black pepper all had a profound inhibitory effect on the growth of HDB isolates atconcentrations of 1% to 5%. There was a large decrease in bacterial growth rate asconcentrations increased. Garlic was the most effective, completely inhibiting growthof Vibrio spp. at 4% concentration and Bacillus spp. and Morganella spp. at 5%concentration. Earlier, workers in Japan (Wendakoon and Sakaguchi, 1992) had tested the effects ofpowdered spices and their ethanol extracts on growth and biogenic amine formation ofEnterobacter aerogenes and M. morganii. Clove and cinnamon were the bestinhibitors of the spices tested (allspice, cardamom, chilli, cinnamon, clove, cumin,black pepper, nutmeg, sage and thyme). 3.3.5.4 Species/strain variation in histidine decarboxylase activity Different bacteria vary significantly in the quantity of histidine decarboxylase (HD)they produce and/or the specific activity of their decarboxylases. According toFerencik (1970), M. morganii (then P. morganii) required histidine levels of 100–200 mg/100 g to induce the production of HD. By contrast, H. alvei decarboxylatedhistidine when it was present at <50 mg/100 g. This suggests that bacteria differaccording to their relative importance as histamine producers on different species offish. Eitenmiller et al (1981, 1982) investigated factors influencing HD production by M.(then Proteus) morganii. Examination of 22 strains of M. morganii revealed that eachpossessed HD activity, although at variable levels. With one high-activity strain(GRMO 6), maximal HD activity occurred at 37oC and pH 6.5. Normal muscle pH offresh yellowfin tuna (6.5) thus corresponds closely to the pH required for optimalactivity of HD, and is low enough to permit rapid enzyme synthesis by M. morganii.Minimal enzyme activity was present when the culture was grown at pH 8.5. HDactivity decreased as the age of the culture increased. Rapid enzyme and histamineformation occurred in yellowfin tuna fillets inoculated with M. morganii and stored at24 and 30oC. Histamine soon reached toxic levels of 520 mg and 608 mg/100 g at 24and 30oC respectively. Little enzyme activity was present in inoculated fillets stored at15oC, and in control fillets that were not inoculated. Olley and Baranowski (1985) pointed out that low-temperature enzyme activity bymicroorganisms that grow at warm temperatures is important in histamine formation,provided sufficient bacterial numbers have been reached before cold storage. Klausenand Huss (1987b) studied growth and histamine production by M. morganii inhistidine-containing broth and in mackerel. Following storage at higher temperatures(10–25oC), large amounts of histamine were formed at low temperatures (0–5oC),when no growth took place. Fujii et al (1994) found that the specific activity of HD ofhalophilic histamine-forming bacteria Photobacterium phosphoreum and P.

Histamine Fish Poisoning
45
histaminum sp. nov. remained at 27% and 53% of the initial value after storage in cellsuspensions for 7 days at 4oC and –20oC respectively, while the viable cells decreasedby 5 log cycles and 8 log cycles of the initial counts respectively. This indicates thatoutbreaks of HFP may be caused by the ingestion of frozen–thawed fish even whenthe viable count of histamine forming bacteria is low. A common feature of studies on bacteria producing HD is that when multiple strainsof the same species are isolated, only one or a few of these strains are prolifichistamine producers (Gale 1946; Voigt and Eitenmiller 1977; Taylor et al 1978a;Taylor et al 1979; Yoshinaga and Frank 1982). Most strains of M. morganii are ableto produce histamine, but only certain strains of K. pneumoniae and H. alvei aredescribed as potent histamine producers (Havelka 1967; Taylor et al 1978a; Taylor etal 1979). Taylor et al (1979) isolated a histamine-producing strain of K. pneumoniaefrom a sample of tuna sashimi implicated in an outbreak of HFP. The strain wascapable of producing 442 mg of histamine per 100 g/tuna in tuna fish infusion broth in7 h under controlled incubation conditions. Only 12/50 other K. pneumoniae strainsisolated from foods, representing five distinct biochemical types, were able to producesuch high levels of histamine in the broth. Behling and Taylor (1982) divided HDB into those species capable of producing largequantities of histamine (>100 mg/100 mL) in tuna fish infusion broth during a shortincubation period (<24 h) at temperatures >15oC and those capable of producingsomewhat lesser quantities (<25 mg/100 mL) after prolonged incubation (>48 h) at30oC or above. M. morganii, K. pneumoniae and Enterobacter aerogenes wereprolific histamine producers, and tested strains of H. alvei, C. freundii and E. coliwere slow producers (Taylor et al 1978a; Behling and Taylor 1982). Taylor et al(1978a) suggested that most of the other bacteria identified as histamine producers inthe scientific literature fall into the slow-producer category. However, in reporting thatM. morganii, K. pneumoniae and H. alvei were the only bacteria that had beenisolated from fish implicated in fish poisoning incidents, Taylor (1985) postulated thatcertain other bacteria may also fall into the prolific-histamine-producer classification,citing as an example the isolation of C. perfringens from decomposing skipjack tunaby Yoshinaga and Frank (1982). Leung (1987) found that, of 36 bacterial isolates from spoiled chub mackerel(Rastrelliger sp.) skin washings, all the Morganella strains (10) produced largequantities of histamine. However, of two strains of Acinetobacter calcoaceticusisolated, one was the strongest producer of all strains tested whereas the other gave atotally negative result. Similarly, only 1/3 K. pneumoniae strains and 1/4 Aeromonashydrophila strains were histamine producers. Thus, for the range of organismsisolated in the study, apart from Morganella spp., the presence of HD was not aconsistent feature exhibited by species. In attempting to explain these results, Leung(1987) postulated that HD may be controlled by a plasmid and that the plasmid maybe transferred from strain to strain, species to species or genus to genus. SubsequentlyBarancin et al (1998) found that the fish pathogen Vibrio anguillarum harbours aplasmid-encoded HD gene. Taylor et al (1979) found no correlation betweenhistamine production and other biochemical characteristics or antibiotic resistance,which may also be linked to plasmid genes.

Histamine Fish Poisoning
46
Many bacteria (especially of the Enterobacteriaceae family) possess either ornithinedecarboxylase or lysine decarboxylase, the enzymes necessary to produce potentialpotentiators of histamine, putrescine and cadaverine, respectively (Taylor and Sumner1986). Since only a few bacteria possess HD responsible for histamine production, itis likely that the bacteria forming histamine differ from those forming putrescine andcadaverine. Leung (1987) found that, of 36 bacterial isolates from spoiled chubmackerel skin washings, none had the ability to decarboxylate all three amino acids,histidine, lysine and ornithine. 3.3.5.5 Bacterial destruction of histamine The histamine levels in a toxic fish depend on free histidine levels in the muscle andthe balance between histamine production and histamine destruction by thecontaminating microflora (Okuzumi et al 1984b). Histaminase (or DAO) activity hasbeen detected in several types of bacteria including M. morganii, Vibrio spp. andKlebsiella spp. (Gale 1942; Satake et al 1953; Ienistea 1971). Gale (1942) found thatbacterial histaminase is best produced under somewhat alkaline conditions (pH 7.5–8), but that moderate activity also occurs under slightly acidic conditions. Yamanaka(1984) showed that in saury, mackerel, yellowtail and big-eyed tuna the overall rate ofhistamine production was greater at 20oC than at 35oC, while in skipjack tuna morehistamine was produced at 35oC. The lower levels of histamine production at hightemperatures in the former species was attributed either to histaminases or to HDBwith a growth optimum of <35oC (Olley and McMeekin 1985). Ferencik (1970) reported that a strain of M. morganii, after being inoculated into asterile tuna flesh homogenate, produced large amounts of histamine. However, asignificant amount of histamine was soon decomposed by the organism. A subsequentsterile addition of histidine to the inoculated homogenate again resulted in histamineformation, followed by histamine destruction. He concluded that the histamineproduction and destruction by the M. morganii strain was determined by theconcentration of free histidine in the fish homogenate. 3.3.6 Histamine levels and toxic dose While the presence of histamine in fish muscle is a good indication thatdecomposition has taken place, its occurrence is extremely variable. As is evidentfrom the above, its production is a function of the species of fish and the individualfish, the part of the fish sampled, time, temperature, and the types and numbers ofbacteria present (Rawles et al 1996). The threshold toxic dose for histamine in foods is not precisely known (Taylor 1986).The variation in histamine levels within a spoiled fish (Lerke et al 1978; Frank et al1981) makes estimates of the toxic threshold difficult to obtain. Lerke et al (1978)showed that the distribution of histamine in spoiling tuna is quite uneven, varyingmore than 4-fold over 3 cm, and being considerably higher near the gut cavity thanelsewhere. Yoshinaga and Frank (1982) attributed this to the non-uniform distributionof spoilage organisms throughout the fish, and noted that muscle levels of histidinewere essentially uniform in fresh skipjack tuna. Simidu and Hibiki (1955) estimated

Histamine Fish Poisoning
47
the threshold toxic dose for histamine in fish at about 60 mg/100 g, but their methodswere not precise. Shalaby (1996) reviewed the oral toxicity to humans of histamine and other biogenicamines in foods. He considered that histamine-induced poisoning is, in general, slightat 8–40 mg, moderate at >40 mg and severe at >100 mg. Based on an analysis ofrecent poisoning episodes, Shalaby (1996) suggested the following guideline levelsfor histamine content of fish:
• <5 mg/100 g (safe for consumption)• 5–20 mg/100 g (possibly toxic)• 20–100 mg/100 g (probably toxic), and• >100 mg/100 g (toxic and unsafe for human consumption).
This assessment correlates with those of Simidu and Hibiki (1955) and Mitchell(1993), that poisoning does occur at lower histamine concentrations than100 mg/100 g fish. There is uncertainty regarding the threshold toxic concentration because potentiatorsof toxicity may be present in fish and lower the effective dosage compared with purehistamine (Institute of Medicine 1991). Different fish could contain differentpotentiators, and the levels of potentiators could also vary considerably from one fishto another. The types and levels of potentiators in a fish would depend on a variety offactors, including the natural constituents of the fish, the contaminating bacteria andtheir metabolic capabilities, and the environmental conditions (mainly temperature).Until the identity, levels and potency of possible potentiators and/or mast celldegranulating factors are elucidated, it is difficult to establish regulatory limits forhistamine in foods on the basis of potential health hazard. FDA guidelines for tuna, mahi-mahi and related fish specify 500 ppm (50 mg/100 g)as the toxicity level, and 50 ppm (5 mg/100 g) as the defect action level becausehistamine is not uniformly distributed in a decomposed fish. Therefore, if 50 ppm isfound in one section, there is a possibility that other units may exceed 500 ppm (FDA1998). These levels, based on years of investigative experience, allow for the non-uniform distribution of histamine in a spoiled fish, but ignore the existence ofsuspected potentiators or other toxins. For control of histamine in fish belonging to the Scombridae and Clupeidae families,European Union Directive No. 91/493 stipulates that nine independent samples fromeach batch should correspond to: 1. An average histamine concentration lower than 100 ppm (10 mg/100 g). 2. No more than 2 samples out of the 9 with a concentration of between 100 and
200 ppm. 3. No sample with a histamine content higher than 200 ppm. The acceptable level set as a manufacturing standard by legislation in the UnitedKingdom in 1992 was 10 mg/100 g (Anon. 1998b).

Histamine Fish Poisoning
48
In Australia and New Zealand, the Australian Food Standards Code (Issue 41,18 November 1998) Standard D1 – Fish (ANZFA 1998a) states: (1B)(a) The level of histamine in a composite sample of fish or fish products, otherthan crustaceans and molluscs, when examined according to Section 977.13 ofAssociation of Official Analytical Chemists (AOAC) 15th Edition (1990) must notexceed 100 mg/kg. (b) For the purpose of this clause, a 'composite sample' is a sample, taken from eachlot, consisting of five portions of equal size taken from five representative samples. This clause came into force in October 1994 (Peter Abbott, ANZFA, pers. comm.1999) and seems conservative and appropriate to protect public health and safety.However, it is currently under review, with a proposal by ANZFA to increase themaximum allowable level of histamine in fish and fish products to 200 mg/kg(ANZFA 1998b). If we regard 10 mg histamine/100 g of fish as the highest level that can be consumedsafely, this must be related to the amount of fish eaten and the weight of the person tocalculate a likely safe dose. If a 60 kg person eats, say, 300 g (wet weight) of this fish,this dose would be 0.5 mg/kg bw. Such a calculation is of limited value, however,considering the variable nature of HFP and the lack of understanding of itspathogenesis.
3.4 Human factors that affect the clinical response 3.4.1 Variation in individual susceptibility The severity of the clinical response will depend on the amount of toxin(s) ingestedand variation in individual susceptibility. The literature would suggest that there is abig variation in individual susceptibility. This was illustrated in the experiment ofMotil and Scrimshaw (1979) mentioned earlier. Oral administration of 100–180 mghistamine in grapefruit juice or in 100 g high-quality tuna caused characteristicsymptoms of mild histamine poisoning in only 1/4 and 4/8 volunteers respectively. According to Taylor et al (1989) attack rates in group outbreaks of histaminepoisoning vary from 50% to 100%. In the human volunteer trials of Van Gelderen etal (1992), only 2/8 subjects (both female) exhibited symptoms after ingesting portionsof spoiled fish containing 90 mg histamine, and these volunteers did not have higherconcentrations of histamine in plasma than the volunteers who did not showsymptoms. In the four cases of 'classic scombrotoxism' described by Blakesley (1983), amountsof histamine ingested were not known. Signs and symptoms varied between thesubjects. Three of four patients exhibited burning skin, headache and nausea, two offour had abdominal cramps and pruritus, and one of four had hive-like skin lesionsand palpitations.

Histamine Fish Poisoning
49
3.4.2 Influence of diet Geiger (1955) suggested that alterations in the intestinal tract caused by seasoned hotdishes prepared from spoiled fish or the simultaneous consumption of alcoholicbeverages might cause histamine to be absorbed at an increased rate, such that itsdetoxification could not keep up with its entry into the circulation. Zee et al (1981)also suggested that histamine may be absorbed more rapidly in the presence of alcoholand exert a more-marked biological effect on the circulatory system. However, in therat, ethanol enhanced DAO and HMT activity to 123% and 111% of normalrespectively (Taylor and Lieber 1979). Mitchell and Code (1954) reported thathistamine taken with a meal (bread, butter and milk) was absorbed to a greater extentthan histamine consumed by itself. Granerus (1968) also studied the effect of diet on histamine metabolism in humans.His studies indicated the variability in urinary excretion of histamine and its mainmetabolite, 1-methyl-4-imidazoleacetic acid. Granerus (1968) quoted a study by Irvineet al (1959) that concluded that some intestinal bacteria could in fact contribute tourinary histamine by decarboxylating histidine in food. While most proteins differ byonly 2–3% in their histidine content, these proteolytic bacteria are favoured by a dietrich in animal protein and inhibited by a diet that favours the acidophilic group ofbacteria (e.g. vegetables, grains, lactose and dextrin). Thus, dietary differences mayexplain the highly varying amounts of histamine metabolite found in the urine of somesubjects studied by Granerus (1968). Foo (1977) cited an observation of Johnson and Overholt (1967) of a significantincrease in the concentration of histamine in the gastric venous blood of dogs whosestomachs had been filled with dilute acetic acid solutions. He suggested that the lemonor vinegar commonly used to enhance the flavour of fish might also enhance theabsorption of histamine from toxic fish. The mechanism may well be that of acids inlemon or vinegar affecting the pH of the intestinal contents and inhibiting the activityof histamine metabolising enzymes. Amines such as histamine, cadaverine, putrescine and tyramine are found in meat andmeat products, cheeses and other fermented foods such as sauerkraut, beverages suchas wine and beer, and fruits and vegetables (Shalaby 1996), and in Chinese foods suchas tamari and soy sauce (Chin et al 1989). Therefore, it would seem that dietary itemsother than fish could theoretically exacerbate HFP if consumed at the same time asspoiled fish. A number of foodborne substances are DAO inhibitors, includinganserine, carnosine, agmatine, thiamine, cadaverine and tyramine (Taylor 1986). Inaddition, the increasing popularity of mixed fish dishes such as seafood marinara maypromote HFP by providing histamine potentiators from different fish species eaten inthe same meal. 3.4.3 Influence of medication Some drugs can inhibit histamine-metabolising enzymes and potentiate histamineactivity when taken in conjunction with food containing high concentrations ofhistamine (Taylor 1986; Chin et al 1989).

Histamine Fish Poisoning
50
HMT is inhibited by analogues of methylmethionine, such as adenosyl-homocysteine,antimalarial drugs (quinacrine, chloroquin and amodiaquin) and numerous agonistsand antagonists of histamine receptors. Some antihistaminic drugs andaminoguanidine are inhibitors of DAO. Several compounds once classified as specificMAO inhibitors are known to inhibit DAO as well. These substances includeisoniazid. (References cited by Taylor 1986.) On the other hand, some individualstaking antihistamines for other reasons may be protected from histamine poisoning tosome extent (Taylor et al 1989). Isoniazid, an anti-tuberculosis medication, has been incriminated most often as anexacerbating factor in HFP (Stratton et al 1991). Scientists in Sri Lanka have reportedseveral incidents of histamine poisoning among patients with tuberculosis whoconsumed tuna while receiving isoniazid therapy (references cited by Taylor 1985). Inone outbreak, 21 patients were affected (Uragoda and Kottegoda 1977) and, inanother, 56 (Senanayake and Vyravanathan 1981). 3.4.4 Disease states and age Histidine metabolism in humans can be altered in certain disease states.Histidinaemia, an inborn error of metabolism resulting from the absence of HAL, isassociated with increased excretion of histamine and its metabolites. Patients with thisdisease or with altered histamine metabolism may be more susceptible to HFP.Exogenous histamine would also exacerbate diseases such as allergies in whichelevated endogenous histamine levels play a role, and mastocytosis (Taylor 1986).Activities of histidine- and histamine-metabolising enzymes may also be affected withage.
3.5 Morbidity and mortality rates HFP is usually a rather mild illness (Taylor 1986). In rare cases it has proved fatal(Eitenmiller 1981). Arnold and Brown (1978) report that morbidity values vary from0.07% to 100% in the literature, but say that such figures are misleading, as differentlots of canned fish may differ greatly in their histamine content. The distribution ofhistamine in individual fish is also highly variable, as is individual susceptibility tohistamine poisoning. Nevertheless, in group outbreaks the attack rate may approach100%. When an outbreak involves raw fish, the attack rate is sometimes substantiallybelow 100% (Taylor 1986; Wu et al 1997). The lack of any visual sign of spoilage in scombrotoxic fish probably accounts for thehigh morbidity rate. With food allergies, it would be unusual for more than one personin a group to experience symptoms caused by a specific food, which assists in thedifferential diagnosis. Allergic reactions to some of the fish commonly incriminated inHFP outbreaks, such as tuna and mahi-mahi, are quite rare (Taylor 1986).

Histamine Fish Poisoning
51
4. EXPOSURE ASSESSMENT 4.1 Definition of exposure assessment Buchanan et al (1998) defines exposure assessment as the probability of consumptionand the amount of the biological agent consumed. Topics that may be consideredunder the heading of exposure assessment include:
• levels of the hazard in food at different stages in its production;• the frequency of exposure;• the duration or extent of exposure;• exposure of sub-populations (including ethnicity, age, geographic location,
consumption patterns). In discussing exposure assessment in relation to HFP, we will examine, in a non-quantitative way, factors affecting the probability of HFP occurring, histamine levelsin fish products in Australia, amounts of fish consumed and at-risk population groups,and future exposure trends.
4.2 Factors affecting the probability of histamine fish poisoning occurring 4.2.1 Post-catching contamination There is uncertainty in the literature regarding the main source of HDB, which havebeen isolated from the skin, gills, intestines and muscle tissues of spoiling fish (Lerkeet al 1978). Kimata (1961) and Yoshinaga and Frank (1982) estimated that histamineformers occupied about 1% of the regular surface microflora of fresh fish and, withextended storage at elevated temperatures, the bacteria invade the muscle and convertfree histidine to histamine. However, in their study of HDB contaminating fishpurchased in retail markets in Spain, López-Sabater et al (1996) found thatPlesiomonas shigelloides was the sole HD isolate frequently associated with themarine environment. Most scientists believe that post-harvesting contamination is the most importantsource of histamine formers. Taylor and Speckhard (1983) devised a method for therecovery of HDB from frozen skipjack tuna (Katsuwonus pelamis) obtained from amajor tuna packer in the United States. Gills, intestines and muscles were sampled.HDB (M. morganii and C. freundii) were isolated from only 3/10 fish, in each casefrom the gills only. The evidence suggested that Enterobacteriaceae are not part of thenormal microflora of tuna and the isolation of these bacteria from the gills, which areusually used as 'hand-holds' in handling, was attributed to post-harvestingcontamination. Taylor et al (1989) went further to say that most of the histamineformers found in fish are common enteric bacteria of humans and animals. However,the status of C. perfringens and N-group bacteria that grow at refrigerationtemperatures cannot be confirmed, since early recovery procedures used did not caterfor their detection. Scoging (1991, 1998) agreed with Taylor (1986) that outbreaks ofHFP may result from fish becoming contaminated after harvesting.

Histamine Fish Poisoning
52
In the United States, most outbreaks of HFP are associated with improperly handledfish from private catches (Lange 1988). HFP has also been linked to restaurant-prepared meals, as well as to canned tuna, both of which involve commercial fishingoperations (Lange 1988). Post-catching contamination with HDB may occur at severallevels — aboard the fishing vessel, at the processing plant, in the distribution system(fresh and frozen fish), and at the level of the user (Taylor 1986). Contaminationduring handling of canned tuna for the preparation of a tuna salad in a restaurant wasimplicated in one German outbreak (Yamani et al 1981). Taylor (1986) suggests thatrestaurant contamination could be particularly important with raw tuna. To understand the sources of contamination of fish with HDB, Subburaj et al (1984)investigated the fish market environment of Mangalore, India, for counts of thesebacteria. They cultured samples from carrying baskets, ice, the market floor, and waterfor wetting fish. They also recorded total plate counts (TPC), HDB counts andhistamine levels in samples of seerfish (Scomberomorus guttatus) and mackerel(Rastrelliger kanagurta). The results demonstrated that HDB (identified with Niven'smedium) were widely distributed in the market environment and water. SinceMorganella spp. (then Proteus spp.) are the most common bacteria associated withHFP, the incidence of these bacteria, in particular, were examined in various marketsamples (see below). Incidence of Morganella spp. in various market samples
Sample No. analysed No. +ve forMorganella
% of Morganella in TPC
Water Ice Basket Floor
3 3 3 4
3 1 3 4
10–15 10
40–80 45–?
The generic composition of HDB in a mackerel sample containing 20 mghistamine/100 g revealed Morganella (52%), Pseudomonas (21%), Plesiomonas(12%), Providencia (8%), Flavobacteria (6%) and Aeromonas (1%). As could beexpected, there was no direct correlation between HDB count and histamine level,confirming that the generic composition of the microflora is more important thannumbers. If fish are stored ungutted, the gut itself may be a source of contamination, particularlyif chilling is delayed. For example, if anaerobic bacteria of the gut such as C.perfringens (a prolific histamine producer) are allowed to proliferate at hightemperatures, they will produce enzymes that are highly active at lower temperatures(Olley et al 1985). 4.2.2 Temperature abuse on fishing vessels At any time between catching and consumption, exposure of certain fish to elevatedtemperature can cause formation of histamine from histidine by HDB, which areinevitably present. Quality assurance on board fishing vessels is the first link in thequality chain that runs from the start of production to final consumption. Formation of

Histamine Fish Poisoning
53
histamine can be induced by short-term temperature abuse early in the chilling stepand detected later (Andersen 1997). The key to the reduction of histamine production is the rapid cooling of the fish aftercatching (Ritchie and Mackie 1979). Initial cooling is important in reducing the rate ofhistamine production, even when temperature rises at a later stage (Mitchell 1993).Many fish species commonly implicated in outbreaks of HFP are caught in warmwater, and fish temperatures at capture frequently exceed 20oC. Fish may remain inpurse-seiner nets or on longlines at this elevated temperature for several hours aftercapture, and fish may die before they leave the water. Tuna have higher body temperatures than most other fish. In the western Pacific,where sea surface temperatures are 27–30oC, tuna often come on board with internaltemperatures of ≥32oC. Spoilage at this temperature is about 30 times faster than at0oC. Because of elevated temperatures, the length of time from death until tuna arechilled is the most critical phase of shipboard handling (Bartram 1997). Once on board the fishing vessel, the fish may or may not be cooled, and methods ofcooling employed vary widely in their efficiency (Taylor 1986; Price 1999). Americanpurse seiners cool the fish in holds filled with refrigerated seawater. Once the hold isfull, the fish can be frozen. The rate of cooling depends on the size of the catch and onthe size of individual fish. The fish may be held at elevated temperatures for sometime, and are not gutted until they reach the processing plant. Variable conditions existfrom boat to boat, and this probably explains the sporadic incidence of histaminepoisoning (Taylor 1986). Any time a fish is held at >4.4oC (40oF) significantly reducesthe expected safe shelf life, but fish that have been handled particularly well on-boardthe harvest vessel may be able to safely withstand somewhat more exposure toelevated temperatures during post-harvest handling (Price 1999). 4.2.3 Inadequate chill-storage procedures Taylor (1986) cites a number of studies on the effect of storage temperature onhistamine formation in various types of fish. While all the studies agreed thathistamine formation is negligible in fish stored at 0oC or below, other results werevariable. Widely varying data exist for both the lower temperature limits for safestorage and for the optimal temperature for producing histamine. This is notsurprising, given the variability of the nature of the bacterial populations on fish.
Low-temperature storage (<10°C) effectively controls the growth of most histamine-producing bacteria, which require a warm temperature for growth. However, bacteriathat grow at refrigeration temperatures can produce smaller amounts of histamine infish stored at temperatures between 0 and 10°C (Ritchie and Mackie 1979; Klausenand Huss 1987b; Stratton and Taylor 1991). Regardless of the species involved,bacteria must grow to a large enough population for significant production ofhistamine to occur. In fish subjected to elevated temperatures, even for short periods, a large population ofbacteria is soon established. During subsequent refrigeration, although bacterialgrowth ceases, residual enzyme activity continues slowly and histamine levels

Histamine Fish Poisoning
54
continue to increase (Klausen and Huss 1987b; Stratton and Taylor 1991; Institute ofMedicine 1991). Klausen and Huss (1987b) found that large amounts of histaminewere formed in histidine-containing broth and in mackerel at low temperatures (0–5oC), where no growth of M. morganii took place, following storage at highertemperatures (10–25oC). Geiger (1944a) found that histamine concentration in scombroid fish increased from0.09 mg/100 g when fresh to 95 mg/100 g when kept at room temperature (20–25oC)for 10 h. According to Gilbert et al (1980), fish packed in melting ice (0oC) remainedible for about 12 days, by which time histamine concentrations are still only 3–4 mg/100 g. At a room temperature (15–25oC), high concentrations of histamine arereached rapidly, and fish become toxic, even though they may appear acceptable to theconsumer. Ritchie and Mackie (1979) monitored the formation of histamine, putrescine,cadaverine, spermine and spermidine in freshly caught mackerel and herring heldungutted on ice for 28 days at 1oC, in an incubator at 10oC, or in an insulated box atambient temperature (25oC). In mackerel held at 1oC, histamine rose slowly from aninitial level of 0.01 mg/100 g at day zero to 5.25 mg/100 g at day 14, and quickly to57.94 mg/100 g at day 28. Cadaverine rose from 0.01 mg/100 g at day zero to43.08 mg/100 g at day 28, while putrescine rose from 0.05 mg/100 g to8.92 mg/100 g. Similar trends occurred for herring. The ratios of the finalconcentration of histamine:cadaverine:putrescine at day 28 for mackerel and herringstored at 1oC were 6.5:4.8:1.0 and 8.3:5.9:1.0 respectively. Thus, the concentrations ofhistamine and other amines, even in putrid fish, were quite low (less than100 mg/100 g/fish) after prolonged low-temperature storage. At the highertemperatures, the amines were produced in relatively large amounts. In an experiment involving nine lots of sardines with undocumented post-harvesthistory bought at a fish market in Morocco, Ababouch et al (1986) demonstrated theeffect of refrigeration on histamine development. Sardines stored at 8oC had a 12-hlonger shelf life than those held at 17oC. A combination of salting and refrigerationwas more effective. Fish held at 8oC and salted at a level of 5% or 8% had a shelf life35 h longer than fish held at 17oC with no salt. Chen and Malison (1987) reported typical scombroid symptoms after consumption ofmackerel stored on ice for 2 days and then kept at room temperature of 30oC for 3–4 hbefore cooking. The incriminated fish contained only 10 mg/100 g histamine. Toxinproduction apparently occurred at a rapid rate, because people who ate lunch from12.30 to 1 pm had a much higher attack rate than those who ate 1–2 h earlier. Otherfish from the same catch did not cause illness after standing at room temperature foronly 1 h. 4.2.4 Inadequate freezing and thawing procedures Ben-Gigirey et al (1998) investigated the changes in biogenic amines, and numbers ofbacteria reported to have decarboxylase activity, in albacore (white tuna) muscleduring frozen storage. Albacore specimens of high quality (either 'extra' or A categoryon sensory analysis) were analysed for their biogenic amine and bacterial contents

Histamine Fish Poisoning
55
after 1, 3, 6 and 9 months of frozen storage at –18oC or –25oC. Thirty-seven intactspecimens of albacore were purchased in October 1995 at the docks of Burela (north-western Spain) and transported on ice and under refrigeration (4oC) to the laboratory.The weight range was 1.2–3.3 kg. Five samples were kept on ice 'until samples couldbe taken for microbiological and biochemical analysis'. The remaining 32 fish wereglazed in cold water, frozen in a freezing tunnel at –40oC, and divided into two groupsof 16 fish each that were stored separately at –18oC and –25oC. The scientists optimised a HPLC method involving a linear elution gradient for theidentification and determination of putrescine, cadaverine, histamine, spermine andspermidine. The initial average contents of all the amines, except for spermidine, were<0.8 mg/100 g. Putrescine decreased during storage after 6 months at eithertemperature, but increased to average concentrations of 5.9 mg/100 g (815% of theinitial level) and 6.8 mg/100 g (942% of the initial level) in the white muscle ofalbacore after 9 months of frozen storage at –18oC or –25oC respectively. Histaminedecreased after 3 months storage, but at –18oC it increased to a final concentration of0.48 mg/100 g (103% of the initial level) after 9 months of storage. This rise did notoccur in samples stored at –25oC. Cadaverine contents tripled or doubled after 3 months of storage at –1oC or –25oCrespectively. Thereafter, cadaverine levels began to decrease. After 9 months' storage,they were higher at –25oC (0.26 mg/100 g, 86% of the initial level) than at –18oC(0.17 mg/100 g, 56% of the initial level). Spermine contents increased as frozenstorage progressed, and reached concentrations of 0.95 mg/100 g (120% of the initiallevel) and 1.0 mg/100 g (129% of the initial level) after 9 months at –18oC and –25oCrespectively. By contrast, spermidine contents persisted in the 11–12.8 mg/100 ginitial range during the first 6 months of frozen storage, and after 9 months the finalconcentrations of this amine were significantly lower. No pathogens responsible for foodborne disease were isolated. Aerobic bacteria thatgrow at ambient temperatures survived frozen storage at –18oC (39%) and –25oC(92%) for 9 months. The survival rate at –25oC of bacteria that grow at refrigerationtemperatures was 4.6% after 9 months. The results suggest that good handling practices are mandatory during the thawing offish before canning, since the absence of certain bacteria potentially involved inspoilage due to amines is not guaranteed, even after 9 months' storage at –25oC, andthe levels of some biogenic amines may increase. 4.2.5 Temperature abuse in the preparation of dried and/or smoked products Toxic levels of histamine have been found in dried and/or smoked products ofmackerel, horse mackerel and sardines (Taylor 1983), as well as in the fishmeal madefrom these fish. Exposure of raw fish to high ambient temperatures accelerates thisreaction. Histamine content increases to a maximum, then decreases with prolongeddrying time. The drying of sardines previously brined in 5–15% sodium chloride for2 h causes pronounced increases in histamine (Pan 1988).

Histamine Fish Poisoning
56
Trinidad et al (1986) reported on histamine production in Spanish mackerel smokedfor 8 h at 45oC and stored at refrigeration temperature (0–5oC) as 3-mm-thick slicespacked in flexible plastic films, under vacuum or not. Histamine increased in bothkinds of packs (see table below), but not to a significant extent because the fish usedfor smoking were very fresh, kept chilled, and contamination during handling andprocessing was kept to a minimum. Vacuum-packed samples had lower microbialcounts at the middle of storage, but at the end (day 30), the difference between the twowas negligible. Changes in histamine content (mg/100 g) of sliced smoked mackerel after storage at 0–5oC
Storage day Vacuum-packed Non-vacuum-packed 0 14 30
1.12 2.18 3.19
1.12 2.98 4.20
Van Spreekens (1987) reported high levels of histamine in vacuum-packed, lightlysalted herring fillets stored at refrigeration temperatures, particularly after low-levelcontamination with histamine-producing photobacteria. The salt concentration of theproduct was a vital factor in their growth (0.5–4%). Van Spreekens (1987) attributed histamine production in salted or hot-smokedmackerel to Photobacterium spp., which grow at refrigeration temperatures. The fishwere exposed to temperature abuse as a result of delayed cooling on board ship and/orduring processing. Van Spreekens (1987) maintains that, because photobacteria arethermolabile and have special requirements for growth, they have often beenoverlooked as HDB and causative agents in HFP. Hot smoking 'practically sterilises' the product and denatures enzymes, thus impartingsome degree of preservation, but does not destroy histamine already formed (Poulter1988). Bremer et al (1998) carried out thermal death trials using a H. alvei strainisolated from a portion of hot-smoked kahawai with a histamine level of166 mg/100 g. Results of the trials, carried out in 0.1% peptone suspension and inkahawai at 54, 55, 56, 57 and 58°C, indicate that hot smoking has the potential toeliminate H. alvei from seafood products. At the same time, histamine is very resistantto heat and, once present in a fish, is not easily destroyed by smoking, cooking orcanning. Fletcher et al (1998) reported the results of a retail survey of the levels of histamine inhot-smoked fish products available in New Zealand, where most cases of HFP areassociated with smoked fish. They purchased 107 samples from Auckland retailmarkets between July 1995 and March 1996 and determined their histamine andbacterial levels. Eight samples (from 5/9 retail markets) had histamine levels thatexceeded 50 mg/kg. In two samples, histamine levels exceeded 200 mg/kg (346 and682 mg/kg). Within or between fish species there were no consistent relationshipsbetween levels of histamine in the samples and either the total aerobic plate counts orthe numbers of histamine-producing bacteria. Some of the samples with elevatedlevels of histamine had relatively low microbial counts. For example, sample 136 hadthe highest histamine level of any of the retail products, yet its microbial counts were

Histamine Fish Poisoning
57
all less than 6 x 103 CFU/g. The evidence showed that histamine had been formedprior to smoking, and that histamine-producing bacteria were eliminated duringsmoking. 4.2.6 Poor canning procedures The rate of histamine formation in fish prior to canning is a function of:
• temperature during storage and preparatory operations;• duration of exposure at critical temperatures;• contamination levels of HDB; and• levels of free histidine in the fish (Warne 1985). Incidents of HFP caused by high levels of histamine in commercially cannedscombroid products have occurred all over the world (Taylor 1983). Improvements inhandling and processing associated with the establishment of HACCP procedures arebecoming widespread. These offer the advantage of in-process control, while avoidingthe need for large-scale terminal analyses of finished product, which is timeconsuming and beyond the capabilities of many small canneries (Warne 1985). 4.2.7 Low-quality fermented products Fermented fish containing high levels of histamine could conceivably present ahazard. In Finland, sugar–salted herring in barrels is more likely to contain high levelsof histamine than canned herring, but has not been implicated as causing illness. Moreresearch is needed to define suitable limits for histamine in fermented products(Taylor 1985). Fermented fish products are usually consumed in small amounts, sohigher concentrations of histamine could possibly be tolerated in these products. 4.2.8 Temperature abuse of raw tuna for the sashimi market The principal seafood safety hazards in the lucrative raw tuna trade are decompositionand histamine formation. Both are linked to temperature abuse. Raw tuna buyers arepresented with so much quality variation that each fish is inspected and graded priorto purchasing. The primary purpose of this evaluation is economic — to direct eachtuna to the most profitable market niche. When decomposition or high histaminelevels occur, they usually involve the lowest grade (Grade 4 or D) of raw tuna. Thehighest quality raw tuna is sold in sushi bars. Grading fulfils the objectives ofHACCP, because raw tuna core temperatures are checked and fish areorganoleptically examined at critical control points (Bartram 1997).

Histamine Fish Poisoning
58
4.3 Histamine levels in fish products in Australia
4.3.1 Survey of histamine in canned tuna
In the absence of information concerning the prevalence of histamine in canned tunaconsumed in Australia, Warne et al (1987) screened cans of tuna purchased fromseveral Melbourne retail markets. Of 104 cans surveyed, 45 were produced inAustralia and the remainder came from Japan, Malaysia, the Philippines and Thailand.Product from each of the cans was screened for histamine at <5 mg/100 g, 5–10 mg/100 g and >10 mg/100 g using TLC based on the method used at the TorryResearch Station in the United Kingdom. Most cans (101/104) contained <5 mg/100 g, and therefore complied with the moststringent of current international standards. In 3/104 cans, histamine levels were in the5–10 mg/100 g range, which is still acceptable. During canning, the HD enzyme isdenatured at retorting temperatures, while bacteria responsible for its production arekilled. The presence of >5 mg/100 g in 3/104 samples suggests that fish weremishandled at some stage in the sequence of operations leading up to retorting. 4.3.2 Survey of biogenic amines in fish products in the ACT Another survey was published as an information paper on the Internet. The AustralianCapital Territory Government published the results of a survey of biogenic amines infish and fish products from April to June 1997 (Rigg 1997). Objectives were todetermine:
• the compliance of fish and fish products (including fresh, canned and dried) forhistamine to the Australian Food Standards Code (ANZFA 1998); and
• the levels of two other biogenic amines, putrescine and cadaverine, which are alsoof concern in fish products.
The paper was poorly presented and did not have a 'Materials and methods' section.Analytical methods were not given, and 'detectable amounts' were not specified. 'Fish'were not adequately defined and included molluscs, prawns and seafood marinara.The paper stated that 58/64 of samples (91%) did not have detectable levels ofhistamine, 5/64 (8%) contained levels below the Food Standards Code level of100 mg/kg and 1/64 (2%) failed to comply with the standard. The latter sample, ofdried fish imported from Asia, contained 653 mg/kg histamine. The paper went on tosay that 3.4% (1/29) of fresh 'fish' compared to 14.3% (5/35) of processed fishsamples were found to contain histamine. These samples were one of fresh chunktuna, one of tinned fish and four of dried fish. Histamine was not detected in any ofthe 20 samples of crustaceans and molluscs surveyed. There was a significantdifference in histamine levels between fresh (3.3%) and processed (14.7%) fishproducts. Putrescine was found in 20/64 samples (31%) and cadaverine in 28/64 (44%). Forputrescine, 10/29 of fresh samples (34%) and 10/35 of processed samples (29%) haddetectable amounts. There were five samples that exceeded 100 mg/kg, whichincluded two of green prawns, one of scallops and two of dried fish. For cadaverine,

Histamine Fish Poisoning
59
13/35 (37%) of fresh samples and 15/35 (43%) of processed samples had detectableamounts. Three contained >100 mg/kg — two of dried fish and one of scallops. 4.3.3 Monitoring by the Australian Government Analytical Laboratories The Australian Government Analytical Laboratories (AGAL), Melbourne, monitorcanned and raw tuna and other fish imports for the Australian Quarantine andInspection Service (AQIS). Random samples are analysed on a regular basis andincluded fish from various Asian countries and the United States. The detectionmethod used is capillary electrophoresis (Trenerry et al 1998), which has proved moresatisfactory than the AOAC fluorometric method (AOAC 1995c). Only a smallpercentage of samples exceed the Australian Food Standards Code (ANZFA 1998)level of 100 mg/kg, and in these few samples concentrations are rarely much greaterthan 100 mg/kg (Philip Marshall, AGAL, pers. comm. 1999). 4.3.4 Monitoring by industry Information on industry monitoring for histamine levels in fish and fish products wassought from Port Lincoln Tuna Processors Pty Ltd, South Australia, the onlyremaining tuna cannery in Australia. Skipjack tuna is canned at Port Lincoln. Allbatches entering the cannery (mainly from Australia, but some from overseas) aremonitored, with five fish from each batch being tested. The factory also monitorshistamine in random samples of its canned product. The ALERTTM ELISA test kit isused (sensitivity 50 ppm). In addition, every 6 months, samples are sent to AGAL inMelbourne for analysis by capillary electrophoresis (sensitivity 10 ppm), to ensurestandardisation of results. The two methods of analysis show good agreement, andhistamine levels routinely do not exceed the Australian Food Standards Codemaximum level of 10 mg/100 g (Lea Traeger, pers. comm. 1999).
4.4 Amounts and types of fish consumed, and at-risk population groups
All population groups are susceptible to HFP, but regional differences undoubtedlyexist in the amounts and types of fish consumed and the way they are processed andstored. The disease still occurs frequently, even in developed countries such as theUnited States, Japan and the United Kingdom (Taylor 1986).
Figures were not found for consumption of various fish species in different countries.Even if they were found, their value would be limited in assessing possible exposureto HFP, because details of handling and storage conditions would not be known.Although fish consumption in Australia is probably increasing, as people arebecoming more health conscious, it is not high by world standards. In 1995, ANZFAconducted an Australian National Nutrition Survey based on a 24-h recall surveymethod for 13 858 people aged 2 years and over. Total mean intake of all species offish for all respondents (consumers and non-consumers of fish) was 12.2 g/day, andfor marine fish the figure was 10.2 g/day. This means that the average Australianconsumes less than 4.5 kg/year fish in his/her diet and less than 4 kg/year of marinefish. Although tuna probably represents a fairly high proportion of the marine fishconsumed, because of its commercial importance, the chance of an Australiancontracting HFP is still very low.

Histamine Fish Poisoning
60
HFP is probably still common in developing countries where fish preserved bytraditional methods is an important part of the diet. Such diets remain extremelypopular and are a major source of inexpensive dietary protein. Their popularity islargely due to desirable flavours produced by chemical changes that occur during thesmoking and drying processes. Traditional African products are rarely salted, butsome may be sun-dried before smoking starts. In the course of sun and air drying,bacteria can multiply in the moist interior of the fish, which has high water activity,causing the inside to spoil while the outside appears satisfactory. The problem wouldbe expected to be greater in tropical countries where higher ambient temperaturespromote the growth of prolific HDBs (Fig. 3).
In tropical countries, fish are cold smoked at 30–40oC, depending on species, toprevent coagulation of protein, which occurs with cooking. This process is designed togive a desired flavour to the product rather than a significant degree of preservation(Poulter 1988). It will not destroy HDB or deactivate HD. Even if hot smokingfollows cold smoking, histamine concentrations may have already become highenough to become hazardous.
Products cured in the traditional way have become less popular in most developedcountries. However, in some non-tropical countries, such as the United Kingdom, fishare cold smoked at low temperatures (<30oC depending on species, Anon. 1988a).The modern, mildly smoked and less extensively dried products are less stable thanthe traditional ones, and this trend has been made possible by chilling and freezing.Neither cold- nor hot-smoked fish will keep for long periods unless supplementarytechniques, such as drying, freezing or chilling, are used (Poulter 1988).
The consumption of raw fish, mainly by the Japanese, probably presents less risk ofHFP because only the highest quality fish are directed to this trade. Also, as theproduct is a delicacy, smaller quantities of fish would be likely to be consumed permeal.
4.5 Future exposure trends
In the past 10 years or so, authorities and producers around the world have becomemore aware of the need for quality assurance in relation to food (Barker andMcKenzie 1997). As a result, standards for seafood quality are improving andexposure to spoiled seafood and thus HFP is likely to become less prevalent. Developing countries have increased their net income from fish and fishery productsfrom about US$3 billion to some US$18 billion (FAO 1999) over the past 10 years.By 1997, developing countries had equalled the production of the developed world,contributing almost 50% of global fishery exports (Karnicki 1997). Great progress hasbeen made in the quality of fish products at the same time as the huge expansion ofinternational trade. This is the result of the introduction of international standards infood hygiene and the application of risk analysis and HACCP principles, as part of acommon, global approach for maximising the quality and safety of all food products(FAO 1999). Thus, HFP is now much less likely to occur as a result of commercial

Histamine Fish Poisoning
61
canning or smoking as HACCP quality assurance measures are increasingly beingadopted. In Australia, the Australian Quarantine and Inspection Service (AQIS) has introducedthe Hazard Analysis and Critical Control Point (HACCP) and International StandardsOrganization 9000 series of standards (ISO 9000) quality management inspectionsystems. These initiatives, together with those of other government and industryagencies, have encouraged the Australian export seafood industry and other sectors ofthe processed food industry to implement quality management systems that addressissues such as good handling practices and temperature control. AQIS, as part of theCommonwealth Department of Agriculture, Fisheries and Forestry, will endeavour toensure that the application of HACCP-based inspection systems in Australia remainsconsistent with overseas regulatory authorities (Aitken 1997).
Predictive microbiology is an emerging area of food microbiology in which microbialresponses to environmental factors are measured under defined and controlledconditions. The potential applications of predictive modelling in HACCP systems arenumerous (Ross and McMeekin 1995). Although predictive modelling cannot beapplied to histamine production directly, for reasons discussed in section 3.3.5.2, ithas an important future role in the potential to control microbiological spoilage of fishand thus will assist indirectly in the control of HFP.
5. RISK CHARACTERISATION
5.1 Definition of risk characterisation
Kindred (1996) defines risk characterisation as ’The description of the nature andoften the magnitude of human risk, including attendant uncertainty’. According toBuchanan et al (1998), this involves integration of the dose–response and exposureassessments into a risk statement, which includes estimation of the likelihood andmagnitude of disease under various conditions of exposure and description ofuncertainties. The risk characterisation should result in a quantitative or qualitativeestimate of the potential for adverse effects of the particular biological hazard on thepopulation.
In this section, an attempt will be made to summarise the nature and magnitude of therisk of HFP, health and other impacts of the disease, and uncertainties and problemareas in understanding and combating the disease. Although much is known aboutHFP, there are still many gaps in our knowledge. Sufficient data are not available atthis stage for an adequate risk assessment. Appropriate strategies of risk managementand risk communication can be put in place only after further data are generated andanalysed.

Histamine Fish Poisoning
62
5.2 Nature and magnitude of risk
5.2.1 Impact on human health
HFP does not have a major impact on human health, at least in Australia. It is a mildbut unpleasant disease, likely to cause discomfort for only a few hours. The morbidityrate may approach 100% following the consumption of toxic fish, but mortality isextremely rare. The disease may be more serious in people taking certain medicines orwith pre-existing pulmonary, cardiac or renal problems.
The reports of HFP in Australia and New Zealand in the scientific literature (Smart1992; Brown 1993; Foo 1975a; Mitchell 1993) are not sufficient to indicate thefrequency of occurrence of HFP in these countries. To rely on these reports forinformation would suggest that the disease is rare. Because HFP is generally mild andof short duration, and responsive to antihistamine therapy, most outbreaks are unlikelyto be reported, even in the daily press.
However, HFP is important from the food safety aspect. It is possible that products,particularly imports, will escape the random monitoring safety net from time to time.Consumers are becoming more demanding, and do not expect food to make them ill.Litigation following food poisoning incidents is becoming more common andproducers, distributors and restaurants will increasingly be held liable for the qualityof the products they handle and sell.
5.2.2 Impact on fishing industries
In 1996–97 Australia produced fishery products worth $1760 million, with exportsworth $1290 million. Total finfish production was about 127 kt worth $454 million(ABS 1999).
If a major outbreak of HFP were to occur in Australia, as they have in Japan and theUnited States in the past, resulting media attention would affect fish consumption andhave a negative impact on the marketing of seafood. An outbreak in another countrycaused by our exports would seriously affect trade. Although such events arebecoming increasingly less likely because of the widespread adoption of HACCPanalysis and quality assurance, they will still occur occasionally owing to incidentssuch as those caused by equipment failure, human error or negligence.
5.3 Uncertainties and problem areas in risk characterisation
5.3.1 Defining histamine fish poisoning and elucidating its pathogenesis
The mechanism of toxicity of HFP is still not clearly understood, which isunsatisfactory. More research is needed to determine the threshold of toxic dose forhistamine and the role that potentiators or other toxins may play in causing thedisease. It is particularly important that HFP be clearly differentiated from syndromescaused by endogenous toxins such as ciguatoxins that may be present in finfish fromtime to time.

Histamine Fish Poisoning
63
As suggested by Mitchell (1993), there may be potential in fractionation of thechemical constituents of toxic fish and assessment of the toxicity of different singleand combined fractions. In addition, it is the view of the authors that little attentionhas been given to the pKa values of the carboxyl, imidazole and amine groups of thecompounds discussed in this review, although pKa greatly influences biologicalproperties. A list of pKa values can be found in any handbook of data for biochemicalresearch (e.g. Dawson et al 1969), and is the pH at which half of the ionisable group isin the dissociated and half is in the unionised form. It is significant that the pKa of theimidazole group is in the normal physiological range. Similarly, in separating thesecompounds found in spoiling fish chromatographically, little consideration has beengiven to the pH of the solvent or to the stabilising effects of solvent counter-ions.
Future regulatory action should take into account the impact of potentiators, and theidentification of potentiators in various foods may be essential (Hui and Taylor 1985).When analysing suspect fish for histamine, there may be advantage in simultaneousdetection of other common putrefactive amines, such as cadaverine and putrescine(Taylor and Sumner 1986).
The safety of urocanic acid has been investigated only in relation to its use in cosmeticproducts and sunscreens (Cosmetic Ingredient Review Expert Panel 1995). Mostanimal experiments have involved topical administration and the toxicity profile isincomplete. Extensive animal data indicated that the substance hasimmunosuppressant properties, but clinical data were inconclusive. The CosmeticIngredient Review Expert Panel reported that ’it cannot be concluded that urocanicacid is safe in cosmetic formulations’. The possible role of urocanic acid in HFP, as apotential mast cell degranulator, may be a suitable research project, beginning withoral dosing studies in laboratory animals.
When the pathogenesis of the disease is understood, clinical HFP will be able to bedefined more accurately and making a differential diagnosis will be easier. There is aneed to assess the actual incidence of fish allergy and to determine what percentage ofcases diagnosed as fish allergy represent misdiagnosis and misclassification of HFP.The latter is differentiated from allergy by the occurrence of clusters of affectedpeople rather than single cases, and by the application of skin tests using non-toxicfish extracts.
5.3.2 Investigating and managing post-harvesting contamination
More research still needs to be done on post-harvesting contamination in order toimprove quality control procedures. Identification of points at which temperatureabuse and/or bacterial contamination occur would include the gathering ofinformation, where this has not been done already, on current fish harvesting,transporting, storage, processing and retailing practices for the species identified asthose most likely to cause HFP. For example, temperature abuse may occur onrecreational boats that lack adequate refrigeration; and Gellert et al (1992) suggestedthat rinsing fish in seawater during handling and cleaning could promotecontamination with microorganisms capable of metabolising histidine to histamine.

Histamine Fish Poisoning
64
Regardless of the origin of the spoilage bacteria, histamine serves as an indicator ofmicrobial spoilage in histidine-containing fish, and many countries includingAustralia have defined maximum histamine levels for fish products. Accumulation ofextensive data on levels of histamine in retail fish and fish products in Australiawould produce a better understanding of the likely incidence of HFP and provide areference point to measure future food hygiene standards. This could be carried outeither in special surveys or the work could be included in routine sampling programsfor other contaminants. Given the variety of species that may potentially contain highlevels of histamine, perhaps all finfish should be monitored. A rapid and cheap assayfor detecting histamine in fish would be of value if made available to the public and,in particular, to recreational fishers. The current ELISA tests are rapid, but not cheap(Lea Traeger, pers. comm. 1999).
Histamine analysis as a means of monitoring spoilage is associated with a number ofproblems. It is time consuming and/or expensive, and samples need to be taken frommultiple sites to compensate for variations in histamine content throughout thedecomposed fish. Perhaps more important is the lack of standardisation of histaminedetection methods employed around the world, and the multitude of tests available.There is a need for global standardisation of histamine detection methods, andlaboratory accreditation and proficiency testing. For all these reasons, together withthe fact that monitoring fish histamine levels may not always ensure protection fromHFP, a method other than direct measurement of histamine may be preferable forquantitative measurement of quality deterioration. Baronowski (1985) suggested thaturocanic acid may be a useful alternative to histamine as a spoilage index inscombroid and other fish that are rich in endogenous histidine. This idea should beinvestigated, especially in the light of new knowledge that urocanic acid is a mast celldegranulator (Wille et al 1999).
A study of the correlation between the amine content and bacterial counts needs to bedone. Ideally, each amine would be correlated with its respective amine-producingbacteria. The detection of histamine, cadaverine and putrescine can be achievedsatisfactorily by TLC or HPLC. As for the development of a medium for detection andenumeration of the amine-producing bacteria, only HDB have received considerableattention. Rapid tests for lysine decarboxylase activity (Brooker et al 1973) andornithine decarboxylase activity (Fay and Barry 1972) could give results within 4 h ofincubation, but a solid medium that would allow direct enumeration of these activitiesis not available (Leung 1987).
Government regulatory agencies require the removal of contaminated fish from themarketplace when human illness occurs and the fish is demonstrated to containunusually high levels of histamine. For this reason, reporting of suspected cases ofHFP to local food authorities can lead to removal of contaminated fish from themarketplace and prevention of additional cases.
Mechanisms need to be in place to allow efficient and complete traceback ofincriminated fish to point of origin, in order to rectify problems leading to spoilage. Atpresent in Australia, tracing fish from commercial outlets is difficult, because of alack of a ticketing system, especially in the middle (wholesaler) stages of distribution

Histamine Fish Poisoning
65
(Rawlin and Herfort 1999). This is a problem area that could be improvedconsiderably, given the necessary resources and cooperation between stakeholders.
Quality control programs operated by the fish processing industry can lead to theremoval of contaminated fish from the marketplace. For example, such programs inthe tuna canning industry have largely eliminated histamine poisoning as a concernwith canned tuna (Taylor et al 1989; Lea Traeger, pers. comm. 1999).

Histamine Fish Poisoning
66
REFERENCES
Ababouch, L (1997) Application of the HACCP principles in the fish-canning industry in Morocco, pp.302–10. In Fish inspection, quality control, and HACCP: A global focus, eds RE Martin et al.Proceedings of the Conference held 19–24 May 1996, Arlington, Virginia. Technomic Publishing Co.Inc., Lancaster, Basel.
Ababouch, L et al (1986) Histamine levels in commercially processed fish in Morocco. Journal ofFood Protection 49(11):904–8.
ABS (1999) Year Book Australia. Australian Bureau of Statistics, Canberra.
Aitken, L (1997) A regulatory approach to HACCP — the Australian perspective, pp. 613–8. In Fishinspection, quality control, and HACCP: A global focus, eds RE Martin et al. Proceedings of theConference held 19–24 May 1996, Arlington, Virginia. Technomic Publishing Co. Inc., Lancaster,Basel.
Andersen, EM (1997) Quality assurance on board fishing vessels, pp. 427–36. In Fish inspection,quality control, and HACCP: A global focus, eds RE Martin et al. Proceedings of the Conference held19–24 May 1996, Arlington, Virginia. Technomic Publishing Co. Inc., Lancaster, Basel.
Anggawati, A (1986) Unpublished results. ACIAR report on visit by RIFT personnel Australia inOctober – December 1986.
Anon. (1988a) Chapter 9. Dried and smoked fishery products: Preparation and composition, Table 10,Cold smoking, p. 156. In: JR Burt (ed) Fish smoking and drying: The effect of smoking and drying onthe nutritional properties of fish. Elsevier Applied Science, London and New York.
Anon. (1988b) Chapter 9. Dried and smoked fishery products: Preparation and composition, Table 8,Hot smoking, p. 148. In: JR Burt (ed) Fish smoking and drying: The effect of smoking and drying onthe nutritional properties of fish. Elsevier Applied Science, London and New York.
Anon. (1998a) New system for seafood safety. Environmental Health Perspectives 106(10):A475−8.
Anon. (1998b) The Food Safety (Fishery Products and Live Shellfish) (Hygiene) Regulations 1998. SINo. 994. HMSO, London. (Cited by Scoging 1998.)
Anon. (1999) Rapid tests for seafood quality: Histamine detection. Food Australia 51(3):102.
ANZFA (1998a) Standard D1 – Fish, in Australian Food Standards Code (Issue 41, 18 November1998). Australian and New Zealand Food Authority, Canberra.
ANZFA (1998b) Development of Joint Australia New Zealand Food Standards as part of the process ofthe Review of the Food Standards Code. Fish and fish products. Explanatory notes, proposal/fullassessment, P183, August 1998.
AOAC (1995a) Histamine in seafood: Biological method, pp. 14–5. Sec. 35.1.30, Method 954.04. InOfficial Methods of Analysis of AOAC International, 16th ed., PA Cunniff, ed. Association of OfficialAnalytical Chemists (AOAC) International, Gaithersburg, MD, USA.
AOAC (1995b) Histamine in seafood: Chemical method, pp. 15–6. Sec 35.5.31, Method 957.07. InOfficial Methods of Analysis of AOAC International, 16th ed., PA Cunniff, ed. Association of OfficialAnalytical Chemists (AOAC) International, Gaithersburg, MD, USA.
AOAC (1995c) Histamine in seafood: Fluorometric method, pp. 16–7. Method 35.1.32, Method977.13. In Official Methods of Analysis of AOAC International, 16th ed., PA Cunniff, ed. Associationof Official Analytical Chemists (AOAC) International, Gaithersburg, MD, USA.

Histamine Fish Poisoning
67
Arnold, SH and Brown, WD (1978) Histamine toxicity from fish products. Advances in Food Research34:113−54.
Arnold, SH et al (1980) Histamine formation by bacteria isolated from skipjack tuna, Katsuwonaspelamis. Bulletin of the Japanese Society of Scientific Fisheries 46:991–5. (Cited by Institute ofMedicine 1991.)
Ascione, A et al (1997) [Two cases of 'scombroid syndrome' with severe cardiovascular compromise.]Cardiologia 42(12):1285–8. (English abstract only.)
Barancin, CE et al (1998) Plasmid-mediated histamine biosynthesis in the bacterial fish pathogenVibrio anguillarum. Plasmid 39(3):235–44. (Abstract only.)
Baranowski, JD (1985) Low-temperature production of urocanic acid by spoilage bacteria isolated frommahimahi (Coryphaena hippurus). Applied and Environmental Microbiology 50(2):546–7.
Barker, J and McKenzie, A (1997) Review of HACCP and HACCP-based food control systems, pp.73–81. In Fish inspection, quality control, and HACCP: A global focus, eds RE Martin et al.Proceedings of the Conference held 19–24 May 1996, Arlington, Virginia. Technomic Publishing Co.Inc., Lancaster, Basel.
Bartholomew, BA et al (1987) Scombrotoxic fish poisoning in Britain: Features of over 250 suspectedincidents from 1976 to 1986. Epidemiology and Infection 99:775–82.
Bartram, PK (1997) Proper processing and handling of raw tuna, pp. 233–7, and Proper shipboardhandling of fresh tuna for the sashimi market, pp. 448–57. In Fish inspection, quality control, andHACCP: A global focus, eds RE Martin et al. Proceedings of the Conference held 19–24 May 1996,Arlington, Virginia. Technomic Publishing Co. Inc., Lancaster, Basel.
Bateman, RC et al (1994) Copper chelation assay for histamine in tuna. Journal of Food Science59(3):517–8. (Cited by Price 1999.)
Behling, AR and Taylor, SL (1982) Bacterial histamine production as a function of temperature andtime of incubation. Journal of Food Science 47:1311-7.
Ben-Gigirey, B et al (1998) Changes in biogenic amines and microbiological analysis in albacore(Thunnus alalunga) muscle during frozen storage. Journal of Food Protection 61(5):608−15.
Bjeldanes, LF et al (1978) On the aetiology of scombroid poisoning: Cadaverine potentiation ofhistamine toxicity in the guinea pig. Food and Cosmetic Toxicology 16(2):157−9.
Blakesley, ML (1983) Scombroid poisoning: Prompt resolution of symptoms with cimitedine. Annals ofEmergency Medicine 12:104.
Bremer, PJ et al (1998) Thermal death times of Hafnia alvei cells in a model suspension and inartificially contaminated hot-smoked kahawai (Arripis trutta). Journal of Food Protection61(8):1047−51.
Brooker, DC et al (1973) Rapid test for lysine decarboxylase activity in Enterobacteriaciae. AppliedMicrobiology 25:622–3. (Cited by Leung 1987.)
Brown, C (1993) Scombroid poisoning (Letter). Medical Journal of Australia 158(6):435−6.
Buchanan, R et al (1998) Principles of risk assessment for illness caused by foodborne biologicalagents. Journal of Food Protection 61(8):1071−4.
Cantoni, C et al (1976) [Tuna flesh deterioration and histamine levels.] Arch. Vet. Ital. 27:60–3. (Citedby Arnold and Brown 1978.)

Histamine Fish Poisoning
68
Caron, JC et al (1982) High-performance liquid chromatographic determination of urocanic acidisomers in biological samples. Journal of Chromatography 230:125–31.
Cavanah, DK and Casale, TB (1993) Chapter 11. Histamine, pp. 321–42. In The mast cell in health anddisease, MA Kaliner and DD Metcalfe, eds. Marcel Dekker, Inc., New York, Basel and Hong Kong.
Chen, K-T and Malison, MD (1987) Outbreak of scombroid fish poisoning, Taiwan. American Journalof Public Health 77(10):1335–6.
Chin, KW et al (1989) The histamine content of oriental foods. Food and Chemical Toxicology27(5):283−7.
Chng, NM and Kuang, HK (eds) (1987) Smoked products, p. 28. In: Southeast Asian fish productsmarine fisheries research and development. Southeast Asian Fisheries Development Center, Singapore.
Chu, CH and Bjeldanes, LF (1981) Effect of diamines, polyamines and tuna fish extracts on the bindingof histamine to mucin in vitro. Journal of Food Science 47 79−88.
Clark, RB (1997) Scombroid poisoning revisited. Letter. Annals of Emergency Medicine 29(3):426–7.
Clifford, MN et al (1989) Studies with volunteers on the role of histamine in suspectedscombrotoxicosis. Journal of the Science of Food and Agriculture 47(3):365–75.
Clifford, MN et al (1991) Is there a role for amines other than histamines in the aetiology ofscombrotoxicosis? Food Additives and Contaminants 8(5):641–52.
Clifford, MN et al (1993) Do saxitoxin-like substances have a role in scombrotoxicosis? FoodAdditives and Contaminants 6:657–67.
Codex Alimentarius Commission (1996) Agenda item 8, CX/FH 96/10: Principles and guidelines forthe application of microbiological risk assessment. Codex Committee on Food Hygiene, 29th Session,Washington, DC, 21–25 October, 1996. Joint FAO/WHO Food Standards Programme.
Cosmetic Ingredient Review Expert Panel (1995) Final report on the safety assessment of urocanic acid.Journal of the American College of Toxicology 14(5):386–423.
Dawson, RMC et al (1969) Data for biochemical research, 2nd ed. Oxford Clarendon Press.
Doe, PE et al (1998) Chapter 5. Methodology for quality measurements, pp. 117–35. In Fish dryingand smoking: Production and quality, PE Doe, ed. Technomic Publishing Co., Inc., Lancaster, Basel.
Downs, M (1997) Scombroid poisoning revisited. Annals of Emergency Medicine 29(3):426−7.
Draisci, R et al (1993) Chromatographia 35:584. (Cited by Draisci et al 1998.)
Draisci, R et al (1998) Improved ion chromatography-integrated pulsed amperometric detection methodfor the evaluation of biogenic amines in food of vegetable or animal origin and in fermented foods.Journal of Chromatography A 798(1−2):109−16.
Edwards, RA et al (1987) Journal of Applied Bacteriology 63:427. (Cited by Draisci et al 1998.)
Eitenmiller, RR et al (1981) Production of histidine decarboxylase and histamine by Proteus morganii.Journal of Food Protection 44(11):815–20.
Eitenmiller, RR et al (1982) Histamine formation in fish: Microbial and biochemical conditions, pp.39–50. In Chemistry and biochemistry of marine food products, RE Martin et al, eds. Avi PublishingCo., Westport, CT.

Histamine Fish Poisoning
69
Eitenmiller, RR and De Souza, SC (1984) Chapter 6. Enzymic mechanisms for amine formation in fish,pp. 431–42. In Seafood toxins, P. Ragelis, ed. American Chemical Society, Washington, DC. (Cited byRawles 1996.)
Etkind, P et al (1987) Bluefish-associated scombroid poisoning. An example of the expanding spectrumof food poisoning from seafood. Journal of the American Medical Association 258(23):3409−10.
Fay, GD and Barry, AL (1972) Rapid ornithine decarboxylase test for the identification ofEnterobacteriaceae. Applied Microbiology 23:710–13.
FAO (1999) FAO Expert Consultation on the Trade Impact of Listeria in Fish Products. University ofMassachusetts, Amhurst, MA, USA, 17–20 May 1999.
FDA (1992) Foodborne pathogenic microorganisms and natural toxins. Center for Food Safety andApplied Nutrition, Rockville, Maryland, 161 pp. (Cited by Lipp and Rose 1997.)
FDA (1998) FDA and EPA guidance levels. Appendix 5, pp. 245–8. In Fish and fishery productshazards and controls guide, 2nd ed. Department of Health and Human Services, Public Health Service,Food and Drug Administration, Center for Food Safety and Applied Nutrition, Office of Seafood,Washington, DC.
FDA (1999) Foodborne Pathogenic Microorganisms and Natural Toxins Handbook. US Center forFood Safety and Applied Nutrition. Food and Drug Administration website http://www-micro.msb.le.ac.uk/others/FDA/~mow/chap38.html, 19/6/99.
Ferencik, M (1970) Formation of histamine during bacterial decarboxylation of histidine in the flesh ofsome marine fishes. Journal of Hygiene, Epidemiology, Microbiology and Immunology 14:52–60.(Cited by Arnold and Brown 1978; Bremer et al 1998.)
Fernández-Salguero, J and Mackie, IM (1979) Histidine metabolism in mackerel (Scomber scombrus).Studies on histidine decarboxylase activity and histamine formation during storage of flesh and liverunder sterile and non-sterile conditions. Journal of Food Technology 14:131–9.
Fletcher, GC et al (1995) Histamine and histidine in New Zealand marine fish and shellfish species,particularly kahawai (Arripis trutta). Journal of Aquatic Food Production Technology 4:53–74. (Citedby Fletcher et al 1998.)
Fletcher, GC et al (1998) Levels of histamine and histamine-producing bacteria in smoked fish fromNew Zealand markets. Journal of Food Protection 61(8):1064−70.
Foo, LY (1975a) Scombroid-type poisoning induced by the ingestion of smoked kahawai. New ZealandMedical Journal 81:476–7.
Foo, LY (1975b) The content of histamine and fish poisoning. New Zealand Medical Journal 82:381–3.
Foo, LY (1976) Scombroid poisoning. Isolation and identification of 'saurine'. Journal of the Science ofFood and Agriculture 27:807–10.
Foo, LY (1977) Scombroid poisoning — recapitulation on the role of histamine. New Zealand MedicalJournal 85:425–7.
Frank, HA (1985) VI. 1. Use of normographs to estimate histamine formation in tuna, pp. 18–20. InHistamine formation in marine products: Production by bacteria, measurement and prediction offormation. FAO Fisheries Technical Paper No. 252. Food and Agriculture Organization of the UnitedNations, Rome.

Histamine Fish Poisoning
70
Frank, HA and Baranowski, JD (1984) Authentic fishery product samples, collected and decomposedunder controlled conditions. Contract 223-80-2295. US Department of Health and Human Services,Washington, DC. (Cited by Baranowski 1985.)
Frank, HA et al (1981) Histamine formation and honeycombing during decomposition of skipjack tuna,Katsuwonus pelamis, at elevated temperatures. Marine Fish Review 43(10):9–14.
Frank, HA et al (1985) Identification and decarboxylase activities of bacteria isolated fromdecomposed mahimahi (Coryphaena hippurus) after incubation at 0 and 32oC. International Journal ofFood Microbiology 2:331–40. (Cited by Rawles et al 1996.)
Fujii, T et al (1994) Viability and histidine decarboxylase activity of halophilic histamine-formingbacteria during frozen storage. Journal of Food Protection 57:611–3.
Fujii, T et al (1997) Identification of the psychrophilic histamine-producing marine bacteria previouslyreferred to as the N-group bacteria. Fish Science 63:807–10.
Furuta, T et al (1996) Pharmacokinetics of stable isotopically labelled L-histidine in humans and theassessment of in vivo histidine ammonia lysase activities. Drug Metabolism and Disposition 24(1)49–54.
Gale, EF (1942) The oxidation of amines by bacteria. Biochemistry Journal 36:54. (Cited by Taylor1986.)
Gale, EF (1946) Bacterial amino acid decarboxylases. In Advances in enzymology 6:1–32. IntersciencePublishers, New York. (Cited by Arnold and Brown 1978; Taylor et al 1978.)
Garland, CD (1985). Unpublished results. (Cited by Leung 1987.)
Geiger, E (1944a) Histamine content of processed and uncanned fish. A tentative method ofquantitative determination of spoilage. Food Research 9:293−7.
Geiger, E (1944b) On the specificity of bacterium decarboxylase. Proceedings of the Society ofExperimental Biology and Medicine 55:11–13. (Cited by Arnold and Brown 1978.)
Geiger, E (1955) Role of histamine in poisoning with spoiled fish. Science 121:865–6. (Cited byArnold and Brown 1978.)
Geiger, E et al (1944) The content and formation of histamine in fish muscle. Archives of Biochemistry3:311–9. (Cited by Arnold and Brown 1978.)
Gellert, GA et al (1992) Scombroid fish poisoning. Underreporting and prevention amongnoncommercial recreational fishers. Western Journal of Medicine 157(6):645−7.
Gessner, B et al (1996) Scombrotoxicosis-like illness following the ingestion of smoked salmon thatdemonstrated low histamine levels and high toxicity on mouse bioassay. Clin. Infect. Dis. 23:1316–8.
Gietzen, K et al (1996) Imidazole fungicides provoke histamine release from mast cells and induceairway contraction. Experimental Toxicology and Pathology 48(6):529–31. (Abstract only.)
Gilbert, RJ et al (1980) Scombrotoxic fish poisoning: Features of the first 50 incidents to be reported inBritain (1976–9). British Medical Journal 281:71–2.
Granerus, G (1968) Effects of oral histamine, histidine and diet on urinary excretion of histamine,methylhistamine, and 1-methyl-4-imidazoleacetic acid in man. Scandinavian Journal of ClinicalLaboratory Investigation 22(Suppl. 104):49–58.
Guss, DA (1998) Scombroid fish poisoning: Successful treatment with cimetidine. Undersea HyperbMed 25(2):123−5.

Histamine Fish Poisoning
71
Hardy, R and Smith, JGM (1976) The storage of mackerel (Scomber scombrus). Development ofhistamine and rancidity. Journal of Science of Food and Agriculture 27:595–9. (Cited by Pan 1985.)
Hart, PH et al (1997) Histamine involvement in UVB-and cis-urocanic acid-induced systemicsuppression of contact hypersensitivity responses. Immunology 91(4):601–8.
Havelka, B (1967) Role of Hafnia bacteria in the rise of histamine in tuna fish meat. Cesk. Hyg.12:343–52. (Cited by Taylor 1985.)
Helander, CG et al (1965) The renal removal of 14C-labelled histamine from the blood in man.Scandinavian Journal of Clinical Laboratory Investigations 17:524-8.
Henderson, PB (1830) Case of poisoning from the bonito (Scomber pelamis). Edinburgh MedicalJournal 34:317–8. (Cited by Taylor 1986; Mitchell 1993.)
Hesterberg, R et al (1984) Histamine content, diamine oxidase activity and histamine methyltransferaseactivity in human tissues: fact or fiction? Agents Actions 14(3/4):325–34.
Hughes, JM et al (1977) Foodborne disease outbreaks of chemical aetiology in the United States,1970–1974. American Journal of Epidemiology 105(3):233–44.
Hughes, RB (1959) Chemical studies on the herring (Culpea harengus). 2. The free amino acids ofherring flesh and their behaviour during post-mortem spoilage. Journal of Science of Food andAgriculture 10:558–64.
Hui, JY and Taylor, SL (1983) High pressure liquid chromatographic determination of putrefactiveamines in foods. Journal of AOAC International 66(4):853–7. (Cited by Wagener 1984.)
Hui, JY and Taylor, SL (1985) Inhibition of in vivo histamine metabolism in rats by foodborne andpharmacologic inhibitors of diamine oxidase, histamine N-methyl transferase, and monoamine oxidase.Toxicology and Applied Pharmacology 8:241–9.
Hungerford, JM et al (1990) Selective determination of histamine by flow injection analysis. AnalyticalChemistry 62:1971–6. (Cited by Price 1999.)
Hungerford, J et al (1997) Rapid chemical tests: Laboratory and field screening methods for seafoodanalysis, pp. 538–50. In Fish inspection, quality control, and HACCP: A global focus, eds RE Martinet al. Proceedings of the Conference held 19–24 May 1996, Arlington, Virginia. Technomic PublishingCo. Inc., Lancaster, Basel.
Husain, SS (1996) Effect of traditional preservatives on the growth of histamine formers. Dissertationsubmitted in partial fulfilment of the requirements for the degree of Master of Science in MarineBiology and Oceanology. Centre of Advanced Study in Marine Biology, Annamalai University,Parangipettai, India.
Ienistea, C (1971) Bacterial production and destruction of histamine in foods, and food poisoningcaused by histamine. Die Nahrung 15:109–13. (Cited by Taylor et al 1978a.)
Ienistea, C (1973) Significance and detection of histamine in food. In The microbiological safety offoods, eds BC Hobbs and JHB Christian. Academic Press, New York. (Cited by Leung, 1987.)
Ijomah, P et al (1991) The importance of endogenous histamine relative to dietary histamine in theaetiology of scombrotoxicosis. Food Additives and Contaminants 8(4):531–42.
Ijomah, P et al (1992) Chapter 17. Further volunteer studies on scombrotoxicosis, pp. 194–9. InPelagic fish: The resource and its exploitation, JR Burt et al, eds. Fishing News Books, Oxford.
Institute of Medicine (1991) Seafood safety. National Academy Press, Washington, DC, 486 pp.

Histamine Fish Poisoning
72
Irvine, WT et al (1959) Urinary output of histamine after a meat meal. Lancet 1:1061. (Cited byGranerus 1968.)
James, D and Olley, J (1985) IX. Summary and future research needs, pp. 47–51. In Histamine inmarine products: Production by bacteria, measurement and prediction of formation. FAO FisheriesTechnical Paper 252. Food and Agriculture Organization of the United Nations, Rome, 1985.
Johnson, LR and Overholt, BF (1967) Release of histamine into gastric venous blood flow followinginjury by acetic or salicyclic acid. Gastroenterology 52:505–9. (Cited by Foo 1977.)
Jung, HPK and Bjeldanes, LF (1979) Effects of cadaverine on histamine transport and metabolism inisolated gut sections of the guinea pig. Food and Cosmetic Toxicology 17:629–32.
Karnicki, ZS (1997) The impact of international trade and trade agreements on fish inspection andquality control, pp. 3–22. In Fish inspection, quality control, and HACCP: A global focus, eds REMartin et al. Proceedings of the Conference held 19–24 May 1996, Arlington, Virginia. TechnomicPublishing Co. Inc., Lancaster, Basel.
Karolus, JJ et al (1985) Presence of histamine in the bluefish, Pomatomus saltatrix. Journal of FoodProtection 48(2):166–8.
Kawabata, T et al (1955) Studies on the allergy-like food poisoning associated with putrefaction ofmarine products. II. Separation of causative substance and some of its chemical characteristics.Japanese Journal of Medical Science and Biology 8:503–19.
Kawai, A and Sakaguchi, M (1968) Histidine metabolism in fish. 2. Formation of urocanic,formimoglutamic, and glutamic acids from histidine in livers of carp and mackerel. Bulletin of theJapanese Society of Scientific Fisheries 34:507–11.
Kerr, GW and Parke, TR (1998) Scombroid poisoning a pseudoallergic syndrome. Journal of theRoyal Society of Medicine 91(2):83−4.
Kim, I-S (1978) Simple analysis of amines in canned tuna fish. MS Thesis, University of California,Berkeley, CA. (Cited by Bjeldanes et al 1978.)
Kim, R (1979) Flushing syndrome due to mahimahi (scombroid fish) poisoning. Archives ofDermatology 115:963−5.
Kimata, M (1961) The histamine problem, pp. 329-52. In Fish as food: Production, biochemistry andmicrobiology, ed. G Borgstrom. Academic Press, New York.
Kindred, TP (1996) Risk analysis and its application in FSIS. Journal of Food Protection 1996Supplement, pp. 24–30.
Klasusen, NK and Lund, E (1986) Formation of biogenic amines in herring and mackerel. Z LebensmUnters Forsch 182(6):459−63.
Klausen, NK and Huss, HH (1987a) A rapid method for detection of histamine-producing bacteria.International Journal of Food Microbiology 5:137–46. (Cited by Stratton and Taylor 1991).
Klausen, NK and Huss, HH (1987b) Growth and histamine production by Morganella morganii undervarious temperature conditions. International Journal of Food Microbiology 5:147–56.
Lange, WR (1988) Scombroid poisoning. American Family Physician 37:163–8.
Lerke, PA and Bell, LD (1976) A rapid fluorometric method for the determination of histamine incanned tuna. Journal of Food Science 41:1282–4. (Cited by Price 1999.)

Histamine Fish Poisoning
73
Lerke, PA et al (1978) Scombroid poisoning. Report of an outbreak. Western Journal of Medicine129:381–6.
Lerke, PA et al (1983) Screening test for histamine in fish. Journal of Food Science 48:155–7.
Leung, E (1987) Amine producing bacteria isolated from chub mackerel (Rastrelliger sp.). A thesis inpartial fulfilment of the requirements for the degree of Bachelor of Agricultural Science with Honours,University of Tasmania.
Lipp, EK and Rose, JB (1997) The role of seafood in foodborne diseases in the United States ofAmerica. Rev. Sci. Tech. 16(2):620−40.
Lopez-Sabater, EI et al (1996) Incidence of histamine-forming bacteria and histamine content inscombroid fish species from retail markets in the Barcelona area. International Journal of FoodMicrobiology 28(3):411−8.
Love, RM (1980) Histidine, p. 427. In The chemical biology of fishes, Vol 2: Advances 1968–77.Academic Press, London, New York, Toronto, Sydney and San Francisco.
Lyons, DE et al (1983) Cadaverine and aminoguanidine potentiate the uptake of histamine in vitro inperfused intestinal segments of rats. Toxicology and Applied Pharmacology 70:445–58.
McInerney, J et al (1996) Scombroid poisoning. Annals of Emergency Medicine 28(2):235−8.
Mackie, IM and Fernández-Salguéro (1977) Histidine metabolism in fish. Urocanic acid in mackerel(Scomber scombrus). Journal of Science of Food and Agriculture 28:935–40.
McMeekin, TA (1986) Personal observation cited by Anggawati (1986).
McMeekin, TA et al (1993) Chapter 10. A thermodynamic approach to bacterial growth, pp. 287–309.In Predictive microbiology: Theory and application. Research Studies Press Ltd and John Wiley &Sons Inc., England and New York.
Maire, R et al (1992) [Incidence and clinical aspects of scombroid fish poisoning.] Schweiz MedWochenschr 122(50):1933–5. (English abstract only.)
Merson, MH et al (1974) Scombroid fish poisoning. Outbreak traced to commercially canned tuna fish.Journal of the American Medical Association 228(10):1268–9.
Mietz, JL and Karmas, E (1977) Chemical quality index of canned tuna as determined by HPLC.Journal of Food Science 42:155−8.
Miller, BM et al (1997) A rapid, easily used test kit to determine histamine concentrations in fish, pp.551–9. In Fish inspection, quality control, and HACCP: A global focus, eds RE Martin et al.Proceedings of the Conference held 19–24 May 1996, Arlington, Virginia. Technomic Publishing Co.Inc., Lancaster, Basel.
Mines, D et al (1997) Poisonings: Food, fish, shellfish. Emergency Medicine Clinics of North America15(1):157−77.
Mitchell, JW (1984) Scombroid food poisoning from fish. New Zealand Medical Journal 97:127.(Cited by Mitchell 1993.)
Mitchell, JW (1993) Scombrotoxic fish poisoning. A report prepared for the Ministry of Health.Institute of Environmental Health and Forensic Sciences Limited, Mt Albert Science Centre, NZ, 18 pp.
Mitchell, JW and O'Brien, G (1992) Food poisoning — and mackerel. New Zealand Medical Journal105:929. (Cited by Mitchell 1993.)

Histamine Fish Poisoning
74
Mitchell, RG and Code, CF (1954) Effect of diet on urinary excretion of histamine. Journal of AppliedPhysiology 6:387–91. (Cited by Rawles et al 1996.)
Molinari, G et al (1989) [Hygiene and health importance of histamine as an unhealthy factor in severalfood products.] Ann Ig 1(3–4):637–46. (English abstract only.)
Mongar, JL (1957) Effect of chain length of aliphatic amines on histamine potentiation and release.British Journal of Pharmacology 12:140–8.
Morii, H et al (1986) Formation of histamine by luminous bacteria isolated from scombroid fish.Bulletin of the Japanese Society of Scientific Fisheries 52:2135–41. (Cited by Stratton et al 1991.)
Morrison, H et al (1980) Analysis of Z and E isomers of urocanic acid by high performance liquidchromatography. Journal of Chromatography 183:83–6.
Mopper, B and Sciacchitano, CJ (1994) Capillary zone electrophoretic determination of histamine infish. Journal of AOAC International 77(4):881−4.
Morrow, JD et al (1991) Evidence that histamine is the causative toxin of scombroid-fish poisoning.New England Journal of Medicine 324(11):716−20.
Motil, KJ and Scrimshaw, NS (1979) The role of exogenous histamine in scombroid poisoning.Toxicology Letters 3:219–23.
Müller, GJ et al (1992) Scombroid poisoning. Case series of 10 incidents involving 22 patients. SouthAfrican Medical Journal 81(8):427−30.
Niven, CF et al (1981) Differential plating medium for quantitative detection of histamine-producingbacteria. Applied Environmental Microbiology 41(1):321−2.
Ohashi, M et al (1994) Oxygen-sensor-based simple assay of histamine in fish using purified amineoxidase. Journal of Food Science 59(3):519–22. (Cited by Price 1999.)
Okuzumi, M et al (1981) Isolation of psychrophilic and halophilic histamine-forming bacteria fromScomber japonicus. Bulletin of the Japanese Society of Scientific Fisheries 47:1591–8.
Okuzumi, M et al (1984a) Occurrence of various histamine-forming bacteria on/in fresh fishes. Bulletinof the Japanese Society of Scientific Fisheries 50(1):161–7.
Okuzumi, M et al (1984b) Changes in numbers of histamine-forming bacteria on/in common mackerelstored at various temperatures. Bulletin of the Japanese Society of Scientific Fisheries 50(4):653–7.
Olley, J (1972) Unconventional sources of fish protein. CSIRO Food Research Quarterly 32(2):27–32.
Olley, J and Baranowski, J (1985) V. Temperature effects on histamine formation, pp. 14–17. InHistamine formation in marine products: Production by bacteria, measurement and prediction offormation. FAO Fisheries Technical Paper No. 252. Food and Agriculture Organization of the UnitedNations, Rome.
Olley, J and McMeekin, TA (1985) VI. 3. Use of temperature function integrators, pp. 24–29. InHistamine formation in marine products: Production by bacteria, measurement and prediction offormation. FAO Fisheries Technical Paper No. 252. Food and Agriculture Organization of the UnitedNations, Rome.
Olley, J, Orejana, F and Pan, BS (1985) VII. 4. Effect of gutting, p. 37. In Histamine formation inmarine products: Production by bacteria, measurement and prediction of formation. FAO FisheriesTechnical Paper No. 252. Food and Agriculture Organization of the United Nations, Rome.

Histamine Fish Poisoning
75
Omura, Y et al (1978) Histamine-forming bacteria isolated from spoiled skipjack tuna and jackmackerel. Journal of Food Science 43:1779–81. (Cited by Baranowski 1985.)
Orejana, FM et al (1983) Histamine production in iced, smoked/dried frigate mackerel. Journal ofFood Science Technology 7(2):30–8.
Pan, BS (1985a) VI. 2. Use of Arrhenius plot to estimate histamine formation in mackerel and bonito,pp. 21–3. In Histamine formation in marine products: Production by bacteria, measurement andprediction of formation. FAO Fisheries Technical Paper No. 252. Food and Agriculture Organization ofthe United Nations, Rome.
Pan, BS (1985b) VIII. 2. Histamine formation in canning processes, pp. 41–44. In Histamine formationin marine products: Production by bacteria, measurement and prediction of formation. FAO FisheriesTechnical Paper No. 252. Food and Agriculture Organization of the United Nations, Rome.
Pan, BS (1988) Chapter 5. Formation of histamine toxicity. Undesirable factors in dried fish products,pp. 66–7. In Fish smoking and drying: The effect of smoking and drying on the nutritional properties offish, JR Burt, ed. Elsevier Applied Science, London and New York.
Parrot, J and Nicot, G (1966) Pharmacology of histamine, pp. 148–61. In Handbuch derExperimeutellin Pharmakologie, Vol. XVIII. Springer-Verlag, New York. (Cited by Taylor 1986;Shalaby 1996.)
Poulter, RG (1988) Chapter 7. Processing and storage of traditional dried and smoked fish products, pp.85–90. In Fish smoking and drying: The effect of smoking and drying on the nutritional properties offish, JR Burt, ed. Elsevier Applied Science, London and New York.
Price, RJ (1999) Compendium of fish and fishery product processes, hazards, and controls. NationalSeafood HACCP Alliance for Training and Education.(Http://seafood.ucdavis.edu/haccp/compendium/compend.htm, updated 8/9/99)
Ramesh, A and Venugopalan, VK (1986) Densities and characteristics of histamine-forming luminousbacteria of marine fish. Food Microbiology 3:103–5.
Rawles, DD et al (1996) Biogenic amines in fish and shellfish. Advances in Food and NutritionResearch 39:329–64.
Rawlin, G and Herfort, A (1999) Ciguatera detection in fish flesh and tracking of possible ciguatera-tainted fish in the marketplace. Internal advice note, Agriculture, Fisheries and Forestry, Australia.
Rice, SL et al (1976) Biologically active amines in food: A review. Journal of Milk and FoodTechnology 39(5):353–8.
Rigg, A (1997) Biogenic amines in fish and fish products. ACT Health Protection Service, Food Safetyand Compliance Program, Food Surveys 1996/97.(Http://www.health.act.gov.au/hps/foodsurvey/biogenic.html, 28/7/99).
Riley, JF (1958) The mechanisms of histamine release from mast cells. J Pharm. Pharmac. 10:271–2.(Cited by Olley 1972.)
Riley, JF (1959). In The mast cells, p. 130. Livingstone, Edinburgh and London. (Cited by Olley 1972.)
Ritchie, AH and Mackie, IM (1979) The formation of diamines and polyamines during storage ofmackerel (Scomber scombrus), pp. 489–94. In Advances in fish science and technology, ed. JJ Connell.Fishing News Books, Surrey, England.
Rogers, PL and Staruszkiewicz, W (1997) Gas chromatographic method for putrescine and cadaverinein canned tuna and mahimahi and fluorometric method for histamine (minor modification of AOAC

Histamine Fish Poisoning
76
Official Method 977.13): Collaborative study. Journal of AOAC International 80(3):591–602. (Abstractonly.)
Ross, T and McMeekin, TA (1995) Chapter 13. Predictive microbiology and HACCP, pp. 330–57. InHACCP in meat, poultry and fish processing, AM Pearson and TR Dutson, eds. Blackie Academic andProfessional, UK.
5XVVHOO��)(�DQG�0DUHWLü��=��������6FRPEURLG�SRLVRQLQJ��0LQL�UHYLHZ�ZLWK�FDVH�KLVWRULHV��Toxicon24(10):967−73.
Ryser, ET et al (1984) Histamine production by psychrotrophic pseudomonads isolated from tuna fish.Journal of Food Protection 47:378–80.
Sabroe, RA and Kobza Black, A (1998) Scombrotoxic fish poisoning. Clinical and ExperimentalDermatology 23(6):258–9. (Abstract only.)
Sánchez-Guerrero, IM et al (1997) Scombroid fish poisoning: A potentially life-threatening allergic-like reaction. Journal of Allergy and Clinical Immunology 100(3):433−4.
Sapin-Jaloustre, H and Sapin-Jaloustre, J (1957) [A little-known food poisoning: Histamine poisoningfrom tuna.] Concours Medical, Paris 79:2705–8. (Cited by Arnold and Brown 1978.)
Satake, K et al (1953) Bacterial oxidation of some primary amines. Journal of Biochemistry 40:299–315. (Cited by Taylor 1986.)
Scoging, AC (1991) Illness associated with seafood. CDR (Lond Eng Rev) 1(11):R117−22.
Scoging, AC (1998) Scombrotoxic (histamine) fish poisoning in the United Kingdom. CommunicableDiseases and Public Health 1(3):204−5.
Senanayake, N and Vyravanathan, S (1981) Histamine reactions due to ingestion of tuna fish (Thunnusargentivittatus) in patients on anti-tuberculosis therapy. Toxicon 19:184–5. (Cited by Taylor 1985.)
Shalaby, AR (1996) Significance of biogenic amines to food safety and human health. Food ResearchInternational 29(7):675−90.
Shewan, JM (1955) The nitrogenous extractives from fresh fish muscle. III. Comparison of several flatfishes and members of the herring–mackerel group. Journal of Science of Food and Agriculture 6:99–104.
Shewan, JM (1970) Bacteriological standards for fish and fishery products. Chemistry and Industry6:193–9.
Shewan, JM (1977) The bacteriology of fresh and spoiling fish and the biochemical changes induced bybacterial action, pp. 51–66. In Proceedings of Conference on the Handling, Processing and Marketingof Tropical Fish, P. Sutcliffe and J Disney, eds. Tropical Products Institute, London. (Cited byYoshinaga and Frank 1982.)
Shibatani, T et al (1974) Enzymatic production of urocanic acid by Achromobacter liquidum. AppliedMicrobiology 27:688–94.
Shultz, DE et al (1976) Rapid thin layer chromatographic method for the determination of histamine infish products. Journal of AOAC International 59(6):1224−5. (Cited by Price 1999.)
Simidu, W and Hibiki, S (1954) Studies on putrefaction of aquatic products. 14. Comparison onputrefaction of different kinds of fish (2). Bulletin of the Japanese Society of Scientific Fisheries20(4):302–4. (Cited by Pan 1985b.)

Histamine Fish Poisoning
77
Simidu, W and Hibiki, S (1955) Studies on putrefaction of aquatic products. 23. On the criticalconcentration of poisoning for histamine. Bulletin of the Japanese Society of Scientific Fisheries21:365–7. (Cited by Taylor 1986.)
Sjaastad, O and Sjaastad, OV (1974) Catabolism of orally administered 14C-histamine in man ActaPharmacol. Toxicol. 34:33. (Cited by Taylor 1986.)
Skovgaard, N and Ellemann, G (1978) Levendsmiddelbaren histaminforgiftning. Ugeskr. Laeg.140:313–6. (Cited by Pan 1985b.)
Smart, DR (1992) Scombroid poisoning. A report of seven cases involving the Western Australiansalmon, Arripis truttaceus. Medical Journal of Australia 157(11−12):748−51.
Sockett, PN (1991) Food poisoning outbreaks associated with manufactured foods in England andWales. Communicable Disease Report 1(10):R105–9.
Sours, HE and Smith, DG (1980) Outbreaks of foodborne disease in the United States. Journal ofInfectious Diseases 142:122–5.
Specht, D (1998) Scombroid fish poisoning. Journal of Emergency Nursing 24(2):118−9.
Stratton, JE and Taylor, SL (1991) Chapter 13. Scombroid poisoning, pp. 331–51. In Microbiology ofmarine food products, D Ward and C Hackney, eds. Spectrum, New York.
Stratton, JE et al (1991) Biogenic amines in cheese and other fermented foods. A review. Journal ofFood Protection 54:460−70.
Stryer, L (1981) In Biochemistry, Chapter 21. Biosynthesis of amino acids and heme, p. 504. WHFreeman and Company, San Francisco.
Subburaj, M et al (1984) Incidence of histidine decarboxylating bacteria in fish and market environs.Food Microbiology 1:263–7.
Sutherland, SK (1983) Australian animal toxins. Oxford University Press, Oxford, Auckland, NewYork. (Source of Kimata reference.)
Suyama, M and Yoshizawa, Y (1973) Free amino acid composition of the skeletal muscle of migratoryfish. Bulletin of the Japanese Society of Scientific Fisheries 39:1339–43. (Cited by Pan 1985b.)
Takagi, M et al (1969) On the formation of histamine during loss of freshness and putrefaction ofvarious marine products. Hokkaido Daigaku Suisan Gakubu Kenkyu Iho 20:227–34. (Cited by Arnoldand Brown 1978; Pan 1985b.)
Taylor, SL (1983) Monograph on histamine poisoning. Codex Alimentarius Commission, FAO/WHO,Rome. 19th Session of the Codex Committee on Food Hygiene, Washington, DC, 26–30 September1983. (Cited by Bremer 1998.)
Taylor, SL (1985) VPH/FOS/85.1. Histamine poisoning associated with fish, cheese, and other foods.World Health Organization, Geneva, Switzerland, 47 pp.
Taylor, SL (1986) Histamine food poisoning: Toxicology and clinical aspects. Critical Reviews inToxicology 17(2):91−128.
Taylor, SL (1988) Marine toxins of microbial origin. Food Technology 42(3):94–8.
Taylor, SL and Lieber, ER (1979) In vivo inhibition of rat intestinal histamine-metabolizing enzymes.Food and Cosmetic Toxicology 17:237–40.

Histamine Fish Poisoning
78
Taylor, SL and Speckhard, MW (1983) Isolation of histamine-producing bacteria from frozen tuna.Marine Fish Review 45(4–6):35–9.
Taylor, SL and Sumner, SS (1986) Determination of histamine, putrescine, and cadaverine, pp. 235–45.In Seafood quality determination, DE Kramer and J Liston, eds. Elsevier, Amsterdam.
Taylor, SL and Woychik, NA (1982) Simple medium for assessing quantitative production of histamineby Enterobacteriaceae. Journal of Food Protection 45(8):747–51.
Taylor, SL et al (1978a) Histamine production by food-borne bacterial species. Journal of Food Safety1:173–87.
Taylor, SL et al (1978b) A simplified method for histamine analysis of foods. Journal of Food Science43:247–50.
Taylor, SL et al (1979) Histamine production by Klebsiella pneumoniae and an incident of scombroidfish poisoning. Applied Environmental Microbiology 37(2):274−8.
Taylor, SL et al (1984) Toxicology of scombroid poisoning, pp. 417–30. In Seafood toxins, EP Ragelis,ed. American Chemical Society, Washington, DC. (Cited by Mitchell 1993.)
Taylor, SL et al (1989) Histamine poisoning (scombroid fish poisoning): An allergy-like intoxication.Journal of Toxicology, Clinical Toxicology 27(4−5):225−40.
Til, HP et al (1997) Acute and subacute toxicity of tyramine, spermidine, spermine, putrescine andcadaverine in rats. Food and Chemical Toxicology 35(3–4):337–48.
Trenerry, VC et al (1998) Method optimisation: Determination of histamine in fish by capillary zoneelectrophoresis. Journal of Capillary Electrophoresis 5(1–2):27–32.
Trinidad, LM et al (1986) Production and storage of smoked Spanish mackerel (Scomberomoruscommerson), pp. 473–5. In The First Asian Fisheries Forum, JL McLean et al, eds. Asian FisheriesSociety, Manila, Philippines.
Todd, EC (1997) Seafood-associated diseases and control in Canada. Rev. Sci. Tech. Off. Int. Epiz.16(2):661−72.
Todd, EC et al (1992) Scombroid poisoning an outbreak in two Ontario communities. CanadianCommunicable Disease Report 18(3):17−9.
Uragoda, CG and Kottegoda, SR (1977) Adverse reactions to isoniazid on ingestion of fish with highhistamine content. Tubercle 58:83–9. (Cited by Taylor 1985.)
Van Gelderen, CEM et al (1992) The effects of histamine administered in fish samples to healthyvolunteers. Clinical Toxicology 30:585–96. (Cited by Fletcher et al 1998.)
Van Spreekens, K (1987) Histamine production by the psychrophilic flora, pp. 309-18. In Seafoodquality determination, DE Kramer and J Liston, eds. Elsvier Science Publishers, Amsterdam.
Voigt, MN and Eitenmiller, RR (1977). Production of tyrosine and histidine decarboxylase in dairy-related bacteria. Journal of Food Protection 40:241–5. (Cited by Taylor et al 1978.)
Wagener, WWD (1984) Biogenic amines in fresh and spoiled fish: High performance liquidchromatography of the dansyl derivatives, pp. 48–50. 38th Annual Report, Fishing Industry ResearchInstitute, University of Capetown, Rondebosch, South Africa.
Warne, D (1985) VIII.3 Application of hazard analysis critical control point concept, pp.45–6. InHistamine formation in marine products: Production by bacteria, measurement and prediction of

Histamine Fish Poisoning
79
formation. FAO Fisheries Technical Paper No. 252. Food and Agriculture Organization of the UnitedNations, Rome.
Warne, D et al (1987) Histamine content survey in canned tuna. A quick and simple screening methodmay be called for in response to increasing international scrutiny of histamine levels inn canned fish.Infofish Marketing Digest 2:40.
Weiss, S et al (1932) The systemic effects of histamine in man. Archives of Internal Medicine 49:360.(Cited by Taylor 1986.)
Wendakoon, CN Sakaguchi, M (1992) Effects of spices on growth and biogenic amine formation bybacteria in fish muscle, pp. 305–13. In Quality assurance in the fish industry, HH Huss et al, eds.Elsevier, Amsterdam. (Cited by Rawles et al 1996.)
White, A et al, eds. (1973) Chapter 5. Amino acid metabolism, p. 697. In Principles of biochemistry,5th ed. McGraw Hill, Inc.
Wille, JJ and Kydonieus, A (1995) Abrogation of contact hypersensitivity in mice by topically appliedmast cell degranulating agents. Journal of Investigative Dermatology 104:679 (abstract). (Cited by Hartet al 1997.)
Wille, JJ et al (1999) Cis-urocanic acid induces mast cell degranulation and release of preformed TNF-alpha: A possible mechanism linking UVB and cis-urocanic acid to immunosupression of contacthypersensitivity. Skin Pharmacology and Applied Skin Physiology 12(1–2):18–27.
Wu, ML et al (1997) Scombroid fish poisoning: An overlooked marine food poisoning. Veterinary andHuman Toxicology 39(4):236−41.
Yamanaka, H (1984) Changes in histamine contents in red meat fish during storage at differenttemperatures. Bulletin of the Japanese Society of Scientific Fisheries 50:695–701. (Cited by Olley andMcMeekin 1985.)
Yamani, MI et al (1981) Histamine poisoning by Proteus spp. from tuna fish. Zbl. Bakt. Hyg., I Abt.Orig. B 173:478–87. (Cited by Taylor 1986.)
Yen, G and Hsieh, C (1991) Simultaneous analysis of biogenic amines in canned fish by HPLC.Journal of Food Science 56(1):158–60. (Cited by Price 1999.)
Yoshinaga, DH and Frank, HA (1982) Histamine-producing bacteria in decomposing skipjack tuna(Katsuwonas pelamis). Applied Environmental Microbiology 44(2):447–52.
Zee, JA et al (1981) Effect of Lactobacillus brevis, Saccharomyces uvarum and grist composition onamine formation in beers. Canadian Institute of Food Science and Technology 14(4):321–5.

Histamine Fish Poisoning
80
Appendix 1: Acknowledgments
Queensland Department of Primary Industries/National Seafood Centre(Brisbane)
Stephen ThrowerSue Poole
Australia and New Zealand Food Authority
Peter AbbottJanis BainesScott Crerar
Australian Government Analytical Laboratories (Melbourne)
Philip Marshall
Agriculture, Fisheries and Forestry Australia
Albert CatonTerry NichollsMike Nunn
Port Lincoln Tuna Processors Pty Ltd
Lindsay GuillotLea Traeger
University of Tasmania
Christian GarlandJohn BowmanTom McMeekinTom Ross
Elisa Systems
Rob Sherlock