2 athyonidium chilensis (semper, 1868) (holothuroidea ... · 2 28 abstract 29 30 athyonidium...
TRANSCRIPT

1
Embryonic development, larval morphology and juvenile growth of the sea cucumber 1
Athyonidium chilensis (Semper, 1868) (Holothuroidea: Dendrochirotida) 2
3
4
CHITA GUISADO1 *, ROBERTO MALTRAIN 1, SERGIO A. CARRASCO1 2, DANIELA DÍAZ-5
GUISADO2 3, HERMAN ROJAS 1, and EDUARDO BUSTOS 3 6
7
8
1Facultad de Ciencias del Mar y de Recursos Naturales, Universidad de Valparaíso, Casilla 5080, 9
Viña del Mar, Chile 10
11
2Present address: School of Biological Sciences and Coastal Ecology Laboratory, Victoria 12
University of Wellington, Wellington 6140, New Zealand 13
14
3Instituto de Ciencia y Tecnología, Universidad Arturo Prat, Ejército 443, Puerto Montt, Chile 15
16
*Corresponding author: Facultad de Ciencias del Mar y de Recursos Naturales, Casilla 5080, 17
Reñaca, Viña del Mar, Chile. Phone: (56) (032) 2507862. Fax: (56) (032) 2507859 18
Email: [email protected] 19
20
21
Running title: Development and growth of Athyonidium chilensis 22
23
24
25
26
27

2
Abstract 28
29
Athyonidium chilensis is the most economically important holothurian in Chile. In spite of the 30
increases in harvest due the high demand from Asian markets, early life stages and juvenile phases 31
of the species have received less attention. The embryonic and larval development of A. chilensis 32
took approximately 5 days and it followed the typical developmental stages described for other sea 33
cucumber species with direct development: the gastrula develops into a vitellaria larva 34
(lecothotrophic stage; 4-5 d pf), which was followed by the pentactula stage (7 d pf). After 35
settlement, individuals were capable of active feeding on micro-algae associated with the sediment. 36
The juveniles (35 d pf) possessed four pairs of ambulacral podia, and after 4 months of cultivation, 37
a substantial number of podia covered their bodies, which were approximately 1.2 mm long. An 38
evident effect of the density during the culture of juvenile was observed, with higher growth rates 39
and survival observed in the lowest density treatment (1 ind/cm2). This study provide the first 40
description of the early life history stages of a sea cucumber in Chilean waters, but also shows that 41
the aquaculture of this species is feasible and could be developed as an alternative to maintain a 42
sustainable harvest and eventually contribute to the restoration of natural populations. 43
44
45
Key words: Athyonidium chilensis, holothuroid, development, juvenile growth, aquaculture 46
47
48
49
50
51
52
53

3
Introduction 54
55
Sea cucumbers are a valuable resource for coastal communities in the Indo-Pacific region, where 56
they have been exploited unsustainably during decades. The fishing pressure exerted on the species 57
has increased over the years fuelled by the rising wholesale price of the product, resulting in the 58
depletion of wild stocks worldwide (Purcell, et al. 2002; Hamel, et al. 2003; Ramofafia, et al. 2003). 59
The recent use of sea cucumbers in the pharmaceutical industry as antibacterial agents have also 60
increased the demand of the species (see Al-Haj, et al. 2009), encouraging an improvement of the 61
artificial culture techniques together with the acquisition of an adequate knowledge of critical 62
phases in captivity. However, in the last decades, only a few commercial holothurians have been 63
successfully reared to settlement, including some tropical and temperate species (e.g., Holothuria 64
scabra, H. fuscogilva, Stichopus japonicus, S. mollis) (reviewed by Ramofafia, et al. 2003). 65
66
Even though the embryonic and larval development of some species of sea cucumbers has been 67
well documented (e.g., Cameron & Fankboner, 1989; Hamel & Mercier, 1996; Battaglene, et al. 68
2002; Hamel, et al. 2003; Laxminarayana, 2005; Asha & Muthiah, 2002; Asha & Muthiah, 2005), 69
most of the species that have been studied belong to the order Asparochidotida, characterized by an 70
indirect development, i.e., the gastrula develops to an auricularia larva (planktotrophic stage), that 71
undergoes metamorphosis to a dololaria larva (lecothotrophic stage) before settlement as a 72
pentactula (Sewell & McEuen, 2002). However, direct development, i.e., gastrula develops to a 73
vitellaria larva (lecothotrophic stage) before settlement (e.g., with no auricularia stage), is the most 74
common form of development in holoturoids and is dominant in 22 of the 25 families (Smiley, et al. 75
1991). 76
77
Athyonidium chilensis is one of the most common species of echinoderms along the Chilean coast, 78
with a geographic distribution that extends from Ancón, Perú (11º 44’ 5” S), to Punta Gaviota in 79

4
Chiloé Island, Chile (42º 03’ 55” S). This species inhabits intertidal and subtidal zones, where can 80
be found in in rocky pools and crevices, below boulders and also buried in the sand (Deichmann, 81
1941; Pawson, 1964; Pawson, 1969, Fernández, 1998). Although A. chilensis is the most 82
commercially important holothurian in Chile, there is no documented information before 1991 83
(SERNAP, 2003). The average harvest of the species from 1998 to 2008 was 269 ton per year, with 84
a maximum extraction of 1,510 ton in 2000 (Renbo & Yuan, 2004; SERNAP, 2008). The 85
unsustainable exploitation of this resource, added to the growing demand from Asian markets, 86
forebodes a drastic decline of the easily-fished wild stocks of A. chilensis in a near future. 87
Therefore, studies focused on reproduction, development and feeding behavior of the species in 88
captivity are extremely important in order to understand the life history of the species, determine its 89
possible aquaculture potential and also take accurate decisions in terms of stock management. 90
91
Despite previous studies have contributed on some aspects of the biology, ecology and culture of A. 92
chilensis (Fernández, 1998; Guisado, et al. 1999; Moreno, 2002; Pérez, 2005; González, 2006; 93
Fernández, 2007; Maltrain, 2007), the early life history of the species has not been completely 94
described and there is a knowledge gap about larval morphology, development, settlement and 95
juvenile growth. In this study we aimed first, to characterize the embryonic and larval development 96
of the sea cucumber A. chilensis under laboratory conditions and second, to evaluate the effects of 97
cultivation density on juvenile growth and survival. 98
99
100
Materials and Methods 101
102
Specimen collection and spawning induction 103
104

5
In May 2006, twenty-five adults of A. chilensis were collected at Maitencillo Beach, Chile (32° 38’ 105
S, 71° 26’ W) by SCUBA diving. Individuals were transported to the wet laboratory of Universidad 106
de Valparaíso, where they were placed in five 67-L tanks filled with micro-filtered seawater (5 107
individuals per tank) and kept in a conditioning period of 48 h. Eighteen sea cucumbers with a body 108
length greater than 15 cm (250-400 g) were further selected for spawning induction, placed into 109
three 67-L tanks filled with micro-filtered seawater (6 individuals per tank) and induced to spawn 110
by means of thermal stimulation (Girapsy & Ivy, 2005; Laxminarayana, 2005) and addition of food 111
ad libitum using the micro-alga Chlorella neustonica (Pérez, 2005). 112
113
Embryo and larval culture 114
115
After spawning, female gametes were deposited in 3-L glass tanks and male gametes were 116
transferred to a 250 ml beaker. In order to carry out fertilization, 2 ml of spermatozoa were added to 117
each 3-L tanks, obtaining a gamete ratio of 1:100 (oocyte - spermatozoa). 118
119
Once fertilization was verified, the eggs were transferred into five 67-L tanks (3 eggs ml-1) and each 120
tank was filled with micro-filtered and UV sterilized seawater. Fifty percent of the seawater was 121
renewed on the second day of cultivation. After 3 or 4 days post-fertilization (pf), only larvae 122
swimming in the water column were selected and then transferred into a 1000-L specially 123
conditioned tank (0.5 larva ml-1). The 50 % of the seawater was renewed daily and the temperature 124
was maintained constant (13 ± 0.3ºC). Two air stones positioned at the bottom of the tank provided 125
continuous aeration. Embryonic and larval development was documented by taking random samples 126
(30 larvae) every 15-30 minutes during the first 24 h pf. Hereafter, samples were taken every hour 127
until the early vitellaria stage was observed (4-5 days). Samples were photographed and measured 128
using an optic microscope (4X magnification) equipped with an ocular micrometer and also where 129
analyzed using a scanning electron microscope. After 10 days of cultivation (i.e., when feeding 130

6
behavior was observed), the larvae were fed daily with the micro-alga Tetraselmis suecica 131
(MLB1094; 1,200 cells ml-1), and after the day 42, 0.5 g of Algamac 2000 TM were also provided 132
once a week. Once settlement was observed, 145 g of fine sand (> 0.5 mm) were added to the tank 133
in order to increase survival (C. Guisado, unpublished data 2006). Size was daily determined by 134
taking measures of the maximum length (ML in mm, defined as the distance between the mouth and 135
the anus) (Hamel & Mercier, 1996; Hamel, et al. 1999) of 30 individuals during the first 8 months 136
of cultivation. Survival was monthly evaluated using 100 cm2 PVC quadrants with six replicates in 137
each measurement. 138
139
Effects of cultivation density on juvenile growth and survival of Athyonidium chilensis 140
141
Three experimental densities were tested in this study. Juveniles obtained in laboratory (75 days pf; 142
0.9 ± 0.1 mm) were placed in 300-ml glass tanks at densities of 1, 3 and 5 individuals per cm2 (i.e., 143
64, 191 and 318 individuals per tank, respectively), with each density being conducted in triplicate. 144
Fine sand was used as substrate based on preliminary experiments (3 g per tank; C. Guisado, 145
unpublished data 2006). Seawater was renewed daily and the temperature was maintained constant 146
(13 ± 0.3 °C). Juveniles were fed daily with the micro-alga T. suecica (1,200 cells/ml) and the 147
supplement Algamac 2000 TM was added once a week. At the beginning and at the end of the 148
experimental period, the ML of 20 randomly selected juveniles per treatment was measured and the 149
absolute growth rate (AGR) was calculated as follow, AGR = MLt-ML0 t-1, where: MLt= final 150
maximum length (μm), ML0 = initial maximum length (μm) and t = experimental period (days). In 151
order to evaluate survival, the total number of individuals in each tank was visually quantified twice 152
a month. 153
154
One- way analyses of variance (ANOVA) were used to test for differences in AGR among the 155
experimental densities examined. Differences in survival of juvenile over time and among different 156

7
experimental densities were tested using a two-way ANOVA. In both cases, significant differences 157
were further examined using multiple pairwise comparison tests (Tukey). Percentage variables were 158
arcsine transformed. Normality and homocedasticity requirements were tested using Lilliefors and 159
Levene tests, respectively. When assumptions were not met, the analyses were performed in rank-160
transformed data (Conover1980). 161
162
163
Results 164
165
Characterization of developmental stages of Athyonidium chilensis 166
167
Spawning and embryonic development 168
169
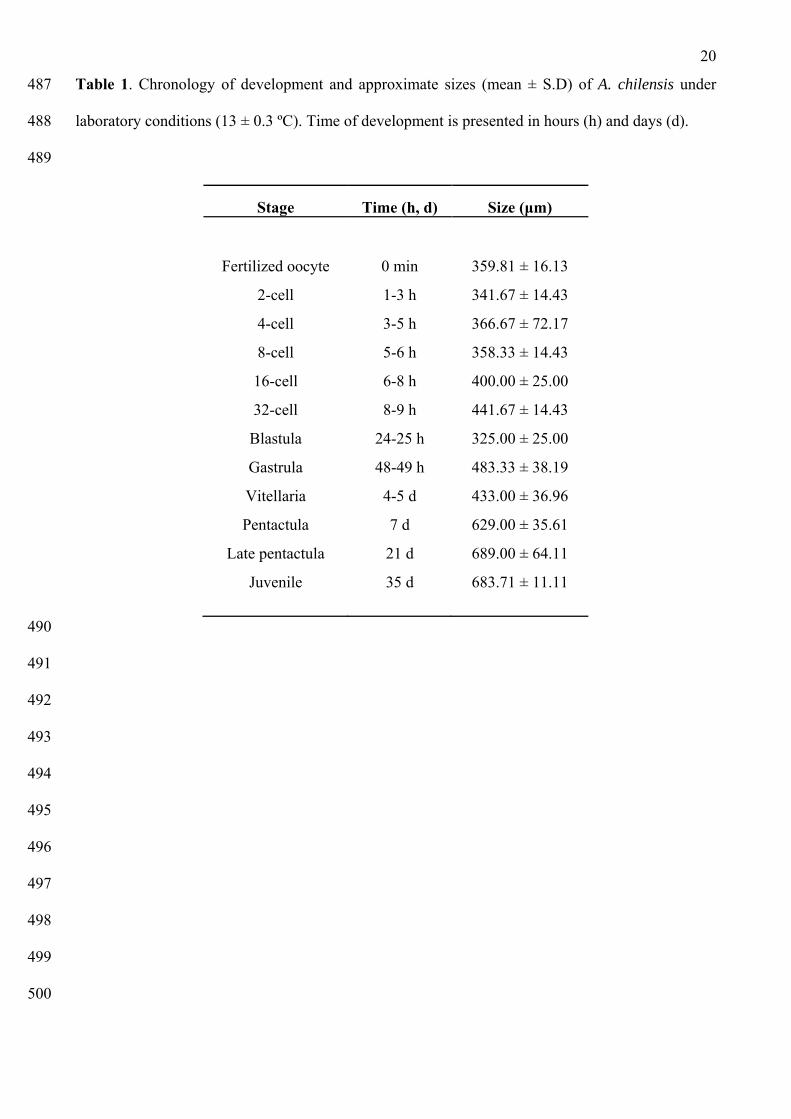
The chronology of development for A. chilensis, with the corresponding size of the embryos and the 170
photographic illustrations of the stages of development, are presented in Table 1 and Figures 1 and 171
2. After spawning induction, only 7 of the 18 individuals spawned (6 males and 1 female). Once 172
fertilization was completed, the estimated quantity of viable eggs was ~ 6 x 105. The eggs were 173
spherical and could be easily seen (359.81 ± 16.13 μm [mean diameter ± S.D]). The fertilization 174
envelope (fe) was observed after 10 min post-fertilization (pf) (Fig. 1A). 175
176
The first cleavage occurred between 1 to 3 h pf. The second (Fig. 1B), third and fourth cleavages 177
took place between 3 to 6 h pf. During these stages blastomeres (b) of equal size and spherical 178
shape were observed, with a clear separation between them. The animal and vegetal poles continued 179
dividing until the 32-cell stage. At this stage the fertilization envelope was still observed (Fig 1C). 180
After 24-25 h pf, a process of embryo compaction, a considerable reduction in the space between 181
the blastomeres and the presence of the blastocele (bc) was observed, being the precursor of the 182

8
blastula stage (Fig. 1D). Rotating movements and constant migration throughout the water column 183
were observed in the embryos, and after 48 h pf the blastula began to elongate and transform into a 184
gastrula (Fig. 1E). The blastopore (bp) and blastopore scams (bs) were clearly visible in the aboral 185
region (Fig. 2A, B) and a small lateral depression at the equatorial region of the embryo’s body 186
began to form, indicating the position of the future vestibule (v) (Fig. 2C). After 4 to 5 d pf the 187
embryo began its transformation into a vitellaria larva (Fig. 1F). 188
189
Larval development 190
191
The vitellaria larva of A. chilensis (433 ± 36.96 μm [mean diameter ± S.D]) was characterized as a 192
free swimming and lecithotrophic larva. The vestibule (v) was clearly visible and the five primary 193
buccal tentacles (pt) were observed inside it (Fig. 1F). After 7 days pf these tentacles began to 194
protrude from the vestibule, process that was the precursor of the pentactula stage (Fig. 1G). The 195
pentactula was characterized by a progressive decrease in the swimming capacity due to the loss of 196
the body cilia. Also, the buccal tentacles (bt) were completely protruded, showing an important role 197
on feeding and locomotion. The anal pore was also observed in the aboral region of the body. At 21 198
days pf the late pentactula possesses, in addition to the buccal tentacles, a pair of ambulacral podia 199
(ap) (Fig. 2 D, E, F). After the settlement of the pentactula, individuals were capable of active 200
feeding on micro-algae associated with the sediment, using their buccal tentacles to collect and 201
ingest food. 202
203
Juvenile growth and survival 204
205
Thirty-five days pf the fourth pair of ambulacral podia appeared, and juvenile with green 206
pigmentation towards the posterior portion of the body were observed (Fig. 1H). During this period, 207
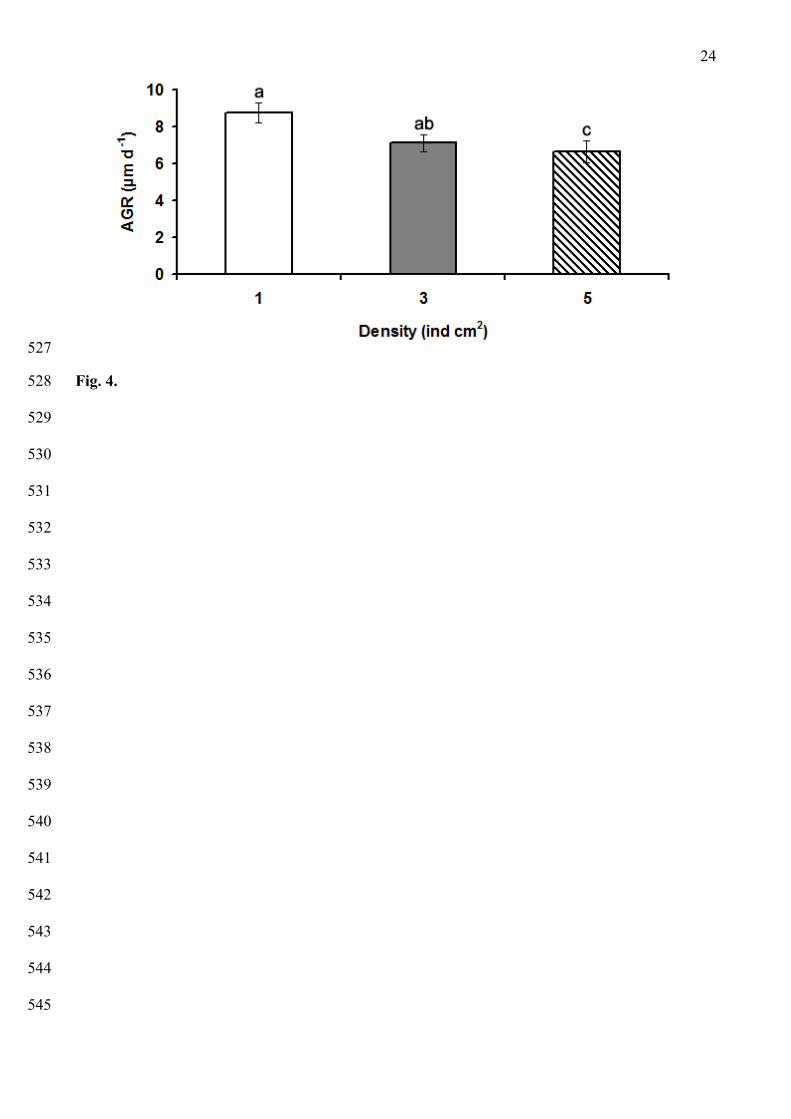
a survival of ~18 % from the total viable larvae was recorded (Fig. 3A). After 4 months, a 208

9
substantial number of completely developed podia and a notable pigmentation on the body wall 209
were observed, and juveniles (1,200 µm ML), began to show some common adult behaviors such as 210
burying and anal pumping movements. During this period, a survival of 1.6 % from the total viable 211
larvae was recorded (Fig 3A). After 8 months of cultivation the survival decreased until 0.24 %, and 212
a maximum size of 1,900 µm was recorded (Fig. 1I, 3A, 3B). 213
214
Effects of cultivation density on growth and survival of juvenile Athyonidium chilensis 215
216
After 3 months, the AGR (µm d-1) of juvenile A. chilensis varied significantly among the three 217
cultivation densities evaluated (one-way ANOVA, F2,177=4.947, P=0.008). The maximum AGR 218
was recorded in the treatment with the lowest density (8.78 ± 4.17 µm d-1 [mean ± S.D]; 1 ind cm2), 219
which was not significantly different from the 7.12 ± 3.57 µm d-1 obtained in the treatment with 3 220
ind cm2 (Tukey, P=0.052); however, differed significantly from the 6.64 ± 4.61 µm d-1 obtained 221
with 5 ind cm2 (Tukey, P=0.008) (Fig. 4). 222
223
The survival of juvenile A. chilensis decreased significantly in all density treatments over the 75 224
days of experiments until a mean value of 39.7 % ± 6.7 % [mean ± S.D] (two-way ANOVA, F5,36= 225
50.204; P < 0.001). Significant differences were detected among treatments (two-way ANOVA, 226
F2,36= 5.101; P = 0.011; Fig. 5), with no interactive effect between time and treatments (two-way 227
ANOVA, interaction time x treatment: F10,36= 1.131; P = 0.368). Overall, the survival of juveniles 228
in the treatment with 1 ind cm2 was not significantly different to the survival in the treatment with 3 229
ind cm2 (Tukey, P=0.378); however, the former differed significantly with the lower values 230
obtained in the treatment with 5 ind cm2 (Tukey, P = 0.008) (Fig. 5). 231
232
233
234

10
Discussion 235
236
Liberation of gametes in A. chilensis was only achieved after stimulation via an increase in the 237
water temperature and the addition of food, a technique previously used to induce spawning in a 238
range of tropical sea cucumbers, including H. scabra, Actinopyga mauritiana (Battaglene, 1999) 239
and H. fuscogilva (Battaglene, et al. 2002; Ramofafia, et al. 2003). Spawning occurred during the 240
night, and invariably, males spawned before the female. This same reproductive behaviour was 241
previously described by Costelloe (1988) in Aslia lefevrei and by Renbo & Yuan (2004) in 242
Apostichopus japonicus. Indeed, some authors have suggested that sperm, or something associated 243
with it, is a proximal signal for synchronizing female to spawn (see Bataglene, et al. 2002). 244
Therefore, in order to successfully carry out fertilization in laboratory is essential to maintain a total 245
control over the broodstocks, because spontaneous spawning could cause polyspermia, a common 246
problem with in vitro fertilizations and a lethal condition in marine invertebrates. 247
248
The average diameter of newly released A. chilensis eggs was 359.81 ± 16.13 μm. Larger sizes have 249
been reported by Costelloe (1988) in the dendrochirote A. lefevrei (400-650 μm) and, Hamel & 250
Mercier (1996) found that the mean diameter of the eggs in Cucumaria frondosa was more than 251
twice the size registered in our study (i.e., 900 μm). By contrast, in species that present indirect 252
development eggs are smaller, as has been recorded for H. scabra (156.4 μm), H. fuscogilva (151.7 253
μm) and Actinopyga mauritiana (109.9 μm) by Ramofafia et al. (2003). These differences in egg 254
size between species could be due to their different reproductive strategies, as species that present 255
direct development only presents a lecitotrophic stage, in which survival and growth rely on 256
endogenous reserves. 257
258
Athionidium chilensis, as other dendrochirotes species (e.g., C. frondosa; Hamel & Mercier 1996) 259
presents direct development, during which the gastrula develops into a lecitotrophic larva (i.e., 260
vitellaria). The mean size recorded in this study for the vitellaria larvae was 433 ± 36.96 μm (mean 261

11
± S.D), while Hamel & Mercier (1996) recorded in C. frondosa a much larger size (1.55 ± 0.3 mm). 262
Apparently, the different larval stages of A. chilensis do not exhibit a significant increase in the 263
maximum length during the larval development, a condition previously observed in C. frondosa as 264
well (Hamel & Mercier, 1996). 265
266
According to several studies, the embryonic and larval development of dendrochirotes is considered 267
brief in comparison with species that present indirect development. For example, in C. frondosa the 268
pentactula stage was recorded between 11-11.5 days at 12 °C (Hamel & Mercier, 1996), in A. 269
chilensis between 3-4 days at 14-16 ºC (Pérez 2005) and according to this study between 6-7 days 270
at 13 ± 0.3 ºC. By contrast, in aspidochirotes the final larval stage is reached later, for example, in 271
A. japonicus it was recorded between 13-17 days at 21.5-23 ºC (Rembo & Yuan, 2004), in 272
Isostichopus fuscus between 22-27 days at 27-29 ºC (Hamel, et al. 2003) and in H. scabra between 273
13-15 days at 25-27 ºC (Ramofafia, et al. 2003). These results suggest that environmental factors 274
such as temperature may play an important role during the developmental stages of several 275
holothurian species and can be especially important as a technique to manage the extension of the 276
larval stage in laboratory reared species (Hamel & Mercier, 1996; Ramofafia et al. 2003). 277
278
Juvenile A. chilensis obtained after 35 days pf were capable of actively use their crown of tentacles 279
for feeding, and their four pairs of ambulacral podia for locomotion and attachment to the substrate. 280
During the first 8 months of cultivation in laboratory, juvenile A. chilensis reached an average 281
length of 1,900 µm, contrasting with the grater sizes recorded by Hamel & Mercier (1996) and 282
Costelloe (1988) during the first year of cultivation in the dendrochirotes C. frondosa and A. 283
lefevrei (10 mm and 3.4 cm, respectively). However, the slow growth rates observed in A. chilensis 284
were more notable when compared with aspidochirotes species. For example, I. fuscus reached a 285
length of 3.5 cm after only 72 days pf (Hamel, et al. 2003), and a similar size was recorded in H. 286
spinifera after 4 months of cultivation (4.8 cm; Asha & Muthiah, 2005). These differences in 287

12
juvenile growth between the orders Aspidochirotida and Dendrochirotida could be due to their 288
completely different biological and ecological characteristics, since aspidochirotes move actively 289
and experience daily cycles of burying, while dendrochirotes are usually buried or covered by 290
substrate (e.g., sand or algal detritus) displaying less mobility and a passive foraging behavior 291
(Giese, et al. 1991). 292
293
As in others sea cucumber species obtained in land-based nursery systems, A. chilensis showed low 294
survival during the first 30 days of cultivation. High mortalities were also recorded during the first 295
20 days post-metamorphosis in C. frondosa by Hamel et al. (2003) and during hatch in H. scabra 296
(Ramofafia, et al. 2003). Few procedures have been proposed in order to avoid high mortalities in 297
early stages of holothurians during cultivation, but one of the most important is the maintenance of 298
an appropriate density of individuals during culture. Past experience has shown that a high density 299
of individuals increases mortality and reduces the availability of space and food, causing 300
malnutrition, slow growth and high variability of size (Xiyin, et al. 2004; Agudo, 2006). Our results 301
suggested that at low density (1 ind/cm2) juveniles grew faster and survive better than conspecifics 302
maintained at high densities (5 ind/cm2), agreeing with the findings of Battaglene et al. (1999) and 303
Xilin (2004), whose observed that the highest growth and survival occurred in the low density 304
treatments (i.e., 1.7 ind/cm2 and 1 ind/cm2 for H. scabra and A. japonicus, respectively). 305
306
In summary, Athyonidium chilensis as well as other holothurian species, undergoes critical phases 307
in captivity, such as fertilization, larval development and settlement. Nevertheless, this species 308
showed a rapid embryonic and larval development. Juvenile growth and survival was affected by 309
the cultivation density, factor that could indirectly mediate the food availability on cultured 310
conditions. This study provide the first data on embryonic development, larval morphology and 311
juvenile growth of A. chilensis, showing that this sea cucumber can be successfully reared in land-312
based nursery systems and that the culture of this species is feasible and can potentially be 313

13
developed as an alternative to maintain sustainable harvest and eventually contribute to the 314
restoration of natural populations. Further research should target to improve the spawning induction 315
and principally, the feeding and growth of juveniles in captivity. 316
317
318
Acknowledgements 319
320
We would like to thank L. Amaro and M. Torres for their help during field collections and 321
laboratory experiments. The comments of anonymous reviewers that improved initial versions of 322
the manuscript are also appreciated. This research was supported by funds of the project FONDEF 323
from CONICYT: DO3I 1072. 324
325
326
References 327
328
Agudo, N. 2006. Sandfish hatchery techniques. Australian Centre for International Agricultural 329
Research (ACIAR), the Secretariat of the Pacific Community (SPC) and the WorldFish Center. 43 330
pp. 331
332
Al-Haj, N. A., Norfarrah, M. A., Shamsudin, M. N., Yusoff, F. M. & Arshad, A. 2009. Novel 333
antibacterial activity of peptide gene extracted from a Malaysian sea cucumber. Res. J. Biol. Sci., 4, 334
482-486. 335
336
Asha, P. S. & Muthiah, P. 2002. Spawning and larval rearing of sea cucumber Holothuria 337
(Theelhothuria) spinifera Theel. SPC Beche-de-mer Information Bulletin, 16, 11-15. 338
339

14
Asha, P. S. & Muthiah, P. 2005. Effects of temperature, salinity and pH on larval growth, survival 340
and development of the sea cucumber Holothuria spinifera Theel. Aquaculture, 250, 823-829. 341
342
Battaglene, S. C., Seymour, E. J. & Ramofafia, C. 1999. Survival and growth of cultured juvenile 343
sea cucumber, Holothuria scabra. Aquaculture, 178, 293-322. 344
345
Battaglene, S. C., Seymour, E. J., Ramofafia, C. & Lane I. 2002. Spawning induction of three 346
tropical sea cucumbers, Holothuria scabra, H. fuscogilva and Actinopyga mauritiana. Aquaculture, 347
207, 29-47. 348
349
Cameron, J. L. & Fankboner, P. V.1989. Reproductive biology of the commercial sea cucumber 350
Parastichopus californicus (Stimpson) (Echinodermata: Holothuroidea). II. Observations on the 351
ecology of development, recruitment and juvenile life stage. J. Exp. Mar. Biol. Ecol., 127, 43-67. 352
353
Conover, W. J. 1980. Practical nonparametric statistics. John Wiley & Sons Inc., New York. 354
355
Costelloe, J. 1988. Reproductive cycle, development and recruitment of two geographically 356
separated populations of the dendrochirote holothurian Aslia lefevrei. Mar. Biol., 99, 535-545. 357
358
Deichmann, E. 1941. The Holothuroidea Collected by the “Velero III” During the years 1932 to 359
1938. Part I, Dendrochirota. Allan Hankock Pacif. Exped., 8, 1-164. 360
361
Fernández, M. 1998. Ecología de Athyonidium chilensis (Semper, 1868) (Echinodermata: 362
Holothuroidea), en tres hábitats de la región de Coquimbo. Tesis para optar al título de Biólogo 363
Marino, Universidad Católica del Norte, pp 73. 364
365

15
Fernandez, V. 2007. Efecto de la proporción de proteína vegetal y animal en la dieta de ejemplares 366
adultos de Athyonidium chilensis (Semper, 1868) (Echinodermata: Holothuridae), sobre el 367
crecimiento y la distribución de los nutrientes alimentarios. Tesis para optar al título de Biólogo 368
Marino, Universidad de Valparaíso, pp 46. 369
370
González, E. 2006. Efecto de dietas naturales sobre el crecimiento y sustratos energéticos de los 371
principales componentes corporales del pepino de mar Athyonidium chilensis. Tesis para optar al 372
título de Biólogo Marino, Universidad Católica del Norte, pp 31. 373
374
Giraspy, D. & Ivy, G. 2005. Australia’s first commercial sea cucumber culture and sea ranching 375
project in Hervey Bay, Queensland, Australia. SPC Beche-de-mer Information Bulletin, 21, 29-31. 376
377
Giese, A., Pearse, J. & Pearse, V. 1991. Reproduction of marine invertebrates. Volume VI. 378
Echinoderms and Lophophorates. The boxwood press Pacific Grove, California, pp 808. 379
380
Guisado, C., Saavedra, J. & Hernández, A. 1999. Inducción al desove y desarrollo larvario de 381
Athyonidium chilensis (Semper, 1868). In: Impresos Universitaria (ed.). Libro de resúmenes XIX 382
Congreso de Ciencias del Mar. Chile. pp 120-121. 383
384
Hamel, J. F. & Mercier, A. 1996. Early development, settlement, growth and spatial distribution of 385
the sea cucumber Cucumaria frondosa (Echinodermata: Holothuroidea). Can. J. Fish. Aquat. Sci., 386
53, 253-271. 387
388
Hamel, J. F., Hycaza-Hidalgo, R. & Mercier, A. 2003. Larval development and juvenile growth of 389
the Galápagos sea cucumber Isostichopus fuscus. SPC Beche-de-mer Information Bulletin, 18, 3-8. 390

16
Laxminarayana, A. 2005. Induced spawning and larval rearing of the sea cucumbers, Bohadschia 391
marmorata and Holothuria atra in Mauritius. SPC Beche-de-mer Information Bulletin, 22, 48-52. 392
393
Maltrain, R. 2007. Crecimiento de Athyonidium chilensis (Semper, 1868) (Echinodermata: 394
Holothuroidea) en cautiverio y policultivo con Eurhomalea lenticularis (Sowerby, 1835). Tesis para 395
optar al título de Biólogo Marino, Universidad Católica del Norte, pp 49. 396
397
Mashanov, V. S. & Dalmatov, I. Y. 2000. Developmental morphology of a holothurian, Cucumaria 398
japonica (Dendrochirota: Holothuroidea), a species with accelerated metamorphosis. Invertebr. 399
Repr. Dev., 37, 137-146. 400
401
Moreno, C. 2002. Aspectos reproductivos de Athyonidium chilensis (Semper, 1868) 402
(Echinodermata: Holothuroidea) en playa el Francés, IV Región, Chile. Tesis para optar al título de 403
Biólogo Marino, Universidad Católica del Norte, pp 38. 404
405
Pawson, D. 1964. The holothuroidea collected by The Royal Society Expedition to Southern Chile, 406
1958–1959. Pac. Sci., 18, 453-470. 407
408
Pawson, D. 1969. Holothuroidea from Chile. Report n° 46 of The Lund University. Chile 409
Expedition . 1948-1949. Sarsia, 38, 121-146. 410
411

17
Pérez, L. 2005. Desarrollo embrionario y larval del pepino de mar Athyonidium chilensis (Semper, 412
1868) (Echinodermata: Holothuroidea). Tesis para optar al titulo de Biólogo Marino. Universidad 413
Católica del Norte, pp 35. 414
415
Purcell, S., Gardner, D. & Bell, J. 2002. Developing optimal strategies for restocking sandfish: a 416
collaborative project in New Caledonia. SPC Beche-de-mer Information Bulletin, 16, 2-4. 417
418
Ramofafia, C., Byrne, M. & Battaglene, C. 2003. Development of three commercial sea cucumbers, 419
Holothuria scabra, H. fuscogilva and Actinopyga mauritiana: larval structure and growth. Mar. 420
Fresh. Res., 54, 657-667. 421
422
Renbo, W. & Yuan, C. 2004. Breeding and culture of the sea cucumber Apostichopus japonicus, 423
Liao. In: Lovattelli, A. (ed.). Advances in sea cucumber aquaculture and management. Session III. 424
Acuaculture advances. FAO Fisheries department. Rome, Italy, pp. 277-286. 425
426
Sewell, M.A. & McEuen, F.S. 2002. Phylum Echinodermata: Holothuroidea. In: C.M. Young, M.A. 427
Sewell and M.E. Rice (eds.). Atlas of marine invertebrate larvae. Academic Press, pp 513-530. 428
429
SERNAP. 2003. Anuario estadístico de pesca. Ministerio de Economía Fomento y Reconstrucción, 430
Chile, pp 156. 431
432
SERNAP. 2008. Anuario estadístico de pesca. Ministerio de Economía Fomento y Reconstrucción, 433
Chile, pp 156. 434
435

18
Smiley, S., McEuen, F.S., Chafee, C. & Krishnan, S. 1991. Echinodermata:Holothuroidea. In: A.C. 436
Giese, J.S. Pearse & V.B. Pearse (eds.). Reproduction of marine invertebrates. Vol. VI, 437
Echinoderms and Lophophorates. Boxwood Press, Pacific Grove, CA, pp. 663-750. 438
439
Xilin, S. 2004. The progress and prospects of studies on artificial propagation and culture of the sea 440
cucumber, Apostichopus japonicus. In: A. Lovatelli, C. Conand, S. Purcell, S. Uthicke, J. F. Hamel 441
& A. Mercier (eds.). Advances in sea cucumber aquaculture and management. FAO Fisheries 442
Technical Paper n°. 463. Rome, Italy, pp. 273-276. 443
444
445
446
447
448
449
450
451
452
453
454
455
456
457
458
459
460
461

19
Figure captions 462
463
Fig.1. Optic microscopy of the embryonic and larval development of A. chilensis. (A) Fertilized 464
oocyte with fertilization envelope (fe) completely elevated. Scale bar = 120 μm. (B) 4-cell stage, 465
where (b) blastomere. Scale bar = 180 μm. (C) 32-cell stage. Scale bar = 150 μm (D) Blastula, 466
where (bc) blastocoele. Scale bar = 100 μm (E) Gastrula, where (v) vestibule. Scale bar = 100 μm 467
(F) Vitellaria, where (pt) primary tentacles. Scale bar = 150 μm (G) Pentactula, where (bt) buccal 468
tentacles. Scale bar = 200 μm. (H) Juvenile 35 d pf, where (a) anus, (bt) buccal tentacles, (ap) 469
ambulacral podia. Scale bar = 230 μm. (I) Juvenile 120 d pf. Scale bar = 275 μm. 470
471
Fig.2. Electronic microscopy of the larval development of A. chilensis. (A) Gastrula with blastopore 472
(bp) in the aboral region. (B) Blastopore scams (bs). (C) Gastrula with the future vestibule (v) in 473
formation. (D and E) Late pentactula, where (bt) buccal tentacles, (ap) ambulacral podia. (F) 474
Buccal tentacles in detail. Scale bar (A, C, D, E) = 100 μm. Scale bar (B) = 10μm. 475
476
Fig. 3. (A) Survival and (B) Size of A. chilensis during the first eight months of cultivation. 477
478
Fig.4. Absolute growth rate (AGR) of juvenile A. chilensis reared in laboratory under three 479
experimental densities (1, 3 and 5 individuals per cm2). Letters indicate significant differences in 480
growth rate. Vertical bars represent S.Es. 481
482
Fig.5. Survival of juvenile A. chilensis reared in laboratory under three experimental densities (1, 3 483
and 5 individuals per cm2). Letters and asterisks indicate significant differences in growth rate. 484
Vertical bars represent S.Es. 485
486
20
Table 1. Chronology of development and approximate sizes (mean ± S.D) of A. chilensis under 487
laboratory conditions (13 ± 0.3 ºC). Time of development is presented in hours (h) and days (d). 488
489
Stage Time (h, d) Size (μm)
Fertilized oocyte 0 min 359.81 ± 16.13
2-cell 1-3 h 341.67 ± 14.43
4-cell 3-5 h 366.67 ± 72.17
8-cell 5-6 h 358.33 ± 14.43
16-cell 6-8 h 400.00 ± 25.00
32-cell 8-9 h 441.67 ± 14.43
Blastula 24-25 h 325.00 ± 25.00
Gastrula 48-49 h 483.33 ± 38.19
Vitellaria 4-5 d 433.00 ± 36.96
Pentactula 7 d 629.00 ± 35.61
Late pentactula 21 d 689.00 ± 64.11
Juvenile 35 d 683.71 ± 11.11
490
491
492
493
494
495
496
497
498
499
500

21
501
502
Fig.1. 503
504
505
506
507

22
508
509
Fig.2. 510
511
512
513

23
514
Fig. 3. 515
516
517
518
519
520
521
522
523
524
525
526
24
527
Fig. 4. 528
529
530
531
532
533
534
535
536
537
538
539
540
541
542
543
544
545

25
546
547
Fig. 5. 548
549