~3--'3 - h.j. andrews experimental forest - oregon state...
TRANSCRIPT

~3--'3
122 J. R. WEBSTERAND J. 1. MEYER (EDITORS) [Volume 16
The apparent rate constant for the tempera-ture effect on specific respiration is too largeby more than an order of magnitude to be asimple metabolic response. Therefore, othercovariates must be amplifying the trend. Poten-tial covariates include a decline in BOC qualitywith increasing latitude caused by shifts in thedominant vegetation, and lower nutrient avail-ability with increasing latitude due to seques-tration and immobilization by stored OM. Jones(1997) reported that the abundance of FBOM iscorrelated with the abundance of CBOM,which in turn is correlated with the abundanceof wood, which is correlated with latitude andprecipitation. These trends suggest a latitudi-nal gradient in mean BOM particle size whichmight amplify the apparent temperature re-sponse of respiration, because, as a first ap-proximation, microbial activity per unit OM isa function of the ratio of surface area to volume
of the particles.It is also possible that sampling biases are
contributing to the respiration trend. I suggestthat in high gradient mountain streams, it is rel-atively "easy" to estimate BOC because the sed-iments are shallow and a high fraction of theBOC consists of woody boles and associated de-bris dams. However, this same heterogeneityalso makes it difficult to accurately estimate sys-tem respiration. Conversely, in more homoge-neous, low-gradient systems, estimating BOC iscomplicated by shoals and deep hyporheicchannels, perhaps leading to underestimates,while estimates of areal respiration may besomewhat easier to acquire.
Even without knowing the mechanisms ofamplification, the large-scale pattern is interest-ing. If interpreted literally, it portends dramaticchanges for stream ecosystems in the exigencyof global warming: a 50% increase in benthicrespiration for each 1°C rise in mean annual wa-ter temperature. Because streams within anecoregion show only weak relationships withtemperature, it appears that saltational changesin stream structure and function are associated
with shifts in major climatic and vegetationzones. Thus, even if significant climate changeoccurs over the next several decades, severalcenturies might be required for a stream to rees-tablish a new equilibrium condition with its wa-tershed.
Although current data suggest that alterationsin stream respiration with temperature will be
modest unless a watershed is pushed into a newbiome, the database for such conclusions issparse and there is good reason to question theadequacy of respiration data from only 22 sites.The apparent contrast between intra-biomeandinter-biome patterns highlights the need forlarge-scale studies that focus on the mecha-nisms controlling the abundance and metabo-lism of organic matte!:
Literature Cited
JONES,J. B. 1997. Benthic organic matter storage instreams: influence of detrital import and export,retention mechanisms, and climate. Pages 109-119 in J. R Webster and J. L. Meyer (editors).Stream organic matter budgets. Journal of theNorth American Benthological Society 16:3-161.
VOET,D., AND J. G. VOET. 1990. Biochemistry. JohnWtley & Sons, New York.
WEBSTER,J. R, AND J. L. MEYER(EDITORS).1997a.Stream organic matter budgets. Journal of theNorth American Benthological Society 16:3-161.
WEBSTER,J. R, AND J. L. MEYER. 1997b. Stream or-ganic matter budgets-introduction. Pages 5-13in J. R Webster and J. L. Meyer (editors). Streamorganic matter budgets. Journal of the NorthAmerican Benthological Society 16:3-161.
WEBSTER,J. R, J. B. WALLACE,AND E. F. BENFIELD.1995. Organic matter processes in streams of theeastern United States. Pages 117-187 in C. E.Cushing, K. W. Cummins, and G. W. Minshall(editors). River and stream ecosystems. Elsevier,Amsterdam.
Suspended particulate organicmatter concentration and
export in streams
STEPHEN W GOLLADAY
J.W.JonesEcological ResearchCenter,Rte 2, Box 2324,
Newton,Georgia31770 USA
From an ecosystem perspective, paM concen-tration and export are significant characteristicsof streams. The transport of material links up-stream and downstream communities trophi-cally into an integrated ecosystem. Most POMtransported in streams is fine material «1 mm)(Sedell et aI. 1978, Cudney and Wallace 1980,


1997] STREAM ORGANIC MATIER BUDGETS 123
Naiman 1982, Wallace et a1. 1982, Hill et a1.1992). Coarser material (Le., leaves and wood)tends to be retained near its point of entry, es-pecially in smaller streams (Webster et a1.1994).POM in transport originates from biological andphysical processing of leaves and wood (Web-ster 1983, Wotton 1984), erosion of soil organicmatter (Meybeck 1982, Hedges et a1. 1986), floc-culation of OOM (Bowen 1984, Wotton 1984),and sloughing or grazing of periphyton (Lam-berti et al. 1987). Concentrations and quantitiesof material exported are detennined by the in-teraction between availability of material to betransported, stream power, and efficiency of in-stream retention devices (Naiman 1982, Golla-day et al. 1987).
Although the importance of material trans-port is well recognized, applying the ecosystemconcept to streams has proved difficult for sev-eral reasons. Longitudinal boundaries of streamecosystems are often indistinct. Lateral bound-aries can vary seasonally as streams expand andcontract across their floodplains. Streams re-ceive substantial quantities of material from theadjacent landscape, but because of the unidirec-tional flow of water, more material typicallyflows through stream segments than is storedor cycled in place. These problems are greatestfor large streams; consequently most ecosystemstudies of streams have focused on headwaters
or individual reaches of larger rivers.Recent studies have suggested that position
within drainages and hydrologic linkages withriparian areas may determine the functional roleof reaches and segments within the largerdrainage (Naiman 1982, Pringle et a1. 1988,Meyer and Edwards 1990, Golladay and Taylor1995). An integrated view of stream ecosystemstructure and function emerges from linkingdiscrete units. If this view of streams is correct,then general trends should be apparent acrossa range of streams. This chapter examines sus-pended POM concentration and POM exportacross the data base of stream study sites de-scribed earlier in this paper.
Methods
The database included 31 streams with values
for average POM concentration and sufficient in'-formation to calculate percent of total organicmatter concentration [%POM = «POM)j(POM+DOM» x 100] (Table 1). Because streambed
area estimates were not available for some (es-pecially larger) streams, it was possible to cal-culate export (g . m-2 streambed area. y-l) foronly 22 of these sites. Storm transport of POMwas not sampled in all streams and this is notedin Table 1. Streams were also classified accordingto presence or absence of riparian forest (forestversus non-forest). The initial step in data anal-ysis was to examine scatter plots of ~ con-centration, %POM, and export. Occasionally, astream (or streams) appeared to be very differentfrom other sites. Subsequent statistical analyseswere performed with and without those "excep-tional" streams and noted in the results. Possible
explanations for exceptional streams, based onsite-description chapters or other information, areprovided in the Discussion.
Differences in POM concentration, % POM,
and POM export in forest and non-foreststreams were analyzed using a Mann-WhitneyRank Sum Test. POM concentration, %POM andPOM export were regressed against physicalfactors listed in the da~abase. In analyses ofstream order and gradient, streams were firstsorted by the presence or absence of riparianforest. Averages were then determined forstreams of the same order or similar gradient.Also, correlations were performed betweenPOM concentration, %POM, and POM exportversus measures of organic matter availability(FBOM, CBOM, total benthic organic matter,and wood standing stocks). Relevant transfor-mations were applied when necessary to meetthe assumptions of the statistical procedure. Sta-tistical analyses were conducted using Sigma-Stat for Windows V. 1.0 (Jandel Scientific Soft-ware, San Raphael, California).
Results
Of the physical factors analyzed, only totalannual precipitation, stream order, and gradientshowed any relationship with POM concentra-tion, %POM, or POM export. There was no cor-relation between measures of organic ma!terstanding stock and POM concentration, %POM,or POM export.
POM concentration
Streams draining forested areas had signifi-cantly higher <PieragePOM concentrations thanthose draining non-forested areas (Mann-Whit-

ney Rank Sum Test, T = 38.0,P = 0.004)(Table1). POM concentrations in forested streamsranged from 0.21 to 15.30mg AFDM/L and innon-forested streams ranged from 0.14 to 1.78mg AFDM/L. White Clay Creek (Pennsylvania)and BeaverCreek (Quebec) had ~e highest av-erage POM concentration, apr.oximately 1.5 to2x greater than any other stream.
With all streams included in the analysis therewas no relationship between POM concentrationand average annual precipitation (rl = 0.001,P =0.8). However, several sites were exceptional (Fig.1). White Clay Creek and Beaver Creek had highPOM concentrations with intermediate precipi-tation. Streams from the McKenzie River drain-
age in Oregon had relatively low POM concen-
124 J. R. WEBSTERAND J. L. MEYER (EDITORS) [Volume 16
TABLE1. Particulate organic matter concentration, contribution of POM to total organic concentration (%POM), and POM export (per unit streambed area) for a number of intensively studied streams. Column 6indicates whether storms were systematically sampled during the process of estimating POM concentrationand export. Stream acronyms are defined by Webster and Meyer (1997).
POM cone. POM export StormsSite Order (mg/L) %POM (g.m-Z.y-I) sampled?
Forest
W30R 1 0.21 28 123 NW40R 1 1.41 44 817 NAGMI 1 5.50 40 2146 NDCOR 1 0.95 37 170 N
FCQB 1 0.63 17 1733 YBBGR 1 2.66 31 687 NBBVA 1 7.17 26 1108 YWBTN 1 3.62 67 1087 YSBNC 1 2.06 67 300 YAKC2 1 1.10 15 586 ?AKC3 1 1.50 7 469 ?HWNC 2 7.22 71 535 y
BCQB 2 11.15 6 - YBBNH 2 1.40 77 267 YCSNC 3 2.06 5 - yMCOR 3 0.46 40 - NWCPA 3 15.30 27 2317 YFRMA 4 1.95 24 - YKCAU 4 5.89 53 1931 YLCOR 5 0.51 42 - NMRQB 5 1.50 7 - YMTQB 6 1.94 6 - YORGA 6 1.14 4 181 YMROR 7 0.68 38 - NMOQB 9 1.21 9 - YMean 3.17 33 904Median 1.50 28 636
Range (25-75%) 1.06-4.09 9-43 283-1420
Non-forest
RSWA 1 0.33 32 75 NCSAN 1 0.33 40 6 ?KPKA 3 0.14 5 2 NKRAK 4 1.78 8 107 YKGKA 5 0.21 8 2 NSCAZ 5 0.47 2 36 YMean 0.55 16 38Median 0.33 8 21
Range (25-75%) 0.21.47 5-32 2-75

"
"
FIG.1. POM concentration versus averageannual precipitation, stream gradient, and stream order. Streamsof similar order or gradient were averaged prior to regression analysis.
trations and high precipitation. With thosestreams excluded from the analysis, POM con-centration was positively correlated with averageannual precipitation (r2 = 0.43,P = 0.0007)(Fig.1). Streams receiving greatest amounts of precip-itation generally occurred in forested areas.
For forest streams, POM concentration wasnegatively correlated with stream gradient (r2=0.87 P = 0.007), i.e. low-gradient streams hadthe highest POM concentrations (Fig. 1). There
were not enough non-forest streams for analy-sis. Generally, non-forest streams had 10werPOM concentrations than forest streams of sim-
ilar gradient.POM concentration in forest streams was
weakly negatively correlated with stream order,(r2 = 0.38,P = 0.1, POM concentration In trans-.formed) (Fig. 1). No relationship was observedbetween POM concentration and stream orderin non-forest streams.
1997] STREAMORGANICMAITER BUDGETS 125
18 , WhiteClayCreek,PA
IForest
15-1 " Non-Forest
Beaver Creek, OBExceptional
121 "...J
9'0E
6 . . f=o.43. p=O.OOO7. .3-1
" MacKenzieRiver0-1 f)" Drainage, OR
I I I I I I I
0 50 100 150 200 250 300
Precipitation (cm)e 60..e- I.eQ) 40e ...J0 0, ...U E I ------ f=o.87. p:0.OO7
20a.. I 0
O0.0 0.1 0.2 0.3 0.4 0.5
Gradient(mIm)I
2 ..f=O.38. p=O.1., --- -
:::r ."00 . .-1 "1 0 0
-2 -I 0I I I
2 3 4 5 6 7
Order

126 J. R. W~BSTERAND J. L. MEYER (EDITORS) [Volume 16
. Foresto Non-Forest
'V Exceptional
.. . . .
1=0.28, p:O.006
i'V'V
MacKenzie RiverDrainage, OR
50
Precipitation (em)
100 150 200 250
FIG. 2. Percent POM versus precipitation.
Percent POM
There was no difference in %POM betweenforest and non-forest streams (Mann-WhitneyRank Sum Test, T = 65, P = 0.13).Percent POMwas weakly correlated with average annual pre-cipitation (r2= 0.20,P = O.Ol).(Fig.2). If streamsof the McKenzieRiver drainage in Oregon wereexcluded, the relationship improved somewhat(r2= 0.28,P = 0.006).No relationshipwas ob-served between percent POM and gradient ororder for either forest or non-forest streams.
POM export
Streams draining forested areas had signifi-cantly higher rates of POM export than thosedraining non-forested areas (Mann-WhitneyRank Sum Test, T = 21.0, P = 0.0001) (Table 1).POM export in forest streams ranged from 123to 2317 g . m-2 streambed area. y-l and in non-forest streams ranged from 2 to 107 g . m-2streambed area. y-l. White Clay Creek had thehighest export.
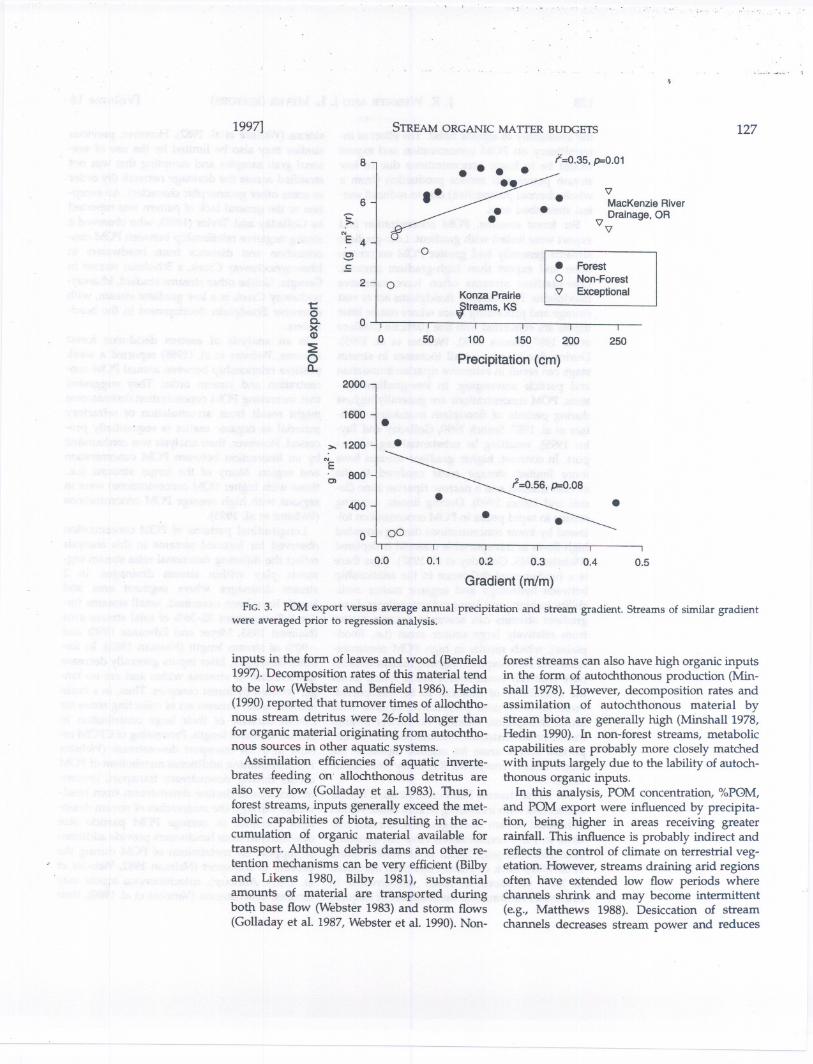
With all streams included in the analysisthere was no relationship between POM exportand average annual precipitation (r2 = 0.11,P =0.14, POM export In transformed). However,several sUes were exceptional (Fig. 3). Streamsfrom the McKenzie River drainage had relative-ly low POM eXfort and received high levels of
precipitation. Streams from Konza prairie, Kan-sas, had low export and received intermediatelevels of precipitation. With the exceptionalstreams excluded from the analysis, POM ex-port was positively correlated with average an-nual precipitation (r2 = 0.35,P = 0.01,POMex-port In transformed).
For forest streams, POM export was negative-ly correlated with stream gradient (r2 = 0.56,P= 0.08), i.e., streams with lowest gradient gen"erally exported more POM than high gradientstreams (Fig. 3). There were not enough non-forest streams for analysis. Generally non-foreststreams had lower export than forest streams ofsimilar gradient. No relationship was found be-tween export and order for forest or non-foreststreams.
Discussion
General patterns across streams
Terrestrial vegetation appears to control or-ganic matter concentration and export across arange of streams. In this study, forest streamshad higher POM concentrations and POM ex-port than non-forest streams. Consistently high-er concentrations and export in forest streamscan be linked to basic differences in organic in-puts and processing between the 2 streamtypes. Forest streams receivehigh aIle-..:hthonous
100
80
0 60a..-c
40Q)
Q)a..
20
o 'T0
1997]
;.
STREAM ORGANIC MATTER BUDGETS 127
8 1"=0.35. p:O.01
FIG.3. POM export versus average annual precipitation and stream gradient. Streams of similar gradientwere averaged prior to regression analysis.
inputs in the form of leaves and wood (Benfield1997).Decomposition rates of this material tendto be low (Webster and Benfield 1986).Hedin(1990)reported that turnover times of allochtho-nous stream detritus were 26-fold longer thanfor organic material originating from autochtho-nous sources in other aquatic systems.
Assimilation efficiencies of aquatic inverte-brates feeding on' allochthonous detritus arealso very low (Golladay et al. 1983).Thus, inforest streams, inputs generally exceed the met-abolic capabilities of biota, resulting in the ac-cumulation of organic material available fortransport. Although debris dams and other re-tention mechanisms can be very efficient (Bilbyand Likens 1980, Bilby 1981), substantialamounts of material are transported duringbo~ base flow (Webster 1983)and storm flows(Golladay et al. 1987,Webster et al. 1990).Non-
forest streams can also have high organic inputsin the form of autochthonous production (Min-shall 1978). However, decomposition rates andassimilation of autochthonous material bystream biota are generally high (Minshall 1978,Hedin 1990). In non-forest streams, metaboliccapabilities are probably more closely matchedwith inputs largely due to the lability of autoch-thonous organic inputs.
In this analysis, POM concentration, %PGM,and POM export were influenced by precipita-tion, being higher in areas receiving greaterrainfall. This influence is probably indirect andreflects the control of climate on terrestrial veg-etation. However, streams draining arid regionsoften have extended low flow periods wherechannels shrink and may become intermittent(e.g., Matthews 1988). Desiccation of streamchannels decreases stream power and reduces
v
61. . MacKenzieRiver.
Drainage.ORVvN
E 4
2L
0
IForestNon-Forest
Konza Prairie Exceptional1:::
o Itreams. KS0
0-)(Q) 0 50 100 150 200 250
Precipitation (em)011..
2000
1600 . .>- 1200 ...J .
NE
C) 800
400
0'.,.-0.0 0.1 0.2 0.3 0.4 0.5
Gradient (m/m)

128 J. R. WEBSTER AND J. 1. MEYER (EDITORS) [Volume 16
the availability of aquatic habit. The effect of in-termittency. on POM concentration and exportwould be to lower concentrations due to low
stream power and reduce production (from awhole channel perspective) due to reduced wet-ted streambed area.
For forest streams, POM concentration andexport were linked with gradient. Low-gradientstreams generally had greater POM concentra-tions and export than high-gradient streams.Low-gradient streams often have extensivefloodplains. Functionally, floodplains act as vaststorage and processing areas where coarse litterinputs are converted into fine particles (Wallaceet al. 1987, Smock 1990, Webster et al. 1995).During floods, even small increases in streamstage can result in extensive riparian inundationand particle scavenging. In low-gradient sys-tems, POM concentrations are generally highestduring periods of floodplain inundation (Wal-lace et al. 1987, Smock 1990, Golladay and Tay-lor 1995), resulting in substantial organic ex-port. In contrast, higher gradient streams havemore limited storage areas confined to thestream channel and a narrow riparian zone (Se-dell and Dahm 1990). During floods, scouringresults in rapid peaks in POM concentration fol-lowed by lower concentrations during extendedhigh flows as transportable material is depleted(Webster 1983, Golladay et al. 1987). Thus thereis a fundamental difference in the relationshipbetween hydrology and organic matter avail-ability in high- and low-gradient systems. Low-gradient streams can scavenge organic matterfrom relatively large source areas (i.e., flood-plains), which results in high POM concentra-tions and POM export. Higher-gradient streamshave relatively small source areas, which limitsthe availability of material to be transported.From an ecosystem perspective, reaches withbroad floodplains, whether headwater swampforests or downstream bottomlands, can be im-portant source areas for organic inputs to thelarger stream ecosystem (Golladay and Taylor1995). .
For forest streams POM concentration tended
to decrease with increasing stream order. Withinindividual stream drainages, downstream pat-terns in POM concentration have not generallybeen observed (e.g., Sedell et al. 1978, Wallacect al. 1982). Lack of pattern has been attributedto local variations in reach reteritiveness in
headwater streams and the influence of frequent
storms (Wallace et al. 1982). However, previousstudies may also be limited by the use of sea-sonal grab samples and sampling that w~s notstratified across the drainage network (by orderor some other geomorphic character). An excep-tion to the general lack of pattern was reportedby Golladay and Taylor (1995), who observed astrong negative relationship between POM con-centration and distance from headwaters in
Ichawaynochaway Creek, a 5th-order stream inGeorgia. Unlike other streams studied, Ichaway-nochaway Creek is a low gradient stream, withextensive floodplain development in the head-waters.
In an analysis of eastern deciduous foreststreams, Webster et al. (1995) reported a weakpositive relationship between annual POM con-centration and stream order. They suggestedthat increasing POM concentration downstreammight result from accumulation of refractorymaterial as organic matter is sequentially pro-cessed. However, their analysis was confoundedby an interaction between POM concentrationand region. Many of the larger streams (i.e.,those with higher POM concentrations) were inregions with high average POM concentrations(Webster et al. 1995).
Longitudinal patterns of POM concentrationobserved for forested streams in this analysisreflect the differing functional roles stream seg-ments play within stream drainages. In 2stream drainages where segment area andlength have been examined, small streams (or-der 1-3) represent 32-36% of total stream area(Naiman 1983, Meyer and Edwards 1990) and.-90% of stream length (Naiman 1983). In for-ested drainages, litter inputs generally decreasedownstream as streams widen and are no lon-
ger covered by forest canopies. Thus, in a basincontext, small streams act as collecting zones forCPOM because of their large contribution tostream area and length. Processing of CPOM re-sults in POM transport downstream (Webster1983). Assuming additional metabolism of POMoccurs during downstream transport, concen-tration should decline downstream from head-
waters through the midreaches of stream drain- .ages. Declines in average POM particle sizedownstream from headwaters provide addition-al support for metabolism of POM during theprocess of transport (Naiman 1982, Webster etal. 1995). Althouj!~1 autochthonous inputs mayincrease downstream (Vannote et al. 1980), their

1997) STREAM ORGANIC MAnER BUDGETS 129
relativelyhigh turnover (Hedin 1990)may min-imize contributions to POM concentration.
Exceptions to general patterns
Many values listed in Table 1 represent flow-weighted annual concentrations or were ob-tained using rating curves derived from exten-sive base-flow and storm sampling. Assumingsampling is adequate, those estimates incorpo-rate the influence of storms, periods of muchorganic transport (Webster et al. 1990).Severalstreams had POM concentrations outside therange reported for most streams. BeaverCreekand White Clay Creek had relativelyhigh POMconcentrations (Table 1). In both cases, stormswere systematically sampled and incorporatedinto estimates of average POM concentration.Streams of the McKenzie River drainage, Ore-gon, had relatively low POM concentrations,es-pecially compared with other forested streams.Those streams were sampled seasonally butstorms were not systematically sampled. Thelow POM export reported from Konza Prairiestreams may also be due to a lack of storm sam-pling. Substantial export of organic matter hasbeen measured during storms in other prairiestreams and has been attributed to low numbersof debris dams or other retention structures(Hill and Gardner 1987).Clearly, intensive sam-pling over a range of hydrologic conditions isessential for accurately determining trends instream ecosystem function. It is not clearwheth-er high POM concentrations in Beaver Creekand White Clay Creek reflect more rigorousstorm sampling than other sites, or perhaps ayear with higher than average organic concen-trations.
Limitations of the data base andgeneral conclusions
This analysis ~learly demonstrates that gen-eral trends in stream ecosystem structure andfunction exist across a range of stream sites.However, the ability to generalize is limited bythe nature of the data. Most of the streams an-
alyzed were smaller than 3rd order. Clearlymore information is needed on larger streams,especially in reg;..:>nslike the southeastern USwhere many la::ger rivers remain relatively un-disturbed and unregulated (Benke 1990). A few
measurements made from streams drainingarid regions (non-forested) suggest more workis needed in the Great Plains and southwesternUS, again, particularly in larger rivers.
Several general trends emerge from the anal-ysis. Terrestrial or riparian vegetation plays animportant role in regulating POMconcentrationand export. Streams draining forested areashave high POM concentrations and export be-cause organic inputs exceed within-reach met-abolic potential; excess material is lost throughdownstream transport. Gradient influencesPOM concentration and export through an in-teraction with riparian geomorphology. Low-gradient streams, i.e., those with broad flood-plains, have greater processing and source areasfor POM than high-gradient and more con-strained streams. In forested streams, positionwithin the drainage influencesPOM concentra-tion. Small streams (orders 1-3),because of theirrelatively large surface area and length, act ascollecting zones for litter inputs within the larg-er stream drainage. This material is subsequent-ly processed, transported, and further metabo-lized as it moves downstream, resulting indownstream declines in POM concentration.General trends emerge across a range of streamstudy sites and suggest that an integrated viewof stream ecosystems results from examiningthe structure and function of individual seg-ments and assembling them in a whole drain-age context.
Acknowledgements
Funding for travel to the Stream OrganicWorkshop was provided by the J. W.Jones Eco-logical Research Center and R. W. WoodruffFoundation. The J. W.Jones EcologicalResearchCenter also provided technical support duringthe preparation of this manuscript. I thank DrsR. J. Mackay, J. R. Webster, and J. L. Meyer, 2anonymous referees, and the participants of theOrganic Matter Workshop for many helpfulcomments.
Literature Cited
BENFIELD,E. F. 1997. Comparisons of litterfall inputto streams. Pages 104-108 in J. R. Webster and J.L. Meyer (editors). Stream organic mltter bud-gets. Journal of the North American ~~nthologicalSociety 16:3-161.

130 J. R. WEBSTER AND J. L. MEYER (EDITORS) [Volume 16
BENKE,A. C. 1990. A perspective on America's van-ishing streams. Journal of the North AmericanBenthological Society 9:77-88.
BILBY,R.E. 1981. Role of organic debris dams in reg-ulating the export of dissolved and particulatematter from a forested watershed. Ecology 62:1234-1243.
BILBY,R. E., ANDG. E. LIKENS. 1980. Importance oforganic debris dams in the structure and functionof stream ecosystems. Ecology 61:1107-1113.
BoWEN,S. H. 1984. Evidence of a detritus food chain
based on consumption of organic precipitates.Bulletin of Marine Science 35:440-448.
CUDNEY,M. D., ANDJ. B. WALLACE.1980. Life cycles,microdistribution and production dynamics of sixspecies of net-spinning caddisflies in a largesoutheastern (U.S.A.) river. Holartic Ecology 3:169-182.
GOLLADAY,S. w., AND B. W. TAYLOR.1995. Charac-
teristics of suspended particulate matter in Icha-waynochaway Creek, a brown-water coastal plainstream. Pages 380-383 in K. J. Hatcher (editor).Proceedings of the 1995 Georgia Water ResourcesConference. The University of Georgia, Athens.
GOLLADAY, S. W., J. R. WEBSTER, AND E. F. BENFIELD.
1983. Factors affecting food utilization by a leafshredding aquatic insect: leaf species and condi-tioning time. Holartic Ecology 6:157-162.
GoLLADAY,S. w., J. R. WEBSTER,AND E. F. BENFIELD.1987. Changes in stream morphology and stormtransport of seston following watershed distur-bance. Journal of the North American Benthol-ogical Society 6:1-11.
HEDGES,J. I., J. R. ERTEL,P. D. QUAY,P. M. GROOTES,J. E. RICHEY,A. H. DEVOL,G. W. FARWELL,F. W.
ScHMIDT,ANDE. SALATI. 1986. Organic carbon-14 in the Amazon River system. Science 231:1129-1131.
HEDIN,L. 0. 1990. Factors controlling sediment com-munity respiration in woodland stream ecosys-tems. Oikos 57:94-105.
HILL, B. H., ANDT. J. GARDNER.1987. Seston dynam-ics in two Texas prairie streams. American Mid-land Naturalist 118:85-93.
HILL, B. H., T. J. GARDNER,AND0. F. EKISOLA.1992.Predictability of streamflow and particulate or-ganic matter concentration as indicators of stabil-ity in prairie streams. Hydrobiologia 242:7-18.
LAMBERTI,G. A., L. R. ASHKENAS,S.A. GREGORY, AND
A. 0. STEINMAN. 1987. Effects of three herbivores
on periphyton communities in laboratorystreams. Journal of the North American Benthol-
ogical Society 6:92-104. .MATTHEWS,W. J. 1988. North American prairie
streams as systems for ecological study. Journalof the North American Benthological Society 7:387--409.
MEYBECK,M. 1982. Carbon, nitrogen, and phospho-
rus transport by world rivers. American Journalof Science 282:401--450.
MEYER,J. L., ANDR. T. EDWARDS. 1990. Ecosystemmetabolism and turnover of organic carbon alonga blackwater river continuum. Ecology 71:668-677.
MINSHALL,G. W. 1978. Autotrophy in stream ecosys-tems. BioScience 28:767-771.
NAIMAN,R. J. 1982. Characteristics of sediment and
organic carbon export from pristine boreal forestwatersheds. Canadian Journal of Fisheries and
Aquatic Sciences 39:1699-1718.NAIMAN,R. J. 1983. The annual pattern and spatial
distribution of aquatic oxygen metabolism in bo-real forest watersheds. Ecological Monographs 53:73-94.
PRiNGLE,C. M., R. J. NAIMAN, G. BRETSCHKO,J. R.KARR,M. W. OsWOOD,J. R. WEBSTER,R.L. WEL-
COMME, AND M. J. WINTERBOURN. 1988. Patch dy-
namics in lotic ecosystems: the stream as a mo-saic. Journal of the North American BenthologicalSociety 7:503-524.
SEDELL,J. R., ANDC. N. DAHM. 1990. Spatial and tem-
poral scales of dissolved organic carbon instreams and rivers. Pages 261-279 in E. M. Perdueand E. T. Gjessing (editors). Organic acids inaquatic ecosystems. John Wiley and Sons, NewYork.
SEDELL,J. R., R. J. NAIMAN, K. W. CUMMINS,G. W.MINSHALL,ANDR. L. VANNOTE.1978. Transport
of particulate organic material in streams as afunction of physical processes. Verhandlugen derInternationalen Vereinigung fur theoretische undandewandte Limnologie 20:1366-1375.
SMOCK,L. A. 1990. Spatial and temporal variation inorganic matter storage in low-gradient, headwa-ter streams. Archiv fur Hydrobiologie 118:169-184.
VANNOTE,R. L., G. W. MINSHALL,K. W. CUMMINS,J.R. SEDELL,AND C. E. CUSHING. 1980. The rivercontinuum concept. Canadian Journal of Fisheriesand Aquatic Sciences 37:130-137.
WALLACE,J. B., A. C. BENKE,A. H. LiNGLE,AND K.PARSONS..1987. Trophic pathways of macroin-vertebrate primary consumers in subtropicalblackwater streams. Archiv fur Hydrobiologie,Supplement 74:423--451.
WALLACE,J. B., D. H. Ross, ANDJ. L. MEYER.1982.Seston and dissolved organic carbon dynamics ina southern Appalachian stream. Ecology 63:824-838.
WEBSTER,J. R. 1983. The role of benthic macroinver-tebrates in detritus dynamics of streams: a com-puter simulation. Ecological Monographs 53:383-4J4.
WLBSTER,J. R., AND E. F. BENFIELD.1986. Vascularplant breakdown in freshwater ecosystems. An-

1997] S1REAM ORGANIC MATTER BUDGETS 131
nual Review of Ecology and Systematics 17:567-594.
WEBSTER, J. R, A. P. COVICH, J. L. TANK, AND T. V.
CROCKEIT. 1994. Retention of coarse organic par-ticles in streams in the southern AppalachianMountains. Journal of the North American Ben-
thological Society 13:140-150.
WEBSTER, J. R, S. W. GOLLADAY, E. F. BENFIELD, D. J.D'ANGELO, AND G. T. PETERS. 1990. Effects of for-
est disturbance on particulate organic matterbudgets of small streams. Journal of the North
American Benthological Society 9:120-140.
WEBSTER,J. R, AND J. L. MEYER. 1997. Stream organicmatter budgets-introduction. Pages 5-13 in J. RWebster and J. L. Meyer (editors). Stream organicmatter budgets. Journal of the North AmericanBenthological Society 16:3-161.
WEBSTER,J. R, J. B. WALLACE,AND E. F. BENFIELD.1995. Organic processes in streams of the easternUnited States. Pages 117-187in C. E. Cushing, K.W. Cummins, and G. W. Minshall (editors). Riverand Stream Ecosystems. Elsevier, Amsterdam.
WOTION, R S. 1984. The importance of identifyingthe origins of microfine particles in aquatic sys-tems. Oikos 43:217-221.
Dissolved organic matterconcentration and flux in streams
PATRICKJ.MULHOLLAND
Environmental Sciences Division,
Oak Ridge Nationall.Jlboratory,P'Q Box 2008,
Oak Ridge, Tennessee37831-6036 USA
Dissolved organic matter (DaM) in streams isimportant as an energy resource for food webs(Bott et al. 1984, Meyer 1990a), a regulator ofnutrient uptake and cycling by heterotrophicmicrobes (Lock 1981, Meyer et al. 1988), a com-plexing agent for metals (McKnight and Bencala1990, Mierle and Ingram 1991)/ and a determi-nant of pH and alkalinity (Oliver et al. 1983).DaM has been shown to be an important met-abolic substrate in 'some streams, with DaM usecontributing significantly to total stream metab-olism (Naiman et al. 1987/ Meyer and Edwards1990). Sources of DaM are both the terrestrialwatershed (McDowell and Likens 1988/ Horn-berger et al. 1994) and instream processes, suchas le'~ching and decomposition of allochthonouspi'l'ticulate organic matter and release by streamalgae (Kaplan and Bott 1982/ Meyer 1990b). For
small streams, the primary site of DOM use isthe stream bottom, including the interstitial wa-ters of streambed sediments (Dahm 1981/ Hy-nes 1983/ Ford and Naiman 1989/ Fiebig andLock 1991). In larger rivers, the water columnmay also be an important site for OOM use(Vannote et al. 1980).
Stream OOM is an important indicator of wa-tershed-scale hydrologic and biogeochemicalprocesses. Large water-soluble pools of DOMare present in the upper soil horizons of forestsand grasslands (litter, 0/ and A horizons) andare sources of OOM to drainage waters; how-ever/ lower soil horizons (B and C horizons) of-ten contain materials (e.g., iron and aluminumoxides) that effectively sorb and immobilizeOOM (McDowell and Wood 1984/ Cronan 1990).Hydrologic pathways involving lateral flow ofwater through or over surface soil layers oftenhave relatively high concentrations of DaM,whereas water flowpaths that pass slowlythrough lower soil layers usually have low DaMconcentrations (Moore and Jackson 1989/ Kaplanand Newbold 1993). The commonly observedincrease in DaM concentrations during periodsof high discharge is at least partly due to shiftsin dominant flowpaths from deeper routes toshallow subsurface or surface pathways (Tateand Meyer 1983/ Mulholland et al. 1990/ Horn-berger et al 1994). Wetlands are also an impor-tant source of DaM to streams, and severalanalyses have shown that differences in base-flow OOM concentrations among streams arestrongly related to the amount of wetland drain-age contributing to streams (Mulholland andKuenzler 1979/ Eckhardt and Moore 1990). Sat-urated riparian soils and wetlands appear to beparticularly important contributors to streamDaM, especially during storms (Fiebig et al.1990/ Hemond 1990).
In this paper I focus on the concentrationsand flux of DaM in streams whose organic mat-ter budgets are presented in the precedingchapters. My objective is to define the variationin annual average DaM concentrations and fluxand to identify what factors might be mosf re-sponsible for this variation.
Methods
The data used in this cnalysis are presentedand described in the sitt:-description chapters inthis paper with the fol1owing exceptions. Values


(
olC~
STREAM ORGANIC MATTER BUDGETS
.
EDIlED BY
1. R. WEBSlER AND JUDY L. MEYER
:
Made in United States of AmericaReprinted from JOURNALOF THENORTH AMERICANBENTHOLOGICALSOCIETY
Vol. 16, No.1, March 1997iO 199"'7by The North American Benthological Society

J. N. Am. Benthol. Soc., 1997, 16(1):J-161~ 1997 by The North American BenthologicaI Society
Stream organic matter budgets1,2
EDITED BY
J. R. WEBSTERAND JUDY L. MEYER
Abstract. This analysis of organic matter dynamics in streams has 3 objectives: 1) to explore therelationships between physical characteristics of streams and their watersheds (climate, geomorphol-ogy) and stream organic matter dynamics using data from a broad geographic area; 2) to comparestream organic matter dynamics in a diverse array of streams in order to suggest determinants ofobserved patterns; and 3) to reveal deficiencies in currently available data on organic matter dynamicsin streams. Streams were included in this analysis not to represent the global diversity of streamtypes but because organic matter data were available. In the introductory chapter we describe thekinds of data included for each stream and provide brief descriptions of previously published organicmatter data for streams included in the comparative analysis but not described in individual chapters.The next 16 chapters present organic matter data for streams from North America, Europe, Australia,and Antarctica. Most of the streams represented are in the temperate zone of North America. Datapresented include climate and geomorphic variables and organic matter inputs, exports, and standingcrops. The chapters on individual streams are followed by 7 chapters analyzing physical features ofthese streams and specific components of the organic matter budgets. Stream size, water temperature,and precipitation were the most important variables setting the physical template for organic matterprocesses occurring in the streams. Watershed area was the best predictor of gross primary produc-tivity (GPP), which increased with increasing watershed area. Watershed area, discharge, and solublereactive phosphorus concentration explained 71% of the variation in GPP. Climate (latitude) andvegetation type were more important than stream order in predicting litter inputs across a broadgeographic range of streams, although, within a river basin, litterfall decreased with increasingstream order. Regression of benthic organic matter (BOM) and latitude and precipitation proveduseful in predicting BOM standing crop in streams at a continental scale, although 80M was alsorelated to channel characteristics such as gradient and woody debris. Benthic respiration increaseddramatically with increasing temperature (Q,o = 7.6), suggesting a response related not only tometabolism but also to changes in BOM quality in response to latitudinal shifts in vegetation. Ter-restrial and riparian vegetation was found to play an important role in regulating suspended partic-ulate organic matter (POM) concentration and export, with higher values observed in forested streamsand in lower gradient streams with extensive floodplains. Channel slope was the best predictor ofdissolved organic matter (DOM) concentration and export, probably because of its relationship withriparian wetlands and hydrologic flowpaths. In the final chapter, a synthesis of the organic matterbudgets, we reached two conclusions: 1) At a global level, stream organic matter dynamics are drivenprimarily by climate through its effect on terrestrial vegetation. 2) Despite significant progress inunderstanding organic matter processes in streams, many of the differences we found among streamsreflect omissions of important components of the budget, especially accurate measures of streambedarea, heterotrophic respiration, standing stock of fine BOM, and groundwater inputs of DOM.
Key loords: stream, organic matter, budget, primary production, litterfall, BOM, DOM, POM, res-piration.
Table of Contents
~lntroduction (J. R. Webster and J. L. Meyer) 5
Canada Stream: a glacial meltwater stream in Taylor Valley,South Victoria Land,Antarctica (0. M. McKnight and C. M. Tate) 14
I Individual chapters should be cited as follows: Author(s). 1997. Chapter title. Pages xxx-xxx in J. R. Websterand J. L. Meyer (editors). Stream organic matter' budgets. Journal of the North American Benthological Society16:3-161.
2 Reprint reC;uests for individual chapters should be sent to the appropriate senior author. (' oJrrespondenceconcerning the entire paper should be directed to: J. R. Webster, Department of Biology, Virginia PolytechnicInstitute and State University, Blacksburg, Virginia 24061 USA.
3