77, 3 2002 the quarterly review - yale university history museum, university of kansas lawrence,...
TRANSCRIPT

Volume 77, No. 3 September 2002
261
The Quarterly Review of Biology, September 2002, Vol. 77, No. 3
Copyright � 2002 by The University of Chicago. All rights reserved.
0033-5770/2002/7703-0001$15.00
The Quarterly Reviewof Biology
THE EVOLUTIONARY ORIGIN AND DIVERSIFICATIONOF FEATHERS
Richard O. Prum
Department of Ecology and Evolutionary Biology, andNatural History Museum, University of Kansas
Lawrence, Kansas 66045 USA
e-mail: [email protected]
Alan H. Brush
92 High Street
Mystic, Connecticut 06355 USA
e-mail: [email protected]
keywordsfeathers, evolution, development, paleontology, birds, dinosaur
Although paleontological evidence is revealing the origin of birds, neither paleontologicalnor developmental evidence yet speaks more than a whisper about the origin of feathers
(Hall 1998:266).
abstractProgress on the evolutionary origin and diversification of feathers has been hampered by conceptual
problems and by the lack of plesiomorphic feather fossils. Recently, both of these limitations have beenovercome by the proposal of the developmental theory of the origin of feathers, and the discovery ofprimitive feather fossils on nonavian theropod dinosaurs. The conceptual problems of previous theoriesof the origin of feathers are reviewed, and the alternative developmental theory is presented and dis-cussed. The developmental theory proposes that feathers evolved through a series of evolutionary nov-elties in developmental mechanisms of the follicle and feather germ. The discovery of primitive andderived fossil feathers on a diversity of coelurosaurian theropod dinosaurs documents that feathersevolved and diversified in nonavian theropods before the origin of birds and before the origin of flight.The morphologies of these primitive feathers are congruent with the predictions of the developmentaltheory. Alternatives to the theropod origin of feathers are critiqued and rejected. Hypotheses for the

262 Volume 77THE QUARTERLY REVIEW OF BIOLOGY
initial function of feathers are reviewed. The aerodynamic theory of feather origins is falsified, butmany other functions remain developmentally and phylogenetically plausible. Whatever their function,feathers evolved by selection for a follicle that would grow an emergent tubular appendage. Feathersare inherently tubular structures. The homology of feathers and scales is weakly supported. Feathersare composed of a suite of evolutionary novelties that evolved by the duplication, hierarchical organi-zation, interaction, dissociation, and differentiation of morphological modules. The unique capacityfor modular subdivision of the tubular feather follicle and germ has fostered the evolution of numerousinnovations that characterize feathers. The evolution of feather keratin and the molecular basis offeather development are also discussed.
FEATHERS are the most complex integu-mentary appendages found in vertebrates
(Lucas and Stettenheim 1972; Bereiter-Hahnet al. 1986). They have complex branchedstructure, and grow from their bases by aunique mechanism (Figures 1 and 2). The evo-lutionary origin of feathers has been a persis-tent and intractable question for more than140 years (Lucas and Stettenheim 1972; Dyck1985; Feduccia 1999; Maderson and Homber-ger 2000). Two important sources have con-tributed to the fundamental difficulty of study-ing this problem: the intellectual limitations ofavailable models, and the lack of any antece-dent fossil feather structures. Over the last fewyears, both problems have been addressed inways that have fundamentally changed ourconception of and answers to these evolution-ary questions. Recent proposals of the devel-opmental theory (Prum 1999; Brush 2000),and startling new paleontological discoveriesof primitive feathers in nonavian theropoddinosaurs (Chen et al. 1998; Ji et al. 1998; Xuet al. 1999a, 1999b, 2000, 2001; Ji et al. 2001),have made it possible to make the first con-crete conclusions about the evolutionary ori-gin of feathers.
The earliest known feathers appear in thefossil record in Archaeopteryx lithographica,known from the 140 million-year-old Soln-hofen Limestone of Germany (de Beer 1954).The discovery of these spectacular fossils inthe 1860s stunned scientists because of theirmosaic of primitive reptilian and modernavian features, including essentially modernfeathers (Griffiths 1996; Martin and Czerkas2000). Most specimens of Archaeopteryx arepreserved with impressions of the remiges andrectrices (flight feathers of the wings and tail,respectively) that exhibit asymmetrical, closedpennaceous vanes indicative of advancedflight capability. The closed pennaceous struc-
ture of the remiges and rectrices of Archaeop-teryx demonstrates an entirely modern mor-phology, however, including differentiateddistal and proximal barbules that interlockbetween neighboring barbs to create the pla-nar vane of modern feathers (Griffiths 1996;Martin and Czerkas 2000). Thus, the oldestknown fossil feathers give no more clues as tothe ancestral morphology and ultimate originof feathers than do the feathers of extant birds.(In this paper, the terms Aves, birds, and avianrefer to members of the most inclusive cladeincluding Archaeopteryx and modern birds. Fora discussion of alternatives, see Gauthier andde Queiroz 2001.)
Research on the origin of feathers requiresa backward extrapolation from the complex,entirely modern feathers of Archaeopteryx andmodern birds to propose plausible ancestralfeather morphologies. Unfortunately, thedevelopment of a heuristic theory of the ori-gin of feathers has been limited by many ofthe same conceptual problems faced by mac-roevolutionary biology over the last century.Early workers attempted to reconstruct prim-itive feather morphologies based on varia-tions in feather structures found among“primitive” lineages of extant birds (reviewedin Dyck 1985). In absence of an explicit con-cept of phylogeny, these theories overlookedthe fact that all modern birds share a com-mon ancestor with Archaeopteryx that alreadyhad fully modern feathers. Therefore, extantvariations were derived, secondarily simpli-fied feather morphologies. Since 1950, manytheories focused on constructing functionaltheories for the origin of feathers (reviewedin Dyck 1985; Feduccia 1999). These theoriesused speculations about the plausible func-tion of ancestral feathers to predict their mor-phology, despite the paucity of evidenceabout the biology of avian ancestors. Func-
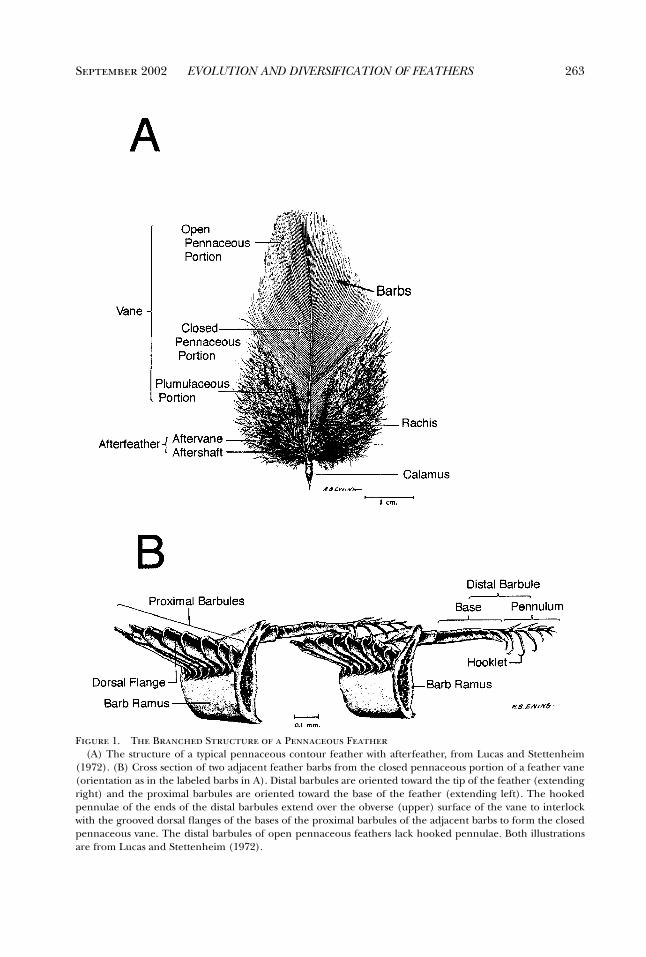
September 2002 263EVOLUTION AND DIVERSIFICATION OF FEATHERS
Figure 1. The Branched Structure of a Pennaceous Feather(A) The structure of a typical pennaceous contour feather with afterfeather, from Lucas and Stettenheim
(1972). (B) Cross section of two adjacent feather barbs from the closed pennaceous portion of a feather vane(orientation as in the labeled barbs in A). Distal barbules are oriented toward the tip of the feather (extendingright) and the proximal barbules are oriented toward the base of the feather (extending left). The hookedpennulae of the ends of the distal barbules extend over the obverse (upper) surface of the vane to interlockwith the grooved dorsal flanges of the bases of the proximal barbules of the adjacent barbs to form the closedpennaceous vane. The distal barbules of open pennaceous feathers lack hooked pennulae. Both illustrationsare from Lucas and Stettenheim (1972).
264 Volume 77THE QUARTERLY REVIEW OF BIOLOGY

September 2002 265EVOLUTION AND DIVERSIFICATION OF FEATHERS
tional theories of the origin of feathers havefailed to establish a consensus on either theoriginal function or original morphology offeathers.
Over the last half of the 20th century, neo-Darwinian approaches to the origin of feath-ers, exemplified by Bock (1965), have hypo-thesized a microevolutionary and functionalcontinuum between feathers and a hypothe-sized antecedent structure (usually an elon-gate scale). Feathers, however, are hierarch-ically complex assemblages of numerousevolutionary novelties—the feather follicle,tubular feather germ, feather branched struc-ture, interacting differentiated barbules—thathave no homolog in any antecedent structures(Brush 1993, 1996, 2000; Prum 1999). Genu-ine evolutionary novelties are distinct fromsimple microevolutionary changes in that theyare qualitatively or categorically different fromany antecedent or homonomous structure(Nitecki 1990; Muller and Wagner 1991; Raff1996). Consequently, Wagner (2000; Wagneret al. 2000; Chiu and Wagner 2001) has arguedthat macroevolutionary research on homologyand the origins of evolutionary noveltiesshould ask different questions that arefocused on uncovering the mechanisms thatgenerate morphological novelties. Tradi-tional neo-Darwinian approaches to the ori-gin of feathers have focused on creatingtheoretical continuity with antecedent struc-tures, and as a consequence, few of these the-ories have adequately appreciated the manynovel aspects of feather morphology andfeather development, and none have formu-lated adequately detailed hypotheses aboutthe origin and evolution of these morpholog-ical and developmental novelties. In contrastto neo-Darwinian approaches, and in congru-
ence with a macroevolutionary concept ofnovelty, the developmental theory of featherorigins is focused specifically on reconstruct-ing the transition of developmental noveltiesrequired for the origin and diversification offeathers (Prum 1999).
Another conceptual problem has been thetendency to propose complex evolutionaryscenarios as intellectual “package deals” thatinclude correlated and interdependenthypotheses about the origins of birds, avianflight, and feathers. One package featuresbirds as an early nondinosaurian lineage ofarchosaurs, the arboreal theory of the originof flight, and the aerodynamic theory of theorigin of feathers (e.g., Feduccia 1999). Analternative package offers birds as a lineageof theropod dinosaurs, the cursorial theory ofthe origin of flight, and the thermal insula-tion theory of the origin of feathers (e.g.,Ostrom 1974). These two packages have beenpromoted inaccurately as reflecting “ornitho-logical” and “paleontological” schools ofthought, respectively (e.g., Feduccia 1999).Clearly, the solutions to these complex ques-tions are ultimately interrelated (i.e., there isonly one history of life). But the fundamentalproblem with these combined scenarios is thatthey conflate the analysis of these complexissues and eliminate many plausible combina-tions. These macroevolutionary questions canonly be productively and rigorously pursuedindependent from one another. By approach-ing the questions of phylogenetic relationshipsand evolutionary functional morphology inde-pendently, emergent historical patterns canbe used to test hypotheses of morphologicalhomology and evolutionary process (Lauderand Liem 1989; Larson and Losos 1996). Per-haps unsurprisingly, the recent theoretical
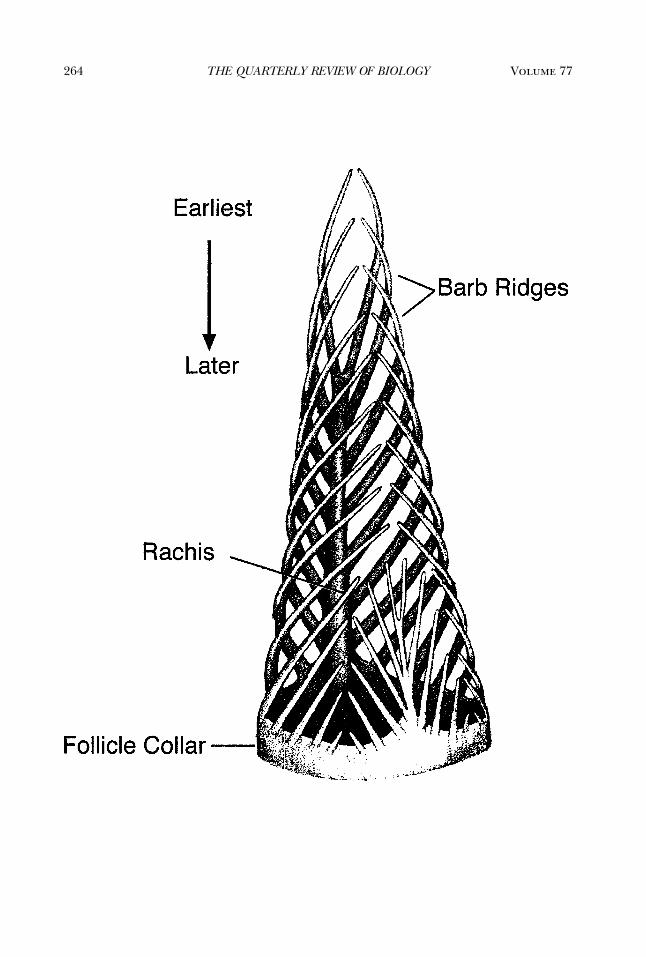
Figure 2. Schematic Diagram of Helical Growth of Barb Ridges of a Pennaceous FeatherThe branched structure of the barbs and the rachis of a feather form by helical growth and fusion of barb
ridges within the tubular feather germ. Feathers grow from the base. Barb ridges form at the new barb locuson the posterior midline of the collar and grow helically around the collar toward the anterior midline wherethey fuse to form the rachis ridge. Subsequent barb ridges fuse to the rachis ridge. In feathers with an after-feather, the new barb locus divides into two laterally displaced new barb loci. Subsequently, new barb ridgesgrow helically both anteriorly to the main rachis and posteriorly to form the hyporachis and vane of theafterfeather. The main vane and the afterfeather form separate vanes united within a single feather by thecalamus (Figure 1A). Pennaceous feathers obtain their planar form only after emerging from the cylindricalfeather sheath when growth is complete. The obverse (upper) and reverse (lower) surfaces of the vane developfrom the outer and inner surfaces of the cylindrical feather germ. Illustration based on Lucas and Stettenheim(1972).

266 Volume 77THE QUARTERLY REVIEW OF BIOLOGY
progress on the evolutionary origin of feathershas come from outside these entrenched sce-narios.
In this paper, we present a conceptualreview of the previous theories of the originof feathers and a synopsis of the recently pro-posed developmental theory of the origin offeathers (Prum 1999; Brush 2000). We reviewrecent paleontological evidence that supportsthe theropod origin of feathers prior to theorigin of birds or flight, and the congruencebetween these newly discovered theropodfeather morphologies and the morphologiespredicted by the developmental theory. Inlight of these findings, we discuss the homol-ogy of feathers and scales, plausible andimplausible functional theories of the originof feathers, feathers as evolutionary novelties,and the molecular basis of feather develop-ment and evolution.
Conceptual Review ofPrevious Theories
Previous theories of the origin of featherscan be categorized in terms of four concep-tual approaches: analyses of the “primitive”feathers of extant birds, functional theories,scale-to-feather transformation theories, andtheories based on the details of keratin bio-chemistry and keratinogenesis. These fourapproaches are not mutually exclusive, buthave often been combined in different waysto advocate a variety of hypotheses. Here wereview the conceptual bases for these theo-ries.
Many early predictions of the ancestralmorphology of feathers were based on anal-yses of the variation in extant feather mor-phology and its distribution among modernbirds (Dyck 1985). These attempts were unsuc-cessful for several reasons. First, researchersattempted comparative evolutionary analy-ses without historically explicit concepts ofmonophyly and phylogeny. This conceptualvacuum led to incongruous arguments thatthe feathers of specific extant groups repre-sent the primitive morphology of all avianfeathers. For example, ratites (Lowe 1935),penguins (Ewart 1921), and megapodes(Becker 1959) were each hypothesized atvarious times to be a basal lineage of birdsthat actually posses primitive feathers. These
authors did not understand explicitly thatan extant lineage within the monophyleticclade of modern birds could not be moreclosely related to the lineage in which feath-ers evolved than Archaeopteryx is, and that thephylogenetic distribution of feather varia-tion in extant bird clades could not be usedto identify primitive feather morphologies.Other researchers argued that the simplerstructure of certain extant feather types, com-monly either plumulaceous downs or openpennaceous contour feathers, supported thehypothesis that such morphologies were likelyto be primitive (Ewart 1921). These hypothe-ses were countered by evidence that some ofthese modern feathers show obviously derivedfeatures (Steiner 1917). For example, somedowns have differentiated distal and proximalbarbules that indicate that they may be sec-ondarily derived from pennaceous feathers inwhich the differentiated barbules intercon-nect to form a closed vane (Dyck 1985). Ofcourse, both arguments incorrectly imaginedthat the ancestral morphology of all featherscould be found within extant feather diversity.Structurally modern feathers have been pres-ent since at least Archaeopteryx, and they havecontinued to evolve, diversify, and simplify invarious ways. The presence of differentiatedbarbules in some modern downs may consti-tute evidence that they are secondarily derivedfrom pennaceous feathers, but this finding isirrelevant to the question of whether plumu-laceous or pennaceous structure was primi-tive to the initial radiation of feather mor-phology. Theories of the origin of feathersbased on nonphylogenetic analyses of extantfeather diversity have confounded the evolu-tion of feathers among extant birds with theinitial origin and diversification of featherspreceding Archaeopteryx.
A second conceptual current has been thedevelopment of functional theories of theevolutionary origin of feathers (reviewed inDyck 1985; Feduccia 1999). These theoriespropose plausible initial functions for ances-tral feathers, and then hypothesize a suitableancestral morphology to fulfill that function.Plausible initial functions have been justifiedbased on notions about the natural historyand ecology of ancestral birds, and notionsabout what functional transitions in mor-

September 2002 267EVOLUTION AND DIVERSIFICATION OF FEATHERS
phology are evolutionarily possible. Recentevidence of biologically convergent, analo-gous instances of the intermediate functionalstates proposed by the models have been usedto support the plausibility of specific func-tional theories (e.g., the hairy arms of Propi-thecus lemurs as an aerodynamic model ofearly feather evolution: Feduccia 1993). Thelist of functional theories of feather originsincludes the hypotheses that feathers evolvedfor flight (e.g., Steiner 1917; Heilmann 1926;Blaszyck 1935; Parkes 1966; Feduccia 1985,1993, 1999), thermal insulation (e.g., Davies1889; Ewart 1921), heat shielding (Regal1975), water repellency (Dyck 1985), com-munication (Mayr 1960), and tactile sensa-tion (Broman 1941).
The conceptual basis of functional theoriesof the origin of feathers is weak because thesetheories rest upon hypotheses about the func-tion of an ancestral structure whose mor-phology is unknown. Biologists can activelydebate the “functional significance” of somewell-known morphological structures fromextant organisms, yet the ability to makeinferences about the function of specific mor-phologies without direct experimental tests islimited. Functional theories of feather originspresume that current knowledge of the iden-tity and natural history of the lineage inwhich feathers initially evolved, and the evo-lutionary mechanisms that shape morphol-ogy and function, are sufficient to permit aconfident reconstruction of the ancestralmorphology of feathers. Given that it can bedifficult to understand the function of knownstructures, it is essentially impossible to con-fidently infer the ancestral function of feath-ers in absence of an independent hypothesison their form. Alternative functional theoriesof the origin of feathers have been justifiedby restating the initial functional specula-tions in absence of supporting evidencefrom the organism’s biology. Over the past20 years, it has been recognized that histori-cal analysis in evolutionary biology requiresan independent documentation of the pat-tern of evolutionary events before testingalternative functional hypotheses about theevolutionary process that explain those events(Lauder 1981; Lauder and Liem 1989;Lauder 1990; Larson and Losos 1996; Padian
2001). Based on nonphylogenetic, neo-Darwinian evolutionary theory (e.g., Bock1965), functional theories of the origin offeathers have failed because they attempt todo exactly the opposite: they use presumedknowledge of adaptive evolutionary processto reconstruct historical pattern.
An additional problem with functional the-ories of the origin of feathers is that func-tional criteria are explicitly incompatible withthe recognition of the homology of struc-tures. Even enthusiastic advocates of the con-cept of the “homology of functions” have rec-ognized that function cannot be used toelucidate structural homology (Lauder 1994;Hall 1999:205). Thus, if a primitive homologof modern feathers were discovered that didnot meet the criteria of a specific functionalmodel, its homology with modern featherswould be rejected based on functional criteriaalone rather than whether its structureshared some unique or special similaritiesthat reflect their historical relationship. Forexample, the homology between feathersand the integumentary appendages of Sino-sauropteryx may have been initially rejectedbecause they did not meet the expectationsof functional theories of the origin of feath-ers (Gibbons 1997; Feduccia 1999). Like-wise, an entirely analogous structure with aconvergent function could be proposed as afeather homolog based entirely on meetingsome of the functional criteria of a proposedtheory. For example, the proposed homol-ogy between feathers and the appendages ofLongisquama ( Jones et al. 2001) are basedentirely on functionally convergent aspects oftheir planar structure. A fundamental aspectof the concept of structural homology is thatit explicitly excludes similarities of function,and thus allows the recognition of homolo-gous structure that have evolved in form as aconsequence of an evolutionary change ortransference in function. Incorporating apriori functional criteria into models for theorigin of feathers has created fundamentalproblems for the field.
A third, important concept in many previ-ous theories of the origin of feathers has beenthe hypothesis that feathers evolved fromscales (reviewed in Lucas and Stettenheim1972). Numerous ancestral feather morphol-

268 Volume 77THE QUARTERLY REVIEW OF BIOLOGY
ogies have been proposed as hypotheticalintermediates between planar reptilian scalesand feathers. These scale-based scenarioshave been incorporated into various func-tional theories, usually in the form of an elon-gate, planar scale (e.g., Heilmann 1926;Parkes 1966; Maderson 1972; Regal 1975;Dyck 1985; Feduccia 1999; Maderson and Ali-bardi 2000). Scale-based hypotheses of theorigin of feathers create a developmental andtopological conundrum that was first pointedout by Davies (1889) and still has not beenadequately addressed by any elongate-scale-to-feather transformation theory. Scales areinherently laminar or planar folds of the skin,whereas feathers are essentially tubular epi-dermal appendages (Prum 1999). A planarpennaceous feather obtains its planar shapeonly after emergence from the cylindricalsheath. The obverse (outer) and reverse(under) surfaces of a pennaceous feathervane develop from the outer and inner sur-faces of the tubular epidermal feather germ,respectively (Figure 2). In contrast, the dorsaland ventral surfaces of a planar scale developfrom the dorsal and ventral surfaces of theoriginal epidermal outgrowth. Thus, the pla-nar surfaces of scales and feathers are nothomologous (Davies 1889; Prum 1999). Oneplanar form cannot be evolutionarily trans-formed into the other while maintaining acontinuously functional planar configuration(Prum 1999). Steiner (1917) attempted toreconcile this contradiction by homologiz-ing the ventral surface of a scale with theafterfeather (Lucas and Stettenheim 1972).This hypothesis, however, is not congruentwith how the afterfeather develops (Lillieand Juhn 1938; Lucas and Stettenheim1972). No other scale-based model for theorigin of feathers has even attempted toresolve this fundamental problem (Heil-mann 1926; Parkes 1966; Maderson 1972;Regal 1975; Dyck 1985; Feduccia 1999; Mad-erson and Alibardi 2000).
Scale-based theories of the origin of feath-ers have also been justified by the generalpremise that pennaceous feathers, with barbsand a rachis in a planar form, constitute theprimitive feather morphology. This conclu-sion has been supported by the observationthat many plumulaceous (downy) feathers of
extant birds have derived structures (Steiner1917; Parkes 1966; Dyck 1985; Feduccia1999). As described above, this conclusion isbased on faulty phylogenetic reasoning, andis contradicted by the more general tubularnature of feathers (see below).
Scale-based models of the origin of feathershave not adequately recognized the suite ofunique evolutionary novelties that character-ize feather structure and development, norhave they attempted to explain their evolu-tionary origins. Any comprehensive theoryof how feathers evolved should also explainthe evolution of how feathers grow (Prum1999). Scale-based models have failed to pro-pose a cogent explanation for the origin ofthe follicle itself. Maderson and Alibardi(2000) accurately describe the function ofthe feather follicle in feather development,but they suggest no mechanism by which itmight have evolved from a scale or whatappendage morphology might grow fromany such intermediate. Furthermore, scale-based models have repeatedly hypothesizedthe evolution of the branched structure offeathers as a process of the division of a pla-nar scale (Heilmann 1926; Regal 1975; Dyck1985; Maderson and Alibardi 2000). Like theproposed homology of the planar surfaces offeathers and scales, these scenarios areincongruent with the basic details of howfeathers grow (Lucas and Stettenheim 1972;Prum 1999). Barb ridges form by the prolif-eration and outfolding of the basal layer ofthe feather germ epithelium, creating thelongitudinal epidermal compartments thatbecome the barbs (Lucas and Stettenheim1972). The proliferation of barb ridges on theventral side of a feather follicle during thegrowth of a pennaceous vane demonstratesclearly that barb ridges do not form by thedivision of a plane but by the proliferation,differentiation, and compartmentalization ofepidermal tissue within the follicle collar(Strong 1902b; Lucas and Stettenheim 1972).In addition, the role of the barb ridge fusionin the creation of the rachis and hyporachisridges during development further demon-strates that the rachis ridge is not homolo-gous with a thickened medial axis of a scale(Lillie and Juhn 1938; Lillie and Wang 1941;Lucas and Stettenheim 1972; Prum 1999).

September 2002 269EVOLUTION AND DIVERSIFICATION OF FEATHERS
Scale-based models of the origin of feathershave proposed only the vaguest scenarios forthe evolution of the complex structure ofbarbs and barbules (Regal 1975). Dyck (1985)made the most explicit attempt but concluded,“When and how the interlocking system of thebarbules formed is difficult to suggest.” Theabsence of these crucial details in scale-basedmodels has frequently been covered with anunsatisfying caveat that feathers did not actu-ally evolve directly from scales (Maderson1972; Regal 1975; Maderson and Alibardi2000). Yet these same authors still maintainthat pennaceous feathers evolved from theelongate, planar, scale-like structure—which isthe source of these conceptual problems.
Recently, Zhang and Zhou (2000) pro-posed that the thick, flat rachi of the elon-gate central rectrices of Protopteryx fengnin-gensis, an Early Cretaceous enantiornithinebird, provide support for a scale-like originof feathers. This hypothesis has two substan-tial problems. First, similar “scale-like” feath-ers occur in living birds, such as penguins(Spheniscidae) and tropicbirds (Phaethon-tidae). These enlarged rachi are formed bythe fusion of barbs and are only superficiallyscale-like. Nearly identical rectrices are alsopresent in the Early Cretaceous confuciusor-nithine birds that are phylogenetically evenmore basal than Protopteryx, but in Confuciu-sornis these feathers are distally branchedjust like the analogous feathers of modernbirds (Chiappe et al. 1999). Unfortunately,the tips of the remiges of the only specimenof Protopteryx are missing, so these criticaldata are unavailable. A second, more criticalproblem with this hypothesis is that Protop-teryx and other enantiornithines are not themost basal birds in avian phylogeny (see Fig-ure 6; Chiappe et al. 1999). Because feathersmust have evolved in a lineage prior to thedifferentiation of Archaeopteryx from otherbirds, the rectrices of the enantiornithineProtopteryx must be secondarily derived. Likevariations in feather morphology amongmodern birds, these fascinating early birdfeathers document the diverse developmen-tal capabilities of ancient feather follicles,but they do not constitute retained plesio-morphic morphologies. Given its phyloge-netic position and the equivocal nature of
the evidence, the rectrices of Protopteryx donot provide evidence of a scale-like origin offeathers.
A fourth approach has been to use detailsof patterns of keratinization in reptiles andbirds to support hypotheses of the origin offeathers. Maderson (1972) and Madersonand Alibardi (2000) have proposed that thesimilarity in vertical alternation of � and b-keratins in feathers and lepidosaur scales jus-tified the use of the lepidosaurian scale as a“model system” for the evolution of feathers.Maderson (1972) made no proposals aboutthe origin of the branched structure of feath-ers, but Maderson and Alibardi (2000) pro-pose that branched feathers evolved throughthe subdivision of an elongate scale into barbsand barbules, as proposed by Regal (1975)and others (reviewed in Dyck 1985). Thisapproach is conceptually superior to purelyfunctional theories because it is based onobservable and testable details of integumen-tary biology and development. Unfortunately,the patterns of �-keratin and b-keratinexpression in the reptilian dermis lead to fewdetailed predictions about the origin of feath-ers and evolution of feather morphogenesisthat are not merely consequences of theassumption that feathers are broadly homol-ogous with scales (including their planar sur-faces), that feathers evolved directly from anelongate scale, and that pennaceous feathersare primitive. Thus, these models have all thepreviously discussed problems inherent instrictly scale-based theories. Furthermore, theproposed relationship between vertical alter-nation of keratin in lepidosaur scales and birdfeathers is based on a misconception of phy-logenetic relationships of birds and reptiles.Birds and crocodylians are archosaurs. Thescales of both crocodylians and birds lack thevertical stratification of �-keratin and b-keratinexpression in the epidermis, and the shed-ding or molt associated with it in both lepi-dosaurs and bird feathers (Maderson andAlibardi 2000). It is not parsimonious to con-clude that molt in lepidosaur scales and birdfeathers is somehow homologous when it isabsent from crocodylian and avian scales. Ifthe vertical alternation of keratin expressionin lepidosaur scales and feathers is evolution-arily convergent, it remains to be demon-

270 Volume 77THE QUARTERLY REVIEW OF BIOLOGY
strated how any analogous aspects of lepido-saur scales can function as an appropriate“model system” for the study of the origin offeathers, as proposed by Maderson and Ali-bardi (2000). The evolution of vertical alter-nation of keratin expression in feathers ulti-mately requires explanation. There is noreason to conclude, however, that the pat-terns of �-keratin expression in lepidosaurscales, crocodilian and avian scale-hingeregions, and the feather sheath are not prox-imate adaptations for their current physicalfunctions.
Brush (1993, 1996) provided an alterna-tive “keratin-based” theory of the origin offeathers. He reviewed the origin of the novelkeratins found in feathers, and the evidenceand theories about their evolution, and con-cluded that feathers are complex hierarch-ical novelties that incorporate numerousunique molecular and developmental fea-tures. Brush (1993, 1996) also directly ques-tioned the homology of scales and feathers,and pointed out the essential novelties of themany details of feather biochemistry anddevelopment. Although he did not proposean explicit scenario for the origin of thecomplex branched structure of feathers,these discussions were based on testabledetails of feather development and antici-pated subsequent models based on featherdevelopment.
A Developmental Theory of theOrigin of Feathers
We have recently proposed an alternativeapproach to the origin of feathers that uses thedetails of feather development to reconstructplausible antecedent morphologies (Prum1999; Brush 2000, 2001). This approach sug-gests that any theory of the origin of feathersshould also provide a complete and consistenttheory of the origin of the complex mecha-nisms of feather development. Furthermore,the details of feather development supportan hypothesis of feather evolution that isindependent of phylogenetic and functionalassumptions. We have each independentlyproposed developmental theories of the ori-gin of feathers. Prum (1999) analyzed thecomplex, hierarchical mechanisms by whichfeathers grow, and hypothesized a transition
series of developmental novelties from thefirst feathers through modern feather mor-phological diversity. Brush (1993, 1996, 2000,2001) used feather biochemistry and the hier-archy of feather development and structureto define the conditions for recognizing theoriginal “protofeather,” and to propose a“phylogeny” of feather types. The implica-tions of these two independent approachesare extensively congruent, and represent aconceptually new direction in the study of theorigin of feathers.
The details of feather development pro-vide a rich source of information aboutfeather biology (reviewed in Lucas and Stet-tenheim 1972, summarized in Prum 1999).Feathers are branched, filamentous keratinstructures uniquely characterized by a tubu-lar follicle which forms by a cylindrical invag-ination around the elongate feather papillaor short bud (Figure 3). Rather than growingfrom bifurcating tips like a plant, feathersform a branched structure from the baseusing two unique mechanisms. The branch-ing of the barbs and rachis is a consequenceof the helical growth of barb ridges aroundthe tubular epidermal cylinder of the featherfollicle and feather germ, and the fusion ofthe barb ridges to the rachis ridge on theanterior side of the follicle (Figure 2). In con-trast, the branched structure of the barbulesand barb rami involves the differentiation ofkeratinocytes within the peripheral barbuleplates of the developing barb ridges into apaired series of cells that connect basally tothe barb ramus.
By focusing on the evolution of the mech-anisms of feather development, Prum (1999)proposed a detailed, testable model of theevolutionary origin of feathers that is in-dependent of functional or phylogeneticassumptions. The model proposed a five-stage transition series in the history offeather diversity as a hypothesized sequenceof novelties in feather development (Figure4). The model hypothesizes that the firstfeather (Stage I) originated with the firstfeather follicle—the cylindrical epidermalinvagination around the initial feather papilla.Subsequent feather diversity evolved througha series of derived developmental noveltieswithin the tubular intermediate epidermal

September 2002 271EVOLUTION AND DIVERSIFICATION OF FEATHERS
layer of the follicle, called the follicle collar,which generates the tubular feather germ.After the origin of the follicle came the differ-entiation of the follicle collar into barb ridgesthat generate the barbs (Stage II). The modelproposes two alternative stages next—the ori-
gin of helical growth (Stage IIIa), or the originof barbule plate differentiation (Stage IIIb).The model cannot differentiate between thetwo alternative orders for these events (i.e.,IIIa before IIIb, or IIIb before IIIa), but fol-lowing the evolution of both of these devel-opmental novelties came the capacity to growboth kinds of branched structure typical ofmodern feathers (Stage IIIa�b). The originof differentiated distal and proximal barbuleplates followed next (Stage IV). Finally, addi-tional developmental mechanisms evolvedand created further diversity in feather struc-ture (Stage Va–f).
This hypothesized series of five develop-mental novelties predicts an explicit transitionseries in the morphologies of the feathersgrown from these follicles (Figure 5; Prum1999). Stage I follicles would produce anunbranched, hollow, tubular feather. Themodel predicts the keratinaceous compositionand essentially tubular geometry of the prim-itive feather, but does not predict the shape,size, stiffness, or other structural qualities. AStage II follicle would grow a tuft of barbsfused basally to a single calamus. A Stage IIIafollicle would grow a feather with a rachis(formed by the initial fusion of feather barbson the anterior side of the follicle) and a seriesof fused barbs. Stage IIIb follicles would pro-duce a tuft of barbs with branched barbules.Stage IIIa�b follicles would grow the firstbipinnate (double-branched) feathers with arachis, barbs, and barbules. In the absence ofdifferentiated barbules, a Stage IIIa�b featherwould be open pennaceous. Stage IV follicleswould grow a pennaceous feather with aclosed, coherent vane created by the interac-tions of the differentiated hooks and grooveson the distal and proximal barbules ofneighboring barbs. Only after Stage IV couldsubsequent novelties in Stages Va–f yieldadditional feather diversity, including anasymmetrical vane (Stage Va) and the after-shaft (Stage Vb).
The justification for the order of the evo-lution of the developmental novelties in themodel comes from the observed causal hier-archy within feather growth mechanisms(Prum 1999). For example, a feather with fil-amentous barbs (Stage II) is hypothesized tohave evolved before the origin of the rachis
Figure 3. Schematic Diagram of the InitialDevelopment of a Feather Follicle
(A) Development of the epidermal feather placodeand dermal condensation. (B) Development of a shortbud or feather papilla. (C) Formation of the featherfollicle through the invagination of a cylinder of epi-dermal tissue around the base of the feather papilla.(D) Cross section of the feather follicle through thehorizontal plane indicated by the dotted line in C. Theinvaginated tubular feather follicle is characterized bya series of tissue layers (from peripheral to central):the dermis of the follicle, the epidermis of the follicle(outer epidermal layer), the follicle cavity or lumen(the space between epidermal layers), the follicle col-lar (inner epidermal layer or ramogenic zone), andthe dermal pulp (tissue at the center of the follicle).The tubular feather germ grows by proliferation anddifferentiation of keratinocytes in the follicle collar.Summarized from Lucas and Stettenheim (1972) andPrum (1999).
272 Volume 77THE QUARTERLY REVIEW OF BIOLOGY

September 2002 273EVOLUTION AND DIVERSIFICATION OF FEATHERS
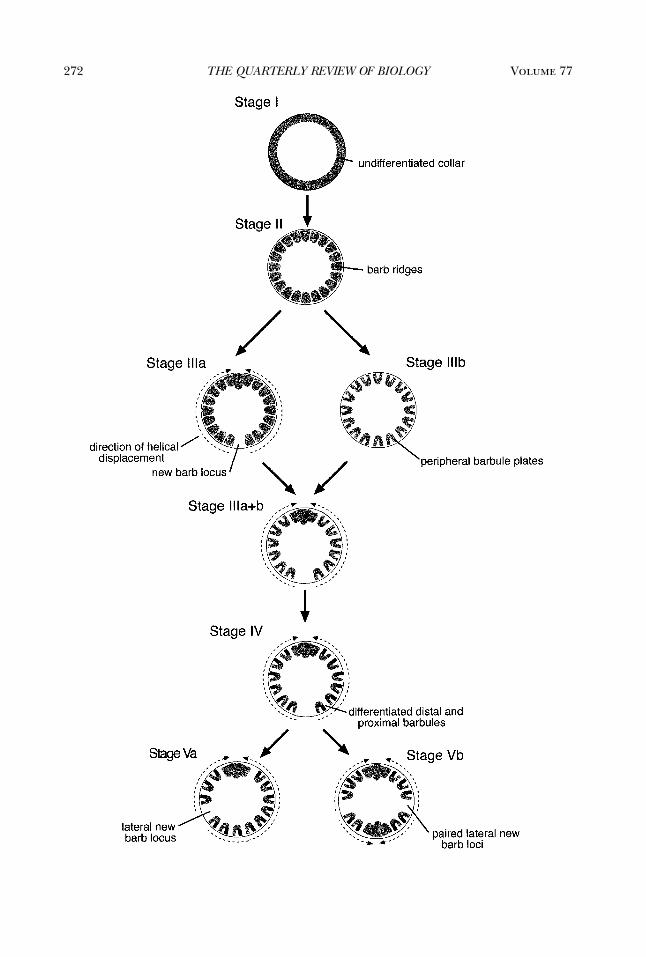
Figure 4. Developmental Model of the Origin and Diversification of Feather FolliclesAn hypothesized transition series of evolutionary novelties in feather development, depicted as a series of
cross sections of the follicle collar—the innermost layer of epidermal tissue in the feather follicle that generatesor develops into the feather (Figure 3D) from Prum (1999). The model is based entirely on the hierarchicaldetails of feather development, and is independent of functional or phylogenetic assumptions. Each diagramis oriented with the anterior surface of the follicle collar upward. The developmental novelties are labeled inthe stages at which they originate. Stage I—Origin of the undifferentiated collar through a cylindrical epidermalinvagination around the base of the feather papilla. Stage II—Origin of the differentiation of the inner layerof the collar into longitudinal barb ridges and the peripheral layer of the collar into the feather sheath. StageIII—Either of these developmental novelties could have occurred first, but both are required before Stage IV.Stage IIIa—Origin of helical displacement of barb ridges and the new barb locus. Stage IIIb—Origin of pairedbarbules from peripheral barb plates within the barb ridges. Stages IIIa�IIIb—Origin of follicle capable ofboth helical displacement and barbule plate differentiation. Stage IV—Origin of differentiated distal and prox-imal barbules within barbule plates of barb ridges. Stage Va—Origin of lateral displacement of the new barbridge locus. Stage Vb—Origin of the division of posterior new barb locus into a pair of laterally displaced loci,and opposing anterior and posterior helical displacement of barb ridges toward the main feather and after-feather of the follicle. See Prum (1999) for details of additional stages in the evolution of feather diversity(Stages Vc–f).
(Stage IIIa) because the rachis of a penna-ceous feather is initially created, or specified,by the fusion of barb ridges (Lillie and Wang1941; Lucas and Stettenheim 1972; Prum1999). Likewise, barbs (Stage II) are hypoth-esized to evolve before barbules (Stage IIIb)because barbules develop within layers ofthe preexisting barb ridges (Prum 1999). Abipinnate feather with a rachis, barbs, andbarbules (Stage IIIa�b) is hypothesized tohave evolved before the differentiated bar-bules (Stage IV) because prior to the originof the rachis, barbules could not have haddistal or proximal orientation relative to thefeather vane. Prum (1999) describes devel-opmental justifications for additional stages.
Although Prum’s (1999) model is based ondevelopmental evidence, it is explicitly notbased on Haeckelian ontogenetic recapitula-tion or von Baer’s Law—the notion that onto-genetically antecedent events are phylogenet-ically primitive. Several stages of the modelare congruent with a recapitulationist sce-nario, but they are not justified on that basis(Prum 1999). Thus, plumulaceous feathersare hypothesized to be primitive to penna-ceous feathers not because the first feathersof extant birds are typically plumulaceous,but because the simplest differentiated folli-cle collar would have grown a plumulaceousfeather (Prum 1999). There are also severalstages of the model that directly conflict withvon Baer’s Law (Prum 1999). For example, inthe development of the first feathers in an
avian embryo, differentiated barb ridgesoriginate in the inner epidermal layer of thefeather germ during the short bud stagebefore the origin of the follicle, whereas inthe model the follicle originates (Stage I)before the barb ridges (Stage II). The initi-ation of barb ridges before the follicle mustbe a derived alteration of the primitiveontogeny, however, because the barbs wouldbe unable to continue to grow without theorganization provided by the follicle. In asecond example, the barbules differentiateand keratinize before the ramus within anyhorizontal section of a barb ridge. A featherwith keratinized barbules that lacked barbrami would simply disintegrate, however. Bar-bules keratinize before the ramus becausethey are peripheral to the ramus within thebarb ridge, and a fully keratinized ramuswould block the diffusion of necessary nutri-ents from the central dermal pulp out to thedeveloping barbule keratinocytes.
Important support for the plausibility ofthe developmental model of the evolutionaryhistory of feathers comes from extant featherdiversity (Prum 1999). All the primitivefeather morphologies proposed by the modelexist among extant avian feathers. For exam-ple, the basic structure of the Stage I featheris present in every known feather in the tubu-lar cylindrical calamus at its base and thecylindrical feather sheath that surrounds it.Specialized tubular feathers that are identicalto the predictions of Stage I grow from the
274 Volume 77THE QUARTERLY REVIEW OF BIOLOGY

September 2002 275EVOLUTION AND DIVERSIFICATION OF FEATHERS
central crown patch of male African peafowl(Afropavo congensis, Phasianidae) and fromthe entire crown of the Bornean bristlehead(Pityriasis gymnocephala, Corvoidea). Addi-tional modern examples of morphologieslike Stages II and III can be found among vari-ous extant avian plumages (Prum 1999). Forexample, certain display plumes in egrets(Egretta, Ardeidae) and birds of paradise (Par-adisaea, Paradisaeidae) correspond well toStage II morphology. Other open penna-ceous contour feathers and semiplumes arecongruent with the predicted Stage IIIa�bmorphology. These modern feathers dem-onstrate that the developmental configura-tions and morphologies predicted by themodel are among the known products offeather follicles. Thus we know that featherfollicles can function in the ways proposed bythe model. These extant feathers are second-arily derived or simplified, however, and theirexistence does not constitute evidence thatprimitive feathers actually had these pre-dicted morphologies. For these inferences,paleontological data are required to docu-ment plesiomorphic feather morphologies(see below).
Brush’s (2000, 2001) approach to the ori-gin of feathers is similar to Prum’s (1999) inthat it also draws on observable details offeather development from extant birds. Afterrecognizing their distinctness from scales,Brush (2000, 2001) hypothesized that feath-ers are essentially cylindrical structures, and
that the follicle and feather keratins are thefundamental feather novelties. Brush (2000,2001) then hypothesized that the first “proto-feather” was an elongate tubular filament. Hereferred to this initial tubular structure as“barb-like,” and envisioned the evolution ofbranched feather structure as the duplicationof these barb-like structures. This hypothesisconfuses the identities of an entire featherand an individual barb. A simple tubularfeather would be homologous with an entiremodern feather, the whole product of growthfrom a follicle. The hollow center of the firstfeather is a consequence of the tubular orga-nization of the follicle and comes from thedermis at the center of the follicle andfeather germ. In contrast, barbs are differ-entiated portions of the tubular feather germ.The evolution of a branched feather mor-phology came not from the duplication of aninitial tubular structure, but from the differ-entiation, proliferation, and organization ofcells within this epidermal tube into barbridges.
The historical transition series of Brush’s(2000) feather phylogram is extensively con-gruent with Prum’s (1999) hypothesis. Prum,however, provided a more explicit develop-mental justification of the stages, and also anexplicit distinction between the evolution ofinitial structural feather diversity and theclasses of extant feathers, many of which arelikely to have evolved through secondary sim-plification of more complex forms.
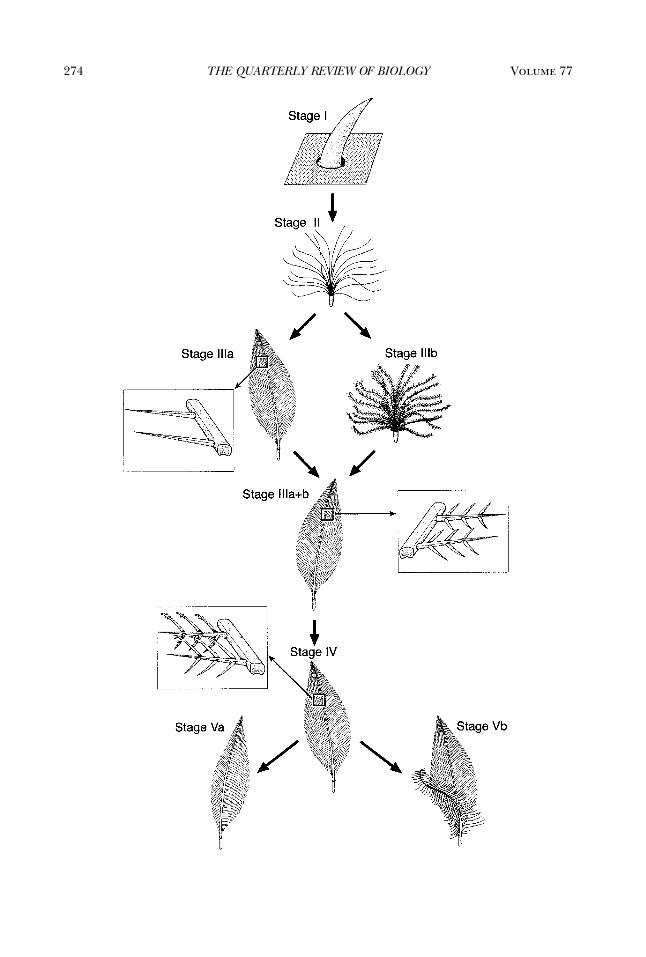
Figure 5. Developmental Model of the Origin and Diversification of FeathersA predicted transition series of feather follicles based on the hypothesized series of evolutionary novelties in
feather developmental mechanisms (Figure 4) from Prum (1999). Stage I—Origin of an undifferentiated tubu-lar collar yields the first feather, a hollow cylinder. Stage II—Origin of a collar with differentiated barb ridgesresults in a mature feather with a tuft of unbranched barbs and a basal calamus emerging from a superficialsheath. Stage IIIa—Origin of helical displacement of barb ridges and the new barb locus results in a pinnatefeather with an indeterminate number of unbranched barbs fused to a central rachis. Stage IIIb—Origin ofperipheral barbule plates within barb ridges yields a feather with numerous branched barbs attached to a basalcalamus. Stages IIIa�IIIb—Origin of a feather with both a rachis and barbs with barbules creates a bipinnate,open pennaceous structure. Stage IV—Origin of differentiated proximal and distal barbules creates the firstclosed pennaceous vane. Distal barbules grew terminally hooked pennulae to attach to the simpler, groovedproximal barbules of the adjacent barb (Figure 1B). Stage Va—Lateral displacement of the new barb locusleads to the growth of a closed pennaceous feather with an asymmetrical vane resembling modern rectricesand remiges. Stage Vb—Division and lateral displacement of the new barb loci yields opposing, anteriorly andposteriorly oriented patterns of helical displacement, producing a main feather and an afterfeather with asingle calamus. The afterfeather could have evolved at any time following Stage IIIb, but likely occurred afterStage IV based on modern aftershaft morphology. See Prum (1999) for details of additional stages in theevolution of feather diversity.

276 Volume 77THE QUARTERLY REVIEW OF BIOLOGY
Paleontological Discovery ofNonavian Dinosaur Feathers
Over the last ten years, the discovery anddescription of the paleofauna from the EarlyCretaceous of the Yixian Formation of Liao-ning Province, northeastern China, have con-tributed numerous breakthroughs in verte-brate paleontology (Stokstad 2001a). Amongthe most exciting and revolutionary findshave been previously unknown radiation ofearly birds and small theropod dinosaurs.The exceptional taphonomic conditions pre-served integumentary structures of many ofthese birds and some nonavian theropoddinosaur specimens. These recently discov-ered theropod integumentary structures pro-vide the first new morphological evidenceabout the origin and evolution of featherssince the description of Archaeopteryx in 1861.Recent analyses have confirmed the diversityof their structure, and have documented sev-eral novel morphological features that theyshare uniquely with avian feathers ( Ji et al.2001; Xu et al. 2001). In combination with theindependent data supporting birds as a line-age of theropod dinosaurs (Gauthier 1986;Holtz 1994; Sereno 1997; Padian and Chiappe1998; Sereno 1999), the new paleontologicalevidence strongly supports the conclusion thatthe novel integumentary structures of thesetheropods are homologous with feathers andconstitute primitive feather morphologies ( Jiet al. 2001; Xu et al. 2001). Furthermore, themorphologies of these dinosaur feathers arecongruent with the predicted stages of thedevelopmental model of the origin of feathers( Ji et al. 2001; Sues 2001; Xu et al. 2001).
The first nonavian theropod described fromthe Yixian Formation was Sinosauropteryx,which was described by Chen et al. (1998) ashaving small (5–6 mm) “protofeathers” alongits back and on its face and belly. Chen et al.(1998) presented a phylogenetic analysis thatindicated that Sinosauropteryx was a basal coe-lurosaur, implying a deeply dinosaurian originfor feathers. Though the homology of thesestructures with feathers was initially rejected bysome (Gibbons 1997; Feduccia 1999), thestructures are cylindrical and emerge fromthe skin in a manner that is nearly unique tofeathers and thus are likely to have grownfrom cylindrical follicles (A H Brush and R O
Prum, personal observation). The append-ages of Sinosauropteryx may have been diverse:some appendages are unbranched, but oth-ers may have been a simple tube with abranched distal tuft (Brush and Prum, per-sonal observation).
Ji et al. (1998) later described two thero-pods with fully pennaceous feathers on theforelimbs and tail—Caudipteryx and Protar-chaeopteryx—and declared them “feathereddinosaurs.” The obvious rachis and herring-bone pattern of barbs have been universallyaccepted as vaned feathers. The phylogeneticanalysis in the original description includedonly a single additional nonavian dinosaur(Velociraptor), however, and Caudipteryx andProtarchaeopteryx were placed as the successivesister groups to birds ( Ji et al. 1998). Thistopology left the possibility open that Caudip-teryx and Protarchaeopteryx could be secondar-ily flightless birds. Accepting the evidence offeathers, critics of the dinosaurian origin ofbirds claimed that this was the case (Feduccia1999; Jones et al. 2000b; Martin and Czerkas2000). Subsequently, independent phyloge-netic analyses have placed Caudipteryx withinthe oviraptoroid theropods (Sereno 1999;Zhou 2000; Zhou and Wang 2000; Zhou et al.2000; Holtz 2001; Norell et al. 2001).
Filamentous integumental appendageshave also been described from the head ofthe alvarezsaurid Shuvuuia deserti from a LateCretaceous deposit in the Gobi Desert ofMongolia (Schweitzer et al. 1999; Schweitzer2001). Schweitzer et al. (1999; Schweitzer2001) used immunological methods and elec-tron microscopy to infer that the filamentousstructures preserved on the head of Shu-vuuia were composed of b-keratin. The alvar-ezsaurids, including Mononychus and Shu-vuuia, have been alternatively hypothesizedto be an early lineage of birds (Chiappe etal. 1996, 1998), or the sister group to theornithomimid theropods (Sereno 1999,2001). The results of Schweitzer et al. (1999)support the hypothesis that the filamentousstructures from Shuvuuia are integumentaryappendages, since b-keratins are onlyexpressed in the integument of reptiles(including birds). Regardless of whether thealvarezsaurids are an early avian lineage orare most closely related to ornithomimids,

September 2002 277EVOLUTION AND DIVERSIFICATION OF FEATHERS
these data substantiate the conclusion thatthe filamentous structures are homologouswith feathers. The initial placement of thealvarezsaurids within birds may have been anartifact of the relatively small samples of non-avian theropods included in the analyses(e.g., Chiappe et al. 1996, 1998). We preferthe hypothesis that is based on more inclusivephylogenetic analyses that places alvarezsaur-ids with the ornithomimids. Although theseanalyses are new and need to be repeated,Schweitzer et al. (1999; Schweitzer 2001) pro-vide unique biochemical, immunologicaldata in support of the origin of feathers intheropod dinosaurs.
Since the description of Caudipteryx andProtarchaeopteryx, filamentous integumentarystructures have been discovered on addi-tional nonavian theropod specimens fromthe Yixian Formation, including long fila-mentous appendages from the back of theulna of a large basal therizinosaur, Beipiao-saurus (Xu et al. 1999a), and covering largeportions of the body of three basal dromaeo-saurs—Sinornithosaurus (Xu et al. 1999b),Microraptor (Xu et al. 2000), and two unknowntaxa closely related to Sinornithosaurus ( Ji et al.2001; Norell et al. 2002). The integumentaryappendages of these taxa consist of filamen-tous structures between 25–190 mm long.
The original descriptions of Sinosauropte-ryx, Sinornithosaurus, and Beipiaosaurus didnot describe these integumentary append-ages in substantial detail or justify explicitly inwhat way they could be homologous withfeathers. With the controversy over the phy-logenetic relationships of the undoubtedlyfeathered Caudipteryx and Protarchaeopteryx(Dalton 2000), confirmation of the preaviantheropod origin of feathers required demon-stration of feather-like structure in the integ-umentary appendages of clearly nonavian the-ropods.
This critical evidence has come from theintegumentary structures of two Liaoningtheropods classified as basal dromaeosaurs.In the original description of Microraptor, Xuet al. (2000) proposed explicitly that theappendages of Microraptor were branched instructure like a rachis and barbs. Subse-quently, Xu et al. (2001) presented an analysisof the integumentary appendages of the type
specimen of Sinornithosaurus that stronglysupported their homology with feathers.They documented that these integumentaryappendages varied in structure over differentparts of the body. Furthermore, the Sinorni-thosaurus integumentary appendages werecomposed of multiple filaments and exhibittwo forms of branched structure: a basal tuftand serial branching off a central shaft. Xuet al. (2001) concluded that Sinornithosaurusprovided little direct evidence for the pres-ence of a rachis. They inferred the presenceof a rachis in one appendage which continuedto add new filaments throughout its lengthwithout increasing in width. Among all knownvertebrate integumentary appendages, thethree features documented in the append-ages of Sinornithosaurus—multiple filaments,basal branching, and serial branching—areunique to avian feathers (Lucas and Stetten-heim 1972; Bereiter-Hahn et al. 1986; Xu etal. 2001). No other vertebrate integumentaryappendages are composed of multiple fila-mentous structures. The hair of chinchillas(Chinchillidae) appears tufted, but actuallygrows from multiple independent roots thatdevelop within a single follicle (Nowak andParadiso 1983; Grau 1994). Furthermore,the basally tufted and the serial branchedmorphologies of Sinornithosaurus are identi-cal in general structure to certain aviandowns and to barbuleless pennaceous feath-ers. These uniquely shared morphologicalfeatures constitute the type of special simi-larity and correspondence of parts thatcharacterize morphological homologs (Pat-terson 1982; Pinna 1991), and their identi-fication provides strong confirmation of thehomology of these theropod integumentarystructures and avian feathers. In combina-tion with the independent phylogeneticevidence that dromaeosaurs and birds aresister groups, the homology with feathers isstrongly supported.
Ji et al. (2001) later described the mor-phology and distribution of the integumen-tary appendages of a basal dromaeosaur thatis probably referable to Sinornithosaurus andis indisputably nonavian. This extraordinaryspecimen exhibits abundant filamentousintegumentary appendages over the entiresurface of its body except for the lower legs

278 Volume 77THE QUARTERLY REVIEW OF BIOLOGY
and feet. As in Sinornithosaurus (Xu et al.2001), these appendages are diverse in mor-phology. The specimen has at least threetypes of feathers, including simple tufts,longer feathers with branches, and indisput-ably vaned feathers with an obvious herring-bone pattern of parallel filaments preservedin the same pattern as the barbs in apennaceous feather. Again, among all ver-tebrate integumentary appendages, thesethree classes of branched structures areunique to feathers.
Most recently, Norell et al. (2002) describedthe integumentary appendages of anotherspecimen of basal dromaeosaur from theYixian Formation that may be distinct fromSinornithosaurus. The specimen shows abun-dant integumentary appendages over thebody, but the appendages of the tail, legs,and wings are particularly well preserved.The specimen has 19 cm long appendagesthat extend from the tip of its tail and 13 cmlong appendages on the legs. Although mostof its integumentary appendages are not wellpreserved, the appendages of the legs andforelimbs clearly demonstrate the rachis andbarbs characteristic of a pennaceous vane. Ina few places, the appendage vanes exhibitgaps that indicate the presence of differen-tiated distal and proximal barbules thatnormally interlock to create the closed pen-naceous structure. This specimen providesdefinitive evidence of the presence of mod-ern feathers in a nonavian dinosaur.
These paleontological discoveries and phy-logenetic analyses essentially demonstrate theorigin of feathers in nonavian terrestrial the-ropod dinosaurs (Padian 1998, 2001; Sereno1999; Sues 2001; Prum 2002). Feathers origi-nated and diversified in structure within thecoelurosaurian theropod dinosaurs beforethe origin of birds or the origin of flight (Fig-ure 6). Integumental appendages that shareunique, derived features with avian feathersare now known from eight different nonaviantheropod taxa: Sinosauropteryx, Shuvuuia, Bei-piaosaurus, Caudipteryx, Protarchaeopteryx, Sinor-nithosaurus, Microraptor, and two unnamedbasal dromaeosaurs from the Yixian Forma-tion. All but the fragmentary Protarchaeopteryxhave been or can be provisionally included ina phylogenetic hypotheses for the theropods.
Feathers are not preserved in Compsognathuslongipes (Ostrom 1978), but since Sinosauropte-ryx is likely closely related to Compsognathus, itis difficult to conclude whether they wereabsent or not. By parsimoniously superimpos-ing these data on one recent phylogenetichypothesis for the theropods (Sereno 1999), itis possible to reconstruct an historical hypoth-esis for the origin and diversification of thefeathers within theropods (Figure 6). Feathersoriginated in a lineage of coelurosaurian the-ropod dinosaurs including both Sinosauropte-ryx and birds, but excluding allosauroids, cer-atosaurids, and coelophysids. Subsequently,feathers with primary branched structure(i.e., tufted), and possibly a rachis, evolvedin a lineage that included the commonancestor of alvarezsaurids, ornithomimids,therizinosaurids, tyrannosaurids, ovirapto-rosaurids, troodontids, dromaeosaurs, andbirds; branched structure may have alreadyevolved if the feathers of Sinosauropteryx ulti-mately reveal genuine branched structure.The rachis, barbules, and the closed penna-ceous vane had all evolved within or beforea lineage that included the oviraptorosaur-ids, troodontids, dromaeosaurs, and birds.Flight and asymmetrical remiges and rectri-ces evolved within the exclusive commonancestor of the birds.
These new paleontological finds also per-mit us to understand some of the early eventsin the evolution of plumage, or the distribu-tion of feathers on the body. Both Caudipteryx(Chen et al. 1998; Gatesy 2001) and anunnamed basal dromaeosaur (Norell et al.2002) exhibit a tuft of long pennaceous feath-ers on the tip of the tail. Furthermore, a diver-sity of Yixian theropods have long feathersattached to the ulna and manus. These obser-vations indicate that aspects of the distribu-tion and pennaceous structure of the “remi-ges” and “rectrices” in birds evolved in acoelurosaurian ancestor long before the ori-gin of flight in birds.
Although it has long been recognized thatbirds are a lineage of theropod dinosaurs,the discovery of “dinosaur feathers” (i.e.,nonavian theropod feathers) has irreversiblychanged our understanding of the evolutionof feathers and birds. Birds are no longer syn-onymous with the possession of feathers, and

September 2002 279EVOLUTION AND DIVERSIFICATION OF FEATHERS
Figure 6. Phylogenetic Hypothesis for the Origin and Diversification of FeathersThis historical hypothesis for the origin and diversification of feathers is based on recent paleontological
discoveries and a proposed phylogeny of the theropod dinosaurs (Sereno 1999). Details of fossil evidence offeathers is displayed at the top: presence (P) or absence (A) of feathers, presence of feather branched structure(Y ), presence of feather diversity (Y ), and the most advanced stage of the developmental model of featherevolution exhibited by that taxon. Fossil evidence of feathers has been reported for eight nonavian theropods.Sinosauropteryx, Shuvuuia, Beipiaosaurus, Caudipteryx, Sinornithosaurus, Microraptor, and the unnamed dromaeo-saur are depicted here as sister taxa to the clades that they have been demonstrated to belong in. The onlyspecimen of the eighth taxon, Protarchaeopteryx, is too fragmentary to be phylogenetically assigned. Feathers arecoded as unknown in the basal coelurosaur Compsognathus. The historical hypothesis for the evolution of featherand feather diversity is a parsimonious reconstruction based on the available data. Optimization of some eventsis equivocal because of missing data (?).
our understanding of the biology of derivedcoelurosaurs will certainly be changed by thisnew appreciation of their integument (Prum2002).
Congruence Between Paleontologyand the Developmental Theory
The evidence that the filamentous integu-mentary structures of nonavian theropoddinosaurs are homologous with avian feathersraises the question of whether these newlydescribed primitive feather morphologies arecongruent with those predicted by the devel-opmental model (Figure 5; Prum 1999). Xu
et al. (2001) showed that the structure of thetufted and serially branched feather mor-phologies found in Sinornithosaurus are con-gruent with Stage II and Stage IIIa of thedevelopmental model. They further pro-posed that the shorter, cylindrical, morerobust integumentary structures in Sinosau-ropteryx were congruent with the hollow tubu-lar morphology of Stage I. Ji et al. (2001)independently concluded that the three typesof feathers documented on the unnamedclose relative of Sinornithosaurus are also con-gruent with Stages II, IIIa, and V of the devel-opmental model (Prum 1999).

280 Volume 77THE QUARTERLY REVIEW OF BIOLOGY
Xu et al. (2001) did not hypothesize thepresence of barbules on the feathers of Sinor-nithosaurus, but Ji et al. (2001) proposed thatthe herringbone pattern of barbs preservedon the forelimb and tail feathers of theunnamed basal dromeosaur constitutes evi-dence of coherent vanes that were main-tained by differentiated interlocking distaland proximal barbules (Stage V: Figure 5;Prum 1999). The difficulty in determiningthe presence of barbules in fossils is that bar-bules consist of a linear series of individualkeratinocytes that are too small to be pre-served in normal fossils. There is no directevidence of barbules on the feathers ofArchaeopteryx (or any other fossil bird as far aswe know), but their existence has beeninferred from the conformation of the barbsinto a coherent, closed vane. But the absenceof a closed pennaceous vane may be the resultof the absence of barbules (Stage IIIa), thepresence of plesiomorphic barbules that lackthe differentiated interlocking morphology(Stage IIIa�b), or the complete separationof the barbs of a closed pennaceous vane dur-ing preservation (Stage IV). Thus, the coher-ent vanes in the feathers of Caudipteryx, Pro-tarchaeopteryx, and the unnamed basaldromeosaur do provide evidence for the pres-ence of differentiated barbules that is identi-cal to the evidence used to support their pres-ence in early bird feathers. We thereforeconclude that the feathers of the forelimbsand tails in Caudipteryx, Protarchaeopteryx, andthe two unnamed basal dromaeosaurs haddifferentiated distal and proximal barbules(Stage IV). It appears, however, that somefeathers in both Sinornithosaurus (Xu et al.2001) and its two unnamed close relatives ( Jiet al. 2001; Norell et al. 2002) also lack closedpennaceous vanes.
As Xu et al. (2001) comment, the currentpaleontological evidence of theropod feath-ers provides some additional phylogenetic sup-port for the transition series predicted by thedevelopmental model for the origin and evo-lution of feathers (Figures 5 and 6). Based onthe current sample, the first feather mor-phology to have evolved was the unbranchedtubular Stage I morphology represented bySinosauropteryx (Figure 6). This indicates thatStage I feathers evolved phylogenetically
prior to any other more complex morphol-ogy. Further phylogenetic congruence withthe predictions of the developmental modelis limited by variable preservation of theintegument of available specimens of coelu-rosaurs, the limited sample of coelurosaursavailable from the highest quality preserva-tion sites (i.e., Liaoning and Solnhofen), andthe bias that fully pennaceous feathers with aprominent rachis are likely to be preservedbetter than earlier stages with finer filamen-tous morphology. Branched feathers clearlyevolved prior to the common ancestor of thetherizinosaur Beipiaosaurus and birds (StageII), but it is not known whether these featherspossessed a rachis (Stage IIIa) or barbules(Stage IIIb). There is tentative support, how-ever, for the conclusion that branched andopen pennaceous structure (Stages II, IIIa,IIIb; e.g., Beipiaosaurus) evolved before differ-entiated proximal and distal barbules and theclosed pennaceous vane (Stage IV; e.g., Cau-dipteryx). Asymmetrically vaned feathersevolved coincident with flight in the commonancestor of birds (Stage Va).
Alternatives to the TheropodOrigin of Feathers
There are few viable alternatives to theconclusion that feathers originated in a non-avian lineage of theropod dinosaurs. One isthat feather-like branched structures evolvedmultiple times among reptilian integumentaryappendages. Convergent filamentous integu-mentary appendages did evolve within thearchosaurian pterosaurs (e.g., Sordes), butthese 5 mm long appendages are notbranched and are independently derivedwithin pterosaurs (Wellnhofer 1996). Thereare additional reports of similar hair-likestructures in additional pterosaurs frominner Mongolia, China (Wang et al. 2002).Also, tantalizing references to long filamen-tous integumentary appendages on the tail ofa specimen of psittacosaur (an ornithischiandinosaur) from the Yixian have also been pub-lished (Stokstad 2001b; Steghaus-Kovac 2002).Although they need to be studied in greaterdepth, the absence of detailed similaritiesshared between these appendages and avianfeathers (e.g., branched or tubular structure)and their distant phylogenetic positions indi-

September 2002 281EVOLUTION AND DIVERSIFICATION OF FEATHERS
cate that these structures are entirely conver-gent with theropod feathers. Convergent evo-lution, however, is highly unlikely to explainthe detailed similarities between the theropodintegumentary structures and avian feathers,given the numerous shared derived similari-ties (i.e., synapomorphies) of these feather-bearing theropod dinosaurs and birds (Gau-thier 1986; Holtz 1994; Sereno 1997, 1999).Diverse feathers including fully pennaceousvanes have now been documented preciselyin the dromeosaur lineage that is hypothe-sized to be most closely related to birds, andin the specimens within that clade that areclosest in age to the oldest indisputable bird,Archaeopteryx. Thus, the paleontological andphylogenetic data leave vanishingly little roomfor two convergent origins of feathers in twolineages that are hypothesized to be immedi-ate sister groups. Yet, hair-like appendagesfrom other nontheropod lineages of archo-saurs do indicate that there was probably aninherent potential for the evolution of diverseintegumentary appendages within all archo-saurs.
Second, it has been proposed that thesefilamentous, nonvaned structures are notintegumentary appendages but some type offibrous connective tissue such as collagen orossified tendons. This hypothesis is simplycontradicted by the abundance of the struc-tures, their obvious preserved relationship tothe integument, their distribution all over thebody, and their undeniable resemblance tofeathers preserved on avian specimens in thesame deposits. In many species, these struc-tures vary in size over the body in the sameway as modern avian feathers. Thus, Beipiao-saurus has an array of 70 mm filamentousstructures coming off the trailing edge of theulna, an unlikely position for any connectivetissue or tendinous structures and identical inposition to avian remiges (Xu et al. 1999a).Sinornithosaurus exhibits a tufted filamentousstructure 35 mm on the tip of its snout—agenuinely unlikely location for an ossifiedtendon (Xu et al. 2001)! The integumentaryappendages of the unnamed relative of Sinor-nithosaurus have a structural variation and dis-tribution over the body that is remarkablyreminiscent of avian plumage ( Ji et al. 2001).The integumentary appendages of these non-
avian theropods are also preserved in essen-tially identical form to many contour featherson indisputably avian fossil preserved in thesame deposits, such as Confuciusornis sanctus,Eoenantiornis buhleri, Changchegnornis hendaozo-iensis (Chiappe et al. 1999), and Protopteryxfengningensis (Zhang and Zhou 2000). It is dif-ficult to argue that the filamentous structuresof these nonavian theropods are entirely dif-ferent in nature from feathers, even thoughthey were accidentally preserved exactly asavian feathers were in the same fossil deposits.There is also the immunological evidencethat the filamentous appendages of the alvar-ezsaurid Shuvuuia are composed of b-keratinwhich is exclusively expressed in the integu-ment of reptiles (Schweitzer et al. 1999).Lastly, the indisputable evidence of the her-ringbone pattern of barbs and the rachis inappendages of basal dromeosaurs confirmsthat these structures are not collagenous orfibrous connective tissues ( Ji et al. 2001;Norell et al. 2002).
Third, various critics have disingenuouslystated that these fossils are too young to pos-sess feathers with ancestral morphologies(e.g., Feduccia 1999). Such statements ignorethe fundamental fact that a feature phyloge-netically generally distributed within a clade(i.e., feathers in higher coelurosaurian dino-saurs) must have evolved prior to another fea-ture that is more restricted to a single, mono-phyletic group within that clade (i.e., flight inbirds). Just as modern monotremes lay eggs,lineages may retain many primitive featurespresent in their ancient ancestors. There isno reason why these younger theropod fossilscannot preserve a morphology that is phylo-genetically primitive to an older avian fossil.Moreover, if Sinosauropteryx is a compsog-nathid, then these integumentary append-ages would presumably have been present inCompsognathus (contra Ostrom 1978), which iscontemporaneous with Archaeopteryx.
Fourth, it is possible to hypothesize that allthese feathered theropods are actually flight-less birds, as critics have argued with Caudip-teryx and Protarcheopteryx (Feduccia 1999;Jones et al. 2000b; Martin and Czerkas 2000).If all these feathered taxa were birds, thenfeathers could still have a single, exclusivelyavian origin. Such a phylogenetic hypothesis

282 Volume 77THE QUARTERLY REVIEW OF BIOLOGY
would be wildly unparsimonious. Given thenumerous primitive and derived features thatthese nonavian theropod taxa share withother theropods (Sereno 1999; Xu et al.1999b, 2000; Zhou and Wang 2000; Zhou etal. 2000), their placement within the birdswould only further contribute to an indisput-able relationship between birds and theropoddinosaurs. It becomes increasingly difficult toargue that various birds and dinosaurs canbe mistaken for each other, but that thesegroups have no phylogenetic relationship toone another.
Rejection of the theropod origin of feath-ers is often accompanied by rejection of clad-istic analysis in general and the theropodancestry of birds in particular (e.g., Feduccia1999; Dodson 2000). Although we will notreview the debate on the theropod ancestryof birds here, we can reiterate that there is noother explicit alternative hypothesis that hasany data to support it (Prum 2002). No criticsof the theropod origin of birds have ever pre-sented an explicit alternative hypothesis ofavian ancestry (Feduccia 1999; Dodson2000). It is insufficient to simplistically dis-miss numerous phylogenetic reconstructionsof independent researchers supporting thetheropod origin of dinosaurs as “garbage in,garbage out” (Stokstad 2000), and then main-tain that there is no available solution to thequestion. To reject the theropod origin ofbirds, one must demonstrate explicitly thatsome other taxon is more closely related tobirds than are theropods. Critics of the the-ropod origin of birds have not even attemptedto do so. In contrast, Feduccia (1999) pro-posed the highest possible standard of evi-dence for the theropod ancestry hypothesisin his challenge to paleontologists to findfeathers on a dromaeosaur—the lineage oftheropods frequently hypothesized to be phy-logenetically most closely related to birds. Inonly a couple of years, fossil integumentaryappendages with several features uniquelyshared with avian feathers have now beenfound in three different taxa from exactly thislineage (Xu et al. 1999b; Xu et al. 2000; Ji etal. 2001; Xu et al. 2001; Norell et al. 2002).With the apparent richness of the Yixian For-mation (Stokstad 2000), it is unlikely thatthese discoveries will end soon.
The Appendages of LONGISQUAMA
One proposed alternative to theropodorigin of feathers—the appendages of Lon-gisquama—requires special attention. Thesmall Triassic reptile Longisquama insigniswas described by Sharov (1970). Longisquamais notable for its extremely long, lamellarintegumentary appendages along its spinethat have been hypothesized to be used in glid-ing (e.g., Feduccia 1999). Sharov (cited as per-sonal communication by Regal 1975) main-tained that Longisquama’s dorsal appendageswere unbranched. Feduccia (1985:76) alsostated that the dorsal appendages of Longis-quama “are not incipient feathers” and thatwhile inspecting the specimens, “I could seeno indication that the elongate scales wereparticularly feather like.” Feduccia (1985:76)concluded that there “is simply no evidencethat they in fact are [feather-like structures].”He later confirmed, “No doubt . . . that thescales of Longisquama were not transmutedinto feathers” (Feduccia 1999:133).
Despite this unanimity of opinion overthree decades, Jones et al. (2000a)—an “icon-oclastic band” (Stokstad 2000) of authorsincluding Feduccia himself—recently pro-posed that the long dorsal appendages of Lon-gisquama were branched structures with arachis and barbs composed of keratinaceousfilaments, and that these structures arehomologous with avian feathers. Subse-quently, the appendages of Longisquama havebeen cited as providing morphological sup-port for the elongate scale hypothesis of theorigin of feathers (Maderson and Alibardi2000). This proposal has received rapid,strong criticism (Reisz and Sues 2000; Prum2001; Unwin and Benton 2001), and nearlycomplete rejection by paleontologists andornithologists familiar with the material(Stokstad 2000). Based on a detailed analysisof the fossils, Reisz and Sues (2000) con-cluded that conditions of deposition of thefossil were inappropriately interpreted andthat many of the proposed feather-like fea-tures were artifacts of preservation. Variousauthors have noted that the characters usedto identify Longisquama as an archosaur—theantorbital and mandibular fenestrae—arepoorly preserved at best and are quite possi-bly artifactual; it is uncertain whether Longis-

September 2002 283EVOLUTION AND DIVERSIFICATION OF FEATHERS
quama is even an archosaur (Stokstad 2000;Unwin and Benton 2001). Prum (2001) com-pared the appendages of Longisquama tofeathers and argued that the proposedbranched structure was illusory (Sharov, ascited by Regal 1975; Feduccia 1985, 1999),that the Longisquama appendages consistedof a continuous membranous lamina, andthat many of the documented details of theproposed branched structure were inconsis-tent with the known mechanisms of feathergrowth. In an energetic but largely semanticrebuttal, Jones et al. (2001) maintained thatthe antorbital fenestra was visible on thepreviously unillustrated counterslab, thatthe feather-like branched structures were“observable facts” (apparently previouslyinvisible to both Sharov and Feduccia), andthat the questions raised about possible arti-facts of deposition, “strain credulity.”
The homology of the Longisquama append-ages with avian feathers is highly disputed.Authors on both sides of the debate are cur-rently preparing additional analyses for pub-lication (P F A Maderson, R Reisz, H-D Sues,personal communication). Given the prob-lematic and controversial evidence itself, thehypothesis that these Longisquama appendagesare homologous with feathers has substantialadditional obstacles. First, this hypothesiswould imply either: (1) that feathers areuniquely shared by Longisquama and birds, andthat birds are unrelated to theropod dino-saurs; or (2) that feathers are primitive to mostarchosaurs and have been lost many times.Alternative (1) would require the rejection ofnumerous derived morphological charactersfrom every part of the body (including mostrecently pennaceous feathers from the integ-ument) that are shared by birds and lineagesof theropod dinosaurs on the basis of thesedisputed impressions. Alternative (2)—oneorigin and numerous losses of feathers—isextremely unparsimonious and completelyunlikely. An additional problem with thehypothesized homology between the append-ages of Longisquama and avian feathers is theimplied scenario for the evolution of avianflight. Feduccia (1999) and others maintainthat Longisquama’s appendages were used forgliding flight. So the hypotheses of homologybetween Longisquama’s appendages and avian
feathers implies that feathers first originatedalong the spine for aerial gliding, and thenspread over the body to create the wings andtail (resulting in the evolution of a secondnovel flying structure while maintaining con-tinuous gliding or flying ability); the originalspinal flying structures were then reducedto become indistinguishable from standardcontour feathers. Thus, the direct implica-tion of Jones et al. (2000a) is that Longis-quama’s “bizarre and unique solution to theproblem of gliding” (Feduccia 1999:95) isactually ancestral to avian flight. This unusualscenario has yet to be defended.
Given the congruence among the phylo-genetic support for the relationship betweenbirds and theropod dinosaurs (e.g., Sereno1999), the fossil evidence of feathers withinlineages of nonavian theropods, and the pre-dictions of primitive feather morphologymade by the developmental model, the alter-native hypothesis that the appendages of Lon-gisquama are homologous with feathers ispoorly supported.
The Tubular Nature of FeathersAn important implication of the develop-
mental model of the origin of feathers is therecognition of the essentially tubular (or hol-low cylindrical) nature of feathers. The cylin-drical follicle and feather germ are generalfeatures that characterize all feathers, there-fore they should be considered the definingfeatures of feathers. Prum (1999) defined afeather as a elongate, cylindrical or tubularepidermal appendage that grows from aninvaginated feather follicle. A feather folliclediffers from a hair follicle in that the follicularinvagination is not merely a depression in theepidermis but a circular trough that encirclesa persistent dermal papilla.
The inherently tubular nature of feathersis manifest in numerous details of featherdevelopment and morphology. All feathersoriginate and grow as tubes of epidermis. Thetubular nature of feathers can be observeddirectly in the growing cylindrical pin feather,the sheath, and the basal calamus character-istic of all known feathers. The familiar planarstructure of a pennaceous feather is only real-ized after the tubular feather emerges fromits superficial sheath. The hypothesis that pla-

284 Volume 77THE QUARTERLY REVIEW OF BIOLOGY
nar, pennaceous feathers are primitive fails toexplain this most general and fundamentalfeature of feathers.
Developmental and genetic perturbationsof feather structure further demonstrate thetubular nature of feathers. Strong (1902a)documented the case of a developmentalanomaly in the contour feathers of a hybriddove in response to temporary food depriva-tion during molt. In the middle of each con-tour feather, the barbs suddenly failed to dif-ferentiate and the entire feather temporarilyreverted to a cylindrical, calamus-like form inthe middle of the feather vane. When foodwas resumed, the barb ridges differentiatedagain, forming a normal vane. The effect wasof a nearly normal pennaceous feather withan anomalous cylindrical calamus in the mid-dle of the vane. Similar anomalies weredescribed by Riddle (1908). Genetic supportfor the essentially cylindrical nature of feath-ers comes from the recessive feather muta-tion Porcupine (pc), that has been describedin pigeons (Cole and Hawkins 1930), chick-ens (Waters 1967; Somes 1990), and Japanesequail (Fulton et al. 1982; Cheng and Brush1984). The Porcupine mutation producesincomplete differentiation and morphogen-esis of the barbs, and results in brittle tubular,quill-like feathers composed of a mass of anas-tomosing barbs that cannot unfold into thetypical planar form. Both of these cases dem-onstrate that perturbations in feather devel-opment reveal the essentially cylindrical,tubular nature of feathers.
Hypothesized primitive feathers are fre-quently referred to as “protofeathers” (e.g.,Brush 2000; Maderson and Alibardi 2000).Given the enormous structural diversity ofextant feathers, it is arbitrary to attempt todistinguish “real” feathers from the varioustheoretically antecedent “protofeathers” onmorphological grounds. In order to empha-size the fundamental importance of the cylin-drical, tubular organization of the follicle asthe inherent feather novelty, we think it ispreferable to recognize all of the diversestructures that likely grow from feather folli-cles as feathers, and we recommend that theterm “protofeather” be abandoned. Primitivestructures that are entirely homologous withfeathers should be called feathers.
Homology of Feathers and ScalesSince the mid-19th century, feathers have
frequently been broadly homologized withscales (reviewed in Lucas and Stettenheim1972). Although feathers and scales sharesome morphological, developmental, andmolecular features, problems with thishypothesis have been known for more than acentury (Davies 1889; see discussion above ofscale-based theories of feather origins). Anumber of fundamental and important dif-ferences between feathers and scales ledBrush (1993, 1996) to question their homol-ogy. Previous simplistic conceptualizations ofthe homology of feathers and scales (with theintellectual goal of evolutionarily transform-ing one into the other) have unnecessarilyobscured the complex, hierarchical relation-ship between these structures. Further, it hasprevented the appropriate recognition of theevolutionary novelties of feathers that requireexplanation.
Feathers and scales are integumentaryappendages that are created by local prolif-eration, differentiation, and keratinization ofepidermal cells (Lucas and Stettenheim 1972;Sawyer et al. 1986). Detailed scrutiny of thedevelopment of scales and feathers indicatesthat there are actually few features sharedbetween the structures (Sawyer et al. 1986).Feathers and scales are appendages producedby local proliferation of epidermal and der-mal cells that creates a folding or protuber-ance on the skin. All feathers and reptilianscales are created by epidermal keratinocytes,but the type of keratin and its distribution inthese structures vary extensively (Baden andMaderson 1970; Maderson 1972; Brush 1985;Sawyer et al. 1986; Brush 1993; Maderson andAlibardi 2000; Sawyer et al. 2000). Feathersand the scutate scales of birds (those on theupper, or dorsal, surface of the legs and feet)begin development with a morphologicallydistinct placode—an epidermal thickeningabove a condensation of dermal cells. In con-trast, avian reticulate scales (those on thelower surfaces of the toes) and all reptilianscales examined to date lack a morphologi-cally definable placode (e.g., Maderson andAlibardi 2000). Avian reticulate scale rudi-ments share temporal and spatial patterns ofexpression of several feather-placode specific

September 2002 285EVOLUTION AND DIVERSIFICATION OF FEATHERS
genes, however (Chuong et al. 1996; Widelitzet al. 1999). Subsequent to placode forma-tion, numerous events in feather develop-ment constitute a series of unique develop-mental mechanisms which have no homologin any other avian or reptilian scales (Brush1993, 1996, 2000; Prum 1999). The differen-tiation of barb ridges from one another, thetubular invagination around the dermalpapilla that creates the follicle, and the sub-sequent developmental mechanisms withinthe follicle collar, the barb ridges, and thetubular feather germ, all of which determinefeather form, are all unique to feathers.
How are feathers homologous with scales?The presence of a derived, morphologicallydistinct placode is only shared by feathers andavian scutellate scales (Maderson and Ali-bardi 2000). The patterns of gene expressionspecific to feather and avian scutate scaleplacodes are also shared with avian reticulatescales, however (Chuong et al. 1996; Widelitzet al. 1999). Thus, a molecularly defined plac-ode is shared by all avian scales and feathersregardless of the presence of the morpholog-ical characteristics of a feather placode.These developmental features support thehomology of avian scales and feathers at thelevel of the placode. Although scutate scaleshave been hypothesized to be secondarilysimplified feathers (Davies 1889), it is moreparsimonious to hypothesize that simpler scu-tate appendages with a morphological plac-ode preceded the origin of feathers, and thatthese scutate structures were retained on theanterior surfaces of the legs in modern birds.
A series of classical (Sengel 1976) and mod-ern developmental experiments (Chuong andWidelitz 1998; Chuong et al. 2000) have doc-umented many commonalities in the signalingmechanisms of mesenchyme-epithelium inter-actions during the development of chick feath-ers and scales. Developmental experimentshave even “transformed” early scutate and rec-ticulate scales into feather short buds inresponse to various chemicals (Dhouailly et al.1980; Zou and Niswander 1996; Widelitz et al.2000). These experiments have reinforced thenotion that there is detailed molecular sup-port for the traditional, broad hypothesis ofhomology between feathers and scales (e.g.,Dyck 1985; Maderson and Alibardi 2000).
While these experiments contribute importantand fundamental details about the mecha-nisms of mesenchyme-epithelium signaling invertebrates, these data do not suggest thatfeathers and scales are broadly homologous(i.e., beyond the level of the placode). Thetransformations between these structures arepurely unidirectional; undifferentiated scalestructures can be induced to produce featherson them, but feather placodes or germs haveyet to be induced to grow into scales (M PHarris, personal communication). This asym-metry is congruent with the hypothesis thatfeather and scale placodes are homologousand that postplacode mechanisms of feathermorphogenesis are all derived.
In conclusion, the morphological andmolecular developmental details shared byavian feather and scales support homologybetween these structures at the level of theplacode. The morphology and developmentof all subsequent structures within the featherare evolutionary novelties that have no hom-ologs in avian or reptilian scales.
Functional ExplanationsReconsidered
Explicit attempts to reconstruct the evolu-tionary history of feathers based on naturalselection for specific functions have failed toidentify an unequivocal, plesiomorphic func-tion of feathers or to accurately predict a tran-sition series of ancestral feather morpholo-gies. We submit that primarily functionalapproaches to reconstruct the evolutionaryhistory of feathers have hindered rather thanadvanced our understanding of the origin offeathers. With the development of phyloge-netic methods, it has been established thatphylogenetic pattern should be determinedbefore the analysis of evolutionary process(e.g., Lauder and Rose 1996). Obviously, it isdifficult to explain why or how some event hasoccurred in evolutionary history withoutactually knowing what has occurred. But thisis exactly what functional theories of featherorigins attempt to do.
Functional explanations of the origin offeathers have received so much attentionbecause the question itself is so compelling,and not because the answers are particularlysatisfying. The function of ancestral feathers

286 Volume 77THE QUARTERLY REVIEW OF BIOLOGY
and the evolutionary mechanisms by whichthey diversified remain fascinating and diffi-cult questions. With the startling discovery ofprimitive feathers in nonavian dinosaurs, wenow have the first data that allow us to reeval-uate proposed functional explanations. Whatdoes the first view of what happened implyabout why and how feathers evolved?
A prominent and oft repeated functionalhypothesis for the origin of feathers is thatthey evolved by natural selection for flight(Steiner 1917; Heilmann 1926; Parkes 1966;Feduccia 1985, 1993, 1999). Recent confir-mation of filamentous and pennaceous feath-ers on nonavian theropod dinosaurs docu-ments that feathers originated in terrestrialcoelurosaurian theropods before the originof birds and before the origin of flight. Con-sequently, aerodynamic hypotheses for theinitial function of feathers have been falsified.It has been repeatedly and strenuously pro-posed that the exquisite adaptation of avianflight feathers for their aerodynamic functionimplies that feathers must have originatedthrough selection for flight (e.g., Feduccia1999), but given the evidence of pennaceousfeathers on the forelimbs and tails of primar-ily flightless theropod dinosaurs, this argu-ment is simply false. Concluding that feathersevolved for flight is like maintaining thatdigits evolved for playing the piano. Only themost derived and complex feather morphol-ogy could yield a feather with advanced aero-dynamic function (Prum 1999), and onlyafter the evolution of a bipinnate, closed pen-naceous vane with differentiated distal andproximal barbules (Stage IV) could therehave been selection for an asymmetrical vanewith aerodynamic properties (Prum 1999).The occurrence of fully pennaceous featherson terrestrial nonavian theropods completelyconfirms this conclusion.
The falsification of the aerodynamic hypoth-esis further corroborates Prum’s (1999) con-clusion that elongate aerodynamic scales weredevelopmentally implausible because the sur-faces of planar scales and feather vanes arenot homologous. Given how branched flightfeathers grow, an evolutionary transformationbetween these two analogous planar append-ages would have had to proceed through inter-mediary cylindrical and filamentous morphol-
ogies that could not have maintained aconsistently planar structure or aerodynamicfunction (Prum 1999). The only feather nov-elty attributable to natural selection for aero-dynamic function is the evolution of the asym-metrical feather vane (Stage Va: Figure 5), theorigin of which exactly co-occurs with the ori-gin of flight and birds (Figures 6).
Recently, another aerodynamic hypothesisfor the origin of feathers has been proposed.Based on the microanatomy and functionalmorphology of feather movement, Homber-ger and de Silva (2000) proposed that feath-ers evolved to produce smooth, adjustablebody contours for “body lift-gliding.” As withtraditional aerodynamic hypotheses, this sug-gestion is falsified by evidence of the originand diversification of feathers within a terres-trial, nongliding lineage of theropod dino-saurs. The feather movement mechanism ofHomberger and de Silva (2000) requires acalamus for the attachment of feather erectorand depressor muscles, however, and a tubu-lar calamus would be available by Stage I ofthe developmental model (Figure 5; Prum1999). Thus the developmental model couldprovide an extremely early potential originfor the feather movement system, as pro-posed by Homberger and de Silva (2000).
Prum (1999) concluded that numerousother proposed initial functions of plesio-morphic feathers are developmentally plau-sible because the simplest possible featherscould have performed these functions. Thesehypotheses include thermal insulation (Davies1889; Ewart 1921), thermal shielding (Regal1975), communication (Mayr 1960), waterrepellency (Dyck 1985), tactile organs (Bro-man 1941), and defense (Prum 1999). Eachhypothesis constitutes a physically plausibleselective advantage of the earliest feather, ahollow tubular filament. Of course, the width,length, and rigidity of the first feathers remainunknown. Based on our current knowledge ofthe biology and natural history of coelurosau-rian theropods, it would be entirely specula-tive to maintain that any of these plausiblefunctions actually was the selective functionthat led to the fixation of the first feathers. Infact, the possibility of gathering morphologi-cal or behavioral information from fossil rep-resentatives of the actual lineages in which

September 2002 287EVOLUTION AND DIVERSIFICATION OF FEATHERS
this event occurred is essentially impossible,and we can rationally abandon hope of iden-tifying a singular functional explanation.
There is, however, an important fundamen-tal commonality among these plausible func-tional hypotheses that is congruent with thedevelopmental theory of the origin of feath-ers (Prum 1999) and that provides a morefundamental explanation of the origin offeathers. Any selection for a substantial integ-umentary appendage that emerges from theskin or extends out of the skin would essen-tially constitute selection for the evolution ofa tubular follicle, the initial event (Stage I) inthe evolution of feathers (Figure 4). Theinvaginated feather follicle creates the oppor-tunity for the continuous proliferation andgrowth of a tubular epidermal appendage upand out of the follicle without demandingequivalent growth of the dermis within theappendage as required by a simple scale (Fig-ure 3D; Prum 1999). The feather germ isnourished by the central dermal pulp, but thepulp does not grow continuously in volumewith the feather germ. Rather, pulp caps con-tinuously seal off the top of the dermal pulpand limit its growth while the pulp providesnutrition for continued epidermal growth(Lucas and Stettenheim 1972). The associa-tion between the follicle and indeterminateepidermal growth is corroborated by the con-vergent evolution of a follicle and extensiveor indeterminate growth in mammalian hair.Whatever it was, the initial functional advan-tage of the earliest feathers constituted natu-ral selection for an emergent appendage thatthen fostered the evolution of the feather fol-licle (Stage I). Although the original func-tional advantage of the first feathers remainsa mystery, the ultimate explanation for theorigin of the feather must have involved selec-tion for epidermal appendages that emergedfrom the skin.
It has frequently been assumed that an evo-lutionary novelty such as feathers must haveevolved for an important reason. For exam-ple, both Regal (1975) and Feduccia (1999)found Mayr’s (1960) hypothesis that feathersevolved by selection for display to be unsatis-fying. Implicit in these criticisms (and in mostfunctional theories) was the notion that afunctionally important key innovation cannot
be explained by an inconsequential mecha-nism (e.g., mate choice), and must haverequired strong direct natural selection for acurrent function. Actually, the origin anddiversification of feathers was likely charac-terized by a diversity of selective forces at dif-ferent stages (Brush 1993). Furthermore,strong natural selection for a specific func-tion can lead directly to functional and devel-opmental integration of a trait within thephenotype that can limit subsequent evolva-bilty (Wagner and Schwenk 2000). Accord-ingly, if early feathers were under strong nat-ural selection for a specific function, suchselection could very well have constrainedthem from continuing to evolve and diversifyin form. It is important to recognize thatdiverse sources of natural or sexual selectionwere likely involved in the evolution of vari-ous stages in feather diversity. By focusing toointensely on singular functional explanations,functional theories of the origin of feathershave obscured the fact that the history offeather evolution is characterized by a contin-ued diversification and novelty in develop-ment, form, and function that cannot beexplained by natural selection for a singlefunction. The most notable features of theevolutionary history of feathers are therepeated origins of the developmental nov-elties that created their increasing hierarchi-cal complexity in form, and provided theopportunities for the evolution of diversity infunction.
Feathers as Evolutionary NoveltiesRecent developments in phylogenetics and
molecular developmental biology have stim-ulated reconsideration of the concept of evo-lutionary novelty (Nitecki 1990; Muller andWagner 1991; Raff 1996; Wagner 2001).Muller and Wagner (1991) defined a mor-phological novelty as a structure that is nei-ther homologous to any ancestral structurenor homonomous (i.e., serially homologous)with any other structure in that organism.
Many features of feathers and feather devel-opment meet this definition and qualify as evo-lutionary novelties. The follicle, the differen-tiated sheath and feather germ, differentiatedbarb ridges, barb rami, barbules, differenti-ated pennulae of the proximal and distal bar-

288 Volume 77THE QUARTERLY REVIEW OF BIOLOGY
bules, and the rachis are all evolutionary nov-elties, as are the derived mechanisms by whichthese novel structures develop. At a molecularlevel, the derived 10 kilodalton �-keratins offeathers are also novel (see below). The mor-phologically distinct placode present in theinitial development of feathers and avian scu-tellate scales is also apparently an evolutionarynovelty, though it may be an extreme in a con-tinuum of changes in cell proliferation anddensity during epidermal appendage devel-opment. If the morphological placode andthe anterior-posterior axis of feather primor-dia were independently evolved from otheravian and reptilian integumental append-ages, then feathers may be entirely novelstructures (but see Homology of Feathersand Scales above).
An explicit, process-independent definitionof evolutionary novelty (Muller and Wagner1991) permits us to examine which of thehypothesized mechanisms for the origin ofevolutionary novelties may have been involvedin the evolution of feathers. Muller andWagner (1991) propose three modes for gen-erating novelties: hierarchical organization,interactivity and dissociability, and equilibriaand thresholds. As documented by bothBrush (1993, 1996, 2000, 2001) and Prum(1999), feather development is extraordinar-ily hierarchical. This hierarchy incorporatesboth nestedness of morphological modulesand the complex interactions among mod-ules. Thus, the tubular epidermal feathergerm includes both the peripheral sheathand the developing feather. The developingfeather is composed of a series of barb ridgesthat develop into many pairs of barbule platesand the ramus. The fusion of barb ridgesearly in the development of a pennaceousfeather determines the identity of the rachisridge, which becomes a major morphologi-cal component of the feather. The presenceof helical growth and the rachis define theproximal and distal sides of the barb ridge,and permit the differentiation of these mod-ules into the morphologies that create theclosed pennaceous vane. The origin and dif-ferentiation of hierarchically nested mor-phological modules in the feather haveresulted in numerous evolutionary noveltiesin structure and development, and consti-
tute the basis of the extraordinary diversityof extant feathers.
Other novelties within feathers are hierar-chical duplications and alterations of theseprocesses. For example, the development ofan afterfeather—a second posteriorly ori-ented vane growing out of the same follicleand attached to the same calamus (Figures 1and 2)—occurs as a consequence of a secondopposing, posteriorly oriented direction ofhelical growth (Lillie and Juhn 1938; Lucasand Stettenheim 1972). As a consequence ofduplication and redirection, the same mech-anisms that produce the main vane of thefeather result in the division of the posteriornew barb locus into two laterally displacednew barb loci, the creation of a second rachisridge (the hyporachis), and ultimately anentire second vane growing simultaneouslyfrom a single follicle.
Interactivity and dissociability refer to theability of components to create new struc-tures through changes in the interactionsamong tissues, or modules, which create theinterconnectedness of ontogenetic networks.Most novelties in feather development ariseas a result of such interactions and dissocia-tions. For example, the interactions of barbridges through fusion initiate the formationof the rachis ridge and, ultimately, determinea fundamental feature of feather morphol-ogy. Further, dissociation of the developmentof the proximal and distal barbule plates andthe independent specification of barbule cellshape within them provided the opportunityfor differentiation of barbules and the evo-lution of the coherent, closed vane of pen-naceous contour and flight feathers.
Equilibrial and threshold effects are in-volved in feather morphogenesis at cellularand molecular levels. Experimental analyseshave determined that chemical thresholdresponses to expression gradients are involvedin the specification of the patterns of featherfollicle location during early development(Chuong et al. 1990; Chuong 1993; Chuongand Widelitz 1998; Jung et al. 1998; Widelitzet al. 1999; Chuong et al. 2000). The molecu-lar understanding of feather morphogenesisbeyond the placode stage is still in its infancy,however, and other examples of the effects ofand responses to molecular gradients on other

September 2002 289EVOLUTION AND DIVERSIFICATION OF FEATHERS
aspects of feather morphogenesis have yet tobe elucidated.
Raff (1996) recognized an additionalmechanism for the origin of evolutionarynovelties: duplication and divergence. Dupli-cation and divergence have played funda-mental roles in the evolution of feather struc-ture and diversity. The duplication of keratingenes is one example at a molecular level(Brush 1978, 1993). At the morphologicallevel, the duplication of numerous featherfollicles over the body surface and subse-quent divergence in morphology of differentfeathers have contributed greatly to the ori-gin and maintenance of feather novelties.
Recently, Wagner and colleagues (Wagner2000; Wagner et al. 2000; Chiu and Wagner2001) proposed that research on the originof evolutionary novelties should be distinctfrom research on standard microevolutionarychange, and should be restructured to askfundamentally different questions that focusdirectly on the mechanisms of the origin ofqualitative innovations. This view under-scores why the traditional neo-Darwinianapproaches to the origin of feathers, as exem-plified by Bock (1965) and Feduccia (1985,1993, 1999), have failed. By emphasizing thereconstruction of a series of functionally andmicroevolutionarily plausible intermediatetransitional states, neo-Darwinian approachesto the origin of feathers have failed to appro-priately recognize the novel features offeather development and morphology, andhave thus failed to adequately explain theirorigins. This failure reveals an inherent weak-ness of neo-Darwinian attempts to synthesizemicro and macroevolution. In contrast, thedevelopmental theory of the origin of feath-ers focuses directly on the explanation of theactual developmental novelties involved inthe origin and diversification of feathers(Prum 1999). Restructuring the inquiry tofocus directly on the explanation of the originof the evolutionary novelties of feathers yieldsa conceptually more appropriate and produc-tive approach.
Why are feathers so complex? Why havefeathers been able to diversify and innovateso extensively? Pending further details on themolecular mechanisms of feather develop-ment (see below), the explanation appears to
come from the innovative structure of thetubular feather follicle and germ, and theirunique potential for interactions between theperipheral epidermis of the developingfeather germ and the central dermal papilla(Figures 3D and 4). There are essentially twobasic kinds of epithelial appendages: thosethat develop from an outgrowth (e.g., scalesand claws), and those that develop fromingrowth (e.g., glands and hair) (Chuong1998). A few appendages, like hair, horn, andfeathers, are distinct in having evolved bothmechanisms. Hair starts with an invaginationand then proliferates from the bottom. In con-trast, a feather starts with an outgrowth (thefirst feather papilla or short bud), and thendevelops its evolutionarily novel ingrowth (thecylindrical follicle). This unique combinationof events creates a tubular juxtaposition of epi-thelial and mesenchymal layers. The tubularorganization of the feather follicle and germfostered the formation and proliferation ofnumerous hierarchically arranged develop-mental modules, such as the barb ridges withinthe entire feather germ and the barbulesplates and ramus within the barb ridges. Thishierarchical, modular organization apparentlyfosters the development of distinct, polarizedmolecular concentration gradients and asym-metries at multiple levels within different mod-ules. The tubular epidermal feather tissue canbe differentiated into barb ridges which them-selves have the potential to differentiate alongthe peripheral-medial, lateral, and proximo-distal axes into the ramus and a series of bar-bules. The evolution of the tubular epidermalfeather germ provided a platform for the evo-lution of an array of subsequent developmen-tal evolutionary novelties that has generatedthe diversity in feather morphology. Interest-ingly, it is actually the constraints, or limitations,on the possible developmental processes per-mitted within this tubular architecture thatcreated the hierarchy in these mechanismsand permitted the identification of the devel-opmental model of the origin of feathers.
Brush (2000, 2001) emphasized the rapid-ity and the inevitability of the evolution ofstructural diversity in feathers subsequent tothe initial origin of the follicle. Althoughobservable mechanisms of feather morpho-genesis are impressive, there are no data yet

290 Volume 77THE QUARTERLY REVIEW OF BIOLOGY
supporting the conclusion that these featuresevolved rapidly or were in any sense inevit-able. Although the fossil record has suddenlyproduced an unexpected treasure of speci-mens relevant to this question, the possibilityof getting sufficient temporal samples toaccurately judge the speed of feather evolu-tion remains remote. Further, historical con-tingency may have played an important rolein the evolution of the developmental andstructural complexity of feathers that insightsinto the molecular mechanisms of feathermorphogenesis may reveal.
Evolution of Feather KeratinsLike other amniote epidermal appendages,
feathers are composed of keratin—an inter-mediate filament protein produced by epi-dermal cells that forms a hard, flexible, andinsoluble polymer. Keratins are extremelydiverse in molecular structure, and two maingroups are recognized: the �-keratins arecharacterized by an �-helical structure, andb-keratins by a b-pleated sheet organization.It is still debated whether the quite distinct �-keratins and b-keratins are historically relatedor entirely convergent in structure and func-tion. There are few sequence similarities andsignificant molecular differences. The �-ker-atins are found in the epidermis of all amni-otes, whereas the b-keratins are found exclu-sively in the epidermis of reptiles, includingbirds. The �-keratins compose all the skinand integumentary appendages of mammals.The �-keratins also compose the soft skin andinterscale areas of reptiles and birds, but theintegumentary appendages of reptiles andbirds can be composed of both �-keratins andb-keratins. All b-keratins are smaller than the�-keratins, and are produced by a family ofclosely related genes. Their basic chemistryis similar, as is their capacity to form fila-ments, although the mechanisms differ. Theb-keratin genes occur as low, tandem repeatsthroughout the genome.
Feathers are composed of a subclass of b-keratins that are referred to as feather kera-tins, or �-keratins (Brush 1978, 1985, 1993;Sawyer et al. 2000). Avian scutate scales, beak,and claw are composed of another subclass offilament, forming �-keratins that are slightlylarger. The feather �-keratins are a family of
smaller (10.4 kd) molecules than the 13.5 kdscutate scale, beak, and claw family of �-keratins. In contrast, avian reticulate scales(on the plantar surfaces of the foot) are com-posed of nonfeather type b-keratins that aremolecularly similar to those of other reptiles.Long thought to be restricted entirely tobirds, recent reports show that alligator clawkeratin is a 14 kd filament molecularly relatedto the avian scutate scale, claw, and beak �-keratins (Sawyer et al. 2000). Thus, �-keratinsare now known to be broadly distributedwithin archosaurs. Based on outgroup com-parison to alligator claw �-keratin, the longer13.5 filament length of the scutate scale, claw,and beak �-keratins is likely primitive. Evo-lution within the two gene families is typifiedby gene duplication and subsequent deletion(Brush 1993).
Various hypotheses about the evolutionaryorigin of �-keratins and the shorter feather�-keratins have been proposed (reviewed inBrush 1993). In summary, current data indi-cate that the �-keratins originated in an archo-saurian ancestor shared by birds and crocodyl-ians. Subsequently, the �-keratins diversifiedinto a longer (13.5 kd) filament class that iscurrently expressed in avian scutate scales,claw, and beak, and a shorter (10.5 kd) classthat is expressed exclusively in feathers.
Brush (1993, 1996, 2000, 2001) has hypoth-esized that the first feather was composed of10.5 kd filament feather �-keratin. Prum(1999) countered that if this protein evolvedby natural selection for some novel function,the morphological novelty of the featherprobably preceded the current molecularnovelty. Only after there was a novel structurewould there likely be opportunity for somenovel function.
Molecular Basis of FeatherDevelopment and Evolution
An important future goal in the study ofthe origin and evolution of feathers is the elu-cidation of the molecular mechanisms offeather morphogenesis. Feather develop-ment has been a classic model system for thestudy of epithelium-mesenchyme interactionsfor many decades (Sengel 1976). Molecularwork on feather development has also accel-erated (reviewed in Chuong and Widelitz

September 2002 291EVOLUTION AND DIVERSIFICATION OF FEATHERS
1998; Chuong et al. 2000). Nearly all of thisresearch has focused upon the very earlieststages of development—the determination ofthe spatial distribution of feather placodeswithin the pterylae, the establishment of ananterior-posterior axis within the featherplacode, and the elongation of feather buds.These data do contribute to support for thehomology of feather and scale placodes (e.g.,Chuong et al. 1996; Widelitz et al. 1999), butthey do not yet contribute to our understand-ing of the development of the many evolution-arily novel features of feathers.
The bias toward the study of early featherdevelopment reflects the fact that feathershave been studied as a model system of gen-eral mechanisms of mesenchyme-epitheliuminteractions rather than as complex assem-blages of evolutionary novelties. With a fewpreliminary exceptions (Nohno et al. 1995;Ting-Berreth and Chuong 1996; Morgan etal. 1998), little research has been done on themolecular mechanisms of the morphogenesisof barb ridges, the differentiation within abarb ridge among the barbule plates and theramus, the differentiation of the cells of theproximal and distal barbule plates, the for-mation of the rachis, the mechanism of heli-cal growth, and the duplication of helicalgrowth within the follicle in the formation ofthe afterfeather. An essential and excitingfuture direction in the investigation of theevolution of feathers will be to describe thenovel molecular developmental mechanismsthat have contributed to the origin of featherstructure and diversity.
ConclusionsThe origin and diversification of feathers
have been intractable questions in evolution-ary biology for more than a century. Progresson these issues has been hampered by con-ceptual problems and the lack of fossils ofprimitive feathers. Both of these limitationshave been overcome by the recent proposal
of a developmental theory of the origin offeathers, and discoveries of primitive featherfossils from nonavian theropod dinosaurs.The developmental theory provides a heuris-tic model of the evolution and diversificationof feathers that is entirely congruent with theknown details of feather development (Prum1999). The discovery of primitive fossil feath-ers documents that feathers evolved anddiversified in nonavian theropod dinosaursbefore the origin of birds and before the ori-gin of flight (Chen et al. 1998; Ji et al. 1998;Xu et al. 1999a, 1999b, 2000, 2001; Ji et al.2001). These primitive feathers are morpho-logically and phylogenetically congruent withthe predictions of the developmental theory( Ji et al. 2001; Sues 2001; Xu et al. 2001). Thebroad homology of feathers and scales hasbeen nearly universally and uncriticallyaccepted, but it is not strongly supported(Brush 1993, 1996). Scales-based theories offeather evolution have failed to explain theorigin of many feather novelties. Feathersand scales are homologous at the level of theplacode, but all subsequent stages of feathermorphology are derived, evolutionary nov-elties. The hypothesis that feathers evolvedby natural selection for flight is falsified, butnumerous other proposed initial functionsof feathers remain plausible. More specifi-cally, feathers likely originated by selectionfor the growth of an integumentary append-age that emerged from the skin, without con-tinuous investment in the dermis, resulting inthe evolution of the novel tubular feather fol-licle. The tubular feather follicle and feathergerm led to subsequent evolution of numer-ous additional morphological and develop-mental novelties. This complexity appears tohave exploited an inherent capacity of theinnovative tubular form of the feather follicleand feather germ. Future research on theevolution of feather keratin and the molecu-lar mechanisms of feather morphogenesiswill provide additional insights into the inno-vative nature of feathers.
REFERENCES
Baden H P, Maderson P F. 1970. Morphological andbiophysical identification of fibrous proteins inamniote epidermis. Journal of Experimental Biology174:224–232.
Becker R. 1959. Die Strukturanalyse der Gefiederfol-gen von Megapodius freyc. reinw. und ihre Bezie-hung zu der Nestlingsdune. Revue suisse Zoologie66:411–527.

292 Volume 77THE QUARTERLY REVIEW OF BIOLOGY
Bereiter-Hahn J, Matolsky A G, Silviva Richards K, edi-tors. 1986. Biology of the Integument. Volume 2: Verte-brates. Berlin: Springer Verlag.
Blaszyck P. 1935. Untersuchungen uber die Stammes-geschichte der Vogelschuppen und Federn unduber die Abhangigkeit ihrer Ausbildung am Vogel-fuss von der Funktion. Gegenbaurs MorphologisheJahrbuch 75:483–567.
Bock W J. 1965. The role of adaptive mechanisms inthe origin of higher levels of organization. System-atic Zoology 14:272–287.
Broman I. 1941. Uber die Entstehung und Bedeutungder Embryonaldunen. Gegenbaurs MorphologischesJahrbuch 86:141–217.
Brush A H. 1978. Feather keratins. Pages 117–139 inChemical Zoology, Volume 10, edited by M Florkinand B T Scheer. New York: Academic Press.
Brush A H. 1985. Convergent evolution of reticulatescales. Journal of Experimental Zoology 236:303–308.
Brush A H. 1993. The origin of feathers. Pages 121–162 in Avian Biology, Volume IX, edited by D S Far-ner et al. London: Academic Press.
Brush A H. 1996. On the origin of feathers. Journal ofEvolutionary Biology 9:131–142.
Brush A H. 2000. Evolving a protofeather and featherdiversity. American Zoologist 40:631–639.
Brush A H. 2001. The beginnings of feathers. Pages171–179 in New Perspectives on the Origin and EarlyEvolution of Birds, edited by J Gauthier and L F Gall.New Haven (CT): Peabody Museum Special Pub-lications.
Cheng K M, Brush A H. 1984. Feather morphology offour different mutations in the Japanese quail.Poultry Science 63:391–400.
Chen P-J, Dong Z-M, Zhen S-N. 1998. An exceptionallywell-preserved theropod dinosaur from the YixianFormation of China. Nature 391:147–152.
Chiappe L M, Ji Q, Ji S, Norell M A. 1999. Anatomyand systematics of the Confuciusornithidae (Ther-opoda: Aves) from the late Mesozoic of northeast-ern China. Bulletin of the American Museum of Nat-ural History 242.
Chiappe L M, Norell M A, Clark J M. 1996. Phyloge-netic position of Mononykus from the Late Creta-ceous of the Gobi Desert. Memoirs of the QueenslandMuseum 39:557–582.
Chiappe L M, Norell M A, Clark J M. 1998. The skullof a new relative of the stem-group bird Mononykus.Nature 392:275–278.
Chiu C-H, Wagner G P. 2001. A genetic perspective onthe origin of evolutionary novelties. Journal of Mor-phology 248:216.
Chuong C-M. 1993. The making of a feather: hom-eoproteins, retinoids, and adhesion molecules.BioEssays 15:513–521.
Chuong C-M. 1998. Morphogenesis of epithelialappendages: variations on top of a common themeand implications in regeneration. Pages 3–10 in
Molecular Basis of Epithelial Appendage Morphogenesis,edited by C M Chuong. Austin (TX): Landes Bio-science.
Chuong C-M, Chodankar R, Widelitz R B, Jiang T-X.2000. Evo-devo of feathers and scales: buildingcomplex epithelial appendages. Current Opinion inGenetics and Development 10:449–456.
Chuong C-M, Oliver G, Ting S A, Jegalian B G, ChenH M, De Robertis E M. 1990. Gradients of hom-eoproteins in developing feather buds. Development110:1021–1030.
Chuong C-M, Widelitz R B. 1998. Feather morphogen-esis: a model of the formation of epithelial append-ages. Pages 57–74 in Molecular Basis of EpithelialAppendage Morphogenesis, edited by C M Chuong.Austin (TX): Landes Bioscience.
Chuong C-M, Widelitz R B, Ting-Berreth S, Jiang T-X.1996. Early events during avian skin appendageregeneration: dependence on epithelial-mesen-chymal interaction and order of molecular reap-pearance. Journal of Investigative Dermatology 107:639–646.
Cole L J, Hawkins L E. 1930. “Porcupine” pigeons.Journal of Heredity 21:50–60.
Dalton R. 2000. Feathers fly in Beijing. Nature 405:992.Davies H R. 1889. Die Entwicklung der Feder und ihre
Beziehungen zu anderen Integumentgebilden.Morphologisches Jahrbuch 15:560–645.
de Beer G. 1954. Archaeopteryx lithographica: A StudyBased on the British Museum Specimen. London: Trust-ees of the British Museum.
Dhouailly D, Hardy M H, Sengel P. 1980. Formationof feathers on chick foot scales: a stage-dependentmorphogenetic response to retinoic acid. Journalof Embryology and Experimental Morphology 58:63–78.
Dodson P. 2000. Origin of birds: the final solution?American Zoologist 40:504–512.
Dyck J. 1985. The evolution of feathers. ZoologicaScripta 14:137–154.
Ewart J C. 1921. Nestling feathers of the mallard withobservations on the composition, origin, and his-tory of feathers. Proceedings of the Zoological Society:609–653.
Feduccia A. 1985. On why the dinosaurs lacked feath-ers. Pages 75–79 in The Beginnings of Birds, editedby M K Hecht et al. Eichstatt: Freunde des Jura-Museums.
Feduccia A. 1993. Aerodynamic model for the earlyevolution of feathers provided by Propithecus (Pri-mates, Lemuridae). Journal of Theoretical Biology160:159–164.
Feduccia A. 1999. The Origin and Evolution of Birds. Sec-ond Edition. New Haven (CT): Yale UniversityPress.
Fulton J E, Roberts C W, Cheng K M. 1982. Porcupine:a feather structure mutation in Japanese quail.Poultry Science 61:429–433.

September 2002 293EVOLUTION AND DIVERSIFICATION OF FEATHERS
Gatesy S M. 2001. The evolutionary history of the the-ropod caudal locomotor module. Pages 333–347in New Perspectives on the Origin and Early Evolutionof Birds, edited by J Gauthier and J F Gall. NewHaven (CT): Peabody Museum of Natural History.
Gauthier J A. 1986. Saurischian monophyly and theorigin of birds. Memoir of the California Academy ofScience 8:1–55.
Gauthier J A, de Queiroz K. 2001. Feathered dino-saurs, flying dinosaurs, crown dinosaurs and thename “Aves”. Pages 7–41 in New Perspectives on theOrigin and Early Evolution of Birds, edited by J Gau-thier and J F Gall. New Haven (CT): PeabodyMuseum of Natural History.
Gibbons A. 1997. Plucking the feathered dinosaur. Sci-ence 278:1229–1230.
Grau V J. 1994. Biologıa y Patologıa de la Chinchilla. San-tiago (Chile): Ediciones OIKOS.
Griffiths P J. 1996. The isolated Archaeopteryx feather.Archaeopteryx 14:1–26.
Hall B K. 1998. Evolutionary Developmental Biology. Sec-ond Edition. London: Chapman & Hall.
Hall B K. 1999. Homology. Chichester (UK): John Wiley& Sons.
Heilmann G. 1926. The Origin of Birds. London:H. F. G. Whitherby.
Holtz T R. 1994. The phylogenetic position of the Tyr-annosauridae: implications for theropod system-atics. Journal of Paleontology 68:1100–1117.
Holtz T R. 2001. Arctometatarsalia revisited: the prob-lem of homoplasy in reconstructing theropod phy-logeny. Pages 99–123 in New Perspectives on the Ori-gin and Early Evolution of Birds, edited by J Gauthierand J F Gall. New Haven (CT): Peabody Museumof Natural History.
Homberger D G, de Silva K N. 2000. Functional micro-anatomy of the feather-bearing integument: impli-cations for the evolution of birds and avian flight.American Zoologist 40:553–574.
Ji Q, Currie P J, Norell M A, Ji S-A. 1998. Two feath-ered dinosaurs from northeastern China. Nature393:753–761.
Ji Q, Norell M A, Gao K-Q, Ji S-A, Ren D. 2001. Thedistribution of integumentary structures in a feath-ered dinosaur. Nature 410:1084–1088.
Jones T D, Ruben J A, Maderson P F A, Martin L D.2001. Longisquama fossil and feather morphology.Science 291:1899.
Jones T D, Ruben J A, Martin L D, Kurochkin E N,Feduccia A, Maderson P F A, Hillenius W J, GeistN R, Alifanov V. 2000a. Nonavian feathers in a lateTriassic archosaur. Science 288:2202–2205.
Jones T D, Farlow J O, Ruben J A, Henderson D M,Hillenius W J. 2000b. Cursoriality in bipedal archo-saurs. Nature 406:716–718.
Jung H-S, Francis-West P H, Widelitz R B, Jiang T-X,Ting-Berreth S A, Tickle C, Wolpert L, Chuong
C-M. 1998. Local inhibitory action of BMPs andtheir relationships with activators in feather for-mation: implications for periodic patterning. Devel-opmental Biology 196:11–23.
Larson A, Losos J A. 1996. Phylogenetic systematics ofadaptation. Pages 187–220 in Adaptation, edited byG V Lauder and M R Rose. San Diego (CA): Aca-demic Press.
Lauder G V. 1981. Form and function: structural anal-ysis in evolutionary morphology. Paleobiology 7:430–442.
Lauder G V. 1990. Functional morphology and system-atics: studying functional patterns in an historicalcontext. Annual Review of Ecology and Systematics 21:317–340.
Lauder G V. 1994. Homology, form, and function.Pages 151–196 in Homology: The Hierarchical Basis ofComparative Biology, edited by B K Hall. San Diego(CA): Academic Press.
Lauder G V, Liem K F. 1989. The role of historicalfactors in the evolution of complex organismalfunctions. Pages 63–78 in Complex Organismal Func-tions: Integration and Evolution in Vertebrates, editedby D B Wake and G Roth. New York: Wiley Inter-science.
Lauder G V, Rose M R. 1996. Adaptation. San Diego(CA): Academic Press.
Lillie F R, Juhn M. 1938. Physiology of developmentof the feather. II. General principles of develop-ment with special reference to the after-feather.Physiological Zoology 11:434–448.
Lillie F R, Wang H. 1941. Physiology of developmentof the feather. V. Experimental morphogenesis.Physiological Zoology 14:103–133.
Lowe P R. 1935. On the relationship of the Struthi-ones to the dinosaurs and to the rest of the avianclass with special reference to the position ofArchaeopteryx. Ibis 5:398–432.
Lucas A M, Stettenheim P R. 1972. Avian Anatomy—Integument. Agricultural Handbook 362. Washing-ton (DC): Government Printing Office.
Maderson P F A. 1972. On how an archosaur scalemight have given rise to an avian feather. AmericanNaturalist 106:424–428.
Maderson P F A, Alibardi L. 2000. The developmentof the sauropsid integument: a contribution to theproblem of the origin and evolution of feathers.American Zoologist 40:513–529.
Maderson P F A, Homberger D G. 2000. Evolutionaryorigin of feathers: a problem demanding interdis-ciplinary communication. American Zoologist 40:455–460.
Martin L D, Czerkas S A. 2000. The fossil record offeather evolution in the Mesozoic. American Zoolo-gist 40:687–694.
Mayr E. 1960. The emergence of evolutionary novel-ties. Pages 349–380 in The Evolution of Life, editedby S Tax. Chicago: University of Chicago Press.

294 Volume 77THE QUARTERLY REVIEW OF BIOLOGY
Morgan B A, Orkin R W, Noramly S, Perez A. 1998.Stage-specific effects of Sonic Hedgehog expressionin the epidermis. Developmental Biology 201:1–12.
Muller G B, Wagner G P. 1991. Novelty in evolution:restructuring the concept. Annual Review of Ecologyand Systematics 22:229–256.
Nitecki M H. 1990. Evolutionary Innovations. Chicago:University of Chicago Press.
Nohno T, Kawakami Y, Ohuchi H, Fujiwara A, Yosh-ioka H, Noji S. 1995. Involvement of the SonicHedgehog gene in chick feather formation. Biochem-ical and Biophysical Research Communications 206:33–39.
Norell M, Clark J M, Makovicky P J. 2001. Phyloge-netic relationships among coelurisaurian dino-saurs. Pages 49–67 in New Perspectives on the Originand Early Evolution of Birds, edited by J Gauthierand J F Gall. New Haven (CT): Peabody Museumof Natural History.
Norell M, Ji Q, Gao K-Q, Yuan C, Zhao Y, Wang L.2002. ‘Modern’ feathers on a non-avian dinosaur.Nature 416:35–37.
Nowak R M, Paradiso J L. 1983. Walker’s Mammals ofthe World. Fourth Edition. Baltimore (MD): JohnsHopkins University Press.
Ostrom J H. 1974. Archaeopteryx and the origin offlight. Quarterly Review of Biology 49:27–47.
Ostrom J H. 1978. Osteology of Compsognathus longi-pes Wagner. Abhandlungen der Bayerischen Staatssa-mmlung fur Palaeontologie und historische Geologie4:73–118.
Padian K. 1998. When is a bird not a bird? Nature 393:423–433.
Padian K. 2001. Cross-testing adaptive hypotheses:phylogenetic analysis and the origin of bird flight.American Zoologist 41:598–607.
Padian K, Chiappe L M. 1998. The origin and earlyevolution of birds. Biological Reviews 73:1–42.
Parkes K C. 1966. Speculations on the origin of feath-ers. Living Bird 5:77–86.
Patterson C. 1982. Morphological characters andhomology. Pages 21–74 in Problems in PhylogeneticReconstruction, edited by K A Joysey and A E Friday.London: Academic Press.
Pinna M C. 1991. Concepts and tests of homology inthe cladistic paradigm. Cladistics 7:367–394.
Prum R O. 1999. Development and evolutionary ori-gin of feathers. Journal of Experimental Zoology 285:291–306.
Prum R O. 2001. Longisquama fossil and feather mor-phology. Science 291:1899–1900.
Prum R O. 2002. Why ornithologists should careabout the theropod origin of birds. Auk 119:1–17.
Raff R A. 1996. The Shape of Life: Genes, Development andthe Evolution of Animal Form. Chicago: University ofChicago Press.
Regal P J. 1975. The evolutionary origin of feathers.Quarterly Review of Biology 50:33–66.
Reisz R R, Sues H-D. 2000. The ‘feathers’ of Longis-quama. Nature 408:428.
Riddle O. 1908. The cause of the production of “down”and other down-like structures in the plumages ofbirds. Biological Bulletin 14:163–176.
Sawyer R H, Glenn T, French J O, Mays B, Shames RB, Barnes G L, Jr, Rhodes W, Ishikawa Y. 2000. Theexpression of beta-keratins in the epidermalappendages of reptiles and birds. American Zoologist40:530–539.
Sawyer R H, Knapp L W, O’Guin W M. 1986. The skinof birds: epidermis, dermis, and appendages.Pages 194–238 in Biology of the Integument 2: Verte-brates, edited by J Bereiter-Hahn et al. Berlin:Springer-Verlag.
Schweitzer M H. 2001. Evolutionary implications ofpossible protofeather structures associated with aspecimen of Shuvuuia deserti. Pages 181–192 in NewPerspectives on the Origin and Early Evolution of Birds,edited by J Gauthier and J F Gall. New Haven (CT):Peabody Museum of Natural History.
Schweitzer M H, Watt J A, Avci R, Knapp L, ChiappeL, Norell M, Marshall M. 1999. Beta-keratin spe-cific immunological reactivity in feather-like struc-tures of the Cretaceous alvarezsaurid, Shuvuuiadeserti. Journal of Experimental Zoology 285:146–157.
Sengel P. 1976. Morphogenesis of Skin. Cambridge: Cam-bridge University Press.
Sereno P. 1997. The origin and evolution of dinosaurs.Annual Review of Earth and Planetary Science 25:435–489.
Serano P. 1999. The evolution of dinosaurs. Science284:2137–2147.
Sereno P. 2001. Alvarezsaurids: birds or ornithomi-mosaurs? Pages 69–98 in New Perspectives on the Ori-gin and Early Evolution of Birds, edited by J Gauthierand J F Gall. New Haven (CT): Peabody Museumof Natural History.
Sharov A G. 1970. An unusual reptile from the lowerTriassic of Fergana. Paleontological Journal 1970:127–130.
Somes R G, Jr. 1990. Mutations and major variants ofplumage and skin in chickens. Pages 169–208 inPoultry Breeding and Genetics, edited by R D Craw-ford. Amsterdam: Elsevier.
Steghaus-Kovac S. 2002. Tug-of-war over mystery fossil.Science 295:1212–1213.
Steiner H. 1917. Das Problem der Diastataxie desVogelflugels. Jenaische Zeitschrift fur Naturwissen-schaft 55:222–496.
Stokstad E. 2000. Feathers, or flight of fancy? Science288:2124–2125.
Stokstad E. 2001a. Exquisite Chinese fossils add newpages to book of life. Science 291:232–236.
Stokstad E. 2001b. Mysterious e-photos vex paleontol-ogists. Science 293:1573.
Strong R M. 1902a. A case of abnormal plumage. Bio-logical Bulletin 3:289–294.

September 2002 295EVOLUTION AND DIVERSIFICATION OF FEATHERS
Strong R M. 1902b. The development of color in thedefinitive feather. Bulletin of the Museum of Compar-ative Zoology 40:147–185.
Sues H-D. 2001. Ruffling feathers. Nature 410:1036–1037.
Ting-Berreth S A, Chuong C-M. 1996. Sonic Hedgehogin feather morphogenesis: induction of mesenchy-mal condensation and association with cell death.Developmental Dynamics 207:157–170.
Unwin D M, Benton M J. 2001. Longisquama fossil andfeather morphology. Science 291:1900–1901.
Wagner G P. 2000. What is the premise of develop-mental evolution? Part I: why is developmentalbiology necessary to explain evolutionary innova-tions? Journal of Experimental Zoology 288:95–98.
Wagner G P. 2001. The Character Concept in EvolutionaryBiology. San Diego (CA): Academic Press.
Wagner G P, Chiu C-H, Laubichler M. 2000. Devel-opmental evolution as a mechanistic science: theinference from developmental mechanisms to evo-lutionary processes. American Zoologist 40:819–831.
Wagner G P, Schwenk K. 2000. Evolutionarily stableconfigurations: functional integration and the evo-lution of phenotypic stability. Pages 155–217 inEvolutionary Biology, Volume 31, edited by M KHecht et al. New York: Kluwer/Plenum.
Wang X, Zhou Z, Zhang F, Xu X. 2002. A nearly com-plete articulated rhamphorhynchoid pterosaurwith exceptionally well-preserved wing membranesand “hairs” from Inner Mongolia, northeast China.Chinese Science Bulletin 47:226–230.
Waters N F. 1967. “Porcupine” chickens. Journal ofHeredity 58:163–164.
Wellnhofer P. 1996. The Illustrated Encyclopedia of Pre-historic Flying Reptiles. New York: Barnes & Noble.
Widelitz R B, Jiang T-X, Chen C J, Stott N S, Jung H-S,
Chuong C-M. 1999. Wnt-7a in feather morphogen-esis: involvement of anterior-posterior asymmetryand proximal-distal elongation demonstrated withan in vitro reconstitution model. Development 126:2577–2587.
Widelitz R B, Jiang T-X, Lu J-F, Chuong C-M. 2000.Beta-catenin in epithelial morphogenesis: con-version of part of avian foot scales into featherbuds with beta-catenin. Developmental Biology 219:98–114.
Xu X, Tang Z-l, Wang X-l. 1999a. A therizinosauroiddinosaur with integumentary structures fromChina. Nature 399:350–354.
Xu X, Wang X-l, Wu X-C. 1999b. A dromaeosauriddinosaur with a filamentous integument from theYixian Formation of China. Nature 401: 262–266.
Xu X, Zhou Z, Prum R O. 2001. Branched integu-mental structures in Sinornithosaurus and the ori-gin of feathers. Nature 410:200–204.
Xu X, Zhou Z, Wang X. 2000. The smallest knownnon-avian theropod dinosaur. Nature 408:705–708.
Zhang F, Zhou Z. 2000. A primitive enantiornithinebird and the origin of feathers. Science 290:1955–1959.
Zhou Z. 2000. Caudipteryx and the origin of flight inbirds. Vertebrata PalAsiatica 38(Supplement):38.
Zhou Z, Wang X-L. 2000. A new species of Caudipteryxfrom the Yixian Formation of Liaoning, northeastChina. Vertebrata PalAsiatica 38:113–127.
Zhou Z, Wang X-L, Xu X. 2000. Important featuers ofCaudipteryx—evidence from two nearly completenew specimens. Vertebrata PalAsiatica 38:241–254.
Zou H, Niswander L. 1996. Requirement of BMP sig-naling in interdigital apoptosis and scale forma-tion. Science 272:738–741.