99nsfgrant
DESCRIPTION
poTRANSCRIPT

INFORMATION ABOUT PRINCIPAL INVESTIGATORS/PROJECT DIRECTORS(PI/PD) andco-PRINCIPAL INVESTIGATORS/co-PROJECT DIRECTORS
Submit only ONE copy of this form for each PI/PD and co-PI/PD identified on the proposal. The form(s) should be attached to the originalproposal as specified in GPG Section II.B. Submission of this information is voluntary and is not a precondition of award. This information willnot be disclosed to external peer reviewers. DO NOT INCLUDE THIS FORM WITH ANY OF THE OTHER COPIES OF YOUR PROPOSAL ASTHIS MAY COMPROMISE THE CONFIDENTIALITY OF THE INFORMATION.
PI/PD Name:
Gender: Male Female
Ethnicity: (Choose one response) Hispanic or Latino Not Hispanic or Latino
Race: (Select one or more)
American Indian or Alaska Native
Asian
Black or African American
Native Hawaiian or Other Pacific Islander
White
Disability Status: (Select one or more)
Hearing Impairment
Visual Impairment
Mobility/Orthopedic Impairment
Other
None
Citizenship: (Choose one) U.S. Citizen Permanent Resident Other non-U.S. Citizen
Check here if you do not wish to provide any or all of the above information (excluding PI/PD name):
REQUIRED: Check here if you are currently serving (or have previously served) as a PI, co-PI or PD on any federally fundedproject
Ethnicity Definition:Hispanic or Latino. A person of Mexican, Puerto Rican, Cuban, South or Central American, or other Spanish culture or origin, regardlessof race.Race Definitions:American Indian or Alaska Native. A person having origins in any of the original peoples of North and South America (including Central America), and who maintains tribal affiliation or community attachment.Asian. A person having origins in any of the original peoples of the Far East, Southeast Asia, or the Indian subcontinent including, for example, Cambodia, China, India, Japan, Korea, Malaysia, Pakistan, the Philippine Islands, Thailand, and Vietnam.Black or African American. A person having origins in any of the black racial groups of Africa.Native Hawaiian or Other Pacific Islander. A person having origins in any of the original peoples of Hawaii, Guam, Samoa,or other Pacific Islands.White. A person having origins in any of the original peoples of Europe, the Middle East, or North Africa.
WHY THIS INFORMATION IS BEING REQUESTED:
The Federal Government has a continuing commitment to monitor the operation of its review and award processes to identify and addressany inequities based on gender, race, ethnicity, or disability of its proposed PIs/PDs. To gather information needed for this importanttask, the proposer should submit a single copy of this form for each identified PI/PD with each proposal. Submission of the requestedinformation is voluntary and will not affect the organization’s eligibility for an award. However, information not submitted will seriously underminethe statistical validity, and therefore the usefulness, of information recieved from others. Any individual not wishing to submit some or all theinformation should check the box provided for this purpose. (The exceptions are the PI/PD name and the information about prior Federal support, thelast question above.)
Collection of this information is authorized by the NSF Act of 1950, as amended, 42 U.S.C. 1861, et seq. Demographic data allows NSF togauge whether our programs and other opportunities in science and technology are fairly reaching and benefiting everyone regardless ofdemographic category; to ensure that those in under-represented groups have the same knowledge of and access to programs and otherresearch and educational oppurtunities; and to assess involvement of international investigators in work supported by NSF. The informationmay be disclosed to government contractors, experts, volunteers and researchers to complete assigned work; and to other governmentagencies in order to coordinate and assess programs. The information may be added to the Reviewer file and used to select potentialcandidates to serve as peer reviewers or advisory committee members. See Systems of Records, NSF-50, "Principal Investigator/ProposalFile and Associated Records", 63 Federal Register 267 (January 5, 1998), and NSF-51, "Reviewer/Proposal File and Associated Records",63 Federal Register 268 (January 5, 1998).
NSF Form 1225(10/99)
Stanley A Cohn

List of Suggested Reviewers or Reviewers Not To Include (optional)
SUGGESTED REVIEWERS:Dr. R. GordonDept. of Botany and RadiologyUniv. of ManitobaWinnepeg R3T 2N2 Canada204-787-1076 (tel)e-mail: [email protected]
Dr. Michael R. GretzDepartment of Biological SciencesMichigan Technological University1400 Townsend Dr. Houghton, MI 49931-1295Phone: 906-487-3175 Fax: 906-487-3167e-mail: [email protected]
Prof. Dr. Donat-P. H derFriedrich-Alexander UniversitatInstitut fur Botanik und Pharmazeutische BiologieStaudtstr. 5, D-91058, ErlangenFRG09131-858216 (tel)09131-858215 (fax)e-mail: [email protected]
Dr. Kyle D. HoaglandUniversity of Nebraska - LincolnSchool of Natural ResourcesDept. of Forestries, Fisheries & WildlifeLincoln, Nebraska 68583-0814Phone: (402) 472-8182Email: [email protected] ([email protected])
Dr. David G. MannDeputy Director - Royal Botanic Garden20A Inverleith RowEndinburgh, EH3 5LR, Scotland UK031-552-7171 (tel)031-552-0382 (fax)e-mail: [email protected]
Dr. E.F. StoermerE. F. StoermerCenter for Great Lakes501 East UniversityUniversity of Michigan

List of Suggested Reviewers or Reviewers Not To Include (optional)
Suggested Reviewers contd...
Ann Arbor, MI 48109-1090Phone 734-764-5238FAX 734-647-2748e-mail: [email protected]
Dr. C. W. SullivanDept. of Biological SciencesUniversity of Southern CaliforniaBovard (ADM) - Room 203 Los Angeles, CA 90089Phone 213-740-6709 FAX 213-740-8919 e-mail: [email protected]
Dr. M. J. SullivanDept. of Biology - Box GYMississippi State Univ.Mississippi State, MS 39762601-325-3120 (tel)e-mail: [email protected]
REVIEWERS NOT TO INCLUDE:

COVER SHEET FOR PROPOSAL TO THE NATIONAL SCIENCE FOUNDATIONFOR NSF USE ONLY
NSF PROPOSAL NUMBER
DATE RECEIVED NUMBER OF COPIES DIVISION ASSIGNED FUND CODE DUNS# (Data Universal Numbering System) FILE LOCATION
FOR CONSIDERATION BY NSF ORGANIZATION UNIT(S) (Indicate the most specific unit known, i.e. program, division, etc.)
PROGRAM ANNOUNCEMENT/SOLICITATION NO./CLOSING DATE/if not in response to a program announcement/solicitation enter NSF 99-2
EMPLOYER IDENTIFICATION NUMBER (EIN) ORTAXPAYER IDENTIFICATION NUMBER (TIN)
SHOW PREVIOUS AWARD NO. IF THIS ISA RENEWALAN ACCOMPLISHMENT-BASED RENEWAL
IS THIS PROPOSAL BEING SUBMITTED TO ANOTHER FEDERALAGENCY? YES NO IF YES, LIST ACRONYMS(S)
NAME OF ORGANIZATION TO WHICH AWARD SHOULD BE MADE ADDRESS OF AWARDEE ORGANIZATION, INCLUDING 9 DIGIT ZIP CODE
AWARDEE ORGANIZATION CODE (IF KNOWN)
IS AWARDEE ORGANIZATION (Check All That Apply)(See GPG II.D.1 For Definitions) FOR-PROFIT ORGANIZATION SMALL BUSINESS MINORITY BUSINESS WOMAN-OWNED BUSINESS
NAME OF PERFORMING ORGANIZATION, IF DIFFERENT FROM ABOVE ADDRESS OF PERFORMING ORGANIZATION, IF DIFFERENT, INCLUDING 9 DIGIT ZIP CODE
PERFORMING ORGANIZATION CODE (IF KNOWN)
TITLE OF PROPOSED PROJECT
REQUESTED AMOUNT
$
PROPOSED DURATION (1-60 MONTHS)
months
REQUESTED STARTING DATE SHOW RELATED PREPROPOSAL NO.,IF APPLICABLE
CHECK APPROPRIATE BOX(ES) IF THIS PROPOSAL INCLUDES ANY OF THE ITEMS LISTED BELOWBEGINNING INVESTIGATOR (GPG 1.A.3)
DISCLOSURE OF LOBBYING ACTIVITIES (GPG II.D.1)
PROPRIETARY & PRIVILEGED INFORMATION (GPG II.D.10)
NATIONAL ENVIRONMENTAL POLICY ACT (GPG II.D.10)
HISTORIC PLACES (GPG II.D.10)
SMALL GRANT FOR EXPLOR. RESEARCH (SGER) (GPG II.D.12)
GROUP PROPOSAL (GPG II.D.12)
VERTEBRATE ANIMALS (GPG II.D.12) IACUC App. Date
HUMAN SUBJECTS (GPG II.D.12)Exemption Subsection or IRB App. Date
INTERNATIONAL COOPERATIVE ACTIVITIES: COUNTRY/COUNTRIES
FACILITATION FOR SCIENTISTS/ENGINEERS WITH DISABILITIES (GPG V.G.)
RESEARCH OPPORTUNITY AWARD (GPG V.H)
PI/PD DEPARTMENT PI/PD POSTAL ADDRESS
PI/PD FAX NUMBER
NAMES (TYPED) High Degree Yr of Degree Telephone Number Electronic Mail Address
PI/PD NAME
CO-PI/PD
CO-PI/PD
CO-PI/PD
CO-PI/PD
NSF Form 1207 (10/98) Page 1 of 2
9982897Program - ECOLOGICAL & EVOLUTIONARY PHYS
NSF94-79 07/10/99
DePaul University
0016717000
DePaul University1 East Jackson BoulevardChicago, IL. 606042218
Physiological Ecology of Diatom Motility and Adhesion
289,784 36 01/01/00
Department of Biological Sciences
773-325-7596
DePaul University2325 N. Clifton AvenueChicago, IL 606143238United States
Stanley A Cohn Ph.D. 1986 773-325-7597 [email protected]
825753379

CERTIFICATION PAGE
Certification for Principal Investigators and Co-Principal Investigators:I certify to the best of my knowledge that: (1) the statements herein (excluding scientific hypotheses and scientific opinions) are true and complete, and(2) the text and graphics herein as well as any accompanying publications or other documents, unless otherwise indicated, are the original work of thesignatories or individuals working under their supervision. I agree to accept responsibility for the scientific conduct of the project and to provide therequired progress reports if an award is made as a result of this application. I understand that the willful provision of false information or concealing a material fact in this proposal or any other communication submitted to NSF is acriminal offense (U.S.Code, Title 18, Section 1001).
Name (Typed) Signature Social Security No.* Date
PI/PD
Co-PI/PD
Co-PI/PD
Co-PI/PD
Co-PI/PD
Certification for Authorized Organizational Representative or Individual Applicant:By signing and submitting this proposal, the individual applicant or the authorized official of the applicant institution is: (1) certifying thatstatements made herein are true and complete to the best of his/her knowledge; and (2) agreeing to accept the obligation to comply with NSFaward terms and conditions if an award is made as a result of this application. Further, the applicant is hereby providing certificationsregarding Federal debt status, debarment and suspension, drug-free workplace, and lobbying activities (see below), as set forth in GrantProposal Guide (GPG), NSF 99-2. Willful provision of false information in this application and its supporting documents or in reports requiredunder an ensuring award is a criminal offense (U. S. Code, Title 18, Section 1001). In addition, if the applicant institution employs more than fifty persons, the authorized official of the applicant institution is certifying that the institution has implemented a written and enforced conflict of interest policy that is consistent with the provisions of Grant Policy Manual Section 510; that to the bestof his/her knowledge, all financial disclosures required by that conflict of interest policy have been made; and that all identified conflicts of interest will havebeen satisfactorily managed, reduced or eliminated prior to the institution’s expenditure of any funds under the award, in accordance with theinstitution’s conflict of interest policy. Conflict which cannot be satisfactorily managed, reduced or eliminated must be disclosed to NSF.
Debt and Debarment Certifications (If answer "yes" to either, please provide explanation.)
Is the organization delinquent on any Federal debt? Yes No Is the organization or its principals presently debarred, suspended, proposed for debarment, declared ineligible, or voluntarily excluded from covered transactions by any Federal department or agency? Yes No
Certification Regarding LobbyingThis certification is required for an award of a Federal contract, grant, or cooperative agreement exceeding $100,000 and for an award of a Federal loan ora commitment providing for the United States to insure or guarantee a loan exceeding $150,000.
Certification for Contracts, Grants, Loans and Cooperative AgreementsThe undersigned certifies, to the best of his or her knowledge and belief, that:
(1) No federal appropriated funds have been paid or will be paid, by or on behalf of the undersigned, to any person for influencing or attempting to influencean officer or employee of any agency, a Member of Congress, an officer or employee of Congress, or an employee of a Member of Congress in connectionwith the awarding of any federal contract, the making of any Federal grant, the making of any Federal loan, the entering into of any cooperative agreement,and the extension, continuation, renewal, amendment, or modification of any Federal contract, grant, loan, or cooperative agreement.
(2) If any funds other than Federal appropriated funds have been paid or will be paid to any person for influencing or attempting to influence an officer oremployee of any agency, a Member of Congress, an officer or employee of Congress, or an employee of a Member of Congress in connection with thisFederal contract, grant, loan, or cooperative agreement, the undersigned shall complete and submit Standard Form-LLL, ‘‘Disclosure Form to ReportLobbying,’’ in accordance with its instructions.
(3) The undersigned shall require that the language of this certification be included in the award documents for all subawards at all tiers includingsubcontracts, subgrants, and contracts under grants, loans, and cooperative agreements and that all subrecipients shall certify and disclose accordingly.
This certification is a material representation of fact upon which reliance was placed when this transaction was made or entered into. Submission of thiscertification is a prerequisite for making or entering into this transaction imposed by section 1352, title 31, U.S. Code. Any person who fails to file therequired certification shall be subject to a civil penalty of not less than $10,000 and not more than $100,000 for each such failure.AUTHORIZED ORGANIZATIONAL REPRESENTATIVE SIGNATURE DATE
NAME/TITLE (TYPED)
TELEPHONE NUMBER ELECTRONIC MAIL ADDRESS FAX NUMBER
*SUBMISSION OF SOCIAL SECURITY NUMBERS IS VOLUNTARY AND WILL NOT AFFECT THE ORGANIZATION’S ELIGIBILITY FOR AN AWARD. HOWEVER, THEY ARE ANINTEGRAL PART OF THE INFORMATION SYSTEM AND ASSIST IN PROCESSING THE PROPOSAL. SSN SOLICITED UNDER NSF ACT OF 1950, AS AMENDED.
Page 2 of 2
Stanley A CohnS
SN
s are confid
ential
and
are not d
isplayed
*ON
FA
ST
LA
NE
SU
BM
ISS
ION
S*
Marjorie P. Piechowski, Ph.D., Director 07/09/99
773-325-2595 [email protected] 773-325-7389

Page 1
Directorate for Biological SciencesDivision of Integrative Organismal Biology
Ecological & Evolutionary Physiology
Proposal Classification FormPI: Cohn, Stanley / Proposal Number: 9982897
CATEGORY I: INVESTIGATOR STATUS (Select ONE) Beginning Investigator - No previous Federal support as PI or Co-PI, excluding fellowships, dissertations, planning grants, etc.
Prior Federal support only
Current Federal support only
Current & prior Federal support
CATEGORY II: FIELDS OF SCIENCE OTHER THAN BIOLOGY INVOLVED IN THIS RESEARCH (Select 1 to 3)
Astronomy
Chemistry
Computer Science
Earth Science
Engineering
Mathematics
Physics
Psychology
Social Sciences
None of the Above
CATEGORY III: SUBSTANTIVE AREA (Select 1 to 4) BEHAVIORAL STUDIES
BIOENGINEERING
BIOGEOGRAPHY
Island Biogeography
Historical/ Evolutionary Biogeography
Phylogeography
Methods/Theory
BIOMATERIALS
BIOTECHNOLOGY
Animal Biotechnology
Plant Biotechnology
Environmental Biotechnology
Marine Biotechnology
Metabolic Engineering
CHRONOBIOLOGY
COGNITIVE NEUROSCIENCE
COMMUNITY ECOLOGY
Community Analysis
Community Structure
Community Stability
Succession
Experimental Microcosms/ Mesocosms
Disturbance
Deforestation
Patch Dynamics
Food Webs/ Trophic Structure
Keystone Species
COMPARATIVE APPROACHES
COMPUTATIONAL BIOLOGY
CONSERVATION & RESTORATION BIOLOGY
CORAL REEFS
CURATION
DATABASES
DEVELOPMENTAL BIOLOGY
ECOSYSTEMS LEVEL
Physical Structure
Decomposition
Biogeochemistry
Limnology/Hydrology
Climate/Microclimate
Whole-System Analysis
Productivity/Biomass
System Energetics
Landscape Dynamics
Chemical & Biochemical Control
Global Change
Climate Change
Regional Studies
Global Studies
Forestry
Resource Management (Wildlife, Fisheries, Range, Other)
Agricultural Ecology
ENDOCRINE DISRUPTORS/ ENVIRONMENTAL ENDOCRINOLOGY
EPIGENETICS
EXTREMOPHILES
GENOMICS (Genome sequence, organization, function)
Viral
Microbial
Fungal
Plant
Animal
HUMAN NUTRITION
INFORMATICS
MARINE MAMMALS
MOLECULAR APPROACHES
Molecular Evolution
NANOSCIENCE
ORGANISMAL SYSTEMS
Physiological Approaches Metabolic Processes
Hormonal Regulation/ Integration
Stress Responses
Sensory Biology
Movement Studies
PALEONTOLOGY
Floristic

Page 2
Faunistic
Paleoecology
Biostratigraphy
Palynology
Micropaleontology
Paleoclimatology
Archeozoic
Paleozoic
Mesozoic
Cenozoic
PHOTOSYNTHESIS
PLANT BIOLOGY
Arabidopsis-Related Plant Research
POPULATION DYNAMICS & LIFE HISTORY
Demography/ Life History
Population Cycles
Distribution/Patchiness/ Marginal Populations
Population Regulation
Intraspecific Competition
Reproductive Strategies
Gender Allocation
Metapopulations
Extinction
POPULATION GENETICS & BREEDING SYSTEMS
Variation
Microevolution
Speciation
Hybridization
Inbreeding/Outbreeding
Gene Flow Measurement
Inheritance/Heritability
Quantitative Genetics/ QTL Analysis
Ecological Genetics
Gender Ratios
Apomixis/ Parthenogenesis
Vegetative Reproduction
REPRODUCTIVE ANIMAL BIOLOGY
SPECIES INTERACTIONS
Predation
Herbivory
Omnivory
Interspecific Competition
Niche Relationships/ Resource Partititioning
Pollination/ Seed Dispersal
Parasitism
Mutualism/ Commensalism
Plant/Fungal/ Microbial Interactions
Mimicry
Animal Pathology
Plant Pathology
Coevolution
Biological Control
SPINAL CORD/ NERVE REGENERATION
STATISTICS & MODELING
Methods/ Instrumentation/ Software
Modeling (general) Modeling of Biological or Molecular Systems
Computational Modeling
Statistics (general) Multivariate Methods
Spatial Statistics & Spatial Modeling
Sampling Design & Analysis
Experimental Design & Analysis
STRUCTURAL BIOLOGY
SYSTEMATICS
Taxonomy/Classification
Nomenclature
Monograph/Revision
Phylogenetics
Phenetics/Cladistics/ Numerical Taxonomy
Macroevolution
NONE OF THE ABOVE
CATEGORY IV: INFRASTRUCTURE (Select 1 to 3) COLLECTIONS/STOCK CULTURES
Collection Enhancement
Collection Refurbishment
Living Organism Stock Cultures
Natural History Collections
DATABASES
Database Initiation
Database Enhancement
Database Maintenance & Curation
Database Methods
FACILITIES
Controlled Environment Facilities
Field Stations Field Facility Structure
Field Facility Equipment
LTER Site
GENOME SEQUENCING Arabidopsis Genome Sequencing
Other Plant Genome Sequencing
INDUSTRY PARTICIPATION
INSTRUMENTATION
Instrument Development
Instrument Acquisition
Computational Hardware Development/Acquisition
TOOLS DEVELOPMENT
Analytical Algorithm Development
Other Software Development
Informatics Tool Development
Technique Development
TRACKING SYSTEMS
Geographic Information Systems
Remote Sensing
TRAINING
Multi-, Cross-, Interdisciplinary Training
NONE OF THE ABOVE
CATEGORY V: HABITAT (Select 1 to 2)
TERRESTRIAL HABITATS GENERAL TERRESTRIAL
TUNDRA
BOREAL FOREST
TEMPERATE
Deciduous Forest
Coniferous Forest
Rain Forest
Mixed Forest
Prairie/Grasslands
Desert
SUBTROPICAL Rain Forest
Seasonal Forest
Savanna
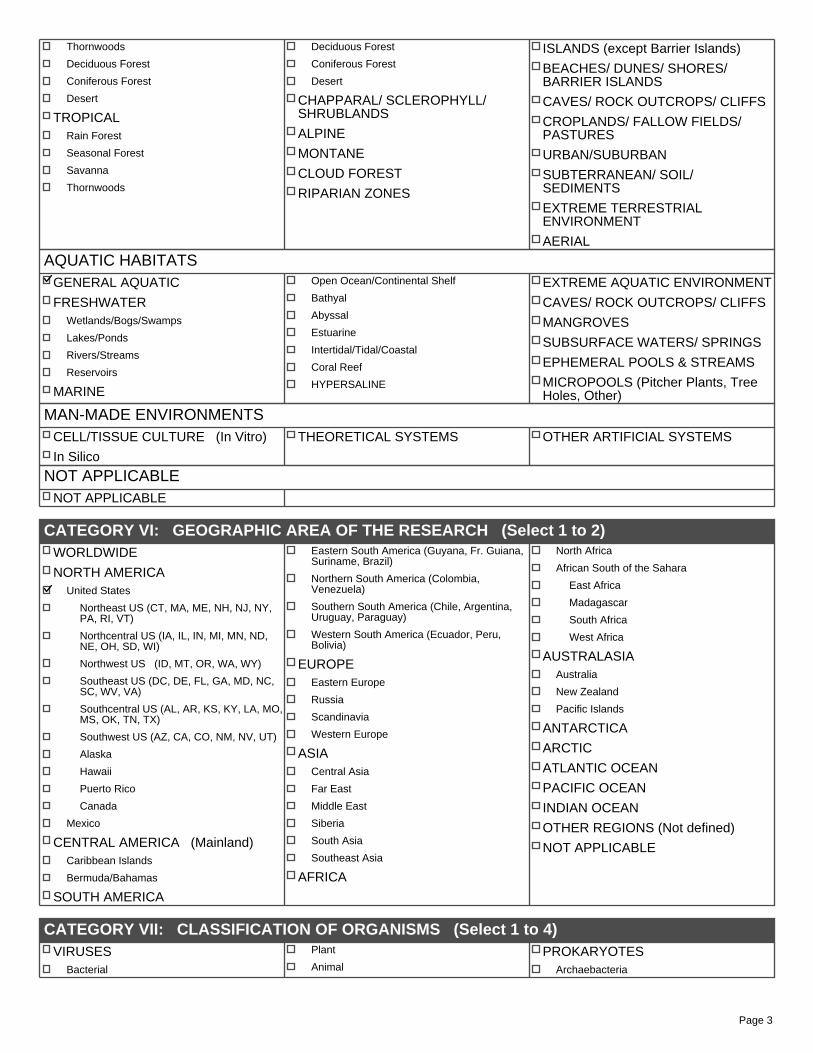
Page 3
Thornwoods
Deciduous Forest
Coniferous Forest
Desert
TROPICAL Rain Forest
Seasonal Forest
Savanna
Thornwoods
Deciduous Forest
Coniferous Forest
Desert
CHAPPARAL/ SCLEROPHYLL/ SHRUBLANDS
ALPINE
MONTANE
CLOUD FOREST
RIPARIAN ZONES
ISLANDS (except Barrier Islands)
BEACHES/ DUNES/ SHORES/ BARRIER ISLANDS
CAVES/ ROCK OUTCROPS/ CLIFFS
CROPLANDS/ FALLOW FIELDS/ PASTURES
URBAN/SUBURBAN
SUBTERRANEAN/ SOIL/ SEDIMENTS
EXTREME TERRESTRIAL ENVIRONMENT
AERIAL
AQUATIC HABITATS GENERAL AQUATIC
FRESHWATER Wetlands/Bogs/Swamps
Lakes/Ponds
Rivers/Streams
Reservoirs
MARINE
Open Ocean/Continental Shelf
Bathyal
Abyssal
Estuarine
Intertidal/Tidal/Coastal
Coral Reef
HYPERSALINE
EXTREME AQUATIC ENVIRONMENT
CAVES/ ROCK OUTCROPS/ CLIFFS
MANGROVES
SUBSURFACE WATERS/ SPRINGS
EPHEMERAL POOLS & STREAMS
MICROPOOLS (Pitcher Plants, Tree Holes, Other)
MAN-MADE ENVIRONMENTS CELL/TISSUE CULTURE (In Vitro)
In Silico
THEORETICAL SYSTEMS OTHER ARTIFICIAL SYSTEMS
NOT APPLICABLE NOT APPLICABLE
CATEGORY VI: GEOGRAPHIC AREA OF THE RESEARCH (Select 1 to 2) WORLDWIDE
NORTH AMERICA United States
Northeast US (CT, MA, ME, NH, NJ, NY, PA, RI, VT)
Northcentral US (IA, IL, IN, MI, MN, ND, NE, OH, SD, WI)
Northwest US (ID, MT, OR, WA, WY)
Southeast US (DC, DE, FL, GA, MD, NC, SC, WV, VA)
Southcentral US (AL, AR, KS, KY, LA, MO,MS, OK, TN, TX)
Southwest US (AZ, CA, CO, NM, NV, UT)
Alaska
Hawaii
Puerto Rico
Canada
Mexico
CENTRAL AMERICA (Mainland) Caribbean Islands
Bermuda/Bahamas
SOUTH AMERICA
Eastern South America (Guyana, Fr. Guiana, Suriname, Brazil)
Northern South America (Colombia, Venezuela)
Southern South America (Chile, Argentina, Uruguay, Paraguay)
Western South America (Ecuador, Peru, Bolivia)
EUROPE Eastern Europe
Russia
Scandinavia
Western Europe
ASIA Central Asia
Far East
Middle East
Siberia
South Asia
Southeast Asia
AFRICA
North Africa
African South of the Sahara
East Africa
Madagascar
South Africa
West Africa
AUSTRALASIA Australia
New Zealand
Pacific Islands
ANTARCTICA
ARCTIC
ATLANTIC OCEAN
PACIFIC OCEAN
INDIAN OCEAN
OTHER REGIONS (Not defined)
NOT APPLICABLE
CATEGORY VII: CLASSIFICATION OF ORGANISMS (Select 1 to 4) VIRUSES
Bacterial
Plant
Animal PROKARYOTES
Archaebacteria

Page 4
Cyanobacteria
Eubacteria
PROTISTA (PROTOZOA) Amoebae
Apicomplexa
Ciliophora
Flagellates
Foraminifera
Microspora
Radiolaria
FUNGI Ascomycota
Basidiomycota
Chytridiomycota
Mitosporic Fungi
Oomycota
Yeasts
Zygomycota
LICHENS
SLIME MOLDS
ALGAE Bacillariophyta (Diatoms)
Charophyta
Chlorophyta
Chrysophyta
Dinoflagellata
Euglenoids
Phaeophyta
Rhodophyta
PLANTS N0N-VASCULAR PLANTS
BRYOPHYTA
Anthocerotae (Hornworts)
Hepaticae (Liverworts)
Musci (Mosses)
VASCULAR PLANTS
FERNS & FERN ALLIES
GYMNOSPERMS
Coniferales (Conifers)
Cycadales (Cycads)
Ginkgoales (Ginkgo)
Gnetales (Gnetophytes)
ANGIOSPERMS
Monocots
Arecaceae (Palmae)
Cyperaceae
Liliaceae
Orchidaceae
Poaceae (Graminae)
Dicots
Apiaceae (Umbelliferae)
Asteraceae (Compositae)
Brassicaceae (Cruciferae)
Fabaceae (Leguminosae)
Lamiaceae (Labiatae)
Rosaceae
Solanaceae
ANIMALS INVERTEBRATES
MESOZOA/PLACOZOA
PORIFERA (Sponges)
CNIDARIA
Hydrozoa (Hydra, etc.)
Scyphozoa (Jellyfish)
Anthozoa (Corals, Sea Anemones)
CTENOPHORA (Comb Jellies)
PLATYHELMINTHES (Flatworms)
Turbellaria (Planarians)
Trematoda (Flukes)
Cestoda (Tapeworms)
Monogenea (Flukes)
GNATHOSTOMULIDA
NEMERTINEA (Rynchocoela) (Ribbon Worms)
ENTOPROCTA (Bryozoa) (Plant-like Animals)
ASCHELMINTHES
Gastrotricha
Kinorhyncha
Loricifera
Nematoda (Roundworms)
Nematomorpha (Horsehair Worms)
Rotifera (Rotatoria)
ACANTHOCEPHALA (Spiny-headed Worms)
PRIAPULOIDEA
BRYOZOA (Ectoprocta) (Plant-like Animals)
PHORONIDEA (Lophophorates)
BRACHIOPODA (Lamp Shells)
MOLLUSCA
Monoplacophora
Aplacophora (Solenogasters)
Polyplacophora (Chitons)
Scaphopoda (Tooth Shells)
Gastropoda (Snails, Slugs, Limpets)
Pelecypoda (Bivalvia) (Clams, Mussels, Oysters, Scallops)
Cephalopoda (Squid, Octopus, Nautilus)
ANNELIDA (Segmented Worms)
Polychaeta (Parapodial Worms)
Oligochaeta (Earthworms)
Hirudinida (Leeches)
POGONOPHORA (Beard Worms)
SIPUNCULOIDEA (Peanut Worms)
ECHIUROIDEA (Spoon Worms)
ARTHROPODA
Cheliceriformes
Merostomata (Horseshoe Crabs)
Pycnogonida (Sea Spiders)
Scorpionida (Scorpions)
Araneae (True Spiders)
Pseudoscorpionida (Pseudoscorpions)
Acarina (Free-living Mites)
Parasitiformes (Parasitic Ticks & Mites)
Crustacea
Branchiopoda (Fairy Shrimp, Water Flea)
Ostracoda (Sea Lice)
Copepoda
Cirripedia (Barnacles)
Amphipoda (Skeleton Shrimp, Whale Lice, Freshwater Shrimp)
Isopoda (Wood Lice, Pillbugs)
Decapoda (Lobster, Crayfish, Crabs, Shrimp)
Hexapoda (Insecta) (Insects)
Apterygota (Springtails, Silverfish, etc.)
Odonata (Dragonflies, Damselflies)
Ephemeroptera (Mayflies)
Orthoptera (Grasshoppers, Crickets)
Dictyoptera (Cockroaches, Mantids, Phasmids)
Isoptera (Termites)
Plecoptera (Stoneflies)
Phthiraptera (Mallophaga & Anoplura) (Lice)
Hemiptera (including Heteroptera) (True Bugs)
Homoptera (Cicadas, Scale Insects,Leafhoppers)
Thysanoptera (Thrips)
Neuroptera (Lacewings, Dobsonflies, Snakeflies)
Trichoptera (Caddisflies)
Lepidoptera (Moths, Butterflies)
Diptera (Flies, Mosquitoes)
Siphonaptera (Fleas)
Coleoptera (Beetles)
Hymenoptera (Ants, Bees, Wasps, Sawflies)
Chilopoda (Centipedes)
Diplopoda (Millipedes)
Pauropoda
Symphyta (Symphyla)
PENTASTOMIDA (Linguatulida) (Tongue Worms)
TARDIGRADA (Tardigrades, Water Bears)
ONYCHOPHORA (Peripatus)
CHAETOGNATHA (Arrow Worms)
ECHINODERMATA
Crinoidea (Sea Lilies, Feather Stars)
Asteroidea (Starfish, Sea Stars)

Page 5
Ophiuroidea (Brittle Stars, Serpent Stars)
Echinoidea (Sea Urchins, Sand Dollars)
Holothuroidea (Sea Cucumbers)
HEMICHORDATA (Acorn Worms, Pterobranchs)
UROCHORDATA (Tunicata) (Tunicates, Sea Squirts, Salps, Ascideans)
CEPHALOCHORDATA (Amphioxus/Lancelet)
VERTEBRATES
AGNATHA (Hagfish, Lamprey)
FISHES
Chondrichthyes (Cartilaginous Fishes) (Sharks, Rays, Ratfish)
Osteichthyes (Bony Fishes)
Sarcopterygia (Lobe-finned Fishes) (Coelacanth, Lungfish)
Actinopterygia (Ray-finned Fishes)
AMPHIBIA
Anura (Frogs, Toads)
Urodela (Salamanders, Newts)
Gymnophiona (Apoda) (Caecilians)
REPTILIA
Chelonia (Turtles, Tortoises)
Serpentes (Snakes)
Sauria (Lizards)
Crocodylia (Crocodilians)
Rhyncocephalia (Tuatara)
AVES (Birds)
Paleognathae (Ratites)
Sphenisciformes (Penguins)
Procellariiformes (Albatrosses, Petrels,Fulmars)
Pelecaniformes (Pelicans, Gannets, Boobies, Tropicbirds)
Ciconiiformes (Herons, Bitterns, Egrets, Storks, Ibis, Flamingo)
Anseriformes (Ducks, Geese, Screamers)
Falconiformes (Vultures, Hawks, Eagles, Condors, Kites, Falcons)
Galliformes (Megapodes, Turkeys, Quail, Pheasants, Peafowl, etc.)
Gruiformes (Cranes, Rails, Gallinules, Coots, Bustards, Crakes)
Charadriiformes (Terns, Gulls, Stilts, Avocets, Plovers, Puffins, etc.)
Columbiformes (Pigeons, Doves)
Psittaciformes (Parrots, Lories, Cockatoos, Kakapo, Conures, etc.)
Cuculiformes (Cuckoos, Turacos, Anis,Coucal, Roadrunner, etc.)
Strigiformes (Owls)
Apodiformes (Hummingbirds, Swifts, Thornbills)
Coraciformes (Kingfishers, Todies, Bee-Eaters, Rollers, Hornbills, etc.)
Piciformes (Woodpeckers, Toucans, Jacamars, Barbets, Honeyguides)
Passeriformes (Passerines)
MAMMALIA
Monotremata (Platypus, Echidna)
Marsupalia (Marsupials)
Eutheria (Placentals)
Insectivora (Hedgehogs, Moles, Shrews, Tenrec, etc.)
Chiroptera (Bats)
Edentata (Anteaters, Sloths, Armadillos)
Primates
Monkeys
Apes (Gibbons, Orang-utan, Gorilla, Chimpanzee)
Humans
Rodentia
Laboratory Rodents (Rat, Mouse,Guinea Pig, Hamster)
Non-Laboratory Rodents
Lagomorphs (Rabbits, Hares, Pikas)
Tubulidenata (Aardvarks)
Carnivora (Bears, Canids, Felids, Mustelids, Viverrids, Hyena, Procyonids)
Ungulates
Perissodactyla (Odd-toed Ungulates) (Horses, Rhinos, Tapirs, etc.)
Artiodactyla (Even-toed Ungulates) (Cattle, Sheep, Deer, Pigs, etc.)
Sirenia (Manatees, Dugongs)
Proboscidea (Elephants)
Marine Mammals (Seals, Walrus, Whales, Otters, Dolphins, Porpoises)
TRANSGENIC ORGANISMS
FOSSIL OR EXTINCT ORGANISMS
NO ORGANISMS
CATEGORY VIII: MODEL ORGANISM (Select ONE) NO MODEL ORGANISM
MODEL ORGANISM (Choose from the list or input up to 9 characters)
VIRUS/BACTERIA Lambda Phage
Rhizobacterium
Escherichia coli
Bacillus subtilis
Cyanobacteria (Selenococcus/Selenobacter)
PROTISTA Acetabularia acetabulum
Chlamydomonas reinhardtii
Paramecium
Tetrahymena
FUNGI Dictyostelium
Neurospora
Saccharomyces cereviseae
Schizosaccharomyces pombe
PLANT
Mouse-Ear Cress (Arabidopsis thaliana)
Ice Plant (Mesembryanthemum spp.)
Barley (Hordeum vulgare)
Corn (Zea mays)
Pea (Pisum sativum)
Tobacco (Nicotiana spp.)
Spinach (Spinacia oleracea)
Alfalfa (Medicago spp.)
Tomato (Lycopersicon spp.)
ANIMAL Nematode (Caenorhabditis elegans)
Sea Slug (Aplysia californica)
Sea Slug (Hermissenda spp.)
Pond Snail (Lymnaea spp.)
Terrestrial Snail (Helix spp.)
Squid/Cuttlefish (Loligo, Sepia, etc.)
Octopus (Octopus spp.)
Leech (Hirudo medicinalis)
Horseshoe Crab (Limulus spp.)
Brine Shrimp (Artemia spp.)
Lobster (Homarus, Panilurus, etc.)
Crayfish (Procambarus, Astacus, etc.)
Dragonfly (Aeschna, etc.)
Grasshopper/Locust (Schistocerca, etc.)
Cockroach (Periplaneta, Blatta, Blatella, etc.)
Mantis (Mantis, Parasphendale, etc.)
Six-Lined Hawk Moth (Manduca sexta)
Fruitfly (Drosophila melanogaster)
Syrphid Fly (Syrphidae)
Apple Maggot (Rhagoletis spp.)
Mosquito (Culex, Aedes, Anopheles, etc.)
Flour Beetle (Tenebrio spp./Tribolium spp.)
Honeybee (Apis mellifera)
Parasitic Wasp (Braconids, Pteromalids, etc.)
Sea Urchin (Diadema, Mellita, etc.)
Ascidian (Boltenia, Molgula, etc.)
Lancelet (Amphioxus spp.)
Lamprey (Petromyzon spp.)
Skate (Raja, Myliobatis, etc.)
Croaker (Sciaenid Fishes)
Electric Fish (Eigenmannia, Sternopygus, etc.)

Page 6
Goldfish (Carassius auratus, etc.)
Perch (Perca spp.)
Zebrafish (Danio (Brachydanio) rerio)
Axolotl (Ambystoma mexicanum)
Mudpuppy (Necturus spp.)
African Clawed Frog (Xenopus laevis)
Bullfrog (Rana catesbeiana)
Grass Frog (Rana pipiens)
Marine Toad (Bufo marinus)
Turtle (Chrysemys, Pseudemys, etc.)
Quail (Coturnix spp.)
Chicken Embryo (Gallus domesticus)
House Sparrow (Passer domesticus)
White-Crowned Sparrow (Zonotrichia leucophrys)
Zebra Finch (Poephila guttata)
Opossum (Monodelphis, Didelphis, etc.)
Bat (Antrozous, Eptesicus, etc.)
Owl Monkey (Aotus spp.)
Rhesus Monkey (Macaca mulatta)
Tamarin (Sanguinus, Leontopithecus spp.)
Chimpanzee (Pan troglodytes)
Human (Homo sapiens)
Chinchilla (Chinchilla laniger)
Deer Mouse (Peromyscus spp.)
Guinea Pig (Cavia porcellus)
Hamster (Mesocricetus, Phodopus, etc.)
Kangaroo Rat (Dipodomys, etc.)
Mouse, Laboratory
Rat, Laboratory
Vole (Microtus spp.)
Domestic Dog (Canis domestica/familiaris)
Domestic Cat (Felis domestica/cattus)
Ferret (Mustelus spp.)
Farm Animals (Horse, Sheep, Pigs, Cattle, Goats)
[Enter your own model organism - up to 9 characters]

A. Project Summary
Title: Physiological Ecology of Diatom Motility and Adhesion
The ability of cells and organisms to perform controlled and directed movements within theirenvironment is critical to their ecological success and survival. This project involves a studydesigned to understand cell movement in diatoms, a ubiquitous group of golden algae abundant inboth freshwater and marine communities. Although diatoms are ecologically important as one ofthe main primary biomass producers in many aquatic communities, little is known about theenvironmental regulators of their motility, and the role that motility plays in their ability tosuccessfully compete (e.g. with other diatoms and with other algae) for light and nutrients.
Many diatoms display active and directed (e.g. phototactic) gliding movements, despite theirenclosure within hardened glass-like cell walls that prohibit direct contact between the cellmembranes and the sediment over which they move. Recent work from our lab has demonstratedthat different diatom species have significantly different motile behaviors, particularly in response toenvironmental light conditions. The experiments outlined in this proposal are designed to help testtwo specific hypotheses: 1) that the differences in diatom motile behaviors are generated bydifferences in the number, distribution and/or translocation speed of cell/substratum contact sites;and 2) species-specific differences in cells' motile responses to both environmental conditions andthe cell type composition in the population generate different sets of optimal ecological conditionsfor each species and population.
This project is designed to test these hypotheses and further understand the ecologicalfactors affecting diatom motility by using video microscopy analysis in three sets of experiments:
1) Quantitatively analyzing the changes in the characteristics of motility and adhesion for severaldiatom species when different combinations of species are placed together under a variety ofenvironmental conditions. This will allow us to determine how inter-species competition alters thetypes of motility characteristics we have already determined for individual species, and the degreeto which light-stimulated behaviors are altered in mixed populations.
2) Using a laboratory flume set-up to measure attributes of diatom adhesion in flowing water. Thiswill allow us to place mixed populations of cells under different flow conditions, allowing us todetermine how population makeup affects or alters the characteristic adhesions of single speciespreviously measured. By altering light and nutrient conditions, as well as water flow, we can beginto determine which ecological conditions are most favorable for which types of diatom populations.
3) Directly analyzing diatom cell/substratum contact sites using reflection interference microscopy.By observing these sites while altering the ecological conditions such light and temperature, we canalso directly observe changes in the contact sites for cells under different conditions. We will thenbe able to correlate the differences in diatom motile behaviors that we have previouslycharacterized with differences in the number, distribution or speed of contact sites.
Moreover, while not directly related to the experimental hypotheses, data accumulated fromthese studies will also help to determine a number of diatom properties useful in understanding theconditions for overall diatom viability:
• the environmental cues which can deleteriously affect diatom motile behavior;• the limits of deleterious environmental conditions (such as nutrient starvation, UV light,
chemical toxins) that can be tolerated by particular diatoms;• the light, nutrient, and substratum conditions most favorable to the growth and vitality of
various diatom species.

TABLE OF CONTENTSFor font size and page formatting specifications, see GPG section II.C.
Section Total No. of Page No.*Pages in Section (Optional)*
Cover Sheet (NSF Form 1207) (Submit Page 2 with original proposal only)
A Project Summary (not to exceed 1 page)
B Table of Contents (NSF Form 1359)
C Project Description (plus Results from Prior
NSF Support) (not to exceed 15 pages) (Exceed only if allowed by aspecific program announcement/solicitation or if approved inadvance by the appropriate NSF Assistant Director or designee)
D References Cited
E Biographical Sketches (Not to exceed 2 pages each)
F Budget (NSF Form 1030, plus up to 3 pages of budget justification)
G Current and Pending Support (NSF Form 1239)
H Facilities, Equipment and Other Resources (NSF Form 1363)
I Special Information/Supplementary Documentation
J Appendix (List below. )
(Include only if allowed by a specific program announcement/solicitation or if approved in advance by the appropriate NSFAssistant Director or designee)
Appendix Items:
*Proposers may select any numbering mechanism for the proposal. The entire proposal however, must be paginated.Complete both columns only if the proposal is numbered consecutively.
NSF Form 1359 (10/99)
1
1
19
9
2
8
1
1
0

C. Project Description
Resubmission Response
This proposal is a resubmission of one previously submitted to NSF in 1997 and 1998 andrevised based on reviewers’ comments. Specific modifications made to this submission include:
• Based on reviewers' comments and discussions with the program director, the number of experimentalsubsections has been reduced. This includes the elimination of the vertical migration experiments,which were the most exploratory of the experiments.
• In order to strengthen the rationale and conceptual framework within the proposal, brief explanationshave been added to each experimental section, stating how the described experiments relate to ourprevious results and to the goals of the proposal.
• Modifications from earlier versions of the proposal that remain are:– measurement of growth rates for the four main species of diatoms under our typical culture
conditions ([10] - all species have doubling times over 2 days)
– control experiments for cell growth rate in all experiments lasting more than several hours
–basic staining techniques to analyze the mucilage secretions produced by the diatoms as part of theflume studies
– additional information on the protocols involved in the flume and adhesion experiments, and on thedistribution of time points used for the observations and measurements used in these experiments
• Earlier reviewers voiced concerns over the level of explanation and activities in the proposal that wouldtie the laboratory experiments proposed and relevant field observations, and were divided betweenexpanding the ecological field experiments in the proposal or eliminating them entirely. In response tothese concerns, the field-based work was cut from the proposal itself, and a section was added at theend of the proposal which outlines the types of field-based work we plan to undertake bothconcurrently and subsequent to accumulating the lab-based data of this proposal. This organizationallows the proposal to more clearly show how we plan to connect the lab results with fieldobservations.
• Earlier reviewers were also split on the degree to which measurement on additional species was needed,with some reviewers feeling they diverted time from focusing more extensively on a fewer number ofspecies. In order to address these concerns, the number of additional new species being tested has beenreduced, with a slightly expanded explanation of why the remaining additional species would be useful.
Introduction and Background
Diatoms are an abundant and ubiquitous group of unicellular algae that are distinctive fortheir highly ornamented and often rigid silica-based cell walls, as well as their golden-brownpigmentation [97]. Diatoms are prevalent in almost all aquatic environments, both freshwater andmarine, and exist in both suspended (planktonic) and sediment-dwelling (benthic) forms. As agroup, diatoms are one of the major sources of primary biomass production. Despite the fact thatthese photosynthetic cells are among the most ecologically important members of aquaticcommunities, much is unknown about their physiology, behavior, ecology, and evolution[43,97,98,110]. In this study we plan to contribute to the understanding of diatom motility andecology by investigating how changes in the environmental conditions (e.g. light, temperature,substratum composition) affect the motile characteristics of different diatom species, particularly asthey compete with multiple species in resource-limited environments.
Diatom StructureDiatom cell walls are composed of two overlapping halves, somewhat like the top and bottom of Petri
plates, which are held together by mucilage. Diatoms secrete large amounts of mucilage, not only for

adhesion of the cell wall components, but for physical protection and adhesion of the cell to the substratumas well [71,97,98,126,127]. Removal of this mucilage by hydrolytic enzymes renders some cells prone torapid osmotic swelling and rupture [9]. The main structural component of each half of the diatom wall, thevalve, is highly ornamented, with species-specific patterns of pores and striations [98,103] that allow the cellto have access to small molecules, ions and nutrients while still offering the cell great physical protectionfrom bacteria, small predators, and osmotic shock. The large variety of wall forms is thought to havederived from the diverse selective pressures on diatoms. Such pressures likely include maximizingsurface/volume ratio for nutrient uptake and adhesive contact, streamlining along the apical axis for reducingresistance to water flow, regulating colonial attachments and sedimentation rates, and efficiently adhering toor moving through sediments and other algal colonies [43,49,78,103]. Additional variations in the cell wallstructure, due to physiological responses to changes in environmental conditions or different stages in lifehistories, make it difficult to assess the actual number of diatom species [43,103].
The relative inflexibility of the silica cell wall also requires that the plane of cell division occursmidway between the two halves [90,92,93], such that each of the daughter cells remains associated with oneof the two halves of the parental wall. Soon after cell division, each daughter cell secretes one new half-wall(valve) from a specialized membranous compartment called the silicalemma, which forms directly beneaththe plasma membrane in the area of the cleavage furrow. The new valves form a protective silica-basedscale over each plasma membrane, after which the two daughter cells enlarge, separating themselves fromone another. In this manner, cells can grow and divide without breaking or rupturing the solid portions of thecell wall. Since each daughter cell produces and secretes one new valve at each division, each vegetative cellis surrounded by a cell wall consisting of one new valve and one valve inherited from the parental cell. Thisarrangement has been useful in studies investigating cell wall morphogenesis, as valves formed duringexperimental treatment can be directly compared to the untreated parental valve present on the same cell[14,102].
A characteristic slit, the raphe, is usually present along the length of one or both of the valves in motilebenthic diatoms. Depending on the species, benthic diatoms may have a raphe on both valves (biraphid),only one of the valves (monoraphid), or no raphe slit at all (araphid). The raphe can be along the flattenedsurface of the valve (known as the valve face), or along an extended edge of the cell wall (known as the keel).The exact shape and orientation of the raphe is different in each species [28,30,98,104], but close proximitybetween the substratum and the raphe is required for motility [41,54].
Diatom MotilityThe structural organization of diatoms, in which the cell is constrained within the confines of a
hardened silica-based cell wall, creates restrictions on cell motility. Major ecological displacements ofdiatoms are no doubt driven by waves, tidal forces, and water currents [79, 108], and possibly amplified bythe loss of adhesion under unfavorable conditions [4]. However, many benthic diatoms are actively motilewhen in contact with a solid or semi-solid surface [41,97,98]. The speed of diatom movements variesdramatically with species, with some forms moving as rapidly as 10-20 µm/sec, while others move at rates <1-2 µm/sec. Motile species can live at or beneath the surface of aquatic sediments, and many types havewell coordinated movements such as diurnal phototactic behavior [54,97] and coordinated alignment of cellsduring sexual reproduction [19,91,126]. Motility also allows diatoms to migrate up within the layers ofsediment or surface algae to absorb more sunlight during the day, and settle into areas with higherconcentrations of organic nutrients at night [34].
The hardened cell wall, which completely surrounds the diatom plasma membrane, does not allow forcytoplasmic extensions or protrusions that could actively propel the cell through the water as cilia or flagellado. Nor can the cell maintain direct membrane-to-substratum contact that would allow the cell to crawl overthe surface in amoeboid-like movement. The mechanisms responsible for generating and controlling thismotility must therefore overcome these physical restrictions [35,41], and recent evidence supports cellularproteoglycan secretions as the material generating cell/substratum attachments [68, 69].
Several lines of evidence indicate that mucilage or proteoglycan material secreted through the raphe isrequired for motility. For example, asymmetric diatoms tend to move in a direction and curvature that matchthat of their raphe [41], strands or filaments extending from the raphe are detected using electron microscopywhen protocols designed to stabilize polysaccharides are used [38,41], and particles adhering to the diatom inthe area of the raphe are observed to be transported bidirectionally along the raphe. In addition, somediatoms are known to deposit mucilage trails from their raphes as they move [35,41], antibodies directedagainst proteoglycan material in the raphe area can inhibit motility and adhesion [68,69], chemicals that

interfere with motility can affect secretion and adhesion [120, 128], and chemicals that inhibit secretion ofextracellular matrix can inhibit motility [119]. Moreover, the motile ability of a diatom is correlated with itsshape and ornamentation (e.g. the shape of its raphe [2,41]). Motility of freshwater diatoms also requires 1-2 atm osmotic pressure of protoplasts against the cell wall [9] suggesting that the pressure may be requiredfor proper extrusion of mucilage material through the raphe.
Several models by which secreted mucilage could generate raphe-based motility have been suggested,including propulsion through rapid mucilage expulsion and/or hydration or directed capillary action[48,50,106]. Mucilage extrusion alone might be able to explain the slower gliding movement of some algaesuch as desmids, photosynthetic bacteria, or some araphid or stalk forming diatoms [52,59,89]. However,the faster and more responsive motility of many raphid diatoms, as illustrated by their phototactic movementsand lateral pairing of cells prior to sexual reproduction [20,21,22,37,41,75,97,98,126] make such amechanism unlikely.
The most extensive model for diatom motility, developed by Edgar and others, is based on several setsof observations: 1) Two bundles of cytoplasmic filaments, associated with the plasma membrane and runningunderneath and parallel to the raphe [39,40], are positively stained with fluorescent phalloidin, a probespecific for actin [42,116,128]; and 2) protocols that stabilize polysaccharides revealed a number of smallfibrous strands of material extending through the raphe to the exterior of the cell [38]. Edgar speculated thatthe fibrous material was composed of adherent mucilage strands secreted from the cell and extruded throughthe raphe. Under this model, motile force could be produced by moving membrane-bound mucilageattachment sites along underlying actin filaments, effectively pulling on the mucilage strands down the lengthof the cell. The mucilage attachment to the substratum would act as an anchoring site against which the cellcould generate force, thus allowing the cell to pull itself along [41].
The mechanisms of such movement could be similar to the cytoplasmic streaming of vesicles orchloroplasts along actin cables in higher plant cells [63,105,129], which is generated by force-producing"motor" proteins such as the myosins [64,65,66,94]. Bidirectional movement could be generated by the useof anti-parallel actin filaments, either by having actin filaments of both orientations contained within eachactin cable, or by having each of the two cables uniformly oriented, but in opposite directions.Microfilaments in diatoms are also involved with other types of intracellular motility, such as cytokinesis andplacement of cell wall material [14,90], so one must be cautious; the actin cables may be associated withsecondary processes, and not directly involved in force generation [48]. Since several forms of microtubule-based motility in diatoms are well-known [14,15,86,130,131], the role of microtubules in motility alsocannot be ruled out. However, latrunculin, a potent actin inhibitor from sea sponge, causes rapid andreversible inhibition of diatom motility [128], further supporting the model of actin cables for motility.
Diatom Ecology and PhysiologyDiatoms have been objects of experimental investigation ever since the intricately sculptured cell walls
and distinct golden pigmentation caught the attention of the early microscopists, and were among the firstorganisms analyzed for mitosis and movement [67,73,106]. These cells are abundant, widespread, and amajor source of primary biomass production in many marine and freshwater aquatic communities [7,97,98].The significance of diatoms at the start of many food chains can be shown, for example, by the toxicpoisonings of humans and brown pelicans [44,82], both of which were ultimately the result of diatomtoxicity. For the pelicans it was due to high levels of domoic acid in anchovies, a principal pelican foodsource, which had fed on a bloom of a toxic diatom, while the human poisonings were the result of domoicacid accumulation in mussels. Such episodes, while rare, graphically demonstrate the importance of diatomsin the maintenance and stability of aquatic food chains and why they are crucial to the ecological stability ofmany aquatic ecosystems [7,98,110]. Their importance in aquatic ecosystems, along with their sensitivity toenvironmental conditions, has made diatoms popular indicators in the study of changes in environmentalwater quality and ecological stress [13,95,96,109].
Diatoms are also considered to be ecologically important contributors to the erodibility and stability ofsome aquatic sediments. The large amount of mucilage secreted by diatoms (which can vary considerably incomposition [57,132]) allows them to adhere strongly to both natural and man-made surfaces [26,27,55],and is likely composed of a complex set of secreted materials [127]. The mucilage secretions can affect theerodibility of some intertidal regions [81], may be responsible for some of the stability of river sediments[70] and is also thought to affect the successful immigration of diatoms into newly available environments[84,108,111,112]. Some studies show an apparent correlation between diatom motility and strength ofadhesion to the substratum, as well as substrate-specific differences in adhesion [1,55,77,113]. Adhesion

may also be correlated with the presence or pre-conditioning of the substrates with bacteria or other biofilms[27,51,85]. Mucilage secretion is also known to be an important factor in inter-species aggregation [31,53].
The ecological regulation of benthic algal communities, and the physiological and competitiveinteractions responsible for generating successful diatom communities remain poorly understood [72].Diatoms can migrate through sediment [80] and during development of algal communities may undergostratification, with particular diatoms species being found at characteristic distances beneath the top surfaceof the community [60,108,112]. Since light can become limited as the density of the community increases,benthic diatom survival must often involve behavioral strategies to compensate for diminishing lightavailability. Such strategies can include movement upward through the surrounding algae, passivedisplacement due to stalk formation or differential adhesion to adjacent material [113], or non-motileresponses such as temporary dormancy or conversion to heterotrophy [115]. The relative contribution ofmotility remains undetermined, but is almost certain to be considerable in many circumstances since non-motile species often remain at the bottom of developing communities [108,112].
Diurnal vertical cell movements may enhance survival of algae such as diatoms by allowing cells tomove upward and capture more light during the day, then resettle downward to the more nutrient rich bottomof the sediment during the night [110,112]. If this is true, there should be environmental cues (e.g. light,ions, surface properties), either within the sediment or within the overlying water, which regulate themovement. However, such regulatory processes are poorly understood in diatoms and other benthic algae[110]. There have been relatively few studies on chemotactic responses in diatoms, although some responsesto sugars have been reported [24]. Nutrient concentrations are often used by protists for determining theorientation of motility [52] and diatoms are also known to be sensitive to nutrient limitation, which can affecttheir adhesion ability [112,118]. Therefore, the available ionic and nutrient concentrations may be importantecological regulators of diatom motility.
In particular, calcium may be important in regulating actomyosin-based force generation and/or thesecretion of mucilage through the raphe, since it can regulate actin-based motility in both muscle and non-muscle cells [6,94,114] as well as fusion of secretory vesicles. Extracellular calcium has been identified asimportant for adhesion and motility in marine diatoms [23,25,26]. However, large external concentrations ofcalcium are not required for freshwater diatom motility [9], although the treatment of freshwater cells withinhibitors of calcium channels [32,45,125] inhibits motility in a dose-dependent manner [9]. Thesedifferences may reflect the fact that marine diatoms live in an environment with a large external calciumconcentration, whereas freshwater diatoms do not, so that freshwater diatoms may use internal (rather thanexternal) stores of calcium for regulation. This may explain why mitochondria (which regulate calcium insome cells [32]) are often observed in the cytoplasm near the raphe [87,88,90]; they could provide both theATP and calcium needed for movement.
Light-stimulated motile responses of diatoms, such as diurnal vertical movements and accumulation ofcells in light spots, have been documented for some time [54,74,76,97,98,99]. Such phototactic movementsof algae usually result from one or more types of behavioral responses to changes in light intensity orwavelength [52]: alterations of cell speed (photokinesis); reversing direction at light-dark boundaries(photophobic response); light-stimulated changes in the direction of cell growth (phototropism); or variationsin the frequency of direction changes. For diatoms, light directed movements seem to be due primarily to aphotophobic response to light/dark boundaries [8,10,20,76,121,122,123,124] detected at the tips of the cells[18]. The presence of actin cables beneath the raphe, and the importance of calcium to motility, suggest thatdiatom light responses may be regulated similarly to the light stimulated chloroplast movement in Mugeotia[56,117] in which calcium regulates chloroplast/actin attachment.
While biraphid pennate diatoms can have considerable differences in motile behaviors and sensitivitiesto light [10,20], it is not yet clear how this relates to their ability to exploit different microenvironments. Forexample, the intensity and spectral quality of light change throughout the various layers of a developing algalcommunity [61,62], so that differential light responses may contribute to the stratification of diatom specieswithin the algal mat [60].
The project described in this proposal outlines a coordinated set of experiments designed to extend ourprevious work and further our understanding of the environmental and ecological conditions affecting diatommotility. Specifically, we plan to: 1) analyze the motility of mixed populations of diatoms to determinechanges in motility and light stimulated effects due to inter-species competition; 2) analyze the changes incell adhesion that are similarly affected by changes in environmental conditions or cell populations; and 3)

analyze the actual cell/substratum contact sites in order to correlate the diatom behaviors with the presence,density, direction and speed of cell contact sites. Such a study should provide significant advances tounderstanding the physiological responses of diatoms to their environment.
Results from Prior NSF Support
Grant Number: IBN-9407279Award Amount/Date: $210,000 - July 1994 to June 1997, extended to 1998Grant Title: Physiology and Ecology of Diatom Motility
Our previous grant allowed us to focus on some ecologically important behavioral differencesbetween diatom species, with emphasis on the motility and adhesion of the cells under variousenvironmental conditions. Most of the work centered on two questions: 1) how do thecharacteristics of diatom movement and adhesion differ between species; and 2) whatenvironmental conditions affect diatom motility and adhesion. Some of our findings aresummarized below.
Characterization of Motile Behavior: We have characterized the motility and adhesion strength offour species of biraphid pennate diatoms [20], developing quantitative assays for cell speed, path curvature,and relative adhesion. Qualitatively, the four species can be characterized as: Craticula cuspidata- fast,weakly adhering, straight path; Pinnularia viridis- slow, moderately adhering, curved path; Stauroneisphoenicenteron - slow, strongly adhering, straight path; Nitzschia linearis- fast, strongly adhering, straightpath (quantitative values reported in [20]).
Environmental Factors Affecting Motility: High concentrations of calcium, reported to berequired for marine diatom motility, are not required for freshwater diatom motility [9], although somecalcium inhibitors do inhibit motility. Freshwater diatoms require very low external osmolarity for motility,suggesting that the 1-2 atm osmotic pressure of the cell protoplast against the cell wall is possibly requiredfor aiding extrusion of mucilage through the narrow raphe.
Correlation of Motility and Adhesion with Temperature: In all four species of diatoms, cellspeed steadily increases with temperature until about 35-37 °C, after which the motility rapidly declines [22].We are now investigating whether this relationship also holds true for adhesion, and whether temperature-dependent changes in adhesion are due to changes in the mucilage itself, or due to changes in its secretion.
Investigation of Diatom Phototaxis: Our experiments have indicated that diatoms collect in lightspots of light levels at about 1-50 µmole/s-m2, using a step-down photophobic response, whereby cellsreverse direction at light/dark boundaries. The diatoms species show differences in both wavelength andintensity sensitivity [10,21]; initial studies show that irradiation of a single diatom culture with spots of lightwith different wavelengths can result in different species distributions in each spot.
In conjunction with the lab of Dr. Jeremy Pickett-Heaps (Univ. of Melbourne) we also investigated therole of localized high intensity light exposure on diatom direction change [18,22]. We determined that high-energy light (about 500 µmole/s-m2 at 500 nm) causes rapid cell reversal in <10 sec when the irradiationoccurs at the leading tip of the cell. In summary, the experiments suggested the following:
• high light level irradiation at the leading tip of a moving diatom causes cell reversal ( > 2 sec exposureswere 100% effective at all wavelengths in our tests)
• irradiation of the trailing end causes no direction change• high-energy and low-energy light responses have the same spectral profile for Craticula• after an initial irradiation-induced direction change, the cell is less responsive to a second direction
change generated by a subsequent irradiation at the new leading end.
Adhesion of Diatoms With/Without Water Current: In conjunction with Dr. Tuchman atLoyola University, Chicago, we designed a flume assay in which diatoms are placed onto a platform overwhich water flow (at variable speed) is generated. Initial results [21] suggest that the relative rate of cell lossdue to water current is species specific, and highly dependent on the substrate to which the cells are adhering[17].
Effect of Toxins and UV Irradiation on Diatom Motility: Since diatoms are such importantcontributors to the primary production in many aquatic ecosystems, we developed assays that use diatom

motility as an indicator of deleterious changes in the ecological conditions. We have begun to investigate theeffects of both UV irradiation and toxic sediments [10,13] on diatom motility and growth rates. Initialresults show that using irradiations of 3 mW/cm2, cells rapidly lose motility with exposures > 30 sec.
In summary we have developed several quantitative assays which have demonstrated considerabledifferences in behavioral responses among several diatom species, and that measurement of these behaviorscan be used as rapid and accurate assays for the presence of environmental stress.
Publications Resulting from NSF Award (Undergraduate Authors marked with asterisk)
Manuscripts†Cohn, S.A. and Disparti, N.C.* (1994). Environmental factors influencing diatom cell motility. J.
Phycol. 30: 818-828.
Cohn, S.A. and Weitzell, R.E. Jr. (1996). Ecological considerations of diatom cell motility: I.Characterization of motility and adhesion in four diatom species. J. Phycol. 32: 928-939.
‡Cohn, S.A., Spurck, T.P., and Pickett-Heaps, J.D. (1999). High energy irradiation at the leading tip ofmoving diatoms causes a rapid change of cell direction. Diatom Research (in press).
‡Cohn, S.A. and McGuire J.R.* (1999). Using diatom motility as an indicator for environmental stress:Effects of toxic sediment elutriates on diatom motility. Diatom Research (submitted, in revision).
§Cohn, S.A. and Sciortino, S. (1999). Examination of motile diatom loss from different substrates dueto the force of water flow. Diatom Research (to be submitted, August/September 1999).
Abstracts†Cohn, S.A. and McGuire, J.R.* (1994). Use of diatom motility assays for toxicity testing. Mol. Biol.
Cell 5(S): 485a.
Cohn, S.A. and Weitzell, R.E. Jr. (1995) Characterization of motility and adhesion in four species ofpennate diatoms. J. Phycol . 31(suppl.): 6.
Cohn, S.A., Weitzell, R.E. Jr., Spurck, T.P., and Pickett-Heaps, J.D. (1995) Characterization of motilityand adhesion in pennate diatoms. Mol. Biol. Cell 6(S): 261a.
Cohn, S.A., Weitzell, R.E. Jr., Norris, A.* and Lazzarotto, M.J.* (1996) Comparative Analysis ofAdhesion and Photo-stimulated Aggregation in Pennate Diatoms. Mol. Biol. Cell 7(S): 232a.
Cohn, S.A., Dunbar, S.A., Skoczylas, C.* and Mucha, J.A.* (1997) Comparative Analysis of DiatomMotility: Phototactic Behavior and Sensitivity to Ultraviolet Radiation. Mol. Biol. Cell 8(S): 386a.
§Cohn, S.A. and Sciortino, S. (1998) Examination of motile diatom loss from different substrates due tothe force of water flow. Mol. Biol. Cell 9(S): 34a.
†The data from this study was predominantly acquired prior to the previous grant, but the grant providedsupport during the submission and/or revision period.
‡The data from this study was predominantly acquired during the period of the previous grant, but thesubmission process was carried out after the grant period was over.
§The data from this study was acquired both during and after the period of the previous grant, usingequipment purchased through the previous grant, but the submission process was carried out after the grantperiod was over.
In addition, another manuscript, Cohn, S.A., Weitzell, R.E. Jr. and Wibisono, B. Effect of temperature ondiatom motility and adhesion. is expected to be ready for submission within 12-18 months. The PI has also

submitted the first draft of a chapter on diatom photo-based motility for an upcoming volume onphotomovement edited by Drs. D-P. Häder and M. Lebert; the manuscript needs to be revised and additionalmaterial will be resubmitted shortly.
Experimental Design
The proposed project is designed to expand upon those experiments carried out under theprevious grant. While our previous work has allowed us to quantitatively characterize the generalmotility and adhesion in four species of pennate diatoms, we now plan to build upon this work byasking two basic questions:
1) How are the characteristics for motility and adhesion we have measured changed by the presence ofparticular combinations of diatoms? That is, how do different diatom species in mixed populations competewith one another for limited resources or space. This includes both looking at changes in cell speed andadhesion as well as changes in cell immigration/emigration onto substrates based on species distributions inthe population.
2) How do the actual cell/substratum contact sites differ between species, and how do their abundance,speed, or distributions change to give rise to the behavioral effects stimulated by environmental changes? Inother words, what are the differences in cell contact sites between the species that gives rise to theirbehavioral differences, and which characteristics of these contact sites change as the cells modify theirbehavior based on changes in environmental stimuli.
We plan to address these questions using three types of approaches:
• Analysis of Motility - As with the previous single species measurements, we plan to characterizethe motility and light-stimulated responses (e.g. high light avoidance, low light accumulation) of diatomsusing computer-assisted video microscopy. However, in this case we will analyze changes in the motilitythat take place when different mixtures of individual species are put together in multi-species populations.We also plan to characterize two additional species with significantly different valve shapes and structuresfrom those previously analyzed, so that we can better assess the role of cell structure in motility andcompetitive success. In addition, we will analyze the motile behaviors of cells in multi-species populationsunder a variety of ecological conditions (e.g. changes in light, nutrients, temperature, substratumcomposition) to better determine which types of conditions are best suited for which diatoms.
• Analysis of Cell Adhesion - Our previous work allowed us to develop assays to measure diatomadhesion in both a static test (inverted coverslip) as well as in a test designed to measure cell loss due to theforce of flowing water. We also measured the effect of different substrates on the relative adhesion ability ofone cell type (Pinnularia viridis). We plan to expand this study to get a better sense of how different cellsreact on different substrates, and how the adhesion of cells may change based on the cell type composition inthe population. Such analysis will allow us to determine the relative immigration and emigration rates of cellsin flowing water as they are removed from different substratum surfaces under conditions where thetemperature, light, and rate of water flow can all be independently altered.
• Analysis of Cell/Substratum Contact Sites - We plan to use reflection interference contrastoptics to directly determine the number, size and speed of cell/substratum contact sites from several differentdiatom species, as well as analyze the alteration of these contact sites during changes in the cell's motility(e.g. direction change) or physical environment (e.g. temperature). We will also analyze changes in thecontact sites that occur as populations with different species distributions are used.
Additional Cell TypesIn the experiments described below, we plan to analyze the four diatom species characterized
in our earlier studies (P. viridis, C. cuspidata, N. linearis, S. phoenicenteron) as well as add twomore cell types to many of the experiments, both of which are typical and widespread [30]. Theadditional cell types are Surirella sp. and Achnanthes sp. Both are cell types with important

structural/functional differences needed to compare with our previous species. In both cases wewill use the largest cells of the form that we can obtain and culture. The reasons for adding each ofthese two types are described below.
• Surirella is a motile form with a peripheral raphe canal that runs circumferentially around anextended rounded keel. This cell type will allow us to answer some critical questions about bothlight stimulation and cell adhesion. Because of the curvature of the raphe canal, analysis of the cellcontact sites in Surirella will allow us to better determine the maximum cell/substratum distancesthat still can generate a substratum contact site. These cells will also give us a better understandingof how cells reorient themselves as cell contact sites move along the length of a raphe. In addition,Surirella can help us answer several questions about light stimulated motile responses. All of thefour species we have previously tested have had raphe branches that extend outward along thelongitudinal axis, away from the cells’ "central areas" where there is a break between the two raphebranches. The two distal tips of the raphe are thus separated on opposite ends of the cell, near theareas where the cells are most sensitive to light. In Surirella, however, the valve and raphe canalsoriginally form after cell division starting at one tip, with the nascent raphe canals extending alongthe periphery of the cell and meeting near the opposite end. Surirella therefore provides a cell typein which the two “tips” of the raphe branches are adjacent to each other, located at the same end ofthe cell. This structural organization thus allows us to ask whether Surirella can in fact respond tothe same light cues as the other cells, and if so if the light receptor sites have been displaced awayfrom the ends of the raphe.
• Achnanthes is a monoraphid form in which several species have shown active motility[5,30]. Such a cell type will allow us to determine the relative motility and adhesion of monoraphidforms (on both the raphe and non-raphe sides) and determine whether there is a difference in cellcontact sites generated on the two sides. If we measure motility in our species, we could determineif the cells would be able to flip over and reassociate from either side, as is easily accomplished inbiraphid forms [20].
If the work outlined in this proposal proves fruitful, we could address a variety of other celland raphe forms in the future (e.g. Nitzschia sigmoidia in which the raphe is sigmoidal,Gomphonema sp. in which one end of the cell is much larger than the other, or Cymbella sp. inwhich the entire cell is curved).
Light Microscopic Analysis of Motility
Our previous work allowed us to develop several assays designed to measure physical andmotile characteristics of cells such as: cell size/shape, average cell speed, path curvature, frequencyof direction change, and attributes of light-stimulated responses. The assay methods are describedbelow, followed by the competition experiments we propose which will use these assays.
Motile Characterization AssaysWork under the previous grant characterized behavioral differences between four species of
pennate diatoms: Craticula cuspidata, Pinnularia viridis, Stauroneis phoenicenteron, andNitzschia linearis [20,21,22 - species determined subsequently]. These species are all large,relatively linear, biraphid diatoms isolated from the same pond (which was artificially fed fromnearby stream water). The cells were all isolated from the outflow region of the pond, where therewas relatively constant water current, in order to obtain species that were likely to exhibit someability to remain adhered in the presence of water flow.
The characterization of cell motility will be accomplished using a computer-assisted videomicroscope set-up as used previously for analysis of diatom motility and kinesin-driven microtubulemotility [9,11,12,16,20,58,101]. The set-up consists of a Zeiss Axioskop microscope equipped withdifferential interference contrast (DIC) optics and a high resolution Dage newvicon video camera.The output of the video camera is directly connected to the video output of an Amiga 2000

computer, resulting in a mixed signal in which the computer cursor is superimposed directly overthe video-microscope image. The computer 'mouse' can then be used to move the computer cursorto desired points on the microscope image as viewed on the video monitor. Using a custom-written program for the Amiga 2000, the computer cursor can be moved to a desired location, themouse button triggered, and the time and relative location of the point marked in the computer. Byfollowing a moving object with the mouse, its velocity can thereby be measured. Initial calibrationbetween screen distances and actual distances on the microscope stage is obtained using a stagemicrometer.
The computer/video microscope set-up can be used to calculate average velocities, intervalvelocities, path lengths, and other motile characteristics, with the measurements made relativelyquickly, in real time. The computer set-up can also take input from a video recorder, allowing thecomputer to calculate velocities from previously recorded images; in this case the calibration ismade from recorded images of a stage micrometer. For those conditions (e.g. movement onopaque surfaces) where transmitted light cannot be used, observations can be made using a high-resolution dissecting stereomicroscope equipped with a video camera and external fiber opticillumination.
Photo-response AssaysPrevious work has clearly shown that the phototactic response in diatoms is strongly due to
photophobic responses of diatoms at light/dark boundaries [20,74,121]. The low/moderate lightlevel step-down response causes diatoms to rapidly change direction and move into the light uponreaching a light/dark boundary, resulting in the accumulation of diatoms in light spots. Thisresponse is sensitive to both light wavelength and intensity, causing the cells to revert to a step-up(out-of-light) response at high light intensity [18,22]. The response is unique for each species, andwe have been able to quantify the response of individual species to change direction at light/darkboundaries and accumulate into light spots. We have developed three assays to measure diatomphoto responses: the boundary assay, the accumulation assay, and the avoidance assay.
In the light/dark boundary assay, previously used to determine the wavelength sensitivity ofthe photophobic response [20], we measure the frequency at which individual cells change directionat light/dark boundaries. Cells from culture are mounted on a slide in dimmed room light and thenplaced in a light-tight box for 1 hour to reduce any transient effects from room lighting duringmounting. Individual cells are then placed on the microscope and exposed to a small (250-300 µmdiameter) spot created by closing down the field diaphragm. Cells in the spot are allowed to moveto the light/dark boundary, during which their speed can be measured, and then scored for whetheror not the cell changes direction at the boundary; this allows us to determine the frequency of cellsexhibiting direction change. The basal rate of light-independent cell reversal can be determined bymeasuring the frequency of cells, illuminated by a fully open diaphragm, reversing direction at a"mock" spot drawn directly on the video screen. The light intensity can be varied to determine theminimum intensity required for a photophobic response, and the intensity at which the responsechanges from into-light to out-of-light.
In the cell accumulation assay, the lower portion of a small petri plate filled with the desiredpopulation of diatoms is placed into a two piece container which blocks all light except a small (5-7mm diameter) spot opening in its bottom, and placed on the illuminated base of a dissectingstereomicroscope. The number of cells within the illuminated spot is counted every 20-30 minutesby transiently opening the top of the container. A marked spot of the same size (blocked from thelight) is placed inside the container in order to count the cells in a "dark" spot for control. Theintensity of the light is controlled using a variable intensity light source, and the effectivewavelengths are varied using either broad bandpass or interference filters. We can thus determinethe rate of cell accumulation for different species, in both unialgal and mixed populations, as afunction of light intensity, light wavelength, cell density, or species composition.

A modification of this assay, in which the petri plate is illuminated using focused fiber opticlighting from above, can be used to illuminate several different areas of a population of cells withdifferent intensities and/or wavelengths of light. We can then determine if light quality can be usedto differentially alter species distributions within the light spots, and thus whether different diatomspecies might use light to segregate themselves into different microenvironments. In all cases, fiberoptic illumination will be used, since this is a cooler form of light and minimizes the changes intemperature during observation; light intensity will be calibrated with a quantum light meter. Ourinitial data [21] indicate that in mixed populations, Craticula cells accumulate much faster inblue/green light, while Stauroneis accumulate much better in red light.
We are currently developing an epi-irradiation avoidance assay, which is an adaptation of theirradiation assay previously used to characterize the C. cuspidata step-up (out of light) response athigh light intensities [18,22]. In this assay, we use the epi-illumination filters of a fluorescentattachment on the Zeiss Axioskop to irradiate the sample slides with 50-100 µm diameter spots ofhigh intensity light of defined intensity and wavelength. In this way, cells can be observed usinglow levels of light from the standard light source from below, and irradiated as desired controllingthe epi-light path from above. Initial tests have shown this method to be able to yield 100%response in stimulating a direction change in moving Craticula cells using a 2 sec irradiation; moreprecise irradiations of < 1 sec will require the shutter system requested in the equipment budget.This assay can be used to determine the light level, wavelength, and exposure time required togenerate an avoidance (step-up) response in cells.
Competition ExperimentsWe plan to extend the previous work we have done on single species, concentrating on the
effects of multi-species populations. Large numbers of the desired cultured cells will be rinsed indiatom medium using serial transfer into spot plate wells using micropipets. Washed cells will thenbe placed on a slide containing a small spacer to keep the cells from getting crushed, and allowed tosettle in the dark for 15 min. Initially, we plan to make samples containing relatively equal numbersof each of the cell types being tested, at a density of about 5-10 cells per mm2. Cells will be placedon the stage of either a compound microscope (boundary assay, avoidance assay, or motilecharacterizations) or a dissecting microscope (accumulation assay) and tested as described above,with each species in the population being measured independently. We plan to carry out a numberof combinations, comparing sets of dual species, triple species, and quadruple species. After ourinitial characterizations of Surirella and Achnanthes, samples containing all six species can also becarried out. Such tests will allow us to determine the behavioral differences between species in amixed population exposed to the same ecological conditions.
By controlling the external environment of the cell populations (e.g. temperature using atemperature-controlled stage) these experiments will allow us to compare the response of cells inthese mixed populations with the responses measured for individual species. They will also allowus to determine which types of temperature and light environments are most conducive for the lightaccumulation of particular species. We can also modify the chemical environment of the cells (e.g.altering the pH or ionic composition) to determine the relative activity and viability of the differentcell types under the altered conditions.
These assays will therefore allow us to answer several key questions about diatom populations:1) Does each diatom species behave differently when part of a mixed population?2) Do diatoms behave differently when in single species vs. multi-species environments?3) Do the differential responses of diatom species (e.g. to light intensity or wavelength) allow cells in a
mixed population to segregate into different sub-populations based on light conditions?
Light Microscopic Analysis of Adhesion
In the course of our previous studies, we developed two main assays to functionally measurediatom adhesion: the inverted coverslip, and the flume assays. These assays allowed us todetermine the relative adhesion of cells by measuring cell loss due to either the force of gravity or

the force of flowing water. We will use these assays to determine if cells in mixed populationsgenerate different adhesion characteristics than cells in single species populations. Previousexperiments on single species have shown that different species have different adhesion abilities[17,20,21]. Diatom species are known to secrete different forms of mucilage [57] which may bemodified by other cells or materials [51,119], so it is important to test the relationship betweenmotility and adhesion, and how differences in the environment and species distributions can alterthe cells’ adhesion ability.
Cell Adhesion on an Inverted CoverslipIn this assay, a small spot of 50-100 cells are placed via micropipet onto a glass coverslip
immersed in diatom medium. The cells are then allowed to settle and adhere for 15 minutes, afterwhich the coverslip (remaining immersed) is inverted so that all of the cells are attached on thelower surface. The cells remaining attached to the coverslip are then counted under a stereo-microscope at 15-30 minute intervals over several hours. Such cells fall off at a rate that decreasesexponentially [20], so that a distinct rate constant can be determined for each species. Anadaptation of this assay places cells on a coverslip in a sealed chamber. Under this method, theentire chamber is inverted, reducing any loss of cells due to media turbulence.
The idea that adhesion and motility are directly coupled has been supported by workindicating that inhibition of cell motility results in changes in cell adhesion [69,120,128]. Theinverted coverslip assay allows us to address the issue directly, by measuring cell adhesion underconditions that alter cell motility. For example, diatoms vary their speed as a function oftemperature [22], and we can determine whether there are similar temperature-dependent changesof adhesion or contact sites. To determine the effect of temperature on adhesion, we will use asmall double-sided cell chamber attached onto a portable temperature controlled stage. In thisassay, 100-200 cells will be placed onto the coverslip on one side of the chamber, which is thenfilled with diatom medium and sealed. The cells are then allowed to adhere onto coverslips atpermissive temperatures (e.g. 25°C), after which the chamber/stage apparatus is inverted. Theinitial fall-off rate and motility of the cells can be observed and measured for 30-90 min, after whichthe temperature can be shifted, during observation of the cells, to a higher temperature (e.g. 35-40°C). The fall-off rate and motility of the cells after the temperature shift can then be comparedwith the initial rates, and with the rates of untreated cell populations. This will allow us todetermine if higher temperatures inhibit the initial adhesion of cells to substrates, or if it reduces theadhesion of previously adhered cells.
Similarly, we can use latrunculin, an actin inhibitor from sea sponge that causes rapid andreversible inhibition of diatom motility [128], to determine the effect of motility inhibition onadhesion. In these experiments, cells will be allowed to settle and adhere onto a coverslip that hadbeen sealed onto a drug perfusion chamber such as the type previously used to determine drugeffects on diatoms in vivo [15]. Once the cells have adhered, the chamber will be inverted and thecells observed in either a stereomicroscope or standard compound microscope to obtain the initialrate of cell detachment. The chamber will then be flushed with 1 µM latrunculin to stop cellmotility, and the new fall-off rate of inhibited cells determined. The latrunculin-based rate can thenbe compared with the pre-treatment fall-off rate, or the rate of untreated cells. This will allow us todetermine the degree to which active cell movement is required for proper cell adhesion.
Cell Adhesion in Water CurrentWe have previously performed initial experiments in which we determined the rate at which
adhered cells are removed from the substratum by the force of flowing water [21]. In this assay,(developed with Dr. N. Tuchman of Loyola University, Chicago) cell adhesion is measured in aflume apparatus in which both the water velocity (measured by a digital flow meter) and the cells'substratum can be varied. The flume set-up consists of a modified aquarium in which an innerblock of the aquarium is sealed off, resulting in a channel of water surrounding the blocked center.A variable speed propeller is immersed into one side of the aquarium, propelling the water aroundthe channel in one direction. A platform is then mounted on the upper surface of the central

compartment (with parallel oriented hollow tubes on either side if needed), so that the water flowsin essentially unidirectional and laminar flow over its surface. Cells are placed onto the platform viamicropipet, allowed to settle and adhere for 15 minutes, then exposed to water flow of variousspeeds from 0-0.5 m/sec (calibrated by a water flow meter). The number of cells remainingadhered to the surface over time is observed and counted using a video stereomicroscope. Initialexperiments suggest that cells detach from the platform at a rate best represented by a power curve(rather than exponential loss), suggesting that cells become increasingly harder to remove from asubstrate over time. Comparative parameters of the power equation can be determined adequatelyby measuring the percentage of cells remaining adhered at 1 min intervals for 5-10 min after theinitiation of water flow, using the average results from at least three replicate trials.
The flume apparatus can also be modified to allow us to change environmental conditionssuch as light or temperature. The water in the flume apparatus can be connected to a temperaturecontrolled circulating water pump, allowing us to vary the water temperature and overhead lightsources can be adjusted and focused onto the platform, in order to determine if the wavelength orintensity of light alters the ability of cells to adhere to the flume platform.
Cells show differential abilities to adhere to various types of surfaces [17,41,113], and ourinitial studies suggest that natural surfaces such as woods are more conducive to diatom adhesionthan synthetic or metal surfaces such as those often contaminated by algal biofouling [26,27]. Thetype and texture of the substratum surface is already known to affect algal establishment anddiversity [36], and our flume set-up allows us to directly test which types of surfaces are mostconducive to cell adhesion and colonization. The flume platform can be composed of any ofvarious materials (e.g. marble, glass, metal, or wood) with the cell loss from each type of surfacemeasured as described above. We can therefore determine differences in adhesion ability of cells ormixed populations that are due to surface properties of the substrate. In addition, the surfaces usedcan be either untreated, or coated with specific additives to change the surface properties (e.g.poly-L-lysine to produce a positively charged surface). Since there is evidence that conditioning ofsurfaces by bacteria or biofilms may affect the ability of diatoms to adhere and grow [27,51,85,98],we can also measure the cell adhesion/loss from surfaces in the presence or absence ofconditioning. Acid cleaned glass coverslips can be used as substrates without modification, or bepre-treated by incubating the coverslips in filtered (non-sterile) pond water in the dark for severaldays to produce a biofilm (similar to [85]). Samples of these conditioned coverslips can beobserved using DIC optics for presence/density of bacteria, and coverslips incubated in sterilizedpond water under axenic conditions can be used for comparison.
Competition and Comparative Analysis ExperimentsSpecies composition has been shown to play a role in the loss of cells due to changes in water
current [3,83], as well as the ability of cells to form multi-species aggregates [31,53], and cells maybe induced to form stronger adhesions from the water flow itself. By analyzing the relative loss ofcells in single and mixed populations, we can observe if the species distributions affect thefunctional adhesion of particular cell types. For example we could determine if a strongly adherentcell type secretes material that allows other types of cells in the population to also adhere morestrongly, or whether multiple species stimulate stronger adhesion in each other. As with thecharacterization studies, we would test populations with mixtures of dual, triple, and quadruplespecies.
We can also test the changes in cellular mucilage secretions by performing simple colorimetricor fluorescent staining procedures with lectins, alcian blue, or ruthenium red (as in [132]). In suchtests, the substrate surfaces will be removed from the flume after being exposed to water flow andthe cells stained with the desired reagent (e.g. 0.2 % w/v Ruthenium Red or 1% w/v Alcian Blue).After incubation and destaining, the surface will be examined under DIC or fluorescent optics. Thiswill allow us to determine the type of mucilage material secreted by the cells under different waterflow regimens and species distributions. By comparing the staining patterns with cells unexposedto water flow, we will be able to determine whether or not the stress of water flow, or the species

composition of the population, significantly change the mucilage material secreted by the cells.This information on mucilage composition/abundance can then be analyzed for any correlationswith the rate of cell detachment.
We also plan to determine relative rates of cell immigration onto substrates. In this case,water flow will be initiated prior to layering any cells onto the platform surface. Diatoms will thenbe inoculated into the flowing water. For short-term experiments, 5-25 ml of pond watercontaining predetermined concentrations of diatoms will be inoculated into the water just upstreamof the platform. The cells will be added using a pipet mounted onto a boom stand and placed intothe water upstream of the platform. The cells will be released using a peristaltic pump mountedonto the pipet to ensure reproducible release of cells. Trials of pipet release of diatoms into petriplates will be used to assess the fraction of cells within the pipet that typically remain adhered to thepipet walls and are not released into the water stream. After the release of cells into the water, thenumber of cells that settle and adhere onto the platform can then be observed by videomicroscopeand counted. For longer-term evaluation, a continuously running flume can be inoculated with alarge concentration of diatoms (approximately 107 cells, enough for a final concentration in theflume water of about 1000 cells/ml) and allowed to incubate (in the presence of illumination for cellgrowth) for several days to several weeks. The number of diatoms settling and adhering onto theplatform surface during this time can then be measured. Changes in cell concentrations due to cellgrowth can be determined by taking samples of the flume water each day and measuring the relativeconcentrations of each cell type present in the flowing water. To test for differential adhesion ontosurfaces, platforms can be made that are composed of two or more surfaces (e.g. half metal, halfglass), so that the relative number and species distribution colonizing onto each of the differentsurfaces can be determined. Such experiments will allow us to compare the emigration/immigrationabilities of diatom species with different surfaces, and find the effective range of water speedswithin which each of these species can remain adhered.
Analysis of Cell/Substratum Contact Sites
Our previous work has demonstrated dramatic differences in the adhesion of different diatomspecies, but did not resolve the direct cause of these differences. That is, differences in adhesioncan result from differences in adhesive strength of mucilage material, cohesion of mucilage strands,or sloughing rate of the strands from the cells. In order to resolve this issue, the number andlocation of cell/substratum contact sites occurring during cell movement must be determined. Weplan to measure contact sites directly using reflection interference contrast microscopy.
Reflection interference contrast (RIC) microscopy is a light microscopic technique that allowsthe determination of cell sites that are in closest contact with the substratum. RIC has been used toaccurately determine adhesion sites for a number of cell types including moving amoebae,fibroblasts, bacteria, and carcinoma cells in culture [e.g. 33,46,47]. The technique uses epi-fluorescent light to illuminate the specimen on the side of the coverslip to which it is adhered.Using the proper filters, RIC generates phase-dependent interference between the epi-illuminatedlight reflecting off of the coverslip/medium boundary, and the light reflected off the surface of thecell [100,107]. Thus, the further away the cell is from the coverslip, the more the phase differencebetween the two sets of light. Such a system results in an image of a gray or light colored cell withdark spots in the areas of closest cell contact with the substratum.
We plan to exploit this method of microscopy by analyzing motile cells in several ways: 1)measuring the number of contact sites per cell generated by different diatom species duringmovement; 2) analyzing the location of these contact sites (e.g. are they generated at one locationin the cell and then translocated down the raphe and removed); 3) determining the alteration in cellcontact sites as a cell changes direction (e.g. are old sites sloughed off as new sites are generatedwhich move in the opposite direction, or do the same contact sites remain and change course); and

4) determining the changes in contact site characteristics in the presence of changes inenvironmental conditions or species compositions in the population.
The analysis of cellular contact sites will enable us to directly answer several questions:
– How many contact sites are present in different types of motile diatoms? Our previous analysis hasshown considerable differences in adhesion between different types of motile diatoms, and using RIC, we willbe able to directly determine the number of contact sites generated by these different cell types. For example,we will be able to determine if strongly adhering species such as Stauroneis have more contact sites, or sitesthat persist longer, than that of the more weakly adhering Craticula.
– What is the pattern of contact sites in cells such as Pinnularia, in which the valve is broadly linear,but the path shape is strongly curved? What is the pattern of contact sites in a cell such as Surirella inwhich the valve/raphe shape is curved? With RIC we will be able to determine the location of cell contactsites relative to the valve and raphe, and better determine how such path orientations are generated.
– What is the difference in the distribution pattern of cell/substratum contacts between the raphe andnon-raphe valves of a monoraphid diatom such as Achnanthes? How do these differences correlate with themeasured adhesion of the two sides?
– How does light affect cell adhesion? Previous work has shown how light stimulation at the tips ofcells can alter cell direction [22]. Using RIC, we will be able to determine if light stimulated reversal of celldirection is driven by reversal of already formed contact sites, loss of particular contact sites, or generationof new sites.
Correlation of Lab and Field Studies
While the majority of the studies outlined in this proposal are based on work with culturedalgal cells under controlled lab conditions, we also recognize the importance of undertaking studiesthat connect the results of the experimental work with natural diatom behavior. To accomplishthis, we propose several stages of field related investigations both concurrently and subsequently tothe experiments proposed in this grant application.
First, we intend to extend our sample collection from our original source pond in order tocorrelate changes in diatom distribution over both time and position within the pond. Over thethree year course of this proposal we plan to take sets of samples from our source pond four timeseach year (Jan, April, July, October), with 6 sediment samples taken at each time period. Thesesamples will include two from the area immediately adjacent to the outflow of approx. 6-12 indepth, two samples from a nearby area of the pond with slower water flow of the same depth, andtwo from a deeper more mid-pond area about 12-24 in depth; the two at each site will be replicatesfor comparison of collection variability. These samples will be collected, fixed, and analyzed forrelative abundance of the main diatom species under study. At each collection site the rate of waterflow will be measured with a flow meter, and the light intensity at both the water surface andunderlying sediment surface will be recorded. In addition, two samples of top water will be takenand fixed for analysis of silicate, total nitrogen, nitrate, and phosphorus concentrations in the water.Some of each water sample will be glass filtered on site to determine the total vs. dissolvedconcentrations of each of the elements/nutrients. The pH and temperature of the pond will also betaken on site. Analysis of these samples will allow us to determine: 1) if there are naturaldifferences in diatom distributions between high water flow and low water flow sites in the pond; 2)if there are natural differences in diatom distributions between sites at different depths in the pond;3) if the relative distributions of the four diatom species changes during the course of the year; 4) ifthere are any changes in species distributions which correlate with changes in water chemistry andtemperature; and 5) whether these changes in spatial distribution in the pond correlate with thespecies-specific adhesion and motility characteristics and sensitivities we observe in the lab studies.
Based on the results of both our lab and initial field studies, I also plan to work withcolleagues both at DePaul and at other nearby institutions to determine local pond or stream sites

with similar types of diatom distributions. These sites will then become objects of futureinvestigations, carried out subsequent to those proposed in this grant, in which we plan toinvestigate diatom behaviors in natural settings. These experiments will include, for example,placing cleaned tiles or surfaces (as in 84,108,111) in several areas of the same stream that havedifferent flow rates. Each of these tiles can then be inoculated with a sample of diatoms from thesame single or multi-species culture, allowed to incubate for several hours, and then analyzed forthe distribution of diatoms remaining as compared to that present in the initial population. Similarexperiments can be performed by placing the tiles in areas of similar stream velocity but differentlight exposures (e.g. open, partially shaded, or fully shaded regions) and then comparing thediatoms remaining on each of the tiles. The cells on the inoculated tiles can also be compared tothe types of diatoms present initially at each of the stream sites, or with the populations colonizingcleaned tiles that are left at the same site but with no inoculation from diatom cultures. In addition,stream conditions can be manipulated (e.g. blocks placed upstream to regulate water flow orshading placed over areas of the stream) to further modify the diatoms' environmental conditions.These experiments will allow us to make more direct comparison to the behaviors of diatoms in labsettings vs. their behavior in natural settings.
Significance and Utility of Proposed Project
The information gleaned from the above experiments will significantly increase ourunderstanding of diatom motile behavior, and will begin to address several types of ecologicallyimportant questions, including:
• What environmental factors (e.g. light, pH, nutrients, temperature, substrate surface composition) moststrongly affect diatom motile behaviors in both single and multi species populations?
• In what way do the characteristics of movement for individual species allow them to successfullycompete for limited resources? Do the motile characteristics of species change when placed in mixedpopulations with diatoms of different speeds, adhesive strength, or other characteristics?
• In what way do cell behaviors correspond to the differences in the distribution and activity of cellcontact sites? In what way do the movement of cell contact sites correlate with differences in cellshape or structure (e.g. with the raphe structure)?
This project is clearly feasible. The PI of this project is well acquainted with the protocolsnecessary for isolating, culturing, growing, and handling diatoms, and has worked previously onmitosis, intracellular motility, reproduction, and physiological responses in diatoms [9,14,15,19,20].The PI has also used computer-assisted video microscopy to analyze motility [11,12,16], and istherefore well prepared to carry out all of the experiments outlined in this project. Additionally,this project can easily be broken up into smaller components that can be performed byundergraduate research assistants, so that they can be responsible for self-contained sets ofexperiments, while still being a part of an overall research project. In addition, this project shouldallow the development of more assays that can rapidly assess and compare the vitality of diatompopulations, and quickly determine the relative stress on diatom populations due to alterations inecological conditions (e.g. acidification, toxins, increased UV irradiation).
In summary, our plan to experimentally investigate the behaviors and physiologicalresponses of diatoms should form a strong foundation for understanding diatom activity and forfuture studies investigating success in natural settings. This approach has been shown to be usefulin integrating laboratory and field studies [29], and should provide a program that both enhancesour understanding of diatoms as well as one that helps carry out the major educational and societalobjectives of NSF.

I-2. RUI Impact Statement
Directly involving undergraduates in laboratory experimentation generates their enthusiasmand engages their minds like no other activity can. I continue to believe that it is incumbent upon allscientists to encourage and foster such opportunities. Over the past five years I have worked withover 20 undergraduates in my lab (see below), most of whom had the opportunity to becomedirectly involved with their own experiments. Through their research, along with related scientificdiscussions at weekly lab meetings and/or national meetings, I have attempted to foster an excitingenvironment where students are encouraged to question, explore and ponder. I encourage anatmosphere where the students are constantly questioning - asking what they are doing, why theyare doing it, and how to devise the next set of new experiments. This atmosphere of scientificenthusiasm is one of the reasons I am committed to the development of undergraduate researchopportunities at DePaul. It is only through ongoing research endeavors, made possible throughgrants such as the RUI program, that such undergraduate research opportunities are available on asteady and continual basis.
It is clear that many of the current problems in our world will require innovative solutionsby people with a sufficient amount of technological and scientific education. For many collegestudents, it is the ability to observe and participate directly in scientific research that provides thecontext for their science courses. Lectures can provide students with the language of science, butwithout active participation in the process of science, they will never learn to speak the language.Through active undergraduate research programs, universities such as DePaul that have a majorfocus on education can help to achieve the goal whereby both science and education can trulybecome integrated.
The project outlined in this proposal will provide an excellent opportunity for students whohave never before considered science to observe, participate, and become excited about answeringnew questions and solving new problems. Most aspects of diatom motility are still virtuallyunknown. As such, students working on this problem will not only have the stimulation of directparticipation in scientific inquiry, but will also have the excitement of making a direct and possiblysignificant contribution to our understanding of a group of ecologically important organisms. Someof my students have already made such contributions, and are authors on published manuscripts,abstracts, or manuscripts in preparation. I believe it is this attribute of science, the thrill ofdiscovering something that no one else in the world knows, that can truly generate the enthusiasmand excitement in students. Given the current apprehensions about science funding, it is likely thatonly through this personal excitement that students will seriously consider science as a personallyfulfilling career. Only by re-initiating the student's sense of curiosity can we show that research andeducation are part of the same process, and that a career in the sciences can lead to an enjoyablelifetime of accomplishment, learning, and exploration.
The Biology Department at DePaul is a fairly young department. Six of the nine full timefaculty have been hired within the past ten years, with three of them hired within the last threeyears. We expect to be replacing two more of the faculty positions within the next two years. Assuch, our department contains a large percentage of recently trained, research oriented, faculty whoare excited about both research and teaching. The enthusiasm is further heightened by our newEnvironmental Sciences/Biology building, which has increased the space for our teaching andresearch labs. Nonetheless, the number of undergraduate majors in our Biology program (approx.

200), along with the institutional commitment of a strong liberal arts education, continues to placeheavy constraints on time and departmental resources for research. These constraints have becomeparticularly evident in the last year, as the University is mid-way through the implementation of anew general education program that includes freshman seminar specialty courses and laboratoryscience courses for all DePaul undergraduates. Such new general education courses, while usefulin promoting scientific principles to students, have put increased burdens on the faculty time anddepartmental resources allocated to research. Therefore, if the excitement of researchopportunities for Biology undergraduates at DePaul is to be maintained, it will be necessary toobtain funding from outside sources.
The previous RUI grant has clearly had an impact on our undergraduate research environmentat DePaul. Below I have listed the DePaul undergraduates who have worked in my lab over thepast four years, along with their current status (if known) and publication &/or ASCB meetingattendance also noted.
(Current Students in Bold)(* = co-authors on abstracts or manuscripts)
(‡ = attended annual American Society for Cell Biology Meeting)
Sarah Corradino - Undergraduate Student - TransferredJoshua Crea - Current DePaul Undergraduate
Nicholas Disparti*‡ - Division Manager at Leica Microscopes Inc.
Brian Hesler‡ - Recent DePaul graduateKarie Jeisel - Former DePaul UndergraduateMelanie Jopek - Current DePaul UndergraduateBill Kosmala - Medical Doctor
Matt Lazzarotto*‡ - Newspaper ReporterTonette Love - Former DePaul Undergraduate
Jim McGuire*‡ - Former DePaul Undergraduate and Master's Student
Joey Mucha‡ - Former DePaul UndergraduateEric Nelson - Abbot LaboratoriesAlex Nesterov - Former Undergraduate Student - TransferredAimee Norris* - Veterinary SchoolMoira Silverman - Current DePaul Undergraduate
Christine Skoczylas‡ - In Univ. of Chicago Neurobiology Program
Jamie Vaeth‡ - Recent DePaul graduateMark Vaselakos - Osteopathic Medical Doctor
Dan Weber‡ - Entering Dental SchoolBernadeta Wibosono - Current DePaul UndergraduateTroy Woodard - Entering Medical School
Ayesha Worsham‡ - Recent DePaul graduate
In addition, I also have had one high school student, Meghan O’Connor, who worked in the lab thispast summer (1999).

Our department graduates 20-40 undergraduate majors per year, many of whom go on toadvanced degrees in Pharmacy, Medicine, Dentistry, and other allied health fields, as well as severaleach year who enter Biology Ph.D. programs. We also educate students from allied fields such asEnvironmental Science, Biochemistry, and Biology Education. With the funding of this proposal,the department will be able to continue its tradition of providing a high quality, broadly basedbackground in Biology, and further advance the opportunities for undergraduates like those aboveto actively engage in scientific research. The current lack of an RUI grant for my lab has alreadybegun to have an impact. Last year, for the first time in several years, no students accompanied meto the annual Cell Biology meeting. This was partially due to the lack of any RUI funding to aidstudent travel.
This proposal will also provide the funding for equipment that is needed for the furtherdevelopment of modern cell biology at DePaul. Equipment purchased from the previous RUI granthas already been used by several of the other faculty members in the department. The larger piecesof equipment requested in this proposal (inverted microscope, video dissecting microscope head)will similarly be available for use by other faculty members and thereby serve students with a widevariety of interests. Such joint usage of resources attests to the close relationship among DePaulBiology faculty, and how improvement of equipment resources for one lab benefits the entiredepartment.
In addition, the equipment requested furthers my ability to film a number of cellular andmicroscopic phenomena for educational purposes. Over the last three years, using the equipmentfrom the last RUI grant, I have recorded a number of video sequences of diatom (and other cell)motility which has been used for demonstration in General Biology, Cell Biology, Cell Motility, andPhycology courses. The equipment has also been used to make instructional videos for a localelementary school. The requested instruments will further our ability to film and edit cellularprocesses for placement on the Internet. We are already in the process of developing a site for ourdiatom research that we hope in the future will include a number of QuickTime video sequences ondiatom movement.
In summary, the past NSF-RUI grant has had a significant impact on the Biology program atDePaul, as well as the overall science community here, in several ways:
• The equipment purchased (e.g. time-lapse video recorder, temperature controlled stage) hasaided several research labs with their work, and has allowed us to film several types ofprocesses that we were previously unable to record such as fish, sea urchin, and frogdevelopment. It has also allowed us to make demonstration videos for several classes.
• The salaries allowed me to fund a number of undergraduates on a part-time basis, several ofwhom have been co-authors on manuscripts or abstracts, as well as assist the travel of severalundergraduates to the annual meeting of the American Society for Cell Biology.
• The grant allowed me the access of a full-time technician, a position that has been instrumentalin the ability to train the undergraduates, and have a continuous research program going. Thetechnicians have also provided unique insights during discussions of research at our weeklylab meetings, and provided strong role models for the undergraduates.

The RUI grant as proposed, would allow us to continue and expand this strong scienceenvironment at DePaul, and continue to encourage students into careers of science and technology.

D - References Cited
1. Becker, K. (1996) Exopolysaccharide production and attachment strength of bacteria and diatoms onsubstrates with different surface tensions. Microb. Ecol. 32, 23-33.
2. Bertrand J. (1992) Mouvements des diatomées. II - Synthèse des mouvements. Cryptogamie Algol. 13,49-71.
3. Biggs B.J.F. and Thomsen (1995) Disturbance of stream periphyton by perturbations in shear stress:Time to structural failure and differences in community resistance. J. Phycol 31, 233-241.
4. Bothwell M.L., Suzuki K.E., Bolin M.K. and Hardy F.J. (1989) Evidence of dark avoidance byphototrophic periphytic diatoms in lotic systems. J. Phycol. 25, 85-94.
5. Boyle J.A., Pickett-Heaps J.D. and Czarnecki, D.B. (1984) Valve morphogenesis in the pennate diatomAchnanthes coarctata. J. Phycol. 20, 563-573.
6. Bray D. (1992) Cell Movements . Garland Pub., New York, 406 pp.
7. Caduto M.J. (1990) Pond and Brook. A guide to nature in freshwater environments. University Pressof New England, Hanover, 276 pp.
8. Cohn S.A. (1993) Light dependent effects on diatom motility. Mol. Biol. Cell 4(S), 168a.
9. Cohn S.A. and Disparti N.C. (1994) Environmental factors influencing diatom cell motility. J. Phycol.30, 818-828.
10. Cohn S.A., Dunbar S.A., Skoczylas C. and Mucha, J.A. (1997) Comparative Analysis of DiatomMotility: Phototactic Behavior and Sensitivity to Ultraviolet Radiation. Mol. Biol. Cell 8(S), 386a.
11. Cohn S.A., Ingold A.L., and Scholey J.M. (1987) Correlation between the ATPase and microtubuletranslocating activities of sea urchin egg kinesin. Nature 328, 160-163.
12. Cohn S.A., Ingold A.L., and Scholey J.M. (1989) Quantitative analysis of sea urchin egg kinesin-driven microtubule motility. J. Biol. Chem. 264, 4290-4297.
13. Cohn, S.A. and McGuire, J.R. (1994) Use of diatom motility assays for toxicity testing. Mol. Biol. Cell5(S), 485a.
14. Cohn S.A., Nash J., and Pickett-Heaps J.D. (1989) The effect of drugs on diatom valve morphogenesis.Protoplasma 149,130-143.
15. Cohn S.A., and Pickett-Heaps J.D. (1988) The effects of colchicine and dinitrophenol on the in vivorates of anaphase A and B in the diatom Surirella. Eur. J. Cell Biol. 46, 523-530.
16. Cohn S.A., Saxton W.M., Lye R.J. and Scholey J.M. (1993) Analyzing Microtubule Motors in RealTime. Meth. Cell. Biol. 39, 75-88.
17. Cohn, S.A. and Sciortino, S. (1998) Examination of motile diatom loss from differentsubstrates due to the force of water flow. Mol. Biol. Cell 9(S): 34a.

18. Cohn S.A., Spurck T.P., and Pickett-Heaps J.D (1999) High energy irradiation at the leading tip ofmoving diatoms causes a rapid change of cell direction. Diatom Research (in press)
19. Cohn S.A., Spurck T.P., Pickett-Heaps J.D. and Edgar L.A. (1989) Perizonium and initial valveformation in the diatom Navicula cuspidata (Bacillariophyceae). J. Phycol. 25, 15-26.
20. Cohn S.A. and Weitzell R.E. Jr. (1996) Ecological considerations of diatom cell motility: I.Characterization of motility and adhesion in four diatom species. J. Phycol. 32, 928-939.
21. Cohn S.A., Weitzell R.E. Jr., Norris A. and Lazzarotto M.J. (1996) Comparative analysis of adhesionand photo-stimulated aggregation in pennate diatoms. Mol. Biol. Cell 7(S), 232a.
22. Cohn S.A., Weitzell R.E. Jr., Spurck T.P., and Pickett-Heaps J.D. (1995) Characterization of motilityand adhesion in pennate diatoms. Mol. Biol. Cell 6(S), 261a.
23. Cooksey B. and Cooksey K.E. (1980) Calcium is necessary for motility in the diatom Amphoracoffaeformis. Plant Physiol. 65, 129-131.
24. Cooksey B. and Cooksey K.E. (1988) Chemical signal-response in diatoms of the genus Amphora. J.Cell Sci. 91, 523-529.
25. Cooksey K.E. (1981) Requirement for calcium in adhesion of a fouling diatom to glass. Appl. & Envir.Microbiol. 41, 1378-1382.
26. Cooksey K.E. and Cooksey B. (1986) Chapt. 3 Adhesion of fouling diatoms to surfaces: somebiochemistry. In Algal Biofouling (Evans L.V. and Hoagland K.D. Eds.) Elsevier, New York.
27. Cooksey K.E. and Wigglesworth-Cooksey B. (1995) Adhesion of bacteria and diatoms to surfaces in thesea: a review. Aquatic Microbial Ecology 9,87-96.
28. Cox E.J. (1977) Raphe structure in naviculoid diatoms as revealed by the scanning electron microscope.Nova Hedwigia Beih. 54, 261-274.
29. Cox E.J. (1993) Freshwater diatom ecology: developing an experimental approach as an aid tointerpreting field data. Hydrobiol. 269/270, 447-452.
30. Cox E.J. (1996) Identification of freshwater diatoms from live material. Chapman and Hall Pub., NewYork. 158 pp.
31. Crocker K.M. and Passow U. (1995) Differential aggregation of diatoms. Marine Ecology Prog. Ser.117, 249-257.
32. Crompton M. (1990) Role of mitochondria in intracellular calcium regulation. In Intracellular Calcium
Regulation. [F. Bronner, ed.] Alan R. Liss, New York. Pp 181-210.
33. Curtis A.S.G. (1964) The mechanism of adhesion of cells to glass. A study by interference reflectionmicroscopy. J. Cell Biol. 20, 199-215.
34. Darley W.M. (1982) Algal Biology: A physiological approach. Blackwell Scientific Pub., London, 168pp.

35. Drum R.W. and Hopkins J.T. (1966) Diatom locomotion: an explanation. Protoplasma 62, 1-32.
36. Dudley T.L. and D'Antonio C.M. (1991) The effects of substrate texture, grazing, and disturbance onmacroalgal establishment in streams. Ecology 72, 297-309.
37. Edgar L.A. (1982) Diatom Locomotion: a consideration of movement in a highly viscous situation. Br.Phycol. J. 17, 243-251.
38. Edgar L.A., and Pickett-Heaps J.D. (1982) Ultrastructural localization of polysaccharides in the motilediatom Navicula cuspidata. Protoplasma 113, 10-22.
39. Edgar L.A. and Pickett-Heaps J.D. (1983) The mechanism of diatom locomotion. I. An ultrastructuralstudy of the motility apparatus. Proc. Roy. Soc. Lond. B. 218, 331-343.
40. Edgar L.A., and Pickett-Heaps J.D. (1984) Valve morphogenesis in the pennate diatom Naviculacuspidata. J. Phycol. 20, 47-61.
41. Edgar L.A., and Pickett-Heaps J.D. (1984) Diatom Locomotion. In Progress in phycological research,Vol 3. [Round, F.E. and Chapman, D.J., eds.] Biopress Ltd, Bristol. Pp 47-88.
42. Edgar L.A. and Zavortink M. (1983) The mechanism of diatom locomotion. II. Identification of actin.Proc. Roy. Soc. Lond. B. 218, 345-348.
43. Edlund M.B. and Stoermer E.F. (1997) Ecological, evolutionary, and systematic significance of diatomlife histories. J. Phycol. 33, 897-918.
44. Fritz L., Quilliam M.A., Wright J.L.C., Beale A.M., and Work T.M. (1992) An outbreak of domoicacid poisoning attributed to the pennate diatom Pseudonitzschia australis. J. Phycol. 28, 439-442.
45. Garrahan P.J. and Rega A.F. (1990) Plasma membrane calcium pump. In Intracellular CalciumRegulation. [F. Bronner, ed.] Alan R. Liss, New York. Pp 271-303.
46. Goldman W.H., Schindl M., Cardozo T.J. and Ezzell R.M. (1995) Motility of vinculin-deficient F9embryonic carcinoma cells analyzed by video, laser confocal, and reflection interference contrastmicroscopy. Exp. Cell Research 221, 311-319
47. Goodwin S.L., Fletcher M. and Burchard R.P. (1989) Interference reflection microscopic study of sitesof association between gliding bacteria and glass substrata. J. Bacteriol. 171, 4589-4594.
48. Gordon R. (1987) A retaliatory role for algal projectiles, with implications for the mechanochemistry ofdiatom gliding motility. J. Theoret. Biol. 126, 419-436.
49. Gordon R., Björklund N.K., Robinson G.G.C. and Kling H.J. (1996) Sheared drops and pennatediatoms. Nova Hedw. Beih. 112, 289-299.
50. Gordon R. and Drum, R.W. (1970) A capillarity mechanism for diatom gliding locomotion. Proc.Natl. Acad. Sci. (USA) 67, 338-44.

51. Guerrini F., Mazzotti A., Boni L and Pistocchi R. (1998) Bacterial-algal interactions in polysaccharideproduction. Aquat. Microbial Ecol. 15, 247-253.
52. Häder D.-P. and Hoiczyk E. (1992) Chapter 1. Gliding Motility. In Algal Cell Motility (M. MelkonianEd.) Chapman and Hall, New York. Pp 1-38.
53. Hansen J.L.S., Timm U., and Kiørboe T. (1995) Adaptive significance of phytoplankton stickiness withemphasis on the diatom Skeletonema costatum. Marine Biology 123, 667-676.
54. Harper M.A. (1977) Chapter 8: Movements. In The Biology of Diatoms. (D.Werner Ed.) Univ. of Calif.Press, Berkeley. Pp 224-249.
55. Harper M.A. and Harper J.F. (1967) Measurements of diatom adhesion and their relationship withmovement. Br. Phycol. Bull. 3,195-207.
56. Haupt W. (1982) Light Mediated Movement of Chloroplasts. Ann Rev. Plant. Physiol. 33, 205-233.
57. Hoagland K.D., Rosowski J.R., Gretz M.R. and Roemer S.C. (1993) Diatom extracellular polymericsubstances: Function, fine structure, chemistry, and physiology. J. Phycol. 29, 537-66.
58. Ingold A.L., Cohn S.A., and Scholey J.M. (1988) Inhibition of kinesin-driven microtubule motility bymonoclonal antibodies to kinesin heavy chains. J. Cell Biol. 107, 2657-2667.
59. Jarosch R. (1962) Gliding. In Physiology and Biochemistry of Algae (R.A. Lewin ed.). Academic Press,NY. Pp 573-581.
60. Johnson R.E., Tuchman N.C. and Peterson C.G. (1996) Changes in the vertical microdistribution ofdiatoms within a developing periphyton mat. J. North Am. Benthol. Soc. 16, 503-519.
61. Jørgensen B.B. and DesMarais D.J. (1988) Optical properties of benthic photosynthetic communities:Fiber optic studies of cyanobacterial mats. Limnol. Oceanogr. 33, 99-113.
62. Jørgensen B.B., Revsbech N.P., and Cohen Y. (1983) Photosynthesis and structure of benthic microbialmats: Microelectrode and SEM studies of four cyanobacterial communities. Limnol. Oceanogr. 28,1075-1093.
63. Kamiya N. (1981) Physical and chemical basis of cytoplasmic streaming Ann. Rev. Plant Physiol. 32,205-236.
64. Korn E.D. and Hammer J.A. (1988) Myosins of nonmuscle cells. Ann. Rev. Biophys. Biophys. Chem.17, 23-45.
65. Korn E.D. and Hammer J.A. (1990) Myosin I. Curr. Opin. Cell Biol. 2, 57-61.
66. LaClaire J.W. II, Chen R. and Herrin D.L. (1995) Identification of a myosin-like protein inChlamydomonas Reinhardtii (Chlorophyta). J. Phycol. 31, 302-306.
67. Lauterborn R. (1896) Untersuchungen über Bau, Kernteilung und Bewegung der Diatomeen. W.Englemann Pub., Leipsig.

68. Lind J.L., Heimann K., Miller E.A., van Vliet C., Hoogenraad N.J., and Wetherbee R. (1997)Substratum adhesion and gliding in a diatom are mediated by extracellular proteoglycans. Planta 203,213-221.
69. Lind J.L, vanVliet C.J., Bacic A. and Wetherbee R. (1996) Characterization of molecules responsiblefor cell substrate adhesion in marine raphid diatoms. Mol. Biol. Cell 7(S), 56a.
70. Lock M.A., Wallace R.R., Costerton J.W., Ventullo R.M. and Charlton S.E. (1984) River epilithon:toward a structural-functional model. OIKOS 42, 10-22.
71. Mann, D.G. and Stickle, A.J. (1991) The genus Craticula. Diatom Res. 6, 79-107.
72. McCormick P.V. and Stevenson R.J. (1991) Mechanisms of benthic algal succession in loticenvironments. Ecology 72, 1835-1848.
73. Müller O. (1893) Die Ortsbewegung der Bacillariaceen betreffend. I. Ber. Dt. Bot. Ges. 11, 571-576.
74. Nultsch W. (1971) Phototactic and photokinetic action spectra of the diatom Nitschia communis.Photochem. Photobiol. 14, 705-712.
75. Nultsch W. (1975) Phototaxis and Photokinesis. In Primitive Sensory and Communication Systems(M.J. Carlile, ed.). Academic Press, NY. Pp 29-90.
76. Nultsch W. and Häder D.-P. (1988) Photomovement in motile microorganisms - II. Photochem.Photobiol. 47, 837-869.
77. Ohtsuka T. (1998) Variation in diatom community structure among habitats within a morphologicalunit in a river. Jap. J. Limnol. 59, 311-328.
78. Pahlow M., Riebesell U., and Wolf-Gladrow D.A. (1997) Impact of cell shape and chain formation onnutrient acquisition by marine diatoms. Limnol. Oceanogr. 42, 1660-1672.
79. Patrick R. (1976) The formation and maintenance of benthic diatom communities. Proc. Am. Phil. Soc.120, 475-84.
80. Patterson D.M. (1986) The migratory behavior of diatom assemblages in a laboratory tidal micro-ecosystem examined by low temperature scanning electron microscopy. Diatom Research 1, 227-239.
81. Patterson D.M. (1989) Short-term changes in the erodibility of intertidal cohesive sediments related tothe migratory behavior of epipelic diatoms. Limnol. Oceanogr. 34, 223-234.
82. Perl T.M., Bédard L., Kosatsky T., Hockin J.C., Todd E.C.D. and Remis, R.S. (1990) An outbreak oftoxic encephalopathy caused by eating mussels contaminated with domoic acid. N. Engl. J. Med. 322,1775-80.
83. Peterson C.G. (1987) Influences of flow regime on development and dessication response of loticdiatom communities. Ecology 68, 946-954.
84. Peterson C.G. and Hoagland K.D. (1990) Effects of wind-induced turbulence and algal mat developmenton epilithic diatom succession in a large reservoir. Arch. Hydrobiol. 118, 47-68.

85. Peterson C.G. and Stevenson R.J.. (1989) Substratum conditioning and diatom colonization in differentcurrent regimes. J. Phycol. 25, 790-793.
86. Pickett-Heaps J.D. (1983) Valve morphogenesis and the microtubule center in three species of thediatom Nitzschia. J. Phycol. 19, 269-281.
87. Pickett-Heaps J.D. (1991) Post-mitotic cellular reorganisation in the diatom Cymatopleura solea: Therole of microtubules and the microtubule center. Cell Motil. Cytoskel. 18, 279-292.
88. Pickett-Heaps J.D., Cohn S., Schmid A-M. and Tippit D.H. (1988) Valve morphogenesis in Surirella(Bacillariophyceae). J. Phycol. 24, 35-49.
89. Pickett-Heaps J.D., Hill D.R.A., and Blaze K.L. (1991) Active gliding motility in an araphid marinediatom Ardissonea (Formerly Synedra) crystallina. J. Phycol. 27, 718-725.
90. Pickett-Heaps J.D., Schmid A.M. and Edgar L.A. (1990) The cell biology and phylogeny of diatomvalve formation. In Progress in phycological research. Vol 7. [Round, F.E. and Chapman, D.J., eds.]Biopress Ltd, Bristol.
91. Pickett-Heaps J.D., Spurck T.P., Cohn S.A., Schoeller A. and Edgar L.A. (1984). Sexual reproductionin the diatom Navicula cuspidata. 16mm color, sound film. 16 min. Cytographics % Dr. J.D. Pickett-Heaps, Dept. of Botany, University of Melbourne, Parkville, Victoria 3052 AUSTRALIA.
92. Pickett-Heaps J.D., Tippit D.H., and Andreozzi J.A. (1979) Cell division in the pennate diatomPinnularia. III. The valve and associated cytoplasmic organelles. Biol. Cell. 35, 195-198.
93. Pickett-Heaps J.D., Tippit D.H., and Andreozzi J.A. (1979) Cell division in the pennate diatomPinnularia. IV. Valve morphogenesis. Biol. Cell. 35, 199-203.
94. Preston T.M., King C.A., and Hyams J.S. (1990) The Cytoskeleton and Cell Motility. Blackie Pub.,London.
95. Reid M.A., Tibby J.C., Penny D. and Gell P.A.. (1995) The use of diatoms to assess past and presentwater quality. Australian J. Ecol. 20, 57-64.
96. Round F.E. (1991) Diatoms in river monitoring studies. J. Appl. Phycol. 3, 129-145.
97. Round F.E. and Crawford R.M. (1990) Chapter 31. The Bacillariophyta in Handbook of Protoctista [L.Margulis and J. Corliss, M. Melkonian, and D.J. Chapman eds.], Jones and Bartlett, Boston, pp. 574-599.
98. Round F.E., Crawford R.M., and Mann D.G. (1990) The Diatoms. Biology and Morphology of theGenera. Cambridge Univ. Press, Cambridge.
99. Round F.E. and Palmer J.D. (1966) Persistent vertical migration rhythms in benthic microflora. II. Fieldand laboratory studies on diatoms from the bank of the river Avon. J. Mar. Biol. Assoc. UK. 46, 191-224.
100. Rubbi C.P. (1994) Light Microscopy: Essential Data. John Wiley & Sons, New York.

101. Saxton W.M., Porter M.E., Cohn S.A., Scholey J.M., Raff E.C., and McIntosh J.R. (1988)Drosophila kinesin: characterization of microtubule motility and ATPase. Proc. Natl. Acad. Sci. USA.85, 1109-1113.
102. Schmid A-M.M. (1980) Valve morphogenesis in diatoms: pattern related filamentous system in pennatesand the effect of APM, colchicine and osmotic pressure. Nova Hedw. 33, 811-847.
103. Schmid A-M. M. (1995) Aspects of morphogenesis and function of diatom cell walls with implicationsfor taxonomy. In Wetherbee, R. Andersen, R.A. & Pickett-Heaps, J.D. [Eds.] The Protistan CellSurface. Springer-Verlag, New York, pp. 43-60.
104. Schrader H.-J. (1974) Types of raphe structures in the diatoms. Nova Hedwigia Beih. 45, 195-217.
105. Sheetz M.P. and Spudich J.A. (1983) Movement of myosin-coated fluorescent beads on actin cables invitro. Nature 303, 31-35.
106. Smith H.L. (1888) A contribution to the life history of the Diatomaceae-part II. Proc. Amer. Soc. Micr.9, 126-167.
107. Slayter E.M. and Slayter H.S. (1992) Light and Electron Microscopy. Cambridge University Press,Cambridge.
108. Stevenson R.J. (1990) Benthic algal community dynamics in a stream during and after a spate. J. N.Amer. Benthol. Soc. 9, 277-288.
109. Stevenson R.J. (1998) Diatom indicators of stream and wetland stressors in a risk managementframework. Envir.. Monitor. and Assess. 51, 107-118.
110. Stevenson R.J., Bothwell M.L. and Lowe R.L. [Eds.] (1996) Algal Ecology. Freshwater BenthicEcosystems. Academic Pres, San Diego. 753 pp.
111. Stevenson R.J. and Peterson C.G. (1989) Variation in benthic diatom (Bacillariophyceae) immigrationwith habitat characteristics and cell morphology. J. Phycol. 25, 120-129.
112. Stevenson R.J., Peterson C.G., Kirschtel D.B., King C.C., and Tuchman N.C. (1991) Density-dependent growth, ecological strategies, and effects of nutrients and shading on benthic diatomsuccession in streams. J. Phycol. 27, 59-69.
113. Tanaka N. (1986) Adhesive strength of epiphytic diatoms on various seaweeds. Bull. Jap. Soc. Sci.Fisheries. 52, 817-821.
114. Tsien R.W. and Tsien R.Y. (1990) Calcium channels, stores, and oscillations. Ann. Rev. Cell Biol. 6,715-760.
115. Tuchman N.C. (1996) Role of heterotrophy in algae. In Algal Ecology. Freshwater BenthicEcosystems. [Stevenson R.J., Bothwell M.L. and Lowe R.L. Eds.] Academic Press, San Diego. Pp 299-319.
116. Vandekerckhove J., Deboben A., Nassal M. and Wieland T. (1986) The phalloidin binding site of F-actin. EMBO J. 4, 22815-22818.

117. Wagner G. and Klein K. (1981) Mechanism of chloroplast movement in Mougeotia. Protoplasma 109,169-185.
118. Waite A., Gallager S. and Dam H.G. (1997) New measurements of phytoplankton aggregation in aflocculator using videography and image analysis. Marine Ecol. Prog. Ser. 155, 77-88.
119. Wang Y., Lu J., Mollet J-C., Gretz M.R. and Hoagland K.D. (1997) Extracellular matrix assembly indiatoms (Bacillariophyceae). II. 2,6, Dichlorobenzonitrile inhibition of motility and stalk production inthe marine diatom Achnanthes longpipes. Plant Physiol. 113, 1071-1080.
120. Webster D.R., Cooksey K.E., and Rubin R.W. (1985) An investigation of the involvement ofcytoskeletal structures and secretion in gliding motility of the marine diatom, Amphora coffaeformis.Cell Motil. 5, 103-122.
121. Wenderoth K. (1982) Photokinese und photophobische Reaktionen der Kieselalge Navicula peregrina.Biol. Film #C1388. Institut für den Wissenschaftlichen Film, Postfach 2351, 37013 Göttingen,Germany.
122. Wenderoth K. (1982) Reaktion der Kieselalge Navicula peregrina auf Belichtung verschiedenerZellabschnitte. Biol. Film #C1466. Institut für den Wissenschaftlichen Film, Postfach 2351, 37013Göttingen, Germany.
123. Wenderoth K. (1983) Phototaxis bei Desmidiaceen und Diatomeen. Biol. Film #C1496. Institut fürden Wissenschaftlichen Film, Postfach 2351, 37013 Göttingen, Germany.
124. Wenderoth K. (1984) Wirkungsspektrum der Step-down-Reaktion bei Diatomeen. Biol. Film#C1520. Institut für den Wissenschaftlichen Film, Postfach 2351, 37013 Göttingen, Germany.
125. Weiss G.B. (1974) Cellular Pharmacology of Lanthanum Ann. Rev. Pharmacol. 14, 343-354.
126. Werner D. (Ed.) (1977) The Biology of Diatoms. University of Calif. Press, Berkeley.
127. Wetherbee R., Lind J.L., Burke J. and Quatrano R.S. (1998) The first kiss: Establishment and control ofinitial adhesion by raphid diatoms. J. Phycol. 34, 9-15.
128. Wetherbee R., Lind J.L., Poulsen N.C. and Spurck T.P. (1996) Cell motility in marine raphid diatomsappears to be actin-based. Mol. Biol. Cell 7(S), 560a.
129. Williamson R.E. (1992) Cytoplasmic streaming in Characean algae: Mechanism, regulation by Ca2+,and organization. In Melkonian, M. [Ed.] Algal Cell Motility , Chapman and Hall Publ., New York,pp. 73-98.
130. Wordeman L. (1992) Chapter 3: The cytoskeleton of the diatoms: The mitotic spindle and cell cycledependent organization. In The Cytoskeleton of the Algae (D. Menzel Ed.) CRC Press, Boca Raton. Pp39-58.
131. Wordeman L., McDonald K.L., and Cande W.Z. (1986) The distribution of cytoplasmic microtubulesthroughout the cell cycle of the centric diatom Stephanopyxis turris: Their role in nuclear migration andpositioning the mitotic spindle during cytokinesis. J. Cell Biol. 102, 1688-1698.

132. Wustman B.A., Gretz M.R. and Hoagland K.D. (1997) Extracellular matrix assembly in diatoms(Bacillariophyceae). I. A model of adhesives based on chemical characterization and localization ofpolysaccharides from the marine diatom Achnanthes longpipesand other diatoms. Plant Physiol. 113,1059-1069.

BIOGRAPHICAL SKETCHA. Vitae
Stanley A. CohnDePaul University - Department of Biological Sciences2325 N. Clifton Ave, Chicago, IL 60614phone: 773-325-7597 fax: 773-325-7596e-mail: [email protected] Sec # 521-90-8230
EducationB.S. Chemistry, with honors, California Institute of Technology (Caltech), 1979Ph.D. Molecular, Cellular, and Developmental Biology, Univ. of Colo. (Boulder), 1986
Thesis: Mechanisms of Mitosis and Valve Morphogenesis in DiatomsThesis Advisor: Dr. Jeremy Pickett-Heaps
Positions HeldDePaul University, Department of Biological Sciences
Associate Professor with Tenure 1996-presentAssistant Professor 1989-1996
Postdoctoral Research Associate, with Dr. Jonathan ScholeyNational Jewish Center for Immunology and Respiratory Medicine, 1986-1989
Graduate Research Assistant, with Dr. Jeremy Pickett-HeapsUniv. of Colo. (Boulder), 1981-1986
Teaching Assistant, Intro. to Molecular, Cellular, and Developmental BiologyUniv. of Colo. (Boulder), 1979-1981.
Fellowships, Awards, and OfficesAmerican Cancer Society Postdoctoral Fellowship, 1987-1989University of Colorado Graduate Fellowship, 1984-85N.I.H. Graduate Training Grant, 1979-80, 1983-85Silver Medal, Royal Society of Arts, 1979Senior Class President, Caltech, 1978-79
MembershipsFellow of the Royal Society of Arts,American Society for Cell BiologyAmerican Association for the Advancement of Science (AAAS)International Society for Diatom ResearchCouncil on Undergraduate Research
Grants AwardedDePaul University
College of Liberal Arts & Sciences Summer Grant (for summer 1990, 1992, 1994, 1997,1999)
University Research Council, Competitive Research Grant (1989,1990,1991,1992, 1998,1999)
Quality of Instruction Council (QIC) Development Grant (1991, 1998)University Research Council Competitive Research Leave (1995)Joint Quality of Instruction Council/University Research Council Grant (1995)
National Science FoundationGrant # IBN-IBN-9407279 - July 1994 to June 1997 - Physiology and Ecology of DiatomMotility - total award $210,000

B. Five Most Relevant Publications (Undergraduate Authors with asterisk):Cohn, S.A. and Weitzell, R.E. Jr. (1996). Ecological considerations of diatom cell motility: I.
Characterization of motility and adhesion in four diatom species. J. Phycol. 32: 928-939.Cohn, S.A. and Disparti, N.C.* (1994). Environmental factors influencing diatom cell motility.
J. Phycol. 30: 818-828.Cohn, S.A., Spurck, T.P., Pickett-Heaps, J.D. and Edgar, L.E. (1989). Perizonium and initial
valve formation in the diatom Navicula cuspidata (Bacillariophyceae). J. Phycol. 25: 15-26.
Cohn, S.A., Nash, J.* and Pickett-Heaps, J.D. (1989). The effect of drugs on diatom valvemorphogenesis. Protoplasma 149: 130-143.
Cohn, S.A. and Pickett-Heaps, J.D. (1988). The effects of colchicine and dinitrophenol on thein vivo rates of anaphase A and B in the diatom Surirella. Eur. J. Cell Biol. 46: 523-530.
Other Significant Publications:Cohn, S.A., Saxton, W.M., Lye, R.J., and Scholey, J.M. (1993) Chapter 5: Analyzing
Microtubule Motors in Real Time. In Meth. Cell. Biol. Vol .39 [J.M. Scholey Ed.],Academic Press, NY, Pp 75-88.
Cohn, S.A. (1990) The mechanochemistry of kinesin: a review. Mol. Chem. Neuropath. 12:83-94.
Cohn, S.A., Ingold, A.L. and Scholey, J.M. (1989). Quantitative analysis of sea urchin eggkinesin-driven microtubule motility. J. Biol. Chem. 264: 4290-4297.
Pickett-Heaps, J.D., Cohn, S., Schmid, A-M. and Tippit, D.H. (1988). Valve morphogenesis inSurirella (Bacillariophyceae). J. Phycol. 24: 35-49.
Cohn, S.A., Ingold, A.L. and Scholey, J.M. (1987). Correlation between the ATPase andmicrotubule translocating activities of sea urchin egg kinesin. Nature 328: 160-163.
C. Collaborators:Dr. Nancy Tuchman, Loyola University Chicago (collaborator on flume experiments)Dr. Donat Häder, Institut fur Botanik und Pharmazeutische Biologie, Germany (submitting a
chapter in a volume he is editing)
D. Graduate Students:Graduate Students Advised (Total = 12):
Dissertation Committee: Lydia Armstrong, PhD, University of Denver, 1989Thesis Committee: Shylaja Muthyala, MS, DePaul University, 1990
Ankita Chitre, MS, DePaul University, 1991Jolie Machota, MS, DePaul University, 1993John Sikora, MS, DePaul University, 1994Mary McCarthy, MS, DePaul University 1994Margaret Liotta, MS, DePaul University, 1998Tim Laurie, MS, DePaul University, 1998Heather Walczak, MS, DePaul University, 1999
Thesis Advisor: Devry Spreitzer, MS, DePaul University, 1995 (Co-advisor)Sam Sciortino, Current MS Student DePaul UniversityJason Weiss, Current MS Student DePaul UniversityDavid Zelner, Current MS Student DePaul University
E. Graduate/Postdoctoral Advisors:Dr. J. R. McIntosh, University of Colorado, Boulder (Dissertation Committee- 2nd reader)Dr. Jeremy D. Pickett-Heaps, University of Melbourne, AUSTRALIA (Graduate Advisor)Dr. Jonathan M. Scholey, University of California, Davis (Postdoctoral Advisor)

SUMMARYPROPOSAL BUDGET
FundsRequested By
proposer
Fundsgranted by NSF
(if different)
Date Checked Date Of Rate Sheet Initials - ORG
NSF FundedPerson-mos.
FOR NSF USE ONLYORGANIZATION PROPOSAL NO. DURATION (months)
Proposed Granted
PRINCIPAL INVESTIGATOR / PROJECT DIRECTOR AWARD NO.
A. SENIOR PERSONNEL: PI/PD, Co-PI’s, Faculty and Other Senior Associates (List each separately with title, A.7. show number in brackets) CAL ACAD SUMR
$ $1.
2.
3.
4.
5.
6. ( ) OTHERS (LIST INDIVIDUALLY ON BUDGET JUSTIFICATION PAGE)
7. ( ) TOTAL SENIOR PERSONNEL (1 - 6)
B. OTHER PERSONNEL (SHOW NUMBERS IN BRACKETS)
1. ( ) POST DOCTORAL ASSOCIATES
2. ( ) OTHER PROFESSIONALS (TECHNICIAN, PROGRAMMER, ETC.)
3. ( ) GRADUATE STUDENTS
4. ( ) UNDERGRADUATE STUDENTS
5. ( ) SECRETARIAL - CLERICAL (IF CHARGED DIRECTLY)
6. ( ) OTHER
TOTAL SALARIES AND WAGES (A + B)
C. FRINGE BENEFITS (IF CHARGED AS DIRECT COSTS)
TOTAL SALARIES, WAGES AND FRINGE BENEFITS (A + B + C)
D. EQUIPMENT (LIST ITEM AND DOLLAR AMOUNT FOR EACH ITEM EXCEEDING $5,000.)
TOTAL EQUIPMENT
E. TRAVEL 1. DOMESTIC (INCL. CANADA, MEXICO AND U.S. POSSESSIONS)
2. FOREIGN
F. PARTICIPANT SUPPORT COSTS
1. STIPENDS $
2. TRAVEL
3. SUBSISTENCE
4. OTHER
TOTAL NUMBER OF PARTICIPANTS ( ) TOTAL PARTICIPANT COSTS
G. OTHER DIRECT COSTS
1. MATERIALS AND SUPPLIES
2. PUBLICATION COSTS/DOCUMENTATION/DISSEMINATION
3. CONSULTANT SERVICES
4. COMPUTER SERVICES
5. SUBAWARDS
6. OTHER
TOTAL OTHER DIRECT COSTS
H. TOTAL DIRECT COSTS (A THROUGH G)
I. INDIRECT COSTS (F&A)(SPECIFY RATE AND BASE)
TOTAL INDIRECT COSTS (F&A)
J. TOTAL DIRECT AND INDIRECT COSTS (H + I)
K. RESIDUAL FUNDS (IF FOR FURTHER SUPPORT OF CURRENT PROJECTS SEE GPG II.D.7.j.)
L. AMOUNT OF THIS REQUEST (J) OR (J MINUS K) $ $
M. COST SHARING PROPOSED LEVEL $ AGREED LEVEL IF DIFFERENT $
PI / PD TYPED NAME & SIGNATURE* DATE FOR NSF USE ONLYINDIRECT COST RATE VERIFICATION
ORG. REP. TYPED NAME & SIGNATURE* DATE
NSF Form 1030 (10/99) Supersedes all previous editions *SIGNATURES REQUIRED ONLY FOR REVISED BUDGET (GPG III.B)
1YEAR
1
DePaul University
Stanley
Stanley
Stanley
A
A
A
Cohn
Cohn
Cohn - Associate Professor 0.00 0.00 2.00 11,889
0 0.00 0.00 0.00 01 0.00 0.00 2.00 11,889
0 0.00 0.00 0.00 01 12.00 0.00 0.00 24,0000 02 6,0000 00 0
41,8898,329
50,218
25,000$Inverted Microscope1,500Shutter/Aperture equipment
26,5002,500
0
00000 0
7,000500
000
50 7,550 86,768
26,113% of SWB (Rate: 52.0000, Base: 50218)
112,8810
112,88125,500

SUMMARYPROPOSAL BUDGET
FundsRequested By
proposer
Fundsgranted by NSF
(if different)
Date Checked Date Of Rate Sheet Initials - ORG
NSF FundedPerson-mos.
FOR NSF USE ONLYORGANIZATION PROPOSAL NO. DURATION (months)
Proposed Granted
PRINCIPAL INVESTIGATOR / PROJECT DIRECTOR AWARD NO.
A. SENIOR PERSONNEL: PI/PD, Co-PI’s, Faculty and Other Senior Associates (List each separately with title, A.7. show number in brackets) CAL ACAD SUMR
$ $1.
2.
3.
4.
5.
6. ( ) OTHERS (LIST INDIVIDUALLY ON BUDGET JUSTIFICATION PAGE)
7. ( ) TOTAL SENIOR PERSONNEL (1 - 6)
B. OTHER PERSONNEL (SHOW NUMBERS IN BRACKETS)
1. ( ) POST DOCTORAL ASSOCIATES
2. ( ) OTHER PROFESSIONALS (TECHNICIAN, PROGRAMMER, ETC.)
3. ( ) GRADUATE STUDENTS
4. ( ) UNDERGRADUATE STUDENTS
5. ( ) SECRETARIAL - CLERICAL (IF CHARGED DIRECTLY)
6. ( ) OTHER
TOTAL SALARIES AND WAGES (A + B)
C. FRINGE BENEFITS (IF CHARGED AS DIRECT COSTS)
TOTAL SALARIES, WAGES AND FRINGE BENEFITS (A + B + C)
D. EQUIPMENT (LIST ITEM AND DOLLAR AMOUNT FOR EACH ITEM EXCEEDING $5,000.)
TOTAL EQUIPMENT
E. TRAVEL 1. DOMESTIC (INCL. CANADA, MEXICO AND U.S. POSSESSIONS)
2. FOREIGN
F. PARTICIPANT SUPPORT COSTS
1. STIPENDS $
2. TRAVEL
3. SUBSISTENCE
4. OTHER
TOTAL NUMBER OF PARTICIPANTS ( ) TOTAL PARTICIPANT COSTS
G. OTHER DIRECT COSTS
1. MATERIALS AND SUPPLIES
2. PUBLICATION COSTS/DOCUMENTATION/DISSEMINATION
3. CONSULTANT SERVICES
4. COMPUTER SERVICES
5. SUBAWARDS
6. OTHER
TOTAL OTHER DIRECT COSTS
H. TOTAL DIRECT COSTS (A THROUGH G)
I. INDIRECT COSTS (F&A)(SPECIFY RATE AND BASE)
TOTAL INDIRECT COSTS (F&A)
J. TOTAL DIRECT AND INDIRECT COSTS (H + I)
K. RESIDUAL FUNDS (IF FOR FURTHER SUPPORT OF CURRENT PROJECTS SEE GPG II.D.7.j.)
L. AMOUNT OF THIS REQUEST (J) OR (J MINUS K) $ $
M. COST SHARING PROPOSED LEVEL $ AGREED LEVEL IF DIFFERENT $
PI / PD TYPED NAME & SIGNATURE* DATE FOR NSF USE ONLYINDIRECT COST RATE VERIFICATION
ORG. REP. TYPED NAME & SIGNATURE* DATE
NSF Form 1030 (10/99) Supersedes all previous editions *SIGNATURES REQUIRED ONLY FOR REVISED BUDGET (GPG III.B)
2YEAR
2
DePaul University
Stanley
Stanley
Stanley
A
A
A
Cohn
Cohn
Cohn - none 0.00 0.00 2.00 12,483
0 0.00 0.00 0.00 01 0.00 0.00 2.00 12,483
0 0.00 0.00 0.00 01 12.00 0.00 0.00 25,2000 02 6,0000 00 0
43,6838,722
52,405
02,500
0
00000 0
4,000500
000
50 4,550 59,455
27,251% of swb (Rate: 52.0000, Base: 52405)
86,7060
86,7060

SUMMARYPROPOSAL BUDGET
FundsRequested By
proposer
Fundsgranted by NSF
(if different)
Date Checked Date Of Rate Sheet Initials - ORG
NSF FundedPerson-mos.
FOR NSF USE ONLYORGANIZATION PROPOSAL NO. DURATION (months)
Proposed Granted
PRINCIPAL INVESTIGATOR / PROJECT DIRECTOR AWARD NO.
A. SENIOR PERSONNEL: PI/PD, Co-PI’s, Faculty and Other Senior Associates (List each separately with title, A.7. show number in brackets) CAL ACAD SUMR
$ $1.
2.
3.
4.
5.
6. ( ) OTHERS (LIST INDIVIDUALLY ON BUDGET JUSTIFICATION PAGE)
7. ( ) TOTAL SENIOR PERSONNEL (1 - 6)
B. OTHER PERSONNEL (SHOW NUMBERS IN BRACKETS)
1. ( ) POST DOCTORAL ASSOCIATES
2. ( ) OTHER PROFESSIONALS (TECHNICIAN, PROGRAMMER, ETC.)
3. ( ) GRADUATE STUDENTS
4. ( ) UNDERGRADUATE STUDENTS
5. ( ) SECRETARIAL - CLERICAL (IF CHARGED DIRECTLY)
6. ( ) OTHER
TOTAL SALARIES AND WAGES (A + B)
C. FRINGE BENEFITS (IF CHARGED AS DIRECT COSTS)
TOTAL SALARIES, WAGES AND FRINGE BENEFITS (A + B + C)
D. EQUIPMENT (LIST ITEM AND DOLLAR AMOUNT FOR EACH ITEM EXCEEDING $5,000.)
TOTAL EQUIPMENT
E. TRAVEL 1. DOMESTIC (INCL. CANADA, MEXICO AND U.S. POSSESSIONS)
2. FOREIGN
F. PARTICIPANT SUPPORT COSTS
1. STIPENDS $
2. TRAVEL
3. SUBSISTENCE
4. OTHER
TOTAL NUMBER OF PARTICIPANTS ( ) TOTAL PARTICIPANT COSTS
G. OTHER DIRECT COSTS
1. MATERIALS AND SUPPLIES
2. PUBLICATION COSTS/DOCUMENTATION/DISSEMINATION
3. CONSULTANT SERVICES
4. COMPUTER SERVICES
5. SUBAWARDS
6. OTHER
TOTAL OTHER DIRECT COSTS
H. TOTAL DIRECT COSTS (A THROUGH G)
I. INDIRECT COSTS (F&A)(SPECIFY RATE AND BASE)
TOTAL INDIRECT COSTS (F&A)
J. TOTAL DIRECT AND INDIRECT COSTS (H + I)
K. RESIDUAL FUNDS (IF FOR FURTHER SUPPORT OF CURRENT PROJECTS SEE GPG II.D.7.j.)
L. AMOUNT OF THIS REQUEST (J) OR (J MINUS K) $ $
M. COST SHARING PROPOSED LEVEL $ AGREED LEVEL IF DIFFERENT $
PI / PD TYPED NAME & SIGNATURE* DATE FOR NSF USE ONLYINDIRECT COST RATE VERIFICATION
ORG. REP. TYPED NAME & SIGNATURE* DATE
NSF Form 1030 (10/99) Supersedes all previous editions *SIGNATURES REQUIRED ONLY FOR REVISED BUDGET (GPG III.B)
3YEAR
3
DePaul University
Stanley
Stanley
Stanley
A
A
A
Cohn
Cohn
Cohn - none 0.00 0.00 2.00 13,107
0 0.00 0.00 0.00 01 0.00 0.00 2.00 13,107
0 0.00 0.00 0.00 01 12.00 0.00 0.00 26,4600 02 6,0000 00 0
45,5679,135
54,702
02,500
0
00000 0
4,000500
000
50 4,550 61,752
28,445% of SWB (Rate: 52.0000, Base: 54702)
90,1970
90,1970

SUMMARY PROPOSAL BUDGET COMMENTS - Year 3
** E- TravelMeeting of the American Society for Cell Biology; PI and one studenttravelexpenses
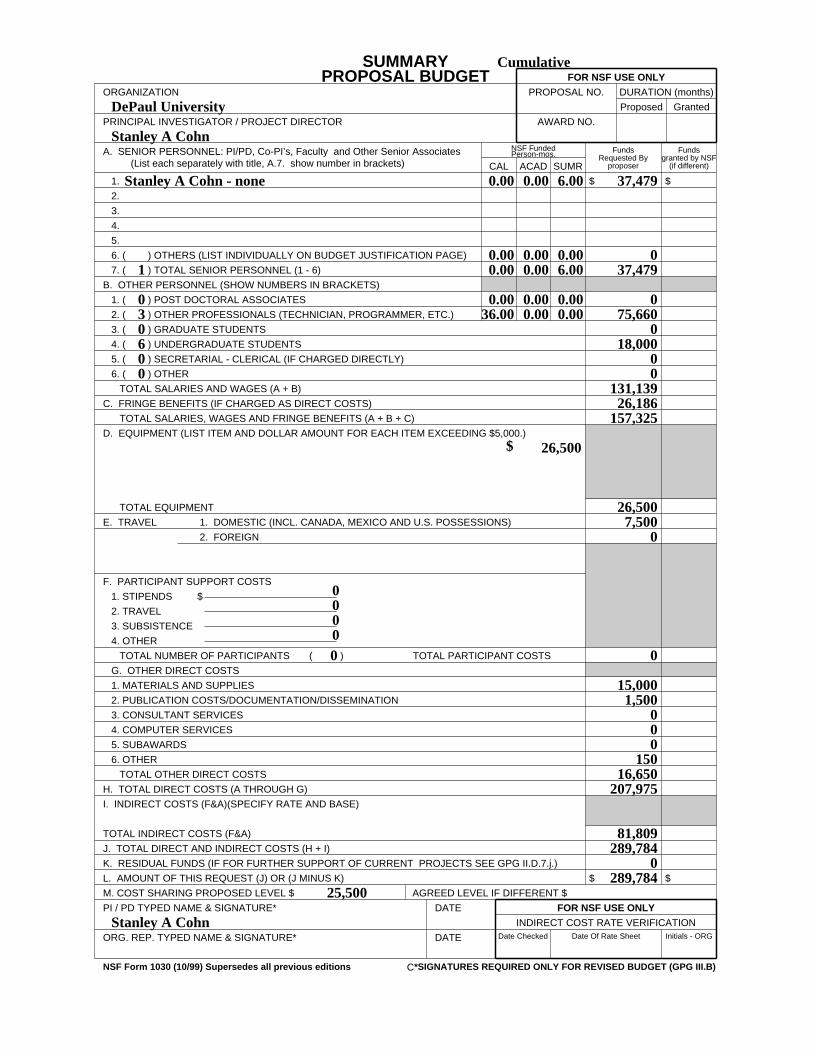
SUMMARYPROPOSAL BUDGET
FundsRequested By
proposer
Fundsgranted by NSF
(if different)
Date Checked Date Of Rate Sheet Initials - ORG
NSF FundedPerson-mos.
FOR NSF USE ONLYORGANIZATION PROPOSAL NO. DURATION (months)
Proposed Granted
PRINCIPAL INVESTIGATOR / PROJECT DIRECTOR AWARD NO.
A. SENIOR PERSONNEL: PI/PD, Co-PI’s, Faculty and Other Senior Associates (List each separately with title, A.7. show number in brackets) CAL ACAD SUMR
$ $1.
2.
3.
4.
5.
6. ( ) OTHERS (LIST INDIVIDUALLY ON BUDGET JUSTIFICATION PAGE)
7. ( ) TOTAL SENIOR PERSONNEL (1 - 6)
B. OTHER PERSONNEL (SHOW NUMBERS IN BRACKETS)
1. ( ) POST DOCTORAL ASSOCIATES
2. ( ) OTHER PROFESSIONALS (TECHNICIAN, PROGRAMMER, ETC.)
3. ( ) GRADUATE STUDENTS
4. ( ) UNDERGRADUATE STUDENTS
5. ( ) SECRETARIAL - CLERICAL (IF CHARGED DIRECTLY)
6. ( ) OTHER
TOTAL SALARIES AND WAGES (A + B)
C. FRINGE BENEFITS (IF CHARGED AS DIRECT COSTS)
TOTAL SALARIES, WAGES AND FRINGE BENEFITS (A + B + C)
D. EQUIPMENT (LIST ITEM AND DOLLAR AMOUNT FOR EACH ITEM EXCEEDING $5,000.)
TOTAL EQUIPMENT
E. TRAVEL 1. DOMESTIC (INCL. CANADA, MEXICO AND U.S. POSSESSIONS)
2. FOREIGN
F. PARTICIPANT SUPPORT COSTS
1. STIPENDS $
2. TRAVEL
3. SUBSISTENCE
4. OTHER
TOTAL NUMBER OF PARTICIPANTS ( ) TOTAL PARTICIPANT COSTS
G. OTHER DIRECT COSTS
1. MATERIALS AND SUPPLIES
2. PUBLICATION COSTS/DOCUMENTATION/DISSEMINATION
3. CONSULTANT SERVICES
4. COMPUTER SERVICES
5. SUBAWARDS
6. OTHER
TOTAL OTHER DIRECT COSTS
H. TOTAL DIRECT COSTS (A THROUGH G)
I. INDIRECT COSTS (F&A)(SPECIFY RATE AND BASE)
TOTAL INDIRECT COSTS (F&A)
J. TOTAL DIRECT AND INDIRECT COSTS (H + I)
K. RESIDUAL FUNDS (IF FOR FURTHER SUPPORT OF CURRENT PROJECTS SEE GPG II.D.7.j.)
L. AMOUNT OF THIS REQUEST (J) OR (J MINUS K) $ $
M. COST SHARING PROPOSED LEVEL $ AGREED LEVEL IF DIFFERENT $
PI / PD TYPED NAME & SIGNATURE* DATE FOR NSF USE ONLYINDIRECT COST RATE VERIFICATION
ORG. REP. TYPED NAME & SIGNATURE* DATE
NSF Form 1030 (10/99) Supersedes all previous editions *SIGNATURES REQUIRED ONLY FOR REVISED BUDGET (GPG III.B)
Cumulative
C
DePaul University
Stanley
Stanley
Stanley
A
A
A
Cohn
Cohn
Cohn - none 0.00 0.00 6.00 37,479
0.00 0.00 0.00 01 0.00 0.00 6.00 37,479
0 0.00 0.00 0.00 03 36.00 0.00 0.00 75,6600 06 18,0000 00 0
131,13926,186
157,325
26,500$
26,5007,500
0
00000 0
15,0001,500
000
150 16,650 207,975
81,809
289,7840
289,78425,500

Budget Justification
Personnel:The salaries requested are for PI summer salary, a full-time technician salary, and salary for two
undergraduate students during the school year [30 weeks at 5 hr/wk during the academic year; 11 weeks at 30 hr/wkduring summer/winter break, at $6.25/hr]. The benefits for the technician position are calculated using DePaul’snormal rate for full-time positions, 29%. This rate includes the following components: FICA 6.20%; Medicare1.45%; Retirement 8.00%; Unemployment 0.20%; Medical insurance 6.50%; Dental insurance 0.50%; Disabilityinsurance 0.35%; Life insurance 0.30%; Tuition 5.5%. Benefits for the PI summer support and the undergraduatestudent include only FICA and Medicare, totaling 7.65%, the normal rate for part-time and summer facultypositions. These positions, as described below, are crucial to the objectives of this proposal.
The presence of a full-time technician in the lab was essential to carrying out the planned work during thelast grant period. During the school year, students have severe time constraints, particularly in terms of the specifichours they have available to do research. At the same time, my teaching responsibilities also constrain the times Ican be present in the lab (e.g. I am teaching a lab course each quarter during the year, and one quarter each year I amresponsible for teaching 3 hr lectures and 4 three-hour laboratory sections each week). It therefore becomesimperative that a full-time technician, independent from the school calendar, be available to the undergraduatesworking in the lab. In this way, we can efficiently train students in the methods of medium preparation and diatomculturing, as well as particular experimental protocols. Moreover, a technician in the lab allows the constantpresence of someone who can help answer technical questions of the students in the lab as they run up againstproblems during an experiment.
In addition, the technicians I hired with the last grant were recent college graduates trying to determinewhether or not to proceed with a career in science. Exposing them to the excitement that is generated within anactive research lab provided, I believe, exactly the extra impetus needed to spur them on to continue in scientificendeavors. The two technicians I hired have both gone on to graduate school - one to a M.S. program, and the otherinto a graduate Pharmacy (Pharm.D.) program.
Similarly, the requested money for undergraduate students is necessary to achieve the goal of increasing thescientific interests and capabilities of the students. I have been a vigorous advocate for including undergraduates inresearch, and have had over 20 undergraduates working in my lab over the past 6 years (see RUI Statement). Mostundergraduate students at DePaul must work to help pay for their tuition, so it is necessary to financially supportthem during the summer if they are to be able to participate in the lab research without suffering the loss of summersalary. A large portion of DePaul students are minority students (about 32% in the 1998 freshman class), and manyof our students are first generation college students receiving financial aid to attend the University. The funding ofthe undergraduate stipend would allow the students to earn summer and school year income to help pay for tuitionwhile still learning science by directly participating in laboratory investigations. The requested funds would allowme to hire two students during each year (30 weeks @ 5 hrs/wk during the academic year and 11 weeks @ 30hrs/wk during the winter/summer academic breaks)
Equipment RequiredIn order to carry out the proposed experiments, a number of pieces of equipment will need to be purchased:
– Additional Stereo-Microscope Head. We currently have both a brightfield/darkfield stand for the stereo-microscope (used in photo-accumulation and standard coverslip inversion tests) and a boom stand for thestereo-microscope head (used for flume and temperature controlled adhesion studies). This requires that thecurrent microscope head be physically moved and aligned each time we need to run a different experiment.More importantly, we cannot run both flume and photo-stimulation experiments simultaneously. Since theseare both important assays that we need to run simultaneously in the lab, particularly during the summer, weneed an additional stereo-microscope head and video adapter.
– Additional Computer. We currently have only one computer in our lab capable of running thespeed/distance/path analysis program. The current computer is connected to the Zeiss Axioskop in a separatemicroscope room. In order to carry out our analysis of motile characteristics for cells being viewed under othermicroscopes (e.g. moving on opaque surfaces, with observation under the dissecting microscope) we needanother computer which can be quickly connected to any of the other microscope set-ups in the lab (i.e. the

head on the boom stand, the head on the darkfield stand, etc.). We plan to purchase a laptop computer withvideo input/output capabilities.
– Reflection Interference Contrast Optics and Inverted Microscope. In order to carry out our experiments on theanalysis of cell contact sites, we will need to have Reflection Interference Contrast optics and an invertedmicroscope. The microscope objective for this type of optics has already been purchased by the BiologyDepartment, but several more lenses and filters are still required. In addition, reflectance interference requiresepi-illumination. Therefore, while the Zeiss Axioskop in the lab (already in use) will be sufficient for tests ofmoving cells on inverted coverslips (i.e. analysis of contact sites on cells in the process of losing attachment),an inverted microscope is required to investigate the standard movement of cells gliding over the top ofsurfaces. By having two Zeiss microscopes, the objectives and filters can be used interchangeably between thetwo microscopes, minimizing the number of optics needed to be purchased.
– Equipment for environmental control of flume set-up. The flume set-up as currently designed operates only atroom temperature and room light conditions. In order to determine the environmental conditions affecting celladhesion as measured by resistance to water current, we need additional light and temperature controlequipment. This will include both an overhead lighting system (an overhead variable fiber optic system forsmall spot lighting and quartz bulbs for longer term lighting), and a temperature regulated water pump.
– Equipment for Photophobic response experiments. Currently, the Zeiss Axioskop in the lab is sufficient forbasic epi-illumination. In order to adapt the microscope for performing more controlled tests on light directedresponses, a controlled shutter and adjustable centerable diaphragm with a small spot size is required.
The costs of these items are as follows (prices reflect estimates from the companies in parentheses):
Shutter/Aperture Equipment for Axioskop for controlled irradiations (Uniblitz) $ 1,500Computer with Video Capabilities $ 3,500Reflection Interference Contrast Equipment (Zeiss) $ 5,000Light/Pump System for Flume (Lauda) $ 12,000Stereo-microscope Head w/ video port and lenses (Leica-Wild)$ 5,000Inverted Microscope for reflection interference contrast (Zeiss)$ 25,000
Total Equipment Costs $ 52,000
Because these equipment costs are extensive, DePaul University has agreed to help share the costs associated withthe needed equipment of this project by purchasing the upgrade equipment for the compound and stereomicroscopes, the Refrigerated Circulating Pump, the Computer, and the Controlled Light Set-up (middle four itemsabove). These items have the greatest potential to be shared with other members of the DePaul community in theirresearch endeavors. Therefore, the breakdown of equipment costs is as follows:
DePaul Equipment Costs $ 25,500Equipment Costs Requested from NSF $ 26,500
Travel:The proposal requests the travel funds for the PI and one student to one scientific meeting per year
($1250/meeting/year/person). This will allow the timely dissemination of the project results in an appropriateforum, as well as detailed discussion of the project with numerous colleagues. In addition, the undergraduatestudents are able to help present their own data, as well as get a much better look of the scientific enterprise at work.Over the past several years (except for the last year, when no funds were available), I have taken at least one studentwith me to the annual Cell Biology meetings. It has always been a mutually exciting and rewarding experience, andallowed me to directly involve the students in the process of scientific discussion.

Other Direct Costs:The supply money as requested is designed for the normal operating costs of the laboratory. The amount
requested for years two and three (4,000/year) is not excessive given the large amount of consumables, glassware,plasticware, and biochemicals that are continually used in making large amounts of diatom medium and culturingthe cells. Based on our spending over the past three years, the annual costs are expected to be broken downapproximately as follows:
Biochemicals and Reagents $ 1,250Glassware/Plasticware $ 1,250Microscopy Supplies $ 1,000Culture Supplies $ 500
The additional money requested for supplies in the first year ($7000 total in supplies) includes additionalcomputer and video material needed to connect the second stereo-microscope head to our measurement andrecording systems. The budget also requests money ($500) for funds to cover the costs of printing/publishing onejournal article per year, along with $50/year for photocopying costs.

Current and Pending Support(See GPG Section II.D.8 for guidance on information to include on this form.)
The following information should be provided for each investigator and other senior personnel. Failure to provide this information may delay consideration of this proposal.
Investigator:Other agencies (including NSF) to which this proposal has been/will be submitted.
Support: Current Pending Submission Planned in Near Future *Transfer of Support
Project/Proposal Title:
Source of Support:Total Award Amount: $ Total Award Period Covered:Location of Project:Person-Months Per Year Committed to the Project. Cal: Acad: Sumr:
Support: Current Pending Submission Planned in Near Future *Transfer of Support
Project/Proposal Title:
Source of Support:Total Award Amount: $ Total Award Period Covered:Location of Project:Person-Months Per Year Committed to the Project. Cal: Acad: Sumr:
Support: Current Pending Submission Planned in Near Future *Transfer of Support
Project/Proposal Title:
Source of Support:Total Award Amount: $ Total Award Period Covered:Location of Project:Person-Months Per Year Committed to the Project. Cal: Acad: Sumr:
Support: Current Pending Submission Planned in Near Future *Transfer of Support
Project/Proposal Title:
Source of Support:Total Award Amount: $ Total Award Period Covered:Location of Project:Person-Months Per Year Committed to the Project. Cal: Acad: Sumr:
Support: Current Pending Submission Planned in Near Future *Transfer of Support
Project/Proposal Title:
Source of Support:Total Award Amount: $ Total Award Period Covered:Location of Project:Person-Months Per Year Committed to the Project. Cal: Acad: Summ:
*If this project has previously been funded by another agency, please list and furnish information for immediately preceding funding period.
NSF Form 1239 (10/99) USE ADDITIONAL SHEETS AS NECESSARYPage G-
Stanley Cohn
Ecological Conditions Affecting Diatom Motility andAdhesion
DePaul University - Liberal Arts & Science Summer Grant5,200 06/15/99 - 09/15/99
DePaul University1.00
11

H. Facilities and Equipment
The Department of Biological Sciences of DePaul University is currently housed at theLincoln Park Campus of DePaul University and has recently moved from the O'Connell ScienceBuilding to the new McGowan Biological and Environmental Sciences building. The laboratoryof Dr. Cohn occupies approximately 750 square feet in the building and already contains severalpieces of equipment to be used for the proposed project. This equipment includes a ZeissAxioskop video microscope fitted with DIC, phase and fluorescence optics and connected via aGenLock device to an Amiga 2000 computer. Additional lab equipment includes a Wild MZ8Stereomicroscope fitted with a video port, a Sony Camera and Video Printer, a Panasonic time-lapse video recorder, an S-Video recorder, a G3 400 MHz computer with video input/output anda DAGE DSP-2000 video processor. Additional equipment includes a refrigerator, freezer,incubators and facilities for cell culturing, pH meter, balances, an electrically controlled thermalcooling plate, a temperature controlled slide holder, and acrylic flume apparatus.
The new McGowan building, opened in July 1998, includes controlled environmentrooms, darkroom, cold room, tissue culture room, PCR room, animal care facilities, dishwashingfacilities, and a greenhouse, all available as shared facilities. There is also high-quality deionizedwater available in each lab. Shared departmental equipment includes autoclaves, anultracentrifuge, scintillation counter, refrigerated low-speed centrifuge, clinical grade phasemicroscopes, an osmometer, respiratory gas analyzer, scanning spectrophotometer, a digital videoprinter, slide printer, and a digital gel scanner/analysis system. There is also a field site van, jointlyused by Biology and Environmental Science. This equipment is all available for joint use by theBiology faculty. Secretarial assistance for typing, as well as funds for copying, postage andtelephones, is also provided.
The University is also committed to continually monitoring and upgrading the sciencecapabilities of the campus. In 1993, DePaul opened a new library building, which is continuing toincrease its science reference facilities and databases, and is set-up for relatively rapid intra-cityacquisition and borrowing from nearby institutions. New Chemistry facilities are planned to bebuilt adjacent to the McGowan building in 3-5 years, giving a significant upgrade to the ChemistryDepartment and its academic program. Moreover, DePaul is moving forward with the planning ofa Food Science Program, which is expected to help foster further growth in the sciences atDePaul.
The location of DePaul near the center of Chicago provides for excellent intellectual andcollaborative opportunities with scientists from the University of Illinois at Chicago, University ofChicago, Northwestern University, Loyola University as well as many other scientific and medicalfacilities within the city. As part of this intellectual exchange, the Biology department maintains aseminar series during the school year that brings in speakers from throughout the Chicago andIllinois region.