a chimeric vesiculo/alphavirus is an effective...
TRANSCRIPT

1
A Chimeric Vesiculo/Alphavirus is an effective Alphavirus Vaccine 1
2
3
Anasuya Chattopadhyay1, Eryu Wang2, Robert Seymour2, Scott C. Weaver2, and John K. Rose1, # 4
5
Author Affiliation: 1Department of Pathology, Yale University School of Medicine, New 6
Haven, CT 06510; 2Institute for Human Infections and Immunity, Sealy Center for Vaccine 7
Development and Department of Pathology, University of Texas Medical Branch, Galveston, 8
TX. 9
10
# Corresponding Author: John K. Rose, Department of Pathology, Yale University School of 11
Medicine, 310 Cedar Street (LH315), New Haven, CT 06510, U.S.A. Phone: 203-785-6794. Fax: 12
203-785-767. E-mail address: [email protected] 13
14
Running Title: VSV∆G Chikungunya Vaccine 15
16
Word count for Abstract: 164 17
18
Word count for text: 4368 (excluding References, figure legends) 19
Copyright © 2012, American Society for Microbiology. All Rights Reserved.J. Virol. doi:10.1128/JVI.01860-12 JVI Accepts, published online ahead of print on 17 October 2012
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

2
ABSTRACT 20
While a large number of mosquito-transmitted alphaviruses are known to cause serious human 21
diseases, there are no licensed vaccines that protect against alphavirus infections. The alphavirus 22
chikungunya virus (CHIKV) has caused multiple recent outbreaks of chikungunya fever. This 23
virus has the potential to cause a worldwide epidemic and has generated strong interest in 24
development of a prophylactic CHIKV vaccine. We report here on the development of a potent 25
experimental vaccine for CHIKV based on a chimeric vesicular stomatitis virus (VSV) 26
expressing the entire CHIKV envelope polyprotein (E3-E2-6K-E1) in place of the VSV 27
glycoprotein (G). These VSVΔG-CHIKV chimeras incorporated functional CHIKV 28
glycoproteins into the viral envelope in place of VSV G. The chimeric viruses were attenuated 29
for growth in tissue culture, but could be propagated to high titers without VSV G 30
complementation. They also generated robust neutralizing antibody and cellular immune 31
responses to CHIKV in mice after a single dose, and protected mice against CHIKV infection. 32
VSV∆G-alphavirus chimeras could have general applicability as alphavirus vaccines. 33
34
35
36
37
38
39
40
41
42
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

3
INTRODUCTION 43
Alphaviruses are found worldwide and many are associated with serious disease in 44
humans and other vertebrates. They are typically transmitted by mosquitoes to vertebrate hosts, 45
and can cause fever, arthritis, and lethal encephalitis (9). There are currently no licensed 46
vaccines that protect against alphavirus infection and disease. Chikungunya virus (CHIKV) is an 47
alphavirus that causes chikungunya fever. The virus was first isolated in 1953 in Tanzania and 48
spread across Africa and Southeast Asia. More recent outbreaks have spread to Europe, and 49
CHIKV has been diagnosed in the United States in travelers returning from endemic areas (25, 50
39). CHIKV has generated global public health concern in part because its spread has been 51
associated with a new mosquito vector, Aedes albopictus, which is widely distributed in Europe 52
and the Americas (25). 53
CHIKV infection in humans results in high fever, headache, vomiting, skin rash and 54
crippling arthritis. While most symptoms resolve by about 10 days, the crippling arthritis can 55
persist for months or even years (25). Recent CHIKV epidemics have shown even more severe 56
symptoms including encephalitis, hemorrhagic disease and mortality (34). 57
CHIKV has a single, positive sense, 11.8 kb RNA genome, which encodes four structural 58
proteins (nsP1-4) and five structural proteins (C, E3, E2, 6K, E1) that are cleaved from a 59
precursor to generate the capsid and envelope glycoproteins (35). Neutralizing antibodies 60
(nAbs) play dominant roles in resolving alphavirus infections and neutralizing epitopes have 61
been mapped within E1 (fusion protein) and E2 (attachment protein) glycoproteins in several 62
alphaviruses (38). However, cellular immunity is also capable of protecting against fatal 63
alphaviral disease (22). 64
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

4
Recent CHIKV epidemics in the La Reunion Island and the Indian subcontinent, 65
southeast Asia and Europe have generated substantial interest in development of CHIKV 66
vaccines. A number of successful vaccine approaches based on expression of the E1 and E2 67
CHIKV glycoproteins have been described recently. These include chimeric alphavirus vectors 68
(42, 43), an adenovirus vector (41), VLP vaccine (1), DNA vaccine (18) and IRES based live-69
attenuated CHIKV vaccine (24). 70
Vesicular stomatitis virus (VSV), a negative-strand RNA virus in the Rhabdoviridae 71
family (Vesiculovirus genus), has been used extensively as an experimental vaccine vector 72
against several viral and bacterial pathogens (4, 12-15, 26, 27, 29, 33). VSV-based vaccine 73
vectors are currently being used in HIV vaccine clinical trials 74
(http://clinicaltrials.gov/ct2/show/NCT01438606). 75
In this study, we initially wanted to determine if we could construct VSV-based vaccine 76
vectors expressing the entire CHIKV E3-E2-6K-E1 precursor polyprotein. We did this in a full-77
length VSV construct as well as in a VSVΔG construct (lacking the VSV glycoprotein gene). 78
Interestingly, we found that the VSVΔG vector expressing CHIKV envelope proteins 79
incorporated the CHIKV glycoproteins efficiently into virus particles and propagated without 80
VSV G complementation. This chimeric VSV/alphavirus vector also induced more potent 81
CHIKV immune responses than the full-length vector with VSV G and protected mice from 82
CHIKV challenge after a single dose. Such chimeric viruses could be generally applicable as 83
alphavirus vaccines. 84
85
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

5
MATERIALS AND METHODS 86
Cells. Baby Hamster Kidney (BHK)-21 cells were maintained in Dulbecco’s modified 87
Eagle’s medium (DMEM) supplemented with 5% Fetal Bovine Serum (FBS). Vero cells, 88
derived from African green monkey kidney cells, were maintained in DMEM containing 10% 89
FBS. 90
Plasmid constructions. To generate pVSV-CHIKV, we first designed a codon 91
optimized synthetic E3-E2-6K-E1 gene (CHIKV S27 prototypic African strain) and had it 92
synthesized with flanking Xho I and Nhe I restriction sites (Genscript, Inc.). This gene was 93
inserted into Xho I-Nhe I digested pVSVXN2 vector (31). pVSVΔG-CHIKV was prepared by 94
deleting the VSV G gene from the pVSV-CHIKV by Mlu I-Xho I digestion, filling-in with T4 95
DNA polymerase (New England Biolabs) and re-ligation. pCAGGS-CHIKV was made by 96
inserting the Xho I-Nhe I digested synthetic E3-E2-6K-E1 fragment into corresponding sites of a 97
modified pCAGGS vector (20) containing these sites. 98
Recombinant virus recovery. Recombinant VSVs (rVSVs) were recovered from 99
pVSV-CHIKV and pVSVΔG-CHIKV as described previously (16, 32). In brief, BHK-21 cells 100
were infected at a multiplicity of infection (MOI) of 10 with vTF-7.3 (7), a vaccinia virus 101
recombinant expressing T7 RNA polymerase. The cells were then transfected with rVSV 102
plasmids (pVSV-) together with support plasmids, pBS-N, pBS-P, pBS-L and pBS-G, encoding 103
VSV proteins. VSV-CHIKV was recovered by transferring the transfected cell supernatants onto 104
fresh BHK-21 cells at 48 h post transfection, and collecting supernatant containing the virus after 105
another 48 h. Virus stock was prepared from individual plaques grown in BHK-21 cells and 106
stored at -70°C. To recover VSV G complemented VSVΔG-CHIKV, transfection supernatant 107
was transferred to cells that were transfected with pCAGGS-G (21) one-day prior, and 108
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

6
supernatant containing the virus was collected after 48 h. The virus was further plaque purified 109
on BHK-G cells (32), and VSV G-complemented stock was prepared and stored at -70°C. A 110
part of VSVΔG-CHIKV recovery supernatant was also plaque-purified without VSV G 111
complementation on BHK-21 cells and further passaged on BHK-21 cells to generate a non-G 112
complemented VSVΔG-CHIKV stock. 113
pCAGGS transfection. 10 μg of the appropriate pCAGGS vector diluted in 0.6 ml of 114
OptiMEM (Invitrogen, CA) was mixed with 30 μl of Lipofectamine Reagent (Invitrogen, CA) 115
also diluted in 0.6 ml of OptiMEM, and incubated at room temperature for 30 min. Confluent 116
monolayers of BHK-21 cells in a 10 cm dishes were washed once with phosphate buffered saline 117
(PBS) and once with OptiMEM. OptiMEM (4.8 ml) was then added followed by addition of the 118
DNA-Lipofectamine mix and incubation for 5 h at 37°C. Subsequently, the transfection mix was 119
removed from the plate and replaced with DMEM containing 10% FBS and incubated overnight 120
at 37°C. 121
Indirect immunofluorescence microscopy. BHK-21 cells on coverslips were infected 122
with recombinant wild-type (rwt)-VSV, VSV-CHIKV or VSVΔG-CHIKV. At 4 h post 123
infection, cells were washed two times with PBS and fixed with 3% paraformaldehyde at room 124
temperature for 20 min. Cells were then washed with PBS containing 10 mM glycine (PBS-125
glycine) and incubated with a 1:200 dilution of either VSV G monoclonal antibodies (MAbs; I1 126
and I14) (17) or an anti-CHIKV antibody [4-week sera from CHIKV-La Reunion (LR) strain 127
infected mouse]. Following this, cells were washed with PBS-Glycine and incubated with 1:500 128
diluted goat anti-mouse AlexaFluor 488 IgG (Molecular probes, Eugene, OR). Cells were then 129
washed twice with PBS-Glycine, mounted on slides using ProLong Gold antifade reagent with 130
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

7
DAPI (Molecular probes, Eugene, OR) and imaged with a Nikon Eclipse 80i fluorescence 131
microscope using a 40X objective. 132
Metabolic labeling and SDS-PAGE of purified viruses and cell lysates. To label 133
infected cells, BHK-21 cells in 3 cm dishes were infected with recombinant viruses or mock 134
infected at an MOI of 20. At 5 h post-infection, cells were washed with methionine-free DMEM 135
followed by labeling with 100 μCi of [35S]-methionine in 1 ml methionine-free DMEM for 30 136
min at 37°C. Cells were then washed twice with PBS and lysed with detergent lysis buffer (1% 137
Nonidet P-40, 0.4% deoxycholate, 50 mM Tris-HCl, pH 8.0, 62.5 mM EDTA) on ice for 5 min. 138
Lysates were collected and clarified at 16,000 x g for 2 min at 4°C. To label rVSVs, BHK-21 139
cells on 6 cm dishes were infected with rwt-VSV, VSV-CHIKV or VSVΔG-CHIKV at an MOI 140
of 100. After adsorption for 1 h, virus was removed and replaced with DMEM containing 5% 141
FBS. After 2 h, cells were washed two-times with pre-warmed methionine-free DMEM. 142
Subsequently, labeling media (90% methionine-free DMEM + 9% serum-free DMEM + 1% 143
FBS) containing 300 μCi of [35S]-methionine was added to each dish and incubated over night at 144
37°C. Virus supernatants were collected after 20 h, clarified at 16,000 x g for 5 min to remove 145
cell debris, layered onto a 20% sucrose gradient (in 10 mM Tris pH 7.4) and centrifuged for 1 h 146
at 38,000 rpm at 4°C. Virus pellet was dissolved in TE. 147
Labeled viruses and cell lysates were analyzed on a 4-12% Bis-Tris NuPAGE gel 148
(Invitrogen, CA) and protein bands were imaged using a Fujifilm BAS 1800 imaging system. 149
One-step growth curve. BHK-21 cells were infected with rwt-VSV, VSV-CHIKV or 150
VSVΔG-CHIKV at a MOI of 10. After virus adsorption for 30 min, cells were washed twice 151
with PBS and DMEM containing 5% FBS was added. Supernatant samples were collected at 0, 152
2, 4, 6, 8 and 24 h post adsorption and titered by plaque assay on BHK-21 cells. 153
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

8
CHIKV pseudotype generation. VSVΔG-eGFP1 pseudotyped with CHIKV envelope 154
proteins was generated by transfecting BHK cells with pCAGGS-CHIKV, followed by infection 155
with VSV G complemented VSVΔG-eGFP1 (15). At 1 h post infection input virus was 156
removed, cells were washed twice with PBS and DMEM containing 5% FBS was added. 157
Supernatant containing VSVΔG-eGFP1 pseudotyped with CHIKV envelope proteins was 158
collected at 24 h and virus titered on BHK-21 cells by counting the number of enhanced green 159
fluorescent protein (eGFP) expressing cells. 160
Animal experiments. Mice, housed under BSL-2 conditions in microisolator cages, 161
were inoculated with recombinant VSV vectors using 106 plaque forming units (PFU) per mouse 162
by the intramuscular (I.M.) route in a volume of 50 μl of serum-free DMEM administered into 163
the right hind leg muscle. For the challenge study, 3 week old female C57Bl/6 mice were 164
obtained from Jackson Laboratories, ME and acclimatized for 4 days before immunization. 165
Blood was collected at 30 days post immunization (dpi) from retro-orbital sinus. These animals 166
were challenged with the wild-type CHIKV-LR strain derived from a cDNA clone (39) using 104 167
PFU per mouse by the subcutaneous (S.C.) route in the left rear footpad. Foot swelling 168
measurements were taken on the day of challenge and for 10 days following using a caliper to 169
measure the height of the foot at the ball. Animals from each immunization group were divided 170
into two subgroups and were bled on alternate days post challenge to measure viral titers by 171
plaque assay on Vero cells. Two animals from each group were sacrificed at 4 days post 172
challenge (38 dpi) for histopathology analysis. UV inactivation of virus was performed using 173
two exposures of virus to 5 x 105 μJoules in a UV Stratalinker 1800 (Stratagene). 174
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

9
To look at cellular immune responses, 6-8-week-old female C57Bl/6 mice were 175
inoculated with virus after 1-week acclimatization, and sacrificed by anesthesia overdose at 7 dpi 176
for isolating splenocytes. 177
Yale University and/or UTMB Institutional Animal Care and Use Committee approved 178
all animal experiments. 179
Virus neutralization assays. The VSVΔG-eGFP1/CHIKV pseudotype neutralization 180
assay was similar to that described previously for Nipah virus pseudotypes (5). Briefly, 181
approximately 50 infectious pseudotype particles were mixed with pooled and serially diluted 182
serum samples from each immunization group, added to monolayer of BHK-21 cells in a 96-well 183
plate and incubated for 20-24 h at 37°C. Infection was determined by visualizing the number of 184
eGFP expressing cells using an Olympus CK-40 microscope equipped with epifluorescence. 185
Absence of infection in duplicate wells for each sample was scored as 100% neutralization. 186
VSV neutralization assay was performed as described previously (29). The VSV 187
neutralization titers were defined as the highest dilution of serum that could completely 188
neutralize infectivity of 100 PFU of VSV on BHK-21 cells 189
CHIKV plaque reduction neutralization tests were performed using standard methods (2) 190
and the CHIKV-LR strain. 191
ELISPOT assay. A gamma interferon (IFN-γ) enzyme linked immunospot assay 192
(ELISPOT) kit (BD Biosciences) was used to quantify T cell activation following immunization. 193
Splenocytes were isolated from immunized mice at 7 dpi by disrupting spleen between frosted 194
ends of two microscope slides. The red blood cells were removed by using red blood cell lysis 195
buffer (eBioscience Inc., San Diego, CA) and spenocytes were collected in RPMI 1640 196
containing 10% FBS after passing through strainer. 2 x 105 splenocytes per well were then 197
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

10
added to a 96-well plate pre-coated with mouse IFN-γantibody and incubated with or without 198
CHIKV E1 (HSMTNAVTI) or E2 (IILYYYELY) peptides at a concentration of 20 μg/ml for 24 199
h at 37°C. Thereafter, cells were first washed with water and then with wash buffer (kit) 200
followed by incubation with biotinylated anti-mouse IFN-γ antibody at 1:2000 dilution for 2 h at 201
room temperature. Following this, cells were again washed with wash buffer; streptavidin-202
horseradish peroxidase (HRP) conjugate (kit) was added at 1:100 dilution and incubated at room 203
temperature for 1 h. The plate was developed according to manufacturer’s protocol using 3-204
amino-9-ethyl-carbazole (AEC) chromogen diluted in AEC substrate buffer. Reaction was 205
stopped by adding water and the plate was air-dried before counting spot forming cells (sfc). 206
207
RESULTS 208
Construction and characterization of recombinant VSVs expressing Chikungunya 209
virus envelope proteins. To generate a VSV recombinant expressing CHIKV surface 210
glycoproteins, a synthetic gene encoding the CHIKV E3-E2-6K-E1 polyprotein was inserted 211
between the G and L genes of VSV (Indiana) in the plasmid vector pVSVXN2 (31) and the 212
resulting plasmid pVSV-CHIKV was used to recover VSV-CHIKV (Fig. 1A) using established 213
procedures (16). A VSV recombinant expressing CHIKV envelope proteins in the absence of 214
VSV G (Fig. 1A) was also recovered by complementation with VSV G (32) and was designated 215
VSVΔG-CHIKV. Initially this recombinant was grown in cells expressing VSV G because we 216
assumed that CHIKV glycoproteins would not be incorporated efficiently into VSV particles or 217
promote efficient infection. As we show below, the VSVΔG-CHIKV recombinant does 218
propagate in the absence of VSV G complementation. 219
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

11
Expression of CHIKV proteins in VSV recombinants. To determine if CHIKV 220
proteins were expressed in cells infected with the VSV/CHIKV recombinants, we initially used 221
indirect immunofluorescence microscopy. BHK-21 cells were infected with VSV-CHIKV, 222
VSVΔG-CHIKV or rwt-VSV (control). Fixed, non-permeablized cells were incubated with 223
MAbs to VSV G or with serum from CHIKV-infected mice, followed by 2° Ab (anti-mouse 224
AlexaFlour-488 conjugate) treatment. As shown in Figure 1B, cells infected with both 225
recombinants showed surface expression of CHIKV proteins, while rwt-VSV infected cells 226
showed no CHIKV protein expression. The CHIKV antiserum recognizes both E1 and E2. VSV 227
G protein was expressed on the cell surface of rwt-VSV and VSV-CHIKV infected cells, while 228
VSVΔG-CHIKV infected cells did not express VSV G. 229
To visualize the expression levels of both E1 and E2 proteins, we performed a 30 min 230
metabolic labeling ([35S]-methionine) of BHK-21 cells infected with VSV-CHIKV, VSVΔG-231
CHIKV, or rwt-VSV. Lysates from infected and mock-infected cells were separated by SDS-232
PAGE and analyzed on a Fujifilm imaging system (Fig. 1C). Because VSV infection shuts off 233
host protein synthesis, the cell lysate from rwt-VSV infected cells shows mainly the viral 234
proteins L, G, N, P and M (Fig 1C, lane 2). VSV-CHIKV infected cells expressed an additional 235
protein with the mobility expected for CHIKV E1 protein (~ 52 kDa) (Fig. 1C, lane 3). Because 236
this was only a 30 minute labeling, the CHIKV E2 would still be in its precursor form of E3-E2 237
(p62). The p62 protein is evident in the VSVΔG-CHIKV infected cells along with E1 (Fig. 1C, 238
lane 4). However, in the VSV-CHIKV infected cells, the p62 protein runs just ahead of and 239
overlapping VSV G, and cannot be discerned clearly. 240
CHIKV glycoproteins are incorporated into VSV recombinants. To determine if the 241
CHIKV glycoproteins were incorporated into VSV particles, BHK-21 cells were infected with 242
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

12
rwt-VSV or either VSV/CHIKV recombinants and labeled for sixteen hours with [35S]-243
methionine. Virus particles purified from the medium were then analyzed by SDS-PAGE (Fig. 244
1D). rwt-VSV and VSV-CHIKV particles contained the five indicated VSV proteins, while 245
VSVΔG-CHIKV particles lacked VSV G. Both VSV-CHIKV and VSVΔG-CHIKV particles 246
contained a single extra protein band with the mobility of a 52 kDa protein, the mobility 247
expected for both E1 and mature E2. The precursor protein p62 present in cells was absent from 248
the VSVΔG-CHIKV virions as expected, because it is cleaved prior to transport to the cell 249
surface. 250
VSVΔG-CHIKV recombinants grow in the absence of VSV G complementation. 251
Although VSV is promiscuous in terms of accepting foreign proteins into its envelope (30, 46), 252
VSVΔG recombinants with foreign viral envelopes often grow to titers 100-fold lower than VSV 253
itself in the absence of complementation with VSV G (3). However, we found that the VSVΔG-254
CHIKV recombinant typically grew to titers only 2-3 fold lower than rwt-VSV and did not 255
require VSV G complementation. To examine the growth of VSVΔG-CHIKV and VSV-CHIKV 256
in more detail we performed a one-step growth curve as shown in Figure 2A. VSV and VSV-257
CHIKV grew at similar rates and reached equivalent titers, while the VSVΔG-CHIKV appeared 258
to lag initially but eventually reached similar titer as VSV by 24 h. VSVΔG-CHIKV also 259
generated smaller plaques in BHK-21 cells compared to VSV-CHIKV (Fig. 2B). The plaques 260
generated by VSV-CHIKV are indistinguishable from VSV plaques (not shown). 261
VSV/CHIKV recombinants induce CHIKV nAbs in C57Bl/6 mice. To determine if 262
the VSV recombinants expressing CHIKV glycoproteins were immunogenic, we initially 263
performed a pilot study to test the immunogenicity of VSV-CHIKV, VSVΔG-CHIKV (G-264
complemented) or VSVΔG-CHIKV (non-complemented) in C57Bl/6 mice. A single I.M. 265
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

13
immunization (106 PFU) with these VSV/CHIKV recombinants induced antibodies that 266
neutralized a VSVΔG-eGFP1/CHIKV pseudotype (not shown). Moreover, the non-267
complemented VSVΔG-CHIKV elicited a CHIKV nAb titer that was at least as good as that of 268
the G-complemented virus or VSV-CHIKV. 269
The immune response requires viral RNA synthesis. To determine if the nAb 270
response to VSVΔG-CHIKV was dependent on viral replication, we examined immune 271
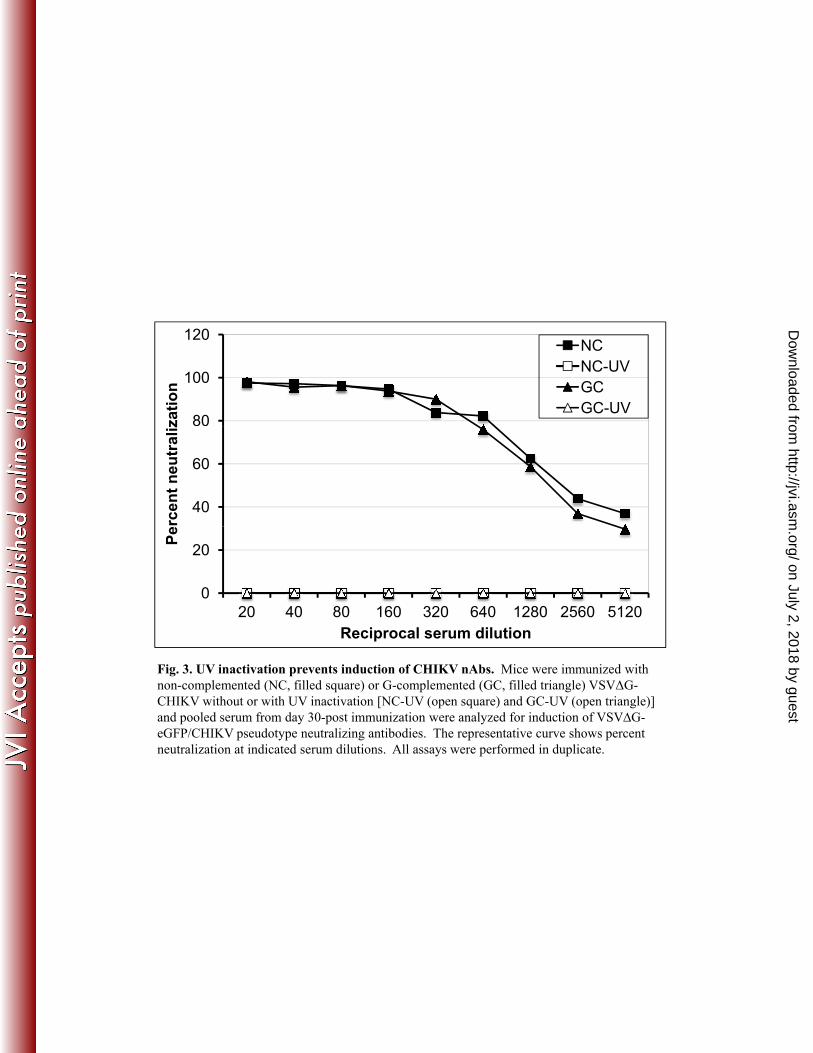
responses to UV inactivated virus particles. As shown in Fig. 3, UV inactivation completely 272
eliminated the antibody response to G-complemented or non-complemented VSVΔG-CHIKV. 273
This result indicates that proteins on incoming virions are not sufficient to generate the immune 274
response and that viral transcription and presumably replication are required to generate 275
sufficient mRNA and protein to induce the immune response. Because the non-complemented 276
VSVΔG-CHIKV would have the major advantage of not inducing anti-G antibodies, we 277
performed a more detailed analysis of this virus as well as VSV-CHIKV to assess induction of 278
immune responses and protective efficacy in a CHIKV challenge model. 279
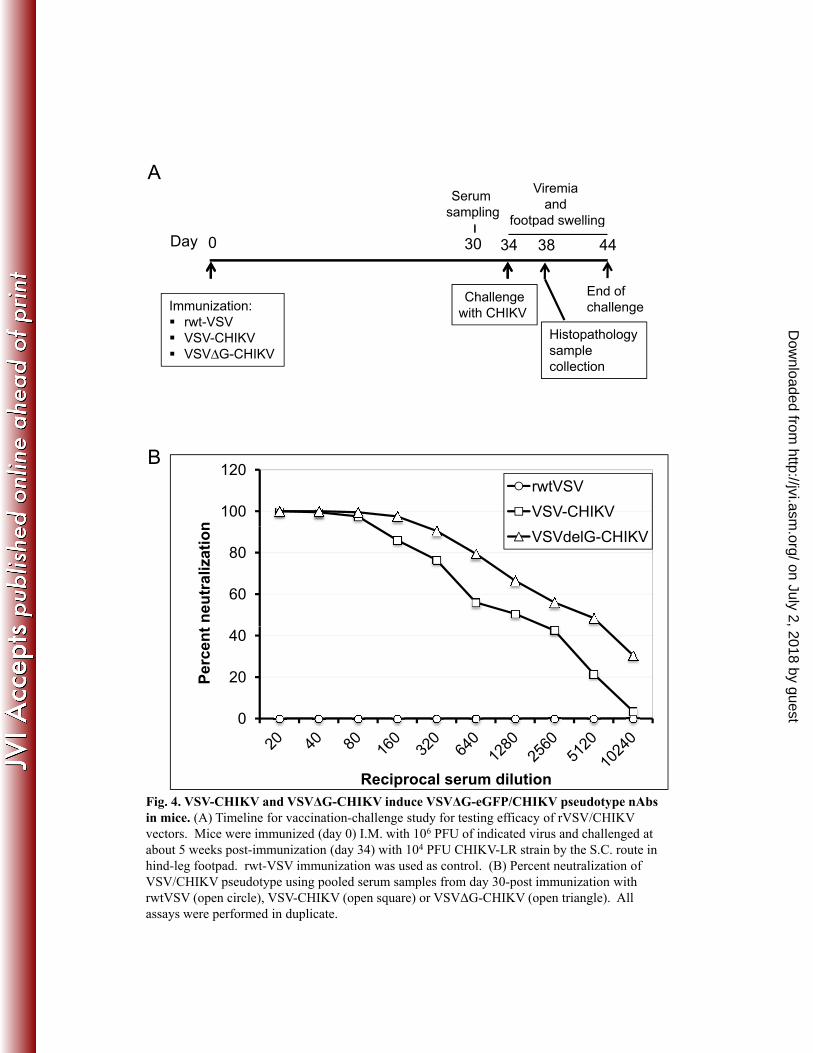
Immunization and challenge study. Fig. 4A shows the timeline of the 280
immunization/challenge study using 3.5 week-old C57Bl/6 mice. Two groups of eight mice each 281
were immunized I.M. with VSV-CHIKV or VSVΔG-CHIKV and a control group (n=8) received 282
rwt-VSV. Blood was collected from the animals at day 30-post immunization and used to 283
determine the CHIKV nAb response from each group. As shown in Fig. 4B, while both 284
VSV/CHIKV recombinants elicited 100% neutralization of VSV/CHIKV pseudotypes 285
(homologous S27 strain) at low dilution, VSVΔG-CHIKV induced a superior response at 286
dilutions greater than 1:160. However, the differences between the two vaccine groups did not 287
reach statistical significance. Control rwt-VSV immunized mice had no CHIKV nAb. To assess 288
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

14
the nAb responses to the heterologous CHIKV-LR strain (challenge virus), plaque reduction 289
neutralization tests (PRNT) were also performed using these serum samples. As shown in Table 290
1, all rVSV/CHIKV immunized animals had PRNT80 titers of ≥80 with the VSVΔG-CHIKV 291
group showing a higher nAb titer on average (ranging from 160 to >640) compared to VSV-292
CHIKV group (titer ranging from 80 to 320). Control mice that received rwt-VSV showed titers 293
below the detection limit (<20). Anti-vector nAb titers directed against VSV G were also 294
measured using pooled serum samples from each immunization group (not shown). Mice 295
immunized with rwt-VSV and VSV-CHIKV had VSV nAb titers of 1: 2,560, while mice 296
immunized with VSVΔG-CHIKV did not generate detectable VSV nAb (<1:20). 297
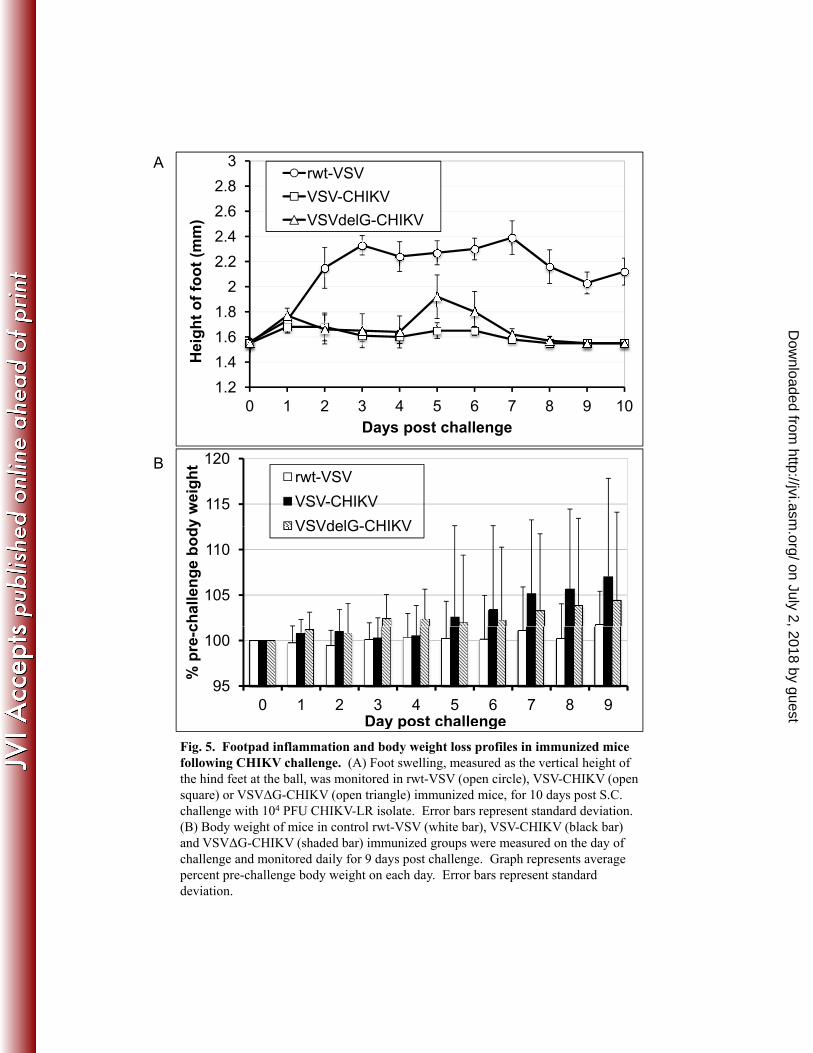
rVSV/CHIKV immunized mice are protected against CHIKV infection. To 298
determine if the VSV/CHIKV recombinants can protect against CHIKV-induced viremia and 299
disease, the immunized C57Bl/6 mice were subjected to S.C. challenge in their rear left footpad 300
with 104 PFU of CHIKV-LR strain at 34 days post immunization and followed for signs of 301
disease over 10 subsequent days. Four mice from each immunization group were bled on 302
alternate days post challenge to assess viremia. Control animals that received rwt-VSV showed 303
viremia during the first two days after challenge while none of the rVSV/CHIKV-vaccinated 304
mice had detectible viremia (Table 2). CHIKV induced disease was also monitored for 10 days 305
post challenge by following signs of local inflammation as measured by swelling of the 306
inoculated foot. As shown in Fig. 5A, rwt-VSV immunized control animals displayed 307
substantial footpad swelling that was sustained for 10 days after challenge. In contrast, mice in 308
both the rVSV/CHIKV-vaccinated groups separately showed significantly less (p<0.001, 309
ANOVA) foot swelling, which returned to pre-challenge levels by day 8 or 9 post challenge (Fig. 310
5A). While the control animals showed a transient weight loss or failed to gain weight after 311
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

15
challenge, VSV/CHIKV-immunized mice did not show any weight loss and gained weight (Fig. 312
5B). However, the differences in weight loss did not reach statistical significance because of the 313
high degree of animal-to-animal variability. 314
Two animals from each group were sacrificed at day 4-post challenge for histopathologic 315
evaluation (not shown). The brain, lung, heart, kidney, bowel, spleen, and muscle, demonstrated 316
no significant histopatholgic differences among the groups. It is possible that we might have 317
noted more significant histopathologic change at later times after infection. This has been noted 318
by others in the muscle at day 7 following challenge (8). We did however note microgranulomas 319
in the liver of all groups, and the vaccine groups showed a 75% reduction in the number of 320
microgranulomas compared to the rwt-VSV control group. These microgranulomas most likely 321
represent a non-specific reaction to hepatic injury. 322
VSV/CHIKV recombinants induce cellular immune responses in C57Bl/6 mice. 323
Because VSV based vaccine vectors are known to be potent inducers of both humoral and 324
cellular immune responses (28, 47), we also looked at the CHIKV specific cellular immune 325
response to the VSV/CHIKV vectors. A previous study, using a multi-dose DNA vaccine 326
expressing consensus CHIKV E1 or E2 sequences, reported induction of CHIKV specific CD8+ 327
CTL responses using peptide pools, and predicted two dominant CD8+ epitopes one in E1 328
protein and one in E2 protein (19). We used synthetic peptides corresponding to these predicted 329
epitopes (HSMTNAVTI on E1 and IILYYYELY on E2) in an IFN-γ ELISPOT assay. Our 330
studies (Fig. 6) showed that the VSV/CHIKV vectors induced CHIKV cellular immune 331
responses by day 7 after a single immunization. Although the response to the E1 epitope was 332
higher than the response to the E2 epitope in both vaccinated groups, VSVΔG-CHIKV induced a 333
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

16
significantly stronger (p<0.01, ANOVA) response to the E2 epitope compared to that of VSV-334
CHIKV. 335
336
DISCUSSION 337
We report here the development of experimental CHIKV vaccines based on VSV vectors 338
expressing the envelope proteins of CHIKV. These were either a full-length VSV-CHIKV 339
vector or a VSVΔG-CHIKV vector that lacks the VSV G gene. Both of these vectors generated 340
robust nAb responses to CHIKV in mice after a single dose, and protected mice from CHIKV 341
challenge. Most interestingly, we found that the VSVΔG-CHIKV recombinant incorporated 342
functional CHIKV envelope proteins into the viral envelope and propagated in the absence of 343
VSV G complementation. Although the VSVΔG-CHIKV recombinant grew more slowly than 344
the full-length vector and made only small plaques, it generated a stronger nAb response to 345
CHIKV than the full-length VSV-CHIKV vector. The stronger antibody responses probably 346
resulted from a combination of greater expression of CHIKV glycoproteins in the absence of 347
VSV G, greater display of the CHIKV proteins on the surface of the viral particles, and/or lack 348
of antigenic competition from VSV G. 349
Chimeric alphaviruses expressing structural protein genes from CHIKV and non-350
structural protein genes of either Venezuelan (VEEV) or eastern equine encephalitis virus 351
(EEEV) have been successfully used as experimental vaccines for CHIKV (43). These viruses 352
also generated potent CHIKV nAbs in mice and protection from challenge after a single dose. It 353
is interesting to note that the titers generated following a single dose VSVΔG-CHIKV vector 354
were comparable to those reported for the VEEV/EEEV-CHIKV chimeras (43). 355
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

17
Passive antibody transfer studies have shown that antibody alone can confer immunity to 356
CHIKV challenge (1, 6, 24). The role of cellular immunity in clearance of CHIKV infection is 357
not well studied. However, recent studies have shown that strong CHIKV-specific T-cell 358
activation occurs early in infection and likely plays a role in early control of viral replication 359
prior to generation of nAb (8, 10, 44). We showed here that a single dose of VSV/CHIKV 360
recombinants induced potent CD8 T-cell responses to dominant epitopes in CHIKV E1 and E2 361
proteins. Recall of memory T-cells generated by vaccination with VSV/CHIKV vectors could 362
therefore complement the robust B-cell responses and aid in viral clearance and protection from 363
CHIKV-induced disease. 364
A major advantage of the VSVΔG vector system is that it does not induce any anti-vector 365
nAb since it lacks the VSV G protein. Hence it could be used in multiple vaccine applications. 366
Based on our experience here with CHIKV, it is likely that VSV∆G vectors could be a general 367
chimeric vaccine platform for other medically important alphaviruses such as VEEV, EEEV and 368
western equine encephalitis virus (WEEV), which are widespread in the Americas. Although the 369
VSVΔG-CHIKV chimeric virus grew more slowly than wild-type VSV, it eventually reached 370
titers nearly equivalent to VSV and thus could easily be scaled up for large-scale vaccine 371
production. We have also found that the VSVΔG-CHIKV recombinant grows in Vero cells to 372
titers equivalent to those obtained in BHK-21 cells. This is important because Vero cells are an 373
approved cell line for vaccine production. 374
VSV-based vaccine vectors given by the I.M. route typically cause no measurable 375
pathogenesis in mice. They are trafficked to draining lymph nodes and rapidly eliminated 376
without causing systemic infection (11, 36). We also observed that the VSV-CHIKV and 377
VSV∆G-CHIKV viruses caused no pathogenesis when given intramuscularly. Interestingly, our 378
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

18
preliminary studies have shown that the VSV∆G-CHIKV vectors are non-pathogenic but 379
immunogenic when given by the intranasal route, while the VSV-CHIKV vector or the VSV 380
vector alone induce substantial pathogenesis (weight loss). A more extensive analysis of 381
pathogenesis and vector spread using multiple routes of inoculation will of course be required 382
before VSV∆G-CHIKV vectors could be moved into clinical trials. 383
Our studies have demonstrated that a VSV-vectored vaccine provides protection against a 384
CHIKV strain (La Reunion) that is about 2% divergent in sequence (37) compared to the vaccine 385
strain (S27). Based on these studies and others showing vaccine cross-protection among more 386
distantly related arthrogenic alphaviruses (8, 23), the VSV-vectored vaccine is likely to provide 387
protection against all three CHIKV clades. Vaccine cross-protection within the encephalitic 388
alphaviruses has also been reported (40, 45). Given the potent immune responses generated by 389
the VSVΔG-CHIKV recombinant, it will be important in the future to examine possible cross 390
protection against other arthrogenic viruses as well as the general applicability of the platform 391
for other alphaviruses. 392
393
ACKNOWLEDGEMENTS 394
This work was supported by NIH grant AI057158. We thank Drs. John Schell and Kapil Bahl 395
for advice on the T-cell studies, and Dr. Alex Ryder for assistance with statistical analysis. 396
397
REFERENCES 398
1. Akahata, W., Z. Y. Yang, H. Andersen, S. Sun, H. A. Holdaway, W. P. Kong, M. G. 399
Lewis, S. Higgs, M. G. Rossmann, S. Rao, and G. J. Nabel. 2010. A virus-like particle 400
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

19
vaccine for epidemic Chikungunya virus protects nonhuman primates against infection. 401
Nat Med 16:334-338. 402
2. Beaty, B. J., Calisher, C.H., Shope, R.E. 1995. Arboviruses, p. 189-212. In E. T. 403
Lennete, Lennete, D.A. (ed.), Diagnostic procedures for viral, rickettsial and chlamydial 404
infections, 7 ed. American Public Health Association, Washington, D.C. 405
3. Boritz, E., J. Gerlach, J. E. Johnson, and J. K. Rose. 1999. Replication-competent 406
rhabdoviruses with human immunodeficiency virus type 1 coats and green fluorescent 407
protein: entry by a pH-independent pathway. J Virol 73:6937-6945. 408
4. Chattopadhyay, A., S. Park, G. Delmas, R. Suresh, S. Senina, D. S. Perlin, and J. K. 409
Rose. 2008. Single-dose, virus-vectored vaccine protection against Yersinia pestis 410
challenge: CD4+ cells are required at the time of challenge for optimal protection. 411
Vaccine 26:6329-6337. 412
5. Chattopadhyay, A., and J. K. Rose. 2011. Complementing defective viruses that 413
express separate paramyxovirus glycoproteins provide a new vaccine vector approach. J 414
Virol 85:2004-2011. 415
6. Couderc, T., N. Khandoudi, M. Grandadam, C. Visse, N. Gangneux, S. Bagot, J. F. 416
Prost, and M. Lecuit. 2009. Prophylaxis and therapy for Chikungunya virus infection. J 417
Infect Dis 200:516-523. 418
7. Fuerst, T. R., E. G. Niles, F. W. Studier, and B. Moss. 1986. Eukaryotic transient-419
expression system based on recombinant vaccinia virus that synthesizes bacteriophage T7 420
RNA polymerase. Proc Natl Acad Sci U S A 83:8122-8126. 421
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

20
8. Gardner, J., I. Anraku, T. T. Le, T. Larcher, L. Major, P. Roques, W. A. Schroder, 422
S. Higgs, and A. Suhrbier. 2010. Chikungunya virus arthritis in adult wild-type mice. J 423
Virol 84:8021-8032. 424
9. Griffin, D. 2006. Alphaviruses, p. 1023-1067. In D. M. Knipe, Howley, P.M. (ed.), 425
Field's Virology, 5th ed. Lippincott, Williams & Wilkins, Philadelphia. 426
10. Hoarau, J. J., M. C. Jaffar Bandjee, P. Krejbich Trotot, T. Das, G. Li-Pat-Yuen, B. 427
Dassa, M. Denizot, E. Guichard, A. Ribera, T. Henni, F. Tallet, M. P. Moiton, B. A. 428
Gauzere, S. Bruniquet, Z. Jaffar Bandjee, P. Morbidelli, G. Martigny, M. Jolivet, F. 429
Gay, M. Grandadam, H. Tolou, V. Vieillard, P. Debre, B. Autran, and P. Gasque. 430
2010. Persistent chronic inflammation and infection by Chikungunya arthritogenic 431
alphavirus in spite of a robust host immune response. J Immunol 184:5914-5927. 432
11. Johnson, J. E., J. W. Coleman, N. K. Kalyan, P. Calderon, K. J. Wright, J. Obregon, 433
E. Ogin-Wilson, R. J. Natuk, D. K. Clarke, S. A. Udem, D. Cooper, and R. M. 434
Hendry. 2009. In vivo biodistribution of a highly attenuated recombinant vesicular 435
stomatitis virus expressing HIV-1 Gag following intramuscular, intranasal, or intravenous 436
inoculation. Vaccine 27:2930-2939. 437
12. Jones, S. M., H. Feldmann, U. Stroher, J. B. Geisbert, L. Fernando, A. Grolla, H. D. 438
Klenk, N. J. Sullivan, V. E. Volchkov, E. A. Fritz, K. M. Daddario, L. E. Hensley, P. 439
B. Jahrling, and T. W. Geisbert. 2005. Live attenuated recombinant vaccine protects 440
nonhuman primates against Ebola and Marburg viruses. Nat Med 11:786-790. 441
13. Kahn, J. S., A. Roberts, C. Weibel, L. Buonocore, and J. K. Rose. 2001. Replication-442
competent or attenuated, nonpropagating vesicular stomatitis viruses expressing 443
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

21
respiratory syncytial virus (RSV) antigens protect mice against RSV challenge. J Virol 444
75:11079-11087. 445
14. Kapadia, S. U., J. K. Rose, E. Lamirande, L. Vogel, K. Subbarao, and A. Roberts. 446
2005. Long-term protection from SARS coronavirus infection conferred by a single 447
immunization with an attenuated VSV-based vaccine. Virology 340:174-182. 448
15. Kapadia, S. U., I. D. Simon, and J. K. Rose. 2008. SARS vaccine based on a 449
replication-defective recombinant vesicular stomatitis virus is more potent than one based 450
on a replication-competent vector. Virology 376:165-172. 451
16. Lawson, N. D., E. A. Stillman, M. A. Whitt, and J. K. Rose. 1995. Recombinant 452
vesicular stomatitis viruses from DNA. Proc Natl Acad Sci U S A 92:4477-4481. 453
17. Lefrancois, L., and D. S. Lyles. 1982. The interaction of antibody with the major 454
surface glycoprotein of vesicular stomatitis virus. I. Analysis of neutralizing epitopes 455
with monoclonal antibodies. Virology 121:157-167. 456
18. Mallilankaraman, K., D. J. Shedlock, H. Bao, O. U. Kawalekar, P. Fagone, A. A. 457
Ramanathan, B. Ferraro, J. Stabenow, P. Vijayachari, S. G. Sundaram, N. 458
Muruganandam, G. Sarangan, P. Srikanth, A. S. Khan, M. G. Lewis, J. J. Kim, N. 459
Y. Sardesai, K. Muthumani, and D. B. Weiner. 2011. A DNA vaccine against 460
chikungunya virus is protective in mice and induces neutralizing antibodies in mice and 461
nonhuman primates. PLoS Negl Trop Dis 5:e928. 462
19. Muthumani, K., K. M. Lankaraman, D. J. Laddy, S. G. Sundaram, C. W. Chung, E. 463
Sako, L. Wu, A. Khan, N. Sardesai, J. J. Kim, P. Vijayachari, and D. B. Weiner. 464
2008. Immunogenicity of novel consensus-based DNA vaccines against Chikungunya 465
virus. Vaccine 26:5128-5134. 466
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

22
20. Niwa, H., K. Yamamura, and J. Miyazaki. 1991. Efficient selection for high-467
expression transfectants with a novel eukaryotic vector. Gene 108:193-199. 468
21. Okuma, K., Y. Matsuura, H. Tatsuo, Y. Inagaki, M. Nakamura, N. Yamamoto, and 469
Y. Yanagi. 2001. Analysis of the molecules involved in human T-cell leukaemia virus 470
type 1 entry by a vesicular stomatitis virus pseudotype bearing its envelope 471
glycoproteins. J Gen Virol 82:821-830. 472
22. Paessler, S., N. E. Yun, B. M. Judy, N. Dziuba, M. A. Zacks, A. H. Grund, I. Frolov, 473
G. A. Campbell, S. C. Weaver, and D. M. Estes. 2007. Alpha-beta T cells provide 474
protection against lethal encephalitis in the murine model of VEEV infection. Virology 475
367:307-323. 476
23. Partidos, C. D., J. Paykel, J. Weger, E. M. Borland, A. M. Powers, R. Seymour, S. C. 477
Weaver, D. T. Stinchcomb, and J. E. Osorio. 2012. Cross-protective immunity against 478
o'nyong-nyong virus afforded by a novel recombinant chikungunya vaccine. Vaccine. 479
24. Plante, K., E. Wang, C. D. Partidos, J. Weger, R. Gorchakov, K. Tsetsarkin, E. M. 480
Borland, A. M. Powers, R. Seymour, D. T. Stinchcomb, J. E. Osorio, I. Frolov, and 481
S. C. Weaver. 2011. Novel chikungunya vaccine candidate with an IRES-based 482
attenuation and host range alteration mechanism. PLoS Pathog 7:e1002142. 483
25. Powers, A. M., and C. H. Logue. 2007. Changing patterns of chikungunya virus: re-484
emergence of a zoonotic arbovirus. J Gen Virol 88:2363-2377. 485
26. Roberts, A., L. Buonocore, R. Price, J. Forman, and J. K. Rose. 1999. Attenuated 486
vesicular stomatitis viruses as vaccine vectors. J Virol 73:3723-3732. 487
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

23
27. Roberts, A., J. D. Reuter, J. H. Wilson, S. Baldwin, and J. K. Rose. 2004. Complete 488
protection from papillomavirus challenge after a single vaccination with a vesicular 489
stomatitis virus vector expressing high levels of L1 protein. J Virol 78:3196-3199. 490
28. Rose, J. K., Whitt, M.A. 2001. Rhabdoviridae: the viruses and their replication. In D. M. 491
Knipe, Howley, P.M., Griffin, D.E., Lamb, R.A.,Martin, M.A., Roizman, B., Straus, S.E. 492
(ed.), Fields' Virology, vol. 4th Ed. Lippincott Williams and Wilkins, Phhiladelphia. 493
29. Rose, N. F., P. A. Marx, A. Luckay, D. F. Nixon, W. J. Moretto, S. M. Donahoe, D. 494
Montefiori, A. Roberts, L. Buonocore, and J. K. Rose. 2001. An effective AIDS 495
vaccine based on live attenuated vesicular stomatitis virus recombinants. Cell 106:539-496
549. 497
30. Schnell, M. J., L. Buonocore, E. Kretzschmar, E. Johnson, and J. K. Rose. 1996. 498
Foreign glycoproteins expressed from recombinant vesicular stomatitis viruses are 499
incorporated efficiently into virus particles. Proc Natl Acad Sci U S A 93:11359-11365. 500
31. Schnell, M. J., L. Buonocore, M. A. Whitt, and J. K. Rose. 1996. The minimal 501
conserved transcription stop-start signal promotes stable expression of a foreign gene in 502
vesicular stomatitis virus. J Virol 70:2318-2323. 503
32. Schnell, M. J., J. E. Johnson, L. Buonocore, and J. K. Rose. 1997. Construction of a 504
novel virus that targets HIV-1-infected cells and controls HIV-1 infection. Cell 90:849-505
857. 506
33. Schwartz, J. A., L. Buonocore, A. Roberts, A. Suguitan, Jr., D. Kobasa, G. 507
Kobinger, H. Feldmann, K. Subbarao, and J. K. Rose. 2007. Vesicular stomatitis 508
virus vectors expressing avian influenza H5 HA induce cross-neutralizing antibodies and 509
long-term protection. Virology 366:166-173. 510
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

24
34. Schwartz, O., and M. L. Albert. 2010. Biology and pathogenesis of chikungunya virus. 511
Nat Rev Microbiol 8:491-500. 512
35. Simizu, B., K. Yamamoto, K. Hashimoto, and T. Ogata. 1984. Structural proteins of 513
Chikungunya virus. J Virol 51:254-258. 514
36. Simon, I. D., J. Publicover, and J. K. Rose. 2007. Replication and propagation of 515
attenuated vesicular stomatitis virus vectors in vivo: vector spread correlates with 516
induction of immune responses and persistence of genomic RNA. J Virol 81:2078-2082. 517
37. Solignat, M., B. Gay, S. Higgs, L. Briant, and C. Devaux. 2009. Replication cycle of 518
chikungunya: a re-emerging arbovirus. Virology 393:183-197. 519
38. Strauss, J. H., and E. G. Strauss. 1994. The alphaviruses: gene expression, replication, 520
and evolution. Microbiol Rev 58:491-562. 521
39. Tsetsarkin, K. A., R. Chen, M. B. Sherman, and S. C. Weaver. 2011. Chikungunya 522
Virus: Evolution and Genetic Determinants of Emergence. Curr Opin Virol 1:310-317. 523
40. Walton, T. E., M. M. Jochim, T. L. Barber, and L. H. Thompson. 1989. Cross-524
protective immunity between equine encephalomyelitis viruses in equids. Am J Vet Res 525
50:1442-1446. 526
41. Wang, D., A. Suhrbier, A. Penn-Nicholson, J. Woraratanadharm, J. Gardner, M. 527
Luo, T. T. Le, I. Anraku, M. Sakalian, D. Einfeld, and J. Y. Dong. 2011. A complex 528
adenovirus vaccine against chikungunya virus provides complete protection against 529
viraemia and arthritis. Vaccine 29:2803-2809. 530
42. Wang, E., D. Y. Kim, S. C. Weaver, and I. Frolov. 2011. Chimeric Chikungunya 531
viruses are nonpathogenic in highly sensitive mouse models but efficiently induce a 532
protective immune response. J Virol 85:9249-9252. 533
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

25
43. Wang, E., E. Volkova, A. P. Adams, N. Forrester, S. Y. Xiao, I. Frolov, and S. C. 534
Weaver. 2008. Chimeric alphavirus vaccine candidates for chikungunya. Vaccine 535
26:5030-5039. 536
44. Wauquier, N., P. Becquart, D. Nkoghe, C. Padilla, A. Ndjoyi-Mbiguino, and E. M. 537
Leroy. 2011. The acute phase of Chikungunya virus infection in humans is associated 538
with strong innate immunity and T CD8 cell activation. J Infect Dis 204:115-123. 539
45. Wust, C. J., R. Crombie, and A. Brown. 1987. Passive protection across subgroups of 540
alphaviruses by hyperimmune non-cross-neutralizing anti-Sindbis serum. Proc Soc Exp 541
Biol Med 184:56-63. 542
46. Zavada, J., Z. Zavadova, G. Russ, K. Polakova, J. Rajcani, J. Stencl, and J. Loksa. 543
1983. Human cell surface proteins selectively assembled into vesicular stomatitis virus 544
virions. Virology 127:345-360. 545
47. Zinkernagel, R. M., B. Adler, and J. J. Holland. 1978. Cell-mediated immunity to 546
vesicular stomatitis virus infections in mice. Exp Cell Biol 46:53-70. 547
548
549
550
551
552
553
554
555
556
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

26
FIGURE LEGENDS 557
Fig. 1. Characterization of recombinant VSVs encoding the CHIKV E3-E2-6K-E1 polyprotein. 558
(A) Diagram of rVSV genomes with the insertion of CHIKV envelope polyprotein in full-length 559
VSV and VSVΔG vectors. The gene order is shown in 3’ to 5’ direction on the negative sense 560
RNA genome. (B) Expression of CHIKV envelope proteins detected by indirect 561
immunofluorescence microscopy using anti-VSV G MAb or anti-CHIKV mouse sera in infected 562
(as indicated) BHK-21 cells. Photomicrographs show cell nuclei stained with DAPI in all fields. 563
(C) [35S]-methionine labeled crude cell lysates of infected BHK-21 cells analyzed by SDS-PAGE 564
showing VSV proteins and CHIKV envelope proteins. VSV proteins are indicated to the left, 565
and CHIKV envelope proteins and molecular mass of VSV proteins are indicated to the right. 566
(D) [35S]-methionine labeled and purified virions analyzed by SDS-PAGE. Proteins 567
incorporated into the virions are indicated. 568
569
Fig. 2. Growth of recombinant VSVs and plaque morphology. (A) BHK-21 cells were infected 570
(MOI=10) for 30 min with rwt-VSV, VSV-CHIKV or VSVΔG-CHIKV and unadsorbed virus 571
was removed by washing cells twice with PBS. Complete growth medium was added and 572
supernatants were collected at indicated times post-adsorption. Virus titers were determined by 573
plaque assay on BHK-21 cells. (B) Plaque morphology of VSV-CHIKV and VSVΔG-CHIKV at 574
2 days post infection of BHK-21 cells. 575
576
Fig. 3. UV inactivation prevents induction of CHIKV nAbs. Mice were immunized with non-577
complemented (NC, filled square) or G-complemented (GC, filled triangle) VSVΔG-CHIKV 578
without or with UV inactivation [NC-UV (open square) and GC-UV (open triangle)] and pooled 579
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

27
serum from day 30-post immunization were analyzed for induction of VSV∆G-eGFP/CHIKV 580
pseudotype nAb. The representative curve shows percent neutralization at indicated serum 581
dilutions. All assays were performed in duplicate. 582
583
Fig. 4. VSV-CHIKV and VSVΔG-CHIKV induce VSVΔG-eGFP/CHIKV pseudotype nAbs in 584
mice. (A) Timeline for vaccination-challenge study for testing efficacy of rVSV/CHIKV 585
vectors. Mice were immunized (day 0) I.M. with 106 PFU of indicated virus and challenged at 586
about 5 weeks post-immunization (day 34) with 104 PFU CHIKV-LR strain by the S.C. route in 587
hind-leg footpad. rwt-VSV immunization was used as control. (B) Percent neutralization of 588
VSV/CHIKV pseudotype using pooled serum samples from day 30-post immunization with 589
rwtVSV (open circle), VSV-CHIKV (open square) or VSVΔG-CHIKV (open triangle). All 590
assays were performed in duplicate. 591
592
Fig. 5. Footpad inflammation and body weight loss profiles in immunized mice following 593
CHIKV challenge. (A) Foot swelling, measured as the vertical height of the hind feet at the ball, 594
was monitored in rwt-VSV (open circle), VSV-CHIKV (open square) or VSVΔG-CHIKV (open 595
triangle) immunized mice, for 10 days post S.C. challenge with 104 PFU CHIKV-LR isolate. 596
Error bars represent standard deviation. (B) Body weight of mice in control rwt-VSV (white 597
bar), VSV-CHIKV (black bar) and VSVΔG-CHIKV (shaded bar) immunized groups were 598
measured on the day of challenge and monitored daily for 9 days post challenge. Graph 599
represents average percent pre-challenge body weight in each group over time. Error bars 600
represent standard deviation. 601
602
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

28
Fig. 6. Induction of cellular immune response in rVSV/CHIKV immunized mice. Splenocytes, 603
harvested from uninfected naïve mice (n=2) or mice immunized with 106 PFU of VSV-CHIKV 604
(n=5) or VSVΔG-CHIKV (n=5), at day 7 post I.M. immunization were stimulated with peptides 605
corresponding to CHIKV E1 (HSMTNAVTI; open bar) or E2 (IILYYYELY; filled bar) CD8+ 606
epitopes for 24 h followed by detection of IFN-γ spot forming cells (SFC) using an Elispot assay. 607
The numbers of SFC from unstimulated control samples have been subtracted. Error bars 608
represent standard deviation. * Indicates a significant difference (p<0.01, ANOVA) between 609
VSV-CHIKV and VSVΔG-CHIKV groups in response to E2 peptide. 610
611
612
613
614
615
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

TABLE 1. Titers of CHIKV specific neutralizing antibodies at day 30 post immunization with
different VSV recombinants
a Groups of 3.5 weeks old C57Bl/6 mice were immunized I.M. with 106 PFU of indicated
viruses. Neutralizing antibody titers were determined in day 30 post-immunization serum
samples using PRNT80.
Mouse Number
PRNT80a
rwt-VSV VSV-CHIKV VSVΔG-CHIKV
1 <20 320 640
2 <20 320 >640
3 <20 160 >640
4 <20 320 320
5 <20 320 160
6 <20 80 320
7 <20 80 160
8 <20 320 320
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

TABLE 2. Viremia in vaccinated and control mice after challenge with CHIKVa
Immunization Group
Virus titer (log10 PFU/ml ± S.D.) (Fraction positive) Day 1b Day 2c Day 3b Day 4c
rwt-VSV 2.55±0.56 (2/4) 2.85±0.47 (4/4) <0.9 (0/4) 2.0±0.0 (1/4)
VSV-CHIKV <0.9 (0/4) <0.9 (0/4) <0.9 (0/4) <0.9 (0/4)
VSVΔG-CHIKV <0.9 (0/4) <0.9 (0/4) <0.9 (0/4) <0.9 (0/4) a S.C. challenge in rear left footpad with 104 PFU of CHIKV La Reunion strain.
b Four mice in each group were bled on day 1 and 3 post challenge.
c Remaining four mice in each group were bled on day 2 and 4 post challenge.
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

VSV-CHIKV(E3-E2-6K-E1)5’3’ LGMPN E1E3 E2 6K
VSVΔG-CHIKV(E3-E2-6K-E1)5’LE1E3 E2 6K3’ MPN
A
rwt-VSV VSV-CHIKV VSVΔG-CHIKV
α-V
SV
GB
α-C
HIK
Vα
C D
p62
240
63
L
G
L
Gp62E1
47
26
GNPM
NPM
E1/E2
Fig. 1. Characterization of recombinant VSVs encoding the CHIKV E3-E2-6K-E1 polyprotein. (A) Diagram of rVSV genomes with insertion of CHIKV envelope polyprotein in full-length VSV and VSVΔG vectors. The gene order is shown in 3’ to 5’ direction on the negative sense RNA genome. (B) Expression of CHIKV envelope proteins detected by indirect immunofluorescence microscopy using anti-VSV G MAb or anti-CHIKV mouse sera in infected (as indicated) BHK-21 cells. Photomicrographs show cell nuclei stained with DAPI in all fields. (C) [35S]-methionine labeled crude cell lysates of infected BHK-21 cells analyzed by SDS-PAGE h i VSV i d CHIKV l i VSV i i di d h l f dshowing VSV proteins and CHIKV envelope proteins. VSV proteins are indicated to the left, and
CHIKV envelope proteins and molecular mass of VSV proteins are indicated to the right. (D) [35S]-methionine labeled and purified virions analyzed by SDS-PAGE. Proteins incorporated into the virions are indicated.
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

109A
1.00E+07
1.00E+08
1.00E+09pf
u/m
l)108
107
109A
1.00E+04
1.00E+05
1.00E+06
Viru
s tit
er (p
rwtVSV
VSV-CHIKV
VSVdelG-CHIKV
106
105
104
1.00E+03
0 2 4 6 8 10 12 14 16 18 20 22 24Hours post adsorption
103
B
VSV-CHIKV VSVΔG-CHIKV
Fig 2 Growth of recombinant VSVs and plaque morphology (A) BHK 21 cellsFig. 2. Growth of recombinant VSVs and plaque morphology. (A) BHK-21 cells were infected (MOI=10) for 30 min with rwt-VSV, VSV-CHIKV or VSVΔG-CHIKV and unadsorbed virus was removed by washing cells twice with PBS. Complete growth medium was added and supernatant samples were collected at indicated times post-adsorption. Virus titers were determined by plaque assay on BHK-21 cells. (B) Plaque morphology of VSV-CHIKV and VSVΔG-CHIKV at 2 days post infection of BHK-21 cells.ce s.
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
80
100
120
zatio
n
NCNC-UVGCGC-UV
40
60
rcen
t neu
tral
iz
0
20
20 40 80 160 320 640 1280 2560 5120
Pe
Fig. 3. UV inactivation prevents induction of CHIKV nAbs. Mice were immunized with non-complemented (NC, filled square) or G-complemented (GC, filled triangle) VSVΔG-CHIKV without or with UV inactivation [NC-UV (open square) and GC-UV (open triangle)] and pooled serum from day 30-post immunization were analyzed for induction of VSV∆G-
Reciprocal serum dilution
eGFP/CHIKV pseudotype neutralizing antibodies. The representative curve shows percent neutralization at indicated serum dilutions. All assays were performed in duplicate.
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
Serum sampling
Viremiaand
footpad swelling
A
Immunization: rwt-VSV
Day 0 30 34 38 44
Challengewith CHIKV
End of challenge
p g
rwt VSV VSV-CHIKV VSVΔG-CHIKV
Histopathology sample collection
B
100
120
n
rwtVSVVSV-CHIKV
60
80
neut
raliz
atio
n
VSVdelG-CHIKV
0
20
40
Perc
ent
Fig. 4. VSV-CHIKV and VSVΔG-CHIKV induce VSVΔG-eGFP/CHIKV pseudotype nAbsin mice. (A) Timeline for vaccination-challenge study for testing efficacy of rVSV/CHIKV
Reciprocal serum dilution
vectors. Mice were immunized (day 0) I.M. with 106 PFU of indicated virus and challenged at about 5 weeks post-immunization (day 34) with 104 PFU CHIKV-LR strain by the S.C. route in hind-leg footpad. rwt-VSV immunization was used as control. (B) Percent neutralization of VSV/CHIKV pseudotype using pooled serum samples from day 30-post immunization with rwtVSV (open circle), VSV-CHIKV (open square) or VSVΔG-CHIKV (open triangle). All assays were performed in duplicate.
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
2.62.83
m)
rwt-VSVVSV-CHIKVVSVdelG-CHIKV
A
1.82
2.22.4
ht o
f foo
t (m
m
1.21.41.6
0 1 2 3 4 5 6 7 8 9 10
Hei
gh
Days post challengeDays post challenge
115
120
y w
eigh
t
rwt-VSVVSV-CHIKVVSVdelG CHIKV
B
105
110
chal
leng
e bo
dy VSVdelG-CHIKV
95
100
0 1 2 3 4 5 6 7 8 9
% p
re-c
Day post challengeFig. 5. Footpad inflammation and body weight loss profiles in immunized mice following CHIKV challenge. (A) Foot swelling, measured as the vertical height of the hind feet at the ball, was monitored in rwt-VSV (open circle), VSV-CHIKV (open square) or VSVΔG-CHIKV (open triangle) immunized mice, for 10 days post S.C. challenge with 104 PFU CHIKV-LR isolate. Error bars represent standard deviation. (B) Body weight of mice in control rwt VSV (white bar) VSV CHIKV (black bar)
y p g
(B) Body weight of mice in control rwt-VSV (white bar), VSV-CHIKV (black bar) and VSVΔG-CHIKV (shaded bar) immunized groups were measured on the day of challenge and monitored daily for 9 days post challenge. Graph represents average percent pre-challenge body weight on each day. Error bars represent standard deviation.
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

800
900
1000
E1
*
500
600
700
FC/106cells E2
100
200
300
400S
0
100
Naïve VSV-CHIKV VSVdelG-CHIKVNaive VSV-CHIKV VSVΔG-CHIKV
Fig. 6. Induction of cellular immune response in rVSV/CHIKV immunized mice. Splenocytes, harvested from uninfected naïve mice (n=2) or mice immunized with 106 PFU of VSV-CHIKV (n=5) or VSVΔG-CHIKV (n=5), at day 7 post I.M. immunization were stimulated with peptides corresponding to CHIKV E1 (HSMTNAVTI; open bar) or E2 (IILYYYELY; filled bar) CD8+ epitopes for 24 h followed by detection of IFN-γ spot forming cells (SFC) using an Elispot assay. The numbers of SFC from unstimulated control samples have been subtracted. Error b t t d d d i ti * I di t i ifi t diff ( <0 01bars represent standard deviation. * Indicates a significant difference (p<0.01, ANOVA) between VSV-CHIKV and VSVΔG-CHIKV groups in response to E2 peptide.
on July 2, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from