a complex of p190rhogap and anillin modulates rhogtp and...
TRANSCRIPT

1
A complex of p190RhoGAP and anillin modulates RhoGTP and the
cytokinetic furrow in human cells
Arkadi Manukyan �,1,5, Kirsten Ludwig �,2,4,5 , Sergio Sanchez-Manchinelly3,4,5, Sarah J.
Parsons4,5,% & P. Todd Stukenberg1,5,%,*
� = These authors contributed equally to this work
%The work was performed equally in both laboratories
1=Department of Biochemistry and Molecular Genetics, University of Virginia Health
System
2= Department of Psychiatry and Behavioral Sciences and Jonsson Cancer Center,
UCLA, Los Angeles,
3=Deallus, Inc., Los Angeles
4=Department of Microbiology, Immunology, and Cancer Biology, University of Virginia
Health System
5=Cancer Center, University of Virginia Health System
PO Box 800733, Charlottesville, VA 22908, USA
*Corresponding Author
TEL: 434-924-5253
FAX: 434-243-7244
Email: [email protected]
Keywords: p190RhoGAP, Anillin, Myosin II, Cytokinesis, Rho
© 2014. Published by The Company of Biologists Ltd.Jo
urna
l of C
ell S
cien
ceA
ccep
ted
man
uscr
ipt
JCS Advance Online Article. Posted on 29 October 2014

2
ABSTRACT
The cytokinetic furrow (CF) is organized by the RhoA GTPase, which recruits actin and
myosin II to the furrow and drives contractility. Here we show a role for the RhoGAP, p190, in
cytokinesis and its involvement in regulating Rho GTP levels and contractility. Cells depleted of
p190RhoGAP (p190) accumulate high levels of RhoGTP and markers of high Rho activity in the
furrow, resulting in failure of the CF to progress to abscission. The loss of p190 can be rescued
by a low dose of the myosin II inhibitor blebbistatin, suggesting that cells fail cytokinesis
because they have too much myosin activity. p190RhoGAP binds the cytokinetic organizer
anillin, and mutants of p190 that are unable to bind anillin or unable to inactivate Rho fail to
rescue cytokinesis defects in p190-depleted cells. Together these data demonstrate that a
complex of p190RhoGAP and anillin modulates RhoGTP levels in the CF to ensure robust
cytokinesis.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

3
INTRODUCTION
Cytokinesis is the final step in cell division where formation and ingression of the
cytokinetic furrow (CF) results in separation of two daughter cells. The process initiates in
anaphase, continues throughout telophase when membrane invagination at the equatorial cell
cortex occurs, and is completed upon membrane abscission (Glotzer, 2001). The contractile
forces required for furrow ingression are provided by a ring of filamentous actin and myosin II
that are juxtaposed to the cell membrane at the equator of the dividing cell. Assembly and
regulation of this contractile ring is critical for achieving proper cell division and is under the
control of the small GTPase RhoA (Piekny et al., 2005).
Early in cytokinesis RhoA localizes to the site of the nascent CF (Nishimura et al., 1998;
Takaishi et al., 1995), where its activity is required for CF formation and contraction, as well as
progression through cytokinesis (Bement et al., 2006). During early anaphase, RhoGTP levels
increase at the contractile ring (Kimura et al., 2000; Maddox and Burridge, 2003; Yoshizaki et
al., 2004), and active RhoA mediates contractile ring assembly through its downstream effectors,
mDia2, and ROCK Kinase (Piekny et al., 2005). mDia2 functions as an actin nucleator that
induces the polymerization of long, unbranched actin filaments (Watanabe et al., 2008), along
with microtubule alignment and stabilization (Narumiya and Yasuda, 2006). ROCK kinase
activates the contractile mechanism of myosin II by phosphorylating Thr18/Ser19 of the
regulatory light chain of myosin II (MLC II) (Matsumura, 2005). Phosphorylation of these sites
on MLC II triggers the motor (ATPase) activity of myosin II, which in turn promotes
crosslinking with newly created actin filaments to form a fully functional contractile ring
(Vavylonis et al., 2008; Wu et al., 2006) that provides the mechanical force required for furrow
contraction and ingression (Bresnick, 1999).
Actomyosin filaments are assembled on a network of cytoskeletal proteins at the cell
cortex, which connect the filaments to the plasma membrane. Anillin, an actin-binding protein, is
a crucial component of this scaffold that is required for cytokinesis (Oegema et al., 2000; Piekny
and Glotzer, 2008; Liu et al., 2012). Anillin has been shown to interact with actin, myosin,
microtubules, septin, mDia, Ect2, MgcRacGAP, Citron Kinase, and RhoA (Gai et al., 2011;
Piekny and Maddox, 2010). Additionally, overexpression of anillin increases Rho activation,
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

4
suggesting that anillin functions in the regulation of Rho during cytokinesis (Suzuki et al., 2005).
Together, these findings support the model that anillin plays a vital role in linking the structural
components of the contractile ring to Rho-regulated signaling proteins, which govern
cytokinesis.
RhoA is a molecular switch that cycles between active (GTP-bound) and inactive (GDP-
bound) states. Transitions between activation states are facilitated by guanine nucleotide
exchange factors (GEFs) (activators), GTPase activating proteins (GAPs) (inactivators), and
guanine nucleotide dissociation inhibitors (GDIs) (inactivators) (Jaffe and Hall, 2005).
MgcRacGAP/Cyk4/RacGAP1 is a major GAP known to function in cytokinesis (Zhao and Fang,
2005; Loria et al. 2012). It binds with the mitotic kinesin like protein, MKLP1/Zen4, to form the
heterotetrameric centralspindlin complex, a key structural component of the spindle midzone
(Mishima et al., 2002). Centralspindlin carries out several critical functions during cell division,
including bundling of antiparallel microtubules to form the central spindle (Jantsch-Plunger et
al., 2000), positioning of the cell division plane, and promoting cycles of RhoA GTPase activity
(D'Avino et al., 2005). The GAP domain of MgcRacGAP limits the amount of RhoGTP at the
furrow and is required for robust cytokinesis (Miller and Bement 2009). In early cytokinesis
MgcRacGAP also activates Rho through complex formation with the guanine nucleotide
exchange factor Ect2 (Somers and Saint, 2003, Yüce et al., 2005, Glotzer, 2009). Recently it has
been shown that MgcRacGAP can bind the plasma membrane through its C1 domain and
promotes cortical contractility and the anchoring of the CF to the midbody (Lekomtsev et al.,
2012). While it is undisputed that MgcRacGAP is a critical cytokinesis regulator it is not clear if
it acts as a RhoGAP or a RacGAP or both, since there is also evidence that MgcRacGAP inhibits
Rac activity during cytokinesis (D’avino et al., 2004, Canman et al., 2008, Bastos et al., 2012).
A second RhoGAP, MP-GAP, has been shown to repress “blebbing” in mitotic cells, and cells
depleted of MPGAP fail in cytokinesis about 18% of the time (Zanin et al., 2013).
Our previous findings suggest that mammalian cells may have an additional level of
RhoA regulation during CF progression, namely p190RhoGAP (p190) (Su et al, 2003; Mikawa
et al, 2008). P190 localizes to the CF (Su et al, 2003), and cells ectopically expressing the protein
exhibit reduced RhoGTP levels in the furrow and become multinucleated, supporting the idea
that proper cycling of RhoA between active and inactive states is required for completion of
cytokinesis (Su et al., 2009). Moreover, in the absence of ectopic expression, it has been
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

5
demonstrated that a transient, proteasome-mediated partial reduction (50%) of endogenous p190
levels during anaphase is required for completion of cytokinesis (Manchinelly et al., 2010; Su et
al., 2003), suggesting that p190 levels and activities are also critical for cytokinesis. During the
current study we overcame a technical challenge that has allowed us to characterize the loss of
function phenotypes of p190RhoGAP in cytokinesis. We report that cells depleted of p190 have
increased levels of RhoGTP in the CF and often fail to divide. Low concentrations of the myosin
II inhibitor blebbistatin can rescue loss of p190 suggesting that cells fail cytokinesis because they
have increased contractility. Additionally, p190 physically associates with anillin, and mutants
of p190 that are unable to interact with anillin fail in cytokinesis. Evidence is presented to
indicate that the interaction between p190 and anillin is regulated by contractile forces, as the
binding is disrupted by the addition of the myosin II inhibitor blebbistatin. Together, these data
demonstrate that p190 acts as a RhoGAP during cytokinesis in mammalian cells and provide
evidence that MLC II, anillin, and p190 act in a feedback loop that modulates Rho GTPase
activity in the CF.
RESULTS
p190RhoGAP is required for proper cytokinesis in HeLa cells.
To determine if depletion of p190 causes failure of cytokinesis, HeLa cells were
transfected with p190-specific small interfering RNAs (siRNAs) for 48 hours, and the number of
multinucleated cells were quantified (Fig. 1A, B). Twelve to fifteen percent of p190 depleted
cells were multinucleated, which is significantly higher compared to control siRNA-treated cells
(n>200). Figure 1, panel C shows the level of p190 depletion after 48 hours of siRNA treatment.
To determine whether exogenous p190 expression could restore normal cell division, we
generated a stable, wild type p190 doxycycline-inducible HeLa cell line that is resistant to
siRNA depletion. Replacement of endogenous p190 with doxycycline-inducible wild type p190
rescued the multinucleation phenotype, thereby addressing concerns about off target siRNA
effects and demonstrating the requirement of p190 for productive cytokinesis (Fig. 1B-C, Fig.
S1A).
To further characterize the cytokinetic defect caused by p190, we followed the behavior
of p190-depleted HeLa cells by live cell differential interference contrast microscopy (DIC) (Fig.
1D, Movie S1/S2). p190 depletion did not affect CF formation and initial ingression, processes
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

6
that are regulated by MgcRacGAP (Zhao and Fang, 2005). However, p190-depleted cells often
failed in abscission, and cells became multinucleated. The most striking phenotype of p190-
depleted cells in telophase was an inability to complete furrow contraction. In addition,
telophase control cells usually migrated in opposite directions and this movement was lost in
p190 depleted cells. Quantification of live cell imaging showed that eight percent of p190
siRNA-treated cells (12/195) failed cytokinesis compared to one percent of control siRNA-
treated cells (1/135), which is similar to the failure rate measured in fixed cells. We conclude that
p190 is required for optimal cytokinesis.
p190RhoGAP regulates RhoGTP during cytokinesis in HeLa cells.
p190 is a RhoGAP that down-regulates RhoGTP levels in interphase cells (Ridley et al.,
1993); however, the effect of loss of p190 on RhoGTP levels in cells undergoing cytokinesis has
not been measured. To address this question p190-depleted HeLa cells were synchronized in
nocodazole for 14-16 hours and released into drug-free medium for 40 minutes to enrich for cells
in late mitosis and undergoing cytokinesis. Lysates were subjected to a RhoA G-LISA assay, a
highly quantitative colorometric assay to measure the amount of RhoAGTP. Figure 2A shows
that p190-depleted cells in cytokinesis exhibited significantly elevated RhoGTP levels compared
to those of siRNA untreated (cytokinetic) cells or cells treated with a control Luciferase siRNA
or with C3, a toxin that efficiently inhibits RhoA activity. Moreover, RhoGTP levels in p190-
depleted cytokinetic cells were similar to those of cells treated with LPA, a potent activator of
RhoA. To determine if RhoA levels are increased at the cytokinetic furrow in p190 depleted cells
we fixed cells with TCA and immunostained for RhoA. TCA precipitation is a validated method
to preserve Rho bound to membranes for immunofluorescence (Yonemura et al., 2004). We
found significantly higher concentrations of RhoA at the CF in late cytokinesing cells after p190
depletion (Fig. 2B-B’). Thus, silencing of p190 effected both localization of Rho and the levels
of RhoGTP in cytokinesing cells. This increase is similar to the level of RhoGTP measured in
the furrow by FRET following ectopic expression of a dominant negative form of p190 in HeLa
cells (Su et al., 2009).
An independent measure of RhoGTP levels is the phosphorylation of myosin II light
chain (MLC II) on Thr18/Ser19, which we assessed by using both anti-phospho-Ser19 MLC II
and anti-phospho-Thr18/Ser19 (pMLC II) antibodies. Both antibodies recognized cytokinetic
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

7
furrows, and have different staining patterns as anti-phospho-Ser19 MLC II antibody stains
pMLC localized only to the contracting membrane, while anti-phospho-Thr18/Ser19 (pMLC II)
antibody also stains pMLC localized to the midzone (Kondo et al., 2012). Cells were stained
with anti-phospho-Ser19 MLC antibody, the intensity of staining at cytokinetic furrows was
quantified (areas quantified are outlined in Fig. S1C), and the ratio of pMLC II at the CF and
outside of the furrow area was determined. We observed an approximate 2-fold increase in
specific MLC II Ser-19 phosphorylation in the furrows of p190-depleted cells as compared to
controls (Fig. 2C-C’). To confirm that the increase in pMLC at the CF is not due to an increase
in total myosin, cells were co-stained with antibodies to anti-phospho-Thr18/Ser19 MLC II and
MLC II, the intensity of staining of both antibodies at the cytokinetic furrow was quantified, and
the ratio of pMLC II/MLC II was determined (Fig. S1D-D’). We see an increase in Rho activity
measured in this manner using pMLC II antibodies against MLC II pSer19 or MLC II pThr18,
pSer19 (Fig. 2C, C’, S1D). Moreover, the increase in phosphorylation of MLC II at the CF due
to p190 depletion could be rescued by exogenously expressing siRNA resistant wild type p190
(Fig. 2C-C’), ruling out siRNA off target effects.
To further investigate if p190 down-regulates RhoGTP levels through its GAP activity,
we generated a stable doxycycline-inducible HeLa cell line that expresses a p190 (R1283A)
point mutant that is defective in GAP activity (Li et al., 1997) (Fig. S1B). Cells expressing this
mutant were analyzed for their ability to rescue the cytokinesis defect associated with depletion
of endogenous p190. The number of multinucleated cells was significantly decreased after rescue
by exogenous wild type (Fig. 1B) but not the GAP mutant p190 (Fig. 2D). The expression level
of p190 (R1283A) was similar to that of the endogenous protein (Fig. 2E). We conclude that
p190 acts as a RhoGAP to down-regulate RhoGTP in the cytokinetic furrow.
Overexpression of p190 decreases RhoGTP at the furrow.
Overexpression of p190 had the opposite effect as depletion on RhoGTP levels in the
furrow. HeLa cells were transiently transfected with GFP-p190 or vector control plasmid,
immuno-stained with Thr18/Ser19 pMLC and MLC antibodies 24 hours later, and examined by
fluorescent confocal microscopy. Approximately 90% of cells showed reduced phosphorylation
of MLC II in their CFs as compared to vector-transfected controls (Fig. 3A-B, Fig. S1E).
Moreover, the ratio of pMLC II/MLC II was approximately 2-fold lower (Fig. 3C-D). These
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

8
effects were similar to those seen after depletion of anillin or addition of C3 toxin, both of which
have been previously shown to affect Rho activation at the CF (Aktories et al., 1989; Piekny and
Glotzer, 2008). Neither p190 overexpression nor C3 treatment affected the levels of total Rho
protein (Fig. S1F), nor did p190 overexpression affect the localization of RhoA, actin, Aurora B,
or microtubules (Fig. S1G), suggesting that p190 specifically affected RhoA GTPase activity
during cytokinesis.
We tested whether constitutively active-MLC II (MLCII (T18D, S19D); CA-MLC-II)
could rescue the multinucleation phenotype generated by p190 overexpression to determine
whether reduced activation of myosin II caused the failure in cytokinesis. Co-transfection of
p190 and CA MLC II significantly decreased the amount of p190-induced multinucleation, while
transfection of CA-MLC II alone had little effect (Fig. 3E). Together, these data demonstrate that
p190 is required for the completion of cytokinesis in HeLa cells, where it acts as a RhoGAP to
down-modulate RhoGTP and pMLC levels to modulate proper CF contraction.
Low doses of Blebbistatin can rescue loss of p190RhoGAP
Since we find higher levels of Rho activity at the CF we tested whether high contractility
caused the cytokinesis failure associated with p190 depletion. Specifically, we asked if low
concentrations of the myosin II inhibitor blebbistatin, which weakens contractility, could rescue
the multinucleation phenotype generated by p190 depletion. HeLa cells were synchronized using
double thymidine block, and endogenous p190 was depleted by siRNA for 48 hours. Eight hours
after release from thymidine HeLa cells were treated for 6 hours with concentrations of
blebbistatin below the amount used to fully inhibit myosin activity. Cells were then fixed and
prepared for confocal microscopy analysis and scored for multinucleation (Fig. 4A). Low
concentrations of blebbistatin rescued p190 depletion in a concentration dependent manner with
0.25 uM giving the best rescue. Higher concentrations of blebbistatin generated multinucleated
cells, as one would expect for full inhibition of myosin. To complement this experiment we also
inhibited ROCK kinase, which is a Rho dependent activator of myosin, with the inhibitor
Y27632. A dose of Y27632 that was below the concentration used to fully inhibit ROCK activity
also partially rescued the generation of multinucleated cells after p190 depletion (Fig S2A). The
fact that low doses of blebbistatin and Y27632 both rescue p190 depletion argues that cells
depleted of p190 fail cytokinesis because of too much Rho stimulated myosin activity.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

9
p190RhoGAP binds with anillin during cytokinesis.
Anillin is a key regulator of cytokinesis in metazoans, thought to function as a scaffold
via its ability to bind Rho, Ect2, MgcRacGAP, actin, and MLC II (Piekny and Maddox, 2010;
Kechad et al., 2012). Anillin localizes at the CF during cytokinesis. It has been shown that p190
also binds Rho and Ect2 (Ludwig et al., 2009; Mikawa et al., 2008) and localizes to the CF
during all stage of cytokinesis (Su et al., 2003). Therefore we tested whether these two proteins
interact with each other. Immunoprecipitation of endogenous p190 co-precipitated endogenous
anillin, and vice versa. The interaction was enriched in lysates from cytokinesing cells (Fig. 4B (-
bleb)).
It has been shown that anillin homology domain (AHD) interacts with RhoA (Piekny and
Glotzer, 2008; Zhaho and Fang, 2005) and Cyk4/MgcRacGAP (D’Avino et al., 2008; Gregory et
al., 2008). We determined whether this same region of Anillin could directly interact with p190.
We expressed and purified His-tagged N-terminal regions of p190 (1-763) (which contains the
anillin binding region that we will identify in the next section) and MBP-tagged C-terminal
region of Anillin (608-1087) from bacteria (Fig. 4C). The purified proteins migrated as
monomers by gel filtration suggesting they are properly folded (Fig. S2B). We bound either
MBP or MBPAnillin(608-1087) protein to amylose beads, which were incubated with p190 (1-
763). The beads were washed and the bound proteins detected by immunoblot. p190 (1-763)
was specifically pulled by beads containing MBP-Anillin(608-1087) (Fig. 4C). We estimate that
between 4 and 0.4% of the input p190 (1-763) protein is pulled down suggesting that the direct
interaction is weak and that additional factors or protein modifications are required for the strong
interaction that we detect in IPs.
Anillin localizes p190RhoGAP to the cleavage furrow.
The interaction of p190 with anillin raised the question of whether anillin was required to
localize p190 to the cleavage furrow during cytokinesis. To test this hypothesis HeLa cells were
depleted of anillin by siRNA, and the subcellular location of p190 was assessed by confocal
microscopy. p190 was no longer enriched at the CF following anillin depletion, while
localization of MLC II and actin were unaffected (Fig. S3A, Fig. S3B). However, MLC II
phosphorylation was markedly decreased and Rho was mislocalized (Fig. S3C), consistent with
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

10
published reports (Piekney and Glotzer, 2008). Together these data suggest that p190 and anillin
interact during cytokinesis and that this interaction enriches p190 at the CF.
The interaction between p190RhoGAP and anillin is required for efficient cytokinesis.
We transfected a series of plasmid constructs expressing different regions of p190 into
HeLa cells to map the anillin interacting domain on p190 (Fig. 5A and Fig. S4A). Constructs
expressing the central area of p190 known as the GBD/S1 and MD regions bound endogenous
anillin (Fig. S4B). The common region of these two constructs contains section 1 (S1) of the
middle domain. An isolated S1 domain of p190 also bound anillin after transient transfection,
defining this domain as an interactive region (Fig. S4C). We then generated constructs of full-
length p190 that contained three similarly sized deletions of sequences within section 1 (Fig.
5A). We assayed their ability to interact with endogenous anillin by transient transfection. p190
lacking the C-terminal third of section 1 (ΔS1C) was poorly precipitated by anillin (Fig. 5B).
p190 mutant proteins lacking the middle section (ΔS1B) also did not bind as well as wild type or
protein lacking the first section of section 1 (ΔS1A), suggesting that the C-terminal portion of the
domain as well as some of S1B mediated anillin binding.
To examine whether the interaction between p190 and anillin is critical for proper
cytokinesis, we generated a stable, doxycycline-inducible HeLa cell line that expresses the p190
mutant ΔS1C (Fig. S2C), which is defective in binding anillin (Fig. 5C). This mutant was then
analyzed for its ability to rescue the increase in phosphorylation of MLC II at the CF and the
cytokinesis defect associated with depletion of endogenous p190. Mutant p190 Dox-inducible
HeLa cell lines were treated with p190 siRNA for forty-eight hours to deplete endogenous
protein, and the siRNA-resistant anillin-binding mutant of p190 was induced to endogenous
levels. Cells were then prepared for confocal microscopy analysis for measuring intensity of
pMLC II at the CF and scoring multinucleation. Replacement of endogenous p190 with similar
levels of the anillin-binding mutant of p190 (Fig. 5D) did not reduce MLC II phosphorylation
(Fig. 5E-E’) at the CF and did not rescue the multinucleation phenotype generated by p190
depletion (Fig. 5F), whereas wild type p190 did (Fig. 1C and 2B’). Together, these results
demonstrate that p190 must interact with anillin to regulate proper Rho levels at the CF for
faithful cytokinesis in HeLa cells
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

11
To exclude the possibility that p190ΔS1C was a null mutant we determined if it
maintained two other identified functions of p190. The cell lines expressing the p190 or
p190ΔS1C were treated with doxycycline to express GFP tagged p190 proteins, lysates were
immunoprecipitated with anti-GFP and blotted for the p190 interacting protein, p120 RasGAP
(Bryant et al. 1995). P190 is also an important substrate of the Src tyrosine kinase so we also
immunoblotted the IPs with anti-phosphotyrosine antibodies (Bouton et al. 1991). Figure 5E
shows that immunoprecipitated p190ΔS1C was tyrosine phosphorylated and bound p120
RasGAP to a similar extent as wild type p190. These results indicate that p190ΔS1C protein
retains two traditional p190 functions and is not a null mutant.
p190 and anillin associate during cytokinesis in a contractility-dependent manner.
HeLa cells in cytokinesis were treated for 40 minutes with 50 uM blebbistatin, a
nonmuscle myosin II inhibitor, which produces a state of weak contractility and blocks
cytokinesis (Straight et al., 2003). The association between p190 and anillin was lost after
blebbistatin treatment (Fig. 4B). This result suggests that the interaction between p190 and
anillin is regulated. The finding that blebbistatin blocks CF progression suggests that the
p190/anillin interaction either requires contractile forces or is a late CF event.
To determine whether high levels of blebbistatin affected RhoGTP levels in a manner
similar to p190 depletion, we visualized MLC II phosphorylation by using a anti-phospho-
Thr18/Ser19 (pMLC II) antibody and confocal microscopy. Blebbistatin treatment resulted in an
almost 2-fold increase in the pMLC II/MLC II ratio, indicating an increase in RhoGTP levels
(Fig. 6A-A’). These changes in Rho activity are similar to those seen after depletion of p190
(Fig. 2A-B’).
DISCUSSION
The small GTPase Rho is a critical regulator of cellular contractility. We suggest that
p190 is required to maintain the proper amount of RhoGTP at the cytokinetic furrow in HeLa
cells. Three independent experiments support this conclusions: in vivo silencing of p190 during
cytokinesis (1) increases the amount of RhoGTP, (Fig. 2A; Fig. 2B-B’); (2) increases the
phosphorylation of MLC II at furrows (Fig. 2C-C’); and (3) causes failure of cells to progress to
abscission, ultimately terminating in multinucleation (Fig. 1). We were also able to rescue the
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

12
loss of p190 by adding low doses of blebbistatin, which suggests that cells fail in cytokinesis
because they have too much myosin II activity. Moreover, a p190 GAP point mutant (p190
R1283A) failed to rescue the multinucleation phenotype.
p190 is one of a growing number of RhoGAPs required for cytokinesis.
MgcRacGAP/Cyk4 is required at an early stage to establish a cytokinetic furrow (Minoshima et
al., 2003; Zhao and Fang, 2005). MgcRacGAP has additional roles including the localization of
the RhoGEF Ect2 to furrow, and it is a matter of debate whether MgcRacGAP acts as a RhoGAP
or RacGAP during CF formation (Bastos et al., 2012; Maddox and Oegema, 2003; Glotzer,
2009; Davies and Canman, 2012). In contrast, p190 action does not appear to be critical for
formation of the furrow, suggesting possible temporal separation of these RhoGAPs. It has also
been proposed that MgcRacGAP/Cyk4 functions in late stages of cytokinesis by linking midzone
microtubules to the plasma membrane (Lekomtsev et al., 2012). MP-GAP also limits RhoA
activity throughout mitosis to stabilize the cortex and limit the Rho zone during cytokinesis
(Zanin et al. 2013). Similar to p190 depletion, Zanin et al. found that depletion of MP-GAP
results in partial cytokinesis failure (15-18% of cells). Thus, multiple RhoGAPs are required to
assure faithful completion of cytokinesis and further defining the roles of the three cytokinetic
GAPs is an important line of future experimentation.
We favor models where MgcRacGAP/Cyk4 establishes furrows and p190 functions to
maintain proper forces during contraction. Consistent with this model are our observations that
all cells initiate furrow formation after p190 depletion that higher levels of MLC phosphorylation
are seen at furrows in cells depleted of p190, and these elevated levels could be rescued by
exogenous expression of wild type p190, or low levels of blebbistatin. We also show that an
interaction between p190 and anillin is required for cytokinesis. The critical experiment is the
replacement of endogenous p190 with a mutant p190 that does not bind anillin. These cells were
not able to decrease MLC phosphorylation at the CF and failed cytokinesis, while wild type p190
was able to rescue these phenotypes. In addition, the interaction between p190 and anillin is
inhibited by blebbistatin, which suggests that contractile forces regulate the action of p190 at the
furrow. Our current experiments could not measure a significant change in contraction rate, and
thus we cannot rule out roles of p190 in abscission. However we prefer a role for p190 in the
furrow, because we measure higher pMLC at furrows in p190 depleted cells.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

13
That RhoGTP levels need to be exquisitely controlled during cytokinesis is underscored,
not only by the identification of three RhoGAPs involved in the process [(MgcRacGAP – (Zhao
and Fang, 2005), MP-GAP (Zanin et al., 2013), and p190RhoGAP (Su et al., 2003)], but also by
our previous finding that levels of p190RhoGAP are reduced approximately 50% in late
cytokinesis (Su et al., 2003) and our current findings that contraction appears to regulate the
proper positioning of p190 (through association with anillin) to reduce levels of activated Rho at
the appropriate intervals. Rho function, like other small GTPases, is highly dynamic, switching
between activated and inactivated states to maintain the proper tension on the myosin-actin
network (Fidyk et al., 2006 Biochemistry 45: 7750-62; Vavylonis et al, 2008 Science 319:97-
100). How the three identified RhoGAPs coordinate with one another and how reduced levels of
p190 and its regulated association with anillin accomplish this, in conjunction with Ect2, is a
major unanswered question requiring further investigation.
Our finding that the interaction between p190 and anillin is inhibited by treatment of the
cells with blebbistatin is an exciting result, but one whose importance is still unclear.
Blebbistatin inhibits the action of class II myosin by inhibiting actin-activated ATPase activity
(Straight et al., 2003). The simplest interpretation of our result is that the interaction between
p190 and anillin is initiated by actin- and myosin-dependent contractility, although we cannot
rule out more complicated interpretations at this time. Our ability to rescue p190 depletion by
partial inhibition of myosin II activity by blebbistatin or Y27632 argues that cells fail cytokinesis
because contractile forces are too high (Fig. 4A, Fig. S2A). Consistent with this interpretation we
see an increase in Rho activity after treatment with higher doses of blebbistatin as measured by
MLC II phosphorylation (Fig. 6A-A’). That MLC II is downstream of p190 is further
emphasized by the fact that the inability to complete cytokinesis due to p190 overexpression (and
decreased RhoGTP levels) (Mikawa et al., 2008; Su et al., 2003; Su et al., 2009) can be rescued
with a constitutively active MLC II.
Figure 7 depicts a speculative model that is consistent with our data. Optimal RhoGTP
levels are ensured by a negative feedback loop induced by the contractility-sensing nature of the
p190-anillin complex. During cytokinesis, RhoGTP is enriched at the CF by specific localization
of Ect2 and MgcRacGAP resulting in activation of ROCK and other effectors of Rho. This in
turn leads to myosin II activation and actin polymerization, resulting in CF contraction. Once
contractility forces reach a threshold, p190 productively associates with anillin at the CF and
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

14
lowers RhoGTP levels. This event in turn results in a decrease in myosin II activation, reduced
actin polymerization, and p190 release, completing the cycle. Since anillin also binds the
RhoGEF Ect2, this model can accommodate reactivation of RhoGTP after release of p190.
Overall, this feedback mechanism would maintain contractility forces at an appropriate level for
completion of cytokinesis. To test this model, it will be important to determine whether and how
contractile forces drive a functional interaction between anillin and p190.
MATERIALS AND METHODS
Cell culture and synchronization:
HeLa cells were obtained from American Tissue Culture Collection (Manassas, VA) and
maintained by serial passage in Dulbecco’s Modified Eagle Medium (DMEM) (Gibco, Carlsbad,
CA) containing 10% fetal calf serum (Gibco) and 1% penicillin/streptomycin (Gibco) at 37ºC in
a 5% CO2, humidified environment. Cells were synchronized for arrest in mitosis by treatment
with 50 ng/mL nocodazole (Cat. Number: M1404, Sigma, St. Louis, MO) for 14-16 hours,
released from the nocodazole block for 40 mins, and harvested by mitotic shake off. Where
indicated, 50µM blebbistatin (Cat. Number: 203390 Calbiochem, San Diego, CA) was added at
the time of nocodazole release. Cells were also synchronized by treatment with 2mM thymidine
for 24 hours, released into fresh media for 12 hours, arrested again in 2mM thymidine for 24
hours, and released for 8-16 hours. 8 hours after release from thymidine, either blebbistatin or
ROCK inhibitor Y27632 was added at the indicated concentrations for an additional 6 hours at
which time cells were fixed for immunofluorescence (IF) analysis.
Mutant constructs:
The p190 deletion mutants were constructed using primers purchased from Integrated
DNA Technologies, Inc. pkH3 triple-HA tagged full-length p190RhoGAP plasmid served as the
template for PCR amplification. Fragments of p190 were amplified and the desired portion of
protein was deleted. Deletion fragments of p190 were replaced using BamH1 and Bmt1
restrictions sites. Amplified deletion mutants were ligated into the pkH3 vector using the T4
DNA Ligase I following standard ligation protocols. All mutants obtained by PCR were
confirmed by sequencing prior to use. Verified plasmids were purified from competent DH5α
E.coli using Maxi-prep (Qiagen) according to manufacturer’s protocol.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

15
Transient transfection:
HeLa cells were transfected with Polyfect Transfection Reagent (Qiagen Inc., Valencia,
CA) according to the manufacturer’s protocol. Plasmids used included the pKH3 plasmid
encoding triple HA-tagged, wild type p190 (gift from Dr. Ian Macara), all mutants of p190
derived from this plasmid, and the GFP plasmid expressing GFP-p190 or GFP-constiutively
active (CA) MLC II (phosphomimetic T18D, S19D) (gift from Dr. Rick Horwitz). Cells were
plated in either 100mm or 6-well dishes (Corning). Mock-transfected cells were treated with
transfection reagents alone. When two or more plasmids were co-transfected, total plasmid levels
were equalized with empty vector. Vector control experiments were performed with empty
pKH3 HA-vectors or empty GFP vectors with equivalent amounts of vector per sample.
siRNA transfection:
Human p190RhoGAP-A was silenced using double-stranded p190 RNAi oligos custom
made by Dharmacon that targeted a unique sequence at the N-terminus of the protein. The
sequence of the double stranded oligos was 5’ AAG AUG CAC AUU GUG GAG CAG 3’.
Human anillin was silenced with the Dharmacon ANLN ON-TARGETplus SMARTpool
(GCAAACAACUAGAAACCAA, GGCGAUGCCUCUUUGAAUA,
GAUCAAGCAUUAGCAGAAA, ACGCAACACUUUUGAAUUA). Glass coverslips were
coated with 1mg/ml poly-L-lysine (Sigma), and cells were treated with p190 oligos (50 pmol)
using the Lipofectamine RNAiMax reagent (Invitrogen), as per the manufacturer’s instructions.
After 24 hrs, cells were synchronized in mitosis by treatment with nocodazole as described
above.
Generation of HeLa Tet-on p190-inducible cell lines and knock down rescue experiments.
The Tet-on expression system obtained from CLONTECH Laboratories, Inc. was used
according to the manufacturer's directions. pcDNA5⁄FRT⁄TO-p190, pcDNA5⁄FRT⁄TO-p190
(R1283A), pcDNA5⁄FRT⁄TO-p190ΔS1C Flp-In™ expression vectors and the Flp recombinase
vector, pOG44, were co-transfected into Flp-In™ HeLa T-REx cells using Lipofectamine 2000
(Invitrogen). Cells were selected in the presence of 0.2mg/ml Hygromycin B and screened for
p190 expression after Dox treatmant. Dox inducible HeLa cell lines were plated at 30% density
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

16
and after 16 hrs, cells were transfected with 75 nM p190 siRNA. 1ug/ml doxycycline was used to
induce expression of full length, the GAP mutant p190 (R1283A) or the anillin binding mutant of
p190. Culture medium was changed after 24 hrs, and the cells were allowed to incubate for 48
hrs post-transfection in the presence of 1ug/ml doxycycline.
RhoGTPase assay:
HeLa cells were treated with control (Luciferase) or p190-siRNA for 24 hrs as described
above, then serum starved for an additional 24 hours. Remaining plates were serum starved for
24 hours, and all plates were then synchronized with nocodazole and released from the block as
described above to capture cells in cytokinesis. Individual plates were then treated with either
10µM LPA (Cat. Number: L7260 Sigma) for 40 mins or 0.5 ug/ml C3 toxin (Cat. Number:
CT04-A Cytoskeleton Inc., Denver, CO) for 2 hours before harvest. RhoGTP levels were
measured at 4°C using the RhoA G-LISA assay from Cytoskeleton Inc. according to
manufacturer’s protocol.
Live-cell imaging and Confocal Microscopy:
To perform live cell imaging, HeLa cells were plated onto poly-L-lysine-coated 1.5
Borosilicate 2-well chambered coverglass dishes (LabTex), and images were acquired using a
20x dry objective lens (numerical aperture = 0.75; Olympus) on a deconvolution microscope
(DeltaVision). Single-plane multipoint acquisitions were captured on a CoolSNAP HQ2 camera
(Photometrics) every 5 minutes within a 37°C chamber. Images of cells treated with control or
p190 siRNAs were collected with identical exposure times and scaled equally. The acquisition
software used was SoftWoRX from Applied Precision.
Cell fixation, Immunofluorescence Microscopy and Quantification:
HeLa cells were grown on poly-L-lysine-coated coverslips, depleted of p190 or anillin,
and synchronized in mitosis as described above for 48 hrs. Cells were fixed for 20 min with 4%
paraformaldehyden and processed for immunofluorescence as described (Bolton et al 2002). For
RhoA immunostaning, cells were fixed with 10% ice cold trichloroacetic acid (TCA) on ice for
15 minutes, than were rinsed three times with 1X PBS containing 30 mM glycine (G-PBS).
(Hayashi et al., 1999; Yonemura et al., 2004)
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

17
Cells were analyzed on a Zeiss Axiovert 200 wide-field fluorescence microscope, fitted
with a confocal scanner using a krypton/argon laser (Perkin Elmer), Hamamatsu EMCCD
camera, a NanoScanZ motor (Prior) and a 63X oil Plan-Apochromat objective. An acousto- optic
tunable filter (AOTF) was used for detection of light at 488, 568 and 647 nm. Photographs were
taken as z-series with 0.4 μm Z- steps. Exposure times were consistent for each channel
throughout individual experiments, and no images were altered after capture. Quantification was
performed using Volocity®5.5 (Perkin Elmer) or ImageJ (NIH) softwares. The areas (region of
interest-ROI) around the CF at two sites were outlined using lasso tool (mask) on a single z-
section to determine the average pixel intensity per square micron, which was then averaged per
cell. This was background corrected by subtracting the average pixel intensity per square micron
measured using the same mask. We measure pixel intensity at regions outside of the CF
similarly. The ratio between these two values was then determined to define the specific
phosphorylation of myosin II light chain using phosphomyosin (Ser19) / (Thr18/Ser19) light
chain II) and Myosin light chain II antibodies (Cell Signaling) (Fig. S1B). Fifteen-thirty cells per
treatment were analyzed for each experiment.
Protein purification, gel filtration and In vitro pull down assay:
6His-tagged p190 (1-763) and MBP-tagged Anillin (608-1087) were bacterially
expressed and purified using amylose beads (NEB) for MBP fusion Anillin and Ni-NTA agarose
beads (Qiagen) for His-tagged p190. The beads were washed, and the 6xHis tagged p190 was
eluted from Ni-NTA agarose with 300 mM imidazole and MBP-tagged Anillin was eluteds with
100mM maltose at 4ºC. Proteins are concentrated and fractionated on the Superdex 200 gel
filtration column in 50 mM Tris (pH=7.6), 150 mM NaCl, 5 mM MgCl2, 1mM DTT buffer.
Protein concentration was determined by Bradford protein assay. To investigate the direct
interaction of the p190 and anillin in vitro binding assay was performed using recombinant 6His-
p190(1-763) and MBP-Anillin(608-1087) proteins in 200 mL 50 mM Tris (pH=7.6), 150 mM
NaCl, 5 mM MgCl2, 0.5% Triton X-100 buffer. 5 ug of p190(1-763) were added to the buffer
containing 5 ug of MBP or MBP-Anillin and proteins were incubated with amylose beads by
rotating at room temperature for 1 hour. Beads where washed 4-5 times with buffer before
adding SDS sample buffer and bound proteins were detected by Western. Blots were probed by
1:250 mouse anti-His (Santa Cruz) and 1:10000 mouse anti-MBP (NEB) antibodies.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

18
Immunoprecipitation:
Cell lysates were prepared as described for Western blotting and incubated with 5 μg of
the indicated immunoprecipitating antibody overnight. After primary antibody incubation,
immunecomplexes were precipitated by protein G agarose (Invitrogen) and separated by
centrifugation at 5,000 rpm for 1 min. After cold PBS washes, SDS sample buffer was added to
each sample, boiled for 5 mins, and subjected to Western blotting analysis, as described above.
ACKNOWLEDGEMENTS
We thank the SJP lab for the p190RhoGAP deletion constructs and Piekny lab for MBP-
Anillin plasmid. We are grateful to J. Pritchard and M. Skalski for technical help and the
Cytokinesis Group for helpful discussions.
The authors declare no potential conflicts of interest with respect to the authorship and/or
publication of this article.
This work was supported by the National Cancer Institute [grant numbers CA39438
(SJP), CA009109 (Institutional T32)], GM063045 for AKM and PTS and UVA Cancer Training
grant [grant number 5 32T CA9109] for AM.
REFERENCES
Aktories, K., Braun, U., Rosener, S., Just, I., and Hall, A. (1989). The rho gene product expressed in E. coli is a substrate of botulinum ADP-ribosyltransferase C3. Biochem Biophys Res Commun 158, 209-213. Bastos, RN., Penate, X., Bates, M., Hammond, D., Barr FA. (2012). CYK4 inhibits Rac1-dependent PAK1 and ARHGEF7 effector pathways during cytokinesis. J Cell Biol. 198(5), 865-80.
Bement, W.M., Miller, A.L., and von Dassow, G. (2006). Rho GTPase activity zones and transient contractile arrays. Bioessays 28, 983-993. Bouton AH, Kanner SB, Vines RR, Wang HC, Gibbs JB, Parsons JT. (1991). Transformation by pp60src or stimulation of cells with epidermal growth factor induces the stable association of tyrosine-phosphorylated cellular proteins with GTPase-activating protein. Mol Cell Biol. 11, 945-53.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

19
Bresnick, A.R. (1999). Molecular mechanisms of nonmuscle myosin-II regulation. Curr Opin Cell Biol 11, 26-33. Bryant SS, Briggs S, Smithgall TE, Martin GA, McCormick F, Chang JH, Parsons SJ, Jove R. (1995). Two SH2 domains of p120 Ras GTPase-activating protein bind synergistically to tyrosine phosphorylated p190 Rho GTPase-activating protein.J Biol Chem. 270, 17947-52. Canman, J. C., Lewellyn, L., Laband, K., Smerdon, S. J., Desai, A., Bowerman, B., Oegema, K. (2008). Inhibition of Rac by the GAP activity of centralspindlin is essential for cytokinesis. Science. 322, 1543-6. Davies, T., and Canman C. J. (2012). Stuck in the middle: Rac, adhesion, and cytokinesis. J. Cell Biol, 198, 769–771 D'Avino P.P., Savoian M.S., Glover D.M. (2004). Mutations in sticky lead to defective organization of the contractile ring during cytokinesis and are enhanced by Rho and suppressed by Rac. J Cell Biol. 166(1), 61-71.
D'Avino, P.P., Savoian, M.S., and Glover, D.M. (2005). Clevage furrow formation and ingression during animal cytokinesis: a microtubule legacy. J Cell Sci 118, 1549-1558. D’Avino PP, Takeda T, Capalbo L, Zhang W, Lilley KS, et al. (2008) Interaction between Anillin and RacGAP50C connects the actomyosin contractile ring with spindle microtubules at the cell division site. J Cell Sci 121, 1151–1158 Gai, M., Camera, P., Dema, A., Bianchi, F., Berto, G., Scarpa, E., Germena, G., and Di Cunto, F. (2011). Citron kinase controls abscission through RhoA and Anillin. Mol Biol Cell. 22(20), 3768-78.
Glotzer, M. (2001). Animal cell cytokinesis. Annu Rev Cell Dev Biol 17, 351-386. Glotzer, M. (2009). Cytokinesis: GAP gap. Curr Biol. 19, R162-5.
Gregory SL, Ebrahimi S, Milverton J, Jones WM, Bejsovec A, et al. (2008) Cell division requires a direct link between microtubule-bound RacGAP and Anillin in the contractile ring. Curr Biol 18, 25–29.
Jaffe, A.B., and Hall, A. (2005). Rho GTPases: biochemistry and biology. Annu Rev Cell Dev Biol 21, 247-269. Jantsch-Plunger, V., Gönczy, P., Romano, A., Schnabel, H., Hamill, D., Schnabel, R., Hyman, AA., Glotzer, M. (2000). CYK-4: A Rho family gtpase activating protein (GAP) required for central spindle formation and cytokinesis. J Cell Biol. 149, 1391-404. Hayashi, K., Yonemura, S., Matsui, T., Tsukita, S. and Tsukita, S.(1999). Immunofluorescence detection of ezrin/radixin/moesin (ERM) proteins with their carboxy-terminal threonine
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

20
phosphorylated in cultured cells and tissues: Application of a novel fixation protocol using trichloroacetic acid (TCA) as a fixative. J. Cell Sci. 112, 1149-1158. Kechad, A., Jananji, S., Ruella, Y., and Hickson, G.R. (2012). Anillin Acts as a Bifunctional Linker Coordinating Midbody Ring Biogenesis during Cytokinesis. Curr Biol. 22, 197-203. Kimura, K., Tsuji, T., Takada, Y., Miki, T., and Narumiya, S. (2000). Accumulation of GTP-bound RhoA during cytokinesis and a critical role of ECT2 in this accumulation. J Biol Chem 275, 17233-17236. Kondo, T., Isoda, R., Uchimura, T., Sugiyama, M., Hamao, K., and Hosoya, H. (2012). Diphosphorylated but not monophosphorylated myosin II regulatory light chain localizes to the midzone without its heavy chain during cytokinesis. Biochem Biophys Res Commun. 417, 686-91.
Lekomtsev, S., Su, K. C., Pye, V. E., Blight, K., Sundaramoorthy, S., Takaki, T., Collinson, L. M., Cherepanov, P., Divecha, N., Petronczki, M. (2012). Centralspindlin links the mitotic spindle to the plasma membrane during cytokinesis. Nature 492, 276-9. Liu, J., Fairn, G.D., Ceccarelli, D.F., Sicheri, F., and Wilde, A. (2012). Clevage Furrow Organization Requires PIP(2)-Mediated Recruitment of Anillin. Curr Biol 22, 64-69. Loria, A., Longhini, K. M., Glotzer, M. (2012). The RhoGAP domain of CYK-4 has an essential role in RhoA activation. Nat Rev Mol Cell Biol. 10, 9-20. Ludwig, K., Manchinelly, S.A., Su, L., Mikawa, M., and Parsons, S.J. (2009). p190RhoGAP-A. UCSD-Nature Molecule Pages. Maddox, A.S., and Burridge, K. (2003). RhoA is required for cortical retraction and rigidity during mitotic cell rounding. J Cell Biol 160, 255-265. Maddox, A.S., and Oegema, K. (2003). Closing the GAP: A Role for a RhoA GAP in Cytokinesis. Molecular Cell 11, 846-848. Manchinelly, S.A., Miller, J.A., Su, L., Miyake, T., Palmer, L., Mikawa, M., and Parsons, S.J. (2010). Mitotic down-regulation of p190RhoGAP is required for the successful completion of cytokinesis. J Biol Chem 285, 26923-26932. Matsumura, F. (2005). Regulation of myosin II during cytokinesis in higher eukaryotes. Trends Cell Biol 15, 371-377. Mikawa, M., Su, L., and Parsons, S.J. (2008). Opposing roles of p190RhoGAP and Ect2 RhoGEF in regulating cytokinesis. Cell Cycle 7, 2003-2012. Miller, A. L., Bement, W. M. (2009). Regulation of cytokinesis by Rho GTPase flux. Nat Cell Biol. 11, 71-7.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

21
Minoshima, Y., Kawashima, T., Hirose, K., Tonozuka, Y., Kawajiri, A., Bao, Y.C., Deng, X., Tatsuka, M., Narumiya, S., May, W.S., Jr., et al. (2003). Phosphorylation by aurora B converts MgcRacGAP to a RhoGAP during cytokinesis. Dev Cell 4, 549-560. Mishima, M., Kaitna, S., Glotzer, M., (2002). Central spindle assembly and cytokinesis require a kinesin-like protein/RhoGAP complex with microtubule bundling activity. Dev Cell. 2, 41-54. Narumiya, S., and Yasuda, S. (2006). Rho GTPases in animal cell mitosis. Curr Opin Cell Biol 18, 199-205. Nishimura, Y., Nakano, K., and Mabuchi, I. (1998). Localization of Rho GTPase in sea urchin eggs. FEBS Lett 441, 121-126. Oegema, K., Savoian, M.S., Mitchison, T.J., and Field, C.M. (2000). Functional analysis of a human homologue of the Drosophila actin binding protein anillin suggests a role in cytokinesis. J Cell Biol 150, 539-552. Piekny, A., Werner, M., and Glotzer, M. (2005). Cytokinesis: welcome to the Rho zone. Trends Cell Biol 15, 651-658. Piekny, A.J., and Glotzer, M. (2008). Anillin is a scaffold protein that links RhoA, actin, and myosin during cytokinesis. Curr Biol 18, 30-36. Piekny, A.J., and Maddox, A.S. (2010). The myriad roles of Anillin during cytokinesis. Semin Cell Dev Biol 21, 881-891. Ridley, A.J., Self, A.J., Kasmi, F., Paterson, H.F., Hall, A., Marshall, C.J., and Ellis, C. (1993). Rho family GTPase activating proteins p190, bcr and rhoGAP show distinct specificities in vitro and in vivo. EMBO J 12, 5151-5160. Somers, W. G., Saint, R. (2003). A RhoGEF and Rho family GTPase-activating protein complex links the contractile ring to cortical microtubules at the onset of cytokinesis. Dev Cell 4, 29-39. Straight, A.F., Cheung, A., Limouze, J., Chen, I., Westwood, N.J., Sellers, J.R., and Mitchison, T.J. (2003). Dissecting temporal and spatial control of cytokinesis with a myosin II Inhibitor. Science 299, 1743-1747. Su, L., Agati, J.M., and Parsons, S.J. (2003). p190RhoGAP is cell cycle regulated and affects cytokinesis. J Cell Biol 163, 571-582. Su, L., Pertz, O., Mikawa, M., Hahn, K., and Parsons, S.J. (2009). p190RhoGAP negatively regulates Rho activity at the clevage furrow of mitotic cells. Exp Cell Res 315, 1347-1359. Suzuki, C., Daigo, Y., Ishikawa, N., Kato, T., Hayama, S., Ito, T., Tsuchiya, E., and Nakamura, Y. (2005). ANLN plays a critical role in human lung carcinogenesis through the activation of
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

22
RHOA and by involvement in the phosphoinositide 3-kinase/AKT pathway. Cancer Res 65, 11314-11325. Takaishi, K., Sasaki, T., Kameyama, T., Tsukita, S., and Takai, Y. (1995). Translocation of activated Rho from the cytoplasm to membrane ruffling area, cell-cell adhesion sites and clevage furrows. Oncogene 11, 39-48. Vavylonis, D., Wu, J.Q., Hao, S., O'Shaughnessy, B., and Pollard, T.D. (2008). Assembly mechanism of the contractile ring for cytokinesis by fission yeast. Science 319, 97-100. Watanabe, S., Ando, Y., Yasuda, S., Hosoya, H., Watanabe, N., Ishizaki, T., and Narumiya, S. (2008). mDia2 induces the actin scaffold for the contractile ring and stabilizes its position during cytokinesis in NIH 3T3 cells. Mol Biol Cell 19, 2328-2338. Wu, X., Sakamoto, T., Zhang, F., Sellers, J.R., and Hammer, J.A., 3rd (2006). In vitro reconstitution of a transport complex containing Rab27a, melanophilin and myosin Va. FEBS Lett 580, 5863-5868. Yoshizaki, H., Ohba, Y., Parrini, M.C., Dulyaninova, N.G., Bresnick, A.R., Mochizuki, N., and Matsuda, M. (2004). Cell type-specific regulation of RhoA activity during cytokinesis. J Biol Chem 279, 44756-44762. Yuce, O., Piekny, A., and Glotzer, M. (2005). An ECT2-centralspindlin complex regulates the localization and function of RhoA. J Cell Biol 170, 571-582. Yonemura, S., Hirao-Minakuchi, K. and Nishimura, Y. (2004). Rho localization in cells and tissues. Exp. Cell Res. 295, 300-314. Zanin, E., Desai, A., Poser, I., Toyoda, Y., Andree, C., Moebius, C., Bickle, M., Conradt, B., Piekny, A. and Oegema, K. (2013). A conserved RhoGTP limits M phase contractility and coordinates with microtubule aster to confine RhoA during cytokinesis. Dev. Cell 26, 496-510
Zhao, W.M., and Fang, G. (2005). MgcRacGAP controls the assembly of the contractile ring and the initiation of cytokinesis. Proc Natl Acad Sci U S A 102, 13158-13163.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

23
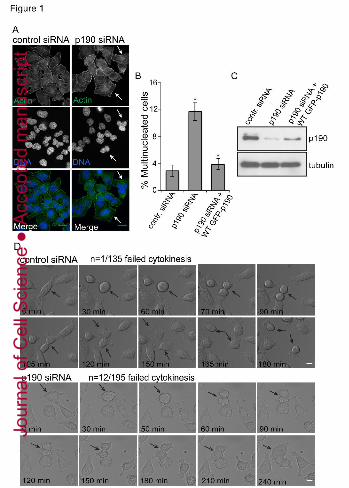
FIGURE LEGENDS Figure 1: p190 is required for cytokinesis.
A: Depletion of p190RhoGAP results in multinucleation because of cytokinesis failure. 48 hrs
after LacZ (control) or p190RhoGAP siRNA treatment HeLa cells were processed for IF for
actin staining with phalloidin (green) and DNA staining with DAPI (blue). Arrows indicate
multinucleated cells. Scale bars, 10 µm. B: Quantification of multinucleation in HeLa cells
depleted of p190 by siRNA. Cells were depleted of p190 and simultaneously were indicated also
treated with 1μg/ml of doxycycline to induce expression of wild type p190. 48 hrs post-
transfection and doxycycline treatment, cells were fixed and stained with phalloidin (actin) or
DAPI (DNA) and scored blindly for bi- or multinucleation (n≥300). Error bars represent standard
deviation from three independent experiments. *=p<0.001 (Student t test (t test)) for both p190
siRNA-treated cells compared to control, and p190 siRNA-treated cells compared to cells
rescued by wild type p190. C: Immunoblot to measure relative p190 levels. D: DIC live-cell
imaging of LacZ or p190 siRNA treated HeLa cells. Recording started after 24 hours transfection
of either siRNA. The top two panels represent LacZ siRNA treated cells, and the bottom two
panels represent p190 siRNA treated cells. Arrows indicate single cell divisions after treatment
with either siRNA. 195 cells treated with p190 siRNA and 135 cells treated with control siRNA
were scored, and the number of multinucleated cells/total number of cells was quantified. Scale
bars, 10 µm.
Figure 2: p190 acts as a RhoGAP in the cytokinesis furrow.
A: p190 depleted cells increase RhoGTP levels in the cytokinetic furrow. RhoGTP levels were
measured with the colorimetric RhoA G-LISA assay. Active RhoGTP protein was used as a
positive control and set at 100%. Results are expressed as the mean percent of RhoGTP +/-
standard error of the mean (SEM), n=4. *=p<0.05 (t test) as compared to cytokinetic cells. B:
Confocal images HeLa cells during cytokinesis stained for RhoA and tubulin 48 hrs after
transfection of siRNA targeted to LacZ (control siRNA) or p190RhoGAP. Images shown are
representative of n>20. Scale bars, 7 µm. B’: Quantification of RhoA intensities at the CF n>20,
*=p=0.0002 (t test). C: Confocal images of pMLC II in wild type p190 doxycycline inducible
HeLa cells during cytokinesis 48 hrs after control (LacZ), p190RhoGAP siRNA and p190 siRNA
with doxycycline treatment. Fixed cells were stained with anti-pMLC II (S19) antibody. Images
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

24
shown are representative of n>20. Scale bars, 4.5 µm. C’: Quantitation of pMLC at the CF. Box
and whisker plot quantifying the ratio of intensity of pMLC II at the CF and outside region of
CF. n>2 with >20 cells/experiment. *, ** =p<0.005 as compared to LacZ siRNA treated cells.
D: A GAP mutant of p190 does not rescue p190-silencing-induced multinucleation. Cells were
depleted of p190 and simultaneously were treated with 1μg/ml of doxycycline to re-express
doxycycline-inducible GAP mutant of p190. Failed cytokinesis was scored as the percentage of
bi- or multinucleated interphase cells (n≥300). Error bars represent standard deviation from three
independent experiments. *=p<0.0001 for both p190 siRNA-treated cells and cells rescued by
p190 GAP mutant (p190 R1283A) compared to control. Note that the experiment in Figure 1B
was performed under identical conditions and at the same time and acts as a control for rescue
with wild type protein. E: The GAP activity of p190 is required for cytokinesis. HeLa cells were
depleted of endogenous p190 and a GAP mutant of p190 (p190 R1283A) was expressed by the
addition of doxycyclin where indicated. Cell lysates were prepared for immunoblot analysis 48
hours post-transfection.
Figure 3: Overexpression of p190 increases RhoGTP levels.
A: Confocal images of total-MLC II and pMLC II in HeLa cells during cytokinesis. HeLa cells
were mock-treated or transiently transfected with empty plasmid or p190 plasmid. Separate
cultures were treated with anillin siRNA or C3 toxin. Cells were then fixed and stained for total
MLC II (red), pMLC II (Thr18/Ser19) (green), and DNA (blue). Images shown are
representative of n>30. Scale bars, 5 µm. B: Quantification of the number of HeLa cells with
enriched pMLC II at the CF during cytokinesis. Results are expressed as the mean percent of
total cells in cytokinesis or cytokinetic cells positive for GFP-p190 exhibiting localization of
pMLC II at the CF +/- SEM, n>3, with >15 cells/experiment. *=p<0.005 (t-test) as compared to
mock. C: HeLa cells were either Mock-treated or transiently transfected with HA-p190 plasmid.
Cells were synchronized by nocodazole block and release and the levels of total MLC II, pMLC
II, and HA-p190 were assessed by Western blot analysis. D: Overexpression of p190 decreases
Rho activity as measured by pMLC II. Quantification of C. Levels were normalized to actin, and
the ratio of pMLC II/total MLC II was then determined and normalized to the ratio in vector-
transfected (VC) cells, which was set to 1. Results are expressed as the mean fold ratio over that
of VC cells +/- SEM, n>3. *=p<0.05 as compared to VC. E: Constitutively active (CA) MLC II
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

25
rescues multinucleation induced by p190 overexpression. HeLa cells were mock-treated or
transiently transfected with HA-p190 or CA MLC II plasmids singly or in combination and
scored for multinucleation. Results are expressed as the mean percent total cells (Mock) or cells
positive for HA-p190 and or CA MLC II that were multinucleated +/- SEM, n>3, with >90
cells/experiment. *=p<0.005 as compared to mock.
Figure 4: p190 and anillin interact during cytokinesis.
A: p190-dependent multinucleation phenotype was rescued by the myosin II inhibitor
blebbistatin. Failed cytokinesis events were measured by the percentage of bi- or multinucleated
interphase cells relative to the total population of cells (n≥300). Error bars represent standard
deviation from three independent experiments. *=p<0.005 for p190 siRNA treated cells compare
to Luc siRNA treated cells, and *=p<0.03 for p190 siRNA compared to p190 siRNA treated cells
in 0.25μM Blebbistatin.
B: Coimmunoprecipitation of anillin and p190 in asynchronous or cytokinetic HeLa cells. HeLa
cells were either untreated (Asyn) or synchronized with nocodazole (Cyto). Cells were then
washed out of nocodazole and either mock-treated or treated with 50 µM blebbistatin for 40 min
and cell lysates generated for immunoprecipitations (IPs). Western blots shown are
representative of n>3. WCL=Whole cell lysate. C: Diagrams of p190 and Anillin constructs
used for recombinant protein expression. MBP and MBP tagged C-terminal anillin was bound to
amylose beads, which were incubated with equal amount of 6His tagged p190(1-763).
Interaction was detected by Western blotting with anti-His and anti-MBP antibodies.
Figure 5: The interaction between p190 and anillin is necessary for proper cytokinesis.
A: Diagram of the p190RhoGAP mutants: Full-length, ΔS1A (Δ379-467), ΔS1B (Δ467-563),
and ΔS1C (Δ563-646). B: Mapping of anillin binding site in p190. HeLa cells were transiently
transfected with HA-tagged variants of p190 (as described in Panel A) or empty vector,
synchronized with nocodazole, and released into mitosis. Extracts were immunoprecipitated as
described in Experimental Procedures and subjected to Western blotting analysis to detect
association of exogenous HA-p190 with anillin. C: Cells expressing p190ΔS1C poorly interact
with Anillin. Cell lines were treated with doxycycline to express either GFP-p190 or GFP-
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

26
p190ΔS1C, lysates were immunoprecipitated with anti-Anillin and blotted for GFP to detect the
interaction between Anillin and p190.
D-G Cytokinesis is not rescued by expression of the anillin-binding mutant of p190. D. p190
protein levels in cells depleted of p190 or after doxycycline expression of p190ΔS1C. E:
RhoGTP activity was indirectly measured by anti-pMLC II (Ser19) antibody. Images shown are
representative of n>30. Scale bars, 4 µm. E’: Quantification of pMLC II at the cleavage furrow.
Box and whisker plot quantifying the ratio of pMLC II intensity at CF and outside region of CF.
n>15 with >20 cells/experiment. *=p<0.005 as compared to Luc siRNA treated cells (t test), NS
= non significance. F: p190ΔS1C does not rescue the multinucletion induced by depletion of
p190 (n≥300). Error bars represent standard deviation from three independent experiments.
*=p<0.005 (t test) for both p190 siRNA-treated cells and cells rescued by anillin binding mutant
of p190 (p190 ΔS1C) compared to LacZ siRNA-treated cells. G: The p190ΔS1C protein retains
two traditional p190 functions. Cell lines were treated with doxycycline to express GFP tagged
p190 or the Anillin binding mutant of p190 (p190 ΔS1C), lysates were immunoprecipitated with
anti-GFP and blotted for p190 interacting protein p120 and pTyr.
Figure 6: p190 and anillin associate during cytokinesis in a contractility-dependent manner
to regulate MLC II activation.
A: Confocal images of total and pMLC in HeLa cells during cytokinesis. HeLa cells were either
mock-treated or treated with 50µM blebbistatin for 40 min and stained for MLC or pMLC
(Thr18/Ser19). The area outlined in red defines the CF (region of interest -ROI), which was
analyzed for pixel intensity. Images shown are representative of n>30. Scale bars, 5 µm. A’:
Quantitation of specific MLC II activity. The ratio of pMLC II to MLC II intensities was
calculated, and results are expressed as the mean pixel intensity ratio +/- SEM, n>3, with >15
cells/experiment. *=p<0.005 (t test) as compared to mock, which was set to 1.
Figure 7. Model for regulation of RhoGTP through p190RhoGAP/anillin complex.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t