a dichotomous species of codium (bryopsidales, chlorophyta ... · escala = 8 y 4 cm,...
TRANSCRIPT

293CODIUM FRAGILE IS COLONIZING NORTHERN CHILERevista Chilena de Historia Natural77: 293-304, 2004
A dichotomous species of Codium (Bryopsidales, Chlorophyta)is colonizing northern Chile
Una especie dicotómica de Codium (Bryopsidales, Chlorophyta)está colonizando el norte de Chile
ALEJANDRA GONZÁLEZ & BERNABÉ SANTELICES
Departamento de Ecología, Facultad de Ciencias Biológicas, Pontificia Universidad Católica de Chile, Casilla 114-D,Santiago, Chile; e-mail: [email protected]
ABSTRACT
In late 2001 and early 2002, a dichotomous species of Codium appeared colonizing the low intertidal andshallow subtidal bottoms of Caldera Bay, northern Chile (27o03’ S, 70o51’ W). Due to the ecological andeconomic impact the species is having in Caldera Bay and its potential spread along the Chilean coastline, westudied the taxonomic identity of the species and examined its relationships with other dichotomous species ofCodium reported for temperate Pacific South America. Morphological analyses suggest that the seaweedsfrom Caldera Bay belong to Codium fragile (Suringar) Hariot. Not only is there strong agreement in internaland external morphological characters, but among all the species reported for Peru and Chile, this is the onlyone exhibiting utricles with rounded, apiculate tip terminating in a mucron. This species has a broadgeographic distribution in temperate waters. In Chile it was known only from the coasts of Valdivia to theStraits of Magellan (39o48’ S, 73o26’ W to 53o10’ S, 73o49’ W). This is the first record of C. fragile innorthern Chile, and this study discusses several alternative hypotheses for the presence of the species into thisarea. The morphological characteristics of the material collected in Caldera partially agree with diagnosticcharacters known for C. fragile subspecies tasmanicum and C. fragile subspecies tomentosoides. However,the rapid population spread of the species in northern Chile, and recent molecular analysis support theidentification of this form as the invasive C. fragile subspecies tomentosoides.
Key words: Codium, introduced species, Chlorophyta, seaweed, northern Chile.
RESUMEN
A fines de 2001 y principio de 2002 apareció en el norte de Chile una especie del género Codium, demorfología dicotómica, colonizando los niveles intermareales y submareales de la Bahía de Caldera (27o03’ S,70o51’ O). Debido al impacto ecológico y económico que ha provocado esta especie en la Bahía de Caldera ya su dispersión potencial a lo largo de la costa de Chile, estudiamos la identidad taxonómica de la especie,examinando su relación con las especies de Codium de hábito erecto descritas para la costa temperada delPacífico Sudamericano. El análisis morfológico sugiere que el alga encontrada en la Bahía de Calderacorresponde a la especie Codium fragile (Suringar) Hariot. Tanto los caracteres morfológicos internos comoexternos del alga son concordantes con la descripción de la especie, incluyendo utrículos de ápicesredondeados apiculados terminados en un mucron puntiagudo. Esta especie posee una amplia distribucióngeográfica en aguas temperadas. En Chile, es conocida solo para las costas entre Valdivia y Magallanes(39o48’ S, 73o26’ O a 53o10’ S, 73o49’ O). Este es el primer registro de C. fragile en el norte de Chile yademás este estudio discute diversas hipótesis alternativas de introducción de la especie en el área. Lascaracterísticas morfológicas de los materiales colectados en Caldera concuerdan parcialmente con loscaracteres diagnósticos conocidos para C. fragile subespecies tasmanicum y C. fragile subespeciestomentosoides. Sin embargo, la expansión poblacional rápida que la especie está mostrando en el norte deChile junto con análisis moleculares recientes apoyan la identificación de estas poblaciones comopertenecientes a la subespecie tomentosoides.
Palabras clave: Codium, especies introducidas, Chlorophyta, algas, norte de Chile.

294 GONZÁLEZ & SANTELICES
INTRODUCTION
Codium Stackhouse (Bryopsidales, Codiaceae)is one of the most common and widespreadseaweed genera in the world (Silva 1951).There are about 100 described species, growingon rocky or sandy habitats in tropical andtemperate waters (Goff et al. 1992, Silva 1992).External morphology includes individuals thathave either a crustose, cushion-like form thatcovers the substratum (“the prostrate species”)or that attach to the substratum at a singlepoint, with the rest of the plant being erect andwith cylindrical dichotomously branched axes(“the dichotomous species”). Bothmorphological types are composed of a tightlywoven mass of coenocytic medullary filamentssurrounded by a cortex of utricles. The utricles,which are the swollen cortical tips of thefilaments, typically have colourless spine-like,rounded or pointed cell wall projections andcell thickenings, frequently used as taxonomiccharacters (Silva 1955, Burrows 1991).
A total of 14 species of Codium have beendescribed for temperate Pacific South America(see Ramirez & Santelices 1991 for listing). Ofthese, eight species are dichotomous and six areprostrate. Not a single dichotomous species hasbeen reported from northern or central Chile(Arica to Concepción; 18o29’ S, 70o19’ W to36o47’ S, 73o14’ W). The dichotomous C.contractum, C. tomentosum and C. fragile havebeen described for southern Chile (Concepciónto the Straits of Magellan), while C.fernandezianum, C. pocokiae and C. unilateralehave been registered for the Chilean oceanicislands only (Isla de Pascua, Juan Fernández yDesventuradas). On the other hand, C.peruvianum and C. foveolatum have beenascribed to the Peruvian coast only (Paita, Piuraand Lima, and Piura respectively).
In late 2001 and early 2002, a dichotomousspecies of Codium appeared colonizing the lowintertidal and shallow subtidal bottoms ofCaldera Bay in northern Chile (27o03’ S, 70o51’W, E. Martínez personal communication). Bythe end of August of 2003 the species hadalready spread to Obispito, 45 km to the north ofCaldera Bay, and to Huasco, 176 km to the southof Caldera Bay (Neill et al. 2003)1. Many ofthese bays in northern Chile are habitat to a
highly endemic native fauna and flora(Santelices 1980, Meneses & Santelices 2000,Santelices & Meneses 2000), and in recentyears, they have become cultivation grounds foreconomically important seaweeds (e.g.,Gracilaria) and shellfish (e.g., Pacific oysters).
Due to the ecological and economic impactthe species may have in Caldera Bay, and in therest of the northern Chilean coastline, here wereport the taxonomic identity of the species,and discuss its potential means of introductionin this area.
MATERIAL AND METHODS
The algal samples were collected by E.Martínez by SCUBA and skin diving atdifferent depths (2-10 m) in Caldera Bay(27o03’ S, 70o51’ W) between January andApril 2002. In total, 19 specimens wereanalyzed and have been deposited in theherbarium of the Sala de Sistemática, PontificiaUniversidad Católica de Chile (SS/UC nos6911-1 to 6911-16 and 6913-1 to 6913-4). Thelength of erect axes and diameter and numberof dichotomies were used to compare theexternal morphology of this species with that ofother species reported for the region. Size,shape and apical structure of utricles were usedas internal taxonomic characters. Utricles werestained with aniline blue and micrographs weretaken using a Nikon Labophot microscope.
RESULTS
Taxonomic identity at the species level
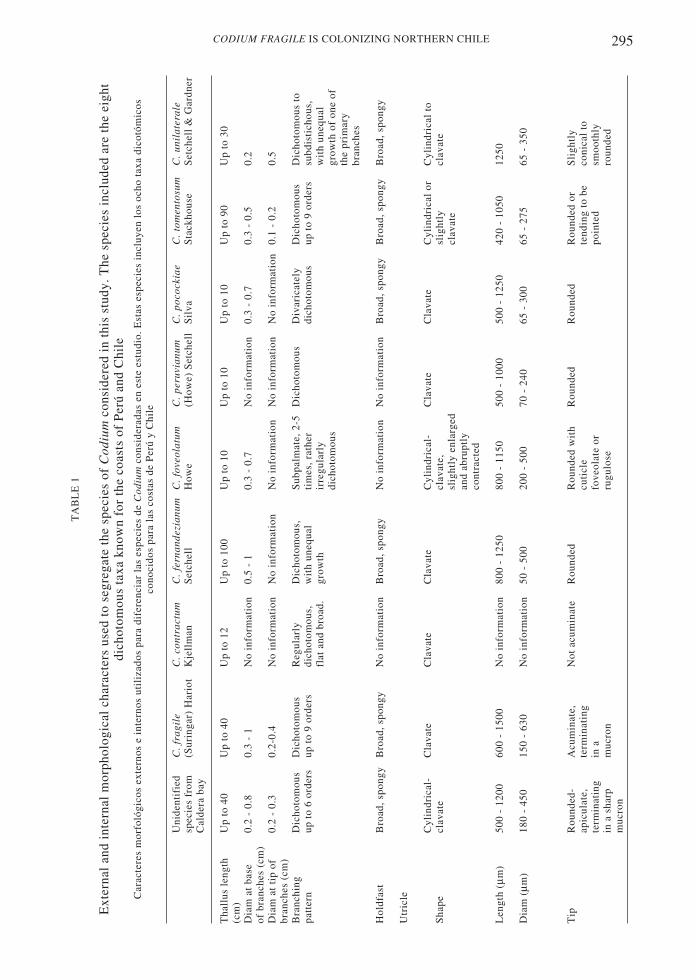
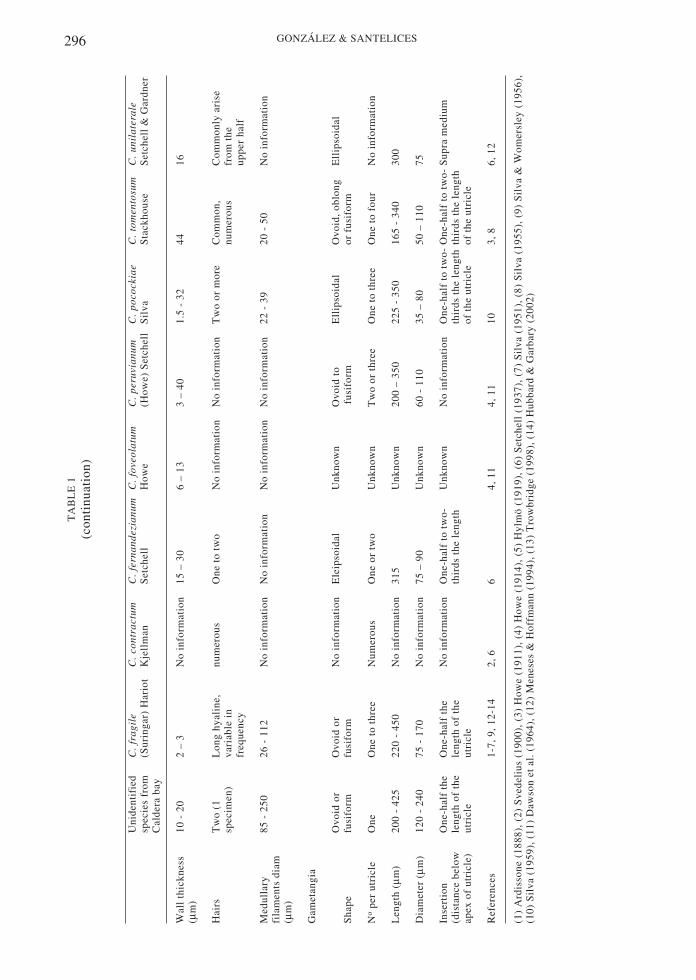
Eleven of the 19 specimens examined exhibitedgametangia, suggesting 57.9 % fertility in thestudied sample. All 19 specimens had similarexternal (Table 1, Fig. 1), and internal (Table 1,Figs. 2 and 3) characters.
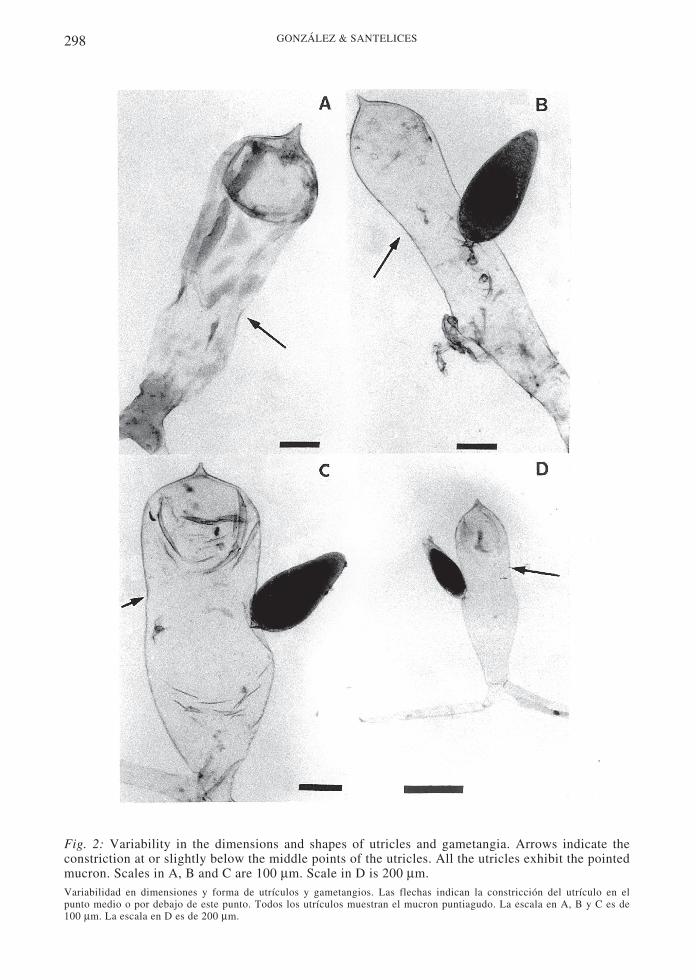
Externally, the plants have severalcylindrical, robust fronds, up to 40 cm long,which arise from a broad, spongy, basal disk(Fig. 1). Branches are blackish green, anddichotomous for up to six orders. Thecylindrical or terete branches are 5 ± 2 mm indiameter at the base, 4 ± 0.5 mm in diameter atthe middle portions and 2.4 ± 0.3 mm indiameter at the tips. The utricles are cylindricalto clavate, frequently with constriction atapproximately around the middle of the utricle(see arrows in Fig. 2). The utricles measure 910± 150 m long and 316 ± 76 µm in diameter.The wall of the utricle is 160 ± 2.6 µm thick,
1 NEILL P, O ALCALDE & J CORREA (2003) Presenciade la especie invasora Codium fragile (Chlorophyta) en elnorte de Chile: efectos potenciales sobre cultivos de Graci-laria chilensis (Rhodophyta). Abstracts of the XLVI Re-unión Anual de la Sociedad de Biología de Chile, XVIReunión Anual de la Sociedad de Ecología de Chile, Puye-hue, Chile: R-56.
295CODIUM FRAGILE IS COLONIZING NORTHERN CHILE
TA
BL
E 1
Ext
erna
l an
d in
tern
al m
orph
olog
ical
cha
ract
ers
used
to
segr
egat
e th
e sp
ecie
s of
Cod
ium
con
side
red
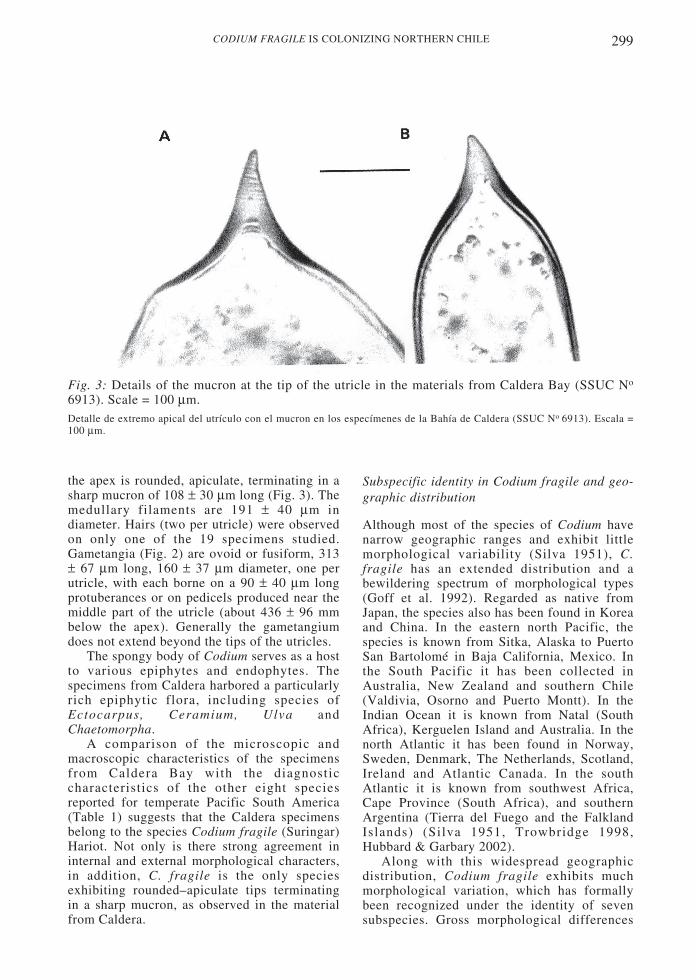
in t
his
stud
y. T
he s
peci
es i
nclu
ded
are
the
eigh
tdi
chot
omou
s ta
xa k
now
n fo
r th
e co
asts
of
Per
ú an
d C
hile
Car
acte
res
mor
foló
gico
s ex
tern
os e
int
erno
s ut
iliz
ados
par
a di
fere
ncia
r la
s es
peci
es d
e C
odiu
m c
onsi
dera
das
en e
ste
estu
dio.
Est
as e
spec
ies
incl
uyen
los
och
o ta
xa d
icot
ómic
osco
noci
dos
para
las
cos
tas
de P
erú
y C
hile
Uni
dent
ifie
dC
. fr
agil
eC
. co
ntra
ctum
C.
fern
ande
zian
umC
. fo
veol
atum
C.
peru
vian
umC
. po
cock
iae
C.
tom
ento
sum
C.
unil
ater
ale
spec
ies
from
(Sur
inga
r) H
ario
tK
jell
man
Set
chel
lH
owe
(How
e) S
etch
ell
Sil
vaS
tack
hous
eS
etch
ell
& G
ardn
erC
alde
ra b
ay
Tha
llus
len
gth
Up
to 4
0U
p to
40
Up
to 1
2U
p to
100
Up
to 1
0U
p to
10
Up
to 1
0U
p to
90
Up
to 3
0(c
m)
Dia
m a
t ba
se0.
2 -
0.8
0.3
- 1
No
info
rmat
ion
0.5
- 1
0.3
- 0.
7N
o in
form
atio
n0.
3 -
0.7
0.3
- 0.
50.
2of
bra
nche
s (c
m)
Dia
m a
t ti
p of
0.2
- 0.
30.
2-0.
4N
o in
form
atio
nN
o in
form
atio
nN
o in
form
atio
nN
o in
form
atio
nN
o in
form
atio
n0.
1 -
0.2
0.5
bran
ches
(cm
)B
ranc
hing
Dic
hoto
mou
sD
icho
tom
ous
Reg
ular
lyD
icho
tom
ous,
Sub
palm
ate,
2-5
Dic
hoto
mou
sD
ivar
icat
ely
Dic
hoto
mou
sD
icho
tom
ous
topa
tter
nup
to
6 or
ders
up t
o 9
orde
rsdi
chot
omou
s,w
ith
uneq
ual
tim
es,
rath
erdi
chot
omou
sup
to
9 or
ders
subd
isti
chou
s,fl
at a
nd b
road
.gr
owth
irre
gula
rly
wit
h un
equa
ldi
chot
omou
sgr
owth
of
one
ofth
e pr
ima r
ybr
a nc h
e s
Hol
dfa s
tB
roa d
, sp
ongy
Bro
ad,
spon
gyN
o in
form
a tio
nB
road
, sp
ongy
No
info
rma t
ion
No
info
rma t
ion
Bro
ad,
spon
gyB
road
, sp
ongy
Bro
ad,
spon
gy
Utr
icle
Cyl
indr
ica l
-C
lava
teC
lava
teC
lava
teC
ylin
dric
a l-
Cla
vate
Cla
vate
Cyl
indr
ica l
or
Cyl
indr
ica l
to
Sha
pec l
a va t
ec l
a va t
e ,sl
ight
lyc l
a va t
esl
ight
ly e
nla r
ged
c la v
a te
a nd
a bru
ptly
c ont
rac t
e d
Len
gth
(µm
)50
0 -
1200
600
- 15
00N
o in
form
a tio
n80
0 -
1250
800
- 11
5050
0 -
1000
500
- 12
5042
0 -
1050
1250
Dia
m (
µm)
180
- 45
015
0 -
630
No
info
rma t
ion
50 -
500
200
- 50
070
- 2
4065
- 3
0065
- 2
7565
- 3
50
Tip
Rou
nded
-A
cum
ina t
e ,N
ot a
c um
ina t
eR
ound
edR
ound
ed w
ith
Rou
nded
Rou
nded
Rou
nded
or
Sli
ghtl
ya p
icul
a te ,
term
ina t
ing
cuti
c le
tend
ing
to b
ec o
nic a
l to
term
ina t
ing
in a
fove
ola t
e or
poin
ted
smoo
thly
in a
sha
rpm
ucro
nru
gulo
sero
unde
dm
ucro
n
296 GONZÁLEZ & SANTELICES
Uni
dent
ifie
dC
. fr
agil
eC
. co
ntra
ctum
C.
fern
ande
zian
umC
. fo
veol
atum
C.
peru
vian
umC
. po
cock
iae
C.
tom
ento
sum
C.
unil
ater
ale
spec
ies
from
(Sur
inga
r) H
ario
tK
jell
man
Set
chel
lH
owe
(How
e) S
etch
ell
Sil
vaS
tack
hous
eS
etch
ell
& G
ardn
erC
alde
ra b
ay
Wal
l th
ickn
ess
10 -
20
2 –
3N
o in
form
atio
n15
– 3
06
– 13
3 –
401.
5 -
3244
16(µ
m)
Hai
rsT
wo
(1L
ong
hyal
ine,
num
erou
sO
ne t
o tw
oN
o in
form
atio
nN
o in
form
atio
nT
wo
or m
ore
Com
mon
,C
omm
only
ari
sesp
ecim
en)
vari
able
in
num
erou
sfr
om t
hefr
eque
ncy
uppe
r ha
lf
Med
ulla
ry85
- 2
5026
- 1
12N
o in
form
atio
nN
o in
form
atio
nN
o in
form
atio
nN
o in
form
atio
n22
- 3
920
- 5
0N
o in
form
atio
nfi
lam
ents
dia
m(µ
m)
Gam
etan
gia
Ovo
id o
rO
void
or
No
info
rmat
ion
Ele
ipso
idal
Unk
now
nO
void
to
Ell
ipso
idal
Ovo
id,
oblo
ngE
llip
soid
alS
hape
fusi
form
fusi
form
fus
ifor
mor
fus
ifor
m
No
per
utri
c le
One
One
to
thre
eN
ume r
ous
One
or
two
Unk
now
nT
wo
or t
hre e
One
to
thre
eO
ne t
o fo
urN
o in
form
a tio
n
Len
gth
(µm
)20
0 -
425
220
- 45
0N
o in
form
a tio
n31
5U
nkno
wn
200
– 35
022
5 -
350
165
- 34
030
0
Dia
me t
e r (
µm)
120
- 24
075
- 1
70N
o in
form
a tio
n75
– 9
0U
nkno
wn
60 -
110
35 –
80
50 –
110
75
Inse
rtio
nO
ne-h
a lf
the
One
-ha l
f th
eN
o in
form
a tio
nO
ne-h
a lf
to t
wo-
Unk
now
nN
o in
form
a tio
nO
ne-h
a lf
to t
wo-
One
-ha l
f to
tw
o-S
upra
med
ium
(dis
tanc
e be
low
leng
th o
f th
ele
ngth
of
the
thir
ds t
he l
e ngt
hth
irds
the
le n
gth
thir
ds t
he l
e ngt
hap
ex o
f ut
ric l
e )ut
ric l
eut
ric l
eof
the
utr
icle
of t
he u
tric
le
Re f
e re n
c es
1-7,
9,
12-1
42,
66
4, 1
14,
11
103,
86,
12
(1)
Ard
isso
ne (
1888
), (
2) S
vede
lius
(19
00),
(3)
How
e (1
911)
, (4
) H
owe
(191
4),
(5)
Hyl
mö
(191
9),
(6)
Se t
c he l
l (1
937)
, (7
) S
ilva
(19
51),
(8)
Sil
va (
1955
), (
9) S
ilva
& W
ome r
sle y
(19
56),
(10)
Sil
va (
1959
), (
11)
Da w
son
e t a
l. (
1964
), (
12)
Men
e se s
& H
offm
ann
(199
4),
(13)
Tro
wbr
idge
(19
98),
(14
) H
ubba
rd &
Ga r
bary
(20
02)
TA
BL
E 1
(con
tinu
atio
n)

297CODIUM FRAGILE IS COLONIZING NORTHERN CHILE
Fig. 1: Specimens of Codium fragile from Caldera Bay. (A) & (B) exhibit the thallus externalmorphology of the species growing on rocky, wave sheltered habitats. Scales = 8 and 4 cm, respec-tively. (C) Elongated herbarium specimen (SSUC Nº 6913) exhibiting the basal disk and the di-chotomously branched erect axes. Scale = 4 cm.Especímenes de Codium fragile recolectados en la Bahía de Caldera. (A) y (B) muestran la morfología externa del talo de laespecie creciendo sobre sustrato rocoso en ambientes protegidos del oleaje. Escala = 8 y 4 cm, respectivamente. (C) Ejemplarde herbario (SSUC Nº 6913) mostrando el disco basal y los ejes erectos con ramificaciones dicotómicas. Escala = 4 cm.
298 GONZÁLEZ & SANTELICES
Fig. 2: Variability in the dimensions and shapes of utricles and gametangia. Arrows indicate theconstriction at or slightly below the middle points of the utricles. All the utricles exhibit the pointedmucron. Scales in A, B and C are 100 µm. Scale in D is 200 µm.Variabilidad en dimensiones y forma de utrículos y gametangios. Las flechas indican la constricción del utrículo en elpunto medio o por debajo de este punto. Todos los utrículos muestran el mucron puntiagudo. La escala en A, B y C es de100 µm. La escala en D es de 200 µm.
299CODIUM FRAGILE IS COLONIZING NORTHERN CHILE
Fig. 3: Details of the mucron at the tip of the utricle in the materials from Caldera Bay (SSUC No
6913). Scale = 100 µm.Detalle de extremo apical del utrículo con el mucron en los especímenes de la Bahía de Caldera (SSUC No 6913). Escala =100 µm.
the apex is rounded, apiculate, terminating in asharp mucron of 108 ± 30 µm long (Fig. 3). Themedullary filaments are 191 ± 40 µm indiameter. Hairs (two per utricle) were observedon only one of the 19 specimens studied.Gametangia (Fig. 2) are ovoid or fusiform, 313± 67 µm long, 160 ± 37 µm diameter, one perutricle, with each borne on a 90 ± 40 µm longprotuberances or on pedicels produced near themiddle part of the utricle (about 436 ± 96 mmbelow the apex). Generally the gametangiumdoes not extend beyond the tips of the utricles.
The spongy body of Codium serves as a hostto various epiphytes and endophytes. Thespecimens from Caldera harbored a particularlyrich epiphytic flora, including species ofEctocarpus, Ceramium, Ulva andChaetomorpha.
A comparison of the microscopic andmacroscopic characteristics of the specimensfrom Caldera Bay with the diagnosticcharacteristics of the other eight speciesreported for temperate Pacific South America(Table 1) suggests that the Caldera specimensbelong to the species Codium fragile (Suringar)Hariot. Not only is there strong agreement ininternal and external morphological characters,in addition, C. fragile is the only speciesexhibiting rounded–apiculate tips terminatingin a sharp mucron, as observed in the materialfrom Caldera.
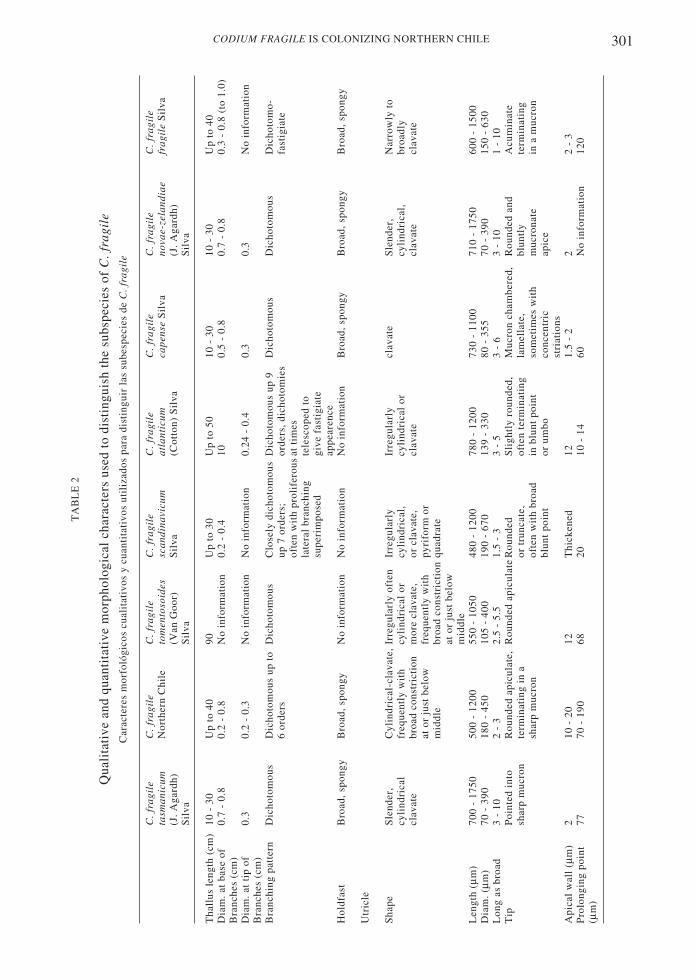
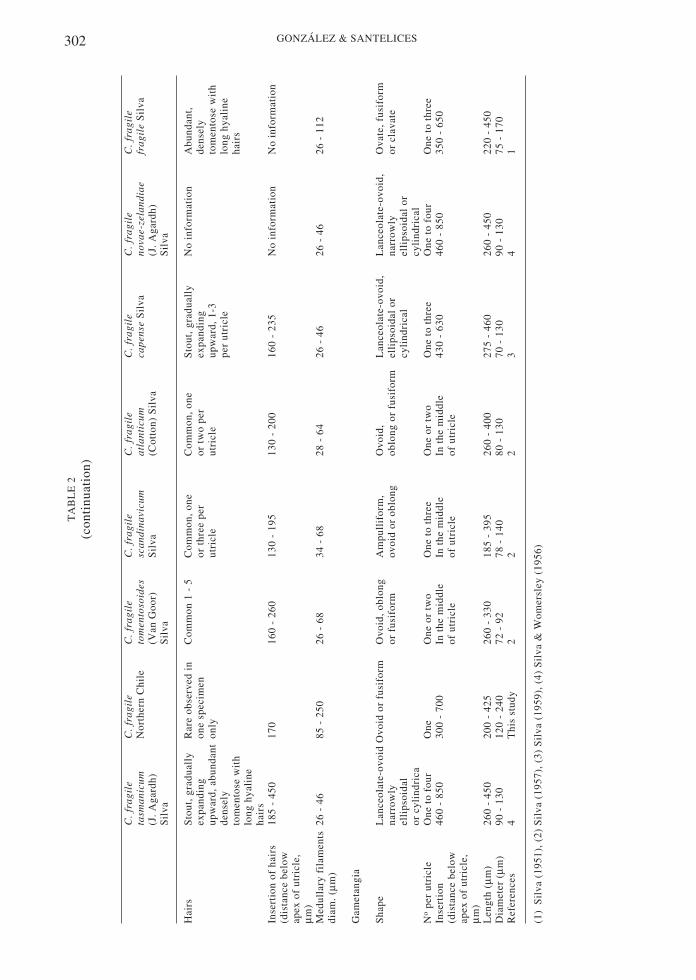
Subspecific identity in Codium fragile and geo-graphic distribution
Although most of the species of Codium havenarrow geographic ranges and exhibit littlemorphological variability (Silva 1951), C.fragile has an extended distribution and abewildering spectrum of morphological types(Goff et al. 1992). Regarded as native fromJapan, the species also has been found in Koreaand China. In the eastern north Pacific, thespecies is known from Sitka, Alaska to PuertoSan Bartolomé in Baja California, Mexico. Inthe South Pacific it has been collected inAustralia, New Zealand and southern Chile(Valdivia, Osorno and Puerto Montt). In theIndian Ocean it is known from Natal (SouthAfrica), Kerguelen Island and Australia. In thenorth Atlantic it has been found in Norway,Sweden, Denmark, The Netherlands, Scotland,Ireland and Atlantic Canada. In the southAtlantic it is known from southwest Africa,Cape Province (South Africa), and southernArgentina (Tierra del Fuego and the FalklandIslands) (Silva 1951, Trowbridge 1998,Hubbard & Garbary 2002).
Along with this widespread geographicdistribution, Codium fragile exhibits muchmorphological variation, which has formallybeen recognized under the identity of sevensubspecies. Gross morphological differences

300 GONZÁLEZ & SANTELICES
among the subspecies of C. fragile are notalways obvious, particularly in areas whereseveral subspecies coexists (Trowbridge 1999).However, the most reliable subspecificcharacters are the size and shape of the utricle,dimensions of the mucron, number of frondsper thallus and branch width. Less reliablediagnostic characters include length andthickness of the thallus and the flatness offronds below the dichotomies (Silva 1951,1957, Silva & Womersley 1956, Trowbridge1996, 1998, Hubbard & Garbary 2002). Itshould be noted, however, that no experimentalstudy has yet evaluated whether the differentmorphologies represent independentsubspecies, or patterns of phenotypic plasticityadapting to local environments (Trowbridge1996, Hubbard & Garbary 2002).
Four of the seven subspecies of Codiumfragile are considered non-invasive and exhibitrestricted geographic distributions. Thus, C.fragile capense occurs on South African shoresonly; C. fragile fragile is found from Sitka,Alaska down to Baja California and the islandsalong that coast (Silva 1951). C. fragile novae-zelandiae is restricted to New Zealand,southeast Australian shores and southernArgentina; while C. fragile tasmanicum isfound on southeast Australian shores(Trowbridge 1996).
The other three subspecies of Codiumfragile are considered invasive. One of them,C. fragile atlanticum , is thought to haveoriginated in Japan and been unintentionallyintroduced with shellfish in southwest Irelandin about 1808 (Silva 1955, Trowbridge & Todd1999, Hubbard & Garbary 2002, Trowbridge2002). Subsequent spreading, most likelythrough rafting or floating, expanded its rangeto now include Norway, the rest of The BritishIsles and eastern Canada (Silva 1955, 1957,Hubbard & Garbary 2002). Codium fragilescandinavicum, was introduced to Denmark in1919, and thereafter spread throughout northernEurope. Now it is found in Sweden and Norwayand other parts of Scandinavia as well asVladivostok, Siberia, and Yokohama (Japan)(Silva 1957). C. fragile tomentosoides is moreor less cosmopolitan throughout temperatewaters. It was first detected in Holland, Europearound 1900, in Denmark in 1919, Sweden in1933, England in 1939, Norway in 1946 andScotland in 1953. The alga rapidly spreadthroughout the Mediterranean during WorldWar II (Silva 1955, 1957, Trowbridge 1999 andreferences therein). C. fragile tomentosoidesnext appeared on northwest Atlantic shores in1957 and is now distributed from North
Carolina to Nova Scotia, Price Edward Island,and eastern Canada (Silva 1957, Carlton &Scanlon 1985, Garbary et al 1997, Garbary &Jess 2000, Hubbard & Garbary 2002). In themid-1970’s, C. fragile tomentosoides appearedin San Francisco, California, on the Pacificcoast of the United States (Coleman 1996). Thefirst appearance of C. fragile tomentosoides inthe Southern Hemisphere was in Aucklandharbor, New Zealand in 1973 (Trowbridge1995, 1996, 1999). In 1995 the alga was firstreported in Australia at Corner Inlet, Victoria,thereafter it has spread some 160 km tosouthwest Australia (Trowbridge 1999 andreferences therein).
The morphological characteristics of thematerial collected in Caldera Bay agree wellwith the diagnostic characters known for C.fragile subspecies tasmanicum and C. fragilesubspecies tomentosoides (Table 2). Bothsubspecies are similar in many external andinternal characters but differ in the presence ofa constriction in the middle parts of the utriclesand in the shape of the tip of the utricle. Thematerial from Caldera Bay resembles C. fragiletomentosoides, having a constriction at or justbelow the middle parts of the utricles (Fig. 2).But they also resembles C. fragile tasmanicumbecause the utricle tips end in a sharp mucron(Fig. 3). The rapid population spread thespecies experiencing in northern Chile addsupport to the identification of this form as theinvasive C. fragile tomentosoides, a conclusionadditionally supported by application ofmolecular techniques to the materials fromnorthern Chile (C. Maggs personalcommunication to Dr. J. Correa). It should bementioned, however, that the Caldera materialsexhibit some morphological variation with C.fragile tomentosoides. The Caldera materialsgenerally lack hairs on the utricles which aredescribed as common in C. fragiletomentosoides. Additional populations shouldbe studied to achieve a more comprehensiveview of the taxonomic importance of thismorphological variation.
DISCUSSION AND CONCLUSIONS
Among the eight dichotomous species ofCodium described for Pacific temperate waters,the materials found in Caldera Bay exhibitmorphological characters typical of the speciesCodium fragile. This finding constitutes a newrecord for the species along the Chilean coast.At a subspecific perspective, the morphologicalcharacters used did not allow discrimination
301CODIUM FRAGILE IS COLONIZING NORTHERN CHILE
TA
BL
E 2
Qua
lita
tive
and
qua
ntit
ativ
e m
orph
olog
ical
cha
ract
ers
used
to
dist
ingu
ish
the
subs
peci
es o
f C
. fr
agil
eC
arac
tere
s m
orfo
lógi
cos
cual
itat
ivos
y c
uant
itat
ivos
uti
liza
dos
para
dis
ting
uir
las
sube
spec
ies
de C
. fr
agil
e
C.
frag
ile
C.
frag
ile
C.
frag
ile
C.
frag
ile
C.
frag
ile
C.
frag
ile
C.
frag
ile
C.
frag
ile
tasm
anic
umN
orth
ern
Chi
leto
men
toso
ides
scan
dina
vicu
mat
lant
icum
cape
nse
Sil
vano
vae-
zela
ndia
efr
agil
e S
ilva
(J.
Aga
rdh)
(Van
Goo
r)S
ilva
(Cot
ton)
Sil
va(J
. A
gard
h)S
ilva
Sil
vaS
ilva
Tha
llus
len
gth
(cm
)10
- 3
0U
p to
40
90U
p to
30
Up
to 5
010
- 3
010
- 3
0U
p to
40
Dia
m.
at b
ase
of0.
7 -
0.8
0.2
- 0.
8N
o in
form
atio
n0.
2 -
0.4
100.
5 -
0.8
0.7
- 0.
80.
3 -
0.8
(to
1.0)
Bra
nche
s (c
m)
Dia
m.
at t
ip o
f0.
30.
2 -
0.3
No
info
rmat
ion
No
info
rmat
ion
0.24
- 0
.40.
30.
3N
o in
form
atio
nB
ranc
hes
(cm
)B
ranc
hing
pat
tern
Dic
hoto
mou
sD
icho
tom
ous
up t
oD
icho
tom
ous
Clo
sely
dic
hoto
mou
sD
icho
tom
ous
up 9
Dic
hoto
mou
sD
icho
tom
ous
Dic
hoto
mo-
6 or
ders
up 7
ord
ers;
orde
rs,
dich
otom
ies
fast
igia
teof
ten
wit
h pr
olif
erou
sat
tim
esla
tera
l br
anch
ing
tele
scop
ed t
osu
peri
mpo
sed
give
fas
tigi
ate
appe
aren
ceH
oldf
a st
Bro
a d,
spon
gyB
road
, sp
ongy
No
info
rma t
ion
No
info
rma t
ion
No
info
rma t
ion
Bro
ad,
spon
gyB
road
, sp
ongy
Bro
ad,
spon
gy
Utr
icle
Sha
peS
lend
e r,
Cyl
indr
ica l
-cla
vate
,Ir
regu
larl
y of
ten
Irre
gula
rly
Irre
gula
rly
c la v
a te
Sle
nde r
,N
a rro
wly
to
c yli
ndri
c al
fre q
uent
ly w
ith
c yli
ndri
c al
orc y
lind
ric a
l,c y
lind
ric a
l or
c yli
ndri
c al,
broa
dly
c la v
a te
broa
d c o
nstr
icti
onm
ore
c la v
a te ,
or c
lava
te,
c la v
a te
c la v
a te
c la v
a te
a t o
r ju
st b
e low
fre q
uent
ly w
ith
pyri
form
or
mid
dle
broa
d c o
nstr
icti
onqu
adra
tea t
or
just
be l
owm
iddl
eL
e ngt
h (µ
m)
700
- 17
5050
0 -
1200
550
- 10
5048
0 -
1200
780
- 12
0073
0 -
1100
710
- 17
5060
0 -
1500
Dia
m.
(µm
)70
- 3
9018
0 -
450
105
- 40
019
0 -
670
139
- 33
080
- 3
5570
- 3
9015
0 -
630
Lon
g a s
bro
a d3
- 10
2 -
32.
5 -
5.5
1.5
- 3
3 -
53
- 6
3 -
101
- 10
Tip
Poi
nte d
int
oR
ound
ed a
pic u
late
,R
ound
ed a
pic u
late
Rou
nded
Sli
ghtl
y ro
unde
d,M
ucro
n c h
ambe
red,
Rou
nded
and
Acu
min
a te
sha r
p m
ucro
nte
rmin
a tin
g in
aor
tru
nca t
e ,of
ten
term
ina t
ing
lam
e lla
te,
blun
tly
term
ina t
ing
sha r
p m
ucro
nof
ten
wit
h br
oad
in b
lunt
poi
ntso
me t
ime s
wit
hm
ucro
nate
in a
muc
ron
blun
t po
int
or u
mbo
conc
e ntr
ica p
ice
stri
a tio
nsA
pic a
l w
a ll
(µm
)2
10 -
20
12T
hic k
e ne d
121.
5 -
22
2 -
3P
rolo
ngin
g po
int
7770
- 1
9068
2010
- 1
460
No
info
rma t
ion
120
(µm
)
302 GONZÁLEZ & SANTELICES
C.
frag
ile
C.
frag
ile
C.
frag
ile
C.
frag
ile
C.
frag
ile
C.
frag
ile
C.
frag
ile
C.
frag
ile
tasm
anic
umN
orth
ern
Chi
leto
men
toso
ides
scan
dina
vicu
mat
lant
icum
cape
nse
Sil
vano
vae-
zela
ndia
efr
agil
e S
ilva
(J.
Aga
rdh)
(Van
Goo
r)S
ilva
(Cot
ton)
Sil
va(J
. A
gard
h)S
ilva
Sil
vaS
ilva
Hai
rsS
tout
, gr
adua
lly
Rar
e ob
serv
ed i
nC
omm
on 1
- 5
Com
mon
, on
eC
omm
on,
one
Sto
ut,
grad
uall
yN
o in
form
atio
nA
bund
ant,
expa
ndin
gon
e sp
ecim
enor
thr
ee p
eror
tw
o pe
rex
pand
ing
dens
ely
upw
ard,
abu
ndan
ton
lyut
ricl
eut
ricl
eup
war
d, 1
-3to
men
tose
wit
hde
nsel
ype
r ut
ricl
elo
ng h
yali
neto
men
tose
wit
hha
irs
long
hya
line
hair
sIn
sert
ion
of h
airs
185
- 45
017
016
0 -
260
130
- 19
513
0 -
200
160
- 23
5N
o in
form
atio
nN
o in
form
atio
n(d
ista
nce
belo
wap
ex o
f ut
ricl
e,µm
)M
edul
lary
fil
amen
ts26
- 4
685
- 2
5026
- 6
834
- 6
828
- 6
426
- 4
626
- 4
626
- 1
12di
am.
(µm
)
Gam
e ta n
gia
Sha
peL
anc e
ola t
e -ov
oid
Ovo
id o
r fu
sifo
rmO
void
, ob
long
Am
pull
ifor
m,
Ovo
id,
Lan
c eol
a te -
ovoi
d,L
anc e
ola t
e -ov
oid,
Ova
te,
fusi
form
narr
owly
or f
usif
orm
ovoi
d or
obl
ong
oblo
ng o
r fu
sifo
rme l
lips
oida
l or
narr
owly
or c
lava
tee l
lips
oida
lc y
lind
ric a
le l
lips
oida
l or
or c
ylin
dric
ac y
lind
ric a
lN
o pe
r ut
ric l
eO
ne t
o fo
urO
neO
ne o
r tw
oO
ne t
o th
ree
One
or
two
One
to
thre
eO
ne t
o fo
urO
ne t
o th
ree
Inse
rtio
n46
0 -
850
300
- 70
0In
the
mid
dle
In t
he m
iddl
eIn
the
mid
dle
430
- 63
046
0 -
850
350
- 65
0(d
ista
nce
belo
wof
utr
icle
of u
tric
leof
utr
icle
a pe x
of
utri
c le ,
µm)
Len
gth
(µm
)26
0 -
450
200
- 42
526
0 -
330
185
- 39
526
0 -
400
275
- 46
026
0 -
450
220
- 45
0D
iam
e te r
(µm
)90
- 1
3012
0 -
240
72 -
92
78 -
140
80 -
130
70 -
130
90 -
130
75 -
170
Re f
e re n
c es
4T
his
stud
y2
22
34
1
(1)
Sil
va (
1951
), (
2) S
ilva
(19
57),
(3)
Sil
va (
1959
), (
4) S
ilva
& W
ome r
sle y
(19
56)
TA
BL
E 2
(con
tinu
atio
n)

303CODIUM FRAGILE IS COLONIZING NORTHERN CHILE
between the subspecies tomentosoides andtasmanicum. It should be noted that somerecent experimental evidence (Trowbridge1996, Hubbard & Garbary 2002) suggests thatsubspecific differences in C. fragile could beattributed in part to environmental factors suchas wave exposure, substrate stability and lightavailability. Future studies with a largernumber of individuals and with additionaltechniques (e.g., molecular markers) may helpto evaluate the validity of the Chilean C. fragilesubspecies. The application of the moleculartechniques (plastid microsatelli tes) addssupport to the identification of this form as theinvasive C. fragile tomentosoides.
One can only speculate about themechanism by which Codium fragile mighthave arrived into Caldera Bay. A firstpossibility would be the transplantation ofshellfish (e.g., Pacific oysters) or seaweed (e.g.,Gracilaria) contaminated with C. fragile eitherfrom southern Chile or abroad (Japan). Thismechanism has been used by several authors toexplain the arrival of C. fragile tomentosoideson the shores of England and Scotland, theMediterranean and Virginia, Maine,Massachusetts, Rhode Island and San Franciscoin the United States (see review by Carlton &Scanlon 1985, Trowbridge 1999).Alternatively, C. fragile may haveinadvertently been introduced via ballast watersor attached to the hull of a ship or pleasurecraft. The Caldera harbor and nearby ports arehome to a commercial fleet that transportsChilean fruit, fishmeal and minerals along theChilean coast and to other countries. A similarmechanism has been used to explain theintroduction of C. fragile on New Zealandshores, because the alga first appeared at thecontainer terminal in the port of Auckland(Trowbridge 1999 and references therein).
As it has occurred in other latitudes, the sizeand abundance of this recent invader of baysand sheltered habitat in northern Chile willprovide a good opportunity to experimentallytest several biological hypotheses related to itsorigin, physiological plasticity and communityeffects. Molecular studies similar to those donewith Caulerpa taxifolia (Vahl) C. Agardh andCodium fragile (Goff et al. 1992, Wiedenmannet al. 2001) should help to trace its geographicorigin and genetic similarity at the intraspecificand interspecific levels. In addition, while inother parts of the world, the economic effectsof the C. fragile invasions have mainly affectedtourism and harbor activities, in northern Chile,it will probably impact more importantly theshellfish and seaweed farming industries.
ACKNOWLEDGMENTS
We are grateful to Drs. Juan Correa, EnriqueMartínez, and Sylvain Faugeron, for thematerials collected and to Paula Neill for thephoto of fresh specimens in Caldera Bay. Thisstudy was supported by FONDECYT (Grant Nº1920855 to B.S.).
LITERATURE CITED
ARDISSONE F (1888) Le alghe della Terre del Fuocoraccolte dal prof Spegazzini. Realle InstitutoLombardo Science e Lettere Rendiconti, ser 2, 21:208-215.
BURROWS EM (1991) Seaweeds of the British Isles.Chlorophyta Natural History Museum, London,United Kingdom. 238 pp.
CARLTON JT & JA SCANLON (1985) Progression anddispersal of an introduced alga: Codium fragile ssptomentosoides (Chlorophyta) on the Atlantic coastof North America. Botanica Marina 28:155-165.
COLEMAN A (1996) DNA analysis methods forrecognizing species invasion: the example ofCodium, and generally applicable methods foralgae. Hydrobiologia 326/327: 29-34.
DAWSON EY, C ACLETO & N FOLDVIK (1964) Theseaweeds of Peru. Nova Hedwigia 13: 1-111.
GARBARY DJ, H VANDERMEULEN & K KIM (1997)Codium fragile ssp tomentosoides (Chlorophyta) in-vades the Gulf of Saint Lawrence, Atlantic Canada.Botanica Marina 40: 537-540.
GARBARY DJ & CB JESS (2000) Current status of theinvasive green alga Codium fragile in eastern Canada.Journal of Phycology (Supplement) 36: 23-24.
GOFF LJ, L LIDDLE, PC SILVA, M VOYTEK & AWCOLEMAN (1992) Tracing species invasion inCodium, a siphonous algae, using molecular tools.American Journal of Botany 79: 1279-1285.
HOWE MA (1911) Phycological studies. V. Some marinealgae of lower California, México. Bulletin of theTorrey Botanical Club 38:489-514, plates 27-34.
HOWE MA (1914) The marine algae of Perú. Memoirs ofthe Torrey Botanical Club 15: 1-185.
HUBBARD CB & DJ GARBARY (2002) Morphologicalvariation of Codium fragile (Chlorophyta) ineastern Canada. Botanica Marina 45: 476-485.
HYLMÖ DE (1919) Zur Kenntniss der subantarktischenund antarktischen Meeresalgen III Chlorophyceen.In: Nordenskjold O (ed) Wissenschaft l icheErgebnisse der schwedischen Sudpolar-Expedition1901-1903, Stockholm, Sweden 4: 1-20, plates 33.
MENESES I & A HOFFMANN (1994) Contribution to themarine algal flora of San Félix Island, DesventuradasArchipelago, Chile. Pacific Science 48: 464-474.
MENESES I & B SANTELICES (2000) Patterns andbreaking points in the distribution of benthic algaealong the temperate Pacific coast of South America.Revista Chilena de Historia Natural 73: 615-623.
RAMÍREZ M & B SANTELICES (1991) Catálogo de lasalgas marinas bentónicas de la costa temperada delPacífico de Sudamérica. Monografías Biológicas,Pontificia Universidad Católica de Chile, Santiago,Chile. 437 pp.
SANTELICES B (1980) Phytogeographic characterizationof the temperate coast of Pacific South America.Phycologia 19:1-12.

304 GONZÁLEZ & SANTELICES
SANTELICES B & I MENESES (2000) A reassessment ofthe phytogeographic characterization of TemperatePacific South America. Revista Chilena de HistoriaNatural 73: 605-614.
SETCHELL WA (1937) The Codiums of the JuanFernández Island. In: Skottsberg C (ed) The naturalhistory of Juan Fernández and Eastern Island 2:587-600, plates 34-48 Uppsala, Sweden.
SILVA P (1951) The genus Codium in California withobservations on the structure of the walls of theutricles. University of California Publications inBotany 25: 79-114.
SILVA P (1955) The dichotomous species of Codium inBritain. Journal of the Marine BiologicalAssociation of the United Kingdom 34: 565-577.
SILVA P (1957) Codium in Scandinavian waters. SvenskBotanisk Tidskrift 51: 117-134.
SILVA P (1959) The genus Codium (Chlorophyta) inSouth Africa. Journal of South African Botany25:103-165.
SILVA P (1992) Geographic patterns of diversity inbenthic marine algae. Pacific Science 46: 429-437.
SILVA P & HBS WOMERSLEY (1956) The genusCodium (Chlorophyta) in southern Australia.Australian Journal of Botany 4: 261-289.
SVEDELIUS N (1900) Algen aus den Landern derMagellansstrasse und Westpatagonien I .Chlorophyceae. In: Nordensjold O (ed)Wissenschaftliche Ergebnisse der schwedischenExpedition nach den Magellanslandern (1895-1897)3: 283-316. Stockholm, Sweden.
TROWBRIDGE CD (1995) Establishment of the greenalga Codium fragile ssp. tomentosoides on New
Associate Editor: Iván GómezReceived April 7, 2003; accepted January 16, 2004
Zealand rocky shores: current distribution andinvertebrate grazers. Journal of Ecology 83: 949-965.
TROWBRIDGE C (1996) Introduction versus nativesubspecies of Codium fragile: how distinctive is theinvasive subspecies tomentosoides. Marine Biology126: 193-204.
TROWBRIDGE C (1998) Ecology of the greenmacroalgae Codium fragile (Suringar) Hariot:invasive and non-invasive subspecies.Oceanography and Marine Biology: an AnnualReview 36: 1-64.
TROWBRIDGE C (1999) An assessment of the potentialspread and options for control of the introducedgreen macroalga Codium fragile ssp. tomentosoideson Australian shores. Codium in Victoria waters.Centre for Research on Introduced Marine Pests,CSIRO, Marine Research. 43 pp.
TROWBRIDGE CD (2002) Coexistence of introduced andnative congeneric algae: Codium fragile and C.tomentosum on Irish rocky intertidal shores. Journalof the Marine Biological Association of the UnitedKingdom 81: 931-937.
TROWBRIDGE CD & C TODD (1999) The familiar isexotic: II Codium fragile ssp tomentosoides onScottish rocky intertidal shores. Botanical Journalof Scotland 50: 161-179.
WIEDENMANN J, A BAUMSTARK, TL PILLEN, AMEINESZ & W VOGEL (2001) DNA fingerprints ofCaulerpa taxifolia provide evidence for theintroduction of an aquarium strain into theMediterranean sea and its close relationship to anAustralian population. Marine Biology 138: 229-234.