a microrna cluster at 14q32 drives aggressive lung...
TRANSCRIPT

Human Cancer Biology
A MicroRNA Cluster at 14q32 Drives Aggressive LungAdenocarcinoma
Ernest Nadal1, Jinjie Zhong1,3, Jules Lin1, Rishindra M. Reddy1, Nithya Ramnath2, Mark B. Orringer1,Andrew C. Chang1, David G. Beer1, and Guoan Chen1
AbstractPurpose:Todeterminewhether different subtypes of lung adenocarcinoma (AC)havedistinctmicroRNA
(miRNA) expression profiles, and to identify miRNAs associated with aggressive subgroups of resected
lung AC.
Experimental Design:miRNA expression profile analysis was performed in 91 resected lung AC and 10
matched nonmalignant lung tissues using a PCR-based array. An independent cohort of 60 lung ACs was
used for validating by quantitative PCR the top 3 prognosticmiRNAs. Gene-expression data from51miRNA
profiled tumors was used for determining transcript-specific miRNA correlations and gene-enrichment
pathway analysis.
Results: Unsupervised hierarchical clustering of 356 miRNAs identified 3 major clusters of lung AC
correlated with stage (P ¼ 0.023), tumor differentiation (P < 0.003), and IASLC histologic subtype of lung
AC (P < 0.005). Patients classified in cluster 3 hadworse survival as compared with the other clusters. Eleven
of 22 miRNAs associated with poor survival were encoded in a large miRNA cluster at 14q32. The top 3
prognostic 14q32miRNAs (miR-411, miR-370, andmiR-376a) were validated in an independent cohort of
60 lung AC. A significant association with cell migration and cell adhesion was found by integrating gene-
expression data with miR-411, miR-370, and miR-376a expression. miR-411 knockdown significantly
reduced cell migration in lung AC cell lines and thismiRNAwas overexpressed in tumors frompatients who
relapsed systemically.
Conclusions:Differentmorphologic subtypes of lung AChave distinctmiRNA expression profiles, and 3
miRNAs encoded at 14q32 (miR-411,miR-370, andmiR-376a)were associatedwithpoor survival after lung
AC resection. Clin Cancer Res; 20(12); 3107–17. �2014 AACR.
IntroductionLung cancer is the leading cause of cancer-related deaths
for both sexes in industrialized countries (1, 2). Adenocar-cinoma (AC) is the most common histologic subtype,accounting for about 40% of lung cancer diagnoses and65,000 deaths each year in the United States. Lung AC is aheterogeneous disease and includes tumors with remark-ably diverse clinical, pathologic, and molecular features. Anew multidisciplinary lung AC classification has beenrecommended based on histopathology as well as clinical,
radiological, and molecular features (3). According to thepredominant histologic pattern, lung AC can be furtherclassified in differentiated (comprising lepidic, invasivemucinous AC, acinar, papillary, and micropapillary) andundifferentiated/solid subtypes.
During the last decade, significant advances in under-standing the critical molecular mechanisms and thetumor heterogeneity in lung AC have provided clinicallyrelevant biomarkers that stratify patients according totheir outcome. In addition, these biomarkers have con-tributed to the development of novel therapeutic strate-gies by identifying new targets as well as predictive mar-kers for specific drugs (4). Analyses using mRNA genomicprofiling from large cohorts of lung ACs have also pro-vided significant information complementing histologicevaluation (5, 6).
MicroRNAs (miRNA) are short noncoding RNAs involvedin many developmental processes that can negatively regu-late gene expression by base pairing to a complementarysequence in the 30 untranslated region of a target mRNA,leading to translational repression. In human cancer, miR-NAs play a pathogenic role in the disease process by acting asoncogenes or tumor suppressor genes (7, 8). Because each
Authors' Affiliations: 1Section of Thoracic Surgery, Department of Sur-gery, University of Michigan Medical School; 2Division of Medical Oncol-ogy, Department of Internal Medicine, University of Michigan, Ann Arbor,Michigan; and 3Xinjiang Medical University, Xinjiang, China
Note: Supplementary data for this article are available at Clinical CancerResearch Online (http://clincancerres.aacrjournals.org/).
Corresponding Authors: Ernest Nadal, University of Michigan, 1500 E.Medical Center Drive, Ann Arbor, MI 48109. E-mail:[email protected]; David G. Beer, [email protected]; and GuoanChen, [email protected]
doi: 10.1158/1078-0432.CCR-13-3348
�2014 American Association for Cancer Research.
ClinicalCancer
Research
www.aacrjournals.org 3107
on May 30, 2018. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 15, 2014; DOI: 10.1158/1078-0432.CCR-13-3348

miRNA can regulate hundreds of targeted genes, miRNAprofiling has been considered superior for classifying cancersubtype, tumor differentiation, or predicting overall survival(OS) compared with expression profiles of protein-codinggenes (9–13). Several studies have identified different miR-NAs associated with lung cancer survival by profiling largesets of non–small cell lung cancer (NSCLC) samples, includ-ing lung AC (14–19).
In this study,we carriedout globalmiRNAprofiling on10nonmalignant lung samples and a cohort of 91 lung ACtumors classified according to the IASLC/ATS/ERS Interna-tional Multidisciplinary Classification of Lung Adenocarci-noma (3) with the aim of identifying relevant miRNAsassociated with survival and with specific morphologicsubtypes of lung AC.
Material and MethodsClinical samples
We used 151 frozen primary tumors and 10 nonma-lignant lung samples matched to the associated tumorfrom patients with lung AC who underwent resection atthe University of Michigan Health System from 1991 to2007. Informed consent was obtained for each subjectand clinical investigations were conducted after approvalby the Institutional Review Board. Tumor specimens wereimmediately frozen following resection and stored at�80�C. Regions containing a minimum of 70% tumorcellularity were used for RNA isolation. Tumor gradeassessment as well as histopathologic analysis of sectionsadjacent to regions used for RNA isolation was performedaccording the IASLC/ATS/ERS International Multidisci-plinary Classification of Lung Adenocarcinoma (3) by 2independent investigators. None of the patients includedin this study received preoperative radiation or chemo-therapy. Clinical data were retrospectively collected fromthe medical records and all cases were staged according to
the revised seventh tumor–node–metastasis classificationcriteria (20). The median follow-up time was 8.12 yearsamong the patients that remained alive. Primary tumorswere randomly assigned to 2 independent sets: trainingand validation set, consisting of 91 and 60 samples,respectively. Patient characteristics are provided in Sup-plementary Table S1.
RNA isolation and miRNA profilingWe profiled 91 lung AC and 10 nonmalignant lung
samples using TaqManOpenArray HumanmicroRNA pan-el (Applied Biosystems), which includes 754miRNAs plus 3controls (U6, RNU44, and RNU48). Details on the RNAextraction, quality control procedures, array preparation,and data normalization are provided in the SupplementaryMaterial.
Validation of miRNA expression by quantitative real-time PCR
Quantitative real-time PCR (qRT-PCR) was performedusing TaqMan microRNA assays (Applied Biosystems) todetermine the expression values of 3 miRNAs (miR-411,miR-370, and miR-376a) in an independent cohort of 60resected lung AC. Details about the qRT-PCR preparationand data normalization are provided in the SupplementaryMaterial. A patient’s risk score was calculated as the sum ofthe expression levels of the 3 prognostic miRNAs in the testset, weighted by the corresponding regression coefficients(b) derived from the Cox regression analysis in the trainingset, as previously reported (21). The risk score was usedto classify patients into high- or low-risk groups, with a highrisk score indicating poorer survival. The distribution ofrisk scores was similar in both sets (Supplementary Fig. S1).In the test set, the median of the risk score was used as thecutoff value.
Lung AC cell lines, transfection, and trans-wellmigration assay
Two lung AC cells endogenously expressing high miR-411 (SK-LU-1 andNCI-H2228)were purchased fromATCC(Manassas) andwere authenticated by genotyping using theIdentifier Plus Kit (Applied Biosystems). These cells weretransfected with miRCURY LNA microRNA power inhibi-tors (Exiqon) either with nontarget control A or antisenseagainst miR-411 using Lipofectamine RNAimax (Invitro-gen), and theirmigration ability was tested by using Boydenchambers (8-mm pore size; BD Biosciences).
Integration of miRNA profile with other genomic dataSNP array data from 216 lung ACs were used for calcu-
lating the copy number of regions encoding selected miR-NAs (22). Available Affymetrix U133A gene expressionmicroarray data from 51 miRNA profiled lung AC tumorswere used from a previous study (5). The original gene setsof embryonic stem cell (ESC), Myc targets, and Notchpathway were obtained from previous publications (23),and average expression of each gene set was calculated foreach tumor. Using a 5% FDR, the correlation between
Translational RelevanceMicroRNAs (miRNA) are small noncoding RNAs
involved in posttranscriptional regulation of geneexpression. Lung cancer miRNA expression profiles haveidentified not only miRNAs differentially expressedamong the distinct histologic subtypes of lung cancer,but also miRNAs that predict prognosis in early-stagenon–small cell lung cancer. In this study, we foundmiRNAs differentially expressed among distinct mor-phologic subtypes of lung adenocarcinoma, which mayhave potential diagnostic utility in the future. In addi-tion, we identified 3 miRNAs encoded at 14q32 region(miR-411, miR-370, and miR-376a) whose expressionwas associated with poor survival, and these miRNAswere validated as independent prognostic markers inpatients with early-stage lung adenocarcinoma. ThesemiRNAs associated with aggressive subtypes of lungadenocarcinomamay be actionable targets in the future.
Nadal et al.
Clin Cancer Res; 20(12) June 15, 2014 Clinical Cancer Research3108
on May 30, 2018. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 15, 2014; DOI: 10.1158/1078-0432.CCR-13-3348

specific miRNAs and mRNA expression was determinedusing 2 approaches: independently of whether they can betargeted by these specificmiRNAs to capture genes that maybe indirectly regulated and restricting the analysis to thepredicted conserved targets downloaded from TargetScanv.6.2 and the Miranda and miRWalk websites (24–26). Toassess biologic processes associatedwith selectedmiRNAs, agene ontology (GO) enrichment analysis was performedbased on significantly correlated genes using DAVID bio-informatics website (27).
Statistical analysisSignificance analysis of microarrays (SAM) algorithms
for paired and unpaired samples were used for identifyingdifferentially expressed miRNAs among tumor and non-malignant samples using 5,000 permutations as previouslydescribed (28). DIANA-miRPath software version 2.0 wasused for pathway enrichment for miRNAs discriminatinglung AC and nonmalignant samples (29). To identifymiRNA expression patterns, an unsupervised hierarchicalcentroid linkage cluster analysis was performed usingCluster v3.0 (30) after mean-centering miRNAs and arraysand heat maps were visualized using the TreeView software(31). Pearson c2 and ANOVA tests were used to determinethe correlation between the clusters and the clinicopath-ologic variables. ANOVA tests were used for comparing themean expression of ESC, Myc, and Notch gene sets amongthe miRNA clusters. A multivariate regression analysisadjusted by tumor grade was performed for miRNAs sig-nificantly up- or downregulated in each specific histologicsubtype for identifying miRNAs differentially expressed insolid, lepidic, and invasive mucinous AC subtypes. Spear-man correlation coefficients and a linear regression anal-ysis, adjusted by gender, were performed to test the asso-ciation between tobacco consumption (measured in packyear) and miRNA expression. Survival curves were plottedusing the Kaplan–Meier method, and survival differenceswere assessed by the log-rank test using the median of eachindividual miRNA as a cutoff. Univariate or multivariateCox proportional hazards were calculated consideringindividual miRNA as a continuous variable. Multivariateanalysis was adjusted by age, gender, and stage. To identifymiRNAs associated with metastatic recurrence, the expres-sion of miRNAs from patients with lung AC who devel-oped metastasis within 5 years of follow-up was comparedwith recurrence-free patients at 5 years.
ResultsIdentification of differentially expressed miRNAs inlung AC versus nonmalignant lungA total of 78miRNAswere differentially expressed in lung
AC as compared with nonmalignant lung tissue by pairedclass-comparison analysis at an FDR of 0.65% (SAM plot isshown in Supplementary Fig. S2). Thirty-seven were foundto be significantly upregulated and 41 downregulated in thetumor tissues (Fig. 1). Using TCGA miRNAseq data from126 lungACversus 78nonmalignant lung samples, 31of 78miRNAs were validated as differentially expressed by class-
Figure 1. Supervised clustering of 78 differentially expressed miRNAsamong 10 lung AC versus 10 matched nonmalignant lung samples.Substantially elevated (yellow) or decreased (blue) expression of themiRNAs is observed for individual tumors.
MicroRNAs at 14q32 Are Prognostic in Lung Adenocarcinoma
www.aacrjournals.org Clin Cancer Res; 20(12) June 15, 2014 3109
on May 30, 2018. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 15, 2014; DOI: 10.1158/1078-0432.CCR-13-3348

comparison analysis in this independent cohort at an FDR<1% (Supplementary Table S2).
The top 10 deregulated miRNAs discriminating lungAC from nonmalignant lung included miR-21, miR-210,miR-183-3p, miR-135b, miR-182 (upregulated) andmiR-144, miR-126, miR30a-3p, miR-195, and miR-145-3p (downregulated). Interestingly, pathway enrich-ment analysis based on the predicted gene targets for thetop 10 discriminating miRNAs yielded pathways relevantin lung cancer biology such as p53, PI3K-Akt, MAPK,TGF-b, WNT, MAPK, and ErbB signaling pathways, aswell as regulation of transcription and posttranslationalprocesses such as ubiquitin-mediated proteolysis (Sup-plementary Fig. S3).
Hierarchical clustering yielded three clusters of lungAC significantly associated with clinicopathologicfeatures and outcome
Unsupervised hierarchical clustering analysis based upon356 expressed miRNAs yielded three major clusters of lungAC, with all nonmalignant lung samples clustering tightlytogether within cluster 1 (Fig. 2A). When nonmalignantsamples were not included in the analysis, we observed thattumors classified in cluster 3 still clustered together and mosttumors overexpressed miRNAs encoded at 14q32 (Supple-mentary Fig. S4). Using unsupervised clustering analysis andthe most variable miRNAs expressed in a large cohort of lungACs from TCGA, a subset of tumors overexpressing 14q32miRNAs was clearly identified (Supplementary Fig. S5).
Cluster 1 Cluster 2 Cluster 3
A
Mucinous
NonmalignantLepidic
PapillaryAcinarSolid
Months after surgery
miR-337miR-127miR-411miR-323-3pmiR-376amiR-410miR-539miR-379cmiR-370miR-487bmiR-409-3pmiR-889miR-485-3pmiR-432miR-758miR-493miR-494miR-655miR-654-3pmiR-409miR-708
DF
S p
rob
abili
ty
B
0 10 20 30 40 50 60
1
0.8
0.6
0.4
0.2
0
Months after surgery0 10 20 30 40 50 60
OS
pro
bab
ility
1
0.8
0.6
0.4
0.2
0
Log-rank P = 0.002
Cluster 3
Cluster 1Cluster 2
Cluster 3
Cluster 1Cluster 2
Log-rank P = 0.001C
Figure 2. Hierarchical clustering ofmiRNA expression in lung ACs. A,three major clusters of tumorswere identified by unsupervisedclustering analysis based on 356miRNAs expressed in 91 tumorsand 10 nonmalignant samples.Samples are depicted in columnsand miRNAs in rows. Lung ACpredominant histologic subtypesof lung AC are displayed bydifferent colors at the top of theheatmap. We highlighted aset of miRNAs noticeablyoverexpressed (yellow) in cluster 3tumors that included severalmiRNAs encoded at the 14q32region. B, the Kaplan–Meier plot ofDFS according to the clustersubgroups. The estimated DFSrate for patients classified incluster 3 was significantly lower (n¼ 34, 32.4% � 0.1%) ascompared with clusters 1 and 2patients (n ¼ 24 and 33, 70.8%� 0.9% and 60.6% � 0.8%,respectively). C, the Kaplan–Meierplot of OS according to the clustersubgroups. The estimated OS ratefor patients classified in cluster 3was significantly lower (n ¼ 34,35.3% � 0.8%) as compared withclusters 1 and 2 patients (n ¼ 24and 33, 75.0%� 0.8% and 63.6%� 0.8%, respectively).
Nadal et al.
Clin Cancer Res; 20(12) June 15, 2014 Clinical Cancer Research3110
on May 30, 2018. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 15, 2014; DOI: 10.1158/1078-0432.CCR-13-3348
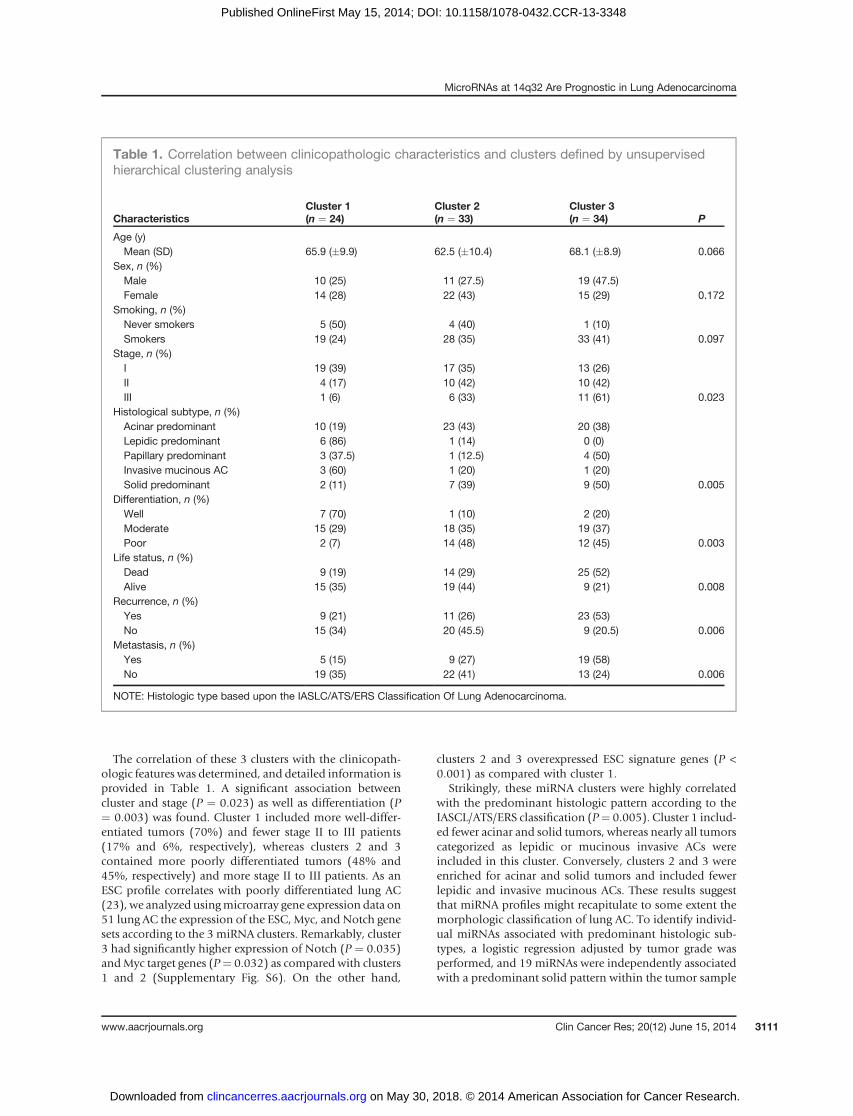
The correlation of these 3 clusters with the clinicopath-ologic features was determined, and detailed information isprovided in Table 1. A significant association betweencluster and stage (P ¼ 0.023) as well as differentiation (P¼ 0.003) was found. Cluster 1 included more well-differ-entiated tumors (70%) and fewer stage II to III patients(17% and 6%, respectively), whereas clusters 2 and 3contained more poorly differentiated tumors (48% and45%, respectively) and more stage II to III patients. As anESC profile correlates with poorly differentiated lung AC(23), we analyzed usingmicroarray gene expression data on51 lung AC the expression of the ESC, Myc, and Notch genesets according to the 3 miRNA clusters. Remarkably, cluster3 had significantly higher expression of Notch (P ¼ 0.035)andMyc target genes (P¼ 0.032) as compared with clusters1 and 2 (Supplementary Fig. S6). On the other hand,
clusters 2 and 3 overexpressed ESC signature genes (P <0.001) as compared with cluster 1.
Strikingly, these miRNA clusters were highly correlatedwith the predominant histologic pattern according to theIASCL/ATS/ERS classification (P¼ 0.005). Cluster 1 includ-ed fewer acinar and solid tumors, whereas nearly all tumorscategorized as lepidic or mucinous invasive ACs wereincluded in this cluster. Conversely, clusters 2 and 3 wereenriched for acinar and solid tumors and included fewerlepidic and invasive mucinous ACs. These results suggestthat miRNA profiles might recapitulate to some extent themorphologic classification of lung AC. To identify individ-ual miRNAs associated with predominant histologic sub-types, a logistic regression adjusted by tumor grade wasperformed, and 19 miRNAs were independently associatedwith a predominant solid pattern within the tumor sample
Table 1. Correlation between clinicopathologic characteristics and clusters defined by unsupervisedhierarchical clustering analysis
CharacteristicsCluster 1(n ¼ 24)
Cluster 2(n ¼ 33)
Cluster 3(n ¼ 34) P
Age (y)Mean (SD) 65.9 (�9.9) 62.5 (�10.4) 68.1 (�8.9) 0.066
Sex, n (%)Male 10 (25) 11 (27.5) 19 (47.5)Female 14 (28) 22 (43) 15 (29) 0.172
Smoking, n (%)Never smokers 5 (50) 4 (40) 1 (10)Smokers 19 (24) 28 (35) 33 (41) 0.097
Stage, n (%)I 19 (39) 17 (35) 13 (26)II 4 (17) 10 (42) 10 (42)III 1 (6) 6 (33) 11 (61) 0.023
Histological subtype, n (%)Acinar predominant 10 (19) 23 (43) 20 (38)Lepidic predominant 6 (86) 1 (14) 0 (0)Papillary predominant 3 (37.5) 1 (12.5) 4 (50)Invasive mucinous AC 3 (60) 1 (20) 1 (20)Solid predominant 2 (11) 7 (39) 9 (50) 0.005
Differentiation, n (%)Well 7 (70) 1 (10) 2 (20)Moderate 15 (29) 18 (35) 19 (37)Poor 2 (7) 14 (48) 12 (45) 0.003
Life status, n (%)Dead 9 (19) 14 (29) 25 (52)Alive 15 (35) 19 (44) 9 (21) 0.008
Recurrence, n (%)Yes 9 (21) 11 (26) 23 (53)No 15 (34) 20 (45.5) 9 (20.5) 0.006
Metastasis, n (%)Yes 5 (15) 9 (27) 19 (58)No 19 (35) 22 (41) 13 (24) 0.006
NOTE: Histologic type based upon the IASLC/ATS/ERS Classification Of Lung Adenocarcinoma.
MicroRNAs at 14q32 Are Prognostic in Lung Adenocarcinoma
www.aacrjournals.org Clin Cancer Res; 20(12) June 15, 2014 3111
on May 30, 2018. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 15, 2014; DOI: 10.1158/1078-0432.CCR-13-3348
and 30 were associated with lepidic pattern or mucinousinvasive AC (Supplementary Table S3).
A borderline association was also found between clusterand patient age (P ¼ 0.066) and between cluster andsmoking history (P ¼ 0.097). Clusters 2 and 3 containedmore heavy smokers than cluster 1. We determined theassociation between miRNA expression and the tobaccopack-year consumed, and 13 miRNAs were significantlyassociated with patient’s smoking intensity: miR-210 waspositively correlated with pack-year, whereas miR-342,miR-151, miR-501-3p, miR-29b, miR-30d, miR-497,miR-222, miR-505, miR-34b, miR-500, and miR-99a-3pwere inversely correlated with pack-year consumed.
Furthermore, we found that the clusters were associatedwith clinical outcome, and patients classified in cluster 3had a significantly worse disease-free survival (DFS) andOS(log-rank test, P¼ 0.002 and 0.001, respectively, Fig. 2B andC) as compared with other patients.
Prognostic miRNAs and validation of the top 3prognostic miRNAs in an independent cohort of lungAC
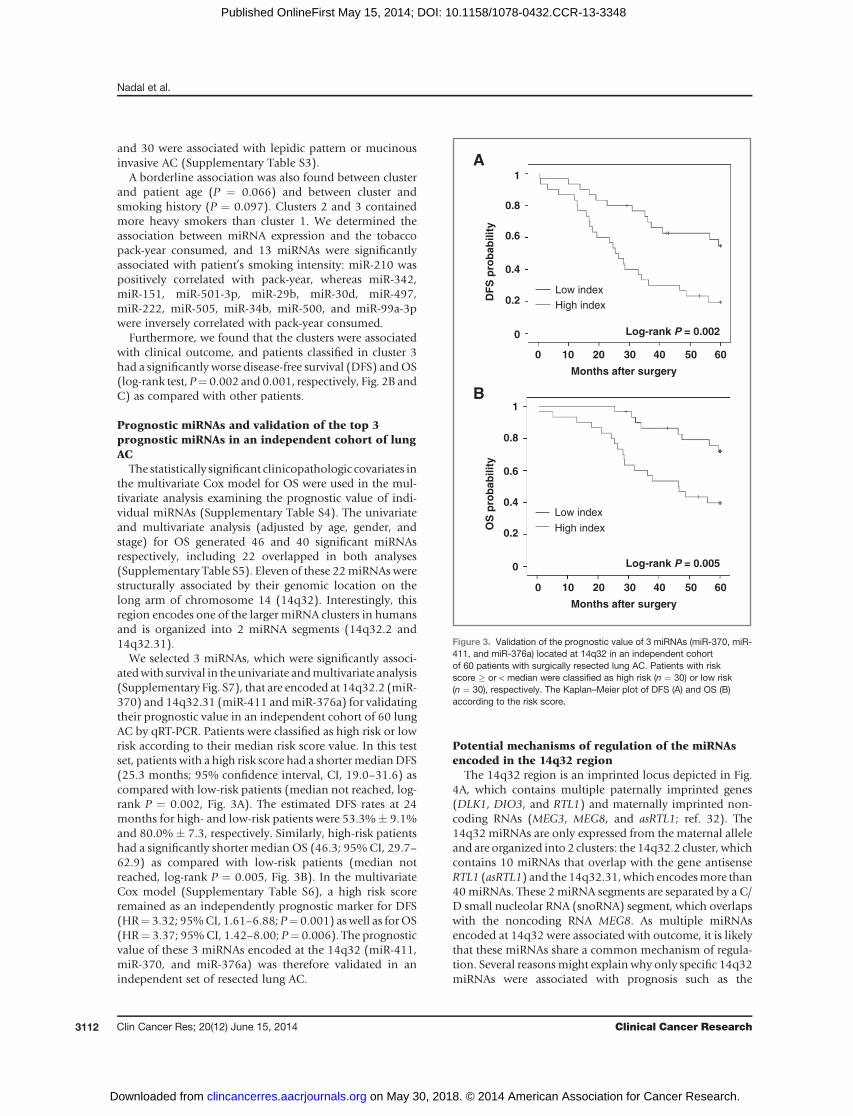
The statistically significant clinicopathologic covariates inthe multivariate Cox model for OS were used in the mul-tivariate analysis examining the prognostic value of indi-vidual miRNAs (Supplementary Table S4). The univariateand multivariate analysis (adjusted by age, gender, andstage) for OS generated 46 and 40 significant miRNAsrespectively, including 22 overlapped in both analyses(Supplementary Table S5). Eleven of these 22miRNAswerestructurally associated by their genomic location on thelong arm of chromosome 14 (14q32). Interestingly, thisregion encodes one of the larger miRNA clusters in humansand is organized into 2 miRNA segments (14q32.2 and14q32.31).
We selected 3 miRNAs, which were significantly associ-atedwith survival in theunivariate andmultivariate analysis(Supplementary Fig. S7), that are encoded at 14q32.2 (miR-370) and 14q32.31 (miR-411 andmiR-376a) for validatingtheir prognostic value in an independent cohort of 60 lungAC by qRT-PCR. Patients were classified as high risk or lowrisk according to their median risk score value. In this testset, patients with a high risk score had a shortermedianDFS(25.3 months; 95% confidence interval, CI, 19.0–31.6) ascompared with low-risk patients (median not reached, log-rank P ¼ 0.002, Fig. 3A). The estimated DFS rates at 24months for high- and low-risk patients were 53.3%� 9.1%and 80.0% � 7.3, respectively. Similarly, high-risk patientshad a significantly shorter median OS (46.3; 95%CI, 29.7–62.9) as compared with low-risk patients (median notreached, log-rank P ¼ 0.005, Fig. 3B). In the multivariateCox model (Supplementary Table S6), a high risk scoreremained as an independently prognostic marker for DFS(HR¼ 3.32; 95%CI, 1.61–6.88; P¼ 0.001) aswell as forOS(HR¼ 3.37; 95%CI, 1.42–8.00; P¼ 0.006). The prognosticvalue of these 3 miRNAs encoded at the 14q32 (miR-411,miR-370, and miR-376a) was therefore validated in anindependent set of resected lung AC.
Potential mechanisms of regulation of the miRNAsencoded in the 14q32 region
The 14q32 region is an imprinted locus depicted in Fig.4A, which contains multiple paternally imprinted genes(DLK1, DIO3, and RTL1) and maternally imprinted non-coding RNAs (MEG3, MEG8, and asRTL1; ref. 32). The14q32 miRNAs are only expressed from the maternal alleleand are organized into 2 clusters: the 14q32.2 cluster, whichcontains 10 miRNAs that overlap with the gene antisenseRTL1 (asRTL1) and the 14q32.31,which encodesmore than40miRNAs. These 2miRNA segments are separated by a C/D small nucleolar RNA (snoRNA) segment, which overlapswith the noncoding RNA MEG8. As multiple miRNAsencoded at 14q32 were associated with outcome, it is likelythat these miRNAs share a common mechanism of regula-tion. Several reasonsmight explain why only specific 14q32miRNAs were associated with prognosis such as the
OS
pro
bab
ility
Months after surgery
DF
S p
rob
abili
ty
B1
0.8
0.6
0.4
0.2
0
0 10 20 30 40 50 60
Months after surgery
0 10 20 30 40 50 60
1
0.8
0.6
0.4
0.2
0
Log-rank P = 0.005
Low indexHigh index
Log-rank P = 0.002
Low index
High index
A
Figure 3. Validation of the prognostic value of 3 miRNAs (miR-370, miR-411, and miR-376a) located at 14q32 in an independent cohortof 60 patients with surgically resected lung AC. Patients with riskscore � or < median were classified as high risk (n ¼ 30) or low risk(n ¼ 30), respectively. The Kaplan–Meier plot of DFS (A) and OS (B)according to the risk score.
Nadal et al.
Clin Cancer Res; 20(12) June 15, 2014 Clinical Cancer Research3112
on May 30, 2018. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 15, 2014; DOI: 10.1158/1078-0432.CCR-13-3348

diversity of target genes regulated by individual miRNAs,differences in the transcript processing or maturation, andthe fact that 14q32 miRNAs are not all consistentlyexpressed in lung AC. Correspondingly, we found that notall the expressed 14q32 miRNAs were significantly corre-lated among them (Supplementary Fig. S8).To examinewhether structural chromosomal changes are
associated with the expression of the miRNAs encoded atthe 14q32 region, we used dense SNP 250K data from 216lung AC (22). The median copy-number value was calcu-lated for the 14q32 region, including both miRNA clusters,and only 1 patient (0.5%) harbored a deletion, whereas 2patients (0.9%) had amplification at this region (Fig. 4B).These results suggest that copy number is not the mainmechanism influencing expression of the miRNAs encoded
at this chromosomal region. Next, we determined thecorrelation of 14q32 miRNAs with other genes encoded at14q32 region using mRNA expression data available fromthe same 51 lungACs included in this study. Themeanof 24expressed miRNAs located at 14q32 was calculated andcorrelated to the expression values of 25 probes correspond-ing to 18 transcripts (coding and noncoding) flanking the14q32 miRNAs. The mean expression of 14q32 miRNAswas significantly correlated to DLK1 and MEG3 expression(P ¼ 0.041 and 0.003, respectively; Fig. 4C). We hypoth-esize that 14q32 miRNAs are epigenetically regulated,because genes and miRNAs encoded within this imprintedlocus might be regulated by DNA methylation of twodifferentially methylated regions located upstream of thetranscription initiation site of MEG3 (32–35).
Figure 4. Regulation of 14q32region. A, genomic map of 14q32miRNA clusters. The paternallyimprinted genes (DLK1, RTL1, andDIO3) are depicted in black and thematernally imprinted noncodingRNAs (MEG3, anti-RTL1, andMEG8) are depicted by diagonallines. The miRNAs at 14q32 regionare organized in 2 segmentsseparated by a C/D snoRNAcluster. The intergenic differentiallymethylated region (IG-DMR) ismethylated in the paternalchromosome. Bold miRNAs wereassociated with poor outcome inthe univariate and multivariateanalysis. B, median copy-numbervalue was calculated using SNP250K data available from 216patients with lung AC for the region100, 445, 178–100, 582, 998.Dashed lines, criteria forconsidering the samples amplified(�3) or deleted (�1). C, scatter plotshowing the significant correlationamong the mean of 14q32miRNAsexpression and MEG3 expression(r ¼ 0.42; n ¼ 51; P ¼ 0.003).
MicroRNAs at 14q32 Are Prognostic in Lung Adenocarcinoma
www.aacrjournals.org Clin Cancer Res; 20(12) June 15, 2014 3113
on May 30, 2018. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 15, 2014; DOI: 10.1158/1078-0432.CCR-13-3348

Genes correlatedwithmiR-370,miR-411, andmiR-376aexpression and GO analysis
We determined the correlation between the expressionof the top 3 prognostic miRNAs (miR-370, miR-411,and miR-376a) and mRNA expression of 22,823 probesin a subset of 51 lung AC, which had both miRNA andmRNA expression data. Using a 5% FDR, we detected asignificant correlation (P < 0.05) among 3,621 uniquemiRNA–mRNA pairs. A gene set enrichment analysiswas performed based upon genes significantly correlatedwith each of the 3 miRNA gene sets, focusing on GOgene set collections. The top 12 GO terms for these 3miRNAs are provided in Supplementary Table S7. Inter-estingly, all 3 miRNAs were significantly associated withregulation of cell migration or cell motion (P < 0.001),whereas miR-370 and miR-411 target genes were alsoassociated with cell adhesion (P � 0.001). In addition,the predicted genes targeted by miR-370, miR-411, andmiR-376a were downloaded from TargetScan v6.2,Miranda, and mirWalk to determine whether the miR-correlated genes are potential direct miRNA targets.When the gene enrichment analysis was restricted togenes that are predicted targets and significantly corre-lated with these miRNAs (959 miRNA–mRNA pairs), thetop terms were involved in the regulation of cell migra-tion and motility as well.
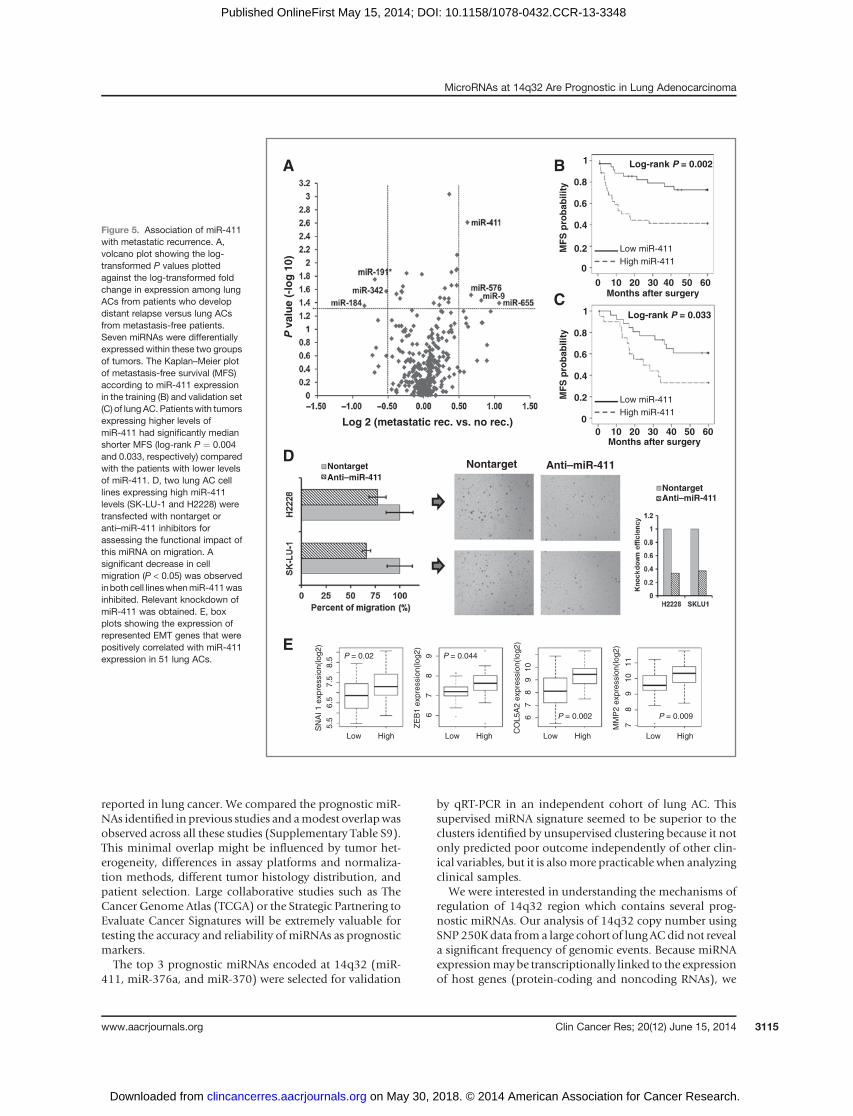
miR-411 expression is associated with metastaticrelapse and EMT genes
As regulation of genes involved in cell motility and celladhesion can contribute to the acquisition of a more met-astatic phenotype, we examined which miRNAs were dif-ferentially expressed in tumors from patients who devel-oped distant relapse as compared with metastasis-freepatients during the follow-up. Strikingly, miR-411 wasidentified among the 7 miRNAs differentially expressed intumors from patients with metastatic disease progression(Fig. 5A). Accordingly, patientswithhighermiR-411 expres-sion had a significantly shorter median metastasis-freesurvival as compared with patients expressing low levels ofmiR-411 in both the training and in the validation set (Fig.5B and C). Next, we determined the functional impact ofmiR-411 knockdown on migration by transfecting 2 lungAC cell lines (SK-LU-1 and NCI-H2228), which endoge-nously overexpress miR-411. A significant reduction in cellmigration was observed in SK-LU-1 and NCI-H2228 cellstransfected with an anti–miR-411 as compared with non-target (Fig. 5D). Interestingly, a significant number of genesoverexpressed during epithelial–mesenchymal transition(EMT; ref. 36) were found positively correlated with miR-411 expression, such as ZEB1, SNAI1, SNAI2, MMP2,COL5A2, or SPARC (Fig. 5E, Supplementary Table S8).
DiscussionmiRNA expression is deregulated in human cancers,
including lung cancer, and contributes to cancer initiationand progression by targeting genes involved in biologic
processes that are hallmarks of cancer such as sustainedproliferative signaling, resisting cell death, activating cellmigration and invasion, inducing angiogenesis, and avoid-ing immunedestructionor deregulating cell energetics (37).The investigation ofmiRNAs associated with amore aggres-sive behavior in lung cancer may provide prognostic anddisease monitoring markers and potentially define noveltherapeutic targets.
In this study, we identified 78 miRNAs differentiallyexpressed in lung AC comparing with nonmalignant lungtissue. Interestingly, 7 of these deregulated miRNAs werefound differentially expressed in lung squamous cell carci-noma (SCC) versus nonmalignant lung in our previousmiRNA study (11): miR-210, miR-200c, miR-200a, miR-106b, miR-182, and miR-183 were upregulated, whereaslet-7e was downregulated. Previous studies demonstratedthat miRNA profiles can discriminate the main histologicsubtypes of lung cancer, small cell lung cancer, and NSCLCas well as between lung AC and SCC (14, 16, 38). However,it is unknown whether different morphologic and molec-ular subtypes of lung AC have specific miRNA expressionprofiles. We identified 3 major clusters of lung AC byunsupervised hierarchical clustering analysis based on theexpression of 356miRNAs. These clusters were significantlyassociated not only with tumor differentiation grade anddisease stage, but also with the predominant morphologicsubtype of lung AC according to the IASLC/ATS/ERS Clas-sification. Cluster 1 was significantly enriched for lepidicandmucinous invasive tumors, whereas cluster 3 containedmore solid lung AC. These results suggest that miRNAexpression profiles may recapitulate to some extent thesignificant heterogeneity of lung AC, and specific miRNAsare dysregulated in distinct morphologic types of lung AC.Themorphologic subtypes are indicative of biologic behav-ior and therefore, miRNA expression profiles, which reflectthe morphologic subtypes of lung AC, may be potentialsurrogate markers of the underlying lung cancer and havepotential clinical utility as diagnostic tools in the future.Further prospective studies will be needed for validatingthese findings. Of note, differential miRNA expressionprofiles have been reported to be reflective of distincthistologic subtypes of breast or ovarian tumors as well(39, 40).
When we examined the effect of tobacco smoking onmiRNA expression, we found several miRNAs were signif-icantly correlated with the amount of tobacco consumed.miR-210, whichwas highly expressed in heavy smokers, hasbeen associated with hypoxia in lung cancer and positivelyregulates HIF-1a (41). Certain miRNAs negatively correlat-ed with smoking status (such as miR-342-5p andmiR-34b)may act as tumor suppressor miRNAs, and their expressionmay be regulated by epigenetic mechanisms (42).
In the survival analysis, we identified a set of overex-pressed miRNAs associated with poor outcome after lungAC resection, which were encoded in the long arm ofchromosome 14 (14q32). Overexpression of 14q32 miR-NAs has been also associated with poor outcome in othersolid tumors (43, 44), but it has not been previously
Nadal et al.
Clin Cancer Res; 20(12) June 15, 2014 Clinical Cancer Research3114
on May 30, 2018. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 15, 2014; DOI: 10.1158/1078-0432.CCR-13-3348
reported in lung cancer. We compared the prognostic miR-NAs identified in previous studies and amodest overlapwasobserved across all these studies (Supplementary Table S9).This minimal overlap might be influenced by tumor het-erogeneity, differences in assay platforms and normaliza-tion methods, different tumor histology distribution, andpatient selection. Large collaborative studies such as TheCancer Genome Atlas (TCGA) or the Strategic Partnering toEvaluate Cancer Signatures will be extremely valuable fortesting the accuracy and reliability of miRNAs as prognosticmarkers.The top 3 prognostic miRNAs encoded at 14q32 (miR-
411, miR-376a, and miR-370) were selected for validation
by qRT-PCR in an independent cohort of lung AC. Thissupervised miRNA signature seemed to be superior to theclusters identified by unsupervised clustering because it notonly predicted poor outcome independently of other clin-ical variables, but it is alsomore practicable when analyzingclinical samples.
We were interested in understanding the mechanisms ofregulation of 14q32 region which contains several prog-nostic miRNAs. Our analysis of 14q32 copy number usingSNP 250Kdata from a large cohort of lung ACdid not reveala significant frequency of genomic events. Because miRNAexpressionmaybe transcriptionally linked to the expressionof host genes (protein-coding and noncoding RNAs), we
A
Log 2 (metastatic rec. vs. no rec.)
P v
alu
e (-
log
10)
Nontarget Anti–miR-411D
MF
S p
rob
abili
ty
0 10 20 30 40 50 60
1
0.8
0.6
0.4
0.2
0
Months after surgery
MF
S p
rob
abili
ty
0 10 20 30 40 50 60
1
0.8
0.6
0.4
0.2
0
Months after surgery
C
B
Low High Low High Low High
P = 0.02 P = 0.044
P = 0.002 P = 0.009
Low High
E
Low miR-411High miR-411
Log-rank P = 0.033
Log-rank P = 0.002
Low miR-411High miR-411
5.5
6.5
7.5
8.5
67
89
67
89
10
78
910
11
SN
AI 1
exp
ress
ion(
log2
)
ZE
B1
expr
essi
on(lo
g2)
CO
L5A
2 ex
pres
sion
(log2
)
MM
P2
expr
essi
on(lo
g2)
Nontarget Anti–miR-411
Nontarget Anti–miR-411
Figure 5. Association of miR-411with metastatic recurrence. A,volcano plot showing the log-transformed P values plottedagainst the log-transformed foldchange in expression among lungACs from patients who developdistant relapse versus lung ACsfrom metastasis-free patients.Seven miRNAs were differentiallyexpressed within these two groupsof tumors. The Kaplan–Meier plotof metastasis-free survival (MFS)according to miR-411 expressionin the training (B) and validation set(C) of lung AC. Patientswith tumorsexpressing higher levels ofmiR-411 had significantly medianshorter MFS (log-rank P ¼ 0.004and 0.033, respectively) comparedwith the patients with lower levelsof miR-411. D, two lung AC celllines expressing high miR-411levels (SK-LU-1 and H2228) weretransfected with nontarget oranti–miR-411 inhibitors forassessing the functional impact ofthis miRNA on migration. Asignificant decrease in cellmigration (P < 0.05) was observedinboth cell lineswhenmiR-411wasinhibited. Relevant knockdown ofmiR-411 was obtained. E, boxplots showing the expression ofrepresented EMT genes that werepositively correlated with miR-411expression in 51 lung ACs.
MicroRNAs at 14q32 Are Prognostic in Lung Adenocarcinoma
www.aacrjournals.org Clin Cancer Res; 20(12) June 15, 2014 3115
on May 30, 2018. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 15, 2014; DOI: 10.1158/1078-0432.CCR-13-3348

determined the correlation of miRNAs encoded at 14q32with other genes located in this region. We found thatexpression of 14q32 miRNAs correlated with two genes(DLK1 and MEG3) located in an imprinted locus, which isregulated by DNA methylation and histone modifications.We hypothesize that 14q32 miRNAs may also be epigenet-ically regulated but further studies are needed to elucidatethe mechanism that controls the expression of this miRNAcluster.miRNAs encoded at 14q32 regionplay an importantrole in stem cell pluripotency as well as lung developmentand differentiation. Indeed, they were overexpressed in thelungs of mice at the late embryonic stage or newborn miceand then decrease dramatically during the first weeks oflife (45, 46). A recent study that performed RNAseq inKRASG12D transgenic tumors found that miRNAs encodedat the Dlk1-Dio3 locus on mouse chromosome 12qF1 wereconsistently increased in tumors as compared with non-malignant lung (47). Interestingly, these miRNAs wererepressed when KRASG12D cells were grown in vitro andwere reactivated when transplanted in vivo.
To investigate the biologic processes related to specific14q32 miRNAs, we determined which genes were corre-lated with specific miRNAs using gene expression datafrom 51 lung ACs. We recognize a limitation of using thisapproach as miRNAs may reduce protein expressionwithout affecting mRNA stability. Nevertheless, thisapproach might be hypothesis generating for identifyingcellular processes that are deregulated in primary tumorsoverexpressing certain miRNAs. We found that severalgene families were consistently correlated with expressionof these miRNAs, such as regulation of cell motility,which may confer a more aggressive phenotype. We alsoobserved that migration of lung AC cells was reduced afterknocking down miR-411. Furthermore, miR-411 wassignificantly overexpressed in tumors from patients whodeveloped a distant recurrence and miR-411 expressioncorrelated with genes involved in EMT. Indeed, it hasbeen recently reported that the expression of variousmiRNAs encoded at 14q32 significantly increased aftertreating lung AC cells with TGF-b, and knockdown ofthese miRNAs inhibited the EMT process (48).
In conclusion, miRNA expression profiles not only iden-tifiedmiRNAs differentially expressed in lung AC compared
with nonmalignant lung, but also were correlated withtumor differentiation and morphologic subtype of lungAC, suggesting that miRNA profiles may subclassify lungAC. We identified specific miRNAs located at 14q32 inde-pendently associated with poor outcome after lung ACresection. We focused on miR-411, which was overex-pressed in patients who develop metastasis after surgeryand was correlated with genes involved in cell migrationand EMT. These findings may have implications for thediagnosis of lung AC, and specific miRNAs might be rele-vant targets in the future usingnovel therapeutic approachessuch as antisense oligonucleotides.
Disclosure of Potential Conflicts of InterestR.M. Reddy reports receiving commercial research support from
GlaxoSmithKine. No potential conflicts of interest were disclosed by theother authors.
Authors' ContributionsConception and design: E. Nadal, R.M. Reddy, A.C. Chang, D.G. Beer,G. ChenDevelopment of methodology: E. Nadal, J. Zhong, D.G. Beer, G. ChenAcquisitionofdata (provided animals, acquired andmanagedpatients,provided facilities, etc.): E. Nadal, J. Zhong, J. Lin, R.M. Reddy,N. Ramnath, M.B. Orringer, D.G. Beer, G. ChenAnalysis and interpretation of data (e.g., statistical analysis, biosta-tistics, computational analysis): E. Nadal, J. Zhong, A.C. Chang,D.G. Beer, G. ChenWriting, review, and/or revision of the manuscript: E. Nadal, J. Lin,R.M. Reddy, N. Ramnath, M.B. Orringer, A.C. Chang, D.G. Beer, G. ChenAdministrative, technical, or material support (i.e., reporting or orga-nizing data, constructing databases): E. Nadal, A.C. Chang, D.G. BeerStudy supervision: D.G. Beer, G. Chen
AcknowledgmentsThe authors thank Dr. Gabriel Capell�a for scientific input and critical
review of this article.
Grant SupportThis study was supported by the Bonnie J. Addario Lung Cancer Foun-
dation and the University of Michigan Comprehensive Cancer Center. E.Nadal was supported by a Spanish Society of Medical Oncology Fellowship.
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received December 11, 2013; revised March 28, 2014; accepted April 1,2014; published OnlineFirst April 14, 2014.
References1. Siegel R, Naishadham D, Jemal A. Cancer statistics, 2012. CA Cancer
J Clin 2012;62:10–29.2. Youlden DR, Cramb SM, Baade PD. The international epidemiology of
lung cancer: geographical distribution and secular trends. J ThoracOncol 2008;3:819–31.
3. Travis WD, Brambilla E, Noguchi M, Nicholson AG, Geisinger KR,Yatabe Y, et al. International association for the study of lung cancer/American thoracic society/European respiratory society internationalmultidisciplinary classification of lung adenocarcinoma. J ThoracOncol 2011;6:244–85.
4. Li T, Kung HJ, Mack PC, Gandara DR. Genotyping and genomicprofiling of non-small-cell lung cancer: implications for current andfuture therapies. J Clin Oncol 2013;31:1039–49.
5. Director's Challenge Consortium for the Molecular Classification ofLung AShedden K, Taylor JM, Enkemann SA, Tsao MS, YeatmanTJ, et al. Gene expression-based survival prediction in lung ade-nocarcinoma: a multi-site, blinded validation study. Nat Med 2008;14:822–7.
6. Bryant CM, Albertus DL, Kim S, Chen G, Brambilla C, Guedj M, et al.Clinically relevant characterization of lung adenocarcinoma subtypesbased on cellular pathways: an international validation study. PLoSONE 2010;5:e11712.
7. Esquela-Kerscher A, Slack FJ. Oncomirs - microRNAs with a role incancer. Nat Rev Cancer 2006;6:259–69.
8. HammondSM.MicroRNAs as tumor suppressors. NatGenet 2007;39:582–3.
Nadal et al.
Clin Cancer Res; 20(12) June 15, 2014 Clinical Cancer Research3116
on May 30, 2018. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 15, 2014; DOI: 10.1158/1078-0432.CCR-13-3348

9. Lu J, Getz G, Miska EA, Alvarez-Saavedra E, Lamb J, Peck D, et al.MicroRNA expression profiles classify human cancers. Nature 2005;435:834–8.
10. Volinia S, Calin GA, Liu CG, Ambs S, Cimmino A, Petrocca F, et al. AmicroRNA expression signature of human solid tumors defines cancergene targets. Proc Natl Acad Sci U S A 2006;103:2257–61.
11. Raponi M, Dossey L, Jatkoe T, Wu X, Chen G, Fan H, et al. MicroRNAclassifiers for predicting prognosis of squamous cell lung cancer.Cancer Res 2009;69:5776–83.
12. Iorio MV, Ferracin M, Liu CG, Veronese A, Spizzo R, Sabbioni S, et al.MicroRNA gene expression deregulation in human breast cancer.Cancer Res 2005;65:7065–70.
13. Sempere LF, Christensen M, Silahtaroglu A, Bak M, Heath CV,Schwartz G, et al. Altered MicroRNA expression confined to specificepithelial cell subpopulations in breast cancer. Cancer Res 2007;67:11612–20.
14. Yanaihara N, Caplen N, Bowman E, Seike M, Kumamoto K, Yi M, et al.Unique microRNA molecular profiles in lung cancer diagnosis andprognosis. Cancer Cell 2006;9:189–98.
15. Yu SL, Chen HY, Chang GC, Chen CY, Chen HW, Singh S, et al.MicroRNA signature predicts survival and relapse in lung cancer.Cancer Cell 2008;13:48–57.
16. Landi MT, Zhao Y, Rotunno M, Koshiol J, Liu H, Bergen AW, et al.MicroRNA expression differentiates histology and predicts survival oflung cancer. Clin Cancer Res 2010;16:430–41.
17. Jang JS, Jeon HS, Sun Z, AubryMC, Tang H, Park CH, et al. IncreasedmiR-708 expression in NSCLC and its association with poor survival inlung adenocarcinoma from never smokers. Clin Cancer Res 2012;18:3658–67.
18. Wang R, Wang ZX, Yang JS, Pan X, De W, Chen LB. MicroRNA-451functions asa tumor suppressor in humannon-small cell lungcancer bytargeting ras-related protein 14 (RAB14). Oncogene 2011;30:2644–58.
19. Boeri M, Verri C, Conte D, Roz L, Modena P, Facchinetti F, et al.MicroRNA signatures in tissues and plasma predict development andprognosis of computed tomography detected lung cancer. Proc NatlAcad Sci U S A 2011;108:3713–8.
20. Edge S, Byrd DR, Compton CC, Fritz AG, Greene FL, Trotti A. AJCCCancer Staging Manual. 7th ed. Chicago: Springer; 2010.
21. Chen HY, Yu SL, Chen CH, Chang GC, Chen CY, Yuan A, et al. A five-gene signature and clinical outcome in non-small-cell lung cancer. NEngl J Med 2007;356:11–20.
22. Weir BA, Woo MS, Getz G, Perner S, Ding L, Beroukhim R, et al.Characterizing the cancer genome in lung adenocarcinoma. Nature2007;450:893–8.
23. Hassan KA, Chen G, Kalemkerian GP, Wicha MS, Beer DG. Anembryonic stem cell-like signature identifies poorly differentiated lungadenocarcinoma but not squamous cell carcinoma. Clin Cancer Res2009;15:6386–90.
24. Lewis BP, Burge CB, Bartel DP. Conserved seed pairing, often flankedby adenosines, indicates that thousands of human genes are micro-RNA targets. Cell 2005;120:15–20.
25. Betel D, Wilson M, Gabow A, Marks DS, Sander C. The microRNA.orgresource: targets and expression. Nucleic AcidsRes 2008;36:D149–53.
26. Dweep H, Sticht C, Pandey P, Gretz N. miRWalk–database: predictionof possible miRNA binding sites by "walking" the genes of threegenomes. J Biomed Inform 2011;44:839–47.
27. Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, et al.Gene ontology: tool for the unification of biology. The Gene OntologyConsortium. Nat Genet 2000;25:25–9.
28. Tusher VG, Tibshirani R, Chu G. Significance analysis of microarraysapplied to the ionizing radiation response. Proc Natl Acad Sci U S A2001;98:5116–21.
29. PapadopoulosGL, AlexiouP,MaragkakisM,ReczkoM,HatzigeorgiouAG. DIANA-mirPath: integrating human and mouse microRNAs inpathways. Bioinformatics 2009;25:1991–3.
30. de Hoon MJ, Imoto S, Nolan J, Miyano S. Open source clusteringsoftware. Bioinformatics 2004;20:1453–4.
31. Eisen MB, Spellman PT, Brown PO, Botstein D. Cluster analysis anddisplay of genome-wide expression patterns. ProcNatl Acad Sci USA1998;95:14863–8.
32. Lin SP, Youngson N, Takada S, Seitz H, Reik W, Paulsen M, et al.Asymmetric regulation of imprinting on the maternal and paternalchromosomes at the Dlk1-Gtl2 imprinted cluster on mouse chromo-some 12. Nat Genet 2003;35:97–102.
33. Takada S, Tevendale M, Baker J, Georgiades P, Campbell E, FreemanT, et al. Delta-like and gtl2 are reciprocally expressed, differentiallymethylated linked imprinted genes on mouse chromosome 12. CurrBiol 2000;10:1135–8.
34. Carr MS, Yevtodiyenko A, Schmidt CL, Schmidt JV. Allele-specifichistone modifications regulate expression of the Dlk1-Gtl2 imprinteddomain. Genomics 2007;89:280–90.
35. Kameswaran V, Bramswig NC, McKenna LB, Penn M, Schug J, HandNJ, et al. Epigenetic regulation of theDLK1-MEG3MicroRNA cluster inhuman type 2 diabetic islets. Cell Metab 2014;19:135–45.
36. Groger CJ, Grubinger M, Waldhor T, Vierlinger K, Mikulits W. Meta-analysis of gene expression signatures defining the epithelial to mes-enchymal transition during cancer progression. PLoS ONE 2012;7:e51136.
37. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation.Cell 2011;144:646–74.
38. Lebanony D, Benjamin H, Gilad S, Ezagouri M, Dov A, Ashkenazi K,et al. Diagnostic assay based on hsa-miR-205 expression distin-guishes squamous from nonsquamous non-small-cell lung carcino-ma. J Clin Oncol 2009;27:2030–7.
39. Blenkiron C, Goldstein LD, Thorne NP, Spiteri I, Chin SF, Dunning MJ,et al. MicroRNA expression profiling of human breast cancer identifiesnew markers of tumor subtype. Genome Biol 2007;8:R214.
40. Iorio MV, Visone R, Di Leva G, Donati V, Petrocca F, Casalini P, et al.MicroRNA signatures in human ovarian cancer. Cancer Res 2007;67:8699–707.
41. Puissegur MP, Mazure NM, Bertero T, Pradelli L, Grosso S, Robbe-Sermesant K, et al. miR-210 is overexpressed in late stages of lungcancer and mediates mitochondrial alterations associated with mod-ulation of HIF-1 activity. Cell Death Differ 2011;18:465–78.
42. Nadal E, Chen G, Gallegos M, Lin L, Ferrer-Torres D, Truini A, et al.Epigenetic inactivation of microRNA-34b/c predicts poor disease-freesurvival in early stage lung adenocarcinoma. Clin Cancer Res 2013;19:6842–52.
43. Luk JM, Burchard J, Zhang C, Liu AM,Wong KF, Shek FH, et al. DLK1-DIO3 genomic imprinted microRNA cluster at 14q32.2 defines a stem-like subtypeof hepatocellular carcinomaassociatedwith poor survival.J Biol Chem 2011;286:30706–13.
44. Costa FF, Bischof JM, Vanin EF, Lulla RR, Wang M, Sredni ST, et al.Identification of microRNAs as potential prognostic markers in epen-dymoma. PLoS ONE 2011;6:e25114.
45. Izzotti A, Calin GA, Steele VE, Croce CM, De Flora S. Relationships ofmicroRNA expression in mouse lung with age and exposure to ciga-rette smoke and light. FASEB J 2009;23:3243–50.
46. Dong J, Jiang G, Asmann YW, Tomaszek S, Jen J, Kislinger T, et al.MicroRNA networks inmouse lung organogenesis. PLoSONE2010;5:e10854.
47. Valdmanis PN, Roy-Chaudhuri B, Kim HK, Sayles LC, Zheng Y,Chuang CH, et al. Upregulation of the microRNA cluster at theDlk1-Dio3 locus in lung adenocarcinoma. Oncogene 2013 Dec 9.[Epub ahead of print].
48. Kitamura K, Seike M, Okano T, Matsuda K, Miyanaga A, Mizutani H,et al. MiR-134/487b/655 cluster regulates TGF-beta-induced epithe-lial-mesenchymal transition and drug resistance to gefitinib by target-ing MAGI2 in lung adenocarcinoma cells. Mol Cancer Ther 2014;13:444–53.
www.aacrjournals.org Clin Cancer Res; 20(12) June 15, 2014 3117
MicroRNAs at 14q32 Are Prognostic in Lung Adenocarcinoma
on May 30, 2018. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 15, 2014; DOI: 10.1158/1078-0432.CCR-13-3348

2014;20:3107-3117. Published OnlineFirst May 15, 2014.Clin Cancer Res Ernest Nadal, Jinjie Zhong, Jules Lin, et al. AdenocarcinomaA MicroRNA Cluster at 14q32 Drives Aggressive Lung
Updated version
10.1158/1078-0432.CCR-13-3348doi:
Access the most recent version of this article at:
Material
Supplementary
http://clincancerres.aacrjournals.org/content/suppl/2014/05/15/1078-0432.CCR-13-3348.DC1
Access the most recent supplemental material at:
Cited articles
http://clincancerres.aacrjournals.org/content/20/12/3107.full#ref-list-1
This article cites 46 articles, 17 of which you can access for free at:
Citing articles
http://clincancerres.aacrjournals.org/content/20/12/3107.full#related-urls
This article has been cited by 1 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://clincancerres.aacrjournals.org/content/20/12/3107To request permission to re-use all or part of this article, use this link
on May 30, 2018. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 15, 2014; DOI: 10.1158/1078-0432.CCR-13-3348