a new subfamily classiï¬cation of the palm family (arecaceae
TRANSCRIPT

Botanical Journal of the Linnean Society
, 2006,
151
, 15–38. With 3 figures
© 2006 The Linnean Society of London,
Botanical Journal of the Linnean Society,
2006,
151
, 15–38
15
Blackwell Publishing LtdOxford, UKBOJ
Botanical Journal of the Linnean Society
0024-4074The Linnean Society of London, 2006? 2006
151
?1538
Original Article
PALM FAMILY PHYLOGENYC. B. ASMUSSEN ET AL.
*Corresponding author. E-mail: [email protected]
The Palms
Guest edited by William J. Baker and Scott Zona
A new subfamily classification of the palm family (Arecaceae): evidence from plastid DNA phylogeny
CONNY B. ASMUSSEN
1
*, JOHN DRANSFIELD
2
, VINNIE DEICKMANN
1
, ANDERS S. BARFOD
3
, JEAN-CHRISTOPHE PINTAUD
4
and WILLIAM J. BAKER
2
1
Department of Ecology, Royal Veterinary and Agricultural University Copenhagen, Rolighedsvej 21, DK-1958 Frederiksberg, Denmark
2
Herbarium, Royal Botanic Gardens, Kew, Surrey TW9 3AE, UK
3
Department of Biological Sciences, University of Aarhus, Ny Munkegade bygn. 541, DK-8000 Århus C, Denmark
4
Institut de Recherche pour le Développement, UMR DGPC/DYNADIV, 911 Avenue Agropolis BP 64501, 34394 Montpellier cedex 5, France
Received June 2005; accepted for publication November 2005
Published phylogeny reconstructions of the palm family (Arecaceae) are based on plastid DNA sequences or restric-tion fragment length polymorphisms (RFLPs), nuclear DNA sequences, morphological characters or a combinationthereof, and include between 33 and 90 palm species. The present study represents all previously recognized sub-families, tribes and subtribes of palms and 161 of the 189 genera. The plastid DNA region
mat
K was sequenced for178 palm species and ten commelinid monocot outgroup species, and was combined with new and previously pub-lished plastid DNA sequences of
trn
L–
trn
F,
rps
16 intron and
rbcL
. The addition of
mat
K sequences and more taxaresulted in a highly resolved and largely well-supported phylogeny. Most importantly, critical basal nodes are nowfully resolved and, in most cases, strongly supported. On the basis of this phylogeny, we have established a new sub-familial classification of the palms, in which five subfamilies are recognized, rather than the six that were includedin the previous classification. The circumscriptions of the subfamilies Calamoideae and Nypoideae were corrobo-rated. The phylogeny supported a new circumscription for the subfamily Coryphoideae, including all taxa previouslyrecognized in Coryphoideae with the addition of the tribe Caryoteae, formerly of the subfamily Arecoideae. Thephylogenetic analysis also supported a new delimitation for the subfamily Ceroxyloideae that contains the tribesCyclospatheae and Ceroxyleae, and all genera formerly included in the subfamily Phytelephantoideae, butexcludes the tribe Hyophorbeae. Finally, the subfamily Arecoideae was modified to exclude the tribe Caryoteae andto include the tribe Hyophorbeae. © 2006 The Linnean Society of London,
Botanical Journal of the Linnean Society
,2006,
151
, 15–38.
ADDITIONAL KEYWORDS:
mat
K – Palmae –
rbc
L –
rps
16 intron –
trn
L–
trn
F.
INTRODUCTION
The palm family (Arecaceae, Palmae) is resolved as amonophyletic group in all higher-level molecularstudies of monocots (e.g. Chase
et al
., 2000; Asmus-sen & Chase, 2001). During the last 10 years, sub-
stantial progress has been made in theunderstanding of the relationships within the fam-ily. Many estimates of palm phylogeny have beenpublished at various taxonomic levels. Nevertheless,numerous ambiguities have persisted, hindering anyattempt to rearrange the formal classification of thefamily, such as, for example, the placements of thetribes Cyclospatheae and Phoeniceae, and of the sub-

16
C. B. ASMUSSEN
ET AL
.
© 2006 The Linnean Society of London,
Botanical Journal of the Linnean Society,
2006,
151
, 15–38
family Phytelephantoideae. In this paper, which isfocused strictly on the circumscription of palm sub-families, we shall refer primarily to those phylogenyreconstructions that explore the systematics of thefamily at the highest level (Uhl
et al
., 1995; Baker
et al
., 1999; Asmussen, Baker & Dransfield, 2000;Asmussen & Chase, 2001; Hahn, 2002a; Lewis &Doyle, 2002). We use the formal subfamily, tribaland subtribal names in the sense of Dransfield &Uhl (1998), who divided the family into six subfami-lies varying in size from one genus (subfamilyNypoideae) to 112 genera (subfamily Arecoideae; seeAppendix). For the subfamily Calamoideae, however,we use the classification of Baker, Dransfield & Hed-derson (2000a). This study provides part of the justi-fication for a forthcoming new classification of palmsbased on phylogenetic data (Dransfield
et al
., 2005);we make references to the new classification, whereappropriate, within the figures and in the discussionsection below.
C
URRENT
STATUS
OF
PALM
FAMILY
PHYLOGENETICS
Subfamilies Calamoideae and Nypoideae
The subfamily Calamoideae is resolved as mono-phyletic in all palm family phylogenies (Uhl
et al
.,1995; Baker
et al
., 1999, 2000a; Baker, Hedderson &Dransfield, 2000b, c; Asmussen
et al
., 2000; Asmussen& Chase, 2001; Lewis & Doyle, 2001; Hahn, 2002a).Baker
et al
. (2000a, b, c) explored the relationshipswithin this subfamily and proposed a new classifica-tion for the Calamoideae with three tribes and ninesubtribes based on a combination of molecular andmorphological data.
Nypa
, the sole representative of the subfamilyNypoideae, is always resolved on an isolated branchwhen maximum parsimony is employed as the opti-mality criterion (Uhl
et al
., 1995; Baker
et al
., 1999;Asmussen
et al
., 2000; Asmussen & Chase, 2001;Lewis & Doyle, 2001; Hahn, 2002a). In a few analysesusing maximum likelihood,
Nypa
is nested in variouspositions among members of the subfamily Calam-oideae or the subfamily Coryphoideae (Hahn, 2002a),but these relationships have received scant support inother systematic studies.
The Calamoideae and the Nypoideae are the princi-pal candidates for the position as the sister taxon tothe remaining members of Arecaceae. In two recentpapers with extensive taxon and nucleotide charactersampling, the subfamily Calamoideae was resolved assister to all other members of the palm family in totalevidence analyses based on parsimony (Asmussen &Chase, 2001; Hahn, 2002a). This finding contrastsmarkedly with the first phylogenetic study of palmsbased on restriction fragment length polymorphisms(RFLPs) and morphology, in which
Nypa
resolved as
sister to the remaining members of Arecoideae, withthe Calamoideae sister to all palms excluding
Nypa
(Uhl
et al
., 1995). However, this result was probablyinfluenced by the use of only one taxon as an outgroup,
Dioscorea
(Dioscoreaceae), which is only distantlyrelated to palms and commelinid monocots as a whole(Chase
et al
., 2000), thereby increasing the potentialfor a spurious rooting. In subsequent studies, Baker
et al
. (1999) and Asmussen
et al
. (2000) did not includenonpalm outgroups, due to alignment problems, root-ing their phylogenies internally on
Nypa
following Uhl
et al
. (1995). Although their methods were explicit,the results are prone to misinterpretation. However,another study, which included nonpalm outgroups,supports
Nypa
as sister to all other palms (Lewis &Doyle, 2001). Unfortunately, in none of these studiesare the relative positions of
Nypa
or the Calamoideaestrongly supported by bootstrap analysis, renderingthe results effectively equivocal.
Subfamily Coryphoideae
More than half the phylogenetic analyses of thepalm family based on DNA sequences do not resolvethe subfamily Coryphoideae as monophyletic (Baker
et al
., 1999; Asmussen
et al
., 2000; Asmussen &Chase, 2001; Hahn, 2002a). However, the plastidRFLP phylogeny of Uhl
et al
. (1995), in which thetaxonomic sampling was heavily biased towardscoryphoids, resolved subfamily Coryphoideae includ-ing the tribe Caryoteae from the subfamilyArecoideae as a monophyletic group. The study ofLewis & Doyle (2001), based on DNA sequences of thenuclear gene, malate synthase, and that of Hahn(2002a), based on a combined, reduced data set,resolved the Coryphoideae as monophyletic. It shouldbe noted, however, that the sample size was small inboth cases. Many data sets group the tribe Caryoteaeof the subfamily Arecoideae together with members ofthe subfamily Coryphoideae, often with close relation-ships to the subtribe Coryphinae or the tribe Boras-seae (Uhl
et al
., 1995; Asmussen
et al
., 2000;Asmussen & Chase, 2001; Hahn, 2002a).
Subfamilies Ceroxyloideae and Phytelephantoideae
It is clear from most phylogenetic analyses that thesubfamily Ceroxyloideae (
sensu
Dransfield & Uhl,1998) is not monophyletic (Uhl
et al
., 1995; Baker
et al
., 1999; Asmussen
et al
., 2000; Asmussen &Chase, 2001; Lewis & Doyle, 2001; Hahn, 2002a, b).One tribe, the Hyophorbeae, is consistently resolvedwith members of the subfamily Arecoideae. The exactrelationships and positions of the tribes Ceroxyleaeand Cyclospatheae are not yet clear, however. In con-trast, the subfamily Phytelephantoideae is alwaysresolved as monophyletic (Uhl
et al
., 1995; Asmussen
et al
., 2000; Asmussen & Chase, 2001; Hahn, 2002a)

PALM FAMILY PHYLOGENY
17
© 2006 The Linnean Society of London,
Botanical Journal of the Linnean Society,
2006,
151
, 15–38
with the exception of Baker
et al
. (1999), where thetwo included species are unresolved in a polytomy atthe base of the Arecoid line (Moore, 1973).
Recent studies provide modest support for a clade ofPhytelephantoideae, Ceroxyloideae, and Arecoideae, agroup that is equivalent to the Arecoid line
sensu
Moore (1973; Asmussen & Chase, 2001; Hahn, 2002a,b). Various studies give indications of potential rela-tionships between the Ceroxyleae, Cyclospatheae, andPhytelephantoideae, or between at least two of thethree groups (Uhl
et al
., 1995; Asmussen & Chase,2001; Hahn, 2002a, b). Most strikingly, Asmussen &Chase (2001) provided evidence, albeit weakly sup-ported, that Phytelephantoideae, Cyclospatheae andCeroxyleae form a monophyletic sister group to thesubfamily Arecoideae.
Subfamily Arecoideae
Most studies point towards a broadly monophyleticsubfamily Arecoideae, with the majority includingthe tribe Hyophorbeae (subfamily Ceroxyloideae) andexcluding the tribe Caryoteae (Uhl
et al
., 1995; Baker
et al
., 1999; Asmussen
et al
., 2000; Asmussen &Chase, 2001; Hahn, 2002a, b).
P
OTENTIAL
FOR
REVISION
OF
THE
CURRENT
CLASSIFICATION
Although much progress has been made towards arobust phylogeny of the palm family, a major revisionof the prevailing classification based on publishedphylogenetic hypotheses is premature. At the highestlevel in particular, the lack of resolution and bootstrapsupport at the basal nodes forming the backbone ofthe phylogeny seriously hinders the production of arobust, lasting, circumscription of subfamilies. Forreasons outlined above, three of the current subfami-lies are in particular need of clarification, namelyCoryphoideae, Ceroxyloideae and Phytelephantoi-deae. The objectives of this study were to explore fur-ther the phylogeny of the palm family by building onprevious studies (Baker
et al
., 1999; Asmussen
et al
.,2000; Asmussen & Chase, 2001) with substantiallyexpanded taxon sampling and by adding the plastidDNA region
mat
K to the pre-existing selection of plas-tid DNA regions (
trn
L–
trn
F,
rps
16 intron and
rbc
L)used in these studies, and to use our findings to pro-pose a formal revision of the subfamily classification ofthe Arecaceae.
MATERIAL AND METHODS
S
AMPLING
This study included 178 palm species, representing162 of the 189 genera recognized in the 1998 treat-ment of Arecaceae (Dransfield & Uhl, 1998). All 36
subtribes, 14 tribes and six subfamilies in the classi-fication of Uhl & Dransfield (1987) and all 36 sub-tribes, 14 tribes and six subfamilies of Dransfield &Uhl (1998; see Appendix) were represented. All tribesand subtribes of the revised classification of theCalamoideae of Baker
et al
. (2000a) were also repre-sented. The
mat
K region was chosen as an additionalplastid DNA region because it has provided many par-simony-informative characters in other monocot stud-ies. Other plastid DNA regions (
rpl
16,
rpo
C, and
ndh
F) were tested on a small sample of species aspotential new plastid DNA markers, but these regionsshowed amplification difficulties, whereas
mat
Kamplified readily in all palm test samples. All
mat
Ksequences were produced for this study and are pub-lished here for the first time. In addition,
rbc
L,
rps
16,and
trnL–trnF sequences for taxa not previouslyincluded in our data sets were generated; all otherdata were recycled from three previous studies (Bakeret al., 1999; Asmussen et al., 2000; Asmussen &Chase, 2001; see Appendix). Ten monocot outgroupspecies were selected from among the clades mostclosely related to the palm family (Chase et al., 2000;see Appendix).
DNA EXTRACTION, POLYMERASE CHAIN REACTION (PCR) AND NUCLEOTIDE SEQUENCING
Total genomic DNA was extracted from fresh or silicagel-dried plant material using the 2× CTAB method ofDoyle & Doyle (1987) or the DNeasy Plant Mini Kit(Qiagen). Some of the 2× CTAB extractions were fol-lowed by purification on caesium chloride/ethidiumbromide gradients (1.55 g ml−1) or with the QIAquickPCR purification kit (Qiagen) with 35% guanidiniumchloride ((NH2)2C:NH.HCl). The DNA concentrationswere measured on a biophotometer (Eppendorf). Allsamples were vouchered with herbarium specimens(see Appendix).
The matK sequences were amplified from totalgenomic DNA using the primer matK-19F with trnK-2R (Table 1; Steele & Vilgalys, 1994). If amplificationwas unsuccessful, reactions were repeated usingmatK-19F with matK-1862R or in two pieces with anycombination of the available primers (Table 1). PCRreactions (100 µl) were prepared on ice by combining65 µl ddH2O, 10 µl 10× DNA polymerase buffer, 8 µl20 µmol l−1 MgCl2, 4 µl 10 mmol l−1 each dNTP, 1 µl10 mg ml−1 bovine serum albumin, 5 µl of each primer(10 µmol l−1), 1 µl 5 µ µl−1 Supertaq DNA polymerase(HT Biotechnology), and 25 ng of template DNA. Theamplifications were conducted on an MJ ResearchPTC-200 thermocycler programmed as follows: onecycle at 94 °C for 3 min, 28 cycles of 94 °C for 1 min,50 °C (or up to 60 °C for problematic DNA samples) for1 min, and 72 °C for 2 min, and a final cycle at 72 °C

18 C. B. ASMUSSEN ET AL.
© 2006 The Linnean Society of London, Botanical Journal of the Linnean Society, 2006, 151, 15–38
for 5 min. The resulting PCR products were checkedon a 0.8% agarose gel with ethidium bromide and puri-fied using the QIAquick PCR purification kit (Qiagen)with 35% guanidinium chloride ((NH2)2C:NH.HCl).The amplification primers and protocols for the rbcLregion were those described in Asmussen & Chase(2001); a new primer, rbcL-1407R was designed andused for DNAs that would not amplify with rbcL-reverse (Fay et al., 1998). The rps16 intron region wasamplified using the primers of Oxelman, Lidén &Berglund (1997) and the protocols of Asmussenet al. (2000). The trnL–trnF region was amplified usingthe primers of Taberlet et al. (1991) and the protocolsfor amplification followed Asmussen et al. (2000)and Baker et al. (1999).
The concentrations of purified PCR products weremeasured on a biophotometer and the products weresequenced using the ABI PRISM BigDye terminatorcycle sequencing ready reaction kit (Perkin-Elmer, ABApplied Biosystems). For matK, the PCR amplificationprimers, matK-19F and trnK-2R, performed poorly assequencing primers and therefore six new primerswere designed as sequencing primers (Table 1). Thesequencing primers for rbcL, rps16 and trnL–trnFwere those described in Asmussen & Chase (2001),Asmussen et al. (2000) and Baker et al. (1999) inaddition to the new rbcL-1407R (Table 1). Cycle-sequencing reactions (10 µl) were prepared by combin-ing 1 µl terminator mix, 3 µl 5× cycle-sequencingbuffer (200 mmol l−1 trizma base, 5 mmol l−1 MgCl2,pH 9.0, from the BigDye terminator kit), 1 µl primer(1 µmol l−1), 25 ng DNA from the cleaned PCR productand ddH2O up to 10 µl. Cycle sequencing was con-ducted on an MJ Research PTC-200 thermocycler pro-grammed as follows: 25 cycles of 96 °C for 30 s, 50 °Cfor 15 s and 60 °C for 4 min
Cycle-sequencing products were cleaned usingDye-Ex Spin columns (Qiagen) or Sephadex G-50(Roche) following the protocol of the manufacturer.The cleaned cycle-sequencing products were analysedon a PE Applied Biosystems 377 automated DNA
sequencer (Perkin-Elmer) or a PE Applied Biosystems3100 capillary automated DNA sequencer (Perkin-Elmer). Each base position in the forward and reversesequences was checked and assembled using the pro-gram SEQUENCHER 3.0 (Gene Codes Corp.).
SEQUENCE ALIGNMENT
Initial automated alignments of consensus sequenceswere performed with the MegAlign program (Laser-gene software package, DNASTAR Inc.) and followedby refinement by hand. The alignment of rbcLsequences was straightforward due to the absence oflength variation. The alignment of matK was also rel-atively straightforward except for a number of indelsat the 3′ end. For the length-variable rps16 intron andtrnL–trnF sequences, the alignments included numer-ous indels, but they were not recoded as additionalcharacters. The aligned matK, rbcL, rps16 intron andtrnL–trnF sequence matrices were combined andanalysed together. For separate analyses of rbcL,rps16 intron and trnL–trnF, see Asmussen & Chase(2001), Asmussen et al. (2000) and Baker et al. (1999).
CLADISTIC ANALYSES
The four data sets were readily combined because theyall originated from plastid DNA and therefore haveidentical evolutionary history, which makes congru-ence tests superfluous. However, the tree statisticsindicate that the individual data sets are compatible,because the number of nodes, the number of supportednodes and the number of highly supported nodesincrease in the result of the analysis of the combineddata set (Table 2). The data sets were analysed byFitch parsimony (Fitch, 1971; unordered, equallyweighted characters) using PAUP* version 4.0 Beta 10(Swofford, 2002). The analyses yielded many trees,principally because of zero-length branches resultingfrom an inadequate number of informative characters.Thus, heuristic searches could not be run to com-pletion. Therefore, the following search strategywas used. One thousand random replicate searcheswere conducted using the tree–bisection–reconnection(TBR) branch-swapping algorithm with steepestdescent and MULPARS in effect, but holding five treesper step to minimize the time spent swapping on sub-optimal trees. A round of TBR swapping was per-formed on the trees collected during the 1000 randomreplicates, collecting 30 000 optimal trees, and thesetrees were swapped to completion. Support for cladeswas calculated by conducting 1000 bootstrap repli-cates, each with five random replicates, subtree prun-ing–regrafting (SPR) swapping, and saving no morethan five trees each replicate. Only groups thatappeared in > 50% of the trees were retained. Jack-
Table 1. Primer sequences designed for this project andnot previously published
Primer name Primer sequence
MatK-300F 5′-AGT TCA GTA CTT GTR AAA CG-3′MatK-445R 5′-GGG AAG ATA CTA ATC GCA GC-3′MatK-809F 5′-CGA TTA ACA TCT TCT GGA GC-3′MatK-971R 5′-ATG CAT GAA GGG ATC CTT GA-3′MatK-1315F 5′-TCG TGT GCT AGA ACT TTG GC-3′MatK-1334R 5′-GCC AAA GTT CTA GCA CAC GA-3′MatK-1862R 5′-CAT TGC ACA CGA CTT TAC C-3′RbcL-1407R 5′-CCA GCT TAT CTA CTG GTT CG-3′
PALM FAMILY PHYLOGENY 19
© 2006 The Linnean Society of London, Botanical Journal of the Linnean Society, 2006, 151, 15–38
knife percentages and Bremer support values werecalculated for the subfamily clades and the majornodes connecting the subfamilies. A 10 000 replicatejackknife analysis was conducted with collapsebranches if the minimum length was zero, jackknifewith 36.79% deletion, emulate ‘Jac’ resampling ineffect, and a full heuristic search of five replicates,saving a maximum of five trees each replicate andnearest-neighbour interchange swapping. Bremersupport was calculated using the ‘load constraint’option, and for each node conducting 1000 randomreplicate searches using the TBR branch-swappingalgorithm with steepest descent and MULPARS ineffect and holding five trees per step. All parsimonyanalyses were performed under DELTRAN due to themalfunction of ACCTRAN in PAUP* 4b version 10.
RESULTS
SEQUENCE VARIATION
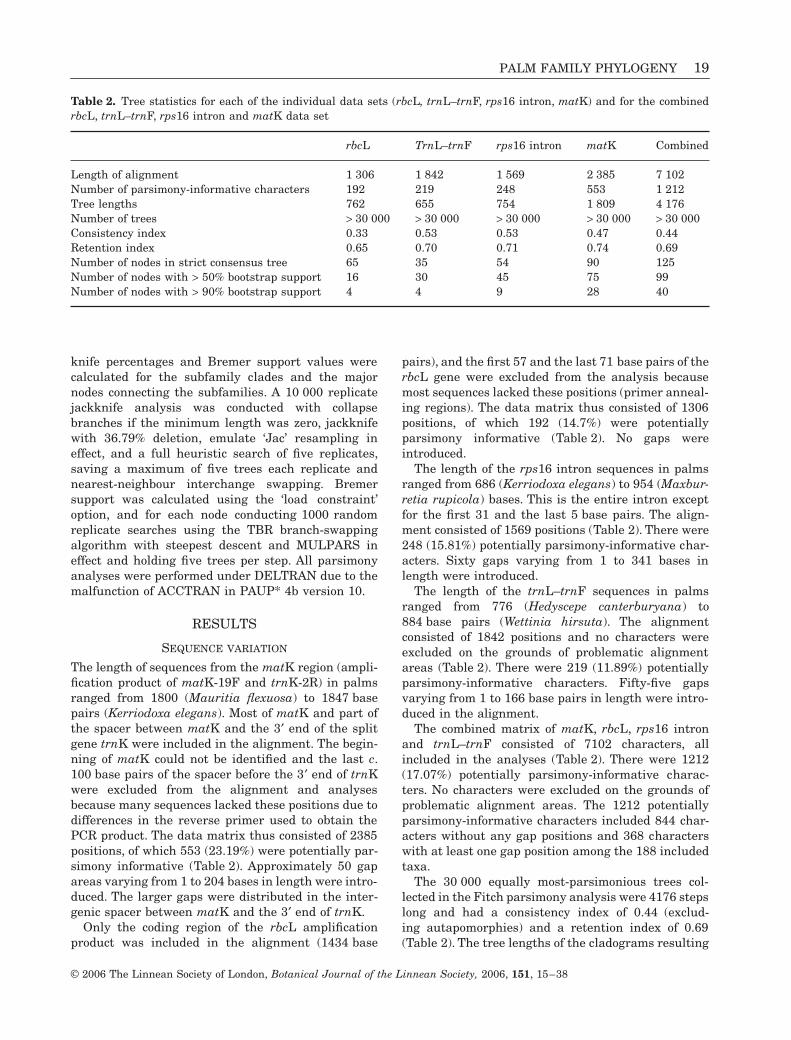
The length of sequences from the matK region (ampli-fication product of matK-19F and trnK-2R) in palmsranged from 1800 (Mauritia flexuosa) to 1847 basepairs (Kerriodoxa elegans). Most of matK and part ofthe spacer between matK and the 3′ end of the splitgene trnK were included in the alignment. The begin-ning of matK could not be identified and the last c.100 base pairs of the spacer before the 3′ end of trnKwere excluded from the alignment and analysesbecause many sequences lacked these positions due todifferences in the reverse primer used to obtain thePCR product. The data matrix thus consisted of 2385positions, of which 553 (23.19%) were potentially par-simony informative (Table 2). Approximately 50 gapareas varying from 1 to 204 bases in length were intro-duced. The larger gaps were distributed in the inter-genic spacer between matK and the 3′ end of trnK.
Only the coding region of the rbcL amplificationproduct was included in the alignment (1434 base
pairs), and the first 57 and the last 71 base pairs of therbcL gene were excluded from the analysis becausemost sequences lacked these positions (primer anneal-ing regions). The data matrix thus consisted of 1306positions, of which 192 (14.7%) were potentiallyparsimony informative (Table 2). No gaps wereintroduced.
The length of the rps16 intron sequences in palmsranged from 686 (Kerriodoxa elegans) to 954 (Maxbur-retia rupicola) bases. This is the entire intron exceptfor the first 31 and the last 5 base pairs. The align-ment consisted of 1569 positions (Table 2). There were248 (15.81%) potentially parsimony-informative char-acters. Sixty gaps varying from 1 to 341 bases inlength were introduced.
The length of the trnL–trnF sequences in palmsranged from 776 (Hedyscepe canterburyana) to884 base pairs (Wettinia hirsuta). The alignmentconsisted of 1842 positions and no characters wereexcluded on the grounds of problematic alignmentareas (Table 2). There were 219 (11.89%) potentiallyparsimony-informative characters. Fifty-five gapsvarying from 1 to 166 base pairs in length were intro-duced in the alignment.
The combined matrix of matK, rbcL, rps16 intronand trnL–trnF consisted of 7102 characters, allincluded in the analyses (Table 2). There were 1212(17.07%) potentially parsimony-informative charac-ters. No characters were excluded on the grounds ofproblematic alignment areas. The 1212 potentiallyparsimony-informative characters included 844 char-acters without any gap positions and 368 characterswith at least one gap position among the 188 includedtaxa.
The 30 000 equally most-parsimonious trees col-lected in the Fitch parsimony analysis were 4176 stepslong and had a consistency index of 0.44 (exclud-ing autapomorphies) and a retention index of 0.69(Table 2). The tree lengths of the cladograms resulting
Table 2. Tree statistics for each of the individual data sets (rbcL, trnL–trnF, rps16 intron, matK) and for the combinedrbcL, trnL–trnF, rps16 intron and matK data set
rbcL TrnL–trnF rps16 intron matK Combined
Length of alignment 1 306 1 842 1 569 2 385 7 102Number of parsimony-informative characters 192 219 248 553 1 212Tree lengths 762 655 754 1 809 4 176Number of trees > 30 000 > 30 000 > 30 000 > 30 000 > 30 000Consistency index 0.33 0.53 0.53 0.47 0.44Retention index 0.65 0.70 0.71 0.74 0.69Number of nodes in strict consensus tree 65 35 54 90 125Number of nodes with > 50% bootstrap support 16 30 45 75 99Number of nodes with > 90% bootstrap support 4 4 9 28 40

20 C. B. ASMUSSEN ET AL.
© 2006 The Linnean Society of London, Botanical Journal of the Linnean Society, 2006, 151, 15–38
from the combined analyses were longer than thecombined lengths of the four individual data sets(762 + 655 + 754 + 1809 = 3980), indicating that thecombined analysis recovered homoplasy not present ineach of the individual analyses.
PHYLOGENETIC ANALYSES
The strict consensus tree of the combined data set waswell resolved and included many well-supportedclades (Fig. 1). The palm family was resolved as mono-phyletic with a bootstrap support of 100%. The mostresolved individual tree had seven polytomies of threetaxa each (Fig. 2). Five of these seven polytomies ofthe most resolved individual tree were present in allindividual trees (Fig. 2, arrows 1–5). Polytomy 6,including four coryphoid taxa, and polytomy 7, com-prising three large clades in the subfamily Arecoideae,were present only in a subgroup of the 30 000 most-parsimonious trees (Fig. 2, arrows 6 and 7). Five of theseven polytomies were positioned in the subfamilyArecoideae, where particularly backbone branchlengths were short compared with backbone branchlengths within the other four subfamilies (Fig. 2).
In the strict consensus tree, the monophyletic (100%bootstrap) subfamily Calamoideae (Fig. 1, clade 1) wasresolved as sister to the rest of the palms. WithinCalamoideae, Eugeissona (tribe Eugeissoneae) wassister to the rest of the Calamoideae (59% bootstrapsupport). Additionally, Calamoideae were divided intotwo large monophyletic groups corresponding to thetwo tribes Lepidocaryeae (91% bootstrap support) andCalameae (70% bootstrap support). The Lepidocar-yeae clade consisted of the African and American taxaMauritia (subtribe Mauritiinae), Raphia (subtribeRaphiinae) and Oncocalamus, Laccosperma and Ere-mospatha (all three from the subtribe Ancistrophylli-nae). The Calameae clade consisted of the largelySouth-east Asian taxa Korthalsia (Korthalsiinae),Salacca (Salaccinae), Calamus (Calaminae), Pigafetta(Pigafettinae), Plectocomia (Plectocomiinae) andMetroxylon (Metroxylinae).
Nypa fruticans, from the monospecific subfamilyNypoideae, was sister to the remaining palms (namelyCalamoideae not included) with a bootstrap support of97% (Fig. 1, clade 2).
The subfamily Coryphoideae, including Caryoteaefrom the subfamily Arecoideae, formed a monophyleticlineage with bootstrap support of 97% (Fig. 1, clade 3).All taxa in this clade have induplicate leaves, exceptfor the anomalous coryphoid genus Guihaia. Threemajor clades received high bootstrap support. The firstof these, with bootstrap support of 98%, consistedentirely of the New World taxa: the genus Sabal (100%bootstrap support; Fig. 1, clade a), sole member of thesubtribe Sabalinae (tribe Corypheae), and the New
World genera of the subtribe Thrinacinae (tribe Cory-pheae; 100% bootstrap support; Fig. 1, clade b). Thesecond major clade, which was weakly supported (61%bootstrap support) as sister to the third (describedbelow), was resolved with 99% bootstrap support andconsisted of exclusively Old World taxa: a highly cor-roborated (100% bootstrap support) monophyleticgroup of three members of the subtribe Coryphinae(Nannorrhops, Kerriodoxa and Chuniophoenix; Fig. 1,clade c), a highly supported (100% bootstrap support)tribe Caryoteae (subfamily Arecoideae, Fig. 1, clade d),the genus Corypha (subtribe Coryphinae; 100% boot-strap support; Fig. 1, clade e) and a highly supported(100% bootstrap support) tribe Borasseae (Fig. 1,clade f). The clade comprising Caryoteae, Corypha andthe Borasseae was monophyletic with bootstrap sup-port of 91%, whereas the support for Corypha as sisterto the Borasseae was low (66% bootstrap support). Thethird major clade received 86% bootstrap support andconsisted of the monogeneric tribe Phoeniceae (100%bootstrap support; Fig. 1, clade g) and a highly sup-ported (99% bootstrap support) clade composed of aparaphyletic subtribe Livistoninae (tribe Corypheae)within which a monophyletic, well-supported (86%bootstrap support) clade of all Old World genera ofThrinacinae (tribe Corypheae) was embedded (Fig. 1,clade h).
There was 85% bootstrap support for the clade cor-responding to Moore’s (1973) Arecoid Line comprisingthe subfamilies Ceroxyloideae, Phytelephantoideaeand Arecoideae, excluding Caryoteae (Fig. 1, clades 4and 5). The subfamily Phytelephantoideae was mono-phyletic (99% bootstrap support) and together withtwo monophyletic tribes, Cyclospatheae (100% boot-strap support) and Ceroxyleae (99% bootstrap sup-port) of the subfamily Ceroxyloideae is denoted asclade 4 on Figure 1 (63% bootstrap support).
The remaining large clade (Fig. 1, clade 5) wasweakly supported (70% bootstrap support) and con-sisted of all genera from the subfamily Arecoideae,except for Caryoteae, with the addition of the tribeHyophorbeae (subfamily Ceroxyloideae). The tribe Iri-arteeae was monophyletic (98% bootstrap support)and sister to a clade (76% bootstrap support) of theremaining members of clade 5. Within this latterclade, the base of which is highly unresolved, Som-mieria and Pelagodoxa (subtribe Iguanurinae) formeda monophyletic group (94% bootstrap support). Themonophyly of the tribe Hyophorbeae (subfamilyCeroxyloideae) was highly supported by bootstrap(100%). The tribe Geonomeae was resolved as non-monophyletic, Welfia and Pholidostachys forming aclade with Manicaria (subtribe Manicariinae; 89%bootstrap support), whereas a clade of Asterogyne,Geonoma, Calyptronoma and Calyptrogyne (80% boot-strap support) resolved elsewhere. The latter group of

PALM FAMILY PHYLOGENY 21
© 2006 The Linnean Society of London, Botanical Journal of the Linnean Society, 2006, 151, 15–38
Geonomeae was sister to a monophyletic subtribeEuterpeinae (56% bootstrap support). The tribeCocoeae was not supported as monophyletic: Beccari-ophoenix (subtribe Beccariophoenicinae) was sister toSclerosperma (Sclerospermatinae) with less than 50%bootstrap support. The remaining members of Cocoeaeformed an unsupported monophyletic group withReinhardtia as the sister group, again without sup-port. The subtribe Elaeidinae (Elaeis) was included ina monophyletic group with the subtribe Bactridinae(Desmoncus, Bactris, Aiphanes and Acrocomia; 73%bootstrap support); and the subtribes Butiinae (Alla-goptera, Syagrus, Cocos, Voanioala, Jubaeopsis) andAttaleinae (Attalea) formed a well-supported mono-phyletic group (91% bootstrap). Most of the speciesrepresenting Indo-Pacific pseudomonomerous generafrom the tribe Areceae resolved in an unsupported andhighly unresolved clade, with some notable exceptions(Pelagodoxa, Sommieria, Iguanura). However, numer-ous smaller groups were resolved within this clade.Two subtribe Arecinae species pairs, Areca triandraand Nenga pumila, and Hydriastele microspadix andH. chaunostachys, the latter representing the recentlysynonymized genus Gronophyllum (Baker & Loo,2004) constituted monophyletic lineages with 84 and78% bootstrap support, respectively, but were notresolved as sister groups. Further clades resolvedwithin the Indo-Pacific pseudomonomerous Areceaeclade and supported by bootstrap include: Rho-palostylis baueri and Hedyscepe canterburyana (sub-tribe Archontophoenicinae; 53% bootstrap support),Masoala (subtribe Masoalinae; 82% bootstrap sup-port), Marojejya (subtribe Masoalinae; 93% bootstrapsupport), Basselinia and Alloschmidia (subtribe Igua-nurinae; 87% bootstrap support), Acanthophoenix,Tectiphiala and Oncosperma (subtribe Oncosper-matinae; 50% bootstrap support), Heterospathe andAlsmithia (subtribe Iguanurinae; 80% bootstrapsupport), Laccospadix and Linospadix (subtribe Lino-spadicinae; 99% bootstrap support), Cyphokentia,Moratia, Lavoixia, Brongniartikentia and Clino-sperma (subtribe Iguanurinae; 63% bootstrap sup-port), and Ptychosperma, Ponapea, Balaka, Veitchia,Carpentaria, Wodyetia, Brassiophoenix and Ptycho-coccus (subtribe Ptychospermatinae; 62% bootstrapsupport).
DISCUSSION
THE SEQUENCES
The matK sequences produced more than twice thenumber of parsimony-informative characters (553) forthe same taxon sample when compared with the otherregions: rbcL (192), trnL–trnF (219) and rps16 intron(248; Table 2). This is in agreement with the resultsfrom other studies where two or more of these plastid
DNA areas were used (Shaw et al., 2005). The rbcLgene produced the fewest parsimony-informativecharacters (192), but these variable charactersresulted in 65 resolved nodes in the palm family,whereas the trnL–trnF region and the rps16 intronproduced only 35 and 54 resolved nodes, respectively,despite supplying more informative characters (219and 248; Table 2). The number of clades with morethan 90% bootstrap support was relatively low in eachof the individual data sets (four, four and nine), exceptfor matK (28), but the combined data set produced thelargest number of highly supported (> 90%) clades (40;Table 2).
THE NEW SUBFAMILY CLASSIFICATION
The dense taxon sampling and the large number ofnucleotide characters included in this study and thehigh levels of resolution and support in the resultingtrees are unprecedented in higher-level palm phyloge-netic research. Our results are sufficiently robust tojustify a formal reclassification of palm subfamiliesand are equally convincing at lower taxonomic levelsin some areas (Fig. 3). Herein, we describe the ratio-nale for recognizing five subfamilies in a forthcomingformal reclassification of the palms (Dransfield et al.,2005).
Subfamily placements for most genera of palmsremain unchanged in the majority of cases withrespect to the previous classification (Uhl & Drans-field, 1987; Dransfield & Uhl, 1998; see Appendix).Nevertheless, the new subfamily classificationrequires three major rearrangements (Figs 1, 2): (1)the tribe Caryoteae from the subfamily Arecoideaesensu Dransfield & Uhl (1998) is moved to a revisedsubfamily Coryphoideae; (2) subfamily Phytelephan-toideae changes rank to tribe Phytelephanteae and isincluded within the new circumscription of the sub-family Ceroxyloideae; and (3) the tribe Hyophorbeaefrom the subfamily Ceroxyloideae (sensu Dransfield &Uhl, 1998) is moved to the subfamily Arecoideae.
SUBFAMILIES CALAMOIDEAE AND NYPOIDEAE
This study strongly supports the monophyly of thesubfamily Calamoideae and firmly positions it as sis-ter to the rest of the palms (Fig. 1, clade 1). Moreover,our results corroborate those of Asmussen & Chase(2001) and the total evidence analyses of Hahn(2002a). The tribal and subtribal classification ofCalamoideae (Baker et al., 2000a) is also corroboratedin this study. The position of the subfamily Nypoideaeas sister to all palms excluding Calamoideae isstrongly supported and its status as a monogenericsubfamily is confirmed, in accordance with all previ-ous studies (Fig. 1, clade 2; Uhl et al., 1995; Baker

22 C. B. ASMUSSEN ET AL.
© 2006 The Linnean Society of London, Botanical Journal of the Linnean Society, 2006, 151, 15–38
Figure 1. The strict consensus tree of 30 000 equally most-parsimonious cladograms resulting from Fitch parsimonyanalyses of combined matK, rbcL, rps16 intron and trnL–trnF data sets. Bootstrap percentages for the clades are givenabove the branches. Five clades are labelled 1–5 for the discussion of the new subfamily classification in the text. Theclades corresponding to the five subfamilies of the new classification are labelled with the subfamily name. Another eightclades are labelled a–h for the discussion of the new tribal classification within the subfamily Coryphoideae. The changesto the classification of Dransfield & Uhl (1998) are indicated with boxes to the right.
Ceroxylon quindiuenseJuania australisOraniopsis appendiculataRavenea louveliiAphandra nataliaPhytelephas aequatorialisAmmandra decaspermaPhytelephas macrocarpaPseudophoenix sargentiiPseudophoenix viniferaMaxburrrtia rupicolaRhapis excelsaRhapidophyllum hystrixGuihaia argyrataTrachycarpus fortuneiChamaerops humilisBrahea berlandieriAcoelorrhaphe wrightiiSerenoa repensColpothrinax wrightiiPritchardia arecinaPritchardia pacificaWashingtonia robustaJohannesteijsmannia altifronsPholidocarpus macrocarpusPritchardiopsis jeanneneyiLicuala kunstleriLivistona chinensisCopernicia pruniferaPhoenix reclinataPhoenix dactyliferaPhoenix canariensisBismarckia nobilisSatranala decussilvaeHyphaene thebaicaMedemia argunBorassodendron machadonisBorassus flabilliferLodoicea maldivicaLatania verschaffeltiiCorypha umbraculiferaCorypha talieraArenga hookerianaArenga undulatifoliaWallichia disticaCaryota mitisCaryota ophiopellisKerriodoxa elegansNannorrhops ritchianaChuniophoenix nanaHemithrinax compactaThrinax morrisiiSchippia concolorTrithrinax campestrisZombia antillarumCoccothrinax argentataThrinax radiataChelyocarpus uleiCryosophila warscewiczianaItaya amicorumSabal bermudanaSabal minorNypa fruticansMetroxylon salomonensePlectocomia mulleriPigafetta elataCalamus aruensisSalacca ramosianaKorthalsia chebEremospatha wendlandianaLaccosperma acutiflorumOncocalamus tuleyiRaphia fariniferaMauritia flexuosaEugeissona tristisHanguana malayanaTradescantia pallidaAnigozanthos manglesiiCanna edulisMusa roseaFargesia sp.Typha angustifoliaVriesia psittacinaDasypogon bromelifoliusKingia australis
SUBFAMILY ARECOIDEAE
100
PALMS
4
3
2
1
5
100
97
97
57
100
100
100
63
8386
100
62
66
10091
7070
9453
100
100
98
70
63100
9896
8710099
61
91
66
100
100
6555
92
7182
58
86
10067
99
100
53
86
75
85
63
100
67
99
9991
58
70
82
100
Caryoteae, formerlyArecoideae
FormerlyPhytelephantoideae
SUBFAMILY CEROXYLOIDEAE
SUBFAMILY CORYPHOIDEAE
SUBFAMILY NYPOIDEAE
SUBFAMILY CALAMOIDEAE
continued
b
a
f
g
h
e
d
c
59
52
52

PALM FAMILY PHYLOGENY 23
© 2006 The Linnean Society of London, Botanical Journal of the Linnean Society, 2006, 151, 15–38
Ptychosperma macarthuriiPonapea ledermannianusBalaka seemanniiVeitchia arecinaCarpentaria arcuminataWodyetia bifurcataBrassiophoenix schumanniiPtychococcus paradoxusCyphokentia macrostachysMoratia ceriferaLavoixia macrocarpaBrongniartikentia lanuginosaClinosperma bractealeLinospadix monostachyaLaccospadix australasicaDransfieldia micranthaHeterospathe elataHeterospathe longipesCarpoxylon macrospermumSatakentia liukiuensisBentinckia nicobaricaClinostigma savoryanumAcanthophoenix rubraTectiphiala feroxOncosperma tigillariumAreca triandraNenga pumilaAlloschmidia glabrataBasselinia velutinaMarojejya darianiiMarojejya insignisMasoala madagascariensisMasoala konaRhopalostylis baueriHedyscepe canterburyanaHydriastele microspadixHydriastele chaunostachys
Actinorhytis calappariaArchontophoenix purpureaActinokentia divaricataChambeyronia macrocarpaKentiopsis oliviformisCampecarpus fulcitusCyphophoenix nucele
Cyphosperma balansaePhysokentia rosea
Veillonia albaNeoveitchia storckiiDypsis lutescensLemurophoenix halleuxiiCalyptrocalyx albertisianaPhoenicophorium borsigianumRoscheria melanochaetesCyrtostachys rendaDictyosperma albumLepidorrhachis mooreanaLoxococcus rupicolaRhopaloblaste augustaAllagoptera arenariaSyagrus smithiiAttalea alleniiCocos nuciferaVoanioala gerardiiJubaeopsis caffraAcrocomia aculeataAcrocomia crispaAiphanes aculeataBactris gasipaesDesmoncus orthacanthosElaeis guineensisReinhardtia simplexPrestoea pubensNeonicholsonia watsoniiEuterpe oleraceaHyospathe macrorhachisCalyptrogyne ghiesbreghtianaCalyptronoma occidentalisGeonoma congestaAsterogyne martianaChamaedorea microspadixGaussia mayaSynechanthus warscewiczianusHyophorbe lagenicaulisWendlandiella gracilisWelfia regiaPholidostachys pulchraManicaria saccifera
Podococcus barteriPodococcus barteriSclerosperma manniiBeccariophoenix madagascariensisPelagodoxa henryanaSommieria leucophyllaOrania lauterbachianaOrania ravakaRoystonea oleracea
Leopoldinia pulchra
Iguanura wallichianaIriartella stenocarpaWettinia hirsutaDictyocaryum lamarckianum
5
Socratea exorrhizaIriartea deltoidea
70
98
77
76
78
80
53
99
52
93
82
62
84
87
81
100
94
92
99
66
8574
8091
60
73
80
91
100
56
8282
6589
56
5094
8763
99
Hyophorbeae, formerlyCeroxyloideae
SUBFAMILY ARECOIDEAE
continued
53
92
51
61
Figure 1. Continued

24 C. B. ASMUSSEN ET AL.
© 2006 The Linnean Society of London, Botanical Journal of the Linnean Society, 2006, 151, 15–38
Ptychosperma macarthuriiPonapea ledmannianus
Balaka seemanniiVeitchia arecina
Carpentaria arcuminataWodyetia bifurcata
Brassiophoenix schumanniiPtychococcus paradoxus
Lepidorrhachis mooreanaArchontophoenix purpurea
Cyphophoenix nuceleVeillonia alba
Rhopalostylis baueriHedyscepe canterburyanaCyphosperma balansae
Physokentia roseaAlloschmidia glabrata
Basselinia velutinaCalyptrocalyx albertisiana
Neoveitchia storckiiCyphokentia macrostachysMoratia cerifera
Lavoixia macrocarpaBrongniartikentia lanuginosaClinosperma bracteale
Linospadix monostachyaLaccospadix australasica
Dransfieldia micranthaHeterospathe elata
Heterospathe longipesActinorhytis calapparia
Chambeyronia macrocarpaKentiopsis oliviformis
Actinokentia divaricataMasoala madagascariensis
Masoala konaRoscheria melanochaetesHydriastele microspadix
Hydriastele chaunostachysCarpoxylon macrospermum
Satakentia liukiuensisBentinckia nicobarica
Clinostigma savoryanumDictyosperma album
Lemurophoenix halleuxiiLoxococcus rupicola
Rhopaloblaste augustaMarojejya darianii
Marojejya insignisDypsis lutescens
Acanthophoenix rubraTectiphiala ferox
Oncosperma tigillariumAreca triandra
Nenga pumilaCyrtostachys renda
Phoenicophorium borsigianumCampecarpus fulcitus
Pelagodoxa henryanaSommieria leucophylla
Iguanura wallichianaPrestoea pubensNeonicholsonia watsonii
Euterpe oleraceaHyospathe macrorhachisCalyptrogyne ghiesbreghtia
Calyptronoma occidentalisGeonoma congestaAsterogyne martiana
Welfia regiaPholidostachys pulchraManicaria sacciferaOrania ravaka
Leopoldinia pulchraOrania lauterbachiana
Allagoptera arenariaSyagrus smithii
Attalea alleniiCocos nucifera
Voanioala gerardiiJubaeopsis caffra
Gastrococos crispaBactris gasipaes
Aiphanes aculeataDesmoncus orthacanthos
Acrocomia aculeataElaeis guineensisReinhardtia simplex
Chamaedorea microspadixGaussia maya
Synechanthus warscezianusHyophorbe lagenicaulis
Wendlandiella gracilisPodococcus barteri
Podococcus barteriRoystonea oleracea
Sclerosperma manniiBeccariophoenix madaensis
Iriartella stenocarpaWettinia hirsuta
Socratea exorrhizaDictyocaryum lamarckianumIriartea deltoidea
Ceroxylon quindiuenseJuania australis
Oraniopsis appendiculataRavenea louvelii
Aphandra nataliaPhytelephas aequatorialis
Ammandra decaspermaPhytelephas macrocarpa
Pseudophoenix sargentiiPseudophoenix vinifera
Maxburrrtia rupicolaRhapis excelsa
Rhapidophyllum hystrixGuihaia argyrataTrachycarpus fortuneiChamaerops humilis
Brahea berlandieriAcoelorraphe wrightii
Serenoa repensColpothrinax wrightii
Pritchardia arecinaPritchardia pacifica
Washingtonia robustaJohannesteijst altifronsPholidocarpus macrocarpus
Pritchardiopsis jeanneneyiLicuala kunstleri
Livistona chinensisCopernicia prunifera
Phoenix reclinataPhoenix dactylifera
Phoenix canariensisBismarckia nobilis
Satranala decussilvaeHyphaene thebaica
Medemia argunBorassodendron machadonis
Borassus flabilliferLodoicea maldivica
Latania verschaffeltiiCorypha umbraculifera
Corypha talieraArenga hookeriana
Arenga undulatifoliaWallichia distica
Caryota mitisCaryota ophiopellis
Kerriodoxa elegansNannorrhops ritchiana
Chuniophoenix nanaHemithrinax compactaThrinax morrisii
Zombia antillarumCoccothrinax argentata
Schippia concolorCryosophila warscewicziana
Itaya amicorumChelyocarpus ulei
Thrinax radiataTrithrinax campestris
Sabal bermudanaSabal minor
Nypa fruticansMetroxylon salomonense
Plectocomia mulleriPigafetta elata
Calamus aruensisSalacca ramosiana
Korthalsia chebEremospatha wendlandiana
Laccosperma acutiflorumOncocalamus tuleyi
Raphia fariniferaMauritia flexuosa
Eugeissona tristis
10 changes
OUTGROUPS
6
1
7
2
SUBFAMILYCALAMOIDEAE
SUBFAMILY NYPOIDEAE
SU
BF
AM
ILY C
OR
YP
HO
IDE
AE
SUBFAMILYCEROXYLOIDEAE
SU
BF
AM
ILY A
RE
CO
I DE
AE
5
3
4

PALM FAMILY PHYLOGENY 25
© 2006 The Linnean Society of London, Botanical Journal of the Linnean Society, 2006, 151, 15–38
et al., 1999; Asmussen et al., 2000; Asmussen &Chase, 2001; Lewis & Doyle, 2001; Hahn, 2002a), not-withstanding a few unusual maximum likelihoodtopologies presented by Hahn (2002a).
SUBFAMILY CORYPHOIDEAE
The matK sequences were particularly useful for theresolution of the subfamily Coryphoideae. However,the addition of more genera probably also contributedto the improved resolution of the relationships com-pared with previous studies (Asmussen & Chase,2001). The new subfamily Coryphoideae (Fig. 1, clade3) is modified only by the inclusion of the tribeCaryoteae, a relationship that can also be foundamong the most-parsimonious solutions emergingfrom many other phylogenetic analyses of moleculardata in the palm family (Uhl et al., 1995; Baker et al.,1999; Asmussen et al., 2000; Asmussen & Chase,2001; Hahn, 2002a). However, until now, a robustmonophyletic group consisting of the subfamilyCoryphoideae and the tribe Caryoteae had onlybeen recovered by Uhl et al. (1995) and Hahn (2002a).
The position of Coryphoideae as sister to all palmsexcept the Calamoideae and Nypa had only previ-ously been recovered by Hahn (2002a) in a highlyreduced taxon sample.
The relationship between the Coryphoid genera iswell resolved, and there are high bootstrap values formany of the subclades (Fig. 1, clade 3). Two of thethree tribes in Dransfield & Uhl’s (1998) classificationof the subfamily Coryphoideae are resolved asmonophyletic: the tribes Borasseae (Fig. 1, clade f)and Phoeniceae (Fig. 1, clade g). The third tribe,Corypheae, is not monophyletic, and just one of thefour constituent subtribes, Sabalinae, is monophyletic(Fig. 1, clade a). The significance of these relationshipsfor classification depends on which nodes are recog-nized and the ranks that they are allocated. In theinterests of nomenclatural stability, we propose thatas many as possible of the current tribes and subtribesare maintained, but major rearrangements of the tribeCorypheae and three of its four subtribes are neededto satisfy the criterion of monophyly (Fig. 1, clade 3).Such a reorganization (Dransfield et al., 2005) resultsin eight tribes (Fig. 1, clades a–h) and a number ofsubtribes, all of which find support among other stud-ies (Uhl et al., 1995; Asmussen et al., 2000; Asmussen& Chase, 2001; Hahn, 2002a).
SUBFAMILIES CEROXYLOIDEAE AND ARECOIDEAE
The bootstrap support for the subfamilies Ceroxy-loideae and Arecoideae is low (63 and 70%, respec-tively). However, the monophyly of both subfamilies isstrongly supported by data from low copy nuclearDNA genes (W. J. Baker, unpubl. data). Furthermore,the Arecoideae is well defined by the floral triad, not-withstanding the floral cluster of the Hyophorbeaeand the presence of triads in Caryoteae. Although theCeroxyloideae is morphologically heterogeneous, it isdefined by all taxa having solitary flowers. The sub-family Phytelephantoideae (sensu Dransfield & Uhl,1998) is highly supported as monophyletic (99% boot-strap support), which is in agreement with other stud-ies (Barfod, 1991; Uhl et al., 1995; Asmussen et al.,2000; Asmussen & Chase, 2001; Hahn, 2002a, b).However, given that Phytelephantoideae is nestedbetween two tribes of Ceroxyloideae, Ceroxyleae and
Figure 2. One cladogram with branch lengths of the 30 000 equally most-parsimonious cladograms resulting from Fitchanalyses of the combined matK, rbcL, rps16 intron and trnL–trnF data sets. A representative of the most resolvedcladograms was chosen. Outgroups were excluded to make the cladogram fit one page. The cladogram is fully resolvedexcept for seven polytomies, which are labelled 1–7. The polytomies labelled 1–5 were present in all 30 000 most-parsimonious cladograms. The clades corresponding to the five subfamilies of the new classification are indicated to theright.
Figure 3. A summary tree showing the relationship of thefive subfamilies in the new classification. The number ofgenera in each subfamily is given below the branches foreach subfamily. Bootstrap percentages that support thesubfamilies are given above the branches and jackknifepercentages and Bremer support values are given belowthe branches.
Calamoideae
Nypoideae
Coryphoideae
Ceroxyloideae
Arecoideae
100
100
97
100
97
85
63
70
21 genera
45 genera
8 genera
112 genera
1 genus100/46
100/28
99/6
100/16
87/3
93/5
63/1
62/1

26 C. B. ASMUSSEN ET AL.
© 2006 The Linnean Society of London, Botanical Journal of the Linnean Society, 2006, 151, 15–38
Cyclospatheae, as sister to the former, the subfamilycan no longer be recognized at the same rank and isplaced as a tribe within the new concept of the sub-family Ceroxyloideae (Fig. 1, clade 4). In the studies ofHahn (2002a, b), the Phytelephantoideae and the tribeCeroxyleae were similarly resolved, but the Cyclos-patheae had a different position. The remainingstudies on palm family phylogenies placed thePhytelephantoideae unresolved as a member of a poly-tomy (Uhl et al., 1995; Baker et al., 1999; Asmussenet al., 2000; Lewis & Doyle, 2002). The inclusion of thetribe Hyophorbeae, formerly of the subfamily Ceroxy-loideae (sensu Dransfield & Uhl, 1998), in the sub-family Arecoideae, as well as the exclusion of the tribeCaryoteae, is in accordance with all previous molecu-lar phylogenies of the palm family (Fig. 1, clade 5). Thelimits of the subfamily Arecoideae require no furtheralterations.
CONCLUSION AND FUTURE PLANS
The addition of matK sequences and more taxa to theprevious palm data sets of Asmussen & Chase (2001)provided the resolution and support required to refinethe subfamily classification of the palm family (Fig. 3).Five subfamilies, all monophyletic, rather than six,are now recognized (Dransfield et al., 2005): (1) thesubfamily Calamoideae, as circumscribed in Drans-field & Uhl (1998); (2) the subfamily Nypoideae, withjust one species, Nypa fruticans; (3) the subfamilyCoryphoideae, comprising those genera included byDransfield & Uhl (1998), with the addition of the tribeCaryoteae; (4) the subfamily Ceroxyloideae, includingthe tribes Cyclospatheae and Ceroxyleae, and thethree phytelephantoid genera; (5) the subfamilyArecoideae, following the concept of Dransfield & Uhl(1998), but with the addition of the tribe Hyophorbeaeand the exclusion of the tribe Caryoteae. This newsubfamily classification will form the backbone of anew edition of Genera Palmarum (Uhl & Dransfield,1987; J. Dransfield, N. W. Uhl, C. B. Asmussen, W. J.Baker, M. M. Harley & C. E. Lewis, unpubl. data).
Within the new subfamilies, high resolution andbootstrap support are recovered in the Calamoideae,Nypoideae, Coryphoideae and, to some extent, theCeroxyloideae. The subfamily Arecoideae is, however,poorly resolved, and the internal nodes generallyreceive low bootstrap support. The low resolution andbootstrap support in Arecoideae are principally aresult of a relatively low number of parsimony-informative characters in this portion of the tree. Themost significant phylogenetic ambiguities remain inthree areas: (1) poorly supported nodes for and somewithin the Ceroxyloideae; (2) poor resolution and sup-port for and within the subfamily Arecoideae; (3) poorsupport and resolution in the clades of coryphoid gen-
era formerly referred to the subtribes Livistoninaeand Thrinacinae. To address these problems and toconsolidate further our findings, we plan to add lowcopy nuclear DNA sequences and additional plastidDNA sequences to this data set and expand the taxonsample to include all genera of palms. Despite theseshortcomings, however, we are confident that the well-supported relationships presented here will be robustto the addition of new data and that our revised sub-family circumscriptions represent significant stepstowards a natural and stable classification of palmsthat will stand the test of time.
ACKNOWLEDGEMENTS
This project was supported by grants from the DanishResearch Council to Conny Asmussen and AndersBarfod. We thank Charlotte Hansen, University ofCopenhagen and Hans Hjort, University of Aarhusfor performing the automated sequencing. We areextremely grateful to all those individuals and insti-tutions who have supported our work by providingmaterial for DNA extraction, especially FairchildTropical Botanic Garden, the Montgomery BotanicalCentre, the Royal Botanic Gardens, Kew, Dr FinnBorchsenius and Phillipp Trénel, University ofAarhus, Dr Carl Lewis, Fairchild Tropical BotanicGarden, and Dr Natalie Uhl, Bailey Hortorium. Wealso wish to thank an anonymous reviewer for helpfulcomments.
REFERENCES
Asmussen CB, Baker WJ, Dransfield J. 2000. Phylogeny ofthe palm family (Arecaceae) based on rps16 intron and trnL–trnF plastid DNA sequences. In: Wilson KL, Morrison DA,eds. Systematics and evolution of monocots. Collingwood,Victoria: CSIRO Publishing, 525–537.
Asmussen CB, Chase MW. 2001. Coding and noncoding plas-tid DNA in palm family systematics. American Journal ofBotany 88: 1103–1117.
Baker WJ, Asmussen CB, Barrow S, Dransfield J,Hedderson TA. 1999. A phylogenetic study of the palmfamily (Palmae) based on chloroplast DNA sequences fromthe trnL–trnF region. Plant Systematics and Evolution 219:111–126.
Baker WJ, Dransfield J, Hedderson TA. 2000a. Phylogeny,character evolution, and a new classification of the Calamoidpalms. Systematic Botany 25: 297–322.
Baker WJ, Hedderson TA, Dransfield J. 2000b. Molecularphylogenetics of subfamily Calamoideae (Palmae) based onnrDNA ITS and cpDNA rps16 intron sequence data. Molec-ular Phylogenetics and Evolution 14: 195–217.
Baker WJ, Hedderson TA, Dransfield J. 2000c. Molecularphylogenetics of Calamus (Palmae) based on 5S nrDNAspacer sequence data. Molecular Phylogenetics and Evolu-tion 14: 218–231.

PALM FAMILY PHYLOGENY 27
© 2006 The Linnean Society of London, Botanical Journal of the Linnean Society, 2006, 151, 15–38
Baker WJ, Loo AHB. 2004. A synopsis of the genus Hydri-astele (Arecaceae). Kew Bulletin 59: 61–68.
Barfod AS. 1991. A monographic study of the subfamilyPhytelephantoideae (Arecaceae). Opera Botanica 105: 1–73.
Chase MW, Soltis DE, Soltis PS, Rudall PJ, Fay MF,Hahn WJ, Sullivan S, Joseph J, Molvray M, Kores PJ,Givnish TJ, Sytsma KJ, Pires JC. 2000. Higher-level sys-tematics of the monocotyledons: an assessment of currentknowledge and a new classification. In: Wilson KL, MorrisonDA, eds. Systematics and evolution of monocots. Colling-wood, Victoria: CSIRO Publishing, 3–16.
Doyle JJ, Doyle JL. 1987. A rapid DNA isolation procedurefrom small quantities of fresh leaf tissue. Phytochemical Bul-letin 19: 11–15.
Dransfield J, Uhl NW. 1998. Palmae. In: Kubitzki K, ed. Thefamilies and genera of vascular plants, IV: Flowering plants,monocotyledons. Berlin: Springer, 306–389.
Dransfield J, Uhl NW, Asmussen CB, Baker WJ,Harley MM, Lewis CE. 2005. A new phylogenetic classifi-cation of the palm family, Arecaceae. Kew Bulletin 60: 559–569.
Fay MF, Bayer C, Alverson WS, de Bruijn AY, Chase MW.1998. Plastid rbcL sequence data indicate a close affinitybetween Diegodendron and Bixa. Taxon 47: 43–50.
Fitch WM. 1971. Toward defining the course of evolution: min-imal change for a specific tree topology. Systematic Zoology20: 406–416.
Hahn WJ. 2002a. A molecular phylogenetic study of thePalmae (Arecaceae) based on atpB, rbcL, and 18S nrDNAsequences. Systematic Biology 51: 92–112.
Hahn WJ. 2002b. A phylogenetic analysis of the Arecoid Lineof palms based on plastid DNA sequence data. MolecularPhylogenetics and Evolution 23: 189–204.
Lewis CE, Doyle JJ. 2001. Phylogenetic utility of the nuclear
gene malate synthase in the palm family (Arecaceae). Molec-ular Phylogenetics and Evolution 19: 409–420.
Lewis CE, Doyle JJ. 2002. A phylogenetic analysis of tribeAreceae (Arecaceae) using two low-copy nuclear genes. PlantSystematics and Evolution 236: 1–17.
Moore HE. 1973. The major groups of palms and their distri-bution. Gentes Herbariorum 11: 27–141.
Oxelman B, Lidén M, Berglund D. 1997. Chloroplast rps16intron phylogeny of the tribe Sileneae (Caryophyllaceae).Plant Systematics and Evolution 206: 393–410.
Shaw J, Lickey EB, Beck JT, Farmer SB, Liu W, Miller J,Siripun KC, Winder CT, Schilling EE, Small RL. 2005.The tortoise and the hare II: relative utility of 21 noncodingchloroplast DNA sequences for phylogenetic analysis. Amer-ican Journal of Botany 91: 142–166.
Steele KP, Vilgalys R. 1994. Phylogenetic analyses ofPolemoniaceae using nucleotide sequences of the plastidgene matK. Systematic Botany 19: 126–142.
Swofford DL. 2002. PAUP*. Phylogenetic analysis using par-simony (*and other methods), Version 4.0. Beta 10. Sunder-land, Massachusetts: Sinauer.
Taberlet P, Gielly L, Pautou G, Bouvet J. 1991. Universalprimers for amplification of three non-coding regions of chlo-roplast DNA. Plant Molecular Biology 17: 1105–1109.
Uhl NW, Dransfield J. 1987. Genera Palmarum: a classifica-tion of palms based on the work of H. E. Moore Jr. Lawrence,Kansas: International Palm Society and L. H. BaileyHortorium.
Uhl NW, Dransfield J, Davis JI, Luckow MA, Hansen KS,Doyle JJ. 1995. Phylogenetic relationships among palms:cladistic analyses of morphological and chloroplast DNArestriction site variation. In: Rudall PJ, Cribb PJ, Cutler DF,Humphries CJ, eds. Monocotyledons: systematics and evolu-tion. Richmond: Royal Botanic Gardens, 623–661.
28 C. B. ASMUSSEN ET AL.
© 2006 The Linnean Society of London, Botanical Journal of the Linnean Society, 2006, 151, 15–38
AP
PE
ND
IX
Vou
cher
an
d da
taba
se i
nfo
rmat
ion
(E
MB
L/G
enB
ank/
DD
BJ
data
base
s) f
or t
he
taxa
use
d in
th
is s
tudy
. H
erba
riu
m a
cron
yms
are
give
n i
n p
aren
thes
es. T
he
firs
tco
lum
n g
ives
th
e su
bfam
ily
clas
sifi
cati
on o
f Dra
nsfi
eld
& U
hl (
1998
) an
d th
e se
con
d co
lum
n g
ives
th
e n
ew s
ubf
amil
y, t
riba
l an
d su
btri
bal c
lass
ifica
tion
of D
ran
sfiel
det
al.
(in
pre
ss)
Su
bfam
ily
clas
sifi
cati
on (
1998
)S
UB
FA
MIL
Y,
trib
e,su
btri
beS
peci
esV
ouch
er i
nfo
rmat
ion
rbcL
rps1
6 in
tron
trn
L–t
rnF
mat
K
CA
LA
MO
IDE
AE
CA
LA
MO
IDE
AE
Eu
geis
son
eae
Eu
geis
son
a tr
isti
s G
riff
.B
aker
501
(K
EP
)A
J404
774
AJ2
4086
9A
J241
278
AM
1145
40L
epid
ocar
yeae
An
cist
roph
ylli
nae
On
coca
lam
us
tule
yiS
un
derl
.S
un
der
lan
d 1
759
(K)
AJ4
0477
6A
J240
871
AJ2
4137
6A
M11
4541
Ere
mos
path
aw
end
lan
dia
na
Bec
c.D
ran
sfiel
d J
D 7
004
(K)
AM
1178
12A
J240
868
AJ2
4127
7A
M11
4542
Lac
cosp
erm
a ac
uti
flor
um
(Bec
c.)
J.D
ran
sf.
Dra
nsfi
eld
JD
700
6 (K
)A
J404
772
AJ2
4086
7A
J241
276
AM
1145
43
Rap
hii
nae
Rap
hia
far
inif
era
(Gae
rtn
.) H
ylan
der
Ru
ther
ford
156
(K
)A
J829
907
AJ2
4218
4A
M11
3612
AM
1145
44
Mau
riti
inae
Mau
riti
a fl
exu
osa
L.f.
Ely
et
al. 1
7 (K
)A
J404
777
AJ2
4087
2A
J241
281
AM
1145
45C
alam
eae
Kor
thal
siin
aeK
orth
alsi
a ch
eb B
ecc.
Bak
er 5
13 (
K)
AM
1101
88A
J242
175
AM
1136
13A
M11
4546
Sal
acci
nae
Sal
acca
ram
osia
na
Mog
ea19
79–4
409
(K)
AM
1101
89A
J242
176
AM
1136
14A
M11
4547
Met
roxy
lin
aeM
etro
xylo
n s
alom
onen
se(W
arb.
) B
ecc.
Zon
a 65
1 (F
TG
)A
M11
0190
AM
1167
69A
M11
3615
AM
1145
48
Pig
afet
tin
aeP
igaf
etta
ela
ta (
Mar
t.)
H.W
endl
.B
aker
508
(K
)A
J829
897
AJ2
4217
1A
M11
3616
AM
1145
49
Ple
ctoc
omii
nae
Ple
ctoc
omia
mu
ller
i B
lum
eB
aker
563
(K
)A
J829
899
AJ2
4216
8A
M11
3617
AM
1145
50C
alam
inea
eC
alam
us
aru
ensi
s B
ecc.
Dra
nsfi
eld
JD
757
1 (K
)A
J404
775
AJ2
4087
0A
J241
279
AM
1145
51N
YP
OID
EA
EN
YP
OID
EA
EN
ypa
fru
tica
ns
Wu
rmb
N.W
. Uh
l s.
n.
(BH
)A
J404
778
AJ2
4087
3A
J241
282
AM
1145
52C
OR
YP
HO
IDE
AE
CO
RY
PH
OID
EA
ES
abal
eae
Sab
al b
erm
ud
ana
L.H
.Bai
ley
1982
–560
2 (K
)A
J404
766
AJ2
4086
2A
J241
271
AM
1145
53
Sab
al m
inor
(N
.J.J
acqu
in)
Per
soon
1964
–454
0 (K
)A
M11
0191
AM
1167
70A
M11
3618
AM
1145
54

PALM FAMILY PHYLOGENY 29
© 2006 The Linnean Society of London, Botanical Journal of the Linnean Society, 2006, 151, 15–38
Cry
osop
hil
eae
Sch
ippi
a co
nco
lor
Bu
rret
Bak
er 9
95 (
FT
G)
AJ4
0474
9A
J404
924
AJ4
0489
1A
M11
4555
Tri
thri
nax
cam
pest
ris
(Bu
rmei
st.)
Dru
de &
G
rise
b.
1938
–270
03 (
K)
AJ4
0474
5A
J240
844
AJ2
4125
3A
M11
4556
Zom
bia
anti
llar
um
(D
esc.
)L
.H.B
aile
yR
. San
der
s 17
63 (
FT
G)
AM
1101
92A
M11
6771
AM
1136
19A
M11
4557
Coc
coth
rin
ax a
rgen
tata
(Jac
q.)
L.H
.Bai
ley
1973
–126
08 (
K)
AJ4
0475
1A
J240
848
AJ2
4125
7A
M11
4558
Hem
ith
rin
ax c
ompa
cta
(Gri
seb.
& H
.Wen
dl.)
H
ook.
f.
C.E
. Lew
is 0
2–07
2 (H
AJB
)A
J829
869
AM
1167
72A
M11
3620
AM
1145
59
Th
rin
ax m
orri
sii
H.W
endl
.J.
Ron
cal
043
(FT
G)
AM
1101
93A
M11
6773
AM
1136
21A
M11
4560
Th
rin
ax r
adia
ta L
odd.
ex
Sch
ult
. & S
chu
lt.f.
1933
–047
031
(K)
AJ4
0475
0A
M11
6774
AJ2
4125
6A
M11
4561
Ch
elyo
carp
us
ule
i D
amm
erB
alsl
ev 6
407
(AA
U)
AJ4
0474
6A
J240
845
AJ2
4125
4A
M11
4562
Cry
osop
hil
aw
arsc
ewic
zian
a B
artl
ett
1991
−109
2 (K
)A
J404
747
AJ2
4084
6A
J241
255
AM
1145
63
Itay
a am
icor
um
H.E
.Moo
reB
aker
990
(F
TG
)A
J404
748
AJ4
0492
3A
J404
890
AM
1145
64P
hoe
nic
eae
Ph
oen
ix r
ecli
nat
a Ja
cq.
Goy
der
et
al. 3
928
(K)
AJ4
0476
7A
J240
863
AJ2
4127
2A
M11
4565
Ph
oen
ix c
anar
ien
sis
Ch
abau
dB
arro
w 7
5 (K
)A
M11
0194
AM
1167
75A
M11
3622
AM
1145
66
Ph
oen
ix d
acty
life
ra L
.B
arro
w 7
7 (K
)A
M11
0195
AM
1167
76A
M11
3623
AM
1145
67L
ivis
ton
eae
Rh
apid
inae
Ch
amae
rops
hu
mil
is L
.B
arro
w 7
6 (K
)A
J404
754
AM
1167
77A
J241
260
AM
1145
68G
uih
aia
argy
rata
(S.
K.L
ee&
F.N
.Wei
) S.
K.L
ee,
F.N
.Wei
& J
.Dra
nsf
.
1984
–447
0 (K
)A
J404
755
AJ2
4085
2A
J241
261
AM
1145
69
Tra
chyc
arpu
s fo
rtu
nei
(Hoo
k.)
H.W
endl
.D
ran
sfiel
d s
.n.
(K)
AJ4
0475
2A
J404
925
AJ4
0489
2A
M11
4570
Rh
apid
oph
yllu
m h
ystr
ix(P
urs
h) H
.Wen
dl. &
Dru
de01
3–67
0130
1 (K
)A
J404
753
AM
1167
78A
J241
259
AM
1145
71
Max
burr
etia
ru
pico
la(R
idle
y) F
urt
ado
C.E
. Lew
is 0
2–08
0 (F
TG
)A
J829
884
AM
1167
79A
M11
3624
AM
1145
72
Rh
apis
exc
elsa
Hen
ry e
xR
ehde
r19
87–2
573
(K)
AJ4
0475
6A
J240
853
AJ2
4126
2A
M11
4573
Su
bfam
ily
clas
sifi
cati
on (
1998
)S
UB
FA
MIL
Y,
trib
e,su
btri
beS
peci
esV
ouch
er i
nfo
rmat
ion
rbcL
rps1
6 in
tron
trn
L–t
rnF
mat
K

30 C. B. ASMUSSEN ET AL.
© 2006 The Linnean Society of London, Botanical Journal of the Linnean Society, 2006, 151, 15–38
Liv
isto
nin
aeL
ivis
ton
a ch
inen
sis
(Jac
q.)
R.B
r. ex
Mar
t.19
82–5
601
(K)
AJ4
0475
7A
J240
854
AJ2
4126
3A
M11
4574
Lic
ual
a ku
nst
leri
Bec
c.19
85–1
497
(K)
AJ4
0475
9A
J240
856
AJ2
4126
5A
M11
4575
Joh
ann
este
ijsm
ann
iaal
tifr
ons
(Rei
chb.
f. &
Z
oll.)
H.E
.Moo
re
1985
–151
5 (K
)A
J404
758
AJ2
4085
5A
J241
264
AM
1145
76
Ph
olid
ocar
pus
mac
roca
rpu
s B
ecc.
B. C
hee
473
32 (
KE
P)
AJ8
2989
4A
M11
6780
AM
1136
25A
M11
4577
Pri
tch
ard
iops
is j
ean
nen
eyi
Bec
c.P
inta
ud
315
(P
)A
M11
0196
AM
1167
81A
M11
3626
AM
1145
78
Un
plac
ed g
ener
a in
Liv
isto
nea
eA
coel
orra
phe
wri
ghti
iH
.Wen
dl.
D. B
ogle
r 12
47 (
FT
G)
AM
1101
97A
M11
6782
AM
1136
27A
M11
4579
Bra
hea
ber
lan
die
ri B
artl
ett
C.E
. Lew
is 0
3–01
1 (F
TG
)A
M11
0198
AM
1167
83A
M11
3628
AM
1145
80C
olpo
thri
nax
wri
ghti
i H
.W
endl
.R
. Rac
ine
41 (
FT
G)
AJ8
2986
2A
M11
6784
AM
1136
29A
M11
4581
Cop
ern
icia
pru
nif
era
(Mil
l.) H
.E.M
oore
C.E
. Lew
is 0
3–01
2 (F
TG
)A
M11
0199
AM
1167
85A
M11
3630
AM
1145
82
Pri
tch
ard
ia a
reci
na
Bec
c.B
aker
118
3 (K
)A
J829
905
AM
1167
86A
M11
3631
AM
1145
83P
ritc
har
dia
pac
ifica
See
m.
& H
.Wen
dl.
1952
–355
01 (
K)
AM
1102
00A
M11
6787
AM
1136
32A
M11
4584
Ser
enoa
rep
ens
(Bar
tram
)J.
K.S
mal
l19
58–6
6102
(K
)A
J404
760
AM
1167
88A
J241
266
AM
1145
85
Was
hin
gton
ia r
obu
sta
H.W
endl
.C
.E. L
ewis
03–
013
(FT
G)
AM
1102
01A
M11
6789
AM
1136
33A
M11
4586
Ch
un
iop
hoe
nic
eae
Ch
un
ioph
oen
ix n
ana
Bu
rret
1986
–301
8 (K
)A
J404
764
AJ2
4086
0A
J241
269
AM
1145
87K
erri
odox
a el
egan
sJ.
Dra
nsf
.19
87–2
685
(K)
AJ4
0476
5A
J240
861
AJ2
4127
0A
M11
4588
Nan
nor
rhop
s ri
tch
ian
aH
.Wen
dl.
1990
–113
2 (K
)A
J404
763
AJ2
4085
9A
J241
268
AM
1145
89
AR
EC
OID
EA
EC
aryo
teae
Car
yota
mit
is L
our.
464–
85.0
5037
(K
)A
J404
790
AJ2
4088
3A
J241
292
AM
1145
90C
aryo
ta o
phio
pell
is D
owe
Bak
er 9
89 (
FT
G)
AJ4
0479
1A
J404
930
AJ4
0489
7A
M11
4591
Are
nga
hoo
keri
ana
(Bec
c.)
Wh
itm
ore
085–
85.0
1492
(K
)A
J404
788
AJ2
4088
2A
J241
291
AM
1145
92
Are
nga
un
du
lati
foli
a B
ecc.
Bak
er 9
87 (
FT
G)
AJ4
0478
9A
J404
929
AJ4
0489
6A
M11
4593
Wal
lich
ia d
isti
cha
T.A
nde
rs.
L-8
0.07
70 (
BH
)A
J404
792
AJ2
4088
4A
J241
293
AM
1145
94
Su
bfam
ily
clas
sifi
cati
on (
1998
)S
UB
FA
MIL
Y,
trib
e,su
btri
beS
peci
esV
ouch
er i
nfo
rmat
ion
rbcL
rps1
6 in
tron
trn
L–t
rnF
mat
K
Ap
pen
dix
Con
tin
ued
PALM FAMILY PHYLOGENY 31
© 2006 The Linnean Society of London, Botanical Journal of the Linnean Society, 2006, 151, 15–38
CO
RY
PH
OID
EA
EC
oryp
hea
eC
oryp
ha
um
brac
uli
fera
L.
1954
–353
01 (
K)
AJ4
0476
1A
J240
858
AJ2
4126
7A
M11
4595
Cor
yph
a ta
lier
a R
oxb.
Bak
er 9
84 (
FT
G)
AJ4
0476
2A
J404
926
AJ4
0489
3A
M11
4596
Bor
asse
aeH
yph
aen
inae
Bis
mar
ckia
nob
ilis
Hil
debr
. & H
.Wen
dl.
Bay
ton
R.P
. 43
(K)
AJ8
2985
2A
M11
6790
AM
1136
34A
M11
4597
Sat
ran
ala
dec
uss
ilva
eB
een
tje
& J
.Dra
nsf
.B
een
tje
& D
ran
sfiel
d 4
810
(K)
AJ4
0477
1A
J240
866
AJ2
4127
5A
M11
4598
Hyp
hae
ne
theb
aica
Mar
t.19
94–3
803
(K)
AJ4
0477
0A
J240
865
AJ2
4127
4A
M11
4599
Med
emia
arg
un
Wu
ert.
ex
H.W
endl
.B
aker
986
(F
TG
)A
J829
885
AM
1167
91A
M11
3635
AM
1146
00
Lat
anii
nae
Lat
ania
ver
sch
affe
ltii
Lem
.J.
Ron
cal
64 (
FT
G)
AF
8298
78A
M11
6792
AM
1136
36A
M11
4601
Lod
oice
a m
ald
ivic
a(J
.F.G
mel
.) P
ers.
1994
–323
1 (K
)A
J404
769
AJ2
4086
4A
J241
273
AM
1146
02
Bor
asso
den
dro
nm
ach
adon
is (
Rid
l.) B
ecc.
1989
–339
4 (K
)A
J404
768
AJ4
0492
7A
J404
894
AM
1146
03
Bor
assu
s fl
abel
life
r L
.W
ilki
n, S
ud
dee
& T
hap
yai
1160
(K
)A
M11
0202
AM
1167
93A
M11
3637
AM
1146
04
CE
RO
XY
LO
IDE
AE
CE
RO
XY
LO
IDE
AE
Cyc
losp
ath
eae
Pse
ud
oph
oen
ix s
arge
nti
iH
.Wen
dl.
FT
G 8
2–44
1C (
BH
)A
J404
779
AJ2
4087
4A
J241
283
AM
1146
05
Pse
ud
oph
oen
ix v
inif
era
(Mar
t.)
Bec
c.B
aker
100
2 (F
TG
)A
J404
780
AJ4
0492
8A
J404
895
AM
1146
06
Cer
oxyl
eae
Cer
oxyl
on q
uin
diu
ense
(H.K
arst
) H
.Wen
dl.
1976
–116
0 (K
)A
J404
781
AJ2
4087
5A
J241
284
AM
1146
07
Juan
ia a
ust
rali
s D
rude
ex
Hoo
k.f.
P. T
rén
el 4
(A
AU
)A
J829
874
AM
1167
94A
M11
3638
AM
1146
08
Ora
nio
psis
app
end
icu
lata
(F.M
.Bai
ley)
J.D
ran
sf.,
A.K
.Irv
ine
& N
.W.U
hl
1988
–227
(K
)A
J404
782
AJ2
4087
6A
J241
285
AM
1146
09
Rav
enea
lou
veli
i B
een
tje
1988
–236
9 (K
)A
J404
783
AJ2
4087
7A
J241
286
AM
1146
10
Su
bfam
ily
clas
sifi
cati
on (
1998
)S
UB
FA
MIL
Y,
trib
e,su
btri
beS
peci
esV
ouch
er i
nfo
rmat
ion
rbcL
rps1
6 in
tron
trn
L–t
rnF
mat
K

32 C. B. ASMUSSEN ET AL.
© 2006 The Linnean Society of London, Botanical Journal of the Linnean Society, 2006, 151, 15–38
PH
YT
EL
EP
HA
NT-
OID
EA
EP
hyt
elep
hea
eA
mm
and
ra d
ecas
perm
aO
.F.C
ook
L-7
7.08
309
(BH
)A
J404
838
AJ4
0495
5A
J404
922
AM
1146
11
Aph
and
ra n
atal
ia (
A.J
.H
ende
rson
& B
alsl
ev)
Bar
fod
Bak
er 9
85 (
K)
AJ4
0483
7A
J404
954
AJ4
0492
1A
M11
4612
Ph
ytel
eph
as a
equ
ator
iali
sS
pru
ce19
93−9
4 (K
)A
J404
835
AJ2
4090
8A
J241
317
AM
1146
13
Ph
ytel
eph
as m
acro
carp
aR
uiz
& P
av.
1992
–248
0 (K
)A
J404
836
AJ2
4090
7A
J241
316
AM
1146
14
AR
EC
OID
EA
EA
RE
CO
IDE
AE
Iria
rtee
aeIr
iart
ella
ste
noc
arpa
Bu
rret
B.M
illá
n,
J.C
.Pin
tau
d,
C.V
egas
728
(U
NM
SM
)A
M11
0203
AM
1167
95A
M11
3639
AM
1146
15
Dic
tyoc
aryu
mla
mar
ckia
nu
m (
Mar
t.)
H.W
endl
.
Asm
uss
en 1
11 (
CP
)A
M11
0204
AM
1167
96A
M11
3640
AM
1146
16
Iria
rtea
del
toid
ea R
uiz
&P
av.
Hen
der
son
042
(B
H)
AJ4
0479
3A
J240
885
AJ2
4129
4A
M11
4617
Soc
rate
a ex
orrh
iza
H.W
endl
.B
aker
992
(F
TG
)A
M11
0205
AM
1167
97A
M11
3641
AM
1146
18
Wet
tin
ia h
irsu
ta B
urr
etB
aker
991
(F
TG
)A
J404
794
AJ4
0493
1A
J404
898
AM
1146
19
CE
RO
XY
LO
IDE
AE
Ch
amae
dor
eeae
Hyo
phor
be l
agen
icau
lis
(L.H
.Bai
ley)
H
.E.M
oore
1983
–674
(K
)A
J404
785
AJ2
4087
9A
J241
288
AM
1146
20
Wen
dla
nd
iell
a gr
acil
isD
amm
. va
r. po
lycl
ada
(Bu
rret
) A
.Hen
ders
on
Zon
a 75
4 (F
TG
)A
M11
0206
AM
1167
98A
M11
3642
AM
1146
21
Syn
ech
anth
us
war
scew
iczi
anu
s H
.Wen
dl.
Kn
ud
sen
& A
smu
ssen
64
0 (A
AU
)A
J404
786
AJ2
4088
0A
J241
787
AM
1146
22
Ch
amae
dor
ea m
icro
spad
ixB
urr
etB
H 6
0–81
1 (B
H)
AJ4
0478
7A
J240
881
AJ2
4129
0A
M11
4623
Gau
ssia
may
a (O
.F.C
ook)
H.J
.Qu
ero
& R
ead
1958
–801
01 (
K)
AJ4
0478
4A
J240
878
AJ2
4128
7A
M11
4624
AR
EC
OID
EA
EP
odoc
occe
aeP
odoc
occu
s ba
rter
i G
.Man
n&
H.W
endl
.R
eits
ma
2840
(B
H)
AM
1102
07A
J240
886
AJ2
4129
5A
M11
4625
Pod
ococ
cus
bart
eri
G.M
ann
& H
.Wen
dl.
Su
nd
erla
nd
180
3 (K
)A
M11
0208
AM
1167
99A
M11
3643
AM
1146
26
Su
bfam
ily
clas
sifi
cati
on (
1998
)S
UB
FA
MIL
Y,
trib
e,su
btri
beS
peci
esV
ouch
er i
nfo
rmat
ion
rbcL
rps1
6 in
tron
trn
L–t
rnF
mat
K
Ap
pen
dix
Con
tin
ued

PALM FAMILY PHYLOGENY 33
© 2006 The Linnean Society of London, Botanical Journal of the Linnean Society, 2006, 151, 15–38
Ora
nie
aeO
ran
ia l
aute
rbac
hia
na
Bec
c.L
-78.
0662
(B
H)
AJ4
0479
6A
J240
887
AJ2
4129
6A
M11
4627
Ora
nia
rav
aka
H.B
een
tje
Dra
nsfi
eld
JD
773
1 (K
)A
M11
0209
AM
1168
00A
M11
3644
AM
1146
28S
cler
ospe
rmea
eS
cler
ospe
rma
man
nii
H.W
endl
.S
un
der
lan
d T
CH
S 1
794
(K)
AJ4
0482
3A
J404
948
AJ4
0491
5A
M11
4629
Roy
ston
eae
Roy
ston
ea o
lera
cea
(Jac
q.)
O.F
.Coo
k19
63–5
7401
(K
)A
J404
805
AJ4
0493
6A
J404
903
AM
1146
30
Rei
nh
ard
tiea
eR
ein
har
dti
a si
mpl
ex(H
.Wen
dl.)
Dru
de e
xD
amm
er
1988
–366
(K
)A
J404
799
AJ4
0493
3A
J404
900
AM
1146
31
Coc
osea
eA
ttal
ein
aeB
ecca
riop
hoe
nix
mad
agas
cari
ensi
s Ju
m.
et H
.Per
rier
1989
–353
2 (K
)A
J404
826
AJ4
0495
1A
J404
918
AM
1146
32
Juba
eops
is c
affr
a B
ecc.
T.B
. Sik
hak
han
e 13
9 (N
H)
AJ8
2987
6A
M11
6801
AM
1136
45A
M11
4633
Voa
nio
ala
gera
rdii
J.
Dra
nsf
.D
ran
sfiel
d J
D 6
389
(K)
AM
1102
10A
M11
6802
AM
1136
46A
M11
4634
All
agop
tera
are
nar
ia(G
omes
) K
un
tze
BH
79.
312
(BH
)A
J404
828
AJ2
4090
2A
J241
311
AM
1146
35
Att
alea
all
enii
H.E
.Moo
reK
nu
dse
n &
Asm
uss
en
612
(AA
U)
AJ4
0482
9A
J240
903
AJ2
4131
2A
M11
4636
Coc
os n
uci
fera
L.
1968
–448
0 (K
)A
M11
0211
AM
1168
03A
M11
3647
AM
1146
37S
yagr
us
smit
hii
(H.E
.Moo
re)
Gla
ssm
.B
alsl
ev 6
404
(AA
U)
AJ4
0482
7A
J240
901
AJ2
4131
0A
M11
4638
Bac
trid
inae
Acr
ocom
ia a
cule
ata
(Jac
q.)
Lod
d. e
x M
art.
Bak
er 1
000
(FT
G)
AM
1102
12A
M11
6804
AM
1136
48A
M11
4639
Acr
ocom
ia c
risp
a (K
un
th)
C.F
.Bak
er e
x B
ecc.
J. R
onca
l 79
(F
TG
)A
M11
0213
AM
1168
05A
M11
3649
AM
1146
40
Aip
han
es a
cule
ata
Wil
ld.
Bor
chse
niu
s 59
9 (A
AU
)A
J404
831
AJ4
0495
3A
J404
920
AM
1146
41B
actr
is g
asip
aes
Ku
nth
C.E
. Lew
is 0
2–02
7 (F
TG
)A
M11
0214
AM
1168
06A
M11
3650
AM
1146
42D
esm
oncu
s or
thac
anth
osM
art.
Zon
a 62
0 (F
TG
)A
M11
0215
AM
1168
07A
M11
3651
AM
1146
43
Ela
eidi
nae
Ela
eis
guin
een
sis
Jacq
.19
87–2
16 (
K)
AJ4
0483
0A
J404
952
AJ4
0491
9A
M11
4644
Man
icar
ieae
Man
icar
ia s
acci
fera
Gae
rtn
.C
.E. L
ewis
03–
010
(FT
G)
AJ4
0479
7A
J240
888
AJ2
4129
7A
M11
4645
Su
bfam
ily
clas
sifi
cati
on (
1998
)S
UB
FA
MIL
Y,
trib
e,su
btri
beS
peci
esV
ouch
er i
nfo
rmat
ion
rbcL
rps1
6 in
tron
trn
L–t
rnF
mat
K

34 C. B. ASMUSSEN ET AL.
© 2006 The Linnean Society of London, Botanical Journal of the Linnean Society, 2006, 151, 15–38
Eu
terp
eae
Hyo
spat
he
mac
rorh
ach
isB
urr
etB
alsl
ev 6
421
(AA
U)
AJ4
0480
4A
J240
891
AJ2
4130
0A
M11
4646
Eu
terp
e ol
erac
ea M
art.
L-7
0.00
17 (
BH
)A
J404
802
AJ2
4088
9A
J241
298
AM
1146
47P
rest
oea
pube
ns
H.E
.Moo
reK
nu
dse
n &
Asm
uss
en
619
(AA
U)
AM
1102
16A
M11
6808
AM
1136
52A
M11
4648
Neo
nic
hol
son
ia w
atso
nii
Dam
mer
L-8
1.03
03 (
BH
)A
J404
803
AJ2
4089
0A
J241
299
AM
1146
49
Geo
nom
atea
eW
elfi
a re
gia
H. W
endl
.K
nu
dse
n &
Asm
uss
en
607
(AA
U)
AJ8
2991
7A
M11
6809
AM
1136
53A
M11
4650
Ph
olid
osta
chys
pu
lch
raH
.Wen
dl. e
x B
urr
etK
nu
dse
n &
Asm
uss
en
613
(AA
U)
AM
1102
17A
M11
6810
AM
1136
54A
M11
4651
Cal
yptr
ogyn
egh
iesb
regh
tian
a (L
inde
n&
H.W
endl
.) H
.Wen
dl.
Kn
ud
sen
& A
smu
ssen
62
7 (A
AU
)A
M11
0218
AM
1168
11A
M11
3655
AM
1146
52
Cal
yptr
onom
a oc
cid
enta
lis
(Sw
.) H
.E.M
oore
FT
G 7
1375
D (
FT
G)
AJ4
0483
2A
J240
904
AJ2
4131
3A
M11
4653
Ast
erog
yne
mar
tian
a(H
.Wen
dl.)
H.W
endl
. ex
Dru
de
L-8
1.02
84 (
BH
)A
J404
833
AJ2
4090
5A
J241
314
AM
1146
54
Geo
nom
a co
nge
sta
H.W
endl
. ex
Spr
uce
Bor
chse
niu
s 34
8 (A
AU
)A
M11
0219
AJ2
4090
6A
J241
315
AM
1146
55
Leo
pol
din
ieae
Leo
pold
inia
pu
lch
ra M
art.
Rom
ero
3060
(V
EN
)A
J404
798
AJ4
0493
2A
J404
899
AM
1146
56P
elag
odox
eae
Pel
agod
oxa
hen
ryan
a B
ecc.
1988
−293
5 (K
)A
J829
892
AM
1168
12A
M11
3656
AM
1146
57S
omm
ieri
a le
uco
phyl
laB
ecc.
1992
–347
7 (K
)A
M11
0220
AM
1168
13A
M11
3657
AM
1146
58
Are
ceae
Arc
hon
toph
oen
icin
aeA
ctin
orh
ytis
cal
appa
ria
H.W
endl
. & D
rude
C.E
. Lew
is 9
7–01
1 (F
TG
)A
J829
847
AM
1168
14A
M11
3658
AM
1146
59
Arc
hon
toph
oen
ix p
urp
ure
aH
odel
& D
owe
Pin
tau
d 4
92 (
TL
)A
J404
806
AJ4
0493
7A
J404
904
AM
1146
60
Act
inok
enti
a d
ivar
icat
a(B
ron
gn.)
Dam
mer
Pin
tau
d 3
51 (
K)
AM
1102
21A
M11
6815
AM
1136
59A
M11
4661
Ch
ambe
yron
ia m
acro
carp
a(B
ron
gn. V
ieil
l. ex
Bec
c.P
inta
ud
361
(K
, NY
)A
M11
0222
AM
1168
16A
M11
3660
AM
1146
62
Ken
tiop
sis
oliv
ifor
mis
(Bro
ngn
. & G
ris)
Bro
ngn
.P
inta
ud
358
(K
, NO
U, N
Y)
AJ4
0480
9A
J240
892
AJ2
4178
8A
M11
4663
Su
bfam
ily
clas
sifi
cati
on (
1998
)S
UB
FA
MIL
Y,
trib
e,su
btri
beS
peci
esV
ouch
er i
nfo
rmat
ion
rbcL
rps1
6 in
tron
trn
L–t
rnF
mat
K
Ap
pen
dix
Con
tin
ued

PALM FAMILY PHYLOGENY 35
© 2006 The Linnean Society of London, Botanical Journal of the Linnean Society, 2006, 151, 15–38
Are
cin
aeA
reca
tri
and
ra R
oxb.
1984
–229
5 (K
)A
J404
819
AJ4
0494
5A
J404
912
AM
1146
64N
enga
pu
mil
a (M
art.
)H
.Wen
dl.
var.
pach
ysta
chys
(B
lum
e)F
ern
ando
Bak
er 9
94 (
FT
G)
AJ4
0481
8A
J404
944
AJ4
0491
1A
M11
4665
Bas
seli
nii
nae
All
osch
mid
ia g
labr
ata
(Bec
c.)
H.E
.Moo
reP
inta
ud
468
(K
)A
J829
849
AM
1168
17A
M11
3661
AM
1146
66
Bas
seli
nia
vel
uti
na
Bec
c.P
inta
ud
365
(P
)A
M11
0223
AM
1168
18A
M11
3662
AM
1146
67C
ampe
carp
us
fulc
itu
s(B
ron
gn.)
H.W
endl
. ex
Bec
c.
Pin
tau
d 4
83 (
TL
)A
M11
0224
AM
1168
19A
M11
3663
AM
1146
68
Cyp
hop
hoe
nix
nu
cele
H.E
.Moo
reP
inta
ud
372
(K
, NO
U, N
Y,
P)
AJ4
0482
1A
M11
6820
AJ2
4130
9A
M11
4669
Cyp
hos
perm
a ba
lan
sae
(Bro
ngn
.) H
.Wen
dl.
ex S
alom
on
Pin
tau
d 4
91 (
TL
)A
M11
0225
AM
1168
21A
M11
3664
AM
1146
70
Ph
ysok
enti
a ro
sea
H.E
.Moo
reP
inta
ud
452
(T
L)
AJ8
2989
6A
M11
6822
AM
1136
65A
M11
4671
Vei
llon
ia a
lba
H.E
.Moo
reP
inta
ud
470
(T
L)
AM
1102
26A
M11
6823
AM
1136
66A
M11
4672
Car
poxy
lin
aeC
arpo
xylo
nm
acro
sper
mu
m H
.Wen
dl.
& D
rude
Zon
a 72
2 (F
TG
)A
J829
859
AM
1168
24A
M11
3667
AM
1146
73
Sat
aken
tia
liu
kiu
ensi
s(H
atu
sim
a) H
.E.M
oore
Pin
tau
d 4
46 (
K)
AM
1102
27A
M11
6825
AM
1136
68A
M11
4674
Neo
veit
chia
sto
rcki
i B
ecc.
Ron
cal
73A
J829
888
AM
1168
26A
M11
3669
AM
1146
75C
lin
ospe
rmat
inae
Cyp
hok
enti
a m
acro
stac
hya
Bro
ngn
.P
inta
ud
482
(T
L)
AJ8
2986
4A
M11
6827
AM
1136
70A
M11
4676
Mor
atia
cer
ifer
aH
.E.M
oore
Pin
tau
d 4
69 (
TL
)A
M11
0228
AM
1168
28A
M11
3671
AM
1146
77
Lav
oixi
a m
acro
carp
aH
.E.M
oore
Pin
tau
d 3
64 (
P)
AJ8
2987
9A
M11
6829
AM
1136
72A
M11
4678
Bro
ngn
iart
iken
tia
lan
ugi
nos
a H
.E.M
oore
Pin
tau
d 3
68 (
P)
AJ8
2985
4A
M11
6830
AM
1136
73A
M11
4679
Cli
nos
perm
a br
acte
ale
(Bro
ngn
.) B
ecc.
Pin
tau
d 3
49 (
K, N
Y)
AJ8
2986
1A
M11
6831
AM
1136
74A
M11
4680
Su
bfam
ily
clas
sifi
cati
on (
1998
)S
UB
FA
MIL
Y,
trib
e,su
btri
beS
peci
esV
ouch
er i
nfo
rmat
ion
rbcL
rps1
6 in
tron
trn
L–t
rnF
mat
K

36 C. B. ASMUSSEN ET AL.
© 2006 The Linnean Society of London, Botanical Journal of the Linnean Society, 2006, 151, 15–38
Dyp
sidi
nae
Dyp
sis
lute
scen
s (H
.Wen
dl.)
Bee
ntj
e &
J.D
ran
sf.
1978
–119
6 (K
)A
J404
800
AJ4
0493
4A
J404
901
AM
1146
81
Lem
uro
phoe
nix
hal
leu
xii
J.D
ran
sf.
Bak
er 1
008
(K)
AM
1102
29A
J404
935
AJ4
0490
2A
M11
4682
Mar
ojej
ya d
aria
nii
J.D
ran
sf. &
N.U
hl
Bak
er 9
98 (
K)
AJ4
0482
5A
J404
950
AJ4
0491
7A
M11
4683
Mar
ojej
ya i
nsi
gnis
Hu
mbe
rtB
aker
101
6 (K
)A
M11
0230
AM
1168
32A
M11
3675
AM
1146
84
Mas
oala
mad
agas
cari
ensi
sJu
m.
1992
–355
2 (K
)A
J404
824
AJ4
0494
9A
J404
916
AM
1146
85
Mas
oala
kon
a B
een
tje
Bak
er 1
038
(K)
AM
1102
31A
M11
6833
AM
1136
76A
M11
4686
Lin
ospa
dici
nae
Cal
yptr
ocal
yxal
bert
isia
nu
s B
ecc.
Bak
er 1
109
(K)
AM
1102
32A
M11
6834
AM
1136
77A
M11
4687
Lin
ospa
dix
mon
osta
chya
(Mar
t.)
H. W
endl
.F
red
riks
en e
t al
. C-2
10 (
C)
AJ4
0481
1A
J404
941
AJ4
0490
8A
M11
4688
Lac
cosp
adix
au
stra
lasi
caH
.Wen
dl. &
Dru
deL
-79.
0850
(B
H)
AJ4
0481
2A
J240
895
AJ2
4130
4A
M11
4689
On
cosp
erm
atin
aeO
nco
sper
ma
tigi
llar
ium
(Jac
k) R
idl.
R. S
and
ers
1768
(F
TG
)A
M11
0233
AM
1168
35A
M11
3678
AM
1146
90
Aca
nth
oph
oen
ix r
ubr
a(B
ory)
H.W
endl
.C
.E. L
ewis
98–
067
(BH
)A
M11
0234
AM
1168
36A
M11
3679
AM
1146
91
Tec
tiph
iala
fer
oxH
.E.M
oore
C.E
. Lew
is 9
8–07
0 (B
H)
AJ8
2991
4A
M11
6837
AM
1136
80A
M11
4692
Pty
chos
perm
atin
aeP
tych
ospe
rma
mac
arth
uri
i(H
.Wen
dl. e
x H
.J.V
eitc
h)
H.W
endl
. ex
Hoo
k. f
.
Zon
a 86
9 (F
TG
)A
M11
0235
AM
1168
38A
M11
3681
AM
1146
93
Pon
apea
led
erm
ann
ian
aB
ecc.
Zon
a 87
8 (F
TG
)A
J829
903
AM
1168
39A
M11
3682
AM
1146
94
Bal
aka
seem
ann
ii B
ecc.
L-6
9.04
04 (
BH
)A
J404
814
AJ2
4089
6A
J241
305
AM
1146
95V
eitc
hia
are
cin
a B
ecc.
Bak
er 1
003
(FT
G)
AJ4
0481
3A
J404
942
AJ4
0490
9A
M11
4696
Car
pen
tari
a ar
cum
inat
a(H
.Wen
dl. &
Dru
de)
Bec
c.B
aker
999
(F
TG
)A
J829
858
AM
1168
40A
M11
3683
AM
1146
97
Wod
yeti
a bi
furc
ata
A.K
.Irv
ine
FT
G 8
5–11
113
(FT
G)
AM
1102
36A
M11
6841
AM
1136
84A
M11
4698
Bra
ssio
phoe
nix
sch
um
ann
ii (
Bec
c.)
Ess
igL
-72.
0031
(B
H)
AJ4
0481
5A
J240
897
AJ2
4130
6A
M11
4699
Pty
choc
occu
s pa
rad
oxu
s(S
chef
f.) B
ecc.
Bak
er 5
72 (
K)
AJ8
2990
6A
M11
6842
AM
1136
85A
M11
4700
Su
bfam
ily
clas
sifi
cati
on (
1998
)S
UB
FA
MIL
Y,
trib
e,su
btri
beS
peci
esV
ouch
er i
nfo
rmat
ion
rbcL
rps1
6 in
tron
trn
L–t
rnF
mat
K
Ap
pen
dix
Con
tin
ued

PALM FAMILY PHYLOGENY 37
© 2006 The Linnean Society of London, Botanical Journal of the Linnean Society, 2006, 151, 15–38
Rh
opal
osty
lidi
nae
Rh
opal
osty
lis
bau
eri
H.W
endl
. & D
rude
va
r. ba
uer
i
Pin
tau
d 3
84 (
NY
)A
J404
808
AJ4
0493
9A
J404
906
AM
1147
01
Hed
ysce
pe c
ante
rbu
ryan
a(C
.Moo
re &
F.M
uel
l.) H
.W
endl
. & D
rude
Pin
tau
d 4
07 (
TL
)A
J404
807
AJ4
0493
8A
J404
905
AM
1147
02
Ver
sch
affe
ltii
nae
Ph
oen
icop
hor
ium
bors
igia
nu
m (
K.K
och
) S
tun
tz
1986
–234
6 (K
)A
M11
0237
AM
1168
43A
M11
3686
AM
1147
03
Ros
cher
ia m
elan
och
aete
s(H
.Wen
dl.)
H.W
endl
.19
85−1
825
(K)
AM
1102
38A
J404
947
AJ4
0491
4A
M11
4704
Un
plac
ed g
ener
a in
Are
ceae
Ben
tin
ckia
nic
obar
ica
Bec
c.P
inta
ud
457
(T
L)
AM
1102
39A
M11
6844
AM
1136
87A
M11
4705
Cli
nos
tigm
a sa
vory
anu
m(R
ehde
r &
E.H
.Wil
son
)H
.E.M
oore
& F
osbe
rg
Pin
tau
d 4
42 (
MA
K)
AM
1102
40A
M11
6845
AM
1136
88A
M11
4706
Cyr
tost
ach
ys r
end
a B
lum
e19
82–5
882
(K)
AJ4
0481
0A
J404
940
AJ4
0490
7A
M11
4707
Dic
tyos
perm
a al
bum
H.W
endl
. & D
rude
C.E
. Lew
is 9
8–06
1 (B
H)
AM
1102
41A
M11
6846
AM
1136
89A
M11
4708
Dra
nsfi
eld
ia m
icra
nth
a(B
ecc.
) W
.J. B
aker
& Z
ona
C.E
. Lew
is s
.n.
(FT
G)
AM
1102
42A
M11
6847
AM
1136
90A
M11
4709
Het
eros
path
e el
ata
(Bec
c.)
F.B
.Ess
ig &
B.E
.You
ng
C.E
. Lew
is 9
9–03
4 (G
UA
M)
AM
1102
43A
M11
6848
AM
1136
91A
M11
4710
Het
eros
path
e lo
ngi
pes
(H.E
.Moo
re)
Nor
up
Pin
tau
d 4
61 (
TL
)A
J829
850
AM
1168
49A
M11
3692
AM
1147
11
Hyd
rias
tele
mic
rosp
adix
(Bec
c.)
Bu
rret
Bak
er 5
73 (
K)
AJ4
0481
7A
J404
943
AJ4
0491
0A
M11
4712
Hyd
rias
tele
ch
aun
osta
chys
(Bu
rret
) H
.E.M
oore
L-7
2.03
59 (
BH
)A
J404
816
AJ2
4089
8A
J241
307
AM
1147
13
Igu
anu
ra w
alli
chia
na
(Mar
t.)
Ben
tham
et
Hoo
k.f.
ex B
ecc.
1985
–148
8 (K
)A
J404
820
AJ4
0494
6A
J404
913
AM
1147
14
Lep
idor
rhac
his
moo
rean
a(F
.Mu
ell.)
O.F
.Coo
kB
aker
116
7 (K
)A
J829
881
AM
1168
50A
M11
3693
AM
1147
15
Lox
ococ
cus
rupi
cola
H.W
endl
.19
90–2
497
(K)
AJ8
2988
2A
M11
6851
AM
1136
94A
M11
4716
Rh
opal
obla
ste
augu
sta
(Ku
rz)
H.E
.Moo
reC
.E. L
ewis
99–
044
(FT
G)
AM
1102
44A
M11
6852
AM
1136
95A
M11
4717
Su
bfam
ily
clas
sifi
cati
on (
1998
)S
UB
FA
MIL
Y,
trib
e,su
btri
beS
peci
esV
ouch
er i
nfo
rmat
ion
rbcL
rps1
6 in
tron
trn
L–t
rnF
mat
K

38 C. B. ASMUSSEN ET AL.
© 2006 The Linnean Society of London, Botanical Journal of the Linnean Society, 2006, 151, 15–38
Ou
tgro
up
s (f
amil
y)D
asyp
ogon
acea
eK
ingi
a au
stra
lis
R.B
r.C
has
e 22
30 (
K)
AM
1102
45A
M11
6853
AM
1136
96A
M11
4718
Das
ypog
onac
eae
Das
ypog
on b
rom
elif
oliu
sR
.Br.
Ch
ase
2229
(K
)A
M11
0246
AM
1168
54A
M11
3697
AM
1147
19
Han
guan
acea
eH
angu
ana
mal
ayan
aM
err.
1998
–147
5 (K
)A
M11
0247
AM
1168
55A
M11
3698
AM
1147
20
Hae
mod
orac
eae
An
igoz
anth
os m
angl
esii
D.D
onA
smu
ssen
109
(C
P)
AM
1102
48A
M11
6856
AM
1136
99A
M11
4721
Poa
ceae
Far
gesi
a sp
.A
smu
ssen
105
(C
P)
AM
1102
49A
M11
6857
AM
1137
00A
M11
4722
Typ
hac
eae
Typ
ha
angu
stif
olia
L.
Asm
uss
en 1
07 (
CP
)A
M11
0250
AM
1168
58A
M11
3701
AM
1147
23C
ann
acea
eC
ann
a ed
uli
s K
er-G
awl.
Asm
uss
en 1
04 (
CP
)A
M11
0251
AM
1168
59A
M11
3702
AM
1147
24M
usa
ceae
Mu
sa r
osea
Bak
erA
smu
ssen
101
(C
P)
AM
1102
52A
M11
6860
AM
1137
03A
M11
4725
Bro
mel
iace
aeV
ries
ia p
sitt
acin
a L
indl
.A
smu
ssen
102
(C
P)
AM
1102
53A
M11
6861
AM
1137
04A
M11
4726
Com
mel
inac
eae
Tra
des
can
tia
pall
ida
(Ros
e) D
.R.H
un
tA
smu
ssen
103
(C
P)
AM
1102
54A
M11
6862
AM
1137
05A
M11
4727
Su
bfam
ily
clas
sifi
cati
on (
1998
)S
UB
FA
MIL
Y,
trib
e,su
btri
beS
peci
esV
ouch
er i
nfo
rmat
ion
rbcL
rps1
6 in
tron
trn
L–t
rnF
mat
K
Ap
pen
dix
Con
tin
ued