a summary of the biostratigraphy of the …polsen/nbcp/cornet_olsen_85_sm.pdf · jurasico temprano...
TRANSCRIPT

Ill CONGRESO L A T I N O A K E R I C A N O D E P A L E O N T O L O G I A . M E X I C O . S I M P O S I O SOBRE F L O R A S D E L T R I A S I C O T A R D I O , S U F I T O G E O G R A F I A Y P A L E O E C O L O G I A . MEt !ORIA. p.67-81, 1 t a b . , 6 f i g . , 1985.
A SUMMARY OF THE BIOSTRATIGRAPHY OF THE NEWARK SUPERGROUP OF EASTERN NORTH AMERICA
WITH COMMENTS ON EARLY MESOZOIC P R O V I N C I A L I T Y
BRUCE CORNET Superior Oil Company Geoscience Lab., 12401 Westheimer Rd. Houston,
Texas 77077 U.S.A.
PAUL E. OLSEN Lamont-Doherty Geological Observatory,Columbia University, Palisades,
New York 10964 U.S.A.
ABSTRACT
The Newark Supergroup of Eastern North America consists of rocks judged to range in age from early Middle Triassic through late Early Jurassic. Dating is principally based on the recognition of seven pa- lynological zones correlated with the European standard stages and on correlation of vertebrates and megafossil plants. The basic floristic trends are a decrease in diversity into the Early Jurassic and the rise to strong dominance of the cheirolepidaceous conifers. Based on publish- ed radiometric scales, the Newark depositional episode lasted roughly 45 million years and was antecedent to the actual separation of the North American and African Plates. Correlation of other Early Mesozoic continental areas with the Newark allows the conclusions that there was a shift from Late Triassic floral and faunal provinciality to Early Jurassic homogeneity, and that this shift was synchronous with a widening of the equatorial arid zone.
RESUMEN
El Supergrupo Newark, del Oriente de America del Norte, consiste en rocas quea juicio de 10s autores tienen un margen de edad entre el ~riasico Medio temprano y el Jurasico Temprano tardlo. La asignaci6n de edad se basa principalmente en el reconocimiento de siete zonas palino- logicas que se correlacionan con 10s pisos estandar europeos y en la correlation mediante vertebrados y megafosiles vegetales. Las tenden- cias floristicas basicas son un decrement0 de la diversidad hacia el Jurasico Temprano y el surgimiento a una dominancia fuerte de las coni- feras del tipo Cheirolepidiaceae. De acuerdo con escalas radiom6tricas publicadas, el episodic de deposit0 Newark dur6 unos 45 millones de aiios y precedi6 la separaci6n de las placas de ~m6rica del Norte y de Africa. La correlaci6n de otras areas continentales del Mesozoico tem- prano con el Newark permite las conclusiones de qua en este lapso suce- di6 un cambio que a partir de la provincialidad de flora y fauna del Triasico Tardio llev6 a la homogeneidad del Jurasico Temprano, y que este cambio fue contemporsneo con el ensanchamiento de la zona arida ecuatorial.
INTRODUCTION AND GEOLOGICAL CONTEXT
The Newark Su~erqroup (Figure 1) con- sists of a series of Early Mesozoic deposits of principally red non-marine elastics and minor thoeliitic igneous rocks exposed in a long series of discrete basins from Nova Scotia, C.anada to South Carolina, U.S.A. (OLSEN, 1978; VAN HOUTEI'I, 1977; FROELICH and OLSEH, 1984). The long axes of these basins
parallel the fabric of the Appalachian Oro- gene and in most cases the basin sequences are preserved as internally faulted and tilt- ed halfgrabens. A long series of closely re- lated basins occur beneath the Atlantic Coast- al Plain and on the Continental Shelf (OLSEM, 1978).
Most Newark Supergroup rocks were ap- barently deposited in asymmetrically subsid-

6 8 CORNET & OLSEN
ing troughs formed along major Paleozoic thrust faults,which were reactivated in the Early Hesozoic as listric normal or normal- oblique faults (LINDHOLM, 1978; PETERSON, et al., 1984). There were at least several kilometers of dip-slip movement during the history of the basins. Whether these basins formed in a simple extensional system acting perpendicular to the axis of the basins or in a wrench system acting oblique to the fabric of the orogene is a matter of ongoing
debate (MANSPEIZER and OLSEN, 1981; MANSPEI- ZER, 1982); however, it is certain that these basins formed just before the onset of actual separation of the North American and African plates and the oldest formation of Atlantic oceanic crust in the liiddle Juras- sic (SUTTER and SMITH, 1979).
As much as 10 km of principally fluvial and lacustrine sediments filled these basins. Host sequences show a bulls-eye facies pat-
SCALE Ikm I
0 800 . . . . . . . . A - EXPOSED
INFERRED
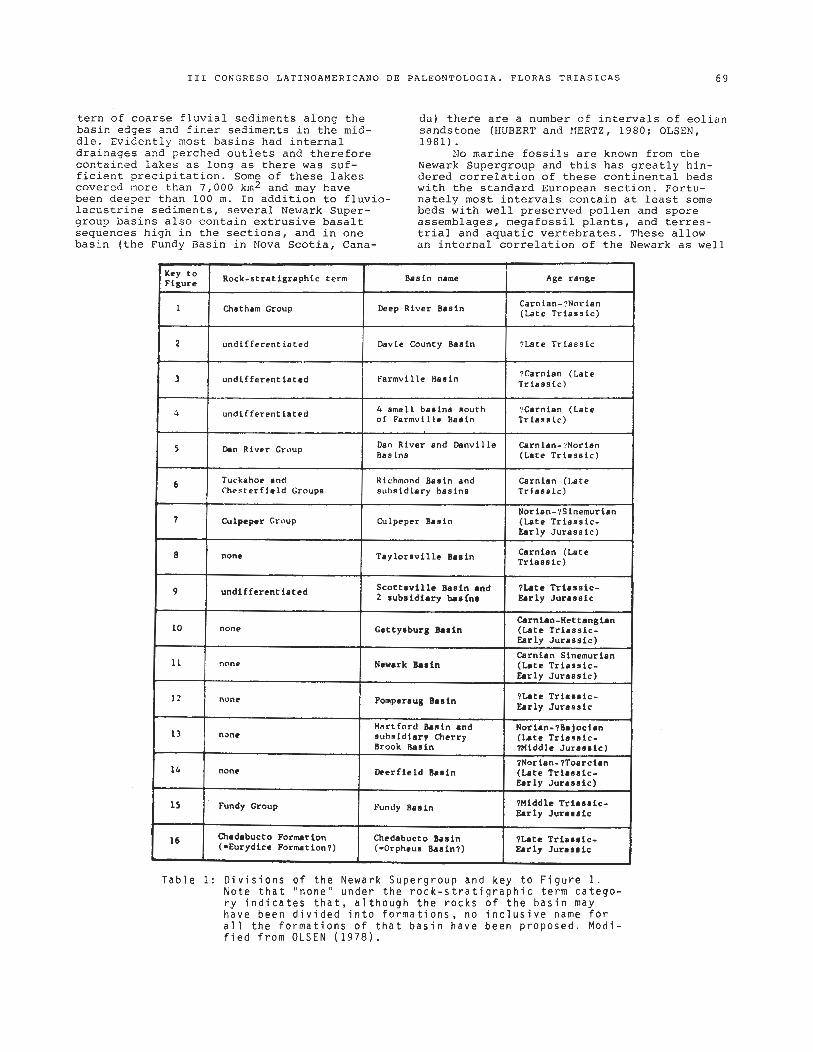
F i g u r e 1: The Newark S u p e r g r o u p o f E a s t e r n N o r t h A m e r i c a . M o d i f i e d f r o m O L S E N ( 1 9 7 8 ) . The i n f e r r e d a r e a s p r o b a b l y o v e r l a p t h e Newark i n a g e b u t some may n o t b e l i t h o l o g i c a l l y s i - m i l a r t o t h e N e w a r k . Key t o F i g u r e i n T a b l e 1.
I11 CONGRESO LATINOAMERICANO DE PALEONTOLOGIA. FLORAS TRIASICAS 5 9
tern of coarse fluvial sediments alonq the basin edges and finer sediments in the mid- dle. Evidently most basins had internal drainages and perched outlets and therefore contained lakes as long as there was suf- ficient precipitation. Some of these lakes covered more than 7,000 km2 and may have been deeper than 100 in. In addition to fluvio- lacustrine sediments, several Newark Super- group basins also contain extrusive basalt sequences high in the sections, and in one basin (the Fundy Basin in Nova Scotia, Cana-
da) there are a number of intervals of eolian sandstone (HUBERT and MERTZ, 1980; OLSEN, 1981).
No marine fossils are known from the Newark Supergroup and this has greatly hin- dered correlation of these continental beds with the standard European section. Fortu- nately most intervals contain at least some beds with well preserved pollen and spore assemblages, megafossil plants, and terres- trial and aquatic vertebrates. These allow an internal correlation of the Newark as well
~~~~0 ~ o c k - s t r a t i g r a p h i c t e r m Basin name Age r a n g e
Carnian-?Norian I 1 1 Chatham Group 1 Deep R i v e r Basin 1 (La te T r i a s s i c )
1 2 1 u n d i f f e r e n t i a t e d 1 Davie County Bas in 1 ?La te T r i a s s i c
u n d i f f e r e n t i a t e d
1 Lkn Rive r and D a n v i l l e Carnian-?Norian 1 5 1 Dan Rive r Group Bas Ins 1 (La te T r i a s s i c )
A
Farmvi l l e Bas in ?Carnian ( L a t e T r i a s s i c )
u n d i f f e r e n t i a t e d
6
I Carn ian ( L a t e T a y l o r s v i l l e Basin T r i a s s i c )
7
1 9 I u n d i f f e r e n t i a t e d I S c o t t s v i l l e Bas in and ?La te T r i a s s i c - 2 s u b s i d i a r y b a s f n s 1 E a r l y J u r a s s i c
4 s m a l l b a s i n s s o u t h o f F a r m v i l l e Basin
Tuckahoe and C h e s t e r f i e l d Groups
Â¥!Carnia ( L a t e T r i a s s i c )
4
Culpeper Group
1 12 1 none Pomperaug Bas in
Richmond Basin and s u b s i d i a r y b a s i n s
10
11
?La te T r i a e s i c - E a r l y J u r a s s i c
Carnian (La te T r j a à § 8 i c
Culpeper Bas in N o r i a n - ~ S i n e m u r i a n ( L a t e T r i a s s i c - E a r l y J u r a s s i c )
none
none
I 1 5 Fundy Croup Funily Baa i n
G e t t y s b u r g Baa in
Newark Bas in
6
?Middle T r i a s s i c - E a r l y J u r a s s i c
Ca rn ian -He t t ang ian ( L a t e T r i a s s i c - E a r l y J u r a s s i c )
Ca rn ian S inemur ian (La te T r i a s s i c - E a r l y J u r a s s i c )
H a r t f o r d Basin and s u b s i d i a r y Cherry Brook Basin
D r e r f i e l d Basin
13
Ill
Nor inn-?Be j o c i a n (La te T r i ~ i ~ i c - ?Middle J u r a s s i c )
7Norian-?Toarctan (La te T r i a s s i c - E a r l y J u r a s s i c )
none
none
T a b l e 1: D i v i s i o n s o f t h e Newark S u p e r g r o u p and k e y t o F i g u r e 1. N o t e t h a t " n o n e " u n d e r t h e r o c k - s t r a t i g r a p h i c t e r m c a t e g o - r y i n d i c a t e s t h a t , a l t h o u g h t h e r o c k s o f t h e b a s i n may h a v e b e e n d i v i d e d i n t o f o r m a t i o n s , no i n c l u s i v e name f o r a l l t h e f o r m a t i o n s o f t h a t b a s i n h a v e b e e n p r o p o s e d . M o d i - f i e d f r o m O L S E N ( 1 9 7 8 ) .
1 6 Chedabucto Format ion ( -Eurydice Format ion?)
Chedabucto Bas in (-Orpheus Bas in? )
?La te T r i a s s i c - E a r l y J u r a s s i c

7 0 CORNET & OLSEN
as correlation with the European standard stages and other Early Mesozoic sections.
We address 4 main goals in this paper: 1) to summarize theage-correlativedata from the Newark, concentrating on the pollen and spore and vertebrate assemblages; 2 ) to re- late this local chronostratigraphic frame- work to other continental Early Mesozoic areas and the European standard stages; 3) to attempt to calibrate the biostratigraphy of the Newark by correlation with published radiometric scales; and 4) to use information from these correlatiorsto briefly examine floral and faunal provinciality during the Early Mesozoic.
PALYNOLOGICAL CORRELATIONS
Plants, especially pollen and spores, are of paramount importance to the biostrati- graphy of the Newark. CORNET (1977a) propos- ed seven pollen and spore zones,which are summarized below and in Figures 2 and 3. The oldest zone, the Chatham-Richmond-Taylors- ville palynoflora, is recognized in the Chatham Group of the Deep River Basin and in the Richmond and Taylorsville basins, and contains about 155 pollen and spore species. This assemblage can be divided into two subzones.
European Standard
tages
TOARCIAN
LIENS- BACHIAf
NORIAN
CARNIAN
LADINIAN
ANISIAN
F i g u r e 2 : I n t e r n a l c o r r e l a t i o n o f t h e Newark , b a s e d on f a u n a l a n d f l o r a l d a t a ( c o r r e l a t i o n s b a s e d on d a t a p r e s e n t e d i n CORNET ( 1 9 7 7 a , 1977 b ) , ULSEN a n d GALTON ( 1 9 7 7 , i n p r e s s ) , OLSEN, McCUNE and THOMSON ( 1 9 8 2 ) a n d OLSEN ( 1 9 8 3 a , 1 9 8 3 b ) .
W i t h i n e a c h c o l u m n r e p r e s e n t i n g a s i n g l e b a s i n , t h e d a r k g r a y zones r e p r e s e n t p r i n c i p a l l y l a c u s t r i n e r o c k s w h i c h a r e d o m i n a n t l y g r a y o r b l a c k ; t h e l i g h t e r g r a y zones r e p r e s e n t l a c u s t r i n e r o c k s w h i c h a r e d o m i n a n t l y r e d b u t w h i c h may h a v e s i g n i f i c a n t g r a y b e d s ; t h e w h i t e zones a r e d o m i n a n t l y f l u v i a l o r unknown; t h e v e r t i c a l l y r u l e d zones r e p r e s e n t e x t r u s i v e b a s a l t f o r m a t i o n s w h i c h o f t e n h a v e i n - t e r b e d d e d s e d i m e n t a r y f o r m a t i o n s w i t h l a c u s t r i n e r o c k s ; a n d t h e zone m a r k e d w i t h " x " s r e p r e s e n t s a l t d e p o s i t s i n t h e o f f s h o r e Orpheus G r a b e n . H o r i z o n t a l l i n e s w i t h i n t h e c o - l umns a r e f o r m a t i o n b o u n d a r i e s .

COROLLINA
MURPHY1
COROLLINA
TOROSUS
COROLLINA
MEYERIANA
MA EUROPEAN - STANDARD
MANASSAS-
UPPER
PASSAIC
LOWER
PASSAIC-
HEIDLERSBURO
190 -
- 200 -
- 208 -
220 -
230 -
NEW OXFORD-
LOCKATONG
STAGES
TOARCIAN
PLIENSBACHIAI
SINEMURIAN
- - - - HETTANGIAN
.
NORIAN
LATE
CARNIAN
MIDDLE CARNIAN RICHMONO-
TAYLORSVILLE
F i g u r e 3 : The d i s t r i b u t i o n o f a g e - d i a g n o s t i c p a l y n o m o r p h t a x a w i t h i n t h e Newark S u p e r g r o u p ( d a t a a d a p t e d f r o m C O R N E T , 1 9 7 7 a ) .

CORNET & OLSEN
A lower subzone is dominated either Chatham-Richmond-Taylorsville 'palynoflora, by crywtoqam spores and gymnospermous all of which indicate that it is of Late monosulcate pollen, or by a diversified Carnian Age. gymnospermous assemblageof non-striate bi- saccate and circumsaccate pollen with minor percentages of monosulcate pollen, circum- polles, tetradopolles, and striate bisaccate pollen. This subzone occurs in the Richmond and Taylorsville basins. A similar pollen- dominated assemblage occurs in the basal Pekin Subbasin of the Chatham Group. A i a t / i i b - p o i i t e b b c a b > i a t u b , A. 6 i m b i i a t u b , CaLamobpo - i a n a t h o h b t i i i L , and La.>n.coid i i teb spp. make up the dominant cryptogram elements of the old- est spore and monosulcate-dominated assem- blage, while O b m u n d a c i d i t e - i i spp. , 8 a c u t a t i . h - ? ~ V i i t e b spp., and G'io.nu. tat- i&po/ i i teb spp. compete with articulate spores (e.g. L a i i c o i - d i t e b sv. and P i t a b o o i i i t e b sp.) for dominance in younger spore-dominated assemblages.
These contrasting cryptogamous and gymnospermous palynofloras alternate with one anotherthrough the Richmond-Taylorsville basin sequences as the lithologic facies alternate from deltaic-lacustrine to fluvio-deltaic. Based on recent well data from the Richmond Basin, at least two upwards-coarseninglacu- strine to fluvial sequences, and the "head and tail" portions of two additional se- quences record the cyclic waxing and waning of a large rift valley lake.
The upper subzone contains a less diver- sified spore assemblage and has more abundant P a i i n c i t i p v t - i t i l i d e n b u b , a circumsaccate mor- photype that ranges down to the oldest Rich- mond Basin strata. The upper subzone also lacks A i a t i - i & p o i i t e t , spp. and contains only rare striate bisaccate pollen, which decrease in abundance and diversity upwardsthrough the lower subzone. This upper sub-assemblaoe occurs in the upper Pekin Formation and Cumnock Formations of the Chatham Group, the lower New Oxford Formation of the Gettysburg Basin, and perhaps the lower Wolfville Forma- tion of the Fundy Group (TRAVERSE, 1983).
Age-diagnostic species of the oldest zone include the overlapping ranges of P a t i - n a b p o i ' i t e i d e n b u i , S t i i a t o a b ' i e - i t e . h a g X u g i i , Th-t .adibpo/ ic( ve i ' i u ca . t . a , P t o X o d i p L o x y p i n u b doubA.nge. ' i i , Pfica-fccAaccub bad - i ub , T ~ L a d i b p o t a cf . T . a u t e c ~ , and L a g e n e t e a m a n . t i n i i , indicat- ing a orohable late Middle Carnian age. How- ever, palynoflorules older than Middle Car- nian are only now becoming well known in Europe and it is possible that the beds of the Richmond and Taylorsville basin may be substantially older than this assessment - perhaps even as old as Middle Triassic.
The upper sub-assemblage of the Chatham- Richmond-Taylorsville palynoflora is replac- ed upwards in the Gettysburg Basin by the New Oxford-Lockatong palynoflora, which has also been recognized in the Newark (CORNET, 1977a) and Dan River basins (ROB- BINS, 1982). This assemblage is dominated by Pa t i nabpon .He . i i d e n b u b , V a t t i b p o h - i t a ig - n a c i i , A t i b poh- i teb p a i v u b , and T i i a d i i b p o i a spp. It still contains C a m e i o b p o i i t e b b e c a - t u b and C. p b e u d o v e ~ i u c a t u ~ , but lacks the Middle Carnian age-diagnostic taxa of the
The New Oxford-Lockatong palynoflora is gradually replaced upwards in the Newark Basin by the Lower Passaic-Heidlersburg palynoflora found thus far in the Culpeper, Gettysburg, Newark, and Hartford basins. This assemblage is distinguished by the pre- sence of C a m e i o b p o i i t e t , u e i i u c o b u b , an abun- dance of large and varied bisaccate species, including the giant A i i h p o f i ^ t u op- iL , and the continued dominance of P a t i n a ~ p o h i t e i d e n h u b , VaHabpo&^ts . f i i g n a c i i , and T i i a d i b - p o i a spp. This assemblage appears to be of Norian age. The upper portion of this zone, in the Newark Basin, contains the first rare occurrences of C o i o t t ^ n a m e y e i i a n a .
The Lower Passaic-Heidlersburg palyno- flora slowly changes upwardsinto the Manas- sas-Upper Passaic palynoflora in the Culpe- per and Newark basins. This zone is distin- guished by the first occurrence of C o i o U h a t o i o b u b and G i a n u t o p e ~ t c u t a t  ¥ ( . p o t i i m d i b and the continuation of taxa common in the pre- vious zone. This zone correlates with what palynostratigraphers have been calling the Rhaetian in Europe. The Rhaetian was first defined in marine rocks in the Austrian Alps, but these rocks have been recently shown to be partly equivalent to marine Late Norian strata elsewhere in Europe (PEARSON, 1970; TOZER, 1979). A number of authors do no longer consider the Rhaetian to represent an independent stage. What its loss does to the palynologically defined Rhaetian is unclear. However, Rhaetian palynofloras probably should be regarded as Late tlorian, while 'typical" Norian assemblages are probably best regarded as Early and Middle Norian. The Manassas-Upper Passaic palynoflora is thus Late Norian in age and is recognized in the Culpeper, Newark, and possibly Fundy basins (The name Manassas was originally chosen for palynoloqical localities near Manassas, Virginia, and does not refer to the Manassas Formation of the Culpeper Basin, which is older).
This latter zone ends upwards with a rapid transition to the C o - t o H i n a meyet^.ana palynoflora, which is characterized by a switch from a high diversity assemblage do- minated by bisaccate pollen, Pat-Lnab , p c i - i t e b , C o i o i i i n a m e y i t i a n a , C. t ~ t ~ b ~ b , and G t a n u - tope. ' icuLat- i iJo/ t ' i^ i u d i b to a low diversity assemblage very strongly dominated (+go%) by the circumpolles C o t o t i i n a m e y e i i a n a . This transition takes place over an 80 m thick "barren" interval below the Orange Mountain Basalt in the Newark Basin (Jack- sonwald Syncline). By the top of this in- terval, all taxa characteristic of the Late Triassic are absent. The high diversity assemblage below this interval is peculiar in having a suite of pollen grains showing all the characters typical of angiosperms, although they are not assignable to any known angiosperm group. According to DOYLE (1978), "...these grains are perhaos the best candidates yet for pre-Cretaceous

I11 CONGRESO LATINOAMERICANO DE PALEONTOLOGIA. FLORAS TRIASICAS 7 3
angiosperms...". An assemblage of somewhat similar grains occurs in the Chatham-Rich- mond-Taylorsville palynoflora.
The C o i o f i i n a meq&ana palynoflora is characterized by the first common appearance of Cci : i !o fu t4 .5po ia h tuk^ f io^ .ma; this zone is the lowest of the Jurassic pollen and spore zones in the Newark. The Triassic-Jurassic boundary is best ?laced just below the Orange Mountain Basalt and the oldest extrusives in other basins (CORNET, 1977a, 1977b) rather than with- in the zone of extrusives as suggested by CORNET, et al. (1973) and CORNET and TRAVERSE (1975). "BIG C o i o i t i u a me.yeh-ia.na palynoflora con- tinues through the entire zone of extrusives and into the overlyinq sediments in all of the Newark Supergroup Basins. Pollen and spore assemblages do not allow a finer subdivision within this zone or a correlation of individu- al sedimentary formations between extrusives in different basins. This zone seems to cover all of the Hettangian and Sinemurian of the Early Jurassic.
The C o i c i f f i n a ¥('0^.0.5u palynoflora succeeds the i.'. i n e y c i i a n a palynoflora upwards with a strong switch in the dominant taxon from C o i o - Pf-ciia mcye..iiana to C . XokoAu.5. This switch oc- curs in the beds some distance above the upper- most extrusives in the Culpeper and Hartford basins. The age of this assemblage appears to be Pliensbachian.
In the Portland Formation of the Hartford Basin there is one zone younger than the Coko- €£<-i t f a o 5ub palynof lora. The C o i o f t i n a mu^.-
.!? . . , , , salynoflora occurs in the highest sampled horizons in the Portland and, hence, in the Newark Supergroup. It is characterized by a dominance of Co?cCC4iti! mu4p11y4 over C . .('Oi0.5ub (52% vs. 39% . The age of this assemblage a?- pears to be Toarcian.
RELATIONSHIP OF PALYNOMORPH ZONES TO MEGAFOSSIL ZONES
Although the megafossil ~lants of the Newark were a major focus of research, par- ticularly with regard to age, in the late 19th and early 20th centuries, much less attention has been paid to them since the 1920's. Recently, ASH (1980) has reviewed North American Early Mesozoic plant assem- blages and divided them into three zones all of which are represented in the Newark Super- qroup (Figure 3).
The oldest is called by ASH the Zone of Eog- ink jo i . te . f i , a bennettitalean gymnosperm. The zone of which it is typical has been identified in the Newark in the Richmond Ba- sin, the Pekin, and the Cumnock Formations of the Deep River Basin, and the Stockton Formation of the Newark Basin (ASH, 1980). Florules of this zone are the most diverse in the Newark Supergroup, with very common ferns and bennettitaleans. Florules in this zone include those described by FONTAINE (1883), HOPE and PATTERSON (1970), DELEVORYAS and HOPE (1971, 1975, 1976). EMHOMS (1856. 1857), WILLARD et al. (1959), BROWN (1911), and BOCK (1952, 1969, in part).
Above the Zone of E o g i n k g o i X e . ~ is ASH'S Zone of V i n o p h y t o n , a conifer-like gymnosperm. This zone has been recognized in the upper member of the Cow Branch Formation of the Dan River Basin (OLSEN it a t . , 1978), and in the middle New Oxford Formation of the Gettysburg Basin (ASH, 1980; WARD, 1900; CORNET, 1977b). Newark assemblages from this zone are some- times dominated by conifers,but are generally still diverse with an abundance of bennettita- leans. These florules are still poorly known.
The youngest zone recognized by ASH is called simply the Upper Zone. This zone is strongly dominated by cheirolepidaceous coni- fers, especially B k a c h y p h y t t u m , Pagio'phqSLtum, and Uiime.te.LLa. The cycadeoid OtozanlUe.5 anJ the fern Cta.t l~, toptn' i . ' i i i are often common. Compared to the older two zones, the florules of this zone are of very low diversity. This zone is represented in the syn- and post-extrusive sediments of the Culpeper, Gettysburg, Newark, Pomperaug, Hartford, and Deerfield basins. Scattered descriptions of elements of these florules were described by EMMONS (1857). BOCK (1969) , and especially NEWBERRY (1888) , and CORNET (1977a), but these are diverse assemblages which definitely can be further subdivided.
The Zone of E o g - i i i k g o i t e . ~ corresponds broadly to CORNET'S (1977a) Chatham-Richmond- Taylorsville palynoflora and it is thus Middle Carnian (or older) in age. The Zone of 'D-in.0- ,3hyton corresponds to the New Oxford-Lockatonq palynoflora and is Late Carnian in age. The Upper Zone corresponds to the Jurassic pollen and spore zones of CORNET (1977a). It should also be noted that this last assemblage clear- ly resembles what is called the H.iime.ie.tta ( C d e L t o t t p i ~ ) assemblage, typical of the Early Jurassic of Great Britain (CORNET , 1977a) and it probably corresponds to Harris's Zone of Thauma . top . t e . i i~ (ASH, 1980) of Het- tangian age. ASH based his zones mostly on deposits in the Western United States. There is as yet no definitive evidence for sedi- ments intermediate in age between the Late Carnian and the Early Jurassic in that area. Therefore, ASH did not recognize megafossil plant zones that might correspond to the Lower Passaic-Heidlersburq palynoflora or the Manassas-Upper Passaie palynoflora in the Newark. NEWBERRY (1888), BOCK (1969, in part) and CORNET (1977a) have described some florules corresponding to these palynomorph zones, notably that of Milford, New Jersey from beds of the Lower Passaic-Heidlersburg palynoflora. In any case, the megafossil assemblaqes from these latter two zones of the Newark are the least well known of the supergroup.
In very broad terms, the most obvious floral trend visible through the entire Newark section is decreasing floral diversity. The oldest beds of the Newark, typified by the Richmond Basin section, have both the largest number of described megafossil spe- cies and the largest number of pollen and spore morphotypes. As important as the number of taxa is, no single taxon is consistent- ly dominant from locality to locality within the Richmond Basin and at some localities the

TRIASSIC JURASSIC
DICTYOPYGE SPP. TANAOCROSSUS SP. CIONICHTHYS SPP. SYNORICHTK~S SP. "DIPLURUS" (PARIOSTEGEOUS) SP. "DIPLURUS" (OSTEOPLEURUS) SP. - TURSEODUS SPP. -. , "PHOLIDOPHORID"
w8 7 CARINACANTHUS JEPSENI
$ "SEMIONOTUS BRAUNII GROUP" SEMIONOTUS SPP. CF. HYBODUS SP. PTYCHOLEPIS SPP. REDFIELDIUS SP. ISCHYPTERUS SPP. "SEMIONOTUS ELEGANS GROUP" DIPLURUS LONGICAUDATUS "ACENTROPHORUS" CHICOPENSIS
"TREMATOSAUR" CF. APHANERAMA SP. CF. ANOMIODON SP. CF. SCLEROSAURUS SP. CF. EXAERETODON SP. CF. PROTOROSAURUS SP. TANYSTROPHEUS SP. "TRILOPHOSAURS" "RAUISUCHIDS" "DICYNODONTS" DOSWELLIA KALENBACHI ~ICTYOCEPHALUS ELEGANS - MICROCONODON TENUIROSTRIS " - DROMETHERIUM SYLVESTRE -.
(T "CAPITOSAURS" (INCL. CALAMOPS) (rt "TYPOTHORAX" SP.
METOPOSAURUS (INCL. EWELOR) SPP. CÃ SCALENODONTOIDES SP.
LEPTOPLEURON SP. "3 NEW PROCOLOPHONIDS" PARADAPEDON ICAROSAURUS SEIFKERI
2. STAGONOLEPIS AETOSAURUS TAOTTRACHELOS SPP.
"THECODONTOSAURUS" GIBBIDENS STECOMUS SPP. SFHODROSAURUS PENNSYLVANICUS AFF. SPHENODON SP. RUTIODON SPP. "PHYTOSAURS" (INCL. CLEPSYSAURUS) HYPSOGNATHUS FENNERI "SMALL SAURISCHIANS" AFF. GLEVOSAURUS SP. "FABROSAURID" STEGOMUSUCHUS SPP. AMNOSAURUS SPP. ANCHISAURUS SOLUS PODOKOSAURUS HOLYOKENSIS
BRACHYCHIROTHERIUM SPP. APATOPUS SPP. GWYNEDDICHNUS SPP. "ANCHISAURIPUS" MILFORDENSIS (N.GEN. l
5 RHYNCIIOSAUROIDES HYPERBATES RHYNCHOSALROIDES BRUKSWICKII BRACHYCHIROTHERIUM EYERMAUI BRACHYCHIROTIIERIUM PARVUM GRALLATOR (GRALUTOR) SPP.
01 RHYNCHOSAUROIDES SPP. GRALLATOR (ANCHISAURIPUS) SPP GRALLATOR (EUBRONTES) SPP. BATRACHOPUS SPP. OTOZOUM SPP. ANOMEOPUS SPP. AMEGHINICIINUS SF.

I11 CONGRESO LATINOAMERICANO DE PALEONTOLOGIA. FLORAS TRIASICAS
relative abundances of taxa are surprisingly evenly distributed. By the Late Carnian, however, conifers often dominate the macro- flora, and conifer pollen is dominant in the microflora. The dominance of one taxon in the microflora rarely exceeds 50%. In the Early Jurassic Newark, however, conifers are very strongly dominant in both macro and microflo- ras and often comprise more than 90% of the assemblages.
VERTEBRATE ZONES
Vertebrate fossils are relatively common in most parts of the Newark Supergroup, espe- cially fishes and reptile footprints. OLSEN, McCUNE, and THOMSON (1982) and OLSEN and GALTON (1977; 1984) have reviewed much of the biostratigraphically useful vertebrate data and recognized a number of informal biostrati- graohic zones. Recent discoveries have shown that the useful biostratigraphic zones are fewer in number and less complex than origi- nally thought(OLSEN, 1983a, 1983b). The dis- tribution of vertebrates in the Newark and the zones they define are shown in Figure 4. Note the correspondence between the upper limit of the A p a - t o p u ~ zone and the base of the C o t o . U i n a me.qe.n.-iana palynoflora and the apparent lack of correspondence of the bound- aries of the other vertebrate zones with the palynoflora zones.
The oldest Newark vertebrate assemblage occurs in a small isolated deposit within the Fundy Basin (A of Figure 4). Vertebrates from this deposit differ from all others in the Newark and indicate an early Middle Triassic (Anisian) age for the deposit. These are the oldest beds in the Newark. No floral data are available from these beds, unfortunately.
The next oldest assemblage occurs in the Richmond, Taylorsville, and Scottsville ba- sins (B of Figure 4) and is characterized by vertebrates found nowhere else. Typical is the- unique refieldiid fish V i c t y o p y g e . This assemblage characterizes the lower part of the Chatham-Richmond-Taylorsville palynoflo- ra zone and appears to be of Middle Carnian age, but it might be older.
The abundance of phytosaur reptiles makes up the next assemblage (C of Figure 4). which is characteristic of Late Triassic vertebrate assemblages from other parts of the world such as the German Middle and Late Keuper. The upper Chatham-Richmond-Taylors- ville through the Lower Passaic-Heidlersburg. palynofloras contain this phytosaur-dominated a~semblaqe~which makes up the bulk of the Newark.
The youngest Newark vertebrate assemblage is characterized by a dominance of the remains of dinosaurs and crocodiles and has no count- erpart in the European Late Triassic (OLSEN and GALTON, 1977, In Press) (D and E of Fig- ure 4). Correspondingly the associated palyno- floras are the Col.oi!t^.nn. meyek iana . through the C o - t o ~ ~ i n a mufipk-i'i palynofloras, themselves correlated with the Early Jurassic. This as-
semblage has its oldest representatives in the sediments above the oldest extrusives in the Newark.
CALIBRATION OF THE NEWARK
Five current and somewhat different ra- diometric scales are available for the Early Hesozoic on a stage by stage level (Figure 5). ~ l l five scales use partially independent data and have rather different age assignments for the boundaries between stages and periods (Figure 5). The lengths of the intervals between stages are. however, closely compar- able. These scales can be tied to the Newark using the palynologically defined age-bound- aries, discussed above. Within the Newark the best defined boundaries are the Middle Car- nian - Late Carnian boundary, the Norian - Hettanqian boundary and the Pliensbachian - Toarcian boundary. To apply the radiometric scales to the Newark we have had to assume that the Late Carnian was 1/3 the duration of the Carnian and, of course, assume that the correlation of the radiometric scales with the European standard stages are correct.
The duration of intervals within most of the Newark calibrated by the available radio- metric scales are in very good agreement with independent calibration by lacustrine sedimen- tary cycles ascribed to climate control by the 21,000 year precession cycle (VAN HOUTEN, 1969, 1980; OLSEN, 1984af 1984b). However, the thicknesses of the palynologically defin- ed Middle Carnian of the Newark is out of proportion with the 1.5 - 3.5 million years alloted to the Middle Carnian in the publish- ed scales. If the sedimentation rates ascrib- ed to the palynologically defined post-Mid- die Carnian of the Newark were similar to those of the Newark Middle Carnian, the du- ration of the pre-Late Carnian of the Newark would be on the order of 10 million years. This suggests either a major problem with assessments of the duration of the Carnian, problems with correlation of the palyno- florules of the Newark with the European section, or the possibility that the sedi- mentation rates for the older beds were significantly higher. The last possibility is in line with sedimentological data from the Richmond Basin.
CORRELATION WITH OTHER AREAS OF CONTINENTAL 'SEDIMENTATION
Important areas of early Mesozoic con- tinental sedimentation outside the Newark include the European Keuper and Infralias, the Newark Supergroup-like basins of Morocco, the Chinle Formation, Dockum Group, and Glen Canyon Group of the southwestern United States, the Stormberg Group of southern Afri- ca, the Ischiqualasto and Los Colorados of Argentina, and finally the Lower Lufeng of China. Correlation between some of these areas and the European standard stages is hampered by a lack of some classes of age-

RADIOMETRIC SCALES
CV m
0)
0
-I <
184 - T
194
P
200 - s
206 - H
213 -
N
225 -, LC
U-E c
232 -
L
238 - A
0, CD
: 0)
- 6 0 " 2 -
< g 0 ) VERTEBRATE
ZONES 187 -
T
193
P
198 - s
204 - 208
225
LC
M-E C
230
L
235
A
240
Newark Supergroup
EUROPEAN PALYNOFLORA M E ~ ~ ~ ~ ~ ' L
STANDARD STAGES
TOARCIAN
s g
SINEMURIAN
- - - - HETTANQIAN
1 182 - T
188
P
197 - s
205
Ti- 208
U N N
225
LC
M-E c
230
235
A
240
LATE
CARNIAN
MIODLE
CARNIAN
200
218
LC
M-E
L W
L
235 A ANISIAN
ZONES ZONES (ASH, 1980)
COROLLINA
MURPHII -T- TOROSUS I
COROLLINA
MEYERIANA UPPER
MANASSAS-
UPPER
LOWER
PASSAIC-
HEIOLERSBIWG
NEW OXFORD
LOCKATONQ DINOpHyTON
FAYLORSVILLE
Ourita Karoo Southwest Yunnan
Basin Basin Argentina China
Morocco S. Africa u. s.
/
GLEN
CANYON
GROUP
! CHINLE - DOCKUM
MOENKOPI
M-C
A D
7
RAKENSBURG
VOLCANICS
f CLARENS
/' UPPER
ELLIOT
. - - -
LOWER
ELLIOT
x MOLTEN0
BEAUFORT
- -7 - -- UPPER LOS
COLORADOS
LOWER LOS
COLORADOS
ISCHIG-
UALASTO
7
UPPER
LOWER
LUFENG - -
LOWER
LOWER
LUFENG
7

I11 COMGRESO LATINOAMERICANO DE PALEONTOLOGIA. FLORAS TRIASICAS
correlative data, such as palynomorphs or marine invertebrates, but correlation by ver- tebrates through the Newark provides the ne- cessary and crucial link. Figure 5 shows an attempted correlation of the above areas with the Newark and the European standard stages.
Of particular interest are the assign- ments of the entire Glen Canyon Group of the southwestern United States, the Upper Storm- berg Group, and the upper Lower Lufeng of China to the Early Jurassic (OLSEN and GALTON, 1977, In Press). Until recently, the prevail- ing view was that all these intervals were Late Triassic in age. With these new age de- signations in mind, we can look at the pat- tern of "climate sensitive" rocks and floral and faunal provinciality for the Late Trias- sic and Early Jurassic (Figure 6).
LATE TRIASSIC PROVINCIALITY
Prominent in the pattern of deposits of Late Triassic Age is a zone between north 40 and south 30 paleolatitude where phytosaur reptiles and metoposaur amphibians are common (the latter only in the Carnian) (Figure 6). They are present even in northern Gondwana- land (Morocco and India) but are absent in the rich continental assemblages of the Ischi- qualasto and related formations of South Amer- ica and the Molteno and Lower Elliot formations of Africa. This equatorial assemblage is do- minated by archosaurs whereas the more south- ern contemporaneous assemblages are dominat- ed by mammal-like reptiles.
The metoposaur-phytosaur biogeographic province has floral counterparts. In the Late Triassic, all vertebrate assemblages south of 30 paleolatitude are associated with 0-tcto-t.d-t.um-type megafossil florules. Only in India do phytosaurs and metoposaurs occur with a P-tcto-tdkum-type florule. In all other areas, phytosaurs and metoposaurs occur with a typical Laurasian bennetitalean-conifer assemblage. The areas which have U-tc~oidium- type florules are dominated by palynoflorules of the Ipswich-Onslow-type (DOLBY and BALME, 1976; TRUSWELL, 1980) , including India (KUMA-
RAN and HAHESWARI, 1980). In the northern hemisphere, phytosaurs and metoposaurs are absent above 30 north; however, there does not seem to be a separate northern floral province in the region where these verte- brates are absent.
Coals are fairly common in Late Trias- sic rocks north of 30 north and south of 30 south latitude (ANDERSON and ANDERSON, 1970; ROBINSON, 1971, 1973). In between are scat- tered occurrences of evaporites and dune sands and much less common coals (Figure 6). Between north and south 30; the coals seem to be restricted toanequatorial belt, but there are so few occurrences, the pattern may not be meaningful. It is worth noting, however, that an arid-to-the-north trend has been noted within the Newark Supergroup which does conform to this postulated pattern (HUBERT and MERTZ, 1980; OLSEN, 1981; MANS- PEIZER, 1982).
EARLY JURASSIC PROVICIALITY
In contrast to the Late Triassic, no continental vertebrate provinciality is vis- ible in sediments of Early Jurassic age (Figure 6). This faunal homogenization extends to the intercontinental distribution of many terrestrial vertebrate genera, especially dinosaurs and mammals (OLSEN and GALTON, In Press). This lack of provinciality also seems to correspond to a reduction in the rate of turnover for faunal assemblages as a whole. A parallel pattern is apparent in Early Juras- sic plants.
It has long been recognized that the strong floral provinciality which characteriz- ed the Late Triassic world gave way in the Early Jurassic to a more hcirogeneous world flora dominated in nany areas by cheirolepidaceous conifers (ALVIN, 1982); this dominance is especially obvious in the preponderance of ConoUwa (Cta.i.iopo.C.C-tb) pollen in many areas. HUGHES (1973) has shown that in the Late Ju- rassic, the relative abundance of Coho!H.ina was greatest in the equatorial region, and it seems clear that this pattern was establish-
F i g u r e 5 : ' C a l i b r a t i o n o f t h e Newark b y p u b l i s h e d r a d i o m e t r i c s c a l e s and c o r r e l a t i o n o f t h e b i o s t r a t i q r a p h i c s u b d i v i s i o n s o f t h e Newark S u p e r g r o u p w i t h o t h e r E a r l y M e s o z o i c c o n t i n e n t a l d e p o s i t s . S o u r c e o f d a t a f o r t h e s e c o r r e l a t i o n s same as F i g u r e 4 a n d i n a d d i t i o n , MANSPEIZER ( 1 9 8 2 ) .
A b b r e v i a t i o n s as f o l l o w s : R a d i o m e t r i c s c a l e s : A, A n i - s i a n ; L , L a d i n i a n ; M-E C , M i d d l e and E a r l y C a r n i a n ; L C, L a t e C a r n i a n ; N, N o r i a n ; H, H e t t a n g i a n ; S, S i n e m u r i a n ; P ? P l i e n s b a c h i a n ; T, T o a r c i a n . V e r t e b r a t e Zones : A - E, d e s i g - n a t i o n s o f v e r t e b r a t e a s s e b l a g e s d i s c u s s e d i n t e x t . O u r i t a B a s i n : AD, a d e n s i t e ; M-C, M i n u t o ~ a c c u ~ - P a t ^ . n a ^ p o ' i H e ~ a s s e m b l a g e ; T , t h o l e i i t j c e x t r u s i v e b a s a l t s ; JC, J u r a s s i c m a r i n e and p a r a l i c c a r b o n a t e s .
N o t e t h a t i n t h i s F i g u r e t h e r a d i o m e t r i c s c a l e s a r e a l l t i e d a t t h e P l i e n s b a c h i a n - T o a r c i a n , N o r i a n - H e t t a n g i a n , and M i d d l e C a r n i a n - L a t e C a r n i a n b o u n d a r i e s as p a l y n o l o - g i c a l l y f i x e d i n t h e Newark .

CORNET G OLSEN
I F i g u r e 6

I11 CONGRESO LATINOAMERICANO DE PALEONTOLOGIA. FLORAS TRIASICAS 7 9
ed by the Early Jurassic. Going from south to north, there is a trend from Co~oHina- dominated (+go%) to CoaoH-ivta-poor (-10%) mi- croflorules (PEDERSON and LUND, 1980) with transitional assemblages occurring around a 25 north ,-ialeolatitude. This pattern is re- flected in the megafossils as well, with conifers strongly dominant in the equatorial region and much less so in the north. Data are as yet insufficient to recognize a simi- lar trend in the southern hemisphere.
Coals are virtually restricted to the high latitudes in the Early Jurassic (PAR- RISH, et al., In Press) (Figure 6) and dune sands are very common in the broad equatori- al humid zone in the data available for the Early Jurassic.
CONCLUSIONS
Rocks of the Newark Supergroup were de- posited during an interval greater than 35 million years, from the Middle Carnian of the Late Triassic through the Toarcian of the Early Jurassic. If the oldest beds da- table by vertebrates are included, the range increases to 45 million years. Within this interval seven pollen and spore zones can be recognized and these allow the Newark to be correlated with the European standard stages and published radiometric scales.
Correlation by vertebrates is especially useful within the Newark and within areas of continental sedimentation where palynomorph data are lacking. Using all the available da- ta, it appears that a number of sequences thought to be Late Triassic in age are actu- ally Early Jurassic.
These correlations permit the conclusion that the transition from Late Triassic to Early Jurassic was marked by a strong decrease in floral and faunal provinciality and an in- crease in indicators of aridity in the equato- rial zone.
ACKNOWLEGEMENTS
The principal palynoloqical research for
this paper was orginally part of the senior author's Ph.D. thesis at The Pennsylvania State University under Dr. ALFRED TRAVERSE and this work was supported in part by National Science Foundation grant number GA36870 to Dr. TRAVER- SE. The junior author's contribution was prod- uced while a Miller Postdoctoral Fellow at University of California at Berkeley and sup- port from the Miller Institute for Basic Re- search is gratefully acknowleqed. We also thank Dr. KEVIN PADIAN for reading the manu- script and for providing many helpful sug- gestions which substantially improved it.
REFERENCES
ALVIN, K.L., 1982. Cheirolepidiaceae. Biology, structure, and paleoecology. Rev. Paleo- hot. Palynol., 37: 71-98.
ANDERSON, H.M. and J.M. ANDERSON, 1970. A preliminary review of the biostratiqraphy of the uppermost Permian, Triassic, and lowermost Jurassic of Gondwanaland. Palaeontographica Africans, supplement to vol. 13: 1-22 (with 22 charts).
ARMSTRONG, R.L., 1982. Late Triassic-Early Jurassic time-scale calibration in British Columbia, Canada, In G.S. ODIN (ed.) Numerical Dating in Stratigraphy, John Wiley and Sons, New York: 509-514.
ASH, S., 1980. Upper Triassic floral zones of North America. 1n:D.L. DILCHER, and T.M. TAYLOR, (eds.) Biostratigraphy of fossil Plants. Dowden, Hutchinson and Ross, Stroudsburg: 153-170.
BOCK, W., 1952. New eastern Triassic Gink- gos. Bull. Wagner Free Inst. Sci., Philadelphia 27: 9-14.
- - - 1969. The American Triassic flora and global correlations. Geol. Center Res. Ser. 3-4, 340 p.
BROWN, AP., 1911. New cycads and conifers from the Triassic of Pennsylvania. Proc. Acad. Nat. Sci. Philadelphia, 63: 7-21.
CORNET, B., 1977a. The Palynostratigraphy and Age of the Newark Supergroup. UnpubL Ph.D. Thesis, Department of Palynology, Hie
F i g u r e 6 : A) P o s i t i o n o f t h e c o n t i n e n t s d u r i n g t h e L a t e T r i a s s i c ( C a r n i a n - N o r i a n ) showi.ng t h e o c c u r r e n c e s o f c l i m a t e s e n s i - t i v e r o c k s (C, c o a l s ; D , dune sands ; E , e v a p o r i t e s ) , t h e p h y t o s a u r - m e t o p o s a u r p r o v i n c e ( p ' , p h y t o s a u r s a n d me topo - s a u r s ; p , o n l y p h y t o s a u r s ; o , good t e t r a p o d a s s e m b l a g e s l a c k i n g p h y t o s a u r s a n d m e t o p o s a u r s ) , a n d t h e Gondwana f l o r a l p r o v i n c e ( 1 , I p s w i c h - O n s l o w m i c r o f l o r a a n d Pic-to- idium- d o m i n a t e d m a c r o f l o r a ) . B) P o s i t i o n o f t h e c o n t i n e n t s d u r i n g t h e E a r l y J u r a s s i c d u r i n g t h e E a r l y J u r a s s i c ( P l i e n s b a c h i a n ) s h o w i n g t h e o c c u r r e n c e s o f t h e " c l i m a t e s e n s i t i v e r o c k s " ( a b b r e v i a - t i o n s as i n A . a b o v e ) .
C i t a t i o n s f o r A and B g i v e n i n OLSEN a n d GALTON ( i n p r e s s ) .

CORNET & OLSEN
Pennsylvania State University: 506 pp. - - - 1977b. Preliminary investigation of
two Late Triassic conifers from York County, Pennsylvania, 1niR.C. ROMANS (ed.) Geobotany, Plenum Publishing, New York. 165-172.
- - - A, TRAVERSE, and N.G. MCDONALD, 1973. Fossil spores, pollen, and fishes from Connecticut indicate Early Jurassic age for part of the Newark Group. Science 182: 1243-1246.
- - - and A. TRAVERSE, 1975. Palynological contribution to the chronology and stratigraphy of the Hartford Basin in Connecticut and Massachusetts. Geosci. Man., 11: 1-33.
DELEVORYAS, T., and R.C. HOPE, 1971. A New Triassic cycad and i'ts phyletic implica- tions. Postilla 150: 22 p.
- - - 1975. Vottzia andfte.wt,Â¥L n. sp., an Up- per Triassic seed cone from North Caro- lina, U.S.A. Rev. Palaeobot. Palynol., 20: 67-74.
- - - 1976. More evidence for a slender growth habit in Mesozoic cycadophytes. Rev. Palaeobot. Palynol., 21: 93-100.
DOLBY, J.H. and B.E. BALME, 1976. Triassic palynology of the Carnarvon Basin, West- ern Australia. Rev. Paleobot. Palynol., 22: 105-168.
DOYLE, J.A., 1978. Origin of Angiosperms. Ann. Rev. Ecol. Syst., 9: 365-392.
EMMONS, E., 1856. Geological report of the midland counties of North Carolina. North Carolina Geol. Surv., 1852-1863, Putnuin, New York: 352 p.
- - - 1857. American Geology, pt 5., Spraugue, Albany: 152 p.
FONTAINE, W.M., 1883. Contributions to the knowleqe of the older Mesozoic flora or Virginia. U.S. Geol. Surv. Mongr., 6: 1- 144.
FROELICH, A.J. and P.E. OLSEN, 1984. Newark Supergroup, a revision of the Newark Group in eastern North America. U.S. Geol. Surv. Bull., 1537-A, 1984: A55- ASS.
HARLAND, W.B., 1982. A Geologic Time Scale, Cambridge University Press, New York: 150 p.
HOPE, R.C. and O.F. PATTERSON 111, 1969. Triassic flora from the Deep River Basin, North Carolina. North Carolina Dept. Conserv. Dev., Spec. Publ., 2: 22 p.
- - - 1 9 7 0 . PrI:iHnpt ' . i : ' i ( 5 i ( Ã § / t i ' n C i i t d : A new plant from the North Carolina Triassic. Jour. Paleo., 44: 1137-1139.
HUBERT, J.F., and K.A. MERTZ, 1980. Eolian dune field of Late Triassic age, Fundy Basin, Nova Scotia. Geology 8: 516-519.
HUGHES, N.F., 1973. Mesozoic and Tertiary distributions and problems of land plant evolution. 1 n: M. F. HUGHES (ed. ) Organ- isms and Continents Through Time, spe-
cial Papers in Palaeont., London, Pa- laeontological Association, 12: 188-198.
KUKERAH, K.P.N. and H.K. MAHESWARI, 1980. Upper Triassic sporae dispersae from the Tiki Formation 2: Miospores from the Janar Nala Section, South Gondwana Ba- sin, India. Palaeontographica Abt., B, 173: 26-84.
LINDHOLM, R.C., 1978. Triassic-Jurassic fault- ing in eastern North America - a model based on pre-Triassic structures. Geo- logy 6: 365-368.
MANSPEIZER, W., 1982. Triassic-Liassic basins and climate of the Atlantic passive margins. Geol. Rund., 71 ( 3 ) , 895-917.
ODIN, G.S. and LETOLLE, R . , 1982. The Trias- sic time scale in 1981. In: G.S. ODIM (ed.) Numerical Dating in Stratigraohy, John Wiley and Sons, New York: 523-536.
OLSEN, P.E., 1978. On the use of the term Newark for Triassic and Early Jurassic rocks of eastern North America. Newsl. Stratigr., 7 (2): 90-95.
- 1980a. A comparison of the vertebrate assemblages from the Newark and Hart- ford Basins (Early Mesozoic, Newark Supergroup) of eastern North America. 1 I : :
L. L. JACOBS, (ed.) Aspects of Vertebrate History: Essays in Honor of Edwin Harris Colbert., Museum of Northern Arizona Press, Flagstaff: 35-53.
- 1980b. Triassic and Jurassic Formations of the Newark Basin. 1n:W. ?'ANSPEIZER, (ed.) Field Studies in Mew Jersey Geol- ogy and Guide to Field Trips, 52nd Ann. Mtg. New York State Geol. Assoc., Newark College of Arts and Sciences, Rutgers University, Newark: 2-39.
- 1980~. Fossil great lakes of the Newark Supergroup in New Jersey. 1t1: W. MANSPEIZER (ed.) Field Studies in New Jersey Geology and Guide to Field Trips, 52nd Ann. Mtg. New York State Geol. Assoc., Newark College of Arts and Sciences, Rutgers University, Newark: 352-398.
- 1981. Comment on "Eolian dune field of Late Triassic age, Fundy Basin, Nova Soctia". Geology 9: 557-561.
- 1983a. Relationship between biostrati- graphic subdivisions and igneous activ- ity in the Newark Supergroup. South- eastern Sect., Geol. Soc. Amer., Abst. 15 (2): 71.
- 1983b. On the non-correlation of the Newark Su?erqroup by fossil fishes: bioqocn11 >. ip lnc , sLruc-LLIL-. i l , . i t n l sril inii.'n-
tological implications. Northeastern Sect., Geol. Soc. Amer., Abstr., 15 (3) : 121.
- 1984a. Comparative Paleolimnoloqy of the Newark Supergroup: A Study of Eco- system Evolution. Unpublished Ph.D. Thesis, Biology Department, Yale Uni- versity: 726 p.
- - - 19S4b. Periodicity of lake-level cycles

I11 C O N G R E S O T . A T I N O A M E R I C A M 0 Dl? P A L E O N T O 1 , O G I A . F L O R A S T R I A S I C A S
in the I.ato Triassic Lockatonq Forma- tion of the Mewark Basin (Newark Super- -:roup, Ilew Jersey and Pennsylvania) . ?ILA?iKOVITCH and Cl inate. liATO symposium. Dordrecht, D. Reidel Publishing.
- - - and P.K. GALLON, 1977. Triassic-Jurassic tetrapod extinctions: are they real, Science 197: 983-986.
- - - In Press. A review of the reptile and amphibian assemblnqos from thr Strom- I*rq of southern Africa, with s:>ecial emphasis on the footprints and the aqe of the Stormberq. Palaeontoloqia Africa- na, HAUGHTON Memorial Volume.
- - - C.L. REYINGTON, 3. CORNET, and K. S. THOMSON, 1978. Cyclic change in Lete Triassic lacustrine communities. Science 201: 729-733.
PARRISH, J.!!., J.T. PARRISH, and A.M. ZIEGLER, In Press. Perrao-Triassic ?aleoqeoqraphy and paleoclimatology and implications for therapsid distributions. 111: HOTFON, . , 111. (ed.).
PEARSON, D.A.B., 1970. Problems of Rhaetian stratigraphy with special reference to the lower boundary of the stage. Quart. Jour. Geol. Soc. Lond., 126: 125-150.
PEDERSEM, K.R. and J.J. LUND, 1980. Palynology of the olant-bearing Rhaetian to Hettan- qian Kap Stewart Formation, Scoresby Sund, East Greenland. Rev. Palaeobot. Palynol., 31: 1-69.
PETERSON, T.A., L.D. BROWN, F.A. COOK, S. KAUF- [.AN, and J.E. OLIVER, 1984. Structure of the Riddleville Basin from COCORP seismic data and implications for reac- tivation tectonics. Jour. Geol., 92: 261-271.
ROBBINS E.I., 1983. "Fossil Lake Danville": The Paleoecology of a Late Triassic Ecosystem on the North Carolina-Virgi- nia Border. Unpubl. Ph.D. Thesis, De- partment of Geosciences, The Pennsvlvania State University: 400 pp.
ROBINSON, P.L., 1971. A problem of faunal replacement on Permo-Triassic conti- nents. Palaeontology 14: 131-153.
- - -
RONA ,
1973. Paleoclimatoloqy and continental drift. 1n:D.H. TARLI~JG, and S.K. RUN- ;:ORN, (eds. ) Implications of Continental Drift to the Earth Sciences, Academic Press, London 451-476.
P.A., 1982. Evaporites at passive mar- gins, Geodynamics Series, Geol. Soc. tAmer., 6: 116-132.
SUTTER, J.F. and T.E. SMITH, 1979. Ar/ Ar aces of diabase intrusions from Newark trend basins i n Connecticut and Mary- land: initiation of central Atlantic Riftinq. Amer. Jour. Sci., 279: 808-831.
TOZEP, E.T., 1979. Latest Triassic ammonoid faunas and biochronoloqy, western Cana- da. Geol. Surv. Can. Pap., 79-aB: 127- 135.
TRAVERSE, A., 1983. Fundy Group palynofloru- les on the northern marqin of the Bay of Fundy, New Brunswick, Canada. North- eastern Sect., Geol. Soc. Amer. Abst., 15 (3): 122.
TRUSWELL, E.M., 1981. Pre-Cenozoic palynology and continental movements. I v t : H.W. Mc- ELHNNY (ed.) Paleoreconstruction of the continents. Geodynamics Series, Geol. SOC. Amer., 2: 13-25.
VAN HIMTE, J.E., 1976. A Jurassic time scale: Amer. Assoc. Petrol. Geol. Bull., 60; 469-497.
VAN HOUTEN, F.B., 1977. Triassic-Liassic de- posits of Morocco and eastern North America. Assoc. Petrol. Geol. Bull., 61: 79-99.
WARD, L.H., 1900. The older Mesozoic, in Sta- tus of the American flora of the Unit.ed States: United States Geological Survey 20th Annual Report, pt. 2: 213-748.
WEBB, J.A., In Press. The radiometric time scale for the Triassic: Jour. Geol. Soc. Australia.
WILLARD, B., J. FREEDMAN, D.B. MCLAUGHLIN, and others, 1959. Geology and mineral resources of Bucks County, Pennsylvania. Penn. Geol. Surv. 4th ser., Bull., C9: 243 p.