a temperature sensitive mutant of heat shock protein 70 ... · a temperature sensitive mutant of...
TRANSCRIPT

Virology 394 (2009) 28–38
Contents lists available at ScienceDirect
Virology
j ourna l homepage: www.e lsev ie r.com/ locate /yv i ro
A temperature sensitive mutant of heat shock protein 70 reveals an essential roleduring the early steps of tombusvirus replication
Robert Yung-Liang Wang 1, Jozsef Stork, Judit Pogany, Peter D. Nagy ⁎Department of Plant Pathology, University of Kentucky, 201F Plant Science Building, Lexington, KY 40546, USA
⁎ Corresponding author. Fax: +1 859 323 1961.E-mail address: [email protected] (P.D. Nagy).
1 Current address: Department of Life Science and GraScience, Chang Gung University, TaoYuan 333, Taiwan.
0042-6822/$ – see front matter © 2009 Elsevier Inc. Adoi:10.1016/j.virol.2009.08.003
a b s t r a c t
a r t i c l e i n f oArticle history:Received 18 February 2009Returned to author for revision21 March 2009Accepted 2 August 2009Available online 12 September 2009
Keywords:TombusvirusYeastViral replicationHsp70Host factorReplicaseTemperature sensitive mutant
By co-opting host proteins for their replication, plus-stranded RNA viruses can support robust replication andsuppress host anti-viral responses. Tomato bushy stunt virus (TBSV) recruit the cellular heat shock protein 70(Hsp70), an abundant cytosolic chaperone, into the replicase complex. By taking advantage of yeast modelhost, we demonstrate that the four-member SSA subfamily of HSP70 genes is essential for TBSV replication.The constitutively expressed SSA1 and SSA2, which are resident proteins in the viral replicase, can becomplemented by the heat-inducible SSA3 and/or SSA4 for TBSV replication. Using a yeast strain carrying atemperature sensitive ssa1ts, but lacking functional SSA2/3/4, we show that inactivation of Ssa1pts led to adefect in membrane localization of the viral replication proteins, resulting in cytosolic distribution of theviral proteins and lack of replicase activity. An in vitro replicase assembly assay with Ssa1pts revealed thatfunctional Ssa1p is required during the replicase assembly process, but not during minus- or plus-strandsynthesis. Temperature shift experiments from nonpermissive to permissive in ssa1ts yeast revealed that there-activated Ssa1pts could promote efficient TBSV replication in the absence of other SSA genes. We alsodemonstrate that the purified recombinant Ssa3p can facilitate the in vitro assembly of the TBSV replicase onyeast membranes, demonstrating that Ssa3p can fully complement the function of Ssa1p. Taken together, thecytosolic SSA subfamily of Hsp70 proteins play essential and multiple roles in TBSV replication.
© 2009 Elsevier Inc. All rights reserved.
Introduction
Due to limiting coding capacity of their genomes, plus-stranded(+)RNA viruses rely extensively on the host during their replication.These viruses hijack subcellular membranes and use the componentsof the host cells to make viral proteins and replicate the viral RNA.Moreover, they co-opt selected host proteins to facilitate viral genomereplication (Ahlquist et al., 2003; Nagy, 2008; Noueiry and Ahlquist,2003; Salonen et al., 2005; Shi and Lai, 2005). Indeed, recent genome-wide screens with Brome mosaic virus (BMV), Tomato bushy stuntvirus (TBSV), Drosophila C virus, hepatitis C virus and West Nile virusrevealed that more than 100 host proteins and many cellularpathways affected replication and infections by each (+)RNA virus(Cherry et al., 2005; Jiang et al., 2006; Krishnan et al., 2008; Kushneret al., 2003; Panavas et al., 2005b; Randall et al., 2007; Serviene et al.,2005). The functions of most of the identified host proteins duringvirus replication, however, are currently unknown.
TBSV, a small (+)RNA virus is used to dissect the roles of hostproteins within the viral replicase, which is the key enzyme for viralgenome replication. The tombusvirus replicase has been intensively
duate Institute of Basic Medical
ll rights reserved.
characterized via proteomics approaches, in vitro replication assaysand development of yeast as a model host (Li et al., 2008, 2009; Nagy,2008; Nagy and Pogany, 2000; Panavas and Nagy, 2003; Panavieneet al., 2004; Pogany and Nagy, 2008; Serva and Nagy, 2006; Wang andNagy, 2008). Previous work revealed that TBSV RNA replicationdepends on two viral-encoded proteins, namely the RNA-dependentRNA polymerase (p92pol RdRp) and p33 replication co-factor, the keyprotein in recruitment of the viral RNA into replication (Monkewichet al., 2005; Pogany and Nagy, 2008; Pogany et al., 2008, 2005). Theperoxisome membrane-bound TBSV replicase also contains 6-to-10host proteins, which are likely involved in most activities of thereplicase (Nagy and Pogany, 2006; White and Nagy, 2004). Theidentified host proteins within the tombusvirus replicase include heatshock protein 70 (Hsp70, coded by the constitutively expressed SSA1and SSA2 genes in yeast) and glyceraldehyde-3-phosphate dehydro-genase (GAPDH, coded by TDH2 and TDH3 genes in yeast), whichbinds to the TBSV (-)RNA and affects plus-strand synthesis (Wang andNagy, 2008). The replicase also contains translation elongation factor1A (eEF1A), which binds to a cis-acting regulatory element in theTBSV (+)RNA as well as to p33 co-factor (Li et al., 2009). Anotherhost-derived component is Cdc34p ubiquitin-conjugating enzyme,which ubiquitinates the p33 replication co-factor (Li et al., 2008).Down-regulation of these host factors inhibited, whereas their over-expression increased TBSV accumulation in yeast model host (Li et al.,2008; Serva and Nagy, 2006; Wang and Nagy, 2008) suggesting that

29R.Y.-L. Wang et al. / Virology 394 (2009) 28–38
they play significant roles in TBSV replication. In addition, Pex19pcytosolic transport protein binds transiently to the viral replicationproteins as well as to the replicase complex, likely facilitating thetransport of the replication proteins to the peroxisomal membranes,the site of replication (Pathak et al., 2008). The functions of the abovehost proteins within the viral replicase are currently under intensiveinvestigations.
The host-coded Hsp70 chaperone family, which represents amajorgroup among the heat shock proteins, and its co-chaperones have beensuggested to promote replication of several (+)RNA and (-)RNAviruses (Brown et al., 2005; Dufresne et al., 2008; Nishikiori et al.,2006; Qanungo et al., 2004; Weeks and Miller, 2008). Based on theknown cellular functions, Hsp70 and other chaperoneswere proposedto stimulate viral RdRp activity (Momose et al., 2002), and participatein the assembly of the viral replicase and enhance replication(Kampmueller and Miller, 2005; Tomita et al., 2003; Weeks andMiller, 2008). Plants infected by various plant viruses express cytosolicHsp70 proteins at elevated levels, indicating that Hsp70 could play animportant role during viral infections (Aparicio et al., 2005; Arandaet al., 1996; Whitham et al., 2003; Whitham et al., 2006). Moreover, aviral-encodedHsp70-like protein is involved in the assembly of virionsand cell-to-cell movement of closteroviruses (Alzhanova et al., 2001;Peremyslov et al., 1999). In addition, Hsp70-like proteins are involvedin nuclear localization, genome replication and cell transformation ofDNA viruses, cell entry, virion assembly and disassembly, envelopeprotein maturation, folding of capsid proteins, and viral transcriptionby various viruses (Mayer, 2005). Additional cellular chaperones, suchasHsp90proteins or the J-domain containingHsp40proteins have alsobeen shown to affect virus replication, including activation of reversetranscriptase for hepadnaviruses (Hu et al., 2004; Stahl et al., 2007;Tavis et al., 1998), or assembly of the BMV replicase (Tomita et al.,2003). Most of the above studies point toward Hsp70 and othercellular chaperones as major players during virus replication.
Host proteins could play different roles during tombusvirusreplication. Currently, tombusvirus replication is divided into sixsequential steps: RNA template selection by p33 replication protein;recruitment of the replication protein-viral RNA complex to the site ofreplication; assembly of the viral replicase complex; synthesis of viralprogeny RNAs, including minus- and plus-strand synthesis; release ofthe newly made plus-strand viral RNAs from the replicase; anddisassembly of the viral replicase (Nagy and Pogany, 2006). Hsp70proteins might play multiple roles/functions in TBSV replication.Accordingly, the two cytosolic Ssa1p and Ssa2p Hsp70 proteinsexpressed constitutively in yeast have been shown to be recruitedfrom the cytosol to the peroxisomal membrane (the site of TBSVreplication) via interaction with the p33 replication co-factor (Servaand Nagy, 2006; Wang et al., 2009). Down-regulation or over-expression of Ssa1/2p in yeast resulted in reduced and elevated levelof TBSV RNA accumulation, respectively, suggesting that these Hsp70proteins are important for TBSV RNA replication (Serva and Nagy,2006). These Hsp70 proteins are likely involved directly in replicationsince Ssa1/2p have been shown as components of the highly purifiedtombusvirus replicase complex (Serva and Nagy, 2006). Using aHSP70 mutant yeast (ssa1ssa2), we found that the viral replicationproteins remained cytosolic at an early time point, suggesting thatHsp70 is involved in subcellular localization of the viral replicationproteins to intracellular membranes (Wang et al., 2009). A novel invitro replication assay also showed that Ssa1/2p are essential for theassembly of the TBSV replicase (Pogany et al., 2008). An in vitromembrane insertion assay demonstrated that Hsp70 promoted theinsertion of the viral replication proteins into the subcellularmembranes (Wang et al., 2009). These functions of Hsp70 are notrestricted to in vitro or yeast-based assays, since knockdown ofcytosolic Hsp70 in plants or inhibition of Hsp70 with a chemicalinhibitor were found to inhibit TBSV replication in a plant host (Wanget al., 2009).
The SSA subfamily of cytosolic Hsp70 consists of four genes andexpression of at least one of the four SSA genes at high level is neededfor yeast viability (Ahsen and Pfanner, 1997). It is currently unknownif the function of Ssa1/2p is essential for TBSV replication andwhetheradditional cellular Hsp70 or other heat shock proteins can comple-ment the functions of Ssa1/2p for supporting TBSV replication. Toaddress these questions and to further dissect the roles of Hsp70 inTBSV replication, in this work, we used a yeast strain lackingfunctional SSA2, SSA3 and SSA4 genes, while the SSA1 gene was eitherwt or temperature sensitive (ts). Based on the ssa1ts strain, wedemonstrate that the Ssa-subfamily of Hsp70 proteins is essential forTBSV replication in yeast. Using biochemical and temperature shiftexperiments, we show that Hsp70 is essential during the early steps ofTBSV replication, but not during minus- and plus-strand synthesis.
Results
TBSV replication can be complemented by a heat shock-inducible hostfactor in ssa1ssa2 mutant yeast
Previous work has shown that ssa1ssa2 mutant yeast can stillsupport TBSV replication, albeit less efficiently and with significantdelay when compared with the wt yeast (Wang et al., 2009). Theseobservations indicate that either Ssa1p/Ssa2p are not essential forTBSV replication or they can be partially complemented possibly byother cellular chaperones. To test if complementation of TBSVreplication can be enhanced by prior induction of heat shock proteins(Boorstein and Craig, 1990), we incubated ssa1ssa2 mutant yeast at42 °C for 30 min prior to the induction of p33/p92pol proteins and theTBSV DI-72 repRNA. In comparison with ssa1ssa2mutant yeast grownat 23 °C all the time (no heat shock), which support only 15% repRNAaccumulation, the ssa1ssa2 mutant yeast treated with short heat-shock reached 80% repRNA accumulation of that measured in wt yeast(no heat shock) (Fig. 1A versus B). The stimulating effect of the shortheat shock treatment prior to TBSV replication in ssa1ssa2 mutantyeast suggests that a heat-inducible host factor, likely a heat-shockprotein, can efficiently complement the missing Ssa1/2p functions forTBSV replication.
The purified heat-inducible Ssa3p Hsp70 can facilitate the assembly ofthe TBSV replicase in vitro
It is possible that the heat-inducible host factor supporting TBSVreplication in ssa1ssa2 mutant yeast, which could also be responsiblefor the partial complementation of TBSV replication at the standardtemperature (Wang et al., 2009), is the cytosolic, stress-inducibleSsa3p and Ssa4p Hsp70 proteins, which are highly similar to oneanother and show 80% sequence identity with Ssa1/Ssa2p (Becker etal., 1996; Lin et al., 2001). Accordingly, Ssa3/4p are expressed at highlevels in ssa1ssa2 cells (Becker et al., 1996) and they can be efficientlyinduced by a short heat-shock treatment (Boorstein and Craig, 1990).
To study if Ssa3p could replace Ssa1p for supporting TBSVreplication, we used the recently developed TBSV replication assaybased on the addition of purified recombinant p33/p92pol and the DI-72 repRNA to a yeast cell-free extract (Pogany et al., 2008). The invitro assembled TBSV replicase is capable of supporting authenticTBSV repRNA replication if ribonucleotides are provided (Pogany andNagy, 2008; Pogany et al., 2008). We have shown previously thatSsa1p is required for the in vitro assembly of the TBSV replicase whenonly the membrane fraction of the cell-free extract is used (Poganyet al., 2008).
To test if Ssa3p can support the assembly of the TBSV replicase invitro, we used purified recombinant Ssa3p (fused with FLAG-tag)together with purified recombinant p33/p92pol and the DI-72 repRNAand mixture of ribonucleotides in combination with the membranefraction of the yeast cell-free extract. The in vitro replication assay

Fig. 1. Increased accumulation of TBSV repRNA in ssa1ssa2 mutant yeast after shortheat shock treatment. (A) Northern blot analysis of TBSV repRNA accumulation level inssa1ssa2mutant yeast cultured continuously at 23 °C. TBSV repRNA accumulates to 15%in ssa1ssa2mutant yeast of that found in wt yeast at the 16 h time point after inductionby galactose. The ribosomal (r)RNA is used as a loading control. (B) After 30 min heatshock treatment of yeast at 42 °C, TBSV repRNA replication was launched by expressionof p33/p92 and DI-72(+)RNA from the GAL1/GAL10 promoters in ssa1ssa2 mutantyeast by using culture media containing galactose, followed by shaking at 23 °C.Northern blot analysis shows that TBSV repRNA accumulated to ∼80% in the heat-treated ssa1ssa2 mutant yeast when compared to the wt yeast at the 16 h time point.(C) Ssa3p can facilitate the in vitro assembly of the TBSV replicase. The purifiedrecombinant FLAG-Ssa3p (lane 2) and FLAG-Ssa1p (lane 1) were added together withpurified TBSV p33 (lanes 1–3) and p92pol (lanes 1–3) to the membrane fraction of acell-free extract prepared from untransformed yeast, which was programmed via theaddition of 1 μg TBSV DI-72 (+)repRNA (lanes 1–3). The newly synthesized repRNA(marked with an arrowhead), representing mostly (+)repRNA due to a full cycle ofreplication, by the in vitro assembled TBSV replicase is analyzed by denaturing PAGEanalysis. The bottom panel shows the Coomassie-blue-stained SDS-PAGE gel with thepurified recombinant FLAG-Ssa3p and FLAG-Ssa1p.
Fig. 2. Lack of TBSV repRNA replication in ssa1ts yeast at the nonpermissivetemperature. (A) Top panels: Northern blot analysis of TBSV repRNA accumulation inssa1ts yeast and SSA1wt yeast grown either at 32 °C or 36 °C, which inactivates Ssa1pts.Bottom panels: Ethidium bromide stained agarose gel shows repRNA and ribosomalrRNA levels in the total RNA samples in ssa1ts and SSA1wt yeast strains 16 h after theinduction of TBSV repRNA replication via galactose. Note that YFP-p33 and CFP-p92each expressed from the GAL1 promoter, whereas DI-72 repRNA was expressed fromthe GAL10 promoter. (B) Western blot analysis to show the accumulation of YFP-p33and CFP-p92 in ssa1ts yeast and SSA1wt yeast grown at 32 °C. The yeast samples werethe same as in panel A. (C) An in vitro replicase assay to test TBSV RNA synthesis inssa1ts yeast and SSA1wt yeast grown at 32 °C. We obtained the membrane fractioncarrying the tombusvirus replicase from ssa1ts yeast and SSA1wt yeast grown at 32 °Cfor 16 h after the addition of galactose to induce TBSV replication. Note that thereplicase contains the endogenous repRNA template. (D) Similar in vitro replicase assaywas used to test TBSV RNA synthesis in ssa1ts yeast and SSA1wt yeast grown at 23 °C. Weobtained the membrane fraction carrying the tombusvirus replicase from ssa1ts yeastand SSA1wt yeast grown at 23 °C for 16 h after the addition of galactose to induce TBSVreplication. The bottom panel shows a Western blot for p33 level in each membrane-bound replicase preparations.
30 R.Y.-L. Wang et al. / Virology 394 (2009) 28–38
revealed that the recombinant Ssa3p was capable of assembling theactive TBSV replicase (Fig. 1C, lane 2), though less efficiently thanSsa1p (lane 1). Since the replicase assay required Ssa3p (Fig. 1C, lanes2 versus 3), these data are consistent with the proposed role of Ssa3pin complementing the missing functions of Ssa1/2p in ssa1ssa2mutant yeast (Fig. 1B).
Expression of Ssa1pts in yeast to study the role of Hsp70 in TBSVreplication
There are four SSA genes in yeast coding for cytosolic Hsp70proteins and deletion of all SSA genes makes yeast nonviable. Todemonstrate if SSA1–4 genes are essential for TBSV replication, we

Fig. 3. Cytosolic localization of p33 replication protein in ssa1ts yeast grown at 36 °C. (A)Confocal laser microscopy shows the localization of YFP-p33 in ssa1ts yeast and SSA1wt
yeast grown at either 23 °C or 36 °C for 14 h after induction from GAL1 promoter. Thepunctate structures show peroxisomal localization, while the “doughnut” -shapestructure indicates cytosolic localization of p33 in yeast cells. The punctate structuresformed in ssa1ts yeast grown at 23 °C are reminiscent of those found in SSA1wt yeast(left side of the panel). (B) Cytosolic localization of peroxisome-targeted GFP, termedGFP-SKL, in ssa1ts yeast grown at 36 °C for 14 h after induction from GAL1 promoter.Note that GFP-SKL carries the peroxisomal targeting sequence “SKL” at the C-terminus.
31R.Y.-L. Wang et al. / Virology 394 (2009) 28–38
used yeast strains with nonfunctional SSA2/3/4 genes, whereas SSA1gene was either wt (SSA1wt ssa2 ssa3 ssa4) or ts (ssa1ts ssa2 ssa3 ssa4).We will refer to these strains as SSA1wt and ssa1ts below. Theadvantage of the ssa1ts is that this yeast strain can be cultured at 29 °Cor below, while Ssa1pts becomes nonfunctional at 35 °C or above,although the cells do not show abnormal morphology or cell cycle-specific arrest at 37 °C (Becker et al., 1996). Ssa1pts was obtained viamutagenesis and it contains P417-L mutation within the peptide-binding domain. The growth inhibition at the elevated temperature inssa1ts cells is cytostatic (not cytocidal) since cells start growing aftertemperature shift to 30 °C even after kept at 37 °C for 3 days (Beckeret al., 1996).
Yeast strains ssa1ts or SSA1wt were transformed with plasmids toexpress p33/p92pol and the repRNA from GAL1/GAL10 promoters andTBSV replication was launched by adding galactose to the culturemedia, followed by switching the temperature of incubation to 32 °C
Fig. 4. The assembled tombusvirus replicase in the membrane fraction of ssa1ts yeast is nproduced by tombusvirus replicase preparations in vitro obtained from ssa1ts yeast and SSA137 °C. The membrane fraction carrying the tombusvirus replicase from ssa1ts yeast and SSA1w
was used in the in vitro replicase assay based on the endogenous repRNA template.
or 36 °C. Northern blot analysis of the RNA samples obtained after 18h of incubation at 32 °C showed accumulation of TBSV repRNA to lessthan 1% in ssa1ts yeast when compared to that of SSA1wt (Fig. 2A, lanes3–4 versus 1–2). The accumulation of TBSV repRNA at 36 °C wasbelow detection limit in ssa1ts yeast when compared to that of SSA1wt
(Fig. 2A, lanes 8–10 versus 5–7). This low level of repRNAaccumulation in ssa1ts yeast grown at 32/36 °C is likely due toplasmid-borne transcription of repRNA (Panavas and Nagy, 2003),indicating the lack of replication of the TBSV repRNA at 32 °C or at36 °C in ssa1ts yeast. Accordingly, isolation of the membrane-boundtombusvirus replicase complexes from the above yeast strains,followed by in vitro replicase assay using the co-purified TBSVrepRNA showed undetectable level of replicase activity from ssa1ts
yeast grown at 32 °C, while the comparable assay from SSA1wt grownat 32 °C showed easily detectable replicase activity in vitro (Fig. 2C). Incontrast, the membrane-bound tombusvirus replicase from ssa1ts
yeast grown at 23 °C was almost as active as that obtained fromSSA1wt grown at 23 °C (Fig. 2D). Thus, as expected, ssa1ts yeast waslikely defective in assembling the tombusvirus replicase at thenonpermissive temperature.
Western blot analysis of total protein extracts form ssa1ts orSSA1wt yeast strains revealed comparable accumulation of p33 (Fig.2B) and p92pol (not shown) replication proteins at 32 °C. Thus, it isunlikely that Ssa1–4p affect the translation or degradation of the viralreplication proteins. Taken together, all these results suggest thatreplicase activity or the assembly of the replicase is completelyblocked at 32/36 °C in the yeast strain carrying ssa1ts and lacking thecomplementing SSA2/3/4 genes. Thus, the SSA subfamily (SSA1/2/3/4) of HSP70 genes is essential for TBSV replication and their functionscannot be complemented by other HSP70 or additional heat shockgenes.
Yeast carrying ssa1ts shows defect in localization of the viral p33replication protein when grown at 36 °C
Since p33 and p92pol replication proteins become cytosolic inssa1ssa2 mutant yeast strain at early time points instead of formingthe characteristic punctate structures on the peroxisomal membranesin wt yeast (Wang et al., 2009), we tested the intracellular localizationof p33 in ssa1ts yeast. The expression of the yellow fluorescentprotein-tagged p33 replication protein (YFP-p33) was initiated fromthe GAL1 promoter by the addition of galactose to the culture media,followed by culturing at either 23 °C or 36 °C. Confocal lasermicroscopy of ssa1ts yeast cells after 18 h of culturing at 36 °Cindicated cytosolic (showing “doughnut”-like distribution) localiza-tion of YFP-p33 (Fig. 3A). This is in contrast with the formation of thepunctate structures by YFP-p33 when studied in ssa1ts yeast grown at23 °C or SSA1wt cultured at either 23 °C or 36 °C (Fig. 2A). These datasuggest a defect in subcellular localization and the transport of p33 tothe peroxisomemembrane when none of the Ssa1–4p was functional.
Since p33, similar to many other peroxisomal membrane proteins,has been shown to be transported to the peroxisome by Pex19p
ot temperature sensitive. Denaturing PAGE analysis shows the radiolabeled repRNAswt yeast grown at 23 °C. The in vitro replication assays were performed at either 25 °C ort yeast grown at 23 °C for 16 h after the addition of galactose to induce TBSV replication

32 R.Y.-L. Wang et al. / Virology 394 (2009) 28–38
cytosolic shuttle protein (Pathak et al., 2008), it is possible that otherperoxisomal proteins might also be mislocalized in ssa1ts yeast at36 °C. To test this question, we expressed GFP-SKL, which carries theC-terminal SKL sequence known to target proteins to peroxisome inwt yeast (Kragler et al., 1993), from the GAL1 promoter. As expected,GFP-SKL was localized to the peroxisomal membrane, forming thepunctate structure, in ssa1ts yeast grown at the permissive 23 °Ctemperature (Fig. 3B). In contrast, GFP-SKL was localized in thecytosol in ssa1ts yeast grown at the nonpermissive 36 °C (Fig. 3B). Thissuggests that the transportation of peroxisomal proteins as well asp33 replication protein to the peroxisome does not occur in theabsence of functional Ssa1–4p proteins.
The assembled replicase from ssa1ts yeast is not sensitive to inactivationof Ssa1pts
To test if the SSA genes are required after the assembly of the viralreplicase, we isolated the tombusvirus replicase bound to themembrane from ssa1ts or SSA1wt yeast cultured at 23 °C, followed byin vitro replicase assay conducted at either 25 °C or the nonpermissive37 °C. These experiments revealed that the tombusvirus replicasefrom ssa1ts yeast was as active at the nonpermissive 37 °C as at 25 °C
Fig. 5. Ssa1p is required during the in vitro assembly of the TBSV replicase, but notduring viral RNA synthesis. (A) A step-wise approach was used to separate the earlysteps, such as TBSV replicase assembly, from the late steps, which include viral RNAsynthesis. The various components used in the procedure are listed. (B) DenaturingPAGE analysis of the radiolabeled in vitro replicase products obtained after the in vitroreconstitution of the TBSV replicase using the conditions shown during step 1 and step3. (C) Similar analysis of the in vitro TBSV replicase assay with Ssa1pts or Ssa1pwt as inpanel B, except using different conditions during step 1 and step 3 as shown. Note thatsamples in lanes 5–8 contained no Ssa1pts or Ssa1pwt during the replicase assembly.
or the replicase derived from SSA1wt yeast (Fig. 4). Thus, the activity ofthe preassembled, membrane-bound tombusvirus replicase contain-ing the endogenous repRNA does not depend on the presence of afunctional Ssa1p.
To test even more rigorously if Ssa1p is required only before/during the assembly of the tombusvirus replicase complex, but notduring RNA synthesis, we used our recently developed in vitroreplicase assembly assay (Pogany et al., 2008). In this assay (Fig. 5A),we added either the purified Ssa1pts or Ssa1pwt in combination withpurified p33/p92pol replication proteins and DI-72(+)repRNA to themembrane fraction of yeast cell-free extract. Importantly, we includedonly rATP and rGTP during the replicase assembly, which is requiredand sufficient to support in vitro assembly, but cannot support RNAsynthesis that needs all four ribonucleotides. After the in vitroassembly, we collected the membrane fraction by centrifugation andremoved all the soluble proteins/RNAs, including those Ssa1p/p33/p92pol molecules not bound to membranes. Then, we added thereplicase buffer and all four ribonucleotides, including 32P-UTP, to theabove membrane fraction to allow the RNA synthesis to occur by thepreassembled TBSV replicase. Importantly, the removal of the solublefraction before the replicase assay prevented the possibility ofreplicase assembly and RNA synthesis to take place simultaneously(Fig. 5A).
When we used 20 °C during the assembly step and either 20 °C or29 °C during the RNA synthesis step, then we observed robust in vitroreplication regardless of using Ssa1pts or Ssa1pwt (Fig. 5B). SinceSsa1pts is partially inactive at 29 °C (Pogany et al., 2008) in the in vitroreplication assay, we propose based on the above results that Ssa1pts
is needed during the replicase assembly process, but not during RNAsynthesis, which include bothminus- and plus-strand synthesis in ourin vitro replication assay (Pogany and Nagy, 2008; Pogany et al.,2008). Accordingly, whenwe used 29 °C during the assembly step and20 °C during the RNA synthesis step, then we observed TBSV repRNAreplication occurring only in case of the Ssa1pwt, but not with theTBSV replicase containing Ssa1pts (Fig. 5C, lanes 2, 4 versus 1 and 3).These data suggest that Ssa1pts was defective in replicase assembly at29 °C in vitro, preventing the subsequent viral RNA synthesis at 20 °Cor 29 °C. Altogether, the obtained results are in agreement with themodel that Ssa1p is required prior/during the replicase assembly, butit is not needed during minus- and plus-strand synthesis in vitro.
Downshift to 23 °C restores peroxisomal localization of p33 replicationprotein in ssa1ts yeast
Since ssa1ts yeast cells survive short periods (up to 3 days) at thenonpermissive temperature (Becker et al., 1996), we used tempera-ture downshift experiments to test how TBSV replication is affectedby the reactivation of Ssa1pts by shifting to the permissivetemperature. First, we cultured ssa1ts yeast expressing YFP-p33 at32 °C, followed by decreasing the temperature to 23 °C and shuttingdown the production of YFP-p33 by changing the carbon-source in themedia from galactose to glucose. Using confocal microscopy, weobserved that about 50% of yeast cells contained the characteristicpunctate structures 6 h after the downshift, suggesting peroxisomallocalization of YFP-p33 (Fig. 6A). The other 50% of yeast cells stillcontained cytosolic YFP-p33, suggesting that the availability of thereactivated Ssa1pts is limited (Fig. 6A). Fractionation of the extracts ofssa1ts yeast cells 6 h after the switch to 23 °C also showed similar ∼50–50% distribution of YFP-p33 between the soluble or membrane-containing fractions (Fig. 6B, lanes 1–3 versus 7–9). In contrast, YFP-p33 expressed in ssa1ts yeast at the nonpermissive temperature wasfound mostly in the soluble fraction (Fig. 6C, 0 h time point). Asexpected, YFP-p33 expressed in SSA1wt yeast was present in themembranous fraction regardless of the growth conditions (Fig. 6B–C,0 and 6 h time points). The cytosolic cellular PGK protein and themembrane-bound cellular ALP protein were found in the expected

Fig. 6. Re-distribution of p33 from the cytosol to the peroxisomes after shifting down the temperature from nonpermissive to permissive in ssa1ts yeast. (A) Confocal lasermicroscopy shows the localization of YFP-p33 in ssa1ts yeast grown at 23 °C in a media containing glucose (to inhibit the synthesis of new YFP-p33 molecules) for 6 h after culturingthe yeast for 24 h at 32 °C in a media containing galactose. Note that the characteristic punctate structures indicating peroxisomal localization (Jonczyk et al., 2007; McCartney et al.,2005; Panavas et al., 2005a) are formed only in ∼50% of ssa1ts yeast cells. (B) Re-localization of p33 to the membrane fraction in ssa1ts yeast after shifting down the temperature to23 °C based on fractionation experiments. Disrupted yeast cells were separated to soluble (cytosolic) and membrane fractions via differential centrifugation, followed by Westernblotting to detect YFP-p33. Top panel: the samples were taken 6 h after downshifting to 23 °C (6 h). Bottom panel: PGK, a soluble yeast protein, and ALPmembrane protein were usedas internal controls in the Western blots. See panel A for further details about the growing conditions for yeast samples. (C) Western blot analysis of localization of YFP-p33 in ssa1ts
yeast cells when the samples were taken prior to downshifting to 23 °C (0 h). Note the partial re-distribution of YFP-p33 from the soluble fraction at the 0 time point (panel C) to themembrane fraction at the 6 h time point (panel B) in ssa1ts yeast.
33R.Y.-L. Wang et al. / Virology 394 (2009) 28–38
fractions (Fig. 6B). Altogether, these data are consistent with themodel that correct peroxisomal localization of YFP-p33 requires afunctional Ssa1p and reactivation of ssa1ts by downshifting to 23 °Cresults in re-localization of p33 from the cytosol to the peroxisomalmembrane in half of the ssa1ts yeast cells by the 6 h time point.
To test how rapidly YFP-p33 is re-localized to the membrane, weused time course experiments based on confocal microscopy and cellfractionation. We found that 30 min after the downshift to 23 °C, asmall number of cells already showed several punctate structures,though the punctate structures became more visible by microscopyand detectable in the membrane fraction after 1–2 h (Fig. 7A and D).The amounts of YFP-p33 and CFP-p92 in the membrane fractionbecame as abundant after 3 h as after 6 h (Fig. 7D, lanes 7–8 versus11–12). Interestingly, the sizes of individual punctate structuresbecame bigger over time after the temperature downshift in theabsence of new YFP-p33 synthesis (Fig. 7A). This suggests on-goingmembrane rearrangement caused by the insertion of p33 into themembrane due to the presence of activated Ssa1pts. Moreover, itseems that it requires 30min to 3 h for the re-activated Ssa1pts to helpre-localize YFP-p33 from the cytosol to the membrane. We observedthe characteristic punctate structures with YFP-p33 in SSA1wt yeastshifted from 32 °C to 23 °C or grown continuously at 23 °C over time(Fig. 7B–C), suggesting that YFP-p33 localized correctly and efficientlyin the presence of functional Ssa1pwt at both temperatures.
Downshift to 23 °C activates the tombusvirus replicase in ssa1ts yeast
To test if the above observed changes in the subcellular localizationof YFP-p33 during downshift to 23 °C correlates with enhancedtombusvirus replicase activity, we measured the replicase activity onthe co-purified repRNA in the membrane fraction of ssa1ts yeast. Weadded either cycloheximide or glucose to the growth media to inhibitthe production of new YFP-p33 and CFP-p92 in ssa1ts yeast afterdownshifting the temperature (Fig. 8A–B). It is important to note thatthe cycloheximide treatment down-regulates the production of allnew host and viral proteins, while glucose selectively inhibits theproduction of new YFP-p33 and CFP-p92, which are driven from theGAL1 promoter. These in vitro experiments revealed that the replicaseactivity continuously increased in samples taken from 30 min to4–6 h, reaching to ∼30% level of the replicase activity obtained withthe preparation from ssa1ts yeast cultured at 23 °C (Fig. 8A). Thus, ittakes from 30min to 3–4 h for the re-activated Ssa1pts to facilitate theassembly/activation of the tombusvirus replicase in the absence ofnew protein synthesis (+cyclehaximide, Fig. 8A, top panel). Similarly,the membrane-localization of YFP-p33 also needed from 60 min to 3–4 h under the same conditions (Fig. 8A, bottom panels). The picturewas somewhat similar in the first 2 h after the downshift to 23 °C inssa1ts yeast cultured in the presence of glucose, which inhibits theproduction of new YFP-p33 and CFP-p92 (Fig. 8B). However, the

34 R.Y.-L. Wang et al. / Virology 394 (2009) 28–38
replicase activity at 3–4 h time points reached up to ∼40% of thereplicase activity obtained with the preparation from ssa1ts yeastcultured at 23 °C (Fig. 8B). This moderate increase in replicase activity
Fig. 7. Time-course experiments to analyze re-distribution of p33 from the cytosol to the pssa1ts yeast. (A) Confocal laser microscopy shows the localization of YFP-p33 in ssa1ts yeast gp92/repRNA) until the shown time point after switching from 32 °C. See the details about ystructures containing YFP-p33 grew over time in ssa1ts yeast cells. (B) Localization of YFP-ppoint after switching from 32 °C. See further details in panel A. (C) Localization of YFP-p33 inblot analysis of fractionation experiments shows the increasing extent of re-localization ofshifting down the temperature to 23 °C. See the details for yeast culturing in panel A. The fracprotein extracts prepared from yeast grown as described in panels A and D.
when compared with the cycloheximide treatment (Fig. 8A) is likelydue to the newly produced active Ssa1pts after the downshift thatseems to affect protein functions by the 4-h time point. Indeed, we
eroxisomes after shifting down the temperature from nonpermissive to permissive inrown at 23 °C in a media containing glucose (to shut down the production of new p33/east culturing in the legend to Fig. 6A. Note that the sizes of the characteristic punctate33 in SSA1wt yeast grown at 23 °C in a media containing glucose until the shown timeSSA1wt yeast grown continuously at 23 °C in a media containing galactose. (D) Westernp33 and p92pol from the soluble fraction to the membrane fraction in ssa1ts yeast aftertions were obtained as described in Fig. 6B. (E)Western blot analysis of p33 level in total
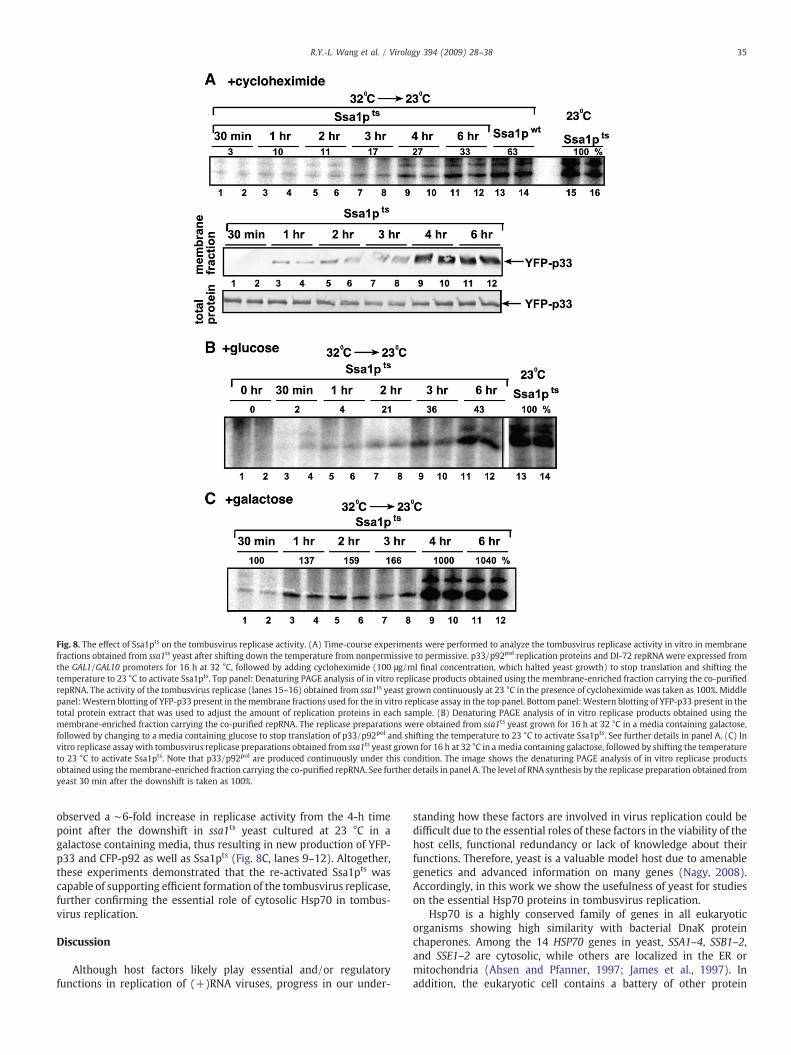
Fig. 8. The effect of Ssa1pts on the tombusvirus replicase activity. (A) Time-course experiments were performed to analyze the tombusvirus replicase activity in vitro in membranefractions obtained from ssa1ts yeast after shifting down the temperature from nonpermissive to permissive. p33/p92pol replication proteins and DI-72 repRNA were expressed fromthe GAL1/GAL10 promoters for 16 h at 32 °C, followed by adding cycloheximide (100 μg/ml final concentration, which halted yeast growth) to stop translation and shifting thetemperature to 23 °C to activate Ssa1pts. Top panel: Denaturing PAGE analysis of in vitro replicase products obtained using the membrane-enriched fraction carrying the co-purifiedrepRNA. The activity of the tombusvirus replicase (lanes 15–16) obtained from ssa1ts yeast grown continuously at 23 °C in the presence of cycloheximide was taken as 100%. Middlepanel: Western blotting of YFP-p33 present in themembrane fractions used for the in vitro replicase assay in the top panel. Bottom panel: Western blotting of YFP-p33 present in thetotal protein extract that was used to adjust the amount of replication proteins in each sample. (B) Denaturing PAGE analysis of in vitro replicase products obtained using themembrane-enriched fraction carrying the co-purified repRNA. The replicase preparations were obtained from ssa1ts yeast grown for 16 h at 32 °C in a media containing galactose,followed by changing to a media containing glucose to stop translation of p33/p92pol and shifting the temperature to 23 °C to activate Ssa1pts. See further details in panel A. (C) Invitro replicase assay with tombusvirus replicase preparations obtained from ssa1ts yeast grown for 16 h at 32 °C in amedia containing galactose, followed by shifting the temperatureto 23 °C to activate Ssa1pts. Note that p33/p92pol are produced continuously under this condition. The image shows the denaturing PAGE analysis of in vitro replicase productsobtained using themembrane-enriched fraction carrying the co-purified repRNA. See further details in panel A. The level of RNA synthesis by the replicase preparation obtained fromyeast 30 min after the downshift is taken as 100%.
35R.Y.-L. Wang et al. / Virology 394 (2009) 28–38
observed a ∼6-fold increase in replicase activity from the 4-h timepoint after the downshift in ssa1ts yeast cultured at 23 °C in agalactose containing media, thus resulting in new production of YFP-p33 and CFP-p92 as well as Ssa1pts (Fig. 8C, lanes 9–12). Altogether,these experiments demonstrated that the re-activated Ssa1pts wascapable of supporting efficient formation of the tombusvirus replicase,further confirming the essential role of cytosolic Hsp70 in tombus-virus replication.
Discussion
Although host factors likely play essential and/or regulatoryfunctions in replication of (+)RNA viruses, progress in our under-
standing how these factors are involved in virus replication could bedifficult due to the essential roles of these factors in the viability of thehost cells, functional redundancy or lack of knowledge about theirfunctions. Therefore, yeast is a valuable model host due to amenablegenetics and advanced information on many genes (Nagy, 2008).Accordingly, in this work we show the usefulness of yeast for studieson the essential Hsp70 proteins in tombusvirus replication.
Hsp70 is a highly conserved family of genes in all eukaryoticorganisms showing high similarity with bacterial DnaK proteinchaperones. Among the 14 HSP70 genes in yeast, SSA1–4, SSB1–2,and SSE1–2 are cytosolic, while others are localized in the ER ormitochondria (Ahsen and Pfanner, 1997; James et al., 1997). Inaddition, the eukaryotic cell contains a battery of other protein

36 R.Y.-L. Wang et al. / Virology 394 (2009) 28–38
chaperones, such as the HSP90 family and the J-domain HSP40 co-chaperones (Brodsky and Chiosis, 2006). However, the purifiedtombusvirus replicase contains only Ssa1p and Ssa2p Hsp70 proteinsat detectable level (Serva and Nagy, 2006), suggesting that tombus-viruses recruit only this group of cytosolic chaperones. Inactivation ofSSA1/2, however, only partially debilitate TBSV repRNA accumulation(Fig. 1A) (Serva and Nagy, 2006), and prior heat shock treatmentenhances TBSV replication, indicating that there is a heat shockinducible factor(s), which is capable of complementing the missingSsa1/2p function for TBSV replication in yeast (Fig. 1B). In this work,we demonstrate that complementation of SSA1/2 is due to the stress-inducible SSA3/4 genes, which are 80% homologous with SSA1/2(Ahsen and Pfanner, 1997). This conclusion is based on (i) thatrecombinant Ssa3p (Fig. 1C) can facilitate the assembly of the TBSVreplicase in vitro; and (ii) we did not observe complementation ofTBSV replication (Fig. 2A) when we used ssa1ts yeast strain lackingfunctional SSA2/3/4 and carrying ssa1ts, which was inactivated at anonpermissive temperature (Becker et al., 1996). Also, the replicasepreparation from ssa1ts yeast grown at the nonpermissive tempera-ture was inactive (Fig. 2C), arguing that the SSA1/2/3/4 subfamily ofHSP70 is essential for TBSV replication, albeit individual members inthe subfamily play redundant function. Thus, the numerous other heatshock proteins of yeast cannot complement the functions provided bySSA subfamily for TBSV replication.
What steps of replication are affected by Hsp70? The datapresented in this and previous papers (Pogany et al., 2008; Wanget al., 2009) indicate that Ssa1p plays a role in the early steps oftombusvirus replication, including subcellular localization of the viralreplication proteins, membrane insertion of the replication proteinsand the assembly of the viral replicase complex. Accordingly, in vitroassembly of the TBSV replicase based on a yeast cell-free extractrevealed that the constitutively expressed Ssa1p (Pogany et al., 2008)as well as the heat-induced Ssa3p (Fig. 1C) could facilitate theassembly event. Also, YFP-p33 is mislocalized to the cytosol in ssa1ts
yeast cultured at the nonpermissive temperature, instead of formingpunctate structures, which are characteristic for p33 associated withthe peroxisome-membrane observed at the permissive temperature(Fig. 3A). Thus, functional Ssa1p is required for correct subcellularlocalization of the viral replication proteins. Interestingly, based ontime course experiments, in which Ssa1pts was re-activated byshifting down to permissive temperature, we found correlationamong accumulation of p33 in the membrane fraction (Fig. 7D), theformation of punctate structures by YFP-p33 from the diffusedcytosolic stage (Fig. 7A), and the in vitro activity of the tombusvirusreplicase inmembrane preparations (Fig. 8A), suggesting that Ssa1p isinvolved in these events. Indeed, we have shown previously thatSsa1p facilitates themembrane-insertion of p33 in vitro (Wang, Stork,and Nagy, 2009). We have also shown that the targeted membranesinclude the peroxisome and ER membranes (Jonczyk et al., 2007;Pathak et al., 2008). Apparently, the membrane insertion step for p33and p92pol is essential for the formation of active replicase complex,since the soluble, cytosolic tombusvirus replicase proteins cannotform functional replicase (Fig. 8B) (McCartney et al., 2005; Panavaset al., 2005a; Wang et al., 2009) and no accumulation of TBSV repRNAis detected in yeast cells showing cytosolic distribution of YFP-p33(Figs. 2A and 6B).
In contrast with the multiple roles for Ssa1p in the early events oftombusvirus replication, we did not find evidence on the role of Ssa1pduring viral RNA synthesis, including minus- or plus-strand synthesis.For example, the isolated replicase complex obtained from ssa1ts
yeast grown at the permissive temperature was as active in viral RNAsynthesis at the nonpermissive 37 °C as at 25 °C in vitro (Fig. 4).Moreover, an in vitro TBSV replicase assembly assay performed withSsa1pts or Ssa1pwt at the permissive 20 °C temperature, followed byRNA synthesis at a partially nonpermissive temperature for Ssa1pts
showed comparable level of activity for the replicase assembled with
Ssa1pts and Ssa1pwt (Fig. 5B), arguing strongly against the role ofSsa1p during viral RNA synthesis in vitro.
To gain insight into the role of Ssa1p in p33 localization and theassembly of the tombusvirus replicase in yeast, we used temperatureshift experiments from nonpermissive to permissive in the absence orpresence of new p33/p92pol production. It took only 30 min to detectthe activity of the tombusvirus replicase after the downshift (Fig. 8A–C), which is similar to the time requirement for tombusvirus replicaseassembly in vitro (Pogany and Nagy, 2008; Pogany et al., 2008).However, the activity of the tombusvirus replicase increased ∼10-foldwhenmeasured 3–4 h after the downshift, suggesting that most of thereplicase assembly or localization of the replication proteins takeplace slowly. We speculate that this slow replicase assembly and/orsubcellular localization processes are due to the limiting amount ofavailable re-activated Ssa1pts since misfolded/mislocalized hostproteins might also compete with the viral replication proteins forthe service of Ssa1pts after the downshift.
This work provides evidence that the heat-inducible SSA3/4 canpartially complement the functions of the constitutively expressedSSA1/2. This is based on the finding that the stimulating effect ofheat shock treatment on TBSV replication in ssa1ssa2 mutant yeast(Fig. 1) was not observed when we used Ssa1pts yeast lackingfunctional SSA2/3/4, suggesting that the heat-inducible host factor islikely Ssa3p and/or Ssa4p that can efficiently complement themissingSsa1/2p functions for TBSV replication in ssa1ssa2 mutant yeast. Inaddition, we also observed that the recombinant Ssa3p was capable ofassembling the active TBSV replicase using an in vitro replicationassay (Fig. 1C).
The selective recruitment of cytosolic Hsp70 for viral TBSVreplication might be more general phenomena involving numerousviruses. Co-opting cellular chaperones could be a very successfulapproach for viruses, since these proteins are abundant and conserved,thus facilitating their recruitment for virus infections. In addition, thecellular chaperones play significant roles in innate resistanceresponses by the host and their sequestration by various virusescould inhibit antiviral responses from the host cell (Mayer, 2005).
Altogether, this work firmly establishes an essential role for SSA1/2/3/4 subfamily of Hsp70 in TBSV replication. The two constitutivelyexpressedmembers, Ssa1p and Ssa2p, play themajor role during TBSVreplication, whereas the heat-inducible Ssa3p and Ssa4p mightcontribute to TBSV accumulation only to a small extent at lowtemperature andmore so at high temperatures.We show that Ssa1p isessential for TBSV accumulation in the absence of the other SSAs. Thefunction of HSP70 is in the intracellular distribution and membraneinsertion of the viral replication proteins, as well as the assembly ofthe viral replicase. Previous work has also shown the importance ofthe cytosolic Hsp70 in TBSV genomic RNA replication in plant cellsand whole plant hosts based on knockdown and inhibition experi-ments (Wang et al., 2009). The obtained data are consistent withHsp70 playing multiple functions during the replication of tombus-virus RNA.
Materials and methods
Yeast strains and expression plasmids
The temperature sensitive ssa1 mutant (ssa1ts ssa2 ssa3 ssa4)strain DS10 (ssa 1–45BKD, ssa2::LEU2 ssa3::TRP1 ssa4::LYS2), the wtSSA1 strain (SSA1wt ssa2 ssa3 ssa4) and the double mutant (ssa1ssa2)strain MW123 (his3 leu2 lys2 trp1 ura3 ssa1::HIS3 ssa2::LEU2) werekindly provided by Elizabeth A. Craig (University of Wisconsin)(Becker et al., 1996). We used pESC-YFP-p33-DI-RNA dual construct(Jonczyk et al., 2007) to express YFP-p33 and DI-72 (+)RNA fromGAL1 and GAL10 promoters, respectively.
To express GFP-SKL carrying the three-amino acid SKL peroxi-somal targeting signal (Kragler et al., 1993), we PCR amplified the GFP

37R.Y.-L. Wang et al. / Virology 394 (2009) 28–38
sequence in combination with the C-terminally fused SKL sequencefrom pYES-GFP (Panavas et al., 2005a) as the template usingthe combination of primers #1292 (5′-CGGCAAGCTTACCATGG-TGAGCAAGGGCGAGGAGCTGT), and #2410 (5′-CGCGGGATCCTTA-TAATTTAGACTTGTACAGCTCGTCCATGC) as well as #1292 (5′-CGGCAAGCTTACCATGGTGAGCAAGGGCGAGGAGCTGT), and #2411(5′-CGCGGGATCCTTACAACTTGGACTTGTACAGCTCGTCCATGC). Thetwo separate PCR products for GFP-SKL were treated with HindIIIand BamHI, followed by their separate ligations into pYES-NT/C. Theobtained two GFP-SKL constructs behaved the same way in ourexperiments (not shown).
Subcellular fractionation
Yeast cells were grown to an optical density (OD600) between 0.8and 1.0. One hundred milligrams of cells was broken by using a cell-homogenizer (Fast prep) in 600 μl of yeast lysis buffer (200 mMsorbitol, 50 mM Tris–HCl, pH 7.5, 15 mM MgCl2, 10 mM KCl, 10 mMβ-mercaptoethanol, yeast protease inhibitormix; Sigma) (Wang et al.,2009). The obtained extracts were centrifuged in amicrocentrifuge for5min at 100×g to pellet cell debris. The supernatantwas subsequentlycentrifuged at 21,000×g to separate cytosolic and membrane-associated proteins into supernatant and pellet fractions, respectively(Wang et al., 2009). Pellets were re-suspended in the lysis buffer andaliquots corresponding to equal OD600 units of the original cell culturewere analyzed by standard sodiumdodecyl sulfate-polyacrylamide gelelectrophoresis (SDS-PAGE) and Western blotting procedures asdescribed previously (Jonczyk et al., 2007; Panaviene et al., 2004).
RNA analysis
Total RNA isolation and Northern blot analysis were performed asdescribed previously (Panavas andNagy, 2003; Panaviene et al., 2004).Briefly, for extraction of total RNA, yeast cells were broken by a cell-homogenizer (Genogrinder) for ∼1 min at room temperature withequal volumes of RNA extraction buffer (50 mM Na-acetate, pH 5.2,10mMEDTA, 1% SDS) andwater-saturated phenol and then incubatedfor 4min at 65 °C, followed by ethanol precipitation. The obtained RNAsamples were separated on a 1.5% agarose gel and transferred toHybond-XL membrane (Amersham) before hybridization with DI-72RNA-specific probe. For detection of plus-strand repRNA, we prepared32P-labeled RIII/IV(minus) probe generated via T7 transcription fromPCR-made template obtained with primers #1165 (AGCGAGTAAGA-CAGACTCTTCA) and #22 (GTAATACGACTCACTATAGGGCTG-CATTTCTGCAATGTTCC) on DI-72 templates (White andMorris, 1994).
Protein analysis
For protein analysis, we used an aliquot of samples used for RNAanalysis. A total of 50 ml yeast culture was harvested, the pelletedcells were re-suspended in 150 μl cold extraction buffer (200 mMsorbitol, 50 mM Tris–HCl, pH 7.5, 15mMMgCl2, 10 mMKCl, 10mM β-mercaptoethanol, yeast protease inhibitor mix; Sigma), and 250 μl ofglass beads was added to each sample. The cells were broken with agenogrinder for 2 min at 1500 rpm. Each sample was further mixedwith 600 μl pre-chilled extraction buffer, and unbroken cells wereremoved by centrifugation at 100×g for 5 min. The supernatant wasmixed with 1/2 volume of 3× SDS-PAGE sample buffer followed bySDS-PAGE and Western blot analysis as described previously(Panaviene et al., 2004).
Tombusvirus replicase assays
The “membrane-enriched” replicase preparations, which aresuitable to test the replicase activity on the endogenous templatespresent within the replicase preparation, were obtained as previously
described (Panaviene et al., 2004, 2005). Briefly, frozen yeast cellswere homogenized with Genogrinder for 2 min at 1500 rpm in 150 μlcold extraction buffer (200 mM sorbitol, 50 mM Tris–HCl, pH 7.5,15 mMMgCl2, 10 mM KCl, 10 mM β-mercaptoethanol, yeast proteaseinhibitor mix; Sigma) plus 250 μl of glass beads. Each sample wasfurther mixed with 600 μl pre-chilled extraction buffer, and unbrokencells were removed by centrifugation at 100×g for 5 min at 4 °C. Thesupernatant was centrifuged for 10 min at 21,000×g at 4 °C, and thenthe pellet was re-suspended and used in a standard tombusvirusreplicase assay. Because no template was added to the in vitroreaction, the replicase preparation could only use the endogenoustemplate present within the enriched membrane fraction. Thereplicase products were phenol-chloroform extracted, precipitatedwith isopropanol-ammonium acetate, and analyzed under denaturingconditions (5% PAGE containing 8 M urea).
Replication assay using the membrane fraction of the yeast cell-freeextract
The replication assay containing 0.5 μg purified recombinant TBSVMBP-p33 and/or TBSV MBP-p92 was done according to Pogany et al.(2008). The membrane fraction of the cell-free extract was obtainedby centrifugation at 4 °C for 10 min to separate the “soluble”(supernatant) and “membrane” (pellet) fraction (21,000×g). Toremove possible contaminating soluble proteins, the pellet waswashed with buffer A (30 mMHEPES-KOH pH 7.4, 100mM potassiumacetate, and 2 mM magnesium acetate) followed by centrifugation at4 °C for 10 min and re-suspension of the pellet in buffer A.
Purification of recombinant tombusvirus replicase proteins from E. coli
TBSV replicase proteins were expressed and purified according toRajendran and Nagy (2003), except using HEPES-KOH buffer (50 mM,pH 7.4) during purification and the elution buffer contained 1 mMDTT instead of the 10 mM β-mercaptoethanol.
Purification of Ssa3p as well as wt and mutant Ssa1p from yeast
The copper-inducible CUP1 promoter was used to express the wtFLAG-tagged Ssa3p, wt FLAG-Ssa1p and the temperature sensitive (ts)FLAG-Ssa1p (Becker et al., 1996) (kindly provided by Elizabeth Craig[University of Wisconsin]) from plasmid pEsc-His/Cup-FLAG/ssa1 wtand pEsc-His/Cup-FLAG/ssa1ts. Ssa1p was affinity purified as de-scribed previously (Pogany et al., 2008).
Confocal laser microscopy
To view yeast cells expressing different fluorescent fusion proteins,yeast strains were transformed with pESC-YFP-p33-DI-RNA, incombination with pYES-p92 or GFP-SKL. The confocal microscopywas performed on an Olympus FV1000 (Olympus America Inc.,Melville, New York) as described (Jonczyk et al., 2007; Wang andNagy, 2008; Wang et al., 2009).
Acknowledgments
We thank Drs. Daniel Barajas and Zhenghe Li for critical reading ofthe manuscript and for very helpful suggestions. This work wassupported by NIH-NIAID (5R21AI072170-02) and by the KentuckyTobacco Research and Development Center at the University ofKentucky, awarded to PDN.
References
Ahlquist, P., Noueiry, A.O., Lee, W.M., Kushner, D.B., Dye, B.T., 2003. Host factors inpositive-strand RNA virus genome replication. J. Virol. 77 (15), 8181–8186.

38 R.Y.-L. Wang et al. / Virology 394 (2009) 28–38
Alzhanova, D.V., Napuli, A.J., Creamer, R., Dolja, V.V., 2001. Cell-to-cell movement andassembly of a plant closterovirus: roles for the capsid proteins and Hsp70 homolog.EMBO J. 20 (24), 6997–7007.
Aparicio, F., Thomas, C.L., Lederer, C., Niu, Y., Wang, D., Maule, A.J., 2005. Virus inductionof heat shock protein 70 reflects a general response to protein accumulation in theplant cytosol. Plant Physiol. 138 (1), 529–536.
Aranda, M.A., Escaler, M., Wang, D., Maule, A.J., 1996. Induction of HSP70 andpolyubiquitin expression associated with plant virus replication. Proc. Natl. Acad.Sci. U. S. A. 93 (26), 15289–15293.
Becker, J., Walter, W., Yan, W., Craig, E.A., 1996. Functional interaction of cytosolichsp70 and a DnaJ-related protein, Ydj1p, in protein translocation in vivo. Mol. Cell.Biol. 16 (8), 4378–4386.
Boorstein, W.R., Craig, E.A., 1990. Transcriptional regulation of SSA3, an HSP70 genefrom Saccharomyces cerevisiae. Mol. Cell. Biol. 10 (6), 3262–3267.
Brodsky, J.L., Chiosis, G., 2006. Hsp70 molecular chaperones: emerging roles in humandisease and identification of small molecule modulators. Curr. Top. Med. Chem. 6(11), 1215–1225.
Brown, G., Rixon, H.W., Steel, J., McDonald, T.P., Pitt, A.R., Graham, S., Sugrue, R.J., 2005.Evidence for an association between heat shock protein 70 and the respiratorysyncytial virus polymerase complex within lipid-raft membranes during virusinfection. Virology 338 (1), 69–80.
Cherry, S., Doukas, T., Armknecht, S., Whelan, S., Wang, H., Sarnow, P., Perrimon, N.,2005. Genome-wide RNAi screen reveals a specific sensitivity of IRES-containingRNA viruses to host translation inhibition. Genes Dev. 19 (4), 445–452.
Dufresne, P.J., Thivierge, K., Cotton, S., Beauchemin, C., Ide, C., Ubalijoro, E., Laliberte, J.F.,Fortin, M.G., 2008. Heat shock 70 protein interaction with Turnipmosaic virus RNA-dependent RNA polymerase within virus-inducedmembrane vesicles. Virology 374(1), 217–227.
Hu, J., Flores, D., Toft, D., Wang, X., Nguyen, D., 2004. Requirement of heat shock protein90 for human hepatitis B virus reverse transcriptase function. J. Virol. 78 (23),13122–13131.
James, P., Pfund, C., Craig, E.A., 1997. Functional specificity among Hsp70 molecularchaperones. Science 275 (5298), 387–389.
Jiang, Y., Serviene, E., Gal, J., Panavas, T., Nagy, P.D., 2006. Identification of essential hostfactors affecting tombusvirus RNA replication based on the yeast Tet promotersHughes Collection. J. Virol. 80 (15), 7394–7404.
Jonczyk, M., Pathak, K.B., Sharma, M., Nagy, P.D., 2007. Exploiting alternative subcellularlocation for replication: tombusvirus replication switches to the endoplasmicreticulum in the absence of peroxisomes. Virology 362 (2), 320–330.
Kampmueller, K.M., Miller, D.J., 2005. The cellular chaperone heat shock protein 90facilitates Flock House virus RNA replication in Drosophila cells. J. Virol. 79 (11),6827–6837.
Kragler, F., Langeder, A., Raupachova, J., Binder, M., Hartig, A., 1993. Two independentperoxisomal targeting signals in catalase A of Saccharomyces cerevisiae. J. Cell. Biol.120 (3), 665–673.
Krishnan, M.N., Ng, A., Sukumaran, B., Gilfoy, F.D., Uchil, P.D., Sultana, H., Brass, A.L.,Adametz, R., Tsui, M., Qian, F., Montgomery, R.R., Lev, S., Mason, P.W., Koski, R.A.,Elledge, S.J., Xavier, R.J., Agaisse, H., Fikrig, E., 2008. RNA interference screen forhuman genes associated with West Nile virus infection. Nature 455 (7210),242–245.
Kushner, D.B., Lindenbach, B.D., Grdzelishvili, V.Z., Noueiry, A.O., Paul, S.M., Ahlquist, P.,2003. Systematic, genome-wide identification of host genes affecting replication ofa positive-strand RNA virus. Proc. Natl. Acad. Sci. U. S. A. 100 (26), 15764–15769.
Lin, B.L., Wang, J.S., Liu, H.C., Chen, R.W., Meyer, Y., Barakat, A., Delseny, M., 2001.Genomic analysis of the Hsp70 superfamily in Arabidopsis thaliana. Cell. StressChaperones 6 (3), 201–208.
Li, Z., Barajas, D., Panavas, T., Herbst, D.A., Nagy, P.D., 2008. Cdc34p ubiquitin-conjugating enzyme is a component of the tombusvirus replicase complex andubiquitinates p33 replication protein. J. Virol. 82 (14), 6911–6926.
Li, Z., Pogany, J., Panavas, T., Xu, K., Esposito, A.M., Kinzy, T.G., Nagy, P.D., 2009.Translation elongation factor 1A is a component of the tombusvirus replicasecomplex and affects the stability of the p33 replication co-factor. Virology 385,245–260.
Mayer, M.P., 2005. Recruitment of Hsp70 chaperones: a crucial part of viral survivalstrategies. Rev. Physiol. Biochem. Pharmacol. 153, 1–46.
McCartney, A.W., Greenwood, J.S., Fabian, M.R., White, K.A., Mullen, R.T., 2005. Locali-zation of the Tomato bushy stunt virus replicationprotein p33 reveals a peroxisome-to-endoplasmic reticulum sorting pathway. Plant Cell. 17 (12), 3513–3531.
Momose, F., Naito, T., Yano, K., Sugimoto, S., Morikawa, Y., Nagata, K., 2002.Identification of Hsp90 as a stimulatory host factor involved in influenza virusRNA synthesis. J. Biol. Chem. 277 (47), 45306–45314.
Monkewich, S., Lin, H.X., Fabian, M.R., Xu, W., Na, H., Ray, D., Chernysheva, O.A., Nagy,P.D., White, K.A., 2005. The p92 polymerase coding region contains an internal RNAelement required at an early step in Tombusvirus genome replication. J. Virol. 79(8), 4848–4858.
Nagy, P.D., 2008. Yeast as a model host to explore plant virus–host interactions. Annu.Rev. Phytopathol. 46, 217–242.
Nagy, P.D., Pogany, J., 2000. Partial purification and characterization of Cucumbernecrosis virus and Tomato bushy stunt virus RNA-dependent RNA polymerases:similarities and differences in template usage between tombusvirus and carmo-virus RNA-dependent RNA polymerases. Virology 276 (2), 279–288.
Nagy, P.D., Pogany, J., 2006. Yeast as a model host to dissect functions of viral and hostfactors in tombusvirus replication. Virology 344 (1), 211–220.
Nishikiori, M., Dohi, K., Mori, M., Meshi, T., Naito, S., Ishikawa, M., 2006. Membrane-bound tomato mosaic virus replication proteins participate in RNA synthesis andare associated with host proteins in a pattern distinct from those that are notmembrane bound. J. Virol. 80 (17), 8459–8468.
Noueiry, A.O., Ahlquist, P., 2003. Bromemosaic virus RNA replication: revealing the roleof the host in RNA virus replication. Annu. Rev. Phytopathol. 41, 77–98.
Panavas, T., Nagy, P.D., 2003. Yeast as a model host to study replication andrecombination of defective interfering RNA of Tomato bushy stunt virus. Virology314 (1), 315–325.
Panavas, T., Hawkins, C.M., Panaviene, Z., Nagy, P.D., 2005a. The role of the p33:p33/p92interaction domain in RNA replication and intracellular localization of p33 and p92proteins of Cucumber necrosis tombusvirus. Virology 338 (1), 81–95.
Panavas, T., Serviene, E., Brasher, J., Nagy, P.D., 2005b. Yeast genome-wide screenreveals dissimilar sets of host genes affecting replication of RNA viruses. Proc. Natl.Acad. Sci. U. S. A. 102 (20), 7326–7331.
Panaviene, Z., Panavas, T., Serva, S., Nagy, P.D., 2004. Purification of the Cucumbernecrosis virus replicase from yeast cells: role of coexpressed viral RNA instimulation of replicase activity. J. Virol. 78 (15), 8254–8263.
Panaviene, Z., Panavas, T., Nagy, P.D., 2005. Role of an internal and two 3′-terminal RNAelements in assembly of tombusvirus replicase. J. Virol. 79 (16), 10608–10618.
Pathak, K.B., Sasvari, Z., Nagy, P.D., 2008. The host Pex19p plays a role in peroxisomallocalization of tombusvirus replication proteins. Virology 379 (2), 294–305.
Peremyslov, V.V., Hagiwara, Y., Dolja, V.V., 1999. HSP70 homolog functions in cell-to-cell movement of a plant virus. Proc. Natl. Acad. Sci. U. S. A. 96 (26), 14771–14776.
Pogany, J., Nagy, P.D., 2008. Authentic replication and recombination of Tomato bushystunt virus RNA in a cell-free extract from yeast. J. Virol. 82 (12), 5967–5980.
Pogany, J., White, K.A., Nagy, P.D., 2005. Specific binding of tombusvirus replicationprotein p33 to an internal replication element in the viral RNA is essential forreplication. J. Virol. 79 (8), 4859–4869.
Pogany, J., Stork, J., Li, Z., Nagy, P.D., 2008. In vitro assembly of the Tomato bushy stuntvirus replicase requires the host Heat shock protein 70. Proc. Natl. Acad. Sci. U. S. A.105 (50), 19956–19961.
Qanungo, K.R., Shaji, D., Mathur, M., Banerjee, A.K., 2004. Two RNA polymerasecomplexes from vesicular stomatitis virus-infected cells that carry out transcriptionand replication of genome RNA. Proc. Natl. Acad. Sci. U. S. A. 101 (16), 5952–5957.
Rajendran, K.S., Nagy, P.D., 2003. Characterization of the RNA-binding domains in thereplicase proteins of Tomato bushy stunt virus. J. Virol. 77 (17), 9244–9258.
Randall, G., Panis, M., Cooper, J.D., Tellinghuisen, T.L., Sukhodolets, K.E., Pfeffer, S.,Landthaler, M., Landgraf, P., Kan, S., Lindenbach, B.D., Chien, M., Weir, D.B., Russo,J.J., Ju, J., Brownstein, M.J., Sheridan, R., Sander, C., Zavolan, M., Tuschl, T., Rice, C.M.,2007. Cellular cofactors affecting hepatitis C virus infection and replication. Proc.Natl. Acad. Sci. U. S. A. 104 (31), 12884–12889.
Salonen, A., Ahola, T., Kaariainen, L., 2005. Viral RNA replication in association withcellular membranes. Curr. Top. Microbiol. Immunol. 285, 139–173.
Serva, S., Nagy, P.D., 2006. Proteomics analysis of the tombusvirus replicase: Hsp70molecular chaperone is associated with the replicase and enhances viral RNAreplication. J. Virol. 80 (5), 2162–2169.
Serviene, E., Shapka, N., Cheng, C.P., Panavas, T., Phuangrat, B., Baker, J., Nagy, P.D., 2005.Genome-wide screen identifies host genes affecting viral RNA recombination. Proc.Natl. Acad. Sci. U. S. A. 102 (30), 10545–10550.
Shi, S.T., Lai, M.M., 2005. Viral and cellular proteins involved in coronavirus replication.Curr. Top. Microbiol. Immunol. 287, 95–131.
Stahl, M., Retzlaff, M., Nassal, M., Beck, J., 2007. Chaperone activation of thehepadnaviral reverse transcriptase for template RNA binding is established bythe Hsp70 and stimulated by the Hsp90 system. Nucleic Acids Res. 35 (18),6124–6136.
Tavis, J.E., Massey, B., Gong, Y., 1998. The duck hepatitis B virus polymerase is activatedby its RNA packaging signal, epsilon. J. Virol. 72 (7), 5789–5796.
Tomita, Y., Mizuno, T., Diez, J., Naito, S., Ahlquist, P., Ishikawa, M., 2003. Mutation of hostDnaJ homolog inhibits brome mosaic virus negative-strand RNA synthesis. J. Virol.77 (5), 2990–2997.
Von Assen, O., Pfanner, N., 1997. Molecular chaperones: towards a characterization ofthe heat-shock protein 70 family. Trends Cell. Biol. 7 (3), 129–133.
Wang, R.Y., Nagy, P.D., 2008. Tomato bushy stunt virus co-opts the RNA-bindingfunction of a host metabolic enzyme for viral genomic RNA synthesis. Cell HostMicrobe. 3 (3), 178–187.
Wang, R.Y., Stork, J., Nagy, P.D., 2009. A key role for heat shock protein 70 in localizationand insertion of the tombusvirus replication proteins to intracellular membranes.J. Virol. 83 (7), 3276–3287.
Weeks, S.A., Miller, D.J., 2008. The heat shock protein 70 cochaperone YDJ1 is requiredfor efficient membrane-specific flock house virus RNA replication complexassembly and function in Saccharomyces cerevisiae. J. Virol. 82 (4), 2004–2012.
White, K.A., Morris, T.J., 1994. Enhanced competitiveness of Tomato bushy stunt virusdefective interfering RNAs by segment duplication or nucleotide insertion. J. Virol.68 (9), 6092–6096.
White, K.A., Nagy, P.D., 2004. Advances in the molecular biology of tombusviruses: geneexpression, genome replication, and recombination. Prog. Nucleic Acid Res. Mol.Biol. 78, 187–226.
Whitham, S.A., Quan, S., Chang, H.S., Cooper, B., Estes, B., Zhu, T., Wang, X., Hou, Y.M.,2003. Diverse RNA viruses elicit the expression of common sets of genes insusceptible Arabidopsis thaliana plants. Plant J. 33 (2), 271–283.
Whitham, S.A., Yang, C., Goodin, M.M., 2006. Global impact: elucidating plant responsesto viral infection. Mol. Plant Microbe Interact. 19 (11), 1207–1215.