a unifying framework for metapopulation dynamics - gu · pdf filevol. 160, no. 2 the american...
TRANSCRIPT

vol. 160, no. 2 the american naturalist august 2002
A Unifying Framework for Metapopulation Dynamics
Karin C. Harding1,* and John M. McNamara2,†
1. Department of Marine Ecology, Goteborg University, Box 461,405 30 Goteborg, Sweden;2. School of Mathematics, University of Bristol, University Walk,Bristol BS8 1TW, United Kingdom
Submitted March 12, 2001; Accepted January 18, 2002
abstract: Many biologically important processes, such as geneticdifferentiation, the spread of disease, and population stability, areaffected by the (natural or enforced) subdivision of populations intonetworks of smaller, partly isolated, subunits. Such “metapopula-tions” can have extremely complex dynamics. We present a newgeneral model that uses only two functions to capture, at the meta-population scale, the main behavior of metapopulations. We showhow complex, structured metapopulation models can be translatedinto our generalized framework. The metapopulation dynamics aris-ing from some important biological processes are illustrated: therescue effect, the Allee effect, and what we term the “antirescueeffect.” The antirescue effect captures instances where high migrationrates are deleterious to population persistence, a phenomenon thathas been largely ignored in metapopulation conservation theory.Management regimes that ignore a significant antirescue effect willbe inadequate and may actually increase extinction risk. Further,consequences of territoriality and conspecific attraction on meta-population-level dynamics are investigated. The new, simplifiedframework can incorporate knowledge from epidemiology, genetics,and population biology in a phenomenological way. It opens up newpossibilities to identify and analyze the factors that are importantfor the evolution and persistence of the many spatially subdividedspecies.
Keywords: metapopulation, rescue effect, multiple equilibria, Alleeeffect, antirescue effect.
The spatial structure of a population can profoundly affectgenetic, epidemiological, and population dynamic pro-cesses within that population. For example, subdivision ofa population can influence gene flow and enhance the rateof loss of genetic diversity in subpopulations (Lacy 1987;
* Corresponding author; e-mail: [email protected].
† E-mail: [email protected].
Am. Nat. 2002. Vol. 160, pp. 173–185. � 2002 by The University of Chicago.0003-0147/2002/16002-0003$15.00. All rights reserved.
Pannell and Charlesworth 1999), which sometimes resultsin harmful levels of inbreeding (Sacceri et al. 1998). How-ever, spatial subdivision is also a requirement for beneficiallocal adaptations to occur (Rolan-Alvarez et al. 1997; Joshiet al. 2001). Within epidemiology, the rate of contact be-tween subpopulations crucially influences the develop-ment of a disease outbreak (Grenfell and Harwood 1997).Population biologists have found a wide variety of caseswhere spatial structure influences the stability of single-and multispecies systems, both in nature and in theoreticalsettings (Tilman and Kareiva 1997; Hanski 1999). All theprocesses exemplified above also have the potential to af-fect the viability or extinction risk of single subpopula-tions. An appreciation of spatial influences on biologicalprocesses has proven important for the understanding ofbasic evolutionary and ecological phenomena. This ap-preciation is also urgently needed if we are to improve themanagement of the many endangered species living infragmented habitats.
However, the construction and analysis of mathematicalmodels of spatially structured populations has turned outto be a formidably complicated task. The processes withina subpopulation can be as complex as in an ordinary singlepopulation and are, in addition, intricately affected by thedynamics of neighboring patches via the migration of in-dividuals. Impressive efforts have been made to modelsuch “metapopulations,” taking into account the size,growth rate, and age structure of each subpopulation (Lev-ins 1969; Hastings and Volin 1989; Allen et al. 1993), patchquality (Hanski 1994), matrix quality (Vandermeer andCarvajal 2001), correlated random events (Harrison andQuinn 1989), and interactions with predators (Taylor1990; Holt 1997). A few models have even attempted topredict the dynamics of real-world metapopulations (Han-ski 1994). These detailed, so-called structured models arenecessary to describe specific populations or specific pro-cesses. Unfortunately, as the models describe specific sit-uations and populations with increasing accuracy, theyinevitably grow in complexity and have been found to beunsuited for establishing general principles concerningmetapopulation dynamics: “This general framework, how-ever, leads to a class of models that are typically too com-plex to analyze” (Hastings and Harrison 1994, p. 175).

174 The American Naturalist
“Although limited mathematical analysis is possible, … itis hard to extract many useful ecological results and pre-dictions without making some further simplifying as-sumptions” (Hanski 1999, p. 64).
Simplified models are needed to address general ques-tions in biology that concern the whole metapopulationscale. However, the simple metapopulation models thathave been available, Levins’s (1969) model and variantsthereof, are too restricted (Gyllenberg et al. 1997). As wewill show in this study, this is because they do not allowfor a range of biological phenomena affecting the dynamicsof metapopulations at the metapopulation level. While agood general model does not have to include all detailsof the real world to give interesting insights, it should beable to express the dynamics of the system, at the level itattempts to capture (Caswell 1988). While it has beenargued that the shortcomings of the unstructured Levinsmodel stem from its many unrealistic assumptions aboutthe local dynamics of subpopulations (Burgman et al.1993; Hanski and Zhang 1993), we show that the limi-tations are instead at the description of the metapopula-tion-level processes themselves. We use the vital core ofLevins’s model to build a new phenomenological modelthat is unstructured but nonetheless captures much moreof the dynamic flexibility of metapopulations.
The new model is used to visualize and classify themetapopulation dynamics that can arise from differentmain types of extinction and colonization processes. Weshow how structured models can be incorporated into oursimplified framework, making it easier to understand theorigin of their behavior. Finally, we give an example onhow extinction times of small metapopulations are influ-enced by the assumed shape of the colonization and ex-tinction functions.
About Levins’s Model
Richard Levins introduced a new approach to the studyof patchy populations (Levins 1969). In his model he as-sumes that the environment is composed of T identicalhabitat patches. Patches are classified as either occupiedor empty; there is no additional local structure. Occupiedpatches send out emigrants at rate m, and these migrantscolonize unoccupied patches. Occupied patches go extinctwith the rate e. When the two processes of extinction andcolonization counterbalance there is an equilibrium num-ber of occupied patches. The characterization of equilibriais important since natural metapopulations are expected,and are sometimes also found, to move toward such “equi-libria of patch occupancy” (Hanski 1999).
However, in Levins’s model, it is assumed that the arrivalof a migrant at an empty patch always turns that patchinto an occupied patch. Thus, the colonization rate of each
patch in Levins’s model is preset; it increases proportion-ally with the number of patches occupied (N). Conse-quently, total colonization rate in the whole metapopu-lation (C) is given by . Further, Levins’sC p mN(1 � N/T)model assumes the extinction rate per patch to be a con-stant, independent of the number of surrounding occupiedpatches. Thus, the total extinction rate (E) in a Levins-type metapopulation is preset to just increase linearly asthe number of occupied patches increases: .E p eN
As we will demonstrate, these preset functions (C andE) leave out important biological processes that act at themetapopulation scale and are the reasons why Levins’smodel cannot act as a good phenomenological model formetapopulations. In order to capture the dynamics ofmetapopulations, we must allow for different types of de-pendencies between migration and colonization rates andbetween migration and extinction rates. This is becauseper patch colonization rate can differ systematically fromthe assumed function of Levins’s model; for example, itmight take several individuals to establish a new subpop-ulation so that the colonization rate can have a nonlineardependence on migration rate (see “Allee Effect” and “TheEffects of Differential Migration”). Also, the influx of mi-grants can significantly affect the subpopulation extinctionrate. This effect can be positive (e.g., by the demographiccontribution of immigrants to small populations) or neg-ative (e.g., by the introduction of a lethal disease); see“Rescue Effect” and “Antirescue Effect.” We develop aframework that can embrace most of the relevant meta-population-level processes (i.e., those that affect patchoccupancy).
A Generalized Metapopulation Model
Faithful to the classic metapopulation scenario of Levins(1969), we let the environment be composed of T discretepatches, where at any given time each patch is either oc-cupied or not. Each occupied patch sends out successfulmigrants at rate m. (By “successful” migrants, we meanthose that survive to reach another patch.) However, incontrast to traditional patch-occupancy models, we willnot assume specific shapes of the functions for the ratesat which patches are colonized (C) or go extinct (E). And,importantly, we shall allow for the possibility that the ex-tinction rate is influenced by the rate of immigration. Wedefine two general functions, the colonization rate perpatch Cpatch(a) and the extinction rate per patch Epatch(b),where a denotes the rate of arrival of colonizers onto anunoccupied patch and b denotes the rate at which im-migrants arrive at an occupied patch. We assume that thesearrival rates for immigrants are functions a(N) andb(N) of the number of occupied patches (N). The rate at

Unifying Metapopulation Model 175
which empty patches are colonized in a metapopulationaccording to this new generalized framework is given by
C (N) p (T � N)C [a(N)]. (1)total patch
The rate at which occupied patches switch to emptypatches Etotal(N) in the new framework is
E (N) p NE [b(N)]. (2)total patch
These two functions (eqq. [1], [2]) are the core of thenew flexible model. By allowing for various dependenciesbetween extinction and colonization on migration we nowcan account for most metapopulation-level dynamics. Tofind the rate at which the number of occupied patcheschanges with time in the metapopulation, we make thesame deterministic approximation to the stochastic changein N as is made in Levins’s model. That is, we assume thatthe number of patches is large enough to make stochasticdeviations in N insignificant. Then the rate of change inpatch occupancy is given by
dNp C (N) � E (N). (3)total totaldt
The immigration rates a(N) and b(N) could be differentand have a complex dependence on the number of oc-cupied patches, N. However, for ease of exposition, weassume that migrants settle at random so that a p b p
. This assumption will be relaxed later. SettingmN/Tin equation (1), we obtaina(N) p mN/T C (N) ptotal
, where denotes the proportionT(1 � x)C (mx) x p N/Tpatch
of occupied patches. Similarly, setting inb(N) p mN/Tequation (2), we obtain . SinceE (N) p TxE (mx)total patch
, equation (3) now reduces todN/dt p Tdx/dt dx/dt p, wheref(x)
f(x) p (1 � x)C (mx) � xE (mx). (4)patch patch
When extinction and colonization rates counterbalance,we have an equilibrium proportion of occupied patches( ). At equilibrium we have , so that satisfies∗ ∗x dx/dt p 0 x
. If , the equilibrium is stable, and the∗ ′ ∗f(x ) p 0 f (x ) ! 0metapopulation will deterministically return to the equi-librium after a small perturbation. If , the equi-′ ∗f (x ) 1 0librium is unstable and a small perturbation away fromthe equilibrium will either make the metapopulation movetoward another equilibrium of patch occupancy or causemetapopulation extinction. We will now look at the meta-population dynamics that arise from several different typesof biologically interesting colonization Cpatch and extinctionfunctions Epatch. However, we first look at the traditionalLevins model again. We will repeatedly return to this
model, but as a specific, basic case of the generalizedframework.
Levins’s Functions Rewritten
In Levins’s model the arrival of an immigrant at an un-occupied patch changes that patch into an occupied patch.This can be written as in our generalizedC (a) p Kapatch
model, where the constant K is the proportion of immi-grants arriving at an unoccupied patch that succeed inestablishing an occupied patch. The total colonization rateCtotal(N) in the metapopulation thus satisfies
C (N) N Ntotal p 1 � Km p (1 � x)Kmx. (5)( )T T T
Levins’s model assumes the patch-specific extinctionrate to be , where e is a constant, which givesE (b) p epatch
E (N) eNtotal p p xe. (6)T T
Equation (4) becomes
f(x) p (1 � x)Kmx � xe p x(Km � e � Kmx). (7)
Thus, the preset shapes of the functions limit Levins’smodel to the single equilibrium . Because∗x p 1 � e/Km
, the equilibrium is stable. The well-known dy-′ ∗f (x ) ! 0namics of Levins’s model is visualized in figure 1. Anequilibrium point occurs where the colonization rateequals the extinction rate (fig. 1A). It can be seen that forany given value of the migration rate (m) the two functionsintersect only once (fig. 1B). A higher migration rate al-ways implies a higher patch occupancy (N/T) at equilib-rium in Levins’s metapopulation scenario. Below we willillustrate what happens with the dynamics of metapopu-lations for a range of examples when migration has adifferent influence on subpopulation extinction andestablishment.
Rescue Effect
The demographic contribution of migrants to small dwin-dling subpopulations may reduce subpopulation extinc-tion risk—the “rescue effect” (Brown and Kodric-Brown1977). Logically, if all subpopulations are of the same sizeand have the same migration rate (as assumed in Levins’smodel), there will be no rescue effect, since subpopulationswill lose as many (or more) migrants as they gain. There-fore, the rescue effect has primarily been included in morecomplex models (Gyllenberg et al. 1997; but see Hanski
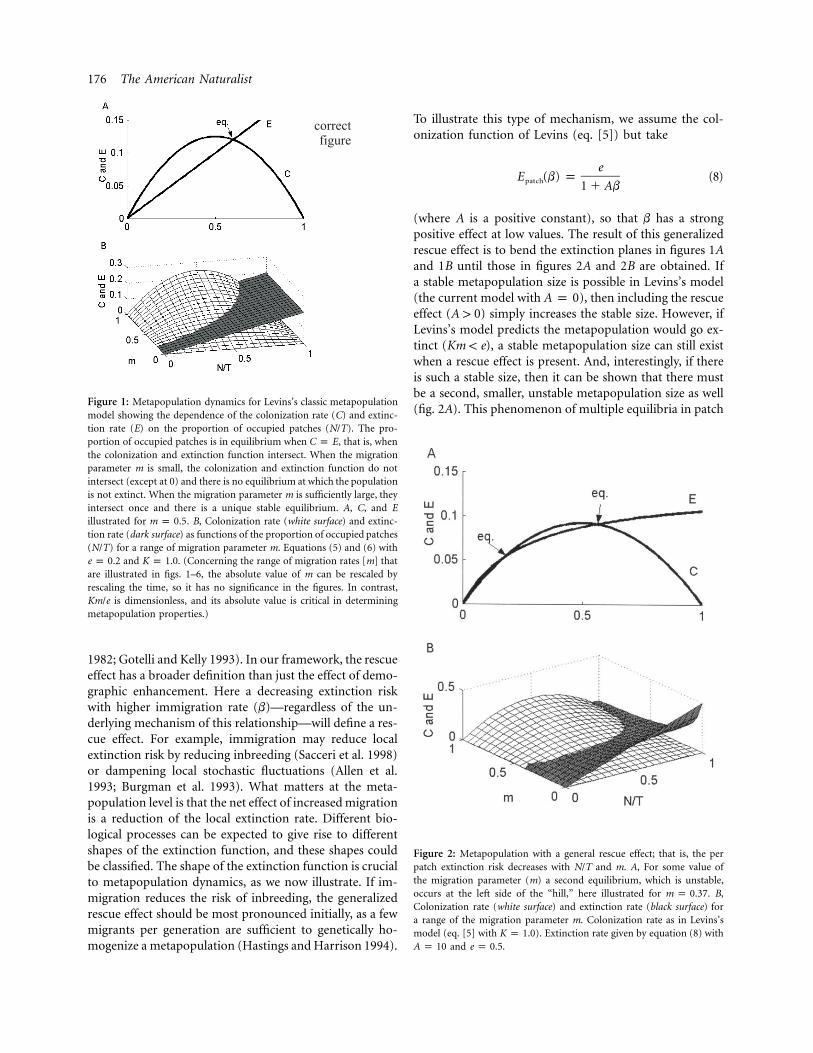
176 The American Naturalist
Figure 1: Metapopulation dynamics for Levins’s classic metapopulationmodel showing the dependence of the colonization rate (C) and extinc-tion rate (E) on the proportion of occupied patches (N/T). The pro-portion of occupied patches is in equilibrium when , that is, whenC p Ethe colonization and extinction function intersect. When the migrationparameter m is small, the colonization and extinction function do notintersect (except at 0) and there is no equilibrium at which the populationis not extinct. When the migration parameter m is sufficiently large, theyintersect once and there is a unique stable equilibrium. A, C, and Eillustrated for . B, Colonization rate (white surface) and extinc-m p 0.5tion rate (dark surface) as functions of the proportion of occupied patches(N/T) for a range of migration parameter m. Equations (5) and (6) with
and . (Concerning the range of migration rates [m] thate p 0.2 K p 1.0are illustrated in figs. 1–6, the absolute value of m can be rescaled byrescaling the time, so it has no significance in the figures. In contrast,
is dimensionless, and its absolute value is critical in determiningKm/emetapopulation properties.)
Figure 2: Metapopulation with a general rescue effect; that is, the perpatch extinction risk decreases with N/T and m. A, For some value ofthe migration parameter (m) a second equilibrium, which is unstable,occurs at the left side of the “hill,” here illustrated for . B,m p 0.37Colonization rate (white surface) and extinction rate (black surface) fora range of the migration parameter m. Colonization rate as in Levins’smodel (eq. [5] with ). Extinction rate given by equation (8) withK p 1.0
and .A p 10 e p 0.5
1982; Gotelli and Kelly 1993). In our framework, the rescueeffect has a broader definition than just the effect of demo-graphic enhancement. Here a decreasing extinction riskwith higher immigration rate (b)—regardless of the un-derlying mechanism of this relationship—will define a res-cue effect. For example, immigration may reduce localextinction risk by reducing inbreeding (Sacceri et al. 1998)or dampening local stochastic fluctuations (Allen et al.1993; Burgman et al. 1993). What matters at the meta-population level is that the net effect of increased migrationis a reduction of the local extinction rate. Different bio-logical processes can be expected to give rise to differentshapes of the extinction function, and these shapes couldbe classified. The shape of the extinction function is crucialto metapopulation dynamics, as we now illustrate. If im-migration reduces the risk of inbreeding, the generalizedrescue effect should be most pronounced initially, as a fewmigrants per generation are sufficient to genetically ho-mogenize a metapopulation (Hastings and Harrison 1994).
To illustrate this type of mechanism, we assume the col-onization function of Levins (eq. [5]) but take
eE (b) p (8)patch 1 � Ab
(where A is a positive constant), so that b has a strongpositive effect at low values. The result of this generalizedrescue effect is to bend the extinction planes in figures 1Aand 1B until those in figures 2A and 2B are obtained. Ifa stable metapopulation size is possible in Levins’s model(the current model with ), then including the rescueA p 0effect ( ) simply increases the stable size. However, ifA 1 0Levins’s model predicts the metapopulation would go ex-tinct ( ), a stable metapopulation size can still existKm ! ewhen a rescue effect is present. And, interestingly, if thereis such a stable size, then it can be shown that there mustbe a second, smaller, unstable metapopulation size as well(fig. 2A). This phenomenon of multiple equilibria in patch
correctfigure
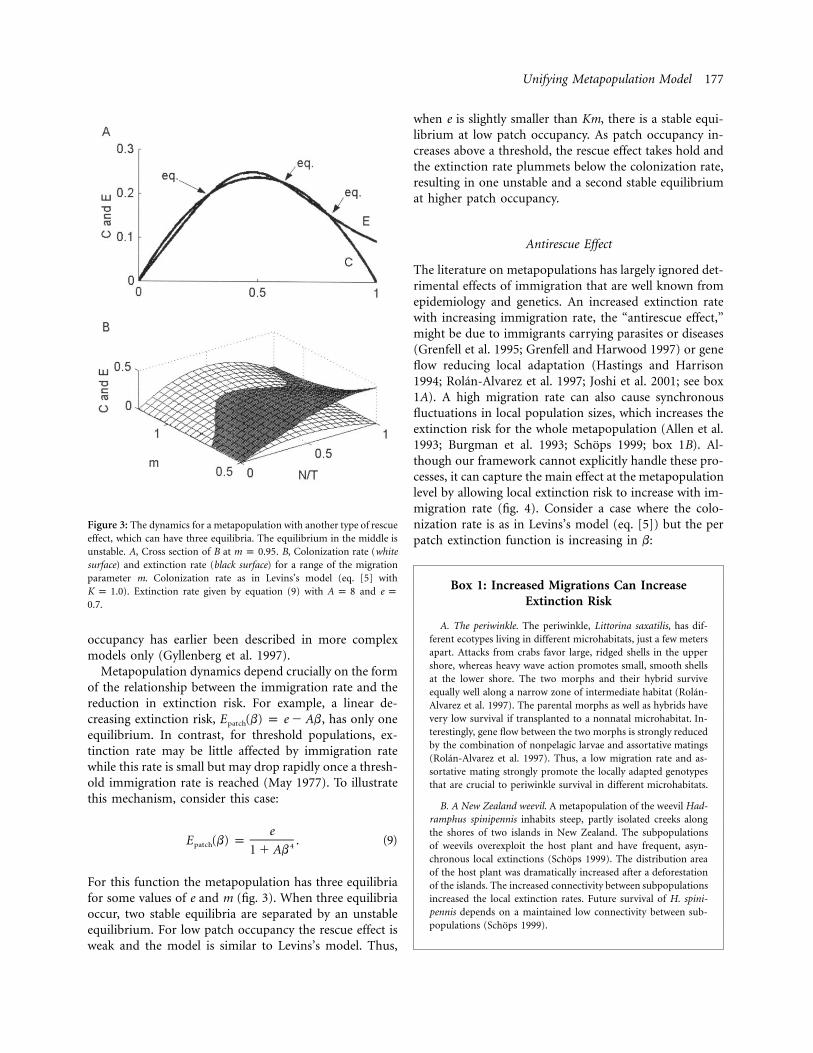
Unifying Metapopulation Model 177
Figure 3: The dynamics for a metapopulation with another type of rescueeffect, which can have three equilibria. The equilibrium in the middle isunstable. A, Cross section of B at . B, Colonization rate (whitem p 0.95surface) and extinction rate (black surface) for a range of the migrationparameter m. Colonization rate as in Levins’s model (eq. [5] with
). Extinction rate given by equation (9) with andK p 1.0 A p 8 e p.0.7
Box 1: Increased Migrations Can IncreaseExtinction Risk
A. The periwinkle. The periwinkle, Littorina saxatilis, has dif-ferent ecotypes living in different microhabitats, just a few metersapart. Attacks from crabs favor large, ridged shells in the uppershore, whereas heavy wave action promotes small, smooth shellsat the lower shore. The two morphs and their hybrid surviveequally well along a narrow zone of intermediate habitat (Rolan-Alvarez et al. 1997). The parental morphs as well as hybrids havevery low survival if transplanted to a nonnatal microhabitat. In-terestingly, gene flow between the two morphs is strongly reducedby the combination of nonpelagic larvae and assortative matings(Rolan-Alvarez et al. 1997). Thus, a low migration rate and as-sortative mating strongly promote the locally adapted genotypesthat are crucial to periwinkle survival in different microhabitats.
B. A New Zealand weevil. A metapopulation of the weevil Had-ramphus spinipennis inhabits steep, partly isolated creeks alongthe shores of two islands in New Zealand. The subpopulationsof weevils overexploit the host plant and have frequent, asyn-chronous local extinctions (Schops 1999). The distribution areaof the host plant was dramatically increased after a deforestationof the islands. The increased connectivity between subpopulationsincreased the local extinction rates. Future survival of H. spini-pennis depends on a maintained low connectivity between sub-populations (Schops 1999).
occupancy has earlier been described in more complexmodels only (Gyllenberg et al. 1997).
Metapopulation dynamics depend crucially on the formof the relationship between the immigration rate and thereduction in extinction risk. For example, a linear de-creasing extinction risk, , has only oneE (b) p e � Abpatch
equilibrium. In contrast, for threshold populations, ex-tinction rate may be little affected by immigration ratewhile this rate is small but may drop rapidly once a thresh-old immigration rate is reached (May 1977). To illustratethis mechanism, consider this case:
eE (b) p . (9)patch 41 � Ab
For this function the metapopulation has three equilibriafor some values of e and m (fig. 3). When three equilibriaoccur, two stable equilibria are separated by an unstableequilibrium. For low patch occupancy the rescue effect isweak and the model is similar to Levins’s model. Thus,
when e is slightly smaller than Km, there is a stable equi-librium at low patch occupancy. As patch occupancy in-creases above a threshold, the rescue effect takes hold andthe extinction rate plummets below the colonization rate,resulting in one unstable and a second stable equilibriumat higher patch occupancy.
Antirescue Effect
The literature on metapopulations has largely ignored det-rimental effects of immigration that are well known fromepidemiology and genetics. An increased extinction ratewith increasing immigration rate, the “antirescue effect,”might be due to immigrants carrying parasites or diseases(Grenfell et al. 1995; Grenfell and Harwood 1997) or geneflow reducing local adaptation (Hastings and Harrison1994; Rolan-Alvarez et al. 1997; Joshi et al. 2001; see box1A). A high migration rate can also cause synchronousfluctuations in local population sizes, which increases theextinction risk for the whole metapopulation (Allen et al.1993; Burgman et al. 1993; Schops 1999; box 1B). Al-though our framework cannot explicitly handle these pro-cesses, it can capture the main effect at the metapopulationlevel by allowing local extinction risk to increase with im-migration rate (fig. 4). Consider a case where the colo-nization rate is as in Levins’s model (eq. [5]) but the perpatch extinction function is increasing in b:
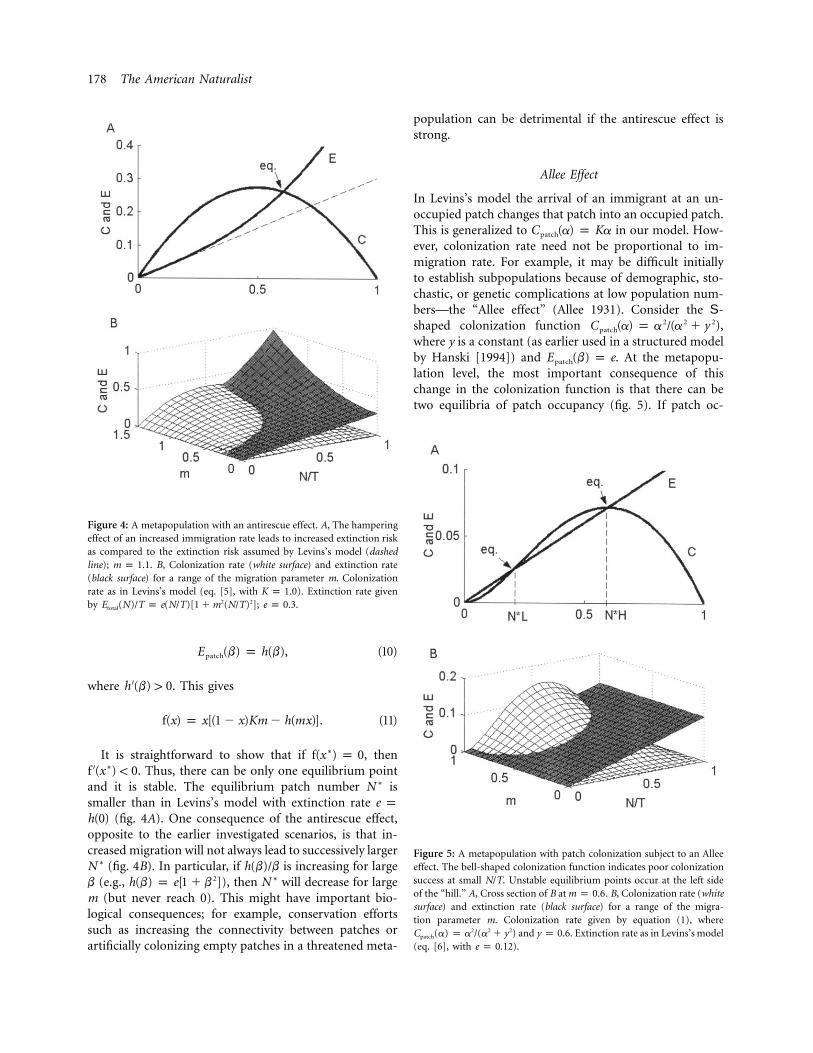
178 The American Naturalist
Figure 4: A metapopulation with an antirescue effect. A, The hamperingeffect of an increased immigration rate leads to increased extinction riskas compared to the extinction risk assumed by Levins’s model (dashedline); . B, Colonization rate (white surface) and extinction ratem p 1.1(black surface) for a range of the migration parameter m. Colonizationrate as in Levins’s model (eq. [5], with ). Extinction rate givenK p 1.0by ; .2 2E (N)/T p e(N/T)[1 � m (N/T) ] e p 0.3total
Figure 5: A metapopulation with patch colonization subject to an Alleeeffect. The bell-shaped colonization function indicates poor colonizationsuccess at small N/T. Unstable equilibrium points occur at the left sideof the “hill.” A, Cross section of B at . B, Colonization rate (whitem p 0.6surface) and extinction rate (black surface) for a range of the migra-tion parameter m. Colonization rate given by equation (1), where
and . Extinction rate as in Levins’s model2 2 2C (a) p a /(a � y ) y p 0.6patch
(eq. [6], with ).e p 0.12
E (b) p h(b), (10)patch
where . This gives′h (b) 1 0
f(x) p x[(1 � x)Km � h(mx)]. (11)
It is straightforward to show that if , then∗f(x ) p 0. Thus, there can be only one equilibrium point′ ∗f (x ) ! 0
and it is stable. The equilibrium patch number is∗Nsmaller than in Levins’s model with extinction rate e p
(fig. 4A). One consequence of the antirescue effect,h(0)opposite to the earlier investigated scenarios, is that in-creased migration will not always lead to successively larger
(fig. 4B). In particular, if h(b)/b is increasing for large∗Nb (e.g., ), then will decrease for large2 ∗h(b) p e[1 � b ] Nm (but never reach 0). This might have important bio-logical consequences; for example, conservation effortssuch as increasing the connectivity between patches orartificially colonizing empty patches in a threatened meta-
population can be detrimental if the antirescue effect isstrong.
Allee Effect
In Levins’s model the arrival of an immigrant at an un-occupied patch changes that patch into an occupied patch.This is generalized to in our model. How-C (a) p Kapatch
ever, colonization rate need not be proportional to im-migration rate. For example, it may be difficult initiallyto establish subpopulations because of demographic, sto-chastic, or genetic complications at low population num-bers—the “Allee effect” (Allee 1931). Consider the S-shaped colonization function ,2 2 2C (a) p a /(a � y )patch
where y is a constant (as earlier used in a structured modelby Hanski [1994]) and . At the metapopu-E (b) p epatch
lation level, the most important consequence of thischange in the colonization function is that there can betwo equilibria of patch occupancy (fig. 5). If patch oc-

Unifying Metapopulation Model 179
cupancy is below , extinction rate exceeds colonization∗NL
and the population rapidly goes extinct. Above and∗NL
below the higher equilibrium colonization rate exceeds∗NH
extinction rate, and the patch occupancy will increase untilis reached (fig. 5A). Other similar S-shaped coloni-∗NH
zation functions give the same qualitative results.
The Effects of Differential Migration
So far we have assumed that migrants settle on a patchchosen at random. Migrants may, however, avoid patchesthat are already occupied in order to avoid competitionfor territories or food. Alternatively, migrants seeking amate, or those at risk from predation while alone, may tryto settle on already occupied patches. When there is suchdifferential migration, the rate of arrival of potential col-onizers is no longer given by , and this willa(N) p mN/Tchange how colonization rate depends on N. To investigatethe effect of differential migration, we take the per patchcolonization and extinction rates to be andC (a) p Kapatch
, respectively, as in Levins’s model. Note thisE (b) p epatch
means that does not depend on N whenC [a(N)]/Npatch
.a(N) p mN/TConsider first the extreme case when migrants avoid
occupied patches and each settles on an empty patch cho-sen at random. Then and hencea(N) p mN/(T � N)
C (N)total p Kmx. (12)T
If , then the metapopulation goes extinct as in Lev-Km ! eins’s model. If , then at the unique equilibrium allKm 1 epatches are occupied. In reality, differential migration isnot likely to be this extreme, particularly since not allmigrants will be able to find an unoccupied patch whenthese are rare. Different degrees of this “repellent behavior”will produce different colonization functions, but all havesteeper initial increase and result in higher equilibriumpatch occupancies (N/T) than the symmetrical coloniza-tion function of Levins’s model.
Now suppose that migrants seek occupied patches. Inthe extreme case in which there is no immigration to un-occupied patches, there is no colonization, and the pop-ulation will go extinct. Suppose instead that migrants settleon an unoccupied patch if they do not soon encounter anoccupied patch. Then a(N)/N will be a decreasing functionof N. Thus, the function Cpatch[a(N)]/N will be decreasingin N. This is in contrast to models in which there isan Allee effect and no differential migration, whereCpatch[a(N)]/N is an increasing function of N. In this sensethere is an “anti-Allee effect” when migrants seek occupiedpatches.
We can also consider the effect of differential migration
on the extinction rate. When migration rate affects per-patch extinction rate, as happens when there is a rescueor antirescue effect, differential migration will interact withthese effects to distort the extinction function Etotal(N). Forexample, suppose that the patch extinction rate increaseswith immigration rate as a result of the spread of disease.Then extinction rate will be lowest when few patches areoccupied. Suppose that migrants also try to avoid occupiedpatches. Since they will be most successful at doing sowhen few patches are occupied, the extinction rate will befurther reduced when few patches are occupied but couldbe little affected when most patches are occupied. Thus,in this example, the differential migration would accen-tuate the antirescue effect in the metapopulation evenmore.
Translating Structured Models
Metapopulation models in which local populations havestructure appear fundamentally more complex than ourgeneralized metapopulation model. However, as the fol-lowing example shows, they can be translated into ourmuch simpler framework while still preserving the im-portant metapopulation-scale features.
Let us first consider a simple structured metapopulationmodel that has been developed to study metapopulationswhere immigration positively affects local populations, in-creasing population density and reducing population ex-tinction risk. Thus, this is a model designed to capturethe rescue effect (Hanski 1985; see also Hanski and Gyl-lenberg 1993; Hanski 1999). Assume that patches are inone of three states: empty (state 0), low population (state1), or high population (state 2). A low-population patchtransforms into a high-population patch at rate r1(a),where a is the rate at which it receives immigrants. Ahigh-population patch changes to a low-population patchat rate r2(a). We assume that r1(a) is an increasing functionof a and r2(a) is a decreasing function. A patch in statei ( ) sends out migrants at rates mi. We assume thati p 1, 2
. A patch in state i ( ) changes into anm ! m i p 1, 21 2
empty patch at rate ei. We assume . An empty patche 1 e1 2
changes into a low-population patch at rate Ka.We now assume that extinction and colonization rates,
e1, e2, and Ka, are small compared with the rates r1 andr2. (Later we comment on the case when this approxi-mation does not hold.) Then the proportion of occupiedpatches that are of each type can be assumed to equilibratebefore there is a change in the number of occupied patchesdue to a colonization or extinction event. Specifically, letthere be N occupied patches, of which a proportion r arehigh population. Then the rate at which patches receiveimmigrants is
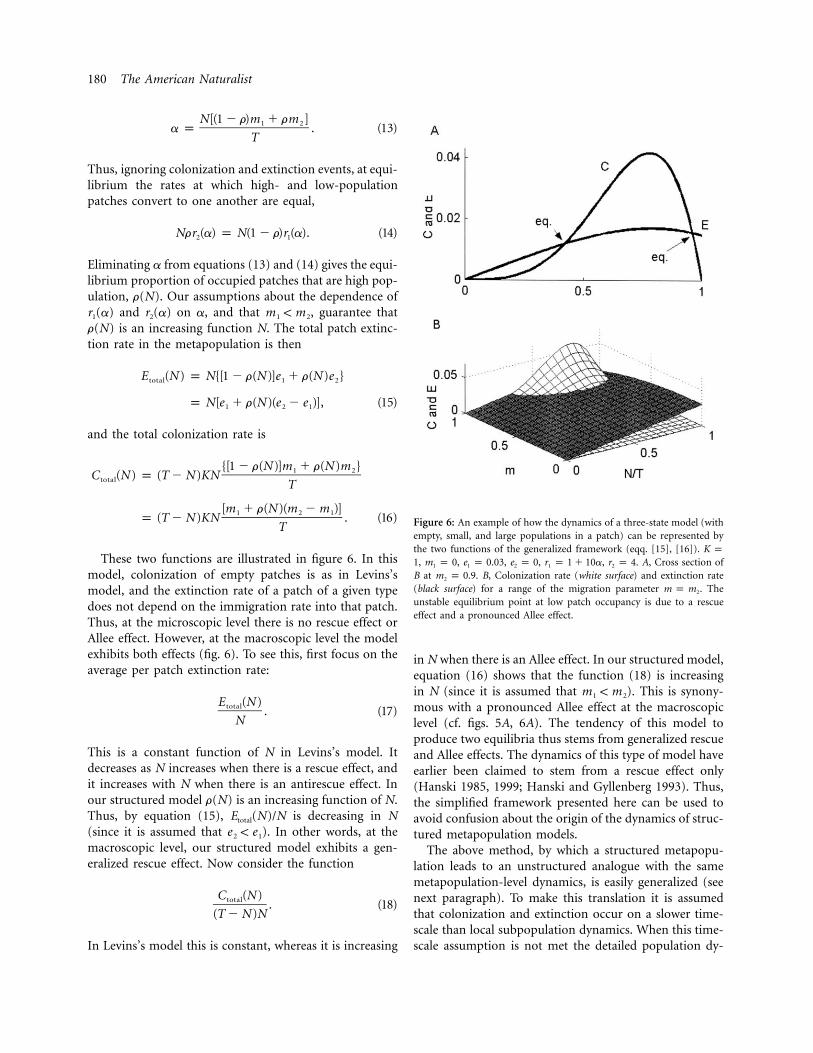
180 The American Naturalist
Figure 6: An example of how the dynamics of a three-state model (withempty, small, and large populations in a patch) can be represented bythe two functions of the generalized framework (eqq. [15], [16]). K p
, , , , , . A, Cross section of1 m p 0 e p 0.03 e p 0 r p 1 � 10a r p 41 1 2 1 2
B at . B, Colonization rate (white surface) and extinction ratem p 0.92
(black surface) for a range of the migration parameter . Them p m2
unstable equilibrium point at low patch occupancy is due to a rescueeffect and a pronounced Allee effect.
N[(1 � r)m � rm ]1 2a p . (13)
T
Thus, ignoring colonization and extinction events, at equi-librium the rates at which high- and low-populationpatches convert to one another are equal,
Nrr (a) p N(1 � r)r (a). (14)2 1
Eliminating a from equations (13) and (14) gives the equi-librium proportion of occupied patches that are high pop-ulation, r(N). Our assumptions about the dependence ofr1(a) and r2(a) on a, and that , guarantee thatm ! m1 2
r(N) is an increasing function N. The total patch extinc-tion rate in the metapopulation is then
E (N) p N{[1 � r(N)]e � r(N)e }total 1 2
p N[e � r(N)(e � e )], (15)1 2 1
and the total colonization rate is
{[1 � r(N)]m � r(N)m }1 2C (N) p (T � N)KNtotal T
[m � r(N)(m � m )]1 2 1p (T � N)KN . (16)T
These two functions are illustrated in figure 6. In thismodel, colonization of empty patches is as in Levins’smodel, and the extinction rate of a patch of a given typedoes not depend on the immigration rate into that patch.Thus, at the microscopic level there is no rescue effect orAllee effect. However, at the macroscopic level the modelexhibits both effects (fig. 6). To see this, first focus on theaverage per patch extinction rate:
E (N)total . (17)N
This is a constant function of N in Levins’s model. Itdecreases as N increases when there is a rescue effect, andit increases with N when there is an antirescue effect. Inour structured model r(N) is an increasing function of N.Thus, by equation (15), Etotal(N)/N is decreasing in N(since it is assumed that ). In other words, at thee ! e2 1
macroscopic level, our structured model exhibits a gen-eralized rescue effect. Now consider the function
C (N)total . (18)(T � N)N
In Levins’s model this is constant, whereas it is increasing
in N when there is an Allee effect. In our structured model,equation (16) shows that the function (18) is increasingin N (since it is assumed that ). This is synony-m ! m1 2
mous with a pronounced Allee effect at the macroscopiclevel (cf. figs. 5A, 6A). The tendency of this model toproduce two equilibria thus stems from generalized rescueand Allee effects. The dynamics of this type of model haveearlier been claimed to stem from a rescue effect only(Hanski 1985, 1999; Hanski and Gyllenberg 1993). Thus,the simplified framework presented here can be used toavoid confusion about the origin of the dynamics of struc-tured metapopulation models.
The above method, by which a structured metapopu-lation leads to an unstructured analogue with the samemetapopulation-level dynamics, is easily generalized (seenext paragraph). To make this translation it is assumedthat colonization and extinction occur on a slower time-scale than local subpopulation dynamics. When this time-scale assumption is not met the detailed population dy-

Unifying Metapopulation Model 181
namics of the structured metapopulation model and ourunstructured analogue will no longer agree. However, ifthe goal is to determine the equilibria of a structured meta-population, the approach outlined above is still valid. Thisis because, at an equilibrium of a metapopulation, time-scales are not important since the system is stationary.Thus, the unstructured analogue, which assumes fast localdynamics, has the same equilibria as the original structuredmetapopulations with slow local dynamics.
More Complex Structured Models
The above approach, in which a three-state model is trans-lated into an unstructured model, generalizes to morecomplex models as follows. Suppose that there are a totalof T patches and that the population on a patch may beof the types , where type 0 correspondsI � 1 0, 1, 2, … , Ito the patch being empty. The other states can describedifferent patch sizes or actual local population sizes. As inour example, we assume that patch dynamics is fast relativeto colonization and extinction rates. The number N ofoccupied patches then determines the proportion ri(N)that are type i. Let a type i population contribute migrantsto a pool at rate mi. Then the total rate at which migrantsenter the pool is Nm(N), where
I
m(N) p r (N)m . (19)� i iip1
If pool members settle on patches at random, the rate atwhich unoccupied patches receive immigrants is a p
. Assuming that the colonization rate of an emptym(N)N/Tpatch is Cpatch(a), we have
C (N)total p (1 � x)C [xm(N)], (20)patchT
where . Let a type i population ( )x p N/T i p 1, … , Ibecome extinct at rate ei. Then the extinction rate is
, whereE (N) p NE (N)total patch
I
E (N) p r (N)e . (21)�patch i iip1
Thus, a structured model (with constant extinction prob-ability for given structure) can be replaced by the gener-alized Levins-type model with no structure but a migra-tion-rate-dependent extinction risk.
Finite Patch Numbers
So far we have assumed that the total number of patches(T) is large. It has also been implicitly assumed that col-onization and extinction events at one location occur in-dependently of what occurs at other patch locations (giventhe current N). It is then reasonable to treat colonizationand extinction events as occurring at a smooth rate. Meta-population dynamics are consequently deterministic, anda metapopulation that reaches a stable equilibrium willremain there indefinitely. Conservation theory is, however,often concerned with the vulnerability to extinction ofmetapopulations that are subject to stochastic effects (Til-man et al. 1994; Hill and Caswell 2001). The details ofhow stochasticity operates are then crucial to questions ofmetapopulation persistence (Lande et al. 1998). For ex-ample, there might be large-scale fluctuations in weatherconditions that affect all patches in a highly correlated way(Harrison and Quinn 1989). Alternatively, patches couldbe independent as in our deterministic model, with thestochasticity due to the low number of patches (Hanski1999; Hill and Caswell 2001). We do not attempt a fullanalysis of the effects of stochasticity in this article. Instead,we illustrate the effects of stochasticity in this latteralternative.
When T is low, the actual events of extinction and col-onization become important. Suppose N patches arecurrently occupied. Before, the functions Ctotal(N) andEtotal(N) were interpreted as rates. Now we assume thatthe time to wait for the next patch colonization (given noextinction occurs before then) is an exponentially distrib-uted random variable with parameter Ctotal(N). Similarly,the time to wait for the next extinction (given no colo-nization occurs before then) is an exponentially distributedrandom variable with parameter Etotal(N). These two ran-dom variables are assumed to be independent. Under theseassumptions the time for the number of occupied patchesto change has an exponential distribution with mean
. After the change, the number�1t(N) p [C(N) � E(N)]of occupied patches is with probabilityN � 1 p(N) p
and is with probabilityC(N)/[C(N) � E(N)] N � 1 1 �.p(N)
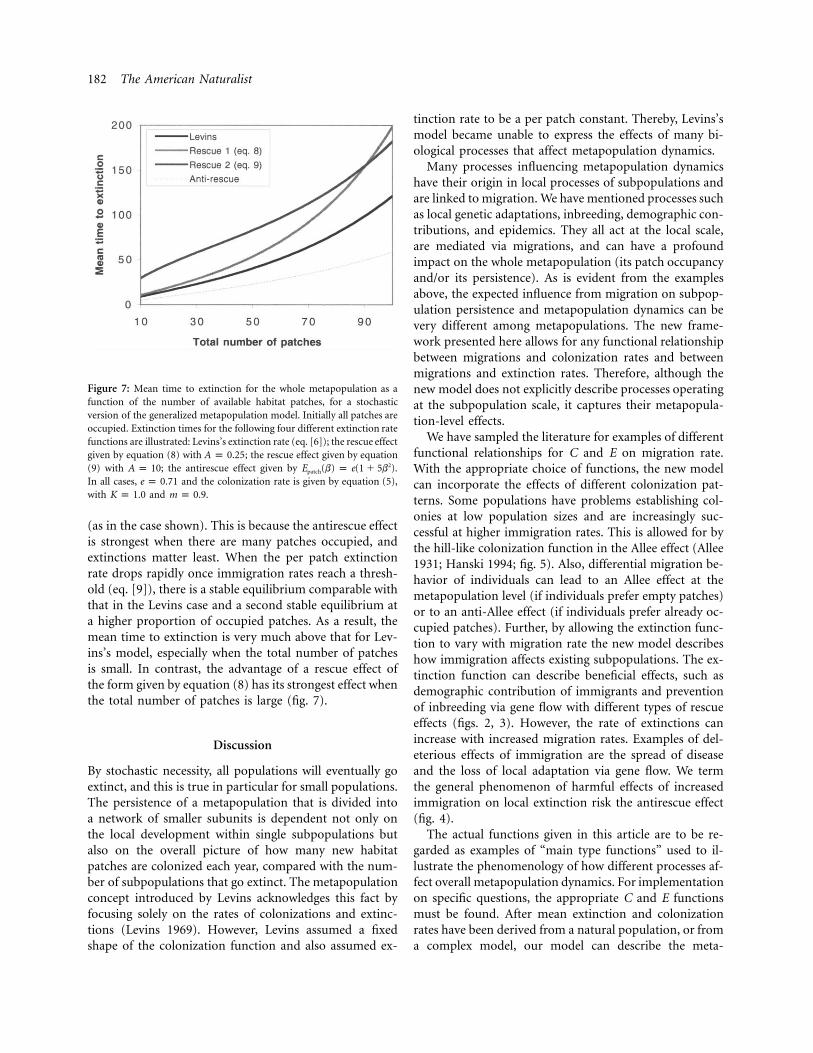
Figure 7 illustrates the use of this stochastic version ofour generalized model. The figure shows how the meantime to population extinction depends on the total numberof patches in the environment. In each of the four casesshown, we have kept the colonization function as in Lev-ins’s model. The extinction functions presented are fortwo different cases of the rescue effect and a case with theantirescue effect, together with the baseline Levins case forcomparison. When there is an antirescue effect, the meantime to extinction is only substantially reduced when thedetrimental effect of increased immigration is very strong
182 The American Naturalist
Figure 7: Mean time to extinction for the whole metapopulation as afunction of the number of available habitat patches, for a stochasticversion of the generalized metapopulation model. Initially all patches areoccupied. Extinction times for the following four different extinction ratefunctions are illustrated: Levins’s extinction rate (eq. [6]); the rescue effectgiven by equation (8) with ; the rescue effect given by equationA p 0.25(9) with ; the antirescue effect given by .2A p 10 E (b) p e(1 � 5b )patch
In all cases, and the colonization rate is given by equation (5),e p 0.71with and .K p 1.0 m p 0.9
(as in the case shown). This is because the antirescue effectis strongest when there are many patches occupied, andextinctions matter least. When the per patch extinctionrate drops rapidly once immigration rates reach a thresh-old (eq. [9]), there is a stable equilibrium comparable withthat in the Levins case and a second stable equilibrium ata higher proportion of occupied patches. As a result, themean time to extinction is very much above that for Lev-ins’s model, especially when the total number of patchesis small. In contrast, the advantage of a rescue effect ofthe form given by equation (8) has its strongest effect whenthe total number of patches is large (fig. 7).
Discussion
By stochastic necessity, all populations will eventually goextinct, and this is true in particular for small populations.The persistence of a metapopulation that is divided intoa network of smaller subunits is dependent not only onthe local development within single subpopulations butalso on the overall picture of how many new habitatpatches are colonized each year, compared with the num-ber of subpopulations that go extinct. The metapopulationconcept introduced by Levins acknowledges this fact byfocusing solely on the rates of colonizations and extinc-tions (Levins 1969). However, Levins assumed a fixedshape of the colonization function and also assumed ex-
tinction rate to be a per patch constant. Thereby, Levins’smodel became unable to express the effects of many bi-ological processes that affect metapopulation dynamics.
Many processes influencing metapopulation dynamicshave their origin in local processes of subpopulations andare linked to migration. We have mentioned processes suchas local genetic adaptations, inbreeding, demographic con-tributions, and epidemics. They all act at the local scale,are mediated via migrations, and can have a profoundimpact on the whole metapopulation (its patch occupancyand/or its persistence). As is evident from the examplesabove, the expected influence from migration on subpop-ulation persistence and metapopulation dynamics can bevery different among metapopulations. The new frame-work presented here allows for any functional relationshipbetween migrations and colonization rates and betweenmigrations and extinction rates. Therefore, although thenew model does not explicitly describe processes operatingat the subpopulation scale, it captures their metapopula-tion-level effects.
We have sampled the literature for examples of differentfunctional relationships for C and E on migration rate.With the appropriate choice of functions, the new modelcan incorporate the effects of different colonization pat-terns. Some populations have problems establishing col-onies at low population sizes and are increasingly suc-cessful at higher immigration rates. This is allowed for bythe hill-like colonization function in the Allee effect (Allee1931; Hanski 1994; fig. 5). Also, differential migration be-havior of individuals can lead to an Allee effect at themetapopulation level (if individuals prefer empty patches)or to an anti-Allee effect (if individuals prefer already oc-cupied patches). Further, by allowing the extinction func-tion to vary with migration rate the new model describeshow immigration affects existing subpopulations. The ex-tinction function can describe beneficial effects, such asdemographic contribution of immigrants and preventionof inbreeding via gene flow with different types of rescueeffects (figs. 2, 3). However, the rate of extinctions canincrease with increased migration rates. Examples of del-eterious effects of immigration are the spread of diseaseand the loss of local adaptation via gene flow. We termthe general phenomenon of harmful effects of increasedimmigration on local extinction risk the antirescue effect(fig. 4).
The actual functions given in this article are to be re-garded as examples of “main type functions” used to il-lustrate the phenomenology of how different processes af-fect overall metapopulation dynamics. For implementationon specific questions, the appropriate C and E functionsmust be found. After mean extinction and colonizationrates have been derived from a natural population, or froma complex model, our model can describe the meta-

Unifying Metapopulation Model 183
population-scale dynamics in a transparent and easily an-alyzed way. This makes it possible to classify and comparethe main behaviors of completely different complexsystems.
So what can we expect the C and E functions to be likein real-world metapopulations? We believe it can be usefulto consider how the metapopulation has been formed.Many populations live on “naturally” patchy habitats.Habitats can be networks of rock pools, islands, the in-testines of birds, or, as a more unusual example, the sunkencarcasses of whales (T. Dahlgren, unpublished manu-script). We suggest that species evolved in such systemscan be expected to have developed optimal dispersal strat-egies related to the benefits and risks of migrations in theirenvironments. Another type of patchy population is ar-tificially created. Modern farming and forestry havechanged the main vegetation of entire landscapes, leavingonly scattered patches of the once dominant vegetationtypes left. Species that have inhabited larger, unbrokenhabitats for perhaps thousands of generations might nothave optimal dispersal strategies to cope with the newconstraints imposed by a sudden fragmentation of theirhabitats.
The focus of metapopulation research has been directedtoward endangered species, living on small remnants oftheir natural habitat. Those populations are likely to ben-efit from increased migration rates to counteract their im-posed isolation. Consequently, a rescue effect is includedin most metapopulation models investigating conservationissues (such as habitat loss). However, it is important torule out other functional relationships since high migra-tion rates can be detrimental for some populations (e.g.,by inducing synchronous fluctuations in local populations[Vandermeer and Carvajal 2001] or by loss of genetic di-versity [Lacy 1987; Pannell and Charlesworth 1999]). Theantirescue effect highlights this possibility in a phenom-enological way (fig. 4). Two examples where extinctionrisk increases as migration rate increases are the periwinkle(Rolan-Alvarez et al. 1997; box 1A) and a New Zealandweevil (Schops 1999; box 1B). Studies of metapopulationswith antirescue effects can give insights into the evolutionof nondispersal strategies, for example, site fidelity, wing-lessness, and territoriality. The awareness of how coloni-zation and extinction functions can differ from what ismostly assumed in models of metapopulations thus alsohas practical significance. The successful managementstrategy of a metapopulation with an antirescue effect willdiffer dramatically from the common scenario. One illus-tration of this is how the expected mean time to extinctionwill change, depending on the assumed shapes of extinc-tion and colonization functions, as the number of habitatpatches decreases (fig. 7). We want to underline the ne-cessity to develop conservation theory for several different
types of metapopulations and to find the relevant colo-nization and extinction functions for real-world meta-population before implementation.
Earlier extensions of Levins’s model that tried to makeit more realistic added a linear rescue effect to Levins’smodel to take into account demographic contributionfrom migration (Hanski 1982, 1985). Also, Gotelli andKelly (1993) suggested that C and E could be any functionof N; however, they only explored the linear rescue effect.The linear rescue effect gives only one stable equilibriumof patch occupancy. The dynamic limitations of Levins’smodel and these extensions with linear rescue effect werebelieved to be an inherent property of all unstructuredmetapopulation models: “The most fundamental differ-ence between the unstructured and structured metapop-ulation models is that with the structured models therecan be multiple equilibria” (Gyllenberg et al. 1997, p. 107;see also Hanski and Gyllenberg 1993). Consequently, theuse of simple Levins-type models in analyzing generalquestions such as consequences of patch destruction onmetapopulation persistence (Tilman et al. 1994) has beencriticized (Gyllenberg et al. 1997). We show that a simplemodel can express most of the complex behaviors de-scribed for structured metapopulation models by varyingthe different functional relationships between migration,colonization, and extinction rates. Although there havepreviously been specific simple extensions to Levins’smodel, there has been no attempt to systematically inves-tigate how the form of the colonization and extinctionfunctions affects metapopulation dynamics. Our article ad-vocates such a study.
For some studies, the desired general C and E functionsfor a specific biological process can be found by first build-ing a structured model, which then is reduced to our sim-plified model. The simplified model can capture the es-sential detail, at the metapopulation level, of the structuredmetapopulation model. Once such a simplified model hasbeen constructed, the dependence of the colonization andextinction functions on the number of occupied patches(cf. formulas [17] and [18]) reveals the metapopulation-scale properties of the original structured model. For ex-ample, it can be seen whether there is an implicit gener-alized Allee effect or a generalized rescue effect. Note thatour definition of these effects differs from those of previousauthors (Brown and Kodric-Brown 1977; Hanski and Gyl-lenberg 1993; Lande et al. 1998). Our definition capturesthe important features at the metapopulation scale ratherthan at the smaller subpopulation scale. Some structuredmodels with complex local dynamics (e.g., due to infec-tious diseases or predator-prey interactions) can exhibitlimit cycles and chaos at the metapopulation level (May1977; Hassell and Wilson 1997). We propose such complexdynamics should be captured by the introduction of time

184 The American Naturalist
lags into our unstructured analogue. Studies that addressspecific processes (such as influence from explicit spatiallocation, time since colonization, and genotype frequen-cies), must obviously utilize more complex models allow-ing for these parameters to be estimated. The generalizedmodel is designed to capture processes that leave a sig-nature at the metapopulation scale.
The generalized model presented here can be used as aphenomenological model in the study of general aspectsof metapopulation structure. Interesting areas for theo-retical applications are environmental stochasticity and ex-tinction risks for metapopulations with different types ofcolonization and extinction functions, evolution of opti-mal migration rates, time-lagged responses in patch oc-cupancy, and investigation of the implicit assumptions ofmore complex models.
Acknowledgments
We wish to thank the following persons for valuable com-ments: I. Hanski, T. Harkonen, R. Holt, A. Houston, H.Kokko, E. Ranta, A. Sih, and D. Wahlstedt. K.C.H. wassupported by the Oscar and Lili Lamm ResearchFoundation.
Literature Cited
Allee, W. C. 1931. Animal aggregations: a study in generalsociology. University of Chicago Press, Chicago.
Allen, J. C., W. M. Schaffer, and D. Rosko. 1993. Chaosreduces species extinction by amplifying local popula-tion noise. Nature 364:229–232.
Brown, J. H., and A. Kodric-Brown. 1977. Turnover ratesin insular biogeography: effect of immigration on ex-tinction. Ecology 58:445–449.
Burgman, M. A., S. Ferson, and H. R. Akcakaya. 1993.Risk assessment in conservation biology. Chapman &Hall, London.
Caswell, H. 1988. Theory and models in ecology: a dif-ferent perspective. Ecological Modelling 43:33–44.
Gotelli, N. J., and W. G. Kelly. 1993. A general model ofmetapopulation dynamics. Oikos 68:36–44.
Grenfell, B. T., and J. Harwood. 1997. (Meta)populationdynamics of infectious diseases. Trends in Ecology &Evolution 12:395–399.
Grenfell, B. T., B. M. Bolker, and A. Kleczkowski. 1995.Seasonality and extinction in chaotic metapopulations.Proceedings of the Royal Society of London B, BiologicalSciences 259:97–103.
Gyllenberg, M., I. Hanski, and A. Hastings. 1997. Struc-tured metapopulation models. Pages 93–122 in I. Han-ski and M. E. Gilpin, eds. Metapopulation biology. Ac-ademic Press, London.
Hanski, I. 1982. Dynamics of regional distribution: thecore and satellite species hypothesis. Oikos 38:210–221.
———. 1985. Single-species spatial dynamics may con-tribute to long-term rarity and commonness. Ecology66:335–343.
———. 1994. A practical model of metapopulation dy-namics. Journal of Animal Ecology 63:151–162.
———. 1999. Metapopulation ecology. Oxford UniversityPress, Oxford.
Hanski, I., and M. Gyllenberg. 1993. Two general meta-population models and the core-satellite species hy-pothesis. American Naturalist 142:17–41.
Hanski, I., and D.-Y. Zhang. 1993. Migration, metapop-ulation dynamics and fugitive co-existence. Journal ofTheoretical Biology 163:491–504.
Harrison, S., and J. F. Quinn. 1989. Correlated environ-ments and the persistence of metapopulatons. Oikos 56:293–298.
Hassell, M. P., and H. B. Wilson. 1997. The dynamics ofspatially distributed host-parasitoid systems. Pages 75–110 in D. Tilman and P. Kareiva, eds. Spatial ecology:the role of space in population dynamics and interspe-cific interactions. Princeton University Press, Princeton,N.J.
Hastings, A., and S. Harrison. 1994. Metapopulation dy-namics and genetics. Annual Review of Ecology andSystematics 25:167–188.
Hastings, A., and C. Volin. 1989. Within-patch dynamicsin a metapopulation. Ecology 70:1261–1266.
Hill, M. F., and H. Caswell. 2001. The effects of habitatdestruction in finite landscapes: a chain-binomial meta-population model. Oikos 93:321–331.
Holt, D. R. 1997. From metapopulation dynamics to com-munity structure. Pages 149–164 in I. Hanski and M.E. Gilpin, eds. Metapopulation biology. Academic Press,San Diego, Calif.
Joshi, J., B. Schmid, M. C. Caldeira, P. G. Dimitrakopoulos,J. Good, R. Harris, A. Hector, et al. 2001. Local adap-tation enhances performance of common plant species.Ecology Letters 4:536–544.
Lacy, R. C. 1987. Loss of genetic diversity from managedpopulations: interacting effects of drift, mutation, im-migration, selection, and population subdivision. Con-servation Biology 1:143–158.
Lande, R., S. Engen, and B.-E. Sæther. 1998. Extinctiontimes in finite metapopulation models with stochasticlocal dynamics. Oikos 83:383–389.
Levins, R. 1969. Some demographic and genetic conse-quences of environmental heterogeneity for biologicalcontrol. Bulletin of the Entomological Society of Amer-ica 15:237–240.
May, R. M. 1977. Thresholds and breakpoints in ecosys-

Unifying Metapopulation Model 185
tems with a multiplicity of stable states. Nature 269:471–477.
Pannell, J. R., and B. Charlesworth. 1999. Neutral geneticdiversity in a metapopulation with recurrent local ex-tinction and recolonization. Evolution 53:664–676.
Rolan-Alvarez, E., K. Johannesson, and J. Erlandsson.1997. The maintenance of a cline in the marine snailLittorina saxatilis: the role of home site advantage andhybrid fitness. Evolution 51:1838–1847.
Sacceri, I., M. Kuussaari, M. Kankare, P. Vikman, W. For-telius, and I. Hanski. 1998. Inbreeding and extinctionin a butterfly metapopulation. Nature 392:491–494.
Schops, K. 1999. Effect of deforestation on the spatialdynamics of an endangered weevil species. Page 45 inHabitat loss: ecological, evolutionary and genetic con-
sequences, Helsinki, September 7–12, 1999. Haka-paino Oy, Helsinki. http://www.helsinki.fi/ml/ekol/se/hleegc1.html
Taylor, A. D. 1990. Metapopulations, dispersal, and pred-ator-prey dynamics: an overview. Ecology 71:429–433.
Tilman, D., and P. Kareiva. 1997. Spatial ecology: the roleof space in population dynamics and interspecific in-teractions. Princeton University Press, Princeton, N.J.
Tilman, D., R. M. May, C. L. Lehman, and M. A. Nowak.1994. Habitat destruction and the extinction debt. Na-ture 371:65–66.
Vandermeer, J., and R. Carvajal. 2001. Metapopulationdynamics and the quality of the matrix. American Nat-uralist 158:211–220.
Associate Editor: Per Lundberg