acomparativeevaluationofanovelvaccineinappps1mousemodelsofalzheimer’sdisease
DESCRIPTION
alzheimer vacunaTRANSCRIPT

Research ArticleA Comparative Evaluation of a Novel Vaccine in APPPS1 MouseModels of Alzheimerrsquos Disease
Ivaacuten Carrera1 Ignacio Etcheverriacutea1 Luciacutea Fernaacutendez-Novoa1
Valter Ruggero Maria Lombardi1 Madepalli Krishnappa Lakshmana2
Ramoacuten Cacabelos3 and Carmen Vigo4
1 Department of Neuroscience EuroEspes Biotechnology Poligono de Bergondo Nave F 15165 A Coruna Spain2Department of Neurobiology Torrey Pines Institute for Molecular Studies 11350 SW Village ParkwayPort Saint Lucie FL 34987 USA
3 EuroEspes Biomedical Research Center Institute for CNS Disorders and Chair of Genomic Medicine University of Camilo Jose Cela15165 Madrid Spain
4Atlas Pharmaceuticals Sunnyvale CA 94089 USA
Correspondence should be addressed to Ivan Carrera neuromorfologiaebioteccom
Received 10 July 2014 Accepted 14 September 2014
Academic Editor Mohammad I Kamboh
Copyright copy 2015 Ivan Carrera et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Immunization against amyloid-beta-peptide (A120573) has been widely investigated as a potential immunotherapeutic approach forAlzheimerrsquos disease (AD) With the aim of developing an active immunogenic vaccine without need of coadjuvant modificationfor human trials and therefore avoiding such side effects we designed the A120573
1ndash42 vaccine (EB101) delivered in a liposomal matrixthat based on our previous studies significantly prevents and reverses the AD neuropathology clearing A120573 plaques while markedlyreducing neuronal degeneration behavioral deficits and minimizing neuroinflammation in APPPS1 transgenic mice Here theefficacy of our immunogenic vaccine EB101 was compared with the original immunization vaccine cocktail A120573
42+ CFAIFA
(Freundrsquos adjuvant) in order to characterize the effect of sphingosine-1-phosphate (S1P) in the immunotherapeutic responseQuantitative analysis of amyloid burden showed a notable decrease in the neuroinflammation reaction against A120573 plaques when S1Pwas compared with other treatments suggesting that S1P plays a key role as a neuroprotective agent Moreover EB101 immunizedmice presented a protective immunogenic reaction resulting in the increase of A120573-specific antibody response and decrease ofreactive glia in the affected brain areas leading to a Th2 immunological reaction
1 Introduction
Alzheimerrsquos disease (AD) is the major cause of demen-tia worldwide and the development of effective therapiesremains a major unmet medical need It is known that it isa heterogeneous and complex disorder in which hundredsof genes distributed across the human genome might beinvolved in close cooperationwith environmental factors andepigenetic phenomena leading to cerebrovascular dysfunc-tion [1] The resulting cognitive impairment is mainly associ-ated with neuronal degeneration at the hippocampal dentategyrus and entorhinal cortex among other brain regionsimplicated in cognitive function involved in learning andmemory processes [2] Based on amyloid cascade hypothesis
elevated levels of A120573 have been correlated with cognitivedecline mediating initial pathogenic events in AD dementia[3 4] such as the accumulation of senile plaques and plaque-associated dystrophic neurites in the brain [5 6] Conse-quently a progressive neuroinflammatory reaction has beenobserved surrounding A120573 plaques resulting in astrogliosisand microglial activation This inflammatory process mayindependently lead to neural dysfunction and cell deaththus establishing a self-perpetuating vicious cycle which fur-ther contributes to neurodegeneration and enhances thepathological hallmarks of the disease [7] Assuming that theneuropathological pathway defined by the amyloid cascadehypothesis causes AD suppression of A120573 in the brains ofpatients in the early phase of dementia should become
Hindawi Publishing CorporationBioMed Research InternationalVolume 2015 Article ID 807146 16 pageshttpdxdoiorg1011552015807146
2 BioMed Research International
a primary therapeutic target In this case A120573 immunothera-pies would become the most promising preclinical strategiesas they have been proven to enhance clearance of A120573 in thebrain of mice models
In order to investigate new therapeutic strategies inAD APPswePS1ΔE9 (APPPS1) mice which overexpress theSwedish mutation of APP together with PS1 deleted in exon 9that rapidly accumulates A120573 plaques at 6 months of age [8 9]have been extensively used in AD research [10] APPPS1mice also develop behavioral and learning deficits [11]plaque-associated neuritic abnormalities [12] inflammationreflected in activated microglia and astrocytes surroundingthe A120573 plaques [13] and deficits in the pre- and postsynap-tic cholinergic transmission [14] In the last decade thesetransgenic mousemodels were extensively used in preclinicalstudies of active immunization [15 16] with preaggregatedA120573 showing high anti-A120573 antibody titers in plasma dramaticreduction of cerebral A120573 burden and reduction in cognitivedecline [17 18] These results in mice did not translate wellin humans Clinical trials conducted by ElanWyeth in 2001using A120573 peptide delivered in QS-21 adjuvant (AN1792)resulted in a meningoencephalitis reaction in 6 of thetreated patients The trial then was immediately stoppedSubsequent studies suggested that these adverse events hadbeen initiated by activation of cytotoxic T cells andorautoimmune reactions [19ndash23] Recently we reported thatimmunotherapeutic treatment with EB101 vaccine consistingof A120573
1ndash42 delivered in a novel immunogen-adjuvant com-posed of liposomes-containing phosphatidylcholine phos-phatidylglycerol cholesterol and sphingosine-1-phosphate(S1P) results in a marked reduction of A120573 plaque burden anddystrophic plaque neurite density diminution of astrocytosisand attenuation of amyloidosis-induced inflammation [2425] In the present study we compare the efficacy of EB101versus the original immunization vaccine cocktail (A120573
42+
CFAIFA) [17] in the APPPS1 mouse model before and afterthe onset of A120573 pathology Moreover we also characterizedthe effect of S1P in the immunotherapeutic response of EB101in this mouse model showing that it plays a key role as aregenerative agent in the central nervous system [26] Thefindings presented were obtained by using immunocyto-chemistry techniques neuronal anatomic mapping and seraantibodycytokines detection by ELISA andmotor behavioraltests suggesting a notable effectiveness of EB101 over A120573
42+
CFAIFA vaccine in clearingA120573 plaques reducing dystrophicplaque neurites preventing inflammation in the entorhinalcortex and hippocampus in this transgenic mouse modelThese results warrant further studies which could prove thatEB101 is a promising vaccine to treat AD patients avoidingadverse effects
2 Materials and Methods
21 Animals Awell-studiedmousemodel of A120573 amyloidosisis the double-transgenic mice B6C3F1J (APPswePS1dE9)expressing a chimeric mousehuman amyloid precursor pro-tein (MoHuAPP695swe) and human presenilin 1 (PS1-ΔE9)mutants both directed to central nervous system (CNS)neurons that exhibits A120573 plaques in the hippocampus and
cortex beginning at 6 months of age (Jackson Laboratory BarHarbor ME) All experimental procedures were conformedto the guidelines established by the European Commu-nities Council Directive (86609EEC) the EU Directive201063EU and the Spanish Royal Decree 12012005 foranimal experimentation and were approved by the EthicalCommittee of the EuroEspes Biotechnology Research Centre(Permit number EE2012-344)
22 Experimental Design Two groups of experimental stud-ies preventive treatment (before amyloid deposition onsetstarting at 7 weeks of age) and therapeutic treatment (afteramyloid deposition onset at 35 weeks of age) were carriedout as described in recent reports [24 25] similar to the long-term protocol reported by Schenk and colleagues [17] andrepresented in Figure 1Mice of both sexes (balanced betweentreatment groups) were randomly assigned to each of thetwo experimental groups and they were divided into fivetreatment groups per study Preventive treatment Group Awas formed by 20 mice (14 transgenic and 6 wild-type mice14 + 6) that were immunized with a cocktail of synthetichumanA120573
42S1P-containing liposomes (EB101) Group Bwas
formed by 20 mice (14 + 6) immunized with A12057342liposome
without S1P Group C was formed by 20 mice (14 + 6)immunized with S1P-containing liposomes Group D wasformed by 20 mice (14 + 6) immunized with A120573
42Freundrsquos
adjuvant complete-incomplete (A12057342+CFAIFA) andGroup
E was formed by 16 mice (12 transgenic and 4 wild-typemice control) inoculated with PBS Therapeutic treatmentthe same treatments were administrated in Groups A B Cand D formed by 20 mice in each group while the GroupE was formed by 16 mice inoculated with PBS as controlMice were immunized with nine injections for sevenmonthsinoculating 100 120583L per injection containing a cocktail of A120573(100 120583g) and phospholipid mix (1mg) (Figure 1)
23 Experimental Procedure The methodological proce-dures to prepare the EB101 and A120573
42+ CFAIFA immuniza-
tions were strictly followed as described by Carrera et al[24] and Webster et al [10] respectively In summary theEB101 vaccine was prepared using 2mg of human A120573
1ndash42(TOCRIS bioscience Tocris Cookson Ltd) corresponding tothe human form of the predominant amyloid A120573 found in thebrains of patients with AD dissolved in 09mL water up to1mL by adding 01mL of 10xPBS then vortexed lyophilizedand stored Liposomes were prepared from 12-dioleoyl-sn-glycero-3-phosphocholine (DOPC) 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphatidylglycerol sodium salt (POPG) choles-terol (CH Northern lipids INC) and plusminus D-erythro-sphingosine-1-phosphate (S1P) (AVANTI at 0303039001molar ratio resp) as described by Lang and colleagues[30] To prepare the A120573
42+ CFAIFA vaccine A120573
1ndash42 wasemulsified 1 1 (vv) with complete Freundrsquos adjuvant (CFA)for the first immunization followed by a boost of incom-plete Freundrsquos adjuvant (IFA) monthly thereafter Once theimmunization period finished mice were kept in the cage for2 months before being euthanized for analysis Mice were 11months old at the end of preventive treatment and 18 monthsold at the end of therapeutic treatment
BioMed Research International 3
Trainingperiod
35 43 51 59 63 71 weeks7 19 2711
Arrived frombreeder
0 9 15 23 31
Therapeutic treatment injectionsand rotarod trials
37 39 47 55 67
Sacrificed(preventive treatment)
Sacrificed(therapeutic treatment)
Group E PBS
3
DOPC
POPG
CHOLS1P
Trainingperiod
Preventive treatment injectionsand rotarod trials
Group A A12057342 + S1P+ L (EB101)Group B A12057342 + LGroup C S1P+ LGroup D A12057342 + Freund ci (A120573FA)
(EB101) A12057342 + S1P+ L
A12057342
Figure 1 Biophysical characterization of the EB101 vaccine and experimental design Biophysical structure of A12057342S1P-containing liposome
(EB101) and schedule of experiments
24 Spleen Cell Preparation Cytokine ELISAs and FlowCytometric Analysis Mice splenocytes were harvested from aclamped spleen tissue as described by Carrera et al [24]TheB-cell-enriched suspensions were obtained from spleen cellsthat were depleted by selective adherence to glass Petri dishesfor 2 h at 37∘C This procedure yielded an enriched B-cellpopulation gt90 CD19+ cells with lt1 CD3+ cells and lt5CD11c+ cells as determined by flow cytometry analysis andgt95 of viable cells as determined by trypan blue exclusionCells were seeded into 96-well tissue culture plates at a densityof approximately 5000 cells per well After stimulation 10 120583Lof MTT solution was added to each well Absorbance wasmeasured on an ELISA plate reader with a test wavelengthof 570 nm The results obtained from triplicate assays wereexpressed as stimulation indexes (ratio between mean ODfrom stimulated and unstimulated cultures)
25 Measurement of Antibody Titers Serum antibody titerswere measured using methods described previously [24 31]Briefly 96-well microtiter plates were coated with 1 120583gmLof A120573
1ndash42 in well-coating buffer and incubated overnight atroom temperature (RT) The sera were added to the wellsat a starting dilution of 1100 in specimen diluent (0014Msodium phosphate pH 74 015M NaCl 06 bovine serumalbumin 005 thimerosal) Four serial dilutions (140011600 16400 and 125600) of the samples were madedirectly in the plates in twofold steps to reach a final dilutionof 1102400 After incubation the second antibody (goatanti-mouse IgG conjugated to horseradish peroxidase fromBoehringer Mannheim) was added to the wells (100 120583Lwellat a dilution of 14000 in specimen diluent) and incubated
for one hour at RT The chromogen reaction (33551015840-tetramethylbenzidine obtained from Pierce Chemicals) wasstopped by the addition of 50 120583L 05M H
2SO4 The color
intensity was then read on an ELISA plate reader (Bio-Rad680 ELISA Reader) Titers were defined as the reciprocalof the dilution of serum giving one half the maximumOD Maximal OD was generally taken from an initial 1100dilution
26 Immunohistochemistry Immunohistochemical data wasanalyzed by using methods described previously [24] Insummary parallel sections were pretreated with H
2O2in
phosphate-buffered saline at 37∘C for 15 minutes to elimi-nate endogenous peroxidase rinsed twice in 005M Trizmabuffered saline (TBS) containing 01 Tween-20 at pH74 (TBS-T) for 10 minutes each pretreated with blockingavidinbiotin kit and then incubated overnight with theprimary antibodies as described in Table 1 The sections weresuccessively rinsed in TBS-T incubated in goat IgG anti-rabbit (Dako) or goat IgG anti-mouse (Dako) dependingon the primary antibody for 1 hour rinsed in TBS-T andthen incubated for 30minutes in ABC kit system (VectastainVector) The labeling was revealed by incubating sectionswith 33-diaminobenzidine as chromogen and hydrogenperoxide as oxidant In several adjacent sections negativecontrols performed by omitting the primary secondary ortertiary antibodies showed no immunostaining Prussianblue iron staining was performed to detect hemosiderin toreveal signs of microhemorrhage although no considerableperivascular microhemorrhages were observed in the brainsections analyzed
4 BioMed Research International
Table1Summaryof
antib
odiesu
sed
Antibod
yDilu
tion
Immun
ogen
Source
Labelin
gClon
enam
eTy
peCatalog
number
Lotn
umber
Ref
B-Amyloid
11000
KLH-con
jugatedsynthetic
peptidec
orrespon
ding
aa36ndash4
2of
human
amyloidBx
-42peptide
Millipore
Amyloid-Bx
-42
polypeptide
12F4
Mou
semon
oclonal
Num
ber0
5-831
DAM1447121
[242527]
GFA
P140
0Antiglialfi
brillaryacid
protein
(mou
seIgG1isotype)
Sigm
aAldric
hAntiglialfi
brillary
acid
protein
G-A-5
Mou
semon
oclonal
Num
berG
3893
078K
4830
[924252829]
DNs
1300
Degeneratingplaque
neurites
Millipore
Degeneratingplaque
neuritesa
ndneurofi
brillarytang
les
mdashRa
bbitpo
lyclo
nal
Num
berA
B1518
PS014344
96[242527]
BioMed Research International 5
27 A120573 Load Calculation Digital images of 7 randomlyselected microscopic transverse sections per treated animalwere analyzed and the percentage of immunostained A120573deposition was determined for all the markers studied asdefined by the stereotaxic Bregma coordinates A total of 7selected sections per animal were evaluated using the NIHImage J program by defining region of interest and setting athreshold to discriminate nonspecific staining Quantitativeanalysis of amyloid burden area was also performed in thehippocampal regions and parietaltemporal cortical regionsWe used areapixels analysis software (Pixcavator 4) toquantify the average number of pixels inside each A120573 plaqueper brain section Therefore Pixcavator intensity thresholdimaging was set to mark only those areas stained with 120573-amyloid antibody (A120573 load) relative to the entire imagearea The immunostaining intensity of each image was firstaveraged per mouse and then per group and expressed inpercentage units
28 Imaging Images were visualized using a microscope(Olympus BX50) and digitized using a digital camera (DP-10Olympus) A120573 burden data was determined by quantitativeimage analysis (Pixcavator 4) and reported as plaques persection (ps) or percentage of immunolabeled area cap-tured within total sample area The microphotographs wereadjusted for brightness and contrast with Corel Photo-Paint(Corel Ottawa Canada) and plates were composed withCorel Draw
29 Sensorimotor and Cognitive Testing After the adminis-tration of the vaccine mice were subjected to sensorimotorand memory testing to measure their coordination andlearning performances during the experimental period andto verify that any treatment-related effects observed in thememory tasks could not be explained by differences in senso-rimotor abilities Prior to testing as previously described [32]the mice were adapted to the room and testing conditions
291 Rotarod The motor strength ability balance andcoordination skills of all the experimental mice in the twotreatment periods were evaluated in a rotarod apparatus(Columbus Instruments Columbus OH) operated at 10 rpmbeginning at 7 weeks of age (preventive group) and 35 weeksof age (therapeutic group) The animals were adapted to theapparatus by receiving training sessions during four weekssufficient to reach a baseline level of performance This pro-cedurewas designed to assessmotor behavior during six trialsof a groupday If themouse remained on the rod for 1minutethe test was completed and scored as 1 Each animal wastested previous to the treatment injection for three sessionswith each session separated by 15 minutes The time thateachmouse remained on the rodwas registered automaticallyand stopped when the animal fell or inverted (by climbing)from the top of the rotating barrel In this experimentone investigator evaluated the rotarod performance and wasblinded to mice treatments and the second one treated themice and scored the test to avoid a bias
292 Fear ConditioningActive Avoidance This fear-moti-vated associative avoidance test is based on electric currentwhere themouse has to learn to predict the occurrence of thisaversive event by actively moving to a different compartment(escape platform) This active avoidance task provides asimple way to assess associative learning and memory bytesting the ability of the mouse to avoid an aversive event inresponse to a stimulus cueThemeasures recorded number ofnonresponses (the mouse failing to reach the escape platformduring the trial) and response latency (latency to avoid orescape) serves as an index of learning and allows memoryto be assessed After habituation for 5min mice received 30randomized escapable footshocks at an intensity of 025mAIn each shock trial the time to reach the escape platformlocated constantly at a central quadrant from the startlocationwas recorded as latency Each trial lasted amaximumof 30 s If the mouse failed to find the platform within 30 s itwas manually guided to the platform and allowed to stay for10 s the latency was recorded 30 s Experimental results areindicated as mean response time for each treatment period
210 Onset and Survival Disease onset and progression wereconfirmed and defined by periodic mice sacrifice and neu-ropathological analysisThemortality rates of APPPS1 trans-genicmice in the course of both sets of experiments were 42for A120573
42+ CFAIFA 17 for EB101 and 12 for the vehicle
211 Statistical Analyses For statistical analyses SPSS (ver-sion 110 SPSS Inc Chicago) was used and a 119875 value lt005 was considered statistically significant The correlationaverage of A120573 plaque density burden area and antibody titerswere analyzed and compared in multiple groups by one-wayanalysis of variance (ANOVA) followed by Fisherrsquos LSD posthocmeans comparison test analysis when relevant All graphsdisplay group mean plusmn SEM
212 Limitations of the Study The ultimate goal of the ADpreclinical research is developing a therapeutic drug strategyin the ideal animal model that should recapitulate most ifnot all the neurobehavioral pathology and underlying thesame mechanisms described in human AD such as therate of amyloid plaque accumulation neurodegenerationand behavioral and cognitive alterations Although such anideal mouse model has not yet been generated we believethat APPswePS1ΔE9 transgenic mice present enough patho-logical features to answer the specific questions addressedby the present study Active A120573 immunotherapy in whichantibodies are produced by the individual upon contact withthe antigen has great potential for preventive AD therapy asshown here in animal models although some aspects suchas an undesirable immune response induced by the adjuvantlong immune response adjustment age-dependent effectsinflammation and microhemorrhages should be addressedbefore its application in clinical trials Being aware of theseissues we designed the present study aiming to solve andovercome all these disadvantages
6 BioMed Research International
3 Results
31 Efficacy of EB101 versus A12057342
+ CFAIFA in PreventingA120573 Burden Development Recently we demonstrated thatimmunization with EB101 vaccine prevents and reverses theAD neuropathology by the marked reduction of A120573 plaquesplaque-associated dystrophic neurites and astrocytosis inmice brain Now we conducted comparative immunizationstudies between EB101 and A120573
42+ FAIFA vaccines in the
prevention ofA120573deposits in themouse brainA120573plaques andvascular amyloid loadswere detected counted andmeasuredas the percent of total surface stained by monoclonal anti-A120573 antibody in the hippocampal and cortical sections Theplaque amyloid load observed in the representative photomi-crographs presented in Figure 2 shows a significant differenceacross the five experimental groups (ANOVA 119865 = 315119875 lt 0001) As a result of the first experimental phase basedon preventive treatment we observed that hippocampal loadwas significantly reduced in the EB101 immunized mice (AFigures 2(a) and 3) when compared with different vaccinecomponents (B-C Figures 2(b)-2(c) and 3) and moderatecompared with A120573
42+ CFAIFA vaccine (D Figures 2(d)
and 3) In fact the load in these treated groups (B-C) wasnot significantly different from PBS treated mice (E Figures2(e) and 3) although there is a positive correlation betweenthe presence of A120573 in treatment regime and the total A120573burden (Figures 2(b) 2(d) and 3) In the EB101 treated micea few compacted A120573 plaques were observed in retrosple-nial granular region of the cortex and at the polymorphdentate gyrus of the hippocampus (Figure 2(a)) Strikinglydifferent was the distribution of A120573 plaques observed inthe other treatment groups where numerous A120573 plaqueswere located at different cortical and hippocampal layers(Figures 2(b)ndash2(e)) This data was also supported by totalA120573 plaques density and size quantification made by imageanalysis software in hippocampal sections stained with anti-A120573 antibody (Figures 3 and 4) The data obtained in thepresent study validate our previous observations demon-strating that the mean burden A120573 plaques (Figures 2(a) and3) of Group A (25 plusmn 5 ps) were significantly different fromthe other treated groups (63 plusmn 6 ps in Figure 2(b) 65 plusmn6 ps in Figure 2(c) 56 plusmn 5 ps in Figure 2(d) 68 plusmn 6 psin Figures 2(e) and 3) This represents a decrease in themean percentage of A120573 plaque area of 301ndash382 in theEB101 immunized mice group relative to other tested groups(Figures 2(a)ndash2(j) and 4)The appearance of amyloid plaquesin the entorhinal (Figures 2(f)ndash2(j)) and piriform cortex wasalso analyzed being detected in low density in Group A(Figure 2(f))moderate density inGroups B (Figure 2(g)) andD (Figure 2(i)) and high density in the entorhinal cortexof Groups C (Figure 2(h)) and E (Figure 2(j)) Thereforethis data shows that in the preventive treatment periodthe clearance effect of the EB101 vaccine on A120573 plaques persection was about 64 (Figures 2(a) and 2(f)) compared topositive controls while in the other treatments regimes 19was detected in A120573
42+ CFAIFA (D) 10 in A120573
42liposome
without S1P (B) and 7 in liposome with S1P treated mice(C) Photomicrographs show that in all groups these plaques
were mainly localized in the external layers of the entorhinalcortex (Figures 2(f)ndash2(j)) In addition we have observedlower stained intensity of the A120573 plaques in the brainregions of EB101 treated mice which contrasts markedlywith the intensely stained A120573-immunoreactivity of theseplaques in each brain section of the other experimental micegroups
32 Efficacy of EB101 versus A12057342
+ CFAIFA in Clearing A120573Plaque Burden In a second set of experiments we comparedthe therapeutic effect of EB101 vaccine with A120573
42+ CFAIFA
in the reduction of A120573 in the brain after the plaques wereestablished in the 35-week-oldAPPPS1 transgenicmice (Fig-ures 2(k)ndash2(t))Themice were treated and analyzed using thesame protocol as in the preventive treatment To determinewhether EB101 vaccine attenuates andor reverses themassivedevelopment of 120573-amyloid plaques brain sections fromwild-type and transgenic mice of all experimental groupswere immunostained with the specific antibody recognizingA1205731ndash42 epitopesThese results show that A120573 deposits (Figures
2(k)ndash2(t)) were notably reduced in brain sections of theEB101 treated mice (A) markedly different from the A120573burden levels observed in the other treated groups (BndashE)As observed in the corresponding photomicrographs EB101treated mice showed a few compacted A120573 plaques mainlylocated at the CA1 layer of the hippocampus (Figure 2(k)) Noreduction in plaque density was observed in the other exper-imental groups (Figures 2(l)ndash2(o)) where numerous largecompacted A120573 plaques were located in almost every layer ofeach brain section The most densely populated section withA120573 plaques were those with no A120573
1ndash42 treatment as shownin Groups C (Figure 2(m)) and E (Figure 2(o)) while a slightreduction of this hallmarks was observed in mice Groups B(Figure 2(l)) and D (Figure 2(n)) The hippocampal sectionsof the therapeutic treatment with EB101 vaccine showed asimilar pattern as that observed in the preventive treatmentThemean burden of A120573 plaques in GroupA (35 plusmn 4 Figure 3)was significantly different from the other treated groups (70plusmn5 in B 74 plusmn 4 in C 65 plusmn 5 in D 80 plusmn 6 in E) The A120573 plaquearea in the EB101 immunized mice group was reduced by297ndash345 relative to the other tested groups (Figure 4) Asobserved in the preventive set of experiments this data showthat in the therapeutic treatment period the clearance effectof the EB101 vaccine on A120573 plaques per section was about59 compared to controls While in mice group treated withA12057342+ CFAIFA the clearance effect was about 20 in Group
D 13 in Group B and 8 in Group C The same pattern ofA120573-immunoreactivity was observed in the entorhinal cortexwhen comparing all experimental treatment groups (Figures2(p)ndash2(t)) Only a few A120573 plaques were observed in theexternal layer of the entorhinal and piriform cortex of micetreated with EB101 (Figure 2(p)) while numerous plaqueswere observed in the corresponding section of Groups B(Figure 2(q)) and D (Figure 2(s)) which show a moderatedensity of A120573 plaques while Groups C (Figure 2(r)) andE (Figure 2(t)) show densely distributed plaques along themedial and external layers of the entorhinal-piriform cortexAs described in the preventive treatment we also detected
BioMed Research International 7
Ect
Cx
CPu
Ect
Gr A
(EB1
01)
(a)
(c)
(b)
(f)
(h)
(g)
Preventive treatment Therapeutic treatment
CA1
PoDGMoDG
CA1
PoDGMoDG
CA1
PoDG
MoDG
DG
DG
Ect
Ect
Cx
Cx
Cx
CPu
CPu
CPuCA1
PoDG
MoDG
DG
CA1
PoDG
MoDG
DG
Ect
Cx
CPu
Ect
Cx
CPu
Cx
Cx
Cx
Cx
Cx
(k)
(l)
(p)
(r)
(q)
CA1
PoDGMoDG
DG
Cx
Ect
Cx
CPuCA1
ControlGr E
(PBS
)
(d)
(e) (j)
Control
PoDG
MoDG
DG
CA1
CA1
PoDG
MoDG
CA1
PoDG
MoDG
DG
DG
Ect
Ect
Cx
Cx
CPu
CPu
CA1
PoDG
MoDG
CA1
PoDGMoDG
DG
Ect
Cx
CPu
PoDGMoDG
DG
Cx
Cx
Cx
(n)
(o)
(s)
(t)
Cx
(i)
Gr B
(A120573
+L)
Gr D
(A120573+
FA)
A120573 A120573 A120573 A120573
A120573A120573A120573
A120573 A120573 A120573 A120573
A120573A120573A120573A120573
A120573 A120573 A120573 A120573
A120573
(m)
Gr C
(S1P
+L)
Figure 2 Comparative effect of EB101 vaccine on beta amyloid deposits Comparative photomicrographs of A120573 immunoreactivity weretaken in the hippocampus ((a)ndash(e) (k)ndash(o)) and cortical ((f)ndash(j) (p)ndash(t)) brain regions of transgenic mice with different treatments beforeA120573 plaques development preventive treatment ((a)ndash(j)) and after the A120573 plaques developed therapeutic treatment ((k)ndash(t)) Preventivetreatment transverse brain sections of 11-month-old mice show reduced number of A120573 deposits in the dentate gyrus hippocampal subregionCA1 (a) and entorhinal cortex (f) following EB101 vaccine immunization (Group A) contrasting with the numerous A120573 immunoreactiveplaques in the correspondent mouse brain sections of Groups B ((b) (g)) C ((c) (h)) D ((d) (i)) and E ((e) (j)) Immunoreactive A120573 in thebrain sections of wild-type (WT) mice is absent (inserts in panels (e) (o)) Therapeutic treatment transverse brain sections of 18-month-oldmice are shown in panels (k)ndash(t) Treatment with EB101 almost completely cleared A120573 load in the dentate gyrus and reduced notably thedensity in hippocampal subregion CA1 (k) and entorhinal cortex (p) compared to the same areas of mouse Groups B ((l) (q)) C ((m)(r)) D((n) (s)) and E ((o) (t)) as determined by number of plaques and staining intensity area Scale bar 100 120583m
8 BioMed Research International
Gr E
(PBS
)
0102030405060708090
Preventive treatmentTherapeutic treatment
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Mea
n of
120573-a
myl
oid
plaq
ues
sect
ion
Gr A
(A120573+
S1P+
L)(E
B101
)
Figure 3 Comparative analysis of A120573 plaque burden in the brainsof APPPS1 mice Mean of A120573 plaque burden in the hippocampaland cortical regions of APPswePS1ΔE9 mice in the five treatmentgroups The mean of the A120573 burden is significantly reduced (119875 lt005) in Group A (EB101) when compared with Groups BndashE duringpreventive treatment and markedly reduced in the therapeutictreatment period Data are presented asmeanplusmn standard error of themean (SEM) Densitymean of A120573 plaques in Tgmice hippocampus
05
1015202530354045
Preventive treatmentTherapeutic treatment
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Gr A
(A120573+
S1P+
L)(E
B101
)
Mea
n of
120573-a
myl
oid
plaq
ues a
rea
Figure 4 Comparative quantification of A120573 burden area in thebrain of APPPS1 mice Quantitative analysis of A120573 burden areain the hippocampal and cortical regions of APPswePS1ΔE9 micetreated with EB101 A120573liposomes S1Pliposomes A120573
42+ FAIFA
and PBS represented by the number of pixels inside the stained areaof each A120573 plaque This graphic shows that A120573 plaques of EB101-treated mice (Group A) are significantly smaller in size (119875 lt 005)than those of the other four treatment groups Data are presented asmean plusmn SEM Mean of A120573 plaques area in Tg mice hippocampus
a reduced staining intensity of the A120573 plaques in the brainregions of mouse Group A contrasting with the intenseA120573-immunoreactivity of plaques in the other experimentalmouse groups as shown in the photomicrographs of Figures2(k)ndash2(t)
33 Immune Response Effects of EB101 versus A12057342+ CFAIFA
Vaccine Astrocyte activation implicated in neuroinflam-mation amyloidogenesis and neuronal cell death in AD
was compared after treatment of the transgenic mice withEB101 and A120573
42+ CFAIFA Immunohistochemical analysis
of GFAP-reactive cells was carried out in the brains ofthe treated mice in the preventive and therapeutic studies(Figure 5) After the preventive treatment (Figures 5(a)ndash5(j))EB101 significantly reduced the density of GFAP-reactive cellclusters in the hippocampal and cortical sections as comparedwith A120573
42+ CFAIFA (D) A120573Lip without S1P (B) Lip with
S1P (C) and PBS (E) The distribution of GFAP immunore-active cells in the transverse section of the mouse brainstreated with EB101 shows a few scattered GFAP-reactiveclusters mainly at the CA1 hippocampal layers contrastingwith numerous dystrophic reactive astrocytes observed indifferent hippocampal areas of mouse brains in Groups BndashE(Figures 5(b)ndash5(e)) Similar neuroinflammation pattern wasobserved in the retrosplenial cortical layers where a fewGFAP-reactive clusters were observed in EB101 treated mice(Figure 5(f)) compared with their profuse presence in theother experimental groups (Figures 5(g)ndash5(j)) After thera-peutic treatment the GFAP immunoreactivity distributionpattern in mouse brains treated with EB101 was similar tocontrol mice (wild-type) mostly devoid of reactive GFAPclusters except a few scattered ones in the CA1 hippocampalexternal layers (Figure 5(k)) In contrast there is a moderatedensity in Group D (Figure 5(n)) and extensive density inGroup B (Figure 5(l)) Group C (Figure 5(m)) and Group Emice (Figure 5(o)) Photomicrographs of retrosplenial cortexshow similar neuroinflammation pattern where a few GFAPclusters detected in the EB101 treated mice (Figure 5(p))contrast with a more extensive density of clusters in the othergroups (Figures 5(q)ndash5(t)) No astrocytosis was observed inwild-type mice during preventive or therapeutic treatment
34 Comparative A120573 Antibody Titers and Th1Th2 CytokineLevels in Treated Mice A120573 antibody titers in the sera weredetermined by ELISA during mouse immunization(Figure 6) Preventive treatment with EB101 resulted in amarked increase of specific IgG A120573
1ndash42 antibody productionin all animals Mice in A and D groups developed andmaintained serum antibody titers between 1 2000 and1 8000 Mice in Groups B C and E showed no significantanti-A120573 antibody production Antibody production and titerin Group B were three and four times lower than those ofGroups D and A respectively After therapeutic treatment allmice treated with EB101 and A120573
42+ FAIFA produced high
levels of IgG antibodies Titer levels of Groups A and D weresignificantly increased to those detected after preventivetreatment (119875 lt 001 versus Group D 119875 lt 0001 versusGroups B C and E) although antibody level in Group D was13-fold lower than in Group A (Figure 6)The effect of EB101(Group A) and A120573
42+ CFAIFA (Group B) vaccines on Th1
andTh2 cytokines was also studied (Figure 7)Measurementsof Th1 and Th2 cytokines in sera of immunized transgenicmice indicate that significant changes occurring in proteinlevels of Th1-related inflammatory mediators followingpreventive (Figures 7(a)ndash7(c)) and therapeutic (Figures 7(d)ndash7(g)) treatment with EB101 vaccine A significant increaseof Th1 cytokine profile was observed in preventive treatment
BioMed Research International 9
Gr E
(PBS
)
(d)
(e)
(i)
(j)
(n)
(o)
(s)
(t)
Cx
GFAP
GFAP GFAP GFAP GFAP
GFAP GFAP GFAP
Cx Cx
Cx
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
GFAP
Gr A
(EB1
01)
(a)
(b)
(c)
(f)
(g)
(h)
Preventive treatment Therapeutic treatment
PoDG
MoDG
(k)
(l)
(m)
(p)
(q)
(r)DG
GFAP GFAP GFAP
GFAPGFAPGFAP GFAP
GFAP GFAP GFAP GFAP
PoDGDG
DG
CA1
Cx
Cx
Cx
Cx
Cx
Cx
PoDG
MoDG
DG
CA1
MoDGDG
CA1
PoDG
MoDG
DG
CA1
Cx
CxCx
MoDG
MoDG
PoDG
CA1CA1
Cx
Gr D
(A120573+
FA)
Gr C
(S1P
+L)
Gr B
(A120573+
L)
Figure 5 Comparative effect of EB101 vaccine on astrocytosis Comparative photomicrographs of GFAP immunoreactivity in thehippocampus ((a)ndash(e) (k)ndash(o)) and cortical ((f)ndash(j) (p)ndash(t)) brain regions at the preventive treatment phase ((a)ndash(j)) and therapeutictreatment group ((k)ndash(t)) Preventive treatment transverse brain sections of 11-month-old mice treated with EB101 show an almost completeprevention of astrocytosis in the detailed sections of the dentate gyrus (a) contrasting with numerous dystrophic reactive astrocytes typicalof an inflammatory reaction observed in different hippocampal corresponding areas of mouse brains in Group B (A120573Lip-treated mice (b))Group C (LipS1P-treated mice (c)) Group D (A120573
42+ CFAIFA-treated mice (d)) and Group E (PBS-treated mice (e)) Immunoreactive
astrocytosis in the brain sections of WT mice is absent (data not shown) Detailed sections of the parietal cortex of treated mice show agradient density of reactive astrocytes immunoreactive to GFAP indicating a neuroinflammation pattern Comparative cortical sections shownumerous astrocytosis areas inmice Groups C and E ((h) (j)) andwide reactive astrocytes clusters inmice Groups B andD ((g) (i)) while justa few of them are observed in Group A (f)Therapeutic treatment transverse brain sections of 18-month-old mice of Group A show a notablereduction of astrocytosis in the hippocampal region (k) after EB101 vaccine immunization Note the astrocytosis contrast density betweenEB101-treatedmice and the other treated groups ((l)ndash(o)) where numerous immunoreactive GFAP clusters are observed in the correspondentbrain sections A few immunoreactive GFAP clusters are observed in the parietal cortex of EB101 treated mice group while a moderate ((q)(s)) to high ((r) (t)) density of these neuropathological inflammation clusters is apparent in the other treatment groups (Groups BndashE) Forabbreviations see list Scale bar 100120583m
10 BioMed Research International
002040608
1121416
Preventive treatmentTherapeutic treatment
Ant
i-120573-a
myl
oid
IgG
antib
odie
s
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Gr A
(A120573+
S1P+
L)(E
B101
)
Figure 6 Quantification of specific anti-120573-amyloid IgG in immu-nized APPPS1 mice The presence of anti-120573-amyloid antibodiesmeasured in the sera of vaccinated and control mice groups duringthe preventive and therapeutic treatments indicates that EB101liposome vaccine was able to induce the highest production ofspecific IgG antibodies Compared to theGroupD immunizedmicethe level of antibodies observed in the Group A was also highersuggesting a more effective immune response (119875 lt 001 versusGroup D 119875 lt 0001 versus Groups B C and E) Quantification ofspecific anti-120573-amyloid IgG in immunized Tg mice
in Group D with respect to all other groups mainlyreported by the high levels of TNF-120572 (Figure 7(b)) andIFN-120574 (Figure 7(c)) cytokine secretions while due to thehigh mortality rate during the therapeutic treatment nosignificant data was obtained from this mice group Inaddition a tendency toTh1 secretion reduction was observedinGroupAwith respect to all other groups in both preventiveand therapeutic treatment The changes in Th2 protein levels(Figure 7(d)) followed the inverse trend where the highestdifferences between Group A and the other treatment groupswere observed in the IL-4 IL-5 and IL-10 cytokine secretions(119875 lt 005 versus Group D 119875 lt 001 versus Group E Figures7(e)ndash7(g))
35 Comparative Motor Coordination and Memory Perfor-mances In order to validate the locomotion integrity duringthe entire immunotherapeutic process as reported in similarprevious studies motor coordination tests have showed thatEB101-treated mice spent significantly more time on theaccelerating rotarodThismotor coordination task shows thatEB101 immunization affected positively their performance(Figure 8(a)) A120573
42liposome (B) S1P-containing liposomes
(C) and PBS (E) (119875 lt 0006) mice performed poorly incomparison to EB101 mice (119875 lt 005) or A120573
42+ CFAIFA
mice (119875 lt 002) in motor abilities and coordination Thefear conditioning-active avoidance test was used to assess thespatial learning and memory functions As expected bothEB101 and A120573
42+ CFAIFA immunized mice readily learned
the location of the platform as shown by a decrease in theescape latency over training days while the control group
mice exhibited no significant changes during 6-day trialsThe average escape latencies of EB101 (68 plusmn 2 s 761 plusmn 22 s)andA120573
42+CFAIFA (116plusmn33 s 123plusmn21 s) immunizedmice
at the end of preventive and therapeutic periods respectivelyduring the consecutive 6 days trial were significantly shorterthan the other treatment groups (A120573
42liposome 163 plusmn 31 s
175 plusmn 28 s S1P-containing liposomes 202 plusmn 19 s 248 plusmn27 s) and the control (2116 plusmn 38 s 268 plusmn 22 s) (119875 lt 001)Therewas no significant difference between EB101 andA120573
42+
CFAIFA immunized group (119875 gt 005) (Figure 8(b)) Theseresults showed that spatial learning was improved in theseimmunized mice
4 Discussion
In this study we have compared a novel active A120573 imm-unotherapy the EB101 vaccine against A120573 with the A120573
42+
CFAIFA vaccine that was first taken into clinical trialswith coadjuvant Qs-21 modification (AN1792 by Orgogozoet al [19]) This type of vaccine design was thought toinduce a strong T-cell mediated inflammatory response (IL-2 and IFN-c positive responses indicative of a Class II(CD4) Th-1 type response [33] leading to acute menin-goencephalitis in the 6 of the patients) The EB101 activeimmunotherapeutic vaccine was primarily designed to avoidundesired inflammatory side effects both in mice andhumans (without need of coadjuvant modification) whileinducing effective antibody titer rates We demonstrated thatlong-term immunization with EB101 induced A120573 antibodiesin the preventive and the therapeutic treatment periodswhich effectively reduced amyloid deposition in both corticaland hippocampal brain regions and prevented neuroinflam-mation probably by blocking toxicity in the surroundingneurons However the AN1792 vaccine was shown to inducein 6 of the patients an inflammatory reaction that mighthave led to the encephalitis type reaction observed in theclinical trial which was responsible for the immediate haltin those clinical studies It seems that AN1792 immunizationdelivered in Qs-21 and polysorbate 80 adjuvant induced aTh1 type response that likely led to autoaggressive T cells andaseptic meningoencephalitis In the present experiments wedemonstrated that a judiciously selected adjuvant produces adifferent humoral response [22] Indeed we have shown thatour novel immunotherapeutic strategy boosts specific A120573
1ndash42antibody generation and induces a Th2 noninflammatoryimmune response This boosted effect is in accordance withprevious immunotherapeutic studies [34] Taking togetherall the present findings these results suggest that EB101vaccine can achieve in a future clinical trial immunogenicresponse rates quite similar to the AN1792 without theinflammatory effects reported The immunohistochemicalanalysis obtained in the present study shows that all the EB101immunized mice resulted in an almost complete preventionof A120573 deposition in the brain with only a few compactA120573 plaques in the retrosplenial and dentate gyrus regionsAlthough there was a slightly increase in death in EB101treated mice compared to controls this significant reductionof A120573 plaques in EB101 treated mice was proven to be more
BioMed Research International 11
Gr A Gr B Gr C Gr D Gr EPreventive treatmentTherapeutic treatment
0005001000150020002500300035004000
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
(a) Th1
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
000
1000
2000
3000
4000
5000TNF-120572
(b) Th1
000500
10001500200025003000
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
IFN-120574
(c) Th1
000500
100015002000250030003500
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
Group A Group B Group C Group D Group EPreventive treatmentTherapeutic treatment
(d) Th2
000
500
1000
1500
2000
2500
3000IL-4
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(e) Th2
000500
100015002000250030003500
IL-5
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(f) Th2
3500IL-10
000500
10001500200025003000
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(g) Th2
Figure 7 Detection of Th1 and Th2 cytokine profile Measurements of the effect of EB101 and A12057342
+ CFAIFA vaccines on Th1 and Th2cytokines profile during preventive and therapeutic treatments ((a)ndash(c)) A significant increase of Th1 cytokine profile was observed inpreventive treatment in Group D with respect to all other groups while a tendency to a reduction inTh1 secretion was observed in Group Awith respect to all other groups in both preventive and therapeutic treatment ((d)ndash(g)) The changes in Th2 protein levels were observed inthe IL-4 IL-5 and IL-10 cytokine secretions of Group A (119875 lt 005 versus Group D 119875 lt 001 versus Group E)
12 BioMed Research International
Gr A
(EB1
01)
05
101520253035
Preventive treatment Therapeutic treatment
Mea
n fa
lling
late
ncy
(s)
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)(a) Graphic of motor coordination and balance
05
1015202530
Mea
n re
spon
se ti
me (
s)
Gr A
(EB1
01)
Preventive treatment Therapeutic treatment
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
(b) Graphic of associative learning and memory
Figure 8 Analysis of motor coordination and balance (a) Rotarod results of motor coordination and balance test after the preventive andtherapeutic treatment showing improved effect of EB101 and A120573
42+ CFAIFA-vaccine compared to other treated mice groups (119875 lt 005)
and (119875 lt 002) versus control mice Latency to fall on the rotarod apparatus was significantly improved in EB101 immunized mice Valuesrepresent means of all six trials of a groupday (b) Spatial learning and memory tested by fear conditioning-active avoidance test The meanresponse time of each treatment group to reach the platform is represented EB101 and A120573
42+ CFAIFA immunized mice had significantly
lower latencies than the other treatment groups (119875 lt 001) versus control mice
efficient than that obtained with A12057342+ CFAIFA immuniza-
tions A comparison of A120573 plaque reduction rates amongtreated groups surprisingly showed that other than EB101and A120573
42+ CFAIFA treated groups some amyloid deposits
were also reduced in Groups B (A1205731ndash42liposome) and C
(S1P-containing liposomes) Although the reduction rates inthese mice were markedly low the subtle lowering effectmight be due to the interaction between the immunizationelements (A120573
1ndash42 and S1P) and the mouse immunogenicsystem as reported by others [35] Based on our presentresults S1P one of the components of the adjuvant used withEB101 might be responsible for the neuroprotective effectobserved in the vulnerable neuronal areas of EB101 treatedmice The advantage in the A120573 load preventionreduction bycombining A120573
1ndash42 and S1P in a unique formulation vaccinehas been also observed in the reduced hippocampal load ofEB101 immunizedmice (Gr A) when comparedwith differentvaccine components (Gr B-C) In fact S1P appears to have ademonstrated therapeutic potential as a regenerative agent inthe nervous system since it controls migration of neuronalstem cells toward a site of spinal cord injury [36] inducecytoskeletal rearrangements to promote transmitter secretion[37] and also trigger diverse cellular effects including angio-genesis cardiac development immunity cell motility andneurite extension [38 39]Therefore it is reasonable to expectthat such a potent biologically active sphingolipid by itselfmay play an important role in the A120573 plaque reduction Inthis study the clearance efficiency rate of each treatment wasanalyzed and results suggested that EB101 (6459) appearsto be more effective than A120573
42+ CFAIFA (1920) vaccine
in both preventive and therapeutic groups The effectivenessof EB101 in older transgenic animals (18 months) showeda clearance effect of 59 suggesting that EB101 may alsohave therapeutic potential inADdiagnosticated patient trials
Similar conclusion was reached byWang and colleagues [40]who demonstrated that combination therapy in aged animalsprovides more valuable insights into a possible translationto humansOur immunologicalmeasurements demonstratedonce again that neuroinflammation develops in an A120573-dependent fashion early in the course of AD [41] and canbe alleviated or reversed by interventional A120573 vaccinationsuch as EB101 Among the possible mechanisms of A120573clearance proposed in AD immunotherapy [42] our resultssuggested that theymay overlap and probably be disease statedependent since the prevention and the therapeutic vaccinewould likely benefit from the transport of A120573 antibodies intothe CNS However some mechanisms of action such as thepreventive effect of A120573 antibodies in the A120573 aggregation [43]microglial phagocytosis induced by the Fc binding portionsbetween A120573 antibodies and microglia receptor [44] and theshift in the gradient of A120573 transport across the blood-brainbarrier due to the increased concentration of A120573 antibodiesin the vascular system resulting in an increase in effluxfrom the brain to blood [45 46] may be playing a role inboth preventive and therapeutic conditions Our findingstherefore suggest that novel treatment strategies aimed atmaintaining or increasing microglial function and restoringthe balance to the immunological system may represent anattractive therapeutic approach even at the advanced stagesof ADThe immunization results obtained with EB101 duringboth preventive and therapeutic treatment showed nomicro-hemorrhages infiltration of lymphocytes or encephalitis-like reaction in the brain suggesting that EB101 does notinduce A120573-specific Th1 reaction possibly due to the neu-roprotective effect of S1P Delivering A120573
1ndash42 in a liposomal-S1P-adjuvant resulted in an effective immunological responseincluding the induction of a Th2 T-cell response whicheffectively prevented neuroinflammation in an animal model
BioMed Research International 13
of AD as reported previously [24 47] Moreover presentresults demonstrated that EB101 did not induce astrogliosisin APPPS1 transgenic mice whereas some GFAP-positiveastrocyte clusters were aggregated in the affected brain areasof mice treated with A120573
42+ CFAIFA These results indicate
that EB101 may result in an effective treatment in AD neu-ropathology due to the absence of peripheral proinflamma-tory cytokine secretion and the absence of astroglial activa-tion in the brains of EB101 treated mice when compared withA12057342
+ CFAIFA immunotherapeutic response In a clinicalsetting however we may expect a wide differential responsein part depending upon the genomic profile of each patient[48 49] In AD behavioural studies in particular amongthe motor coordination tests the rotarod performance hasbeen reported to be impaired in some AD transgenic micemodels bearing A120573 plaques [50 51] This result was found inmutants prior to A120573 plaque formation at 3 and 6 months ofage but was not reproduced after plaques had been formedat 16 and 24 months of age [52] perhaps because of thedeclining abilities of older normal mice Here we presentthe effects of vaccine treatments in APPPS1 mice on motorcoordination a neglected aspect of the AD experimentalbehavior but justified on the basis of the deficient posturalcontrol reported in patients with Alzheimerrsquos disease [53]sometimes even during early stages [54] Our longitudinalmotor behavioral results showed no significant differencesin the ambulatory movement grip strength or coordinationaddressed by their performance in the rotating rod betweenAPPPS1 and wild-type mice for any age group studiedin contrast with other treated groups demonstrating thatlongitudinal motor assessments of AD transgenic mousemodels can provide valuable information with regard tothe effect of immunization on the progression of behavioraldeficits and overall motor activity As shown the EB101-treated group of mice exhibited longer latencies and betterbalancing skills indicating the absence of any negative effectof EB101 immunization on these mice The classical fearconditioning test is assumed to reveal the functional integrityof the hippocampus [55] representing a special memorytask to evaluate the AD models and its improvement afterimmunization periods In this APPPS1 mouse model thefirst associative learning and spatial workingmemory deficitsare shown at 5-6 months of age [56] although its impair-ment progression is relatively slow compared to most othermodels At the end of preventive immunization period EB101and A120573
42+ CFAIFA immunized mice have significantly
shorter escape latencies than the other treatment groups(A12057342liposome S1P-containing liposomes) and the controls
in the fear conditioning test Shorter escape latencies duringspatial learning indicated that deficits of spatial memorywere attenuated in EB101 and A120573
42+ CFAIFA immunized
mice Previous studies have showed that by 15 months ofage the spatial working memory is consistently impairedthroughout the rest of the life span [31 57] At the end oftherapeutic immunization period both EB101 and A120573
42+
CFAIFA immunized mice showed significantly improvedresults on the hippocampus-dependent spatial learning andmemory test when compared with other treatment groupsand the controls Taken together these observations are
in agreement with the impairment in conditioned learningin response to a tone stimulus of aged APPPS1 micealthough when immunized with EB101 during preventiveor therapeutic periods such impairments are significantlyattenuated
In summary the present results corroborate our previouspublished findings showing that EB101 immunization reducesamyloid accumulation in the APPPS1 mouse brain [2425] The strongest reduction in the preventive immunizationwas observed in the retrosplenial cortex followed by thehippocampus and entorhinal-piriform cortex Our resultsfurther support that EB101 ismore effectivewhen themice areimmunized prior to developing plaques in the hippocampaland cortical brain regions There is extensive evidence inmouse preclinical studies [58] and human clinical trials [5960] that the adverse effects of active and passive A120573 vaccinesresult in microhemorrhages and encephalitic reactions [60ndash63] yet themechanismbywhich they occur has not been fullyelucidated In the treatment sections analyzed we found nomicrohemorrhages or acute inflammatory processes in all thesurvived mice of the group indicating that EB101 therapeuticapproach presents no adverse inflammatory effects in thisAD mice models Thus our present results unequivocallydemonstrate that EB101 vaccinemay not only halt AD-relatedpathology as a preventive treatment but may also reverse it inestablished cases related to the improvement of the learningandmemory in both cases leading to a robust immunologicaltherapeutic response Present experiments also demonstratethat EB101 exerts more significant and substantial effects onthe reduction of A120573 deposition than A120573
42+CFAIFA vaccine
in A120573 mice models of AD However further exploration ofthe contribution of genetic environmental and physiologicalfactors in the development resistance to endogenous A120573-antibody is needed in order to optimize and modulate thetherapeutic strategies currently being developed
5 Conclusions
The present study carried out in APPPS1 transgenic mousemodel of AD compared the A120573 burden reduction andcellular immune responses produced by two immunizationdelivery strategies of A120573 in Freund adjuvant (described byWebster et al [10]) and in liposomeS1P matrix (describedby Schenk et al [17])The EB101 immunotherapeutic strategyboosted antibody generation A120573 plaque reduction and ananti-inflammatory response more effectively than A120573
42+
CFAIFA especially if given prior to or in the early stages ofpathology The present results also showed that vaccinationwith EB101 prevented the formation of new plaques andpresumably led to a reduction in established plaques whileinducing aTh2 immune reactionwithout encephalitogenic T-cell responses or brain inflammation related with astrogliosisHowever despite the efficacy demonstrated of this novelvaccine approach further preclinical and clinical studieson EB101 are warranted to evaluate its efficacy and themechanism for the future utility of such vaccine for ADtherapy
14 BioMed Research International
List of Abbreviations
AD Alzheimerrsquos diseaseA120573 120573-Amyloid proteinAPP Amyloid precursor proteinAPT Anterior pretectal nucleusCA1 Field CA1 hippocampusCA3 Field CA3 hippocampusCPu Caudate putamen (striatum)Cx CortexDG Dentate gyrusDN Dystrophic neuriteEct Entorhinal cortexCFA Complete Freundrsquos adjuvantGrDG Granular dentate gyrusIFA Incomplete Freundrsquos adjuvantIL InterleukinLPMR Lateral posterior thalamic nucleus (mediorostral)MoDG Molecular dentate gyrusPir Piriform cortexPoDG Polymorph dentate gyrusRSGb Retrosplenial granular cortex b regionTg TransgenicTh T helper
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Ivan Carrera Ignacio Etcheverrıa Lucıa Fernandez-NovoaValter Lombardi Ramon Cacabelos and Carmen Vigo con-ceived and designed the experiments Ivan Carrera and Igna-cio Etcheverrıa performed the experiments Ivan CarreraIgnacio Etcheverrıa and Valter Lombardi analyzed the dataIvan Carrera Ignacio Etcheverrıa Lucıa Fernandez-Novoaand Valter Lombardi contributed reagentsmaterialsanalysistools Ivan Carrera Carmen Vigo andMadepalli KrishnappaLakshmana wrote the paper
Acknowledgments
This work was supported by National Institute of Aging(NIA)NIH Grant no 1R01AG036859-01 (Madepalli Krish-nappa Lakshmana) and the financial support of the ProgramaOperativo Feder Galicia 2007ndash2013 (IN841C 201254 to PhDCarrera I)
References
[1] R Cacabelos L Fernandez-Novoa V Lombardi Y Kubotaand M Takeda ldquoMolecular genetics of Alzheimerrsquos diseaseand agingrdquoMethods and Findings in Experimental and ClinicalPharmacology vol 27 pp 1ndash573 2005
[2] P T Nelson I Alafuzoff E H Bigio et al ldquoCorrelationof alzheimer disease neuropathologic changes with cognitive
status a review of the literaturerdquo Journal of Neuropathology andExperimental Neurology vol 71 no 5 pp 362ndash381 2012
[3] JNaslundVHaroutunian RMohs et al ldquoCorrelation betweenelevated levels of amyloid 120573-peptide in the brain and cognitivedeclinerdquo Journal of the American Medical Association vol 283no 12 pp 1571ndash1577 2000
[4] A Koyama O I Okereke T Yang D Blacker D J Selkoeand F Grodstein ldquoPlasma amyloid-120573 as a predictor of dementiaand cognitive decline a systematic review and meta-analysisrdquoArchives of Neurology vol 69 no 7 pp 824ndash831 2012
[5] A M Fagan C M Roe C Xiong M A Mintun J C Morrisand D M Holtzman ldquoCerebrospinal fluid tau120573-amyloid42ratio as a prediction of cognitive decline in nondemented olderadultsrdquo Archives of Neurology vol 64 no 3 pp 343ndash349 2007
[6] S Parvathy P Davies V Haroutunian et al ldquoCorrelationbetween A120573-40- A120573-42- and A120573x-43-containing amyloidplaques and cognitive declinerdquo Archives of Neurology vol 58no 12 pp 2025ndash2032 2001
[7] E Karran M Mercken and B D Strooper ldquoThe amyloidcascade hypothesis for Alzheimerrsquos disease an appraisal for thedevelopment of therapeuticsrdquo Nature Reviews Drug Discoveryvol 10 no 9 pp 698ndash712 2011
[8] J L Jankowsky D J Fadale J Anderson et al ldquoMutantpresenilins specifically elevate the levels of the 42 residue 120573-amyloid peptide in vivo evidence for augmentation of a 42-specific 120574 secretaserdquo Human Molecular Genetics vol 13 no 2pp 159ndash170 2004
[9] L Holcomb M N Gordon E Mcgowan et al ldquoAcceleratedAlzheimer-type phenotype in transgenic mice carrying bothmutant amyloid precursor protein and presenilin 1 transgenesrdquoNature Medicine vol 4 no 1 pp 97ndash100 1998
[10] S JWebster A D Bachstetter and L J van Eldik ldquoComprehen-sive behavioral characterization of an APPPS-1 double knock-in mouse model of Alzheimerrsquos diseaserdquo Alzheimerrsquos ResearchandTherapy vol 5 no 3 article 28 2013
[11] H Xiong D Callaghan J Wodzinska et al ldquoBiochemical andbehavioral characterization of the double transgenic mousemodel (APPswePS1dE9) of Alzheimerrsquos diseaserdquo NeuroscienceBulletin vol 27 no 4 pp 221ndash232 2011
[12] M Garcia-Alloza L A Borrelli B T Hyman and B J BacskaildquoAntioxidants have a rapid and long-lasting effect on neuriticabnormalities in APPPS1 micerdquo Neurobiology of Aging vol 31no 12 pp 2058ndash2068 2010
[13] M Meyer-Luehmann M Mielke T L Spires-Jones et al ldquoAreporter of local dendritic translocation shows plaque-relatedloss of neural system function in APP-transgenic micerdquo Journalof Neuroscience vol 29 no 40 pp 12636ndash12640 2009
[14] E Machova V Rudajev H Smyckova H Koivisto H Tanilaand V Dolezal ldquoFunctional cholinergic damage develops withamyloid accumulation in young adult APPswePS1dE9 trans-genic micerdquo Neurobiology of Disease vol 38 no 1 pp 27ndash352010
[15] T Malm J Koistinaho and K Kanninen ldquoUtilization of APP-swePS1dE9 transgenic mice in research of Alzheimerrsquos diseasefocus on gene therapy and cell-based therapy applicationsrdquoInternational Journal of Alzheimerrsquos Disease vol 2011 Article ID517160 8 pages 2011
[16] J Lee and P Han ldquoAn update of animal models of alzheimerdisease with a reevaluation of plaque depositionsrdquoExperimentalNeurobiology vol 22 pp 84ndash95 2013
BioMed Research International 15
[17] D Schenk R Barbour W Dunn et al ldquoImmunization withamyloid-120573 attenuates Alzheimer disease-like pathology in thePDAPP mouserdquo Nature vol 400 no 6740 pp 173ndash177 1999
[18] H J Fu B Liu J L Frost and C A Lemere ldquoAmyloid-120573immunotherapy for Alzheimerrsquos diseaserdquoCNS and NeurologicalDisorders-Drug Targets vol 9 no 2 pp 197ndash206 2010
[19] J-M Orgogozo S Gilman J-F Dartigues et al ldquoSubacutemeningoencephalitis in a subset of patients with AD after A120573
42
immunizationrdquo Neurology vol 61 no 1 pp 46ndash54 2003[20] J A R Nicolll DWilkinson CHolmes P Steart HMarkham
andROWeller ldquoNeuropathology of humanAlzheimer diseaseafter immunization with amyloid-120573 peptide a case reportrdquoNature Medicine vol 9 no 4 pp 448ndash452 2003
[21] I Ferrer M Boada Rovira M L Sanchez Guerra M JRey and F Costa-Jussa ldquoNeuropathology and pathogenesisof encephalitis following amyloid-Neuropathology and patho-genesis of encephalitis following amyloid-beta immunizationin Alzheimerrsquos disease immunization in Alzheimerrsquos diseaserdquoBrain Pathology vol 14 no 1 pp 11ndash20 2004
[22] D H Cribbs A Ghochikyan V Vasilevko et al ldquoAdjuvant-dependent modulation ofTh1 andTh2 responses to immuniza-tion with 120573-amyloidrdquo International Immunology vol 15 no 4pp 505ndash514 2003
[23] C Holmes D Boche D Wilkinson et al ldquoLong-term effectsof A120573
42immunisation in Alzheimerrsquos disease follow-up of a
randomised placebo-controlled phase I trialrdquo The Lancet vol372 no 9634 pp 216ndash223 2008
[24] I Carrera I Etcheverrıa L Fernandez-Novoa V LombardiR Cacabelos and C Vigo ldquoVaccine development to treatAlzheimerrsquos disease neuropathology in APPPS1 transgenicmicerdquo International Journal of Alzheimerrsquos Disease vol 2012Article ID 376138 17 pages 2012
[25] I Carrera I Etcheverrıa Y Li et al ldquoImmunocytochemicalcharacterization of Alzheimer disease hallmarks in APPPS1transgenic mice treated with a new anti-amyloid-120573 vaccinerdquoBioMed Research International vol 2013 Article ID 709145 12pages 2013
[26] V Anelli R Bassi G Tettamanti P Viani and L Riboni ldquoExtra-cellular release of newly synthesized sphingosine-1-phosphateby cerebellar granule cells and astrocytesrdquo Journal of Neuro-chemistry vol 92 no 5 pp 1204ndash1215 2005
[27] S Schroeter K Khan R Barbour et al ldquoImmunotherapyreduces vascular amyloid-120573 in PDAPP micerdquo Journal of Neu-roscience vol 28 no 27 pp 6787ndash6793 2008
[28] Z Suo A Cox N Bartelli et al ldquoGRK5 deficiency leads to earlyAlzheimer-like pathology and working memory impairmentrdquoNeurobiology of Aging vol 28 no 12 pp 1873ndash1888 2007
[29] W Kamphuis C Mamber MMoeton et al ldquoGFAP isoforms inadult mouse brain with a focus on neurogenic astrocytes andreactive astrogliosis in mouse models of Alzheimer diseaserdquoPLoS ONE vol 7 no 8 Article ID e42823 2012
[30] J Lang C Vigo-Pelfrey and F Martin ldquoLiposomes composedof partially hydrogenated egg phosphatidylcholines fatty acidcomposition thermal phase behavior and oxidative stabilityrdquoChemistry and Physics of Lipids vol 53 no 1 pp 91ndash101 1990
[31] D Morgan D M Diamond P E Gottschall et al ldquoA 120573peptide vaccination prevents memory loss in an animal modelof Alzheimerrsquos diseaserdquo Nature vol 408 no 6815 pp 982ndash9852000
[32] M L Brownlow L Benner D DrsquoAgostino M N Gordon andD Morgan ldquoKetogenic diet improves motor performance but
not cognition in two mouse models of Alzheimerrsquos pathologyrdquoPLoS ONE vol 8 no 9 Article ID e75713 2013
[33] M Pride P Seubert M Grundman M Hagen J Eldridgeand R S Black ldquoProgress in the active immunotherapeu-tic approach to Alzheimerrsquos disease clinical investigationsinto AN1792-associated meningoencephalitisrdquo Neurodegenera-tive Diseases vol 5 no 3-4 pp 194ndash196 2008
[34] CWiessner K-HWiederhold A C Tissot et al ldquoThe second-generation activeA120573 immunotherapyCAD106 reduces amyloidaccumulation in APP transgenic mice while minimizing poten-tial side effectsrdquoThe Journal of Neuroscience vol 31 no 25 pp9323ndash9331 2011
[35] D Lambracht-Washington and R Rosenberg ldquoAdvances in thedevelopment of vaccines for Alzheimerrsquos diseaserdquo DiscoveryMedicine vol 15 no 84 pp 319ndash326 2013
[36] A Kimura T Ohmori R Ohkawa et al ldquoEssential roles ofsphingosine 1-phosphateS1P
1receptor axis in the migration of
neural stem cells toward a site of spinal cord injuryrdquo Stem Cellsvol 25 no 1 pp 115ndash124 2007
[37] S S Lee C H Woo J D Chang and J H Kim ldquoRoles of Racand cytosolic phospholipase A2 in the intracellular signalling inresponse to titanium particlesrdquo Cellular Signalling vol 15 no 3pp 339ndash345 2003
[38] E Masliah L Hansen A Adame et al ldquoA120573 vaccination effectson plaque pathology in the absence of encephalitis inAlzheimerdiseaserdquo Neurology vol 64 no 1 pp 129ndash131 2005
[39] A Pilorget B Annabi F Bouzeghrane J Marvaldi J Luisand R Beliveau ldquoInhibition of angiogenic properties of brainendothelial cells by platelet-derived sphingosine-1-phosphaterdquoJournal of Cerebral Blood Flow and Metabolism vol 25 no 9pp 1171ndash1182 2005
[40] A Wang P Das R C Switzer III T E Golde and J LJankowsky ldquoRobust amyloid clearance in a mouse model ofAlzheimerrsquos disease provides novel insights into themechanismof amyloid-120573 immunotherapyrdquo Journal of Neuroscience vol 31no 11 pp 4124ndash4136 2011
[41] G Krabbe A Halle V Matyash et al ldquoFunctional impairmentof microglia coincides with Beta-amyloid deposition in micewith Alzheimer-like pathologyrdquo PLoS ONE vol 8 no 4 ArticleID e60921 2013
[42] Y H Liu B Giunta H-D Zhou J Tan and Y J WangldquoImmunotherapy for Alzheimer disease-the challenge ofadverse effectsrdquo Nature Reviews Neurology vol 8 no 8 pp465ndash469 2012
[43] B Solomon R Koppel D Frankel and E Hanan-AharonldquoDisaggregation of Alzheimer120573-amyloid by site-directedmAbrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 94 no 8 pp 4109ndash4112 1997
[44] F Bard C Cannon R Barbour et al ldquoPeripherally adminis-tered antibodies against amyloid 120573-peptide enter the centralnervous system and reduce pathology in a mouse model ofAlzheimer diseaserdquo Nature Medicine vol 6 no 8 pp 916ndash9192000
[45] R B DeMattos K R Bales D J Cummins J C Dodart S MPaul and D M Holtzman ldquoPeripheral anti-A120573 antibody altersCNS and plasma A120573 clearance and decreases brain A120573 burdenin a mouse model of Alzheimerrsquos diseaserdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 98 no 15 pp 8850ndash8855 2001
[46] C A Lemere E T Spooner J LaFrancois et al ldquoEvidence forperipheral clearance of cerebral A120573 protein following chronic
16 BioMed Research International
active A120573 immunization in PSAPP micerdquo Neurobiology ofDisease vol 14 no 1 pp 10ndash18 2003
[47] T Town M Vendrame A Patel et al ldquoReduced Th1 andenhanced Th2 immunity after immunization with Alzheimerrsquos120573-amyloid
1minus42rdquo Journal of Neuroimmunology vol 132 no 1-2
pp 49ndash59 2002[48] R Cacabelos ldquoPharmacogenomics in Alzheimerrsquos diseaserdquo
Methods in Molecular Biology vol 448 pp 213ndash357 2008[49] R Cacabelos ldquoPharmacogenomics and therapeutic strategies
for dementiardquo Expert Review of Molecular Diagnostics vol 9no 6 pp 567ndash611 2009
[50] D van Dam R DrsquoHooge M Staufenbiel C van Ginneken Fvan Meir and P P de Deyn ldquoAge-dependent cognitive declinein the APP23 model precedes amyloid depositionrdquo EuropeanJournal of Neuroscience vol 17 no 2 pp 388ndash396 2003
[51] M Dumont C Strazielle M Staufenbiel and R LalondeldquoSpatial learning and exploration of environmental stimuli in24-month-old female APP23 transgenic mice with the Swedishmutationrdquo Brain Research vol 1024 no 1-2 pp 113ndash121 2004
[52] R Lalonde M Dumont M Staufenbiel C Sturchler-Pierratand C Strazielle ldquoSpatial learning exploration anxiety andmotor coordination in female APP23 transgenic mice with theSwedish mutationrdquo Brain Research vol 956 no 1 pp 36ndash442002
[53] F J Huff F Boller F Lucchelli R Querriera J Beyer and SBelle ldquoThe neurologic examination in patients with probableAlzheimerrsquos diseaserdquo Archives of Neurology vol 44 no 9 pp929ndash932 1987
[54] A F Pettersson M Engardt and L O Wahlund ldquoActivitylevel and balance in subjects with mild Alzheimerrsquos diseaserdquoDementia and Geriatric Cognitive Disorders vol 13 no 4 pp213ndash216 2002
[55] L Calandreau B Desgranges R Jaffard and A DesmedtldquoSwitching from contextual to tone fear conditioning and viceversa the key role of the glutamatergic hippocampal-lateralseptal neurotransmissionrdquo Learning amp Memory vol 17 no 9pp 440ndash443 2010
[56] F Trinchese S Liu F Battaglia SWalter PMMathews andOArancio ldquoProgressive age-related development of Alzheimer-like pathology in APPPS1 micerdquo Annals of Neurology vol 55no 6 pp 801ndash814 2004
[57] D M Wilcock A Rojiani A Rosenthal et al ldquoPassiveimmunotherapy against A120573 in aged APP-transgenic micereverses cognitive deficits and depletes parenchymal amyloiddeposits in spite of increased vascular amyloid and microhem-orrhagerdquo Journal of Neuroinflammation vol 1 article 24 2004
[58] S A Ferguson S Sarkar and L C Schmued ldquoLongitudinalbehavioral changes in the APPPS1 transgenic AlzheimerrsquosDisease modelrdquo Behavioural Brain Research vol 242 no 1 pp125ndash134 2013
[59] D Wilcock and C Colton ldquoImmunotherapy vascular pathol-ogy and microhemorrhages in transgenic micerdquo CNS andNeurological DisordersmdashDrug Targets vol 8 no 1 pp 50ndash642009
[60] M Grundman S Gilman R S Black et al ldquoAn alternativemethod for estimating efficacy of the AN1792 vaccine foralzheimer diseaserdquo Neurology vol 71 no 9 pp 697ndash698 2008
[61] D Boche E Zotova R O Weller et al ldquoConsequence of A120573immunization on the vasculature of human Alzheimerrsquos diseasebrainrdquo Brain vol 131 no 12 pp 3299ndash3310 2008
[62] M Pfeifer S Boncristiano L Bondolfi et al ldquoCerebral hemor-rhage after passive anti-a120573 immunotherapyrdquo Science vol 298no 5597 p 1379 2002
[63] M M Racke L I Boone D L Hepburn et al ldquoExacerbationof cerebral amyloid angiopathy-associatedmicrohemorrhage inamyloid precursor protein transgenic mice by immunotherapyis dependent on antibody recognition of deposited forms ofamyloid 120573rdquo Journal of Neuroscience vol 25 no 3 pp 629ndash6362005
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

2 BioMed Research International
a primary therapeutic target In this case A120573 immunothera-pies would become the most promising preclinical strategiesas they have been proven to enhance clearance of A120573 in thebrain of mice models
In order to investigate new therapeutic strategies inAD APPswePS1ΔE9 (APPPS1) mice which overexpress theSwedish mutation of APP together with PS1 deleted in exon 9that rapidly accumulates A120573 plaques at 6 months of age [8 9]have been extensively used in AD research [10] APPPS1mice also develop behavioral and learning deficits [11]plaque-associated neuritic abnormalities [12] inflammationreflected in activated microglia and astrocytes surroundingthe A120573 plaques [13] and deficits in the pre- and postsynap-tic cholinergic transmission [14] In the last decade thesetransgenic mousemodels were extensively used in preclinicalstudies of active immunization [15 16] with preaggregatedA120573 showing high anti-A120573 antibody titers in plasma dramaticreduction of cerebral A120573 burden and reduction in cognitivedecline [17 18] These results in mice did not translate wellin humans Clinical trials conducted by ElanWyeth in 2001using A120573 peptide delivered in QS-21 adjuvant (AN1792)resulted in a meningoencephalitis reaction in 6 of thetreated patients The trial then was immediately stoppedSubsequent studies suggested that these adverse events hadbeen initiated by activation of cytotoxic T cells andorautoimmune reactions [19ndash23] Recently we reported thatimmunotherapeutic treatment with EB101 vaccine consistingof A120573
1ndash42 delivered in a novel immunogen-adjuvant com-posed of liposomes-containing phosphatidylcholine phos-phatidylglycerol cholesterol and sphingosine-1-phosphate(S1P) results in a marked reduction of A120573 plaque burden anddystrophic plaque neurite density diminution of astrocytosisand attenuation of amyloidosis-induced inflammation [2425] In the present study we compare the efficacy of EB101versus the original immunization vaccine cocktail (A120573
42+
CFAIFA) [17] in the APPPS1 mouse model before and afterthe onset of A120573 pathology Moreover we also characterizedthe effect of S1P in the immunotherapeutic response of EB101in this mouse model showing that it plays a key role as aregenerative agent in the central nervous system [26] Thefindings presented were obtained by using immunocyto-chemistry techniques neuronal anatomic mapping and seraantibodycytokines detection by ELISA andmotor behavioraltests suggesting a notable effectiveness of EB101 over A120573
42+
CFAIFA vaccine in clearingA120573 plaques reducing dystrophicplaque neurites preventing inflammation in the entorhinalcortex and hippocampus in this transgenic mouse modelThese results warrant further studies which could prove thatEB101 is a promising vaccine to treat AD patients avoidingadverse effects
2 Materials and Methods
21 Animals Awell-studiedmousemodel of A120573 amyloidosisis the double-transgenic mice B6C3F1J (APPswePS1dE9)expressing a chimeric mousehuman amyloid precursor pro-tein (MoHuAPP695swe) and human presenilin 1 (PS1-ΔE9)mutants both directed to central nervous system (CNS)neurons that exhibits A120573 plaques in the hippocampus and
cortex beginning at 6 months of age (Jackson Laboratory BarHarbor ME) All experimental procedures were conformedto the guidelines established by the European Commu-nities Council Directive (86609EEC) the EU Directive201063EU and the Spanish Royal Decree 12012005 foranimal experimentation and were approved by the EthicalCommittee of the EuroEspes Biotechnology Research Centre(Permit number EE2012-344)
22 Experimental Design Two groups of experimental stud-ies preventive treatment (before amyloid deposition onsetstarting at 7 weeks of age) and therapeutic treatment (afteramyloid deposition onset at 35 weeks of age) were carriedout as described in recent reports [24 25] similar to the long-term protocol reported by Schenk and colleagues [17] andrepresented in Figure 1Mice of both sexes (balanced betweentreatment groups) were randomly assigned to each of thetwo experimental groups and they were divided into fivetreatment groups per study Preventive treatment Group Awas formed by 20 mice (14 transgenic and 6 wild-type mice14 + 6) that were immunized with a cocktail of synthetichumanA120573
42S1P-containing liposomes (EB101) Group Bwas
formed by 20 mice (14 + 6) immunized with A12057342liposome
without S1P Group C was formed by 20 mice (14 + 6)immunized with S1P-containing liposomes Group D wasformed by 20 mice (14 + 6) immunized with A120573
42Freundrsquos
adjuvant complete-incomplete (A12057342+CFAIFA) andGroup
E was formed by 16 mice (12 transgenic and 4 wild-typemice control) inoculated with PBS Therapeutic treatmentthe same treatments were administrated in Groups A B Cand D formed by 20 mice in each group while the GroupE was formed by 16 mice inoculated with PBS as controlMice were immunized with nine injections for sevenmonthsinoculating 100 120583L per injection containing a cocktail of A120573(100 120583g) and phospholipid mix (1mg) (Figure 1)
23 Experimental Procedure The methodological proce-dures to prepare the EB101 and A120573
42+ CFAIFA immuniza-
tions were strictly followed as described by Carrera et al[24] and Webster et al [10] respectively In summary theEB101 vaccine was prepared using 2mg of human A120573
1ndash42(TOCRIS bioscience Tocris Cookson Ltd) corresponding tothe human form of the predominant amyloid A120573 found in thebrains of patients with AD dissolved in 09mL water up to1mL by adding 01mL of 10xPBS then vortexed lyophilizedand stored Liposomes were prepared from 12-dioleoyl-sn-glycero-3-phosphocholine (DOPC) 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphatidylglycerol sodium salt (POPG) choles-terol (CH Northern lipids INC) and plusminus D-erythro-sphingosine-1-phosphate (S1P) (AVANTI at 0303039001molar ratio resp) as described by Lang and colleagues[30] To prepare the A120573
42+ CFAIFA vaccine A120573
1ndash42 wasemulsified 1 1 (vv) with complete Freundrsquos adjuvant (CFA)for the first immunization followed by a boost of incom-plete Freundrsquos adjuvant (IFA) monthly thereafter Once theimmunization period finished mice were kept in the cage for2 months before being euthanized for analysis Mice were 11months old at the end of preventive treatment and 18 monthsold at the end of therapeutic treatment
BioMed Research International 3
Trainingperiod
35 43 51 59 63 71 weeks7 19 2711
Arrived frombreeder
0 9 15 23 31
Therapeutic treatment injectionsand rotarod trials
37 39 47 55 67
Sacrificed(preventive treatment)
Sacrificed(therapeutic treatment)
Group E PBS
3
DOPC
POPG
CHOLS1P
Trainingperiod
Preventive treatment injectionsand rotarod trials
Group A A12057342 + S1P+ L (EB101)Group B A12057342 + LGroup C S1P+ LGroup D A12057342 + Freund ci (A120573FA)
(EB101) A12057342 + S1P+ L
A12057342
Figure 1 Biophysical characterization of the EB101 vaccine and experimental design Biophysical structure of A12057342S1P-containing liposome
(EB101) and schedule of experiments
24 Spleen Cell Preparation Cytokine ELISAs and FlowCytometric Analysis Mice splenocytes were harvested from aclamped spleen tissue as described by Carrera et al [24]TheB-cell-enriched suspensions were obtained from spleen cellsthat were depleted by selective adherence to glass Petri dishesfor 2 h at 37∘C This procedure yielded an enriched B-cellpopulation gt90 CD19+ cells with lt1 CD3+ cells and lt5CD11c+ cells as determined by flow cytometry analysis andgt95 of viable cells as determined by trypan blue exclusionCells were seeded into 96-well tissue culture plates at a densityof approximately 5000 cells per well After stimulation 10 120583Lof MTT solution was added to each well Absorbance wasmeasured on an ELISA plate reader with a test wavelengthof 570 nm The results obtained from triplicate assays wereexpressed as stimulation indexes (ratio between mean ODfrom stimulated and unstimulated cultures)
25 Measurement of Antibody Titers Serum antibody titerswere measured using methods described previously [24 31]Briefly 96-well microtiter plates were coated with 1 120583gmLof A120573
1ndash42 in well-coating buffer and incubated overnight atroom temperature (RT) The sera were added to the wellsat a starting dilution of 1100 in specimen diluent (0014Msodium phosphate pH 74 015M NaCl 06 bovine serumalbumin 005 thimerosal) Four serial dilutions (140011600 16400 and 125600) of the samples were madedirectly in the plates in twofold steps to reach a final dilutionof 1102400 After incubation the second antibody (goatanti-mouse IgG conjugated to horseradish peroxidase fromBoehringer Mannheim) was added to the wells (100 120583Lwellat a dilution of 14000 in specimen diluent) and incubated
for one hour at RT The chromogen reaction (33551015840-tetramethylbenzidine obtained from Pierce Chemicals) wasstopped by the addition of 50 120583L 05M H
2SO4 The color
intensity was then read on an ELISA plate reader (Bio-Rad680 ELISA Reader) Titers were defined as the reciprocalof the dilution of serum giving one half the maximumOD Maximal OD was generally taken from an initial 1100dilution
26 Immunohistochemistry Immunohistochemical data wasanalyzed by using methods described previously [24] Insummary parallel sections were pretreated with H
2O2in
phosphate-buffered saline at 37∘C for 15 minutes to elimi-nate endogenous peroxidase rinsed twice in 005M Trizmabuffered saline (TBS) containing 01 Tween-20 at pH74 (TBS-T) for 10 minutes each pretreated with blockingavidinbiotin kit and then incubated overnight with theprimary antibodies as described in Table 1 The sections weresuccessively rinsed in TBS-T incubated in goat IgG anti-rabbit (Dako) or goat IgG anti-mouse (Dako) dependingon the primary antibody for 1 hour rinsed in TBS-T andthen incubated for 30minutes in ABC kit system (VectastainVector) The labeling was revealed by incubating sectionswith 33-diaminobenzidine as chromogen and hydrogenperoxide as oxidant In several adjacent sections negativecontrols performed by omitting the primary secondary ortertiary antibodies showed no immunostaining Prussianblue iron staining was performed to detect hemosiderin toreveal signs of microhemorrhage although no considerableperivascular microhemorrhages were observed in the brainsections analyzed
4 BioMed Research International
Table1Summaryof
antib
odiesu
sed
Antibod
yDilu
tion
Immun
ogen
Source
Labelin
gClon
enam
eTy
peCatalog
number
Lotn
umber
Ref
B-Amyloid
11000
KLH-con
jugatedsynthetic
peptidec
orrespon
ding
aa36ndash4
2of
human
amyloidBx
-42peptide
Millipore
Amyloid-Bx
-42
polypeptide
12F4
Mou
semon
oclonal
Num
ber0
5-831
DAM1447121
[242527]
GFA
P140
0Antiglialfi
brillaryacid
protein
(mou
seIgG1isotype)
Sigm
aAldric
hAntiglialfi
brillary
acid
protein
G-A-5
Mou
semon
oclonal
Num
berG
3893
078K
4830
[924252829]
DNs
1300
Degeneratingplaque
neurites
Millipore
Degeneratingplaque
neuritesa
ndneurofi
brillarytang
les
mdashRa
bbitpo
lyclo
nal
Num
berA
B1518
PS014344
96[242527]
BioMed Research International 5
27 A120573 Load Calculation Digital images of 7 randomlyselected microscopic transverse sections per treated animalwere analyzed and the percentage of immunostained A120573deposition was determined for all the markers studied asdefined by the stereotaxic Bregma coordinates A total of 7selected sections per animal were evaluated using the NIHImage J program by defining region of interest and setting athreshold to discriminate nonspecific staining Quantitativeanalysis of amyloid burden area was also performed in thehippocampal regions and parietaltemporal cortical regionsWe used areapixels analysis software (Pixcavator 4) toquantify the average number of pixels inside each A120573 plaqueper brain section Therefore Pixcavator intensity thresholdimaging was set to mark only those areas stained with 120573-amyloid antibody (A120573 load) relative to the entire imagearea The immunostaining intensity of each image was firstaveraged per mouse and then per group and expressed inpercentage units
28 Imaging Images were visualized using a microscope(Olympus BX50) and digitized using a digital camera (DP-10Olympus) A120573 burden data was determined by quantitativeimage analysis (Pixcavator 4) and reported as plaques persection (ps) or percentage of immunolabeled area cap-tured within total sample area The microphotographs wereadjusted for brightness and contrast with Corel Photo-Paint(Corel Ottawa Canada) and plates were composed withCorel Draw
29 Sensorimotor and Cognitive Testing After the adminis-tration of the vaccine mice were subjected to sensorimotorand memory testing to measure their coordination andlearning performances during the experimental period andto verify that any treatment-related effects observed in thememory tasks could not be explained by differences in senso-rimotor abilities Prior to testing as previously described [32]the mice were adapted to the room and testing conditions
291 Rotarod The motor strength ability balance andcoordination skills of all the experimental mice in the twotreatment periods were evaluated in a rotarod apparatus(Columbus Instruments Columbus OH) operated at 10 rpmbeginning at 7 weeks of age (preventive group) and 35 weeksof age (therapeutic group) The animals were adapted to theapparatus by receiving training sessions during four weekssufficient to reach a baseline level of performance This pro-cedurewas designed to assessmotor behavior during six trialsof a groupday If themouse remained on the rod for 1minutethe test was completed and scored as 1 Each animal wastested previous to the treatment injection for three sessionswith each session separated by 15 minutes The time thateachmouse remained on the rodwas registered automaticallyand stopped when the animal fell or inverted (by climbing)from the top of the rotating barrel In this experimentone investigator evaluated the rotarod performance and wasblinded to mice treatments and the second one treated themice and scored the test to avoid a bias
292 Fear ConditioningActive Avoidance This fear-moti-vated associative avoidance test is based on electric currentwhere themouse has to learn to predict the occurrence of thisaversive event by actively moving to a different compartment(escape platform) This active avoidance task provides asimple way to assess associative learning and memory bytesting the ability of the mouse to avoid an aversive event inresponse to a stimulus cueThemeasures recorded number ofnonresponses (the mouse failing to reach the escape platformduring the trial) and response latency (latency to avoid orescape) serves as an index of learning and allows memoryto be assessed After habituation for 5min mice received 30randomized escapable footshocks at an intensity of 025mAIn each shock trial the time to reach the escape platformlocated constantly at a central quadrant from the startlocationwas recorded as latency Each trial lasted amaximumof 30 s If the mouse failed to find the platform within 30 s itwas manually guided to the platform and allowed to stay for10 s the latency was recorded 30 s Experimental results areindicated as mean response time for each treatment period
210 Onset and Survival Disease onset and progression wereconfirmed and defined by periodic mice sacrifice and neu-ropathological analysisThemortality rates of APPPS1 trans-genicmice in the course of both sets of experiments were 42for A120573
42+ CFAIFA 17 for EB101 and 12 for the vehicle
211 Statistical Analyses For statistical analyses SPSS (ver-sion 110 SPSS Inc Chicago) was used and a 119875 value lt005 was considered statistically significant The correlationaverage of A120573 plaque density burden area and antibody titerswere analyzed and compared in multiple groups by one-wayanalysis of variance (ANOVA) followed by Fisherrsquos LSD posthocmeans comparison test analysis when relevant All graphsdisplay group mean plusmn SEM
212 Limitations of the Study The ultimate goal of the ADpreclinical research is developing a therapeutic drug strategyin the ideal animal model that should recapitulate most ifnot all the neurobehavioral pathology and underlying thesame mechanisms described in human AD such as therate of amyloid plaque accumulation neurodegenerationand behavioral and cognitive alterations Although such anideal mouse model has not yet been generated we believethat APPswePS1ΔE9 transgenic mice present enough patho-logical features to answer the specific questions addressedby the present study Active A120573 immunotherapy in whichantibodies are produced by the individual upon contact withthe antigen has great potential for preventive AD therapy asshown here in animal models although some aspects suchas an undesirable immune response induced by the adjuvantlong immune response adjustment age-dependent effectsinflammation and microhemorrhages should be addressedbefore its application in clinical trials Being aware of theseissues we designed the present study aiming to solve andovercome all these disadvantages
6 BioMed Research International
3 Results
31 Efficacy of EB101 versus A12057342
+ CFAIFA in PreventingA120573 Burden Development Recently we demonstrated thatimmunization with EB101 vaccine prevents and reverses theAD neuropathology by the marked reduction of A120573 plaquesplaque-associated dystrophic neurites and astrocytosis inmice brain Now we conducted comparative immunizationstudies between EB101 and A120573
42+ FAIFA vaccines in the
prevention ofA120573deposits in themouse brainA120573plaques andvascular amyloid loadswere detected counted andmeasuredas the percent of total surface stained by monoclonal anti-A120573 antibody in the hippocampal and cortical sections Theplaque amyloid load observed in the representative photomi-crographs presented in Figure 2 shows a significant differenceacross the five experimental groups (ANOVA 119865 = 315119875 lt 0001) As a result of the first experimental phase basedon preventive treatment we observed that hippocampal loadwas significantly reduced in the EB101 immunized mice (AFigures 2(a) and 3) when compared with different vaccinecomponents (B-C Figures 2(b)-2(c) and 3) and moderatecompared with A120573
42+ CFAIFA vaccine (D Figures 2(d)
and 3) In fact the load in these treated groups (B-C) wasnot significantly different from PBS treated mice (E Figures2(e) and 3) although there is a positive correlation betweenthe presence of A120573 in treatment regime and the total A120573burden (Figures 2(b) 2(d) and 3) In the EB101 treated micea few compacted A120573 plaques were observed in retrosple-nial granular region of the cortex and at the polymorphdentate gyrus of the hippocampus (Figure 2(a)) Strikinglydifferent was the distribution of A120573 plaques observed inthe other treatment groups where numerous A120573 plaqueswere located at different cortical and hippocampal layers(Figures 2(b)ndash2(e)) This data was also supported by totalA120573 plaques density and size quantification made by imageanalysis software in hippocampal sections stained with anti-A120573 antibody (Figures 3 and 4) The data obtained in thepresent study validate our previous observations demon-strating that the mean burden A120573 plaques (Figures 2(a) and3) of Group A (25 plusmn 5 ps) were significantly different fromthe other treated groups (63 plusmn 6 ps in Figure 2(b) 65 plusmn6 ps in Figure 2(c) 56 plusmn 5 ps in Figure 2(d) 68 plusmn 6 psin Figures 2(e) and 3) This represents a decrease in themean percentage of A120573 plaque area of 301ndash382 in theEB101 immunized mice group relative to other tested groups(Figures 2(a)ndash2(j) and 4)The appearance of amyloid plaquesin the entorhinal (Figures 2(f)ndash2(j)) and piriform cortex wasalso analyzed being detected in low density in Group A(Figure 2(f))moderate density inGroups B (Figure 2(g)) andD (Figure 2(i)) and high density in the entorhinal cortexof Groups C (Figure 2(h)) and E (Figure 2(j)) Thereforethis data shows that in the preventive treatment periodthe clearance effect of the EB101 vaccine on A120573 plaques persection was about 64 (Figures 2(a) and 2(f)) compared topositive controls while in the other treatments regimes 19was detected in A120573
42+ CFAIFA (D) 10 in A120573
42liposome
without S1P (B) and 7 in liposome with S1P treated mice(C) Photomicrographs show that in all groups these plaques
were mainly localized in the external layers of the entorhinalcortex (Figures 2(f)ndash2(j)) In addition we have observedlower stained intensity of the A120573 plaques in the brainregions of EB101 treated mice which contrasts markedlywith the intensely stained A120573-immunoreactivity of theseplaques in each brain section of the other experimental micegroups
32 Efficacy of EB101 versus A12057342
+ CFAIFA in Clearing A120573Plaque Burden In a second set of experiments we comparedthe therapeutic effect of EB101 vaccine with A120573
42+ CFAIFA
in the reduction of A120573 in the brain after the plaques wereestablished in the 35-week-oldAPPPS1 transgenicmice (Fig-ures 2(k)ndash2(t))Themice were treated and analyzed using thesame protocol as in the preventive treatment To determinewhether EB101 vaccine attenuates andor reverses themassivedevelopment of 120573-amyloid plaques brain sections fromwild-type and transgenic mice of all experimental groupswere immunostained with the specific antibody recognizingA1205731ndash42 epitopesThese results show that A120573 deposits (Figures
2(k)ndash2(t)) were notably reduced in brain sections of theEB101 treated mice (A) markedly different from the A120573burden levels observed in the other treated groups (BndashE)As observed in the corresponding photomicrographs EB101treated mice showed a few compacted A120573 plaques mainlylocated at the CA1 layer of the hippocampus (Figure 2(k)) Noreduction in plaque density was observed in the other exper-imental groups (Figures 2(l)ndash2(o)) where numerous largecompacted A120573 plaques were located in almost every layer ofeach brain section The most densely populated section withA120573 plaques were those with no A120573
1ndash42 treatment as shownin Groups C (Figure 2(m)) and E (Figure 2(o)) while a slightreduction of this hallmarks was observed in mice Groups B(Figure 2(l)) and D (Figure 2(n)) The hippocampal sectionsof the therapeutic treatment with EB101 vaccine showed asimilar pattern as that observed in the preventive treatmentThemean burden of A120573 plaques in GroupA (35 plusmn 4 Figure 3)was significantly different from the other treated groups (70plusmn5 in B 74 plusmn 4 in C 65 plusmn 5 in D 80 plusmn 6 in E) The A120573 plaquearea in the EB101 immunized mice group was reduced by297ndash345 relative to the other tested groups (Figure 4) Asobserved in the preventive set of experiments this data showthat in the therapeutic treatment period the clearance effectof the EB101 vaccine on A120573 plaques per section was about59 compared to controls While in mice group treated withA12057342+ CFAIFA the clearance effect was about 20 in Group
D 13 in Group B and 8 in Group C The same pattern ofA120573-immunoreactivity was observed in the entorhinal cortexwhen comparing all experimental treatment groups (Figures2(p)ndash2(t)) Only a few A120573 plaques were observed in theexternal layer of the entorhinal and piriform cortex of micetreated with EB101 (Figure 2(p)) while numerous plaqueswere observed in the corresponding section of Groups B(Figure 2(q)) and D (Figure 2(s)) which show a moderatedensity of A120573 plaques while Groups C (Figure 2(r)) andE (Figure 2(t)) show densely distributed plaques along themedial and external layers of the entorhinal-piriform cortexAs described in the preventive treatment we also detected
BioMed Research International 7
Ect
Cx
CPu
Ect
Gr A
(EB1
01)
(a)
(c)
(b)
(f)
(h)
(g)
Preventive treatment Therapeutic treatment
CA1
PoDGMoDG
CA1
PoDGMoDG
CA1
PoDG
MoDG
DG
DG
Ect
Ect
Cx
Cx
Cx
CPu
CPu
CPuCA1
PoDG
MoDG
DG
CA1
PoDG
MoDG
DG
Ect
Cx
CPu
Ect
Cx
CPu
Cx
Cx
Cx
Cx
Cx
(k)
(l)
(p)
(r)
(q)
CA1
PoDGMoDG
DG
Cx
Ect
Cx
CPuCA1
ControlGr E
(PBS
)
(d)
(e) (j)
Control
PoDG
MoDG
DG
CA1
CA1
PoDG
MoDG
CA1
PoDG
MoDG
DG
DG
Ect
Ect
Cx
Cx
CPu
CPu
CA1
PoDG
MoDG
CA1
PoDGMoDG
DG
Ect
Cx
CPu
PoDGMoDG
DG
Cx
Cx
Cx
(n)
(o)
(s)
(t)
Cx
(i)
Gr B
(A120573
+L)
Gr D
(A120573+
FA)
A120573 A120573 A120573 A120573
A120573A120573A120573
A120573 A120573 A120573 A120573
A120573A120573A120573A120573
A120573 A120573 A120573 A120573
A120573
(m)
Gr C
(S1P
+L)
Figure 2 Comparative effect of EB101 vaccine on beta amyloid deposits Comparative photomicrographs of A120573 immunoreactivity weretaken in the hippocampus ((a)ndash(e) (k)ndash(o)) and cortical ((f)ndash(j) (p)ndash(t)) brain regions of transgenic mice with different treatments beforeA120573 plaques development preventive treatment ((a)ndash(j)) and after the A120573 plaques developed therapeutic treatment ((k)ndash(t)) Preventivetreatment transverse brain sections of 11-month-old mice show reduced number of A120573 deposits in the dentate gyrus hippocampal subregionCA1 (a) and entorhinal cortex (f) following EB101 vaccine immunization (Group A) contrasting with the numerous A120573 immunoreactiveplaques in the correspondent mouse brain sections of Groups B ((b) (g)) C ((c) (h)) D ((d) (i)) and E ((e) (j)) Immunoreactive A120573 in thebrain sections of wild-type (WT) mice is absent (inserts in panels (e) (o)) Therapeutic treatment transverse brain sections of 18-month-oldmice are shown in panels (k)ndash(t) Treatment with EB101 almost completely cleared A120573 load in the dentate gyrus and reduced notably thedensity in hippocampal subregion CA1 (k) and entorhinal cortex (p) compared to the same areas of mouse Groups B ((l) (q)) C ((m)(r)) D((n) (s)) and E ((o) (t)) as determined by number of plaques and staining intensity area Scale bar 100 120583m
8 BioMed Research International
Gr E
(PBS
)
0102030405060708090
Preventive treatmentTherapeutic treatment
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Mea
n of
120573-a
myl
oid
plaq
ues
sect
ion
Gr A
(A120573+
S1P+
L)(E
B101
)
Figure 3 Comparative analysis of A120573 plaque burden in the brainsof APPPS1 mice Mean of A120573 plaque burden in the hippocampaland cortical regions of APPswePS1ΔE9 mice in the five treatmentgroups The mean of the A120573 burden is significantly reduced (119875 lt005) in Group A (EB101) when compared with Groups BndashE duringpreventive treatment and markedly reduced in the therapeutictreatment period Data are presented asmeanplusmn standard error of themean (SEM) Densitymean of A120573 plaques in Tgmice hippocampus
05
1015202530354045
Preventive treatmentTherapeutic treatment
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Gr A
(A120573+
S1P+
L)(E
B101
)
Mea
n of
120573-a
myl
oid
plaq
ues a
rea
Figure 4 Comparative quantification of A120573 burden area in thebrain of APPPS1 mice Quantitative analysis of A120573 burden areain the hippocampal and cortical regions of APPswePS1ΔE9 micetreated with EB101 A120573liposomes S1Pliposomes A120573
42+ FAIFA
and PBS represented by the number of pixels inside the stained areaof each A120573 plaque This graphic shows that A120573 plaques of EB101-treated mice (Group A) are significantly smaller in size (119875 lt 005)than those of the other four treatment groups Data are presented asmean plusmn SEM Mean of A120573 plaques area in Tg mice hippocampus
a reduced staining intensity of the A120573 plaques in the brainregions of mouse Group A contrasting with the intenseA120573-immunoreactivity of plaques in the other experimentalmouse groups as shown in the photomicrographs of Figures2(k)ndash2(t)
33 Immune Response Effects of EB101 versus A12057342+ CFAIFA
Vaccine Astrocyte activation implicated in neuroinflam-mation amyloidogenesis and neuronal cell death in AD
was compared after treatment of the transgenic mice withEB101 and A120573
42+ CFAIFA Immunohistochemical analysis
of GFAP-reactive cells was carried out in the brains ofthe treated mice in the preventive and therapeutic studies(Figure 5) After the preventive treatment (Figures 5(a)ndash5(j))EB101 significantly reduced the density of GFAP-reactive cellclusters in the hippocampal and cortical sections as comparedwith A120573
42+ CFAIFA (D) A120573Lip without S1P (B) Lip with
S1P (C) and PBS (E) The distribution of GFAP immunore-active cells in the transverse section of the mouse brainstreated with EB101 shows a few scattered GFAP-reactiveclusters mainly at the CA1 hippocampal layers contrastingwith numerous dystrophic reactive astrocytes observed indifferent hippocampal areas of mouse brains in Groups BndashE(Figures 5(b)ndash5(e)) Similar neuroinflammation pattern wasobserved in the retrosplenial cortical layers where a fewGFAP-reactive clusters were observed in EB101 treated mice(Figure 5(f)) compared with their profuse presence in theother experimental groups (Figures 5(g)ndash5(j)) After thera-peutic treatment the GFAP immunoreactivity distributionpattern in mouse brains treated with EB101 was similar tocontrol mice (wild-type) mostly devoid of reactive GFAPclusters except a few scattered ones in the CA1 hippocampalexternal layers (Figure 5(k)) In contrast there is a moderatedensity in Group D (Figure 5(n)) and extensive density inGroup B (Figure 5(l)) Group C (Figure 5(m)) and Group Emice (Figure 5(o)) Photomicrographs of retrosplenial cortexshow similar neuroinflammation pattern where a few GFAPclusters detected in the EB101 treated mice (Figure 5(p))contrast with a more extensive density of clusters in the othergroups (Figures 5(q)ndash5(t)) No astrocytosis was observed inwild-type mice during preventive or therapeutic treatment
34 Comparative A120573 Antibody Titers and Th1Th2 CytokineLevels in Treated Mice A120573 antibody titers in the sera weredetermined by ELISA during mouse immunization(Figure 6) Preventive treatment with EB101 resulted in amarked increase of specific IgG A120573
1ndash42 antibody productionin all animals Mice in A and D groups developed andmaintained serum antibody titers between 1 2000 and1 8000 Mice in Groups B C and E showed no significantanti-A120573 antibody production Antibody production and titerin Group B were three and four times lower than those ofGroups D and A respectively After therapeutic treatment allmice treated with EB101 and A120573
42+ FAIFA produced high
levels of IgG antibodies Titer levels of Groups A and D weresignificantly increased to those detected after preventivetreatment (119875 lt 001 versus Group D 119875 lt 0001 versusGroups B C and E) although antibody level in Group D was13-fold lower than in Group A (Figure 6)The effect of EB101(Group A) and A120573
42+ CFAIFA (Group B) vaccines on Th1
andTh2 cytokines was also studied (Figure 7)Measurementsof Th1 and Th2 cytokines in sera of immunized transgenicmice indicate that significant changes occurring in proteinlevels of Th1-related inflammatory mediators followingpreventive (Figures 7(a)ndash7(c)) and therapeutic (Figures 7(d)ndash7(g)) treatment with EB101 vaccine A significant increaseof Th1 cytokine profile was observed in preventive treatment
BioMed Research International 9
Gr E
(PBS
)
(d)
(e)
(i)
(j)
(n)
(o)
(s)
(t)
Cx
GFAP
GFAP GFAP GFAP GFAP
GFAP GFAP GFAP
Cx Cx
Cx
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
GFAP
Gr A
(EB1
01)
(a)
(b)
(c)
(f)
(g)
(h)
Preventive treatment Therapeutic treatment
PoDG
MoDG
(k)
(l)
(m)
(p)
(q)
(r)DG
GFAP GFAP GFAP
GFAPGFAPGFAP GFAP
GFAP GFAP GFAP GFAP
PoDGDG
DG
CA1
Cx
Cx
Cx
Cx
Cx
Cx
PoDG
MoDG
DG
CA1
MoDGDG
CA1
PoDG
MoDG
DG
CA1
Cx
CxCx
MoDG
MoDG
PoDG
CA1CA1
Cx
Gr D
(A120573+
FA)
Gr C
(S1P
+L)
Gr B
(A120573+
L)
Figure 5 Comparative effect of EB101 vaccine on astrocytosis Comparative photomicrographs of GFAP immunoreactivity in thehippocampus ((a)ndash(e) (k)ndash(o)) and cortical ((f)ndash(j) (p)ndash(t)) brain regions at the preventive treatment phase ((a)ndash(j)) and therapeutictreatment group ((k)ndash(t)) Preventive treatment transverse brain sections of 11-month-old mice treated with EB101 show an almost completeprevention of astrocytosis in the detailed sections of the dentate gyrus (a) contrasting with numerous dystrophic reactive astrocytes typicalof an inflammatory reaction observed in different hippocampal corresponding areas of mouse brains in Group B (A120573Lip-treated mice (b))Group C (LipS1P-treated mice (c)) Group D (A120573
42+ CFAIFA-treated mice (d)) and Group E (PBS-treated mice (e)) Immunoreactive
astrocytosis in the brain sections of WT mice is absent (data not shown) Detailed sections of the parietal cortex of treated mice show agradient density of reactive astrocytes immunoreactive to GFAP indicating a neuroinflammation pattern Comparative cortical sections shownumerous astrocytosis areas inmice Groups C and E ((h) (j)) andwide reactive astrocytes clusters inmice Groups B andD ((g) (i)) while justa few of them are observed in Group A (f)Therapeutic treatment transverse brain sections of 18-month-old mice of Group A show a notablereduction of astrocytosis in the hippocampal region (k) after EB101 vaccine immunization Note the astrocytosis contrast density betweenEB101-treatedmice and the other treated groups ((l)ndash(o)) where numerous immunoreactive GFAP clusters are observed in the correspondentbrain sections A few immunoreactive GFAP clusters are observed in the parietal cortex of EB101 treated mice group while a moderate ((q)(s)) to high ((r) (t)) density of these neuropathological inflammation clusters is apparent in the other treatment groups (Groups BndashE) Forabbreviations see list Scale bar 100120583m
10 BioMed Research International
002040608
1121416
Preventive treatmentTherapeutic treatment
Ant
i-120573-a
myl
oid
IgG
antib
odie
s
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Gr A
(A120573+
S1P+
L)(E
B101
)
Figure 6 Quantification of specific anti-120573-amyloid IgG in immu-nized APPPS1 mice The presence of anti-120573-amyloid antibodiesmeasured in the sera of vaccinated and control mice groups duringthe preventive and therapeutic treatments indicates that EB101liposome vaccine was able to induce the highest production ofspecific IgG antibodies Compared to theGroupD immunizedmicethe level of antibodies observed in the Group A was also highersuggesting a more effective immune response (119875 lt 001 versusGroup D 119875 lt 0001 versus Groups B C and E) Quantification ofspecific anti-120573-amyloid IgG in immunized Tg mice
in Group D with respect to all other groups mainlyreported by the high levels of TNF-120572 (Figure 7(b)) andIFN-120574 (Figure 7(c)) cytokine secretions while due to thehigh mortality rate during the therapeutic treatment nosignificant data was obtained from this mice group Inaddition a tendency toTh1 secretion reduction was observedinGroupAwith respect to all other groups in both preventiveand therapeutic treatment The changes in Th2 protein levels(Figure 7(d)) followed the inverse trend where the highestdifferences between Group A and the other treatment groupswere observed in the IL-4 IL-5 and IL-10 cytokine secretions(119875 lt 005 versus Group D 119875 lt 001 versus Group E Figures7(e)ndash7(g))
35 Comparative Motor Coordination and Memory Perfor-mances In order to validate the locomotion integrity duringthe entire immunotherapeutic process as reported in similarprevious studies motor coordination tests have showed thatEB101-treated mice spent significantly more time on theaccelerating rotarodThismotor coordination task shows thatEB101 immunization affected positively their performance(Figure 8(a)) A120573
42liposome (B) S1P-containing liposomes
(C) and PBS (E) (119875 lt 0006) mice performed poorly incomparison to EB101 mice (119875 lt 005) or A120573
42+ CFAIFA
mice (119875 lt 002) in motor abilities and coordination Thefear conditioning-active avoidance test was used to assess thespatial learning and memory functions As expected bothEB101 and A120573
42+ CFAIFA immunized mice readily learned
the location of the platform as shown by a decrease in theescape latency over training days while the control group
mice exhibited no significant changes during 6-day trialsThe average escape latencies of EB101 (68 plusmn 2 s 761 plusmn 22 s)andA120573
42+CFAIFA (116plusmn33 s 123plusmn21 s) immunizedmice
at the end of preventive and therapeutic periods respectivelyduring the consecutive 6 days trial were significantly shorterthan the other treatment groups (A120573
42liposome 163 plusmn 31 s
175 plusmn 28 s S1P-containing liposomes 202 plusmn 19 s 248 plusmn27 s) and the control (2116 plusmn 38 s 268 plusmn 22 s) (119875 lt 001)Therewas no significant difference between EB101 andA120573
42+
CFAIFA immunized group (119875 gt 005) (Figure 8(b)) Theseresults showed that spatial learning was improved in theseimmunized mice
4 Discussion
In this study we have compared a novel active A120573 imm-unotherapy the EB101 vaccine against A120573 with the A120573
42+
CFAIFA vaccine that was first taken into clinical trialswith coadjuvant Qs-21 modification (AN1792 by Orgogozoet al [19]) This type of vaccine design was thought toinduce a strong T-cell mediated inflammatory response (IL-2 and IFN-c positive responses indicative of a Class II(CD4) Th-1 type response [33] leading to acute menin-goencephalitis in the 6 of the patients) The EB101 activeimmunotherapeutic vaccine was primarily designed to avoidundesired inflammatory side effects both in mice andhumans (without need of coadjuvant modification) whileinducing effective antibody titer rates We demonstrated thatlong-term immunization with EB101 induced A120573 antibodiesin the preventive and the therapeutic treatment periodswhich effectively reduced amyloid deposition in both corticaland hippocampal brain regions and prevented neuroinflam-mation probably by blocking toxicity in the surroundingneurons However the AN1792 vaccine was shown to inducein 6 of the patients an inflammatory reaction that mighthave led to the encephalitis type reaction observed in theclinical trial which was responsible for the immediate haltin those clinical studies It seems that AN1792 immunizationdelivered in Qs-21 and polysorbate 80 adjuvant induced aTh1 type response that likely led to autoaggressive T cells andaseptic meningoencephalitis In the present experiments wedemonstrated that a judiciously selected adjuvant produces adifferent humoral response [22] Indeed we have shown thatour novel immunotherapeutic strategy boosts specific A120573
1ndash42antibody generation and induces a Th2 noninflammatoryimmune response This boosted effect is in accordance withprevious immunotherapeutic studies [34] Taking togetherall the present findings these results suggest that EB101vaccine can achieve in a future clinical trial immunogenicresponse rates quite similar to the AN1792 without theinflammatory effects reported The immunohistochemicalanalysis obtained in the present study shows that all the EB101immunized mice resulted in an almost complete preventionof A120573 deposition in the brain with only a few compactA120573 plaques in the retrosplenial and dentate gyrus regionsAlthough there was a slightly increase in death in EB101treated mice compared to controls this significant reductionof A120573 plaques in EB101 treated mice was proven to be more
BioMed Research International 11
Gr A Gr B Gr C Gr D Gr EPreventive treatmentTherapeutic treatment
0005001000150020002500300035004000
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
(a) Th1
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
000
1000
2000
3000
4000
5000TNF-120572
(b) Th1
000500
10001500200025003000
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
IFN-120574
(c) Th1
000500
100015002000250030003500
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
Group A Group B Group C Group D Group EPreventive treatmentTherapeutic treatment
(d) Th2
000
500
1000
1500
2000
2500
3000IL-4
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(e) Th2
000500
100015002000250030003500
IL-5
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(f) Th2
3500IL-10
000500
10001500200025003000
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(g) Th2
Figure 7 Detection of Th1 and Th2 cytokine profile Measurements of the effect of EB101 and A12057342
+ CFAIFA vaccines on Th1 and Th2cytokines profile during preventive and therapeutic treatments ((a)ndash(c)) A significant increase of Th1 cytokine profile was observed inpreventive treatment in Group D with respect to all other groups while a tendency to a reduction inTh1 secretion was observed in Group Awith respect to all other groups in both preventive and therapeutic treatment ((d)ndash(g)) The changes in Th2 protein levels were observed inthe IL-4 IL-5 and IL-10 cytokine secretions of Group A (119875 lt 005 versus Group D 119875 lt 001 versus Group E)
12 BioMed Research International
Gr A
(EB1
01)
05
101520253035
Preventive treatment Therapeutic treatment
Mea
n fa
lling
late
ncy
(s)
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)(a) Graphic of motor coordination and balance
05
1015202530
Mea
n re
spon
se ti
me (
s)
Gr A
(EB1
01)
Preventive treatment Therapeutic treatment
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
(b) Graphic of associative learning and memory
Figure 8 Analysis of motor coordination and balance (a) Rotarod results of motor coordination and balance test after the preventive andtherapeutic treatment showing improved effect of EB101 and A120573
42+ CFAIFA-vaccine compared to other treated mice groups (119875 lt 005)
and (119875 lt 002) versus control mice Latency to fall on the rotarod apparatus was significantly improved in EB101 immunized mice Valuesrepresent means of all six trials of a groupday (b) Spatial learning and memory tested by fear conditioning-active avoidance test The meanresponse time of each treatment group to reach the platform is represented EB101 and A120573
42+ CFAIFA immunized mice had significantly
lower latencies than the other treatment groups (119875 lt 001) versus control mice
efficient than that obtained with A12057342+ CFAIFA immuniza-
tions A comparison of A120573 plaque reduction rates amongtreated groups surprisingly showed that other than EB101and A120573
42+ CFAIFA treated groups some amyloid deposits
were also reduced in Groups B (A1205731ndash42liposome) and C
(S1P-containing liposomes) Although the reduction rates inthese mice were markedly low the subtle lowering effectmight be due to the interaction between the immunizationelements (A120573
1ndash42 and S1P) and the mouse immunogenicsystem as reported by others [35] Based on our presentresults S1P one of the components of the adjuvant used withEB101 might be responsible for the neuroprotective effectobserved in the vulnerable neuronal areas of EB101 treatedmice The advantage in the A120573 load preventionreduction bycombining A120573
1ndash42 and S1P in a unique formulation vaccinehas been also observed in the reduced hippocampal load ofEB101 immunizedmice (Gr A) when comparedwith differentvaccine components (Gr B-C) In fact S1P appears to have ademonstrated therapeutic potential as a regenerative agent inthe nervous system since it controls migration of neuronalstem cells toward a site of spinal cord injury [36] inducecytoskeletal rearrangements to promote transmitter secretion[37] and also trigger diverse cellular effects including angio-genesis cardiac development immunity cell motility andneurite extension [38 39]Therefore it is reasonable to expectthat such a potent biologically active sphingolipid by itselfmay play an important role in the A120573 plaque reduction Inthis study the clearance efficiency rate of each treatment wasanalyzed and results suggested that EB101 (6459) appearsto be more effective than A120573
42+ CFAIFA (1920) vaccine
in both preventive and therapeutic groups The effectivenessof EB101 in older transgenic animals (18 months) showeda clearance effect of 59 suggesting that EB101 may alsohave therapeutic potential inADdiagnosticated patient trials
Similar conclusion was reached byWang and colleagues [40]who demonstrated that combination therapy in aged animalsprovides more valuable insights into a possible translationto humansOur immunologicalmeasurements demonstratedonce again that neuroinflammation develops in an A120573-dependent fashion early in the course of AD [41] and canbe alleviated or reversed by interventional A120573 vaccinationsuch as EB101 Among the possible mechanisms of A120573clearance proposed in AD immunotherapy [42] our resultssuggested that theymay overlap and probably be disease statedependent since the prevention and the therapeutic vaccinewould likely benefit from the transport of A120573 antibodies intothe CNS However some mechanisms of action such as thepreventive effect of A120573 antibodies in the A120573 aggregation [43]microglial phagocytosis induced by the Fc binding portionsbetween A120573 antibodies and microglia receptor [44] and theshift in the gradient of A120573 transport across the blood-brainbarrier due to the increased concentration of A120573 antibodiesin the vascular system resulting in an increase in effluxfrom the brain to blood [45 46] may be playing a role inboth preventive and therapeutic conditions Our findingstherefore suggest that novel treatment strategies aimed atmaintaining or increasing microglial function and restoringthe balance to the immunological system may represent anattractive therapeutic approach even at the advanced stagesof ADThe immunization results obtained with EB101 duringboth preventive and therapeutic treatment showed nomicro-hemorrhages infiltration of lymphocytes or encephalitis-like reaction in the brain suggesting that EB101 does notinduce A120573-specific Th1 reaction possibly due to the neu-roprotective effect of S1P Delivering A120573
1ndash42 in a liposomal-S1P-adjuvant resulted in an effective immunological responseincluding the induction of a Th2 T-cell response whicheffectively prevented neuroinflammation in an animal model
BioMed Research International 13
of AD as reported previously [24 47] Moreover presentresults demonstrated that EB101 did not induce astrogliosisin APPPS1 transgenic mice whereas some GFAP-positiveastrocyte clusters were aggregated in the affected brain areasof mice treated with A120573
42+ CFAIFA These results indicate
that EB101 may result in an effective treatment in AD neu-ropathology due to the absence of peripheral proinflamma-tory cytokine secretion and the absence of astroglial activa-tion in the brains of EB101 treated mice when compared withA12057342
+ CFAIFA immunotherapeutic response In a clinicalsetting however we may expect a wide differential responsein part depending upon the genomic profile of each patient[48 49] In AD behavioural studies in particular amongthe motor coordination tests the rotarod performance hasbeen reported to be impaired in some AD transgenic micemodels bearing A120573 plaques [50 51] This result was found inmutants prior to A120573 plaque formation at 3 and 6 months ofage but was not reproduced after plaques had been formedat 16 and 24 months of age [52] perhaps because of thedeclining abilities of older normal mice Here we presentthe effects of vaccine treatments in APPPS1 mice on motorcoordination a neglected aspect of the AD experimentalbehavior but justified on the basis of the deficient posturalcontrol reported in patients with Alzheimerrsquos disease [53]sometimes even during early stages [54] Our longitudinalmotor behavioral results showed no significant differencesin the ambulatory movement grip strength or coordinationaddressed by their performance in the rotating rod betweenAPPPS1 and wild-type mice for any age group studiedin contrast with other treated groups demonstrating thatlongitudinal motor assessments of AD transgenic mousemodels can provide valuable information with regard tothe effect of immunization on the progression of behavioraldeficits and overall motor activity As shown the EB101-treated group of mice exhibited longer latencies and betterbalancing skills indicating the absence of any negative effectof EB101 immunization on these mice The classical fearconditioning test is assumed to reveal the functional integrityof the hippocampus [55] representing a special memorytask to evaluate the AD models and its improvement afterimmunization periods In this APPPS1 mouse model thefirst associative learning and spatial workingmemory deficitsare shown at 5-6 months of age [56] although its impair-ment progression is relatively slow compared to most othermodels At the end of preventive immunization period EB101and A120573
42+ CFAIFA immunized mice have significantly
shorter escape latencies than the other treatment groups(A12057342liposome S1P-containing liposomes) and the controls
in the fear conditioning test Shorter escape latencies duringspatial learning indicated that deficits of spatial memorywere attenuated in EB101 and A120573
42+ CFAIFA immunized
mice Previous studies have showed that by 15 months ofage the spatial working memory is consistently impairedthroughout the rest of the life span [31 57] At the end oftherapeutic immunization period both EB101 and A120573
42+
CFAIFA immunized mice showed significantly improvedresults on the hippocampus-dependent spatial learning andmemory test when compared with other treatment groupsand the controls Taken together these observations are
in agreement with the impairment in conditioned learningin response to a tone stimulus of aged APPPS1 micealthough when immunized with EB101 during preventiveor therapeutic periods such impairments are significantlyattenuated
In summary the present results corroborate our previouspublished findings showing that EB101 immunization reducesamyloid accumulation in the APPPS1 mouse brain [2425] The strongest reduction in the preventive immunizationwas observed in the retrosplenial cortex followed by thehippocampus and entorhinal-piriform cortex Our resultsfurther support that EB101 ismore effectivewhen themice areimmunized prior to developing plaques in the hippocampaland cortical brain regions There is extensive evidence inmouse preclinical studies [58] and human clinical trials [5960] that the adverse effects of active and passive A120573 vaccinesresult in microhemorrhages and encephalitic reactions [60ndash63] yet themechanismbywhich they occur has not been fullyelucidated In the treatment sections analyzed we found nomicrohemorrhages or acute inflammatory processes in all thesurvived mice of the group indicating that EB101 therapeuticapproach presents no adverse inflammatory effects in thisAD mice models Thus our present results unequivocallydemonstrate that EB101 vaccinemay not only halt AD-relatedpathology as a preventive treatment but may also reverse it inestablished cases related to the improvement of the learningandmemory in both cases leading to a robust immunologicaltherapeutic response Present experiments also demonstratethat EB101 exerts more significant and substantial effects onthe reduction of A120573 deposition than A120573
42+CFAIFA vaccine
in A120573 mice models of AD However further exploration ofthe contribution of genetic environmental and physiologicalfactors in the development resistance to endogenous A120573-antibody is needed in order to optimize and modulate thetherapeutic strategies currently being developed
5 Conclusions
The present study carried out in APPPS1 transgenic mousemodel of AD compared the A120573 burden reduction andcellular immune responses produced by two immunizationdelivery strategies of A120573 in Freund adjuvant (described byWebster et al [10]) and in liposomeS1P matrix (describedby Schenk et al [17])The EB101 immunotherapeutic strategyboosted antibody generation A120573 plaque reduction and ananti-inflammatory response more effectively than A120573
42+
CFAIFA especially if given prior to or in the early stages ofpathology The present results also showed that vaccinationwith EB101 prevented the formation of new plaques andpresumably led to a reduction in established plaques whileinducing aTh2 immune reactionwithout encephalitogenic T-cell responses or brain inflammation related with astrogliosisHowever despite the efficacy demonstrated of this novelvaccine approach further preclinical and clinical studieson EB101 are warranted to evaluate its efficacy and themechanism for the future utility of such vaccine for ADtherapy
14 BioMed Research International
List of Abbreviations
AD Alzheimerrsquos diseaseA120573 120573-Amyloid proteinAPP Amyloid precursor proteinAPT Anterior pretectal nucleusCA1 Field CA1 hippocampusCA3 Field CA3 hippocampusCPu Caudate putamen (striatum)Cx CortexDG Dentate gyrusDN Dystrophic neuriteEct Entorhinal cortexCFA Complete Freundrsquos adjuvantGrDG Granular dentate gyrusIFA Incomplete Freundrsquos adjuvantIL InterleukinLPMR Lateral posterior thalamic nucleus (mediorostral)MoDG Molecular dentate gyrusPir Piriform cortexPoDG Polymorph dentate gyrusRSGb Retrosplenial granular cortex b regionTg TransgenicTh T helper
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Ivan Carrera Ignacio Etcheverrıa Lucıa Fernandez-NovoaValter Lombardi Ramon Cacabelos and Carmen Vigo con-ceived and designed the experiments Ivan Carrera and Igna-cio Etcheverrıa performed the experiments Ivan CarreraIgnacio Etcheverrıa and Valter Lombardi analyzed the dataIvan Carrera Ignacio Etcheverrıa Lucıa Fernandez-Novoaand Valter Lombardi contributed reagentsmaterialsanalysistools Ivan Carrera Carmen Vigo andMadepalli KrishnappaLakshmana wrote the paper
Acknowledgments
This work was supported by National Institute of Aging(NIA)NIH Grant no 1R01AG036859-01 (Madepalli Krish-nappa Lakshmana) and the financial support of the ProgramaOperativo Feder Galicia 2007ndash2013 (IN841C 201254 to PhDCarrera I)
References
[1] R Cacabelos L Fernandez-Novoa V Lombardi Y Kubotaand M Takeda ldquoMolecular genetics of Alzheimerrsquos diseaseand agingrdquoMethods and Findings in Experimental and ClinicalPharmacology vol 27 pp 1ndash573 2005
[2] P T Nelson I Alafuzoff E H Bigio et al ldquoCorrelationof alzheimer disease neuropathologic changes with cognitive
status a review of the literaturerdquo Journal of Neuropathology andExperimental Neurology vol 71 no 5 pp 362ndash381 2012
[3] JNaslundVHaroutunian RMohs et al ldquoCorrelation betweenelevated levels of amyloid 120573-peptide in the brain and cognitivedeclinerdquo Journal of the American Medical Association vol 283no 12 pp 1571ndash1577 2000
[4] A Koyama O I Okereke T Yang D Blacker D J Selkoeand F Grodstein ldquoPlasma amyloid-120573 as a predictor of dementiaand cognitive decline a systematic review and meta-analysisrdquoArchives of Neurology vol 69 no 7 pp 824ndash831 2012
[5] A M Fagan C M Roe C Xiong M A Mintun J C Morrisand D M Holtzman ldquoCerebrospinal fluid tau120573-amyloid42ratio as a prediction of cognitive decline in nondemented olderadultsrdquo Archives of Neurology vol 64 no 3 pp 343ndash349 2007
[6] S Parvathy P Davies V Haroutunian et al ldquoCorrelationbetween A120573-40- A120573-42- and A120573x-43-containing amyloidplaques and cognitive declinerdquo Archives of Neurology vol 58no 12 pp 2025ndash2032 2001
[7] E Karran M Mercken and B D Strooper ldquoThe amyloidcascade hypothesis for Alzheimerrsquos disease an appraisal for thedevelopment of therapeuticsrdquo Nature Reviews Drug Discoveryvol 10 no 9 pp 698ndash712 2011
[8] J L Jankowsky D J Fadale J Anderson et al ldquoMutantpresenilins specifically elevate the levels of the 42 residue 120573-amyloid peptide in vivo evidence for augmentation of a 42-specific 120574 secretaserdquo Human Molecular Genetics vol 13 no 2pp 159ndash170 2004
[9] L Holcomb M N Gordon E Mcgowan et al ldquoAcceleratedAlzheimer-type phenotype in transgenic mice carrying bothmutant amyloid precursor protein and presenilin 1 transgenesrdquoNature Medicine vol 4 no 1 pp 97ndash100 1998
[10] S JWebster A D Bachstetter and L J van Eldik ldquoComprehen-sive behavioral characterization of an APPPS-1 double knock-in mouse model of Alzheimerrsquos diseaserdquo Alzheimerrsquos ResearchandTherapy vol 5 no 3 article 28 2013
[11] H Xiong D Callaghan J Wodzinska et al ldquoBiochemical andbehavioral characterization of the double transgenic mousemodel (APPswePS1dE9) of Alzheimerrsquos diseaserdquo NeuroscienceBulletin vol 27 no 4 pp 221ndash232 2011
[12] M Garcia-Alloza L A Borrelli B T Hyman and B J BacskaildquoAntioxidants have a rapid and long-lasting effect on neuriticabnormalities in APPPS1 micerdquo Neurobiology of Aging vol 31no 12 pp 2058ndash2068 2010
[13] M Meyer-Luehmann M Mielke T L Spires-Jones et al ldquoAreporter of local dendritic translocation shows plaque-relatedloss of neural system function in APP-transgenic micerdquo Journalof Neuroscience vol 29 no 40 pp 12636ndash12640 2009
[14] E Machova V Rudajev H Smyckova H Koivisto H Tanilaand V Dolezal ldquoFunctional cholinergic damage develops withamyloid accumulation in young adult APPswePS1dE9 trans-genic micerdquo Neurobiology of Disease vol 38 no 1 pp 27ndash352010
[15] T Malm J Koistinaho and K Kanninen ldquoUtilization of APP-swePS1dE9 transgenic mice in research of Alzheimerrsquos diseasefocus on gene therapy and cell-based therapy applicationsrdquoInternational Journal of Alzheimerrsquos Disease vol 2011 Article ID517160 8 pages 2011
[16] J Lee and P Han ldquoAn update of animal models of alzheimerdisease with a reevaluation of plaque depositionsrdquoExperimentalNeurobiology vol 22 pp 84ndash95 2013
BioMed Research International 15
[17] D Schenk R Barbour W Dunn et al ldquoImmunization withamyloid-120573 attenuates Alzheimer disease-like pathology in thePDAPP mouserdquo Nature vol 400 no 6740 pp 173ndash177 1999
[18] H J Fu B Liu J L Frost and C A Lemere ldquoAmyloid-120573immunotherapy for Alzheimerrsquos diseaserdquoCNS and NeurologicalDisorders-Drug Targets vol 9 no 2 pp 197ndash206 2010
[19] J-M Orgogozo S Gilman J-F Dartigues et al ldquoSubacutemeningoencephalitis in a subset of patients with AD after A120573
42
immunizationrdquo Neurology vol 61 no 1 pp 46ndash54 2003[20] J A R Nicolll DWilkinson CHolmes P Steart HMarkham
andROWeller ldquoNeuropathology of humanAlzheimer diseaseafter immunization with amyloid-120573 peptide a case reportrdquoNature Medicine vol 9 no 4 pp 448ndash452 2003
[21] I Ferrer M Boada Rovira M L Sanchez Guerra M JRey and F Costa-Jussa ldquoNeuropathology and pathogenesisof encephalitis following amyloid-Neuropathology and patho-genesis of encephalitis following amyloid-beta immunizationin Alzheimerrsquos disease immunization in Alzheimerrsquos diseaserdquoBrain Pathology vol 14 no 1 pp 11ndash20 2004
[22] D H Cribbs A Ghochikyan V Vasilevko et al ldquoAdjuvant-dependent modulation ofTh1 andTh2 responses to immuniza-tion with 120573-amyloidrdquo International Immunology vol 15 no 4pp 505ndash514 2003
[23] C Holmes D Boche D Wilkinson et al ldquoLong-term effectsof A120573
42immunisation in Alzheimerrsquos disease follow-up of a
randomised placebo-controlled phase I trialrdquo The Lancet vol372 no 9634 pp 216ndash223 2008
[24] I Carrera I Etcheverrıa L Fernandez-Novoa V LombardiR Cacabelos and C Vigo ldquoVaccine development to treatAlzheimerrsquos disease neuropathology in APPPS1 transgenicmicerdquo International Journal of Alzheimerrsquos Disease vol 2012Article ID 376138 17 pages 2012
[25] I Carrera I Etcheverrıa Y Li et al ldquoImmunocytochemicalcharacterization of Alzheimer disease hallmarks in APPPS1transgenic mice treated with a new anti-amyloid-120573 vaccinerdquoBioMed Research International vol 2013 Article ID 709145 12pages 2013
[26] V Anelli R Bassi G Tettamanti P Viani and L Riboni ldquoExtra-cellular release of newly synthesized sphingosine-1-phosphateby cerebellar granule cells and astrocytesrdquo Journal of Neuro-chemistry vol 92 no 5 pp 1204ndash1215 2005
[27] S Schroeter K Khan R Barbour et al ldquoImmunotherapyreduces vascular amyloid-120573 in PDAPP micerdquo Journal of Neu-roscience vol 28 no 27 pp 6787ndash6793 2008
[28] Z Suo A Cox N Bartelli et al ldquoGRK5 deficiency leads to earlyAlzheimer-like pathology and working memory impairmentrdquoNeurobiology of Aging vol 28 no 12 pp 1873ndash1888 2007
[29] W Kamphuis C Mamber MMoeton et al ldquoGFAP isoforms inadult mouse brain with a focus on neurogenic astrocytes andreactive astrogliosis in mouse models of Alzheimer diseaserdquoPLoS ONE vol 7 no 8 Article ID e42823 2012
[30] J Lang C Vigo-Pelfrey and F Martin ldquoLiposomes composedof partially hydrogenated egg phosphatidylcholines fatty acidcomposition thermal phase behavior and oxidative stabilityrdquoChemistry and Physics of Lipids vol 53 no 1 pp 91ndash101 1990
[31] D Morgan D M Diamond P E Gottschall et al ldquoA 120573peptide vaccination prevents memory loss in an animal modelof Alzheimerrsquos diseaserdquo Nature vol 408 no 6815 pp 982ndash9852000
[32] M L Brownlow L Benner D DrsquoAgostino M N Gordon andD Morgan ldquoKetogenic diet improves motor performance but
not cognition in two mouse models of Alzheimerrsquos pathologyrdquoPLoS ONE vol 8 no 9 Article ID e75713 2013
[33] M Pride P Seubert M Grundman M Hagen J Eldridgeand R S Black ldquoProgress in the active immunotherapeu-tic approach to Alzheimerrsquos disease clinical investigationsinto AN1792-associated meningoencephalitisrdquo Neurodegenera-tive Diseases vol 5 no 3-4 pp 194ndash196 2008
[34] CWiessner K-HWiederhold A C Tissot et al ldquoThe second-generation activeA120573 immunotherapyCAD106 reduces amyloidaccumulation in APP transgenic mice while minimizing poten-tial side effectsrdquoThe Journal of Neuroscience vol 31 no 25 pp9323ndash9331 2011
[35] D Lambracht-Washington and R Rosenberg ldquoAdvances in thedevelopment of vaccines for Alzheimerrsquos diseaserdquo DiscoveryMedicine vol 15 no 84 pp 319ndash326 2013
[36] A Kimura T Ohmori R Ohkawa et al ldquoEssential roles ofsphingosine 1-phosphateS1P
1receptor axis in the migration of
neural stem cells toward a site of spinal cord injuryrdquo Stem Cellsvol 25 no 1 pp 115ndash124 2007
[37] S S Lee C H Woo J D Chang and J H Kim ldquoRoles of Racand cytosolic phospholipase A2 in the intracellular signalling inresponse to titanium particlesrdquo Cellular Signalling vol 15 no 3pp 339ndash345 2003
[38] E Masliah L Hansen A Adame et al ldquoA120573 vaccination effectson plaque pathology in the absence of encephalitis inAlzheimerdiseaserdquo Neurology vol 64 no 1 pp 129ndash131 2005
[39] A Pilorget B Annabi F Bouzeghrane J Marvaldi J Luisand R Beliveau ldquoInhibition of angiogenic properties of brainendothelial cells by platelet-derived sphingosine-1-phosphaterdquoJournal of Cerebral Blood Flow and Metabolism vol 25 no 9pp 1171ndash1182 2005
[40] A Wang P Das R C Switzer III T E Golde and J LJankowsky ldquoRobust amyloid clearance in a mouse model ofAlzheimerrsquos disease provides novel insights into themechanismof amyloid-120573 immunotherapyrdquo Journal of Neuroscience vol 31no 11 pp 4124ndash4136 2011
[41] G Krabbe A Halle V Matyash et al ldquoFunctional impairmentof microglia coincides with Beta-amyloid deposition in micewith Alzheimer-like pathologyrdquo PLoS ONE vol 8 no 4 ArticleID e60921 2013
[42] Y H Liu B Giunta H-D Zhou J Tan and Y J WangldquoImmunotherapy for Alzheimer disease-the challenge ofadverse effectsrdquo Nature Reviews Neurology vol 8 no 8 pp465ndash469 2012
[43] B Solomon R Koppel D Frankel and E Hanan-AharonldquoDisaggregation of Alzheimer120573-amyloid by site-directedmAbrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 94 no 8 pp 4109ndash4112 1997
[44] F Bard C Cannon R Barbour et al ldquoPeripherally adminis-tered antibodies against amyloid 120573-peptide enter the centralnervous system and reduce pathology in a mouse model ofAlzheimer diseaserdquo Nature Medicine vol 6 no 8 pp 916ndash9192000
[45] R B DeMattos K R Bales D J Cummins J C Dodart S MPaul and D M Holtzman ldquoPeripheral anti-A120573 antibody altersCNS and plasma A120573 clearance and decreases brain A120573 burdenin a mouse model of Alzheimerrsquos diseaserdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 98 no 15 pp 8850ndash8855 2001
[46] C A Lemere E T Spooner J LaFrancois et al ldquoEvidence forperipheral clearance of cerebral A120573 protein following chronic
16 BioMed Research International
active A120573 immunization in PSAPP micerdquo Neurobiology ofDisease vol 14 no 1 pp 10ndash18 2003
[47] T Town M Vendrame A Patel et al ldquoReduced Th1 andenhanced Th2 immunity after immunization with Alzheimerrsquos120573-amyloid
1minus42rdquo Journal of Neuroimmunology vol 132 no 1-2
pp 49ndash59 2002[48] R Cacabelos ldquoPharmacogenomics in Alzheimerrsquos diseaserdquo
Methods in Molecular Biology vol 448 pp 213ndash357 2008[49] R Cacabelos ldquoPharmacogenomics and therapeutic strategies
for dementiardquo Expert Review of Molecular Diagnostics vol 9no 6 pp 567ndash611 2009
[50] D van Dam R DrsquoHooge M Staufenbiel C van Ginneken Fvan Meir and P P de Deyn ldquoAge-dependent cognitive declinein the APP23 model precedes amyloid depositionrdquo EuropeanJournal of Neuroscience vol 17 no 2 pp 388ndash396 2003
[51] M Dumont C Strazielle M Staufenbiel and R LalondeldquoSpatial learning and exploration of environmental stimuli in24-month-old female APP23 transgenic mice with the Swedishmutationrdquo Brain Research vol 1024 no 1-2 pp 113ndash121 2004
[52] R Lalonde M Dumont M Staufenbiel C Sturchler-Pierratand C Strazielle ldquoSpatial learning exploration anxiety andmotor coordination in female APP23 transgenic mice with theSwedish mutationrdquo Brain Research vol 956 no 1 pp 36ndash442002
[53] F J Huff F Boller F Lucchelli R Querriera J Beyer and SBelle ldquoThe neurologic examination in patients with probableAlzheimerrsquos diseaserdquo Archives of Neurology vol 44 no 9 pp929ndash932 1987
[54] A F Pettersson M Engardt and L O Wahlund ldquoActivitylevel and balance in subjects with mild Alzheimerrsquos diseaserdquoDementia and Geriatric Cognitive Disorders vol 13 no 4 pp213ndash216 2002
[55] L Calandreau B Desgranges R Jaffard and A DesmedtldquoSwitching from contextual to tone fear conditioning and viceversa the key role of the glutamatergic hippocampal-lateralseptal neurotransmissionrdquo Learning amp Memory vol 17 no 9pp 440ndash443 2010
[56] F Trinchese S Liu F Battaglia SWalter PMMathews andOArancio ldquoProgressive age-related development of Alzheimer-like pathology in APPPS1 micerdquo Annals of Neurology vol 55no 6 pp 801ndash814 2004
[57] D M Wilcock A Rojiani A Rosenthal et al ldquoPassiveimmunotherapy against A120573 in aged APP-transgenic micereverses cognitive deficits and depletes parenchymal amyloiddeposits in spite of increased vascular amyloid and microhem-orrhagerdquo Journal of Neuroinflammation vol 1 article 24 2004
[58] S A Ferguson S Sarkar and L C Schmued ldquoLongitudinalbehavioral changes in the APPPS1 transgenic AlzheimerrsquosDisease modelrdquo Behavioural Brain Research vol 242 no 1 pp125ndash134 2013
[59] D Wilcock and C Colton ldquoImmunotherapy vascular pathol-ogy and microhemorrhages in transgenic micerdquo CNS andNeurological DisordersmdashDrug Targets vol 8 no 1 pp 50ndash642009
[60] M Grundman S Gilman R S Black et al ldquoAn alternativemethod for estimating efficacy of the AN1792 vaccine foralzheimer diseaserdquo Neurology vol 71 no 9 pp 697ndash698 2008
[61] D Boche E Zotova R O Weller et al ldquoConsequence of A120573immunization on the vasculature of human Alzheimerrsquos diseasebrainrdquo Brain vol 131 no 12 pp 3299ndash3310 2008
[62] M Pfeifer S Boncristiano L Bondolfi et al ldquoCerebral hemor-rhage after passive anti-a120573 immunotherapyrdquo Science vol 298no 5597 p 1379 2002
[63] M M Racke L I Boone D L Hepburn et al ldquoExacerbationof cerebral amyloid angiopathy-associatedmicrohemorrhage inamyloid precursor protein transgenic mice by immunotherapyis dependent on antibody recognition of deposited forms ofamyloid 120573rdquo Journal of Neuroscience vol 25 no 3 pp 629ndash6362005
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

BioMed Research International 3
Trainingperiod
35 43 51 59 63 71 weeks7 19 2711
Arrived frombreeder
0 9 15 23 31
Therapeutic treatment injectionsand rotarod trials
37 39 47 55 67
Sacrificed(preventive treatment)
Sacrificed(therapeutic treatment)
Group E PBS
3
DOPC
POPG
CHOLS1P
Trainingperiod
Preventive treatment injectionsand rotarod trials
Group A A12057342 + S1P+ L (EB101)Group B A12057342 + LGroup C S1P+ LGroup D A12057342 + Freund ci (A120573FA)
(EB101) A12057342 + S1P+ L
A12057342
Figure 1 Biophysical characterization of the EB101 vaccine and experimental design Biophysical structure of A12057342S1P-containing liposome
(EB101) and schedule of experiments
24 Spleen Cell Preparation Cytokine ELISAs and FlowCytometric Analysis Mice splenocytes were harvested from aclamped spleen tissue as described by Carrera et al [24]TheB-cell-enriched suspensions were obtained from spleen cellsthat were depleted by selective adherence to glass Petri dishesfor 2 h at 37∘C This procedure yielded an enriched B-cellpopulation gt90 CD19+ cells with lt1 CD3+ cells and lt5CD11c+ cells as determined by flow cytometry analysis andgt95 of viable cells as determined by trypan blue exclusionCells were seeded into 96-well tissue culture plates at a densityof approximately 5000 cells per well After stimulation 10 120583Lof MTT solution was added to each well Absorbance wasmeasured on an ELISA plate reader with a test wavelengthof 570 nm The results obtained from triplicate assays wereexpressed as stimulation indexes (ratio between mean ODfrom stimulated and unstimulated cultures)
25 Measurement of Antibody Titers Serum antibody titerswere measured using methods described previously [24 31]Briefly 96-well microtiter plates were coated with 1 120583gmLof A120573
1ndash42 in well-coating buffer and incubated overnight atroom temperature (RT) The sera were added to the wellsat a starting dilution of 1100 in specimen diluent (0014Msodium phosphate pH 74 015M NaCl 06 bovine serumalbumin 005 thimerosal) Four serial dilutions (140011600 16400 and 125600) of the samples were madedirectly in the plates in twofold steps to reach a final dilutionof 1102400 After incubation the second antibody (goatanti-mouse IgG conjugated to horseradish peroxidase fromBoehringer Mannheim) was added to the wells (100 120583Lwellat a dilution of 14000 in specimen diluent) and incubated
for one hour at RT The chromogen reaction (33551015840-tetramethylbenzidine obtained from Pierce Chemicals) wasstopped by the addition of 50 120583L 05M H
2SO4 The color
intensity was then read on an ELISA plate reader (Bio-Rad680 ELISA Reader) Titers were defined as the reciprocalof the dilution of serum giving one half the maximumOD Maximal OD was generally taken from an initial 1100dilution
26 Immunohistochemistry Immunohistochemical data wasanalyzed by using methods described previously [24] Insummary parallel sections were pretreated with H
2O2in
phosphate-buffered saline at 37∘C for 15 minutes to elimi-nate endogenous peroxidase rinsed twice in 005M Trizmabuffered saline (TBS) containing 01 Tween-20 at pH74 (TBS-T) for 10 minutes each pretreated with blockingavidinbiotin kit and then incubated overnight with theprimary antibodies as described in Table 1 The sections weresuccessively rinsed in TBS-T incubated in goat IgG anti-rabbit (Dako) or goat IgG anti-mouse (Dako) dependingon the primary antibody for 1 hour rinsed in TBS-T andthen incubated for 30minutes in ABC kit system (VectastainVector) The labeling was revealed by incubating sectionswith 33-diaminobenzidine as chromogen and hydrogenperoxide as oxidant In several adjacent sections negativecontrols performed by omitting the primary secondary ortertiary antibodies showed no immunostaining Prussianblue iron staining was performed to detect hemosiderin toreveal signs of microhemorrhage although no considerableperivascular microhemorrhages were observed in the brainsections analyzed
4 BioMed Research International
Table1Summaryof
antib
odiesu
sed
Antibod
yDilu
tion
Immun
ogen
Source
Labelin
gClon
enam
eTy
peCatalog
number
Lotn
umber
Ref
B-Amyloid
11000
KLH-con
jugatedsynthetic
peptidec
orrespon
ding
aa36ndash4
2of
human
amyloidBx
-42peptide
Millipore
Amyloid-Bx
-42
polypeptide
12F4
Mou
semon
oclonal
Num
ber0
5-831
DAM1447121
[242527]
GFA
P140
0Antiglialfi
brillaryacid
protein
(mou
seIgG1isotype)
Sigm
aAldric
hAntiglialfi
brillary
acid
protein
G-A-5
Mou
semon
oclonal
Num
berG
3893
078K
4830
[924252829]
DNs
1300
Degeneratingplaque
neurites
Millipore
Degeneratingplaque
neuritesa
ndneurofi
brillarytang
les
mdashRa
bbitpo
lyclo
nal
Num
berA
B1518
PS014344
96[242527]
BioMed Research International 5
27 A120573 Load Calculation Digital images of 7 randomlyselected microscopic transverse sections per treated animalwere analyzed and the percentage of immunostained A120573deposition was determined for all the markers studied asdefined by the stereotaxic Bregma coordinates A total of 7selected sections per animal were evaluated using the NIHImage J program by defining region of interest and setting athreshold to discriminate nonspecific staining Quantitativeanalysis of amyloid burden area was also performed in thehippocampal regions and parietaltemporal cortical regionsWe used areapixels analysis software (Pixcavator 4) toquantify the average number of pixels inside each A120573 plaqueper brain section Therefore Pixcavator intensity thresholdimaging was set to mark only those areas stained with 120573-amyloid antibody (A120573 load) relative to the entire imagearea The immunostaining intensity of each image was firstaveraged per mouse and then per group and expressed inpercentage units
28 Imaging Images were visualized using a microscope(Olympus BX50) and digitized using a digital camera (DP-10Olympus) A120573 burden data was determined by quantitativeimage analysis (Pixcavator 4) and reported as plaques persection (ps) or percentage of immunolabeled area cap-tured within total sample area The microphotographs wereadjusted for brightness and contrast with Corel Photo-Paint(Corel Ottawa Canada) and plates were composed withCorel Draw
29 Sensorimotor and Cognitive Testing After the adminis-tration of the vaccine mice were subjected to sensorimotorand memory testing to measure their coordination andlearning performances during the experimental period andto verify that any treatment-related effects observed in thememory tasks could not be explained by differences in senso-rimotor abilities Prior to testing as previously described [32]the mice were adapted to the room and testing conditions
291 Rotarod The motor strength ability balance andcoordination skills of all the experimental mice in the twotreatment periods were evaluated in a rotarod apparatus(Columbus Instruments Columbus OH) operated at 10 rpmbeginning at 7 weeks of age (preventive group) and 35 weeksof age (therapeutic group) The animals were adapted to theapparatus by receiving training sessions during four weekssufficient to reach a baseline level of performance This pro-cedurewas designed to assessmotor behavior during six trialsof a groupday If themouse remained on the rod for 1minutethe test was completed and scored as 1 Each animal wastested previous to the treatment injection for three sessionswith each session separated by 15 minutes The time thateachmouse remained on the rodwas registered automaticallyand stopped when the animal fell or inverted (by climbing)from the top of the rotating barrel In this experimentone investigator evaluated the rotarod performance and wasblinded to mice treatments and the second one treated themice and scored the test to avoid a bias
292 Fear ConditioningActive Avoidance This fear-moti-vated associative avoidance test is based on electric currentwhere themouse has to learn to predict the occurrence of thisaversive event by actively moving to a different compartment(escape platform) This active avoidance task provides asimple way to assess associative learning and memory bytesting the ability of the mouse to avoid an aversive event inresponse to a stimulus cueThemeasures recorded number ofnonresponses (the mouse failing to reach the escape platformduring the trial) and response latency (latency to avoid orescape) serves as an index of learning and allows memoryto be assessed After habituation for 5min mice received 30randomized escapable footshocks at an intensity of 025mAIn each shock trial the time to reach the escape platformlocated constantly at a central quadrant from the startlocationwas recorded as latency Each trial lasted amaximumof 30 s If the mouse failed to find the platform within 30 s itwas manually guided to the platform and allowed to stay for10 s the latency was recorded 30 s Experimental results areindicated as mean response time for each treatment period
210 Onset and Survival Disease onset and progression wereconfirmed and defined by periodic mice sacrifice and neu-ropathological analysisThemortality rates of APPPS1 trans-genicmice in the course of both sets of experiments were 42for A120573
42+ CFAIFA 17 for EB101 and 12 for the vehicle
211 Statistical Analyses For statistical analyses SPSS (ver-sion 110 SPSS Inc Chicago) was used and a 119875 value lt005 was considered statistically significant The correlationaverage of A120573 plaque density burden area and antibody titerswere analyzed and compared in multiple groups by one-wayanalysis of variance (ANOVA) followed by Fisherrsquos LSD posthocmeans comparison test analysis when relevant All graphsdisplay group mean plusmn SEM
212 Limitations of the Study The ultimate goal of the ADpreclinical research is developing a therapeutic drug strategyin the ideal animal model that should recapitulate most ifnot all the neurobehavioral pathology and underlying thesame mechanisms described in human AD such as therate of amyloid plaque accumulation neurodegenerationand behavioral and cognitive alterations Although such anideal mouse model has not yet been generated we believethat APPswePS1ΔE9 transgenic mice present enough patho-logical features to answer the specific questions addressedby the present study Active A120573 immunotherapy in whichantibodies are produced by the individual upon contact withthe antigen has great potential for preventive AD therapy asshown here in animal models although some aspects suchas an undesirable immune response induced by the adjuvantlong immune response adjustment age-dependent effectsinflammation and microhemorrhages should be addressedbefore its application in clinical trials Being aware of theseissues we designed the present study aiming to solve andovercome all these disadvantages
6 BioMed Research International
3 Results
31 Efficacy of EB101 versus A12057342
+ CFAIFA in PreventingA120573 Burden Development Recently we demonstrated thatimmunization with EB101 vaccine prevents and reverses theAD neuropathology by the marked reduction of A120573 plaquesplaque-associated dystrophic neurites and astrocytosis inmice brain Now we conducted comparative immunizationstudies between EB101 and A120573
42+ FAIFA vaccines in the
prevention ofA120573deposits in themouse brainA120573plaques andvascular amyloid loadswere detected counted andmeasuredas the percent of total surface stained by monoclonal anti-A120573 antibody in the hippocampal and cortical sections Theplaque amyloid load observed in the representative photomi-crographs presented in Figure 2 shows a significant differenceacross the five experimental groups (ANOVA 119865 = 315119875 lt 0001) As a result of the first experimental phase basedon preventive treatment we observed that hippocampal loadwas significantly reduced in the EB101 immunized mice (AFigures 2(a) and 3) when compared with different vaccinecomponents (B-C Figures 2(b)-2(c) and 3) and moderatecompared with A120573
42+ CFAIFA vaccine (D Figures 2(d)
and 3) In fact the load in these treated groups (B-C) wasnot significantly different from PBS treated mice (E Figures2(e) and 3) although there is a positive correlation betweenthe presence of A120573 in treatment regime and the total A120573burden (Figures 2(b) 2(d) and 3) In the EB101 treated micea few compacted A120573 plaques were observed in retrosple-nial granular region of the cortex and at the polymorphdentate gyrus of the hippocampus (Figure 2(a)) Strikinglydifferent was the distribution of A120573 plaques observed inthe other treatment groups where numerous A120573 plaqueswere located at different cortical and hippocampal layers(Figures 2(b)ndash2(e)) This data was also supported by totalA120573 plaques density and size quantification made by imageanalysis software in hippocampal sections stained with anti-A120573 antibody (Figures 3 and 4) The data obtained in thepresent study validate our previous observations demon-strating that the mean burden A120573 plaques (Figures 2(a) and3) of Group A (25 plusmn 5 ps) were significantly different fromthe other treated groups (63 plusmn 6 ps in Figure 2(b) 65 plusmn6 ps in Figure 2(c) 56 plusmn 5 ps in Figure 2(d) 68 plusmn 6 psin Figures 2(e) and 3) This represents a decrease in themean percentage of A120573 plaque area of 301ndash382 in theEB101 immunized mice group relative to other tested groups(Figures 2(a)ndash2(j) and 4)The appearance of amyloid plaquesin the entorhinal (Figures 2(f)ndash2(j)) and piriform cortex wasalso analyzed being detected in low density in Group A(Figure 2(f))moderate density inGroups B (Figure 2(g)) andD (Figure 2(i)) and high density in the entorhinal cortexof Groups C (Figure 2(h)) and E (Figure 2(j)) Thereforethis data shows that in the preventive treatment periodthe clearance effect of the EB101 vaccine on A120573 plaques persection was about 64 (Figures 2(a) and 2(f)) compared topositive controls while in the other treatments regimes 19was detected in A120573
42+ CFAIFA (D) 10 in A120573
42liposome
without S1P (B) and 7 in liposome with S1P treated mice(C) Photomicrographs show that in all groups these plaques
were mainly localized in the external layers of the entorhinalcortex (Figures 2(f)ndash2(j)) In addition we have observedlower stained intensity of the A120573 plaques in the brainregions of EB101 treated mice which contrasts markedlywith the intensely stained A120573-immunoreactivity of theseplaques in each brain section of the other experimental micegroups
32 Efficacy of EB101 versus A12057342
+ CFAIFA in Clearing A120573Plaque Burden In a second set of experiments we comparedthe therapeutic effect of EB101 vaccine with A120573
42+ CFAIFA
in the reduction of A120573 in the brain after the plaques wereestablished in the 35-week-oldAPPPS1 transgenicmice (Fig-ures 2(k)ndash2(t))Themice were treated and analyzed using thesame protocol as in the preventive treatment To determinewhether EB101 vaccine attenuates andor reverses themassivedevelopment of 120573-amyloid plaques brain sections fromwild-type and transgenic mice of all experimental groupswere immunostained with the specific antibody recognizingA1205731ndash42 epitopesThese results show that A120573 deposits (Figures
2(k)ndash2(t)) were notably reduced in brain sections of theEB101 treated mice (A) markedly different from the A120573burden levels observed in the other treated groups (BndashE)As observed in the corresponding photomicrographs EB101treated mice showed a few compacted A120573 plaques mainlylocated at the CA1 layer of the hippocampus (Figure 2(k)) Noreduction in plaque density was observed in the other exper-imental groups (Figures 2(l)ndash2(o)) where numerous largecompacted A120573 plaques were located in almost every layer ofeach brain section The most densely populated section withA120573 plaques were those with no A120573
1ndash42 treatment as shownin Groups C (Figure 2(m)) and E (Figure 2(o)) while a slightreduction of this hallmarks was observed in mice Groups B(Figure 2(l)) and D (Figure 2(n)) The hippocampal sectionsof the therapeutic treatment with EB101 vaccine showed asimilar pattern as that observed in the preventive treatmentThemean burden of A120573 plaques in GroupA (35 plusmn 4 Figure 3)was significantly different from the other treated groups (70plusmn5 in B 74 plusmn 4 in C 65 plusmn 5 in D 80 plusmn 6 in E) The A120573 plaquearea in the EB101 immunized mice group was reduced by297ndash345 relative to the other tested groups (Figure 4) Asobserved in the preventive set of experiments this data showthat in the therapeutic treatment period the clearance effectof the EB101 vaccine on A120573 plaques per section was about59 compared to controls While in mice group treated withA12057342+ CFAIFA the clearance effect was about 20 in Group
D 13 in Group B and 8 in Group C The same pattern ofA120573-immunoreactivity was observed in the entorhinal cortexwhen comparing all experimental treatment groups (Figures2(p)ndash2(t)) Only a few A120573 plaques were observed in theexternal layer of the entorhinal and piriform cortex of micetreated with EB101 (Figure 2(p)) while numerous plaqueswere observed in the corresponding section of Groups B(Figure 2(q)) and D (Figure 2(s)) which show a moderatedensity of A120573 plaques while Groups C (Figure 2(r)) andE (Figure 2(t)) show densely distributed plaques along themedial and external layers of the entorhinal-piriform cortexAs described in the preventive treatment we also detected
BioMed Research International 7
Ect
Cx
CPu
Ect
Gr A
(EB1
01)
(a)
(c)
(b)
(f)
(h)
(g)
Preventive treatment Therapeutic treatment
CA1
PoDGMoDG
CA1
PoDGMoDG
CA1
PoDG
MoDG
DG
DG
Ect
Ect
Cx
Cx
Cx
CPu
CPu
CPuCA1
PoDG
MoDG
DG
CA1
PoDG
MoDG
DG
Ect
Cx
CPu
Ect
Cx
CPu
Cx
Cx
Cx
Cx
Cx
(k)
(l)
(p)
(r)
(q)
CA1
PoDGMoDG
DG
Cx
Ect
Cx
CPuCA1
ControlGr E
(PBS
)
(d)
(e) (j)
Control
PoDG
MoDG
DG
CA1
CA1
PoDG
MoDG
CA1
PoDG
MoDG
DG
DG
Ect
Ect
Cx
Cx
CPu
CPu
CA1
PoDG
MoDG
CA1
PoDGMoDG
DG
Ect
Cx
CPu
PoDGMoDG
DG
Cx
Cx
Cx
(n)
(o)
(s)
(t)
Cx
(i)
Gr B
(A120573
+L)
Gr D
(A120573+
FA)
A120573 A120573 A120573 A120573
A120573A120573A120573
A120573 A120573 A120573 A120573
A120573A120573A120573A120573
A120573 A120573 A120573 A120573
A120573
(m)
Gr C
(S1P
+L)
Figure 2 Comparative effect of EB101 vaccine on beta amyloid deposits Comparative photomicrographs of A120573 immunoreactivity weretaken in the hippocampus ((a)ndash(e) (k)ndash(o)) and cortical ((f)ndash(j) (p)ndash(t)) brain regions of transgenic mice with different treatments beforeA120573 plaques development preventive treatment ((a)ndash(j)) and after the A120573 plaques developed therapeutic treatment ((k)ndash(t)) Preventivetreatment transverse brain sections of 11-month-old mice show reduced number of A120573 deposits in the dentate gyrus hippocampal subregionCA1 (a) and entorhinal cortex (f) following EB101 vaccine immunization (Group A) contrasting with the numerous A120573 immunoreactiveplaques in the correspondent mouse brain sections of Groups B ((b) (g)) C ((c) (h)) D ((d) (i)) and E ((e) (j)) Immunoreactive A120573 in thebrain sections of wild-type (WT) mice is absent (inserts in panels (e) (o)) Therapeutic treatment transverse brain sections of 18-month-oldmice are shown in panels (k)ndash(t) Treatment with EB101 almost completely cleared A120573 load in the dentate gyrus and reduced notably thedensity in hippocampal subregion CA1 (k) and entorhinal cortex (p) compared to the same areas of mouse Groups B ((l) (q)) C ((m)(r)) D((n) (s)) and E ((o) (t)) as determined by number of plaques and staining intensity area Scale bar 100 120583m
8 BioMed Research International
Gr E
(PBS
)
0102030405060708090
Preventive treatmentTherapeutic treatment
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Mea
n of
120573-a
myl
oid
plaq
ues
sect
ion
Gr A
(A120573+
S1P+
L)(E
B101
)
Figure 3 Comparative analysis of A120573 plaque burden in the brainsof APPPS1 mice Mean of A120573 plaque burden in the hippocampaland cortical regions of APPswePS1ΔE9 mice in the five treatmentgroups The mean of the A120573 burden is significantly reduced (119875 lt005) in Group A (EB101) when compared with Groups BndashE duringpreventive treatment and markedly reduced in the therapeutictreatment period Data are presented asmeanplusmn standard error of themean (SEM) Densitymean of A120573 plaques in Tgmice hippocampus
05
1015202530354045
Preventive treatmentTherapeutic treatment
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Gr A
(A120573+
S1P+
L)(E
B101
)
Mea
n of
120573-a
myl
oid
plaq
ues a
rea
Figure 4 Comparative quantification of A120573 burden area in thebrain of APPPS1 mice Quantitative analysis of A120573 burden areain the hippocampal and cortical regions of APPswePS1ΔE9 micetreated with EB101 A120573liposomes S1Pliposomes A120573
42+ FAIFA
and PBS represented by the number of pixels inside the stained areaof each A120573 plaque This graphic shows that A120573 plaques of EB101-treated mice (Group A) are significantly smaller in size (119875 lt 005)than those of the other four treatment groups Data are presented asmean plusmn SEM Mean of A120573 plaques area in Tg mice hippocampus
a reduced staining intensity of the A120573 plaques in the brainregions of mouse Group A contrasting with the intenseA120573-immunoreactivity of plaques in the other experimentalmouse groups as shown in the photomicrographs of Figures2(k)ndash2(t)
33 Immune Response Effects of EB101 versus A12057342+ CFAIFA
Vaccine Astrocyte activation implicated in neuroinflam-mation amyloidogenesis and neuronal cell death in AD
was compared after treatment of the transgenic mice withEB101 and A120573
42+ CFAIFA Immunohistochemical analysis
of GFAP-reactive cells was carried out in the brains ofthe treated mice in the preventive and therapeutic studies(Figure 5) After the preventive treatment (Figures 5(a)ndash5(j))EB101 significantly reduced the density of GFAP-reactive cellclusters in the hippocampal and cortical sections as comparedwith A120573
42+ CFAIFA (D) A120573Lip without S1P (B) Lip with
S1P (C) and PBS (E) The distribution of GFAP immunore-active cells in the transverse section of the mouse brainstreated with EB101 shows a few scattered GFAP-reactiveclusters mainly at the CA1 hippocampal layers contrastingwith numerous dystrophic reactive astrocytes observed indifferent hippocampal areas of mouse brains in Groups BndashE(Figures 5(b)ndash5(e)) Similar neuroinflammation pattern wasobserved in the retrosplenial cortical layers where a fewGFAP-reactive clusters were observed in EB101 treated mice(Figure 5(f)) compared with their profuse presence in theother experimental groups (Figures 5(g)ndash5(j)) After thera-peutic treatment the GFAP immunoreactivity distributionpattern in mouse brains treated with EB101 was similar tocontrol mice (wild-type) mostly devoid of reactive GFAPclusters except a few scattered ones in the CA1 hippocampalexternal layers (Figure 5(k)) In contrast there is a moderatedensity in Group D (Figure 5(n)) and extensive density inGroup B (Figure 5(l)) Group C (Figure 5(m)) and Group Emice (Figure 5(o)) Photomicrographs of retrosplenial cortexshow similar neuroinflammation pattern where a few GFAPclusters detected in the EB101 treated mice (Figure 5(p))contrast with a more extensive density of clusters in the othergroups (Figures 5(q)ndash5(t)) No astrocytosis was observed inwild-type mice during preventive or therapeutic treatment
34 Comparative A120573 Antibody Titers and Th1Th2 CytokineLevels in Treated Mice A120573 antibody titers in the sera weredetermined by ELISA during mouse immunization(Figure 6) Preventive treatment with EB101 resulted in amarked increase of specific IgG A120573
1ndash42 antibody productionin all animals Mice in A and D groups developed andmaintained serum antibody titers between 1 2000 and1 8000 Mice in Groups B C and E showed no significantanti-A120573 antibody production Antibody production and titerin Group B were three and four times lower than those ofGroups D and A respectively After therapeutic treatment allmice treated with EB101 and A120573
42+ FAIFA produced high
levels of IgG antibodies Titer levels of Groups A and D weresignificantly increased to those detected after preventivetreatment (119875 lt 001 versus Group D 119875 lt 0001 versusGroups B C and E) although antibody level in Group D was13-fold lower than in Group A (Figure 6)The effect of EB101(Group A) and A120573
42+ CFAIFA (Group B) vaccines on Th1
andTh2 cytokines was also studied (Figure 7)Measurementsof Th1 and Th2 cytokines in sera of immunized transgenicmice indicate that significant changes occurring in proteinlevels of Th1-related inflammatory mediators followingpreventive (Figures 7(a)ndash7(c)) and therapeutic (Figures 7(d)ndash7(g)) treatment with EB101 vaccine A significant increaseof Th1 cytokine profile was observed in preventive treatment
BioMed Research International 9
Gr E
(PBS
)
(d)
(e)
(i)
(j)
(n)
(o)
(s)
(t)
Cx
GFAP
GFAP GFAP GFAP GFAP
GFAP GFAP GFAP
Cx Cx
Cx
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
GFAP
Gr A
(EB1
01)
(a)
(b)
(c)
(f)
(g)
(h)
Preventive treatment Therapeutic treatment
PoDG
MoDG
(k)
(l)
(m)
(p)
(q)
(r)DG
GFAP GFAP GFAP
GFAPGFAPGFAP GFAP
GFAP GFAP GFAP GFAP
PoDGDG
DG
CA1
Cx
Cx
Cx
Cx
Cx
Cx
PoDG
MoDG
DG
CA1
MoDGDG
CA1
PoDG
MoDG
DG
CA1
Cx
CxCx
MoDG
MoDG
PoDG
CA1CA1
Cx
Gr D
(A120573+
FA)
Gr C
(S1P
+L)
Gr B
(A120573+
L)
Figure 5 Comparative effect of EB101 vaccine on astrocytosis Comparative photomicrographs of GFAP immunoreactivity in thehippocampus ((a)ndash(e) (k)ndash(o)) and cortical ((f)ndash(j) (p)ndash(t)) brain regions at the preventive treatment phase ((a)ndash(j)) and therapeutictreatment group ((k)ndash(t)) Preventive treatment transverse brain sections of 11-month-old mice treated with EB101 show an almost completeprevention of astrocytosis in the detailed sections of the dentate gyrus (a) contrasting with numerous dystrophic reactive astrocytes typicalof an inflammatory reaction observed in different hippocampal corresponding areas of mouse brains in Group B (A120573Lip-treated mice (b))Group C (LipS1P-treated mice (c)) Group D (A120573
42+ CFAIFA-treated mice (d)) and Group E (PBS-treated mice (e)) Immunoreactive
astrocytosis in the brain sections of WT mice is absent (data not shown) Detailed sections of the parietal cortex of treated mice show agradient density of reactive astrocytes immunoreactive to GFAP indicating a neuroinflammation pattern Comparative cortical sections shownumerous astrocytosis areas inmice Groups C and E ((h) (j)) andwide reactive astrocytes clusters inmice Groups B andD ((g) (i)) while justa few of them are observed in Group A (f)Therapeutic treatment transverse brain sections of 18-month-old mice of Group A show a notablereduction of astrocytosis in the hippocampal region (k) after EB101 vaccine immunization Note the astrocytosis contrast density betweenEB101-treatedmice and the other treated groups ((l)ndash(o)) where numerous immunoreactive GFAP clusters are observed in the correspondentbrain sections A few immunoreactive GFAP clusters are observed in the parietal cortex of EB101 treated mice group while a moderate ((q)(s)) to high ((r) (t)) density of these neuropathological inflammation clusters is apparent in the other treatment groups (Groups BndashE) Forabbreviations see list Scale bar 100120583m
10 BioMed Research International
002040608
1121416
Preventive treatmentTherapeutic treatment
Ant
i-120573-a
myl
oid
IgG
antib
odie
s
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Gr A
(A120573+
S1P+
L)(E
B101
)
Figure 6 Quantification of specific anti-120573-amyloid IgG in immu-nized APPPS1 mice The presence of anti-120573-amyloid antibodiesmeasured in the sera of vaccinated and control mice groups duringthe preventive and therapeutic treatments indicates that EB101liposome vaccine was able to induce the highest production ofspecific IgG antibodies Compared to theGroupD immunizedmicethe level of antibodies observed in the Group A was also highersuggesting a more effective immune response (119875 lt 001 versusGroup D 119875 lt 0001 versus Groups B C and E) Quantification ofspecific anti-120573-amyloid IgG in immunized Tg mice
in Group D with respect to all other groups mainlyreported by the high levels of TNF-120572 (Figure 7(b)) andIFN-120574 (Figure 7(c)) cytokine secretions while due to thehigh mortality rate during the therapeutic treatment nosignificant data was obtained from this mice group Inaddition a tendency toTh1 secretion reduction was observedinGroupAwith respect to all other groups in both preventiveand therapeutic treatment The changes in Th2 protein levels(Figure 7(d)) followed the inverse trend where the highestdifferences between Group A and the other treatment groupswere observed in the IL-4 IL-5 and IL-10 cytokine secretions(119875 lt 005 versus Group D 119875 lt 001 versus Group E Figures7(e)ndash7(g))
35 Comparative Motor Coordination and Memory Perfor-mances In order to validate the locomotion integrity duringthe entire immunotherapeutic process as reported in similarprevious studies motor coordination tests have showed thatEB101-treated mice spent significantly more time on theaccelerating rotarodThismotor coordination task shows thatEB101 immunization affected positively their performance(Figure 8(a)) A120573
42liposome (B) S1P-containing liposomes
(C) and PBS (E) (119875 lt 0006) mice performed poorly incomparison to EB101 mice (119875 lt 005) or A120573
42+ CFAIFA
mice (119875 lt 002) in motor abilities and coordination Thefear conditioning-active avoidance test was used to assess thespatial learning and memory functions As expected bothEB101 and A120573
42+ CFAIFA immunized mice readily learned
the location of the platform as shown by a decrease in theescape latency over training days while the control group
mice exhibited no significant changes during 6-day trialsThe average escape latencies of EB101 (68 plusmn 2 s 761 plusmn 22 s)andA120573
42+CFAIFA (116plusmn33 s 123plusmn21 s) immunizedmice
at the end of preventive and therapeutic periods respectivelyduring the consecutive 6 days trial were significantly shorterthan the other treatment groups (A120573
42liposome 163 plusmn 31 s
175 plusmn 28 s S1P-containing liposomes 202 plusmn 19 s 248 plusmn27 s) and the control (2116 plusmn 38 s 268 plusmn 22 s) (119875 lt 001)Therewas no significant difference between EB101 andA120573
42+
CFAIFA immunized group (119875 gt 005) (Figure 8(b)) Theseresults showed that spatial learning was improved in theseimmunized mice
4 Discussion
In this study we have compared a novel active A120573 imm-unotherapy the EB101 vaccine against A120573 with the A120573
42+
CFAIFA vaccine that was first taken into clinical trialswith coadjuvant Qs-21 modification (AN1792 by Orgogozoet al [19]) This type of vaccine design was thought toinduce a strong T-cell mediated inflammatory response (IL-2 and IFN-c positive responses indicative of a Class II(CD4) Th-1 type response [33] leading to acute menin-goencephalitis in the 6 of the patients) The EB101 activeimmunotherapeutic vaccine was primarily designed to avoidundesired inflammatory side effects both in mice andhumans (without need of coadjuvant modification) whileinducing effective antibody titer rates We demonstrated thatlong-term immunization with EB101 induced A120573 antibodiesin the preventive and the therapeutic treatment periodswhich effectively reduced amyloid deposition in both corticaland hippocampal brain regions and prevented neuroinflam-mation probably by blocking toxicity in the surroundingneurons However the AN1792 vaccine was shown to inducein 6 of the patients an inflammatory reaction that mighthave led to the encephalitis type reaction observed in theclinical trial which was responsible for the immediate haltin those clinical studies It seems that AN1792 immunizationdelivered in Qs-21 and polysorbate 80 adjuvant induced aTh1 type response that likely led to autoaggressive T cells andaseptic meningoencephalitis In the present experiments wedemonstrated that a judiciously selected adjuvant produces adifferent humoral response [22] Indeed we have shown thatour novel immunotherapeutic strategy boosts specific A120573
1ndash42antibody generation and induces a Th2 noninflammatoryimmune response This boosted effect is in accordance withprevious immunotherapeutic studies [34] Taking togetherall the present findings these results suggest that EB101vaccine can achieve in a future clinical trial immunogenicresponse rates quite similar to the AN1792 without theinflammatory effects reported The immunohistochemicalanalysis obtained in the present study shows that all the EB101immunized mice resulted in an almost complete preventionof A120573 deposition in the brain with only a few compactA120573 plaques in the retrosplenial and dentate gyrus regionsAlthough there was a slightly increase in death in EB101treated mice compared to controls this significant reductionof A120573 plaques in EB101 treated mice was proven to be more
BioMed Research International 11
Gr A Gr B Gr C Gr D Gr EPreventive treatmentTherapeutic treatment
0005001000150020002500300035004000
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
(a) Th1
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
000
1000
2000
3000
4000
5000TNF-120572
(b) Th1
000500
10001500200025003000
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
IFN-120574
(c) Th1
000500
100015002000250030003500
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
Group A Group B Group C Group D Group EPreventive treatmentTherapeutic treatment
(d) Th2
000
500
1000
1500
2000
2500
3000IL-4
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(e) Th2
000500
100015002000250030003500
IL-5
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(f) Th2
3500IL-10
000500
10001500200025003000
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(g) Th2
Figure 7 Detection of Th1 and Th2 cytokine profile Measurements of the effect of EB101 and A12057342
+ CFAIFA vaccines on Th1 and Th2cytokines profile during preventive and therapeutic treatments ((a)ndash(c)) A significant increase of Th1 cytokine profile was observed inpreventive treatment in Group D with respect to all other groups while a tendency to a reduction inTh1 secretion was observed in Group Awith respect to all other groups in both preventive and therapeutic treatment ((d)ndash(g)) The changes in Th2 protein levels were observed inthe IL-4 IL-5 and IL-10 cytokine secretions of Group A (119875 lt 005 versus Group D 119875 lt 001 versus Group E)
12 BioMed Research International
Gr A
(EB1
01)
05
101520253035
Preventive treatment Therapeutic treatment
Mea
n fa
lling
late
ncy
(s)
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)(a) Graphic of motor coordination and balance
05
1015202530
Mea
n re
spon
se ti
me (
s)
Gr A
(EB1
01)
Preventive treatment Therapeutic treatment
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
(b) Graphic of associative learning and memory
Figure 8 Analysis of motor coordination and balance (a) Rotarod results of motor coordination and balance test after the preventive andtherapeutic treatment showing improved effect of EB101 and A120573
42+ CFAIFA-vaccine compared to other treated mice groups (119875 lt 005)
and (119875 lt 002) versus control mice Latency to fall on the rotarod apparatus was significantly improved in EB101 immunized mice Valuesrepresent means of all six trials of a groupday (b) Spatial learning and memory tested by fear conditioning-active avoidance test The meanresponse time of each treatment group to reach the platform is represented EB101 and A120573
42+ CFAIFA immunized mice had significantly
lower latencies than the other treatment groups (119875 lt 001) versus control mice
efficient than that obtained with A12057342+ CFAIFA immuniza-
tions A comparison of A120573 plaque reduction rates amongtreated groups surprisingly showed that other than EB101and A120573
42+ CFAIFA treated groups some amyloid deposits
were also reduced in Groups B (A1205731ndash42liposome) and C
(S1P-containing liposomes) Although the reduction rates inthese mice were markedly low the subtle lowering effectmight be due to the interaction between the immunizationelements (A120573
1ndash42 and S1P) and the mouse immunogenicsystem as reported by others [35] Based on our presentresults S1P one of the components of the adjuvant used withEB101 might be responsible for the neuroprotective effectobserved in the vulnerable neuronal areas of EB101 treatedmice The advantage in the A120573 load preventionreduction bycombining A120573
1ndash42 and S1P in a unique formulation vaccinehas been also observed in the reduced hippocampal load ofEB101 immunizedmice (Gr A) when comparedwith differentvaccine components (Gr B-C) In fact S1P appears to have ademonstrated therapeutic potential as a regenerative agent inthe nervous system since it controls migration of neuronalstem cells toward a site of spinal cord injury [36] inducecytoskeletal rearrangements to promote transmitter secretion[37] and also trigger diverse cellular effects including angio-genesis cardiac development immunity cell motility andneurite extension [38 39]Therefore it is reasonable to expectthat such a potent biologically active sphingolipid by itselfmay play an important role in the A120573 plaque reduction Inthis study the clearance efficiency rate of each treatment wasanalyzed and results suggested that EB101 (6459) appearsto be more effective than A120573
42+ CFAIFA (1920) vaccine
in both preventive and therapeutic groups The effectivenessof EB101 in older transgenic animals (18 months) showeda clearance effect of 59 suggesting that EB101 may alsohave therapeutic potential inADdiagnosticated patient trials
Similar conclusion was reached byWang and colleagues [40]who demonstrated that combination therapy in aged animalsprovides more valuable insights into a possible translationto humansOur immunologicalmeasurements demonstratedonce again that neuroinflammation develops in an A120573-dependent fashion early in the course of AD [41] and canbe alleviated or reversed by interventional A120573 vaccinationsuch as EB101 Among the possible mechanisms of A120573clearance proposed in AD immunotherapy [42] our resultssuggested that theymay overlap and probably be disease statedependent since the prevention and the therapeutic vaccinewould likely benefit from the transport of A120573 antibodies intothe CNS However some mechanisms of action such as thepreventive effect of A120573 antibodies in the A120573 aggregation [43]microglial phagocytosis induced by the Fc binding portionsbetween A120573 antibodies and microglia receptor [44] and theshift in the gradient of A120573 transport across the blood-brainbarrier due to the increased concentration of A120573 antibodiesin the vascular system resulting in an increase in effluxfrom the brain to blood [45 46] may be playing a role inboth preventive and therapeutic conditions Our findingstherefore suggest that novel treatment strategies aimed atmaintaining or increasing microglial function and restoringthe balance to the immunological system may represent anattractive therapeutic approach even at the advanced stagesof ADThe immunization results obtained with EB101 duringboth preventive and therapeutic treatment showed nomicro-hemorrhages infiltration of lymphocytes or encephalitis-like reaction in the brain suggesting that EB101 does notinduce A120573-specific Th1 reaction possibly due to the neu-roprotective effect of S1P Delivering A120573
1ndash42 in a liposomal-S1P-adjuvant resulted in an effective immunological responseincluding the induction of a Th2 T-cell response whicheffectively prevented neuroinflammation in an animal model
BioMed Research International 13
of AD as reported previously [24 47] Moreover presentresults demonstrated that EB101 did not induce astrogliosisin APPPS1 transgenic mice whereas some GFAP-positiveastrocyte clusters were aggregated in the affected brain areasof mice treated with A120573
42+ CFAIFA These results indicate
that EB101 may result in an effective treatment in AD neu-ropathology due to the absence of peripheral proinflamma-tory cytokine secretion and the absence of astroglial activa-tion in the brains of EB101 treated mice when compared withA12057342
+ CFAIFA immunotherapeutic response In a clinicalsetting however we may expect a wide differential responsein part depending upon the genomic profile of each patient[48 49] In AD behavioural studies in particular amongthe motor coordination tests the rotarod performance hasbeen reported to be impaired in some AD transgenic micemodels bearing A120573 plaques [50 51] This result was found inmutants prior to A120573 plaque formation at 3 and 6 months ofage but was not reproduced after plaques had been formedat 16 and 24 months of age [52] perhaps because of thedeclining abilities of older normal mice Here we presentthe effects of vaccine treatments in APPPS1 mice on motorcoordination a neglected aspect of the AD experimentalbehavior but justified on the basis of the deficient posturalcontrol reported in patients with Alzheimerrsquos disease [53]sometimes even during early stages [54] Our longitudinalmotor behavioral results showed no significant differencesin the ambulatory movement grip strength or coordinationaddressed by their performance in the rotating rod betweenAPPPS1 and wild-type mice for any age group studiedin contrast with other treated groups demonstrating thatlongitudinal motor assessments of AD transgenic mousemodels can provide valuable information with regard tothe effect of immunization on the progression of behavioraldeficits and overall motor activity As shown the EB101-treated group of mice exhibited longer latencies and betterbalancing skills indicating the absence of any negative effectof EB101 immunization on these mice The classical fearconditioning test is assumed to reveal the functional integrityof the hippocampus [55] representing a special memorytask to evaluate the AD models and its improvement afterimmunization periods In this APPPS1 mouse model thefirst associative learning and spatial workingmemory deficitsare shown at 5-6 months of age [56] although its impair-ment progression is relatively slow compared to most othermodels At the end of preventive immunization period EB101and A120573
42+ CFAIFA immunized mice have significantly
shorter escape latencies than the other treatment groups(A12057342liposome S1P-containing liposomes) and the controls
in the fear conditioning test Shorter escape latencies duringspatial learning indicated that deficits of spatial memorywere attenuated in EB101 and A120573
42+ CFAIFA immunized
mice Previous studies have showed that by 15 months ofage the spatial working memory is consistently impairedthroughout the rest of the life span [31 57] At the end oftherapeutic immunization period both EB101 and A120573
42+
CFAIFA immunized mice showed significantly improvedresults on the hippocampus-dependent spatial learning andmemory test when compared with other treatment groupsand the controls Taken together these observations are
in agreement with the impairment in conditioned learningin response to a tone stimulus of aged APPPS1 micealthough when immunized with EB101 during preventiveor therapeutic periods such impairments are significantlyattenuated
In summary the present results corroborate our previouspublished findings showing that EB101 immunization reducesamyloid accumulation in the APPPS1 mouse brain [2425] The strongest reduction in the preventive immunizationwas observed in the retrosplenial cortex followed by thehippocampus and entorhinal-piriform cortex Our resultsfurther support that EB101 ismore effectivewhen themice areimmunized prior to developing plaques in the hippocampaland cortical brain regions There is extensive evidence inmouse preclinical studies [58] and human clinical trials [5960] that the adverse effects of active and passive A120573 vaccinesresult in microhemorrhages and encephalitic reactions [60ndash63] yet themechanismbywhich they occur has not been fullyelucidated In the treatment sections analyzed we found nomicrohemorrhages or acute inflammatory processes in all thesurvived mice of the group indicating that EB101 therapeuticapproach presents no adverse inflammatory effects in thisAD mice models Thus our present results unequivocallydemonstrate that EB101 vaccinemay not only halt AD-relatedpathology as a preventive treatment but may also reverse it inestablished cases related to the improvement of the learningandmemory in both cases leading to a robust immunologicaltherapeutic response Present experiments also demonstratethat EB101 exerts more significant and substantial effects onthe reduction of A120573 deposition than A120573
42+CFAIFA vaccine
in A120573 mice models of AD However further exploration ofthe contribution of genetic environmental and physiologicalfactors in the development resistance to endogenous A120573-antibody is needed in order to optimize and modulate thetherapeutic strategies currently being developed
5 Conclusions
The present study carried out in APPPS1 transgenic mousemodel of AD compared the A120573 burden reduction andcellular immune responses produced by two immunizationdelivery strategies of A120573 in Freund adjuvant (described byWebster et al [10]) and in liposomeS1P matrix (describedby Schenk et al [17])The EB101 immunotherapeutic strategyboosted antibody generation A120573 plaque reduction and ananti-inflammatory response more effectively than A120573
42+
CFAIFA especially if given prior to or in the early stages ofpathology The present results also showed that vaccinationwith EB101 prevented the formation of new plaques andpresumably led to a reduction in established plaques whileinducing aTh2 immune reactionwithout encephalitogenic T-cell responses or brain inflammation related with astrogliosisHowever despite the efficacy demonstrated of this novelvaccine approach further preclinical and clinical studieson EB101 are warranted to evaluate its efficacy and themechanism for the future utility of such vaccine for ADtherapy
14 BioMed Research International
List of Abbreviations
AD Alzheimerrsquos diseaseA120573 120573-Amyloid proteinAPP Amyloid precursor proteinAPT Anterior pretectal nucleusCA1 Field CA1 hippocampusCA3 Field CA3 hippocampusCPu Caudate putamen (striatum)Cx CortexDG Dentate gyrusDN Dystrophic neuriteEct Entorhinal cortexCFA Complete Freundrsquos adjuvantGrDG Granular dentate gyrusIFA Incomplete Freundrsquos adjuvantIL InterleukinLPMR Lateral posterior thalamic nucleus (mediorostral)MoDG Molecular dentate gyrusPir Piriform cortexPoDG Polymorph dentate gyrusRSGb Retrosplenial granular cortex b regionTg TransgenicTh T helper
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Ivan Carrera Ignacio Etcheverrıa Lucıa Fernandez-NovoaValter Lombardi Ramon Cacabelos and Carmen Vigo con-ceived and designed the experiments Ivan Carrera and Igna-cio Etcheverrıa performed the experiments Ivan CarreraIgnacio Etcheverrıa and Valter Lombardi analyzed the dataIvan Carrera Ignacio Etcheverrıa Lucıa Fernandez-Novoaand Valter Lombardi contributed reagentsmaterialsanalysistools Ivan Carrera Carmen Vigo andMadepalli KrishnappaLakshmana wrote the paper
Acknowledgments
This work was supported by National Institute of Aging(NIA)NIH Grant no 1R01AG036859-01 (Madepalli Krish-nappa Lakshmana) and the financial support of the ProgramaOperativo Feder Galicia 2007ndash2013 (IN841C 201254 to PhDCarrera I)
References
[1] R Cacabelos L Fernandez-Novoa V Lombardi Y Kubotaand M Takeda ldquoMolecular genetics of Alzheimerrsquos diseaseand agingrdquoMethods and Findings in Experimental and ClinicalPharmacology vol 27 pp 1ndash573 2005
[2] P T Nelson I Alafuzoff E H Bigio et al ldquoCorrelationof alzheimer disease neuropathologic changes with cognitive
status a review of the literaturerdquo Journal of Neuropathology andExperimental Neurology vol 71 no 5 pp 362ndash381 2012
[3] JNaslundVHaroutunian RMohs et al ldquoCorrelation betweenelevated levels of amyloid 120573-peptide in the brain and cognitivedeclinerdquo Journal of the American Medical Association vol 283no 12 pp 1571ndash1577 2000
[4] A Koyama O I Okereke T Yang D Blacker D J Selkoeand F Grodstein ldquoPlasma amyloid-120573 as a predictor of dementiaand cognitive decline a systematic review and meta-analysisrdquoArchives of Neurology vol 69 no 7 pp 824ndash831 2012
[5] A M Fagan C M Roe C Xiong M A Mintun J C Morrisand D M Holtzman ldquoCerebrospinal fluid tau120573-amyloid42ratio as a prediction of cognitive decline in nondemented olderadultsrdquo Archives of Neurology vol 64 no 3 pp 343ndash349 2007
[6] S Parvathy P Davies V Haroutunian et al ldquoCorrelationbetween A120573-40- A120573-42- and A120573x-43-containing amyloidplaques and cognitive declinerdquo Archives of Neurology vol 58no 12 pp 2025ndash2032 2001
[7] E Karran M Mercken and B D Strooper ldquoThe amyloidcascade hypothesis for Alzheimerrsquos disease an appraisal for thedevelopment of therapeuticsrdquo Nature Reviews Drug Discoveryvol 10 no 9 pp 698ndash712 2011
[8] J L Jankowsky D J Fadale J Anderson et al ldquoMutantpresenilins specifically elevate the levels of the 42 residue 120573-amyloid peptide in vivo evidence for augmentation of a 42-specific 120574 secretaserdquo Human Molecular Genetics vol 13 no 2pp 159ndash170 2004
[9] L Holcomb M N Gordon E Mcgowan et al ldquoAcceleratedAlzheimer-type phenotype in transgenic mice carrying bothmutant amyloid precursor protein and presenilin 1 transgenesrdquoNature Medicine vol 4 no 1 pp 97ndash100 1998
[10] S JWebster A D Bachstetter and L J van Eldik ldquoComprehen-sive behavioral characterization of an APPPS-1 double knock-in mouse model of Alzheimerrsquos diseaserdquo Alzheimerrsquos ResearchandTherapy vol 5 no 3 article 28 2013
[11] H Xiong D Callaghan J Wodzinska et al ldquoBiochemical andbehavioral characterization of the double transgenic mousemodel (APPswePS1dE9) of Alzheimerrsquos diseaserdquo NeuroscienceBulletin vol 27 no 4 pp 221ndash232 2011
[12] M Garcia-Alloza L A Borrelli B T Hyman and B J BacskaildquoAntioxidants have a rapid and long-lasting effect on neuriticabnormalities in APPPS1 micerdquo Neurobiology of Aging vol 31no 12 pp 2058ndash2068 2010
[13] M Meyer-Luehmann M Mielke T L Spires-Jones et al ldquoAreporter of local dendritic translocation shows plaque-relatedloss of neural system function in APP-transgenic micerdquo Journalof Neuroscience vol 29 no 40 pp 12636ndash12640 2009
[14] E Machova V Rudajev H Smyckova H Koivisto H Tanilaand V Dolezal ldquoFunctional cholinergic damage develops withamyloid accumulation in young adult APPswePS1dE9 trans-genic micerdquo Neurobiology of Disease vol 38 no 1 pp 27ndash352010
[15] T Malm J Koistinaho and K Kanninen ldquoUtilization of APP-swePS1dE9 transgenic mice in research of Alzheimerrsquos diseasefocus on gene therapy and cell-based therapy applicationsrdquoInternational Journal of Alzheimerrsquos Disease vol 2011 Article ID517160 8 pages 2011
[16] J Lee and P Han ldquoAn update of animal models of alzheimerdisease with a reevaluation of plaque depositionsrdquoExperimentalNeurobiology vol 22 pp 84ndash95 2013
BioMed Research International 15
[17] D Schenk R Barbour W Dunn et al ldquoImmunization withamyloid-120573 attenuates Alzheimer disease-like pathology in thePDAPP mouserdquo Nature vol 400 no 6740 pp 173ndash177 1999
[18] H J Fu B Liu J L Frost and C A Lemere ldquoAmyloid-120573immunotherapy for Alzheimerrsquos diseaserdquoCNS and NeurologicalDisorders-Drug Targets vol 9 no 2 pp 197ndash206 2010
[19] J-M Orgogozo S Gilman J-F Dartigues et al ldquoSubacutemeningoencephalitis in a subset of patients with AD after A120573
42
immunizationrdquo Neurology vol 61 no 1 pp 46ndash54 2003[20] J A R Nicolll DWilkinson CHolmes P Steart HMarkham
andROWeller ldquoNeuropathology of humanAlzheimer diseaseafter immunization with amyloid-120573 peptide a case reportrdquoNature Medicine vol 9 no 4 pp 448ndash452 2003
[21] I Ferrer M Boada Rovira M L Sanchez Guerra M JRey and F Costa-Jussa ldquoNeuropathology and pathogenesisof encephalitis following amyloid-Neuropathology and patho-genesis of encephalitis following amyloid-beta immunizationin Alzheimerrsquos disease immunization in Alzheimerrsquos diseaserdquoBrain Pathology vol 14 no 1 pp 11ndash20 2004
[22] D H Cribbs A Ghochikyan V Vasilevko et al ldquoAdjuvant-dependent modulation ofTh1 andTh2 responses to immuniza-tion with 120573-amyloidrdquo International Immunology vol 15 no 4pp 505ndash514 2003
[23] C Holmes D Boche D Wilkinson et al ldquoLong-term effectsof A120573
42immunisation in Alzheimerrsquos disease follow-up of a
randomised placebo-controlled phase I trialrdquo The Lancet vol372 no 9634 pp 216ndash223 2008
[24] I Carrera I Etcheverrıa L Fernandez-Novoa V LombardiR Cacabelos and C Vigo ldquoVaccine development to treatAlzheimerrsquos disease neuropathology in APPPS1 transgenicmicerdquo International Journal of Alzheimerrsquos Disease vol 2012Article ID 376138 17 pages 2012
[25] I Carrera I Etcheverrıa Y Li et al ldquoImmunocytochemicalcharacterization of Alzheimer disease hallmarks in APPPS1transgenic mice treated with a new anti-amyloid-120573 vaccinerdquoBioMed Research International vol 2013 Article ID 709145 12pages 2013
[26] V Anelli R Bassi G Tettamanti P Viani and L Riboni ldquoExtra-cellular release of newly synthesized sphingosine-1-phosphateby cerebellar granule cells and astrocytesrdquo Journal of Neuro-chemistry vol 92 no 5 pp 1204ndash1215 2005
[27] S Schroeter K Khan R Barbour et al ldquoImmunotherapyreduces vascular amyloid-120573 in PDAPP micerdquo Journal of Neu-roscience vol 28 no 27 pp 6787ndash6793 2008
[28] Z Suo A Cox N Bartelli et al ldquoGRK5 deficiency leads to earlyAlzheimer-like pathology and working memory impairmentrdquoNeurobiology of Aging vol 28 no 12 pp 1873ndash1888 2007
[29] W Kamphuis C Mamber MMoeton et al ldquoGFAP isoforms inadult mouse brain with a focus on neurogenic astrocytes andreactive astrogliosis in mouse models of Alzheimer diseaserdquoPLoS ONE vol 7 no 8 Article ID e42823 2012
[30] J Lang C Vigo-Pelfrey and F Martin ldquoLiposomes composedof partially hydrogenated egg phosphatidylcholines fatty acidcomposition thermal phase behavior and oxidative stabilityrdquoChemistry and Physics of Lipids vol 53 no 1 pp 91ndash101 1990
[31] D Morgan D M Diamond P E Gottschall et al ldquoA 120573peptide vaccination prevents memory loss in an animal modelof Alzheimerrsquos diseaserdquo Nature vol 408 no 6815 pp 982ndash9852000
[32] M L Brownlow L Benner D DrsquoAgostino M N Gordon andD Morgan ldquoKetogenic diet improves motor performance but
not cognition in two mouse models of Alzheimerrsquos pathologyrdquoPLoS ONE vol 8 no 9 Article ID e75713 2013
[33] M Pride P Seubert M Grundman M Hagen J Eldridgeand R S Black ldquoProgress in the active immunotherapeu-tic approach to Alzheimerrsquos disease clinical investigationsinto AN1792-associated meningoencephalitisrdquo Neurodegenera-tive Diseases vol 5 no 3-4 pp 194ndash196 2008
[34] CWiessner K-HWiederhold A C Tissot et al ldquoThe second-generation activeA120573 immunotherapyCAD106 reduces amyloidaccumulation in APP transgenic mice while minimizing poten-tial side effectsrdquoThe Journal of Neuroscience vol 31 no 25 pp9323ndash9331 2011
[35] D Lambracht-Washington and R Rosenberg ldquoAdvances in thedevelopment of vaccines for Alzheimerrsquos diseaserdquo DiscoveryMedicine vol 15 no 84 pp 319ndash326 2013
[36] A Kimura T Ohmori R Ohkawa et al ldquoEssential roles ofsphingosine 1-phosphateS1P
1receptor axis in the migration of
neural stem cells toward a site of spinal cord injuryrdquo Stem Cellsvol 25 no 1 pp 115ndash124 2007
[37] S S Lee C H Woo J D Chang and J H Kim ldquoRoles of Racand cytosolic phospholipase A2 in the intracellular signalling inresponse to titanium particlesrdquo Cellular Signalling vol 15 no 3pp 339ndash345 2003
[38] E Masliah L Hansen A Adame et al ldquoA120573 vaccination effectson plaque pathology in the absence of encephalitis inAlzheimerdiseaserdquo Neurology vol 64 no 1 pp 129ndash131 2005
[39] A Pilorget B Annabi F Bouzeghrane J Marvaldi J Luisand R Beliveau ldquoInhibition of angiogenic properties of brainendothelial cells by platelet-derived sphingosine-1-phosphaterdquoJournal of Cerebral Blood Flow and Metabolism vol 25 no 9pp 1171ndash1182 2005
[40] A Wang P Das R C Switzer III T E Golde and J LJankowsky ldquoRobust amyloid clearance in a mouse model ofAlzheimerrsquos disease provides novel insights into themechanismof amyloid-120573 immunotherapyrdquo Journal of Neuroscience vol 31no 11 pp 4124ndash4136 2011
[41] G Krabbe A Halle V Matyash et al ldquoFunctional impairmentof microglia coincides with Beta-amyloid deposition in micewith Alzheimer-like pathologyrdquo PLoS ONE vol 8 no 4 ArticleID e60921 2013
[42] Y H Liu B Giunta H-D Zhou J Tan and Y J WangldquoImmunotherapy for Alzheimer disease-the challenge ofadverse effectsrdquo Nature Reviews Neurology vol 8 no 8 pp465ndash469 2012
[43] B Solomon R Koppel D Frankel and E Hanan-AharonldquoDisaggregation of Alzheimer120573-amyloid by site-directedmAbrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 94 no 8 pp 4109ndash4112 1997
[44] F Bard C Cannon R Barbour et al ldquoPeripherally adminis-tered antibodies against amyloid 120573-peptide enter the centralnervous system and reduce pathology in a mouse model ofAlzheimer diseaserdquo Nature Medicine vol 6 no 8 pp 916ndash9192000
[45] R B DeMattos K R Bales D J Cummins J C Dodart S MPaul and D M Holtzman ldquoPeripheral anti-A120573 antibody altersCNS and plasma A120573 clearance and decreases brain A120573 burdenin a mouse model of Alzheimerrsquos diseaserdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 98 no 15 pp 8850ndash8855 2001
[46] C A Lemere E T Spooner J LaFrancois et al ldquoEvidence forperipheral clearance of cerebral A120573 protein following chronic
16 BioMed Research International
active A120573 immunization in PSAPP micerdquo Neurobiology ofDisease vol 14 no 1 pp 10ndash18 2003
[47] T Town M Vendrame A Patel et al ldquoReduced Th1 andenhanced Th2 immunity after immunization with Alzheimerrsquos120573-amyloid
1minus42rdquo Journal of Neuroimmunology vol 132 no 1-2
pp 49ndash59 2002[48] R Cacabelos ldquoPharmacogenomics in Alzheimerrsquos diseaserdquo
Methods in Molecular Biology vol 448 pp 213ndash357 2008[49] R Cacabelos ldquoPharmacogenomics and therapeutic strategies
for dementiardquo Expert Review of Molecular Diagnostics vol 9no 6 pp 567ndash611 2009
[50] D van Dam R DrsquoHooge M Staufenbiel C van Ginneken Fvan Meir and P P de Deyn ldquoAge-dependent cognitive declinein the APP23 model precedes amyloid depositionrdquo EuropeanJournal of Neuroscience vol 17 no 2 pp 388ndash396 2003
[51] M Dumont C Strazielle M Staufenbiel and R LalondeldquoSpatial learning and exploration of environmental stimuli in24-month-old female APP23 transgenic mice with the Swedishmutationrdquo Brain Research vol 1024 no 1-2 pp 113ndash121 2004
[52] R Lalonde M Dumont M Staufenbiel C Sturchler-Pierratand C Strazielle ldquoSpatial learning exploration anxiety andmotor coordination in female APP23 transgenic mice with theSwedish mutationrdquo Brain Research vol 956 no 1 pp 36ndash442002
[53] F J Huff F Boller F Lucchelli R Querriera J Beyer and SBelle ldquoThe neurologic examination in patients with probableAlzheimerrsquos diseaserdquo Archives of Neurology vol 44 no 9 pp929ndash932 1987
[54] A F Pettersson M Engardt and L O Wahlund ldquoActivitylevel and balance in subjects with mild Alzheimerrsquos diseaserdquoDementia and Geriatric Cognitive Disorders vol 13 no 4 pp213ndash216 2002
[55] L Calandreau B Desgranges R Jaffard and A DesmedtldquoSwitching from contextual to tone fear conditioning and viceversa the key role of the glutamatergic hippocampal-lateralseptal neurotransmissionrdquo Learning amp Memory vol 17 no 9pp 440ndash443 2010
[56] F Trinchese S Liu F Battaglia SWalter PMMathews andOArancio ldquoProgressive age-related development of Alzheimer-like pathology in APPPS1 micerdquo Annals of Neurology vol 55no 6 pp 801ndash814 2004
[57] D M Wilcock A Rojiani A Rosenthal et al ldquoPassiveimmunotherapy against A120573 in aged APP-transgenic micereverses cognitive deficits and depletes parenchymal amyloiddeposits in spite of increased vascular amyloid and microhem-orrhagerdquo Journal of Neuroinflammation vol 1 article 24 2004
[58] S A Ferguson S Sarkar and L C Schmued ldquoLongitudinalbehavioral changes in the APPPS1 transgenic AlzheimerrsquosDisease modelrdquo Behavioural Brain Research vol 242 no 1 pp125ndash134 2013
[59] D Wilcock and C Colton ldquoImmunotherapy vascular pathol-ogy and microhemorrhages in transgenic micerdquo CNS andNeurological DisordersmdashDrug Targets vol 8 no 1 pp 50ndash642009
[60] M Grundman S Gilman R S Black et al ldquoAn alternativemethod for estimating efficacy of the AN1792 vaccine foralzheimer diseaserdquo Neurology vol 71 no 9 pp 697ndash698 2008
[61] D Boche E Zotova R O Weller et al ldquoConsequence of A120573immunization on the vasculature of human Alzheimerrsquos diseasebrainrdquo Brain vol 131 no 12 pp 3299ndash3310 2008
[62] M Pfeifer S Boncristiano L Bondolfi et al ldquoCerebral hemor-rhage after passive anti-a120573 immunotherapyrdquo Science vol 298no 5597 p 1379 2002
[63] M M Racke L I Boone D L Hepburn et al ldquoExacerbationof cerebral amyloid angiopathy-associatedmicrohemorrhage inamyloid precursor protein transgenic mice by immunotherapyis dependent on antibody recognition of deposited forms ofamyloid 120573rdquo Journal of Neuroscience vol 25 no 3 pp 629ndash6362005
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

4 BioMed Research International
Table1Summaryof
antib
odiesu
sed
Antibod
yDilu
tion
Immun
ogen
Source
Labelin
gClon
enam
eTy
peCatalog
number
Lotn
umber
Ref
B-Amyloid
11000
KLH-con
jugatedsynthetic
peptidec
orrespon
ding
aa36ndash4
2of
human
amyloidBx
-42peptide
Millipore
Amyloid-Bx
-42
polypeptide
12F4
Mou
semon
oclonal
Num
ber0
5-831
DAM1447121
[242527]
GFA
P140
0Antiglialfi
brillaryacid
protein
(mou
seIgG1isotype)
Sigm
aAldric
hAntiglialfi
brillary
acid
protein
G-A-5
Mou
semon
oclonal
Num
berG
3893
078K
4830
[924252829]
DNs
1300
Degeneratingplaque
neurites
Millipore
Degeneratingplaque
neuritesa
ndneurofi
brillarytang
les
mdashRa
bbitpo
lyclo
nal
Num
berA
B1518
PS014344
96[242527]
BioMed Research International 5
27 A120573 Load Calculation Digital images of 7 randomlyselected microscopic transverse sections per treated animalwere analyzed and the percentage of immunostained A120573deposition was determined for all the markers studied asdefined by the stereotaxic Bregma coordinates A total of 7selected sections per animal were evaluated using the NIHImage J program by defining region of interest and setting athreshold to discriminate nonspecific staining Quantitativeanalysis of amyloid burden area was also performed in thehippocampal regions and parietaltemporal cortical regionsWe used areapixels analysis software (Pixcavator 4) toquantify the average number of pixels inside each A120573 plaqueper brain section Therefore Pixcavator intensity thresholdimaging was set to mark only those areas stained with 120573-amyloid antibody (A120573 load) relative to the entire imagearea The immunostaining intensity of each image was firstaveraged per mouse and then per group and expressed inpercentage units
28 Imaging Images were visualized using a microscope(Olympus BX50) and digitized using a digital camera (DP-10Olympus) A120573 burden data was determined by quantitativeimage analysis (Pixcavator 4) and reported as plaques persection (ps) or percentage of immunolabeled area cap-tured within total sample area The microphotographs wereadjusted for brightness and contrast with Corel Photo-Paint(Corel Ottawa Canada) and plates were composed withCorel Draw
29 Sensorimotor and Cognitive Testing After the adminis-tration of the vaccine mice were subjected to sensorimotorand memory testing to measure their coordination andlearning performances during the experimental period andto verify that any treatment-related effects observed in thememory tasks could not be explained by differences in senso-rimotor abilities Prior to testing as previously described [32]the mice were adapted to the room and testing conditions
291 Rotarod The motor strength ability balance andcoordination skills of all the experimental mice in the twotreatment periods were evaluated in a rotarod apparatus(Columbus Instruments Columbus OH) operated at 10 rpmbeginning at 7 weeks of age (preventive group) and 35 weeksof age (therapeutic group) The animals were adapted to theapparatus by receiving training sessions during four weekssufficient to reach a baseline level of performance This pro-cedurewas designed to assessmotor behavior during six trialsof a groupday If themouse remained on the rod for 1minutethe test was completed and scored as 1 Each animal wastested previous to the treatment injection for three sessionswith each session separated by 15 minutes The time thateachmouse remained on the rodwas registered automaticallyand stopped when the animal fell or inverted (by climbing)from the top of the rotating barrel In this experimentone investigator evaluated the rotarod performance and wasblinded to mice treatments and the second one treated themice and scored the test to avoid a bias
292 Fear ConditioningActive Avoidance This fear-moti-vated associative avoidance test is based on electric currentwhere themouse has to learn to predict the occurrence of thisaversive event by actively moving to a different compartment(escape platform) This active avoidance task provides asimple way to assess associative learning and memory bytesting the ability of the mouse to avoid an aversive event inresponse to a stimulus cueThemeasures recorded number ofnonresponses (the mouse failing to reach the escape platformduring the trial) and response latency (latency to avoid orescape) serves as an index of learning and allows memoryto be assessed After habituation for 5min mice received 30randomized escapable footshocks at an intensity of 025mAIn each shock trial the time to reach the escape platformlocated constantly at a central quadrant from the startlocationwas recorded as latency Each trial lasted amaximumof 30 s If the mouse failed to find the platform within 30 s itwas manually guided to the platform and allowed to stay for10 s the latency was recorded 30 s Experimental results areindicated as mean response time for each treatment period
210 Onset and Survival Disease onset and progression wereconfirmed and defined by periodic mice sacrifice and neu-ropathological analysisThemortality rates of APPPS1 trans-genicmice in the course of both sets of experiments were 42for A120573
42+ CFAIFA 17 for EB101 and 12 for the vehicle
211 Statistical Analyses For statistical analyses SPSS (ver-sion 110 SPSS Inc Chicago) was used and a 119875 value lt005 was considered statistically significant The correlationaverage of A120573 plaque density burden area and antibody titerswere analyzed and compared in multiple groups by one-wayanalysis of variance (ANOVA) followed by Fisherrsquos LSD posthocmeans comparison test analysis when relevant All graphsdisplay group mean plusmn SEM
212 Limitations of the Study The ultimate goal of the ADpreclinical research is developing a therapeutic drug strategyin the ideal animal model that should recapitulate most ifnot all the neurobehavioral pathology and underlying thesame mechanisms described in human AD such as therate of amyloid plaque accumulation neurodegenerationand behavioral and cognitive alterations Although such anideal mouse model has not yet been generated we believethat APPswePS1ΔE9 transgenic mice present enough patho-logical features to answer the specific questions addressedby the present study Active A120573 immunotherapy in whichantibodies are produced by the individual upon contact withthe antigen has great potential for preventive AD therapy asshown here in animal models although some aspects suchas an undesirable immune response induced by the adjuvantlong immune response adjustment age-dependent effectsinflammation and microhemorrhages should be addressedbefore its application in clinical trials Being aware of theseissues we designed the present study aiming to solve andovercome all these disadvantages
6 BioMed Research International
3 Results
31 Efficacy of EB101 versus A12057342
+ CFAIFA in PreventingA120573 Burden Development Recently we demonstrated thatimmunization with EB101 vaccine prevents and reverses theAD neuropathology by the marked reduction of A120573 plaquesplaque-associated dystrophic neurites and astrocytosis inmice brain Now we conducted comparative immunizationstudies between EB101 and A120573
42+ FAIFA vaccines in the
prevention ofA120573deposits in themouse brainA120573plaques andvascular amyloid loadswere detected counted andmeasuredas the percent of total surface stained by monoclonal anti-A120573 antibody in the hippocampal and cortical sections Theplaque amyloid load observed in the representative photomi-crographs presented in Figure 2 shows a significant differenceacross the five experimental groups (ANOVA 119865 = 315119875 lt 0001) As a result of the first experimental phase basedon preventive treatment we observed that hippocampal loadwas significantly reduced in the EB101 immunized mice (AFigures 2(a) and 3) when compared with different vaccinecomponents (B-C Figures 2(b)-2(c) and 3) and moderatecompared with A120573
42+ CFAIFA vaccine (D Figures 2(d)
and 3) In fact the load in these treated groups (B-C) wasnot significantly different from PBS treated mice (E Figures2(e) and 3) although there is a positive correlation betweenthe presence of A120573 in treatment regime and the total A120573burden (Figures 2(b) 2(d) and 3) In the EB101 treated micea few compacted A120573 plaques were observed in retrosple-nial granular region of the cortex and at the polymorphdentate gyrus of the hippocampus (Figure 2(a)) Strikinglydifferent was the distribution of A120573 plaques observed inthe other treatment groups where numerous A120573 plaqueswere located at different cortical and hippocampal layers(Figures 2(b)ndash2(e)) This data was also supported by totalA120573 plaques density and size quantification made by imageanalysis software in hippocampal sections stained with anti-A120573 antibody (Figures 3 and 4) The data obtained in thepresent study validate our previous observations demon-strating that the mean burden A120573 plaques (Figures 2(a) and3) of Group A (25 plusmn 5 ps) were significantly different fromthe other treated groups (63 plusmn 6 ps in Figure 2(b) 65 plusmn6 ps in Figure 2(c) 56 plusmn 5 ps in Figure 2(d) 68 plusmn 6 psin Figures 2(e) and 3) This represents a decrease in themean percentage of A120573 plaque area of 301ndash382 in theEB101 immunized mice group relative to other tested groups(Figures 2(a)ndash2(j) and 4)The appearance of amyloid plaquesin the entorhinal (Figures 2(f)ndash2(j)) and piriform cortex wasalso analyzed being detected in low density in Group A(Figure 2(f))moderate density inGroups B (Figure 2(g)) andD (Figure 2(i)) and high density in the entorhinal cortexof Groups C (Figure 2(h)) and E (Figure 2(j)) Thereforethis data shows that in the preventive treatment periodthe clearance effect of the EB101 vaccine on A120573 plaques persection was about 64 (Figures 2(a) and 2(f)) compared topositive controls while in the other treatments regimes 19was detected in A120573
42+ CFAIFA (D) 10 in A120573
42liposome
without S1P (B) and 7 in liposome with S1P treated mice(C) Photomicrographs show that in all groups these plaques
were mainly localized in the external layers of the entorhinalcortex (Figures 2(f)ndash2(j)) In addition we have observedlower stained intensity of the A120573 plaques in the brainregions of EB101 treated mice which contrasts markedlywith the intensely stained A120573-immunoreactivity of theseplaques in each brain section of the other experimental micegroups
32 Efficacy of EB101 versus A12057342
+ CFAIFA in Clearing A120573Plaque Burden In a second set of experiments we comparedthe therapeutic effect of EB101 vaccine with A120573
42+ CFAIFA
in the reduction of A120573 in the brain after the plaques wereestablished in the 35-week-oldAPPPS1 transgenicmice (Fig-ures 2(k)ndash2(t))Themice were treated and analyzed using thesame protocol as in the preventive treatment To determinewhether EB101 vaccine attenuates andor reverses themassivedevelopment of 120573-amyloid plaques brain sections fromwild-type and transgenic mice of all experimental groupswere immunostained with the specific antibody recognizingA1205731ndash42 epitopesThese results show that A120573 deposits (Figures
2(k)ndash2(t)) were notably reduced in brain sections of theEB101 treated mice (A) markedly different from the A120573burden levels observed in the other treated groups (BndashE)As observed in the corresponding photomicrographs EB101treated mice showed a few compacted A120573 plaques mainlylocated at the CA1 layer of the hippocampus (Figure 2(k)) Noreduction in plaque density was observed in the other exper-imental groups (Figures 2(l)ndash2(o)) where numerous largecompacted A120573 plaques were located in almost every layer ofeach brain section The most densely populated section withA120573 plaques were those with no A120573
1ndash42 treatment as shownin Groups C (Figure 2(m)) and E (Figure 2(o)) while a slightreduction of this hallmarks was observed in mice Groups B(Figure 2(l)) and D (Figure 2(n)) The hippocampal sectionsof the therapeutic treatment with EB101 vaccine showed asimilar pattern as that observed in the preventive treatmentThemean burden of A120573 plaques in GroupA (35 plusmn 4 Figure 3)was significantly different from the other treated groups (70plusmn5 in B 74 plusmn 4 in C 65 plusmn 5 in D 80 plusmn 6 in E) The A120573 plaquearea in the EB101 immunized mice group was reduced by297ndash345 relative to the other tested groups (Figure 4) Asobserved in the preventive set of experiments this data showthat in the therapeutic treatment period the clearance effectof the EB101 vaccine on A120573 plaques per section was about59 compared to controls While in mice group treated withA12057342+ CFAIFA the clearance effect was about 20 in Group
D 13 in Group B and 8 in Group C The same pattern ofA120573-immunoreactivity was observed in the entorhinal cortexwhen comparing all experimental treatment groups (Figures2(p)ndash2(t)) Only a few A120573 plaques were observed in theexternal layer of the entorhinal and piriform cortex of micetreated with EB101 (Figure 2(p)) while numerous plaqueswere observed in the corresponding section of Groups B(Figure 2(q)) and D (Figure 2(s)) which show a moderatedensity of A120573 plaques while Groups C (Figure 2(r)) andE (Figure 2(t)) show densely distributed plaques along themedial and external layers of the entorhinal-piriform cortexAs described in the preventive treatment we also detected
BioMed Research International 7
Ect
Cx
CPu
Ect
Gr A
(EB1
01)
(a)
(c)
(b)
(f)
(h)
(g)
Preventive treatment Therapeutic treatment
CA1
PoDGMoDG
CA1
PoDGMoDG
CA1
PoDG
MoDG
DG
DG
Ect
Ect
Cx
Cx
Cx
CPu
CPu
CPuCA1
PoDG
MoDG
DG
CA1
PoDG
MoDG
DG
Ect
Cx
CPu
Ect
Cx
CPu
Cx
Cx
Cx
Cx
Cx
(k)
(l)
(p)
(r)
(q)
CA1
PoDGMoDG
DG
Cx
Ect
Cx
CPuCA1
ControlGr E
(PBS
)
(d)
(e) (j)
Control
PoDG
MoDG
DG
CA1
CA1
PoDG
MoDG
CA1
PoDG
MoDG
DG
DG
Ect
Ect
Cx
Cx
CPu
CPu
CA1
PoDG
MoDG
CA1
PoDGMoDG
DG
Ect
Cx
CPu
PoDGMoDG
DG
Cx
Cx
Cx
(n)
(o)
(s)
(t)
Cx
(i)
Gr B
(A120573
+L)
Gr D
(A120573+
FA)
A120573 A120573 A120573 A120573
A120573A120573A120573
A120573 A120573 A120573 A120573
A120573A120573A120573A120573
A120573 A120573 A120573 A120573
A120573
(m)
Gr C
(S1P
+L)
Figure 2 Comparative effect of EB101 vaccine on beta amyloid deposits Comparative photomicrographs of A120573 immunoreactivity weretaken in the hippocampus ((a)ndash(e) (k)ndash(o)) and cortical ((f)ndash(j) (p)ndash(t)) brain regions of transgenic mice with different treatments beforeA120573 plaques development preventive treatment ((a)ndash(j)) and after the A120573 plaques developed therapeutic treatment ((k)ndash(t)) Preventivetreatment transverse brain sections of 11-month-old mice show reduced number of A120573 deposits in the dentate gyrus hippocampal subregionCA1 (a) and entorhinal cortex (f) following EB101 vaccine immunization (Group A) contrasting with the numerous A120573 immunoreactiveplaques in the correspondent mouse brain sections of Groups B ((b) (g)) C ((c) (h)) D ((d) (i)) and E ((e) (j)) Immunoreactive A120573 in thebrain sections of wild-type (WT) mice is absent (inserts in panels (e) (o)) Therapeutic treatment transverse brain sections of 18-month-oldmice are shown in panels (k)ndash(t) Treatment with EB101 almost completely cleared A120573 load in the dentate gyrus and reduced notably thedensity in hippocampal subregion CA1 (k) and entorhinal cortex (p) compared to the same areas of mouse Groups B ((l) (q)) C ((m)(r)) D((n) (s)) and E ((o) (t)) as determined by number of plaques and staining intensity area Scale bar 100 120583m
8 BioMed Research International
Gr E
(PBS
)
0102030405060708090
Preventive treatmentTherapeutic treatment
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Mea
n of
120573-a
myl
oid
plaq
ues
sect
ion
Gr A
(A120573+
S1P+
L)(E
B101
)
Figure 3 Comparative analysis of A120573 plaque burden in the brainsof APPPS1 mice Mean of A120573 plaque burden in the hippocampaland cortical regions of APPswePS1ΔE9 mice in the five treatmentgroups The mean of the A120573 burden is significantly reduced (119875 lt005) in Group A (EB101) when compared with Groups BndashE duringpreventive treatment and markedly reduced in the therapeutictreatment period Data are presented asmeanplusmn standard error of themean (SEM) Densitymean of A120573 plaques in Tgmice hippocampus
05
1015202530354045
Preventive treatmentTherapeutic treatment
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Gr A
(A120573+
S1P+
L)(E
B101
)
Mea
n of
120573-a
myl
oid
plaq
ues a
rea
Figure 4 Comparative quantification of A120573 burden area in thebrain of APPPS1 mice Quantitative analysis of A120573 burden areain the hippocampal and cortical regions of APPswePS1ΔE9 micetreated with EB101 A120573liposomes S1Pliposomes A120573
42+ FAIFA
and PBS represented by the number of pixels inside the stained areaof each A120573 plaque This graphic shows that A120573 plaques of EB101-treated mice (Group A) are significantly smaller in size (119875 lt 005)than those of the other four treatment groups Data are presented asmean plusmn SEM Mean of A120573 plaques area in Tg mice hippocampus
a reduced staining intensity of the A120573 plaques in the brainregions of mouse Group A contrasting with the intenseA120573-immunoreactivity of plaques in the other experimentalmouse groups as shown in the photomicrographs of Figures2(k)ndash2(t)
33 Immune Response Effects of EB101 versus A12057342+ CFAIFA
Vaccine Astrocyte activation implicated in neuroinflam-mation amyloidogenesis and neuronal cell death in AD
was compared after treatment of the transgenic mice withEB101 and A120573
42+ CFAIFA Immunohistochemical analysis
of GFAP-reactive cells was carried out in the brains ofthe treated mice in the preventive and therapeutic studies(Figure 5) After the preventive treatment (Figures 5(a)ndash5(j))EB101 significantly reduced the density of GFAP-reactive cellclusters in the hippocampal and cortical sections as comparedwith A120573
42+ CFAIFA (D) A120573Lip without S1P (B) Lip with
S1P (C) and PBS (E) The distribution of GFAP immunore-active cells in the transverse section of the mouse brainstreated with EB101 shows a few scattered GFAP-reactiveclusters mainly at the CA1 hippocampal layers contrastingwith numerous dystrophic reactive astrocytes observed indifferent hippocampal areas of mouse brains in Groups BndashE(Figures 5(b)ndash5(e)) Similar neuroinflammation pattern wasobserved in the retrosplenial cortical layers where a fewGFAP-reactive clusters were observed in EB101 treated mice(Figure 5(f)) compared with their profuse presence in theother experimental groups (Figures 5(g)ndash5(j)) After thera-peutic treatment the GFAP immunoreactivity distributionpattern in mouse brains treated with EB101 was similar tocontrol mice (wild-type) mostly devoid of reactive GFAPclusters except a few scattered ones in the CA1 hippocampalexternal layers (Figure 5(k)) In contrast there is a moderatedensity in Group D (Figure 5(n)) and extensive density inGroup B (Figure 5(l)) Group C (Figure 5(m)) and Group Emice (Figure 5(o)) Photomicrographs of retrosplenial cortexshow similar neuroinflammation pattern where a few GFAPclusters detected in the EB101 treated mice (Figure 5(p))contrast with a more extensive density of clusters in the othergroups (Figures 5(q)ndash5(t)) No astrocytosis was observed inwild-type mice during preventive or therapeutic treatment
34 Comparative A120573 Antibody Titers and Th1Th2 CytokineLevels in Treated Mice A120573 antibody titers in the sera weredetermined by ELISA during mouse immunization(Figure 6) Preventive treatment with EB101 resulted in amarked increase of specific IgG A120573
1ndash42 antibody productionin all animals Mice in A and D groups developed andmaintained serum antibody titers between 1 2000 and1 8000 Mice in Groups B C and E showed no significantanti-A120573 antibody production Antibody production and titerin Group B were three and four times lower than those ofGroups D and A respectively After therapeutic treatment allmice treated with EB101 and A120573
42+ FAIFA produced high
levels of IgG antibodies Titer levels of Groups A and D weresignificantly increased to those detected after preventivetreatment (119875 lt 001 versus Group D 119875 lt 0001 versusGroups B C and E) although antibody level in Group D was13-fold lower than in Group A (Figure 6)The effect of EB101(Group A) and A120573
42+ CFAIFA (Group B) vaccines on Th1
andTh2 cytokines was also studied (Figure 7)Measurementsof Th1 and Th2 cytokines in sera of immunized transgenicmice indicate that significant changes occurring in proteinlevels of Th1-related inflammatory mediators followingpreventive (Figures 7(a)ndash7(c)) and therapeutic (Figures 7(d)ndash7(g)) treatment with EB101 vaccine A significant increaseof Th1 cytokine profile was observed in preventive treatment
BioMed Research International 9
Gr E
(PBS
)
(d)
(e)
(i)
(j)
(n)
(o)
(s)
(t)
Cx
GFAP
GFAP GFAP GFAP GFAP
GFAP GFAP GFAP
Cx Cx
Cx
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
GFAP
Gr A
(EB1
01)
(a)
(b)
(c)
(f)
(g)
(h)
Preventive treatment Therapeutic treatment
PoDG
MoDG
(k)
(l)
(m)
(p)
(q)
(r)DG
GFAP GFAP GFAP
GFAPGFAPGFAP GFAP
GFAP GFAP GFAP GFAP
PoDGDG
DG
CA1
Cx
Cx
Cx
Cx
Cx
Cx
PoDG
MoDG
DG
CA1
MoDGDG
CA1
PoDG
MoDG
DG
CA1
Cx
CxCx
MoDG
MoDG
PoDG
CA1CA1
Cx
Gr D
(A120573+
FA)
Gr C
(S1P
+L)
Gr B
(A120573+
L)
Figure 5 Comparative effect of EB101 vaccine on astrocytosis Comparative photomicrographs of GFAP immunoreactivity in thehippocampus ((a)ndash(e) (k)ndash(o)) and cortical ((f)ndash(j) (p)ndash(t)) brain regions at the preventive treatment phase ((a)ndash(j)) and therapeutictreatment group ((k)ndash(t)) Preventive treatment transverse brain sections of 11-month-old mice treated with EB101 show an almost completeprevention of astrocytosis in the detailed sections of the dentate gyrus (a) contrasting with numerous dystrophic reactive astrocytes typicalof an inflammatory reaction observed in different hippocampal corresponding areas of mouse brains in Group B (A120573Lip-treated mice (b))Group C (LipS1P-treated mice (c)) Group D (A120573
42+ CFAIFA-treated mice (d)) and Group E (PBS-treated mice (e)) Immunoreactive
astrocytosis in the brain sections of WT mice is absent (data not shown) Detailed sections of the parietal cortex of treated mice show agradient density of reactive astrocytes immunoreactive to GFAP indicating a neuroinflammation pattern Comparative cortical sections shownumerous astrocytosis areas inmice Groups C and E ((h) (j)) andwide reactive astrocytes clusters inmice Groups B andD ((g) (i)) while justa few of them are observed in Group A (f)Therapeutic treatment transverse brain sections of 18-month-old mice of Group A show a notablereduction of astrocytosis in the hippocampal region (k) after EB101 vaccine immunization Note the astrocytosis contrast density betweenEB101-treatedmice and the other treated groups ((l)ndash(o)) where numerous immunoreactive GFAP clusters are observed in the correspondentbrain sections A few immunoreactive GFAP clusters are observed in the parietal cortex of EB101 treated mice group while a moderate ((q)(s)) to high ((r) (t)) density of these neuropathological inflammation clusters is apparent in the other treatment groups (Groups BndashE) Forabbreviations see list Scale bar 100120583m
10 BioMed Research International
002040608
1121416
Preventive treatmentTherapeutic treatment
Ant
i-120573-a
myl
oid
IgG
antib
odie
s
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Gr A
(A120573+
S1P+
L)(E
B101
)
Figure 6 Quantification of specific anti-120573-amyloid IgG in immu-nized APPPS1 mice The presence of anti-120573-amyloid antibodiesmeasured in the sera of vaccinated and control mice groups duringthe preventive and therapeutic treatments indicates that EB101liposome vaccine was able to induce the highest production ofspecific IgG antibodies Compared to theGroupD immunizedmicethe level of antibodies observed in the Group A was also highersuggesting a more effective immune response (119875 lt 001 versusGroup D 119875 lt 0001 versus Groups B C and E) Quantification ofspecific anti-120573-amyloid IgG in immunized Tg mice
in Group D with respect to all other groups mainlyreported by the high levels of TNF-120572 (Figure 7(b)) andIFN-120574 (Figure 7(c)) cytokine secretions while due to thehigh mortality rate during the therapeutic treatment nosignificant data was obtained from this mice group Inaddition a tendency toTh1 secretion reduction was observedinGroupAwith respect to all other groups in both preventiveand therapeutic treatment The changes in Th2 protein levels(Figure 7(d)) followed the inverse trend where the highestdifferences between Group A and the other treatment groupswere observed in the IL-4 IL-5 and IL-10 cytokine secretions(119875 lt 005 versus Group D 119875 lt 001 versus Group E Figures7(e)ndash7(g))
35 Comparative Motor Coordination and Memory Perfor-mances In order to validate the locomotion integrity duringthe entire immunotherapeutic process as reported in similarprevious studies motor coordination tests have showed thatEB101-treated mice spent significantly more time on theaccelerating rotarodThismotor coordination task shows thatEB101 immunization affected positively their performance(Figure 8(a)) A120573
42liposome (B) S1P-containing liposomes
(C) and PBS (E) (119875 lt 0006) mice performed poorly incomparison to EB101 mice (119875 lt 005) or A120573
42+ CFAIFA
mice (119875 lt 002) in motor abilities and coordination Thefear conditioning-active avoidance test was used to assess thespatial learning and memory functions As expected bothEB101 and A120573
42+ CFAIFA immunized mice readily learned
the location of the platform as shown by a decrease in theescape latency over training days while the control group
mice exhibited no significant changes during 6-day trialsThe average escape latencies of EB101 (68 plusmn 2 s 761 plusmn 22 s)andA120573
42+CFAIFA (116plusmn33 s 123plusmn21 s) immunizedmice
at the end of preventive and therapeutic periods respectivelyduring the consecutive 6 days trial were significantly shorterthan the other treatment groups (A120573
42liposome 163 plusmn 31 s
175 plusmn 28 s S1P-containing liposomes 202 plusmn 19 s 248 plusmn27 s) and the control (2116 plusmn 38 s 268 plusmn 22 s) (119875 lt 001)Therewas no significant difference between EB101 andA120573
42+
CFAIFA immunized group (119875 gt 005) (Figure 8(b)) Theseresults showed that spatial learning was improved in theseimmunized mice
4 Discussion
In this study we have compared a novel active A120573 imm-unotherapy the EB101 vaccine against A120573 with the A120573
42+
CFAIFA vaccine that was first taken into clinical trialswith coadjuvant Qs-21 modification (AN1792 by Orgogozoet al [19]) This type of vaccine design was thought toinduce a strong T-cell mediated inflammatory response (IL-2 and IFN-c positive responses indicative of a Class II(CD4) Th-1 type response [33] leading to acute menin-goencephalitis in the 6 of the patients) The EB101 activeimmunotherapeutic vaccine was primarily designed to avoidundesired inflammatory side effects both in mice andhumans (without need of coadjuvant modification) whileinducing effective antibody titer rates We demonstrated thatlong-term immunization with EB101 induced A120573 antibodiesin the preventive and the therapeutic treatment periodswhich effectively reduced amyloid deposition in both corticaland hippocampal brain regions and prevented neuroinflam-mation probably by blocking toxicity in the surroundingneurons However the AN1792 vaccine was shown to inducein 6 of the patients an inflammatory reaction that mighthave led to the encephalitis type reaction observed in theclinical trial which was responsible for the immediate haltin those clinical studies It seems that AN1792 immunizationdelivered in Qs-21 and polysorbate 80 adjuvant induced aTh1 type response that likely led to autoaggressive T cells andaseptic meningoencephalitis In the present experiments wedemonstrated that a judiciously selected adjuvant produces adifferent humoral response [22] Indeed we have shown thatour novel immunotherapeutic strategy boosts specific A120573
1ndash42antibody generation and induces a Th2 noninflammatoryimmune response This boosted effect is in accordance withprevious immunotherapeutic studies [34] Taking togetherall the present findings these results suggest that EB101vaccine can achieve in a future clinical trial immunogenicresponse rates quite similar to the AN1792 without theinflammatory effects reported The immunohistochemicalanalysis obtained in the present study shows that all the EB101immunized mice resulted in an almost complete preventionof A120573 deposition in the brain with only a few compactA120573 plaques in the retrosplenial and dentate gyrus regionsAlthough there was a slightly increase in death in EB101treated mice compared to controls this significant reductionof A120573 plaques in EB101 treated mice was proven to be more
BioMed Research International 11
Gr A Gr B Gr C Gr D Gr EPreventive treatmentTherapeutic treatment
0005001000150020002500300035004000
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
(a) Th1
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
000
1000
2000
3000
4000
5000TNF-120572
(b) Th1
000500
10001500200025003000
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
IFN-120574
(c) Th1
000500
100015002000250030003500
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
Group A Group B Group C Group D Group EPreventive treatmentTherapeutic treatment
(d) Th2
000
500
1000
1500
2000
2500
3000IL-4
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(e) Th2
000500
100015002000250030003500
IL-5
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(f) Th2
3500IL-10
000500
10001500200025003000
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(g) Th2
Figure 7 Detection of Th1 and Th2 cytokine profile Measurements of the effect of EB101 and A12057342
+ CFAIFA vaccines on Th1 and Th2cytokines profile during preventive and therapeutic treatments ((a)ndash(c)) A significant increase of Th1 cytokine profile was observed inpreventive treatment in Group D with respect to all other groups while a tendency to a reduction inTh1 secretion was observed in Group Awith respect to all other groups in both preventive and therapeutic treatment ((d)ndash(g)) The changes in Th2 protein levels were observed inthe IL-4 IL-5 and IL-10 cytokine secretions of Group A (119875 lt 005 versus Group D 119875 lt 001 versus Group E)
12 BioMed Research International
Gr A
(EB1
01)
05
101520253035
Preventive treatment Therapeutic treatment
Mea
n fa
lling
late
ncy
(s)
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)(a) Graphic of motor coordination and balance
05
1015202530
Mea
n re
spon
se ti
me (
s)
Gr A
(EB1
01)
Preventive treatment Therapeutic treatment
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
(b) Graphic of associative learning and memory
Figure 8 Analysis of motor coordination and balance (a) Rotarod results of motor coordination and balance test after the preventive andtherapeutic treatment showing improved effect of EB101 and A120573
42+ CFAIFA-vaccine compared to other treated mice groups (119875 lt 005)
and (119875 lt 002) versus control mice Latency to fall on the rotarod apparatus was significantly improved in EB101 immunized mice Valuesrepresent means of all six trials of a groupday (b) Spatial learning and memory tested by fear conditioning-active avoidance test The meanresponse time of each treatment group to reach the platform is represented EB101 and A120573
42+ CFAIFA immunized mice had significantly
lower latencies than the other treatment groups (119875 lt 001) versus control mice
efficient than that obtained with A12057342+ CFAIFA immuniza-
tions A comparison of A120573 plaque reduction rates amongtreated groups surprisingly showed that other than EB101and A120573
42+ CFAIFA treated groups some amyloid deposits
were also reduced in Groups B (A1205731ndash42liposome) and C
(S1P-containing liposomes) Although the reduction rates inthese mice were markedly low the subtle lowering effectmight be due to the interaction between the immunizationelements (A120573
1ndash42 and S1P) and the mouse immunogenicsystem as reported by others [35] Based on our presentresults S1P one of the components of the adjuvant used withEB101 might be responsible for the neuroprotective effectobserved in the vulnerable neuronal areas of EB101 treatedmice The advantage in the A120573 load preventionreduction bycombining A120573
1ndash42 and S1P in a unique formulation vaccinehas been also observed in the reduced hippocampal load ofEB101 immunizedmice (Gr A) when comparedwith differentvaccine components (Gr B-C) In fact S1P appears to have ademonstrated therapeutic potential as a regenerative agent inthe nervous system since it controls migration of neuronalstem cells toward a site of spinal cord injury [36] inducecytoskeletal rearrangements to promote transmitter secretion[37] and also trigger diverse cellular effects including angio-genesis cardiac development immunity cell motility andneurite extension [38 39]Therefore it is reasonable to expectthat such a potent biologically active sphingolipid by itselfmay play an important role in the A120573 plaque reduction Inthis study the clearance efficiency rate of each treatment wasanalyzed and results suggested that EB101 (6459) appearsto be more effective than A120573
42+ CFAIFA (1920) vaccine
in both preventive and therapeutic groups The effectivenessof EB101 in older transgenic animals (18 months) showeda clearance effect of 59 suggesting that EB101 may alsohave therapeutic potential inADdiagnosticated patient trials
Similar conclusion was reached byWang and colleagues [40]who demonstrated that combination therapy in aged animalsprovides more valuable insights into a possible translationto humansOur immunologicalmeasurements demonstratedonce again that neuroinflammation develops in an A120573-dependent fashion early in the course of AD [41] and canbe alleviated or reversed by interventional A120573 vaccinationsuch as EB101 Among the possible mechanisms of A120573clearance proposed in AD immunotherapy [42] our resultssuggested that theymay overlap and probably be disease statedependent since the prevention and the therapeutic vaccinewould likely benefit from the transport of A120573 antibodies intothe CNS However some mechanisms of action such as thepreventive effect of A120573 antibodies in the A120573 aggregation [43]microglial phagocytosis induced by the Fc binding portionsbetween A120573 antibodies and microglia receptor [44] and theshift in the gradient of A120573 transport across the blood-brainbarrier due to the increased concentration of A120573 antibodiesin the vascular system resulting in an increase in effluxfrom the brain to blood [45 46] may be playing a role inboth preventive and therapeutic conditions Our findingstherefore suggest that novel treatment strategies aimed atmaintaining or increasing microglial function and restoringthe balance to the immunological system may represent anattractive therapeutic approach even at the advanced stagesof ADThe immunization results obtained with EB101 duringboth preventive and therapeutic treatment showed nomicro-hemorrhages infiltration of lymphocytes or encephalitis-like reaction in the brain suggesting that EB101 does notinduce A120573-specific Th1 reaction possibly due to the neu-roprotective effect of S1P Delivering A120573
1ndash42 in a liposomal-S1P-adjuvant resulted in an effective immunological responseincluding the induction of a Th2 T-cell response whicheffectively prevented neuroinflammation in an animal model
BioMed Research International 13
of AD as reported previously [24 47] Moreover presentresults demonstrated that EB101 did not induce astrogliosisin APPPS1 transgenic mice whereas some GFAP-positiveastrocyte clusters were aggregated in the affected brain areasof mice treated with A120573
42+ CFAIFA These results indicate
that EB101 may result in an effective treatment in AD neu-ropathology due to the absence of peripheral proinflamma-tory cytokine secretion and the absence of astroglial activa-tion in the brains of EB101 treated mice when compared withA12057342
+ CFAIFA immunotherapeutic response In a clinicalsetting however we may expect a wide differential responsein part depending upon the genomic profile of each patient[48 49] In AD behavioural studies in particular amongthe motor coordination tests the rotarod performance hasbeen reported to be impaired in some AD transgenic micemodels bearing A120573 plaques [50 51] This result was found inmutants prior to A120573 plaque formation at 3 and 6 months ofage but was not reproduced after plaques had been formedat 16 and 24 months of age [52] perhaps because of thedeclining abilities of older normal mice Here we presentthe effects of vaccine treatments in APPPS1 mice on motorcoordination a neglected aspect of the AD experimentalbehavior but justified on the basis of the deficient posturalcontrol reported in patients with Alzheimerrsquos disease [53]sometimes even during early stages [54] Our longitudinalmotor behavioral results showed no significant differencesin the ambulatory movement grip strength or coordinationaddressed by their performance in the rotating rod betweenAPPPS1 and wild-type mice for any age group studiedin contrast with other treated groups demonstrating thatlongitudinal motor assessments of AD transgenic mousemodels can provide valuable information with regard tothe effect of immunization on the progression of behavioraldeficits and overall motor activity As shown the EB101-treated group of mice exhibited longer latencies and betterbalancing skills indicating the absence of any negative effectof EB101 immunization on these mice The classical fearconditioning test is assumed to reveal the functional integrityof the hippocampus [55] representing a special memorytask to evaluate the AD models and its improvement afterimmunization periods In this APPPS1 mouse model thefirst associative learning and spatial workingmemory deficitsare shown at 5-6 months of age [56] although its impair-ment progression is relatively slow compared to most othermodels At the end of preventive immunization period EB101and A120573
42+ CFAIFA immunized mice have significantly
shorter escape latencies than the other treatment groups(A12057342liposome S1P-containing liposomes) and the controls
in the fear conditioning test Shorter escape latencies duringspatial learning indicated that deficits of spatial memorywere attenuated in EB101 and A120573
42+ CFAIFA immunized
mice Previous studies have showed that by 15 months ofage the spatial working memory is consistently impairedthroughout the rest of the life span [31 57] At the end oftherapeutic immunization period both EB101 and A120573
42+
CFAIFA immunized mice showed significantly improvedresults on the hippocampus-dependent spatial learning andmemory test when compared with other treatment groupsand the controls Taken together these observations are
in agreement with the impairment in conditioned learningin response to a tone stimulus of aged APPPS1 micealthough when immunized with EB101 during preventiveor therapeutic periods such impairments are significantlyattenuated
In summary the present results corroborate our previouspublished findings showing that EB101 immunization reducesamyloid accumulation in the APPPS1 mouse brain [2425] The strongest reduction in the preventive immunizationwas observed in the retrosplenial cortex followed by thehippocampus and entorhinal-piriform cortex Our resultsfurther support that EB101 ismore effectivewhen themice areimmunized prior to developing plaques in the hippocampaland cortical brain regions There is extensive evidence inmouse preclinical studies [58] and human clinical trials [5960] that the adverse effects of active and passive A120573 vaccinesresult in microhemorrhages and encephalitic reactions [60ndash63] yet themechanismbywhich they occur has not been fullyelucidated In the treatment sections analyzed we found nomicrohemorrhages or acute inflammatory processes in all thesurvived mice of the group indicating that EB101 therapeuticapproach presents no adverse inflammatory effects in thisAD mice models Thus our present results unequivocallydemonstrate that EB101 vaccinemay not only halt AD-relatedpathology as a preventive treatment but may also reverse it inestablished cases related to the improvement of the learningandmemory in both cases leading to a robust immunologicaltherapeutic response Present experiments also demonstratethat EB101 exerts more significant and substantial effects onthe reduction of A120573 deposition than A120573
42+CFAIFA vaccine
in A120573 mice models of AD However further exploration ofthe contribution of genetic environmental and physiologicalfactors in the development resistance to endogenous A120573-antibody is needed in order to optimize and modulate thetherapeutic strategies currently being developed
5 Conclusions
The present study carried out in APPPS1 transgenic mousemodel of AD compared the A120573 burden reduction andcellular immune responses produced by two immunizationdelivery strategies of A120573 in Freund adjuvant (described byWebster et al [10]) and in liposomeS1P matrix (describedby Schenk et al [17])The EB101 immunotherapeutic strategyboosted antibody generation A120573 plaque reduction and ananti-inflammatory response more effectively than A120573
42+
CFAIFA especially if given prior to or in the early stages ofpathology The present results also showed that vaccinationwith EB101 prevented the formation of new plaques andpresumably led to a reduction in established plaques whileinducing aTh2 immune reactionwithout encephalitogenic T-cell responses or brain inflammation related with astrogliosisHowever despite the efficacy demonstrated of this novelvaccine approach further preclinical and clinical studieson EB101 are warranted to evaluate its efficacy and themechanism for the future utility of such vaccine for ADtherapy
14 BioMed Research International
List of Abbreviations
AD Alzheimerrsquos diseaseA120573 120573-Amyloid proteinAPP Amyloid precursor proteinAPT Anterior pretectal nucleusCA1 Field CA1 hippocampusCA3 Field CA3 hippocampusCPu Caudate putamen (striatum)Cx CortexDG Dentate gyrusDN Dystrophic neuriteEct Entorhinal cortexCFA Complete Freundrsquos adjuvantGrDG Granular dentate gyrusIFA Incomplete Freundrsquos adjuvantIL InterleukinLPMR Lateral posterior thalamic nucleus (mediorostral)MoDG Molecular dentate gyrusPir Piriform cortexPoDG Polymorph dentate gyrusRSGb Retrosplenial granular cortex b regionTg TransgenicTh T helper
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Ivan Carrera Ignacio Etcheverrıa Lucıa Fernandez-NovoaValter Lombardi Ramon Cacabelos and Carmen Vigo con-ceived and designed the experiments Ivan Carrera and Igna-cio Etcheverrıa performed the experiments Ivan CarreraIgnacio Etcheverrıa and Valter Lombardi analyzed the dataIvan Carrera Ignacio Etcheverrıa Lucıa Fernandez-Novoaand Valter Lombardi contributed reagentsmaterialsanalysistools Ivan Carrera Carmen Vigo andMadepalli KrishnappaLakshmana wrote the paper
Acknowledgments
This work was supported by National Institute of Aging(NIA)NIH Grant no 1R01AG036859-01 (Madepalli Krish-nappa Lakshmana) and the financial support of the ProgramaOperativo Feder Galicia 2007ndash2013 (IN841C 201254 to PhDCarrera I)
References
[1] R Cacabelos L Fernandez-Novoa V Lombardi Y Kubotaand M Takeda ldquoMolecular genetics of Alzheimerrsquos diseaseand agingrdquoMethods and Findings in Experimental and ClinicalPharmacology vol 27 pp 1ndash573 2005
[2] P T Nelson I Alafuzoff E H Bigio et al ldquoCorrelationof alzheimer disease neuropathologic changes with cognitive
status a review of the literaturerdquo Journal of Neuropathology andExperimental Neurology vol 71 no 5 pp 362ndash381 2012
[3] JNaslundVHaroutunian RMohs et al ldquoCorrelation betweenelevated levels of amyloid 120573-peptide in the brain and cognitivedeclinerdquo Journal of the American Medical Association vol 283no 12 pp 1571ndash1577 2000
[4] A Koyama O I Okereke T Yang D Blacker D J Selkoeand F Grodstein ldquoPlasma amyloid-120573 as a predictor of dementiaand cognitive decline a systematic review and meta-analysisrdquoArchives of Neurology vol 69 no 7 pp 824ndash831 2012
[5] A M Fagan C M Roe C Xiong M A Mintun J C Morrisand D M Holtzman ldquoCerebrospinal fluid tau120573-amyloid42ratio as a prediction of cognitive decline in nondemented olderadultsrdquo Archives of Neurology vol 64 no 3 pp 343ndash349 2007
[6] S Parvathy P Davies V Haroutunian et al ldquoCorrelationbetween A120573-40- A120573-42- and A120573x-43-containing amyloidplaques and cognitive declinerdquo Archives of Neurology vol 58no 12 pp 2025ndash2032 2001
[7] E Karran M Mercken and B D Strooper ldquoThe amyloidcascade hypothesis for Alzheimerrsquos disease an appraisal for thedevelopment of therapeuticsrdquo Nature Reviews Drug Discoveryvol 10 no 9 pp 698ndash712 2011
[8] J L Jankowsky D J Fadale J Anderson et al ldquoMutantpresenilins specifically elevate the levels of the 42 residue 120573-amyloid peptide in vivo evidence for augmentation of a 42-specific 120574 secretaserdquo Human Molecular Genetics vol 13 no 2pp 159ndash170 2004
[9] L Holcomb M N Gordon E Mcgowan et al ldquoAcceleratedAlzheimer-type phenotype in transgenic mice carrying bothmutant amyloid precursor protein and presenilin 1 transgenesrdquoNature Medicine vol 4 no 1 pp 97ndash100 1998
[10] S JWebster A D Bachstetter and L J van Eldik ldquoComprehen-sive behavioral characterization of an APPPS-1 double knock-in mouse model of Alzheimerrsquos diseaserdquo Alzheimerrsquos ResearchandTherapy vol 5 no 3 article 28 2013
[11] H Xiong D Callaghan J Wodzinska et al ldquoBiochemical andbehavioral characterization of the double transgenic mousemodel (APPswePS1dE9) of Alzheimerrsquos diseaserdquo NeuroscienceBulletin vol 27 no 4 pp 221ndash232 2011
[12] M Garcia-Alloza L A Borrelli B T Hyman and B J BacskaildquoAntioxidants have a rapid and long-lasting effect on neuriticabnormalities in APPPS1 micerdquo Neurobiology of Aging vol 31no 12 pp 2058ndash2068 2010
[13] M Meyer-Luehmann M Mielke T L Spires-Jones et al ldquoAreporter of local dendritic translocation shows plaque-relatedloss of neural system function in APP-transgenic micerdquo Journalof Neuroscience vol 29 no 40 pp 12636ndash12640 2009
[14] E Machova V Rudajev H Smyckova H Koivisto H Tanilaand V Dolezal ldquoFunctional cholinergic damage develops withamyloid accumulation in young adult APPswePS1dE9 trans-genic micerdquo Neurobiology of Disease vol 38 no 1 pp 27ndash352010
[15] T Malm J Koistinaho and K Kanninen ldquoUtilization of APP-swePS1dE9 transgenic mice in research of Alzheimerrsquos diseasefocus on gene therapy and cell-based therapy applicationsrdquoInternational Journal of Alzheimerrsquos Disease vol 2011 Article ID517160 8 pages 2011
[16] J Lee and P Han ldquoAn update of animal models of alzheimerdisease with a reevaluation of plaque depositionsrdquoExperimentalNeurobiology vol 22 pp 84ndash95 2013
BioMed Research International 15
[17] D Schenk R Barbour W Dunn et al ldquoImmunization withamyloid-120573 attenuates Alzheimer disease-like pathology in thePDAPP mouserdquo Nature vol 400 no 6740 pp 173ndash177 1999
[18] H J Fu B Liu J L Frost and C A Lemere ldquoAmyloid-120573immunotherapy for Alzheimerrsquos diseaserdquoCNS and NeurologicalDisorders-Drug Targets vol 9 no 2 pp 197ndash206 2010
[19] J-M Orgogozo S Gilman J-F Dartigues et al ldquoSubacutemeningoencephalitis in a subset of patients with AD after A120573
42
immunizationrdquo Neurology vol 61 no 1 pp 46ndash54 2003[20] J A R Nicolll DWilkinson CHolmes P Steart HMarkham
andROWeller ldquoNeuropathology of humanAlzheimer diseaseafter immunization with amyloid-120573 peptide a case reportrdquoNature Medicine vol 9 no 4 pp 448ndash452 2003
[21] I Ferrer M Boada Rovira M L Sanchez Guerra M JRey and F Costa-Jussa ldquoNeuropathology and pathogenesisof encephalitis following amyloid-Neuropathology and patho-genesis of encephalitis following amyloid-beta immunizationin Alzheimerrsquos disease immunization in Alzheimerrsquos diseaserdquoBrain Pathology vol 14 no 1 pp 11ndash20 2004
[22] D H Cribbs A Ghochikyan V Vasilevko et al ldquoAdjuvant-dependent modulation ofTh1 andTh2 responses to immuniza-tion with 120573-amyloidrdquo International Immunology vol 15 no 4pp 505ndash514 2003
[23] C Holmes D Boche D Wilkinson et al ldquoLong-term effectsof A120573
42immunisation in Alzheimerrsquos disease follow-up of a
randomised placebo-controlled phase I trialrdquo The Lancet vol372 no 9634 pp 216ndash223 2008
[24] I Carrera I Etcheverrıa L Fernandez-Novoa V LombardiR Cacabelos and C Vigo ldquoVaccine development to treatAlzheimerrsquos disease neuropathology in APPPS1 transgenicmicerdquo International Journal of Alzheimerrsquos Disease vol 2012Article ID 376138 17 pages 2012
[25] I Carrera I Etcheverrıa Y Li et al ldquoImmunocytochemicalcharacterization of Alzheimer disease hallmarks in APPPS1transgenic mice treated with a new anti-amyloid-120573 vaccinerdquoBioMed Research International vol 2013 Article ID 709145 12pages 2013
[26] V Anelli R Bassi G Tettamanti P Viani and L Riboni ldquoExtra-cellular release of newly synthesized sphingosine-1-phosphateby cerebellar granule cells and astrocytesrdquo Journal of Neuro-chemistry vol 92 no 5 pp 1204ndash1215 2005
[27] S Schroeter K Khan R Barbour et al ldquoImmunotherapyreduces vascular amyloid-120573 in PDAPP micerdquo Journal of Neu-roscience vol 28 no 27 pp 6787ndash6793 2008
[28] Z Suo A Cox N Bartelli et al ldquoGRK5 deficiency leads to earlyAlzheimer-like pathology and working memory impairmentrdquoNeurobiology of Aging vol 28 no 12 pp 1873ndash1888 2007
[29] W Kamphuis C Mamber MMoeton et al ldquoGFAP isoforms inadult mouse brain with a focus on neurogenic astrocytes andreactive astrogliosis in mouse models of Alzheimer diseaserdquoPLoS ONE vol 7 no 8 Article ID e42823 2012
[30] J Lang C Vigo-Pelfrey and F Martin ldquoLiposomes composedof partially hydrogenated egg phosphatidylcholines fatty acidcomposition thermal phase behavior and oxidative stabilityrdquoChemistry and Physics of Lipids vol 53 no 1 pp 91ndash101 1990
[31] D Morgan D M Diamond P E Gottschall et al ldquoA 120573peptide vaccination prevents memory loss in an animal modelof Alzheimerrsquos diseaserdquo Nature vol 408 no 6815 pp 982ndash9852000
[32] M L Brownlow L Benner D DrsquoAgostino M N Gordon andD Morgan ldquoKetogenic diet improves motor performance but
not cognition in two mouse models of Alzheimerrsquos pathologyrdquoPLoS ONE vol 8 no 9 Article ID e75713 2013
[33] M Pride P Seubert M Grundman M Hagen J Eldridgeand R S Black ldquoProgress in the active immunotherapeu-tic approach to Alzheimerrsquos disease clinical investigationsinto AN1792-associated meningoencephalitisrdquo Neurodegenera-tive Diseases vol 5 no 3-4 pp 194ndash196 2008
[34] CWiessner K-HWiederhold A C Tissot et al ldquoThe second-generation activeA120573 immunotherapyCAD106 reduces amyloidaccumulation in APP transgenic mice while minimizing poten-tial side effectsrdquoThe Journal of Neuroscience vol 31 no 25 pp9323ndash9331 2011
[35] D Lambracht-Washington and R Rosenberg ldquoAdvances in thedevelopment of vaccines for Alzheimerrsquos diseaserdquo DiscoveryMedicine vol 15 no 84 pp 319ndash326 2013
[36] A Kimura T Ohmori R Ohkawa et al ldquoEssential roles ofsphingosine 1-phosphateS1P
1receptor axis in the migration of
neural stem cells toward a site of spinal cord injuryrdquo Stem Cellsvol 25 no 1 pp 115ndash124 2007
[37] S S Lee C H Woo J D Chang and J H Kim ldquoRoles of Racand cytosolic phospholipase A2 in the intracellular signalling inresponse to titanium particlesrdquo Cellular Signalling vol 15 no 3pp 339ndash345 2003
[38] E Masliah L Hansen A Adame et al ldquoA120573 vaccination effectson plaque pathology in the absence of encephalitis inAlzheimerdiseaserdquo Neurology vol 64 no 1 pp 129ndash131 2005
[39] A Pilorget B Annabi F Bouzeghrane J Marvaldi J Luisand R Beliveau ldquoInhibition of angiogenic properties of brainendothelial cells by platelet-derived sphingosine-1-phosphaterdquoJournal of Cerebral Blood Flow and Metabolism vol 25 no 9pp 1171ndash1182 2005
[40] A Wang P Das R C Switzer III T E Golde and J LJankowsky ldquoRobust amyloid clearance in a mouse model ofAlzheimerrsquos disease provides novel insights into themechanismof amyloid-120573 immunotherapyrdquo Journal of Neuroscience vol 31no 11 pp 4124ndash4136 2011
[41] G Krabbe A Halle V Matyash et al ldquoFunctional impairmentof microglia coincides with Beta-amyloid deposition in micewith Alzheimer-like pathologyrdquo PLoS ONE vol 8 no 4 ArticleID e60921 2013
[42] Y H Liu B Giunta H-D Zhou J Tan and Y J WangldquoImmunotherapy for Alzheimer disease-the challenge ofadverse effectsrdquo Nature Reviews Neurology vol 8 no 8 pp465ndash469 2012
[43] B Solomon R Koppel D Frankel and E Hanan-AharonldquoDisaggregation of Alzheimer120573-amyloid by site-directedmAbrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 94 no 8 pp 4109ndash4112 1997
[44] F Bard C Cannon R Barbour et al ldquoPeripherally adminis-tered antibodies against amyloid 120573-peptide enter the centralnervous system and reduce pathology in a mouse model ofAlzheimer diseaserdquo Nature Medicine vol 6 no 8 pp 916ndash9192000
[45] R B DeMattos K R Bales D J Cummins J C Dodart S MPaul and D M Holtzman ldquoPeripheral anti-A120573 antibody altersCNS and plasma A120573 clearance and decreases brain A120573 burdenin a mouse model of Alzheimerrsquos diseaserdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 98 no 15 pp 8850ndash8855 2001
[46] C A Lemere E T Spooner J LaFrancois et al ldquoEvidence forperipheral clearance of cerebral A120573 protein following chronic
16 BioMed Research International
active A120573 immunization in PSAPP micerdquo Neurobiology ofDisease vol 14 no 1 pp 10ndash18 2003
[47] T Town M Vendrame A Patel et al ldquoReduced Th1 andenhanced Th2 immunity after immunization with Alzheimerrsquos120573-amyloid
1minus42rdquo Journal of Neuroimmunology vol 132 no 1-2
pp 49ndash59 2002[48] R Cacabelos ldquoPharmacogenomics in Alzheimerrsquos diseaserdquo
Methods in Molecular Biology vol 448 pp 213ndash357 2008[49] R Cacabelos ldquoPharmacogenomics and therapeutic strategies
for dementiardquo Expert Review of Molecular Diagnostics vol 9no 6 pp 567ndash611 2009
[50] D van Dam R DrsquoHooge M Staufenbiel C van Ginneken Fvan Meir and P P de Deyn ldquoAge-dependent cognitive declinein the APP23 model precedes amyloid depositionrdquo EuropeanJournal of Neuroscience vol 17 no 2 pp 388ndash396 2003
[51] M Dumont C Strazielle M Staufenbiel and R LalondeldquoSpatial learning and exploration of environmental stimuli in24-month-old female APP23 transgenic mice with the Swedishmutationrdquo Brain Research vol 1024 no 1-2 pp 113ndash121 2004
[52] R Lalonde M Dumont M Staufenbiel C Sturchler-Pierratand C Strazielle ldquoSpatial learning exploration anxiety andmotor coordination in female APP23 transgenic mice with theSwedish mutationrdquo Brain Research vol 956 no 1 pp 36ndash442002
[53] F J Huff F Boller F Lucchelli R Querriera J Beyer and SBelle ldquoThe neurologic examination in patients with probableAlzheimerrsquos diseaserdquo Archives of Neurology vol 44 no 9 pp929ndash932 1987
[54] A F Pettersson M Engardt and L O Wahlund ldquoActivitylevel and balance in subjects with mild Alzheimerrsquos diseaserdquoDementia and Geriatric Cognitive Disorders vol 13 no 4 pp213ndash216 2002
[55] L Calandreau B Desgranges R Jaffard and A DesmedtldquoSwitching from contextual to tone fear conditioning and viceversa the key role of the glutamatergic hippocampal-lateralseptal neurotransmissionrdquo Learning amp Memory vol 17 no 9pp 440ndash443 2010
[56] F Trinchese S Liu F Battaglia SWalter PMMathews andOArancio ldquoProgressive age-related development of Alzheimer-like pathology in APPPS1 micerdquo Annals of Neurology vol 55no 6 pp 801ndash814 2004
[57] D M Wilcock A Rojiani A Rosenthal et al ldquoPassiveimmunotherapy against A120573 in aged APP-transgenic micereverses cognitive deficits and depletes parenchymal amyloiddeposits in spite of increased vascular amyloid and microhem-orrhagerdquo Journal of Neuroinflammation vol 1 article 24 2004
[58] S A Ferguson S Sarkar and L C Schmued ldquoLongitudinalbehavioral changes in the APPPS1 transgenic AlzheimerrsquosDisease modelrdquo Behavioural Brain Research vol 242 no 1 pp125ndash134 2013
[59] D Wilcock and C Colton ldquoImmunotherapy vascular pathol-ogy and microhemorrhages in transgenic micerdquo CNS andNeurological DisordersmdashDrug Targets vol 8 no 1 pp 50ndash642009
[60] M Grundman S Gilman R S Black et al ldquoAn alternativemethod for estimating efficacy of the AN1792 vaccine foralzheimer diseaserdquo Neurology vol 71 no 9 pp 697ndash698 2008
[61] D Boche E Zotova R O Weller et al ldquoConsequence of A120573immunization on the vasculature of human Alzheimerrsquos diseasebrainrdquo Brain vol 131 no 12 pp 3299ndash3310 2008
[62] M Pfeifer S Boncristiano L Bondolfi et al ldquoCerebral hemor-rhage after passive anti-a120573 immunotherapyrdquo Science vol 298no 5597 p 1379 2002
[63] M M Racke L I Boone D L Hepburn et al ldquoExacerbationof cerebral amyloid angiopathy-associatedmicrohemorrhage inamyloid precursor protein transgenic mice by immunotherapyis dependent on antibody recognition of deposited forms ofamyloid 120573rdquo Journal of Neuroscience vol 25 no 3 pp 629ndash6362005
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

BioMed Research International 5
27 A120573 Load Calculation Digital images of 7 randomlyselected microscopic transverse sections per treated animalwere analyzed and the percentage of immunostained A120573deposition was determined for all the markers studied asdefined by the stereotaxic Bregma coordinates A total of 7selected sections per animal were evaluated using the NIHImage J program by defining region of interest and setting athreshold to discriminate nonspecific staining Quantitativeanalysis of amyloid burden area was also performed in thehippocampal regions and parietaltemporal cortical regionsWe used areapixels analysis software (Pixcavator 4) toquantify the average number of pixels inside each A120573 plaqueper brain section Therefore Pixcavator intensity thresholdimaging was set to mark only those areas stained with 120573-amyloid antibody (A120573 load) relative to the entire imagearea The immunostaining intensity of each image was firstaveraged per mouse and then per group and expressed inpercentage units
28 Imaging Images were visualized using a microscope(Olympus BX50) and digitized using a digital camera (DP-10Olympus) A120573 burden data was determined by quantitativeimage analysis (Pixcavator 4) and reported as plaques persection (ps) or percentage of immunolabeled area cap-tured within total sample area The microphotographs wereadjusted for brightness and contrast with Corel Photo-Paint(Corel Ottawa Canada) and plates were composed withCorel Draw
29 Sensorimotor and Cognitive Testing After the adminis-tration of the vaccine mice were subjected to sensorimotorand memory testing to measure their coordination andlearning performances during the experimental period andto verify that any treatment-related effects observed in thememory tasks could not be explained by differences in senso-rimotor abilities Prior to testing as previously described [32]the mice were adapted to the room and testing conditions
291 Rotarod The motor strength ability balance andcoordination skills of all the experimental mice in the twotreatment periods were evaluated in a rotarod apparatus(Columbus Instruments Columbus OH) operated at 10 rpmbeginning at 7 weeks of age (preventive group) and 35 weeksof age (therapeutic group) The animals were adapted to theapparatus by receiving training sessions during four weekssufficient to reach a baseline level of performance This pro-cedurewas designed to assessmotor behavior during six trialsof a groupday If themouse remained on the rod for 1minutethe test was completed and scored as 1 Each animal wastested previous to the treatment injection for three sessionswith each session separated by 15 minutes The time thateachmouse remained on the rodwas registered automaticallyand stopped when the animal fell or inverted (by climbing)from the top of the rotating barrel In this experimentone investigator evaluated the rotarod performance and wasblinded to mice treatments and the second one treated themice and scored the test to avoid a bias
292 Fear ConditioningActive Avoidance This fear-moti-vated associative avoidance test is based on electric currentwhere themouse has to learn to predict the occurrence of thisaversive event by actively moving to a different compartment(escape platform) This active avoidance task provides asimple way to assess associative learning and memory bytesting the ability of the mouse to avoid an aversive event inresponse to a stimulus cueThemeasures recorded number ofnonresponses (the mouse failing to reach the escape platformduring the trial) and response latency (latency to avoid orescape) serves as an index of learning and allows memoryto be assessed After habituation for 5min mice received 30randomized escapable footshocks at an intensity of 025mAIn each shock trial the time to reach the escape platformlocated constantly at a central quadrant from the startlocationwas recorded as latency Each trial lasted amaximumof 30 s If the mouse failed to find the platform within 30 s itwas manually guided to the platform and allowed to stay for10 s the latency was recorded 30 s Experimental results areindicated as mean response time for each treatment period
210 Onset and Survival Disease onset and progression wereconfirmed and defined by periodic mice sacrifice and neu-ropathological analysisThemortality rates of APPPS1 trans-genicmice in the course of both sets of experiments were 42for A120573
42+ CFAIFA 17 for EB101 and 12 for the vehicle
211 Statistical Analyses For statistical analyses SPSS (ver-sion 110 SPSS Inc Chicago) was used and a 119875 value lt005 was considered statistically significant The correlationaverage of A120573 plaque density burden area and antibody titerswere analyzed and compared in multiple groups by one-wayanalysis of variance (ANOVA) followed by Fisherrsquos LSD posthocmeans comparison test analysis when relevant All graphsdisplay group mean plusmn SEM
212 Limitations of the Study The ultimate goal of the ADpreclinical research is developing a therapeutic drug strategyin the ideal animal model that should recapitulate most ifnot all the neurobehavioral pathology and underlying thesame mechanisms described in human AD such as therate of amyloid plaque accumulation neurodegenerationand behavioral and cognitive alterations Although such anideal mouse model has not yet been generated we believethat APPswePS1ΔE9 transgenic mice present enough patho-logical features to answer the specific questions addressedby the present study Active A120573 immunotherapy in whichantibodies are produced by the individual upon contact withthe antigen has great potential for preventive AD therapy asshown here in animal models although some aspects suchas an undesirable immune response induced by the adjuvantlong immune response adjustment age-dependent effectsinflammation and microhemorrhages should be addressedbefore its application in clinical trials Being aware of theseissues we designed the present study aiming to solve andovercome all these disadvantages
6 BioMed Research International
3 Results
31 Efficacy of EB101 versus A12057342
+ CFAIFA in PreventingA120573 Burden Development Recently we demonstrated thatimmunization with EB101 vaccine prevents and reverses theAD neuropathology by the marked reduction of A120573 plaquesplaque-associated dystrophic neurites and astrocytosis inmice brain Now we conducted comparative immunizationstudies between EB101 and A120573
42+ FAIFA vaccines in the
prevention ofA120573deposits in themouse brainA120573plaques andvascular amyloid loadswere detected counted andmeasuredas the percent of total surface stained by monoclonal anti-A120573 antibody in the hippocampal and cortical sections Theplaque amyloid load observed in the representative photomi-crographs presented in Figure 2 shows a significant differenceacross the five experimental groups (ANOVA 119865 = 315119875 lt 0001) As a result of the first experimental phase basedon preventive treatment we observed that hippocampal loadwas significantly reduced in the EB101 immunized mice (AFigures 2(a) and 3) when compared with different vaccinecomponents (B-C Figures 2(b)-2(c) and 3) and moderatecompared with A120573
42+ CFAIFA vaccine (D Figures 2(d)
and 3) In fact the load in these treated groups (B-C) wasnot significantly different from PBS treated mice (E Figures2(e) and 3) although there is a positive correlation betweenthe presence of A120573 in treatment regime and the total A120573burden (Figures 2(b) 2(d) and 3) In the EB101 treated micea few compacted A120573 plaques were observed in retrosple-nial granular region of the cortex and at the polymorphdentate gyrus of the hippocampus (Figure 2(a)) Strikinglydifferent was the distribution of A120573 plaques observed inthe other treatment groups where numerous A120573 plaqueswere located at different cortical and hippocampal layers(Figures 2(b)ndash2(e)) This data was also supported by totalA120573 plaques density and size quantification made by imageanalysis software in hippocampal sections stained with anti-A120573 antibody (Figures 3 and 4) The data obtained in thepresent study validate our previous observations demon-strating that the mean burden A120573 plaques (Figures 2(a) and3) of Group A (25 plusmn 5 ps) were significantly different fromthe other treated groups (63 plusmn 6 ps in Figure 2(b) 65 plusmn6 ps in Figure 2(c) 56 plusmn 5 ps in Figure 2(d) 68 plusmn 6 psin Figures 2(e) and 3) This represents a decrease in themean percentage of A120573 plaque area of 301ndash382 in theEB101 immunized mice group relative to other tested groups(Figures 2(a)ndash2(j) and 4)The appearance of amyloid plaquesin the entorhinal (Figures 2(f)ndash2(j)) and piriform cortex wasalso analyzed being detected in low density in Group A(Figure 2(f))moderate density inGroups B (Figure 2(g)) andD (Figure 2(i)) and high density in the entorhinal cortexof Groups C (Figure 2(h)) and E (Figure 2(j)) Thereforethis data shows that in the preventive treatment periodthe clearance effect of the EB101 vaccine on A120573 plaques persection was about 64 (Figures 2(a) and 2(f)) compared topositive controls while in the other treatments regimes 19was detected in A120573
42+ CFAIFA (D) 10 in A120573
42liposome
without S1P (B) and 7 in liposome with S1P treated mice(C) Photomicrographs show that in all groups these plaques
were mainly localized in the external layers of the entorhinalcortex (Figures 2(f)ndash2(j)) In addition we have observedlower stained intensity of the A120573 plaques in the brainregions of EB101 treated mice which contrasts markedlywith the intensely stained A120573-immunoreactivity of theseplaques in each brain section of the other experimental micegroups
32 Efficacy of EB101 versus A12057342
+ CFAIFA in Clearing A120573Plaque Burden In a second set of experiments we comparedthe therapeutic effect of EB101 vaccine with A120573
42+ CFAIFA
in the reduction of A120573 in the brain after the plaques wereestablished in the 35-week-oldAPPPS1 transgenicmice (Fig-ures 2(k)ndash2(t))Themice were treated and analyzed using thesame protocol as in the preventive treatment To determinewhether EB101 vaccine attenuates andor reverses themassivedevelopment of 120573-amyloid plaques brain sections fromwild-type and transgenic mice of all experimental groupswere immunostained with the specific antibody recognizingA1205731ndash42 epitopesThese results show that A120573 deposits (Figures
2(k)ndash2(t)) were notably reduced in brain sections of theEB101 treated mice (A) markedly different from the A120573burden levels observed in the other treated groups (BndashE)As observed in the corresponding photomicrographs EB101treated mice showed a few compacted A120573 plaques mainlylocated at the CA1 layer of the hippocampus (Figure 2(k)) Noreduction in plaque density was observed in the other exper-imental groups (Figures 2(l)ndash2(o)) where numerous largecompacted A120573 plaques were located in almost every layer ofeach brain section The most densely populated section withA120573 plaques were those with no A120573
1ndash42 treatment as shownin Groups C (Figure 2(m)) and E (Figure 2(o)) while a slightreduction of this hallmarks was observed in mice Groups B(Figure 2(l)) and D (Figure 2(n)) The hippocampal sectionsof the therapeutic treatment with EB101 vaccine showed asimilar pattern as that observed in the preventive treatmentThemean burden of A120573 plaques in GroupA (35 plusmn 4 Figure 3)was significantly different from the other treated groups (70plusmn5 in B 74 plusmn 4 in C 65 plusmn 5 in D 80 plusmn 6 in E) The A120573 plaquearea in the EB101 immunized mice group was reduced by297ndash345 relative to the other tested groups (Figure 4) Asobserved in the preventive set of experiments this data showthat in the therapeutic treatment period the clearance effectof the EB101 vaccine on A120573 plaques per section was about59 compared to controls While in mice group treated withA12057342+ CFAIFA the clearance effect was about 20 in Group
D 13 in Group B and 8 in Group C The same pattern ofA120573-immunoreactivity was observed in the entorhinal cortexwhen comparing all experimental treatment groups (Figures2(p)ndash2(t)) Only a few A120573 plaques were observed in theexternal layer of the entorhinal and piriform cortex of micetreated with EB101 (Figure 2(p)) while numerous plaqueswere observed in the corresponding section of Groups B(Figure 2(q)) and D (Figure 2(s)) which show a moderatedensity of A120573 plaques while Groups C (Figure 2(r)) andE (Figure 2(t)) show densely distributed plaques along themedial and external layers of the entorhinal-piriform cortexAs described in the preventive treatment we also detected
BioMed Research International 7
Ect
Cx
CPu
Ect
Gr A
(EB1
01)
(a)
(c)
(b)
(f)
(h)
(g)
Preventive treatment Therapeutic treatment
CA1
PoDGMoDG
CA1
PoDGMoDG
CA1
PoDG
MoDG
DG
DG
Ect
Ect
Cx
Cx
Cx
CPu
CPu
CPuCA1
PoDG
MoDG
DG
CA1
PoDG
MoDG
DG
Ect
Cx
CPu
Ect
Cx
CPu
Cx
Cx
Cx
Cx
Cx
(k)
(l)
(p)
(r)
(q)
CA1
PoDGMoDG
DG
Cx
Ect
Cx
CPuCA1
ControlGr E
(PBS
)
(d)
(e) (j)
Control
PoDG
MoDG
DG
CA1
CA1
PoDG
MoDG
CA1
PoDG
MoDG
DG
DG
Ect
Ect
Cx
Cx
CPu
CPu
CA1
PoDG
MoDG
CA1
PoDGMoDG
DG
Ect
Cx
CPu
PoDGMoDG
DG
Cx
Cx
Cx
(n)
(o)
(s)
(t)
Cx
(i)
Gr B
(A120573
+L)
Gr D
(A120573+
FA)
A120573 A120573 A120573 A120573
A120573A120573A120573
A120573 A120573 A120573 A120573
A120573A120573A120573A120573
A120573 A120573 A120573 A120573
A120573
(m)
Gr C
(S1P
+L)
Figure 2 Comparative effect of EB101 vaccine on beta amyloid deposits Comparative photomicrographs of A120573 immunoreactivity weretaken in the hippocampus ((a)ndash(e) (k)ndash(o)) and cortical ((f)ndash(j) (p)ndash(t)) brain regions of transgenic mice with different treatments beforeA120573 plaques development preventive treatment ((a)ndash(j)) and after the A120573 plaques developed therapeutic treatment ((k)ndash(t)) Preventivetreatment transverse brain sections of 11-month-old mice show reduced number of A120573 deposits in the dentate gyrus hippocampal subregionCA1 (a) and entorhinal cortex (f) following EB101 vaccine immunization (Group A) contrasting with the numerous A120573 immunoreactiveplaques in the correspondent mouse brain sections of Groups B ((b) (g)) C ((c) (h)) D ((d) (i)) and E ((e) (j)) Immunoreactive A120573 in thebrain sections of wild-type (WT) mice is absent (inserts in panels (e) (o)) Therapeutic treatment transverse brain sections of 18-month-oldmice are shown in panels (k)ndash(t) Treatment with EB101 almost completely cleared A120573 load in the dentate gyrus and reduced notably thedensity in hippocampal subregion CA1 (k) and entorhinal cortex (p) compared to the same areas of mouse Groups B ((l) (q)) C ((m)(r)) D((n) (s)) and E ((o) (t)) as determined by number of plaques and staining intensity area Scale bar 100 120583m
8 BioMed Research International
Gr E
(PBS
)
0102030405060708090
Preventive treatmentTherapeutic treatment
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Mea
n of
120573-a
myl
oid
plaq
ues
sect
ion
Gr A
(A120573+
S1P+
L)(E
B101
)
Figure 3 Comparative analysis of A120573 plaque burden in the brainsof APPPS1 mice Mean of A120573 plaque burden in the hippocampaland cortical regions of APPswePS1ΔE9 mice in the five treatmentgroups The mean of the A120573 burden is significantly reduced (119875 lt005) in Group A (EB101) when compared with Groups BndashE duringpreventive treatment and markedly reduced in the therapeutictreatment period Data are presented asmeanplusmn standard error of themean (SEM) Densitymean of A120573 plaques in Tgmice hippocampus
05
1015202530354045
Preventive treatmentTherapeutic treatment
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Gr A
(A120573+
S1P+
L)(E
B101
)
Mea
n of
120573-a
myl
oid
plaq
ues a
rea
Figure 4 Comparative quantification of A120573 burden area in thebrain of APPPS1 mice Quantitative analysis of A120573 burden areain the hippocampal and cortical regions of APPswePS1ΔE9 micetreated with EB101 A120573liposomes S1Pliposomes A120573
42+ FAIFA
and PBS represented by the number of pixels inside the stained areaof each A120573 plaque This graphic shows that A120573 plaques of EB101-treated mice (Group A) are significantly smaller in size (119875 lt 005)than those of the other four treatment groups Data are presented asmean plusmn SEM Mean of A120573 plaques area in Tg mice hippocampus
a reduced staining intensity of the A120573 plaques in the brainregions of mouse Group A contrasting with the intenseA120573-immunoreactivity of plaques in the other experimentalmouse groups as shown in the photomicrographs of Figures2(k)ndash2(t)
33 Immune Response Effects of EB101 versus A12057342+ CFAIFA
Vaccine Astrocyte activation implicated in neuroinflam-mation amyloidogenesis and neuronal cell death in AD
was compared after treatment of the transgenic mice withEB101 and A120573
42+ CFAIFA Immunohistochemical analysis
of GFAP-reactive cells was carried out in the brains ofthe treated mice in the preventive and therapeutic studies(Figure 5) After the preventive treatment (Figures 5(a)ndash5(j))EB101 significantly reduced the density of GFAP-reactive cellclusters in the hippocampal and cortical sections as comparedwith A120573
42+ CFAIFA (D) A120573Lip without S1P (B) Lip with
S1P (C) and PBS (E) The distribution of GFAP immunore-active cells in the transverse section of the mouse brainstreated with EB101 shows a few scattered GFAP-reactiveclusters mainly at the CA1 hippocampal layers contrastingwith numerous dystrophic reactive astrocytes observed indifferent hippocampal areas of mouse brains in Groups BndashE(Figures 5(b)ndash5(e)) Similar neuroinflammation pattern wasobserved in the retrosplenial cortical layers where a fewGFAP-reactive clusters were observed in EB101 treated mice(Figure 5(f)) compared with their profuse presence in theother experimental groups (Figures 5(g)ndash5(j)) After thera-peutic treatment the GFAP immunoreactivity distributionpattern in mouse brains treated with EB101 was similar tocontrol mice (wild-type) mostly devoid of reactive GFAPclusters except a few scattered ones in the CA1 hippocampalexternal layers (Figure 5(k)) In contrast there is a moderatedensity in Group D (Figure 5(n)) and extensive density inGroup B (Figure 5(l)) Group C (Figure 5(m)) and Group Emice (Figure 5(o)) Photomicrographs of retrosplenial cortexshow similar neuroinflammation pattern where a few GFAPclusters detected in the EB101 treated mice (Figure 5(p))contrast with a more extensive density of clusters in the othergroups (Figures 5(q)ndash5(t)) No astrocytosis was observed inwild-type mice during preventive or therapeutic treatment
34 Comparative A120573 Antibody Titers and Th1Th2 CytokineLevels in Treated Mice A120573 antibody titers in the sera weredetermined by ELISA during mouse immunization(Figure 6) Preventive treatment with EB101 resulted in amarked increase of specific IgG A120573
1ndash42 antibody productionin all animals Mice in A and D groups developed andmaintained serum antibody titers between 1 2000 and1 8000 Mice in Groups B C and E showed no significantanti-A120573 antibody production Antibody production and titerin Group B were three and four times lower than those ofGroups D and A respectively After therapeutic treatment allmice treated with EB101 and A120573
42+ FAIFA produced high
levels of IgG antibodies Titer levels of Groups A and D weresignificantly increased to those detected after preventivetreatment (119875 lt 001 versus Group D 119875 lt 0001 versusGroups B C and E) although antibody level in Group D was13-fold lower than in Group A (Figure 6)The effect of EB101(Group A) and A120573
42+ CFAIFA (Group B) vaccines on Th1
andTh2 cytokines was also studied (Figure 7)Measurementsof Th1 and Th2 cytokines in sera of immunized transgenicmice indicate that significant changes occurring in proteinlevels of Th1-related inflammatory mediators followingpreventive (Figures 7(a)ndash7(c)) and therapeutic (Figures 7(d)ndash7(g)) treatment with EB101 vaccine A significant increaseof Th1 cytokine profile was observed in preventive treatment
BioMed Research International 9
Gr E
(PBS
)
(d)
(e)
(i)
(j)
(n)
(o)
(s)
(t)
Cx
GFAP
GFAP GFAP GFAP GFAP
GFAP GFAP GFAP
Cx Cx
Cx
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
GFAP
Gr A
(EB1
01)
(a)
(b)
(c)
(f)
(g)
(h)
Preventive treatment Therapeutic treatment
PoDG
MoDG
(k)
(l)
(m)
(p)
(q)
(r)DG
GFAP GFAP GFAP
GFAPGFAPGFAP GFAP
GFAP GFAP GFAP GFAP
PoDGDG
DG
CA1
Cx
Cx
Cx
Cx
Cx
Cx
PoDG
MoDG
DG
CA1
MoDGDG
CA1
PoDG
MoDG
DG
CA1
Cx
CxCx
MoDG
MoDG
PoDG
CA1CA1
Cx
Gr D
(A120573+
FA)
Gr C
(S1P
+L)
Gr B
(A120573+
L)
Figure 5 Comparative effect of EB101 vaccine on astrocytosis Comparative photomicrographs of GFAP immunoreactivity in thehippocampus ((a)ndash(e) (k)ndash(o)) and cortical ((f)ndash(j) (p)ndash(t)) brain regions at the preventive treatment phase ((a)ndash(j)) and therapeutictreatment group ((k)ndash(t)) Preventive treatment transverse brain sections of 11-month-old mice treated with EB101 show an almost completeprevention of astrocytosis in the detailed sections of the dentate gyrus (a) contrasting with numerous dystrophic reactive astrocytes typicalof an inflammatory reaction observed in different hippocampal corresponding areas of mouse brains in Group B (A120573Lip-treated mice (b))Group C (LipS1P-treated mice (c)) Group D (A120573
42+ CFAIFA-treated mice (d)) and Group E (PBS-treated mice (e)) Immunoreactive
astrocytosis in the brain sections of WT mice is absent (data not shown) Detailed sections of the parietal cortex of treated mice show agradient density of reactive astrocytes immunoreactive to GFAP indicating a neuroinflammation pattern Comparative cortical sections shownumerous astrocytosis areas inmice Groups C and E ((h) (j)) andwide reactive astrocytes clusters inmice Groups B andD ((g) (i)) while justa few of them are observed in Group A (f)Therapeutic treatment transverse brain sections of 18-month-old mice of Group A show a notablereduction of astrocytosis in the hippocampal region (k) after EB101 vaccine immunization Note the astrocytosis contrast density betweenEB101-treatedmice and the other treated groups ((l)ndash(o)) where numerous immunoreactive GFAP clusters are observed in the correspondentbrain sections A few immunoreactive GFAP clusters are observed in the parietal cortex of EB101 treated mice group while a moderate ((q)(s)) to high ((r) (t)) density of these neuropathological inflammation clusters is apparent in the other treatment groups (Groups BndashE) Forabbreviations see list Scale bar 100120583m
10 BioMed Research International
002040608
1121416
Preventive treatmentTherapeutic treatment
Ant
i-120573-a
myl
oid
IgG
antib
odie
s
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Gr A
(A120573+
S1P+
L)(E
B101
)
Figure 6 Quantification of specific anti-120573-amyloid IgG in immu-nized APPPS1 mice The presence of anti-120573-amyloid antibodiesmeasured in the sera of vaccinated and control mice groups duringthe preventive and therapeutic treatments indicates that EB101liposome vaccine was able to induce the highest production ofspecific IgG antibodies Compared to theGroupD immunizedmicethe level of antibodies observed in the Group A was also highersuggesting a more effective immune response (119875 lt 001 versusGroup D 119875 lt 0001 versus Groups B C and E) Quantification ofspecific anti-120573-amyloid IgG in immunized Tg mice
in Group D with respect to all other groups mainlyreported by the high levels of TNF-120572 (Figure 7(b)) andIFN-120574 (Figure 7(c)) cytokine secretions while due to thehigh mortality rate during the therapeutic treatment nosignificant data was obtained from this mice group Inaddition a tendency toTh1 secretion reduction was observedinGroupAwith respect to all other groups in both preventiveand therapeutic treatment The changes in Th2 protein levels(Figure 7(d)) followed the inverse trend where the highestdifferences between Group A and the other treatment groupswere observed in the IL-4 IL-5 and IL-10 cytokine secretions(119875 lt 005 versus Group D 119875 lt 001 versus Group E Figures7(e)ndash7(g))
35 Comparative Motor Coordination and Memory Perfor-mances In order to validate the locomotion integrity duringthe entire immunotherapeutic process as reported in similarprevious studies motor coordination tests have showed thatEB101-treated mice spent significantly more time on theaccelerating rotarodThismotor coordination task shows thatEB101 immunization affected positively their performance(Figure 8(a)) A120573
42liposome (B) S1P-containing liposomes
(C) and PBS (E) (119875 lt 0006) mice performed poorly incomparison to EB101 mice (119875 lt 005) or A120573
42+ CFAIFA
mice (119875 lt 002) in motor abilities and coordination Thefear conditioning-active avoidance test was used to assess thespatial learning and memory functions As expected bothEB101 and A120573
42+ CFAIFA immunized mice readily learned
the location of the platform as shown by a decrease in theescape latency over training days while the control group
mice exhibited no significant changes during 6-day trialsThe average escape latencies of EB101 (68 plusmn 2 s 761 plusmn 22 s)andA120573
42+CFAIFA (116plusmn33 s 123plusmn21 s) immunizedmice
at the end of preventive and therapeutic periods respectivelyduring the consecutive 6 days trial were significantly shorterthan the other treatment groups (A120573
42liposome 163 plusmn 31 s
175 plusmn 28 s S1P-containing liposomes 202 plusmn 19 s 248 plusmn27 s) and the control (2116 plusmn 38 s 268 plusmn 22 s) (119875 lt 001)Therewas no significant difference between EB101 andA120573
42+
CFAIFA immunized group (119875 gt 005) (Figure 8(b)) Theseresults showed that spatial learning was improved in theseimmunized mice
4 Discussion
In this study we have compared a novel active A120573 imm-unotherapy the EB101 vaccine against A120573 with the A120573
42+
CFAIFA vaccine that was first taken into clinical trialswith coadjuvant Qs-21 modification (AN1792 by Orgogozoet al [19]) This type of vaccine design was thought toinduce a strong T-cell mediated inflammatory response (IL-2 and IFN-c positive responses indicative of a Class II(CD4) Th-1 type response [33] leading to acute menin-goencephalitis in the 6 of the patients) The EB101 activeimmunotherapeutic vaccine was primarily designed to avoidundesired inflammatory side effects both in mice andhumans (without need of coadjuvant modification) whileinducing effective antibody titer rates We demonstrated thatlong-term immunization with EB101 induced A120573 antibodiesin the preventive and the therapeutic treatment periodswhich effectively reduced amyloid deposition in both corticaland hippocampal brain regions and prevented neuroinflam-mation probably by blocking toxicity in the surroundingneurons However the AN1792 vaccine was shown to inducein 6 of the patients an inflammatory reaction that mighthave led to the encephalitis type reaction observed in theclinical trial which was responsible for the immediate haltin those clinical studies It seems that AN1792 immunizationdelivered in Qs-21 and polysorbate 80 adjuvant induced aTh1 type response that likely led to autoaggressive T cells andaseptic meningoencephalitis In the present experiments wedemonstrated that a judiciously selected adjuvant produces adifferent humoral response [22] Indeed we have shown thatour novel immunotherapeutic strategy boosts specific A120573
1ndash42antibody generation and induces a Th2 noninflammatoryimmune response This boosted effect is in accordance withprevious immunotherapeutic studies [34] Taking togetherall the present findings these results suggest that EB101vaccine can achieve in a future clinical trial immunogenicresponse rates quite similar to the AN1792 without theinflammatory effects reported The immunohistochemicalanalysis obtained in the present study shows that all the EB101immunized mice resulted in an almost complete preventionof A120573 deposition in the brain with only a few compactA120573 plaques in the retrosplenial and dentate gyrus regionsAlthough there was a slightly increase in death in EB101treated mice compared to controls this significant reductionof A120573 plaques in EB101 treated mice was proven to be more
BioMed Research International 11
Gr A Gr B Gr C Gr D Gr EPreventive treatmentTherapeutic treatment
0005001000150020002500300035004000
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
(a) Th1
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
000
1000
2000
3000
4000
5000TNF-120572
(b) Th1
000500
10001500200025003000
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
IFN-120574
(c) Th1
000500
100015002000250030003500
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
Group A Group B Group C Group D Group EPreventive treatmentTherapeutic treatment
(d) Th2
000
500
1000
1500
2000
2500
3000IL-4
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(e) Th2
000500
100015002000250030003500
IL-5
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(f) Th2
3500IL-10
000500
10001500200025003000
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(g) Th2
Figure 7 Detection of Th1 and Th2 cytokine profile Measurements of the effect of EB101 and A12057342
+ CFAIFA vaccines on Th1 and Th2cytokines profile during preventive and therapeutic treatments ((a)ndash(c)) A significant increase of Th1 cytokine profile was observed inpreventive treatment in Group D with respect to all other groups while a tendency to a reduction inTh1 secretion was observed in Group Awith respect to all other groups in both preventive and therapeutic treatment ((d)ndash(g)) The changes in Th2 protein levels were observed inthe IL-4 IL-5 and IL-10 cytokine secretions of Group A (119875 lt 005 versus Group D 119875 lt 001 versus Group E)
12 BioMed Research International
Gr A
(EB1
01)
05
101520253035
Preventive treatment Therapeutic treatment
Mea
n fa
lling
late
ncy
(s)
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)(a) Graphic of motor coordination and balance
05
1015202530
Mea
n re
spon
se ti
me (
s)
Gr A
(EB1
01)
Preventive treatment Therapeutic treatment
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
(b) Graphic of associative learning and memory
Figure 8 Analysis of motor coordination and balance (a) Rotarod results of motor coordination and balance test after the preventive andtherapeutic treatment showing improved effect of EB101 and A120573
42+ CFAIFA-vaccine compared to other treated mice groups (119875 lt 005)
and (119875 lt 002) versus control mice Latency to fall on the rotarod apparatus was significantly improved in EB101 immunized mice Valuesrepresent means of all six trials of a groupday (b) Spatial learning and memory tested by fear conditioning-active avoidance test The meanresponse time of each treatment group to reach the platform is represented EB101 and A120573
42+ CFAIFA immunized mice had significantly
lower latencies than the other treatment groups (119875 lt 001) versus control mice
efficient than that obtained with A12057342+ CFAIFA immuniza-
tions A comparison of A120573 plaque reduction rates amongtreated groups surprisingly showed that other than EB101and A120573
42+ CFAIFA treated groups some amyloid deposits
were also reduced in Groups B (A1205731ndash42liposome) and C
(S1P-containing liposomes) Although the reduction rates inthese mice were markedly low the subtle lowering effectmight be due to the interaction between the immunizationelements (A120573
1ndash42 and S1P) and the mouse immunogenicsystem as reported by others [35] Based on our presentresults S1P one of the components of the adjuvant used withEB101 might be responsible for the neuroprotective effectobserved in the vulnerable neuronal areas of EB101 treatedmice The advantage in the A120573 load preventionreduction bycombining A120573
1ndash42 and S1P in a unique formulation vaccinehas been also observed in the reduced hippocampal load ofEB101 immunizedmice (Gr A) when comparedwith differentvaccine components (Gr B-C) In fact S1P appears to have ademonstrated therapeutic potential as a regenerative agent inthe nervous system since it controls migration of neuronalstem cells toward a site of spinal cord injury [36] inducecytoskeletal rearrangements to promote transmitter secretion[37] and also trigger diverse cellular effects including angio-genesis cardiac development immunity cell motility andneurite extension [38 39]Therefore it is reasonable to expectthat such a potent biologically active sphingolipid by itselfmay play an important role in the A120573 plaque reduction Inthis study the clearance efficiency rate of each treatment wasanalyzed and results suggested that EB101 (6459) appearsto be more effective than A120573
42+ CFAIFA (1920) vaccine
in both preventive and therapeutic groups The effectivenessof EB101 in older transgenic animals (18 months) showeda clearance effect of 59 suggesting that EB101 may alsohave therapeutic potential inADdiagnosticated patient trials
Similar conclusion was reached byWang and colleagues [40]who demonstrated that combination therapy in aged animalsprovides more valuable insights into a possible translationto humansOur immunologicalmeasurements demonstratedonce again that neuroinflammation develops in an A120573-dependent fashion early in the course of AD [41] and canbe alleviated or reversed by interventional A120573 vaccinationsuch as EB101 Among the possible mechanisms of A120573clearance proposed in AD immunotherapy [42] our resultssuggested that theymay overlap and probably be disease statedependent since the prevention and the therapeutic vaccinewould likely benefit from the transport of A120573 antibodies intothe CNS However some mechanisms of action such as thepreventive effect of A120573 antibodies in the A120573 aggregation [43]microglial phagocytosis induced by the Fc binding portionsbetween A120573 antibodies and microglia receptor [44] and theshift in the gradient of A120573 transport across the blood-brainbarrier due to the increased concentration of A120573 antibodiesin the vascular system resulting in an increase in effluxfrom the brain to blood [45 46] may be playing a role inboth preventive and therapeutic conditions Our findingstherefore suggest that novel treatment strategies aimed atmaintaining or increasing microglial function and restoringthe balance to the immunological system may represent anattractive therapeutic approach even at the advanced stagesof ADThe immunization results obtained with EB101 duringboth preventive and therapeutic treatment showed nomicro-hemorrhages infiltration of lymphocytes or encephalitis-like reaction in the brain suggesting that EB101 does notinduce A120573-specific Th1 reaction possibly due to the neu-roprotective effect of S1P Delivering A120573
1ndash42 in a liposomal-S1P-adjuvant resulted in an effective immunological responseincluding the induction of a Th2 T-cell response whicheffectively prevented neuroinflammation in an animal model
BioMed Research International 13
of AD as reported previously [24 47] Moreover presentresults demonstrated that EB101 did not induce astrogliosisin APPPS1 transgenic mice whereas some GFAP-positiveastrocyte clusters were aggregated in the affected brain areasof mice treated with A120573
42+ CFAIFA These results indicate
that EB101 may result in an effective treatment in AD neu-ropathology due to the absence of peripheral proinflamma-tory cytokine secretion and the absence of astroglial activa-tion in the brains of EB101 treated mice when compared withA12057342
+ CFAIFA immunotherapeutic response In a clinicalsetting however we may expect a wide differential responsein part depending upon the genomic profile of each patient[48 49] In AD behavioural studies in particular amongthe motor coordination tests the rotarod performance hasbeen reported to be impaired in some AD transgenic micemodels bearing A120573 plaques [50 51] This result was found inmutants prior to A120573 plaque formation at 3 and 6 months ofage but was not reproduced after plaques had been formedat 16 and 24 months of age [52] perhaps because of thedeclining abilities of older normal mice Here we presentthe effects of vaccine treatments in APPPS1 mice on motorcoordination a neglected aspect of the AD experimentalbehavior but justified on the basis of the deficient posturalcontrol reported in patients with Alzheimerrsquos disease [53]sometimes even during early stages [54] Our longitudinalmotor behavioral results showed no significant differencesin the ambulatory movement grip strength or coordinationaddressed by their performance in the rotating rod betweenAPPPS1 and wild-type mice for any age group studiedin contrast with other treated groups demonstrating thatlongitudinal motor assessments of AD transgenic mousemodels can provide valuable information with regard tothe effect of immunization on the progression of behavioraldeficits and overall motor activity As shown the EB101-treated group of mice exhibited longer latencies and betterbalancing skills indicating the absence of any negative effectof EB101 immunization on these mice The classical fearconditioning test is assumed to reveal the functional integrityof the hippocampus [55] representing a special memorytask to evaluate the AD models and its improvement afterimmunization periods In this APPPS1 mouse model thefirst associative learning and spatial workingmemory deficitsare shown at 5-6 months of age [56] although its impair-ment progression is relatively slow compared to most othermodels At the end of preventive immunization period EB101and A120573
42+ CFAIFA immunized mice have significantly
shorter escape latencies than the other treatment groups(A12057342liposome S1P-containing liposomes) and the controls
in the fear conditioning test Shorter escape latencies duringspatial learning indicated that deficits of spatial memorywere attenuated in EB101 and A120573
42+ CFAIFA immunized
mice Previous studies have showed that by 15 months ofage the spatial working memory is consistently impairedthroughout the rest of the life span [31 57] At the end oftherapeutic immunization period both EB101 and A120573
42+
CFAIFA immunized mice showed significantly improvedresults on the hippocampus-dependent spatial learning andmemory test when compared with other treatment groupsand the controls Taken together these observations are
in agreement with the impairment in conditioned learningin response to a tone stimulus of aged APPPS1 micealthough when immunized with EB101 during preventiveor therapeutic periods such impairments are significantlyattenuated
In summary the present results corroborate our previouspublished findings showing that EB101 immunization reducesamyloid accumulation in the APPPS1 mouse brain [2425] The strongest reduction in the preventive immunizationwas observed in the retrosplenial cortex followed by thehippocampus and entorhinal-piriform cortex Our resultsfurther support that EB101 ismore effectivewhen themice areimmunized prior to developing plaques in the hippocampaland cortical brain regions There is extensive evidence inmouse preclinical studies [58] and human clinical trials [5960] that the adverse effects of active and passive A120573 vaccinesresult in microhemorrhages and encephalitic reactions [60ndash63] yet themechanismbywhich they occur has not been fullyelucidated In the treatment sections analyzed we found nomicrohemorrhages or acute inflammatory processes in all thesurvived mice of the group indicating that EB101 therapeuticapproach presents no adverse inflammatory effects in thisAD mice models Thus our present results unequivocallydemonstrate that EB101 vaccinemay not only halt AD-relatedpathology as a preventive treatment but may also reverse it inestablished cases related to the improvement of the learningandmemory in both cases leading to a robust immunologicaltherapeutic response Present experiments also demonstratethat EB101 exerts more significant and substantial effects onthe reduction of A120573 deposition than A120573
42+CFAIFA vaccine
in A120573 mice models of AD However further exploration ofthe contribution of genetic environmental and physiologicalfactors in the development resistance to endogenous A120573-antibody is needed in order to optimize and modulate thetherapeutic strategies currently being developed
5 Conclusions
The present study carried out in APPPS1 transgenic mousemodel of AD compared the A120573 burden reduction andcellular immune responses produced by two immunizationdelivery strategies of A120573 in Freund adjuvant (described byWebster et al [10]) and in liposomeS1P matrix (describedby Schenk et al [17])The EB101 immunotherapeutic strategyboosted antibody generation A120573 plaque reduction and ananti-inflammatory response more effectively than A120573
42+
CFAIFA especially if given prior to or in the early stages ofpathology The present results also showed that vaccinationwith EB101 prevented the formation of new plaques andpresumably led to a reduction in established plaques whileinducing aTh2 immune reactionwithout encephalitogenic T-cell responses or brain inflammation related with astrogliosisHowever despite the efficacy demonstrated of this novelvaccine approach further preclinical and clinical studieson EB101 are warranted to evaluate its efficacy and themechanism for the future utility of such vaccine for ADtherapy
14 BioMed Research International
List of Abbreviations
AD Alzheimerrsquos diseaseA120573 120573-Amyloid proteinAPP Amyloid precursor proteinAPT Anterior pretectal nucleusCA1 Field CA1 hippocampusCA3 Field CA3 hippocampusCPu Caudate putamen (striatum)Cx CortexDG Dentate gyrusDN Dystrophic neuriteEct Entorhinal cortexCFA Complete Freundrsquos adjuvantGrDG Granular dentate gyrusIFA Incomplete Freundrsquos adjuvantIL InterleukinLPMR Lateral posterior thalamic nucleus (mediorostral)MoDG Molecular dentate gyrusPir Piriform cortexPoDG Polymorph dentate gyrusRSGb Retrosplenial granular cortex b regionTg TransgenicTh T helper
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Ivan Carrera Ignacio Etcheverrıa Lucıa Fernandez-NovoaValter Lombardi Ramon Cacabelos and Carmen Vigo con-ceived and designed the experiments Ivan Carrera and Igna-cio Etcheverrıa performed the experiments Ivan CarreraIgnacio Etcheverrıa and Valter Lombardi analyzed the dataIvan Carrera Ignacio Etcheverrıa Lucıa Fernandez-Novoaand Valter Lombardi contributed reagentsmaterialsanalysistools Ivan Carrera Carmen Vigo andMadepalli KrishnappaLakshmana wrote the paper
Acknowledgments
This work was supported by National Institute of Aging(NIA)NIH Grant no 1R01AG036859-01 (Madepalli Krish-nappa Lakshmana) and the financial support of the ProgramaOperativo Feder Galicia 2007ndash2013 (IN841C 201254 to PhDCarrera I)
References
[1] R Cacabelos L Fernandez-Novoa V Lombardi Y Kubotaand M Takeda ldquoMolecular genetics of Alzheimerrsquos diseaseand agingrdquoMethods and Findings in Experimental and ClinicalPharmacology vol 27 pp 1ndash573 2005
[2] P T Nelson I Alafuzoff E H Bigio et al ldquoCorrelationof alzheimer disease neuropathologic changes with cognitive
status a review of the literaturerdquo Journal of Neuropathology andExperimental Neurology vol 71 no 5 pp 362ndash381 2012
[3] JNaslundVHaroutunian RMohs et al ldquoCorrelation betweenelevated levels of amyloid 120573-peptide in the brain and cognitivedeclinerdquo Journal of the American Medical Association vol 283no 12 pp 1571ndash1577 2000
[4] A Koyama O I Okereke T Yang D Blacker D J Selkoeand F Grodstein ldquoPlasma amyloid-120573 as a predictor of dementiaand cognitive decline a systematic review and meta-analysisrdquoArchives of Neurology vol 69 no 7 pp 824ndash831 2012
[5] A M Fagan C M Roe C Xiong M A Mintun J C Morrisand D M Holtzman ldquoCerebrospinal fluid tau120573-amyloid42ratio as a prediction of cognitive decline in nondemented olderadultsrdquo Archives of Neurology vol 64 no 3 pp 343ndash349 2007
[6] S Parvathy P Davies V Haroutunian et al ldquoCorrelationbetween A120573-40- A120573-42- and A120573x-43-containing amyloidplaques and cognitive declinerdquo Archives of Neurology vol 58no 12 pp 2025ndash2032 2001
[7] E Karran M Mercken and B D Strooper ldquoThe amyloidcascade hypothesis for Alzheimerrsquos disease an appraisal for thedevelopment of therapeuticsrdquo Nature Reviews Drug Discoveryvol 10 no 9 pp 698ndash712 2011
[8] J L Jankowsky D J Fadale J Anderson et al ldquoMutantpresenilins specifically elevate the levels of the 42 residue 120573-amyloid peptide in vivo evidence for augmentation of a 42-specific 120574 secretaserdquo Human Molecular Genetics vol 13 no 2pp 159ndash170 2004
[9] L Holcomb M N Gordon E Mcgowan et al ldquoAcceleratedAlzheimer-type phenotype in transgenic mice carrying bothmutant amyloid precursor protein and presenilin 1 transgenesrdquoNature Medicine vol 4 no 1 pp 97ndash100 1998
[10] S JWebster A D Bachstetter and L J van Eldik ldquoComprehen-sive behavioral characterization of an APPPS-1 double knock-in mouse model of Alzheimerrsquos diseaserdquo Alzheimerrsquos ResearchandTherapy vol 5 no 3 article 28 2013
[11] H Xiong D Callaghan J Wodzinska et al ldquoBiochemical andbehavioral characterization of the double transgenic mousemodel (APPswePS1dE9) of Alzheimerrsquos diseaserdquo NeuroscienceBulletin vol 27 no 4 pp 221ndash232 2011
[12] M Garcia-Alloza L A Borrelli B T Hyman and B J BacskaildquoAntioxidants have a rapid and long-lasting effect on neuriticabnormalities in APPPS1 micerdquo Neurobiology of Aging vol 31no 12 pp 2058ndash2068 2010
[13] M Meyer-Luehmann M Mielke T L Spires-Jones et al ldquoAreporter of local dendritic translocation shows plaque-relatedloss of neural system function in APP-transgenic micerdquo Journalof Neuroscience vol 29 no 40 pp 12636ndash12640 2009
[14] E Machova V Rudajev H Smyckova H Koivisto H Tanilaand V Dolezal ldquoFunctional cholinergic damage develops withamyloid accumulation in young adult APPswePS1dE9 trans-genic micerdquo Neurobiology of Disease vol 38 no 1 pp 27ndash352010
[15] T Malm J Koistinaho and K Kanninen ldquoUtilization of APP-swePS1dE9 transgenic mice in research of Alzheimerrsquos diseasefocus on gene therapy and cell-based therapy applicationsrdquoInternational Journal of Alzheimerrsquos Disease vol 2011 Article ID517160 8 pages 2011
[16] J Lee and P Han ldquoAn update of animal models of alzheimerdisease with a reevaluation of plaque depositionsrdquoExperimentalNeurobiology vol 22 pp 84ndash95 2013
BioMed Research International 15
[17] D Schenk R Barbour W Dunn et al ldquoImmunization withamyloid-120573 attenuates Alzheimer disease-like pathology in thePDAPP mouserdquo Nature vol 400 no 6740 pp 173ndash177 1999
[18] H J Fu B Liu J L Frost and C A Lemere ldquoAmyloid-120573immunotherapy for Alzheimerrsquos diseaserdquoCNS and NeurologicalDisorders-Drug Targets vol 9 no 2 pp 197ndash206 2010
[19] J-M Orgogozo S Gilman J-F Dartigues et al ldquoSubacutemeningoencephalitis in a subset of patients with AD after A120573
42
immunizationrdquo Neurology vol 61 no 1 pp 46ndash54 2003[20] J A R Nicolll DWilkinson CHolmes P Steart HMarkham
andROWeller ldquoNeuropathology of humanAlzheimer diseaseafter immunization with amyloid-120573 peptide a case reportrdquoNature Medicine vol 9 no 4 pp 448ndash452 2003
[21] I Ferrer M Boada Rovira M L Sanchez Guerra M JRey and F Costa-Jussa ldquoNeuropathology and pathogenesisof encephalitis following amyloid-Neuropathology and patho-genesis of encephalitis following amyloid-beta immunizationin Alzheimerrsquos disease immunization in Alzheimerrsquos diseaserdquoBrain Pathology vol 14 no 1 pp 11ndash20 2004
[22] D H Cribbs A Ghochikyan V Vasilevko et al ldquoAdjuvant-dependent modulation ofTh1 andTh2 responses to immuniza-tion with 120573-amyloidrdquo International Immunology vol 15 no 4pp 505ndash514 2003
[23] C Holmes D Boche D Wilkinson et al ldquoLong-term effectsof A120573
42immunisation in Alzheimerrsquos disease follow-up of a
randomised placebo-controlled phase I trialrdquo The Lancet vol372 no 9634 pp 216ndash223 2008
[24] I Carrera I Etcheverrıa L Fernandez-Novoa V LombardiR Cacabelos and C Vigo ldquoVaccine development to treatAlzheimerrsquos disease neuropathology in APPPS1 transgenicmicerdquo International Journal of Alzheimerrsquos Disease vol 2012Article ID 376138 17 pages 2012
[25] I Carrera I Etcheverrıa Y Li et al ldquoImmunocytochemicalcharacterization of Alzheimer disease hallmarks in APPPS1transgenic mice treated with a new anti-amyloid-120573 vaccinerdquoBioMed Research International vol 2013 Article ID 709145 12pages 2013
[26] V Anelli R Bassi G Tettamanti P Viani and L Riboni ldquoExtra-cellular release of newly synthesized sphingosine-1-phosphateby cerebellar granule cells and astrocytesrdquo Journal of Neuro-chemistry vol 92 no 5 pp 1204ndash1215 2005
[27] S Schroeter K Khan R Barbour et al ldquoImmunotherapyreduces vascular amyloid-120573 in PDAPP micerdquo Journal of Neu-roscience vol 28 no 27 pp 6787ndash6793 2008
[28] Z Suo A Cox N Bartelli et al ldquoGRK5 deficiency leads to earlyAlzheimer-like pathology and working memory impairmentrdquoNeurobiology of Aging vol 28 no 12 pp 1873ndash1888 2007
[29] W Kamphuis C Mamber MMoeton et al ldquoGFAP isoforms inadult mouse brain with a focus on neurogenic astrocytes andreactive astrogliosis in mouse models of Alzheimer diseaserdquoPLoS ONE vol 7 no 8 Article ID e42823 2012
[30] J Lang C Vigo-Pelfrey and F Martin ldquoLiposomes composedof partially hydrogenated egg phosphatidylcholines fatty acidcomposition thermal phase behavior and oxidative stabilityrdquoChemistry and Physics of Lipids vol 53 no 1 pp 91ndash101 1990
[31] D Morgan D M Diamond P E Gottschall et al ldquoA 120573peptide vaccination prevents memory loss in an animal modelof Alzheimerrsquos diseaserdquo Nature vol 408 no 6815 pp 982ndash9852000
[32] M L Brownlow L Benner D DrsquoAgostino M N Gordon andD Morgan ldquoKetogenic diet improves motor performance but
not cognition in two mouse models of Alzheimerrsquos pathologyrdquoPLoS ONE vol 8 no 9 Article ID e75713 2013
[33] M Pride P Seubert M Grundman M Hagen J Eldridgeand R S Black ldquoProgress in the active immunotherapeu-tic approach to Alzheimerrsquos disease clinical investigationsinto AN1792-associated meningoencephalitisrdquo Neurodegenera-tive Diseases vol 5 no 3-4 pp 194ndash196 2008
[34] CWiessner K-HWiederhold A C Tissot et al ldquoThe second-generation activeA120573 immunotherapyCAD106 reduces amyloidaccumulation in APP transgenic mice while minimizing poten-tial side effectsrdquoThe Journal of Neuroscience vol 31 no 25 pp9323ndash9331 2011
[35] D Lambracht-Washington and R Rosenberg ldquoAdvances in thedevelopment of vaccines for Alzheimerrsquos diseaserdquo DiscoveryMedicine vol 15 no 84 pp 319ndash326 2013
[36] A Kimura T Ohmori R Ohkawa et al ldquoEssential roles ofsphingosine 1-phosphateS1P
1receptor axis in the migration of
neural stem cells toward a site of spinal cord injuryrdquo Stem Cellsvol 25 no 1 pp 115ndash124 2007
[37] S S Lee C H Woo J D Chang and J H Kim ldquoRoles of Racand cytosolic phospholipase A2 in the intracellular signalling inresponse to titanium particlesrdquo Cellular Signalling vol 15 no 3pp 339ndash345 2003
[38] E Masliah L Hansen A Adame et al ldquoA120573 vaccination effectson plaque pathology in the absence of encephalitis inAlzheimerdiseaserdquo Neurology vol 64 no 1 pp 129ndash131 2005
[39] A Pilorget B Annabi F Bouzeghrane J Marvaldi J Luisand R Beliveau ldquoInhibition of angiogenic properties of brainendothelial cells by platelet-derived sphingosine-1-phosphaterdquoJournal of Cerebral Blood Flow and Metabolism vol 25 no 9pp 1171ndash1182 2005
[40] A Wang P Das R C Switzer III T E Golde and J LJankowsky ldquoRobust amyloid clearance in a mouse model ofAlzheimerrsquos disease provides novel insights into themechanismof amyloid-120573 immunotherapyrdquo Journal of Neuroscience vol 31no 11 pp 4124ndash4136 2011
[41] G Krabbe A Halle V Matyash et al ldquoFunctional impairmentof microglia coincides with Beta-amyloid deposition in micewith Alzheimer-like pathologyrdquo PLoS ONE vol 8 no 4 ArticleID e60921 2013
[42] Y H Liu B Giunta H-D Zhou J Tan and Y J WangldquoImmunotherapy for Alzheimer disease-the challenge ofadverse effectsrdquo Nature Reviews Neurology vol 8 no 8 pp465ndash469 2012
[43] B Solomon R Koppel D Frankel and E Hanan-AharonldquoDisaggregation of Alzheimer120573-amyloid by site-directedmAbrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 94 no 8 pp 4109ndash4112 1997
[44] F Bard C Cannon R Barbour et al ldquoPeripherally adminis-tered antibodies against amyloid 120573-peptide enter the centralnervous system and reduce pathology in a mouse model ofAlzheimer diseaserdquo Nature Medicine vol 6 no 8 pp 916ndash9192000
[45] R B DeMattos K R Bales D J Cummins J C Dodart S MPaul and D M Holtzman ldquoPeripheral anti-A120573 antibody altersCNS and plasma A120573 clearance and decreases brain A120573 burdenin a mouse model of Alzheimerrsquos diseaserdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 98 no 15 pp 8850ndash8855 2001
[46] C A Lemere E T Spooner J LaFrancois et al ldquoEvidence forperipheral clearance of cerebral A120573 protein following chronic
16 BioMed Research International
active A120573 immunization in PSAPP micerdquo Neurobiology ofDisease vol 14 no 1 pp 10ndash18 2003
[47] T Town M Vendrame A Patel et al ldquoReduced Th1 andenhanced Th2 immunity after immunization with Alzheimerrsquos120573-amyloid
1minus42rdquo Journal of Neuroimmunology vol 132 no 1-2
pp 49ndash59 2002[48] R Cacabelos ldquoPharmacogenomics in Alzheimerrsquos diseaserdquo
Methods in Molecular Biology vol 448 pp 213ndash357 2008[49] R Cacabelos ldquoPharmacogenomics and therapeutic strategies
for dementiardquo Expert Review of Molecular Diagnostics vol 9no 6 pp 567ndash611 2009
[50] D van Dam R DrsquoHooge M Staufenbiel C van Ginneken Fvan Meir and P P de Deyn ldquoAge-dependent cognitive declinein the APP23 model precedes amyloid depositionrdquo EuropeanJournal of Neuroscience vol 17 no 2 pp 388ndash396 2003
[51] M Dumont C Strazielle M Staufenbiel and R LalondeldquoSpatial learning and exploration of environmental stimuli in24-month-old female APP23 transgenic mice with the Swedishmutationrdquo Brain Research vol 1024 no 1-2 pp 113ndash121 2004
[52] R Lalonde M Dumont M Staufenbiel C Sturchler-Pierratand C Strazielle ldquoSpatial learning exploration anxiety andmotor coordination in female APP23 transgenic mice with theSwedish mutationrdquo Brain Research vol 956 no 1 pp 36ndash442002
[53] F J Huff F Boller F Lucchelli R Querriera J Beyer and SBelle ldquoThe neurologic examination in patients with probableAlzheimerrsquos diseaserdquo Archives of Neurology vol 44 no 9 pp929ndash932 1987
[54] A F Pettersson M Engardt and L O Wahlund ldquoActivitylevel and balance in subjects with mild Alzheimerrsquos diseaserdquoDementia and Geriatric Cognitive Disorders vol 13 no 4 pp213ndash216 2002
[55] L Calandreau B Desgranges R Jaffard and A DesmedtldquoSwitching from contextual to tone fear conditioning and viceversa the key role of the glutamatergic hippocampal-lateralseptal neurotransmissionrdquo Learning amp Memory vol 17 no 9pp 440ndash443 2010
[56] F Trinchese S Liu F Battaglia SWalter PMMathews andOArancio ldquoProgressive age-related development of Alzheimer-like pathology in APPPS1 micerdquo Annals of Neurology vol 55no 6 pp 801ndash814 2004
[57] D M Wilcock A Rojiani A Rosenthal et al ldquoPassiveimmunotherapy against A120573 in aged APP-transgenic micereverses cognitive deficits and depletes parenchymal amyloiddeposits in spite of increased vascular amyloid and microhem-orrhagerdquo Journal of Neuroinflammation vol 1 article 24 2004
[58] S A Ferguson S Sarkar and L C Schmued ldquoLongitudinalbehavioral changes in the APPPS1 transgenic AlzheimerrsquosDisease modelrdquo Behavioural Brain Research vol 242 no 1 pp125ndash134 2013
[59] D Wilcock and C Colton ldquoImmunotherapy vascular pathol-ogy and microhemorrhages in transgenic micerdquo CNS andNeurological DisordersmdashDrug Targets vol 8 no 1 pp 50ndash642009
[60] M Grundman S Gilman R S Black et al ldquoAn alternativemethod for estimating efficacy of the AN1792 vaccine foralzheimer diseaserdquo Neurology vol 71 no 9 pp 697ndash698 2008
[61] D Boche E Zotova R O Weller et al ldquoConsequence of A120573immunization on the vasculature of human Alzheimerrsquos diseasebrainrdquo Brain vol 131 no 12 pp 3299ndash3310 2008
[62] M Pfeifer S Boncristiano L Bondolfi et al ldquoCerebral hemor-rhage after passive anti-a120573 immunotherapyrdquo Science vol 298no 5597 p 1379 2002
[63] M M Racke L I Boone D L Hepburn et al ldquoExacerbationof cerebral amyloid angiopathy-associatedmicrohemorrhage inamyloid precursor protein transgenic mice by immunotherapyis dependent on antibody recognition of deposited forms ofamyloid 120573rdquo Journal of Neuroscience vol 25 no 3 pp 629ndash6362005
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

6 BioMed Research International
3 Results
31 Efficacy of EB101 versus A12057342
+ CFAIFA in PreventingA120573 Burden Development Recently we demonstrated thatimmunization with EB101 vaccine prevents and reverses theAD neuropathology by the marked reduction of A120573 plaquesplaque-associated dystrophic neurites and astrocytosis inmice brain Now we conducted comparative immunizationstudies between EB101 and A120573
42+ FAIFA vaccines in the
prevention ofA120573deposits in themouse brainA120573plaques andvascular amyloid loadswere detected counted andmeasuredas the percent of total surface stained by monoclonal anti-A120573 antibody in the hippocampal and cortical sections Theplaque amyloid load observed in the representative photomi-crographs presented in Figure 2 shows a significant differenceacross the five experimental groups (ANOVA 119865 = 315119875 lt 0001) As a result of the first experimental phase basedon preventive treatment we observed that hippocampal loadwas significantly reduced in the EB101 immunized mice (AFigures 2(a) and 3) when compared with different vaccinecomponents (B-C Figures 2(b)-2(c) and 3) and moderatecompared with A120573
42+ CFAIFA vaccine (D Figures 2(d)
and 3) In fact the load in these treated groups (B-C) wasnot significantly different from PBS treated mice (E Figures2(e) and 3) although there is a positive correlation betweenthe presence of A120573 in treatment regime and the total A120573burden (Figures 2(b) 2(d) and 3) In the EB101 treated micea few compacted A120573 plaques were observed in retrosple-nial granular region of the cortex and at the polymorphdentate gyrus of the hippocampus (Figure 2(a)) Strikinglydifferent was the distribution of A120573 plaques observed inthe other treatment groups where numerous A120573 plaqueswere located at different cortical and hippocampal layers(Figures 2(b)ndash2(e)) This data was also supported by totalA120573 plaques density and size quantification made by imageanalysis software in hippocampal sections stained with anti-A120573 antibody (Figures 3 and 4) The data obtained in thepresent study validate our previous observations demon-strating that the mean burden A120573 plaques (Figures 2(a) and3) of Group A (25 plusmn 5 ps) were significantly different fromthe other treated groups (63 plusmn 6 ps in Figure 2(b) 65 plusmn6 ps in Figure 2(c) 56 plusmn 5 ps in Figure 2(d) 68 plusmn 6 psin Figures 2(e) and 3) This represents a decrease in themean percentage of A120573 plaque area of 301ndash382 in theEB101 immunized mice group relative to other tested groups(Figures 2(a)ndash2(j) and 4)The appearance of amyloid plaquesin the entorhinal (Figures 2(f)ndash2(j)) and piriform cortex wasalso analyzed being detected in low density in Group A(Figure 2(f))moderate density inGroups B (Figure 2(g)) andD (Figure 2(i)) and high density in the entorhinal cortexof Groups C (Figure 2(h)) and E (Figure 2(j)) Thereforethis data shows that in the preventive treatment periodthe clearance effect of the EB101 vaccine on A120573 plaques persection was about 64 (Figures 2(a) and 2(f)) compared topositive controls while in the other treatments regimes 19was detected in A120573
42+ CFAIFA (D) 10 in A120573
42liposome
without S1P (B) and 7 in liposome with S1P treated mice(C) Photomicrographs show that in all groups these plaques
were mainly localized in the external layers of the entorhinalcortex (Figures 2(f)ndash2(j)) In addition we have observedlower stained intensity of the A120573 plaques in the brainregions of EB101 treated mice which contrasts markedlywith the intensely stained A120573-immunoreactivity of theseplaques in each brain section of the other experimental micegroups
32 Efficacy of EB101 versus A12057342
+ CFAIFA in Clearing A120573Plaque Burden In a second set of experiments we comparedthe therapeutic effect of EB101 vaccine with A120573
42+ CFAIFA
in the reduction of A120573 in the brain after the plaques wereestablished in the 35-week-oldAPPPS1 transgenicmice (Fig-ures 2(k)ndash2(t))Themice were treated and analyzed using thesame protocol as in the preventive treatment To determinewhether EB101 vaccine attenuates andor reverses themassivedevelopment of 120573-amyloid plaques brain sections fromwild-type and transgenic mice of all experimental groupswere immunostained with the specific antibody recognizingA1205731ndash42 epitopesThese results show that A120573 deposits (Figures
2(k)ndash2(t)) were notably reduced in brain sections of theEB101 treated mice (A) markedly different from the A120573burden levels observed in the other treated groups (BndashE)As observed in the corresponding photomicrographs EB101treated mice showed a few compacted A120573 plaques mainlylocated at the CA1 layer of the hippocampus (Figure 2(k)) Noreduction in plaque density was observed in the other exper-imental groups (Figures 2(l)ndash2(o)) where numerous largecompacted A120573 plaques were located in almost every layer ofeach brain section The most densely populated section withA120573 plaques were those with no A120573
1ndash42 treatment as shownin Groups C (Figure 2(m)) and E (Figure 2(o)) while a slightreduction of this hallmarks was observed in mice Groups B(Figure 2(l)) and D (Figure 2(n)) The hippocampal sectionsof the therapeutic treatment with EB101 vaccine showed asimilar pattern as that observed in the preventive treatmentThemean burden of A120573 plaques in GroupA (35 plusmn 4 Figure 3)was significantly different from the other treated groups (70plusmn5 in B 74 plusmn 4 in C 65 plusmn 5 in D 80 plusmn 6 in E) The A120573 plaquearea in the EB101 immunized mice group was reduced by297ndash345 relative to the other tested groups (Figure 4) Asobserved in the preventive set of experiments this data showthat in the therapeutic treatment period the clearance effectof the EB101 vaccine on A120573 plaques per section was about59 compared to controls While in mice group treated withA12057342+ CFAIFA the clearance effect was about 20 in Group
D 13 in Group B and 8 in Group C The same pattern ofA120573-immunoreactivity was observed in the entorhinal cortexwhen comparing all experimental treatment groups (Figures2(p)ndash2(t)) Only a few A120573 plaques were observed in theexternal layer of the entorhinal and piriform cortex of micetreated with EB101 (Figure 2(p)) while numerous plaqueswere observed in the corresponding section of Groups B(Figure 2(q)) and D (Figure 2(s)) which show a moderatedensity of A120573 plaques while Groups C (Figure 2(r)) andE (Figure 2(t)) show densely distributed plaques along themedial and external layers of the entorhinal-piriform cortexAs described in the preventive treatment we also detected
BioMed Research International 7
Ect
Cx
CPu
Ect
Gr A
(EB1
01)
(a)
(c)
(b)
(f)
(h)
(g)
Preventive treatment Therapeutic treatment
CA1
PoDGMoDG
CA1
PoDGMoDG
CA1
PoDG
MoDG
DG
DG
Ect
Ect
Cx
Cx
Cx
CPu
CPu
CPuCA1
PoDG
MoDG
DG
CA1
PoDG
MoDG
DG
Ect
Cx
CPu
Ect
Cx
CPu
Cx
Cx
Cx
Cx
Cx
(k)
(l)
(p)
(r)
(q)
CA1
PoDGMoDG
DG
Cx
Ect
Cx
CPuCA1
ControlGr E
(PBS
)
(d)
(e) (j)
Control
PoDG
MoDG
DG
CA1
CA1
PoDG
MoDG
CA1
PoDG
MoDG
DG
DG
Ect
Ect
Cx
Cx
CPu
CPu
CA1
PoDG
MoDG
CA1
PoDGMoDG
DG
Ect
Cx
CPu
PoDGMoDG
DG
Cx
Cx
Cx
(n)
(o)
(s)
(t)
Cx
(i)
Gr B
(A120573
+L)
Gr D
(A120573+
FA)
A120573 A120573 A120573 A120573
A120573A120573A120573
A120573 A120573 A120573 A120573
A120573A120573A120573A120573
A120573 A120573 A120573 A120573
A120573
(m)
Gr C
(S1P
+L)
Figure 2 Comparative effect of EB101 vaccine on beta amyloid deposits Comparative photomicrographs of A120573 immunoreactivity weretaken in the hippocampus ((a)ndash(e) (k)ndash(o)) and cortical ((f)ndash(j) (p)ndash(t)) brain regions of transgenic mice with different treatments beforeA120573 plaques development preventive treatment ((a)ndash(j)) and after the A120573 plaques developed therapeutic treatment ((k)ndash(t)) Preventivetreatment transverse brain sections of 11-month-old mice show reduced number of A120573 deposits in the dentate gyrus hippocampal subregionCA1 (a) and entorhinal cortex (f) following EB101 vaccine immunization (Group A) contrasting with the numerous A120573 immunoreactiveplaques in the correspondent mouse brain sections of Groups B ((b) (g)) C ((c) (h)) D ((d) (i)) and E ((e) (j)) Immunoreactive A120573 in thebrain sections of wild-type (WT) mice is absent (inserts in panels (e) (o)) Therapeutic treatment transverse brain sections of 18-month-oldmice are shown in panels (k)ndash(t) Treatment with EB101 almost completely cleared A120573 load in the dentate gyrus and reduced notably thedensity in hippocampal subregion CA1 (k) and entorhinal cortex (p) compared to the same areas of mouse Groups B ((l) (q)) C ((m)(r)) D((n) (s)) and E ((o) (t)) as determined by number of plaques and staining intensity area Scale bar 100 120583m
8 BioMed Research International
Gr E
(PBS
)
0102030405060708090
Preventive treatmentTherapeutic treatment
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Mea
n of
120573-a
myl
oid
plaq
ues
sect
ion
Gr A
(A120573+
S1P+
L)(E
B101
)
Figure 3 Comparative analysis of A120573 plaque burden in the brainsof APPPS1 mice Mean of A120573 plaque burden in the hippocampaland cortical regions of APPswePS1ΔE9 mice in the five treatmentgroups The mean of the A120573 burden is significantly reduced (119875 lt005) in Group A (EB101) when compared with Groups BndashE duringpreventive treatment and markedly reduced in the therapeutictreatment period Data are presented asmeanplusmn standard error of themean (SEM) Densitymean of A120573 plaques in Tgmice hippocampus
05
1015202530354045
Preventive treatmentTherapeutic treatment
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Gr A
(A120573+
S1P+
L)(E
B101
)
Mea
n of
120573-a
myl
oid
plaq
ues a
rea
Figure 4 Comparative quantification of A120573 burden area in thebrain of APPPS1 mice Quantitative analysis of A120573 burden areain the hippocampal and cortical regions of APPswePS1ΔE9 micetreated with EB101 A120573liposomes S1Pliposomes A120573
42+ FAIFA
and PBS represented by the number of pixels inside the stained areaof each A120573 plaque This graphic shows that A120573 plaques of EB101-treated mice (Group A) are significantly smaller in size (119875 lt 005)than those of the other four treatment groups Data are presented asmean plusmn SEM Mean of A120573 plaques area in Tg mice hippocampus
a reduced staining intensity of the A120573 plaques in the brainregions of mouse Group A contrasting with the intenseA120573-immunoreactivity of plaques in the other experimentalmouse groups as shown in the photomicrographs of Figures2(k)ndash2(t)
33 Immune Response Effects of EB101 versus A12057342+ CFAIFA
Vaccine Astrocyte activation implicated in neuroinflam-mation amyloidogenesis and neuronal cell death in AD
was compared after treatment of the transgenic mice withEB101 and A120573
42+ CFAIFA Immunohistochemical analysis
of GFAP-reactive cells was carried out in the brains ofthe treated mice in the preventive and therapeutic studies(Figure 5) After the preventive treatment (Figures 5(a)ndash5(j))EB101 significantly reduced the density of GFAP-reactive cellclusters in the hippocampal and cortical sections as comparedwith A120573
42+ CFAIFA (D) A120573Lip without S1P (B) Lip with
S1P (C) and PBS (E) The distribution of GFAP immunore-active cells in the transverse section of the mouse brainstreated with EB101 shows a few scattered GFAP-reactiveclusters mainly at the CA1 hippocampal layers contrastingwith numerous dystrophic reactive astrocytes observed indifferent hippocampal areas of mouse brains in Groups BndashE(Figures 5(b)ndash5(e)) Similar neuroinflammation pattern wasobserved in the retrosplenial cortical layers where a fewGFAP-reactive clusters were observed in EB101 treated mice(Figure 5(f)) compared with their profuse presence in theother experimental groups (Figures 5(g)ndash5(j)) After thera-peutic treatment the GFAP immunoreactivity distributionpattern in mouse brains treated with EB101 was similar tocontrol mice (wild-type) mostly devoid of reactive GFAPclusters except a few scattered ones in the CA1 hippocampalexternal layers (Figure 5(k)) In contrast there is a moderatedensity in Group D (Figure 5(n)) and extensive density inGroup B (Figure 5(l)) Group C (Figure 5(m)) and Group Emice (Figure 5(o)) Photomicrographs of retrosplenial cortexshow similar neuroinflammation pattern where a few GFAPclusters detected in the EB101 treated mice (Figure 5(p))contrast with a more extensive density of clusters in the othergroups (Figures 5(q)ndash5(t)) No astrocytosis was observed inwild-type mice during preventive or therapeutic treatment
34 Comparative A120573 Antibody Titers and Th1Th2 CytokineLevels in Treated Mice A120573 antibody titers in the sera weredetermined by ELISA during mouse immunization(Figure 6) Preventive treatment with EB101 resulted in amarked increase of specific IgG A120573
1ndash42 antibody productionin all animals Mice in A and D groups developed andmaintained serum antibody titers between 1 2000 and1 8000 Mice in Groups B C and E showed no significantanti-A120573 antibody production Antibody production and titerin Group B were three and four times lower than those ofGroups D and A respectively After therapeutic treatment allmice treated with EB101 and A120573
42+ FAIFA produced high
levels of IgG antibodies Titer levels of Groups A and D weresignificantly increased to those detected after preventivetreatment (119875 lt 001 versus Group D 119875 lt 0001 versusGroups B C and E) although antibody level in Group D was13-fold lower than in Group A (Figure 6)The effect of EB101(Group A) and A120573
42+ CFAIFA (Group B) vaccines on Th1
andTh2 cytokines was also studied (Figure 7)Measurementsof Th1 and Th2 cytokines in sera of immunized transgenicmice indicate that significant changes occurring in proteinlevels of Th1-related inflammatory mediators followingpreventive (Figures 7(a)ndash7(c)) and therapeutic (Figures 7(d)ndash7(g)) treatment with EB101 vaccine A significant increaseof Th1 cytokine profile was observed in preventive treatment
BioMed Research International 9
Gr E
(PBS
)
(d)
(e)
(i)
(j)
(n)
(o)
(s)
(t)
Cx
GFAP
GFAP GFAP GFAP GFAP
GFAP GFAP GFAP
Cx Cx
Cx
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
GFAP
Gr A
(EB1
01)
(a)
(b)
(c)
(f)
(g)
(h)
Preventive treatment Therapeutic treatment
PoDG
MoDG
(k)
(l)
(m)
(p)
(q)
(r)DG
GFAP GFAP GFAP
GFAPGFAPGFAP GFAP
GFAP GFAP GFAP GFAP
PoDGDG
DG
CA1
Cx
Cx
Cx
Cx
Cx
Cx
PoDG
MoDG
DG
CA1
MoDGDG
CA1
PoDG
MoDG
DG
CA1
Cx
CxCx
MoDG
MoDG
PoDG
CA1CA1
Cx
Gr D
(A120573+
FA)
Gr C
(S1P
+L)
Gr B
(A120573+
L)
Figure 5 Comparative effect of EB101 vaccine on astrocytosis Comparative photomicrographs of GFAP immunoreactivity in thehippocampus ((a)ndash(e) (k)ndash(o)) and cortical ((f)ndash(j) (p)ndash(t)) brain regions at the preventive treatment phase ((a)ndash(j)) and therapeutictreatment group ((k)ndash(t)) Preventive treatment transverse brain sections of 11-month-old mice treated with EB101 show an almost completeprevention of astrocytosis in the detailed sections of the dentate gyrus (a) contrasting with numerous dystrophic reactive astrocytes typicalof an inflammatory reaction observed in different hippocampal corresponding areas of mouse brains in Group B (A120573Lip-treated mice (b))Group C (LipS1P-treated mice (c)) Group D (A120573
42+ CFAIFA-treated mice (d)) and Group E (PBS-treated mice (e)) Immunoreactive
astrocytosis in the brain sections of WT mice is absent (data not shown) Detailed sections of the parietal cortex of treated mice show agradient density of reactive astrocytes immunoreactive to GFAP indicating a neuroinflammation pattern Comparative cortical sections shownumerous astrocytosis areas inmice Groups C and E ((h) (j)) andwide reactive astrocytes clusters inmice Groups B andD ((g) (i)) while justa few of them are observed in Group A (f)Therapeutic treatment transverse brain sections of 18-month-old mice of Group A show a notablereduction of astrocytosis in the hippocampal region (k) after EB101 vaccine immunization Note the astrocytosis contrast density betweenEB101-treatedmice and the other treated groups ((l)ndash(o)) where numerous immunoreactive GFAP clusters are observed in the correspondentbrain sections A few immunoreactive GFAP clusters are observed in the parietal cortex of EB101 treated mice group while a moderate ((q)(s)) to high ((r) (t)) density of these neuropathological inflammation clusters is apparent in the other treatment groups (Groups BndashE) Forabbreviations see list Scale bar 100120583m
10 BioMed Research International
002040608
1121416
Preventive treatmentTherapeutic treatment
Ant
i-120573-a
myl
oid
IgG
antib
odie
s
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Gr A
(A120573+
S1P+
L)(E
B101
)
Figure 6 Quantification of specific anti-120573-amyloid IgG in immu-nized APPPS1 mice The presence of anti-120573-amyloid antibodiesmeasured in the sera of vaccinated and control mice groups duringthe preventive and therapeutic treatments indicates that EB101liposome vaccine was able to induce the highest production ofspecific IgG antibodies Compared to theGroupD immunizedmicethe level of antibodies observed in the Group A was also highersuggesting a more effective immune response (119875 lt 001 versusGroup D 119875 lt 0001 versus Groups B C and E) Quantification ofspecific anti-120573-amyloid IgG in immunized Tg mice
in Group D with respect to all other groups mainlyreported by the high levels of TNF-120572 (Figure 7(b)) andIFN-120574 (Figure 7(c)) cytokine secretions while due to thehigh mortality rate during the therapeutic treatment nosignificant data was obtained from this mice group Inaddition a tendency toTh1 secretion reduction was observedinGroupAwith respect to all other groups in both preventiveand therapeutic treatment The changes in Th2 protein levels(Figure 7(d)) followed the inverse trend where the highestdifferences between Group A and the other treatment groupswere observed in the IL-4 IL-5 and IL-10 cytokine secretions(119875 lt 005 versus Group D 119875 lt 001 versus Group E Figures7(e)ndash7(g))
35 Comparative Motor Coordination and Memory Perfor-mances In order to validate the locomotion integrity duringthe entire immunotherapeutic process as reported in similarprevious studies motor coordination tests have showed thatEB101-treated mice spent significantly more time on theaccelerating rotarodThismotor coordination task shows thatEB101 immunization affected positively their performance(Figure 8(a)) A120573
42liposome (B) S1P-containing liposomes
(C) and PBS (E) (119875 lt 0006) mice performed poorly incomparison to EB101 mice (119875 lt 005) or A120573
42+ CFAIFA
mice (119875 lt 002) in motor abilities and coordination Thefear conditioning-active avoidance test was used to assess thespatial learning and memory functions As expected bothEB101 and A120573
42+ CFAIFA immunized mice readily learned
the location of the platform as shown by a decrease in theescape latency over training days while the control group
mice exhibited no significant changes during 6-day trialsThe average escape latencies of EB101 (68 plusmn 2 s 761 plusmn 22 s)andA120573
42+CFAIFA (116plusmn33 s 123plusmn21 s) immunizedmice
at the end of preventive and therapeutic periods respectivelyduring the consecutive 6 days trial were significantly shorterthan the other treatment groups (A120573
42liposome 163 plusmn 31 s
175 plusmn 28 s S1P-containing liposomes 202 plusmn 19 s 248 plusmn27 s) and the control (2116 plusmn 38 s 268 plusmn 22 s) (119875 lt 001)Therewas no significant difference between EB101 andA120573
42+
CFAIFA immunized group (119875 gt 005) (Figure 8(b)) Theseresults showed that spatial learning was improved in theseimmunized mice
4 Discussion
In this study we have compared a novel active A120573 imm-unotherapy the EB101 vaccine against A120573 with the A120573
42+
CFAIFA vaccine that was first taken into clinical trialswith coadjuvant Qs-21 modification (AN1792 by Orgogozoet al [19]) This type of vaccine design was thought toinduce a strong T-cell mediated inflammatory response (IL-2 and IFN-c positive responses indicative of a Class II(CD4) Th-1 type response [33] leading to acute menin-goencephalitis in the 6 of the patients) The EB101 activeimmunotherapeutic vaccine was primarily designed to avoidundesired inflammatory side effects both in mice andhumans (without need of coadjuvant modification) whileinducing effective antibody titer rates We demonstrated thatlong-term immunization with EB101 induced A120573 antibodiesin the preventive and the therapeutic treatment periodswhich effectively reduced amyloid deposition in both corticaland hippocampal brain regions and prevented neuroinflam-mation probably by blocking toxicity in the surroundingneurons However the AN1792 vaccine was shown to inducein 6 of the patients an inflammatory reaction that mighthave led to the encephalitis type reaction observed in theclinical trial which was responsible for the immediate haltin those clinical studies It seems that AN1792 immunizationdelivered in Qs-21 and polysorbate 80 adjuvant induced aTh1 type response that likely led to autoaggressive T cells andaseptic meningoencephalitis In the present experiments wedemonstrated that a judiciously selected adjuvant produces adifferent humoral response [22] Indeed we have shown thatour novel immunotherapeutic strategy boosts specific A120573
1ndash42antibody generation and induces a Th2 noninflammatoryimmune response This boosted effect is in accordance withprevious immunotherapeutic studies [34] Taking togetherall the present findings these results suggest that EB101vaccine can achieve in a future clinical trial immunogenicresponse rates quite similar to the AN1792 without theinflammatory effects reported The immunohistochemicalanalysis obtained in the present study shows that all the EB101immunized mice resulted in an almost complete preventionof A120573 deposition in the brain with only a few compactA120573 plaques in the retrosplenial and dentate gyrus regionsAlthough there was a slightly increase in death in EB101treated mice compared to controls this significant reductionof A120573 plaques in EB101 treated mice was proven to be more
BioMed Research International 11
Gr A Gr B Gr C Gr D Gr EPreventive treatmentTherapeutic treatment
0005001000150020002500300035004000
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
(a) Th1
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
000
1000
2000
3000
4000
5000TNF-120572
(b) Th1
000500
10001500200025003000
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
IFN-120574
(c) Th1
000500
100015002000250030003500
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
Group A Group B Group C Group D Group EPreventive treatmentTherapeutic treatment
(d) Th2
000
500
1000
1500
2000
2500
3000IL-4
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(e) Th2
000500
100015002000250030003500
IL-5
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(f) Th2
3500IL-10
000500
10001500200025003000
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(g) Th2
Figure 7 Detection of Th1 and Th2 cytokine profile Measurements of the effect of EB101 and A12057342
+ CFAIFA vaccines on Th1 and Th2cytokines profile during preventive and therapeutic treatments ((a)ndash(c)) A significant increase of Th1 cytokine profile was observed inpreventive treatment in Group D with respect to all other groups while a tendency to a reduction inTh1 secretion was observed in Group Awith respect to all other groups in both preventive and therapeutic treatment ((d)ndash(g)) The changes in Th2 protein levels were observed inthe IL-4 IL-5 and IL-10 cytokine secretions of Group A (119875 lt 005 versus Group D 119875 lt 001 versus Group E)
12 BioMed Research International
Gr A
(EB1
01)
05
101520253035
Preventive treatment Therapeutic treatment
Mea
n fa
lling
late
ncy
(s)
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)(a) Graphic of motor coordination and balance
05
1015202530
Mea
n re
spon
se ti
me (
s)
Gr A
(EB1
01)
Preventive treatment Therapeutic treatment
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
(b) Graphic of associative learning and memory
Figure 8 Analysis of motor coordination and balance (a) Rotarod results of motor coordination and balance test after the preventive andtherapeutic treatment showing improved effect of EB101 and A120573
42+ CFAIFA-vaccine compared to other treated mice groups (119875 lt 005)
and (119875 lt 002) versus control mice Latency to fall on the rotarod apparatus was significantly improved in EB101 immunized mice Valuesrepresent means of all six trials of a groupday (b) Spatial learning and memory tested by fear conditioning-active avoidance test The meanresponse time of each treatment group to reach the platform is represented EB101 and A120573
42+ CFAIFA immunized mice had significantly
lower latencies than the other treatment groups (119875 lt 001) versus control mice
efficient than that obtained with A12057342+ CFAIFA immuniza-
tions A comparison of A120573 plaque reduction rates amongtreated groups surprisingly showed that other than EB101and A120573
42+ CFAIFA treated groups some amyloid deposits
were also reduced in Groups B (A1205731ndash42liposome) and C
(S1P-containing liposomes) Although the reduction rates inthese mice were markedly low the subtle lowering effectmight be due to the interaction between the immunizationelements (A120573
1ndash42 and S1P) and the mouse immunogenicsystem as reported by others [35] Based on our presentresults S1P one of the components of the adjuvant used withEB101 might be responsible for the neuroprotective effectobserved in the vulnerable neuronal areas of EB101 treatedmice The advantage in the A120573 load preventionreduction bycombining A120573
1ndash42 and S1P in a unique formulation vaccinehas been also observed in the reduced hippocampal load ofEB101 immunizedmice (Gr A) when comparedwith differentvaccine components (Gr B-C) In fact S1P appears to have ademonstrated therapeutic potential as a regenerative agent inthe nervous system since it controls migration of neuronalstem cells toward a site of spinal cord injury [36] inducecytoskeletal rearrangements to promote transmitter secretion[37] and also trigger diverse cellular effects including angio-genesis cardiac development immunity cell motility andneurite extension [38 39]Therefore it is reasonable to expectthat such a potent biologically active sphingolipid by itselfmay play an important role in the A120573 plaque reduction Inthis study the clearance efficiency rate of each treatment wasanalyzed and results suggested that EB101 (6459) appearsto be more effective than A120573
42+ CFAIFA (1920) vaccine
in both preventive and therapeutic groups The effectivenessof EB101 in older transgenic animals (18 months) showeda clearance effect of 59 suggesting that EB101 may alsohave therapeutic potential inADdiagnosticated patient trials
Similar conclusion was reached byWang and colleagues [40]who demonstrated that combination therapy in aged animalsprovides more valuable insights into a possible translationto humansOur immunologicalmeasurements demonstratedonce again that neuroinflammation develops in an A120573-dependent fashion early in the course of AD [41] and canbe alleviated or reversed by interventional A120573 vaccinationsuch as EB101 Among the possible mechanisms of A120573clearance proposed in AD immunotherapy [42] our resultssuggested that theymay overlap and probably be disease statedependent since the prevention and the therapeutic vaccinewould likely benefit from the transport of A120573 antibodies intothe CNS However some mechanisms of action such as thepreventive effect of A120573 antibodies in the A120573 aggregation [43]microglial phagocytosis induced by the Fc binding portionsbetween A120573 antibodies and microglia receptor [44] and theshift in the gradient of A120573 transport across the blood-brainbarrier due to the increased concentration of A120573 antibodiesin the vascular system resulting in an increase in effluxfrom the brain to blood [45 46] may be playing a role inboth preventive and therapeutic conditions Our findingstherefore suggest that novel treatment strategies aimed atmaintaining or increasing microglial function and restoringthe balance to the immunological system may represent anattractive therapeutic approach even at the advanced stagesof ADThe immunization results obtained with EB101 duringboth preventive and therapeutic treatment showed nomicro-hemorrhages infiltration of lymphocytes or encephalitis-like reaction in the brain suggesting that EB101 does notinduce A120573-specific Th1 reaction possibly due to the neu-roprotective effect of S1P Delivering A120573
1ndash42 in a liposomal-S1P-adjuvant resulted in an effective immunological responseincluding the induction of a Th2 T-cell response whicheffectively prevented neuroinflammation in an animal model
BioMed Research International 13
of AD as reported previously [24 47] Moreover presentresults demonstrated that EB101 did not induce astrogliosisin APPPS1 transgenic mice whereas some GFAP-positiveastrocyte clusters were aggregated in the affected brain areasof mice treated with A120573
42+ CFAIFA These results indicate
that EB101 may result in an effective treatment in AD neu-ropathology due to the absence of peripheral proinflamma-tory cytokine secretion and the absence of astroglial activa-tion in the brains of EB101 treated mice when compared withA12057342
+ CFAIFA immunotherapeutic response In a clinicalsetting however we may expect a wide differential responsein part depending upon the genomic profile of each patient[48 49] In AD behavioural studies in particular amongthe motor coordination tests the rotarod performance hasbeen reported to be impaired in some AD transgenic micemodels bearing A120573 plaques [50 51] This result was found inmutants prior to A120573 plaque formation at 3 and 6 months ofage but was not reproduced after plaques had been formedat 16 and 24 months of age [52] perhaps because of thedeclining abilities of older normal mice Here we presentthe effects of vaccine treatments in APPPS1 mice on motorcoordination a neglected aspect of the AD experimentalbehavior but justified on the basis of the deficient posturalcontrol reported in patients with Alzheimerrsquos disease [53]sometimes even during early stages [54] Our longitudinalmotor behavioral results showed no significant differencesin the ambulatory movement grip strength or coordinationaddressed by their performance in the rotating rod betweenAPPPS1 and wild-type mice for any age group studiedin contrast with other treated groups demonstrating thatlongitudinal motor assessments of AD transgenic mousemodels can provide valuable information with regard tothe effect of immunization on the progression of behavioraldeficits and overall motor activity As shown the EB101-treated group of mice exhibited longer latencies and betterbalancing skills indicating the absence of any negative effectof EB101 immunization on these mice The classical fearconditioning test is assumed to reveal the functional integrityof the hippocampus [55] representing a special memorytask to evaluate the AD models and its improvement afterimmunization periods In this APPPS1 mouse model thefirst associative learning and spatial workingmemory deficitsare shown at 5-6 months of age [56] although its impair-ment progression is relatively slow compared to most othermodels At the end of preventive immunization period EB101and A120573
42+ CFAIFA immunized mice have significantly
shorter escape latencies than the other treatment groups(A12057342liposome S1P-containing liposomes) and the controls
in the fear conditioning test Shorter escape latencies duringspatial learning indicated that deficits of spatial memorywere attenuated in EB101 and A120573
42+ CFAIFA immunized
mice Previous studies have showed that by 15 months ofage the spatial working memory is consistently impairedthroughout the rest of the life span [31 57] At the end oftherapeutic immunization period both EB101 and A120573
42+
CFAIFA immunized mice showed significantly improvedresults on the hippocampus-dependent spatial learning andmemory test when compared with other treatment groupsand the controls Taken together these observations are
in agreement with the impairment in conditioned learningin response to a tone stimulus of aged APPPS1 micealthough when immunized with EB101 during preventiveor therapeutic periods such impairments are significantlyattenuated
In summary the present results corroborate our previouspublished findings showing that EB101 immunization reducesamyloid accumulation in the APPPS1 mouse brain [2425] The strongest reduction in the preventive immunizationwas observed in the retrosplenial cortex followed by thehippocampus and entorhinal-piriform cortex Our resultsfurther support that EB101 ismore effectivewhen themice areimmunized prior to developing plaques in the hippocampaland cortical brain regions There is extensive evidence inmouse preclinical studies [58] and human clinical trials [5960] that the adverse effects of active and passive A120573 vaccinesresult in microhemorrhages and encephalitic reactions [60ndash63] yet themechanismbywhich they occur has not been fullyelucidated In the treatment sections analyzed we found nomicrohemorrhages or acute inflammatory processes in all thesurvived mice of the group indicating that EB101 therapeuticapproach presents no adverse inflammatory effects in thisAD mice models Thus our present results unequivocallydemonstrate that EB101 vaccinemay not only halt AD-relatedpathology as a preventive treatment but may also reverse it inestablished cases related to the improvement of the learningandmemory in both cases leading to a robust immunologicaltherapeutic response Present experiments also demonstratethat EB101 exerts more significant and substantial effects onthe reduction of A120573 deposition than A120573
42+CFAIFA vaccine
in A120573 mice models of AD However further exploration ofthe contribution of genetic environmental and physiologicalfactors in the development resistance to endogenous A120573-antibody is needed in order to optimize and modulate thetherapeutic strategies currently being developed
5 Conclusions
The present study carried out in APPPS1 transgenic mousemodel of AD compared the A120573 burden reduction andcellular immune responses produced by two immunizationdelivery strategies of A120573 in Freund adjuvant (described byWebster et al [10]) and in liposomeS1P matrix (describedby Schenk et al [17])The EB101 immunotherapeutic strategyboosted antibody generation A120573 plaque reduction and ananti-inflammatory response more effectively than A120573
42+
CFAIFA especially if given prior to or in the early stages ofpathology The present results also showed that vaccinationwith EB101 prevented the formation of new plaques andpresumably led to a reduction in established plaques whileinducing aTh2 immune reactionwithout encephalitogenic T-cell responses or brain inflammation related with astrogliosisHowever despite the efficacy demonstrated of this novelvaccine approach further preclinical and clinical studieson EB101 are warranted to evaluate its efficacy and themechanism for the future utility of such vaccine for ADtherapy
14 BioMed Research International
List of Abbreviations
AD Alzheimerrsquos diseaseA120573 120573-Amyloid proteinAPP Amyloid precursor proteinAPT Anterior pretectal nucleusCA1 Field CA1 hippocampusCA3 Field CA3 hippocampusCPu Caudate putamen (striatum)Cx CortexDG Dentate gyrusDN Dystrophic neuriteEct Entorhinal cortexCFA Complete Freundrsquos adjuvantGrDG Granular dentate gyrusIFA Incomplete Freundrsquos adjuvantIL InterleukinLPMR Lateral posterior thalamic nucleus (mediorostral)MoDG Molecular dentate gyrusPir Piriform cortexPoDG Polymorph dentate gyrusRSGb Retrosplenial granular cortex b regionTg TransgenicTh T helper
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Ivan Carrera Ignacio Etcheverrıa Lucıa Fernandez-NovoaValter Lombardi Ramon Cacabelos and Carmen Vigo con-ceived and designed the experiments Ivan Carrera and Igna-cio Etcheverrıa performed the experiments Ivan CarreraIgnacio Etcheverrıa and Valter Lombardi analyzed the dataIvan Carrera Ignacio Etcheverrıa Lucıa Fernandez-Novoaand Valter Lombardi contributed reagentsmaterialsanalysistools Ivan Carrera Carmen Vigo andMadepalli KrishnappaLakshmana wrote the paper
Acknowledgments
This work was supported by National Institute of Aging(NIA)NIH Grant no 1R01AG036859-01 (Madepalli Krish-nappa Lakshmana) and the financial support of the ProgramaOperativo Feder Galicia 2007ndash2013 (IN841C 201254 to PhDCarrera I)
References
[1] R Cacabelos L Fernandez-Novoa V Lombardi Y Kubotaand M Takeda ldquoMolecular genetics of Alzheimerrsquos diseaseand agingrdquoMethods and Findings in Experimental and ClinicalPharmacology vol 27 pp 1ndash573 2005
[2] P T Nelson I Alafuzoff E H Bigio et al ldquoCorrelationof alzheimer disease neuropathologic changes with cognitive
status a review of the literaturerdquo Journal of Neuropathology andExperimental Neurology vol 71 no 5 pp 362ndash381 2012
[3] JNaslundVHaroutunian RMohs et al ldquoCorrelation betweenelevated levels of amyloid 120573-peptide in the brain and cognitivedeclinerdquo Journal of the American Medical Association vol 283no 12 pp 1571ndash1577 2000
[4] A Koyama O I Okereke T Yang D Blacker D J Selkoeand F Grodstein ldquoPlasma amyloid-120573 as a predictor of dementiaand cognitive decline a systematic review and meta-analysisrdquoArchives of Neurology vol 69 no 7 pp 824ndash831 2012
[5] A M Fagan C M Roe C Xiong M A Mintun J C Morrisand D M Holtzman ldquoCerebrospinal fluid tau120573-amyloid42ratio as a prediction of cognitive decline in nondemented olderadultsrdquo Archives of Neurology vol 64 no 3 pp 343ndash349 2007
[6] S Parvathy P Davies V Haroutunian et al ldquoCorrelationbetween A120573-40- A120573-42- and A120573x-43-containing amyloidplaques and cognitive declinerdquo Archives of Neurology vol 58no 12 pp 2025ndash2032 2001
[7] E Karran M Mercken and B D Strooper ldquoThe amyloidcascade hypothesis for Alzheimerrsquos disease an appraisal for thedevelopment of therapeuticsrdquo Nature Reviews Drug Discoveryvol 10 no 9 pp 698ndash712 2011
[8] J L Jankowsky D J Fadale J Anderson et al ldquoMutantpresenilins specifically elevate the levels of the 42 residue 120573-amyloid peptide in vivo evidence for augmentation of a 42-specific 120574 secretaserdquo Human Molecular Genetics vol 13 no 2pp 159ndash170 2004
[9] L Holcomb M N Gordon E Mcgowan et al ldquoAcceleratedAlzheimer-type phenotype in transgenic mice carrying bothmutant amyloid precursor protein and presenilin 1 transgenesrdquoNature Medicine vol 4 no 1 pp 97ndash100 1998
[10] S JWebster A D Bachstetter and L J van Eldik ldquoComprehen-sive behavioral characterization of an APPPS-1 double knock-in mouse model of Alzheimerrsquos diseaserdquo Alzheimerrsquos ResearchandTherapy vol 5 no 3 article 28 2013
[11] H Xiong D Callaghan J Wodzinska et al ldquoBiochemical andbehavioral characterization of the double transgenic mousemodel (APPswePS1dE9) of Alzheimerrsquos diseaserdquo NeuroscienceBulletin vol 27 no 4 pp 221ndash232 2011
[12] M Garcia-Alloza L A Borrelli B T Hyman and B J BacskaildquoAntioxidants have a rapid and long-lasting effect on neuriticabnormalities in APPPS1 micerdquo Neurobiology of Aging vol 31no 12 pp 2058ndash2068 2010
[13] M Meyer-Luehmann M Mielke T L Spires-Jones et al ldquoAreporter of local dendritic translocation shows plaque-relatedloss of neural system function in APP-transgenic micerdquo Journalof Neuroscience vol 29 no 40 pp 12636ndash12640 2009
[14] E Machova V Rudajev H Smyckova H Koivisto H Tanilaand V Dolezal ldquoFunctional cholinergic damage develops withamyloid accumulation in young adult APPswePS1dE9 trans-genic micerdquo Neurobiology of Disease vol 38 no 1 pp 27ndash352010
[15] T Malm J Koistinaho and K Kanninen ldquoUtilization of APP-swePS1dE9 transgenic mice in research of Alzheimerrsquos diseasefocus on gene therapy and cell-based therapy applicationsrdquoInternational Journal of Alzheimerrsquos Disease vol 2011 Article ID517160 8 pages 2011
[16] J Lee and P Han ldquoAn update of animal models of alzheimerdisease with a reevaluation of plaque depositionsrdquoExperimentalNeurobiology vol 22 pp 84ndash95 2013
BioMed Research International 15
[17] D Schenk R Barbour W Dunn et al ldquoImmunization withamyloid-120573 attenuates Alzheimer disease-like pathology in thePDAPP mouserdquo Nature vol 400 no 6740 pp 173ndash177 1999
[18] H J Fu B Liu J L Frost and C A Lemere ldquoAmyloid-120573immunotherapy for Alzheimerrsquos diseaserdquoCNS and NeurologicalDisorders-Drug Targets vol 9 no 2 pp 197ndash206 2010
[19] J-M Orgogozo S Gilman J-F Dartigues et al ldquoSubacutemeningoencephalitis in a subset of patients with AD after A120573
42
immunizationrdquo Neurology vol 61 no 1 pp 46ndash54 2003[20] J A R Nicolll DWilkinson CHolmes P Steart HMarkham
andROWeller ldquoNeuropathology of humanAlzheimer diseaseafter immunization with amyloid-120573 peptide a case reportrdquoNature Medicine vol 9 no 4 pp 448ndash452 2003
[21] I Ferrer M Boada Rovira M L Sanchez Guerra M JRey and F Costa-Jussa ldquoNeuropathology and pathogenesisof encephalitis following amyloid-Neuropathology and patho-genesis of encephalitis following amyloid-beta immunizationin Alzheimerrsquos disease immunization in Alzheimerrsquos diseaserdquoBrain Pathology vol 14 no 1 pp 11ndash20 2004
[22] D H Cribbs A Ghochikyan V Vasilevko et al ldquoAdjuvant-dependent modulation ofTh1 andTh2 responses to immuniza-tion with 120573-amyloidrdquo International Immunology vol 15 no 4pp 505ndash514 2003
[23] C Holmes D Boche D Wilkinson et al ldquoLong-term effectsof A120573
42immunisation in Alzheimerrsquos disease follow-up of a
randomised placebo-controlled phase I trialrdquo The Lancet vol372 no 9634 pp 216ndash223 2008
[24] I Carrera I Etcheverrıa L Fernandez-Novoa V LombardiR Cacabelos and C Vigo ldquoVaccine development to treatAlzheimerrsquos disease neuropathology in APPPS1 transgenicmicerdquo International Journal of Alzheimerrsquos Disease vol 2012Article ID 376138 17 pages 2012
[25] I Carrera I Etcheverrıa Y Li et al ldquoImmunocytochemicalcharacterization of Alzheimer disease hallmarks in APPPS1transgenic mice treated with a new anti-amyloid-120573 vaccinerdquoBioMed Research International vol 2013 Article ID 709145 12pages 2013
[26] V Anelli R Bassi G Tettamanti P Viani and L Riboni ldquoExtra-cellular release of newly synthesized sphingosine-1-phosphateby cerebellar granule cells and astrocytesrdquo Journal of Neuro-chemistry vol 92 no 5 pp 1204ndash1215 2005
[27] S Schroeter K Khan R Barbour et al ldquoImmunotherapyreduces vascular amyloid-120573 in PDAPP micerdquo Journal of Neu-roscience vol 28 no 27 pp 6787ndash6793 2008
[28] Z Suo A Cox N Bartelli et al ldquoGRK5 deficiency leads to earlyAlzheimer-like pathology and working memory impairmentrdquoNeurobiology of Aging vol 28 no 12 pp 1873ndash1888 2007
[29] W Kamphuis C Mamber MMoeton et al ldquoGFAP isoforms inadult mouse brain with a focus on neurogenic astrocytes andreactive astrogliosis in mouse models of Alzheimer diseaserdquoPLoS ONE vol 7 no 8 Article ID e42823 2012
[30] J Lang C Vigo-Pelfrey and F Martin ldquoLiposomes composedof partially hydrogenated egg phosphatidylcholines fatty acidcomposition thermal phase behavior and oxidative stabilityrdquoChemistry and Physics of Lipids vol 53 no 1 pp 91ndash101 1990
[31] D Morgan D M Diamond P E Gottschall et al ldquoA 120573peptide vaccination prevents memory loss in an animal modelof Alzheimerrsquos diseaserdquo Nature vol 408 no 6815 pp 982ndash9852000
[32] M L Brownlow L Benner D DrsquoAgostino M N Gordon andD Morgan ldquoKetogenic diet improves motor performance but
not cognition in two mouse models of Alzheimerrsquos pathologyrdquoPLoS ONE vol 8 no 9 Article ID e75713 2013
[33] M Pride P Seubert M Grundman M Hagen J Eldridgeand R S Black ldquoProgress in the active immunotherapeu-tic approach to Alzheimerrsquos disease clinical investigationsinto AN1792-associated meningoencephalitisrdquo Neurodegenera-tive Diseases vol 5 no 3-4 pp 194ndash196 2008
[34] CWiessner K-HWiederhold A C Tissot et al ldquoThe second-generation activeA120573 immunotherapyCAD106 reduces amyloidaccumulation in APP transgenic mice while minimizing poten-tial side effectsrdquoThe Journal of Neuroscience vol 31 no 25 pp9323ndash9331 2011
[35] D Lambracht-Washington and R Rosenberg ldquoAdvances in thedevelopment of vaccines for Alzheimerrsquos diseaserdquo DiscoveryMedicine vol 15 no 84 pp 319ndash326 2013
[36] A Kimura T Ohmori R Ohkawa et al ldquoEssential roles ofsphingosine 1-phosphateS1P
1receptor axis in the migration of
neural stem cells toward a site of spinal cord injuryrdquo Stem Cellsvol 25 no 1 pp 115ndash124 2007
[37] S S Lee C H Woo J D Chang and J H Kim ldquoRoles of Racand cytosolic phospholipase A2 in the intracellular signalling inresponse to titanium particlesrdquo Cellular Signalling vol 15 no 3pp 339ndash345 2003
[38] E Masliah L Hansen A Adame et al ldquoA120573 vaccination effectson plaque pathology in the absence of encephalitis inAlzheimerdiseaserdquo Neurology vol 64 no 1 pp 129ndash131 2005
[39] A Pilorget B Annabi F Bouzeghrane J Marvaldi J Luisand R Beliveau ldquoInhibition of angiogenic properties of brainendothelial cells by platelet-derived sphingosine-1-phosphaterdquoJournal of Cerebral Blood Flow and Metabolism vol 25 no 9pp 1171ndash1182 2005
[40] A Wang P Das R C Switzer III T E Golde and J LJankowsky ldquoRobust amyloid clearance in a mouse model ofAlzheimerrsquos disease provides novel insights into themechanismof amyloid-120573 immunotherapyrdquo Journal of Neuroscience vol 31no 11 pp 4124ndash4136 2011
[41] G Krabbe A Halle V Matyash et al ldquoFunctional impairmentof microglia coincides with Beta-amyloid deposition in micewith Alzheimer-like pathologyrdquo PLoS ONE vol 8 no 4 ArticleID e60921 2013
[42] Y H Liu B Giunta H-D Zhou J Tan and Y J WangldquoImmunotherapy for Alzheimer disease-the challenge ofadverse effectsrdquo Nature Reviews Neurology vol 8 no 8 pp465ndash469 2012
[43] B Solomon R Koppel D Frankel and E Hanan-AharonldquoDisaggregation of Alzheimer120573-amyloid by site-directedmAbrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 94 no 8 pp 4109ndash4112 1997
[44] F Bard C Cannon R Barbour et al ldquoPeripherally adminis-tered antibodies against amyloid 120573-peptide enter the centralnervous system and reduce pathology in a mouse model ofAlzheimer diseaserdquo Nature Medicine vol 6 no 8 pp 916ndash9192000
[45] R B DeMattos K R Bales D J Cummins J C Dodart S MPaul and D M Holtzman ldquoPeripheral anti-A120573 antibody altersCNS and plasma A120573 clearance and decreases brain A120573 burdenin a mouse model of Alzheimerrsquos diseaserdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 98 no 15 pp 8850ndash8855 2001
[46] C A Lemere E T Spooner J LaFrancois et al ldquoEvidence forperipheral clearance of cerebral A120573 protein following chronic
16 BioMed Research International
active A120573 immunization in PSAPP micerdquo Neurobiology ofDisease vol 14 no 1 pp 10ndash18 2003
[47] T Town M Vendrame A Patel et al ldquoReduced Th1 andenhanced Th2 immunity after immunization with Alzheimerrsquos120573-amyloid
1minus42rdquo Journal of Neuroimmunology vol 132 no 1-2
pp 49ndash59 2002[48] R Cacabelos ldquoPharmacogenomics in Alzheimerrsquos diseaserdquo
Methods in Molecular Biology vol 448 pp 213ndash357 2008[49] R Cacabelos ldquoPharmacogenomics and therapeutic strategies
for dementiardquo Expert Review of Molecular Diagnostics vol 9no 6 pp 567ndash611 2009
[50] D van Dam R DrsquoHooge M Staufenbiel C van Ginneken Fvan Meir and P P de Deyn ldquoAge-dependent cognitive declinein the APP23 model precedes amyloid depositionrdquo EuropeanJournal of Neuroscience vol 17 no 2 pp 388ndash396 2003
[51] M Dumont C Strazielle M Staufenbiel and R LalondeldquoSpatial learning and exploration of environmental stimuli in24-month-old female APP23 transgenic mice with the Swedishmutationrdquo Brain Research vol 1024 no 1-2 pp 113ndash121 2004
[52] R Lalonde M Dumont M Staufenbiel C Sturchler-Pierratand C Strazielle ldquoSpatial learning exploration anxiety andmotor coordination in female APP23 transgenic mice with theSwedish mutationrdquo Brain Research vol 956 no 1 pp 36ndash442002
[53] F J Huff F Boller F Lucchelli R Querriera J Beyer and SBelle ldquoThe neurologic examination in patients with probableAlzheimerrsquos diseaserdquo Archives of Neurology vol 44 no 9 pp929ndash932 1987
[54] A F Pettersson M Engardt and L O Wahlund ldquoActivitylevel and balance in subjects with mild Alzheimerrsquos diseaserdquoDementia and Geriatric Cognitive Disorders vol 13 no 4 pp213ndash216 2002
[55] L Calandreau B Desgranges R Jaffard and A DesmedtldquoSwitching from contextual to tone fear conditioning and viceversa the key role of the glutamatergic hippocampal-lateralseptal neurotransmissionrdquo Learning amp Memory vol 17 no 9pp 440ndash443 2010
[56] F Trinchese S Liu F Battaglia SWalter PMMathews andOArancio ldquoProgressive age-related development of Alzheimer-like pathology in APPPS1 micerdquo Annals of Neurology vol 55no 6 pp 801ndash814 2004
[57] D M Wilcock A Rojiani A Rosenthal et al ldquoPassiveimmunotherapy against A120573 in aged APP-transgenic micereverses cognitive deficits and depletes parenchymal amyloiddeposits in spite of increased vascular amyloid and microhem-orrhagerdquo Journal of Neuroinflammation vol 1 article 24 2004
[58] S A Ferguson S Sarkar and L C Schmued ldquoLongitudinalbehavioral changes in the APPPS1 transgenic AlzheimerrsquosDisease modelrdquo Behavioural Brain Research vol 242 no 1 pp125ndash134 2013
[59] D Wilcock and C Colton ldquoImmunotherapy vascular pathol-ogy and microhemorrhages in transgenic micerdquo CNS andNeurological DisordersmdashDrug Targets vol 8 no 1 pp 50ndash642009
[60] M Grundman S Gilman R S Black et al ldquoAn alternativemethod for estimating efficacy of the AN1792 vaccine foralzheimer diseaserdquo Neurology vol 71 no 9 pp 697ndash698 2008
[61] D Boche E Zotova R O Weller et al ldquoConsequence of A120573immunization on the vasculature of human Alzheimerrsquos diseasebrainrdquo Brain vol 131 no 12 pp 3299ndash3310 2008
[62] M Pfeifer S Boncristiano L Bondolfi et al ldquoCerebral hemor-rhage after passive anti-a120573 immunotherapyrdquo Science vol 298no 5597 p 1379 2002
[63] M M Racke L I Boone D L Hepburn et al ldquoExacerbationof cerebral amyloid angiopathy-associatedmicrohemorrhage inamyloid precursor protein transgenic mice by immunotherapyis dependent on antibody recognition of deposited forms ofamyloid 120573rdquo Journal of Neuroscience vol 25 no 3 pp 629ndash6362005
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
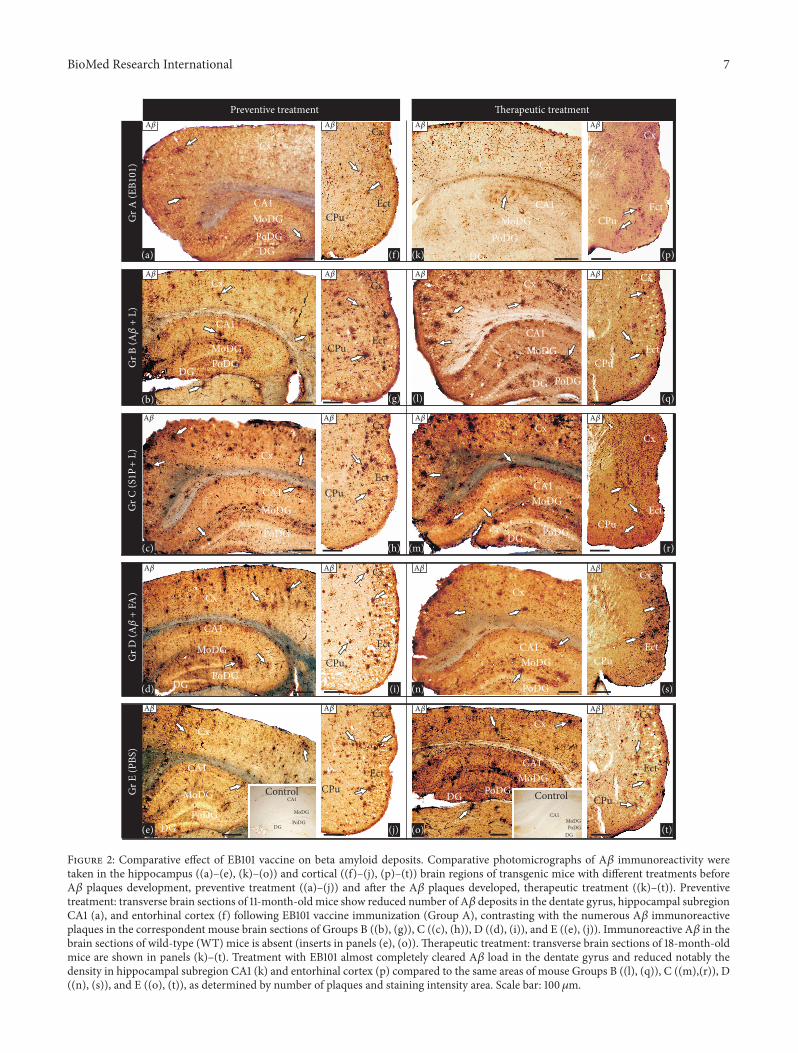
BioMed Research International 7
Ect
Cx
CPu
Ect
Gr A
(EB1
01)
(a)
(c)
(b)
(f)
(h)
(g)
Preventive treatment Therapeutic treatment
CA1
PoDGMoDG
CA1
PoDGMoDG
CA1
PoDG
MoDG
DG
DG
Ect
Ect
Cx
Cx
Cx
CPu
CPu
CPuCA1
PoDG
MoDG
DG
CA1
PoDG
MoDG
DG
Ect
Cx
CPu
Ect
Cx
CPu
Cx
Cx
Cx
Cx
Cx
(k)
(l)
(p)
(r)
(q)
CA1
PoDGMoDG
DG
Cx
Ect
Cx
CPuCA1
ControlGr E
(PBS
)
(d)
(e) (j)
Control
PoDG
MoDG
DG
CA1
CA1
PoDG
MoDG
CA1
PoDG
MoDG
DG
DG
Ect
Ect
Cx
Cx
CPu
CPu
CA1
PoDG
MoDG
CA1
PoDGMoDG
DG
Ect
Cx
CPu
PoDGMoDG
DG
Cx
Cx
Cx
(n)
(o)
(s)
(t)
Cx
(i)
Gr B
(A120573
+L)
Gr D
(A120573+
FA)
A120573 A120573 A120573 A120573
A120573A120573A120573
A120573 A120573 A120573 A120573
A120573A120573A120573A120573
A120573 A120573 A120573 A120573
A120573
(m)
Gr C
(S1P
+L)
Figure 2 Comparative effect of EB101 vaccine on beta amyloid deposits Comparative photomicrographs of A120573 immunoreactivity weretaken in the hippocampus ((a)ndash(e) (k)ndash(o)) and cortical ((f)ndash(j) (p)ndash(t)) brain regions of transgenic mice with different treatments beforeA120573 plaques development preventive treatment ((a)ndash(j)) and after the A120573 plaques developed therapeutic treatment ((k)ndash(t)) Preventivetreatment transverse brain sections of 11-month-old mice show reduced number of A120573 deposits in the dentate gyrus hippocampal subregionCA1 (a) and entorhinal cortex (f) following EB101 vaccine immunization (Group A) contrasting with the numerous A120573 immunoreactiveplaques in the correspondent mouse brain sections of Groups B ((b) (g)) C ((c) (h)) D ((d) (i)) and E ((e) (j)) Immunoreactive A120573 in thebrain sections of wild-type (WT) mice is absent (inserts in panels (e) (o)) Therapeutic treatment transverse brain sections of 18-month-oldmice are shown in panels (k)ndash(t) Treatment with EB101 almost completely cleared A120573 load in the dentate gyrus and reduced notably thedensity in hippocampal subregion CA1 (k) and entorhinal cortex (p) compared to the same areas of mouse Groups B ((l) (q)) C ((m)(r)) D((n) (s)) and E ((o) (t)) as determined by number of plaques and staining intensity area Scale bar 100 120583m
8 BioMed Research International
Gr E
(PBS
)
0102030405060708090
Preventive treatmentTherapeutic treatment
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Mea
n of
120573-a
myl
oid
plaq
ues
sect
ion
Gr A
(A120573+
S1P+
L)(E
B101
)
Figure 3 Comparative analysis of A120573 plaque burden in the brainsof APPPS1 mice Mean of A120573 plaque burden in the hippocampaland cortical regions of APPswePS1ΔE9 mice in the five treatmentgroups The mean of the A120573 burden is significantly reduced (119875 lt005) in Group A (EB101) when compared with Groups BndashE duringpreventive treatment and markedly reduced in the therapeutictreatment period Data are presented asmeanplusmn standard error of themean (SEM) Densitymean of A120573 plaques in Tgmice hippocampus
05
1015202530354045
Preventive treatmentTherapeutic treatment
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Gr A
(A120573+
S1P+
L)(E
B101
)
Mea
n of
120573-a
myl
oid
plaq
ues a
rea
Figure 4 Comparative quantification of A120573 burden area in thebrain of APPPS1 mice Quantitative analysis of A120573 burden areain the hippocampal and cortical regions of APPswePS1ΔE9 micetreated with EB101 A120573liposomes S1Pliposomes A120573
42+ FAIFA
and PBS represented by the number of pixels inside the stained areaof each A120573 plaque This graphic shows that A120573 plaques of EB101-treated mice (Group A) are significantly smaller in size (119875 lt 005)than those of the other four treatment groups Data are presented asmean plusmn SEM Mean of A120573 plaques area in Tg mice hippocampus
a reduced staining intensity of the A120573 plaques in the brainregions of mouse Group A contrasting with the intenseA120573-immunoreactivity of plaques in the other experimentalmouse groups as shown in the photomicrographs of Figures2(k)ndash2(t)
33 Immune Response Effects of EB101 versus A12057342+ CFAIFA
Vaccine Astrocyte activation implicated in neuroinflam-mation amyloidogenesis and neuronal cell death in AD
was compared after treatment of the transgenic mice withEB101 and A120573
42+ CFAIFA Immunohistochemical analysis
of GFAP-reactive cells was carried out in the brains ofthe treated mice in the preventive and therapeutic studies(Figure 5) After the preventive treatment (Figures 5(a)ndash5(j))EB101 significantly reduced the density of GFAP-reactive cellclusters in the hippocampal and cortical sections as comparedwith A120573
42+ CFAIFA (D) A120573Lip without S1P (B) Lip with
S1P (C) and PBS (E) The distribution of GFAP immunore-active cells in the transverse section of the mouse brainstreated with EB101 shows a few scattered GFAP-reactiveclusters mainly at the CA1 hippocampal layers contrastingwith numerous dystrophic reactive astrocytes observed indifferent hippocampal areas of mouse brains in Groups BndashE(Figures 5(b)ndash5(e)) Similar neuroinflammation pattern wasobserved in the retrosplenial cortical layers where a fewGFAP-reactive clusters were observed in EB101 treated mice(Figure 5(f)) compared with their profuse presence in theother experimental groups (Figures 5(g)ndash5(j)) After thera-peutic treatment the GFAP immunoreactivity distributionpattern in mouse brains treated with EB101 was similar tocontrol mice (wild-type) mostly devoid of reactive GFAPclusters except a few scattered ones in the CA1 hippocampalexternal layers (Figure 5(k)) In contrast there is a moderatedensity in Group D (Figure 5(n)) and extensive density inGroup B (Figure 5(l)) Group C (Figure 5(m)) and Group Emice (Figure 5(o)) Photomicrographs of retrosplenial cortexshow similar neuroinflammation pattern where a few GFAPclusters detected in the EB101 treated mice (Figure 5(p))contrast with a more extensive density of clusters in the othergroups (Figures 5(q)ndash5(t)) No astrocytosis was observed inwild-type mice during preventive or therapeutic treatment
34 Comparative A120573 Antibody Titers and Th1Th2 CytokineLevels in Treated Mice A120573 antibody titers in the sera weredetermined by ELISA during mouse immunization(Figure 6) Preventive treatment with EB101 resulted in amarked increase of specific IgG A120573
1ndash42 antibody productionin all animals Mice in A and D groups developed andmaintained serum antibody titers between 1 2000 and1 8000 Mice in Groups B C and E showed no significantanti-A120573 antibody production Antibody production and titerin Group B were three and four times lower than those ofGroups D and A respectively After therapeutic treatment allmice treated with EB101 and A120573
42+ FAIFA produced high
levels of IgG antibodies Titer levels of Groups A and D weresignificantly increased to those detected after preventivetreatment (119875 lt 001 versus Group D 119875 lt 0001 versusGroups B C and E) although antibody level in Group D was13-fold lower than in Group A (Figure 6)The effect of EB101(Group A) and A120573
42+ CFAIFA (Group B) vaccines on Th1
andTh2 cytokines was also studied (Figure 7)Measurementsof Th1 and Th2 cytokines in sera of immunized transgenicmice indicate that significant changes occurring in proteinlevels of Th1-related inflammatory mediators followingpreventive (Figures 7(a)ndash7(c)) and therapeutic (Figures 7(d)ndash7(g)) treatment with EB101 vaccine A significant increaseof Th1 cytokine profile was observed in preventive treatment
BioMed Research International 9
Gr E
(PBS
)
(d)
(e)
(i)
(j)
(n)
(o)
(s)
(t)
Cx
GFAP
GFAP GFAP GFAP GFAP
GFAP GFAP GFAP
Cx Cx
Cx
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
GFAP
Gr A
(EB1
01)
(a)
(b)
(c)
(f)
(g)
(h)
Preventive treatment Therapeutic treatment
PoDG
MoDG
(k)
(l)
(m)
(p)
(q)
(r)DG
GFAP GFAP GFAP
GFAPGFAPGFAP GFAP
GFAP GFAP GFAP GFAP
PoDGDG
DG
CA1
Cx
Cx
Cx
Cx
Cx
Cx
PoDG
MoDG
DG
CA1
MoDGDG
CA1
PoDG
MoDG
DG
CA1
Cx
CxCx
MoDG
MoDG
PoDG
CA1CA1
Cx
Gr D
(A120573+
FA)
Gr C
(S1P
+L)
Gr B
(A120573+
L)
Figure 5 Comparative effect of EB101 vaccine on astrocytosis Comparative photomicrographs of GFAP immunoreactivity in thehippocampus ((a)ndash(e) (k)ndash(o)) and cortical ((f)ndash(j) (p)ndash(t)) brain regions at the preventive treatment phase ((a)ndash(j)) and therapeutictreatment group ((k)ndash(t)) Preventive treatment transverse brain sections of 11-month-old mice treated with EB101 show an almost completeprevention of astrocytosis in the detailed sections of the dentate gyrus (a) contrasting with numerous dystrophic reactive astrocytes typicalof an inflammatory reaction observed in different hippocampal corresponding areas of mouse brains in Group B (A120573Lip-treated mice (b))Group C (LipS1P-treated mice (c)) Group D (A120573
42+ CFAIFA-treated mice (d)) and Group E (PBS-treated mice (e)) Immunoreactive
astrocytosis in the brain sections of WT mice is absent (data not shown) Detailed sections of the parietal cortex of treated mice show agradient density of reactive astrocytes immunoreactive to GFAP indicating a neuroinflammation pattern Comparative cortical sections shownumerous astrocytosis areas inmice Groups C and E ((h) (j)) andwide reactive astrocytes clusters inmice Groups B andD ((g) (i)) while justa few of them are observed in Group A (f)Therapeutic treatment transverse brain sections of 18-month-old mice of Group A show a notablereduction of astrocytosis in the hippocampal region (k) after EB101 vaccine immunization Note the astrocytosis contrast density betweenEB101-treatedmice and the other treated groups ((l)ndash(o)) where numerous immunoreactive GFAP clusters are observed in the correspondentbrain sections A few immunoreactive GFAP clusters are observed in the parietal cortex of EB101 treated mice group while a moderate ((q)(s)) to high ((r) (t)) density of these neuropathological inflammation clusters is apparent in the other treatment groups (Groups BndashE) Forabbreviations see list Scale bar 100120583m
10 BioMed Research International
002040608
1121416
Preventive treatmentTherapeutic treatment
Ant
i-120573-a
myl
oid
IgG
antib
odie
s
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Gr A
(A120573+
S1P+
L)(E
B101
)
Figure 6 Quantification of specific anti-120573-amyloid IgG in immu-nized APPPS1 mice The presence of anti-120573-amyloid antibodiesmeasured in the sera of vaccinated and control mice groups duringthe preventive and therapeutic treatments indicates that EB101liposome vaccine was able to induce the highest production ofspecific IgG antibodies Compared to theGroupD immunizedmicethe level of antibodies observed in the Group A was also highersuggesting a more effective immune response (119875 lt 001 versusGroup D 119875 lt 0001 versus Groups B C and E) Quantification ofspecific anti-120573-amyloid IgG in immunized Tg mice
in Group D with respect to all other groups mainlyreported by the high levels of TNF-120572 (Figure 7(b)) andIFN-120574 (Figure 7(c)) cytokine secretions while due to thehigh mortality rate during the therapeutic treatment nosignificant data was obtained from this mice group Inaddition a tendency toTh1 secretion reduction was observedinGroupAwith respect to all other groups in both preventiveand therapeutic treatment The changes in Th2 protein levels(Figure 7(d)) followed the inverse trend where the highestdifferences between Group A and the other treatment groupswere observed in the IL-4 IL-5 and IL-10 cytokine secretions(119875 lt 005 versus Group D 119875 lt 001 versus Group E Figures7(e)ndash7(g))
35 Comparative Motor Coordination and Memory Perfor-mances In order to validate the locomotion integrity duringthe entire immunotherapeutic process as reported in similarprevious studies motor coordination tests have showed thatEB101-treated mice spent significantly more time on theaccelerating rotarodThismotor coordination task shows thatEB101 immunization affected positively their performance(Figure 8(a)) A120573
42liposome (B) S1P-containing liposomes
(C) and PBS (E) (119875 lt 0006) mice performed poorly incomparison to EB101 mice (119875 lt 005) or A120573
42+ CFAIFA
mice (119875 lt 002) in motor abilities and coordination Thefear conditioning-active avoidance test was used to assess thespatial learning and memory functions As expected bothEB101 and A120573
42+ CFAIFA immunized mice readily learned
the location of the platform as shown by a decrease in theescape latency over training days while the control group
mice exhibited no significant changes during 6-day trialsThe average escape latencies of EB101 (68 plusmn 2 s 761 plusmn 22 s)andA120573
42+CFAIFA (116plusmn33 s 123plusmn21 s) immunizedmice
at the end of preventive and therapeutic periods respectivelyduring the consecutive 6 days trial were significantly shorterthan the other treatment groups (A120573
42liposome 163 plusmn 31 s
175 plusmn 28 s S1P-containing liposomes 202 plusmn 19 s 248 plusmn27 s) and the control (2116 plusmn 38 s 268 plusmn 22 s) (119875 lt 001)Therewas no significant difference between EB101 andA120573
42+
CFAIFA immunized group (119875 gt 005) (Figure 8(b)) Theseresults showed that spatial learning was improved in theseimmunized mice
4 Discussion
In this study we have compared a novel active A120573 imm-unotherapy the EB101 vaccine against A120573 with the A120573
42+
CFAIFA vaccine that was first taken into clinical trialswith coadjuvant Qs-21 modification (AN1792 by Orgogozoet al [19]) This type of vaccine design was thought toinduce a strong T-cell mediated inflammatory response (IL-2 and IFN-c positive responses indicative of a Class II(CD4) Th-1 type response [33] leading to acute menin-goencephalitis in the 6 of the patients) The EB101 activeimmunotherapeutic vaccine was primarily designed to avoidundesired inflammatory side effects both in mice andhumans (without need of coadjuvant modification) whileinducing effective antibody titer rates We demonstrated thatlong-term immunization with EB101 induced A120573 antibodiesin the preventive and the therapeutic treatment periodswhich effectively reduced amyloid deposition in both corticaland hippocampal brain regions and prevented neuroinflam-mation probably by blocking toxicity in the surroundingneurons However the AN1792 vaccine was shown to inducein 6 of the patients an inflammatory reaction that mighthave led to the encephalitis type reaction observed in theclinical trial which was responsible for the immediate haltin those clinical studies It seems that AN1792 immunizationdelivered in Qs-21 and polysorbate 80 adjuvant induced aTh1 type response that likely led to autoaggressive T cells andaseptic meningoencephalitis In the present experiments wedemonstrated that a judiciously selected adjuvant produces adifferent humoral response [22] Indeed we have shown thatour novel immunotherapeutic strategy boosts specific A120573
1ndash42antibody generation and induces a Th2 noninflammatoryimmune response This boosted effect is in accordance withprevious immunotherapeutic studies [34] Taking togetherall the present findings these results suggest that EB101vaccine can achieve in a future clinical trial immunogenicresponse rates quite similar to the AN1792 without theinflammatory effects reported The immunohistochemicalanalysis obtained in the present study shows that all the EB101immunized mice resulted in an almost complete preventionof A120573 deposition in the brain with only a few compactA120573 plaques in the retrosplenial and dentate gyrus regionsAlthough there was a slightly increase in death in EB101treated mice compared to controls this significant reductionof A120573 plaques in EB101 treated mice was proven to be more
BioMed Research International 11
Gr A Gr B Gr C Gr D Gr EPreventive treatmentTherapeutic treatment
0005001000150020002500300035004000
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
(a) Th1
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
000
1000
2000
3000
4000
5000TNF-120572
(b) Th1
000500
10001500200025003000
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
IFN-120574
(c) Th1
000500
100015002000250030003500
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
Group A Group B Group C Group D Group EPreventive treatmentTherapeutic treatment
(d) Th2
000
500
1000
1500
2000
2500
3000IL-4
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(e) Th2
000500
100015002000250030003500
IL-5
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(f) Th2
3500IL-10
000500
10001500200025003000
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(g) Th2
Figure 7 Detection of Th1 and Th2 cytokine profile Measurements of the effect of EB101 and A12057342
+ CFAIFA vaccines on Th1 and Th2cytokines profile during preventive and therapeutic treatments ((a)ndash(c)) A significant increase of Th1 cytokine profile was observed inpreventive treatment in Group D with respect to all other groups while a tendency to a reduction inTh1 secretion was observed in Group Awith respect to all other groups in both preventive and therapeutic treatment ((d)ndash(g)) The changes in Th2 protein levels were observed inthe IL-4 IL-5 and IL-10 cytokine secretions of Group A (119875 lt 005 versus Group D 119875 lt 001 versus Group E)
12 BioMed Research International
Gr A
(EB1
01)
05
101520253035
Preventive treatment Therapeutic treatment
Mea
n fa
lling
late
ncy
(s)
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)(a) Graphic of motor coordination and balance
05
1015202530
Mea
n re
spon
se ti
me (
s)
Gr A
(EB1
01)
Preventive treatment Therapeutic treatment
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
(b) Graphic of associative learning and memory
Figure 8 Analysis of motor coordination and balance (a) Rotarod results of motor coordination and balance test after the preventive andtherapeutic treatment showing improved effect of EB101 and A120573
42+ CFAIFA-vaccine compared to other treated mice groups (119875 lt 005)
and (119875 lt 002) versus control mice Latency to fall on the rotarod apparatus was significantly improved in EB101 immunized mice Valuesrepresent means of all six trials of a groupday (b) Spatial learning and memory tested by fear conditioning-active avoidance test The meanresponse time of each treatment group to reach the platform is represented EB101 and A120573
42+ CFAIFA immunized mice had significantly
lower latencies than the other treatment groups (119875 lt 001) versus control mice
efficient than that obtained with A12057342+ CFAIFA immuniza-
tions A comparison of A120573 plaque reduction rates amongtreated groups surprisingly showed that other than EB101and A120573
42+ CFAIFA treated groups some amyloid deposits
were also reduced in Groups B (A1205731ndash42liposome) and C
(S1P-containing liposomes) Although the reduction rates inthese mice were markedly low the subtle lowering effectmight be due to the interaction between the immunizationelements (A120573
1ndash42 and S1P) and the mouse immunogenicsystem as reported by others [35] Based on our presentresults S1P one of the components of the adjuvant used withEB101 might be responsible for the neuroprotective effectobserved in the vulnerable neuronal areas of EB101 treatedmice The advantage in the A120573 load preventionreduction bycombining A120573
1ndash42 and S1P in a unique formulation vaccinehas been also observed in the reduced hippocampal load ofEB101 immunizedmice (Gr A) when comparedwith differentvaccine components (Gr B-C) In fact S1P appears to have ademonstrated therapeutic potential as a regenerative agent inthe nervous system since it controls migration of neuronalstem cells toward a site of spinal cord injury [36] inducecytoskeletal rearrangements to promote transmitter secretion[37] and also trigger diverse cellular effects including angio-genesis cardiac development immunity cell motility andneurite extension [38 39]Therefore it is reasonable to expectthat such a potent biologically active sphingolipid by itselfmay play an important role in the A120573 plaque reduction Inthis study the clearance efficiency rate of each treatment wasanalyzed and results suggested that EB101 (6459) appearsto be more effective than A120573
42+ CFAIFA (1920) vaccine
in both preventive and therapeutic groups The effectivenessof EB101 in older transgenic animals (18 months) showeda clearance effect of 59 suggesting that EB101 may alsohave therapeutic potential inADdiagnosticated patient trials
Similar conclusion was reached byWang and colleagues [40]who demonstrated that combination therapy in aged animalsprovides more valuable insights into a possible translationto humansOur immunologicalmeasurements demonstratedonce again that neuroinflammation develops in an A120573-dependent fashion early in the course of AD [41] and canbe alleviated or reversed by interventional A120573 vaccinationsuch as EB101 Among the possible mechanisms of A120573clearance proposed in AD immunotherapy [42] our resultssuggested that theymay overlap and probably be disease statedependent since the prevention and the therapeutic vaccinewould likely benefit from the transport of A120573 antibodies intothe CNS However some mechanisms of action such as thepreventive effect of A120573 antibodies in the A120573 aggregation [43]microglial phagocytosis induced by the Fc binding portionsbetween A120573 antibodies and microglia receptor [44] and theshift in the gradient of A120573 transport across the blood-brainbarrier due to the increased concentration of A120573 antibodiesin the vascular system resulting in an increase in effluxfrom the brain to blood [45 46] may be playing a role inboth preventive and therapeutic conditions Our findingstherefore suggest that novel treatment strategies aimed atmaintaining or increasing microglial function and restoringthe balance to the immunological system may represent anattractive therapeutic approach even at the advanced stagesof ADThe immunization results obtained with EB101 duringboth preventive and therapeutic treatment showed nomicro-hemorrhages infiltration of lymphocytes or encephalitis-like reaction in the brain suggesting that EB101 does notinduce A120573-specific Th1 reaction possibly due to the neu-roprotective effect of S1P Delivering A120573
1ndash42 in a liposomal-S1P-adjuvant resulted in an effective immunological responseincluding the induction of a Th2 T-cell response whicheffectively prevented neuroinflammation in an animal model
BioMed Research International 13
of AD as reported previously [24 47] Moreover presentresults demonstrated that EB101 did not induce astrogliosisin APPPS1 transgenic mice whereas some GFAP-positiveastrocyte clusters were aggregated in the affected brain areasof mice treated with A120573
42+ CFAIFA These results indicate
that EB101 may result in an effective treatment in AD neu-ropathology due to the absence of peripheral proinflamma-tory cytokine secretion and the absence of astroglial activa-tion in the brains of EB101 treated mice when compared withA12057342
+ CFAIFA immunotherapeutic response In a clinicalsetting however we may expect a wide differential responsein part depending upon the genomic profile of each patient[48 49] In AD behavioural studies in particular amongthe motor coordination tests the rotarod performance hasbeen reported to be impaired in some AD transgenic micemodels bearing A120573 plaques [50 51] This result was found inmutants prior to A120573 plaque formation at 3 and 6 months ofage but was not reproduced after plaques had been formedat 16 and 24 months of age [52] perhaps because of thedeclining abilities of older normal mice Here we presentthe effects of vaccine treatments in APPPS1 mice on motorcoordination a neglected aspect of the AD experimentalbehavior but justified on the basis of the deficient posturalcontrol reported in patients with Alzheimerrsquos disease [53]sometimes even during early stages [54] Our longitudinalmotor behavioral results showed no significant differencesin the ambulatory movement grip strength or coordinationaddressed by their performance in the rotating rod betweenAPPPS1 and wild-type mice for any age group studiedin contrast with other treated groups demonstrating thatlongitudinal motor assessments of AD transgenic mousemodels can provide valuable information with regard tothe effect of immunization on the progression of behavioraldeficits and overall motor activity As shown the EB101-treated group of mice exhibited longer latencies and betterbalancing skills indicating the absence of any negative effectof EB101 immunization on these mice The classical fearconditioning test is assumed to reveal the functional integrityof the hippocampus [55] representing a special memorytask to evaluate the AD models and its improvement afterimmunization periods In this APPPS1 mouse model thefirst associative learning and spatial workingmemory deficitsare shown at 5-6 months of age [56] although its impair-ment progression is relatively slow compared to most othermodels At the end of preventive immunization period EB101and A120573
42+ CFAIFA immunized mice have significantly
shorter escape latencies than the other treatment groups(A12057342liposome S1P-containing liposomes) and the controls
in the fear conditioning test Shorter escape latencies duringspatial learning indicated that deficits of spatial memorywere attenuated in EB101 and A120573
42+ CFAIFA immunized
mice Previous studies have showed that by 15 months ofage the spatial working memory is consistently impairedthroughout the rest of the life span [31 57] At the end oftherapeutic immunization period both EB101 and A120573
42+
CFAIFA immunized mice showed significantly improvedresults on the hippocampus-dependent spatial learning andmemory test when compared with other treatment groupsand the controls Taken together these observations are
in agreement with the impairment in conditioned learningin response to a tone stimulus of aged APPPS1 micealthough when immunized with EB101 during preventiveor therapeutic periods such impairments are significantlyattenuated
In summary the present results corroborate our previouspublished findings showing that EB101 immunization reducesamyloid accumulation in the APPPS1 mouse brain [2425] The strongest reduction in the preventive immunizationwas observed in the retrosplenial cortex followed by thehippocampus and entorhinal-piriform cortex Our resultsfurther support that EB101 ismore effectivewhen themice areimmunized prior to developing plaques in the hippocampaland cortical brain regions There is extensive evidence inmouse preclinical studies [58] and human clinical trials [5960] that the adverse effects of active and passive A120573 vaccinesresult in microhemorrhages and encephalitic reactions [60ndash63] yet themechanismbywhich they occur has not been fullyelucidated In the treatment sections analyzed we found nomicrohemorrhages or acute inflammatory processes in all thesurvived mice of the group indicating that EB101 therapeuticapproach presents no adverse inflammatory effects in thisAD mice models Thus our present results unequivocallydemonstrate that EB101 vaccinemay not only halt AD-relatedpathology as a preventive treatment but may also reverse it inestablished cases related to the improvement of the learningandmemory in both cases leading to a robust immunologicaltherapeutic response Present experiments also demonstratethat EB101 exerts more significant and substantial effects onthe reduction of A120573 deposition than A120573
42+CFAIFA vaccine
in A120573 mice models of AD However further exploration ofthe contribution of genetic environmental and physiologicalfactors in the development resistance to endogenous A120573-antibody is needed in order to optimize and modulate thetherapeutic strategies currently being developed
5 Conclusions
The present study carried out in APPPS1 transgenic mousemodel of AD compared the A120573 burden reduction andcellular immune responses produced by two immunizationdelivery strategies of A120573 in Freund adjuvant (described byWebster et al [10]) and in liposomeS1P matrix (describedby Schenk et al [17])The EB101 immunotherapeutic strategyboosted antibody generation A120573 plaque reduction and ananti-inflammatory response more effectively than A120573
42+
CFAIFA especially if given prior to or in the early stages ofpathology The present results also showed that vaccinationwith EB101 prevented the formation of new plaques andpresumably led to a reduction in established plaques whileinducing aTh2 immune reactionwithout encephalitogenic T-cell responses or brain inflammation related with astrogliosisHowever despite the efficacy demonstrated of this novelvaccine approach further preclinical and clinical studieson EB101 are warranted to evaluate its efficacy and themechanism for the future utility of such vaccine for ADtherapy
14 BioMed Research International
List of Abbreviations
AD Alzheimerrsquos diseaseA120573 120573-Amyloid proteinAPP Amyloid precursor proteinAPT Anterior pretectal nucleusCA1 Field CA1 hippocampusCA3 Field CA3 hippocampusCPu Caudate putamen (striatum)Cx CortexDG Dentate gyrusDN Dystrophic neuriteEct Entorhinal cortexCFA Complete Freundrsquos adjuvantGrDG Granular dentate gyrusIFA Incomplete Freundrsquos adjuvantIL InterleukinLPMR Lateral posterior thalamic nucleus (mediorostral)MoDG Molecular dentate gyrusPir Piriform cortexPoDG Polymorph dentate gyrusRSGb Retrosplenial granular cortex b regionTg TransgenicTh T helper
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Ivan Carrera Ignacio Etcheverrıa Lucıa Fernandez-NovoaValter Lombardi Ramon Cacabelos and Carmen Vigo con-ceived and designed the experiments Ivan Carrera and Igna-cio Etcheverrıa performed the experiments Ivan CarreraIgnacio Etcheverrıa and Valter Lombardi analyzed the dataIvan Carrera Ignacio Etcheverrıa Lucıa Fernandez-Novoaand Valter Lombardi contributed reagentsmaterialsanalysistools Ivan Carrera Carmen Vigo andMadepalli KrishnappaLakshmana wrote the paper
Acknowledgments
This work was supported by National Institute of Aging(NIA)NIH Grant no 1R01AG036859-01 (Madepalli Krish-nappa Lakshmana) and the financial support of the ProgramaOperativo Feder Galicia 2007ndash2013 (IN841C 201254 to PhDCarrera I)
References
[1] R Cacabelos L Fernandez-Novoa V Lombardi Y Kubotaand M Takeda ldquoMolecular genetics of Alzheimerrsquos diseaseand agingrdquoMethods and Findings in Experimental and ClinicalPharmacology vol 27 pp 1ndash573 2005
[2] P T Nelson I Alafuzoff E H Bigio et al ldquoCorrelationof alzheimer disease neuropathologic changes with cognitive
status a review of the literaturerdquo Journal of Neuropathology andExperimental Neurology vol 71 no 5 pp 362ndash381 2012
[3] JNaslundVHaroutunian RMohs et al ldquoCorrelation betweenelevated levels of amyloid 120573-peptide in the brain and cognitivedeclinerdquo Journal of the American Medical Association vol 283no 12 pp 1571ndash1577 2000
[4] A Koyama O I Okereke T Yang D Blacker D J Selkoeand F Grodstein ldquoPlasma amyloid-120573 as a predictor of dementiaand cognitive decline a systematic review and meta-analysisrdquoArchives of Neurology vol 69 no 7 pp 824ndash831 2012
[5] A M Fagan C M Roe C Xiong M A Mintun J C Morrisand D M Holtzman ldquoCerebrospinal fluid tau120573-amyloid42ratio as a prediction of cognitive decline in nondemented olderadultsrdquo Archives of Neurology vol 64 no 3 pp 343ndash349 2007
[6] S Parvathy P Davies V Haroutunian et al ldquoCorrelationbetween A120573-40- A120573-42- and A120573x-43-containing amyloidplaques and cognitive declinerdquo Archives of Neurology vol 58no 12 pp 2025ndash2032 2001
[7] E Karran M Mercken and B D Strooper ldquoThe amyloidcascade hypothesis for Alzheimerrsquos disease an appraisal for thedevelopment of therapeuticsrdquo Nature Reviews Drug Discoveryvol 10 no 9 pp 698ndash712 2011
[8] J L Jankowsky D J Fadale J Anderson et al ldquoMutantpresenilins specifically elevate the levels of the 42 residue 120573-amyloid peptide in vivo evidence for augmentation of a 42-specific 120574 secretaserdquo Human Molecular Genetics vol 13 no 2pp 159ndash170 2004
[9] L Holcomb M N Gordon E Mcgowan et al ldquoAcceleratedAlzheimer-type phenotype in transgenic mice carrying bothmutant amyloid precursor protein and presenilin 1 transgenesrdquoNature Medicine vol 4 no 1 pp 97ndash100 1998
[10] S JWebster A D Bachstetter and L J van Eldik ldquoComprehen-sive behavioral characterization of an APPPS-1 double knock-in mouse model of Alzheimerrsquos diseaserdquo Alzheimerrsquos ResearchandTherapy vol 5 no 3 article 28 2013
[11] H Xiong D Callaghan J Wodzinska et al ldquoBiochemical andbehavioral characterization of the double transgenic mousemodel (APPswePS1dE9) of Alzheimerrsquos diseaserdquo NeuroscienceBulletin vol 27 no 4 pp 221ndash232 2011
[12] M Garcia-Alloza L A Borrelli B T Hyman and B J BacskaildquoAntioxidants have a rapid and long-lasting effect on neuriticabnormalities in APPPS1 micerdquo Neurobiology of Aging vol 31no 12 pp 2058ndash2068 2010
[13] M Meyer-Luehmann M Mielke T L Spires-Jones et al ldquoAreporter of local dendritic translocation shows plaque-relatedloss of neural system function in APP-transgenic micerdquo Journalof Neuroscience vol 29 no 40 pp 12636ndash12640 2009
[14] E Machova V Rudajev H Smyckova H Koivisto H Tanilaand V Dolezal ldquoFunctional cholinergic damage develops withamyloid accumulation in young adult APPswePS1dE9 trans-genic micerdquo Neurobiology of Disease vol 38 no 1 pp 27ndash352010
[15] T Malm J Koistinaho and K Kanninen ldquoUtilization of APP-swePS1dE9 transgenic mice in research of Alzheimerrsquos diseasefocus on gene therapy and cell-based therapy applicationsrdquoInternational Journal of Alzheimerrsquos Disease vol 2011 Article ID517160 8 pages 2011
[16] J Lee and P Han ldquoAn update of animal models of alzheimerdisease with a reevaluation of plaque depositionsrdquoExperimentalNeurobiology vol 22 pp 84ndash95 2013
BioMed Research International 15
[17] D Schenk R Barbour W Dunn et al ldquoImmunization withamyloid-120573 attenuates Alzheimer disease-like pathology in thePDAPP mouserdquo Nature vol 400 no 6740 pp 173ndash177 1999
[18] H J Fu B Liu J L Frost and C A Lemere ldquoAmyloid-120573immunotherapy for Alzheimerrsquos diseaserdquoCNS and NeurologicalDisorders-Drug Targets vol 9 no 2 pp 197ndash206 2010
[19] J-M Orgogozo S Gilman J-F Dartigues et al ldquoSubacutemeningoencephalitis in a subset of patients with AD after A120573
42
immunizationrdquo Neurology vol 61 no 1 pp 46ndash54 2003[20] J A R Nicolll DWilkinson CHolmes P Steart HMarkham
andROWeller ldquoNeuropathology of humanAlzheimer diseaseafter immunization with amyloid-120573 peptide a case reportrdquoNature Medicine vol 9 no 4 pp 448ndash452 2003
[21] I Ferrer M Boada Rovira M L Sanchez Guerra M JRey and F Costa-Jussa ldquoNeuropathology and pathogenesisof encephalitis following amyloid-Neuropathology and patho-genesis of encephalitis following amyloid-beta immunizationin Alzheimerrsquos disease immunization in Alzheimerrsquos diseaserdquoBrain Pathology vol 14 no 1 pp 11ndash20 2004
[22] D H Cribbs A Ghochikyan V Vasilevko et al ldquoAdjuvant-dependent modulation ofTh1 andTh2 responses to immuniza-tion with 120573-amyloidrdquo International Immunology vol 15 no 4pp 505ndash514 2003
[23] C Holmes D Boche D Wilkinson et al ldquoLong-term effectsof A120573
42immunisation in Alzheimerrsquos disease follow-up of a
randomised placebo-controlled phase I trialrdquo The Lancet vol372 no 9634 pp 216ndash223 2008
[24] I Carrera I Etcheverrıa L Fernandez-Novoa V LombardiR Cacabelos and C Vigo ldquoVaccine development to treatAlzheimerrsquos disease neuropathology in APPPS1 transgenicmicerdquo International Journal of Alzheimerrsquos Disease vol 2012Article ID 376138 17 pages 2012
[25] I Carrera I Etcheverrıa Y Li et al ldquoImmunocytochemicalcharacterization of Alzheimer disease hallmarks in APPPS1transgenic mice treated with a new anti-amyloid-120573 vaccinerdquoBioMed Research International vol 2013 Article ID 709145 12pages 2013
[26] V Anelli R Bassi G Tettamanti P Viani and L Riboni ldquoExtra-cellular release of newly synthesized sphingosine-1-phosphateby cerebellar granule cells and astrocytesrdquo Journal of Neuro-chemistry vol 92 no 5 pp 1204ndash1215 2005
[27] S Schroeter K Khan R Barbour et al ldquoImmunotherapyreduces vascular amyloid-120573 in PDAPP micerdquo Journal of Neu-roscience vol 28 no 27 pp 6787ndash6793 2008
[28] Z Suo A Cox N Bartelli et al ldquoGRK5 deficiency leads to earlyAlzheimer-like pathology and working memory impairmentrdquoNeurobiology of Aging vol 28 no 12 pp 1873ndash1888 2007
[29] W Kamphuis C Mamber MMoeton et al ldquoGFAP isoforms inadult mouse brain with a focus on neurogenic astrocytes andreactive astrogliosis in mouse models of Alzheimer diseaserdquoPLoS ONE vol 7 no 8 Article ID e42823 2012
[30] J Lang C Vigo-Pelfrey and F Martin ldquoLiposomes composedof partially hydrogenated egg phosphatidylcholines fatty acidcomposition thermal phase behavior and oxidative stabilityrdquoChemistry and Physics of Lipids vol 53 no 1 pp 91ndash101 1990
[31] D Morgan D M Diamond P E Gottschall et al ldquoA 120573peptide vaccination prevents memory loss in an animal modelof Alzheimerrsquos diseaserdquo Nature vol 408 no 6815 pp 982ndash9852000
[32] M L Brownlow L Benner D DrsquoAgostino M N Gordon andD Morgan ldquoKetogenic diet improves motor performance but
not cognition in two mouse models of Alzheimerrsquos pathologyrdquoPLoS ONE vol 8 no 9 Article ID e75713 2013
[33] M Pride P Seubert M Grundman M Hagen J Eldridgeand R S Black ldquoProgress in the active immunotherapeu-tic approach to Alzheimerrsquos disease clinical investigationsinto AN1792-associated meningoencephalitisrdquo Neurodegenera-tive Diseases vol 5 no 3-4 pp 194ndash196 2008
[34] CWiessner K-HWiederhold A C Tissot et al ldquoThe second-generation activeA120573 immunotherapyCAD106 reduces amyloidaccumulation in APP transgenic mice while minimizing poten-tial side effectsrdquoThe Journal of Neuroscience vol 31 no 25 pp9323ndash9331 2011
[35] D Lambracht-Washington and R Rosenberg ldquoAdvances in thedevelopment of vaccines for Alzheimerrsquos diseaserdquo DiscoveryMedicine vol 15 no 84 pp 319ndash326 2013
[36] A Kimura T Ohmori R Ohkawa et al ldquoEssential roles ofsphingosine 1-phosphateS1P
1receptor axis in the migration of
neural stem cells toward a site of spinal cord injuryrdquo Stem Cellsvol 25 no 1 pp 115ndash124 2007
[37] S S Lee C H Woo J D Chang and J H Kim ldquoRoles of Racand cytosolic phospholipase A2 in the intracellular signalling inresponse to titanium particlesrdquo Cellular Signalling vol 15 no 3pp 339ndash345 2003
[38] E Masliah L Hansen A Adame et al ldquoA120573 vaccination effectson plaque pathology in the absence of encephalitis inAlzheimerdiseaserdquo Neurology vol 64 no 1 pp 129ndash131 2005
[39] A Pilorget B Annabi F Bouzeghrane J Marvaldi J Luisand R Beliveau ldquoInhibition of angiogenic properties of brainendothelial cells by platelet-derived sphingosine-1-phosphaterdquoJournal of Cerebral Blood Flow and Metabolism vol 25 no 9pp 1171ndash1182 2005
[40] A Wang P Das R C Switzer III T E Golde and J LJankowsky ldquoRobust amyloid clearance in a mouse model ofAlzheimerrsquos disease provides novel insights into themechanismof amyloid-120573 immunotherapyrdquo Journal of Neuroscience vol 31no 11 pp 4124ndash4136 2011
[41] G Krabbe A Halle V Matyash et al ldquoFunctional impairmentof microglia coincides with Beta-amyloid deposition in micewith Alzheimer-like pathologyrdquo PLoS ONE vol 8 no 4 ArticleID e60921 2013
[42] Y H Liu B Giunta H-D Zhou J Tan and Y J WangldquoImmunotherapy for Alzheimer disease-the challenge ofadverse effectsrdquo Nature Reviews Neurology vol 8 no 8 pp465ndash469 2012
[43] B Solomon R Koppel D Frankel and E Hanan-AharonldquoDisaggregation of Alzheimer120573-amyloid by site-directedmAbrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 94 no 8 pp 4109ndash4112 1997
[44] F Bard C Cannon R Barbour et al ldquoPeripherally adminis-tered antibodies against amyloid 120573-peptide enter the centralnervous system and reduce pathology in a mouse model ofAlzheimer diseaserdquo Nature Medicine vol 6 no 8 pp 916ndash9192000
[45] R B DeMattos K R Bales D J Cummins J C Dodart S MPaul and D M Holtzman ldquoPeripheral anti-A120573 antibody altersCNS and plasma A120573 clearance and decreases brain A120573 burdenin a mouse model of Alzheimerrsquos diseaserdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 98 no 15 pp 8850ndash8855 2001
[46] C A Lemere E T Spooner J LaFrancois et al ldquoEvidence forperipheral clearance of cerebral A120573 protein following chronic
16 BioMed Research International
active A120573 immunization in PSAPP micerdquo Neurobiology ofDisease vol 14 no 1 pp 10ndash18 2003
[47] T Town M Vendrame A Patel et al ldquoReduced Th1 andenhanced Th2 immunity after immunization with Alzheimerrsquos120573-amyloid
1minus42rdquo Journal of Neuroimmunology vol 132 no 1-2
pp 49ndash59 2002[48] R Cacabelos ldquoPharmacogenomics in Alzheimerrsquos diseaserdquo
Methods in Molecular Biology vol 448 pp 213ndash357 2008[49] R Cacabelos ldquoPharmacogenomics and therapeutic strategies
for dementiardquo Expert Review of Molecular Diagnostics vol 9no 6 pp 567ndash611 2009
[50] D van Dam R DrsquoHooge M Staufenbiel C van Ginneken Fvan Meir and P P de Deyn ldquoAge-dependent cognitive declinein the APP23 model precedes amyloid depositionrdquo EuropeanJournal of Neuroscience vol 17 no 2 pp 388ndash396 2003
[51] M Dumont C Strazielle M Staufenbiel and R LalondeldquoSpatial learning and exploration of environmental stimuli in24-month-old female APP23 transgenic mice with the Swedishmutationrdquo Brain Research vol 1024 no 1-2 pp 113ndash121 2004
[52] R Lalonde M Dumont M Staufenbiel C Sturchler-Pierratand C Strazielle ldquoSpatial learning exploration anxiety andmotor coordination in female APP23 transgenic mice with theSwedish mutationrdquo Brain Research vol 956 no 1 pp 36ndash442002
[53] F J Huff F Boller F Lucchelli R Querriera J Beyer and SBelle ldquoThe neurologic examination in patients with probableAlzheimerrsquos diseaserdquo Archives of Neurology vol 44 no 9 pp929ndash932 1987
[54] A F Pettersson M Engardt and L O Wahlund ldquoActivitylevel and balance in subjects with mild Alzheimerrsquos diseaserdquoDementia and Geriatric Cognitive Disorders vol 13 no 4 pp213ndash216 2002
[55] L Calandreau B Desgranges R Jaffard and A DesmedtldquoSwitching from contextual to tone fear conditioning and viceversa the key role of the glutamatergic hippocampal-lateralseptal neurotransmissionrdquo Learning amp Memory vol 17 no 9pp 440ndash443 2010
[56] F Trinchese S Liu F Battaglia SWalter PMMathews andOArancio ldquoProgressive age-related development of Alzheimer-like pathology in APPPS1 micerdquo Annals of Neurology vol 55no 6 pp 801ndash814 2004
[57] D M Wilcock A Rojiani A Rosenthal et al ldquoPassiveimmunotherapy against A120573 in aged APP-transgenic micereverses cognitive deficits and depletes parenchymal amyloiddeposits in spite of increased vascular amyloid and microhem-orrhagerdquo Journal of Neuroinflammation vol 1 article 24 2004
[58] S A Ferguson S Sarkar and L C Schmued ldquoLongitudinalbehavioral changes in the APPPS1 transgenic AlzheimerrsquosDisease modelrdquo Behavioural Brain Research vol 242 no 1 pp125ndash134 2013
[59] D Wilcock and C Colton ldquoImmunotherapy vascular pathol-ogy and microhemorrhages in transgenic micerdquo CNS andNeurological DisordersmdashDrug Targets vol 8 no 1 pp 50ndash642009
[60] M Grundman S Gilman R S Black et al ldquoAn alternativemethod for estimating efficacy of the AN1792 vaccine foralzheimer diseaserdquo Neurology vol 71 no 9 pp 697ndash698 2008
[61] D Boche E Zotova R O Weller et al ldquoConsequence of A120573immunization on the vasculature of human Alzheimerrsquos diseasebrainrdquo Brain vol 131 no 12 pp 3299ndash3310 2008
[62] M Pfeifer S Boncristiano L Bondolfi et al ldquoCerebral hemor-rhage after passive anti-a120573 immunotherapyrdquo Science vol 298no 5597 p 1379 2002
[63] M M Racke L I Boone D L Hepburn et al ldquoExacerbationof cerebral amyloid angiopathy-associatedmicrohemorrhage inamyloid precursor protein transgenic mice by immunotherapyis dependent on antibody recognition of deposited forms ofamyloid 120573rdquo Journal of Neuroscience vol 25 no 3 pp 629ndash6362005
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

8 BioMed Research International
Gr E
(PBS
)
0102030405060708090
Preventive treatmentTherapeutic treatment
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Mea
n of
120573-a
myl
oid
plaq
ues
sect
ion
Gr A
(A120573+
S1P+
L)(E
B101
)
Figure 3 Comparative analysis of A120573 plaque burden in the brainsof APPPS1 mice Mean of A120573 plaque burden in the hippocampaland cortical regions of APPswePS1ΔE9 mice in the five treatmentgroups The mean of the A120573 burden is significantly reduced (119875 lt005) in Group A (EB101) when compared with Groups BndashE duringpreventive treatment and markedly reduced in the therapeutictreatment period Data are presented asmeanplusmn standard error of themean (SEM) Densitymean of A120573 plaques in Tgmice hippocampus
05
1015202530354045
Preventive treatmentTherapeutic treatment
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Gr A
(A120573+
S1P+
L)(E
B101
)
Mea
n of
120573-a
myl
oid
plaq
ues a
rea
Figure 4 Comparative quantification of A120573 burden area in thebrain of APPPS1 mice Quantitative analysis of A120573 burden areain the hippocampal and cortical regions of APPswePS1ΔE9 micetreated with EB101 A120573liposomes S1Pliposomes A120573
42+ FAIFA
and PBS represented by the number of pixels inside the stained areaof each A120573 plaque This graphic shows that A120573 plaques of EB101-treated mice (Group A) are significantly smaller in size (119875 lt 005)than those of the other four treatment groups Data are presented asmean plusmn SEM Mean of A120573 plaques area in Tg mice hippocampus
a reduced staining intensity of the A120573 plaques in the brainregions of mouse Group A contrasting with the intenseA120573-immunoreactivity of plaques in the other experimentalmouse groups as shown in the photomicrographs of Figures2(k)ndash2(t)
33 Immune Response Effects of EB101 versus A12057342+ CFAIFA
Vaccine Astrocyte activation implicated in neuroinflam-mation amyloidogenesis and neuronal cell death in AD
was compared after treatment of the transgenic mice withEB101 and A120573
42+ CFAIFA Immunohistochemical analysis
of GFAP-reactive cells was carried out in the brains ofthe treated mice in the preventive and therapeutic studies(Figure 5) After the preventive treatment (Figures 5(a)ndash5(j))EB101 significantly reduced the density of GFAP-reactive cellclusters in the hippocampal and cortical sections as comparedwith A120573
42+ CFAIFA (D) A120573Lip without S1P (B) Lip with
S1P (C) and PBS (E) The distribution of GFAP immunore-active cells in the transverse section of the mouse brainstreated with EB101 shows a few scattered GFAP-reactiveclusters mainly at the CA1 hippocampal layers contrastingwith numerous dystrophic reactive astrocytes observed indifferent hippocampal areas of mouse brains in Groups BndashE(Figures 5(b)ndash5(e)) Similar neuroinflammation pattern wasobserved in the retrosplenial cortical layers where a fewGFAP-reactive clusters were observed in EB101 treated mice(Figure 5(f)) compared with their profuse presence in theother experimental groups (Figures 5(g)ndash5(j)) After thera-peutic treatment the GFAP immunoreactivity distributionpattern in mouse brains treated with EB101 was similar tocontrol mice (wild-type) mostly devoid of reactive GFAPclusters except a few scattered ones in the CA1 hippocampalexternal layers (Figure 5(k)) In contrast there is a moderatedensity in Group D (Figure 5(n)) and extensive density inGroup B (Figure 5(l)) Group C (Figure 5(m)) and Group Emice (Figure 5(o)) Photomicrographs of retrosplenial cortexshow similar neuroinflammation pattern where a few GFAPclusters detected in the EB101 treated mice (Figure 5(p))contrast with a more extensive density of clusters in the othergroups (Figures 5(q)ndash5(t)) No astrocytosis was observed inwild-type mice during preventive or therapeutic treatment
34 Comparative A120573 Antibody Titers and Th1Th2 CytokineLevels in Treated Mice A120573 antibody titers in the sera weredetermined by ELISA during mouse immunization(Figure 6) Preventive treatment with EB101 resulted in amarked increase of specific IgG A120573
1ndash42 antibody productionin all animals Mice in A and D groups developed andmaintained serum antibody titers between 1 2000 and1 8000 Mice in Groups B C and E showed no significantanti-A120573 antibody production Antibody production and titerin Group B were three and four times lower than those ofGroups D and A respectively After therapeutic treatment allmice treated with EB101 and A120573
42+ FAIFA produced high
levels of IgG antibodies Titer levels of Groups A and D weresignificantly increased to those detected after preventivetreatment (119875 lt 001 versus Group D 119875 lt 0001 versusGroups B C and E) although antibody level in Group D was13-fold lower than in Group A (Figure 6)The effect of EB101(Group A) and A120573
42+ CFAIFA (Group B) vaccines on Th1
andTh2 cytokines was also studied (Figure 7)Measurementsof Th1 and Th2 cytokines in sera of immunized transgenicmice indicate that significant changes occurring in proteinlevels of Th1-related inflammatory mediators followingpreventive (Figures 7(a)ndash7(c)) and therapeutic (Figures 7(d)ndash7(g)) treatment with EB101 vaccine A significant increaseof Th1 cytokine profile was observed in preventive treatment
BioMed Research International 9
Gr E
(PBS
)
(d)
(e)
(i)
(j)
(n)
(o)
(s)
(t)
Cx
GFAP
GFAP GFAP GFAP GFAP
GFAP GFAP GFAP
Cx Cx
Cx
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
GFAP
Gr A
(EB1
01)
(a)
(b)
(c)
(f)
(g)
(h)
Preventive treatment Therapeutic treatment
PoDG
MoDG
(k)
(l)
(m)
(p)
(q)
(r)DG
GFAP GFAP GFAP
GFAPGFAPGFAP GFAP
GFAP GFAP GFAP GFAP
PoDGDG
DG
CA1
Cx
Cx
Cx
Cx
Cx
Cx
PoDG
MoDG
DG
CA1
MoDGDG
CA1
PoDG
MoDG
DG
CA1
Cx
CxCx
MoDG
MoDG
PoDG
CA1CA1
Cx
Gr D
(A120573+
FA)
Gr C
(S1P
+L)
Gr B
(A120573+
L)
Figure 5 Comparative effect of EB101 vaccine on astrocytosis Comparative photomicrographs of GFAP immunoreactivity in thehippocampus ((a)ndash(e) (k)ndash(o)) and cortical ((f)ndash(j) (p)ndash(t)) brain regions at the preventive treatment phase ((a)ndash(j)) and therapeutictreatment group ((k)ndash(t)) Preventive treatment transverse brain sections of 11-month-old mice treated with EB101 show an almost completeprevention of astrocytosis in the detailed sections of the dentate gyrus (a) contrasting with numerous dystrophic reactive astrocytes typicalof an inflammatory reaction observed in different hippocampal corresponding areas of mouse brains in Group B (A120573Lip-treated mice (b))Group C (LipS1P-treated mice (c)) Group D (A120573
42+ CFAIFA-treated mice (d)) and Group E (PBS-treated mice (e)) Immunoreactive
astrocytosis in the brain sections of WT mice is absent (data not shown) Detailed sections of the parietal cortex of treated mice show agradient density of reactive astrocytes immunoreactive to GFAP indicating a neuroinflammation pattern Comparative cortical sections shownumerous astrocytosis areas inmice Groups C and E ((h) (j)) andwide reactive astrocytes clusters inmice Groups B andD ((g) (i)) while justa few of them are observed in Group A (f)Therapeutic treatment transverse brain sections of 18-month-old mice of Group A show a notablereduction of astrocytosis in the hippocampal region (k) after EB101 vaccine immunization Note the astrocytosis contrast density betweenEB101-treatedmice and the other treated groups ((l)ndash(o)) where numerous immunoreactive GFAP clusters are observed in the correspondentbrain sections A few immunoreactive GFAP clusters are observed in the parietal cortex of EB101 treated mice group while a moderate ((q)(s)) to high ((r) (t)) density of these neuropathological inflammation clusters is apparent in the other treatment groups (Groups BndashE) Forabbreviations see list Scale bar 100120583m
10 BioMed Research International
002040608
1121416
Preventive treatmentTherapeutic treatment
Ant
i-120573-a
myl
oid
IgG
antib
odie
s
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Gr A
(A120573+
S1P+
L)(E
B101
)
Figure 6 Quantification of specific anti-120573-amyloid IgG in immu-nized APPPS1 mice The presence of anti-120573-amyloid antibodiesmeasured in the sera of vaccinated and control mice groups duringthe preventive and therapeutic treatments indicates that EB101liposome vaccine was able to induce the highest production ofspecific IgG antibodies Compared to theGroupD immunizedmicethe level of antibodies observed in the Group A was also highersuggesting a more effective immune response (119875 lt 001 versusGroup D 119875 lt 0001 versus Groups B C and E) Quantification ofspecific anti-120573-amyloid IgG in immunized Tg mice
in Group D with respect to all other groups mainlyreported by the high levels of TNF-120572 (Figure 7(b)) andIFN-120574 (Figure 7(c)) cytokine secretions while due to thehigh mortality rate during the therapeutic treatment nosignificant data was obtained from this mice group Inaddition a tendency toTh1 secretion reduction was observedinGroupAwith respect to all other groups in both preventiveand therapeutic treatment The changes in Th2 protein levels(Figure 7(d)) followed the inverse trend where the highestdifferences between Group A and the other treatment groupswere observed in the IL-4 IL-5 and IL-10 cytokine secretions(119875 lt 005 versus Group D 119875 lt 001 versus Group E Figures7(e)ndash7(g))
35 Comparative Motor Coordination and Memory Perfor-mances In order to validate the locomotion integrity duringthe entire immunotherapeutic process as reported in similarprevious studies motor coordination tests have showed thatEB101-treated mice spent significantly more time on theaccelerating rotarodThismotor coordination task shows thatEB101 immunization affected positively their performance(Figure 8(a)) A120573
42liposome (B) S1P-containing liposomes
(C) and PBS (E) (119875 lt 0006) mice performed poorly incomparison to EB101 mice (119875 lt 005) or A120573
42+ CFAIFA
mice (119875 lt 002) in motor abilities and coordination Thefear conditioning-active avoidance test was used to assess thespatial learning and memory functions As expected bothEB101 and A120573
42+ CFAIFA immunized mice readily learned
the location of the platform as shown by a decrease in theescape latency over training days while the control group
mice exhibited no significant changes during 6-day trialsThe average escape latencies of EB101 (68 plusmn 2 s 761 plusmn 22 s)andA120573
42+CFAIFA (116plusmn33 s 123plusmn21 s) immunizedmice
at the end of preventive and therapeutic periods respectivelyduring the consecutive 6 days trial were significantly shorterthan the other treatment groups (A120573
42liposome 163 plusmn 31 s
175 plusmn 28 s S1P-containing liposomes 202 plusmn 19 s 248 plusmn27 s) and the control (2116 plusmn 38 s 268 plusmn 22 s) (119875 lt 001)Therewas no significant difference between EB101 andA120573
42+
CFAIFA immunized group (119875 gt 005) (Figure 8(b)) Theseresults showed that spatial learning was improved in theseimmunized mice
4 Discussion
In this study we have compared a novel active A120573 imm-unotherapy the EB101 vaccine against A120573 with the A120573
42+
CFAIFA vaccine that was first taken into clinical trialswith coadjuvant Qs-21 modification (AN1792 by Orgogozoet al [19]) This type of vaccine design was thought toinduce a strong T-cell mediated inflammatory response (IL-2 and IFN-c positive responses indicative of a Class II(CD4) Th-1 type response [33] leading to acute menin-goencephalitis in the 6 of the patients) The EB101 activeimmunotherapeutic vaccine was primarily designed to avoidundesired inflammatory side effects both in mice andhumans (without need of coadjuvant modification) whileinducing effective antibody titer rates We demonstrated thatlong-term immunization with EB101 induced A120573 antibodiesin the preventive and the therapeutic treatment periodswhich effectively reduced amyloid deposition in both corticaland hippocampal brain regions and prevented neuroinflam-mation probably by blocking toxicity in the surroundingneurons However the AN1792 vaccine was shown to inducein 6 of the patients an inflammatory reaction that mighthave led to the encephalitis type reaction observed in theclinical trial which was responsible for the immediate haltin those clinical studies It seems that AN1792 immunizationdelivered in Qs-21 and polysorbate 80 adjuvant induced aTh1 type response that likely led to autoaggressive T cells andaseptic meningoencephalitis In the present experiments wedemonstrated that a judiciously selected adjuvant produces adifferent humoral response [22] Indeed we have shown thatour novel immunotherapeutic strategy boosts specific A120573
1ndash42antibody generation and induces a Th2 noninflammatoryimmune response This boosted effect is in accordance withprevious immunotherapeutic studies [34] Taking togetherall the present findings these results suggest that EB101vaccine can achieve in a future clinical trial immunogenicresponse rates quite similar to the AN1792 without theinflammatory effects reported The immunohistochemicalanalysis obtained in the present study shows that all the EB101immunized mice resulted in an almost complete preventionof A120573 deposition in the brain with only a few compactA120573 plaques in the retrosplenial and dentate gyrus regionsAlthough there was a slightly increase in death in EB101treated mice compared to controls this significant reductionof A120573 plaques in EB101 treated mice was proven to be more
BioMed Research International 11
Gr A Gr B Gr C Gr D Gr EPreventive treatmentTherapeutic treatment
0005001000150020002500300035004000
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
(a) Th1
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
000
1000
2000
3000
4000
5000TNF-120572
(b) Th1
000500
10001500200025003000
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
IFN-120574
(c) Th1
000500
100015002000250030003500
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
Group A Group B Group C Group D Group EPreventive treatmentTherapeutic treatment
(d) Th2
000
500
1000
1500
2000
2500
3000IL-4
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(e) Th2
000500
100015002000250030003500
IL-5
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(f) Th2
3500IL-10
000500
10001500200025003000
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(g) Th2
Figure 7 Detection of Th1 and Th2 cytokine profile Measurements of the effect of EB101 and A12057342
+ CFAIFA vaccines on Th1 and Th2cytokines profile during preventive and therapeutic treatments ((a)ndash(c)) A significant increase of Th1 cytokine profile was observed inpreventive treatment in Group D with respect to all other groups while a tendency to a reduction inTh1 secretion was observed in Group Awith respect to all other groups in both preventive and therapeutic treatment ((d)ndash(g)) The changes in Th2 protein levels were observed inthe IL-4 IL-5 and IL-10 cytokine secretions of Group A (119875 lt 005 versus Group D 119875 lt 001 versus Group E)
12 BioMed Research International
Gr A
(EB1
01)
05
101520253035
Preventive treatment Therapeutic treatment
Mea
n fa
lling
late
ncy
(s)
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)(a) Graphic of motor coordination and balance
05
1015202530
Mea
n re
spon
se ti
me (
s)
Gr A
(EB1
01)
Preventive treatment Therapeutic treatment
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
(b) Graphic of associative learning and memory
Figure 8 Analysis of motor coordination and balance (a) Rotarod results of motor coordination and balance test after the preventive andtherapeutic treatment showing improved effect of EB101 and A120573
42+ CFAIFA-vaccine compared to other treated mice groups (119875 lt 005)
and (119875 lt 002) versus control mice Latency to fall on the rotarod apparatus was significantly improved in EB101 immunized mice Valuesrepresent means of all six trials of a groupday (b) Spatial learning and memory tested by fear conditioning-active avoidance test The meanresponse time of each treatment group to reach the platform is represented EB101 and A120573
42+ CFAIFA immunized mice had significantly
lower latencies than the other treatment groups (119875 lt 001) versus control mice
efficient than that obtained with A12057342+ CFAIFA immuniza-
tions A comparison of A120573 plaque reduction rates amongtreated groups surprisingly showed that other than EB101and A120573
42+ CFAIFA treated groups some amyloid deposits
were also reduced in Groups B (A1205731ndash42liposome) and C
(S1P-containing liposomes) Although the reduction rates inthese mice were markedly low the subtle lowering effectmight be due to the interaction between the immunizationelements (A120573
1ndash42 and S1P) and the mouse immunogenicsystem as reported by others [35] Based on our presentresults S1P one of the components of the adjuvant used withEB101 might be responsible for the neuroprotective effectobserved in the vulnerable neuronal areas of EB101 treatedmice The advantage in the A120573 load preventionreduction bycombining A120573
1ndash42 and S1P in a unique formulation vaccinehas been also observed in the reduced hippocampal load ofEB101 immunizedmice (Gr A) when comparedwith differentvaccine components (Gr B-C) In fact S1P appears to have ademonstrated therapeutic potential as a regenerative agent inthe nervous system since it controls migration of neuronalstem cells toward a site of spinal cord injury [36] inducecytoskeletal rearrangements to promote transmitter secretion[37] and also trigger diverse cellular effects including angio-genesis cardiac development immunity cell motility andneurite extension [38 39]Therefore it is reasonable to expectthat such a potent biologically active sphingolipid by itselfmay play an important role in the A120573 plaque reduction Inthis study the clearance efficiency rate of each treatment wasanalyzed and results suggested that EB101 (6459) appearsto be more effective than A120573
42+ CFAIFA (1920) vaccine
in both preventive and therapeutic groups The effectivenessof EB101 in older transgenic animals (18 months) showeda clearance effect of 59 suggesting that EB101 may alsohave therapeutic potential inADdiagnosticated patient trials
Similar conclusion was reached byWang and colleagues [40]who demonstrated that combination therapy in aged animalsprovides more valuable insights into a possible translationto humansOur immunologicalmeasurements demonstratedonce again that neuroinflammation develops in an A120573-dependent fashion early in the course of AD [41] and canbe alleviated or reversed by interventional A120573 vaccinationsuch as EB101 Among the possible mechanisms of A120573clearance proposed in AD immunotherapy [42] our resultssuggested that theymay overlap and probably be disease statedependent since the prevention and the therapeutic vaccinewould likely benefit from the transport of A120573 antibodies intothe CNS However some mechanisms of action such as thepreventive effect of A120573 antibodies in the A120573 aggregation [43]microglial phagocytosis induced by the Fc binding portionsbetween A120573 antibodies and microglia receptor [44] and theshift in the gradient of A120573 transport across the blood-brainbarrier due to the increased concentration of A120573 antibodiesin the vascular system resulting in an increase in effluxfrom the brain to blood [45 46] may be playing a role inboth preventive and therapeutic conditions Our findingstherefore suggest that novel treatment strategies aimed atmaintaining or increasing microglial function and restoringthe balance to the immunological system may represent anattractive therapeutic approach even at the advanced stagesof ADThe immunization results obtained with EB101 duringboth preventive and therapeutic treatment showed nomicro-hemorrhages infiltration of lymphocytes or encephalitis-like reaction in the brain suggesting that EB101 does notinduce A120573-specific Th1 reaction possibly due to the neu-roprotective effect of S1P Delivering A120573
1ndash42 in a liposomal-S1P-adjuvant resulted in an effective immunological responseincluding the induction of a Th2 T-cell response whicheffectively prevented neuroinflammation in an animal model
BioMed Research International 13
of AD as reported previously [24 47] Moreover presentresults demonstrated that EB101 did not induce astrogliosisin APPPS1 transgenic mice whereas some GFAP-positiveastrocyte clusters were aggregated in the affected brain areasof mice treated with A120573
42+ CFAIFA These results indicate
that EB101 may result in an effective treatment in AD neu-ropathology due to the absence of peripheral proinflamma-tory cytokine secretion and the absence of astroglial activa-tion in the brains of EB101 treated mice when compared withA12057342
+ CFAIFA immunotherapeutic response In a clinicalsetting however we may expect a wide differential responsein part depending upon the genomic profile of each patient[48 49] In AD behavioural studies in particular amongthe motor coordination tests the rotarod performance hasbeen reported to be impaired in some AD transgenic micemodels bearing A120573 plaques [50 51] This result was found inmutants prior to A120573 plaque formation at 3 and 6 months ofage but was not reproduced after plaques had been formedat 16 and 24 months of age [52] perhaps because of thedeclining abilities of older normal mice Here we presentthe effects of vaccine treatments in APPPS1 mice on motorcoordination a neglected aspect of the AD experimentalbehavior but justified on the basis of the deficient posturalcontrol reported in patients with Alzheimerrsquos disease [53]sometimes even during early stages [54] Our longitudinalmotor behavioral results showed no significant differencesin the ambulatory movement grip strength or coordinationaddressed by their performance in the rotating rod betweenAPPPS1 and wild-type mice for any age group studiedin contrast with other treated groups demonstrating thatlongitudinal motor assessments of AD transgenic mousemodels can provide valuable information with regard tothe effect of immunization on the progression of behavioraldeficits and overall motor activity As shown the EB101-treated group of mice exhibited longer latencies and betterbalancing skills indicating the absence of any negative effectof EB101 immunization on these mice The classical fearconditioning test is assumed to reveal the functional integrityof the hippocampus [55] representing a special memorytask to evaluate the AD models and its improvement afterimmunization periods In this APPPS1 mouse model thefirst associative learning and spatial workingmemory deficitsare shown at 5-6 months of age [56] although its impair-ment progression is relatively slow compared to most othermodels At the end of preventive immunization period EB101and A120573
42+ CFAIFA immunized mice have significantly
shorter escape latencies than the other treatment groups(A12057342liposome S1P-containing liposomes) and the controls
in the fear conditioning test Shorter escape latencies duringspatial learning indicated that deficits of spatial memorywere attenuated in EB101 and A120573
42+ CFAIFA immunized
mice Previous studies have showed that by 15 months ofage the spatial working memory is consistently impairedthroughout the rest of the life span [31 57] At the end oftherapeutic immunization period both EB101 and A120573
42+
CFAIFA immunized mice showed significantly improvedresults on the hippocampus-dependent spatial learning andmemory test when compared with other treatment groupsand the controls Taken together these observations are
in agreement with the impairment in conditioned learningin response to a tone stimulus of aged APPPS1 micealthough when immunized with EB101 during preventiveor therapeutic periods such impairments are significantlyattenuated
In summary the present results corroborate our previouspublished findings showing that EB101 immunization reducesamyloid accumulation in the APPPS1 mouse brain [2425] The strongest reduction in the preventive immunizationwas observed in the retrosplenial cortex followed by thehippocampus and entorhinal-piriform cortex Our resultsfurther support that EB101 ismore effectivewhen themice areimmunized prior to developing plaques in the hippocampaland cortical brain regions There is extensive evidence inmouse preclinical studies [58] and human clinical trials [5960] that the adverse effects of active and passive A120573 vaccinesresult in microhemorrhages and encephalitic reactions [60ndash63] yet themechanismbywhich they occur has not been fullyelucidated In the treatment sections analyzed we found nomicrohemorrhages or acute inflammatory processes in all thesurvived mice of the group indicating that EB101 therapeuticapproach presents no adverse inflammatory effects in thisAD mice models Thus our present results unequivocallydemonstrate that EB101 vaccinemay not only halt AD-relatedpathology as a preventive treatment but may also reverse it inestablished cases related to the improvement of the learningandmemory in both cases leading to a robust immunologicaltherapeutic response Present experiments also demonstratethat EB101 exerts more significant and substantial effects onthe reduction of A120573 deposition than A120573
42+CFAIFA vaccine
in A120573 mice models of AD However further exploration ofthe contribution of genetic environmental and physiologicalfactors in the development resistance to endogenous A120573-antibody is needed in order to optimize and modulate thetherapeutic strategies currently being developed
5 Conclusions
The present study carried out in APPPS1 transgenic mousemodel of AD compared the A120573 burden reduction andcellular immune responses produced by two immunizationdelivery strategies of A120573 in Freund adjuvant (described byWebster et al [10]) and in liposomeS1P matrix (describedby Schenk et al [17])The EB101 immunotherapeutic strategyboosted antibody generation A120573 plaque reduction and ananti-inflammatory response more effectively than A120573
42+
CFAIFA especially if given prior to or in the early stages ofpathology The present results also showed that vaccinationwith EB101 prevented the formation of new plaques andpresumably led to a reduction in established plaques whileinducing aTh2 immune reactionwithout encephalitogenic T-cell responses or brain inflammation related with astrogliosisHowever despite the efficacy demonstrated of this novelvaccine approach further preclinical and clinical studieson EB101 are warranted to evaluate its efficacy and themechanism for the future utility of such vaccine for ADtherapy
14 BioMed Research International
List of Abbreviations
AD Alzheimerrsquos diseaseA120573 120573-Amyloid proteinAPP Amyloid precursor proteinAPT Anterior pretectal nucleusCA1 Field CA1 hippocampusCA3 Field CA3 hippocampusCPu Caudate putamen (striatum)Cx CortexDG Dentate gyrusDN Dystrophic neuriteEct Entorhinal cortexCFA Complete Freundrsquos adjuvantGrDG Granular dentate gyrusIFA Incomplete Freundrsquos adjuvantIL InterleukinLPMR Lateral posterior thalamic nucleus (mediorostral)MoDG Molecular dentate gyrusPir Piriform cortexPoDG Polymorph dentate gyrusRSGb Retrosplenial granular cortex b regionTg TransgenicTh T helper
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Ivan Carrera Ignacio Etcheverrıa Lucıa Fernandez-NovoaValter Lombardi Ramon Cacabelos and Carmen Vigo con-ceived and designed the experiments Ivan Carrera and Igna-cio Etcheverrıa performed the experiments Ivan CarreraIgnacio Etcheverrıa and Valter Lombardi analyzed the dataIvan Carrera Ignacio Etcheverrıa Lucıa Fernandez-Novoaand Valter Lombardi contributed reagentsmaterialsanalysistools Ivan Carrera Carmen Vigo andMadepalli KrishnappaLakshmana wrote the paper
Acknowledgments
This work was supported by National Institute of Aging(NIA)NIH Grant no 1R01AG036859-01 (Madepalli Krish-nappa Lakshmana) and the financial support of the ProgramaOperativo Feder Galicia 2007ndash2013 (IN841C 201254 to PhDCarrera I)
References
[1] R Cacabelos L Fernandez-Novoa V Lombardi Y Kubotaand M Takeda ldquoMolecular genetics of Alzheimerrsquos diseaseand agingrdquoMethods and Findings in Experimental and ClinicalPharmacology vol 27 pp 1ndash573 2005
[2] P T Nelson I Alafuzoff E H Bigio et al ldquoCorrelationof alzheimer disease neuropathologic changes with cognitive
status a review of the literaturerdquo Journal of Neuropathology andExperimental Neurology vol 71 no 5 pp 362ndash381 2012
[3] JNaslundVHaroutunian RMohs et al ldquoCorrelation betweenelevated levels of amyloid 120573-peptide in the brain and cognitivedeclinerdquo Journal of the American Medical Association vol 283no 12 pp 1571ndash1577 2000
[4] A Koyama O I Okereke T Yang D Blacker D J Selkoeand F Grodstein ldquoPlasma amyloid-120573 as a predictor of dementiaand cognitive decline a systematic review and meta-analysisrdquoArchives of Neurology vol 69 no 7 pp 824ndash831 2012
[5] A M Fagan C M Roe C Xiong M A Mintun J C Morrisand D M Holtzman ldquoCerebrospinal fluid tau120573-amyloid42ratio as a prediction of cognitive decline in nondemented olderadultsrdquo Archives of Neurology vol 64 no 3 pp 343ndash349 2007
[6] S Parvathy P Davies V Haroutunian et al ldquoCorrelationbetween A120573-40- A120573-42- and A120573x-43-containing amyloidplaques and cognitive declinerdquo Archives of Neurology vol 58no 12 pp 2025ndash2032 2001
[7] E Karran M Mercken and B D Strooper ldquoThe amyloidcascade hypothesis for Alzheimerrsquos disease an appraisal for thedevelopment of therapeuticsrdquo Nature Reviews Drug Discoveryvol 10 no 9 pp 698ndash712 2011
[8] J L Jankowsky D J Fadale J Anderson et al ldquoMutantpresenilins specifically elevate the levels of the 42 residue 120573-amyloid peptide in vivo evidence for augmentation of a 42-specific 120574 secretaserdquo Human Molecular Genetics vol 13 no 2pp 159ndash170 2004
[9] L Holcomb M N Gordon E Mcgowan et al ldquoAcceleratedAlzheimer-type phenotype in transgenic mice carrying bothmutant amyloid precursor protein and presenilin 1 transgenesrdquoNature Medicine vol 4 no 1 pp 97ndash100 1998
[10] S JWebster A D Bachstetter and L J van Eldik ldquoComprehen-sive behavioral characterization of an APPPS-1 double knock-in mouse model of Alzheimerrsquos diseaserdquo Alzheimerrsquos ResearchandTherapy vol 5 no 3 article 28 2013
[11] H Xiong D Callaghan J Wodzinska et al ldquoBiochemical andbehavioral characterization of the double transgenic mousemodel (APPswePS1dE9) of Alzheimerrsquos diseaserdquo NeuroscienceBulletin vol 27 no 4 pp 221ndash232 2011
[12] M Garcia-Alloza L A Borrelli B T Hyman and B J BacskaildquoAntioxidants have a rapid and long-lasting effect on neuriticabnormalities in APPPS1 micerdquo Neurobiology of Aging vol 31no 12 pp 2058ndash2068 2010
[13] M Meyer-Luehmann M Mielke T L Spires-Jones et al ldquoAreporter of local dendritic translocation shows plaque-relatedloss of neural system function in APP-transgenic micerdquo Journalof Neuroscience vol 29 no 40 pp 12636ndash12640 2009
[14] E Machova V Rudajev H Smyckova H Koivisto H Tanilaand V Dolezal ldquoFunctional cholinergic damage develops withamyloid accumulation in young adult APPswePS1dE9 trans-genic micerdquo Neurobiology of Disease vol 38 no 1 pp 27ndash352010
[15] T Malm J Koistinaho and K Kanninen ldquoUtilization of APP-swePS1dE9 transgenic mice in research of Alzheimerrsquos diseasefocus on gene therapy and cell-based therapy applicationsrdquoInternational Journal of Alzheimerrsquos Disease vol 2011 Article ID517160 8 pages 2011
[16] J Lee and P Han ldquoAn update of animal models of alzheimerdisease with a reevaluation of plaque depositionsrdquoExperimentalNeurobiology vol 22 pp 84ndash95 2013
BioMed Research International 15
[17] D Schenk R Barbour W Dunn et al ldquoImmunization withamyloid-120573 attenuates Alzheimer disease-like pathology in thePDAPP mouserdquo Nature vol 400 no 6740 pp 173ndash177 1999
[18] H J Fu B Liu J L Frost and C A Lemere ldquoAmyloid-120573immunotherapy for Alzheimerrsquos diseaserdquoCNS and NeurologicalDisorders-Drug Targets vol 9 no 2 pp 197ndash206 2010
[19] J-M Orgogozo S Gilman J-F Dartigues et al ldquoSubacutemeningoencephalitis in a subset of patients with AD after A120573
42
immunizationrdquo Neurology vol 61 no 1 pp 46ndash54 2003[20] J A R Nicolll DWilkinson CHolmes P Steart HMarkham
andROWeller ldquoNeuropathology of humanAlzheimer diseaseafter immunization with amyloid-120573 peptide a case reportrdquoNature Medicine vol 9 no 4 pp 448ndash452 2003
[21] I Ferrer M Boada Rovira M L Sanchez Guerra M JRey and F Costa-Jussa ldquoNeuropathology and pathogenesisof encephalitis following amyloid-Neuropathology and patho-genesis of encephalitis following amyloid-beta immunizationin Alzheimerrsquos disease immunization in Alzheimerrsquos diseaserdquoBrain Pathology vol 14 no 1 pp 11ndash20 2004
[22] D H Cribbs A Ghochikyan V Vasilevko et al ldquoAdjuvant-dependent modulation ofTh1 andTh2 responses to immuniza-tion with 120573-amyloidrdquo International Immunology vol 15 no 4pp 505ndash514 2003
[23] C Holmes D Boche D Wilkinson et al ldquoLong-term effectsof A120573
42immunisation in Alzheimerrsquos disease follow-up of a
randomised placebo-controlled phase I trialrdquo The Lancet vol372 no 9634 pp 216ndash223 2008
[24] I Carrera I Etcheverrıa L Fernandez-Novoa V LombardiR Cacabelos and C Vigo ldquoVaccine development to treatAlzheimerrsquos disease neuropathology in APPPS1 transgenicmicerdquo International Journal of Alzheimerrsquos Disease vol 2012Article ID 376138 17 pages 2012
[25] I Carrera I Etcheverrıa Y Li et al ldquoImmunocytochemicalcharacterization of Alzheimer disease hallmarks in APPPS1transgenic mice treated with a new anti-amyloid-120573 vaccinerdquoBioMed Research International vol 2013 Article ID 709145 12pages 2013
[26] V Anelli R Bassi G Tettamanti P Viani and L Riboni ldquoExtra-cellular release of newly synthesized sphingosine-1-phosphateby cerebellar granule cells and astrocytesrdquo Journal of Neuro-chemistry vol 92 no 5 pp 1204ndash1215 2005
[27] S Schroeter K Khan R Barbour et al ldquoImmunotherapyreduces vascular amyloid-120573 in PDAPP micerdquo Journal of Neu-roscience vol 28 no 27 pp 6787ndash6793 2008
[28] Z Suo A Cox N Bartelli et al ldquoGRK5 deficiency leads to earlyAlzheimer-like pathology and working memory impairmentrdquoNeurobiology of Aging vol 28 no 12 pp 1873ndash1888 2007
[29] W Kamphuis C Mamber MMoeton et al ldquoGFAP isoforms inadult mouse brain with a focus on neurogenic astrocytes andreactive astrogliosis in mouse models of Alzheimer diseaserdquoPLoS ONE vol 7 no 8 Article ID e42823 2012
[30] J Lang C Vigo-Pelfrey and F Martin ldquoLiposomes composedof partially hydrogenated egg phosphatidylcholines fatty acidcomposition thermal phase behavior and oxidative stabilityrdquoChemistry and Physics of Lipids vol 53 no 1 pp 91ndash101 1990
[31] D Morgan D M Diamond P E Gottschall et al ldquoA 120573peptide vaccination prevents memory loss in an animal modelof Alzheimerrsquos diseaserdquo Nature vol 408 no 6815 pp 982ndash9852000
[32] M L Brownlow L Benner D DrsquoAgostino M N Gordon andD Morgan ldquoKetogenic diet improves motor performance but
not cognition in two mouse models of Alzheimerrsquos pathologyrdquoPLoS ONE vol 8 no 9 Article ID e75713 2013
[33] M Pride P Seubert M Grundman M Hagen J Eldridgeand R S Black ldquoProgress in the active immunotherapeu-tic approach to Alzheimerrsquos disease clinical investigationsinto AN1792-associated meningoencephalitisrdquo Neurodegenera-tive Diseases vol 5 no 3-4 pp 194ndash196 2008
[34] CWiessner K-HWiederhold A C Tissot et al ldquoThe second-generation activeA120573 immunotherapyCAD106 reduces amyloidaccumulation in APP transgenic mice while minimizing poten-tial side effectsrdquoThe Journal of Neuroscience vol 31 no 25 pp9323ndash9331 2011
[35] D Lambracht-Washington and R Rosenberg ldquoAdvances in thedevelopment of vaccines for Alzheimerrsquos diseaserdquo DiscoveryMedicine vol 15 no 84 pp 319ndash326 2013
[36] A Kimura T Ohmori R Ohkawa et al ldquoEssential roles ofsphingosine 1-phosphateS1P
1receptor axis in the migration of
neural stem cells toward a site of spinal cord injuryrdquo Stem Cellsvol 25 no 1 pp 115ndash124 2007
[37] S S Lee C H Woo J D Chang and J H Kim ldquoRoles of Racand cytosolic phospholipase A2 in the intracellular signalling inresponse to titanium particlesrdquo Cellular Signalling vol 15 no 3pp 339ndash345 2003
[38] E Masliah L Hansen A Adame et al ldquoA120573 vaccination effectson plaque pathology in the absence of encephalitis inAlzheimerdiseaserdquo Neurology vol 64 no 1 pp 129ndash131 2005
[39] A Pilorget B Annabi F Bouzeghrane J Marvaldi J Luisand R Beliveau ldquoInhibition of angiogenic properties of brainendothelial cells by platelet-derived sphingosine-1-phosphaterdquoJournal of Cerebral Blood Flow and Metabolism vol 25 no 9pp 1171ndash1182 2005
[40] A Wang P Das R C Switzer III T E Golde and J LJankowsky ldquoRobust amyloid clearance in a mouse model ofAlzheimerrsquos disease provides novel insights into themechanismof amyloid-120573 immunotherapyrdquo Journal of Neuroscience vol 31no 11 pp 4124ndash4136 2011
[41] G Krabbe A Halle V Matyash et al ldquoFunctional impairmentof microglia coincides with Beta-amyloid deposition in micewith Alzheimer-like pathologyrdquo PLoS ONE vol 8 no 4 ArticleID e60921 2013
[42] Y H Liu B Giunta H-D Zhou J Tan and Y J WangldquoImmunotherapy for Alzheimer disease-the challenge ofadverse effectsrdquo Nature Reviews Neurology vol 8 no 8 pp465ndash469 2012
[43] B Solomon R Koppel D Frankel and E Hanan-AharonldquoDisaggregation of Alzheimer120573-amyloid by site-directedmAbrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 94 no 8 pp 4109ndash4112 1997
[44] F Bard C Cannon R Barbour et al ldquoPeripherally adminis-tered antibodies against amyloid 120573-peptide enter the centralnervous system and reduce pathology in a mouse model ofAlzheimer diseaserdquo Nature Medicine vol 6 no 8 pp 916ndash9192000
[45] R B DeMattos K R Bales D J Cummins J C Dodart S MPaul and D M Holtzman ldquoPeripheral anti-A120573 antibody altersCNS and plasma A120573 clearance and decreases brain A120573 burdenin a mouse model of Alzheimerrsquos diseaserdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 98 no 15 pp 8850ndash8855 2001
[46] C A Lemere E T Spooner J LaFrancois et al ldquoEvidence forperipheral clearance of cerebral A120573 protein following chronic
16 BioMed Research International
active A120573 immunization in PSAPP micerdquo Neurobiology ofDisease vol 14 no 1 pp 10ndash18 2003
[47] T Town M Vendrame A Patel et al ldquoReduced Th1 andenhanced Th2 immunity after immunization with Alzheimerrsquos120573-amyloid
1minus42rdquo Journal of Neuroimmunology vol 132 no 1-2
pp 49ndash59 2002[48] R Cacabelos ldquoPharmacogenomics in Alzheimerrsquos diseaserdquo
Methods in Molecular Biology vol 448 pp 213ndash357 2008[49] R Cacabelos ldquoPharmacogenomics and therapeutic strategies
for dementiardquo Expert Review of Molecular Diagnostics vol 9no 6 pp 567ndash611 2009
[50] D van Dam R DrsquoHooge M Staufenbiel C van Ginneken Fvan Meir and P P de Deyn ldquoAge-dependent cognitive declinein the APP23 model precedes amyloid depositionrdquo EuropeanJournal of Neuroscience vol 17 no 2 pp 388ndash396 2003
[51] M Dumont C Strazielle M Staufenbiel and R LalondeldquoSpatial learning and exploration of environmental stimuli in24-month-old female APP23 transgenic mice with the Swedishmutationrdquo Brain Research vol 1024 no 1-2 pp 113ndash121 2004
[52] R Lalonde M Dumont M Staufenbiel C Sturchler-Pierratand C Strazielle ldquoSpatial learning exploration anxiety andmotor coordination in female APP23 transgenic mice with theSwedish mutationrdquo Brain Research vol 956 no 1 pp 36ndash442002
[53] F J Huff F Boller F Lucchelli R Querriera J Beyer and SBelle ldquoThe neurologic examination in patients with probableAlzheimerrsquos diseaserdquo Archives of Neurology vol 44 no 9 pp929ndash932 1987
[54] A F Pettersson M Engardt and L O Wahlund ldquoActivitylevel and balance in subjects with mild Alzheimerrsquos diseaserdquoDementia and Geriatric Cognitive Disorders vol 13 no 4 pp213ndash216 2002
[55] L Calandreau B Desgranges R Jaffard and A DesmedtldquoSwitching from contextual to tone fear conditioning and viceversa the key role of the glutamatergic hippocampal-lateralseptal neurotransmissionrdquo Learning amp Memory vol 17 no 9pp 440ndash443 2010
[56] F Trinchese S Liu F Battaglia SWalter PMMathews andOArancio ldquoProgressive age-related development of Alzheimer-like pathology in APPPS1 micerdquo Annals of Neurology vol 55no 6 pp 801ndash814 2004
[57] D M Wilcock A Rojiani A Rosenthal et al ldquoPassiveimmunotherapy against A120573 in aged APP-transgenic micereverses cognitive deficits and depletes parenchymal amyloiddeposits in spite of increased vascular amyloid and microhem-orrhagerdquo Journal of Neuroinflammation vol 1 article 24 2004
[58] S A Ferguson S Sarkar and L C Schmued ldquoLongitudinalbehavioral changes in the APPPS1 transgenic AlzheimerrsquosDisease modelrdquo Behavioural Brain Research vol 242 no 1 pp125ndash134 2013
[59] D Wilcock and C Colton ldquoImmunotherapy vascular pathol-ogy and microhemorrhages in transgenic micerdquo CNS andNeurological DisordersmdashDrug Targets vol 8 no 1 pp 50ndash642009
[60] M Grundman S Gilman R S Black et al ldquoAn alternativemethod for estimating efficacy of the AN1792 vaccine foralzheimer diseaserdquo Neurology vol 71 no 9 pp 697ndash698 2008
[61] D Boche E Zotova R O Weller et al ldquoConsequence of A120573immunization on the vasculature of human Alzheimerrsquos diseasebrainrdquo Brain vol 131 no 12 pp 3299ndash3310 2008
[62] M Pfeifer S Boncristiano L Bondolfi et al ldquoCerebral hemor-rhage after passive anti-a120573 immunotherapyrdquo Science vol 298no 5597 p 1379 2002
[63] M M Racke L I Boone D L Hepburn et al ldquoExacerbationof cerebral amyloid angiopathy-associatedmicrohemorrhage inamyloid precursor protein transgenic mice by immunotherapyis dependent on antibody recognition of deposited forms ofamyloid 120573rdquo Journal of Neuroscience vol 25 no 3 pp 629ndash6362005
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

BioMed Research International 9
Gr E
(PBS
)
(d)
(e)
(i)
(j)
(n)
(o)
(s)
(t)
Cx
GFAP
GFAP GFAP GFAP GFAP
GFAP GFAP GFAP
Cx Cx
Cx
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
PoDGDG
MoDG
Cx
CA1
GFAP
Gr A
(EB1
01)
(a)
(b)
(c)
(f)
(g)
(h)
Preventive treatment Therapeutic treatment
PoDG
MoDG
(k)
(l)
(m)
(p)
(q)
(r)DG
GFAP GFAP GFAP
GFAPGFAPGFAP GFAP
GFAP GFAP GFAP GFAP
PoDGDG
DG
CA1
Cx
Cx
Cx
Cx
Cx
Cx
PoDG
MoDG
DG
CA1
MoDGDG
CA1
PoDG
MoDG
DG
CA1
Cx
CxCx
MoDG
MoDG
PoDG
CA1CA1
Cx
Gr D
(A120573+
FA)
Gr C
(S1P
+L)
Gr B
(A120573+
L)
Figure 5 Comparative effect of EB101 vaccine on astrocytosis Comparative photomicrographs of GFAP immunoreactivity in thehippocampus ((a)ndash(e) (k)ndash(o)) and cortical ((f)ndash(j) (p)ndash(t)) brain regions at the preventive treatment phase ((a)ndash(j)) and therapeutictreatment group ((k)ndash(t)) Preventive treatment transverse brain sections of 11-month-old mice treated with EB101 show an almost completeprevention of astrocytosis in the detailed sections of the dentate gyrus (a) contrasting with numerous dystrophic reactive astrocytes typicalof an inflammatory reaction observed in different hippocampal corresponding areas of mouse brains in Group B (A120573Lip-treated mice (b))Group C (LipS1P-treated mice (c)) Group D (A120573
42+ CFAIFA-treated mice (d)) and Group E (PBS-treated mice (e)) Immunoreactive
astrocytosis in the brain sections of WT mice is absent (data not shown) Detailed sections of the parietal cortex of treated mice show agradient density of reactive astrocytes immunoreactive to GFAP indicating a neuroinflammation pattern Comparative cortical sections shownumerous astrocytosis areas inmice Groups C and E ((h) (j)) andwide reactive astrocytes clusters inmice Groups B andD ((g) (i)) while justa few of them are observed in Group A (f)Therapeutic treatment transverse brain sections of 18-month-old mice of Group A show a notablereduction of astrocytosis in the hippocampal region (k) after EB101 vaccine immunization Note the astrocytosis contrast density betweenEB101-treatedmice and the other treated groups ((l)ndash(o)) where numerous immunoreactive GFAP clusters are observed in the correspondentbrain sections A few immunoreactive GFAP clusters are observed in the parietal cortex of EB101 treated mice group while a moderate ((q)(s)) to high ((r) (t)) density of these neuropathological inflammation clusters is apparent in the other treatment groups (Groups BndashE) Forabbreviations see list Scale bar 100120583m
10 BioMed Research International
002040608
1121416
Preventive treatmentTherapeutic treatment
Ant
i-120573-a
myl
oid
IgG
antib
odie
s
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Gr A
(A120573+
S1P+
L)(E
B101
)
Figure 6 Quantification of specific anti-120573-amyloid IgG in immu-nized APPPS1 mice The presence of anti-120573-amyloid antibodiesmeasured in the sera of vaccinated and control mice groups duringthe preventive and therapeutic treatments indicates that EB101liposome vaccine was able to induce the highest production ofspecific IgG antibodies Compared to theGroupD immunizedmicethe level of antibodies observed in the Group A was also highersuggesting a more effective immune response (119875 lt 001 versusGroup D 119875 lt 0001 versus Groups B C and E) Quantification ofspecific anti-120573-amyloid IgG in immunized Tg mice
in Group D with respect to all other groups mainlyreported by the high levels of TNF-120572 (Figure 7(b)) andIFN-120574 (Figure 7(c)) cytokine secretions while due to thehigh mortality rate during the therapeutic treatment nosignificant data was obtained from this mice group Inaddition a tendency toTh1 secretion reduction was observedinGroupAwith respect to all other groups in both preventiveand therapeutic treatment The changes in Th2 protein levels(Figure 7(d)) followed the inverse trend where the highestdifferences between Group A and the other treatment groupswere observed in the IL-4 IL-5 and IL-10 cytokine secretions(119875 lt 005 versus Group D 119875 lt 001 versus Group E Figures7(e)ndash7(g))
35 Comparative Motor Coordination and Memory Perfor-mances In order to validate the locomotion integrity duringthe entire immunotherapeutic process as reported in similarprevious studies motor coordination tests have showed thatEB101-treated mice spent significantly more time on theaccelerating rotarodThismotor coordination task shows thatEB101 immunization affected positively their performance(Figure 8(a)) A120573
42liposome (B) S1P-containing liposomes
(C) and PBS (E) (119875 lt 0006) mice performed poorly incomparison to EB101 mice (119875 lt 005) or A120573
42+ CFAIFA
mice (119875 lt 002) in motor abilities and coordination Thefear conditioning-active avoidance test was used to assess thespatial learning and memory functions As expected bothEB101 and A120573
42+ CFAIFA immunized mice readily learned
the location of the platform as shown by a decrease in theescape latency over training days while the control group
mice exhibited no significant changes during 6-day trialsThe average escape latencies of EB101 (68 plusmn 2 s 761 plusmn 22 s)andA120573
42+CFAIFA (116plusmn33 s 123plusmn21 s) immunizedmice
at the end of preventive and therapeutic periods respectivelyduring the consecutive 6 days trial were significantly shorterthan the other treatment groups (A120573
42liposome 163 plusmn 31 s
175 plusmn 28 s S1P-containing liposomes 202 plusmn 19 s 248 plusmn27 s) and the control (2116 plusmn 38 s 268 plusmn 22 s) (119875 lt 001)Therewas no significant difference between EB101 andA120573
42+
CFAIFA immunized group (119875 gt 005) (Figure 8(b)) Theseresults showed that spatial learning was improved in theseimmunized mice
4 Discussion
In this study we have compared a novel active A120573 imm-unotherapy the EB101 vaccine against A120573 with the A120573
42+
CFAIFA vaccine that was first taken into clinical trialswith coadjuvant Qs-21 modification (AN1792 by Orgogozoet al [19]) This type of vaccine design was thought toinduce a strong T-cell mediated inflammatory response (IL-2 and IFN-c positive responses indicative of a Class II(CD4) Th-1 type response [33] leading to acute menin-goencephalitis in the 6 of the patients) The EB101 activeimmunotherapeutic vaccine was primarily designed to avoidundesired inflammatory side effects both in mice andhumans (without need of coadjuvant modification) whileinducing effective antibody titer rates We demonstrated thatlong-term immunization with EB101 induced A120573 antibodiesin the preventive and the therapeutic treatment periodswhich effectively reduced amyloid deposition in both corticaland hippocampal brain regions and prevented neuroinflam-mation probably by blocking toxicity in the surroundingneurons However the AN1792 vaccine was shown to inducein 6 of the patients an inflammatory reaction that mighthave led to the encephalitis type reaction observed in theclinical trial which was responsible for the immediate haltin those clinical studies It seems that AN1792 immunizationdelivered in Qs-21 and polysorbate 80 adjuvant induced aTh1 type response that likely led to autoaggressive T cells andaseptic meningoencephalitis In the present experiments wedemonstrated that a judiciously selected adjuvant produces adifferent humoral response [22] Indeed we have shown thatour novel immunotherapeutic strategy boosts specific A120573
1ndash42antibody generation and induces a Th2 noninflammatoryimmune response This boosted effect is in accordance withprevious immunotherapeutic studies [34] Taking togetherall the present findings these results suggest that EB101vaccine can achieve in a future clinical trial immunogenicresponse rates quite similar to the AN1792 without theinflammatory effects reported The immunohistochemicalanalysis obtained in the present study shows that all the EB101immunized mice resulted in an almost complete preventionof A120573 deposition in the brain with only a few compactA120573 plaques in the retrosplenial and dentate gyrus regionsAlthough there was a slightly increase in death in EB101treated mice compared to controls this significant reductionof A120573 plaques in EB101 treated mice was proven to be more
BioMed Research International 11
Gr A Gr B Gr C Gr D Gr EPreventive treatmentTherapeutic treatment
0005001000150020002500300035004000
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
(a) Th1
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
000
1000
2000
3000
4000
5000TNF-120572
(b) Th1
000500
10001500200025003000
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
IFN-120574
(c) Th1
000500
100015002000250030003500
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
Group A Group B Group C Group D Group EPreventive treatmentTherapeutic treatment
(d) Th2
000
500
1000
1500
2000
2500
3000IL-4
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(e) Th2
000500
100015002000250030003500
IL-5
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(f) Th2
3500IL-10
000500
10001500200025003000
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(g) Th2
Figure 7 Detection of Th1 and Th2 cytokine profile Measurements of the effect of EB101 and A12057342
+ CFAIFA vaccines on Th1 and Th2cytokines profile during preventive and therapeutic treatments ((a)ndash(c)) A significant increase of Th1 cytokine profile was observed inpreventive treatment in Group D with respect to all other groups while a tendency to a reduction inTh1 secretion was observed in Group Awith respect to all other groups in both preventive and therapeutic treatment ((d)ndash(g)) The changes in Th2 protein levels were observed inthe IL-4 IL-5 and IL-10 cytokine secretions of Group A (119875 lt 005 versus Group D 119875 lt 001 versus Group E)
12 BioMed Research International
Gr A
(EB1
01)
05
101520253035
Preventive treatment Therapeutic treatment
Mea
n fa
lling
late
ncy
(s)
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)(a) Graphic of motor coordination and balance
05
1015202530
Mea
n re
spon
se ti
me (
s)
Gr A
(EB1
01)
Preventive treatment Therapeutic treatment
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
(b) Graphic of associative learning and memory
Figure 8 Analysis of motor coordination and balance (a) Rotarod results of motor coordination and balance test after the preventive andtherapeutic treatment showing improved effect of EB101 and A120573
42+ CFAIFA-vaccine compared to other treated mice groups (119875 lt 005)
and (119875 lt 002) versus control mice Latency to fall on the rotarod apparatus was significantly improved in EB101 immunized mice Valuesrepresent means of all six trials of a groupday (b) Spatial learning and memory tested by fear conditioning-active avoidance test The meanresponse time of each treatment group to reach the platform is represented EB101 and A120573
42+ CFAIFA immunized mice had significantly
lower latencies than the other treatment groups (119875 lt 001) versus control mice
efficient than that obtained with A12057342+ CFAIFA immuniza-
tions A comparison of A120573 plaque reduction rates amongtreated groups surprisingly showed that other than EB101and A120573
42+ CFAIFA treated groups some amyloid deposits
were also reduced in Groups B (A1205731ndash42liposome) and C
(S1P-containing liposomes) Although the reduction rates inthese mice were markedly low the subtle lowering effectmight be due to the interaction between the immunizationelements (A120573
1ndash42 and S1P) and the mouse immunogenicsystem as reported by others [35] Based on our presentresults S1P one of the components of the adjuvant used withEB101 might be responsible for the neuroprotective effectobserved in the vulnerable neuronal areas of EB101 treatedmice The advantage in the A120573 load preventionreduction bycombining A120573
1ndash42 and S1P in a unique formulation vaccinehas been also observed in the reduced hippocampal load ofEB101 immunizedmice (Gr A) when comparedwith differentvaccine components (Gr B-C) In fact S1P appears to have ademonstrated therapeutic potential as a regenerative agent inthe nervous system since it controls migration of neuronalstem cells toward a site of spinal cord injury [36] inducecytoskeletal rearrangements to promote transmitter secretion[37] and also trigger diverse cellular effects including angio-genesis cardiac development immunity cell motility andneurite extension [38 39]Therefore it is reasonable to expectthat such a potent biologically active sphingolipid by itselfmay play an important role in the A120573 plaque reduction Inthis study the clearance efficiency rate of each treatment wasanalyzed and results suggested that EB101 (6459) appearsto be more effective than A120573
42+ CFAIFA (1920) vaccine
in both preventive and therapeutic groups The effectivenessof EB101 in older transgenic animals (18 months) showeda clearance effect of 59 suggesting that EB101 may alsohave therapeutic potential inADdiagnosticated patient trials
Similar conclusion was reached byWang and colleagues [40]who demonstrated that combination therapy in aged animalsprovides more valuable insights into a possible translationto humansOur immunologicalmeasurements demonstratedonce again that neuroinflammation develops in an A120573-dependent fashion early in the course of AD [41] and canbe alleviated or reversed by interventional A120573 vaccinationsuch as EB101 Among the possible mechanisms of A120573clearance proposed in AD immunotherapy [42] our resultssuggested that theymay overlap and probably be disease statedependent since the prevention and the therapeutic vaccinewould likely benefit from the transport of A120573 antibodies intothe CNS However some mechanisms of action such as thepreventive effect of A120573 antibodies in the A120573 aggregation [43]microglial phagocytosis induced by the Fc binding portionsbetween A120573 antibodies and microglia receptor [44] and theshift in the gradient of A120573 transport across the blood-brainbarrier due to the increased concentration of A120573 antibodiesin the vascular system resulting in an increase in effluxfrom the brain to blood [45 46] may be playing a role inboth preventive and therapeutic conditions Our findingstherefore suggest that novel treatment strategies aimed atmaintaining or increasing microglial function and restoringthe balance to the immunological system may represent anattractive therapeutic approach even at the advanced stagesof ADThe immunization results obtained with EB101 duringboth preventive and therapeutic treatment showed nomicro-hemorrhages infiltration of lymphocytes or encephalitis-like reaction in the brain suggesting that EB101 does notinduce A120573-specific Th1 reaction possibly due to the neu-roprotective effect of S1P Delivering A120573
1ndash42 in a liposomal-S1P-adjuvant resulted in an effective immunological responseincluding the induction of a Th2 T-cell response whicheffectively prevented neuroinflammation in an animal model
BioMed Research International 13
of AD as reported previously [24 47] Moreover presentresults demonstrated that EB101 did not induce astrogliosisin APPPS1 transgenic mice whereas some GFAP-positiveastrocyte clusters were aggregated in the affected brain areasof mice treated with A120573
42+ CFAIFA These results indicate
that EB101 may result in an effective treatment in AD neu-ropathology due to the absence of peripheral proinflamma-tory cytokine secretion and the absence of astroglial activa-tion in the brains of EB101 treated mice when compared withA12057342
+ CFAIFA immunotherapeutic response In a clinicalsetting however we may expect a wide differential responsein part depending upon the genomic profile of each patient[48 49] In AD behavioural studies in particular amongthe motor coordination tests the rotarod performance hasbeen reported to be impaired in some AD transgenic micemodels bearing A120573 plaques [50 51] This result was found inmutants prior to A120573 plaque formation at 3 and 6 months ofage but was not reproduced after plaques had been formedat 16 and 24 months of age [52] perhaps because of thedeclining abilities of older normal mice Here we presentthe effects of vaccine treatments in APPPS1 mice on motorcoordination a neglected aspect of the AD experimentalbehavior but justified on the basis of the deficient posturalcontrol reported in patients with Alzheimerrsquos disease [53]sometimes even during early stages [54] Our longitudinalmotor behavioral results showed no significant differencesin the ambulatory movement grip strength or coordinationaddressed by their performance in the rotating rod betweenAPPPS1 and wild-type mice for any age group studiedin contrast with other treated groups demonstrating thatlongitudinal motor assessments of AD transgenic mousemodels can provide valuable information with regard tothe effect of immunization on the progression of behavioraldeficits and overall motor activity As shown the EB101-treated group of mice exhibited longer latencies and betterbalancing skills indicating the absence of any negative effectof EB101 immunization on these mice The classical fearconditioning test is assumed to reveal the functional integrityof the hippocampus [55] representing a special memorytask to evaluate the AD models and its improvement afterimmunization periods In this APPPS1 mouse model thefirst associative learning and spatial workingmemory deficitsare shown at 5-6 months of age [56] although its impair-ment progression is relatively slow compared to most othermodels At the end of preventive immunization period EB101and A120573
42+ CFAIFA immunized mice have significantly
shorter escape latencies than the other treatment groups(A12057342liposome S1P-containing liposomes) and the controls
in the fear conditioning test Shorter escape latencies duringspatial learning indicated that deficits of spatial memorywere attenuated in EB101 and A120573
42+ CFAIFA immunized
mice Previous studies have showed that by 15 months ofage the spatial working memory is consistently impairedthroughout the rest of the life span [31 57] At the end oftherapeutic immunization period both EB101 and A120573
42+
CFAIFA immunized mice showed significantly improvedresults on the hippocampus-dependent spatial learning andmemory test when compared with other treatment groupsand the controls Taken together these observations are
in agreement with the impairment in conditioned learningin response to a tone stimulus of aged APPPS1 micealthough when immunized with EB101 during preventiveor therapeutic periods such impairments are significantlyattenuated
In summary the present results corroborate our previouspublished findings showing that EB101 immunization reducesamyloid accumulation in the APPPS1 mouse brain [2425] The strongest reduction in the preventive immunizationwas observed in the retrosplenial cortex followed by thehippocampus and entorhinal-piriform cortex Our resultsfurther support that EB101 ismore effectivewhen themice areimmunized prior to developing plaques in the hippocampaland cortical brain regions There is extensive evidence inmouse preclinical studies [58] and human clinical trials [5960] that the adverse effects of active and passive A120573 vaccinesresult in microhemorrhages and encephalitic reactions [60ndash63] yet themechanismbywhich they occur has not been fullyelucidated In the treatment sections analyzed we found nomicrohemorrhages or acute inflammatory processes in all thesurvived mice of the group indicating that EB101 therapeuticapproach presents no adverse inflammatory effects in thisAD mice models Thus our present results unequivocallydemonstrate that EB101 vaccinemay not only halt AD-relatedpathology as a preventive treatment but may also reverse it inestablished cases related to the improvement of the learningandmemory in both cases leading to a robust immunologicaltherapeutic response Present experiments also demonstratethat EB101 exerts more significant and substantial effects onthe reduction of A120573 deposition than A120573
42+CFAIFA vaccine
in A120573 mice models of AD However further exploration ofthe contribution of genetic environmental and physiologicalfactors in the development resistance to endogenous A120573-antibody is needed in order to optimize and modulate thetherapeutic strategies currently being developed
5 Conclusions
The present study carried out in APPPS1 transgenic mousemodel of AD compared the A120573 burden reduction andcellular immune responses produced by two immunizationdelivery strategies of A120573 in Freund adjuvant (described byWebster et al [10]) and in liposomeS1P matrix (describedby Schenk et al [17])The EB101 immunotherapeutic strategyboosted antibody generation A120573 plaque reduction and ananti-inflammatory response more effectively than A120573
42+
CFAIFA especially if given prior to or in the early stages ofpathology The present results also showed that vaccinationwith EB101 prevented the formation of new plaques andpresumably led to a reduction in established plaques whileinducing aTh2 immune reactionwithout encephalitogenic T-cell responses or brain inflammation related with astrogliosisHowever despite the efficacy demonstrated of this novelvaccine approach further preclinical and clinical studieson EB101 are warranted to evaluate its efficacy and themechanism for the future utility of such vaccine for ADtherapy
14 BioMed Research International
List of Abbreviations
AD Alzheimerrsquos diseaseA120573 120573-Amyloid proteinAPP Amyloid precursor proteinAPT Anterior pretectal nucleusCA1 Field CA1 hippocampusCA3 Field CA3 hippocampusCPu Caudate putamen (striatum)Cx CortexDG Dentate gyrusDN Dystrophic neuriteEct Entorhinal cortexCFA Complete Freundrsquos adjuvantGrDG Granular dentate gyrusIFA Incomplete Freundrsquos adjuvantIL InterleukinLPMR Lateral posterior thalamic nucleus (mediorostral)MoDG Molecular dentate gyrusPir Piriform cortexPoDG Polymorph dentate gyrusRSGb Retrosplenial granular cortex b regionTg TransgenicTh T helper
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Ivan Carrera Ignacio Etcheverrıa Lucıa Fernandez-NovoaValter Lombardi Ramon Cacabelos and Carmen Vigo con-ceived and designed the experiments Ivan Carrera and Igna-cio Etcheverrıa performed the experiments Ivan CarreraIgnacio Etcheverrıa and Valter Lombardi analyzed the dataIvan Carrera Ignacio Etcheverrıa Lucıa Fernandez-Novoaand Valter Lombardi contributed reagentsmaterialsanalysistools Ivan Carrera Carmen Vigo andMadepalli KrishnappaLakshmana wrote the paper
Acknowledgments
This work was supported by National Institute of Aging(NIA)NIH Grant no 1R01AG036859-01 (Madepalli Krish-nappa Lakshmana) and the financial support of the ProgramaOperativo Feder Galicia 2007ndash2013 (IN841C 201254 to PhDCarrera I)
References
[1] R Cacabelos L Fernandez-Novoa V Lombardi Y Kubotaand M Takeda ldquoMolecular genetics of Alzheimerrsquos diseaseand agingrdquoMethods and Findings in Experimental and ClinicalPharmacology vol 27 pp 1ndash573 2005
[2] P T Nelson I Alafuzoff E H Bigio et al ldquoCorrelationof alzheimer disease neuropathologic changes with cognitive
status a review of the literaturerdquo Journal of Neuropathology andExperimental Neurology vol 71 no 5 pp 362ndash381 2012
[3] JNaslundVHaroutunian RMohs et al ldquoCorrelation betweenelevated levels of amyloid 120573-peptide in the brain and cognitivedeclinerdquo Journal of the American Medical Association vol 283no 12 pp 1571ndash1577 2000
[4] A Koyama O I Okereke T Yang D Blacker D J Selkoeand F Grodstein ldquoPlasma amyloid-120573 as a predictor of dementiaand cognitive decline a systematic review and meta-analysisrdquoArchives of Neurology vol 69 no 7 pp 824ndash831 2012
[5] A M Fagan C M Roe C Xiong M A Mintun J C Morrisand D M Holtzman ldquoCerebrospinal fluid tau120573-amyloid42ratio as a prediction of cognitive decline in nondemented olderadultsrdquo Archives of Neurology vol 64 no 3 pp 343ndash349 2007
[6] S Parvathy P Davies V Haroutunian et al ldquoCorrelationbetween A120573-40- A120573-42- and A120573x-43-containing amyloidplaques and cognitive declinerdquo Archives of Neurology vol 58no 12 pp 2025ndash2032 2001
[7] E Karran M Mercken and B D Strooper ldquoThe amyloidcascade hypothesis for Alzheimerrsquos disease an appraisal for thedevelopment of therapeuticsrdquo Nature Reviews Drug Discoveryvol 10 no 9 pp 698ndash712 2011
[8] J L Jankowsky D J Fadale J Anderson et al ldquoMutantpresenilins specifically elevate the levels of the 42 residue 120573-amyloid peptide in vivo evidence for augmentation of a 42-specific 120574 secretaserdquo Human Molecular Genetics vol 13 no 2pp 159ndash170 2004
[9] L Holcomb M N Gordon E Mcgowan et al ldquoAcceleratedAlzheimer-type phenotype in transgenic mice carrying bothmutant amyloid precursor protein and presenilin 1 transgenesrdquoNature Medicine vol 4 no 1 pp 97ndash100 1998
[10] S JWebster A D Bachstetter and L J van Eldik ldquoComprehen-sive behavioral characterization of an APPPS-1 double knock-in mouse model of Alzheimerrsquos diseaserdquo Alzheimerrsquos ResearchandTherapy vol 5 no 3 article 28 2013
[11] H Xiong D Callaghan J Wodzinska et al ldquoBiochemical andbehavioral characterization of the double transgenic mousemodel (APPswePS1dE9) of Alzheimerrsquos diseaserdquo NeuroscienceBulletin vol 27 no 4 pp 221ndash232 2011
[12] M Garcia-Alloza L A Borrelli B T Hyman and B J BacskaildquoAntioxidants have a rapid and long-lasting effect on neuriticabnormalities in APPPS1 micerdquo Neurobiology of Aging vol 31no 12 pp 2058ndash2068 2010
[13] M Meyer-Luehmann M Mielke T L Spires-Jones et al ldquoAreporter of local dendritic translocation shows plaque-relatedloss of neural system function in APP-transgenic micerdquo Journalof Neuroscience vol 29 no 40 pp 12636ndash12640 2009
[14] E Machova V Rudajev H Smyckova H Koivisto H Tanilaand V Dolezal ldquoFunctional cholinergic damage develops withamyloid accumulation in young adult APPswePS1dE9 trans-genic micerdquo Neurobiology of Disease vol 38 no 1 pp 27ndash352010
[15] T Malm J Koistinaho and K Kanninen ldquoUtilization of APP-swePS1dE9 transgenic mice in research of Alzheimerrsquos diseasefocus on gene therapy and cell-based therapy applicationsrdquoInternational Journal of Alzheimerrsquos Disease vol 2011 Article ID517160 8 pages 2011
[16] J Lee and P Han ldquoAn update of animal models of alzheimerdisease with a reevaluation of plaque depositionsrdquoExperimentalNeurobiology vol 22 pp 84ndash95 2013
BioMed Research International 15
[17] D Schenk R Barbour W Dunn et al ldquoImmunization withamyloid-120573 attenuates Alzheimer disease-like pathology in thePDAPP mouserdquo Nature vol 400 no 6740 pp 173ndash177 1999
[18] H J Fu B Liu J L Frost and C A Lemere ldquoAmyloid-120573immunotherapy for Alzheimerrsquos diseaserdquoCNS and NeurologicalDisorders-Drug Targets vol 9 no 2 pp 197ndash206 2010
[19] J-M Orgogozo S Gilman J-F Dartigues et al ldquoSubacutemeningoencephalitis in a subset of patients with AD after A120573
42
immunizationrdquo Neurology vol 61 no 1 pp 46ndash54 2003[20] J A R Nicolll DWilkinson CHolmes P Steart HMarkham
andROWeller ldquoNeuropathology of humanAlzheimer diseaseafter immunization with amyloid-120573 peptide a case reportrdquoNature Medicine vol 9 no 4 pp 448ndash452 2003
[21] I Ferrer M Boada Rovira M L Sanchez Guerra M JRey and F Costa-Jussa ldquoNeuropathology and pathogenesisof encephalitis following amyloid-Neuropathology and patho-genesis of encephalitis following amyloid-beta immunizationin Alzheimerrsquos disease immunization in Alzheimerrsquos diseaserdquoBrain Pathology vol 14 no 1 pp 11ndash20 2004
[22] D H Cribbs A Ghochikyan V Vasilevko et al ldquoAdjuvant-dependent modulation ofTh1 andTh2 responses to immuniza-tion with 120573-amyloidrdquo International Immunology vol 15 no 4pp 505ndash514 2003
[23] C Holmes D Boche D Wilkinson et al ldquoLong-term effectsof A120573
42immunisation in Alzheimerrsquos disease follow-up of a
randomised placebo-controlled phase I trialrdquo The Lancet vol372 no 9634 pp 216ndash223 2008
[24] I Carrera I Etcheverrıa L Fernandez-Novoa V LombardiR Cacabelos and C Vigo ldquoVaccine development to treatAlzheimerrsquos disease neuropathology in APPPS1 transgenicmicerdquo International Journal of Alzheimerrsquos Disease vol 2012Article ID 376138 17 pages 2012
[25] I Carrera I Etcheverrıa Y Li et al ldquoImmunocytochemicalcharacterization of Alzheimer disease hallmarks in APPPS1transgenic mice treated with a new anti-amyloid-120573 vaccinerdquoBioMed Research International vol 2013 Article ID 709145 12pages 2013
[26] V Anelli R Bassi G Tettamanti P Viani and L Riboni ldquoExtra-cellular release of newly synthesized sphingosine-1-phosphateby cerebellar granule cells and astrocytesrdquo Journal of Neuro-chemistry vol 92 no 5 pp 1204ndash1215 2005
[27] S Schroeter K Khan R Barbour et al ldquoImmunotherapyreduces vascular amyloid-120573 in PDAPP micerdquo Journal of Neu-roscience vol 28 no 27 pp 6787ndash6793 2008
[28] Z Suo A Cox N Bartelli et al ldquoGRK5 deficiency leads to earlyAlzheimer-like pathology and working memory impairmentrdquoNeurobiology of Aging vol 28 no 12 pp 1873ndash1888 2007
[29] W Kamphuis C Mamber MMoeton et al ldquoGFAP isoforms inadult mouse brain with a focus on neurogenic astrocytes andreactive astrogliosis in mouse models of Alzheimer diseaserdquoPLoS ONE vol 7 no 8 Article ID e42823 2012
[30] J Lang C Vigo-Pelfrey and F Martin ldquoLiposomes composedof partially hydrogenated egg phosphatidylcholines fatty acidcomposition thermal phase behavior and oxidative stabilityrdquoChemistry and Physics of Lipids vol 53 no 1 pp 91ndash101 1990
[31] D Morgan D M Diamond P E Gottschall et al ldquoA 120573peptide vaccination prevents memory loss in an animal modelof Alzheimerrsquos diseaserdquo Nature vol 408 no 6815 pp 982ndash9852000
[32] M L Brownlow L Benner D DrsquoAgostino M N Gordon andD Morgan ldquoKetogenic diet improves motor performance but
not cognition in two mouse models of Alzheimerrsquos pathologyrdquoPLoS ONE vol 8 no 9 Article ID e75713 2013
[33] M Pride P Seubert M Grundman M Hagen J Eldridgeand R S Black ldquoProgress in the active immunotherapeu-tic approach to Alzheimerrsquos disease clinical investigationsinto AN1792-associated meningoencephalitisrdquo Neurodegenera-tive Diseases vol 5 no 3-4 pp 194ndash196 2008
[34] CWiessner K-HWiederhold A C Tissot et al ldquoThe second-generation activeA120573 immunotherapyCAD106 reduces amyloidaccumulation in APP transgenic mice while minimizing poten-tial side effectsrdquoThe Journal of Neuroscience vol 31 no 25 pp9323ndash9331 2011
[35] D Lambracht-Washington and R Rosenberg ldquoAdvances in thedevelopment of vaccines for Alzheimerrsquos diseaserdquo DiscoveryMedicine vol 15 no 84 pp 319ndash326 2013
[36] A Kimura T Ohmori R Ohkawa et al ldquoEssential roles ofsphingosine 1-phosphateS1P
1receptor axis in the migration of
neural stem cells toward a site of spinal cord injuryrdquo Stem Cellsvol 25 no 1 pp 115ndash124 2007
[37] S S Lee C H Woo J D Chang and J H Kim ldquoRoles of Racand cytosolic phospholipase A2 in the intracellular signalling inresponse to titanium particlesrdquo Cellular Signalling vol 15 no 3pp 339ndash345 2003
[38] E Masliah L Hansen A Adame et al ldquoA120573 vaccination effectson plaque pathology in the absence of encephalitis inAlzheimerdiseaserdquo Neurology vol 64 no 1 pp 129ndash131 2005
[39] A Pilorget B Annabi F Bouzeghrane J Marvaldi J Luisand R Beliveau ldquoInhibition of angiogenic properties of brainendothelial cells by platelet-derived sphingosine-1-phosphaterdquoJournal of Cerebral Blood Flow and Metabolism vol 25 no 9pp 1171ndash1182 2005
[40] A Wang P Das R C Switzer III T E Golde and J LJankowsky ldquoRobust amyloid clearance in a mouse model ofAlzheimerrsquos disease provides novel insights into themechanismof amyloid-120573 immunotherapyrdquo Journal of Neuroscience vol 31no 11 pp 4124ndash4136 2011
[41] G Krabbe A Halle V Matyash et al ldquoFunctional impairmentof microglia coincides with Beta-amyloid deposition in micewith Alzheimer-like pathologyrdquo PLoS ONE vol 8 no 4 ArticleID e60921 2013
[42] Y H Liu B Giunta H-D Zhou J Tan and Y J WangldquoImmunotherapy for Alzheimer disease-the challenge ofadverse effectsrdquo Nature Reviews Neurology vol 8 no 8 pp465ndash469 2012
[43] B Solomon R Koppel D Frankel and E Hanan-AharonldquoDisaggregation of Alzheimer120573-amyloid by site-directedmAbrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 94 no 8 pp 4109ndash4112 1997
[44] F Bard C Cannon R Barbour et al ldquoPeripherally adminis-tered antibodies against amyloid 120573-peptide enter the centralnervous system and reduce pathology in a mouse model ofAlzheimer diseaserdquo Nature Medicine vol 6 no 8 pp 916ndash9192000
[45] R B DeMattos K R Bales D J Cummins J C Dodart S MPaul and D M Holtzman ldquoPeripheral anti-A120573 antibody altersCNS and plasma A120573 clearance and decreases brain A120573 burdenin a mouse model of Alzheimerrsquos diseaserdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 98 no 15 pp 8850ndash8855 2001
[46] C A Lemere E T Spooner J LaFrancois et al ldquoEvidence forperipheral clearance of cerebral A120573 protein following chronic
16 BioMed Research International
active A120573 immunization in PSAPP micerdquo Neurobiology ofDisease vol 14 no 1 pp 10ndash18 2003
[47] T Town M Vendrame A Patel et al ldquoReduced Th1 andenhanced Th2 immunity after immunization with Alzheimerrsquos120573-amyloid
1minus42rdquo Journal of Neuroimmunology vol 132 no 1-2
pp 49ndash59 2002[48] R Cacabelos ldquoPharmacogenomics in Alzheimerrsquos diseaserdquo
Methods in Molecular Biology vol 448 pp 213ndash357 2008[49] R Cacabelos ldquoPharmacogenomics and therapeutic strategies
for dementiardquo Expert Review of Molecular Diagnostics vol 9no 6 pp 567ndash611 2009
[50] D van Dam R DrsquoHooge M Staufenbiel C van Ginneken Fvan Meir and P P de Deyn ldquoAge-dependent cognitive declinein the APP23 model precedes amyloid depositionrdquo EuropeanJournal of Neuroscience vol 17 no 2 pp 388ndash396 2003
[51] M Dumont C Strazielle M Staufenbiel and R LalondeldquoSpatial learning and exploration of environmental stimuli in24-month-old female APP23 transgenic mice with the Swedishmutationrdquo Brain Research vol 1024 no 1-2 pp 113ndash121 2004
[52] R Lalonde M Dumont M Staufenbiel C Sturchler-Pierratand C Strazielle ldquoSpatial learning exploration anxiety andmotor coordination in female APP23 transgenic mice with theSwedish mutationrdquo Brain Research vol 956 no 1 pp 36ndash442002
[53] F J Huff F Boller F Lucchelli R Querriera J Beyer and SBelle ldquoThe neurologic examination in patients with probableAlzheimerrsquos diseaserdquo Archives of Neurology vol 44 no 9 pp929ndash932 1987
[54] A F Pettersson M Engardt and L O Wahlund ldquoActivitylevel and balance in subjects with mild Alzheimerrsquos diseaserdquoDementia and Geriatric Cognitive Disorders vol 13 no 4 pp213ndash216 2002
[55] L Calandreau B Desgranges R Jaffard and A DesmedtldquoSwitching from contextual to tone fear conditioning and viceversa the key role of the glutamatergic hippocampal-lateralseptal neurotransmissionrdquo Learning amp Memory vol 17 no 9pp 440ndash443 2010
[56] F Trinchese S Liu F Battaglia SWalter PMMathews andOArancio ldquoProgressive age-related development of Alzheimer-like pathology in APPPS1 micerdquo Annals of Neurology vol 55no 6 pp 801ndash814 2004
[57] D M Wilcock A Rojiani A Rosenthal et al ldquoPassiveimmunotherapy against A120573 in aged APP-transgenic micereverses cognitive deficits and depletes parenchymal amyloiddeposits in spite of increased vascular amyloid and microhem-orrhagerdquo Journal of Neuroinflammation vol 1 article 24 2004
[58] S A Ferguson S Sarkar and L C Schmued ldquoLongitudinalbehavioral changes in the APPPS1 transgenic AlzheimerrsquosDisease modelrdquo Behavioural Brain Research vol 242 no 1 pp125ndash134 2013
[59] D Wilcock and C Colton ldquoImmunotherapy vascular pathol-ogy and microhemorrhages in transgenic micerdquo CNS andNeurological DisordersmdashDrug Targets vol 8 no 1 pp 50ndash642009
[60] M Grundman S Gilman R S Black et al ldquoAn alternativemethod for estimating efficacy of the AN1792 vaccine foralzheimer diseaserdquo Neurology vol 71 no 9 pp 697ndash698 2008
[61] D Boche E Zotova R O Weller et al ldquoConsequence of A120573immunization on the vasculature of human Alzheimerrsquos diseasebrainrdquo Brain vol 131 no 12 pp 3299ndash3310 2008
[62] M Pfeifer S Boncristiano L Bondolfi et al ldquoCerebral hemor-rhage after passive anti-a120573 immunotherapyrdquo Science vol 298no 5597 p 1379 2002
[63] M M Racke L I Boone D L Hepburn et al ldquoExacerbationof cerebral amyloid angiopathy-associatedmicrohemorrhage inamyloid precursor protein transgenic mice by immunotherapyis dependent on antibody recognition of deposited forms ofamyloid 120573rdquo Journal of Neuroscience vol 25 no 3 pp 629ndash6362005
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

10 BioMed Research International
002040608
1121416
Preventive treatmentTherapeutic treatment
Ant
i-120573-a
myl
oid
IgG
antib
odie
s
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
Gr A
(A120573+
S1P+
L)(E
B101
)
Figure 6 Quantification of specific anti-120573-amyloid IgG in immu-nized APPPS1 mice The presence of anti-120573-amyloid antibodiesmeasured in the sera of vaccinated and control mice groups duringthe preventive and therapeutic treatments indicates that EB101liposome vaccine was able to induce the highest production ofspecific IgG antibodies Compared to theGroupD immunizedmicethe level of antibodies observed in the Group A was also highersuggesting a more effective immune response (119875 lt 001 versusGroup D 119875 lt 0001 versus Groups B C and E) Quantification ofspecific anti-120573-amyloid IgG in immunized Tg mice
in Group D with respect to all other groups mainlyreported by the high levels of TNF-120572 (Figure 7(b)) andIFN-120574 (Figure 7(c)) cytokine secretions while due to thehigh mortality rate during the therapeutic treatment nosignificant data was obtained from this mice group Inaddition a tendency toTh1 secretion reduction was observedinGroupAwith respect to all other groups in both preventiveand therapeutic treatment The changes in Th2 protein levels(Figure 7(d)) followed the inverse trend where the highestdifferences between Group A and the other treatment groupswere observed in the IL-4 IL-5 and IL-10 cytokine secretions(119875 lt 005 versus Group D 119875 lt 001 versus Group E Figures7(e)ndash7(g))
35 Comparative Motor Coordination and Memory Perfor-mances In order to validate the locomotion integrity duringthe entire immunotherapeutic process as reported in similarprevious studies motor coordination tests have showed thatEB101-treated mice spent significantly more time on theaccelerating rotarodThismotor coordination task shows thatEB101 immunization affected positively their performance(Figure 8(a)) A120573
42liposome (B) S1P-containing liposomes
(C) and PBS (E) (119875 lt 0006) mice performed poorly incomparison to EB101 mice (119875 lt 005) or A120573
42+ CFAIFA
mice (119875 lt 002) in motor abilities and coordination Thefear conditioning-active avoidance test was used to assess thespatial learning and memory functions As expected bothEB101 and A120573
42+ CFAIFA immunized mice readily learned
the location of the platform as shown by a decrease in theescape latency over training days while the control group
mice exhibited no significant changes during 6-day trialsThe average escape latencies of EB101 (68 plusmn 2 s 761 plusmn 22 s)andA120573
42+CFAIFA (116plusmn33 s 123plusmn21 s) immunizedmice
at the end of preventive and therapeutic periods respectivelyduring the consecutive 6 days trial were significantly shorterthan the other treatment groups (A120573
42liposome 163 plusmn 31 s
175 plusmn 28 s S1P-containing liposomes 202 plusmn 19 s 248 plusmn27 s) and the control (2116 plusmn 38 s 268 plusmn 22 s) (119875 lt 001)Therewas no significant difference between EB101 andA120573
42+
CFAIFA immunized group (119875 gt 005) (Figure 8(b)) Theseresults showed that spatial learning was improved in theseimmunized mice
4 Discussion
In this study we have compared a novel active A120573 imm-unotherapy the EB101 vaccine against A120573 with the A120573
42+
CFAIFA vaccine that was first taken into clinical trialswith coadjuvant Qs-21 modification (AN1792 by Orgogozoet al [19]) This type of vaccine design was thought toinduce a strong T-cell mediated inflammatory response (IL-2 and IFN-c positive responses indicative of a Class II(CD4) Th-1 type response [33] leading to acute menin-goencephalitis in the 6 of the patients) The EB101 activeimmunotherapeutic vaccine was primarily designed to avoidundesired inflammatory side effects both in mice andhumans (without need of coadjuvant modification) whileinducing effective antibody titer rates We demonstrated thatlong-term immunization with EB101 induced A120573 antibodiesin the preventive and the therapeutic treatment periodswhich effectively reduced amyloid deposition in both corticaland hippocampal brain regions and prevented neuroinflam-mation probably by blocking toxicity in the surroundingneurons However the AN1792 vaccine was shown to inducein 6 of the patients an inflammatory reaction that mighthave led to the encephalitis type reaction observed in theclinical trial which was responsible for the immediate haltin those clinical studies It seems that AN1792 immunizationdelivered in Qs-21 and polysorbate 80 adjuvant induced aTh1 type response that likely led to autoaggressive T cells andaseptic meningoencephalitis In the present experiments wedemonstrated that a judiciously selected adjuvant produces adifferent humoral response [22] Indeed we have shown thatour novel immunotherapeutic strategy boosts specific A120573
1ndash42antibody generation and induces a Th2 noninflammatoryimmune response This boosted effect is in accordance withprevious immunotherapeutic studies [34] Taking togetherall the present findings these results suggest that EB101vaccine can achieve in a future clinical trial immunogenicresponse rates quite similar to the AN1792 without theinflammatory effects reported The immunohistochemicalanalysis obtained in the present study shows that all the EB101immunized mice resulted in an almost complete preventionof A120573 deposition in the brain with only a few compactA120573 plaques in the retrosplenial and dentate gyrus regionsAlthough there was a slightly increase in death in EB101treated mice compared to controls this significant reductionof A120573 plaques in EB101 treated mice was proven to be more
BioMed Research International 11
Gr A Gr B Gr C Gr D Gr EPreventive treatmentTherapeutic treatment
0005001000150020002500300035004000
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
(a) Th1
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
000
1000
2000
3000
4000
5000TNF-120572
(b) Th1
000500
10001500200025003000
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
IFN-120574
(c) Th1
000500
100015002000250030003500
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
Group A Group B Group C Group D Group EPreventive treatmentTherapeutic treatment
(d) Th2
000
500
1000
1500
2000
2500
3000IL-4
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(e) Th2
000500
100015002000250030003500
IL-5
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(f) Th2
3500IL-10
000500
10001500200025003000
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(g) Th2
Figure 7 Detection of Th1 and Th2 cytokine profile Measurements of the effect of EB101 and A12057342
+ CFAIFA vaccines on Th1 and Th2cytokines profile during preventive and therapeutic treatments ((a)ndash(c)) A significant increase of Th1 cytokine profile was observed inpreventive treatment in Group D with respect to all other groups while a tendency to a reduction inTh1 secretion was observed in Group Awith respect to all other groups in both preventive and therapeutic treatment ((d)ndash(g)) The changes in Th2 protein levels were observed inthe IL-4 IL-5 and IL-10 cytokine secretions of Group A (119875 lt 005 versus Group D 119875 lt 001 versus Group E)
12 BioMed Research International
Gr A
(EB1
01)
05
101520253035
Preventive treatment Therapeutic treatment
Mea
n fa
lling
late
ncy
(s)
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)(a) Graphic of motor coordination and balance
05
1015202530
Mea
n re
spon
se ti
me (
s)
Gr A
(EB1
01)
Preventive treatment Therapeutic treatment
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
(b) Graphic of associative learning and memory
Figure 8 Analysis of motor coordination and balance (a) Rotarod results of motor coordination and balance test after the preventive andtherapeutic treatment showing improved effect of EB101 and A120573
42+ CFAIFA-vaccine compared to other treated mice groups (119875 lt 005)
and (119875 lt 002) versus control mice Latency to fall on the rotarod apparatus was significantly improved in EB101 immunized mice Valuesrepresent means of all six trials of a groupday (b) Spatial learning and memory tested by fear conditioning-active avoidance test The meanresponse time of each treatment group to reach the platform is represented EB101 and A120573
42+ CFAIFA immunized mice had significantly
lower latencies than the other treatment groups (119875 lt 001) versus control mice
efficient than that obtained with A12057342+ CFAIFA immuniza-
tions A comparison of A120573 plaque reduction rates amongtreated groups surprisingly showed that other than EB101and A120573
42+ CFAIFA treated groups some amyloid deposits
were also reduced in Groups B (A1205731ndash42liposome) and C
(S1P-containing liposomes) Although the reduction rates inthese mice were markedly low the subtle lowering effectmight be due to the interaction between the immunizationelements (A120573
1ndash42 and S1P) and the mouse immunogenicsystem as reported by others [35] Based on our presentresults S1P one of the components of the adjuvant used withEB101 might be responsible for the neuroprotective effectobserved in the vulnerable neuronal areas of EB101 treatedmice The advantage in the A120573 load preventionreduction bycombining A120573
1ndash42 and S1P in a unique formulation vaccinehas been also observed in the reduced hippocampal load ofEB101 immunizedmice (Gr A) when comparedwith differentvaccine components (Gr B-C) In fact S1P appears to have ademonstrated therapeutic potential as a regenerative agent inthe nervous system since it controls migration of neuronalstem cells toward a site of spinal cord injury [36] inducecytoskeletal rearrangements to promote transmitter secretion[37] and also trigger diverse cellular effects including angio-genesis cardiac development immunity cell motility andneurite extension [38 39]Therefore it is reasonable to expectthat such a potent biologically active sphingolipid by itselfmay play an important role in the A120573 plaque reduction Inthis study the clearance efficiency rate of each treatment wasanalyzed and results suggested that EB101 (6459) appearsto be more effective than A120573
42+ CFAIFA (1920) vaccine
in both preventive and therapeutic groups The effectivenessof EB101 in older transgenic animals (18 months) showeda clearance effect of 59 suggesting that EB101 may alsohave therapeutic potential inADdiagnosticated patient trials
Similar conclusion was reached byWang and colleagues [40]who demonstrated that combination therapy in aged animalsprovides more valuable insights into a possible translationto humansOur immunologicalmeasurements demonstratedonce again that neuroinflammation develops in an A120573-dependent fashion early in the course of AD [41] and canbe alleviated or reversed by interventional A120573 vaccinationsuch as EB101 Among the possible mechanisms of A120573clearance proposed in AD immunotherapy [42] our resultssuggested that theymay overlap and probably be disease statedependent since the prevention and the therapeutic vaccinewould likely benefit from the transport of A120573 antibodies intothe CNS However some mechanisms of action such as thepreventive effect of A120573 antibodies in the A120573 aggregation [43]microglial phagocytosis induced by the Fc binding portionsbetween A120573 antibodies and microglia receptor [44] and theshift in the gradient of A120573 transport across the blood-brainbarrier due to the increased concentration of A120573 antibodiesin the vascular system resulting in an increase in effluxfrom the brain to blood [45 46] may be playing a role inboth preventive and therapeutic conditions Our findingstherefore suggest that novel treatment strategies aimed atmaintaining or increasing microglial function and restoringthe balance to the immunological system may represent anattractive therapeutic approach even at the advanced stagesof ADThe immunization results obtained with EB101 duringboth preventive and therapeutic treatment showed nomicro-hemorrhages infiltration of lymphocytes or encephalitis-like reaction in the brain suggesting that EB101 does notinduce A120573-specific Th1 reaction possibly due to the neu-roprotective effect of S1P Delivering A120573
1ndash42 in a liposomal-S1P-adjuvant resulted in an effective immunological responseincluding the induction of a Th2 T-cell response whicheffectively prevented neuroinflammation in an animal model
BioMed Research International 13
of AD as reported previously [24 47] Moreover presentresults demonstrated that EB101 did not induce astrogliosisin APPPS1 transgenic mice whereas some GFAP-positiveastrocyte clusters were aggregated in the affected brain areasof mice treated with A120573
42+ CFAIFA These results indicate
that EB101 may result in an effective treatment in AD neu-ropathology due to the absence of peripheral proinflamma-tory cytokine secretion and the absence of astroglial activa-tion in the brains of EB101 treated mice when compared withA12057342
+ CFAIFA immunotherapeutic response In a clinicalsetting however we may expect a wide differential responsein part depending upon the genomic profile of each patient[48 49] In AD behavioural studies in particular amongthe motor coordination tests the rotarod performance hasbeen reported to be impaired in some AD transgenic micemodels bearing A120573 plaques [50 51] This result was found inmutants prior to A120573 plaque formation at 3 and 6 months ofage but was not reproduced after plaques had been formedat 16 and 24 months of age [52] perhaps because of thedeclining abilities of older normal mice Here we presentthe effects of vaccine treatments in APPPS1 mice on motorcoordination a neglected aspect of the AD experimentalbehavior but justified on the basis of the deficient posturalcontrol reported in patients with Alzheimerrsquos disease [53]sometimes even during early stages [54] Our longitudinalmotor behavioral results showed no significant differencesin the ambulatory movement grip strength or coordinationaddressed by their performance in the rotating rod betweenAPPPS1 and wild-type mice for any age group studiedin contrast with other treated groups demonstrating thatlongitudinal motor assessments of AD transgenic mousemodels can provide valuable information with regard tothe effect of immunization on the progression of behavioraldeficits and overall motor activity As shown the EB101-treated group of mice exhibited longer latencies and betterbalancing skills indicating the absence of any negative effectof EB101 immunization on these mice The classical fearconditioning test is assumed to reveal the functional integrityof the hippocampus [55] representing a special memorytask to evaluate the AD models and its improvement afterimmunization periods In this APPPS1 mouse model thefirst associative learning and spatial workingmemory deficitsare shown at 5-6 months of age [56] although its impair-ment progression is relatively slow compared to most othermodels At the end of preventive immunization period EB101and A120573
42+ CFAIFA immunized mice have significantly
shorter escape latencies than the other treatment groups(A12057342liposome S1P-containing liposomes) and the controls
in the fear conditioning test Shorter escape latencies duringspatial learning indicated that deficits of spatial memorywere attenuated in EB101 and A120573
42+ CFAIFA immunized
mice Previous studies have showed that by 15 months ofage the spatial working memory is consistently impairedthroughout the rest of the life span [31 57] At the end oftherapeutic immunization period both EB101 and A120573
42+
CFAIFA immunized mice showed significantly improvedresults on the hippocampus-dependent spatial learning andmemory test when compared with other treatment groupsand the controls Taken together these observations are
in agreement with the impairment in conditioned learningin response to a tone stimulus of aged APPPS1 micealthough when immunized with EB101 during preventiveor therapeutic periods such impairments are significantlyattenuated
In summary the present results corroborate our previouspublished findings showing that EB101 immunization reducesamyloid accumulation in the APPPS1 mouse brain [2425] The strongest reduction in the preventive immunizationwas observed in the retrosplenial cortex followed by thehippocampus and entorhinal-piriform cortex Our resultsfurther support that EB101 ismore effectivewhen themice areimmunized prior to developing plaques in the hippocampaland cortical brain regions There is extensive evidence inmouse preclinical studies [58] and human clinical trials [5960] that the adverse effects of active and passive A120573 vaccinesresult in microhemorrhages and encephalitic reactions [60ndash63] yet themechanismbywhich they occur has not been fullyelucidated In the treatment sections analyzed we found nomicrohemorrhages or acute inflammatory processes in all thesurvived mice of the group indicating that EB101 therapeuticapproach presents no adverse inflammatory effects in thisAD mice models Thus our present results unequivocallydemonstrate that EB101 vaccinemay not only halt AD-relatedpathology as a preventive treatment but may also reverse it inestablished cases related to the improvement of the learningandmemory in both cases leading to a robust immunologicaltherapeutic response Present experiments also demonstratethat EB101 exerts more significant and substantial effects onthe reduction of A120573 deposition than A120573
42+CFAIFA vaccine
in A120573 mice models of AD However further exploration ofthe contribution of genetic environmental and physiologicalfactors in the development resistance to endogenous A120573-antibody is needed in order to optimize and modulate thetherapeutic strategies currently being developed
5 Conclusions
The present study carried out in APPPS1 transgenic mousemodel of AD compared the A120573 burden reduction andcellular immune responses produced by two immunizationdelivery strategies of A120573 in Freund adjuvant (described byWebster et al [10]) and in liposomeS1P matrix (describedby Schenk et al [17])The EB101 immunotherapeutic strategyboosted antibody generation A120573 plaque reduction and ananti-inflammatory response more effectively than A120573
42+
CFAIFA especially if given prior to or in the early stages ofpathology The present results also showed that vaccinationwith EB101 prevented the formation of new plaques andpresumably led to a reduction in established plaques whileinducing aTh2 immune reactionwithout encephalitogenic T-cell responses or brain inflammation related with astrogliosisHowever despite the efficacy demonstrated of this novelvaccine approach further preclinical and clinical studieson EB101 are warranted to evaluate its efficacy and themechanism for the future utility of such vaccine for ADtherapy
14 BioMed Research International
List of Abbreviations
AD Alzheimerrsquos diseaseA120573 120573-Amyloid proteinAPP Amyloid precursor proteinAPT Anterior pretectal nucleusCA1 Field CA1 hippocampusCA3 Field CA3 hippocampusCPu Caudate putamen (striatum)Cx CortexDG Dentate gyrusDN Dystrophic neuriteEct Entorhinal cortexCFA Complete Freundrsquos adjuvantGrDG Granular dentate gyrusIFA Incomplete Freundrsquos adjuvantIL InterleukinLPMR Lateral posterior thalamic nucleus (mediorostral)MoDG Molecular dentate gyrusPir Piriform cortexPoDG Polymorph dentate gyrusRSGb Retrosplenial granular cortex b regionTg TransgenicTh T helper
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Ivan Carrera Ignacio Etcheverrıa Lucıa Fernandez-NovoaValter Lombardi Ramon Cacabelos and Carmen Vigo con-ceived and designed the experiments Ivan Carrera and Igna-cio Etcheverrıa performed the experiments Ivan CarreraIgnacio Etcheverrıa and Valter Lombardi analyzed the dataIvan Carrera Ignacio Etcheverrıa Lucıa Fernandez-Novoaand Valter Lombardi contributed reagentsmaterialsanalysistools Ivan Carrera Carmen Vigo andMadepalli KrishnappaLakshmana wrote the paper
Acknowledgments
This work was supported by National Institute of Aging(NIA)NIH Grant no 1R01AG036859-01 (Madepalli Krish-nappa Lakshmana) and the financial support of the ProgramaOperativo Feder Galicia 2007ndash2013 (IN841C 201254 to PhDCarrera I)
References
[1] R Cacabelos L Fernandez-Novoa V Lombardi Y Kubotaand M Takeda ldquoMolecular genetics of Alzheimerrsquos diseaseand agingrdquoMethods and Findings in Experimental and ClinicalPharmacology vol 27 pp 1ndash573 2005
[2] P T Nelson I Alafuzoff E H Bigio et al ldquoCorrelationof alzheimer disease neuropathologic changes with cognitive
status a review of the literaturerdquo Journal of Neuropathology andExperimental Neurology vol 71 no 5 pp 362ndash381 2012
[3] JNaslundVHaroutunian RMohs et al ldquoCorrelation betweenelevated levels of amyloid 120573-peptide in the brain and cognitivedeclinerdquo Journal of the American Medical Association vol 283no 12 pp 1571ndash1577 2000
[4] A Koyama O I Okereke T Yang D Blacker D J Selkoeand F Grodstein ldquoPlasma amyloid-120573 as a predictor of dementiaand cognitive decline a systematic review and meta-analysisrdquoArchives of Neurology vol 69 no 7 pp 824ndash831 2012
[5] A M Fagan C M Roe C Xiong M A Mintun J C Morrisand D M Holtzman ldquoCerebrospinal fluid tau120573-amyloid42ratio as a prediction of cognitive decline in nondemented olderadultsrdquo Archives of Neurology vol 64 no 3 pp 343ndash349 2007
[6] S Parvathy P Davies V Haroutunian et al ldquoCorrelationbetween A120573-40- A120573-42- and A120573x-43-containing amyloidplaques and cognitive declinerdquo Archives of Neurology vol 58no 12 pp 2025ndash2032 2001
[7] E Karran M Mercken and B D Strooper ldquoThe amyloidcascade hypothesis for Alzheimerrsquos disease an appraisal for thedevelopment of therapeuticsrdquo Nature Reviews Drug Discoveryvol 10 no 9 pp 698ndash712 2011
[8] J L Jankowsky D J Fadale J Anderson et al ldquoMutantpresenilins specifically elevate the levels of the 42 residue 120573-amyloid peptide in vivo evidence for augmentation of a 42-specific 120574 secretaserdquo Human Molecular Genetics vol 13 no 2pp 159ndash170 2004
[9] L Holcomb M N Gordon E Mcgowan et al ldquoAcceleratedAlzheimer-type phenotype in transgenic mice carrying bothmutant amyloid precursor protein and presenilin 1 transgenesrdquoNature Medicine vol 4 no 1 pp 97ndash100 1998
[10] S JWebster A D Bachstetter and L J van Eldik ldquoComprehen-sive behavioral characterization of an APPPS-1 double knock-in mouse model of Alzheimerrsquos diseaserdquo Alzheimerrsquos ResearchandTherapy vol 5 no 3 article 28 2013
[11] H Xiong D Callaghan J Wodzinska et al ldquoBiochemical andbehavioral characterization of the double transgenic mousemodel (APPswePS1dE9) of Alzheimerrsquos diseaserdquo NeuroscienceBulletin vol 27 no 4 pp 221ndash232 2011
[12] M Garcia-Alloza L A Borrelli B T Hyman and B J BacskaildquoAntioxidants have a rapid and long-lasting effect on neuriticabnormalities in APPPS1 micerdquo Neurobiology of Aging vol 31no 12 pp 2058ndash2068 2010
[13] M Meyer-Luehmann M Mielke T L Spires-Jones et al ldquoAreporter of local dendritic translocation shows plaque-relatedloss of neural system function in APP-transgenic micerdquo Journalof Neuroscience vol 29 no 40 pp 12636ndash12640 2009
[14] E Machova V Rudajev H Smyckova H Koivisto H Tanilaand V Dolezal ldquoFunctional cholinergic damage develops withamyloid accumulation in young adult APPswePS1dE9 trans-genic micerdquo Neurobiology of Disease vol 38 no 1 pp 27ndash352010
[15] T Malm J Koistinaho and K Kanninen ldquoUtilization of APP-swePS1dE9 transgenic mice in research of Alzheimerrsquos diseasefocus on gene therapy and cell-based therapy applicationsrdquoInternational Journal of Alzheimerrsquos Disease vol 2011 Article ID517160 8 pages 2011
[16] J Lee and P Han ldquoAn update of animal models of alzheimerdisease with a reevaluation of plaque depositionsrdquoExperimentalNeurobiology vol 22 pp 84ndash95 2013
BioMed Research International 15
[17] D Schenk R Barbour W Dunn et al ldquoImmunization withamyloid-120573 attenuates Alzheimer disease-like pathology in thePDAPP mouserdquo Nature vol 400 no 6740 pp 173ndash177 1999
[18] H J Fu B Liu J L Frost and C A Lemere ldquoAmyloid-120573immunotherapy for Alzheimerrsquos diseaserdquoCNS and NeurologicalDisorders-Drug Targets vol 9 no 2 pp 197ndash206 2010
[19] J-M Orgogozo S Gilman J-F Dartigues et al ldquoSubacutemeningoencephalitis in a subset of patients with AD after A120573
42
immunizationrdquo Neurology vol 61 no 1 pp 46ndash54 2003[20] J A R Nicolll DWilkinson CHolmes P Steart HMarkham
andROWeller ldquoNeuropathology of humanAlzheimer diseaseafter immunization with amyloid-120573 peptide a case reportrdquoNature Medicine vol 9 no 4 pp 448ndash452 2003
[21] I Ferrer M Boada Rovira M L Sanchez Guerra M JRey and F Costa-Jussa ldquoNeuropathology and pathogenesisof encephalitis following amyloid-Neuropathology and patho-genesis of encephalitis following amyloid-beta immunizationin Alzheimerrsquos disease immunization in Alzheimerrsquos diseaserdquoBrain Pathology vol 14 no 1 pp 11ndash20 2004
[22] D H Cribbs A Ghochikyan V Vasilevko et al ldquoAdjuvant-dependent modulation ofTh1 andTh2 responses to immuniza-tion with 120573-amyloidrdquo International Immunology vol 15 no 4pp 505ndash514 2003
[23] C Holmes D Boche D Wilkinson et al ldquoLong-term effectsof A120573
42immunisation in Alzheimerrsquos disease follow-up of a
randomised placebo-controlled phase I trialrdquo The Lancet vol372 no 9634 pp 216ndash223 2008
[24] I Carrera I Etcheverrıa L Fernandez-Novoa V LombardiR Cacabelos and C Vigo ldquoVaccine development to treatAlzheimerrsquos disease neuropathology in APPPS1 transgenicmicerdquo International Journal of Alzheimerrsquos Disease vol 2012Article ID 376138 17 pages 2012
[25] I Carrera I Etcheverrıa Y Li et al ldquoImmunocytochemicalcharacterization of Alzheimer disease hallmarks in APPPS1transgenic mice treated with a new anti-amyloid-120573 vaccinerdquoBioMed Research International vol 2013 Article ID 709145 12pages 2013
[26] V Anelli R Bassi G Tettamanti P Viani and L Riboni ldquoExtra-cellular release of newly synthesized sphingosine-1-phosphateby cerebellar granule cells and astrocytesrdquo Journal of Neuro-chemistry vol 92 no 5 pp 1204ndash1215 2005
[27] S Schroeter K Khan R Barbour et al ldquoImmunotherapyreduces vascular amyloid-120573 in PDAPP micerdquo Journal of Neu-roscience vol 28 no 27 pp 6787ndash6793 2008
[28] Z Suo A Cox N Bartelli et al ldquoGRK5 deficiency leads to earlyAlzheimer-like pathology and working memory impairmentrdquoNeurobiology of Aging vol 28 no 12 pp 1873ndash1888 2007
[29] W Kamphuis C Mamber MMoeton et al ldquoGFAP isoforms inadult mouse brain with a focus on neurogenic astrocytes andreactive astrogliosis in mouse models of Alzheimer diseaserdquoPLoS ONE vol 7 no 8 Article ID e42823 2012
[30] J Lang C Vigo-Pelfrey and F Martin ldquoLiposomes composedof partially hydrogenated egg phosphatidylcholines fatty acidcomposition thermal phase behavior and oxidative stabilityrdquoChemistry and Physics of Lipids vol 53 no 1 pp 91ndash101 1990
[31] D Morgan D M Diamond P E Gottschall et al ldquoA 120573peptide vaccination prevents memory loss in an animal modelof Alzheimerrsquos diseaserdquo Nature vol 408 no 6815 pp 982ndash9852000
[32] M L Brownlow L Benner D DrsquoAgostino M N Gordon andD Morgan ldquoKetogenic diet improves motor performance but
not cognition in two mouse models of Alzheimerrsquos pathologyrdquoPLoS ONE vol 8 no 9 Article ID e75713 2013
[33] M Pride P Seubert M Grundman M Hagen J Eldridgeand R S Black ldquoProgress in the active immunotherapeu-tic approach to Alzheimerrsquos disease clinical investigationsinto AN1792-associated meningoencephalitisrdquo Neurodegenera-tive Diseases vol 5 no 3-4 pp 194ndash196 2008
[34] CWiessner K-HWiederhold A C Tissot et al ldquoThe second-generation activeA120573 immunotherapyCAD106 reduces amyloidaccumulation in APP transgenic mice while minimizing poten-tial side effectsrdquoThe Journal of Neuroscience vol 31 no 25 pp9323ndash9331 2011
[35] D Lambracht-Washington and R Rosenberg ldquoAdvances in thedevelopment of vaccines for Alzheimerrsquos diseaserdquo DiscoveryMedicine vol 15 no 84 pp 319ndash326 2013
[36] A Kimura T Ohmori R Ohkawa et al ldquoEssential roles ofsphingosine 1-phosphateS1P
1receptor axis in the migration of
neural stem cells toward a site of spinal cord injuryrdquo Stem Cellsvol 25 no 1 pp 115ndash124 2007
[37] S S Lee C H Woo J D Chang and J H Kim ldquoRoles of Racand cytosolic phospholipase A2 in the intracellular signalling inresponse to titanium particlesrdquo Cellular Signalling vol 15 no 3pp 339ndash345 2003
[38] E Masliah L Hansen A Adame et al ldquoA120573 vaccination effectson plaque pathology in the absence of encephalitis inAlzheimerdiseaserdquo Neurology vol 64 no 1 pp 129ndash131 2005
[39] A Pilorget B Annabi F Bouzeghrane J Marvaldi J Luisand R Beliveau ldquoInhibition of angiogenic properties of brainendothelial cells by platelet-derived sphingosine-1-phosphaterdquoJournal of Cerebral Blood Flow and Metabolism vol 25 no 9pp 1171ndash1182 2005
[40] A Wang P Das R C Switzer III T E Golde and J LJankowsky ldquoRobust amyloid clearance in a mouse model ofAlzheimerrsquos disease provides novel insights into themechanismof amyloid-120573 immunotherapyrdquo Journal of Neuroscience vol 31no 11 pp 4124ndash4136 2011
[41] G Krabbe A Halle V Matyash et al ldquoFunctional impairmentof microglia coincides with Beta-amyloid deposition in micewith Alzheimer-like pathologyrdquo PLoS ONE vol 8 no 4 ArticleID e60921 2013
[42] Y H Liu B Giunta H-D Zhou J Tan and Y J WangldquoImmunotherapy for Alzheimer disease-the challenge ofadverse effectsrdquo Nature Reviews Neurology vol 8 no 8 pp465ndash469 2012
[43] B Solomon R Koppel D Frankel and E Hanan-AharonldquoDisaggregation of Alzheimer120573-amyloid by site-directedmAbrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 94 no 8 pp 4109ndash4112 1997
[44] F Bard C Cannon R Barbour et al ldquoPeripherally adminis-tered antibodies against amyloid 120573-peptide enter the centralnervous system and reduce pathology in a mouse model ofAlzheimer diseaserdquo Nature Medicine vol 6 no 8 pp 916ndash9192000
[45] R B DeMattos K R Bales D J Cummins J C Dodart S MPaul and D M Holtzman ldquoPeripheral anti-A120573 antibody altersCNS and plasma A120573 clearance and decreases brain A120573 burdenin a mouse model of Alzheimerrsquos diseaserdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 98 no 15 pp 8850ndash8855 2001
[46] C A Lemere E T Spooner J LaFrancois et al ldquoEvidence forperipheral clearance of cerebral A120573 protein following chronic
16 BioMed Research International
active A120573 immunization in PSAPP micerdquo Neurobiology ofDisease vol 14 no 1 pp 10ndash18 2003
[47] T Town M Vendrame A Patel et al ldquoReduced Th1 andenhanced Th2 immunity after immunization with Alzheimerrsquos120573-amyloid
1minus42rdquo Journal of Neuroimmunology vol 132 no 1-2
pp 49ndash59 2002[48] R Cacabelos ldquoPharmacogenomics in Alzheimerrsquos diseaserdquo
Methods in Molecular Biology vol 448 pp 213ndash357 2008[49] R Cacabelos ldquoPharmacogenomics and therapeutic strategies
for dementiardquo Expert Review of Molecular Diagnostics vol 9no 6 pp 567ndash611 2009
[50] D van Dam R DrsquoHooge M Staufenbiel C van Ginneken Fvan Meir and P P de Deyn ldquoAge-dependent cognitive declinein the APP23 model precedes amyloid depositionrdquo EuropeanJournal of Neuroscience vol 17 no 2 pp 388ndash396 2003
[51] M Dumont C Strazielle M Staufenbiel and R LalondeldquoSpatial learning and exploration of environmental stimuli in24-month-old female APP23 transgenic mice with the Swedishmutationrdquo Brain Research vol 1024 no 1-2 pp 113ndash121 2004
[52] R Lalonde M Dumont M Staufenbiel C Sturchler-Pierratand C Strazielle ldquoSpatial learning exploration anxiety andmotor coordination in female APP23 transgenic mice with theSwedish mutationrdquo Brain Research vol 956 no 1 pp 36ndash442002
[53] F J Huff F Boller F Lucchelli R Querriera J Beyer and SBelle ldquoThe neurologic examination in patients with probableAlzheimerrsquos diseaserdquo Archives of Neurology vol 44 no 9 pp929ndash932 1987
[54] A F Pettersson M Engardt and L O Wahlund ldquoActivitylevel and balance in subjects with mild Alzheimerrsquos diseaserdquoDementia and Geriatric Cognitive Disorders vol 13 no 4 pp213ndash216 2002
[55] L Calandreau B Desgranges R Jaffard and A DesmedtldquoSwitching from contextual to tone fear conditioning and viceversa the key role of the glutamatergic hippocampal-lateralseptal neurotransmissionrdquo Learning amp Memory vol 17 no 9pp 440ndash443 2010
[56] F Trinchese S Liu F Battaglia SWalter PMMathews andOArancio ldquoProgressive age-related development of Alzheimer-like pathology in APPPS1 micerdquo Annals of Neurology vol 55no 6 pp 801ndash814 2004
[57] D M Wilcock A Rojiani A Rosenthal et al ldquoPassiveimmunotherapy against A120573 in aged APP-transgenic micereverses cognitive deficits and depletes parenchymal amyloiddeposits in spite of increased vascular amyloid and microhem-orrhagerdquo Journal of Neuroinflammation vol 1 article 24 2004
[58] S A Ferguson S Sarkar and L C Schmued ldquoLongitudinalbehavioral changes in the APPPS1 transgenic AlzheimerrsquosDisease modelrdquo Behavioural Brain Research vol 242 no 1 pp125ndash134 2013
[59] D Wilcock and C Colton ldquoImmunotherapy vascular pathol-ogy and microhemorrhages in transgenic micerdquo CNS andNeurological DisordersmdashDrug Targets vol 8 no 1 pp 50ndash642009
[60] M Grundman S Gilman R S Black et al ldquoAn alternativemethod for estimating efficacy of the AN1792 vaccine foralzheimer diseaserdquo Neurology vol 71 no 9 pp 697ndash698 2008
[61] D Boche E Zotova R O Weller et al ldquoConsequence of A120573immunization on the vasculature of human Alzheimerrsquos diseasebrainrdquo Brain vol 131 no 12 pp 3299ndash3310 2008
[62] M Pfeifer S Boncristiano L Bondolfi et al ldquoCerebral hemor-rhage after passive anti-a120573 immunotherapyrdquo Science vol 298no 5597 p 1379 2002
[63] M M Racke L I Boone D L Hepburn et al ldquoExacerbationof cerebral amyloid angiopathy-associatedmicrohemorrhage inamyloid precursor protein transgenic mice by immunotherapyis dependent on antibody recognition of deposited forms ofamyloid 120573rdquo Journal of Neuroscience vol 25 no 3 pp 629ndash6362005
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

BioMed Research International 11
Gr A Gr B Gr C Gr D Gr EPreventive treatmentTherapeutic treatment
0005001000150020002500300035004000
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
IL-1120572
IFN
-120574TN
F-120572
IL-2
(a) Th1
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
000
1000
2000
3000
4000
5000TNF-120572
(b) Th1
000500
10001500200025003000
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
IFN-120574
(c) Th1
000500
100015002000250030003500
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
IL-4
IL-5
IL-6
IL-1
0G
M-C
SFIL
-17
Group A Group B Group C Group D Group EPreventive treatmentTherapeutic treatment
(d) Th2
000
500
1000
1500
2000
2500
3000IL-4
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(e) Th2
000500
100015002000250030003500
IL-5
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(f) Th2
3500IL-10
000500
10001500200025003000
Group A Group B Group C Group D Group E
Preventive treatmentTherapeutic treatment
(g) Th2
Figure 7 Detection of Th1 and Th2 cytokine profile Measurements of the effect of EB101 and A12057342
+ CFAIFA vaccines on Th1 and Th2cytokines profile during preventive and therapeutic treatments ((a)ndash(c)) A significant increase of Th1 cytokine profile was observed inpreventive treatment in Group D with respect to all other groups while a tendency to a reduction inTh1 secretion was observed in Group Awith respect to all other groups in both preventive and therapeutic treatment ((d)ndash(g)) The changes in Th2 protein levels were observed inthe IL-4 IL-5 and IL-10 cytokine secretions of Group A (119875 lt 005 versus Group D 119875 lt 001 versus Group E)
12 BioMed Research International
Gr A
(EB1
01)
05
101520253035
Preventive treatment Therapeutic treatment
Mea
n fa
lling
late
ncy
(s)
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)(a) Graphic of motor coordination and balance
05
1015202530
Mea
n re
spon
se ti
me (
s)
Gr A
(EB1
01)
Preventive treatment Therapeutic treatment
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
(b) Graphic of associative learning and memory
Figure 8 Analysis of motor coordination and balance (a) Rotarod results of motor coordination and balance test after the preventive andtherapeutic treatment showing improved effect of EB101 and A120573
42+ CFAIFA-vaccine compared to other treated mice groups (119875 lt 005)
and (119875 lt 002) versus control mice Latency to fall on the rotarod apparatus was significantly improved in EB101 immunized mice Valuesrepresent means of all six trials of a groupday (b) Spatial learning and memory tested by fear conditioning-active avoidance test The meanresponse time of each treatment group to reach the platform is represented EB101 and A120573
42+ CFAIFA immunized mice had significantly
lower latencies than the other treatment groups (119875 lt 001) versus control mice
efficient than that obtained with A12057342+ CFAIFA immuniza-
tions A comparison of A120573 plaque reduction rates amongtreated groups surprisingly showed that other than EB101and A120573
42+ CFAIFA treated groups some amyloid deposits
were also reduced in Groups B (A1205731ndash42liposome) and C
(S1P-containing liposomes) Although the reduction rates inthese mice were markedly low the subtle lowering effectmight be due to the interaction between the immunizationelements (A120573
1ndash42 and S1P) and the mouse immunogenicsystem as reported by others [35] Based on our presentresults S1P one of the components of the adjuvant used withEB101 might be responsible for the neuroprotective effectobserved in the vulnerable neuronal areas of EB101 treatedmice The advantage in the A120573 load preventionreduction bycombining A120573
1ndash42 and S1P in a unique formulation vaccinehas been also observed in the reduced hippocampal load ofEB101 immunizedmice (Gr A) when comparedwith differentvaccine components (Gr B-C) In fact S1P appears to have ademonstrated therapeutic potential as a regenerative agent inthe nervous system since it controls migration of neuronalstem cells toward a site of spinal cord injury [36] inducecytoskeletal rearrangements to promote transmitter secretion[37] and also trigger diverse cellular effects including angio-genesis cardiac development immunity cell motility andneurite extension [38 39]Therefore it is reasonable to expectthat such a potent biologically active sphingolipid by itselfmay play an important role in the A120573 plaque reduction Inthis study the clearance efficiency rate of each treatment wasanalyzed and results suggested that EB101 (6459) appearsto be more effective than A120573
42+ CFAIFA (1920) vaccine
in both preventive and therapeutic groups The effectivenessof EB101 in older transgenic animals (18 months) showeda clearance effect of 59 suggesting that EB101 may alsohave therapeutic potential inADdiagnosticated patient trials
Similar conclusion was reached byWang and colleagues [40]who demonstrated that combination therapy in aged animalsprovides more valuable insights into a possible translationto humansOur immunologicalmeasurements demonstratedonce again that neuroinflammation develops in an A120573-dependent fashion early in the course of AD [41] and canbe alleviated or reversed by interventional A120573 vaccinationsuch as EB101 Among the possible mechanisms of A120573clearance proposed in AD immunotherapy [42] our resultssuggested that theymay overlap and probably be disease statedependent since the prevention and the therapeutic vaccinewould likely benefit from the transport of A120573 antibodies intothe CNS However some mechanisms of action such as thepreventive effect of A120573 antibodies in the A120573 aggregation [43]microglial phagocytosis induced by the Fc binding portionsbetween A120573 antibodies and microglia receptor [44] and theshift in the gradient of A120573 transport across the blood-brainbarrier due to the increased concentration of A120573 antibodiesin the vascular system resulting in an increase in effluxfrom the brain to blood [45 46] may be playing a role inboth preventive and therapeutic conditions Our findingstherefore suggest that novel treatment strategies aimed atmaintaining or increasing microglial function and restoringthe balance to the immunological system may represent anattractive therapeutic approach even at the advanced stagesof ADThe immunization results obtained with EB101 duringboth preventive and therapeutic treatment showed nomicro-hemorrhages infiltration of lymphocytes or encephalitis-like reaction in the brain suggesting that EB101 does notinduce A120573-specific Th1 reaction possibly due to the neu-roprotective effect of S1P Delivering A120573
1ndash42 in a liposomal-S1P-adjuvant resulted in an effective immunological responseincluding the induction of a Th2 T-cell response whicheffectively prevented neuroinflammation in an animal model
BioMed Research International 13
of AD as reported previously [24 47] Moreover presentresults demonstrated that EB101 did not induce astrogliosisin APPPS1 transgenic mice whereas some GFAP-positiveastrocyte clusters were aggregated in the affected brain areasof mice treated with A120573
42+ CFAIFA These results indicate
that EB101 may result in an effective treatment in AD neu-ropathology due to the absence of peripheral proinflamma-tory cytokine secretion and the absence of astroglial activa-tion in the brains of EB101 treated mice when compared withA12057342
+ CFAIFA immunotherapeutic response In a clinicalsetting however we may expect a wide differential responsein part depending upon the genomic profile of each patient[48 49] In AD behavioural studies in particular amongthe motor coordination tests the rotarod performance hasbeen reported to be impaired in some AD transgenic micemodels bearing A120573 plaques [50 51] This result was found inmutants prior to A120573 plaque formation at 3 and 6 months ofage but was not reproduced after plaques had been formedat 16 and 24 months of age [52] perhaps because of thedeclining abilities of older normal mice Here we presentthe effects of vaccine treatments in APPPS1 mice on motorcoordination a neglected aspect of the AD experimentalbehavior but justified on the basis of the deficient posturalcontrol reported in patients with Alzheimerrsquos disease [53]sometimes even during early stages [54] Our longitudinalmotor behavioral results showed no significant differencesin the ambulatory movement grip strength or coordinationaddressed by their performance in the rotating rod betweenAPPPS1 and wild-type mice for any age group studiedin contrast with other treated groups demonstrating thatlongitudinal motor assessments of AD transgenic mousemodels can provide valuable information with regard tothe effect of immunization on the progression of behavioraldeficits and overall motor activity As shown the EB101-treated group of mice exhibited longer latencies and betterbalancing skills indicating the absence of any negative effectof EB101 immunization on these mice The classical fearconditioning test is assumed to reveal the functional integrityof the hippocampus [55] representing a special memorytask to evaluate the AD models and its improvement afterimmunization periods In this APPPS1 mouse model thefirst associative learning and spatial workingmemory deficitsare shown at 5-6 months of age [56] although its impair-ment progression is relatively slow compared to most othermodels At the end of preventive immunization period EB101and A120573
42+ CFAIFA immunized mice have significantly
shorter escape latencies than the other treatment groups(A12057342liposome S1P-containing liposomes) and the controls
in the fear conditioning test Shorter escape latencies duringspatial learning indicated that deficits of spatial memorywere attenuated in EB101 and A120573
42+ CFAIFA immunized
mice Previous studies have showed that by 15 months ofage the spatial working memory is consistently impairedthroughout the rest of the life span [31 57] At the end oftherapeutic immunization period both EB101 and A120573
42+
CFAIFA immunized mice showed significantly improvedresults on the hippocampus-dependent spatial learning andmemory test when compared with other treatment groupsand the controls Taken together these observations are
in agreement with the impairment in conditioned learningin response to a tone stimulus of aged APPPS1 micealthough when immunized with EB101 during preventiveor therapeutic periods such impairments are significantlyattenuated
In summary the present results corroborate our previouspublished findings showing that EB101 immunization reducesamyloid accumulation in the APPPS1 mouse brain [2425] The strongest reduction in the preventive immunizationwas observed in the retrosplenial cortex followed by thehippocampus and entorhinal-piriform cortex Our resultsfurther support that EB101 ismore effectivewhen themice areimmunized prior to developing plaques in the hippocampaland cortical brain regions There is extensive evidence inmouse preclinical studies [58] and human clinical trials [5960] that the adverse effects of active and passive A120573 vaccinesresult in microhemorrhages and encephalitic reactions [60ndash63] yet themechanismbywhich they occur has not been fullyelucidated In the treatment sections analyzed we found nomicrohemorrhages or acute inflammatory processes in all thesurvived mice of the group indicating that EB101 therapeuticapproach presents no adverse inflammatory effects in thisAD mice models Thus our present results unequivocallydemonstrate that EB101 vaccinemay not only halt AD-relatedpathology as a preventive treatment but may also reverse it inestablished cases related to the improvement of the learningandmemory in both cases leading to a robust immunologicaltherapeutic response Present experiments also demonstratethat EB101 exerts more significant and substantial effects onthe reduction of A120573 deposition than A120573
42+CFAIFA vaccine
in A120573 mice models of AD However further exploration ofthe contribution of genetic environmental and physiologicalfactors in the development resistance to endogenous A120573-antibody is needed in order to optimize and modulate thetherapeutic strategies currently being developed
5 Conclusions
The present study carried out in APPPS1 transgenic mousemodel of AD compared the A120573 burden reduction andcellular immune responses produced by two immunizationdelivery strategies of A120573 in Freund adjuvant (described byWebster et al [10]) and in liposomeS1P matrix (describedby Schenk et al [17])The EB101 immunotherapeutic strategyboosted antibody generation A120573 plaque reduction and ananti-inflammatory response more effectively than A120573
42+
CFAIFA especially if given prior to or in the early stages ofpathology The present results also showed that vaccinationwith EB101 prevented the formation of new plaques andpresumably led to a reduction in established plaques whileinducing aTh2 immune reactionwithout encephalitogenic T-cell responses or brain inflammation related with astrogliosisHowever despite the efficacy demonstrated of this novelvaccine approach further preclinical and clinical studieson EB101 are warranted to evaluate its efficacy and themechanism for the future utility of such vaccine for ADtherapy
14 BioMed Research International
List of Abbreviations
AD Alzheimerrsquos diseaseA120573 120573-Amyloid proteinAPP Amyloid precursor proteinAPT Anterior pretectal nucleusCA1 Field CA1 hippocampusCA3 Field CA3 hippocampusCPu Caudate putamen (striatum)Cx CortexDG Dentate gyrusDN Dystrophic neuriteEct Entorhinal cortexCFA Complete Freundrsquos adjuvantGrDG Granular dentate gyrusIFA Incomplete Freundrsquos adjuvantIL InterleukinLPMR Lateral posterior thalamic nucleus (mediorostral)MoDG Molecular dentate gyrusPir Piriform cortexPoDG Polymorph dentate gyrusRSGb Retrosplenial granular cortex b regionTg TransgenicTh T helper
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Ivan Carrera Ignacio Etcheverrıa Lucıa Fernandez-NovoaValter Lombardi Ramon Cacabelos and Carmen Vigo con-ceived and designed the experiments Ivan Carrera and Igna-cio Etcheverrıa performed the experiments Ivan CarreraIgnacio Etcheverrıa and Valter Lombardi analyzed the dataIvan Carrera Ignacio Etcheverrıa Lucıa Fernandez-Novoaand Valter Lombardi contributed reagentsmaterialsanalysistools Ivan Carrera Carmen Vigo andMadepalli KrishnappaLakshmana wrote the paper
Acknowledgments
This work was supported by National Institute of Aging(NIA)NIH Grant no 1R01AG036859-01 (Madepalli Krish-nappa Lakshmana) and the financial support of the ProgramaOperativo Feder Galicia 2007ndash2013 (IN841C 201254 to PhDCarrera I)
References
[1] R Cacabelos L Fernandez-Novoa V Lombardi Y Kubotaand M Takeda ldquoMolecular genetics of Alzheimerrsquos diseaseand agingrdquoMethods and Findings in Experimental and ClinicalPharmacology vol 27 pp 1ndash573 2005
[2] P T Nelson I Alafuzoff E H Bigio et al ldquoCorrelationof alzheimer disease neuropathologic changes with cognitive
status a review of the literaturerdquo Journal of Neuropathology andExperimental Neurology vol 71 no 5 pp 362ndash381 2012
[3] JNaslundVHaroutunian RMohs et al ldquoCorrelation betweenelevated levels of amyloid 120573-peptide in the brain and cognitivedeclinerdquo Journal of the American Medical Association vol 283no 12 pp 1571ndash1577 2000
[4] A Koyama O I Okereke T Yang D Blacker D J Selkoeand F Grodstein ldquoPlasma amyloid-120573 as a predictor of dementiaand cognitive decline a systematic review and meta-analysisrdquoArchives of Neurology vol 69 no 7 pp 824ndash831 2012
[5] A M Fagan C M Roe C Xiong M A Mintun J C Morrisand D M Holtzman ldquoCerebrospinal fluid tau120573-amyloid42ratio as a prediction of cognitive decline in nondemented olderadultsrdquo Archives of Neurology vol 64 no 3 pp 343ndash349 2007
[6] S Parvathy P Davies V Haroutunian et al ldquoCorrelationbetween A120573-40- A120573-42- and A120573x-43-containing amyloidplaques and cognitive declinerdquo Archives of Neurology vol 58no 12 pp 2025ndash2032 2001
[7] E Karran M Mercken and B D Strooper ldquoThe amyloidcascade hypothesis for Alzheimerrsquos disease an appraisal for thedevelopment of therapeuticsrdquo Nature Reviews Drug Discoveryvol 10 no 9 pp 698ndash712 2011
[8] J L Jankowsky D J Fadale J Anderson et al ldquoMutantpresenilins specifically elevate the levels of the 42 residue 120573-amyloid peptide in vivo evidence for augmentation of a 42-specific 120574 secretaserdquo Human Molecular Genetics vol 13 no 2pp 159ndash170 2004
[9] L Holcomb M N Gordon E Mcgowan et al ldquoAcceleratedAlzheimer-type phenotype in transgenic mice carrying bothmutant amyloid precursor protein and presenilin 1 transgenesrdquoNature Medicine vol 4 no 1 pp 97ndash100 1998
[10] S JWebster A D Bachstetter and L J van Eldik ldquoComprehen-sive behavioral characterization of an APPPS-1 double knock-in mouse model of Alzheimerrsquos diseaserdquo Alzheimerrsquos ResearchandTherapy vol 5 no 3 article 28 2013
[11] H Xiong D Callaghan J Wodzinska et al ldquoBiochemical andbehavioral characterization of the double transgenic mousemodel (APPswePS1dE9) of Alzheimerrsquos diseaserdquo NeuroscienceBulletin vol 27 no 4 pp 221ndash232 2011
[12] M Garcia-Alloza L A Borrelli B T Hyman and B J BacskaildquoAntioxidants have a rapid and long-lasting effect on neuriticabnormalities in APPPS1 micerdquo Neurobiology of Aging vol 31no 12 pp 2058ndash2068 2010
[13] M Meyer-Luehmann M Mielke T L Spires-Jones et al ldquoAreporter of local dendritic translocation shows plaque-relatedloss of neural system function in APP-transgenic micerdquo Journalof Neuroscience vol 29 no 40 pp 12636ndash12640 2009
[14] E Machova V Rudajev H Smyckova H Koivisto H Tanilaand V Dolezal ldquoFunctional cholinergic damage develops withamyloid accumulation in young adult APPswePS1dE9 trans-genic micerdquo Neurobiology of Disease vol 38 no 1 pp 27ndash352010
[15] T Malm J Koistinaho and K Kanninen ldquoUtilization of APP-swePS1dE9 transgenic mice in research of Alzheimerrsquos diseasefocus on gene therapy and cell-based therapy applicationsrdquoInternational Journal of Alzheimerrsquos Disease vol 2011 Article ID517160 8 pages 2011
[16] J Lee and P Han ldquoAn update of animal models of alzheimerdisease with a reevaluation of plaque depositionsrdquoExperimentalNeurobiology vol 22 pp 84ndash95 2013
BioMed Research International 15
[17] D Schenk R Barbour W Dunn et al ldquoImmunization withamyloid-120573 attenuates Alzheimer disease-like pathology in thePDAPP mouserdquo Nature vol 400 no 6740 pp 173ndash177 1999
[18] H J Fu B Liu J L Frost and C A Lemere ldquoAmyloid-120573immunotherapy for Alzheimerrsquos diseaserdquoCNS and NeurologicalDisorders-Drug Targets vol 9 no 2 pp 197ndash206 2010
[19] J-M Orgogozo S Gilman J-F Dartigues et al ldquoSubacutemeningoencephalitis in a subset of patients with AD after A120573
42
immunizationrdquo Neurology vol 61 no 1 pp 46ndash54 2003[20] J A R Nicolll DWilkinson CHolmes P Steart HMarkham
andROWeller ldquoNeuropathology of humanAlzheimer diseaseafter immunization with amyloid-120573 peptide a case reportrdquoNature Medicine vol 9 no 4 pp 448ndash452 2003
[21] I Ferrer M Boada Rovira M L Sanchez Guerra M JRey and F Costa-Jussa ldquoNeuropathology and pathogenesisof encephalitis following amyloid-Neuropathology and patho-genesis of encephalitis following amyloid-beta immunizationin Alzheimerrsquos disease immunization in Alzheimerrsquos diseaserdquoBrain Pathology vol 14 no 1 pp 11ndash20 2004
[22] D H Cribbs A Ghochikyan V Vasilevko et al ldquoAdjuvant-dependent modulation ofTh1 andTh2 responses to immuniza-tion with 120573-amyloidrdquo International Immunology vol 15 no 4pp 505ndash514 2003
[23] C Holmes D Boche D Wilkinson et al ldquoLong-term effectsof A120573
42immunisation in Alzheimerrsquos disease follow-up of a
randomised placebo-controlled phase I trialrdquo The Lancet vol372 no 9634 pp 216ndash223 2008
[24] I Carrera I Etcheverrıa L Fernandez-Novoa V LombardiR Cacabelos and C Vigo ldquoVaccine development to treatAlzheimerrsquos disease neuropathology in APPPS1 transgenicmicerdquo International Journal of Alzheimerrsquos Disease vol 2012Article ID 376138 17 pages 2012
[25] I Carrera I Etcheverrıa Y Li et al ldquoImmunocytochemicalcharacterization of Alzheimer disease hallmarks in APPPS1transgenic mice treated with a new anti-amyloid-120573 vaccinerdquoBioMed Research International vol 2013 Article ID 709145 12pages 2013
[26] V Anelli R Bassi G Tettamanti P Viani and L Riboni ldquoExtra-cellular release of newly synthesized sphingosine-1-phosphateby cerebellar granule cells and astrocytesrdquo Journal of Neuro-chemistry vol 92 no 5 pp 1204ndash1215 2005
[27] S Schroeter K Khan R Barbour et al ldquoImmunotherapyreduces vascular amyloid-120573 in PDAPP micerdquo Journal of Neu-roscience vol 28 no 27 pp 6787ndash6793 2008
[28] Z Suo A Cox N Bartelli et al ldquoGRK5 deficiency leads to earlyAlzheimer-like pathology and working memory impairmentrdquoNeurobiology of Aging vol 28 no 12 pp 1873ndash1888 2007
[29] W Kamphuis C Mamber MMoeton et al ldquoGFAP isoforms inadult mouse brain with a focus on neurogenic astrocytes andreactive astrogliosis in mouse models of Alzheimer diseaserdquoPLoS ONE vol 7 no 8 Article ID e42823 2012
[30] J Lang C Vigo-Pelfrey and F Martin ldquoLiposomes composedof partially hydrogenated egg phosphatidylcholines fatty acidcomposition thermal phase behavior and oxidative stabilityrdquoChemistry and Physics of Lipids vol 53 no 1 pp 91ndash101 1990
[31] D Morgan D M Diamond P E Gottschall et al ldquoA 120573peptide vaccination prevents memory loss in an animal modelof Alzheimerrsquos diseaserdquo Nature vol 408 no 6815 pp 982ndash9852000
[32] M L Brownlow L Benner D DrsquoAgostino M N Gordon andD Morgan ldquoKetogenic diet improves motor performance but
not cognition in two mouse models of Alzheimerrsquos pathologyrdquoPLoS ONE vol 8 no 9 Article ID e75713 2013
[33] M Pride P Seubert M Grundman M Hagen J Eldridgeand R S Black ldquoProgress in the active immunotherapeu-tic approach to Alzheimerrsquos disease clinical investigationsinto AN1792-associated meningoencephalitisrdquo Neurodegenera-tive Diseases vol 5 no 3-4 pp 194ndash196 2008
[34] CWiessner K-HWiederhold A C Tissot et al ldquoThe second-generation activeA120573 immunotherapyCAD106 reduces amyloidaccumulation in APP transgenic mice while minimizing poten-tial side effectsrdquoThe Journal of Neuroscience vol 31 no 25 pp9323ndash9331 2011
[35] D Lambracht-Washington and R Rosenberg ldquoAdvances in thedevelopment of vaccines for Alzheimerrsquos diseaserdquo DiscoveryMedicine vol 15 no 84 pp 319ndash326 2013
[36] A Kimura T Ohmori R Ohkawa et al ldquoEssential roles ofsphingosine 1-phosphateS1P
1receptor axis in the migration of
neural stem cells toward a site of spinal cord injuryrdquo Stem Cellsvol 25 no 1 pp 115ndash124 2007
[37] S S Lee C H Woo J D Chang and J H Kim ldquoRoles of Racand cytosolic phospholipase A2 in the intracellular signalling inresponse to titanium particlesrdquo Cellular Signalling vol 15 no 3pp 339ndash345 2003
[38] E Masliah L Hansen A Adame et al ldquoA120573 vaccination effectson plaque pathology in the absence of encephalitis inAlzheimerdiseaserdquo Neurology vol 64 no 1 pp 129ndash131 2005
[39] A Pilorget B Annabi F Bouzeghrane J Marvaldi J Luisand R Beliveau ldquoInhibition of angiogenic properties of brainendothelial cells by platelet-derived sphingosine-1-phosphaterdquoJournal of Cerebral Blood Flow and Metabolism vol 25 no 9pp 1171ndash1182 2005
[40] A Wang P Das R C Switzer III T E Golde and J LJankowsky ldquoRobust amyloid clearance in a mouse model ofAlzheimerrsquos disease provides novel insights into themechanismof amyloid-120573 immunotherapyrdquo Journal of Neuroscience vol 31no 11 pp 4124ndash4136 2011
[41] G Krabbe A Halle V Matyash et al ldquoFunctional impairmentof microglia coincides with Beta-amyloid deposition in micewith Alzheimer-like pathologyrdquo PLoS ONE vol 8 no 4 ArticleID e60921 2013
[42] Y H Liu B Giunta H-D Zhou J Tan and Y J WangldquoImmunotherapy for Alzheimer disease-the challenge ofadverse effectsrdquo Nature Reviews Neurology vol 8 no 8 pp465ndash469 2012
[43] B Solomon R Koppel D Frankel and E Hanan-AharonldquoDisaggregation of Alzheimer120573-amyloid by site-directedmAbrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 94 no 8 pp 4109ndash4112 1997
[44] F Bard C Cannon R Barbour et al ldquoPeripherally adminis-tered antibodies against amyloid 120573-peptide enter the centralnervous system and reduce pathology in a mouse model ofAlzheimer diseaserdquo Nature Medicine vol 6 no 8 pp 916ndash9192000
[45] R B DeMattos K R Bales D J Cummins J C Dodart S MPaul and D M Holtzman ldquoPeripheral anti-A120573 antibody altersCNS and plasma A120573 clearance and decreases brain A120573 burdenin a mouse model of Alzheimerrsquos diseaserdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 98 no 15 pp 8850ndash8855 2001
[46] C A Lemere E T Spooner J LaFrancois et al ldquoEvidence forperipheral clearance of cerebral A120573 protein following chronic
16 BioMed Research International
active A120573 immunization in PSAPP micerdquo Neurobiology ofDisease vol 14 no 1 pp 10ndash18 2003
[47] T Town M Vendrame A Patel et al ldquoReduced Th1 andenhanced Th2 immunity after immunization with Alzheimerrsquos120573-amyloid
1minus42rdquo Journal of Neuroimmunology vol 132 no 1-2
pp 49ndash59 2002[48] R Cacabelos ldquoPharmacogenomics in Alzheimerrsquos diseaserdquo
Methods in Molecular Biology vol 448 pp 213ndash357 2008[49] R Cacabelos ldquoPharmacogenomics and therapeutic strategies
for dementiardquo Expert Review of Molecular Diagnostics vol 9no 6 pp 567ndash611 2009
[50] D van Dam R DrsquoHooge M Staufenbiel C van Ginneken Fvan Meir and P P de Deyn ldquoAge-dependent cognitive declinein the APP23 model precedes amyloid depositionrdquo EuropeanJournal of Neuroscience vol 17 no 2 pp 388ndash396 2003
[51] M Dumont C Strazielle M Staufenbiel and R LalondeldquoSpatial learning and exploration of environmental stimuli in24-month-old female APP23 transgenic mice with the Swedishmutationrdquo Brain Research vol 1024 no 1-2 pp 113ndash121 2004
[52] R Lalonde M Dumont M Staufenbiel C Sturchler-Pierratand C Strazielle ldquoSpatial learning exploration anxiety andmotor coordination in female APP23 transgenic mice with theSwedish mutationrdquo Brain Research vol 956 no 1 pp 36ndash442002
[53] F J Huff F Boller F Lucchelli R Querriera J Beyer and SBelle ldquoThe neurologic examination in patients with probableAlzheimerrsquos diseaserdquo Archives of Neurology vol 44 no 9 pp929ndash932 1987
[54] A F Pettersson M Engardt and L O Wahlund ldquoActivitylevel and balance in subjects with mild Alzheimerrsquos diseaserdquoDementia and Geriatric Cognitive Disorders vol 13 no 4 pp213ndash216 2002
[55] L Calandreau B Desgranges R Jaffard and A DesmedtldquoSwitching from contextual to tone fear conditioning and viceversa the key role of the glutamatergic hippocampal-lateralseptal neurotransmissionrdquo Learning amp Memory vol 17 no 9pp 440ndash443 2010
[56] F Trinchese S Liu F Battaglia SWalter PMMathews andOArancio ldquoProgressive age-related development of Alzheimer-like pathology in APPPS1 micerdquo Annals of Neurology vol 55no 6 pp 801ndash814 2004
[57] D M Wilcock A Rojiani A Rosenthal et al ldquoPassiveimmunotherapy against A120573 in aged APP-transgenic micereverses cognitive deficits and depletes parenchymal amyloiddeposits in spite of increased vascular amyloid and microhem-orrhagerdquo Journal of Neuroinflammation vol 1 article 24 2004
[58] S A Ferguson S Sarkar and L C Schmued ldquoLongitudinalbehavioral changes in the APPPS1 transgenic AlzheimerrsquosDisease modelrdquo Behavioural Brain Research vol 242 no 1 pp125ndash134 2013
[59] D Wilcock and C Colton ldquoImmunotherapy vascular pathol-ogy and microhemorrhages in transgenic micerdquo CNS andNeurological DisordersmdashDrug Targets vol 8 no 1 pp 50ndash642009
[60] M Grundman S Gilman R S Black et al ldquoAn alternativemethod for estimating efficacy of the AN1792 vaccine foralzheimer diseaserdquo Neurology vol 71 no 9 pp 697ndash698 2008
[61] D Boche E Zotova R O Weller et al ldquoConsequence of A120573immunization on the vasculature of human Alzheimerrsquos diseasebrainrdquo Brain vol 131 no 12 pp 3299ndash3310 2008
[62] M Pfeifer S Boncristiano L Bondolfi et al ldquoCerebral hemor-rhage after passive anti-a120573 immunotherapyrdquo Science vol 298no 5597 p 1379 2002
[63] M M Racke L I Boone D L Hepburn et al ldquoExacerbationof cerebral amyloid angiopathy-associatedmicrohemorrhage inamyloid precursor protein transgenic mice by immunotherapyis dependent on antibody recognition of deposited forms ofamyloid 120573rdquo Journal of Neuroscience vol 25 no 3 pp 629ndash6362005
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

12 BioMed Research International
Gr A
(EB1
01)
05
101520253035
Preventive treatment Therapeutic treatment
Mea
n fa
lling
late
ncy
(s)
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)(a) Graphic of motor coordination and balance
05
1015202530
Mea
n re
spon
se ti
me (
s)
Gr A
(EB1
01)
Preventive treatment Therapeutic treatment
Gr E
(PBS
)
Gr B
(A120573+
L)
Gr C
(S1P
+L)
Gr D
(A120573+
CFA
IFA
)
(b) Graphic of associative learning and memory
Figure 8 Analysis of motor coordination and balance (a) Rotarod results of motor coordination and balance test after the preventive andtherapeutic treatment showing improved effect of EB101 and A120573
42+ CFAIFA-vaccine compared to other treated mice groups (119875 lt 005)
and (119875 lt 002) versus control mice Latency to fall on the rotarod apparatus was significantly improved in EB101 immunized mice Valuesrepresent means of all six trials of a groupday (b) Spatial learning and memory tested by fear conditioning-active avoidance test The meanresponse time of each treatment group to reach the platform is represented EB101 and A120573
42+ CFAIFA immunized mice had significantly
lower latencies than the other treatment groups (119875 lt 001) versus control mice
efficient than that obtained with A12057342+ CFAIFA immuniza-
tions A comparison of A120573 plaque reduction rates amongtreated groups surprisingly showed that other than EB101and A120573
42+ CFAIFA treated groups some amyloid deposits
were also reduced in Groups B (A1205731ndash42liposome) and C
(S1P-containing liposomes) Although the reduction rates inthese mice were markedly low the subtle lowering effectmight be due to the interaction between the immunizationelements (A120573
1ndash42 and S1P) and the mouse immunogenicsystem as reported by others [35] Based on our presentresults S1P one of the components of the adjuvant used withEB101 might be responsible for the neuroprotective effectobserved in the vulnerable neuronal areas of EB101 treatedmice The advantage in the A120573 load preventionreduction bycombining A120573
1ndash42 and S1P in a unique formulation vaccinehas been also observed in the reduced hippocampal load ofEB101 immunizedmice (Gr A) when comparedwith differentvaccine components (Gr B-C) In fact S1P appears to have ademonstrated therapeutic potential as a regenerative agent inthe nervous system since it controls migration of neuronalstem cells toward a site of spinal cord injury [36] inducecytoskeletal rearrangements to promote transmitter secretion[37] and also trigger diverse cellular effects including angio-genesis cardiac development immunity cell motility andneurite extension [38 39]Therefore it is reasonable to expectthat such a potent biologically active sphingolipid by itselfmay play an important role in the A120573 plaque reduction Inthis study the clearance efficiency rate of each treatment wasanalyzed and results suggested that EB101 (6459) appearsto be more effective than A120573
42+ CFAIFA (1920) vaccine
in both preventive and therapeutic groups The effectivenessof EB101 in older transgenic animals (18 months) showeda clearance effect of 59 suggesting that EB101 may alsohave therapeutic potential inADdiagnosticated patient trials
Similar conclusion was reached byWang and colleagues [40]who demonstrated that combination therapy in aged animalsprovides more valuable insights into a possible translationto humansOur immunologicalmeasurements demonstratedonce again that neuroinflammation develops in an A120573-dependent fashion early in the course of AD [41] and canbe alleviated or reversed by interventional A120573 vaccinationsuch as EB101 Among the possible mechanisms of A120573clearance proposed in AD immunotherapy [42] our resultssuggested that theymay overlap and probably be disease statedependent since the prevention and the therapeutic vaccinewould likely benefit from the transport of A120573 antibodies intothe CNS However some mechanisms of action such as thepreventive effect of A120573 antibodies in the A120573 aggregation [43]microglial phagocytosis induced by the Fc binding portionsbetween A120573 antibodies and microglia receptor [44] and theshift in the gradient of A120573 transport across the blood-brainbarrier due to the increased concentration of A120573 antibodiesin the vascular system resulting in an increase in effluxfrom the brain to blood [45 46] may be playing a role inboth preventive and therapeutic conditions Our findingstherefore suggest that novel treatment strategies aimed atmaintaining or increasing microglial function and restoringthe balance to the immunological system may represent anattractive therapeutic approach even at the advanced stagesof ADThe immunization results obtained with EB101 duringboth preventive and therapeutic treatment showed nomicro-hemorrhages infiltration of lymphocytes or encephalitis-like reaction in the brain suggesting that EB101 does notinduce A120573-specific Th1 reaction possibly due to the neu-roprotective effect of S1P Delivering A120573
1ndash42 in a liposomal-S1P-adjuvant resulted in an effective immunological responseincluding the induction of a Th2 T-cell response whicheffectively prevented neuroinflammation in an animal model
BioMed Research International 13
of AD as reported previously [24 47] Moreover presentresults demonstrated that EB101 did not induce astrogliosisin APPPS1 transgenic mice whereas some GFAP-positiveastrocyte clusters were aggregated in the affected brain areasof mice treated with A120573
42+ CFAIFA These results indicate
that EB101 may result in an effective treatment in AD neu-ropathology due to the absence of peripheral proinflamma-tory cytokine secretion and the absence of astroglial activa-tion in the brains of EB101 treated mice when compared withA12057342
+ CFAIFA immunotherapeutic response In a clinicalsetting however we may expect a wide differential responsein part depending upon the genomic profile of each patient[48 49] In AD behavioural studies in particular amongthe motor coordination tests the rotarod performance hasbeen reported to be impaired in some AD transgenic micemodels bearing A120573 plaques [50 51] This result was found inmutants prior to A120573 plaque formation at 3 and 6 months ofage but was not reproduced after plaques had been formedat 16 and 24 months of age [52] perhaps because of thedeclining abilities of older normal mice Here we presentthe effects of vaccine treatments in APPPS1 mice on motorcoordination a neglected aspect of the AD experimentalbehavior but justified on the basis of the deficient posturalcontrol reported in patients with Alzheimerrsquos disease [53]sometimes even during early stages [54] Our longitudinalmotor behavioral results showed no significant differencesin the ambulatory movement grip strength or coordinationaddressed by their performance in the rotating rod betweenAPPPS1 and wild-type mice for any age group studiedin contrast with other treated groups demonstrating thatlongitudinal motor assessments of AD transgenic mousemodels can provide valuable information with regard tothe effect of immunization on the progression of behavioraldeficits and overall motor activity As shown the EB101-treated group of mice exhibited longer latencies and betterbalancing skills indicating the absence of any negative effectof EB101 immunization on these mice The classical fearconditioning test is assumed to reveal the functional integrityof the hippocampus [55] representing a special memorytask to evaluate the AD models and its improvement afterimmunization periods In this APPPS1 mouse model thefirst associative learning and spatial workingmemory deficitsare shown at 5-6 months of age [56] although its impair-ment progression is relatively slow compared to most othermodels At the end of preventive immunization period EB101and A120573
42+ CFAIFA immunized mice have significantly
shorter escape latencies than the other treatment groups(A12057342liposome S1P-containing liposomes) and the controls
in the fear conditioning test Shorter escape latencies duringspatial learning indicated that deficits of spatial memorywere attenuated in EB101 and A120573
42+ CFAIFA immunized
mice Previous studies have showed that by 15 months ofage the spatial working memory is consistently impairedthroughout the rest of the life span [31 57] At the end oftherapeutic immunization period both EB101 and A120573
42+
CFAIFA immunized mice showed significantly improvedresults on the hippocampus-dependent spatial learning andmemory test when compared with other treatment groupsand the controls Taken together these observations are
in agreement with the impairment in conditioned learningin response to a tone stimulus of aged APPPS1 micealthough when immunized with EB101 during preventiveor therapeutic periods such impairments are significantlyattenuated
In summary the present results corroborate our previouspublished findings showing that EB101 immunization reducesamyloid accumulation in the APPPS1 mouse brain [2425] The strongest reduction in the preventive immunizationwas observed in the retrosplenial cortex followed by thehippocampus and entorhinal-piriform cortex Our resultsfurther support that EB101 ismore effectivewhen themice areimmunized prior to developing plaques in the hippocampaland cortical brain regions There is extensive evidence inmouse preclinical studies [58] and human clinical trials [5960] that the adverse effects of active and passive A120573 vaccinesresult in microhemorrhages and encephalitic reactions [60ndash63] yet themechanismbywhich they occur has not been fullyelucidated In the treatment sections analyzed we found nomicrohemorrhages or acute inflammatory processes in all thesurvived mice of the group indicating that EB101 therapeuticapproach presents no adverse inflammatory effects in thisAD mice models Thus our present results unequivocallydemonstrate that EB101 vaccinemay not only halt AD-relatedpathology as a preventive treatment but may also reverse it inestablished cases related to the improvement of the learningandmemory in both cases leading to a robust immunologicaltherapeutic response Present experiments also demonstratethat EB101 exerts more significant and substantial effects onthe reduction of A120573 deposition than A120573
42+CFAIFA vaccine
in A120573 mice models of AD However further exploration ofthe contribution of genetic environmental and physiologicalfactors in the development resistance to endogenous A120573-antibody is needed in order to optimize and modulate thetherapeutic strategies currently being developed
5 Conclusions
The present study carried out in APPPS1 transgenic mousemodel of AD compared the A120573 burden reduction andcellular immune responses produced by two immunizationdelivery strategies of A120573 in Freund adjuvant (described byWebster et al [10]) and in liposomeS1P matrix (describedby Schenk et al [17])The EB101 immunotherapeutic strategyboosted antibody generation A120573 plaque reduction and ananti-inflammatory response more effectively than A120573
42+
CFAIFA especially if given prior to or in the early stages ofpathology The present results also showed that vaccinationwith EB101 prevented the formation of new plaques andpresumably led to a reduction in established plaques whileinducing aTh2 immune reactionwithout encephalitogenic T-cell responses or brain inflammation related with astrogliosisHowever despite the efficacy demonstrated of this novelvaccine approach further preclinical and clinical studieson EB101 are warranted to evaluate its efficacy and themechanism for the future utility of such vaccine for ADtherapy
14 BioMed Research International
List of Abbreviations
AD Alzheimerrsquos diseaseA120573 120573-Amyloid proteinAPP Amyloid precursor proteinAPT Anterior pretectal nucleusCA1 Field CA1 hippocampusCA3 Field CA3 hippocampusCPu Caudate putamen (striatum)Cx CortexDG Dentate gyrusDN Dystrophic neuriteEct Entorhinal cortexCFA Complete Freundrsquos adjuvantGrDG Granular dentate gyrusIFA Incomplete Freundrsquos adjuvantIL InterleukinLPMR Lateral posterior thalamic nucleus (mediorostral)MoDG Molecular dentate gyrusPir Piriform cortexPoDG Polymorph dentate gyrusRSGb Retrosplenial granular cortex b regionTg TransgenicTh T helper
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Ivan Carrera Ignacio Etcheverrıa Lucıa Fernandez-NovoaValter Lombardi Ramon Cacabelos and Carmen Vigo con-ceived and designed the experiments Ivan Carrera and Igna-cio Etcheverrıa performed the experiments Ivan CarreraIgnacio Etcheverrıa and Valter Lombardi analyzed the dataIvan Carrera Ignacio Etcheverrıa Lucıa Fernandez-Novoaand Valter Lombardi contributed reagentsmaterialsanalysistools Ivan Carrera Carmen Vigo andMadepalli KrishnappaLakshmana wrote the paper
Acknowledgments
This work was supported by National Institute of Aging(NIA)NIH Grant no 1R01AG036859-01 (Madepalli Krish-nappa Lakshmana) and the financial support of the ProgramaOperativo Feder Galicia 2007ndash2013 (IN841C 201254 to PhDCarrera I)
References
[1] R Cacabelos L Fernandez-Novoa V Lombardi Y Kubotaand M Takeda ldquoMolecular genetics of Alzheimerrsquos diseaseand agingrdquoMethods and Findings in Experimental and ClinicalPharmacology vol 27 pp 1ndash573 2005
[2] P T Nelson I Alafuzoff E H Bigio et al ldquoCorrelationof alzheimer disease neuropathologic changes with cognitive
status a review of the literaturerdquo Journal of Neuropathology andExperimental Neurology vol 71 no 5 pp 362ndash381 2012
[3] JNaslundVHaroutunian RMohs et al ldquoCorrelation betweenelevated levels of amyloid 120573-peptide in the brain and cognitivedeclinerdquo Journal of the American Medical Association vol 283no 12 pp 1571ndash1577 2000
[4] A Koyama O I Okereke T Yang D Blacker D J Selkoeand F Grodstein ldquoPlasma amyloid-120573 as a predictor of dementiaand cognitive decline a systematic review and meta-analysisrdquoArchives of Neurology vol 69 no 7 pp 824ndash831 2012
[5] A M Fagan C M Roe C Xiong M A Mintun J C Morrisand D M Holtzman ldquoCerebrospinal fluid tau120573-amyloid42ratio as a prediction of cognitive decline in nondemented olderadultsrdquo Archives of Neurology vol 64 no 3 pp 343ndash349 2007
[6] S Parvathy P Davies V Haroutunian et al ldquoCorrelationbetween A120573-40- A120573-42- and A120573x-43-containing amyloidplaques and cognitive declinerdquo Archives of Neurology vol 58no 12 pp 2025ndash2032 2001
[7] E Karran M Mercken and B D Strooper ldquoThe amyloidcascade hypothesis for Alzheimerrsquos disease an appraisal for thedevelopment of therapeuticsrdquo Nature Reviews Drug Discoveryvol 10 no 9 pp 698ndash712 2011
[8] J L Jankowsky D J Fadale J Anderson et al ldquoMutantpresenilins specifically elevate the levels of the 42 residue 120573-amyloid peptide in vivo evidence for augmentation of a 42-specific 120574 secretaserdquo Human Molecular Genetics vol 13 no 2pp 159ndash170 2004
[9] L Holcomb M N Gordon E Mcgowan et al ldquoAcceleratedAlzheimer-type phenotype in transgenic mice carrying bothmutant amyloid precursor protein and presenilin 1 transgenesrdquoNature Medicine vol 4 no 1 pp 97ndash100 1998
[10] S JWebster A D Bachstetter and L J van Eldik ldquoComprehen-sive behavioral characterization of an APPPS-1 double knock-in mouse model of Alzheimerrsquos diseaserdquo Alzheimerrsquos ResearchandTherapy vol 5 no 3 article 28 2013
[11] H Xiong D Callaghan J Wodzinska et al ldquoBiochemical andbehavioral characterization of the double transgenic mousemodel (APPswePS1dE9) of Alzheimerrsquos diseaserdquo NeuroscienceBulletin vol 27 no 4 pp 221ndash232 2011
[12] M Garcia-Alloza L A Borrelli B T Hyman and B J BacskaildquoAntioxidants have a rapid and long-lasting effect on neuriticabnormalities in APPPS1 micerdquo Neurobiology of Aging vol 31no 12 pp 2058ndash2068 2010
[13] M Meyer-Luehmann M Mielke T L Spires-Jones et al ldquoAreporter of local dendritic translocation shows plaque-relatedloss of neural system function in APP-transgenic micerdquo Journalof Neuroscience vol 29 no 40 pp 12636ndash12640 2009
[14] E Machova V Rudajev H Smyckova H Koivisto H Tanilaand V Dolezal ldquoFunctional cholinergic damage develops withamyloid accumulation in young adult APPswePS1dE9 trans-genic micerdquo Neurobiology of Disease vol 38 no 1 pp 27ndash352010
[15] T Malm J Koistinaho and K Kanninen ldquoUtilization of APP-swePS1dE9 transgenic mice in research of Alzheimerrsquos diseasefocus on gene therapy and cell-based therapy applicationsrdquoInternational Journal of Alzheimerrsquos Disease vol 2011 Article ID517160 8 pages 2011
[16] J Lee and P Han ldquoAn update of animal models of alzheimerdisease with a reevaluation of plaque depositionsrdquoExperimentalNeurobiology vol 22 pp 84ndash95 2013
BioMed Research International 15
[17] D Schenk R Barbour W Dunn et al ldquoImmunization withamyloid-120573 attenuates Alzheimer disease-like pathology in thePDAPP mouserdquo Nature vol 400 no 6740 pp 173ndash177 1999
[18] H J Fu B Liu J L Frost and C A Lemere ldquoAmyloid-120573immunotherapy for Alzheimerrsquos diseaserdquoCNS and NeurologicalDisorders-Drug Targets vol 9 no 2 pp 197ndash206 2010
[19] J-M Orgogozo S Gilman J-F Dartigues et al ldquoSubacutemeningoencephalitis in a subset of patients with AD after A120573
42
immunizationrdquo Neurology vol 61 no 1 pp 46ndash54 2003[20] J A R Nicolll DWilkinson CHolmes P Steart HMarkham
andROWeller ldquoNeuropathology of humanAlzheimer diseaseafter immunization with amyloid-120573 peptide a case reportrdquoNature Medicine vol 9 no 4 pp 448ndash452 2003
[21] I Ferrer M Boada Rovira M L Sanchez Guerra M JRey and F Costa-Jussa ldquoNeuropathology and pathogenesisof encephalitis following amyloid-Neuropathology and patho-genesis of encephalitis following amyloid-beta immunizationin Alzheimerrsquos disease immunization in Alzheimerrsquos diseaserdquoBrain Pathology vol 14 no 1 pp 11ndash20 2004
[22] D H Cribbs A Ghochikyan V Vasilevko et al ldquoAdjuvant-dependent modulation ofTh1 andTh2 responses to immuniza-tion with 120573-amyloidrdquo International Immunology vol 15 no 4pp 505ndash514 2003
[23] C Holmes D Boche D Wilkinson et al ldquoLong-term effectsof A120573
42immunisation in Alzheimerrsquos disease follow-up of a
randomised placebo-controlled phase I trialrdquo The Lancet vol372 no 9634 pp 216ndash223 2008
[24] I Carrera I Etcheverrıa L Fernandez-Novoa V LombardiR Cacabelos and C Vigo ldquoVaccine development to treatAlzheimerrsquos disease neuropathology in APPPS1 transgenicmicerdquo International Journal of Alzheimerrsquos Disease vol 2012Article ID 376138 17 pages 2012
[25] I Carrera I Etcheverrıa Y Li et al ldquoImmunocytochemicalcharacterization of Alzheimer disease hallmarks in APPPS1transgenic mice treated with a new anti-amyloid-120573 vaccinerdquoBioMed Research International vol 2013 Article ID 709145 12pages 2013
[26] V Anelli R Bassi G Tettamanti P Viani and L Riboni ldquoExtra-cellular release of newly synthesized sphingosine-1-phosphateby cerebellar granule cells and astrocytesrdquo Journal of Neuro-chemistry vol 92 no 5 pp 1204ndash1215 2005
[27] S Schroeter K Khan R Barbour et al ldquoImmunotherapyreduces vascular amyloid-120573 in PDAPP micerdquo Journal of Neu-roscience vol 28 no 27 pp 6787ndash6793 2008
[28] Z Suo A Cox N Bartelli et al ldquoGRK5 deficiency leads to earlyAlzheimer-like pathology and working memory impairmentrdquoNeurobiology of Aging vol 28 no 12 pp 1873ndash1888 2007
[29] W Kamphuis C Mamber MMoeton et al ldquoGFAP isoforms inadult mouse brain with a focus on neurogenic astrocytes andreactive astrogliosis in mouse models of Alzheimer diseaserdquoPLoS ONE vol 7 no 8 Article ID e42823 2012
[30] J Lang C Vigo-Pelfrey and F Martin ldquoLiposomes composedof partially hydrogenated egg phosphatidylcholines fatty acidcomposition thermal phase behavior and oxidative stabilityrdquoChemistry and Physics of Lipids vol 53 no 1 pp 91ndash101 1990
[31] D Morgan D M Diamond P E Gottschall et al ldquoA 120573peptide vaccination prevents memory loss in an animal modelof Alzheimerrsquos diseaserdquo Nature vol 408 no 6815 pp 982ndash9852000
[32] M L Brownlow L Benner D DrsquoAgostino M N Gordon andD Morgan ldquoKetogenic diet improves motor performance but
not cognition in two mouse models of Alzheimerrsquos pathologyrdquoPLoS ONE vol 8 no 9 Article ID e75713 2013
[33] M Pride P Seubert M Grundman M Hagen J Eldridgeand R S Black ldquoProgress in the active immunotherapeu-tic approach to Alzheimerrsquos disease clinical investigationsinto AN1792-associated meningoencephalitisrdquo Neurodegenera-tive Diseases vol 5 no 3-4 pp 194ndash196 2008
[34] CWiessner K-HWiederhold A C Tissot et al ldquoThe second-generation activeA120573 immunotherapyCAD106 reduces amyloidaccumulation in APP transgenic mice while minimizing poten-tial side effectsrdquoThe Journal of Neuroscience vol 31 no 25 pp9323ndash9331 2011
[35] D Lambracht-Washington and R Rosenberg ldquoAdvances in thedevelopment of vaccines for Alzheimerrsquos diseaserdquo DiscoveryMedicine vol 15 no 84 pp 319ndash326 2013
[36] A Kimura T Ohmori R Ohkawa et al ldquoEssential roles ofsphingosine 1-phosphateS1P
1receptor axis in the migration of
neural stem cells toward a site of spinal cord injuryrdquo Stem Cellsvol 25 no 1 pp 115ndash124 2007
[37] S S Lee C H Woo J D Chang and J H Kim ldquoRoles of Racand cytosolic phospholipase A2 in the intracellular signalling inresponse to titanium particlesrdquo Cellular Signalling vol 15 no 3pp 339ndash345 2003
[38] E Masliah L Hansen A Adame et al ldquoA120573 vaccination effectson plaque pathology in the absence of encephalitis inAlzheimerdiseaserdquo Neurology vol 64 no 1 pp 129ndash131 2005
[39] A Pilorget B Annabi F Bouzeghrane J Marvaldi J Luisand R Beliveau ldquoInhibition of angiogenic properties of brainendothelial cells by platelet-derived sphingosine-1-phosphaterdquoJournal of Cerebral Blood Flow and Metabolism vol 25 no 9pp 1171ndash1182 2005
[40] A Wang P Das R C Switzer III T E Golde and J LJankowsky ldquoRobust amyloid clearance in a mouse model ofAlzheimerrsquos disease provides novel insights into themechanismof amyloid-120573 immunotherapyrdquo Journal of Neuroscience vol 31no 11 pp 4124ndash4136 2011
[41] G Krabbe A Halle V Matyash et al ldquoFunctional impairmentof microglia coincides with Beta-amyloid deposition in micewith Alzheimer-like pathologyrdquo PLoS ONE vol 8 no 4 ArticleID e60921 2013
[42] Y H Liu B Giunta H-D Zhou J Tan and Y J WangldquoImmunotherapy for Alzheimer disease-the challenge ofadverse effectsrdquo Nature Reviews Neurology vol 8 no 8 pp465ndash469 2012
[43] B Solomon R Koppel D Frankel and E Hanan-AharonldquoDisaggregation of Alzheimer120573-amyloid by site-directedmAbrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 94 no 8 pp 4109ndash4112 1997
[44] F Bard C Cannon R Barbour et al ldquoPeripherally adminis-tered antibodies against amyloid 120573-peptide enter the centralnervous system and reduce pathology in a mouse model ofAlzheimer diseaserdquo Nature Medicine vol 6 no 8 pp 916ndash9192000
[45] R B DeMattos K R Bales D J Cummins J C Dodart S MPaul and D M Holtzman ldquoPeripheral anti-A120573 antibody altersCNS and plasma A120573 clearance and decreases brain A120573 burdenin a mouse model of Alzheimerrsquos diseaserdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 98 no 15 pp 8850ndash8855 2001
[46] C A Lemere E T Spooner J LaFrancois et al ldquoEvidence forperipheral clearance of cerebral A120573 protein following chronic
16 BioMed Research International
active A120573 immunization in PSAPP micerdquo Neurobiology ofDisease vol 14 no 1 pp 10ndash18 2003
[47] T Town M Vendrame A Patel et al ldquoReduced Th1 andenhanced Th2 immunity after immunization with Alzheimerrsquos120573-amyloid
1minus42rdquo Journal of Neuroimmunology vol 132 no 1-2
pp 49ndash59 2002[48] R Cacabelos ldquoPharmacogenomics in Alzheimerrsquos diseaserdquo
Methods in Molecular Biology vol 448 pp 213ndash357 2008[49] R Cacabelos ldquoPharmacogenomics and therapeutic strategies
for dementiardquo Expert Review of Molecular Diagnostics vol 9no 6 pp 567ndash611 2009
[50] D van Dam R DrsquoHooge M Staufenbiel C van Ginneken Fvan Meir and P P de Deyn ldquoAge-dependent cognitive declinein the APP23 model precedes amyloid depositionrdquo EuropeanJournal of Neuroscience vol 17 no 2 pp 388ndash396 2003
[51] M Dumont C Strazielle M Staufenbiel and R LalondeldquoSpatial learning and exploration of environmental stimuli in24-month-old female APP23 transgenic mice with the Swedishmutationrdquo Brain Research vol 1024 no 1-2 pp 113ndash121 2004
[52] R Lalonde M Dumont M Staufenbiel C Sturchler-Pierratand C Strazielle ldquoSpatial learning exploration anxiety andmotor coordination in female APP23 transgenic mice with theSwedish mutationrdquo Brain Research vol 956 no 1 pp 36ndash442002
[53] F J Huff F Boller F Lucchelli R Querriera J Beyer and SBelle ldquoThe neurologic examination in patients with probableAlzheimerrsquos diseaserdquo Archives of Neurology vol 44 no 9 pp929ndash932 1987
[54] A F Pettersson M Engardt and L O Wahlund ldquoActivitylevel and balance in subjects with mild Alzheimerrsquos diseaserdquoDementia and Geriatric Cognitive Disorders vol 13 no 4 pp213ndash216 2002
[55] L Calandreau B Desgranges R Jaffard and A DesmedtldquoSwitching from contextual to tone fear conditioning and viceversa the key role of the glutamatergic hippocampal-lateralseptal neurotransmissionrdquo Learning amp Memory vol 17 no 9pp 440ndash443 2010
[56] F Trinchese S Liu F Battaglia SWalter PMMathews andOArancio ldquoProgressive age-related development of Alzheimer-like pathology in APPPS1 micerdquo Annals of Neurology vol 55no 6 pp 801ndash814 2004
[57] D M Wilcock A Rojiani A Rosenthal et al ldquoPassiveimmunotherapy against A120573 in aged APP-transgenic micereverses cognitive deficits and depletes parenchymal amyloiddeposits in spite of increased vascular amyloid and microhem-orrhagerdquo Journal of Neuroinflammation vol 1 article 24 2004
[58] S A Ferguson S Sarkar and L C Schmued ldquoLongitudinalbehavioral changes in the APPPS1 transgenic AlzheimerrsquosDisease modelrdquo Behavioural Brain Research vol 242 no 1 pp125ndash134 2013
[59] D Wilcock and C Colton ldquoImmunotherapy vascular pathol-ogy and microhemorrhages in transgenic micerdquo CNS andNeurological DisordersmdashDrug Targets vol 8 no 1 pp 50ndash642009
[60] M Grundman S Gilman R S Black et al ldquoAn alternativemethod for estimating efficacy of the AN1792 vaccine foralzheimer diseaserdquo Neurology vol 71 no 9 pp 697ndash698 2008
[61] D Boche E Zotova R O Weller et al ldquoConsequence of A120573immunization on the vasculature of human Alzheimerrsquos diseasebrainrdquo Brain vol 131 no 12 pp 3299ndash3310 2008
[62] M Pfeifer S Boncristiano L Bondolfi et al ldquoCerebral hemor-rhage after passive anti-a120573 immunotherapyrdquo Science vol 298no 5597 p 1379 2002
[63] M M Racke L I Boone D L Hepburn et al ldquoExacerbationof cerebral amyloid angiopathy-associatedmicrohemorrhage inamyloid precursor protein transgenic mice by immunotherapyis dependent on antibody recognition of deposited forms ofamyloid 120573rdquo Journal of Neuroscience vol 25 no 3 pp 629ndash6362005
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

BioMed Research International 13
of AD as reported previously [24 47] Moreover presentresults demonstrated that EB101 did not induce astrogliosisin APPPS1 transgenic mice whereas some GFAP-positiveastrocyte clusters were aggregated in the affected brain areasof mice treated with A120573
42+ CFAIFA These results indicate
that EB101 may result in an effective treatment in AD neu-ropathology due to the absence of peripheral proinflamma-tory cytokine secretion and the absence of astroglial activa-tion in the brains of EB101 treated mice when compared withA12057342
+ CFAIFA immunotherapeutic response In a clinicalsetting however we may expect a wide differential responsein part depending upon the genomic profile of each patient[48 49] In AD behavioural studies in particular amongthe motor coordination tests the rotarod performance hasbeen reported to be impaired in some AD transgenic micemodels bearing A120573 plaques [50 51] This result was found inmutants prior to A120573 plaque formation at 3 and 6 months ofage but was not reproduced after plaques had been formedat 16 and 24 months of age [52] perhaps because of thedeclining abilities of older normal mice Here we presentthe effects of vaccine treatments in APPPS1 mice on motorcoordination a neglected aspect of the AD experimentalbehavior but justified on the basis of the deficient posturalcontrol reported in patients with Alzheimerrsquos disease [53]sometimes even during early stages [54] Our longitudinalmotor behavioral results showed no significant differencesin the ambulatory movement grip strength or coordinationaddressed by their performance in the rotating rod betweenAPPPS1 and wild-type mice for any age group studiedin contrast with other treated groups demonstrating thatlongitudinal motor assessments of AD transgenic mousemodels can provide valuable information with regard tothe effect of immunization on the progression of behavioraldeficits and overall motor activity As shown the EB101-treated group of mice exhibited longer latencies and betterbalancing skills indicating the absence of any negative effectof EB101 immunization on these mice The classical fearconditioning test is assumed to reveal the functional integrityof the hippocampus [55] representing a special memorytask to evaluate the AD models and its improvement afterimmunization periods In this APPPS1 mouse model thefirst associative learning and spatial workingmemory deficitsare shown at 5-6 months of age [56] although its impair-ment progression is relatively slow compared to most othermodels At the end of preventive immunization period EB101and A120573
42+ CFAIFA immunized mice have significantly
shorter escape latencies than the other treatment groups(A12057342liposome S1P-containing liposomes) and the controls
in the fear conditioning test Shorter escape latencies duringspatial learning indicated that deficits of spatial memorywere attenuated in EB101 and A120573
42+ CFAIFA immunized
mice Previous studies have showed that by 15 months ofage the spatial working memory is consistently impairedthroughout the rest of the life span [31 57] At the end oftherapeutic immunization period both EB101 and A120573
42+
CFAIFA immunized mice showed significantly improvedresults on the hippocampus-dependent spatial learning andmemory test when compared with other treatment groupsand the controls Taken together these observations are
in agreement with the impairment in conditioned learningin response to a tone stimulus of aged APPPS1 micealthough when immunized with EB101 during preventiveor therapeutic periods such impairments are significantlyattenuated
In summary the present results corroborate our previouspublished findings showing that EB101 immunization reducesamyloid accumulation in the APPPS1 mouse brain [2425] The strongest reduction in the preventive immunizationwas observed in the retrosplenial cortex followed by thehippocampus and entorhinal-piriform cortex Our resultsfurther support that EB101 ismore effectivewhen themice areimmunized prior to developing plaques in the hippocampaland cortical brain regions There is extensive evidence inmouse preclinical studies [58] and human clinical trials [5960] that the adverse effects of active and passive A120573 vaccinesresult in microhemorrhages and encephalitic reactions [60ndash63] yet themechanismbywhich they occur has not been fullyelucidated In the treatment sections analyzed we found nomicrohemorrhages or acute inflammatory processes in all thesurvived mice of the group indicating that EB101 therapeuticapproach presents no adverse inflammatory effects in thisAD mice models Thus our present results unequivocallydemonstrate that EB101 vaccinemay not only halt AD-relatedpathology as a preventive treatment but may also reverse it inestablished cases related to the improvement of the learningandmemory in both cases leading to a robust immunologicaltherapeutic response Present experiments also demonstratethat EB101 exerts more significant and substantial effects onthe reduction of A120573 deposition than A120573
42+CFAIFA vaccine
in A120573 mice models of AD However further exploration ofthe contribution of genetic environmental and physiologicalfactors in the development resistance to endogenous A120573-antibody is needed in order to optimize and modulate thetherapeutic strategies currently being developed
5 Conclusions
The present study carried out in APPPS1 transgenic mousemodel of AD compared the A120573 burden reduction andcellular immune responses produced by two immunizationdelivery strategies of A120573 in Freund adjuvant (described byWebster et al [10]) and in liposomeS1P matrix (describedby Schenk et al [17])The EB101 immunotherapeutic strategyboosted antibody generation A120573 plaque reduction and ananti-inflammatory response more effectively than A120573
42+
CFAIFA especially if given prior to or in the early stages ofpathology The present results also showed that vaccinationwith EB101 prevented the formation of new plaques andpresumably led to a reduction in established plaques whileinducing aTh2 immune reactionwithout encephalitogenic T-cell responses or brain inflammation related with astrogliosisHowever despite the efficacy demonstrated of this novelvaccine approach further preclinical and clinical studieson EB101 are warranted to evaluate its efficacy and themechanism for the future utility of such vaccine for ADtherapy
14 BioMed Research International
List of Abbreviations
AD Alzheimerrsquos diseaseA120573 120573-Amyloid proteinAPP Amyloid precursor proteinAPT Anterior pretectal nucleusCA1 Field CA1 hippocampusCA3 Field CA3 hippocampusCPu Caudate putamen (striatum)Cx CortexDG Dentate gyrusDN Dystrophic neuriteEct Entorhinal cortexCFA Complete Freundrsquos adjuvantGrDG Granular dentate gyrusIFA Incomplete Freundrsquos adjuvantIL InterleukinLPMR Lateral posterior thalamic nucleus (mediorostral)MoDG Molecular dentate gyrusPir Piriform cortexPoDG Polymorph dentate gyrusRSGb Retrosplenial granular cortex b regionTg TransgenicTh T helper
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Ivan Carrera Ignacio Etcheverrıa Lucıa Fernandez-NovoaValter Lombardi Ramon Cacabelos and Carmen Vigo con-ceived and designed the experiments Ivan Carrera and Igna-cio Etcheverrıa performed the experiments Ivan CarreraIgnacio Etcheverrıa and Valter Lombardi analyzed the dataIvan Carrera Ignacio Etcheverrıa Lucıa Fernandez-Novoaand Valter Lombardi contributed reagentsmaterialsanalysistools Ivan Carrera Carmen Vigo andMadepalli KrishnappaLakshmana wrote the paper
Acknowledgments
This work was supported by National Institute of Aging(NIA)NIH Grant no 1R01AG036859-01 (Madepalli Krish-nappa Lakshmana) and the financial support of the ProgramaOperativo Feder Galicia 2007ndash2013 (IN841C 201254 to PhDCarrera I)
References
[1] R Cacabelos L Fernandez-Novoa V Lombardi Y Kubotaand M Takeda ldquoMolecular genetics of Alzheimerrsquos diseaseand agingrdquoMethods and Findings in Experimental and ClinicalPharmacology vol 27 pp 1ndash573 2005
[2] P T Nelson I Alafuzoff E H Bigio et al ldquoCorrelationof alzheimer disease neuropathologic changes with cognitive
status a review of the literaturerdquo Journal of Neuropathology andExperimental Neurology vol 71 no 5 pp 362ndash381 2012
[3] JNaslundVHaroutunian RMohs et al ldquoCorrelation betweenelevated levels of amyloid 120573-peptide in the brain and cognitivedeclinerdquo Journal of the American Medical Association vol 283no 12 pp 1571ndash1577 2000
[4] A Koyama O I Okereke T Yang D Blacker D J Selkoeand F Grodstein ldquoPlasma amyloid-120573 as a predictor of dementiaand cognitive decline a systematic review and meta-analysisrdquoArchives of Neurology vol 69 no 7 pp 824ndash831 2012
[5] A M Fagan C M Roe C Xiong M A Mintun J C Morrisand D M Holtzman ldquoCerebrospinal fluid tau120573-amyloid42ratio as a prediction of cognitive decline in nondemented olderadultsrdquo Archives of Neurology vol 64 no 3 pp 343ndash349 2007
[6] S Parvathy P Davies V Haroutunian et al ldquoCorrelationbetween A120573-40- A120573-42- and A120573x-43-containing amyloidplaques and cognitive declinerdquo Archives of Neurology vol 58no 12 pp 2025ndash2032 2001
[7] E Karran M Mercken and B D Strooper ldquoThe amyloidcascade hypothesis for Alzheimerrsquos disease an appraisal for thedevelopment of therapeuticsrdquo Nature Reviews Drug Discoveryvol 10 no 9 pp 698ndash712 2011
[8] J L Jankowsky D J Fadale J Anderson et al ldquoMutantpresenilins specifically elevate the levels of the 42 residue 120573-amyloid peptide in vivo evidence for augmentation of a 42-specific 120574 secretaserdquo Human Molecular Genetics vol 13 no 2pp 159ndash170 2004
[9] L Holcomb M N Gordon E Mcgowan et al ldquoAcceleratedAlzheimer-type phenotype in transgenic mice carrying bothmutant amyloid precursor protein and presenilin 1 transgenesrdquoNature Medicine vol 4 no 1 pp 97ndash100 1998
[10] S JWebster A D Bachstetter and L J van Eldik ldquoComprehen-sive behavioral characterization of an APPPS-1 double knock-in mouse model of Alzheimerrsquos diseaserdquo Alzheimerrsquos ResearchandTherapy vol 5 no 3 article 28 2013
[11] H Xiong D Callaghan J Wodzinska et al ldquoBiochemical andbehavioral characterization of the double transgenic mousemodel (APPswePS1dE9) of Alzheimerrsquos diseaserdquo NeuroscienceBulletin vol 27 no 4 pp 221ndash232 2011
[12] M Garcia-Alloza L A Borrelli B T Hyman and B J BacskaildquoAntioxidants have a rapid and long-lasting effect on neuriticabnormalities in APPPS1 micerdquo Neurobiology of Aging vol 31no 12 pp 2058ndash2068 2010
[13] M Meyer-Luehmann M Mielke T L Spires-Jones et al ldquoAreporter of local dendritic translocation shows plaque-relatedloss of neural system function in APP-transgenic micerdquo Journalof Neuroscience vol 29 no 40 pp 12636ndash12640 2009
[14] E Machova V Rudajev H Smyckova H Koivisto H Tanilaand V Dolezal ldquoFunctional cholinergic damage develops withamyloid accumulation in young adult APPswePS1dE9 trans-genic micerdquo Neurobiology of Disease vol 38 no 1 pp 27ndash352010
[15] T Malm J Koistinaho and K Kanninen ldquoUtilization of APP-swePS1dE9 transgenic mice in research of Alzheimerrsquos diseasefocus on gene therapy and cell-based therapy applicationsrdquoInternational Journal of Alzheimerrsquos Disease vol 2011 Article ID517160 8 pages 2011
[16] J Lee and P Han ldquoAn update of animal models of alzheimerdisease with a reevaluation of plaque depositionsrdquoExperimentalNeurobiology vol 22 pp 84ndash95 2013
BioMed Research International 15
[17] D Schenk R Barbour W Dunn et al ldquoImmunization withamyloid-120573 attenuates Alzheimer disease-like pathology in thePDAPP mouserdquo Nature vol 400 no 6740 pp 173ndash177 1999
[18] H J Fu B Liu J L Frost and C A Lemere ldquoAmyloid-120573immunotherapy for Alzheimerrsquos diseaserdquoCNS and NeurologicalDisorders-Drug Targets vol 9 no 2 pp 197ndash206 2010
[19] J-M Orgogozo S Gilman J-F Dartigues et al ldquoSubacutemeningoencephalitis in a subset of patients with AD after A120573
42
immunizationrdquo Neurology vol 61 no 1 pp 46ndash54 2003[20] J A R Nicolll DWilkinson CHolmes P Steart HMarkham
andROWeller ldquoNeuropathology of humanAlzheimer diseaseafter immunization with amyloid-120573 peptide a case reportrdquoNature Medicine vol 9 no 4 pp 448ndash452 2003
[21] I Ferrer M Boada Rovira M L Sanchez Guerra M JRey and F Costa-Jussa ldquoNeuropathology and pathogenesisof encephalitis following amyloid-Neuropathology and patho-genesis of encephalitis following amyloid-beta immunizationin Alzheimerrsquos disease immunization in Alzheimerrsquos diseaserdquoBrain Pathology vol 14 no 1 pp 11ndash20 2004
[22] D H Cribbs A Ghochikyan V Vasilevko et al ldquoAdjuvant-dependent modulation ofTh1 andTh2 responses to immuniza-tion with 120573-amyloidrdquo International Immunology vol 15 no 4pp 505ndash514 2003
[23] C Holmes D Boche D Wilkinson et al ldquoLong-term effectsof A120573
42immunisation in Alzheimerrsquos disease follow-up of a
randomised placebo-controlled phase I trialrdquo The Lancet vol372 no 9634 pp 216ndash223 2008
[24] I Carrera I Etcheverrıa L Fernandez-Novoa V LombardiR Cacabelos and C Vigo ldquoVaccine development to treatAlzheimerrsquos disease neuropathology in APPPS1 transgenicmicerdquo International Journal of Alzheimerrsquos Disease vol 2012Article ID 376138 17 pages 2012
[25] I Carrera I Etcheverrıa Y Li et al ldquoImmunocytochemicalcharacterization of Alzheimer disease hallmarks in APPPS1transgenic mice treated with a new anti-amyloid-120573 vaccinerdquoBioMed Research International vol 2013 Article ID 709145 12pages 2013
[26] V Anelli R Bassi G Tettamanti P Viani and L Riboni ldquoExtra-cellular release of newly synthesized sphingosine-1-phosphateby cerebellar granule cells and astrocytesrdquo Journal of Neuro-chemistry vol 92 no 5 pp 1204ndash1215 2005
[27] S Schroeter K Khan R Barbour et al ldquoImmunotherapyreduces vascular amyloid-120573 in PDAPP micerdquo Journal of Neu-roscience vol 28 no 27 pp 6787ndash6793 2008
[28] Z Suo A Cox N Bartelli et al ldquoGRK5 deficiency leads to earlyAlzheimer-like pathology and working memory impairmentrdquoNeurobiology of Aging vol 28 no 12 pp 1873ndash1888 2007
[29] W Kamphuis C Mamber MMoeton et al ldquoGFAP isoforms inadult mouse brain with a focus on neurogenic astrocytes andreactive astrogliosis in mouse models of Alzheimer diseaserdquoPLoS ONE vol 7 no 8 Article ID e42823 2012
[30] J Lang C Vigo-Pelfrey and F Martin ldquoLiposomes composedof partially hydrogenated egg phosphatidylcholines fatty acidcomposition thermal phase behavior and oxidative stabilityrdquoChemistry and Physics of Lipids vol 53 no 1 pp 91ndash101 1990
[31] D Morgan D M Diamond P E Gottschall et al ldquoA 120573peptide vaccination prevents memory loss in an animal modelof Alzheimerrsquos diseaserdquo Nature vol 408 no 6815 pp 982ndash9852000
[32] M L Brownlow L Benner D DrsquoAgostino M N Gordon andD Morgan ldquoKetogenic diet improves motor performance but
not cognition in two mouse models of Alzheimerrsquos pathologyrdquoPLoS ONE vol 8 no 9 Article ID e75713 2013
[33] M Pride P Seubert M Grundman M Hagen J Eldridgeand R S Black ldquoProgress in the active immunotherapeu-tic approach to Alzheimerrsquos disease clinical investigationsinto AN1792-associated meningoencephalitisrdquo Neurodegenera-tive Diseases vol 5 no 3-4 pp 194ndash196 2008
[34] CWiessner K-HWiederhold A C Tissot et al ldquoThe second-generation activeA120573 immunotherapyCAD106 reduces amyloidaccumulation in APP transgenic mice while minimizing poten-tial side effectsrdquoThe Journal of Neuroscience vol 31 no 25 pp9323ndash9331 2011
[35] D Lambracht-Washington and R Rosenberg ldquoAdvances in thedevelopment of vaccines for Alzheimerrsquos diseaserdquo DiscoveryMedicine vol 15 no 84 pp 319ndash326 2013
[36] A Kimura T Ohmori R Ohkawa et al ldquoEssential roles ofsphingosine 1-phosphateS1P
1receptor axis in the migration of
neural stem cells toward a site of spinal cord injuryrdquo Stem Cellsvol 25 no 1 pp 115ndash124 2007
[37] S S Lee C H Woo J D Chang and J H Kim ldquoRoles of Racand cytosolic phospholipase A2 in the intracellular signalling inresponse to titanium particlesrdquo Cellular Signalling vol 15 no 3pp 339ndash345 2003
[38] E Masliah L Hansen A Adame et al ldquoA120573 vaccination effectson plaque pathology in the absence of encephalitis inAlzheimerdiseaserdquo Neurology vol 64 no 1 pp 129ndash131 2005
[39] A Pilorget B Annabi F Bouzeghrane J Marvaldi J Luisand R Beliveau ldquoInhibition of angiogenic properties of brainendothelial cells by platelet-derived sphingosine-1-phosphaterdquoJournal of Cerebral Blood Flow and Metabolism vol 25 no 9pp 1171ndash1182 2005
[40] A Wang P Das R C Switzer III T E Golde and J LJankowsky ldquoRobust amyloid clearance in a mouse model ofAlzheimerrsquos disease provides novel insights into themechanismof amyloid-120573 immunotherapyrdquo Journal of Neuroscience vol 31no 11 pp 4124ndash4136 2011
[41] G Krabbe A Halle V Matyash et al ldquoFunctional impairmentof microglia coincides with Beta-amyloid deposition in micewith Alzheimer-like pathologyrdquo PLoS ONE vol 8 no 4 ArticleID e60921 2013
[42] Y H Liu B Giunta H-D Zhou J Tan and Y J WangldquoImmunotherapy for Alzheimer disease-the challenge ofadverse effectsrdquo Nature Reviews Neurology vol 8 no 8 pp465ndash469 2012
[43] B Solomon R Koppel D Frankel and E Hanan-AharonldquoDisaggregation of Alzheimer120573-amyloid by site-directedmAbrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 94 no 8 pp 4109ndash4112 1997
[44] F Bard C Cannon R Barbour et al ldquoPeripherally adminis-tered antibodies against amyloid 120573-peptide enter the centralnervous system and reduce pathology in a mouse model ofAlzheimer diseaserdquo Nature Medicine vol 6 no 8 pp 916ndash9192000
[45] R B DeMattos K R Bales D J Cummins J C Dodart S MPaul and D M Holtzman ldquoPeripheral anti-A120573 antibody altersCNS and plasma A120573 clearance and decreases brain A120573 burdenin a mouse model of Alzheimerrsquos diseaserdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 98 no 15 pp 8850ndash8855 2001
[46] C A Lemere E T Spooner J LaFrancois et al ldquoEvidence forperipheral clearance of cerebral A120573 protein following chronic
16 BioMed Research International
active A120573 immunization in PSAPP micerdquo Neurobiology ofDisease vol 14 no 1 pp 10ndash18 2003
[47] T Town M Vendrame A Patel et al ldquoReduced Th1 andenhanced Th2 immunity after immunization with Alzheimerrsquos120573-amyloid
1minus42rdquo Journal of Neuroimmunology vol 132 no 1-2
pp 49ndash59 2002[48] R Cacabelos ldquoPharmacogenomics in Alzheimerrsquos diseaserdquo
Methods in Molecular Biology vol 448 pp 213ndash357 2008[49] R Cacabelos ldquoPharmacogenomics and therapeutic strategies
for dementiardquo Expert Review of Molecular Diagnostics vol 9no 6 pp 567ndash611 2009
[50] D van Dam R DrsquoHooge M Staufenbiel C van Ginneken Fvan Meir and P P de Deyn ldquoAge-dependent cognitive declinein the APP23 model precedes amyloid depositionrdquo EuropeanJournal of Neuroscience vol 17 no 2 pp 388ndash396 2003
[51] M Dumont C Strazielle M Staufenbiel and R LalondeldquoSpatial learning and exploration of environmental stimuli in24-month-old female APP23 transgenic mice with the Swedishmutationrdquo Brain Research vol 1024 no 1-2 pp 113ndash121 2004
[52] R Lalonde M Dumont M Staufenbiel C Sturchler-Pierratand C Strazielle ldquoSpatial learning exploration anxiety andmotor coordination in female APP23 transgenic mice with theSwedish mutationrdquo Brain Research vol 956 no 1 pp 36ndash442002
[53] F J Huff F Boller F Lucchelli R Querriera J Beyer and SBelle ldquoThe neurologic examination in patients with probableAlzheimerrsquos diseaserdquo Archives of Neurology vol 44 no 9 pp929ndash932 1987
[54] A F Pettersson M Engardt and L O Wahlund ldquoActivitylevel and balance in subjects with mild Alzheimerrsquos diseaserdquoDementia and Geriatric Cognitive Disorders vol 13 no 4 pp213ndash216 2002
[55] L Calandreau B Desgranges R Jaffard and A DesmedtldquoSwitching from contextual to tone fear conditioning and viceversa the key role of the glutamatergic hippocampal-lateralseptal neurotransmissionrdquo Learning amp Memory vol 17 no 9pp 440ndash443 2010
[56] F Trinchese S Liu F Battaglia SWalter PMMathews andOArancio ldquoProgressive age-related development of Alzheimer-like pathology in APPPS1 micerdquo Annals of Neurology vol 55no 6 pp 801ndash814 2004
[57] D M Wilcock A Rojiani A Rosenthal et al ldquoPassiveimmunotherapy against A120573 in aged APP-transgenic micereverses cognitive deficits and depletes parenchymal amyloiddeposits in spite of increased vascular amyloid and microhem-orrhagerdquo Journal of Neuroinflammation vol 1 article 24 2004
[58] S A Ferguson S Sarkar and L C Schmued ldquoLongitudinalbehavioral changes in the APPPS1 transgenic AlzheimerrsquosDisease modelrdquo Behavioural Brain Research vol 242 no 1 pp125ndash134 2013
[59] D Wilcock and C Colton ldquoImmunotherapy vascular pathol-ogy and microhemorrhages in transgenic micerdquo CNS andNeurological DisordersmdashDrug Targets vol 8 no 1 pp 50ndash642009
[60] M Grundman S Gilman R S Black et al ldquoAn alternativemethod for estimating efficacy of the AN1792 vaccine foralzheimer diseaserdquo Neurology vol 71 no 9 pp 697ndash698 2008
[61] D Boche E Zotova R O Weller et al ldquoConsequence of A120573immunization on the vasculature of human Alzheimerrsquos diseasebrainrdquo Brain vol 131 no 12 pp 3299ndash3310 2008
[62] M Pfeifer S Boncristiano L Bondolfi et al ldquoCerebral hemor-rhage after passive anti-a120573 immunotherapyrdquo Science vol 298no 5597 p 1379 2002
[63] M M Racke L I Boone D L Hepburn et al ldquoExacerbationof cerebral amyloid angiopathy-associatedmicrohemorrhage inamyloid precursor protein transgenic mice by immunotherapyis dependent on antibody recognition of deposited forms ofamyloid 120573rdquo Journal of Neuroscience vol 25 no 3 pp 629ndash6362005
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

14 BioMed Research International
List of Abbreviations
AD Alzheimerrsquos diseaseA120573 120573-Amyloid proteinAPP Amyloid precursor proteinAPT Anterior pretectal nucleusCA1 Field CA1 hippocampusCA3 Field CA3 hippocampusCPu Caudate putamen (striatum)Cx CortexDG Dentate gyrusDN Dystrophic neuriteEct Entorhinal cortexCFA Complete Freundrsquos adjuvantGrDG Granular dentate gyrusIFA Incomplete Freundrsquos adjuvantIL InterleukinLPMR Lateral posterior thalamic nucleus (mediorostral)MoDG Molecular dentate gyrusPir Piriform cortexPoDG Polymorph dentate gyrusRSGb Retrosplenial granular cortex b regionTg TransgenicTh T helper
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Ivan Carrera Ignacio Etcheverrıa Lucıa Fernandez-NovoaValter Lombardi Ramon Cacabelos and Carmen Vigo con-ceived and designed the experiments Ivan Carrera and Igna-cio Etcheverrıa performed the experiments Ivan CarreraIgnacio Etcheverrıa and Valter Lombardi analyzed the dataIvan Carrera Ignacio Etcheverrıa Lucıa Fernandez-Novoaand Valter Lombardi contributed reagentsmaterialsanalysistools Ivan Carrera Carmen Vigo andMadepalli KrishnappaLakshmana wrote the paper
Acknowledgments
This work was supported by National Institute of Aging(NIA)NIH Grant no 1R01AG036859-01 (Madepalli Krish-nappa Lakshmana) and the financial support of the ProgramaOperativo Feder Galicia 2007ndash2013 (IN841C 201254 to PhDCarrera I)
References
[1] R Cacabelos L Fernandez-Novoa V Lombardi Y Kubotaand M Takeda ldquoMolecular genetics of Alzheimerrsquos diseaseand agingrdquoMethods and Findings in Experimental and ClinicalPharmacology vol 27 pp 1ndash573 2005
[2] P T Nelson I Alafuzoff E H Bigio et al ldquoCorrelationof alzheimer disease neuropathologic changes with cognitive
status a review of the literaturerdquo Journal of Neuropathology andExperimental Neurology vol 71 no 5 pp 362ndash381 2012
[3] JNaslundVHaroutunian RMohs et al ldquoCorrelation betweenelevated levels of amyloid 120573-peptide in the brain and cognitivedeclinerdquo Journal of the American Medical Association vol 283no 12 pp 1571ndash1577 2000
[4] A Koyama O I Okereke T Yang D Blacker D J Selkoeand F Grodstein ldquoPlasma amyloid-120573 as a predictor of dementiaand cognitive decline a systematic review and meta-analysisrdquoArchives of Neurology vol 69 no 7 pp 824ndash831 2012
[5] A M Fagan C M Roe C Xiong M A Mintun J C Morrisand D M Holtzman ldquoCerebrospinal fluid tau120573-amyloid42ratio as a prediction of cognitive decline in nondemented olderadultsrdquo Archives of Neurology vol 64 no 3 pp 343ndash349 2007
[6] S Parvathy P Davies V Haroutunian et al ldquoCorrelationbetween A120573-40- A120573-42- and A120573x-43-containing amyloidplaques and cognitive declinerdquo Archives of Neurology vol 58no 12 pp 2025ndash2032 2001
[7] E Karran M Mercken and B D Strooper ldquoThe amyloidcascade hypothesis for Alzheimerrsquos disease an appraisal for thedevelopment of therapeuticsrdquo Nature Reviews Drug Discoveryvol 10 no 9 pp 698ndash712 2011
[8] J L Jankowsky D J Fadale J Anderson et al ldquoMutantpresenilins specifically elevate the levels of the 42 residue 120573-amyloid peptide in vivo evidence for augmentation of a 42-specific 120574 secretaserdquo Human Molecular Genetics vol 13 no 2pp 159ndash170 2004
[9] L Holcomb M N Gordon E Mcgowan et al ldquoAcceleratedAlzheimer-type phenotype in transgenic mice carrying bothmutant amyloid precursor protein and presenilin 1 transgenesrdquoNature Medicine vol 4 no 1 pp 97ndash100 1998
[10] S JWebster A D Bachstetter and L J van Eldik ldquoComprehen-sive behavioral characterization of an APPPS-1 double knock-in mouse model of Alzheimerrsquos diseaserdquo Alzheimerrsquos ResearchandTherapy vol 5 no 3 article 28 2013
[11] H Xiong D Callaghan J Wodzinska et al ldquoBiochemical andbehavioral characterization of the double transgenic mousemodel (APPswePS1dE9) of Alzheimerrsquos diseaserdquo NeuroscienceBulletin vol 27 no 4 pp 221ndash232 2011
[12] M Garcia-Alloza L A Borrelli B T Hyman and B J BacskaildquoAntioxidants have a rapid and long-lasting effect on neuriticabnormalities in APPPS1 micerdquo Neurobiology of Aging vol 31no 12 pp 2058ndash2068 2010
[13] M Meyer-Luehmann M Mielke T L Spires-Jones et al ldquoAreporter of local dendritic translocation shows plaque-relatedloss of neural system function in APP-transgenic micerdquo Journalof Neuroscience vol 29 no 40 pp 12636ndash12640 2009
[14] E Machova V Rudajev H Smyckova H Koivisto H Tanilaand V Dolezal ldquoFunctional cholinergic damage develops withamyloid accumulation in young adult APPswePS1dE9 trans-genic micerdquo Neurobiology of Disease vol 38 no 1 pp 27ndash352010
[15] T Malm J Koistinaho and K Kanninen ldquoUtilization of APP-swePS1dE9 transgenic mice in research of Alzheimerrsquos diseasefocus on gene therapy and cell-based therapy applicationsrdquoInternational Journal of Alzheimerrsquos Disease vol 2011 Article ID517160 8 pages 2011
[16] J Lee and P Han ldquoAn update of animal models of alzheimerdisease with a reevaluation of plaque depositionsrdquoExperimentalNeurobiology vol 22 pp 84ndash95 2013
BioMed Research International 15
[17] D Schenk R Barbour W Dunn et al ldquoImmunization withamyloid-120573 attenuates Alzheimer disease-like pathology in thePDAPP mouserdquo Nature vol 400 no 6740 pp 173ndash177 1999
[18] H J Fu B Liu J L Frost and C A Lemere ldquoAmyloid-120573immunotherapy for Alzheimerrsquos diseaserdquoCNS and NeurologicalDisorders-Drug Targets vol 9 no 2 pp 197ndash206 2010
[19] J-M Orgogozo S Gilman J-F Dartigues et al ldquoSubacutemeningoencephalitis in a subset of patients with AD after A120573
42
immunizationrdquo Neurology vol 61 no 1 pp 46ndash54 2003[20] J A R Nicolll DWilkinson CHolmes P Steart HMarkham
andROWeller ldquoNeuropathology of humanAlzheimer diseaseafter immunization with amyloid-120573 peptide a case reportrdquoNature Medicine vol 9 no 4 pp 448ndash452 2003
[21] I Ferrer M Boada Rovira M L Sanchez Guerra M JRey and F Costa-Jussa ldquoNeuropathology and pathogenesisof encephalitis following amyloid-Neuropathology and patho-genesis of encephalitis following amyloid-beta immunizationin Alzheimerrsquos disease immunization in Alzheimerrsquos diseaserdquoBrain Pathology vol 14 no 1 pp 11ndash20 2004
[22] D H Cribbs A Ghochikyan V Vasilevko et al ldquoAdjuvant-dependent modulation ofTh1 andTh2 responses to immuniza-tion with 120573-amyloidrdquo International Immunology vol 15 no 4pp 505ndash514 2003
[23] C Holmes D Boche D Wilkinson et al ldquoLong-term effectsof A120573
42immunisation in Alzheimerrsquos disease follow-up of a
randomised placebo-controlled phase I trialrdquo The Lancet vol372 no 9634 pp 216ndash223 2008
[24] I Carrera I Etcheverrıa L Fernandez-Novoa V LombardiR Cacabelos and C Vigo ldquoVaccine development to treatAlzheimerrsquos disease neuropathology in APPPS1 transgenicmicerdquo International Journal of Alzheimerrsquos Disease vol 2012Article ID 376138 17 pages 2012
[25] I Carrera I Etcheverrıa Y Li et al ldquoImmunocytochemicalcharacterization of Alzheimer disease hallmarks in APPPS1transgenic mice treated with a new anti-amyloid-120573 vaccinerdquoBioMed Research International vol 2013 Article ID 709145 12pages 2013
[26] V Anelli R Bassi G Tettamanti P Viani and L Riboni ldquoExtra-cellular release of newly synthesized sphingosine-1-phosphateby cerebellar granule cells and astrocytesrdquo Journal of Neuro-chemistry vol 92 no 5 pp 1204ndash1215 2005
[27] S Schroeter K Khan R Barbour et al ldquoImmunotherapyreduces vascular amyloid-120573 in PDAPP micerdquo Journal of Neu-roscience vol 28 no 27 pp 6787ndash6793 2008
[28] Z Suo A Cox N Bartelli et al ldquoGRK5 deficiency leads to earlyAlzheimer-like pathology and working memory impairmentrdquoNeurobiology of Aging vol 28 no 12 pp 1873ndash1888 2007
[29] W Kamphuis C Mamber MMoeton et al ldquoGFAP isoforms inadult mouse brain with a focus on neurogenic astrocytes andreactive astrogliosis in mouse models of Alzheimer diseaserdquoPLoS ONE vol 7 no 8 Article ID e42823 2012
[30] J Lang C Vigo-Pelfrey and F Martin ldquoLiposomes composedof partially hydrogenated egg phosphatidylcholines fatty acidcomposition thermal phase behavior and oxidative stabilityrdquoChemistry and Physics of Lipids vol 53 no 1 pp 91ndash101 1990
[31] D Morgan D M Diamond P E Gottschall et al ldquoA 120573peptide vaccination prevents memory loss in an animal modelof Alzheimerrsquos diseaserdquo Nature vol 408 no 6815 pp 982ndash9852000
[32] M L Brownlow L Benner D DrsquoAgostino M N Gordon andD Morgan ldquoKetogenic diet improves motor performance but
not cognition in two mouse models of Alzheimerrsquos pathologyrdquoPLoS ONE vol 8 no 9 Article ID e75713 2013
[33] M Pride P Seubert M Grundman M Hagen J Eldridgeand R S Black ldquoProgress in the active immunotherapeu-tic approach to Alzheimerrsquos disease clinical investigationsinto AN1792-associated meningoencephalitisrdquo Neurodegenera-tive Diseases vol 5 no 3-4 pp 194ndash196 2008
[34] CWiessner K-HWiederhold A C Tissot et al ldquoThe second-generation activeA120573 immunotherapyCAD106 reduces amyloidaccumulation in APP transgenic mice while minimizing poten-tial side effectsrdquoThe Journal of Neuroscience vol 31 no 25 pp9323ndash9331 2011
[35] D Lambracht-Washington and R Rosenberg ldquoAdvances in thedevelopment of vaccines for Alzheimerrsquos diseaserdquo DiscoveryMedicine vol 15 no 84 pp 319ndash326 2013
[36] A Kimura T Ohmori R Ohkawa et al ldquoEssential roles ofsphingosine 1-phosphateS1P
1receptor axis in the migration of
neural stem cells toward a site of spinal cord injuryrdquo Stem Cellsvol 25 no 1 pp 115ndash124 2007
[37] S S Lee C H Woo J D Chang and J H Kim ldquoRoles of Racand cytosolic phospholipase A2 in the intracellular signalling inresponse to titanium particlesrdquo Cellular Signalling vol 15 no 3pp 339ndash345 2003
[38] E Masliah L Hansen A Adame et al ldquoA120573 vaccination effectson plaque pathology in the absence of encephalitis inAlzheimerdiseaserdquo Neurology vol 64 no 1 pp 129ndash131 2005
[39] A Pilorget B Annabi F Bouzeghrane J Marvaldi J Luisand R Beliveau ldquoInhibition of angiogenic properties of brainendothelial cells by platelet-derived sphingosine-1-phosphaterdquoJournal of Cerebral Blood Flow and Metabolism vol 25 no 9pp 1171ndash1182 2005
[40] A Wang P Das R C Switzer III T E Golde and J LJankowsky ldquoRobust amyloid clearance in a mouse model ofAlzheimerrsquos disease provides novel insights into themechanismof amyloid-120573 immunotherapyrdquo Journal of Neuroscience vol 31no 11 pp 4124ndash4136 2011
[41] G Krabbe A Halle V Matyash et al ldquoFunctional impairmentof microglia coincides with Beta-amyloid deposition in micewith Alzheimer-like pathologyrdquo PLoS ONE vol 8 no 4 ArticleID e60921 2013
[42] Y H Liu B Giunta H-D Zhou J Tan and Y J WangldquoImmunotherapy for Alzheimer disease-the challenge ofadverse effectsrdquo Nature Reviews Neurology vol 8 no 8 pp465ndash469 2012
[43] B Solomon R Koppel D Frankel and E Hanan-AharonldquoDisaggregation of Alzheimer120573-amyloid by site-directedmAbrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 94 no 8 pp 4109ndash4112 1997
[44] F Bard C Cannon R Barbour et al ldquoPeripherally adminis-tered antibodies against amyloid 120573-peptide enter the centralnervous system and reduce pathology in a mouse model ofAlzheimer diseaserdquo Nature Medicine vol 6 no 8 pp 916ndash9192000
[45] R B DeMattos K R Bales D J Cummins J C Dodart S MPaul and D M Holtzman ldquoPeripheral anti-A120573 antibody altersCNS and plasma A120573 clearance and decreases brain A120573 burdenin a mouse model of Alzheimerrsquos diseaserdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 98 no 15 pp 8850ndash8855 2001
[46] C A Lemere E T Spooner J LaFrancois et al ldquoEvidence forperipheral clearance of cerebral A120573 protein following chronic
16 BioMed Research International
active A120573 immunization in PSAPP micerdquo Neurobiology ofDisease vol 14 no 1 pp 10ndash18 2003
[47] T Town M Vendrame A Patel et al ldquoReduced Th1 andenhanced Th2 immunity after immunization with Alzheimerrsquos120573-amyloid
1minus42rdquo Journal of Neuroimmunology vol 132 no 1-2
pp 49ndash59 2002[48] R Cacabelos ldquoPharmacogenomics in Alzheimerrsquos diseaserdquo
Methods in Molecular Biology vol 448 pp 213ndash357 2008[49] R Cacabelos ldquoPharmacogenomics and therapeutic strategies
for dementiardquo Expert Review of Molecular Diagnostics vol 9no 6 pp 567ndash611 2009
[50] D van Dam R DrsquoHooge M Staufenbiel C van Ginneken Fvan Meir and P P de Deyn ldquoAge-dependent cognitive declinein the APP23 model precedes amyloid depositionrdquo EuropeanJournal of Neuroscience vol 17 no 2 pp 388ndash396 2003
[51] M Dumont C Strazielle M Staufenbiel and R LalondeldquoSpatial learning and exploration of environmental stimuli in24-month-old female APP23 transgenic mice with the Swedishmutationrdquo Brain Research vol 1024 no 1-2 pp 113ndash121 2004
[52] R Lalonde M Dumont M Staufenbiel C Sturchler-Pierratand C Strazielle ldquoSpatial learning exploration anxiety andmotor coordination in female APP23 transgenic mice with theSwedish mutationrdquo Brain Research vol 956 no 1 pp 36ndash442002
[53] F J Huff F Boller F Lucchelli R Querriera J Beyer and SBelle ldquoThe neurologic examination in patients with probableAlzheimerrsquos diseaserdquo Archives of Neurology vol 44 no 9 pp929ndash932 1987
[54] A F Pettersson M Engardt and L O Wahlund ldquoActivitylevel and balance in subjects with mild Alzheimerrsquos diseaserdquoDementia and Geriatric Cognitive Disorders vol 13 no 4 pp213ndash216 2002
[55] L Calandreau B Desgranges R Jaffard and A DesmedtldquoSwitching from contextual to tone fear conditioning and viceversa the key role of the glutamatergic hippocampal-lateralseptal neurotransmissionrdquo Learning amp Memory vol 17 no 9pp 440ndash443 2010
[56] F Trinchese S Liu F Battaglia SWalter PMMathews andOArancio ldquoProgressive age-related development of Alzheimer-like pathology in APPPS1 micerdquo Annals of Neurology vol 55no 6 pp 801ndash814 2004
[57] D M Wilcock A Rojiani A Rosenthal et al ldquoPassiveimmunotherapy against A120573 in aged APP-transgenic micereverses cognitive deficits and depletes parenchymal amyloiddeposits in spite of increased vascular amyloid and microhem-orrhagerdquo Journal of Neuroinflammation vol 1 article 24 2004
[58] S A Ferguson S Sarkar and L C Schmued ldquoLongitudinalbehavioral changes in the APPPS1 transgenic AlzheimerrsquosDisease modelrdquo Behavioural Brain Research vol 242 no 1 pp125ndash134 2013
[59] D Wilcock and C Colton ldquoImmunotherapy vascular pathol-ogy and microhemorrhages in transgenic micerdquo CNS andNeurological DisordersmdashDrug Targets vol 8 no 1 pp 50ndash642009
[60] M Grundman S Gilman R S Black et al ldquoAn alternativemethod for estimating efficacy of the AN1792 vaccine foralzheimer diseaserdquo Neurology vol 71 no 9 pp 697ndash698 2008
[61] D Boche E Zotova R O Weller et al ldquoConsequence of A120573immunization on the vasculature of human Alzheimerrsquos diseasebrainrdquo Brain vol 131 no 12 pp 3299ndash3310 2008
[62] M Pfeifer S Boncristiano L Bondolfi et al ldquoCerebral hemor-rhage after passive anti-a120573 immunotherapyrdquo Science vol 298no 5597 p 1379 2002
[63] M M Racke L I Boone D L Hepburn et al ldquoExacerbationof cerebral amyloid angiopathy-associatedmicrohemorrhage inamyloid precursor protein transgenic mice by immunotherapyis dependent on antibody recognition of deposited forms ofamyloid 120573rdquo Journal of Neuroscience vol 25 no 3 pp 629ndash6362005
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

BioMed Research International 15
[17] D Schenk R Barbour W Dunn et al ldquoImmunization withamyloid-120573 attenuates Alzheimer disease-like pathology in thePDAPP mouserdquo Nature vol 400 no 6740 pp 173ndash177 1999
[18] H J Fu B Liu J L Frost and C A Lemere ldquoAmyloid-120573immunotherapy for Alzheimerrsquos diseaserdquoCNS and NeurologicalDisorders-Drug Targets vol 9 no 2 pp 197ndash206 2010
[19] J-M Orgogozo S Gilman J-F Dartigues et al ldquoSubacutemeningoencephalitis in a subset of patients with AD after A120573
42
immunizationrdquo Neurology vol 61 no 1 pp 46ndash54 2003[20] J A R Nicolll DWilkinson CHolmes P Steart HMarkham
andROWeller ldquoNeuropathology of humanAlzheimer diseaseafter immunization with amyloid-120573 peptide a case reportrdquoNature Medicine vol 9 no 4 pp 448ndash452 2003
[21] I Ferrer M Boada Rovira M L Sanchez Guerra M JRey and F Costa-Jussa ldquoNeuropathology and pathogenesisof encephalitis following amyloid-Neuropathology and patho-genesis of encephalitis following amyloid-beta immunizationin Alzheimerrsquos disease immunization in Alzheimerrsquos diseaserdquoBrain Pathology vol 14 no 1 pp 11ndash20 2004
[22] D H Cribbs A Ghochikyan V Vasilevko et al ldquoAdjuvant-dependent modulation ofTh1 andTh2 responses to immuniza-tion with 120573-amyloidrdquo International Immunology vol 15 no 4pp 505ndash514 2003
[23] C Holmes D Boche D Wilkinson et al ldquoLong-term effectsof A120573
42immunisation in Alzheimerrsquos disease follow-up of a
randomised placebo-controlled phase I trialrdquo The Lancet vol372 no 9634 pp 216ndash223 2008
[24] I Carrera I Etcheverrıa L Fernandez-Novoa V LombardiR Cacabelos and C Vigo ldquoVaccine development to treatAlzheimerrsquos disease neuropathology in APPPS1 transgenicmicerdquo International Journal of Alzheimerrsquos Disease vol 2012Article ID 376138 17 pages 2012
[25] I Carrera I Etcheverrıa Y Li et al ldquoImmunocytochemicalcharacterization of Alzheimer disease hallmarks in APPPS1transgenic mice treated with a new anti-amyloid-120573 vaccinerdquoBioMed Research International vol 2013 Article ID 709145 12pages 2013
[26] V Anelli R Bassi G Tettamanti P Viani and L Riboni ldquoExtra-cellular release of newly synthesized sphingosine-1-phosphateby cerebellar granule cells and astrocytesrdquo Journal of Neuro-chemistry vol 92 no 5 pp 1204ndash1215 2005
[27] S Schroeter K Khan R Barbour et al ldquoImmunotherapyreduces vascular amyloid-120573 in PDAPP micerdquo Journal of Neu-roscience vol 28 no 27 pp 6787ndash6793 2008
[28] Z Suo A Cox N Bartelli et al ldquoGRK5 deficiency leads to earlyAlzheimer-like pathology and working memory impairmentrdquoNeurobiology of Aging vol 28 no 12 pp 1873ndash1888 2007
[29] W Kamphuis C Mamber MMoeton et al ldquoGFAP isoforms inadult mouse brain with a focus on neurogenic astrocytes andreactive astrogliosis in mouse models of Alzheimer diseaserdquoPLoS ONE vol 7 no 8 Article ID e42823 2012
[30] J Lang C Vigo-Pelfrey and F Martin ldquoLiposomes composedof partially hydrogenated egg phosphatidylcholines fatty acidcomposition thermal phase behavior and oxidative stabilityrdquoChemistry and Physics of Lipids vol 53 no 1 pp 91ndash101 1990
[31] D Morgan D M Diamond P E Gottschall et al ldquoA 120573peptide vaccination prevents memory loss in an animal modelof Alzheimerrsquos diseaserdquo Nature vol 408 no 6815 pp 982ndash9852000
[32] M L Brownlow L Benner D DrsquoAgostino M N Gordon andD Morgan ldquoKetogenic diet improves motor performance but
not cognition in two mouse models of Alzheimerrsquos pathologyrdquoPLoS ONE vol 8 no 9 Article ID e75713 2013
[33] M Pride P Seubert M Grundman M Hagen J Eldridgeand R S Black ldquoProgress in the active immunotherapeu-tic approach to Alzheimerrsquos disease clinical investigationsinto AN1792-associated meningoencephalitisrdquo Neurodegenera-tive Diseases vol 5 no 3-4 pp 194ndash196 2008
[34] CWiessner K-HWiederhold A C Tissot et al ldquoThe second-generation activeA120573 immunotherapyCAD106 reduces amyloidaccumulation in APP transgenic mice while minimizing poten-tial side effectsrdquoThe Journal of Neuroscience vol 31 no 25 pp9323ndash9331 2011
[35] D Lambracht-Washington and R Rosenberg ldquoAdvances in thedevelopment of vaccines for Alzheimerrsquos diseaserdquo DiscoveryMedicine vol 15 no 84 pp 319ndash326 2013
[36] A Kimura T Ohmori R Ohkawa et al ldquoEssential roles ofsphingosine 1-phosphateS1P
1receptor axis in the migration of
neural stem cells toward a site of spinal cord injuryrdquo Stem Cellsvol 25 no 1 pp 115ndash124 2007
[37] S S Lee C H Woo J D Chang and J H Kim ldquoRoles of Racand cytosolic phospholipase A2 in the intracellular signalling inresponse to titanium particlesrdquo Cellular Signalling vol 15 no 3pp 339ndash345 2003
[38] E Masliah L Hansen A Adame et al ldquoA120573 vaccination effectson plaque pathology in the absence of encephalitis inAlzheimerdiseaserdquo Neurology vol 64 no 1 pp 129ndash131 2005
[39] A Pilorget B Annabi F Bouzeghrane J Marvaldi J Luisand R Beliveau ldquoInhibition of angiogenic properties of brainendothelial cells by platelet-derived sphingosine-1-phosphaterdquoJournal of Cerebral Blood Flow and Metabolism vol 25 no 9pp 1171ndash1182 2005
[40] A Wang P Das R C Switzer III T E Golde and J LJankowsky ldquoRobust amyloid clearance in a mouse model ofAlzheimerrsquos disease provides novel insights into themechanismof amyloid-120573 immunotherapyrdquo Journal of Neuroscience vol 31no 11 pp 4124ndash4136 2011
[41] G Krabbe A Halle V Matyash et al ldquoFunctional impairmentof microglia coincides with Beta-amyloid deposition in micewith Alzheimer-like pathologyrdquo PLoS ONE vol 8 no 4 ArticleID e60921 2013
[42] Y H Liu B Giunta H-D Zhou J Tan and Y J WangldquoImmunotherapy for Alzheimer disease-the challenge ofadverse effectsrdquo Nature Reviews Neurology vol 8 no 8 pp465ndash469 2012
[43] B Solomon R Koppel D Frankel and E Hanan-AharonldquoDisaggregation of Alzheimer120573-amyloid by site-directedmAbrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 94 no 8 pp 4109ndash4112 1997
[44] F Bard C Cannon R Barbour et al ldquoPeripherally adminis-tered antibodies against amyloid 120573-peptide enter the centralnervous system and reduce pathology in a mouse model ofAlzheimer diseaserdquo Nature Medicine vol 6 no 8 pp 916ndash9192000
[45] R B DeMattos K R Bales D J Cummins J C Dodart S MPaul and D M Holtzman ldquoPeripheral anti-A120573 antibody altersCNS and plasma A120573 clearance and decreases brain A120573 burdenin a mouse model of Alzheimerrsquos diseaserdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 98 no 15 pp 8850ndash8855 2001
[46] C A Lemere E T Spooner J LaFrancois et al ldquoEvidence forperipheral clearance of cerebral A120573 protein following chronic
16 BioMed Research International
active A120573 immunization in PSAPP micerdquo Neurobiology ofDisease vol 14 no 1 pp 10ndash18 2003
[47] T Town M Vendrame A Patel et al ldquoReduced Th1 andenhanced Th2 immunity after immunization with Alzheimerrsquos120573-amyloid
1minus42rdquo Journal of Neuroimmunology vol 132 no 1-2
pp 49ndash59 2002[48] R Cacabelos ldquoPharmacogenomics in Alzheimerrsquos diseaserdquo
Methods in Molecular Biology vol 448 pp 213ndash357 2008[49] R Cacabelos ldquoPharmacogenomics and therapeutic strategies
for dementiardquo Expert Review of Molecular Diagnostics vol 9no 6 pp 567ndash611 2009
[50] D van Dam R DrsquoHooge M Staufenbiel C van Ginneken Fvan Meir and P P de Deyn ldquoAge-dependent cognitive declinein the APP23 model precedes amyloid depositionrdquo EuropeanJournal of Neuroscience vol 17 no 2 pp 388ndash396 2003
[51] M Dumont C Strazielle M Staufenbiel and R LalondeldquoSpatial learning and exploration of environmental stimuli in24-month-old female APP23 transgenic mice with the Swedishmutationrdquo Brain Research vol 1024 no 1-2 pp 113ndash121 2004
[52] R Lalonde M Dumont M Staufenbiel C Sturchler-Pierratand C Strazielle ldquoSpatial learning exploration anxiety andmotor coordination in female APP23 transgenic mice with theSwedish mutationrdquo Brain Research vol 956 no 1 pp 36ndash442002
[53] F J Huff F Boller F Lucchelli R Querriera J Beyer and SBelle ldquoThe neurologic examination in patients with probableAlzheimerrsquos diseaserdquo Archives of Neurology vol 44 no 9 pp929ndash932 1987
[54] A F Pettersson M Engardt and L O Wahlund ldquoActivitylevel and balance in subjects with mild Alzheimerrsquos diseaserdquoDementia and Geriatric Cognitive Disorders vol 13 no 4 pp213ndash216 2002
[55] L Calandreau B Desgranges R Jaffard and A DesmedtldquoSwitching from contextual to tone fear conditioning and viceversa the key role of the glutamatergic hippocampal-lateralseptal neurotransmissionrdquo Learning amp Memory vol 17 no 9pp 440ndash443 2010
[56] F Trinchese S Liu F Battaglia SWalter PMMathews andOArancio ldquoProgressive age-related development of Alzheimer-like pathology in APPPS1 micerdquo Annals of Neurology vol 55no 6 pp 801ndash814 2004
[57] D M Wilcock A Rojiani A Rosenthal et al ldquoPassiveimmunotherapy against A120573 in aged APP-transgenic micereverses cognitive deficits and depletes parenchymal amyloiddeposits in spite of increased vascular amyloid and microhem-orrhagerdquo Journal of Neuroinflammation vol 1 article 24 2004
[58] S A Ferguson S Sarkar and L C Schmued ldquoLongitudinalbehavioral changes in the APPPS1 transgenic AlzheimerrsquosDisease modelrdquo Behavioural Brain Research vol 242 no 1 pp125ndash134 2013
[59] D Wilcock and C Colton ldquoImmunotherapy vascular pathol-ogy and microhemorrhages in transgenic micerdquo CNS andNeurological DisordersmdashDrug Targets vol 8 no 1 pp 50ndash642009
[60] M Grundman S Gilman R S Black et al ldquoAn alternativemethod for estimating efficacy of the AN1792 vaccine foralzheimer diseaserdquo Neurology vol 71 no 9 pp 697ndash698 2008
[61] D Boche E Zotova R O Weller et al ldquoConsequence of A120573immunization on the vasculature of human Alzheimerrsquos diseasebrainrdquo Brain vol 131 no 12 pp 3299ndash3310 2008
[62] M Pfeifer S Boncristiano L Bondolfi et al ldquoCerebral hemor-rhage after passive anti-a120573 immunotherapyrdquo Science vol 298no 5597 p 1379 2002
[63] M M Racke L I Boone D L Hepburn et al ldquoExacerbationof cerebral amyloid angiopathy-associatedmicrohemorrhage inamyloid precursor protein transgenic mice by immunotherapyis dependent on antibody recognition of deposited forms ofamyloid 120573rdquo Journal of Neuroscience vol 25 no 3 pp 629ndash6362005
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

16 BioMed Research International
active A120573 immunization in PSAPP micerdquo Neurobiology ofDisease vol 14 no 1 pp 10ndash18 2003
[47] T Town M Vendrame A Patel et al ldquoReduced Th1 andenhanced Th2 immunity after immunization with Alzheimerrsquos120573-amyloid
1minus42rdquo Journal of Neuroimmunology vol 132 no 1-2
pp 49ndash59 2002[48] R Cacabelos ldquoPharmacogenomics in Alzheimerrsquos diseaserdquo
Methods in Molecular Biology vol 448 pp 213ndash357 2008[49] R Cacabelos ldquoPharmacogenomics and therapeutic strategies
for dementiardquo Expert Review of Molecular Diagnostics vol 9no 6 pp 567ndash611 2009
[50] D van Dam R DrsquoHooge M Staufenbiel C van Ginneken Fvan Meir and P P de Deyn ldquoAge-dependent cognitive declinein the APP23 model precedes amyloid depositionrdquo EuropeanJournal of Neuroscience vol 17 no 2 pp 388ndash396 2003
[51] M Dumont C Strazielle M Staufenbiel and R LalondeldquoSpatial learning and exploration of environmental stimuli in24-month-old female APP23 transgenic mice with the Swedishmutationrdquo Brain Research vol 1024 no 1-2 pp 113ndash121 2004
[52] R Lalonde M Dumont M Staufenbiel C Sturchler-Pierratand C Strazielle ldquoSpatial learning exploration anxiety andmotor coordination in female APP23 transgenic mice with theSwedish mutationrdquo Brain Research vol 956 no 1 pp 36ndash442002
[53] F J Huff F Boller F Lucchelli R Querriera J Beyer and SBelle ldquoThe neurologic examination in patients with probableAlzheimerrsquos diseaserdquo Archives of Neurology vol 44 no 9 pp929ndash932 1987
[54] A F Pettersson M Engardt and L O Wahlund ldquoActivitylevel and balance in subjects with mild Alzheimerrsquos diseaserdquoDementia and Geriatric Cognitive Disorders vol 13 no 4 pp213ndash216 2002
[55] L Calandreau B Desgranges R Jaffard and A DesmedtldquoSwitching from contextual to tone fear conditioning and viceversa the key role of the glutamatergic hippocampal-lateralseptal neurotransmissionrdquo Learning amp Memory vol 17 no 9pp 440ndash443 2010
[56] F Trinchese S Liu F Battaglia SWalter PMMathews andOArancio ldquoProgressive age-related development of Alzheimer-like pathology in APPPS1 micerdquo Annals of Neurology vol 55no 6 pp 801ndash814 2004
[57] D M Wilcock A Rojiani A Rosenthal et al ldquoPassiveimmunotherapy against A120573 in aged APP-transgenic micereverses cognitive deficits and depletes parenchymal amyloiddeposits in spite of increased vascular amyloid and microhem-orrhagerdquo Journal of Neuroinflammation vol 1 article 24 2004
[58] S A Ferguson S Sarkar and L C Schmued ldquoLongitudinalbehavioral changes in the APPPS1 transgenic AlzheimerrsquosDisease modelrdquo Behavioural Brain Research vol 242 no 1 pp125ndash134 2013
[59] D Wilcock and C Colton ldquoImmunotherapy vascular pathol-ogy and microhemorrhages in transgenic micerdquo CNS andNeurological DisordersmdashDrug Targets vol 8 no 1 pp 50ndash642009
[60] M Grundman S Gilman R S Black et al ldquoAn alternativemethod for estimating efficacy of the AN1792 vaccine foralzheimer diseaserdquo Neurology vol 71 no 9 pp 697ndash698 2008
[61] D Boche E Zotova R O Weller et al ldquoConsequence of A120573immunization on the vasculature of human Alzheimerrsquos diseasebrainrdquo Brain vol 131 no 12 pp 3299ndash3310 2008
[62] M Pfeifer S Boncristiano L Bondolfi et al ldquoCerebral hemor-rhage after passive anti-a120573 immunotherapyrdquo Science vol 298no 5597 p 1379 2002
[63] M M Racke L I Boone D L Hepburn et al ldquoExacerbationof cerebral amyloid angiopathy-associatedmicrohemorrhage inamyloid precursor protein transgenic mice by immunotherapyis dependent on antibody recognition of deposited forms ofamyloid 120573rdquo Journal of Neuroscience vol 25 no 3 pp 629ndash6362005
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014