age validation of juvenile shortfi n mako isurus oxy ... · cerna and licandeo, 2009; okamura and...
TRANSCRIPT

147
Age validation of juvenile Shortfi n Mako (Isurus oxyrinchus) tagged and marked with oxytetracycline off southern California
R. J. David Wells (contact author)1,2
Susan E. Smith1
Suzanne Kohin1
Ellen Freund1
Natalie Spear1
Darlene A. Ramon1
Email address for contact author: [email protected]
1 Fisheries Resources Division Southwest Fisheries Science Center National Marine Fisheries Service, NOAA 8901 La Jolla Shores Drive La Jolla, California 92037
Present address for contact author: Department of Marine Biology Texas A&M University at Galveston 1001 Texas Clipper Rd. Galveston, Texas 77553
Manuscript submitted 23 August 2012.Manuscript accepted 20 February 2013.Fish. Bull 111:147–160 (2013).doi 10.7755/FB.111.2.3
The views and opinions expressed or implied in this article are those of the author (or authors) and do not necessar-ily refl ect the position of the National Marine Fisheries Service, NOAA.
Abstract—The purpose of this study was to validate aging results of ju-venile Shortfin Mako (Isurus oxy-rinchus) by vertebral band counts. Vertebrae of 29 juvenile Shortfin Mako marked with oxytetracycline (OTC) were obtained from tag-re-capture activities to determine cen-trum growth-band deposition. Tag-ging occurred off southern Califor-nia from 1996 to 2010, and time at liberty of the 29 sharks ranged from 4 months to 4.4 years (mean=1.3 years). Growth information also was obtained from length-frequency modal analyses (MULTIFAN and MIXDIST) by using a 29-year data set of commercial and research catch data, in addition to a tag-recapture growth model (e.g, the GROTAG model). For vertebrae samples used for age validation, shark size at time of release ranged from 79 to 142 cm fork length (FL) and from 98 to 200 cm FL at recapture. Results from band counts of vertebrae distal to OTC marks indicate 2 band pairs (2 translucent and 2 opaque) are formed each year for Shortfin Mako of the size range examined. Length-frequency analyses identified 3 age-class modes. Growth rate estimates from 26.5 to 35.5 cm/year were cal-culated for the first age-class mode (85 cm FL) and from 22.4 to 28.6 cm/year for the second age-class mode (130 cm FL). Results from the tag-recapture growth model revealed fast growth during time at liberty for tagged fish of the 2 youngest age classes. Collectively, these methods suggest rapid growth of juvenile Shortfin Mako in the southern Cali-fornia study area and indicate bian-nual deposition of growth bands in vertebrae for the first 5 years.
For almost 3 decades, researchers in various parts of the world have fo-cused on the problem of accurately interpreting age and growth in the Shortfi n Mako (Isurus oxyrinchus) (e.g., Cailliet and Bedford, 1983; Cailliet et al., 1983; Pratt and Casey, 1983; Casey and Kohler, 1992; Cam-pana et al., 2002; Bishop et al.1; Ri-bot-Carballal et al., 2005; Bishop et al., 2006; Natanson et al., 2006; Ar-dizzone et al., 2006; Maia et al., 2007; Cerna and Licandeo, 2009; Okamura and Semba, 2009; Semba et al., 2009). Driving these efforts is the need to better assess the vulnerability of Shortfi n Mako to harvest in commer-cial and recreational fi sheries and as bycatch in longline and driftnet fi sh-eries from high-seas fl eets (Stevens, 2008). Studies of the demographic
1 Bishop, S. D. H., M. P. Francis, and C. Duffy. 2004. Age, growth, maturity, longevity and natural mortality of the Shortfi n Mako shark (Isurus oxyrinchus) in New Zealand waters. Working Paper SCTB17, BIO-4, National Institute of Water and Atmospheric Research, New Zealand, Ministry of Fisheries, NZ, 34 p. Presented at the 17th Meeting of the Standing Committee on Tuna and Bill-fi sh (SCTB), Majuro, Marshall Islands, 9–18 August 2004.
dynamics of sharks have shown that average age at fi rst maturity and the relative rate of growth that deter-mines this parameter is one of the leading factors that affect the abil-ity of most sharks to rebound from harvest pressures (Smith et al., 1998; Cortés, 2002; Garcia et al., 2008). Ac-curate age determinations also are necessary for calculations of growth and mortality rates, age at recruit-ment, and longevity.
The Shortfi n Mako is an epipe-lagic species distributed in temper-ate and tropical seas worldwide (Compagno, 2001) and seldom found in water temperatures lower than 13–17°C (Casey and Kohler, 1992; Stevens, 2008). Shortfi n Mako occur off the U.S. West Coast principally off California and Oregon, and catches are associated primarily with warm sea-surface temperatures from 15° to 25°C (PFMC, 2011). Most Shortfi n Mako off the U.S. West Coast are sex-ually immature, and high recapture rates for tagged juveniles show that young individuals remain for about 2 years in nearshore California waters (Taylor and Bedford, 2001), where they are more frequently taken in the summer months and are only
2 Department of Marine Biology Texas A&M University at Galveston 1001 Texas Clipper Rd. Galveston, Texas 77553

148 Fishery Bulletin 111(2)
rarely found below the thermocline (Holts and Bed-ford, 1993). Large fi sh approaching 200 cm fork length (FL) disappear from the catch and presumably move offshore to a more oceanic and highly migratory exis-tence, where most males are mature but females are not (Mollet et al., 2000; Joung and Hsu, 2005; Semba et al., 2011).
Researchers who study the age and growth of this species have used pairs of alternating bands of dif-ferent mineralization in vertebral centra to estimate age. These estimates differ depending on assumptions about the number of band pairs that represent a single year of growth. Bishop et al. (2006) noted that despite different conclusions in these studies, all of them ap-pear to have produced relatively similar growth curves if the same rate of band deposition was assumed. This fi nding indicates that techniques used by different research groups to identify countable band pairs are generally alike, with the greatest difference being as-sumptions about rates of band deposition. Smaller dis-parities may be due to ontogeny, geographic variability, divergent methods of preparing and interpreting verte-bral banding patterns, diffi culties in interpreting band-ing patterns, differences in sample sizes, or limitations of the growth models used (Bishop et al., 2006).
Although considerable advances have been made in determination of age and growth of Shortfi n Mako, questions remain as to why current length-at-age models underestimate growth of young fi sh, especial-ly fi sh ≤200 cm FL (Pratt and Casey, 1983; Bishop et al.1; Maia et al., 2007). Bishop et al. (2006) suggested that this discrepancy may be due to the inability of commonly used growth models to reconcile such rap-id juvenile growth with the slow growth predicted for subadults and adult sharks. Direct age-validation techniques, such as oxytetracycline (OTC) marking and recapture of an adequate sample size of Shortfi n Mako ≤200 cm FL, therefore, are needed to help resolve the issue of band-deposition rates in juvenile Shortfi n Mako vertebrae.
In this study, we examined OTC-marked vertebrae of 29 juvenile Shortfi n Mako (≤200 cm FL) released off California and later recaptured from 2000 to 2010 (at liberty from 145 days to 4.4 years). OTC is deposited at sites of active calcifi cation, such as vertebral centra, and is known to remain distinct for at least 20 years in certain fi nfi sh, such as the Sablefi sh (Anoplopoma fi m-bria) (Beamish and McFarlane, 2000), and sharks, such as the Leopard Shark (Triakis semifasciata) (Smith et al., 2003). The combination of tag-recapture and chemi-cal marking is thought to be the most robust method of age validation (Campana, 2001; Goldman, 2005). These methods test the accuracy of the counts of vertebral band pairs as annual indicators through observation of the banding pattern deposited distally to the OTC mark during the known time at liberty. To supplement this information, we also conducted analyses of at-lib-erty growth of tagged-recaptured sharks and length-
frequency data from 3 decades of length data from commercial and research sources.
Materials and methods
Tagging methods
Sharks for tagging and OTC injection were captured in the Southern California Bight (SCB) (Fig. 1) with baited pelagic longlines and identifi ed as Shortfi n Mako through the method described by Compagno (2001). Leaders were unsnapped from the main line, and sharks were guided into a semisubmerged metal tagging cradle at the stern of the vessel. The cradle was then raised to facilitate tagging, measuring, and OTC injection, while the eyes of the shark were cov-ered with a wet chamois cloth and a saltwater ven-tilation hose continuously ran water over the shark’s gills. Each shark was tagged on the dorsal fi n with a plastic Rototag2 (Dalton ID, Henley-on-Thames, UK) la-beled with contact and reward information in English and Spanish and with instructions to measure the fi sh and save the vertebrae. Most sharks also were double-tagged with a spaghetti tag placed in the dorsal mus-culature beneath the fi rst dorsal fi n.
At tagging, the sex of each shark was determined and each shark was measured (straight line FL or to-tal length [TL]) to the nearest centimeter with a sta-tionary meter stick fi tted to the shark tagging cradle. Sharks were given an intraperitoneal injection of OTC at a dose rate of 25 mg/kg of body weight; the dose was estimated with a length-weight dose table developed from length-weight measurements of Shortfi n Mako measured by NOAA scientifi c observers for the Califor-nia drift gillnet fi shery (Rasmussen3).
Laboratory methods for processing vertebrae
Past studies have shown that band counts are consis-tent throughout the vertebral column in the Shortfi n Mako, indicating that vertebrae from any region can be used in age analysis, although the larger central ver-tebrae are easier to read because of wider band spac-ing (Bishop et al.1; Bishop et al., 2006; Natanson et al., 2006). OTC-marked vertebrae were obtained from Shortfi n Mako recaptured on research cruises and com-mercial and recreational fi shing vessels between 2000 and 2010. The widest diameter vertebral centra in a given sample were chosen for sectioning. We used only OTC-marked vertebrae from tag recaptures at liberty longer than 124 days (or ~4 months) to incorporate one full season into band-pair counts.
2 Mention of trade names or commercial companies is for iden-tifi cation purposes only and does not imply endorsement by the National Marine Fisheries Service, NOAA.
3 Rasmussen, R. 1995. Unpubl. data. Southwest Fisheries Science Center, NOAA, La Jolla, CA.
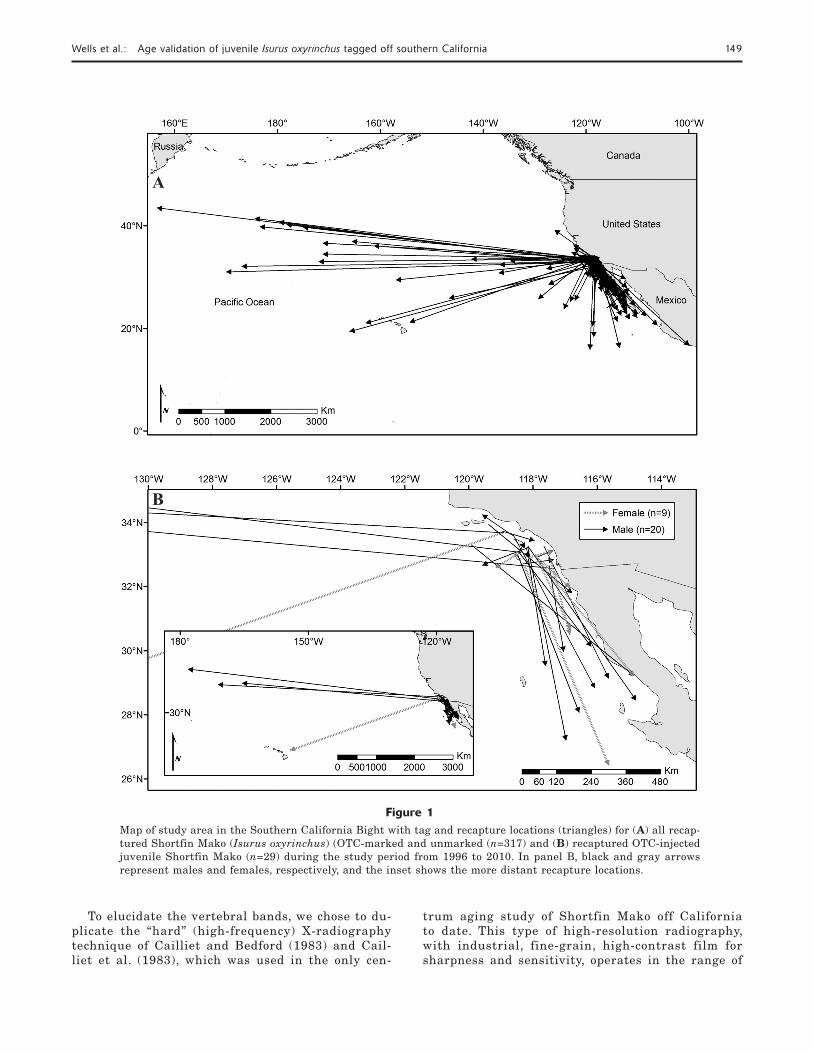
Wells et al.: Age validation of juvenile Isurus oxyrinchus tagged off southern California 149
Figure 1Map of study area in the Southern California Bight with tag and recapture locations (triangles) for (A) all recap-tured Shortfin Mako (Isurus oxyrinchus) (OTC-marked and unmarked (n=317) and (B) recaptured OTC-injected juvenile Shortfin Mako (n=29) during the study period from 1996 to 2010. In panel B, black and gray arrows represent males and females, respectively, and the inset shows the more distant recapture locations.
A
B
To elucidate the vertebral bands, we chose to du-plicate the “hard” (high-frequency) X-radiography technique of Cailliet and Bedford (1983) and Cail-liet et al. (1983), which was used in the only cen-
trum aging study of Shortfin Mako off California to date. This type of high-resolution radiography, with industrial, fine-grain, high-contrast film for sharpness and sensitivity, operates in the range of

150 Fishery Bulletin 111(2)
10–110 kV at 3 mA and penetrates the highly calci-fied vertebrae.
Samples were stored frozen until processed and kept from light and excessive UV exposure to preserve the OTC time mark. Whole centra were separated, cleaned of excess tissue, rinsed and air-dried. Once dry, 2 centra from each sample were chosen, and 2 types of sections were prepared with a low-speed circular saw (IsoMet, Buehler, Lake Bluff, IL). To duplicate the Cailliet et al. (1983) method of X-raying the whole centrum face and to avoid the double image of the anterior and poste-rior face superimposed in X-ray images, a transverse cut was made through the centrum to create 2 equal halves of the centrum face. Additionally, longitudinal (sagittal) sections, 0.3–1.0 mm thick, were taken from the center of the second vertebrae to further elucidate banding patterns, especially growth zones along the centrum edge that are sometimes diffi cult to read on centrum face X-rays (Cailliet et al., 1983). Sections were then mounted and examined under a dissecting microscope at 7.5–10.0× magnifi cation, with refl ected long-wave UV light (365 nm) to illuminate the OTC mark. A metal pin was glued into place to visualize the position of the brightest, most distal edge of the OTC mark before it was X-rayed. Samples were X-rayed with a General Electric (Fairfi eld, CT) Mobile 100-15 X-ray unit for exposures between 15 and 45 s at 5 mA and 20–40 kV by using Kodak Industrex M100 fi lm (Readypack II; Eastman Kodak Co., Rochester, NY). X-radiographs were photographed with a Leica Z16 APO dissecting microscope with substage illumination and a Leica DFC420 digital camera (Leica Microsystems, Wetzlar, Germany).
Standardization of band-reading techniques and terminology
We examined data collected by commercial drift gill-net observers and from research longline surveys and determined the peak in abundance of postpartum-size Shortfi n Mako <70 cm FL to identify peak parturition time off southern California. Most individuals (90%) were collected between August and November, provid-ing a tentative parturition time in our study region. Size at birth has been estimated at approximately 63 cm FL (70 cm TL), with parturition occurring year-round according to Mollet et al. (2000); parturition may occur primarily in summer in the South Pacifi c (Duffy and Francis, 2001) and eastern North Atlantic (Maia et al., 2007). The birth band for counting purposes was identifi ed as the most pronounced calcifi ed fi rst band distal to the centrum focus and indicated by a change in the angle of the centra (Bishop et al., 2006). Seasons were defi ned according to solstice and equinox periods in the Northern Hemisphere.
Band-pair counts
Band pairs were counted from digital images of X-ray photographs on a computer screen. We referred to the
original X-rays if more detail was desired. As in Bishop et al.1 and Bishop et al. (2006), counts excluded the birth band, which represents age 0. Alternating pairs of translucent bands (hypomineralized; appearing dark in X-ray) and opaque bands (hypermineralized; appearing light in X-ray) were assumed to represent one complete band pair. Two separate band counts were made: 1) to-tal band pairs, or bands distal to the presumed birth band, and 2) band pairs distal to the OTC mark. Band-pair counting began for the former at the distal edge of the fi rst translucent zone beyond the birth band and for the latter at the distal edge of the fi rst translucent zone beyond the OTC mark. In many cases, the OTC mark was directly on a translucent zone, but this zone was not included in the distal-to-OTC counts because only partial growth occurred during this period. This method results in counts that are conservative; counts are lower than they would be if this zone were includ-ed. If a count ended with a partial band pair (observed when the centrum edge was opaque), a plus sign was appended to the count number. For statistical analy-ses, the plus sign was converted to an arbitrary par-tial band count of 0.5. The corpus calcareum (centrum “arm”) was used as a primary counting surface, and bands in the intermedialia were used for confi rmation of a band pair, although we were not always successful in acquisition of sections with the fragile intermedialia intact (Goldman, 2005; Branstetter and Musick, 1994).
Each sample was examined and counted indepen-dently by 3 readers. Bands were “blind “counted with-out knowledge of Shortfi n Mako length, sex, or time at liberty. Readers consulted with each other on cri-teria for counts before readings. Samples for which there was disagreement were counted a second time with X-ray images from which corresponding sample numbers had been removed and placed in a random order. Counts with similar readings among readers were deemed fi nal; however, several samples did not have similar counts. For those samples without similar counts, the average number of band pairs was reported because differences were minor and no readings were deemed irreconcilable.
Differences among fi nal readings of each of the 3 in-dependent readers were examined through an analysis of variance (ANOVA) with readers as the dependent variable. A least-squares linear regression analysis was performed, and the null hypothesis that the slope (b) of the relationship between the number of band pairs and time was 1:1 (a situation that occurred if one opaque and one translucent band were deposited each year) was tested with a two-tailed t-test (Kusher et al., 1992). Age bias was investigated with age-bias plots and chi-square tests of symmetry by using the contingency table methods of Bowker (1948) and Hoe-nig et al. (1995). Differences in band-pair counts among readers were evaluated by the average percent error (APE) (Beamish and Fournier, 1981) and coeffi cient of variation (CV) (Chang, 1982) for readings distal to both the OTC mark and birth band.

Wells et al.: Age validation of juvenile Isurus oxyrinchus tagged off southern California 151
Length-frequency analysis
Length-frequency distributions of juvenile Shortfi n Mako were analyzed to estimate size at age of the fi rst 3 age classes and to compare to vertebrae readings. The MULTIFAN model (Fournier et al., 1990) was one of 2 techniques used to analyze length-frequency data on the basis of size distributions. This model simultane-ously analyzes multiple length-frequency distributions using a maximum likelihood method to estimate the number of age classes and proportions of fi sh at age. Tests for signifi cance (best-fi t growth parameters) were made through the use of likelihood ratio tests. First, a systematic search was performed to estimate the num-ber of age classes and to hold the standard deviations of length constant across all age classes. Next, stan-dard deviations of length were allowed to vary across age classes. The MIXDIST package (MacDonald and Pitcher, 1979) in R, vers. 2.8.0 (R Development Core Team, 2008) was the second length-frequency analysis used. This analysis uses a maximum likelihood method to estimate proportions of fi sh at age with the added benefi t of fi tting non-normally distributed data.
Data for both techniques were analyzed annually and came from 2 sources: 1) fi shery-dependent data from the California drift gillnet fi shery (1981–2009), which operates between May and January, and 2) fi sh-ery-independent data from longline research surveys of juvenile Shortfi n Mako conducted by the NOAA South-west Fisheries Science Center (1993–2009) between June and August of each year. A mixture of length measurements taken across study years: TL, FL and alternate length (AL, straight line distance between the origins of the fi rst and second dorsal fi ns) were standardized into FL for this study to allow for com-parison of our results with the results of other studies. The following length conversions were obtained from the 2 sources and used to standardize data for subse-quent length-frequency analyses:
FL = 0.913 × (TL) – 0.397, coeffi cient of determination (r2) = 0.986 (n=2177)
FL = 2.402 × (AL) + 9.996, r2=0.957 (n=3250).
Size data were combined between sexes because no signifi cant difference existed (P=0.769), and size bins of 5 cm, ranging from 55 to 265 cm FL, were used.
Growth of tagged and recaptured mako sharks
Growth rates were calculated for 1) recaptured, OTC-marked sharks and 2) angler- and research-re-leased, unlabeled sharks for which reliable length es-timates were available (n=62 of 317 returns) (Fig. 1A). Growth was estimated with the tag-recapture growth model GROTAG (Francis 1988a, 1988b) on the basis of length and time-at-liberty. This model was chosen be-cause age-based and length-based growth models often differ, and it is a useful alternative for comparison of
growth rates at particular sizes (Francis, 1988a; Na-tanson et al., 2006; Claisse et al., 2009). We used a maximum likelihood approach with this model to esti-mate growth rates (gα and gβ) at 2 selected lengths (α and β), a CV of growth variability, mean measurement error and standard deviation, and outlier probability. Therefore, estimated growth of a Shortfi n Mako, i, was calculated with the following equation:
�Li = ((βgα – αgβ) / (gα –gβ) – Li) / (1 – (1 + (gα –gβ) / α –β)�Ti),
where Li = length at release; and �Ti = the tag deployment time.
Results
Tagging and recapturing oxytetracycline-marked sharks
Off southern California from 1996 to 2010, 940 OTC-injected Shortfi n Mako were tagged and released (Fig. 1, A and B). Of the subset of released sharks for which sexes were determined, 67% were males and 33% were females. Average size at release was 110 cm FL (±0.80 SE). Of the released OTC-marked sharks, 35 were re-captured from 2000 to 2010. Of these 35 sharks, 29 fi sh were selected for this study because they had been at liberty for ≥4 months and OTC marks in the vertebrae fl uoresced (Table 1, Fig. 1B). Five samples were ex-cluded from analysis because of a short time-at-liberty (within a range from 24 to 60 days), provided no infor-mation on band-pair progression, and in the case of 1 shark, the vertebrae did not fl uoresce.
For the 29 fi sh used in this study, average time-at-liberty was 522 days (±71.0 SE), within a range from 145 to 1594 days, and average size was 109 cm FL (±2.9 SE) at tagging and 148 cm FL (±5.0 SE) at re-capture (Table 1). The average displacement distance (great-circle distance from tagging to recapture loca-tion) for the 29 OTC-marked Shortfi n Mako was 902 km (±291 km SE), within a range from 2 to 5295 km (Fig. 1B). According to results from analyses with lin-ear regressions, no signifi cant effects of time-at-liberty or size-at-tagging existed in relation to total displace-ment distance (P>0.05).
Age validation results
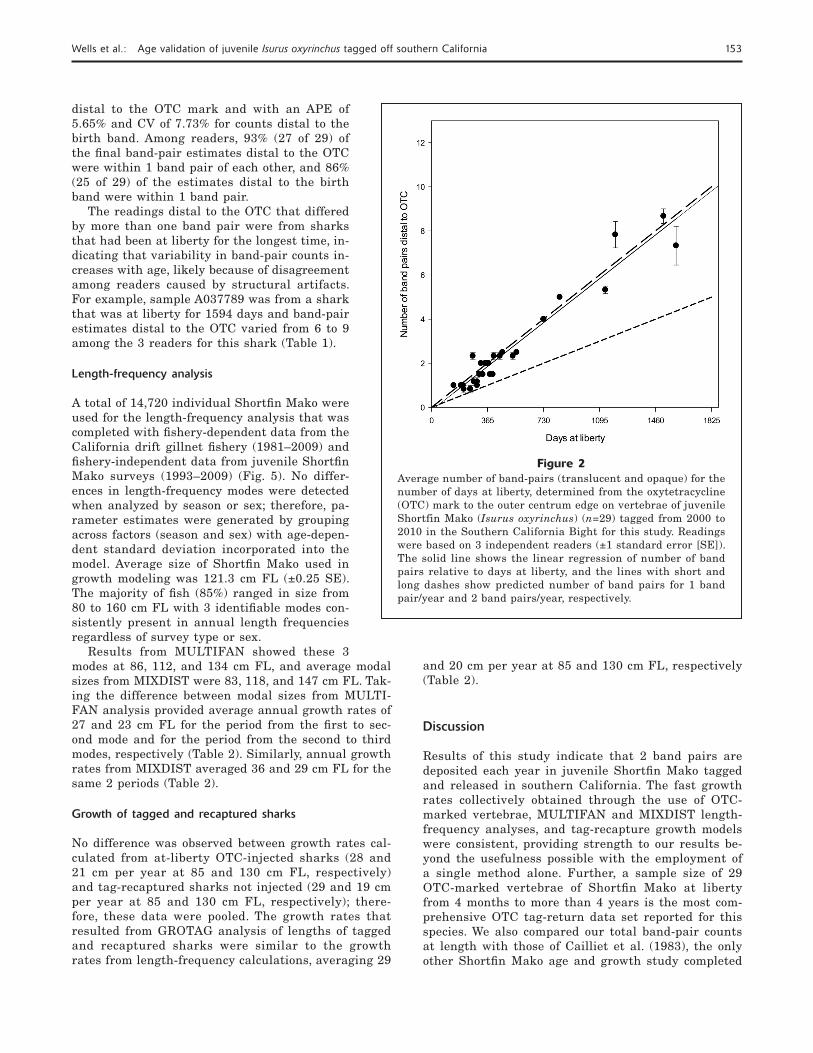
Results from readings of the OTC-marked vertebrae indicated that 2 band pairs are deposited each year in samples analyzed in this study. The observed slope of the relationship between the number of band pairs each year and years-at-liberty signifi cantly differed from the 1:1 relationship (P<0.01). Specifi cally, the av-erage number of band pairs predicted each year was modeled with the following linear regression: average number of band pairs = 1.988 × (number of years-at-large) – 0.136, r2=0.942 (P<0.05) (Fig. 2).
152 Fishery Bulletin 111(2)
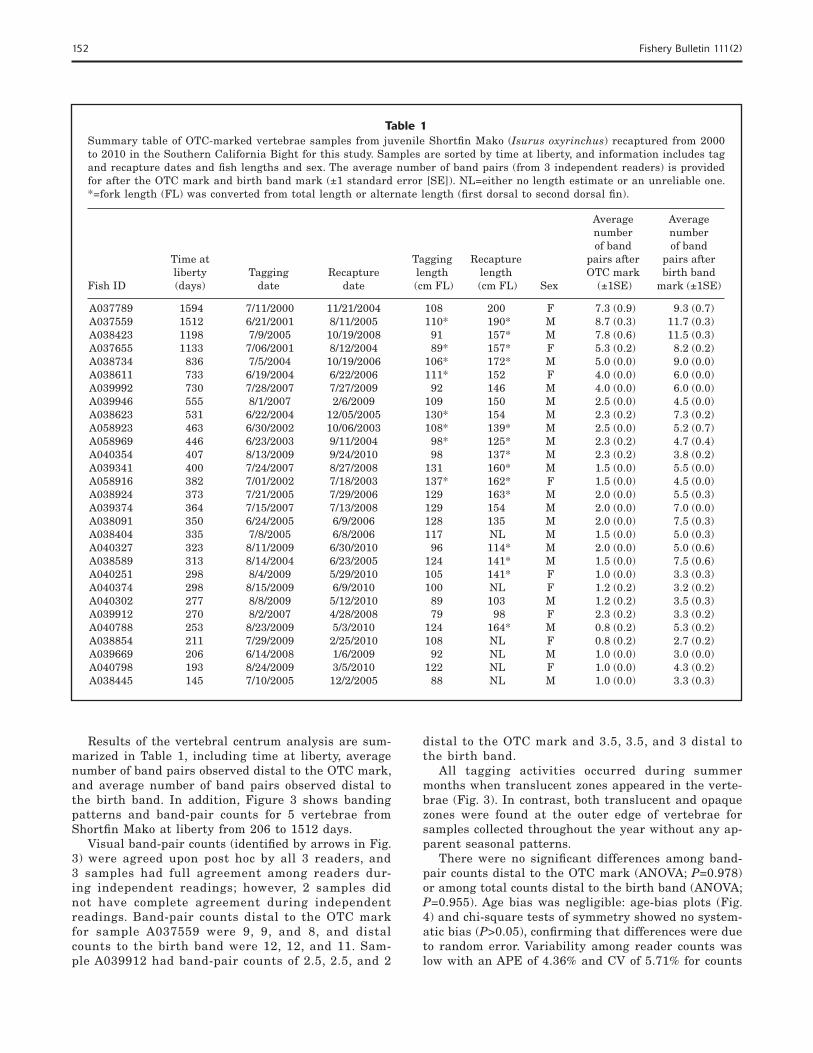
Table 1Summary table of OTC-marked vertebrae samples from juvenile Shortfi n Mako (Isurus oxyrinchus) recaptured from 2000 to 2010 in the Southern California Bight for this study. Samples are sorted by time at liberty, and information includes tag and recapture dates and fi sh lengths and sex. The average number of band pairs (from 3 independent readers) is provided for after the OTC mark and birth band mark (±1 standard error [SE]). NL=either no length estimate or an unreliable one. *=fork length (FL) was converted from total length or alternate length (fi rst dorsal to second dorsal fi n).
Average Average number number of band of band Time at Tagging Recapture pairs after pairs after liberty Tagging Recapture length length OTC mark birth bandFish ID (days) date date (cm FL) (cm FL) Sex (±1SE) mark (±1SE)
A037789 1594 7/11/2000 11/21/2004 108 200 F 7.3 (0.9) 9.3 (0.7)A037559 1512 6/21/2001 8/11/2005 110* 190* M 8.7 (0.3) 11.7 (0.3)A038423 1198 7/9/2005 10/19/2008 91 157* M 7.8 (0.6) 11.5 (0.3)A037655 1133 7/06/2001 8/12/2004 89* 157* F 5.3 (0.2) 8.2 (0.2)A038734 836 7/5/2004 10/19/2006 106* 172* M 5.0 (0.0) 9.0 (0.0)A038611 733 6/19/2004 6/22/2006 111* 152 F 4.0 (0.0) 6.0 (0.0)A039992 730 7/28/2007 7/27/2009 92 146 M 4.0 (0.0) 6.0 (0.0)A039946 555 8/1/2007 2/6/2009 109 150 M 2.5 (0.0) 4.5 (0.0)A038623 531 6/22/2004 12/05/2005 130* 154 M 2.3 (0.2) 7.3 (0.2)A058923 463 6/30/2002 10/06/2003 108* 139* M 2.5 (0.0) 5.2 (0.7)A058969 446 6/23/2003 9/11/2004 98* 125* M 2.3 (0.2) 4.7 (0.4)A040354 407 8/13/2009 9/24/2010 98 137* M 2.3 (0.2) 3.8 (0.2)A039341 400 7/24/2007 8/27/2008 131 160* M 1.5 (0.0) 5.5 (0.0)A058916 382 7/01/2002 7/18/2003 137* 162* F 1.5 (0.0) 4.5 (0.0)A038924 373 7/21/2005 7/29/2006 129 163* M 2.0 (0.0) 5.5 (0.3)A039374 364 7/15/2007 7/13/2008 129 154 M 2.0 (0.0) 7.0 (0.0)A038091 350 6/24/2005 6/9/2006 128 135 M 2.0 (0.0) 7.5 (0.3)A038404 335 7/8/2005 6/8/2006 117 NL M 1.5 (0.0) 5.0 (0.3)A040327 323 8/11/2009 6/30/2010 96 114* M 2.0 (0.0) 5.0 (0.6)A038589 313 8/14/2004 6/23/2005 124 141* M 1.5 (0.0) 7.5 (0.6)A040251 298 8/4/2009 5/29/2010 105 141* F 1.0 (0.0) 3.3 (0.3)A040374 298 8/15/2009 6/9/2010 100 NL F 1.2 (0.2) 3.2 (0.2)A040302 277 8/8/2009 5/12/2010 89 103 M 1.2 (0.2) 3.5 (0.3)A039912 270 8/2/2007 4/28/2008 79 98 F 2.3 (0.2) 3.3 (0.2)A040788 253 8/23/2009 5/3/2010 124 164* M 0.8 (0.2) 5.3 (0.2)A038854 211 7/29/2009 2/25/2010 108 NL F 0.8 (0.2) 2.7 (0.2)A039669 206 6/14/2008 1/6/2009 92 NL M 1.0 (0.0) 3.0 (0.0)A040798 193 8/24/2009 3/5/2010 122 NL F 1.0 (0.0) 4.3 (0.2)A038445 145 7/10/2005 12/2/2005 88 NL M 1.0 (0.0) 3.3 (0.3)
Results of the vertebral centrum analysis are sum-marized in Table 1, including time at liberty, average number of band pairs observed distal to the OTC mark, and average number of band pairs observed distal to the birth band. In addition, Figure 3 shows banding patterns and band-pair counts for 5 vertebrae from Shortfi n Mako at liberty from 206 to 1512 days.
Visual band-pair counts (identifi ed by arrows in Fig. 3) were agreed upon post hoc by all 3 readers, and 3 samples had full agreement among readers dur-ing independent readings; however, 2 samples did not have complete agreement during independent readings. Band-pair counts distal to the OTC mark for sample A037559 were 9, 9, and 8, and distal counts to the birth band were 12, 12, and 11. Sam-ple A039912 had band-pair counts of 2.5, 2.5, and 2
distal to the OTC mark and 3.5, 3.5, and 3 distal to the birth band.
All tagging activities occurred during summer months when translucent zones appeared in the verte-brae (Fig. 3). In contrast, both translucent and opaque zones were found at the outer edge of vertebrae for samples collected throughout the year without any ap-parent seasonal patterns.
There were no signifi cant differences among band-pair counts distal to the OTC mark (ANOVA; P=0.978) or among total counts distal to the birth band (ANOVA; P=0.955). Age bias was negligible: age-bias plots (Fig. 4) and chi-square tests of symmetry showed no system-atic bias (P>0.05), confi rming that differences were due to random error. Variability among reader counts was low with an APE of 4.36% and CV of 5.71% for counts
Wells et al.: Age validation of juvenile Isurus oxyrinchus tagged off southern California 153
Figure 2Average number of band-pairs (translucent and opaque) for the number of days at liberty, determined from the oxytetracycline (OTC) mark to the outer centrum edge on vertebrae of juvenile Shortfin Mako (Isurus oxyrinchus) (n=29) tagged from 2000 to 2010 in the Southern California Bight for this study. Readings were based on 3 independent readers (±1 standard error [SE]). The solid line shows the linear regression of number of band pairs relative to days at liberty, and the lines with short and long dashes show predicted number of band pairs for 1 band pair/year and 2 band pairs/year, respectively.
distal to the OTC mark and with an APE of 5.65% and CV of 7.73% for counts distal to the birth band. Among readers, 93% (27 of 29) of the fi nal band-pair estimates distal to the OTC were within 1 band pair of each other, and 86% (25 of 29) of the estimates distal to the birth band were within 1 band pair.
The readings distal to the OTC that differed by more than one band pair were from sharks that had been at liberty for the longest time, in-dicating that variability in band-pair counts in-creases with age, likely because of disagreement among readers caused by structural artifacts. For example, sample A037789 was from a shark that was at liberty for 1594 days and band-pair estimates distal to the OTC varied from 6 to 9 among the 3 readers for this shark (Table 1).
Length-frequency analysis
A total of 14,720 individual Shortfi n Mako were used for the length-frequency analysis that was completed with fi shery-dependent data from the California drift gillnet fi shery (1981–2009) and fi shery-independent data from juvenile Shortfi n Mako surveys (1993–2009) (Fig. 5). No differ-ences in length-frequency modes were detected when analyzed by season or sex; therefore, pa-rameter estimates were generated by grouping across factors (season and sex) with age-depen-dent standard deviation incorporated into the model. Average size of Shortfi n Mako used in growth modeling was 121.3 cm FL (±0.25 SE). The majority of fi sh (85%) ranged in size from 80 to 160 cm FL with 3 identifi able modes con-sistently present in annual length frequencies regardless of survey type or sex.
Results from MULTIFAN showed these 3 modes at 86, 112, and 134 cm FL, and average modal sizes from MIXDIST were 83, 118, and 147 cm FL. Tak-ing the difference between modal sizes from MULTI-FAN analysis provided average annual growth rates of 27 and 23 cm FL for the period from the fi rst to sec-ond mode and for the period from the second to third modes, respectively (Table 2). Similarly, annual growth rates from MIXDIST averaged 36 and 29 cm FL for the same 2 periods (Table 2).
Growth of tagged and recaptured sharks
No difference was observed between growth rates cal-culated from at-liberty OTC-injected sharks (28 and 21 cm per year at 85 and 130 cm FL, respectively) and tag-recaptured sharks not injected (29 and 19 cm per year at 85 and 130 cm FL, respectively); there-fore, these data were pooled. The growth rates that resulted from GROTAG analysis of lengths of tagged and recaptured sharks were similar to the growth rates from length-frequency calculations, averaging 29
and 20 cm per year at 85 and 130 cm FL, respectively (Table 2).
Discussion
Results of this study indicate that 2 band pairs are deposited each year in juvenile Shortfi n Mako tagged and released in southern California. The fast growth rates collectively obtained through the use of OTC-marked vertebrae, MULTIFAN and MIXDIST length-frequency analyses, and tag-recapture growth models were consistent, providing strength to our results be-yond the usefulness possible with the employment of a single method alone. Further, a sample size of 29 OTC-marked vertebrae of Shortfi n Mako at liberty from 4 months to more than 4 years is the most com-prehensive OTC tag-return data set reported for this species. We also compared our total band-pair counts at length with those of Cailliet et al. (1983), the only other Shortfi n Mako age and growth study completed

154 Fishery Bulletin 111(2)
Figure 3X-ray images of vertebrae sections that show band-pair progression of 5 OTC-marked juvenile Shortfin Mako (Isurus oxy-rinchus) recaptured during this study off southern California. The small inset in each image shows the OTC fluorescence under UV light. Translucent or hyaline (rapid growth) bands or darker areas of vertebrae show periods of rapid growth, and opaque bands or lighter areas of vertebrae show periods of slow growth. Arrows indicate band-pair counts.

Wells et al.: Age validation of juvenile Isurus oxyrinchus tagged off southern California 155
Figure 4Age-bias plots for all OTC-marked juvenile Shortfin Mako (Isurus oxyrinchus) independently aged by 3 readers during this study. Graphs in the left column represent total band-pair counts distal to the OTC mark, and graphs in the right column represent total band-pair counts distal to the birth band.
Counts after the OTC mark Counts after the birth mark
in this region, to determine if vertebral readings or as-sumptions about growth-band deposition differed. Total band-pair counts relative to fi sh size in this study were very similar to the average band-pair counts, relative to fi sh size, obtained by Cailliet et al. (1983) (Fig. 6).
This fi nding indicates that vertebral readings were similar between the 2 studies and only the assumption about deposition rates differed.
Given differences in the rates of band-pair deposi-tion of Shortfi n Mako in other studies, our fi ndings
Reader 2 Reader 2
Reader 3 Reader 3
Reader 3 Reader 3
R
ead
er 2
R
ead
er 1
R
ead
er 1
R
ead
er 2
R
ead
er 1
R
ead
er 1

156 Fishery Bulletin 111(2)
should not be extrapolated beyond our size range and geographic area; however, our results indicate that growth patterns of juvenile and adult Shortfi n Mako merit additional research.
The differences in rates of band-pair deposition of Shortfi n Mako among studies may be due to a number of factors, including study location, methods, and on-togeny. Rates of band-pair deposition of the Common Carp (Cyprinus carpio) vary geographically among sim-
ilar age classes of the same species; biannual deposition occurs in South Africa (Winker et al., 2010) and an-nual deposition occurs in Australia (Brown et al., 2004), exemplifying the importance of regional age validation.
In one of the fi rst Shortfi n Mako aging studies, Pratt and Casey (1983) read silver-nitrate-stained, whole ver-tebral centra collected in the north-west Atlantic and suggested that 2 band pairs were deposited annually, after noting that calculated growth under this assumption closely matched direct observations of growth over time for this species (especially for fi sh ≤200 cm FL). Pratt and Casey’s (1983) growth curves were generated from tag-recapture growth data col-lected in the fi eld, temporal analysis of length-month information, and a vertebral age-based growth curve cor-roborated and compared with length-frequency mode analyses.
In contrast, Cailliet and Bedford (1983) and Cailliet et al. (1983) used hard (high-frequency) X-radiography and silver nitrate staining of whole centra to elucidate banding patterns of Shortfi n Mako in the northeast Pacifi c. The authors of both studies assumed that only 1 band pair was
deposited annually and obtained a growth rate of about half the rate of that of Pratt and Casey (1983), but they did not test the assumption of 1 versus 2 band pairs per year.
Campana et al. (2002) concluded that the hypothesis of a single band pair was the most consistent with the banding pattern obtained from a 21-year-old Shortfi n Mako (328 cm FL) collected from the northeast Atlantic and aged with bomb-radiocarbon techniques. Ardizzone
Table 2Estimates from the tag-recapture growth model with GROTAG for juvenile Shortfi n Mako (Isurus oxyrinchus) in the eastern Pacifi c (n=62). Growth estimates from the length-frequency techniques (MULTIFAN and MIXDIST) also are included for the fi rst 2 age classes. K=Brody growth parameter, L∞=mean asymptotic length.
Tag-recapture Length frequency Length frequencyParameter Symbol (unit) growth model (MULTIFAN) (MIXDIST)
Growth rate g85 (cm/year) 29 27 36 g130 (cm/year) 20 23 29K (per year) 0.19 0.17 L∞ (cm) 231.03 252.12 Growth variability ν 0.31 Mean measurement error m (cm) 2.87 Standard deviation measurement error s 4.46 Log likelihood λ –235.8
Figure 5Total percent length-frequency distribution of Shortfin Mako (Isurus oxy-rinchus) collected from fishery-dependent (1981–2009) and fishery-inde-pendent (1993–2009) surveys conducted in the Southern California Bight. Vertical bars are placed in length bins of 5 cm along the x-axis, showing the range of fork lengths (FL) from 55 to 265 cm FL. Arrows show the approximate size at 50% maturity for males and females determined by Semba et al. (2011).

Wells et al.: Age validation of juvenile Isurus oxyrinchus tagged off southern California 157
Figure 6Individual band-pair counts relative to size, shown in fork length (cm), of juvenile Shortfin Mako (Isurus oxyrinchus) from this study (n=29) com-pared with the average number of band-pair counts relative to size by Cailliet et al. (1983). Vertebral readings were similar between these 2 studies, and only the assumption about deposition rates differed. Sharks for both studies were captured off southern California.
et al. (2006) expanded on this study with 54 samples used for radiocarbon chronologies. Their results supported the hypothesis of annual band-pair deposition for older ages, but they did not rule out biannual band-pair depo-sition for young fi sh. Similarly, Natan-son et al. (2006) reiterated the afore-mentioned results and presented evi-dence of annual band-pair formation from an OTC-injected, 241-cm-FL, male Shortfi n Mako at liberty for just over 1 year. However, growth curves generated from their tag-recapture and length-frequency data showed a much faster growth rate for young Shortfi n Mako than the rate calcu-lated from vertebrae data, and the authors suggested the possibility that rates of band deposition may change with ontogeny.
Bishop et al.1 and Bishop et al. (2006) examined age and growth of Shortfi n Mako from New Zealand wa-ters and assumed a deposition rate of 1 band pair per year on the basis of Campana et al. (2002) and Ribot-Carballal et al. (2005). They obtained a growth curve similar to that of Cail-liet et al. (1983), and their best-fi t Schnute growth model predicted that growth was fast for the fi rst few years of life (39 cm during year 1), then slowed for older ages to a rate that is similar to juvenile growth rates found in our study. Like Pratt and Casey (1983), they found that length-frequency data indicated considerably faster growth than did estimates from vertebral ages for the younger age classes with the assumption of 1 band pair per year. Through the use of cohort analysis, Maia et al. (2007) also reported fast growth in juve-nile Shortfi n Mako collected from the longline fi shery in the eastern North Atlantic (average growth rate of 61.1 cm/year for the fi rst year, and 40.6 cm/year for the second year).
More recently, Okamura and Semba (2009) and Semba et al. (2009) examined monthly centrum edge patterns with a new statistical method and surface shadow technique to determine periodicity of band-pair formation. Their results indicated that the deposition of band pairs in Shortfi n Mako in the western and cen-tral North Pacifi c has an annual cycle, but they also warned about sources of error from inaccuracies in cen-trum margin readings and from a large variability in the timing and duration of band deposition.
Biannual band-pair deposition is not species-specifi c to Shortfi n Mako; it reportedly has occurred in other shark species as well. Chen et al. (1990) and Anislado-Tolentino et al. (2008) suggested biannual band-pair deposition in the Scalloped Hammerhead (Sphyrna
lewini). In addition, Parker and Stott (1965) suggest-ed that 2 band pairs were laid down annually in the Basking Shark (Cetorhinus maximus); however, those conclusions were questioned by Pauly4 (2002). Further, Natanson et al. (2008) examined vertebral growth pat-terns in C. maximus as a function of ontogeny and ques-tioned the feasibility of aging this species with vertebral band-pair counts.
Ontogenetic differences may play a critical role in band-pair formation for Shortfi n Mako, as well. Unfor-tunately, none of our OTC-marked vertebrae were from sharks >200 cm FL at tagging or recapture, but the 3 largest sharks at recapture (172, 190, and 200 cm FL) exhibited a pattern consistent with biannual deposition. Two of these sharks (200 and 190 cm FL) were at liberty for more than 4 years and averaged 7.3 and 8.7 band pairs distal to the OTC mark, respectively, indicating that the biannual pattern continues in fi sh of this size. Both had similar total band-pair counts, averaging 9.3 and 11.7, respectively.
If one assumes a biannual pattern, as indicated by our OTC marking results, these fi sh would range from
4 Pauly, D. 1978. A critique of some literature data on the growth, reproduction and mortality of the lamnid shark, Ce-torhinus maximus (Gunnerus). ICES Coucil Meeting Doc. 1978/H:17, 10 p.

158 Fishery Bulletin 111(2)
4.5 to 6 years of age when collected. Examination of vertebral centra from some of our larger, non-OTC-tagged, recaptured Shortfi n Mako reveals that as fi sh approach maturity, banding patterns appear to become more distinct, with both fast and slow growth zones becoming more regular and evenly spaced. Therefore, it is possible that these larger fi sh, on entering a more oceanic realm, change the periodicity of their band-ing pattern. On the basis of these results, it appears that Shortfi n Mako ≤200 cm FL and found off Cali-fornia grow much faster than previously thought, with observed rapid growth (biannual band deposition) for approximately the fi rst 6 years of life. Slower growth (annual deposition) thereafter may occur but cannot be confi rmed by this study.
Mechanisms that drive the observed biannual band-pair deposition may be linked to seasonal migration patterns and subsequent food availability. Through the use of similar OTC tag-recapture methods, Murphy et al. (1998) determined that Black Drum (Pogonias cromis) begin to deposit biannual band pairs after 4 years of age because of a shift in migration patterns. Similarly, Shortfi n Mako tagged in southern California appear to exhibit a biannual growth cycle, with fast-growth periods (wide, translucent band formation) in summer and winter and slow-growth periods (narrow, opaque band formation) in spring and fall. One possible explanation is that, as coastal waters warm and cool seasonally, juvenile Shortfi n Mako move between rich feeding grounds off California in the summer and off Mexico in the winter and spend spring and fall migrat-ing between these feeding areas.
Of the 15 OTC-marked sharks recaptured in Baja Mexico waters (Fig. 1B), only 2 sharks were recaptured during summer months. In contrast, 70% of the returns of OTC-marked sharks in California coastal waters oc-curred during summer and there were no returns in the winter. In addition, on the basis of data from all tag recaptures (OTC and non-OTC-marked sharks, n=317, senior author, unpubl. data), 40% of all shark recaptures in Mexico (south of 30°N) occurred dur-ing winter, but only 7% of sharks were recaptured in Mexico during summer. Likewise, 58% and 7% of all recaptures in California coastal waters occurred during summer and winter seasons, respectively. Preliminary analysis of movements of juvenile (<150 cm FL) Short-fi n Mako tracked with satellite tags (2003–10, n=26) also shows high relative densities of Shortfi n Mako at reported locations off Baja California, Mexico, in the winter and off California in the summer and more dis-persive offshore movements in the spring and fall (S. Kohin, unpubl. data).
Tag-recapture techniques with OTC are among the most powerful methods for validation of age and growth patterns in fi shes, but there are diffi culties with this method. One disadvantage in the use of chemical marking and recaptures is that the number of band pairs formed for short times between tagging and recapture is often low, resulting in a potentially large
relative error if one of the band pairs (such as on the growing edge) is misinterpreted (Campana, 2001). For example, misinterpretation of a single growth zone in a fi sh at liberty for 2 years would result in a 50% error, but the same misinterpretation in a fi sh at liberty for 10 years would produce an error of only 10%. Fortunately, the relatively large sample size of 15 sharks at liberty for more than 1 year and 4 sharks at liberty for more than 3 years (2 of them for more than 4 years) allowed us to confi rm the consistent banding patterns between both short- and long-term deployments.
An additional problem with the use of chemical marking techniques, such as with OTC, is that growth may be inhibited (Pfi zer,5 1975; Monaghan, 1993); how-ever, other researchers that have worked with elasmo-branchs have shown OTC to have little adverse effects on growth (Tanaka [1990] for the Japanese Wobbegong [Orectolobus japonicus] and Gelsleichter et al. [1998] for the Nurse Shark [Ginglymostoma cirratum]). Further, this problem does not appear relevant in this study be-cause fast (i.e., not inhibited) growth was observed with our OTC-marked juveniles.
Conclusions
Direct and indirect methods used in this study indi-cate rapid growth of juvenile Shortfi n Mako in south-ern California and the OTC-marked vertebrae support a pattern of biannual deposition for the fi rst 5 years of life. Accurate age and growth parameters in stock assessment models are important for fi sheries man-agement and are necessary for calculations of growth and mortality rates, age at recruitment, and longevity. This article highlights the need to continue life history studies of elasmobranch species throughout different regions and ocean basins.
Acknowledgments
We thank personnel of the Southwest Fisheries Sci-ence Center for assistance in tagging and logistical operations to make this study possible: specifi cally, D. Holts, R. Rasmussen, R. Vetter, and J. Wraith. We also thank NOAA Fisheries observers for their diligent work in collecting specimens and length data and to the many anglers and commercial operators for return-ing OTC-marked vertebrae. We especially thank O. So-sa-Nishizaki for help with collection and shipment of samples from Mexico, G. Morse for sharing microscope and imaging equipment, and both A. Andrews and M. Francis for valuable suggestions and comments on ear-lier drafts.
5 Pfi zer, Inc. 1975. Oxytetracycline intramuscular solution leafl et 60-1051-00-8, 2 p. Pfi zer Lab. Div., New York.

Wells et al.: Age validation of juvenile Isurus oxyrinchus tagged off southern California 159
Literature cited
Anislado-Tolentino, V., M. Gallardo-Cabello, F. Amezcua-Lin-ares, and C. Robinson-Mendoza.
2008. Age and growth of the scalloped hammerhead shark, Sphyrna lewini (Griffi th & Smith, 1834) from the Southern coast of Sinaloa, México. Hidrobiológica 18:31–40.
Ardizzone, D., G. M. Cailliet, L. J. Natanson, A. H. Andrews, L. A. Kerr, and T. A. Brown.
2006. Application of bomb radiocarbon chronologies to shortfi n mako (Isurus oxyrinchus) age validation. En-viron. Biol. Fishes 77:355–366.
Beamish, R. J., and D. A. Fournier. 1981. A method for comparing the precision of a set
of age determinations. Can. J. Fish. Aquat. Sci. 38:982–983.
Beamish, R. J., and G. A. McFarlane. 2000. Reevaluation of the interpretation of annuli from
otoliths of a long-lived fi sh, Anoplopoma fi mbria. Fish. Res. 46:105–111.
Bishop, S. D. H., M. P. Francis, C. Duffy, and J. C. Montgomery. 2006. Age, growth, maturity, longevity and natural mor-
tality of the shortfi n mako shark (Isurus oxyrinchus) in New Zealand waters. Mar. Freshw. Res. 57:143–154.
Bowker, A. H. 1948. A test for symmetry in contingency tables. J. Am.
Stat. Assoc. 43:572–574.Branstetter, S., and J. A. Musick.
1994. Age and growth estimates for the sand tiger in the northwestern Atlantic Ocean. Trans. Am. Fish. Soc. 123:242–254.
Brown, P., C. Green, K. P. Sivakumaran, D. Stoessel, and A. Giles.
2004. Validating otolith annuli for annual age de-termination of common carp. Trans. Am. Fish. Soc. 133:133–196.
Cailliet, G. M., and D. S. Bedford. 1983. The biology of three pelagic sharks from Cali-
fornia waters, and their emerging fi sheries: a review. CalCOFI Rep. 24:57–69.
Cailliet, G. M., L. K. Martin, J. T. Harvey, D. Kusher, and B. A. Welden.
1983. Preliminary studies on the age and growth of blue, Prionace glauca, common thresher, Alopias vulpi-nus, and shortfi n mako, Isurus oxyrinchus, sharks from California waters. In Proceedings of the international workshop on age determination of oceanic pelagic fi sh-es: tunas, billfi shes, and sharks; 15–18 February 1982 (E. D. Prince and L. M. Pulos, eds.), p. 179–188. NOAA Tech. Rep. NMFS 8.
Campana, S. E. 2001. Accuracy, precision and quality control in age de-
termination, including a review of the use and abuse of age validation methods. J. Fish Biol. 59:197–242.
Campana, S. E., L. J. Natanson, and S. Myklevoll. 2002. Bomb dating and age determination of the large
pelagic sharks. Can. J. Fish. Aquat. Sci. 59:450–455. Casey, J. G., and N. E. Kohler.
1992. Tagging studies on the shortfi n mako shark (Isu-rus oxyrinchus) in the western North Atlantic. Aust. J. Mar. Freshw. Res. 43:45–60.
Cerna, F., and R. Licandeo. 2009. Age and growth of the shortfi n mako (Isurus oxy-
rinchus) in the south-eastern Pacifi c off Chile. Mar. Freshw. Res. 60:394–403.
Chang, W. Y. B. 1982. A statistical method for evaluating the reproduc-
ibility of age determination. Can. J. Fish. Aquat. Sci. 39:1208–1210.
Chen, C. T., T. C. Leu, S. J. Joung, and N. C. H. Lo. 1990. Age and growth of the scalloped hammerhead,
Sphyrna lewini, in northeastern Taiwan waters. Pac. Sci. 44:156–170.
Claisse, J. T., M. Kienzle, M. E. Bushnell, D. J. Schafer, and J. D. Parrish.
2009. Habitat- and sex-specifi c life history patterns of yellow tang Zebrasoma fl avescens in Hawaii, USA. Mar. Ecol. Prog. Ser. 389:245–255.
Compagno, L. J. V. 2001. Sharks of the world. An annotated and illustrated
catalogue of shark species known to date. Vol. 2: Bull-head, mackerel and carpet sharks (Heterodontiformes, Lamniformes and Orectolobiformes). FAO Species Cat-alogue for Fishery Purposes 1, 269 p. FAO, Rome.
Cortés, E. 2002. Incorporating uncertainty into demographic mod-
eling: application to shark populations and their conser-vation. Conserv. Biol. 16:1048–1062.
Duffy, C., and M. P. Francis. 2001. Evidence for summer parturition in shortfi n mako
(Isurus oxyrinchus) sharks from New Zealand waters. Z. J. Mar. Freshw. Res. 35:319–324.
Fournier, D. A., J. R. Sibert, J. Majkowski, and J. Hampton. 1990. MULTIFAN: a likelihood-based method for esti-
mating growth parameters and age composition from multiple length frequency data sets illustrated us-ing data for southern bluefi n tuna (Thunnus mac-coyii). Can. J. Fish. Aquat. Sci. 47:301–317.
Francis, R. I. C. C. 1988a. Are growth parameters estimated from tagging
and age-at-length data comparable? Can. J. Fish. Aquat. Sci. 45:936–942.
1988b. Maximum likelihood estimation of growth and growth variability from tagging data. N. Z. J. Mar. Freshw. Res 22:43–51.
García, V. B., L. O. Lucifora, and R. A. Myers. 2008. The importance of habitat and life history to ex-
tinction risk in sharks, skates, rays and chimeras. Proc. R. Soc. Lond., Ser. B: Biol. Sci. 275:83–89.
Gelsleichter, J., E. Cortés, C. A. Manire, R. E. Heuter, and J. A. Musick.
1998. Evaluation of oxytetracycline on growth of captive nurse sharks, Ginglymostoma cirratum. Fish. Bull. 96:624–627.
Goldman, K. J. 2005. Age and growth of elasmobranch fi shes. In Man-
agement techniques for elasmobranch fi sheries (J. A. Musick and R. Bonfi l, eds.), p. 76–102. FAO Fish. Tech. Pap. 474. FAO, Rome.
Hoenig, J. M., J. M. Morgan, and C. A. Brown. 1995. Analyzing differences between two age determi-
nation methods by tests of symmetry. Can. J. Fish. Aquat. Sci. 52:364–368.

160 Fishery Bulletin 111(2)
Holts, D. B., and D. W. Bedford. 1993. Horizontal and vertical movements of the shortfi n
mako shark, Isurus oxyrinchus, in the southern Califor-nia bight. Aust. J. Mar. Freshw. Res. 44:901–909.
Joung, S. J., and H. H Hsu.2005. Reproduction and embryonic development of the
shortfi n mako, Isurus oxyrinchus Rafi nesque, 1810, in the northwestern Pacifi c. Zool. Stud. 44:487–496.
Kusher, D. I., S. E. Smith, and G. M. Cailliet. 1992. Validated age and growth of the leopard shark,
Triakis semifasciata, with comments on reproduc-tion. Environ. Biol. Fishes 35:187–203.
MacDonald, P. D. M., and T. J. Pitcher. 1979. Age-groups from size-frequency data: a versatile
effi cient method of analyzing distribution mixtures. J. Fish. Res. Board Can. 36:987–1001.
Maia, A., N. Queiroz, H. N. Cabral, A. M. Santos, and J. P. Correia.
2007. Reproductive biology and population dynamics of the shortfi n mako, Isurus oxyrinchus Rafi nesque, 1810, off the southwest Portuguese coast, eastern North At-lantic. J. Appl. Ichthyol. 23:246–251.
Mollet, H. F., G. Cliff, H. L. Pratt, and J. D. Stevens. 2000. Reproductive biology of the female shortfi n mako,
Isurus oxyrinchus Rafi nesque, 1810, with comments on the embryonic development of lamnoids. Fish. Bull. 98:299–318.
Monaghan, J. P. 1993. Comparison of calcein and tetracycline as chemi-
cal markers in summer fl ounder. Trans. Am. Fish. Soc. 122:298–301.
Murphy, M. D., D. H. Adams, D. M. Tremain, and B. L. Winner. 1998. Direct validation of ages determined for adult
black drum, Pogonias cromis, in east-central Flori-da, with notes on black drum migration. Fish. Bull. 96:382–387.
Natanson, L. J., N. E. Kohler, D. Ardizzone, G. M. Cailliet, S. P. Wintner, and H. F. Mollet.
2006. Validated age and growth estimates for the short-fi n mako, Isurus oxyrinchus, in the North Atlantic Ocean. Environ. Biol. Fishes 77:367–383.
Natanson, L. J., S. P. Wintner, F. Johansson, A. Piercy, P. Campbell, A. D. Maddalena, S. J. B. Gulak, B. Human, F. C. Fulgosi, D. A. Ebert, F. Hemida, F. H. Mollen, S. Vanni, G. H. Burgess, L. J. V. Compagno, and A. Wedderburn-Maxwell.
2008. Ontogenetic vertebral growth patterns in the basking shark Cetorhinus maximus. Mar. Ecol. Prog. Ser. 361:267–278.
Okamura, H., and Y. Semba. 2009. A novel statistical method for validating the peri-
odicity of vertebral growth band formation in elasmo-branch fi shes. Can. J. Fish. Aquat. Sci. 66:771–780.
PFMC (Pacifi c Fishery Management Council). 2011. Fishery management plan for U.S. West Coast
fisheries for highly migratory species. As amended through Amendment 2, 106 p. Pacifi c Fishery Man-agement Council, Portland, OR. [Available from http://www.pcouncil.org/highly-migratory-species/fi shery-management-plan-and-amendments/.]
Parker, H. W., and F. C. Stott. 1965. Age, size and vertebral calcifi cation in the basking
shark, Cetorhinus maximus (Gunnerus). Zool. Meded. 40:305–319.
Pauly, D.2002. Growth and mortality of the basking shark Ceto-
rhinus maximus and their implications for management of whale sharks Rhincodon typus. In Elasmobranch biodiversity, conservation and management. Proceed-ings of the international seminar and workshop; Sabah, Malaysia, July 1997 (S. L. Fowler, T. Reid, and F. A. Dip-per, eds.), p. 199–208. Occasional Papers of the IUCN Survival Commission, no. 25. Int. Union Conserv. Na-ture, Gland, Switzerland.
Pratt, H. L., Jr., and J. G. Casey. 1983. Age and growth of shortfi n mako, Isurus oxyrin-
chus, using four methods. Can. J. Fish. Aquat. Sci. 40:1944–1957.
R Development Core Team.2008. R: a language and environment for statistical
computing. R Foundation for Statistical Computing. [Available from: http://R-project.org, accessed November 2008.]
Ribot-Carballal, M. C., F. Galván-Magana, and C. Quinonez-Velazquez.
2005. Age and growth of the shortfi n mako shark, Isurus oxyrinchus, from the western coast of Baja California Sur, Mexico. Fish. Res. 75:14–21.
Semba, Y., H. Nakano, and I. Aoki. 2009. Age and growth analysis of the shortfi n mako
shark, Isurus oxyrinchus, in the western and central North Pacifi c Ocean. Environ. Biol. Fishes 84:377–391.
Semba, Y. I. Aoki, and K. Yokawa. 2011. Size at maturity and reproductive traits of short-
fi n mako, Isurus oxyrinchus, in the western and central North Pacifi c. Mar. Freshw. Res. 62:20–29.
Smith, S. E., D. W. Au, and C. Show. 1998. Intrinsic rebound potentials of 26 species of Pacifi c
sharks. Mar. Freshw. Res. 49:663–678. Smith, S. E., R. A. Mitchell, and D. Fuller.
2003. Age-validation of a leopard shark recaptured after 20 years. Fish. Bull. 101:194–198.
Stevens, J. D. 2008. The biology and ecology of the shortfi n mako
shark, Isurus oxyrinchus. In Sharks of the open ocean: biology, fi sheries and conservation (M. D. Camhi, E. K. Pikitch, and E. A. Babcock, eds), p. 87–94. Blackwell Publ., Ltd., Oxford.
Tanaka, S. 1990. Age and growth studies on the calcifi ed struc-
tures of newborn sharks in the laboratory aquaria us-ing tetracycline. In Elasmobranchs as living resources: advances in the biology, ecology, systematics, and the status of the fi sheries (H. L. Pratt Jr., S. H. Gruber, and T. Taniuchi, eds.), p. 189–202. NOAA Tech. Rep. NMFS 90.
Taylor, V. B., and D. W. Bedford. 2001. Shortfi n mako shark. In California living marine
resources: a status report (W. S. Leet, C. M. DeWees, R. Klingbeil, and E. J. Larson, eds), p. 336–337. Publica-tion SG01-11, California Dep. Fish Game, Sacramento, CA.
Winker, H., O. L. F. Weyl, A. J. Booth, and B. R. Ellender. 2010. Validating and corroborating the deposition of two
annual growth zones in asteriscus otoliths of common carp Cyprinus carpio from South Africa’s largest im-poundment. J. Fish Biol. 77:2210–2228.