alien animals in hawai ifs native ecosystems: … animals in hawai!ifs native ecosystems: toward...
TRANSCRIPT

ALIEN ANIMALS INHAWAI!IfS NATIVE ECOSYSTEMS:
TOWARD CONTROLLING THE ADVERSE EFFECTSOF INTRODUCED VERTEBRATES
Charles P. Stone
ABSTRACT
The adverse effects of introduced birds and mam-mals on native taxa and ecosystems in Hawaifi have beenlong term, widespread, and severe. Impacts began atleast 1,500 years ago with colonization by the Poly-nesians and their flora and fauna, and continued withtheir increasingly severe disturbance to the landscape,especially below 500 m elevation. Problems acceleratedwith the arrival of continental man in 1778, and con-tinue to the present day with suspected deliberate re-leases of birds that threaten native species as re-cently as 1982. Alien vertebrates can affect nativebiota through predation, competition, depredation, andhabitat degradation. Negative impacts can be subtle ordramatic, but evidence of importance is manifested inlarge percentages of extinct and rare taxa. In thispaper, adverse effects of major bird and mammal intro-ductions are outlined where possible for islands, vege-tation zones, and rare taxa. Although much remains tobe learned, suggestions for reducing negative effectsof alien vertebrates can be made. These include: En-forcement of efficient quarantine procedures; suffi-cient support for enduring and complete vertebrate dam-age control programs (including research, management,and monitoring) on lands managed for preservation ofnative Hawaiian ecosystems and taxa; development ofmultiple and adaptable methods of vertebrate damagereduction; preservation and management of the most in-tact areas remaining in Hawai'i; and cooperation andcommunication among the agencies and special interestgroups (including developers and conservationists) inland use planning on regional bases. Cooperative ap-proaches, effective education and communication aboutthe value of protected areas, and the increased useful-ness of such areas for all citizens, are seen as par-ticularly important in reducing continued damage toHawai'ifs remaining native ecosystems by alien verte-brates.
251

INTRODUCTION
Adverse effects of introduced or alien vertebrateson native Hawaiian ecosystems and organisms may be di-vided into 2 categories: those that occurred largely inthe past (including the introductions of the earlyPolynesians), and those that occur at present. Thefirst are of interest because they provide examples ofthe devastating effects that aliens can cause over timeto native species with small population sizes and in-adequate defenses against intruders. Such early dis-ruptions, when viewed in accurately constructed andcomprehensive historical contexts, can provide insightsinto compositions and functions of the Hawaiian biotaat a time when it was more intact. The effects of pastintroductions also provide sufficient warning to pro-voke caution about future introductions or transloca-tions. The adverse impacts of some of these introduc-tions are unknown, but the effects of others are clear-ly visible today.
Ongoing and active disturbances by alien verte-brates are of interest for 2 additional reasons.First, they allow direct observation and manipulationthrough which one can obtain understandings of the eco-logy of aliens and interrelationships within ecosys-tems. Secondly, through reduction, exclusion, or elim-ination of problem animals, entire ecosystems (or im-portant components thereof) can be rehabilitated tosome degree (see Lamoureux, this volume). Where eco-system deterioration caused by introduced animals canbe stopped or slowed, systems may at least functionwith primarily native components; and natural processes(including nutrient cycling, gene flow, succession, andevolution), although not pristine, may also be less in-fluenced by man and his induced changes.
Damage caused by alien vertebrates to native floraand fauna in Hawai'i may be categorized as predation,competition, depredation, or habitat degradation.Predation is the killing of one animal by another, andincludes such important examples as the eating of eggsand young of birds by mammals, and consumption of in-sects and snails by mammals or birds. Competition isthe negative influence of one animal on another when aresource needed by both is somehow limited. Relation-ships among native and exotic birds and a common foodsupply, and relationships among native birds, intro-duced rats and common food or nest sites, are exam-ples. Depredation is the eating or otherwise destroy-ing of plants by animals. Consumption, trampling, oruprooting of native plants (or plant parts) by alienmammals are examples. Depredations on plants influenceother animals through impacts on their habitats, andthese will be discussed under "Habitat Degradation."
252

Introduced vertebrates can have additional, subtleeffects on native ecosystems and biota. Disease can betransmitted to native species through alien vertebratereservoirs (see van Riper and van Riper, this volume).Introduced animals can disperse alien plants and thusincrease plant densities, rates of spread, and distri-butional limits. Vertebrates sometimes also enhancegermination of introduced plants through seed scarifi-cation in digestive tracts, or through digging up and/or fertilization with feces of potential seedbeds.Serious long-term damage can be caused by some alienvertebrates that disrupt nutrient cycling, initiate andaccelerate erosion, radically change compositions ofplant and animal communities, and alter evolution ofother species through disruption of natural selection.
The purpose of this paper is to address some ofthe complexities of damage caused to native terrestrialecosystems by birds and mammals introduced to Hawai'iby man. Introduced reptiles and amphibians also affectnative species, particularly invertebrates (Howarth,this volume), but there is little information availableabout what are probably less severe impacts, at leastcurrently. A brief summary of adverse effects prior tothe arrival of European man will be presented, and thedistribution of different kinds of impacts will be out-lined by island and by general vegetation type. Man-agement strategies for different animals and situationswill be discussed and research needs identified. Acoordinated approach to the management and research ofintroduced mammals and birds in Hawai'i's threatenedecosystems will be suggested.
POLYNESIAN IMPACTS (400 A.D. — 1778 A.D.)
Beginning at least in 400 A.D., human colonizersof the Hawaiian Islands began arriving with their at-tendant land use practices, flora, and fauna. Verte-brate introductions included domestic pigs (Sus scro-fa), red junglefowl (Gallus gallus), dogs (Canis famil-iaris), Polynesian or Pacific rats (Rattus exulans),and various stowaway reptiles (Kirch 1982). Althoughdocumentation is lacking, rats probably began to affectlowland invertebrates and plants shortly after humanarrival; predation on low- or ground-nesting forestbirds and seabirds would not be unexpected based onsimilar activity in historical time (Kepler 1967; At-kinson 1977; Imber 1978). Junglefowl, rats, and theother small vertebrates spread into lowland forests atrates dependent mostly upon genetic and behavioraladaptation to changing Hawaiian environments. Highreproductive rates, low predation, and shortage of com-petition may have favored these small vertebrates insuitable habitat, especially that modified by man.However, on the whole, the continuing land modification
253

fllfll HiniiW jlft have overwhelmed
fufci s* thai? iBttflduoofl inlialQ on ttiQ biota (Kirch1982) .
According to one estimate, about 80 percent of thelowland forest (below 500 m) was drastically altered bywidespread use of fire, by agricultural and aquacul-tural development, and by forest clearing (Kirch1982). Evidence of agriculture has been found as highas 1200 m in elevation (McEldowney 1983), and slash andburn agriculture was perhaps used to maintain piligrasslands (Kirch 1982) for thatching huts; Sadleriaspp. and arrowroot (Tacca leontopetaloides) were usedas pig fodder and famine food (McEldowney 1983). Des-criptions of the early landscape were given by Europeanexplorers such as Cook and King, Vancouver and Menzies,and Chamisso (Olson and James 1982), and the impacts ofHawaiians on native vegetation are usually inferredfrom this.
A human population of perhaps 200,000-250,000 atthe time of European contact (Schmitt 1971) may repre-sent a decline resulting from reduced carrying capacity(Kirch 1982). The effects of such large numbers ofpeople and their alterations of the native vegetationwere undoubtedly large. Impacts on avian species alonewere enormous, with 39 [now 45] of Hawai'i's 80 [now86] known species of birds eliminated prior to the ar-rival of Western man in 1778 (Olson and James 1982).Most of the 13-15 flightless species of birds wereprobably hunted to varying degrees and eventually ex-tirpated. Development of irrigated fields may haveresulted in larger populations of the endemic Hawaiianduck (Anas wyvilliana), common moorhen (Gallinulachloropus), American coot (Fulica americana) and black-necked stilt (Himantopus mexicanus) as residents.Establishment of the short-eared owl (Asio flammeus)may have been aided by manfs introduction of the Poly-nesian rat (Olson and James 1982).
Dogs and pigs were valued by Hawaiians as scaven-gers and as sources of animal protein (Handy and Handy1972; Kirch 1979), and were probably usually kept undercontrol. Junglefowl and their eggs may have been lessfavored for food (Handy and Handy 1972) than some othersources, and may therefore have been less controlled;they formerly ranged from sea level to 2,100 m onHawai'i (Schwartz and Schwartz 1949). Junglefowl alsomay have arrived later than other domestics (Tuggle1979), but this is uncertain. Handy and Handy (1972)noted that pigs were allowed to "run about the kauhale(homestead) and gardens while they were young, but whenthey were sizable and ready for fattening they werepenned inside enclosures of heaped-up stones." Whetherthis was always the case is unknown. Recent evidence
254

suggests that pigs sometimes were separated from gar-dens and irrigated fields by keeping them within vil-lage walls (D.B. Barrere, pers. comm.). Pigs probablyexisted throughout most of the islands by the time ofEuropean contact (McEldowney 1979), and feral popula-tions in "moist" forests could have been "an olderutilized resource" (McEldowney 1983). Whether theywere important invaders of Hawaiian rain forests isunknown. As on other Pacific islands such as New Gui-nea, some pigs may have been tightly managed and herdedwith dogs; some may have been caught at times fromnearby forests and fallow fields; and some may havebeen semi-wild or pariah (Diong 1983) populations, fedand even called in by people as needed (H. McEldowney,pers. comm.).
The reportedly small size of Polynesian pigs mayhave been less a function of genetics than a diet lowin protein and the rapid use of younger pigs for food.P.V. Kirch (pers. comm.) noted that most pigs recoveredfrom archaeological sites are young and that pigs werefed coconut, sweet potato, and breadfruit. Pigs alsoate human wastes and thus served as a means of villagesanitation. Handy and Handy (1972) suggested thatferal pigs in upland areas subsisted on fruits, nuts,seeds, and various ferns, and that they "grubbed forroots." Considerable acreages of fallow land and sec-ondary growth (McEldowney 1983) may have served to at-tract foraging feral pigs more than dense rain forest.
Animal protein is required for maximum growth andreproduction of pigs (Pond and Houpt 1978; NationalResearch Council 1979), and other than turtle eggs,fish, and beached marine mammals from a sometimes cap-ricious ocean, there were few protein sources in na-tive, as opposed to man-influenced, ecosystems. (Ani-mal protein may have declined as human populationsincreased, as it has elsewhere, e.g. Kirch and Yen1982.) Thus, feral pig populations that existed inrain forests prior to Cook's arrival may have beencomprised of small-sized pigs at low densities. Whatis known about nutrition, pig husbandry practices, andagricultural land use would seem to suggest that modi-fications of the interior rain forest vegetation byPolynesian pigs was not great.
EFFECTS ON ISLANDS AND ECOSYSTEMS
Although Polynesian introductions and land useresulted in changes that continue to the present day,subsequent deliberate and accidental introductions andpractices of early explorers, settlers, agriculturists,hunting groups, bird clubs, and governments caused ad-ditional modifications. More comprehensive historiesof vertebrate introductions and distributions are given
255

elsewhere (Tomich 1969; Kramer 1971f van Riper and vanl In I I |
Riper 1982! Berger 197/1, 1975, 1981), and will not berepeated here.
The effects of the alien vertebrates now presentare complex, varied, and subtle. Some of the directeffects on different vegetational types and the biotaof the Hawaiian Islands will be mentioned in this sec-tion. As indicated earlier, the general headings willbe depredation, predation, competition, and habitatdegradation. A complete treatment is not possible, butwhat is considered most important will be stressed.Illustrations of indirect effects and of the complexi-ties involved in reducing alien influences will then bepresented. More succinct statements about problems andongoing and proposed solutions will be found in a sub-sequent section. A diagram of general impact of impor-tant vertebrate taxa is presented in fig. 1 (afterScott et al., in press). A much condensed table ofrare plant taxa threatened by alien vertebrates (table1) was adapted from the more complete treatment byWagner, Herbst, and Yee (this volume).
DepredationDomestic and feral cattle (Bos taurus). Cattle
were "historically abundant on Kauaif Oahu, Molokai,Maui, and Hawaii" and heavily overstocked on Lana'i(Tomich 1969). Feral cattle were eliminated by the mid1900's on ©•ahu, and probably Moloka'i more recently,but still exist on Maui (L.L. Loope, pers. comm.) andHawai'i. Present distribution on Hawai'i includes theSouth Kona District, Mauna Loaf and Hamakua and Punaforests. Cattle are present in remote forests, in sub-alpine scrub, or on inaccessible lava flows on ranch-lands (Tomich 1969; R.L. Walker, pers. comm.).
These animals have serious negative impacts on thevegetation of dry, mesic, and wet forests at low andhigh elevations. According to Scott et al. (in press):
Domestic and feral cattle have beenoverall the single most destructiveagent to Hawaiian ecosystems, particu-larly to mesic forests.... Koa repro-duction is completely suppressed bygrazing (Baldwin and Fagerlund 1943),and cattle are mostly responsible forconverting large tracts of forest toopen pasture on south and northwestHaleakala, lower elevations of west Mauiand Lanai, much of Molokai, the dry sideof Kohala Mountain..., the Waimeaplains, the north side of Mauna Loabelow 2200 m elevation, the mesic andwest slopes of Hualalai, most of south
256
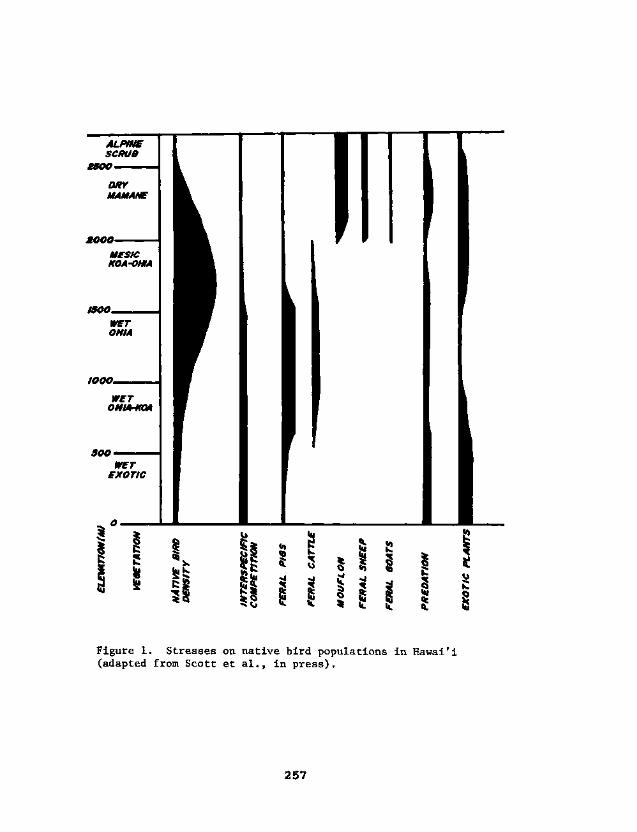
Figure 1. Stresses on native bird populations in HawaiTi(adapted from Scott et al., in press).
257

Table 1. Native Hawaiian plants listed as endangered (L), proposed for listing (P),or candidates for review by the U.S. Fish and Wildlife Service (C), that are alsothought to be harmed by alien vertebrates.
Taxon
Haplostachys haplostachyavar. angustifolia
Vicia menziesii
Gouania hillebrandi
Kokia drynarioides
Abutilon menziesii
Argyroxiphium sandwicensevar. sandwicense
Cyanea superba
2Distribution
H
H
M
H
HrL
H
0
Status
L
L
L
L
C
C
C
Threats3
Feral goats, sheep, fire,alien plants, military
Cattle/ logging, pigs,reforestation, rodents
Feral and domestic livestock,alien plants, insects
Livestock, fountain grass,rodents, fire
Axis deer, cattle, goats,insects, fire
Feral animals, especiallymouflon; removal of fruitfor propagation
Feral cattle, goats, pigs,alien plants, fire, military

2 Adapted from Wagner, Herbst, and Yee (this volume).Hawaii i, Lanaxi, Oxahu, MolokaNi, Maui, Kaua%i.Botanists can also threaten some species.

and Klauea , A consistent pattern ofcattle invading wet forests from adja-cent mesic areas occurs at ecotones onMaui and Hawaii, and formerly occurredon Kauai before control in the 1920 fs to1930fs. . .
Cattle were involved (with goats and sheep) inremoving much of the native dryland forest on Nifihauand Kahofolawe (Wagner, Herbst, and Yee, this volume).They have severely reduced mamane (Sophora chryso-phylla) trees on Parker Ranch on Hawai'i (Scowcroft1983). McEldowney (1983) summarized some of the dras-tic impacts on vegetation, soil and water supplies inthe Waimea area in the mid to late 1800 fs but suggestedthat the native forests were reduced by Hawaiian landuse practices as well as cattle depredations.
In addition to causing forest destruction andfragmentation, cattle have seriously threatened andprobably caused the extinction of many native planttaxa. Cattle currently threaten rare plants such asVicia menziesii. Abut i Ion menziesii, and Cyanea superba(table 1) . Cattle reduce native plant diversity andsimplify structure, composition, and function of vege-tation near the ground. The effects of severe grazingpressure on plant gene pools, nutrient cycling, and,plant evolution (through natural selection) are un-doubtedly severe.
Feral sheep (Qvis aries) . This species occursfrom 600-3,600 m on Hawai'i. It is present chiefly onMauna Kea and Hualalai, although it descends to sealevel in the Kafu District (Tomich 1969; van Riper andvan Riper 1982) . Feral sheep have not been seen onKahofolawe since the early 1980 fs (R.L. Walker, pers.comm. ) .
Sheep mainly damage dry areas above 1,000 m onHawaifi. Their effects on mamane reproduction can beextreme, especially near tree line (van Riper 1980;Scowcroft 1983; Scowcroft and Giffin 1983). Feralsheep compact soil and increase erosion throughestablishment and repeated use of trails. Sheep wereimplicated with goats and cattle in the destruction ofmuch of the native vegetation of Ni'ihau and Kaho'olawe(Wagner, Herbst, and Yee, this volume) . Feral sheepare a threat to the Federally endangered Haplostachyshaplostachya var. angustifolia, (table 1) , Mauna Keasilversword (Argyroxiphium sandwicense var. sandwi-cense) , Stenogyne diffusa, S. microphylla, and probablyother rare taxa in Hawai'i fs dry uplands.
260

MoufIon (Ovis musimon). Mouflon are establishedon upper Mauna Loa and Mauna Kea on Hawai1^ and in thedry kiawe (Prosopis chilensis) forests of Lanafi(Tomich 1969; van Riper and van Riper 1982). In themamane forests of Mauna Kea, they have effects on thevegetation similar to those of feral sheep (Giffin1982). In subalpine fohifa (Metrosideros polvmorpha)and alpine scrub in the Kafu District on Hawai!i, theyalso damage Kafu silversword (Argyroxiphium kauense)and other native plants; on Mauna Kea, they threaten A.sandwicense var. sandwicense. On Lana'i, mouflonapparently do not penetrate native forest (Scott etal., in press).
Feral goats (Capra hircus). Goats in Hawaifi nowoccur on all main islands except Ni'ihau and Lana*i,where they have been probably eliminated. They damagedrier open ecosystems from low to high elevations. OnMaui and Hawai1!, they are found in subalpine woodlandand alpine grassland. Goats commonly enter upperelevation wet forest on Maui and in the past alsodestroyed wet forest remnants on Lanafi. On Moloka'i,Hawai'i, and Maui, goats degrade low elevation dry for-est. On Kaua'i, they damage the edge of the AlakafiSwamp and enter wet habitats in dry periods. Goatshave had a major role in the destruction of dryland andmesophytic forest on Ni'ihau and Kahofolawe in the past(Tomich 1969; Scott et al., in press).
Feral goats are implicated as threats to Haplo-stachys haplostachya. AbutiIon menziesii, Cvanea super-ba, Hibiscadelphus distans (table 1), and other rareplants (Wagner, Herbst, and Yee, this volume). Effectson koa (Acacia koa) (Spatz and Mueller-Dombois 1973),silver-swords, Haleakala sandalwood (Santalum hale-akalae), and other rare trees have been reported (Loope1982, 1983a; Loope and Scowcroft, this volume). Feralgoats had probably eliminated all traces of Canavaliakauensis except the seedbank in the lowlands of Hawai'iVolcanoes National Park. When goats were removed, dor-mant seeds sprouted, and the plant reappeared (Mueller-Dombois 1981). Goats limit mamane reproduction inHaleakala National Park, and apparently negativelyaffect reproduction of woody native species such asCanthium odoratum, Dodonaea viscosa, and Osteomelesanthyllidiofolia in Hawai'i Volcanoes National Park(Williams 1980).
Black-tailed and axis deer (Qdocoileus hemionusand Axis axis). Blacktails are found on western Kauafiin dry native and alien forests (Tomich 1969). [Koaand 'ohifa are native elements of these forests; kukui(Aleurites moluccana) is a Polynesian introduction; andsilk oak (Grevillea robusta) and several species ofeucalyptus are aliens (Kramer 1971).] Axis deer are
261

forests and range into higher "ohi'a forests on Molo-ka'i, Lanafi, and Maui (van Riper and van Riper 1982),with a small remnant on O*ahu. Blacktails have threat-ened rain forest and rare bird habitat in the AlakafiSwamp in the past. Axis deer and pigs have badly de-graded vegetation and soils of East Molokafi, with at-tendant damage to coral reefs on the southern coastthrough siltation (Scott et al., in press). Axis deerare viewed as the most serious existing threat to La-na'i's remaining forest and are a potential threat tomesic and rain forest on Haleakala (Scott et al., inpress). Invasion of adjacent rain forest by axis deermay follow creation of open forest by cattle, pigs, orgoats.
Feral pigs (Sus scrofa). Feral pigs occur on allmajor islands except Lana'i and Kaho^lawe (Tomich1969; van Riper and van Riper 1982). Pigs were appar-ently present on Laysan Island and Sand Island at Mid-way Atoll in the late 1800fs and early 1900!s (Kramer1971). On Kauafi, Ofahu and Maui, pigs are restrictedto rain forest and grasslands on ranches (Giffin 1978)or in the subalpine forests of Haleakala National Park(Jacobi 1976). According to Giffin (1978) and Kramer(1971), pigs are abundant on Ni'ihau in arid and openareas with kiawe (Zone A of Ripperton and Hosaka1942). On Hawai'i, pigs are found from coastal dry-lands through rain forest to the upper slopes of MaunaLoa and Mauna Kea. They have been observed at 3,030 m,according to van Riper and van Riper (1982). In mamanewoodlands of Mauna Kea, densities are much lower thanin rain forest (Giffin 1978), where pigs apparentlyreach levels (19-79 /km2) unmatched elsewhere (Singer1981).
Feral pigs are the major current modifiers ofHawaiian forests, probably even exceeding damage doneby man. Pig damage has reached extreme levels in thiscentury, perhaps as a result of increasing densities aswell as expanding distributions. The reasons for thisare unclear, although it has been postulated that ani-mal protein in the form of earthworms, and mutualisticrelationships with certain dominant alien plants (e.g.Psidium cattleianum, Passiflora mollissima, Myricafaya), have made conditions more favorable for pigsthan previously (Diong 1983). Man and pigs togetherseem to be enhancing pig habitat over time, favoringintroduction of alien plants and resulting in acceler-ating damage in many areas. The role of wild dog con-trol (through poisoning and other means) in recentcreation of more favorable pig habitat in some mesicand dry forests is unstudied.
262

Pigs compound and intensify the problem of alienplant ingress by creating open habitats through diggingup, eating, and trampling native species, and by in-creasing soil fertility. Pig activity thus worksagainst reestablishment of many native plants adaptedto poor soils, and favors establishment of alien plants(this will be discussed further in the section entitled"Nutrient Cycling"). Pigs also transport plant propa-gules in their feces and pelage. The spread of alienweeds since 1945 in Kipahulu Valley seems well corre-lated with pig range extension (C.W. Smith and L.L.Loope, pers. comm.). Large areas can be affected bypig activity. It has been calculated that feral pigsin the Kilauea Forest Reserve on Hawai'i dig up overhalf of the diggable area in a year"s time (Cooray andMueller-Dombois 1981).
Alien plants enhanced by pig activity in Hawaifi,in addition to those noted above, include: Ageratinariparia, Rubus penetrans, Hedychium spp., Sonchus spp.,Buddleia asiatica, Phaius tankervilliae. Anemonehupehensis, Andropogon spp., Paspalum spp., Setariapalmifolia, Solanum pseudo-capsicum, Psidium guajava,and Passiflora ligularis (J.K. Baker, unpubl. data).
That the exclusion or removal of pigs can resultin recovery of native vegetation and changed plant com-position has been quantified through exclosure studiesin rain forests (Katahira 1980; Higashino and Stone1982), and in subalpine grasslands (Spatz and Mueller-Dombois 1975; Jacobi 1976). Some native species seemable to remain stable or increase in the absence ofpigs over varying periods of time in different areas,but continued disturbance often favors aliens. In sub-alpine grasslands in Hawai!i Volcanoes and HaleakalaNational Parks where pig disturbance continued, nativeDeschampsia australis and Panicum tenuifolium were re-placed by Holcus lanataus (Spatz and Mueller-Dombois1975; Jacobi 1976). Pigs severely damage fragile andlimited communities, such as Haleakala greensword(Argyroxiphium virescens) and Qreobolus furcatus andCarex svenonis bogs on Hawai'i, Maui, and Kaua'i (Gagne1982; Loope 1983a).
Feral pigs selectively take certain native plantspecies, thereby reducing already limited populationsor confining them to high epiphytic strata (e.g.Astelia). Tree ferns (Cibotium spp.) and Sadleria spp.are preferred items, as are other ferns (e.g. Marattiaand PteridiuitO , Astelia spp., Freycinetia, and variousLobeliaceae and Labiatae. Pigs trample Peperomia andbreak weak stems of native Cvrtandra, mints (Phvllo-stegia spp. and Stenogvne spp.), and orchids (Loope1983b) and are specifically identified as a threat tothe candidate endangered species Cyanea superba and the
263

liiiH 1)plants suck
spp.) may sometimes be taken in numbers great enough toaffect forest composition, growth forms, and successionover large areas (Diong 1983; J.K. Baker, unpubl.data).
Black and Polynesian rats (Rattus rattus and R.exulans). Black (roof or ship) and Polynesian rats arefound on all major Hawaiian islands (Tomich 1969; vanRiper and van Riper 1982), often in the same vegetationtype. Tomich (1969) indicated that black rats are alsopresent on Sand and Eastern Islands at Midway and pos-sibly other islands, whereas Polynesian rats may alsoinhabit Ni'ihau, Kafula (off Ni1ihau), Kure Atoll (atnortheastern end of the chain) and some other islands.Although R. exulans characteristically favors lowlandsand R. rattus low and mid elevations (Tomich 1969; vanRiper and van Riper 1982), both species are adaptableand found at higher elevations, sometimes in substan-tial numbers. The black rat occurs to 2,970 m aroundbuildings at Haleakala National Park (Tomich 1969) andto at least 2,440 m in Hawai*i Volcanoes National Park(Tomich 1981). The Polynesian rat has been trapped at2,060 m in Kipahulu Valley on Maui, and both speciesare common in wet koa and wet "oh^a and are present inscrub 'oh^a vegetation types there (Stone et al.1984). In Hawai'i Volcanoes National Park, black ratswere 3 times as trappable in rain forest as in mesicforest (C.A. Russell and C.P. Stone, unpubl. data).
Black rats damage flowers, fruit, and bark ofHibiscadelphus (Baker and Allen 1976; Russell 1980),and bark of Qsmanthus sandwicensisf Acacia koa, Copros-ma rhynchocarpa, and Pittosporum spp. Damage to fruitof Pittosporum hosmeri and Santalum paniculatum hasalso been noted. The importance of rats as depredatorsand pollinators of Freycinetia (Perkins 1903) has re-cently been questioned (Cox 1983), but probably pre-maturely .
Black rats in rain forests at Hawaifi VolcanoesNational Park consume green plants and seeds more fre-quently than adult or immature insects. Plants takeninclude Fragaria vesca, Rubus rosaefolius, Phvsalisperuviana, Vaccinium calycinumf and V. reticulatum.Seeds of Carex wahuensis, Microlaena stipoides, Pas-palum dilatatum, and Sacciolepis indica were eaten(C.A. Russell and C.P. Stone, unpubl. data).
Black rats in mesic forests at Hawaifi VolcanoesNational Park ate green plant material more frequentlythan insect larvae. Adult insects were taken even lessfrequently. Seeds dominated in summer diets and greenplants in winter (C.A. Russell and C.P. Stone, unpubl.
264

data). Rat food habits and densities in different veg-etation types are being further analyzed for popula-tions in native forests of Kipahulu Valley and Hawai'iVolcanoes National Park.
House Mice (Mus musculus). This species has oneof the most widespread geographical and ecological dis-tributions of any of the alien mammals in Hawaifi. Itis abundant over a wide range of vegetation types onall of the main islands, at Midway Islands, and on manyislets. Mice range from sea level to 3,920 m (Tomich1969; van Riper and van Riper 1982). Data on compara-tive densities in vegetational zones are not available,but mice are probably most abundant in lowland habi-tats, and populations are known to sometimes erupt in"drier beach, grassland, scrub, and forest areas," forexample on Maui, Hawai'i, and Kahofolawe (Tomich 1969;R.L. Walker, pers. comm.). On Maui, substantial popu-lations may exist in wet forests over 1,200 m in someareas, but not in others; possible reasons for this areunder investigation (Stone et al. 1984). Little infor-mation on Mus food habits in Hawai'i is available, butin the lowlands, invertebrates, grass seeds, fruit, andother items are taken (Kami 1966). Much more needs tobe known about the impacts of this ubiquitous alien onnative Hawaiian systems.
PredationSmall Indian mongooses (Herpestes auropunctatus).
Mongooses are present on all main islands except Kaua'i(possibly), Lanafi, Kaho'olawe, and Nifihau (Tomich1969). The species ranges from sea level to abovetimberline and is generally most abundant at lower ele-vations (less than 600 m) on windward coasts. Small tomoderate populations occur from 600-1,200 m in mesicvegetation (Baldwin, Schwartz, and Schwartz 1952) andin subalpine vegetation (Banko and Manuwal 1982; C.P.Stone, unpubl. data). However, at favorable locations,a strict elevational gradient may not hold. Limiteddata from Hawai'i Volcanoes National Park suggest twiceas many mongooses in mesic forests at about 1,880 m asin rain forest at 1,160 m and one to several times asmany animals at 30 m in coastal grasslands with aliengrasses and shrubs as at higher elevations (C.P. Stone,unpubl. data; C.A. Russell and C.P. Stone, unpubl.data).
Mongooses undoubtedly disperse strawberry guavaand other alien plants (Baldwin, Schwartz, and Schwartz1952) and prey upon colonial seabirds (King -and Gould1967; Simons 1983) and marine organisms (LaRivers 1948;Baldwin, Schwartz, and Schwartz 1952), but for purposesof this discussion, predation on native land animalswill be emphasized.
265

Predation upon Hawaiian crow or falala (Corvus
nawaiiensis) rleagiings (fiirrin i$85) and nene (Neso-chen sandvicensis) eggs and incubating females (Banko1982) are the most important obvious effects of mon-gooses on native animals in Hawai*i at present. Preda-tion on nene in subalpine nesting areas where mongoosesare often scarce may be less frequent than predation inthe lowlands where mongooses are more abundant, but itis still a major mortality factor for geese. However,vegetational cover is also important: mongoose preda-tion on pheasant nests was higher in areas of sparsevegetation at low altitudes than in areas of densercover at higher altitudes, even though mongoose popula-tions were apparently lower in the lower elevation/sparse vegetation area (Smith and Woodworth 1951).Re-establishment of lowland breeding nene populationsprobably depends on mongoose reduction there, althoughother limiting factors such as nutrition may also beimportant (Banko 1982; Stone et al. 1983). Mongoosecontrol in relatively undisturbed mesic koa-'oh^a for-est currently used by the 'alala (Burr et al. 1982)seems necessary. The rarity of red junglefowl on manyislands has been attributed to cat and mongoose preda-tion to a varying degree (Berger 1981). Effects ofmongooses on low nesting forest passerines are unknown.
Baldwin, Schwartz, and Schwartz (1952) consideredinvertebrates, including Lepidoptera, Orthoptera, Co-leoptera, Hymenoptera, Diptera, Isopoda, and Arachnida,to be important in the mongoose diet. That many of thetaxa taken are alien, may be more a function of loca-tion of collections than an indication of little impacton native invertebrates. More information on effectsof this opportunistic carnivore on birds, plants, andinvertebrates in Hawaififs native ecosystems is needed.
Feral cats (Felis catus). Feral cats occur on allmain islands and on other islands in the chain. Theywere common in the forests of some islands by the mid1800!s (Scott et al., in press). Cats are probablymost abundant at low and middle elevations, near humanhabitations, and in drier areas (Tomich 1969; van Riperand van Riper 1982), but they are found wild in remoteareas such as Kipahulu Valley rain forests and in mesicforests at high elevations on Mauna Loa (C.P. Stone,unpubl. data).
Cats may have contributed to the extinction of theHawaiian rail (Porzana sandwichensis) and are presentlyimportant predators on sea bird nesting colonies (Ber-ger 1981; Simons 1983; J.L. Sincock, pers. comm.). Ad-verse effects on native terrestrial birds probably arelimited to those that nest on the ground or low in theunderstory. Red junglefowl may have been extirpated bycats and mongooses on some islands, although other
266

factors were undoubtedly involved (Berger 1981). Ef-fects on nene populations are unknown, but the noctur-nal habits of cats might facilitate killing of femaleson nests as well as goslings. Forest birds that wouldlikely be vulnerable because they forage in the under-story include f elepaio (Chasiempis sandwichensis),Hawaiian thrush or 'oma'o (Phaeornis obscurus). puaiohi(P. palmeri), Maui parrotbill (Pseudonestor xantho-phrvs^, and po'ouli (Melamprosops phaeosoma), accordingto Scott et al. (in press). Young ^lala, as mentionedearlier, are also vulnerable when on or near theground. Tomich (1969) reported lepidopteran remains inthe tracts of 2 cats collected on Mauna Kea, but it isunlikely that the impact on native invertebrates isgreat. C. van Riper III (unpubl. data) found birds in6 of 9 cat stomachs from Mauna Kea (5 with passerines)and rodents and snails in the other 3 stomachs. Catstomachs and scats collected in native forests shouldbe saved and analyzed so that a data base can be accu-mulated.
Black and Polynesian rats. The distributions ofblack and Polynesian rats are given above. Black ratswere thought to have caused the extinction of trans-planted Laysan rails (Porzanula palmeri) and Laysanfinches (Telespyza cantans) at Midway Islands (Tomich1969); to have been partly responsible for extinctionof the Hawaiian rail (Berger 1981); and to adverselyaffect dark-rumped petrel (Pterodroma phaeopygia)colonies in Haleakala National Park (Simons 1983).Polynesian rats prey on Laysan albatrosses (Diomedeaimmutabilis) and other seabirds (Kepler 1967; Tomich1969).
Past impacts of arboreal black rats on native for-est birds are assumed to have been enormous (Atkinson1977). Reductions of forest birds elsewhere in thePacific followed introduction of black rats, and de-creases of such forest birds as Maui parrotbill, *ofu(Psittirostra psittacea), crested honeycreeper (Pal-meria dolei), and fakiapolafau (Hemignathus munroi) asa result of black rat irruptions in Hawaifi were sug-gested by Atkinson (1977). Rat predation (probably R.rattus) may have helped reduce populations of thecavity-nesting Kaua'i 'o1© (Moho braccatus). accordingto Scott et al. (in press). Probably other cavity-nesting species have also been affected. !Apapane(Himatione sanguinea) feathers were found in one of 86stomachs from black rats taken in native rain forest onHawaifi (C.A. Russell and C.P. Stone, unpubl. data).
Black rats were reported to prey on land mollusksby Perkins (1903). In the montane rain forest onHawai'i, adult insects occurred in 32%, insect larvaein 17%r and annelids in 9% of 86 stomachs collected
267

Russell and c.r, stone, unpubi. data), Mult analarval insects occurred more frequently in winter, andannelids in summer. Adult insects taken were in theorders Diptera, Hemiptera, and Hymenoptera, while lar-vae were in the orders Coleoptera and Lepidoptera.Further work on diets of both species of rats in Ki-pahulu Valley is under way under National Park Servicecontract with B.P. Bishop Museum (Stone et al. 1984).
Interspecific CompetitionNative and alien birds. Banko and Banko (1976)
concluded that 3 introduced avian species—the commonmyna (Acridotheres tristis), Japanese white-eye(Zosterops japonica), and red-billed leiothrix (Leio-thrix lutea)—had potential roles in reducing popula-tions of native forest birds, on the basis of habitatoverlap and food habits. They traced the introductionsand histories of populations of each species inHawai'i. The barn owl, Tyto alba, was not believed tobe a serious threat to the "io, Buteo solitaris, or theshort-eared owl, Asio flammeus, in the past or at pres-ent (Banko and Banko, n.d.). Competition for, and re-duction of, such important lepidopteran taxa as Geo-metridae, Pyralidae and other food resources, espe-cially during the nesting season of native birds, ispresumed. Banko (1978) and Gagne (1980) also outlinedthe deleterious effects of continental flies and waspson insects that were formerly important foods for 'o'u,Hawai'i creeper (Qreomystis mana), palila (Loxioidesbailleui), and ^ma1©, among other species. Competi-tion could have been particularly important at theheight of alien bird population expansion (Banko andBanko, n.d.), especially if coupled with a diminishedprey base resulting from insect predation, parasitismand diseases. Atkinson (1977), using a similar his-torical/ecological approach, also concluded that themelodious laughing-thrush or hwa-mei, Garrulax canorus,might be a factor in competition with native birds,based on widespread distribution of this species inHawaiian forests. However, he judged that the intro-duction was too late to implicate the hwa-mei in majordeclines of native birds.
Ralph (1978) suggested that among 10 common pas-serines in Hawaifi Island forests, "interspecific ac-tions appear to play a secondary role at most times."His data have not yet been fully analyzed or published.
Conant (1981) studied distributions and densitiesof 8 alien and 9 native birds on Mauna Loa and con-cluded that Leiothrix may have displaced the fomafo orHawaiian thrush in mountain parkland, savanna andfohifa dry forest. Although Japanese white-eyes in-vaded closed rain forest in considerable numbers, com-petition was thought to be of little importance in
268

controlling distribution of natives. Conant noted,however, that species densities could be limited to thedetriment of less aggressive competitors around scarcenectar sources.
Van Riper (1976) hypothesized competition betweenthe common famakihi, Hemignathus virens, and Japanesewhite-eye in !amakihi territories. He found that al-though lack of mamane nectar could prevent breeding,excess nectar (provided by him) resulted in increasedterritorial defense and low breeding success (van Riper1984). The precise influence of Zosterops was not de-termined in his study, but it could be crucial depend-ing on food resources and population levels. Conant(1976) provided some evidence that common famakihi and"apapane affect each otherfs densities depending uponwhether or not nectar is the main food resource (if itis, fapapane densities exceed those of ^makihi); how-ever, she noted that rainfall and other parameters[perhaps often including competition with Zosterops]may also define optimum habitats for both species.
Pimm and Pimm (1982) suggested that more dominantnative species such as the 'i^wi, Vestiaria coccinea,and several extinct and endangered honeycreepers wereless able to subsist on marginal resources than lessaggressive competitors such as 'apapane and 'amakihi.Alien species such as Zosterops might adversely affectsuch dominants by forcing increased territorial defenseat good nectar sites (preferred species with manyblooms); by relegation to less favorable sites (pre-ferred species with fewer blooms or secondary tree spe-cies) ; or through reducing and further dispersing analready dispersed nectar supply.
Scott et al. (in press) discussed distributions,habitat occurrences, and densities of alien bird spe-cies in Hawai!i. Space is not available here to coverpossible competitive interactions species by species,but alien bird richness was highest in dry woodlandsbelow 1,500 m with introduced plant understories. Itwas noted that few introduced gamebirds penetrateclosed forests, and that disturbed habitats containmore alien bird species than undisturbed. Disturbedforests allow avenues of ingress for alien birds, ac-cording to the authors.
Mountainspring and Scott (in press) used partialcorrelation matrices for paired species (with habitateffects removed), to infer competition (for food) amongalien and native forest passerines. The bulk of signi-ficant correlations were positive (67% of total), sug-gesting species association rather than competitiveavoidance. The authors noted that such species as theomnivorous Zosteropsf Leiothrixr and Garrulax; the
269

insectivorous 'elepaic, Kaua'i creeperbairdil, ' akepa fLoxops coccineusl; and coiion 'ama-kihi; and the nectarivorous fapapane and fifiwi showedpositive associations. Increasingly or decreasinglyfavorable habitat for a particular guild was evidentlysimilarly responded to by all species in the guildacross a variety of locations on each island. The onlyconsistent negative partial correlations (presumed toindicate competitive avoidance) were for Zosterops/Chasiempsis on windward Hawaifi and Zosterops/Vestiariain montane forests on Hawaifi. Scott et al. (in press)hypothesized competition for understory insects to ex-plain the first relationship and competition for lowerquality (dispersed?) nectar to explain the second. Asnoted earlier, increased territorial defense by Vesti-aria may be another (not exclusive) explanation fornegative associations over the long term.
Native birds and rats. Atkinson (1977) noted thatmany Hawaiian forest birds depend on nectar or insectsfor food, but thought that many insects are unavailableto rats because they are too small or impossible fprrats to reach. This generalization has not been inves-tigated. Perkins (1903) pointed to the depredation ofrats (probably the more arboreal R. rattus) on Frey-cinetia arborea fruit. Since ̂ 'u and falala also de-pended heavily on this fruit, rats may have especiallycompeted with them when and where rats and ̂ 'u and/orfalala were abundant (Banko and Banko, n.d.).
Based on our knowledge of rat food habits inHawai'i (see previous discussions) it seems that directcompetition would be likely for native birds that spe-cialize on fruit and large conspicuous invertebratesthat are active day and night. However, insect eggs,pupae, and many small invertebrates on and above groundlevel might be vulnerable to both birds and rats.Thrushes and perhaps pofouli (a specialist on snails,according to Baldwin and Casey 1983) are among nativebirds that may compete with rats, in addition to spe-cies already mentioned. However, almost nothing isknown of dynamic interrelationships among rats (bothspecies), plants and invertebrates' in thrush andpo'ouli habitats. (The same interrelationships shouldalso be studied in similar areas not frequented bythose avian species, for comparison.) Information onrats and their adverse effects in other important for-est bird habitat is also lacking, but preliminary dataon rat densities and food habits and bird abundancefrom several areas in Kipahulu Valley are being ana-lyzed (Stone et al. 1984).
Habitat DegradationOptimum habitat for a native animal may be
considered as the natural complex of physical and
270

biological factors in which a population is at a peakthat can be sustained over time without damage to thehabitat (carrying capacity). Introduction of mostalien elements usually degrades the optimum habitat fornative species. Alien diseases, predators, and compet-itors of vertebrates have already been discussed, orwill be discussed elsewhere (van Riper and van Riper,this volume). Additional man-influenced changes suchas grazing, clearing land for agriculture, burning, andlumbering (cow, plow, fire, and ax) also destroy or de-grade habitat for many natives, but these effects arenot the subject of this paper. Direct effects of alienanimal depredations on native plants and ecosystemswere discussed above, but not in the context of nativeanimal habitat. This will be touched on here, largelyin relation to native birds (Gagne and Christensen,this volume, address invertebrate habitat). Informa-tion is largely taken from Banko and Banko (n.d.), andthe large volume of data collected by Scott et al. (inpress) during the U.S. Fish and Wildlife ServiceHawai'i Forest Bird Survey.
Effects of feral sheep and goats on nene habitatwere judged important in local population impacts byBaldwin (1947). Cattle, goats, and possibly pigs werebelieved highly important factors in nene habitat de-gradation in the mid-1850fs and later (Banko and Banko,n.d.). Effects begun then, together with invasion ofalien plants, may have permanently altered considerablehabitat.
Grazing by sheep and cattle was partly responsiblefor eliminating, fragmenting, and degrading high-elevation akiapola'au habitat (Scott et al., inpress). Effects of feral sheep, goats, and mouflon onmamane habitat of the Federally endangered palila wereserious enough to force a court order to remove theferal sheep and goats from Mauna Kea (Kobayashi 1979;Scowcroft 1983). Cattle grazing and lumber harvest onleeward Hawai!i severely affected habitat of 'akepa,Hawai!i creeper, and falala. According to Giffin,Scott, and Mountainspring (in prep.), nearly all of theundisturbed and none of the disturbed koa-fohifa for-ests in Kona were once occupied by crows. However,P.C. Banko (pers. comm.) has found breeding 'alala inkoa-»ohifa forests that were highly disturbed by cattleand other activities. Scott et al. (in press) saidthat cattle damage to forest understories on theirKohala study area on Hawaifi correlated with low densi-ties of 'elepaio.
Feral ungulates were thought responsible for thelower famakihi densities in Maui dry forests than insimilar dry forest habitat on Hawai*i. On Moloka'i,•apapane are absent or present in low densities as a
271

9f wmifiwn t[ iiiii m^i PWi pio Qfflcattle. On East Moloka'i, loss af lamakihi is tiad toaxis deer depredations.
Feral pigs are important in the reduction of cer-tain Hawaiian lobeliads, especially in wet forests(Diong 1983). Native birds that use these taxa includeBishop!s 'o'o, Mono bishopi (if extant); the fi!iwi;and the fo fu (Scott et al., in press). Bishop's !o!omay be especially sensitive to habitat degradationcaused by pigs, partly because of lobeliad reduction.Pigs (and rats) also consume Freycinetia, favored byfo fu and 'alala, and probably lower habitat quality ofthese birds accordingly.
Feral pigs may have negative effects on the Feder-ally endangered Pseudonestor and Melamprosops. Bothfavor dense forest understories for foraging, and feralpigs are effective in simplifying understory composi-tion and structure. Casey, Mountainspring and Scott(in prep.) showed that pofouli habitat had "light pigdamage and well developed herb, ground fern, and mosslayers." They hypothesized destruction by pigs ofmicrohabitat needed for land snails and other inverte-brates favored by pofouli.
Habitat degradation by feral pigs is also thoughtto affect Hawaifi creeper, fakepaf and felepaio onHawaifi; and Kauafi thrush, small Kaua'i thrush, andthe Kauafi '0*0 on Kauafi. Foraging of crested honey-creepers on Maui on understory nectar producers such asRubus hawaiiensis when Metrosideros polvmorpha bloomsare scarce, may also be reduced by feral pig activity(Scott et al., in press).
The role of alien birds in degradation of habitatof native birds may be important. Peak populations ofbirds that invade forest ecosystems might affect foods(insects and plants) severely enough to permanently re-duce habitat quality for natives. Such species as theJapanese white-eye, red-billed leiothrix, and commonmyna may have already reduced habitat quality by thismeans in many areas.
Indirect EffectsImpacts on other aliens. The important, and some-
times mutualistic, role of feral pigs in dispersing andencouraging weedy alien plants has been discussed pre-viously. Mongooses, rats, and mice are also respon-sible for spreading alien plants, but the effects areless obvious and the magnitude uncertain. Alien birdsdisperse or are closely associated with numerous alienplants, of which Passiflora, Myrica, Schinus, Lantana,and Clidemia are probably the most important in nativesystems. Warshauer et al. (1983), Lewin (1971), and
272

Scott et al. (in press) noted associations of the fol-lowing introduced birds with Passiflora; black fran-colin (Francolinus francolinus), Erckelfs francolin (F.erckelii), gray francolin '(F. pondicerianus), kalijpheasant (Lophura leucomelana), common peafowl (Pavocristatus), wild turkey (Meleagris gallopavo), Leio-thrix lutea, mockingbird (Mimus polyglottos), Japanesewhite-eye, northern cardinal (Cardinalis cardinalis),house finch (Carpodacus mexicanus), and spotted dove(Streptopelia chinensis). Fortunately, many of thesespecies do not penetrate intact forest, but most willuse forest openings created by man or feral animals foringress.
A number of avian species in addition to thosementioned above are associated with alien grasses,herbs or shrubs (Scott et al., in press). They includeCalifornia quail (Callipeopla californica), ring-neckedpheasant (Phasianus colchicus), zebra dove (Geopeliastriata), Eurasian skylark (Alauda arvensis) (a goodindicator of degraded, fragmented forests), melodiouslaughing thrush, and nutmeg mannikin (Lonchura punctu-lata). Overall alien species diversity is highest inbroken woodland and is heavily influenced by game birdoccurrence. Cardinalis distributes Psidium spp. andSchinus terebinthifolius in lowland forests (Scott etal., in press). In a more subtle relationship amongaliens, van Riper (1980) noted that alien birds mightbe contributing to the spread of naio (Myoporum sandwi-cense) on Mauna Kea through dispersal of seeds whileferal sheep reduce competition from mamane throughbrowsing.
Areas disturbed by feral ungulates contain agreater variety of alien bird species than undisturbedareas (Scott et al., in press). Stock pond and otherwater sources created by or for cattle are sought outby various game birds, house finches and common mynas,among others. Cattle and pig wallows also attractalien insect vectors of malaria or pox (van Riper andvan Riper, this volume) that may thus reduce nativebird populations. In some instances, continued pres-ence of ungulates can perpetuate alien plants thatwould eventually give way to natives in their absence(Loope and Scowcroft, this volume). The establishmentof alien plants after disturbance by feral animals,which are then able to maintain permanent populationsin the absence of animals through allelopathy, mono-typic stands, or altered nutrient cycles, has also beennoted (Smith, this volume). The encouragement of in-troduced vegetation via alien animal distribution andcultivation contributes an alien fauna of snails, in-sects, spiders, other invertebrates, and plant patho-gens (Howarth, this volume). Ecosystem modificationscaused by the negative effects of these associated
273

aliens on native vertebrates and invertebrates could be
tremendous.Forest openings do not always need to be caused by
man or his associates to encourage alien birds. Theapparently natural phenomenon in Hawaiian forestscalled fohifa dieback (Mueller-Dombois, this volume) iscorrelated with reduced native and increased introducedbirds. In scattered dieback sites in the Hamakua areaof Hawai'i, 'apapane, 'i^wi, "oma1©, and felepaio num-bers were 70, 77, 47, and 93% lower than in tall,closed canopy forest. Red-billed leiothrix and Japa-nese white-eye numbers were 30 and 34% higher (Scott etal., in press). Patchiness of vegetation of differentsorts can favor native and non-native alike. For exam-ple, 'i'iwi and melodious laughing thrush both occur ingreater numbers where understory diversity is high; na-tive tree falls may help create such diversity (Scottet al., in press).
Nutrient cycling. Smith (this volume) stated thatthe effects of alien plants on nutrients in Hawaiianecosystems deserve special consideration. The dangerthat aliens such as Myrica can, through nitrogen fixa-tion, create their own favorable environment onHawaii's young and nitrogen-poor soils was empha-sized. A study to determine the role of Mvrica faya inaltering primary succession in Hawaifi Volcanoes Na-tional Park has been proposed and funded (P.M. Vitou-sek, pers. comm.).
Feral pigs also play an important role in alteringnutrient cycling in Hawaiian systems. They are distri-butors of Myrica faya, the berries of which average 12%of the total volume of food taken in the Puhimau Unitof Hawai*i Volcanoes National Park (n = 54 adults takenby hunting) on a year-long basis (C.P. Stone, unpubl.data). But probably more importantly, pigs innitrogen-limited areas can also modify nutrient sinks,availability, and dynamics through their rooting ac-tivities. Where pigs are absent, organic material(and nutrients) build up (Higashino and Stone 1982) andultimately affect soil formation. Where pigs root, theavailability of "nitrogen [and other nutrients] toplants and the potential for nitrogen losses to thesite (in leaching or denitrification) is greatly in-creased" (P.M. Vitousek, pers. comm.; Vitousek et al.1979; Vitousek et al. 1981). Short-term nitrogenavailability (favoring alien plant establishment) andlong-term nitrogen loss (preventing the usual succes-sion of natives) may be the pattern on such sites, es-pecially if pig disturbance continues. Parent soilmaterials (ash, pahoehoe, or fa fa) also affect nutri-ents, and the long-term advantages of nitrogen-fixerson some sites may not always hold as nutrient
274

availability changes (Vitousek, Van Cleve, and Bala-krishnan, in press). However, the continual influx ofalien plant species is likely on pig-altered sites.The effects on plant and animal succession and commun-ity composition as a result of altered nutrient avail-ability alone could be substantial. Pigs also alterecosystem structure and processes through trampling,alien propagule introduction and enhancement, reproduc-tive reduction of natives, and soil erosion, as notedabove. Pig rooting and defecation favor alien soilinvertebrates, further altering nutrient cycling(Howarth, this volume).
REDUCTION OF ALIEN IMPACTS
It should go without saying that extreme care mustbe exercised in the introduction and translocation ofalien birds and mammals in Hawai'i. The best way tomanage aliens is to prevent new species from enteringthe State and to prevent aliens that are present fromaffecting additional areas. As indicated in table 2,introductions that have recently succeeded are occur-ring at a much more rapid rate than introductions byeither Polynesians or nature prior to man. The Hawai'iDepartment of Agriculture (DOA) and the Department ofLand and Natural Resources (DLNR) have responsibilityfor controlling the introduction of vertebrates underChapter 150A, Part II of Hawaifi Revised Statutes (HRS)and Chapter 187, Sect 1.2, HRS (Burr 1984). DOA admin-isters Chapter 150A and maintains a list of prohibitedentry species. It is expected to confiscate animalsand charge the owners for expenses. DLNR administersChapter 187, which covers translocation from one areain the State to another as well as importation. How-ever, only the introductions proposed by DLNR arecovered; other proposals are not addressed. If ananimal escapes, it is classified as "wild" and a permitis required under Chapter 191 to control or eliminateit when it is involved in agricultural damage, is anuisance, or is a health hazard (Burr 1984). The DLNRand DOA cooperate closely on updating the DOA list ofrestricted or prohibited entry species. There havebeen recommendations to develop a list of species thatshould be exempted from the wild bird protection provi-sion.
Complexities of Damage Control"Ecology may not only be more complex than we
think; it may be more complex than we can think" (F.Egler). Perhaps the major problem in understanding thetrue impacts of aliens and devising management strate-gies to reduce them is the number of confounded para-meters involved. Before damage can be understood, pre-dicted, and overcome, one may need to know how inter-actions occur among: soil fertility, weather, land use
275

Land Mammals
Years perSuccessful Species
Land Birds
Years perSuccessful Species
1 3
20,000,000 500
20* 1
1,000,000 1500
18
11
45
4
These underwent considerable adaptive radiation to form 35extant species and many now extinct species (see Olson andJames 1982).
276

history, plant phenology, predation, competition andother interrelationships with many species; populationstructure, density, distribution, and dynamics; animalmovements, nutrition, and the vegetation complex. Suchknowledge does not come cheaply or in a short time orfrom one study area, because problem vertebrates arelong-lived, mobile, adaptable, and difficult to ob-serve. Good information on one parameter, such aspopulation density, may be misleading (Van Home1983) . For example, high densities of young animalsmight be indicative of poor habitat, which is occupiedby young because of social dominance by adults else-where, or because of high reproductive success and tem-porary survival. Attempting to reduce vulnerable prob-lem animals in such situations might not be effica-cious.
Other information required for good control pro-grams includes the effects of different populationlevels upon the resource at risk, the cost of conduct-ing the control, the value of the resource protected,effects of control on nontarget animals, effects ofcontrol on the resources being protected and on theecosystem, the amount and length of followup needed,and feedback as to the effect on the problem animals.Managers need to establish long- and short-term priori-ties to determine what resources can be brought to bearon problem animals in different ecosystems and situa-tions. To predict effectiveness of control under es-tablished time and budget constraints, animal popula-tions and productivity should be compared (throughresponsive models) to determine the optimum efforts toachieve the desired reduction in damage. Proper moni-toring of control effects necessitates systematic sam-pling of both animal populations and the resources thatthey damage. This should really be done pre and postcontrol at a minimum.
The goal of reducing the effects of introducedvertebrates upon Hawaiian ecosystems is not accepted byeveryone. Some believe that nothing can be done;others that it is too expensive and/or time consuming;and others consider that alien vertebrates add scien-tific, aesthetic, social, recreational, or even cul-tural and religious values that are more important thanthe values of native ecosystems. Studies of the impor-tance of these values in the minds of Hawaii's citi-zens, and of the economics involved, have not yet beenmade; however, most informed people (probably a dis-tressingly small portion of any population!) wouldprobably agree that preservation and management of na-tive ecosystems in some areas is necessary and desir-able. There is now overwhelming evidence that inHawai'i this depends upon reduction or elimination ofalien vertebrate impacts. To eliminate the adverse
277

it if iiifiiiiuT to MiOitior man? or cuoiuring the aliens, at least "tempararily." The fact thatindividual animals can't just be killed "temporarily"raises a conflict about the value of life. So a deci-sion to reduce the adverse effects of alien vertebratesbecomes a complicated economic, social, political,ethical, aesthetic, recreational, scientific, and landuse decision. It also involves a worldwide, as well asa local, community of interest in Hawaiian ecosystems.Letfs assume we have somehow made the decision to con-trol alien vertebrates to benefit native Hawaiian eco-systems in some areas. Then what?
Depredations"Good fences make good neighbors" (R. Frost). If
opposing land uses such as sustained yield recreationalhunting and native ecosystem preservation and manage-ment are to be supported and perpetuated (and thisseems realistic), feral ungulates must be excluded fromsome areas and perpetuated in others. Fence construc-tion is costly and maintenance is continual, but thereis no other way to sustain adjacent land uses withthese opposing objectives. Programs in Hawaifi Volca-noes and Haleakala National Parks to reduce and eventu-ally eliminate feral goats and pigs are examples ofungulate control programs that depend upon fences. Thequestion of who should build and maintain fences de-serves further discussion elsewhere, but in this casethe Federal government is assuming the sole responsi-bility for perpetuating Hawai!ifs native ecosystems.
Once fences are constructed, reduction of the ani-mals by hunting, snaring, trapping, or poisoning ispossible. Removal of live animals for use elsewhere isusually expensive and time consuming, requires a placeto put the animals, generally costs more than it isworth biologically, and does not usually result inelimination of the all-important last animals that canquickly repopulate the area with offspring. There are,of course, political and social considerations. Driftfences may be used to restrict animal movements withinfenced areas or to direct animals toward accessibleareas for removal. Internal barriers consisting ofcombinations of fences and topographic features may benecessary to delimit areas from which animals can beeffectively removed. Such areas should be large enoughto reduce major fencing costs and avoid artificialpaddock-like situations, but small enough to allow ef-ficient elimination of animals with the resourcesavailable.
Public hunting may be used as a tool to reduce de-predations of ungulates in some instances, but manage-ment by public hunting alone usually results in sus-tained yield. Hunting as a control method is most
278

effective where habitat is limited and accessible. Re-moval of sheep from Mauna Kea and goats and pigs fromparts of Hawaifi Volcanoes National Park has taken anorganized, sustained use of agency "shooters" in con-junction with public "hunters." Barrett and Stone(1983) found that the Deputy Ranger or Citizen HunterProgram active in Hawaifi Volcanoes since 1972 was noteffective in reducing feral pig populations except inhighly accessible areas (within 500 m of a road). Theaverage removal rate by citizen hunters (percentage ofestimated carrying capacity) was 3% of the adult pigpopulation every 6 months; 30-40% should be removed ifpopulations are to be reduced to extinction in a 3-5year period. Through use of agency hunters and dogs,we have achieved an estimated removal rate (as of May1984) of 25% in one rain forest unit and over 50% in amesic forest (Stone and Taylor 1984).
The Hawai'i Department of Land and Natural Re-sources recognized the need to manage ungulates withinnative ecosystems in the Hawai'i Wildlife Plan (HawaifiDivision of Forestry and Wildlife 1983:63):
Where the presence of big game popu-lations within sensitive native ecosys-tems is destructive, elimination can beaccomplished through public or staffhunting opportunities.... Where thepublic hunting is inadequate in removingexcess game animals, drives and trappingshould be employed, but only as a lastresort due to the high costs.... Feralpig, goat and sheep populations shouldbe monitored and maintained at levelsresulting in minimum damage to water-sheds and native ecosystem protectionand should be controlled by publichunting whenever possible.... In NaturalArea Reserves, the objective should beto reduce feral game mammals to the low-est possible levels using public hunt-ing.
When combined with fencing and critical evaluation, andif organized properly, such programs might keep somepopulations in check. However, in most valuable natu-ral areas, the goal should be elimination of ungulatesrather than sustained yield, even at low populationlevels. The potential for increase when even a fewanimals remain is large.
Control of depredating ungulates with chemicaltoxicants is a controversial and expensive proposition,but it deserves further consideration in Hawaii, con-sidering the magnitude of the problem and the scarcity
279

of secondary and non-target animals. No toxicants are
currently registered for use witn ungulates?use entails a complicated process of registration,classification, labeling, and certification under theFederal Insecticide, Fungicide, and Rodenticide Act(FIFRA) of 1974 and the Federal Environmental PesticideControl Act of 1972, administered by the EnvironmentalProtection Agency (EPA) (Hood 1978). Use of toxicantsfor pig control in remote areas in Hawaifi would prob-ably necessitate registration under Section 24 ofFIFRA, which provides that a state may register pesti-cides formulated for distribution and use within thatstate to meet special local needs, if the state is cer-tified by EPA as capable of administering the Act. Al-ternative emergency provisions under Section 18 ofFIFRA, or Experimental Use Permits (EUP) for chemicaluse also exist under more limited conditions of re-search and/or area of application.
The use of chemicals is neither a panacea nor anenvironmental disaster. Chemicals can be used safelyand effectively in some cases and can have certainadvantages such as low cost (once registered), targetspecies specificity, non-specificity within a popula-tion (all sexes and ages are vulnerable), and ease ofuse in remote areas. However, chemical effectivenessusually depends upon bait acceptance, and some animalsmay be difficult to attract to bait—e.g. feral pigs(J. Hone, pers. comm.; R.H. Barrett, pers. comm.; C.P.Stone, unpubl. data). Chemicals, like other animalcontrol tools, must be repeatedly used as long as re-producing animals remain. Complete reliance on chemi-cals, biocontrol, fences, hunting, or any one tool isunrealistic, but all appropriate control methods shouldbe developed and improved for use where needed. Al-though registering a toxicant is costly, informationgathered in responsible research/management programswill be useful in meeting eventual registration re-quirements. Toxicants will likely be needed to controlsome species in some situations (see discussion below).
A number of research/management emphases are es-sential in well-conducted depredation control programs,and considerable research information and continualfeedback are usually necessary. Some idea of ungulatepopulation size, sex, and age structure, reproductiverate, and distribution in the control unit is necessaryto determine whether rate of removal is sufficient toeffect reduction. Information on rate of removalshould be continually recorded and compared with theo-retical or known reproduction and mortality. Indexesto the population being removed and to the resourcesbeing damaged are required to determine progress andadjust control strategies. Supplemental information onanimal movements through radio telemetry is useful in
280

improving and adjusting control strategies to changinganimal behavior and densities. Data on food habits mayalso provide cues to vulnerability or shifting behaviorpatterns as control progresses.
Primary emphases in ungulate control programsshould be:
1. The necessity for efforts lasting many years.2. Continual learning and feedback about success
of control (monitoring).3. Provision for sufficient resources to effect
reduction.4. The development of multiple methods to reduce
animals.It is likely that what will work in one area at onepopulation level will not be as useful under differentconditions. To summarize alliteratively, multiples[years and methods], money, monitoring, and modifica-tion are keys to controlling depredations by ungulatesin Hawaifi.
Predation"Well, so much for the unicorns" [said Noah,
standing over 2 carcasses]. "From now on all carni-vores will be confined to C deck" ("The Far Side").Some things just don't mix, and the best way to pre-serve one is to get rid of the other. It is generallytrue that when predators need to be "controlled" topreserve and manage native Hawaiian ecosystems, thebest way is to kill them. Mongooses, cats, and ratssimply cannot be effectively excluded or repelled ortrapped and transplanted in most situations in thewild. Again, this is a controversial matter with manyramifications beyond the ecological, but letfs assumewe have opted for controlling predator impacts.
Predator damage may sometimes be controlled bytrapping where the plant or animal to be protected isrestricted in distribution. Examples are protection ofdark-rumped petrels from mongooses, cats, and rats atbreeding colonies (Simons 1983); protection of nene inrelease pens in backcountry situations (N. Santos,pers. comm.; H. Hoshide, pers. comm.); removal of mon-gooses around an 'alala nest (Giffin 1983); or protec-tion of a group of rare trees from rats (C. Zimmer,pers. comm.). Shooting and trapping have been effec-tive in removing cats from somewhat limited areas (e.g.415 ha Jarvis Island) (Rauzon 1983), if enough manpoweris available. However, chemical toxicants are neces-sary in most cases where the resource to be protectedfrom predators is dispersed over a large area or overtime.
Several rodenticides have been registered foragricultural use and some laboratory and field work has
281

tone on mongoose and eat toxicants (iroMvoitti anaWoodside 1953; Kridler 1966; L. Pank, pers. comm.).Risk to other animals (nontarget hazards) and to ani-mals that eat poisoned animals (secondary hazards) areminimal in Hawai1!, at least insofar as native speciesare concerned. Nevertheless, we need quantitative in-formation about these hazards; about baiting techniquesin native ecosystems (including bait spacing, timing,substrate, and doses); more lab research on toxicitylevels, acceptance, and appropriate chemicals; and moreinformation on predator ecology and potential for con-trol in Hawai'i^ ecosystems. Basic practical ques-tions of feasibility in terms of reinvasion rates andsizes of areas that can be effectively treated in dif-ferent situations are largely unaddressed.
A 3-year program headed by the U.S. Fish and Wild-life Service (FWS), with the National Park Service(NFS), and the Hawai'i Division of Forestry and Wild-life (DOFAW) cooperating, has been designed to developa "drop-bait" toxicant to eliminate or reduce mongoosesseasonally in nene and falala breeding areas, waterbirdhabitat, colonial nesting sites for seabirds, and per-haps native forest other than that used by crows. Theobjective is to obtain a special local need registra-tion under Section 24 of FIFRA (J. Keith, pers.comm.). Current research on rat ecology and control innative forests is under way, with limited efforts inHaleakala and Hawai'i Volcanoes National Parks (Stoneet al. 1984; C. Stone, unpubl. data). Funding for amore substantial program of rat and mongoose researchmay be available from NFS and could be coordinated withthe mongoose project headed by FWS. The policy of theDLNR on small alien mammals as stated in the HawaifiWildlife Plan is to support research by others on pred-ators and rodents; to implement control only when it isneeded and will be effective; and to control only wherecost/benefit ratios are favorable (Hawai'i Division ofForestry and Wildlife 1983).
Interspecific Competition"New questions arise when many populations and
entire biomes are being fragmented and reduced on sucha scale and at such rates. These questions are a greatchallenge to the ingenuity of biologists .... Unlesswe solve them, we will end up with less than we intend,struggling in our ignorance to protect geneticallyeroding populations and decaying ecosystems" (T. Love-joy) . Despite a number of introductions of alien birdsinto Hawai'i in the last century (Moulton and Pimm1983) and documented increases and extinctions of thespecies and populations involved (Banko and Banko,n.d.)/ we do not have good quantitative and qualitativeinformation about adverse impacts of introduced birdson the native avifauna. Banko and Banko (n.d.) used an
282

historical/ecological approach to infer competitionamong different species and in different areas. Iffood habits of species pairs were similar and alien in-creases coincided in time with native decreases, compe-tition was assumed to have played a role. Mountain-spring and Scott (in press) and Conant (1981) examineddensities of various species in different ecosystems atone point in time. Relationships between species pairsthat were negative were assumed indicative of competi-tion, and those that were positive, of association, forthe purpose of using a common food base or other mutualresource. Variation in relationships with time of yearor over a period of years was not considered.
Although both approaches have value, a full under-standing of interspecific competition among native andintroduced birds in Hawaifi awaits further study. Dy-namic relationships throughout the year over a periodof years need to be explored. Densities, reproductiverates, mortalities, and movements of potential competi-tors should be studied in relation to habitat variation(e.g. bloom or insect abundance, rainfall, temperature,population levels of other avian species, impacts ofalien mammals, diseases, and other biota) over time.The effects on forest resources (especially during peakalien population levels) need to be determined and thepermanence ascertained. Activity budgets, densities,food habits, and reproductive success of native birdsin habitats with and without alien birds should be de-termined.
The potential for additional species of introducedbirds to compete with native species still exists.Williams (1983a, b) suggested that red-vented bulbuls(Pvcnonotus cafer) may have been deliberately releasedon Ofahu and on Hawaifi in 1966 and 1982, respective-ly. The red-whiskered bulbul (P. jacosus) was appar-ently accidentally released on O'ahu in about 1965.Neither species is noted for colonizing across openwater, but both seem to be increasing on Ofahu and maydisperse to neighbor islands (Williams 1983b; Conant1983). Bulbuls are known to be rapid colonizers, agri-cultural pests, and potential threats to native forestbirds. The possibility that these species and otherintroductions will further reduce forest invertebratesand plants must be taken seriously; initial populationsshould be removed where possible or closely monitoredwhere not possible.
An important species such as the Japanese white-eye or an irrupting species that cannot be controlledshould be followed through introduction or buildup,population irruption, and population decline in severalareas, and its impacts on forest resources, includingnative avian species, should be determined. Japanese
283

white-eyes are currently widespread in native forests
but may be declining or on the verge of doing so Insome areas already (Dunmire, 1962). The possible in-crease of the Japanese white-eye and melodious laughingthrush in Kipahulu Valley (Scott et al., in press); thepossible expansion of the laughing thrush at lower ele-vations in Kafu and Kona; the decline of Leiothrix atlow elevations especially on O'ahu (Shallenberger1981); the close association of Leiothrix and thelaughing thrush with increasing naio (van Riper 1980)on Mauna Kea; and the increase of Leiothrix on north-west Haleakala in Kula, are some areas of interest. Weneed a better understanding of the dynamics of alienbird populations in Hawaififs forests. More informa-tion on rat population dynamics and behavior in dif-ferent vegetation types, areas, seasons, and years isalso needed, to better understand the importance ofrats as competitors with the native avifauna.
In addition to better understanding the phenomenonof competition, we need to give further emphasis to theimportance of large, intact areas in preserve design.As indicated by Scott et al. (in press), native birdsneed large undisturbed tracts of native forest to pro-vide buffers against irruptions of alien birds and theperturbations caused by alien birds and mammals. Na-tive invertebrates are also well served by intact na-tive forests (Gagne and Christensen, this volume).Many introduced species in the past, and likely in thefuture, will be stopped near the forest edges unlessthere are roads, clearings, or trails into native for-ests. Activities (including feral animal control)that open and fragment forests, thus enhancing furtherinvasion of alien birds, invertebrates, and plants,need to be carefully evaluated. Optimum sizes of"alien resistant" tracts in different vegetation typesshould be determined. Vertebrates such as the melodi-ous laughing thrush, Leiothrix, and Zosterops that dopresently penetrate largely unmodified forests can havepotentially devastating effects on native plant, inver-tebrate, and avian forms, at least during peak popula-tion phases.
Management-Research Coordination"#!%?$ Ivory tower research!" "#!%?$ seat-of-the-
pants management!" (Subdued conversation between a nat-ural resource manager and a researcher). In practicewe often take shortcuts in solving problems in naturalresource conservation. We donft have enough informa-tion, but we must begin to "do something.ff In actingprematurely we risk making mistakes, but often theproblems are severe and call for immediate attention.Waiting for more facts isnft good enough, especiallywhen they accumulate slowly. It is important that ifwe are to manage without many of the facts needed,
284

however, that we proceed cautiously and learn from ourmistakes. Trial and error management of alien verte-brates must be designed to collect data on as manyaspects of the ecology of damage as possible. Manage-ment efforts should be treated as experiments with rep-licates, controls, proper experimental design, statis-tical testing, and reporting (McNab 1983). Researchpersonnel should be directly involved in coordinatingwith Management to increase the usable information fromsuch "experiments." We cannot afford to further upsetHawaifi!s badly disturbed native systems through im-proper and unguided management experiments. Nor can weafford to waste time in reinventing ineffective strate-gies and not taking advantage of the knowledge gainedthrough considerable effort elsewhere. Continual andwidespread communication of research and managementfindings is mandatory.
Multiple Approaches and Persistence"There is no free lunch11 (B. Commoner) . This is
especially true for those of us in Hawaifi who are con-cerned about managing disturbed and deteriorating eco-systems. History reveals no panaceas for reducingalien animal populations and impacts. Biocontrol,chemical toxicants, hunting, fences, and habitat im-provement (for natives) are all useful tools, but allhave characteristic disadvantages and varying degreesof impermanence (e.g. Howarth 1983). We need to knowmore about the entire arsenal of management tools, be-cause different approaches are likely to be effectiveunder different conditions. For example, we are chang-ing feral goat and pig control methods in Hawai'i Vol-canoes National Park as population distribution anddensities and animal movements and wariness change.The ecological complexities touched upon above make theuse of varied combinations of damage reduction methodsnecessary if management is to be effective. The cur-rent approach to damage control, called Integrated PestManagement (IPM), while more generally applied to in-vertebrates, recognizes that flexible approaches andmultiple methods are usually needed to solve agricul-tural problems far less complicated than those we facein Hawaiian ecosystems (Huffaker 1975; Wilson andHuffaker 1976; Flint and van den Bosch 1981; Ruggieroand Johnston 1984).
Control efforts must be coordinated at research,management, and administrative levels. Realistic long-term goals must be set and receive continued support toattain objectives. Planning and prioritizing for thefuture by research, management and administration mustbe carefully done on the basis of facts at hand, andshould allow for the continued accumulation of betterinformation and adjusting priorities as we learn.Long-term planning and persistent coordinated efforts
285

TOWARD COOPERATIVE EFFORTS
"If you're not part of the solution, you're partof the problem" (Eldridge Cleaver/Bobby Seale). "Wehave met the enemy, and he is us11 (Pogo) . The problemof preserving and managing the remaining examples ofHawaifi!s native ecosystems, especially through reduc-tion and elimination of alien vertebrates, goes beyondwhat any one landowner, manager, or other special in-terest group (including conservationists) can accom-plish. A local and international network of conserva-tionists, developers or consumptive users, land usespecialists, natural scientists, businessmen, educa-tors, politicians, sociologists, economists, and otherexperts must be brought into the process of relatingnative ecosystems to community, national, and globalconcerns. The Nature Conservancy of Hawaifi has madeconsiderable progress in this direction, although theinitial focus was preservation and management of nativebird habitat (a valid indicator for an initial actionprogram), and the areas protected by The Conservancyare small and few, with budgets and staff to match(Holt and Fox, this volume). The State Natural AreaReserves System, now about 20 sites (P.Q. Tomich, pers.comm.), is also appropriate, but again, areas are smalland few, and active on-site management is needed.
If the problem of alien animals in Hawai'i1s exem-plary ecosystems is to be dealt with effectively, a co-operative approach should be implemented. We will needto cooperatively choose important and representativeecosystems upon which to concentrate limited coopera-tive resources. Our choices should be based on consid-erations of preserve design, protection, and manage-ment. We will need to decide which systems are mostintact, most in need of preservation, and least likelyto be influenced by aliens and other threats in thefuture. Increased communication among conservationistsand developers would help both groups to better planfor the future. Zones of cooperation and buffer zonesaround protected areas should be seriously consideredin planning (Gregg 1984). We will need to apply inten-sive, sustained, interdisciplinary research to protect-ed and used areas alike, to understand the ecosystemswe are dealing with in biological, socioeconomic, andother contexts. The ecology of aliens in systems man-aged for multiple use or sustained yield hunting de-serves further study in this regard. We will need toknow the consequences of periodic irruptions and de-clines of alien animals and the population biology ofnatives in protected and unprotected areas, and whatgoverns the dynamics. We will need to fully understand
286

the effects of our management actions on alien animalsand on Hawaifi*s ecosystems. We will need to explainthe rationale and impacts of alien control programs toa wide variety of interested parties for a long time,and invite them to participate in decisions that affectthem. We will have to seek a role in decisions made byothers that impinge upon our responsiblity of managingprotected areas through control of aliens to preventdegredation of ecosystems.
The tasks are enormously complicated, enduring,and time-consuming. Our knowledge of how to proceed,our progress in doing so, and even our communicationsabout the problems involved (except within scientific/management circles), are rudimentary. But progress isbeing made, and the end result, reducing the impacts ofaliens in Hawaii's irreplaceable ecosystems, is wellworth the effort.
ACKNOWLEDGEMENTS
I would like to thank C.W. Smith, L.L. Loope, J.M.Scott, P.K. Higashino, P.C. Banko, S. Mountainspring,J.D. Jacobi, R.L. Walker, F. Howarth, and P.Q. Tomichfor reviewing the manuscript and for their helpful sug-gestions. J.M. Scott, W.L. Wagner, P.C. Banko, andW.E. Banko generously allowed me to adapt material fromtheir unpublished papers elsewhere (including table 1and the figure) for this paper, and I am grateful. Ma-halo to S.M. Graves for drafting the figure. I am alsoindebted to D.B. Stone for encouragement, feedback, andtyping and updating many versions of the manuscript onthe microcomputer. I appreciate her patience, support,and judgement.
287

LITERATURE CITED
Atkinson, I.A.E. 1977. A reassessment of factors, par-ticularly Rattus rattus L., that influenced thedecline of endemic forest birds in the HawaiianIslands. Pac. Sci. 31(2):109-133.
Baker, J.K., and S. Allen. 1976. Studies on the endemicHawaiian genus Hibiscadelphus (hau-kuahiwi).Proc. 1st Hawaii Volcanoes Natl. Park Nat. Sci.Conf., 19-22. Honolulu: Univ. Hawaii.
Baldwin, P.H. 1947. Foods of the Hawaiian goose.Condor 49(3):108-120.
Baldwin, P.H., and T.L.C. Casey. 1983. A preliminarylist of foods of the poouli. Elepaio 43(7):53-56.
Baldwin, P.H., and G.O. Fagerlund. 1943. The effect ofcattle grazing on koa reproduction in HawaiiNational Park. Ecology 24(1):118-122.
Baldwin, P.H., C.W. Schwartz, and E.R. Schwartz. 1952.Life history and economic status of the mongoosein Hawaii. J. Mammal. 33(3):335-356.
Banko, P.C. 1982. Productivity of wild and captive nenepopulations. Proc. 4th Hawaii Volcanoes Natl.Park Nat. Sci. Conf., 12-32. Honolulu: Univ.Hawaii.
Banko, P.C., and D.A. Manuwal. 1982. Life history,ecology, and management of nene £Brant.a sandvi-censisjL in Hawaii Volcanoes and Haleakala NationalParks. Final Rep., Natl. Park Serv. ContractCX-8000-8-0005. Seattle: Univ. Wash. Coop. Natl.Park Resour. Stud. Unit.
Banko, W.E. 1978. Some limiting factors and researchneeds of endangered Hawaiian forest birds. Proc.2nd Hawaii Volcanoes Natl. Park Nat. Sci. Conf.,17-25. Honolulu: Univ. Hawaii.
Banko, W.E., and P.C. Banko. 1976. Role of fooddepletion by foreign organisms in historicaldecline of Hawaiian forest birds. Proc. 1stHawaii Volcanoes Natl. Park Nat. Sci. Conf.,29-34. Honolulu: Univ. Hawaii.
Banko, W.E., and P.C. Banko. n.d. Decline and extinc-tion of endemic Hawaiian avifauna: role of fooddepletion and competition by foreign organisms.U.S. Fish Wildl. Serv., Hawaii Volcanoes Natl.Park. Typescript.
288

Barrett, R.H., III, and C.P. Stone. 1983. Hunting as acontrol method for wild pigs in Hawaii VolcanoesNational Park: a report for Resource Management[Division]. Hawaii Volcanoes Natl. Park.Typescript.
Berger, A.J. 1974. History of exotic birds in Hawaii:first of two installments. Elepaio 35(6):60-65.
Berger, A.J. 1975. History of exotic birds in Hawaii:second and final installment. Elepaio 35(7):72-80.
Berger, A.J. 1981. Hawaiian birdlife. 2nd ed. Honolulu:Univ. Pr. Hawaii.
Burr, T.A. 1984. Introduced birds in Hawaii and someassociated State management problems. Trans. 24thAnn. Hawaii For. Wildl. Conf., 92-97. Honolulu.
Burr, T., P.Q. Tomich, E. Kosaka, W. Kramer, J.M.Scott, E. Kridler, J. Giffin, D. Woodside, and R.Bachman. 1982. Alala recovery plan. U.S. FishWildl. Serv., Portland, Ore.
Casey, T.L.C., S. Mountainspring, and J.M. Scott. Inprep. Behavioral ecology and limiting factors ofthe poouli. U.S. Fish Wildl. Serv.
Conant, S. 1976. Bird distribution and abundance above3000 feet in Hawaii Volcanoes National Park.Proc. 1st Hawaii Volcanoes Natl. Park Nat. Sci.Conf., 63-74. Honolulu: Univ. Hawaii.
Conant, S. 1981. Birds. (Altitudinal distribution oforganisms along an island mountain transect). InIsland ecosystems; biological organization inselected Hawaiian communities, ed. D. Mueller-Dombois, R.W. Bridges, and H.L. Carson., 97-105.Stroudsburg, Penn.: Hutchinson Ross Pub. Co.
289

290

Hawaii Division of Forestry and Wildlife. 1983. Hawaiiwildlife plan. Honolulu: Hawaii Dep. Land Nat.Resour.
Higashino, P.K., and C.P. Stone. 1982. The fern jungleexclosure in Hawaii Volcanoes National Park: 13years without feral pigs in a rain forest.[Abstract] Proc. 4th Hawaii Volcanoes Natl. ParkNat. Sci. Conf., 86. Honolulu: Univ. Hawaii.
Holt, R.A., and B. Fox. Protection status of thenative Hawaiian biota. [This volume]
Hood, G.A. 1978. Vertebrate control chemicals: currentstatus of registrations, rebuttable presumptionsagainst registrations, and effects on users.Proc. 8th Vert. Pest Control Conf., 170-176.
Howarth, F.G. 1983. Classical biocontrol: panacea orPandora!s box. Proc. Hawaii. Entomol. Soc. 24(2&3):239-244.
Howarth, F.G. Impacts of alien land arthropods andmollusks on native plants and animals in Hawaii.[This volume]
Huffaker, C.B. 1975. Biological control in themanagement of pests. Agro-Ecosystems 2:15-31.
Imber, M.J. 1978. The effects of rats on breeding suc-cess of petrels. In The ecology and control ofrodents in New Zealand nature reserves, ed. P.R.Dingwall, I.A.E. Atkinson, and C. Hay, 67-72. NewZealand Dep. Lands Surv. Info. Ser. 4.Wellington.
Jacobi, J.D. 1976. The influence of feral pigs on anative alpine grassland in Haleakala NationalPark. Proc. 1st Hawaii Volcanoes Natl. Park Nat.Sci. Conf., 107-112. Honolulu: Univ. Hawaii.
Jacobi, J.D. 1980. Changes in a native grassland inHaleakala National Park following disturbance byferal pigs. Proc. 2nd Conf. Sci. Res. Natl.Parks. Vol. 8, 294-308. San Francisco, Calif.:Natl. Park Serv.
Kami, H.T. 1966. Foods of rodents in the HonokaaDistrict, Hawaii. Pac. Sci. 20(3):367-373.
Katahira, L. 1980. The effects of feral pigs on a mon-tane rain forest in Hawaii Volcanoes NationalPark. Proc. 3rd Hawaii Volcanoes Natl. Park Nat.Sci. Conf., 173-178. Honolulu: Univ. Hawaii.
291

292

Loope, L.L., and P.G. Scowcroft. Vegetation responsewithin exclosures in Hawaii: a review. [Thisvolume]
McEldowney, H. 1979. Archaeological and historicalliterature search and research design. Lava flowcontrol study, Hilo, Hawaii. Prep. U.S. ArmyCorp. Engineers, Contract DACW84-77-C-0019. B.P.Bishop Mus. Anthropol. Dep., Honolulu.Typescript.
McEldowney, H. 1983. A description of major vegetationpatterns in the Waimea-Kawaihae region during theearly historic period. Rep. 16. In Archaeologicalinvestigations of the Mudlane-Waimea-Kawaihae roadcorridor, island of Hawaii; an interdisciplinarystudy of an environmental transect, ed. J.T. Clarkand P.V. Kirch, 407-448. B.P. Bishop Museum Dep.Anthropol. Rep. Ser. 83-1.
McNab, J. 1983. Wildlife management as scientificexperimentation. Wildl. Soc. Bull. 11(4):397-401.
Moulton, M.P., and S.L. Pimm. 1983. The introducedHawaiian avifauna: biogeographic evidence for com-petition. Am. Nat. 121(5):669-690.
Mountainspring, S., and J.M. Scott. In press. Inter-specific competition among Hawaiian forest birds.Ecol. Monogr..
Mueller-Dombois, D. 1981. Vegetation dynamics in acoastal grassland of Hawaii. Vegetatio46:131-140.
293

Simons, T.R. 1983. Biology and conservation of theendangered Hawaiian dark-rumped petrel (Pterodromaphaeopygia sandwichensis). Univ. Wash. Coop.Natl. Park Resour. Stud. Unit Tech. Rep. 83-2.Seattle: Univ. Wash.
294

Singer, F.J. 1981. Wild pig populations in the NationalParks. Environ. Manage. 51(3):263-270.
Smith, C.W. Impact of alien plants on Hawaii's nativebiota. [This volume]
Smith, J.D., and J.R. Woodworth. 1951. A study of thepheasant, California quail, and lace-necked dovein Hawaii. Hawaii Fish Game Div. Spec. Bull. 3.
Spatz, G., and D. Mueller-Dombois. 1973. The influenceof feral goats on koa tree reproduction in HawaiiVolcanoes National Park. Ecology 54(4):870-876.
Spatz, G., and D. Mueller-Dombois. 1975. Successionpatterns after pig diggings in grasslandcommunities on Mauna Loa, Hawaii. Phvtocoenologia3(2/3):346-373.
Stone, C.P. 1984. National Park Service research andmanagement to control introduced animals inHawaii. Trans. 24th Ann. For. Wildl. Conf.,68-80. Honolulu.
Stone, C.P., and D.D. Taylor. 1984. Status of feralpig management and research in Hawaii VolcanoesNational Park. Proc. 5th Hawaii Volcanoes Natl.Park Nat. Sci. Conf.. 106-117. Honolulu: Univ.Hawaii.
Stone, C.P., P.C. Banko, P.K. Higashino, and F.G.Howarth. 1984. Interrelationships of alien andnative plants and animals in Kipahulu Valley,Haleakala National Park: a preliminary report.Proc. 5th Hawaii Volcanoes Natl. Park Nat. Sci.Conf.. 91-105. Honolulu: Univ. Hawaii.
Stone, C.P., R.L. Walker, J.M. Scott, and P.C. Banko.1983. Hawaiian goose management and research:where do we go from here? Elepaio 44(2):11-15.
Tomich, P.Q. 1969. Mammals in Hawaii; a synopsis andnotational bibliography. Honolulu: Bishop Mus. Pr.
Tomich, P.Q. 1981. Rodents. (Altitudinal distributionof organisms along an island mountain transect).In Island ecosystems; Biological organization inselected Hawaiian communities. ed. D. Mueller-Dombois, R.W. Bridges, and H.L. Carson, 105-110.Stroudsburg, Penn.: Hutchinson Ross Pub. Co.
Tuggle, H.D. 1979. Hawaii. In The prehistory of Poly-nesia, ed. J.D. Jennings, 167-199. Cambridge,Mass.: Harvard Univ. Pr.
295

296

Williams, J. 1980. Native vs exotic woody vegetationrecovery following goat removal in the easterncoastal lowlands on Hawaii Volcanoes NationalPark. Proc. 3rd Hawaii Volcanoes Natl. Park Nat.Sci. Conf.. 373-382. Honolulu: Univ. Hawaii.
Williams, R.N. 1983a. Bulbul introductions on Oahu.Elepaio 43(11):89-90.
Williams, R.N. 1983b. The red-vented bulbul on theisland of Hawaii. Elepaio 43(12):101-103.
Wilson, F., and C.B. Huffaker. 1976. The philosophy,scope and importance of biological control. InTheory and practice of biological control, ed.C.B. Huffaker and P.S. Messenger, 3-15. Inter-natl. Organ. Biol. Control, Internatl. Ctr. Biol.Control. New York: Academic Pr.
Woodworth, J.R., and D.H. Woodside. 1953. Mongoosepoison experiment. Terr. Hawaii Pittman-RobertsonJob Complet. Rep. Proj. 5-R-4, Work Plan 4, Job 29(4), 1-16. Typescript.
297