amelioration of neurological and biochemical deficits by peroxynitrite decomposition catalysts in...
TRANSCRIPT

European Journal of Pharmacology 596 (2008) 77–83
Contents lists available at ScienceDirect
European Journal of Pharmacology
j ourna l homepage: www.e lsev ie r.com/ locate /e jphar
Amelioration of neurological and biochemical deficits by peroxynitritedecomposition catalysts in experimental diabetic neuropathy
Manish Arora, Ashutosh Kumar, Ravinder K. Kaundal, Shyam S. Sharma ⁎Molecular Neuropharmacology Laboratory, Department of Pharmacology and Toxicology, National Institute of Pharmaceutical Education and Research (NIPER),Sector-67, S.A.S. Nagar, Mohali, Punjab 160062, India
⁎ Corresponding author. Tel.: +91 172 2214683 87; faxE-mail address: [email protected] (S.S. Sharma).
0014-2999/$ – see front matter © 2008 Elsevier B.V. Aldoi:10.1016/j.ejphar.2008.08.003
a b s t r a c t
a r t i c l e i n f oArticle history:
Diabetic neuropathy, a majo Received 10 April 2008Received in revised form 31 July 2008Accepted 7 August 2008Available online 16 August 2008Keywords:Diabetic neuropathyStreptozotocinPeroxynitriteOxidative stressPARPNerve blood flowMotor nerve conduction velocityNociceptionAllodynia
r complication of diabetes, affects more than 60% of diabetic patients. Recently,involvement of peroxynitrite has been postulated in diabetic neuropathy. In the present study, we have studiedthe effects of peroxynitrite decomposition catalysts (PDC's)—5,10,15,20-tetrakis(4-sulfonatophenyl) porphyr-inato iron(III) [FeTPPS] and 5,10,15,20-tetrakis(N-methyl-4-pyridyl)porphyrinato iron(III) [FeTMPyP]—in experimental diabetic neuropathy. Male Sprague–Dawley rats, with six weeks of untreated diabetes weretreated for two weeks with peroxynitrite decomposition catalysts. Diabetic animals showed a significantdecrease in motor nerve conduction velocity and nerve blood flow, nociception as evident from decreasedtail flick latency (hyperalgesia) and increased paw withdrawal pressure (mechanical allodynia) alongwith elevation in peroxynitrite and reduction in nerve glutathione levels. Two weeks treatment withPDC's significantly improved all the above stated functional and biochemical deficits. Aftermath of thisstudy advocates the beneficial effects of peroxynitrite decomposition catalysts in experimental diabeticneuropathy.
© 2008 Elsevier B.V. All rights reserved.
1. Introduction
Diabetic neuropathy is a heterogeneous disorder whose enigmaticetiology is characterized by a number of interwoven pathwayssubsuming increased flux of glucose to polyol pathway, increasedhexosamine shunt, aldose reductase activation, decrease in nervemyoinositol content, formation of advanced glycation end products(AGE), impaired neurotrophic support, activation of protein kinase C(PKC), activation of poly (ADP-ribose) polymerase (PARP), impairedinsulin/C peptide action etc (Evans et al., 2002; Obrosova et al., 2005a).All these factors result in the formation of a variety of reactive oxygenspecies (ROS) which are the ultimate malefactor involved in most ofthe diabetic complications (Pop-Busui et al., 2002; Sima, 2006).Amongst this class of species, the notorious ones are superoxide (O2
−·),hydrogen peroxide (H2O2), peroxynitrite (ONOO−) and hydroxylradical (OH−). Superoxide though performs some of the house keepingfunctions and is an audacious defense against microbial invasion, butunder pathological conditions, unfettered production of nitric oxide(Fujimoto et al., 2004) coupled with deficient body's antioxidantdefense results in reaction of nitric oxide with superoxide to formperoxynitrite which is several times multiple of its parents (super-oxide and nitric oxide) in terms of tissue toxicity (Patel et al., 1999).
: +91 172 2214692.
l rights reserved.
Peroxynitrite can cause hydroxylation and nitration of the aromaticresidues of proteins and nucleotides (Stamler, 1994). Peroxynitrite isalso responsible for the abrasion of the body's antioxidant defense byinactivation of enzyme superoxide dismutase (Bartosz, 1996; Szabo,2003). Generation of superoxide and peroxynitrite impairs vascularfunction in diabetic rats, which precedes the slowing of nerveconduction velocity (Coppey et al., 2002). In addition to these effects,peroxynitrite induced DNA damage leads to over-activation of PARP, anuclear enzyme involved in DNA repair (Szabo, 2005). Ramificationsof PARP over-activation include energy depletion, failure of membraneassociated ATP dependent ion pumps and disruption of membranepotential. The deleterious effects of peroxynitrite have been reportedin a number of inflammatory conditions as well as in cardiovascularand central nervous system disorders (Cuzzocrea et al., 2006; Mableyet al., 2002). Peroxynitrite is also involved in the pathogenesis ofcomplications associated with diabetes which includes diabeticneuropathy, vasculopathy, retinopathy, and nephropathy (Obrosovaet al., 2005b).
Peroxynitrite decomposition catalysts act by promoting thecleavage of peroxynitrite to nitrate thereby reducing the levels ofnotorious reactive intermediates such as nitrogen dioxide andhydroxyl radicals (Bartosz, 1996; Patel et al., 1999). Neuroprotectivepotential of peroxynitrite decomposition catalysts in cerebral ischemicreperfusion injury has been demonstrated earlier (Sharma et al.,2004; Thiyagarajan et al., 2004). In the present study we have inves-tigated the neuroprotective potential of peroxynitrite decomposition

78 M. Arora et al. / European Journal of Pharmacology 596 (2008) 77–83
catalysts (FeTPPS and FeTMPyP) in experimental diabetic neuropathyby assessing various functional, behavioral, biochemical and immu-nohistological parameters.
2. Materials and methods
2.1. Reagents
FeTPPS and FeTMPyP were purchased from Calbiochem, Germanyand dissolved in saline. Streptozotocin (STZ) was procured fromSigma-Aldrich, U.S.A and dissolved in citrate buffer (pH 4.4). Glucoseoxidase–peroxidase (GOD–POD) kit was purchased from Accurex,India. All other chemicals of analytical grade were purchased locally.
2.2. Animals
Healthy male Sprague–Dawley rats (250–270 g) were obtainedfrom the Central Animal Facility (CAF), National Institute of Pharma-ceutical Education and Research (NIPER), Punjab, India. Animals wereprovided with standard diet and water ad libitum. They were housedin plastic cages (two in each) at a controlled temperature 22±2 °C andhumidity 55±5%, with 12 hour light and dark cycle. All the experi-mental protocols were approved by the Institutional Animal EthicsCommittee of NIPER.
2.3. Induction of diabetes
Diabetes was induced using a single dose of streptozotocin (55 mg/kg, i.p.). Age matched control rats received an equal volume of vehicle(citrate buffer). Diabetes was confirmed after 48 h of streptozotocininjection by estimating plasma glucose levels using glucose oxidase–peroxidase (GOD–POD) and the rats with plasma glucose levelN250 mg/dl were used for further studies.
2.4. Treatment schedule
Diabetic neuropathy was well developed after six weeks ofstreptozotocin treatment as reported earlier (Sayyed et al., 2006).The twoweeks treatment with FeTPPS (1, 3 and 10mg/kg/day, i.p.) andFeTMPyP (1 and 3 mg/kg/day, i.p.) was started after the sixth week ofdiabetes induction and continued till the eighth week. All theparameters (motor nerve conduction velocity, nerve blood flow,nociception and biochemical parameters) were measured 24 h afterthe last dose. The number of animals used for motor nerve conductionvelocity, nerve blood flow and thermal hyperalgesia was 6–8 and forbiochemical (lipid peroxidation, peroxynitrite and glutathione levels)estimations 3–6.
2.5. Motor nerve conduction velocity
Power Lab 8sp instrument (AD Instruments, Australia) was usedfor the measurement of motor nerve conduction velocity. Briefly theanimals were anesthetized by 4% halothane in a mixture of nitrousoxide and oxygen and anesthesia was maintained with 1% halothane,using gaseous anesthesia system (Harvard apparatus, UK). Motornerve conduction velocity was measured by stimulating the sciatic(proximal to sciatic notch) and tibial (distally to ankle) nerve usingbipolar needle (261/2 gauge) electrodes with 3 volt, single stimulus asdescribed previously (Kumar et al., 2007). Body temperature of ratswas maintained at 37 °C using homoeothermic blanket system(Harvard, UK) throughout the experiment. Motor nerve conductionvelocity was calculated by following formula:
Motor nerve conduction velocity m=sð Þ¼ Distance between sciatic and tibial nerve stimulation pointð Þ
= Sciatic M wave latency−Tibial M wave latencyð Þ:
2.6. Composite nerve blood flow
Nerve blood flow was measured using Laser Doppler flowmeter(Perimed, Sweden) as described earlier by Kumar et al. (2007). TheLaser Doppler has been compared to other methods and has beenreported to be equivalent to other available methods like hydrogenclearance (Takeuchi and Low, 1987) and [14C] iodo antipyrine method(Rundquist et al., 1985) of blood flow measurement. Briefly, theanimals were anesthetized by 4% halothane in a mixture of nitrousoxide and oxygen and anesthesia was maintained with 1% halothane,using gaseous anesthesia system (Harvard apparatus, UK). Anesthe-tized animals were placed on stereotaxic equipment to achieveuniform positioning of Laser Doppler probe each time. Sciatic nerve ofthe left flank was exposed and laser probe was placed just above thenerve. Nerve blood flow was recorded for 10 min after 15 min ofstabilization period. Averages of 10 min continuous recordings wererepresented as arbitrary perfusion units (PU). During the recording ofnerve blood flow, body temperature of the rats was maintained at37 °C using homoeothermic blanket system (Harvard apparatus, UK).
2.7. Thermal hyperalgesia
Hyperalgesia was assessed using tail immersion test. Rats weretrained for three days prior to final measurement. The rat tail wasimmersed in warm (45 °C) water and the tail flick response latency(withdrawal response of tail) or any signs of strugglewere observed asthe end point response. Cut-off time was kept at 15 s.
2.8. Mechanical allodynia
To assess the mechanical allodynia, the Electrovonfrey Anaesthe-siometer was used (IITC, Woodland Hills CA). The animals were kepton a perforated platform and vonfrey hairs were applied to theunderside of their paw. The pressure (g) at which the rat withdraws itsright paw was recorded (Sweitzer et al., 2004). Five consecutivereadings were taken at an interval of 5 min.
2.9. Peroxynitrite estimation
Peroxynitrite formation was estimated by using a fluorescent dyedihydrorhodamine123 (Szabo et al., 1995; Thiyagarajan et al., 2004).Dihydrorhodamine123 gets oxidized to rhodamine123 in a peroxyni-trite dependent manner. Rats were injected dihydrorhodamine123through the femoral vein (2×10−6 M/ml/kg in saline). After 2 h ofinjection, blood was withdrawn and fluorescence in the plasma wasmeasured using the spectrofluorometer at an excitationwavelength of500 nm and emission wavelength of 536 nm. The plasma level ofrhodamine123 was calculated from the standard curve obtained fromauthentic rhodamine123 in a concentration of 0–10 nM prepared inplasma obtained from the untreated rat.
2.10. Glutathione estimation
Sciatic nerve homogenate was used for measuring reducedglutathione (GSH) content as described by Moron et al. (1979) withsome modification. Nerve was homogenized in phosphate buffersaline (pH 7.4). An aliquot (0.1 ml) of homogenate was mixed with 10%sulphosalicylic acid (0.5 ml) and kept in ice for 20 min. Then theresulting solutionwas centrifuged at 10,000 g for 5min at 4 °C, 50 μl ofthe supernatant was mixed with 450 μl of phosphate buffer and 1.5 mlof 5,5′-dithiobis (2-nitro benzoic acid) in 0.1 M phosphate buffer, (pH8.0), incubated for 10 min at 37 °C followed by measurement ofabsorbance at 412 nm spectrophotometrically, using reduced glu-tathione as an external standard (Moron et al., 1979; Tietze, 1969).Protein content in tissue homogenate supernatant was determined bythe Lowry method (Lowry et al., 1951).

Fig. 1. Effect of the two weeks treatment with FeTPPS and FeTMPyP on motornerve conduction velocity measured in eight week diabetic rats with or withouttreatment. ⁎⁎⁎Pb0.001 vs agematched control rats, ##Pb0.01 vs vehicle treated diabeticrats, n=6–8.
79M. Arora et al. / European Journal of Pharmacology 596 (2008) 77–83
2.11. Poly (ADP-ribose) immunohistochemistry
In situ fixed sciatic nerves were kept in 30% sucrose buffer (0.2 Mphosphate buffer pH 7.2) overnight and 10 μm thin sections wereobtained using Cryotome (Thermo Shandon, Europe). The sectionswere washed with tris buffered saline (TBS,pH 7.4) followed byincubation with 20 μg/ml of proteinase K for 20 min for antigenretrieval. After washing with TBS, the sections were incubated withblocking buffer (5% normal goat serum in PBS) for 2 h. Endogenousbiotin binding sites were blocked by sequential incubation of avidinand biotin for 30 min each. The sections were then incubated withprimary antibodies (anti-PAR mouse IgG from Alexis Biochemicals,USA), in blocking buffer at 4 °C overnight. The sections were washedthree times with PBS and then incubated with biotin conjugatedsecondary antibodies (anti-mouse IgG—Sigma-Aldrich Inc. USA) inblocking buffer for 2 h. The specific labeling was detected by avidinconjugated fluorescein isothiocyanate (FITC). The sections weremounted in 4,6,-Di Amindino-2-Phenyl Indole (DAPI) containingmounting media and were observed under fluorescent microscope(Leica, Germany) and images were acquired using a CCD camera (LeicaDC300F) (Sharma et al., 2008).
2.12. Nitrotyrosine immunohistochemistry
Nerve cryosections were incubated with anti-nitro tyrosineprimary antibodies, in blocking buffer at 4°C overnight after quench-ing the endogenous peroxidase with H2O2. Nitrotyrosine positive cellson nerve sections were detected using vecta stain ABC kit (VectorLaboratories). Nerve sections were then counterstained withhaematoxylin.
2.13. Statistical analysis
Results are expressed as mean±S.E.M. Jandel Sigma Stat Version 2software was used for statistical analysis. Significance of differencebetween the two groups was evaluated using Student's t-test. For themultiple comparisons, one way analysis of variance (ANOVA) wasused. When ANOVA showed significant difference, post hoc analysiswas performed with Tukey's test. Pb0.05 was considered statisticallysignificant. Individual between comparisons were made using theStudent–Newman–Keuls multiple range test.
3. Results
3.1. Plasma glucose level and body weight
After streptozotocin administration, the animals showed 4–5 foldincrease in plasma glucose levels as compared to the age matchedcontrol rats (vehicle treated). Treatment with FeTPPS (1, 3 and 10 mg/kg) and FeTMPyP (1 mg/kg and 3 mg/kg) did not show effect on bloodglucose levels. Progressive weight loss was also observed in thediabetic animals which were not reversed by any of the treatment(Table 1).
Table 1Effect of two weeks treatment with FeTPPS and FeTMPyP on body weight and plasmaglucose levels
Groups Body weight (g) Plasma glucose level (mg/dl)
Control 432±5 110±5Diabetic control 210±12a 430±8a
Diabetic+FP 1 mg/kg 218±10 421±7Diabetic+FP 3 mg/kg 220±8 410±12Diabetic+FP 10 mg/kg 216±11 398±10Diabetic+FM 1 mg/kg 224±10 416±12Diabetic+FM 3 mg/kg 219±7 426±8
Results are expressed as mean±S.E.M.a Pb0.001 vs age matched control rats.
3.2. Motor nerve conduction velocity
Motor nerve conduction velocity was reduced significantly in eightweek diabetic rats when compared to age matched control rats.FeTMPyP (3 mg/kg) and FeTPPS (10 mg/kg) treatment showedsignificant reversal. Other doses fail to provide any reversal of motornerve conduction velocity (Fig. 1).
3.3. Sciatic nerve blood flow
In eight weeks old diabetic rats, a significant reduction incomposite nerve blood flow was observed as compared to agematched control rats. Treatment with peroxynitrite decompositioncatalysts showed a significant improvement in nerve blood flow(Fig. 2).
3.4. Nociception: hot immersion performance
When subjected to the hot immersion performance test aftereight weeks of streptozotocin treatment, diabetic rats exhibited asignificant reduction in tail flick latency as compared to the controlrats. Treatment with FeTPPS (1 and 3 mg/kg) and FeTMPyP (1 mg/kg)did not reversed thermal hyperalgesia significantly, while FeTMPyP(3 mg/kg) and FeTPPS (10 mg/kg) have shown a significant protectiveeffect (Fig. 3).
Fig. 2. Effect of the twoweeks treatment with FeTPPS and FeTMPyP on composite nerveblood flowmeasured in eight week diabetic rats with or without treatment ⁎⁎⁎Pb0.001vs age matched control rats, ##Pb0.01, ###Pb0.001 vs vehicle treated diabetic rats,n=6–8.
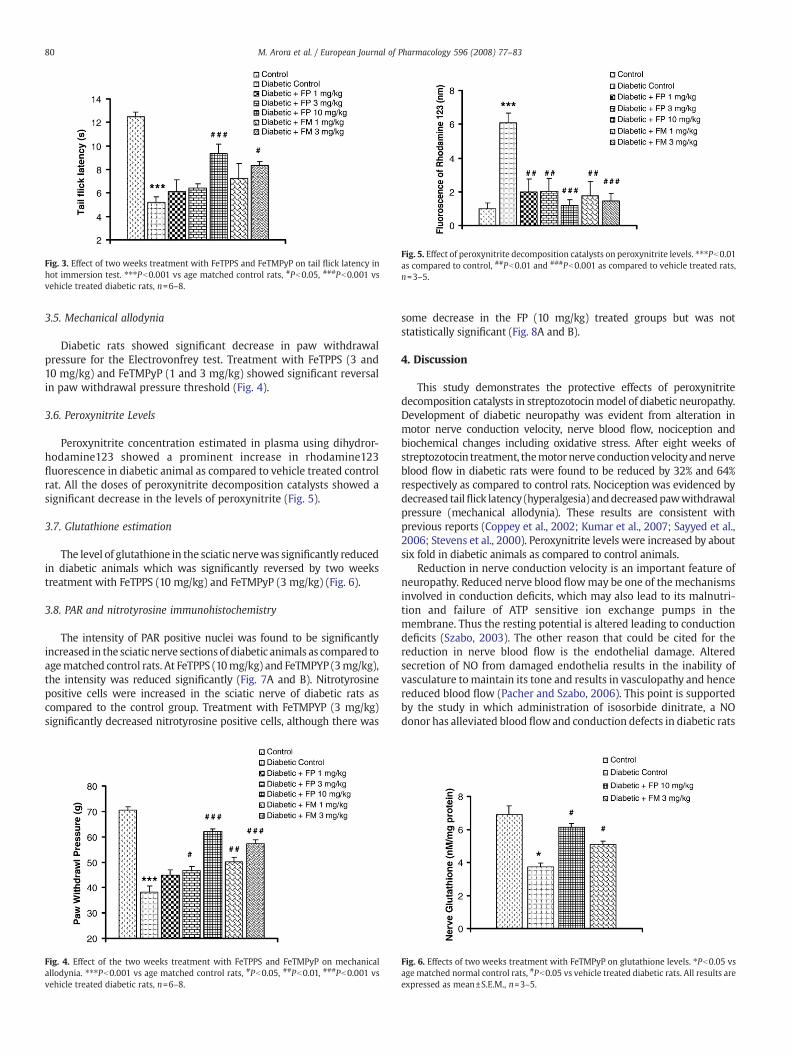
Fig. 5. Effect of peroxynitrite decomposition catalysts on peroxynitrite levels. ⁎⁎⁎Pb0.01as compared to control, ##Pb0.01 and ###Pb0.001 as compared to vehicle treated rats,n=3–5.
Fig. 3. Effect of two weeks treatment with FeTPPS and FeTMPyP on tail flick latency inhot immersion test. ⁎⁎⁎Pb0.001 vs age matched control rats, #Pb0.05, ###Pb0.001 vsvehicle treated diabetic rats, n=6–8.
80 M. Arora et al. / European Journal of Pharmacology 596 (2008) 77–83
3.5. Mechanical allodynia
Diabetic rats showed significant decrease in paw withdrawalpressure for the Electrovonfrey test. Treatment with FeTPPS (3 and10 mg/kg) and FeTMPyP (1 and 3 mg/kg) showed significant reversalin paw withdrawal pressure threshold (Fig. 4).
3.6. Peroxynitrite Levels
Peroxynitrite concentration estimated in plasma using dihydror-hodamine123 showed a prominent increase in rhodamine123fluorescence in diabetic animal as compared to vehicle treated controlrat. All the doses of peroxynitrite decomposition catalysts showed asignificant decrease in the levels of peroxynitrite (Fig. 5).
3.7. Glutathione estimation
The level of glutathione in the sciatic nervewas significantly reducedin diabetic animals which was significantly reversed by two weekstreatment with FeTPPS (10 mg/kg) and FeTMPyP (3 mg/kg) (Fig. 6).
3.8. PAR and nitrotyrosine immunohistochemistry
The intensity of PAR positive nuclei was found to be significantlyincreased in the sciatic nerve sectionsof diabetic animals as compared toagematched control rats. At FeTPPS (10mg/kg) and FeTMPYP (3mg/kg),the intensity was reduced significantly (Fig. 7A and B). Nitrotyrosinepositive cells were increased in the sciatic nerve of diabetic rats ascompared to the control group. Treatment with FeTMPYP (3 mg/kg)significantly decreased nitrotyrosine positive cells, although there was
Fig. 4. Effect of the two weeks treatment with FeTPPS and FeTMPyP on mechanicalallodynia. ⁎⁎⁎Pb0.001 vs age matched control rats, #Pb0.05, ##Pb0.01, ###Pb0.001 vsvehicle treated diabetic rats, n=6–8.
some decrease in the FP (10 mg/kg) treated groups but was notstatistically significant (Fig. 8A and B).
4. Discussion
This study demonstrates the protective effects of peroxynitritedecomposition catalysts in streptozotocinmodel of diabetic neuropathy.Development of diabetic neuropathy was evident from alteration inmotor nerve conduction velocity, nerve blood flow, nociception andbiochemical changes including oxidative stress. After eight weeks ofstreptozotocin treatment, themotornerve conductionvelocityandnerveblood flow in diabetic rats were found to be reduced by 32% and 64%respectively as compared to control rats. Nociception was evidenced bydecreased tailflick latency (hyperalgesia) anddecreasedpawwithdrawalpressure (mechanical allodynia). These results are consistent withprevious reports (Coppey et al., 2002; Kumar et al., 2007; Sayyed et al.,2006; Stevens et al., 2000). Peroxynitrite levels were increased by aboutsix fold in diabetic animals as compared to control animals.
Reduction in nerve conduction velocity is an important feature ofneuropathy. Reduced nerve blood flowmay be one of the mechanismsinvolved in conduction deficits, which may also lead to its malnutri-tion and failure of ATP sensitive ion exchange pumps in themembrane. Thus the resting potential is altered leading to conductiondeficits (Szabo, 2003). The other reason that could be cited for thereduction in nerve blood flow is the endothelial damage. Alteredsecretion of NO from damaged endothelia results in the inability ofvasculature to maintain its tone and results in vasculopathy and hencereduced blood flow (Pacher and Szabo, 2006). This point is supportedby the study in which administration of isosorbide dinitrate, a NOdonor has alleviated blood flowand conduction defects in diabetic rats
Fig. 6. Effects of two weeks treatment with FeTMPyP on glutathione levels. ⁎Pb0.05 vsage matched normal control rats, #Pb0.05 vs vehicle treated diabetic rats. All results areexpressed as mean±S.E.M., n=3–5.

Fig. 7. A: Effect of two weeks treatment with peroxynitrite decomposition catalysts on PAR immunoreactivity. Upper panel shows PAR immunopositive cells and lower panel showsthe total cell population. Bar shows a length of 50 μm. B: Effects of two weeks treatment with FeTPPS and FeTMPyP on nerve PAR immunopositivity. ⁎⁎⁎Pb0.001 vs age matchednormal control rats, ##Pb0.01 vs vehicle treated diabetic rats.
81M. Arora et al. / European Journal of Pharmacology 596 (2008) 77–83
(Cameron and Cotter, 1995). In our study, we have observed asignificant improvement in nerve blood flow and conduction deficitsafter 2 weeks of peroxynitrite decomposition catalysts treatment.
We have estimated peroxynitrite levels by using dihydrorhoda-mine oxidation method flourimetrically. Oxidation of dihydrorhoda-mine to fluorescent rhodamine is a marker of cellular oxidantproduction. Studies reveal that this method can be efficiently usedas a probe for peroxynitrite production (Kooy et al., 1994). Peroxyni-trite decomposition catalysts reduced the levels of peroxynitrite inplasma of diabetic rats. Treatment with FeTPPS 10 mg/kg andFeTMPyP 3 mg/kg also restored the intracellular glutathione levelsto a significant level. Attenuating elevated peroxynitrite levels andrestoring depleted glutathione levels hence decreasing oxidativestress may be one of the probable reasons for neuroprotectivebeneficiaries provided by peroxynitrite decomposition catalysts.
Neuropathic pain is a common symptom of diabetic neuropathy;hence we studied the effect of peroxynitrite decomposition catalystson sensori-motor alterations in experimental diabetic rats. FeTMPyPshowed significant improvement of mechanical allodynia and hyper-algesia. However, FeTPPS did not show any effect at lower dose levels.Multiple mechanisms are known to be involved in the etiology ofdiabetic neuropathic pain. The role of COX has beenwell established inpain and peroxynitrite is known tomodulate the activation of COX andvarious other inflammatory mediators (Fujimoto et al., 2004). COX-2inhibition has been shown to reduce the neuropathic pain indicatinginflammation as a major contributor (Freshwater et al., 2002). Theanalgesic action of peroxynitrite decomposition catalysts could beattributed to their various actions such as direct effect on peroxynitritelevels (Misko et al., 1998), indirectly reducing COX activation (Nedelec
et al., 2001) and anti-inflammatory activity as shown by some inves-tigators (Misko et al., 1998; Salvemini et al., 1998).
Due to its higher lipid permeability, peroxynitrite easily permeatesthe cell membrane (Marla et al., 1997) and then nuclear membranewhere it induces DNA damage. Extensive DNA damage leads to PARPactivation and cellular depletion of NAD+ (Kiss and Szabo, 2005; Patelet al., 1999). The amount of PAR is minimal in the normal cells and itslevel increases extensively during PARP over-activation. Hence, nerveimmunohistochemistry for PAR was studied to reveal the effect ofperoxynitrite decomposition catalysts on its over-activation. Nervesections from peroxynitrite decomposition catalysts treated ratsshowed reduced immunopositivity towards PAR when compared tocontrol animals. Since poly ADP-ribosylation of various transcriptionalfactors leads to expression of various inflammatory mediators in-volved in pain perception, the protection afforded by peroxynitritedecomposition catalysts towards mechanical allodynia and thermalhyperalgesia could be attributed to inhibition of PARP over-activation.Thus it can be concluded that nitrosative stress is a major pathway inPARP activation and peroxynitrite decomposition catalysts affordedprotection by inhibiting the over-activation of PARP.
Peroxynitrite ill effects are also manifested by nitrotyrosine (NT)accumulation in the sciatic nerve which is one of the important factorscontributing to nerve dysfunction in experimental diabetic neuropathy.To access the effect of peroxynitrite decomposition catalysts weinvestigated nitrotyrosine levels in sciatic nerve.We found a substantialincrease in nitrotyrosine levels in nerve microsections of diabeticanimals as compared to control animals. Treatmentwith FeTMPyP3mg/kg decreased nitrotyrosine levels in nerve microsections of treatedanimals. The beneficial role of peroxynitrite decomposition catalysts in

Fig. 8. A: Effect of two weeks treatment with peroxynitrite decomposition catalysts onnitrotyrosine levels. Representative micrographs showing nitrotyrosine positive cells insciatic nerve of control, diabetic control and treated animals. Micron bar shows a lengthof 50 μm. B: Effects of two weeks treatment with FeTPPS and FeTMPyP on nervenitrotyrosine immunopositivity. ⁎⁎⁎Pb0.001 vs age matched normal control rats,##Pb0.01 vs vehicle treated diabetic rats.
82 M. Arora et al. / European Journal of Pharmacology 596 (2008) 77–83
treated animals indicates the role of peroxynitrite in impaired nerves inanimals with experimental diabetic neuropathy.
The results from the present study indicate that peroxynitrite doplay a major role in the pathogenesis of diabetic neuropathy andperoxynitrite decomposition catalysts can be promising agents in thefuture to counteract the progression of neuropathy and neuropathicpain. Since peroxynitrite is also involved in the destruction of isletcells of pancreas, counteracting nitrosative stress during early stagesof diabetes could prevent development of hyperglycemia itself andthus hyperglycemia induced oxidative stress.
Manypresent reports (Drel et al., 2007; Obrosova et al., 2007) provideconcrete evidence of the potential of some of the peroxynitrite decom-position catalysts against developmentof variousdeficits associatedwithdiabetic neuropathy. Our treatment regimendiffers from those studies aswe have treated rats for twoweeks after six weeks of untreated diabetesand is targeted against attained diabetic complication.
In conclusion, it can be said that peroxynitrite decompositioncatalysts showed protection against diabetic neuropathy as evidentfrom the improvement in motor nerve conduction velocity and nerveblood flow, hyperalgesia and biochemical deficits. This study canprovide rationale for designing pharmacological interventions againstperoxynitrite formation in diabetic neuropathy.
Acknowledgement
This study was supported by a grant from the Council of Scientificand Industrial Research (CSIR) India to Dr. Shyam S. Sharma.
References
Bartosz, G., 1996. Peroxynitrite: mediator of the toxic action of nitric oxide. ActaBiochim. Pol. 43, 645–659.
Cameron, N.E., Cotter, M.A., 1995. Effects of chronic treatment with a nitric oxide donoron nerve conduction abnormalities and endoneurial blood flow in streptozotocin-diabetic rats. Eur. J. Clin. Investig. 25, 19–24.
Coppey, L.J., Gellett, J.S., Davidson, E.P., Dunlap, J.A., Yorek, M.A., 2002. Changes inendoneurial blood flow, motor nerve conduction velocity and vascular relaxation ofepineurial arterioles of the sciatic nerve in ZDF-obese diabetic rats. Diabetes/Metab.Res. Rev. 18, 49–56.
Cuzzocrea, S., Mazzon, E., Di Paola, R., Esposito, E., Macarthur, H., Matuschak, G.M.,Salvemini, D., 2006. A role for nitric oxide-mediated peroxynitrite formation in amodel of endotoxin-induced shock. J. Pharmacol. Exp. Ther. 319, 73–81.
Drel, V.R., Pacher, P., Vareniuk, I., Pavlov, I., Ilnytska, O., Lyzogubov, V.V., Tibrewala, J.,Groves, J.T., Obrosova, I.G., 2007. A peroxynitrite decomposition catalyst counteractssensory neuropathy in streptozotocin-diabetic mice. Eur. J. Pharmacol. 569, 48–58.
Evans, J.L., Goldfine, I.D., Maddux, B.A., Grodsky, G.M., 2002. Oxidative stress and stress-activated signaling pathways: a unifying hypothesis of type 2 diabetes. Endocr. Rev.23, 599–622.
Freshwater, J.D., Svensson, C.I., Malmberg, A.B., Calcutt, N.A., 2002. Elevated spinalcyclooxygenase and prostaglandin release during hyperalgesia in diabetic rats.Diabetes 51, 2249–2255.
Fujimoto, Y., Uno, E., Sakuma, S., 2004. Effects of reactive oxygen and nitrogen species oncyclooxygenase-1 and -2 activities. Prostaglandins Leukot. Essent. Fat. Acids 71,335–340.
Kiss, L., Szabo, C., 2005. The pathogenesis of diabetic complications: the role of DNAinjury and poly(ADP-ribose) polymerase activation in peroxynitrite-mediatedcytotoxicity. Mem. Inst. Oswaldo Cruz 100 (Suppl 1), 29–37.
Kooy, N.W., Royall, J.A., Ischiropoulos, H., Beckman, J.S., 1994. Peroxynitrite-mediatedoxidation of dihydrorhodamine 123. Free Radic. Biol. Med. 16, 149–156.
Kumar, A., Kaundal, R.K., Iyer, S., Sharma, S.S., 2007. Effects of resveratrol on nervefunctions, oxidative stress and DNA fragmentation in experimental diabeticneuropathy. Life Sci. 80, 1236–1244.
Lowry, O.H., Rosebrough, N.J., Farr, A.L., Randall, R.J., 1951. Protein measurement withthe Folin phenol reagent. J. Biol. Chem. 193, 265–275.
Mabley, J.G., Liaudet, L., Pacher, P., Southan, G.J., Groves, J.T., Salzman, A.L., Szabo, C.,2002. Part II: beneficial effects of the peroxynitrite decomposition catalyst FP15 inmurine models of arthritis and colitis. Mol. Med. 8, 581–590.
Marla, S.S., Lee, J., Groves, J.T., 1997. Peroxynitrite rapidly permeates phospholipidmembranes. Proc. Natl. Acad. Sci. U. S. A. 94, 14243–14248.
Misko, T.P., Highkin, M.K., Veenhuizen, A.W., Manning, P.T., Stern, M.K., Currie, M.G.,Salvemini, D., 1998. Characterization of the cytoprotective action of peroxynitritedecomposition catalysts. J. Biol. Chem. 273, 15646–15653.
Moron, M.S., Depierre, J.W., Mannervik, B., 1979. Levels of glutathione, glutathionereductase and glutathione S-transferase activities in rat lung and liver. Biochim.Biophys. Acta 582, 67–78.
Nedelec, E., Abid, A., Cipolletta, C., Presle, N., Terlain, B., Netter, P., Jouzeau, J., 2001.Stimulation of cyclooxygenase-2-activity by nitric oxide-derived species in ratchondrocyte: lack of contribution to loss of cartilage anabolism. Biochem.Pharmacol. 61, 965–978.
Obrosova, I.G., Drel, V.R., Pacher, P., Ilnytska, O., Wang, Z.Q., Stevens, M.J., Yorek, M.A.,2005a. Oxidative-nitrosative stress and poly(ADP-ribose) polymerase (PARP)activation in experimental diabetic neuropathy: the relation is revisited. Diabetes54, 3435–3441.
Obrosova, I.G., Mabley, J.G., Zsengeller, Z., Charniauskaya, T., Abatan, O.I., Groves, J.T.,Szabo, C., 2005b. Role for nitrosative stress in diabetic neuropathy: evidence fromstudies with a peroxynitrite decomposition catalyst. FASEB. J. 19, 401–403.
Obrosova, I.G., Drel, V.R., Oltman, C.L., Mashtalir, N., Tibrewala, J., Groves, J.T., Yorek,M.A.,2007. Role of nitrosative stress in early neuropathy and vascular dysfunction instreptozotocin-diabetic rats. Am. J. Physiol., Endocrinol. Metab. 293, E1645–1655.
Pacher, P., Szabo, C., 2006. Role of peroxynitrite in the pathogenesis of cardiovascularcomplications of diabetes. Curr. Opin. Pharmacol. 6, 136–141.
Patel, R.P., McAndrew, J., Sellak, H., White, C.R., Jo, H., Freeman, B.A., Darley-Usmar, V.M.,1999. Biological aspects of reactive nitrogen species. Biochim. Biophys. Acta 1411,385–400.
Pop-Busui, R., Marinescu, V., Van Huysen, C., Li, F., Sullivan, K., Greene, D.A., Larkin, D.,Stevens, M.J., 2002. Dissection of metabolic, vascular, and nerve conductioninterrelationships in experimental diabetic neuropathy by cyclooxygenase inhibi-tion and acetyl-L-carnitine administration. Diabetes 51, 2619–2628.
Rundquist, I., Smith, Q.R., Michel, M.E., Ask, P., Oberg, P.A., Rapoport, S.I., 1985. Sciaticnerve blood flow measured by laser Doppler flowmetry and [14C]iodoantipyrine.Am. J. Physiol. 248, H311–317.
Salvemini, D., Wang, Z.Q., Stern, M.K., Currie, M.G., Misko, T.P., 1998. Peroxynitritedecomposition catalysts: therapeutics for peroxynitrite-mediated pathology. Proc.Natl. Acad. Sci. U. S. A. 95, 2659–2663.
Sayyed, S.G., Kumar, A., Sharma, S.S., 2006. Effects of U83836E on nerve functions,hyperalgesia and oxidative stress in experimental diabetic neuropathy. Life Sci. 79,777–783.
Sharma, S.S., Munusamy, S., Thiyagarajan, M., Kaul, C.L., 2004. Neuroprotective effect ofperoxynitrite decomposition catalyst and poly(adenosine diphosphate-ribose)polymerase inhibitor alone and in combination in rats with focal cerebral ischemia.J. Neurosurg. 101, 669–675.
Sharma, S.S., Kumar, A., Kaundal, R.K., 2008. Protective effects of 4-amino1,8-napthalimide, a poly (ADP-ribose) polymerase inhibitor in experimental diabeticneuropathy. Life Sci. 82, 570–576.

83M. Arora et al. / European Journal of Pharmacology 596 (2008) 77–83
Sima, A.A., 2006. Pathological mechanisms involved in diabetic neuropathy: can weslow the process? Curr. Opin. Investig. Drugs 7, 324–337.
Stamler, J.S., 1994. Redox signaling: nitrosylation and related target interactions of nitricoxide. Cell 78, 931–936.
Stevens, M.J., Obrosova, I., Cao, X., Van Huysen, C., Greene, D.A., 2000. Effects of DL-alpha-lipoic acid on peripheral nerve conduction, blood flow, energy metabolism,and oxidative stress in experimental diabetic neuropathy. Diabetes 49, 1006–1015.
Sweitzer, S.M., Medicherla, S., Almirez, R., Dugar, S., Chakravarty, S., Shumilla, J.A.,Yeomans, D.C., Protter, A.A., 2004. Antinociceptive action of a p38alpha MAPKinhibitor, SD-282, in a diabetic neuropathy model. Pain 109, 409–419.
Szabo, C., 2003. Multiple pathways of peroxynitrite cytotoxicity. Toxicol. Lett. 140–141,105–112.
Szabo, C., 2005. Roles of poly(ADP-ribose) polymerase activation in the pathogenesis ofdiabetes mellitus and its complications. Pharmacol. Res. 52, 60–71.
Szabo, C., Salzman, A.L., Ischiropoulos, D., 1995. Peroxynitrite-mediated oxidation ofdihydrorhodamine123 occurs in early stages of endotoxic and hemorrhagic shockand ischemia–reperfusion injury. FEBS Lett. 372, 229–232.
Takeuchi, M., Low, P.A., 1987. Dynamic peripheral nerve metabolic and vascularresponses to exsanguination. Am. J. Physiol. 253, E349–353.
Thiyagarajan, M., Kaul, C.L., Sharma, S.S., 2004. Neuroprotective efficacy and therapeutictime window of peroxynitrite decomposition catalysts in focal cerebral ischemia inrats. Br. J. Pharmacol. 142, 899–911.
Tietze, F., 1969. Enzymic method for quantitative determination of nanogram amountsof total and oxidized glutathione: applications to mammalian blood and othertissues. Anal. Biochem. 27, 502–522.