an introduction to marmosops (marsupialia: didelphidae...
TRANSCRIPT

Copyright q American Museum of Natural History 2004 ISSN 0003-0082
P U B L I S H E D B Y T H E A M E R I C A N M U S E U M O F N AT U R A L H I S T O RY
CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024
Number 3466, 40 pp., 12 figures, 13 tables December 30, 2004
An Introduction to Marmosops (Marsupialia:Didelphidae), with the Description of a New
Species from Bolivia and Notes on the Taxonomyand Distribution of Other Bolivian Forms
ROBERT S. VOSS,1 TERESA TARIFA,2,3 AND ERIC YENSEN4
CONTENTS
Abstract . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2Taxonomic Accounts . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
Marmosops Matschie, 1916 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5Marmosops creightoni, new species . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11
Notes on the Taxonomy and Distribution of Other Bolivian Forms . . . . . . . . . . . . . . . . . . 24Marmosops bishopi . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24Marmosops impavidus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 25Marmosops noctivagus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 27Marmosops ocellatus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29Unidentified Marmosops . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
Discussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32Acknowledgments . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 34Appendix 1: Type localities of nominal species of Marmosops . . . . . . . . . . . . . . . . . . . . . . 37Appendix 2: Gazetteer of examined specimens of Bolivian Marmosops . . . . . . . . . . . . . . 39
1 Division of Vertebrate Zoology (Mammalogy), American Museum of Natural History.2 Coleccion Boliviana de Fauna, La Paz.3 Current address: Museum of Natural History, Albertson College, Caldwell, ID 83605.4 Department of Biology, Albertson College, Caldwell, ID 83605.

2 NO. 3466AMERICAN MUSEUM NOVITATES
ABSTRACT
In order to facilitate much-needed revisionary research on Marmosops, we summarize thecurrently accepted species-level taxonomy, provide full bibliographic citations for originaldescriptions of all 36 included nominal taxa, map their type localities, and list their typematerial (if known). We rediagnose the genus Marmosops, compare it with three other didel-phid genera to which misidentified specimens of Marmosops have often been referred, andreview the phylogenetic evidence that Marmosops is monophyletic. After describing a newspecies from the eastern-slope montane forests of Bolivia, we review the taxonomy of otherBolivian congeners based on morphological characters and published cytochrome-b gene se-quences. Among our taxonomic results, we synonymize albiventris Tate (1931), dorotheaThomas (1911), and yungasensis Tate (1931) with M. noctivagus (Tschudi, 1845). By contrast,M. ocellatus (Tate, 1931), currently considered a synonym of dorothea, appears to be a validspecies. Whereas published range maps of Bolivian species of Marmosops are demonstrablybased on misidentified material and show little correspondence with known environmentalfactors, locality records based on specimens examined for this report make much more eco-geographic sense.
INTRODUCTION
Didelphid marsupials represent the onlysubstantially intact radiation of metatherianmammals in the New World, where they con-stitute a distinctive component of terrestrialvertebrate faunas in many tropical and sub-tropical habitats. Although certain aspects ofdidelphid classification have remained stablefor many years, notably the generic taxono-my of the larger opossums, recent discover-ies have underscored our still very incom-plete knowledge of phylogenetic diversityand relationships among the smaller forms,especially the so-called ‘‘marmosines’’ (for-merly classified as, or allied with, MarmosaGray, 1821). Such discoveries include highlydivergent new clades (Voss et al., 2001; Vossand Jansa, 2003; Voss et al., 2004) and un-suspected species-level diversity concealedby traditional synonymies (Mustrangi andPatton, 1997; Voss et al., 2001; Patton andCosta, 2003). Both phenomena suggest thatthe current classification of small didelphidsrequires critical attention.
The didelphid marsupial genus Marmo-sops was originally named by Matschie(1916) to contain 11 species, of which M.incanus (Lund, 1840) was designated as thetype. Although Tate (1933) treated Marmo-sops as a junior synonym of Marmosa, heacknowledged that Matschie’s taxon corre-sponded to a diagnosable cluster of species(the ‘‘Noctivaga Group’’) that could logicallybe recognized as a subgenus. In the decadesthat followed, however, Marmosops contin-
ued to be treated as a junior synonym ofMarmosa, even in classifications that usedsubgeneric categories (e.g., Cabrera, 1958;Reig et al., 1985). The current recognition ofMarmosops as a distinct taxon dates fromGardner and Creighton (1989), who raised allof Tate’s species groups of Marmosa to ge-neric rank.
As currently recognized in the systematicliterature, Marmosops includes 36 nominaltaxa, of which 14 are considered to be validspecies and 22 to be subspecies or juniorsynonyms (table 1). The geographic disper-sion of type localities (fig. 1) approximatesthe distribution of the genus itself, whichranges from Panama to southeastern Brazil.Species of Marmosops occur in lowland rainforests, lowland dry forests, and in montane(‘‘cloud’’) forests to about 3000 m above sealevel. Trapping studies suggest that thesesmall (20–120 g) opossums are predomi-nantly understory species that are often takenon the ground or a few meters above theground (e.g., on logs, branches, and lianas),but apparently never in the canopy (Mal-colm, 1991; Patton et al., 2000; Voss et al.,2001; Cunha and Vieira, 2002; Vieira andMonteiro-Filho, 2003). Although alleged tobe insectivorous or omnivorous (Emmons,1997; Mustrangi and Patton, 1997; Cordero,2001; Voss et al., 2001), no analyses of rel-evant dietary data have been published. Infact, the natural history of Marmosops re-mains almost completely unstudied despitethe local abundance of some species in rel-atively accessible habitats.

2004 3VOSS ET AL.: BOLIVIAN MARMOSOPS
TABLE 1Valid Species and Synonyms Currently Referred to the Genus Marmosopsa

4 NO. 3466AMERICAN MUSEUM NOVITATES
Fig. 1. Type localities of nominal species referred to the genus Marmosops. Numbers are keyed togazetteer entries in appendix 1.
Museum specimens of Marmosops are fre-quently misidentified due to the absence ofany comprehensive current reference foridentifications. Although progress has beenmade in sorting out the taxonomy of Mar-mosops in southeastern Brazil (Mustrangiand Patton, 1997), western Amazonia (Pattonet al., 2000), and the Guiana region (Voss etal., 2001), no revisionary literature has ad-dressed the species-level systematics of thegenus in other parts of its enormous geo-graphic range, especially along the Andes.Even a superficial familiarity with the largecollections that have accumulated sinceTate’s (1933) revision suggests the urgentneed for testing traditionally accepted speciesconcepts with newly available material.
Herein we formally rediagnose Marmo-
sops to facilitate its recognition as a distinctclade and to provide a foundation for sub-sequent revisionary work. We next describea new species from Bolivia and summarizewhat little is known about its ecogeographicdistribution on the eastern slopes of the An-des. Lastly, we review the taxonomy and dis-tribution of other Bolivian congeners in orderto assess the regional diversity of the genusbased on museum specimens examined in thecourse of this study.
MATERIALS AND METHODS
Specimens cited in our tables and text arepreserved in the following collections (listedin order of their standard institutional abbre-viations): AMNH, American Museum of

2004 5VOSS ET AL.: BOLIVIAN MARMOSOPS
Natural History (New York); BMNH, Natu-ral History Museum (London); CBF, Colec-cion Boliviana de Fauna (La Paz); FMNH,Field Museum of Natural History (Chicago);INPA, Instituto Nacional de Pesquisas daAmazonia (Manaus); MNCN, Museo Na-cional de Ciencias Naturales (Madrid);MNRJ, Museu Nacional (Rio de Janeiro);MSB, Museum of Southwestern Biology(University of New Mexico, Albuquerque);MVZ, Museum of Vertebrate Zoology (Uni-versity of California, Berkeley); ROM, RoyalOntario Museum (Toronto); UMMZ, Univer-sity of Michigan Museum of Zoology (AnnArbor); USNM, National Museum of NaturalHistory (Washington, D.C.); ZMB, Museumfur Naturkunde der Humboldt-Universitat zuBerlin (Berlin); and ZMUC, Zoological Mu-seum of the University of Copenhagen (Co-penhagen). In addition, we examined a smallnumber of uncataloged specimens (cited byfield number: LHE, Louise H. Emmons;MJS, Matthew J. Swarner) that will eventu-ally be transferred to the USNM.
We transcribed total length (nose to fleshytail-tip, TL) and length of tail (basal flexureto fleshy tip, LT) from specimen labels orfield notes, and we computed head-and-bodylength (HBL) by subtracting LT from TL. Wealso transcribed length of hind foot (heel totip of longest claw, HF), length of ear (fromnotch, Ear), and weight from specimen labelsor field notes, but we sometimes remeasuredHF on fluid-preserved specimens to checkthe accuracy of values recorded by the col-lector, and we used our values wheneverlarge discrepancies were found. All externalmeasurements are reported to the nearest mil-limeter, and all weights are reported to thenearest gram.
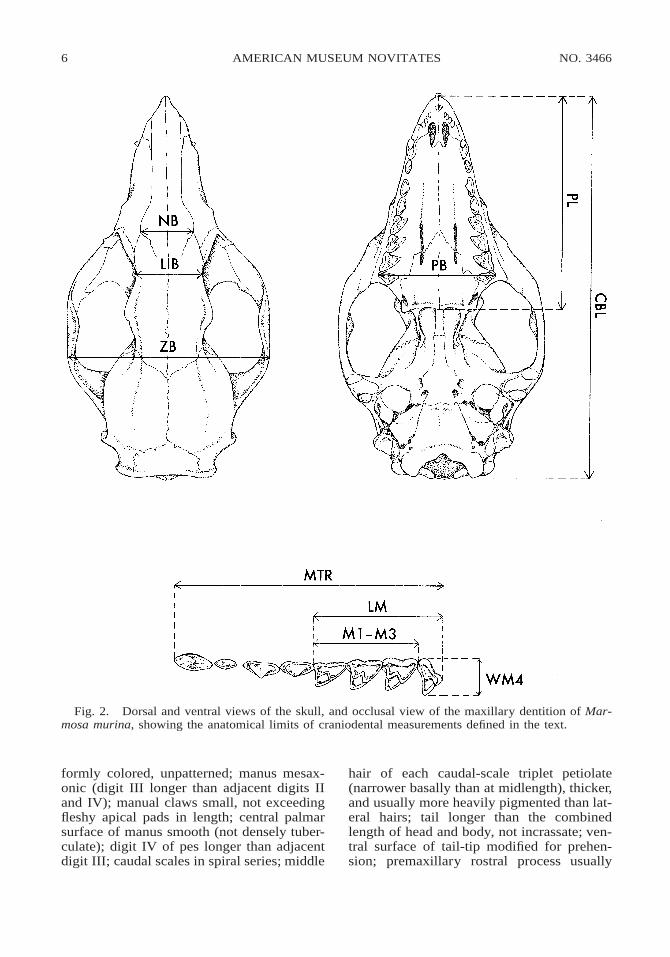
Craniodental measurements were takenwith digital calipers and recorded to the near-est 0.01 mm, but values reported herein arerounded to the nearest 0.1 mm. The followingwere measured as illustrated in figure 2:
Condylo-Basal Length (CBL): Measured from theoccipital condyles to the anteriormost point ofthe premaxillae.
Nasal Breadth (NB): Measured across the triple-point sutures of the nasal, frontal, and maxillarybones on each side.
Least Interorbital Breadth (LIB): Measured at thenarrowest point across the frontals between the
orbits, even when the postorbital constriction(between the temporal fossae) is narrower.
Zygomatic Breadth (ZB): Measured at the widestpoint across both zygomatic arches.
Palatal Length (PL): Measured from the anterior-most point of the premaxillae to the postpala-tine torus, including the postpalatine spine (ifpresent).
Palatal Breadth (PB): Measured across the labialmargins of the fourth molar (M4) crowns, at ornear the stylar A position.
Maxillary Toothrow Length (MTR): Measuredfrom the anterior margin of the canine (C1) tothe posterior margin of the fourth molar (M4).
Length of Molars (LM): Measured from the an-teriormost labial margin of M1 to the posteri-ormost point on M4.
Length of M1–M3 (M1–M3): Measured from theanteriormost labial margin of M1 to the poste-riormost point on M3.
Width of M4 (WM4): Measured from the labialmargin of the crown at or near the stylar Aposition to the lingual apex of the protocone.
Except as noted below, all analyzed char-acter data were obtained from adult speci-mens as determined by dental criteria. Fol-lowing Voss et al. (2001), a specimen wasjudged to be juvenile if dP3 is still in place;subadult if dP3 has been shed but P3 and/orM4 are still incompletely errupted; and adultif the permanent dentition is complete. Qual-itative character variation is described hereinusing terminology that is explained or ref-erenced by Voss and Jansa (2003) and Wible(2003). An exception (not defined by thoseauthors) is the prefix ‘‘self-’’ as used in com-bination with descriptors of ventral pelagecolor, such as self-white or self-cream. Thisusage applies to hairs that have the same col-oration from base to tip, as opposed to hairsthat are basally gray and distally white orcream (for example).
TAXONOMIC ACCOUNTS
Marmosops Matschie, 1916
TYPE SPECIES: Marmosops incanus (Lund,1840) by original designation.
DIAGNOSIS: Species of Marmosops can bedistinguished from other small didelphidmarsupials by the following combination ofcharacter states: eye surrounded by mask ofblackish fur contrasting in color with palerfur of crown and cheeks; dorsal pelage uni-
6 NO. 3466AMERICAN MUSEUM NOVITATES
Fig. 2. Dorsal and ventral views of the skull, and occlusal view of the maxillary dentition of Mar-mosa murina, showing the anatomical limits of craniodental measurements defined in the text.
formly colored, unpatterned; manus mesax-onic (digit III longer than adjacent digits IIand IV); manual claws small, not exceedingfleshy apical pads in length; central palmarsurface of manus smooth (not densely tuber-culate); digit IV of pes longer than adjacentdigit III; caudal scales in spiral series; middle
hair of each caudal-scale triplet petiolate(narrower basally than at midlength), thicker,and usually more heavily pigmented than lat-eral hairs; tail longer than the combinedlength of head and body, not incrassate; ven-tral surface of tail-tip modified for prehen-sion; premaxillary rostral process usually

2004 7VOSS ET AL.: BOLIVIAN MARMOSOPS
well developed; postorbital processes usuallyabsent; petrosal usually exposed on lateralsurface of braincase through fenestra be-tween squamosal and parietal bones; maxil-lary fenestrae absent; posterolateral palatalforamina small, not extending lingual to M4protocones; maxillary and alisphenoid not incontact on floor of orbit; extracranial courseof mandibular nerve (V3) enclosed by an-teromedial strut of alisphenoid bulla (second-ary foramen ovale present); fenestra cochleaeexposed laterally (not enclosed in a bony si-nus); I2–I5 with approximately symmetricalrhomboidal crowns increasing in breadthfrom front to back; P2 and P3 subequal inheight; molar dentition highly carnassialized;centrocrista strongly V-shaped, its apex highabove trigon basin; lower canine (c1) pro-cumbent and premolariform (always with aflattened, bladelike apex and usually with asmall posterior accessory cusp); p2 tallerthan p3; dp3 fully molariform; m3 hypoconidlabially salient; entoconid large and well de-veloped on m1–m3.
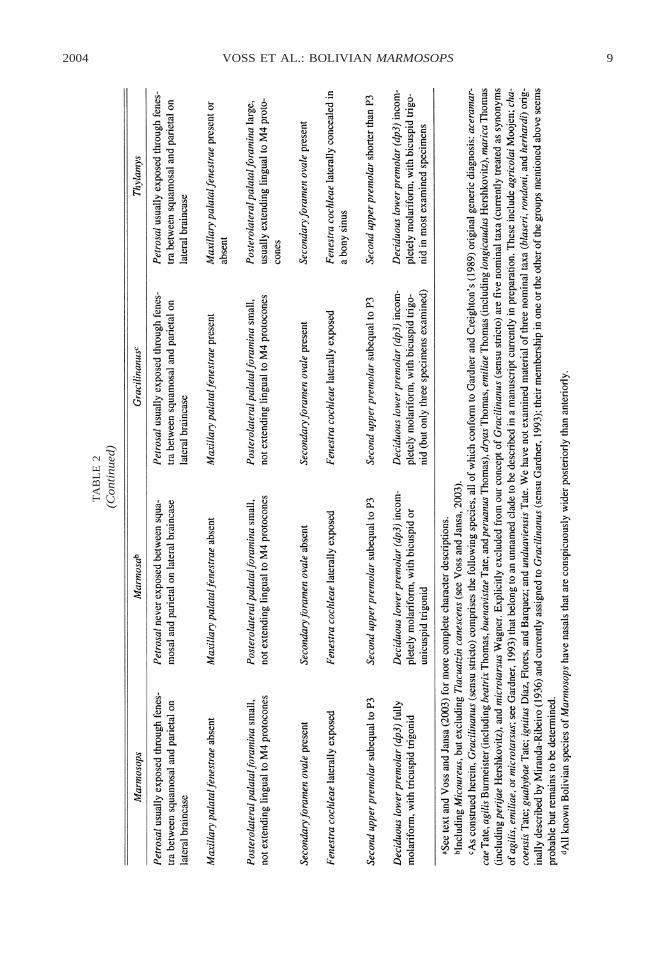
COMPARISONS WITH MARMOSA: Species ofMarmosops have long been confused withMarmosa, which they superficially resemblein size and external appearance, but thesetaxa are only distantly related (Jansa andVoss, 2000; Voss and Jansa, 2003) and canreadily be distinguished by qualitative integ-umental and craniodental characters (table 2).In the field, two external characters are usefulfor generic identification. Whereas the manusof Marmosops is mesaxonic, with digit (d)III distinctly longer than the adjacent digits(dII and dIV; fig. 3A), the manus of Mar-mosa is paraxonic, with dIII and dIV sube-qual in length (fig. 3B). Easily determinedfrom live specimens (or properly fluid-pre-served material) with elastic phalangealjoints that can be manipulated to align andstraighten adjacent digits for relative lengthcomparisons, this character is often hard toscore from dried skins with hard, bent, twist-ed, or otherwise distorted fingers (see Vossand Jansa, 2003: character 10). A second dis-tinguishing external trait is the morphologyof the caudal pelage, of which the centralhair in each caudal-scale triplet is petiolate(narrower basally than at mid-length), thick-er, and usually more darkly pigmented thanthe lateral hairs in Marmosops; by contrast,
the three hairs of each caudal-scale triplet aresubequal in thickness and similarly shapedand pigmented in Marmosa (Voss and Jansa,2003: character 27).
In dorsal cranial comparisons, these taxaare distinguished by the usual absence of dis-tinct postorbital processes of the frontal bonesin Marmosops (fig. 4B) versus the usual pres-ence of distinct postorbital processes in adultsof most species of Marmosa (fig. 4A). In lat-eral cranial view, the petrosal capsule that en-closes the paraflocculus and semicircular ca-nals (5 pars canalicularis or pars mastoideus)is usually exposed through a fenestra betweenthe squamosal and parietal bones in Marmo-sops (fig. 5A), but there is no such lateral pe-trosal exposure in Marmosa (fig. 5B). In ven-tral cranial view, Marmosops is distinguishedby the invariant presence of a secondary fo-ramen ovale (fig. 6B), which is just as con-sistently absent in Marmosa (fig. 6A).
Marmosops and Marmosa are similar inmost dental traits, but the morphology of thedeciduous lower third premolar (dp3) ap-pears to provide a consistent difference. Inall examined juvenile specimens of Marmo-sops (fig. 7, upper panel) this tooth is fullymolariform because the trigonid includes adistinct paraconid, protoconid, and metaco-nid. By contrast, the dp3 of juvenile Mar-mosa (fig. 7, lower panel) is not fully mo-lariform because the bicuspid trigonid lacksa distinct metaconid.
COMPARISONS WITH GRACILINANUS: Museumspecimens of Marmosops are sometimes mis-identified as Gracilinanus, apparently the re-sult of using inaccurate published keys.5 De-spite such confusion, the two genera can bedistinguished unambiguously by several ex-ternal morphological differences (table 2).Unlike Marmosops, which has a mesaxonicmanus with dIII longer than the other digits,Gracilinanus (like Marmosa) has a paraxonicmanus with dIII and dIV subequal in length.Also, whereas Marmosops has spirally ar-ranged caudal scales, the caudal scales ofGracilinanus are arranged in predominantly
5 Among other characters erroneously used in relevantkey couplets by Hershkovitz (1992: 6) and Anderson(1997: 30), presence/absence of gular glands, plantar padmorphology, nasal proportions, and presence/absence ofpostorbital processes do not reliably distinguish speciesof Marmosops from Gracilinanus.

8 NO. 3466AMERICAN MUSEUM NOVITATES
TA
BL
E2
Dia
gnos
tic
Mor
ph
olog
ical
Com
par
ison
sam
ong
Mar
mos
ops,
Mar
mos
a,G
raci
lin
anu
s,an
dT
hyl
amys
a
2004 9VOSS ET AL.: BOLIVIAN MARMOSOPS
TA
BL
E2
(Con
tinu
ed)

10 NO. 3466AMERICAN MUSEUM NOVITATES
Fig. 3. Dorsal views of right forefeet of Marmosops incanus (A, MVZ 197629) and Marmosarobinsoni (B, AMNH 259983) illustrating generic differences in digital proportions. Species of Mar-mosops have mesaxonic forefeet in which digit III is longer than all of the other manual digits and digitV is short (about equal in length to digit I). By contrast, species of Marmosa (and Gracilinanus) haveparaxonic forefeet in which digits III and IV are subequal and digit V is long.
annular series (Voss and Jansa, 2003: char-acter 23). Lastly, the central hair of each cau-dal-scale triplet is petiolate, thicker, and usu-ally more heavily pigmented than the lateralhairs in Marmosops, whereas the central hairis not conspicuously differentiated from thelateral hairs in Gracilinanus (Voss and Jansa,2003: character 27).
Cranially, Marmosops consistently lacksmaxillary fenestrae, palatal openings that areconsistently present in Gracilinanus on eachside of the maxillopalatine openings near theM1/M2 commissure (fig. 8). The morpholo-gy of the deciduous lower premolar (dp3)might also distinguish these taxa, but wehave examined so few juvenile Gracilinanus(two specimens of G. agilis and one G. em-iliae, all with incomplete trigonids) that thediagnostic value of this trait is unclear.
COMPARISONS WITH THYLAMYS: Specimensof Marmosops have occasionally been mis-identified as Thylamys despite the large num-ber of consistent differences that distinguishthese very dissimilar taxa. Whereas the dor-sal body pelage of Marmosops is uniformlycolored and unpatterned, the middorsum ofThylamys is distinctly darker than the flanks,
with a more-or-less sharp line of transitionbetween the two colors on either side (Vossand Jansa, 2003: character 7). Additional ex-ternal differences (table 2) include the mor-phology of the central palmar surface of themanus (Voss and Jansa, 2003: character 11),the occurrence of lateral carpal tubercles(Voss and Jansa, 2003: character 12), occur-rence of plantar pelage on the tarsus (Vossand Jansa, 2003: character 16), caudal hairmorphology (Voss and Jansa, 2003: character27), and caudal incrassation (Voss and Jansa,2003: character 28). Craniodental differencesbetween these genera are likewise numerous,including the presence/absence of a rostralpremaxillary process (Voss and Jansa, 2003:character 29), shape of the nasal bones (Vossand Jansa, 2003: character 33), size and ex-tent of the posterolateral palatal foramina(Voss and Jansa, 2003: character 41), lateralexposure of the fenestra cochleae (Voss andJansa, 2003: character 47), and upper pre-molar proportions (Voss and Jansa, 2003:character 55).
PHYLOGENETIC STATUS AND RELATIONSHIPS:Although distinguished from certain other‘‘marmosines’’ by a relatively small number

2004 11VOSS ET AL.: BOLIVIAN MARMOSOPS
Fig. 4. Dorsal views of the interorbital region in Marmosa murina (A, AMNH 267368), Marmosopsnoctivagus (B, AMNH 262402), and Gracilinanus agilis (C, MVZ 197439) illustrating generic differ-ences in the development of postorbital processes. Distinct postorbital processes of the frontals (ppf)are normally present in fully adult specimens of most Marmosa species, but they are usually absent inMarmosops and Gracilinanus (indistinct processes are sometimes developed in old adults). Supraorbitalbeads (b), consisting of upturned dorsally-grooved ridges, are present in some but not all species ofMarmosops. Scale bars equal 5 mm.
of morphological characters, Marmosops ap-pears to be a natural group based on recentphylogenetic analyses in which genericmonophyly was tested rather than assumed(Jansa and Voss, 2000; Voss and Jansa,2003). The most taxonomically completeanalysis to date (Voss and Jansa, 2003) in-cluded only five species of Marmosops, butamong these were members of the diminu-tive M. parvidens complex as well as largeAmazonian and southeastern Brazilianforms; generic monophyly was supported inseparate parsimony analyses of both non-molecular (morphological 1 karyotypic)characters and sequences from exon 1 of thenuclear gene encoding the InterphotoreceptorRetinoid Binding Protein (IRBP), in a like-lihood analysis of the IRBP data, and in aparsimony analysis of a combined-data (non-molecular 1 IRBP) supermatrix. The onlydiscrepant result concerning generic mono-phyly, Kirsch and Palma’s (1995) report thatGracilinanus agilis and Marmosops doro-thea are almost indistinguishable by thermalelution of scnDNA heteroduplexes, is nowknown to be an artifact of specimen mis-identification (see Voss and Jansa, 2003: 57).
Several phylogenetic analyses of molecu-lar data (e.g., Kirsch and Palma, 1995; Jansaand Voss, 2000; Voss and Jansa, 2003) sug-
gest that Marmosops belongs to a didelphineclade that also includes Gracilinanus, Les-todelphys, and Thylamys. The recently de-scribed genus Chacodelphys may also belongto this group (Voss et al., 2004). AlthoughPatton et al. (1996) recovered Marmosops asthe sister taxon of Caluromys, the cyto-chrome-b sequences they analyzed appear tobe substitution-saturated at this level of tax-onomic comparison (Jansa and Voss, 2000:fig. 12), and the relationship in question(Marmosops 1 Caluromys) had only trivialbootstrap support in their parsimony results.Palma and Spotorno’s (1999) report of a sis-ter-group relationship between Marmosopsand Metachirus (supported by very largebootstrap values in their neighbor-joiningand parsimony trees) is an unexplainedanomaly that is not consistent with any otheranalytic results or character data of which weare aware.
Marmosops creightoni, new speciesFigure 9
HOLOTYPE: An adult male specimen in theColeccion Boliviana de Fauna (CBF 6552)collected by Eric Yensen (original number1705) on 5 August 1999 near the Saynanihydroelectric generating station (ca. 168079S,

12 NO. 3466AMERICAN MUSEUM NOVITATES
Fig. 5. Lateral view of posterior braincase in Marmosops impavidus (A, AMNH 272709) and Mar-mosa murina (B, AMNH 272816) illustrating the presence of a fenestra (fen) that exposes the petrosal(pet) between the parietal (par) and squamosal (sq) in the former taxon. The petrosal is not laterallyexposed by a fenestra between the parietal and squamosal in Marmosa.
688059W; 2500 m above sea level) in the val-ley of the Rıo Zongo, Departamento La Paz,Bolivia. The holotype is a whole (unskinned)fluid-preserved specimen with an extractedskull and mandibles.
PARATYPES: Five additional specimens(listed as unidentified ‘‘Marmosa’’ by An-derson, 1997: 151) were collected by G.K.Creighton in 1979 at two adjacent localitiesin the valley of the Rıo Zongo, DepartmentoLa Paz, Bolivia. Two of these (UMMZ155999, 156000) were taken above the typelocality near the Cuticucho hydroelectricgenerating station (ca. 168089S, 688079W) ata recorded elevation of 2967 m, whereasthree others (UMMZ 156001–156003) weretrapped below the type locality at a recordedelevation of 2000 m. All five paratypes areadult animals preserved as whole (unskin-ned) fluids with extracted skulls and mandi-bles.
DISTRIBUTION: Known only from wet mon-tane forest in the valley of the Rıo Zongo,Departamento La Paz, Bolivia, between 2000and 3000 m above sea level (fig. 10).
ETYMOLOGY: For G. Ken Creighton, col-lector of the first known specimens of thisspecies and author of the first modern anal-ysis of didelphid phylogeny based on mor-phological characters (Creighton, 1984).
DESCRIPTION: Marmosops creightoni is amedium-sized species (see table 3 for mea-surements and weights), much larger thanmembers of the M. parvidens group (ca. 20–35 g adult weight; Voss et al., 2001: tables9, 10) but substantially smaller than somelowland forms like M. incanus (adults ofwhich can weigh as much as 120 g; Loriniet al., 1994). The body pelage is almost en-tirely dark, the colors probably somewhatfaded in our fluid-preserved specimens, butstill a rich chocolate-brown dorsally on theholotype (collected in 1999) and only a littlemore bleached on the 24-year-old paratypes.The ventral body coloration is slightly palerbrown, but there is no sharp distinction be-tween dorsal and ventral color zones, and allof the thoracic and abdominal fur is gray-based. The chin is self-white in all of thematerial at hand, and white fur extends pos-teriorly onto the throat of one paratype(UMMZ 155601). The blackish mask thatsurrounds the eye on each side of the headdoes not extend anteriorly to include themystacial pads (which are covered with shortbrownish fur) or posteriorly to the base ofthe ears. The mystacial and genal vibrissaeare blackish, whereas the submental and in-terramal hairs are white. The pinnae are uni-formly dark and appear naked (a sparse au-

2004 13VOSS ET AL.: BOLIVIAN MARMOSOPS
Fig. 6. Ventral view of left ear region in Marmosa murina (A, AMNH 267368) and Marmosopspinheiroi (B, AMNH 267346) illustrating generic differences in the presence/absence of a secondaryforamen ovale. In Marmosa, the mandibular branch of the trigeminal nerve (V3, reconstructed courseshown by heavy arrow) emerges from the endocranial lumen via the foramen ovale (fo), which isbordered by the alisphenoid (als) and the petrosal (pet); the extracranial course of the nerve is unen-closed in this taxon. In Marmosops, however, the extracranial course of V3 is partially enclosed by abony strut (st) that extends from the anteromedial surface of the tympanic wing of the alisphenoid (twa)across the transverse canal foramen (tcf); the nerve then emerges from a so-called secondary foramenovale. Other abbreviations: bs, basisphenoid; bo, basioccipital; cc, carotid canal; ect, ectotympanic.
ricular pelage is only microscopically visi-ble). Laid back alongside the head, the shortmystacial vibrissae do not extend to the tipsof the pinnae. A small patch of almost-nakedskin on the midline of the throat suggests thepresence of a weakly developed gular glandin all of the male specimens examined. Themetacarpals and metatarsals are covered withdark (brownish) hairs, but the digits of bothmanus and pes are abruptly whitish. All ofthe males we examined have a knob-like gla-brous tubercle on the lateral aspect of thewrist that is absent in both females. The scro-tal epithelium is pigmented (mottled-grayishin preservative) and sparsely covered bybrownish fur. Both female specimens have4–1–4 5 9 abdominal-inguinal mammae.Body fur extends onto a short (ca. 1 cm) seg-ment of the tail base, but the remainder(.90%) of the tail is virtually naked and
covered by epidermal scales in predominant-ly spiral series. Under high magnification(25–503), each caudal scale is seen to havethree hairs projecting from its posterior mar-gin; the central hair is visibly thicker than thelateral hairs, but perhaps not quite twice aswide. On the dorsal surface of the tail, thesehairs are no more than one to two scale rowslong, but they are longer (three to four scalerows) on the ventral caudal surface. The ex-posed caudal epithelium is dark (grayish)above and below basally, but it is indistinctlybicolored (dark above, pale below) for atleast half the length of that organ, and thetail-tip is entirely white for the last 1–3 cm.
In dorsal view the skull is distinctivelyproportioned with small orbits, narrow zy-gomatic arches, and a very broad interorbitalregion. The nasal bones are long and con-spicuously widened posteriorly near the max-

14 NO. 3466AMERICAN MUSEUM NOVITATES
Fig. 7. Occlusal views of right dp3 and m1 illustrating the generic difference in trigonid morphologyof the deciduous tooth. Top, Marmosops impavidus (MUSM 13286) with a complete (tricuspid) dp3trigonid. Bottom, Marmosa murina (MUSM 15297) with an incomplete (bicuspid) dp3 trigonid. Notethat whereas taxonomic variation in this trait was correctly described by Voss and Jansa (2003: character64), the accompanying plate (Voss and Jansa, 2003: fig. 15) was mislabeled.
illary/premaxillary suture. The broad inter-orbital region is due to the considerable in-flation of the cranial capsule that encloses theolfactory bulb, behind which there is a shal-low postorbital constriction; the supraorbitalmargins are rounded, with only indistinctbeading even in the largest male specimenexamined.6 The braincase is inflated andsmooth, without conspicuous temporal scars.In lateral cranial view the premaxillary/max-illary suture is lateral to (rather than behind)
6 The terms ‘‘bead’’ and ‘‘beading’’ as traditionallyused by systematic mammalogists to describe the dor-sally-grooved supraorbital ridges developed in somemarsupials and rodents (fig. 4B) are derived by analogyfrom the upturned rims used to strengthen sheet metalor other flat surfaces in manufacture (i.e., the last of fourdefinitions listed for the noun ‘‘bead’’ in Webster’s Sev-enth New Collegiate Dictionary, 1963).
the alveolus of I5 in most examined speci-mens (the suture is anterior to the alveoluson the left side of UMMZ 155999, and it isbehind the alveolus on the right side ofUMMZ 156000). The infraorbital foramen isvery large (its anterior opening approximate-ly as high as P3 is tall), and the ventral de-flection of the zygomatic arch beneath theorbit is shallow. The lacrimal foramina arecontained within the anterior orbital margin.The petrosal is laterally exposed through afenestra between the squamosal and parietalbones. In ventral cranial view the incisive fo-ramina extend between (but not behind) thecanine alveoli. The maxillopalatine fenestraeare long and narrow, extending on each sidefrom a point opposite P3 to M3 or to the M3/M4 commissure. The palatine fenestrae are
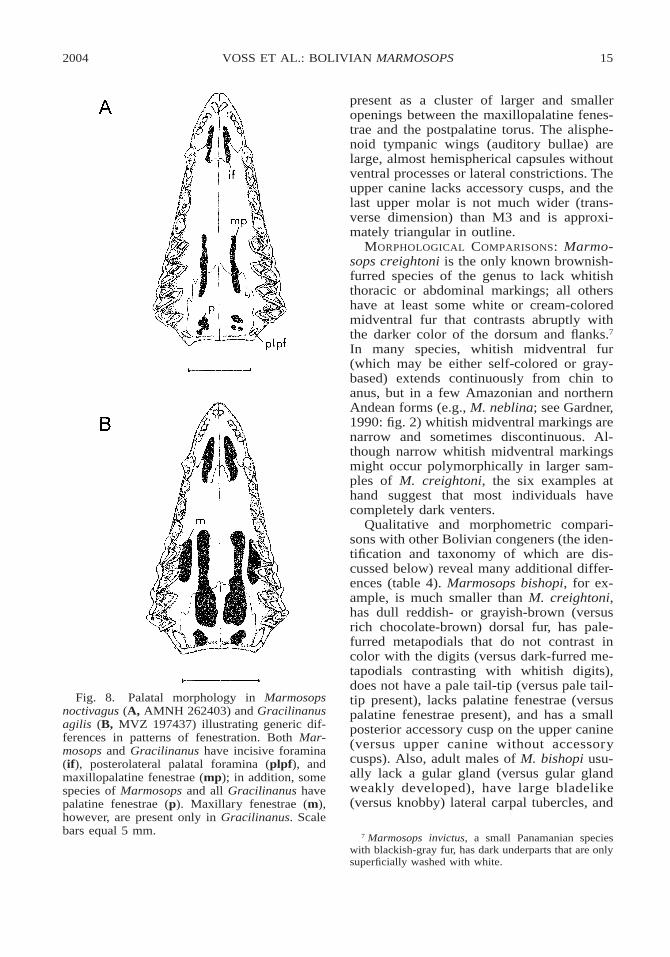
2004 15VOSS ET AL.: BOLIVIAN MARMOSOPS
Fig. 8. Palatal morphology in Marmosopsnoctivagus (A, AMNH 262403) and Gracilinanusagilis (B, MVZ 197437) illustrating generic dif-ferences in patterns of fenestration. Both Mar-mosops and Gracilinanus have incisive foramina(if), posterolateral palatal foramina (plpf), andmaxillopalatine fenestrae (mp); in addition, somespecies of Marmosops and all Gracilinanus havepalatine fenestrae (p). Maxillary fenestrae (m),however, are present only in Gracilinanus. Scalebars equal 5 mm.
present as a cluster of larger and smalleropenings between the maxillopalatine fenes-trae and the postpalatine torus. The alisphe-noid tympanic wings (auditory bullae) arelarge, almost hemispherical capsules withoutventral processes or lateral constrictions. Theupper canine lacks accessory cusps, and thelast upper molar is not much wider (trans-verse dimension) than M3 and is approxi-mately triangular in outline.
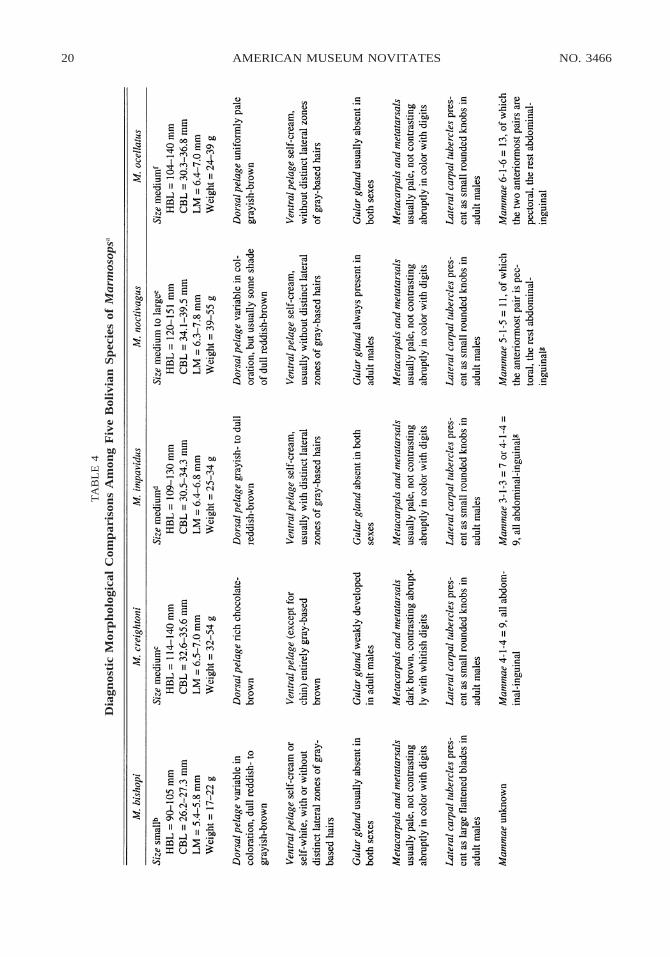
MORPHOLOGICAL COMPARISONS: Marmo-sops creightoni is the only known brownish-furred species of the genus to lack whitishthoracic or abdominal markings; all othershave at least some white or cream-coloredmidventral fur that contrasts abruptly withthe darker color of the dorsum and flanks.7
In many species, whitish midventral fur(which may be either self-colored or gray-based) extends continuously from chin toanus, but in a few Amazonian and northernAndean forms (e.g., M. neblina; see Gardner,1990: fig. 2) whitish midventral markings arenarrow and sometimes discontinuous. Al-though narrow whitish midventral markingsmight occur polymorphically in larger sam-ples of M. creightoni, the six examples athand suggest that most individuals havecompletely dark venters.
Qualitative and morphometric compari-sons with other Bolivian congeners (the iden-tification and taxonomy of which are dis-cussed below) reveal many additional differ-ences (table 4). Marmosops bishopi, for ex-ample, is much smaller than M. creightoni,has dull reddish- or grayish-brown (versusrich chocolate-brown) dorsal fur, has pale-furred metapodials that do not contrast incolor with the digits (versus dark-furred me-tapodials contrasting with whitish digits),does not have a pale tail-tip (versus pale tail-tip present), lacks palatine fenestrae (versuspalatine fenestrae present), and has a smallposterior accessory cusp on the upper canine(versus upper canine without accessorycusps). Also, adult males of M. bishopi usu-ally lack a gular gland (versus gular glandweakly developed), have large bladelike(versus knobby) lateral carpal tubercles, and
7 Marmosops invictus, a small Panamanian specieswith blackish-gray fur, has dark underparts that are onlysuperficially washed with white.

16 NO. 3466AMERICAN MUSEUM NOVITATES
Fig. 9. Dorsal, ventral, and lateral views of the skull, and lateral view of the left hemimandible ofthe holotype of Marmosops creightoni (CBF 6552); however, some missing details (e.g., the nasal tips,broken in the holotype) are reconstructed from UMMZ 155999 (a paratype). Note that the illustratedleft hemimandible of CBF 6552 is unusual in possessing only a single mental foramen; the right hem-imandible has two foramina, as do both hemimandibles of all five paratypes. Scale bar equals 10 mm.

2004 17VOSS ET AL.: BOLIVIAN MARMOSOPS
Fig. 10. Collection localities of Marmosops bishopi and M. creightoni in Bolivia. Numbers are keyedto gazetteer entries in appendix 2.
have a pale scrotum with white fur and un-pigmented skin (versus scrotum dark withbrown fur and mottled-grayish skin).
Marmosops impavidus, another Bolivianspecies, overlaps with M. creightoni in manymeasurements, although it averages smallerin most dimensions. However, M. impavidushas dull grayish- or reddish-brown (versusrich chocolate-brown) dorsal fur; pale-furredmetapodials that do not contrast in color with
the digits (versus dark-furred metapodialscontrasting with whitish digits); a much nar-rower interorbital region (LIB 5 5.6–6.3 ver-sus 6.6–7.2 mm); and much smaller bullae.In addition, adult males of M. impavidus ap-pear to have no trace of a gular gland (versusgular gland weakly developed), and theyhave a pale scrotum with white fur and un-pigmented skin (versus scrotum dark withbrownish fur and mottled-grayish skin).

18 NO. 3466AMERICAN MUSEUM NOVITATES
TABLE 3Measurements (mm) and Weights (g) of Marmosops creightoni
Bolivian specimens of Marmosops nocti-vagus overlap broadly with M. creightoni inmost measured dimensions, but they usuallyhave dull reddish-brown (versus rich choco-late-brown) dorsal fur; pale-furred metapo-dials that do not contrast in color with thedigits (versus dark-furred metapodials con-trasting with whitish digits); distinct supra-orbital beads that conceal the postorbital con-striction from dorsal view (versus supraor-bital beads absent or indistinct and postor-bital constriction visible dorsally); andsmaller, conical or flask-shaped bullae (ver-sus bullae larger and hemispherical). In ad-dition, adult males of M. noctivagus have apale scrotum with white fur and unpigmentedskin (versus scrotum dark with brownish furand mottled-grayish skin), and adult femaleshave 5–1–5 5 11 mammae, of which the an-teriormost pair is pectoral (versus 4–1–4 59 abdominal-inguinal teats).
Marmosops ocellatus also overlaps broad-ly with M. creightoni in most measured di-mensions, but it has much paler grayish-brown (versus rich chocolate-brown) dorsalfur; pale-furred metapodials that do not con-trast in color with the digits (versus dark-furred metapodials contrasting with whitish
digits); distinctly bi- and particolored tails,with the terminal one-third or more com-pletely pale (versus tail indistinctly bicoloredand only the tip completely pale); a muchnarrower interorbital region (LIB 5 5.5–6.3mm versus 6.6–7.2 mm); and proportionatelywider zygomatic arches (ZB/CBL 3 100 553–54% versus 48–50%). In addition, mostadult males of M. ocellatus lack a gular gland(versus gular gland weakly developed), andadult females have 6–1–6 5 13 mammae, ofwhich the anteriormost two pairs are pectoral(versus 4–1–4 5 9 abdominal-inguinal mam-mae).
Although we examined representative ma-terial of every species of Marmosops listedas valid in table 1, additional pairwise com-parisons are unnecessary because other cur-rently recognized species exhibit no specialsimilarity with M. creightoni. Indeed, mostextralimital (nonBolivian) species have strik-ingly divergent traits. Marmosops cracens,M. fuscatus, and M. incanus, for example,have uniformly narrow nasals with subpar-allel lateral margins (quite unlike the poste-riorly expanded nasals of Bolivian forms);M. pinheiroi has lacrimal foramina that arelaterally exposed anterior to the orbit (the

2004 19VOSS ET AL.: BOLIVIAN MARMOSOPS
lacrimal foramina of Bolivian species areusually concealed from lateral view insidethe orbit); and M. paulensis has very longincisive foramina that extend posteriorly be-hind the canine alveoli (the incisive foraminaare substantially shorter in all Bolivian taxa).These and other taxonomically informativecharacters merit uniform scoring for phylo-genetic analysis in any comprehensive at-tempt to revise the contents of Marmosops,but such exercises are beyond the scope ofthis preliminary report.
MOLECULAR COMPARISONS: The first 680base-pairs of the mitochondrial cytochrome-b gene were sequenced from the holotype ofMarmosops creightoni by J.L. Patton, whosegenerosity in sharing these data allowed usto make preliminary comparisons with ho-mologous didelphid sequences previouslyobtained by his laboratory (Patton et al.,1996; Mustrangi and Patton, 1997; Patton etal., 2000). In order to assess the relationshipsof M. creightoni with other congenericforms, we assembled a small dataset repre-senting nine species of Marmosops, and weincluded Gracilinanus microtarsus as an out-group (table 5). We selected exemplar hap-lotypes based on sequence length (the lon-gest available sequence for each species waschosen), previous representation in publishedreports (Patton et al., 1996; Mustrangi andPatton, 1997; Patton et al., 2000), and acces-sibility of cataloged morphological vouchermaterial in museums. Most species are rep-resented by single haplotypes in our analysis,but we included three sequences from geo-graphically divergent samples of M. nocti-vagus to assess molecular variation within ageographically widespread taxon.
As quantified by pairwise uncorrected se-quence divergence (table 6), Marmosopscreightoni is phenetically most similar to M.noctivagus, from which it differs on averageby about 9.7%. Interestingly, of the three an-alyzed haplotypes of the latter species, M.creightoni is most divergent (10.9%) fromthe geographically proximal Bolivian sampleand least divergent (9.2%) from each of thePeruvian and Brazilian specimens. All otherpairwise interspecific comparisons in theseresults are .10% divergent, with the excep-tion of a relatively small value (5.7%) be-tween M. impavidus and M. ocellatus.
A branch-and-bound parsimony analysisof the nucleotide sequence data (implement-ed with PAUP 4.0b10; Swofford, 1998) re-covered a single minimum-length tree withtwo main clades (fig. 11), one of which con-sists of Marmosops creightoni, M. impavi-dus, M. noctivagus, and M. ocellatus in thesequence (noctivagus (creightoni (impavidus1 ocellatus))). The other clade containsmembers of the parvidens complex (M. bish-opi and M. parvidens), two southeastern Bra-zilian forms (M. incanus and M. paulensis),and the Amazonian species M. neblina. Un-fortunately, there is only weak Bremer andbootstrap support for several branches in thistopology, notably including all of those thatresolve the relationships of creightoni withother congeneric forms.
NATURAL HISTORY: All of our material ofMarmosops creightoni was collected in thevalley of the Rıo Zongo, one of the principalheadwaters of the Rıo Beni on the easternslopes of the Cordillera Real. The Valle deZongo descends precipitously from 4800 to,900 m above sea level, with flanking slopesthat are often in excess of 608. The climateis very humid, with clouds and mist eachmorning and afternoon for most of the year.Between 2000 and 3000 m above sea level—the known elevational range of M. creighto-ni—the natural vegetation of the Valle deZongo is humid montane (‘‘cloud’’) forest.8
Habitat information associated with speci-mens of M. creightoni taken at three locali-ties in the Valle de Zongo is summarized be-low, in order of their collection dates.
The five paratypes that G.K. Creightoncollected in 1979 were taken at two localitiesalong a road that descends the valley andprovides access to a series of hydroelectricgenerating stations, power lines, and supportfacilities. His first collecting site was at Cu-
8 We use the nontechnical term ‘‘cloud forest’’ formontane rainforests generally, including the formationsthat Grubb (1977) usefully defined as Lower MontaneRain Forest, Upper Montane Rain Forest, and SubalpineRain Forest. Synonyms used by Bolivian botanists in-clude ‘‘Ceja de Monte de Yungas’’ (corresponding toSubalpine Rainforest; e.g., at Cuticucho) and ‘‘BosqueHumedo Montanoso de Yungas’’ (corresponding to Up-per Montane Rainforest; e.g., at Saynani). For a criticalreview of ‘‘cloud forest’’ and its technical synonymsfrom a field zoologist’s point of view, see Myers (1969);for another (botanical) perspective, see Webster (1995).
20 NO. 3466AMERICAN MUSEUM NOVITATES
TA
BL
E4
Dia
gnos
tic
Mor
ph
olog
ical
Com
par
ison
sA
mon
gF
ive
Bol
ivia
nS
pec
ies
ofM
arm
osop
sa

2004 21VOSS ET AL.: BOLIVIAN MARMOSOPS
TA
BL
E4
(Con
tinu
ed)

22 NO. 3466AMERICAN MUSEUM NOVITATES
TABLE 5Exemplar Specimens Included in Molecular Analysis
TABLE 6Uncorrected Percent Sequence Divergence Among Cytochrome-b Haplotypes
from Nine Species of Marmosops
ticucho (5 Cuti Khuchu on recent topo-graphic maps), which he recorded as havingan elevation of 2967 m above sea level (butsee appendix 2: locality 16). The site is de-scribed in his fieldnotes as follows:
Since our arrival [six days ago] we have not seen thesun for more than an hour a day. Clouds move up thevalley in the morning at around 8:30AM and formjust below our camp. Intermittent rain and mist arethe order of the day. The temperature here stays rel-atively constant, between 55 and 658 F [13–188 C].The forest here is very damp—many lichens, mossand ferns on branches of trees. We have trapped inthree habitats here: cloud forest (relatively undis-turbed), 2nd growth forest—with much bamboo; and
artificial clearings under power lines . . . We are verynear treeline here; as one looks up the valley the treesthin out and disappear at a fairly distinct ‘‘line’’ about300 m above camp. Looking down the valley thetrees get noticeably higher . . . [3 October 1979]
Both specimens of Marmosops creightonifrom Cuticucho were taken in the cloud for-est with snap traps: one (UMMZ 155999) atthe base of a tree and the second (UMMZ156000) on a low branch. Creighton also col-lected five species of sigmodontine rodentsat this site, of which Akodon mimus, Oxy-mycterus nigrifrons, and two undescribedforms of Thomasomys (‘‘sp. 1’’ and ‘‘sp. 3’’

2004 23VOSS ET AL.: BOLIVIAN MARMOSOPS
Fig. 11. The unique minimum-length tree recovered by a branch-and-bound parsimony analysis ofcytochrome-b nucleotide sequences representing 11 ingroup haplotypes from 9 Marmosops species and1 outgroup haplotype from Gracilinanus microtarsus (used to root the tree, not shown). Bremer supportand bootstrap values (.50%) are provided above and below each branch, respectively. The bootstrapvalues were obtained from branch-and-bound analyses of 1000 pseudoreplicated datasets. Tree length(including autapomorphies) 5 689 steps, consistency index (excluding autapomorphies) 5 0.48, retentionindex 5 0.44.
of Pacheco, 2003) were trapped in the cloudforest; Oryzomys levipes was trapped in near-by secondary growth. Additional species ofsmall mammals subsequently trapped nearCuticucho by other researchers include Gra-cilinanus aceramarcae (see Salazar-Bravo etal., 2002), Akodon cf. boliviensis, and an un-described form of Oligoryzomys (‘‘sp. B’’ ofthe flavescens group; Carleton and Musser,1989).
Farther down the valley G.K. Creightondescribed his camp at a locality he called‘‘Cement Mine’’ (not identifiable on our to-pographic maps), with a recorded elevationof 2000 m, as being much warmer than atCuticucho and with taller forest. However,
Much of the forest has been cut or burned at one timeor another in the past 30 years, hence most . . . is2nd growth except in the higher valleys intersectingthis one at more or less right angles. Lichens & mosson trees are solid in places but not as ‘‘lush’’ as nearCuticucho. There are more broadleaf trees and ‘‘pa-paya-like’’ trees here. Also large (;10 m) tree ferns.Bamboo undergrowth is extensive and very dense
near artificial clearings. Little undergrowth beneaththe bamboo except for a few ferns. [6 October 1979]
The first four nights of trapping (5–8 Octo-ber) at this site yielded only rodents (thesame species as those found in the cloud for-est near Cuticucho plus Microryzomys minu-tus and Thomasomys cf. oreas), but twospecimens of Marmosops creightoni weretaken on 9 October and another on 10 Oc-tober. Creighton’s notes for these dates indi-cate that the marsupials were trapped in ‘‘hu-mid, dark, mossy forest’’ on horizontalbranches near large trees ‘‘covered withmoss, ferns, bromeliads & vines’’. Appar-ently, they were ‘‘not caught coming to thebait but, rather, blundering into the traps asthey tiptoe along their accustomed routes.’’The area where all three Marmosops weretrapped included ‘‘about the only placeswhere high, undisturbed natural forest is ac-cessible enough to set a trapline’’, the onlyother primary forest being restricted to verysteep slopes above the cultivated valley floor.

24 NO. 3466AMERICAN MUSEUM NOVITATES
TABLE 7Measurements (mm) and Weights (g)
of the Brazilian Holotype and ReferredBolivian Material of Marmosops bishopi
The holotype of Marmosops creightoniwas collected by EY near the Saynani hy-droelectric generating station at 2500 m onthe southeastern side of the valley. The ob-viously disturbed forest at this site consistedof patches with a closed canopy but alsoopen areas with isolated trees. The trees wereladen with epiphytes, and the ground coverconsisted of vines, small bamboo, grasses,moss, and ferns. The holotype was taken onthe ground, in a snap trap set in dense grassunder a small tree. All of the other smallmammals trapped at Saynani by EY and oth-er researchers are sigmodontine rodents, in-cluding Akodon mimus, Chibchanomys sp.,Microryzomys minutus, Oryzomys levipes,Oxymycterus nigrifrons, and the same unde-scribed forms of Oligoryzomys and Thoma-somys that occur at Cuticucho.
In summary, Marmosops creightoni hasbeen collected in both primary cloud forestand disturbed vegetation in the Valle de Zon-go, where it occurs sympatrically with Gra-cilinanus aceramarcae and several species ofsigmodontine rodents between 2000 and3000 m above sea level. A different speciesof Marmosops, however, inhabits lower ele-vations in the Zongo valley. Two specimens,one from El Vertigo at 1800 m (CBF 4003)and another from Kahua at 1150 m (CBF3880), are both referable to M. noctivagus aswe recognize that species below. Future col-lecting in the elevational interval between2000 and 1000 m would be helpful in pin-pointing just where these congeneric speciesoccur sympatrically or parapatrically alongthe forested eastern slopes of the CordilleraReal.
NOTES ON THE TAXONOMY ANDDISTRIBUTION OF OTHER
BOLIVIAN FORMS
In the course of searching museum collec-tions for additional examples of Marmosopscreightoni, we examined many other Boli-vian specimens of Marmosops and sortedthem into phenotypically distinctive groupsthat we provisionally recognize as species.The following accounts summarize our con-clusions about the taxonomy and distributionof these forms based on currently availableinformation. For each species recognized be-
low, we explain the morphological and othercriteria that were used to assign names, andwe note conflicting identifications of thesame material reported by Anderson (1997),the most authoritative available reference onBolivian mammalogy.9
Marmosops bishopi
This appears to be the appropriate namefor Bolivian specimens of the Marmosopsparvidens group, of which we have exam-ined eight examples from widely scatteredlocalities in Beni, Cochabamba, La Paz, andSanta Cruz (fig. 10). These are all very smallanimals (table 7) with much shorter molarseries (LM # 5.8 mm) than any other Boli-vian congener. The skulls have rounded su-praorbital margins that lack distinct beading,shallow postorbital constrictions are dorsallyvisible, there are no palatine fenestrae, and
9 Because Anderson (1997) seldom cited museum cat-alog numbers, we consulted his card file of examinedspecimens (preserved in the AMNH Department ofMammalogy archives) to determine the empirical basisfor mapped collection localities of Bolivian Marmosops(Anderson, 1997: figs. 493–495).

2004 25VOSS ET AL.: BOLIVIAN MARMOSOPS
the upper canines have small posterior ac-cessory cusps. Although one adult male(MSB 55843) has an almost-naked patch ofskin and a streak of red-stained fur on themidline of the throat, two others (CBF 7531,LHE 1541) have no trace of a gular gland.All three adult males have well-developed,blade-like lateral carpal tubercles, and thescrotum is pale, with white fur and unpig-mented skin. None of the females we ex-amined had well-developed mammae, so it isnot known whether or not this taxon has pec-toral teats.
There is noteworthy pelage color variationamong the material at hand. All of the spec-imens we examined from Beni, Cochabamba,La Paz, and western Santa Cruz have dullreddish-brown dorsal fur, self-whitish ventralfur bordered by more-or-less distinct lateralzones of gray-based hairs, and unicoloredtails. By contrast, the single available skinfrom eastern Santa Cruz (LHE 1541)10 haspale grayish-brown dorsal fur, self-creamventral fur (without well- defined lateralzones of gray-based hairs), and an indistinct-ly bicolored tail. Despite these pigmental dif-ferences, specimens from eastern Santa Cruzdo not differ from the rest of our Bolivianmaterial in other respects (e.g., measure-ments and qualitative craniodental traits),and partial cytochrome-b sequences fromrepresentative exemplars (LHE 1541 andAMNH 268938) differ by only about 3.8%(J.L. Patton, personal commun.).
Specimens that we identify as Marmosopsbishopi were included among the materialthat Anderson (1997) reported as M. impav-idus (e.g., UMMZ 156014), M. noctivaguskeaysi (USNM 579249), and M. parvidens(AMNH 268938).
BOLIVIAN SPECIMENS EXAMINED: Beni, 1 km ELa Embocada (UMMZ 156014); Cochabamba,Cordillera de Mosetenes (CBF 7531); La Paz,Alto Rıo Madidi (USNM 579249), La Reserva(AMNH 268938); Santa Cruz, El Refugio (LHE1541, 1553, 1556), San Rafael de Amboro (MSB55843).
10 Unfortunately, both of the other specimens we ex-amined from eastern Santa Cruz (LHE 1553, 1556) con-sist of skulls extracted from rotted specimens found inpitfall traps.
Marmosops impavidus
The application of the name impavidus isproblematic because no type material isknown to exist, and because Tschudi’s (1845)original description is diagnostically inade-quate (Tate, 1933: 25, 182). Current usagedates from Cabrera (1958), who used the bi-nomen M. impavidus for the taxon that Tate(1933) recognized as M. caucae. Althoughthe lost type material is assumed to havebeen collected in the eastern foothills of thePeruvian Andes, no exact location was ex-plicitly stated in the original description. Forthe purposes of this report, we assume that asmall series of specimens (AMNH 230017,230018, 230020, 230022–230027) collectednear San Pablo (ca. 108279S, 748529W; 275m above sea level) in the Peruvian depart-ment of Pasco represents the typical mor-phology.
This a medium-size species (table 8), larg-er than Marmosops bishopi but averagingsmaller than typical specimens of M. nocti-vagus (see Patton et al. [2000] and below).The dorsal pelage is dull grayish-brown toreddish-brown, with drab (grayish) hues pre-dominating among younger specimens andwarmer (reddish) tones predominatingamong older adults. The ventral pelage ofmost Peruvian and western Brazilian speci-mens consists of a median strip of self-whiteor cream-colored fur from chin to anus, bor-dered by lateral zones of gray-based fur be-tween the fore- and hindlimbs. However, inthe Peruvian material at hand (which in-cludes specimens from Ucayali and Loreto inaddition to the Pasco sample mentionedabove) the relative widths of the median self-colored and lateral gray-based zones exhibitconsiderable variation. The metapodials arecovered with pale fur that does not contrastabruptly in color with the digits. Tails aresometimes indistinctly bicolored (paler be-low than above; e.g., AMNH 230017) andparticolored (paler distally than proximally;e.g., AMNH 230027), but other individualshave almost unicolored tails (e.g., AMNH76531). Adult males have knob-like lateralcarpal tubercles, but none that we examinedhas a gular gland; the scrotal fur is white andthe scrotal epithelium is unpigmented. Onelactating female from Loreto (AMNH

26 NO. 3466AMERICAN MUSEUM NOVITATES
TABLE 8Measurements (mm) and Weights (g) of Peruvian and Bolivian Specimens of Marmosops impavidus
272709) has 3–1–3 5 7 mammae, but anoth-er from the same locality (AMNH 273151)has 4–1–4 5 9; all of the teats in both in-dividuals are abdominal/inguinal.
In craniodental morphology, Marmosopsimpavidus can be distinguished from otherBolivian congeners by the absence of supra-orbital beads (usually distinct in M. noctiva-gus); presence of a shallow but usually dis-tinct postorbital constriction (usually absent inM. noctivagus); presence of palatine fenestrae(absent in M. bishopi); absence of accessoryupper canine cusps (present in M. bishopi);and very small, usually conical bullae.
The sole Bolivian specimen that we iden-tify as Marmosops impavidus is MSB 57002,which was collected sympatrically with M.noctivagus at Palmira in the lowlands of Pan-do department (fig. 12). This specimen hasredder dorsal fur and narrower nasals than domost Peruvian specimens, but its morpholo-gy is otherwise a good match with topotyp-ical (Peruvian) material. Other Bolivianspecimens previously referred to this species,however, appear to represent different taxa.
Anderson (1997: 153) reported two spec-imens identified as Marmosops impavidusfrom Incachaca in Cochabamba department,but cited only one by catalog number(AMNH 38718); the specimen in question is
a juvenile whose broken skull (originallymiscataloged as 38711) exhibits incipient su-praorbital beading, very short canines, andsmall molars (LM 5 6.3 mm), traits thatclosely match Tate’s (1933) description ofyungasensis—a taxon that we consider to bea junior synonym of M. noctivagus (see be-low). Two other Bolivian specimens explic-itly identified by Anderson as M. impavidusare UMMZ 126680 (from 20 km NNE Car-anavi, in La Paz department) and UMMZ156014 (from 1 km E La Embocada, in Benidepartment). The former is a subadult whoselack of supraorbital beading is possibly dueto immaturity, but whose lack of a postorbitalconstriction more closely resembles the ma-terial we refer to M. noctivagus (see below).UMMZ 156014, however, is an example ofM. bishopi (see above). The only additionalmaterial that Anderson referred to M. impav-idus consists of MSB 57000 (from Indepen-dencia in Pando department) and MSB58508 (from 2 km S Caranda in Santa Cruzdepartment); both are examples of ‘‘Graci-linanus’’ unduaviensis, a taxon that is cur-rently treated (e.g., by Gardner, 1993) as ajunior synonym of G. agilis.
BOLIVIAN SPECIMENS EXAMINED: Pando, Palmira(MSB 57002).

2004 27VOSS ET AL.: BOLIVIAN MARMOSOPS
Fig. 12. Collection localities of Marmosops impavidus, M. noctivagus, and M. ocellatus in Bolivia.Numbers are keyed to gazetteer entries in appendix 2.
Marmosops noctivagus
This appears to be the appropriate namefor the most-frequently collected species ofMarmosops in north-central Bolivia (fig. 12);we consider albiventris, dorothea, keaysi,and yungasensis to be synonyms, as ex-plained below. The name noctivagus is basedon Peruvian type material that we have notexamined, but a series of AMNH specimensfrom the Andean piedmont (,1000 m) near
Tarma in the department of Junın are more-or-less topotypical, and we have a large se-ries of very similar specimens from Loreto.One Bolivian specimen from La Paz (USNM579250) and three from Pando (AMNH262402–262404) closely resemble this low-land Peruvian material and could be referredto the nominotypical subspecies (M. n. noc-tivagus) if a trinomial nomenclature wereneeded. This material comprises relatively

28 NO. 3466AMERICAN MUSEUM NOVITATES
TABLE 9Measurements (mm) and Weights (g) of Lowland Specimens of Marmosops noctivagus
from Peru and Bolivia
large specimens (table 9) with dull reddish-brown dorsal fur, mostly self-whitish ventralfur, pale-furred metapodials that do not con-trast abruptly in color with the digits, andmore-or-less unicolored (all-dark) tails. Allof the adult males we examined have well-developed gular glands, knobby (not blade-like) lateral carpal tubercles, and pale scro-tums with whitish fur and unpigmented skin.Although we have not seen fluid-preservedparous adult females from Bolivia, two Pe-ruvian specimens (AMNH 272782, 273051)each have 5–1–5 5 11 mammae, of whichthe anteriormost pair is pectoral. Associatedskulls have distinct supraorbital beads (bestdeveloped in fully adult individuals; fig. 4B)and lack dorsally visible postorbital constric-tions; palatine fenestrae are present, and theauditory bullae are small and conical. Theupper canine lacks accessory cusps, and M4is very wide.
The remaining Bolivian material that werefer to Marmosops noctivagus is from thefoothills and lower montane regions of Beni,
Cochabamba, and La Paz. The specimens inquestion include the type material of doro-thea and yungasensis, but an older name forthe same phenotype is keaysi (from southernPeru; appendix 1).11 Measurements of rep-resentative examples (including relevant typematerial; table 10) indicate that these high-land specimens are about the same size astypical (lowland) noctivagus, but they have(on average) slightly broader interorbital re-gions, shorter molar rows, and narrowerfourth upper molars. In addition, the dorsalfur tends to be slightly longer, the palatal fe-nestrae to be somewhat larger, and the bullaeto be more inflated in highland examplesthan in lowland specimens. Also, whereas
11 Although Anderson (1997: 152–154) reported‘‘Marmosops dorothea’’ and ‘‘Marmosops noctivaguskeaysi’’ as co-occuring at Chijchijpa in the departmentof La Paz, all of the specimens in question (at AMNHand MSB) are taxonomically indistinguishable. Ander-son’s records of these taxa from Chijchijpa therefore rep-resent alternative identifications (apparently copied fromskin labels), not sympatry.

2004 29VOSS ET AL.: BOLIVIAN MARMOSOPS
most lowland specimens have more-or-lessunicolored (all dark) tails, tails of highlandspecimens are usually indistinctly bicolored(dark above, pale below) and particolored(paler distally than basally). Despite such el-evation-correlated phenotypic variation, cy-tochrome-b sequences analyzed by Mustran-gi and Patton (1997) and by Patton et al.(2000) indicate only minor divergence (about3%) among lowland and highland haplotypesof noctivagus-like Marmosops, includingspecimens from the Amazonian lowlands ofwestern Brazil (e.g., INPA 2931), the An-dean slopes of southeastern Peru (e.g., MVZ171408), and northern Bolivia (e.g., AMNH268936). The nominal taxon that Tate (1931)described as albiventris, currently synony-mized with M. impavidus, is morphologicallyindistinguishable from M. noctivagus and hassimilar mtDNA sequences (Mustrangi andPatton, 1997).12
Traditional taxonomic distinctions amongforms that we consider conspecific with Mar-mosops noctivagus are almost entirely basedon coat color. Tate (1933: 153–161), for ex-ample, described the dorsal fur as ‘‘varyingfrom natal brown . . . to Mars brown’’ intypical noctivagus, by contrast with ‘‘bonebrown . . . or warm sepia’’ in keaysi and‘‘pecan brown’’ in dorothea. These chromat-ic differences are real, but similarities amongthese nominal taxa in other morphologicalcharacters, the existence of intermediate pel-age hues in larger series of noctivagus-likeeastern-slope specimens obtained in recentyears, and the modest range of mtDNA var-iation among geographically widespreadsamples suggest that such color contrasts arenot taxonomically significant.
Paradoxically, several holotypes in thiscomplex exhibit atypical craniodental mea-surements, a factor that may have contributedto the traditional view that these specimensrepresent distinct taxa. For example, the ho-
12 We examined the holotype of Marmosa caucae al-biventris (USNM 194378) and the specimens that Mus-trangi and Patton (1997) identified as Marmosops albi-ventris (MVZ 171408, 173930–173935). All of this ma-terial fits our working diagnosis of M. noctivagus (table4) and exhibits no close similarity with M. impavidus.In particular, the ventral fur is mostly self-white, the su-praorbital margins are distinctly beaded in mature adults,and all of the adult male skins exhibit well-developedgular glands.
lotype of keaysi is an exceptionally largemale with a very broad interorbit that is notmatched by any paratype or topotype that weexamined. By contrast, the holotype of do-rothea has an unusually narrow interorbit,and the holotype of yungasensis has a con-spicuously short molar toothrow. Unfortu-nately, there are no large series from any lo-cality to serve as a relevant standard bywhich such morphometric variation might beassessed. Just among the 11 specimens of‘‘yungasensis’’ from Pitiguaya, however, in-terorbital breadth varies from 5.6 to 6.4 mm,and length of the molar series varies from 6.3to 7.1 mm. Therefore, none of the exception-al values for these dimensions in table 10would appear inconsistent with the hypoth-esis that the taxa represent one geographi-cally variable species.
Most of the specimens that we refer toMarmosops noctivagus were identified byAnderson (1997) as M. dorothea or M. noc-tivagus keaysi, but some were identified asM. impavidus as explained in the precedingaccount, and three others (UMMZ 155829,156006, 156007) were identified as Gracili-nanus agilis unduaviensis.
BOLIVIAN SPECIMENS EXAMINED: Beni, 1 km ELa Embocada (UMMZ 156008, 156013), Rurren-abaque (AMNH 247651); Cochabamba, Incacha-ca (AMNH 38718), Cordillera de Mosetenes(CBF 7527, 7560, 7573, 7577), Tablas Monte(AMNH 275451–275458; MSB 70276–70280,87080); La Paz, 20 km NNE Caranavi (UMMZ126680), Chijchijpa (AMNH 268936, 268937,275459; MSB 57004, 68333, 68334, 87092,87110), El Vertigo (CBF 4003), 13 km SW Ixia-mas (USNM 579250), Kahua (CBF 3880), La Re-serva (MSB 68335), Pitiguaya (AMNH 72550,72558–72562, 72564, 72566; UMMZ 155829,156006, 156007), Rıo Solacama (BMNH1.2.1.36, 1.6.7.79), Serranıa Bella Vista (AMNH275460), Ticunhuaya (AMNH 72567); Pando,Palmira (AMNH 262402, 262403), Santa Rosa(AMNH 262404).
Marmosops ocellatus
Judging from the large numbers of speci-mens in museum collections, Marmosopsocellatus is the commonest (or most fre-quently trapped) small marsupial throughoutmost of Santa Cruz department, from thebase of the Andes to the Brazilian frontier(fig. 12). Currently considered to be a junior

30 NO. 3466AMERICAN MUSEUM NOVITATES
TABLE 10Measurements (mm) and Weights (g) of Highland Specimens of Marmosops noctivagus,
Including the Type Material of Nominal Taxa Originally Described as keaysi, dorothea, and yungasensis
synonym of dorothea (5 M. noctivagus inour usage; see above), M. ocellatus is an un-ambiguously diagnosable taxon that moreclosely resembles M. impavidus in morphol-ogy and mtDNA sequences (tables 4, 6).
Externally, Marmosops ocellatus is dis-tinctively paler than other Bolivian species,with sandy grayish-brown dorsal fur andself-cream underparts that lack distinct lat-eral zones of gray-based hairs. The metapo-dials are covered dorsally with pale (whitish)hairs that do not contrast abruptly in colorwith the digits. The tail is distinctly bicolored(dark above, pale below) and particolored(paler distally than proximally), such that thedistal one-third or more of the organ is com-pletely pale in most specimens. Most exam-ined males have no trace of a gular gland,the only exception being the holotype (an un-usually large specimen; table 11), in whichthe gland appears to be present. The lateralcarpal tubercles of adult males are bulbousor knoblike, and the scrotum is pale (with
whitish fur and unpigmented skin). Threeparous adult female specimens prepared inthe field by L.H. Emmons (LHE 1468, 1596,1808) each had 6–1–6 5 13 mammae, ofwhich the anteriormost two pairs appear tobe pectoral in digital images that we exam-ined. The supraorbital margins of the skullare rounded, lacking distinct beads even inthe largest specimens, and a shallow postor-bital constriction is dorsally visible. Palatinefenestrae are consistently present, the uppercanine lacks accessory cusps, and the audi-tory bullae are moderately large.
Marmosops ocellatus differs from M. im-pavidus by its paler and grayer dorsal col-oration, absence of distinct lateral zones ofgray-based ventral hairs, possession of pec-toral mammae, and more distinctly bi- andparticolored tail. In addition, the palate ismore extensively fenestrated and the audito-ry bullae are much larger in M. ocellatusthan in M. impavidus. Although the differ-ence between exemplar sequences of these

2004 31VOSS ET AL.: BOLIVIAN MARMOSOPS
TABLE 11Measurements (mm) and Weights (g) of Selected Specimens of Marmosops ocellatusa
taxa (about 5.7%; table 6) is not as large asthe differences observed among other speciesrecognized by us, the phenotypic distinctive-ness of M. ocellatus and its apparently dis-crete geographic distribution suggest that itrepresents a unique evolutionary lineage thatshould be known by its own name.
Most of the material that we refer to Mar-mosops ocellatus was identified by Anderson(1997) as M. dorothea, but two specimens(MSB 67020, 87094) were identified as Gra-cilinanus agilis buenavistae, and another(AMNH 263549) was identified as Thylamysmacrurus.
BOLIVIAN SPECIMENS EXAMINED: Santa Cruz, 6km by road W Ascencion (AMNH 261265), As-erradero Pontons (CBF 6163), Ayacucho (USNM390571, 390572), Buenavista (BMNH 26.1.5.25[holotype], 28.2.9.87, 28.2.9.90), 4.5 km N and1.5 km E Cerro Amboro (MSB 55844), El Refu-gio (LHE 1569, 1573, 1577, 1596), 3.5 km WEstacion El Pailon (AMNH 260026–260028;MSB 55070), Hacienda el Pelicano (AMNH275462), 7 km E and 3 km N Ingeniero Mora(AMNH 247652), Lago Caiman (USNM 581979),2 km SW Las Cruces (AMNH 263549; MSB63274), Mangabalito (MJS 005), 3 km SE Mon-tero (MSB 87094), Palmar (USNM 390569), San
Miguel Rincon (AMNH 260029), 10 km N SanRamon (AMNH 261266, 261267), 15 km S SantaCruz (MSB 58510–58514, 59886, 67020), 27 kmSE Santa Cruz (59884, 59885), Santa Rosita(USNM 390022), Tita (MSB 55071).
Unidentified Marmosops
Among the Bolivian specimens we ex-amined are three that we are unable to con-fidently identify with any of the taxa recog-nized in this report. Two (AMNH 275461,MSB 67021) were collected 7 km SE Ari-ruma at an elevation of 1750 m on the lowerAndean slopes of western Santa Cruz de-partment. Unlike Marmosops ocellatus,which occurs in the adjacent lowlands ofSanta Cruz, these specimens have darker dor-sal fur, paler (self-white) underparts, and lackdistinct postorbital constrictions. They differfrom M. impavidus by their grayer dorsal fur,by lacking lateral zones of gray-based ventralfur, and by lacking distinct postorbital con-strictions. They differ from all of the formsherein referred to M. noctivagus by lackinginterorbital beads. They differ from M. bish-opi by having grayer dorsal pelage, well-de-

32 NO. 3466AMERICAN MUSEUM NOVITATES
TABLE 12Geographic and Elevational Distribution of Small Nonvolant Cloud-Forest Mammals
Collected Sympatrically with Marmosops creightoni
veloped palatine fenestrae, much larger teeth(LM 5 6.7 mm), and by lacking accessorycusps on C1.
The other unidentified specimen is MSB63275, a young adult female from the RıoLimon at 1300 m on the lower Andeanslopes of northeastern Chuquisaca. It hasbrowner and somewhat paler dorsal fur thandoes the Rıo Ariruma material, and self-cream ventral fur with broad lateral bands ofgray-based hairs (versus underparts entirelyself-white). However, the crania of all threespecimens are not dissimilar, and they differfrom those of recognized Bolivian taxa byessentially the same characters.
DISCUSSION
Bolivia encompasses a diversity of land-scapes with different vegetation types thatmight be expected to support distinct faunas.As currently recognized in the literature,however, Bolivian species of Marmosopshave broadly overlapping distributions thatdo not coincide with any obvious biogeo-graphic regions or macrohabitats. Althoughwe have not examined all of the material rep-resented in published range maps, the spec-imens identified in this report suggest thatsome species have much more geographical-ly restricted and ecologically comprehensibledistributions than do those illustrated by An-derson (1997) and Brown (2004).
Specimens that fit our working diagnosisof Marmosops noctivagus (a total of 53 in-dividuals from 17 localities) are from thelowland rain forests and lower-montanecloud forests of northern and central Bolivia,a region that approximately corresponds to
the Amazonian (‘‘Acre y Madre de Dios’’)and ‘‘Yungas’’ biogeographic provinces ofNavarro and Maldonado (2002). This speciesis also widespread in eastern Peru and west-ern Brazil, where it has been collected insimilar habitats (Voss and Emmons, 1996;Patton et al., 2000). Future collecting in Bo-livia will probably show that M. noctivagusoccurs throughout Pando department, in theforested parts of La Paz up to about 2000 mon the eastern Andean slopes, along the for-ested Andean foothills of southernmost Beni,and on the Andean piedmont and lower-mon-tane slopes of Cochabamba to at least 188S.
The other Bolivian species for which wehave examined enough material to character-ize its ecogeographic distribution (a total of37 individuals from 20 localities) is Mar-mosops ocellatus, which seems to be wide-spread in the ‘‘Cerrado’’ biogeographic prov-ince of Navarro and Maldonado (2002).Within this region, however, M. ocellatus hasonly been trapped in closed-canopy forests,not in the savanna woodlands or shrubbyhabitats that characterize Cerrado sensu stric-to (L.H. Emmons, in litt.). We expect thatfuture collecting will show that M. ocellatusoccurs throughout those parts of Santa Cruzreceiving about 1000–1500 mm of annualrainfall and in similar habitats of southwest-ern Brazil (Mato Grosso).
Although currently known from only threeadjacent localities in La Paz, Marmosopscreightoni is almost certainly more wide-spread. Based on the known geographicranges of other small nonvolant cloud-forestmammals that occur in the same elevationalinterval in the Valle de Zongo (table 12), we

2004 33VOSS ET AL.: BOLIVIAN MARMOSOPS
TABLE 13Records of Sympatry Among Bolivian
Species of Marmosops(Based on material examined for this report)
predict that M. creightoni will eventually befound in wet montane forests from centralBolivia to southeastern Peru. Whether or notM. creightoni is really co-distributed withany of the taxa listed in table 12 remains tobe demonstrated by more intensive collect-ing, but it is noteworthy that this assemblageof species is largely restricted to humid east-ern-Andean slopes with few convenientpoints of access for inventory fieldwork ex-cept along roads where colonization has al-ready caused extensive deforestation. There-fore, it may be a long time before significantrange extensions of M. creightoni are report-ed.
The material that we refer to Marmosopsbishopi (a total of eight individuals from sixlocalities) is an interesting exception to theecogeographically restricted distributions ofother Bolivian forms. Instead, specimens ofM. bishopi have been taken in a diversity oflandscapes, where they account for most ofthe known examples of sympatry document-ed by congeneric material examined for thisreport (table 13). Because strikingly differentmacrohabitats can sometimes occur in closeproximity, we consulted field notes associ-ated with Bolivian specimens of M. bishopiseeking some common ecological circum-stance in which they might have been taken,but we found none: captures have been re-corded in primary lowland rainforest(UMMZ 156014, USNM 579249), lowlanddry forest (LHE 1541), seasonally floodedevergreen gallery forest (LHE 1553, 1556),and cloud forest (CBF 7531). Either M. bish-
opi is an unusually eurytopic species, or thematerial that we consider conspecific mayconsist of two or more morphologicallycryptic taxa.
Some of the obvious gaps in our distri-bution maps of Bolivian Marmosops are cer-tainly artifactual, but others are probably not.Despite considerable collecting effort in theseasonally flooded savannas and gallery for-ests of central and northern Beni, for exam-ple, no specimens of Marmosops have beencollected there. The genus is likewise un-known from substantial collections made inthe arid thorn-scrub of southern Santa Cruz,eastern Chuquisaca, and eastern Tarija, norhave we seen any material among the manyhundreds of small mammals collected in thealtiplano habitats of southwestern Bolivia.By contrast, the eastern-slope lower-montaneforests of Chuquisaca and Tarija remain verypoorly sampled by mammalogists, and it ispossible that the genus is more widely dis-tributed there than current records suggest.
ACKNOWLEDGMENTS
We thank the curators and collection sup-port staffs that hosted our visits and pro-cessed the loans that made this study possi-ble, especially Paula Jenkins (BMNH); Julie-ta Vargas (CBF); Jorge Salazar-Bravo, BillGannon, and Joe Cook (MSB); Phil Myersand Steve Hinshaw (UMMZ); and Al Gard-ner, Linda Gordon, and Helen Kafka(USNM). Bolivian fieldwork was partiallyfunded by a grant from the American Societyof Mammalogists (to TT); and by the Na-tional Geographic Society, the Weeden Foun-dation, and BIOPAT (through grants to M.Kessler and S. Herzog). Logistical support inthe Valle de Zongo was graciously providedby the Companıa Boliviana de Energıa Elec-trica (COBEE). Cristian Olivo helped EYwith fieldwork in the Saynani area. Permitsto collect in Bolivia were issued by the Di-reccion General de Biodiversidad, Ministeriode Desarrollo Sostenible y Medio Ambiente.
Louise Emmons provided useful observa-tions and digital images from her recentfieldwork in eastern Santa Cruz and allowedus to examine uncataloged material that shecollected there. At the AMNH, Neil Duncanexpertly extracted and cleaned skulls from

34 NO. 3466AMERICAN MUSEUM NOVITATES
fluid-preserved specimens, and Pat Brunauerprovided essential bibliographic support. JimPatton set new standards for professionalgenerosity in permitting us unrestricted ac-cess to the cytochrome-b sequences obtainedby his lab, and Marcelo Weksler helped usanalyze them. We gratefully acknowledge afamiliar debt to Pat Wynne, whose artisticskills did so much to enhance our descrip-tions of didelphid morphology, and we thankRob Anderson for his technical assistance inthe production of figures 10 and 12. Ourmanuscript was substantially improved byhelpful suggestions provided by Rob Ander-son, Louise Emmons, David Flores, Al Gard-ner, and Rogerio Rossi, who generously reada preliminary draft when they had many oth-er things to do: we thank them for their time,effort, and expertise.
REFERENCES
Allen, J.A. 1900. Descriptions of new Americanmarsupials. Bulletin of the American Museumof Natural History 13: 191–199.
Allen, J.A. 1901. On a further collection of mam-mals from southeastern Peru, collected by Mr.H.H. Keays, with descriptions of new species.Bulletin of the American Museum of NaturalHistory 14: 41–46.
Allen, J.A., and F.M. Chapman. 1897. On a sec-ond collection of mammals from the island ofTrinidad, with descriptions of new species, anda note on some mammals from the island ofDominica, W.I. Bulletin of the American Mu-seum of Natural History 9: 13–30.
Anderson, R.P. 1999. Preliminary review of thesystematics and biogeography of the spinypocket mice (Heteromys) of Colombia. Revistade la Academia Colombiana de Ciencias Ex-actas, Fısicas y Naturales 23(suplemento espe-cial): 613–630.
Anderson, S. 1997. Mammals of Bolivia, taxon-omy and distribution. Bulletin of the AmericanMuseum of Natural History 231: 1–652.
Anthony, H.E. 1922. Preliminary report on Ecua-dorean mammals. No. 2. American MuseumNovitates 32: 1–6, map.
Brown, B.E. 2004. Atlas of New World marsu-pials. Fieldiana Zoology (new series) 102: i–vii,1–308.
Burmeister, H. 1856. Erlauterung zur Fauna Bras-iliens, enthaltend Abbildungen und ausfuhrli-che Beschreibungen neuer oder ungenugendbekannter Thier-Arten. Berlin: Druck und Ver-lag von Georg Reimer.
Cabrera, A. 1913. Dos mamıferos nuevos de lafauna neotropical. Trabajos del Museo de Cien-cias Naturales (Serie Zoologica) 9: 1–15 1 1pl.
Cabrera, A. 1958[‘‘1957’’]. Catalogo de los mam-ıferos de America del Sur [part 1]. Revista delMuseo Argentino de Ciencias Naturales ‘‘Ber-nardino Rivadavia’’ (Ciencias Zoologicas) 4:1–307.
Carleton, M.D., and G.G. Musser. 1989. System-atic studies of oryzomyine rodents (Muridae,Sigmodontinae): a synopsis of Microryzomys.Bulletin of the American Museum of NaturalHistory 191: 1–83.
Chubb, C. 1919. Notes on collections of birds inthe British Museum, from Ecuador, Peru, Bo-livia, and Argentina. Part 1. Tinamidae–Ralli-dae. Ibis 11: 1–55.
Cordero R., G.A. 2001. Ecological data on Mar-mosops fuscatus in a lowland tropical forest innorthern Venezuela. Mammalia 65: 228–231.
Creighton, G.K. 1984. Systematic studies onopossums (Didelphidae) and rodents (Criceti-dae). Ph.D. diss., University of Michigan. AnnArbor, MI: University Microfilms.
Cunha, A.A., and M.V. Vieira. 2002. Support di-ameter, incline, and vertical movements of fourdidelphid marsupials in the Atlantic forest ofBrazil. Journal of Zoology (London) 258: 419–426.
DMA. 1992. Gazetteer of Bolivia, 2nd ed. Wash-ington, DC: Defense Mapping Agency.
Emmons, L.H. 1997. Neotropical rainforest mam-mals, 2nd ed. Chicago: University of ChicagoPress.
Fairchild, G.B., and C.O. Handley, Jr. 1966. Gaz-etteer of collecting localities in Panama. In R.L.Wenzel and V.J. Tipton (editors), Ectoparasitesof Panama: 9–22, folding map.Chicago: FieldMuseum of Natural History.
Flores, D.A., M.M. Dıaz, and R.M. Barquez.2000. Mouse opossums (Didelphimorphia, Di-delphidae) of northwestern Argentina: system-atics and distribution. Zeitschrift fur Saugetier-kunde 65: 321–339.
Gardner, A.L. 1990 [‘‘1989’’]. Two new mammalsfrom southern Venezuela and comments on theaffinities of the highland fauna of Cerro de laNeblina. In K.H. Redford and J.F. Eisenberg(editors), Advances in Neotropical mammalo-gy: 411–424. Gainesville, FL: Sandhill CranePress. [According to A.L. Gardner (personalcommun.), this volume was published on 6February 1990]
Gardner, A.L. 1993. Order Didelphimorphia. InD.E. Wilson and D.M. Reeder (editors), Mam-mal species of the world, 2nd ed: 15–23.Wash-ington, DC: Smithsonian Institution Press.

2004 35VOSS ET AL.: BOLIVIAN MARMOSOPS
Gardner, A.L., and G.K. Creighton. 1989. A newgeneric name for Tate’s microtarsus group ofSouth American mouse opossums (Marsupialia:Didelphidae). Proceedings of the Biological So-ciety of Washington 102: 3–7.
Goldman, E.A. 1912. New mammals from easternPanama. Smithsonian Miscellaneous Collec-tions 60: 1–18.
Goodwin, G.G., and A.M. Greenhall. 1961. A re-view of the bats of Trinidad and Tobago. Bul-letin of the American Museum of Natural His-tory 122: 187–302 1 40 pls.
Grubb, P.J. 1977. Control of forest growth anddistribution on wet tropical mountains: withspecial reference to mineral nutrition. AnnualReview of Ecology and Systematics 8: 83–107.
Handley, C.O., Jr., and L. Gordon. 1979. Newspecies of mammals from northern SouthAmerica: mouse possums, genus MarmosaGray. In J.F. Eisenberg (editor), Vertebrate ecol-ogy in the northern Neotropics: 65–72. Wash-ington, D.C.: Smithsonian Institution Press.
Hershkovitz, P. 1992. The South American gracilemouse opossums, genus Gracilinanus Gardnerand Creighton, 1989 (Marmosidae, Marsupia-lia): a taxonomic review with notes on generalmorphology and relationships. Fieldiana Zool-ogy, new series 39: 1–56.
IGM. 1967. Zongo, 1: 50,000, Hoja 5945 I, SerieH731 [topographic map]. La Paz: InstitutoGeografico Militar.
Jansa, S.A., and R.S. Voss. 2000. Phylogeneticstudies on didelphid marsupials I. Introductionand preliminary results from nuclear IRBPgene sequences. Journal of Mammalian Evo-lution 7: 43–77.
Killeen, T.J., and T.S. Schulenberg (editors). 1998.A biological assessment of Parque NacionalNoel Kempff Mercado, Bolivia. RAP WorkingPapers 10: 1–372.
Kirsch, J.A.W., and R.E. Palma. 1995. DNA/DNAhybridization studies of carnivorous marsupi-als. V. A further estimate of relationshipsamong opossums (Marsupialia: Didelphidae).Mammalia 59: 403–425.
Langguth, A., V.L.A.G. Limeira, and S. Franco.1997. Novo catalogo do material-tipo da cole-cao de mamıferos do Museu Nacional. Publi-cacoes Avulsas do Museu Nacional 70: 1–29.
Lorini, M.L., J.A. de Oliveira, and V.G. Persson.1994. Annual age structure and reproductivepatterns in Marmosa incana (Lund, 1840) (Di-delphidae, Marsupialia). Zeitschrift fur Sauge-tierkunde 59: 65–73.
Lund, P.W. 1840. [Preprint of] Blik paa BrasiliensDyreverden for sidste Jordomvaeltning. TredieAfhandling: Fortsaettelse af Pattedyrene. Kon-gelige Danske Videnskabernes Selskabs Na-
turvidenskabelige og Matematiske Afhandlin-ger 8: 219–272 1 pls. xiv–xxiv. [Preprint 56pp., repaginated; not seen. Journal issue pagi-nated as above published 1841]
Malcolm, J.R. 1991. Comparative abundances ofNeotropical small mammals by trap height.Journal of Mammalogy 72: 188–192.
Matschie, P. 1916. Bemerkungen uber die GattungDidelphis L. Sitzungsberichte der GesellschaftNaturforschender Freunde, Berlin 1916: 259–272.
Miller, G.S. 1913. Five new mammals from trop-ical America. Proceedings of the Biological So-ciety of Washington 26: 31–34.
Miranda-Ribeiro, A. de. 1936. Didelphia ouMammalia-Ovovivipara. Revista do MuseuPaulista 20: 245–424.
Mustrangi, M.A., and J.L. Patton. 1997. Phylo-geography and systematics of the slendermouse opossum Marmosops (Marsupialia, Di-delphidae). University of California Publica-tions in Zoology 130: i–x, 1–86.
Myers, C.W. 1969. The ecological geography ofcloud forest in Panama. American MuseumNovitates 2396: 1–52.
Myers, P., J.L. Patton, and M.F. Smith. 1990. Areview of the Boliviensis Group of Akodon(Muridae: Sigmodontinae), with emphasis onPeru and Bolivia. Miscellaneous PublicationsMuseum of Zoology University of Michigan177: i–iv, 1–104.
Navarro, G., and M. Maldonado. 2002. Geografıaecologica de Bolivia, vegetacion y ambientesacuaticos. Cochabamba: Centro de Ecologıa Si-mon I. Patino.
Oliveira, J. de. 1998. Morphometric assessment ofspecies groups in the South American rodentgenus Oxymycterus (Sigmodontinae), with tax-onomic notes based on the analysis of type ma-terial. Unpublished Ph.D. diss., Texas TechUniversity.
Osgood, W.H. 1913. New Peruvian mammals.Field Museum of Natural History ZoologicalSeries 10: 93–100.
Osgood, W.H. 1915. New mammals from Braziland Peru. Field Museum of Natural HistoryZoological Series 10: 187–198.
Pacheco, V.R. 2003. Phylogenetic analyses of theThomasomyini (Muroidea: Sigmodontinae)based on morphological data. UnpublishedPh.D. diss., City University of New York.
Palma, R. E., and A. E. Spotorno. 1999. Molec-ular systematics of marsupials based on therRNA 12S mitochondrial gene: the phylogenyof Didelphimorphia and of the living fossil mi-crobiotheriid Dromiciops gliroides. MolecularPhylogenetics and Evolution 13: 525–535.
Patton, J.L., and L.P. Costa. 2003. Molecular phy-

36 NO. 3466AMERICAN MUSEUM NOVITATES
logeography and species limits in rainforest di-delphid marsupials of South America. In M.E.Jones, C.R. Dickman, and M. Archer (editors),Predators with pouches: the biology of carniv-orous marsupials: 63–81.Melbourne: CSIROPress.
Patton, J.L., S.F. dos Reis, and M.N.F. da Silva.1996. Relationships among didelphid marsupi-als based on sequence variation in the mito-chondrial cytochrome b gene. Journal of Mam-malian Evolution 3: 3–29.
Patton, J.L., M.N.F. da Silva, and J.R. Malcolm.2000. Mammals of the Rio Jurua and the evo-lutionary and ecological diversification ofAmazonia. Bulletin of the American Museumof Natural History 244: 1–306.
Paynter, R.A., Jr. 1982. Ornithological gazetteerof Venezuela. Cambridge, MA: Museum ofComparative Zoology (Harvard University).
Paynter, R.A., Jr. 1992. Ornithological gazetteerof Bolivia, 2nd ed. Cambridge, MA: Museumof Comparative Zoology (Harvard University).
Paynter, R.A., Jr. 1993. Ornithological gazetteerof Ecuador, 2nd ed. Cambridge, MA: Museumof Comparative Zoology (Harvard University).
Paynter, R.A., Jr. 1997. Ornithological gazetteerof Colombia, 2nd ed. Cambridge, MA: Muse-um of Comparative Zoology (Harvard Univer-sity).
Paynter, R.A., Jr., and M.L. Traylor, Jr. 1991. Or-nithological gazetteer of Brazil [2 vols.]. Cam-bridge, MA: Museum of Comparative Zoology(Harvard University).
Pine, R.H. 1981. Reviews of the mouse opossumsMarmosa parvidens Tate and Marmosa invictaGoldman (Mammalia: Marsupialia: Didelphi-dae) with description of a new species. Mam-malia 45: 55–70.
Reig, O.A., J.A.W. Kirsch, and L.G. Marshall.1985. New conclusions on the relationships ofthe opossum-like marsupials, with an annotatedclassification of the Didelphimorphia. Amegh-iniana 21: 335–343.
Salazar-Bravo, J., E. Yensen, T. Tarifa, and T.L.Yates. 2002. Distributional records of BolivianMammals. Mastozoologıa Neotropical 9: 70–78.
Sherborn, C.D. 1922. Index animalium [section 2,part 1]. London: Bitish Museum (Natural His-tory).
Stephens, L., and M.L. Traylor, Jr. 1983. Ornitho-logical gazetteer of Peru. Cambridge, MA: Mu-seum of Comparative Zoology (Harvard Uni-versity).
Stephens, L., and M.L. Traylor, Jr. 1985. Ornitho-logical gazetteer of the Guianas. Cambridge,MA: Museum of Comparative Zoology (Har-vard University).
Swofford, D.L. 1998. PAUP*, Phylogenetic anal-ysis using parsimony, beta version 4.0b10. Sun-derland, MA: Sinauer Associates.
Tate, G.H.H. 1931. Brief diagnoses of twenty-sixapparently new forms of Marmosa (Marsupia-lia) from South America. American MuseumNovitates 493: 1–14.
Tate, G.H.H. 1933. A systematic revision of themarsupial genus Marmosa with a discussion ofthe adaptive radiation of the murine opossums(Marmosa). Bulletin of the American Museumof Natural History 66: 1–250 1 26 pls.
Thomas, O. 1896. On new small mammals fromthe Neotropical region. Annals and Magazineof Natural History, Series 6, 18: 301–314.
Thomas, O. 1900. Descriptions of new Neotropi-cal mammals. Annals and Magazine of NaturalHistory, Series 7, 5: 217–222.
Thomas, O. 1911. New mammals from tropicalSouth America. Annals and Magazine of Nat-ural History, Series 8, 7: 513–517.
Thomas, O. 1913. New mammals from SouthAmerica. Annals and Magazine of Natural His-tory, Series 8, 12: 567–574.
Thomas, O. 1920. On mammals from the lowerAmazons in the Goeldi Museum, Para. Annalsand Magazine of Natural History, Series 9, 6:266–283.
Thomas, O. 1924. New South American mam-mals. Annals and Magazine of Natural History,Series 9, 13: 234–237.
Thomas, O. 1927a. The Godman-Thomas Expe-dition to Peru.—V. On mammals collected byMr. R.W. Hendee in the Province of San Mar-tin, N. Peru, mostly at Yurac Yacu. Annals andMagazine of Natural History, Series 9, 19:361–375.
Thomas, O. 1927b. The Godman-Thomas Expe-dition to Peru.—VI. On mammals from the up-per Huallaga and neighboring highlands. An-nals and Magazine of Natural History, Series 9,20: 594–608.
Tschudi, J. J. von. 1844–1846. Untersuchungenuber die Fauna Peruana. St. Gallen: Scheitlinund Zollikofer. [Published in parts; pp. 133–188, in which the descriptions of Marmosopsimpavidus and M. noctivagus occur, appearedin 1845 according to Sherborn, 1922: cxxiv]
Vieira, E.M., and E.L.A. Monteiro-Filho. 2003.Vertical stratification of small mammals in theAtlantic rainforest of south-eastern Brazil. Jour-nal of Tropical Ecology 19: 501–507.
Voss, R.S., and L.H. Emmons. 1996. Mammaliandiversity in Neotropical lowland rainforests: apreliminary assessment. Bulletin of the Amer-ican Museum of Natural History 230: 1–115.
Voss, R.S., and S.A. Jansa. 2003. Phylogeneticstudies on didelphid marsupials II. Nonmolec-

2004 37VOSS ET AL.: BOLIVIAN MARMOSOPS
ular data and new IRBP sequences: separateand combined analyses of didelphine relation-ships with denser taxon sampling. Bulletin ofthe American Museum of Natural History 276:1–82.
Voss, R.S., D.P. Lunde, and N.B. Simmons. 2001.The mammals of Paracou, French Guiana: aNeotropical rainforest fauna. Part 2. Nonvolantspecies. Bulletin of the American Museum ofNatural History 263: 1–236.
Voss, R.S., A.L. Gardner, and S.A. Jansa. 2004.On the relationships of ‘‘Marmosa’’ formosaShamel, 1930 (Marsupialia: Didelphidae), a
phylogenetic puzzle from the Chaco of northernArgentina. American Museum Novitates 3442:1–18.
Webster, G.L. 1995. The panorama of Neotropicalcloud forests. In S.P. Churchill, H. Balslev, E.Forero, and J.L. Luteyn (editors), Biodiversityand conservation of Neotropical montane for-ests: 53–77.New York: New York BotanicalGarden.
Wible, J.R. 2003. On the cranial osteology of theshort-tailed opossum Monodelphis brevicauda-ta (Didelphidae, Marsupialia). Annals of theCarnegie Museum 72: 137–202.
APPENDIX 1
TYPE LOCALITIES OF NOMINAL SPECIES
OF MARMOSOPS
Below we list the type localities of all nominaltaxa currently referred to the genus Marmosops,including the new species described herein. Itali-cized place names are those of the largest politicaldivision (state/department/province) within eachcountry. Unless reported by the original describer,geographic coordinates and elevation above sealevel are provided in square brackets with a citedsecondary source for these data. Numbered local-ities are mapped in figure 1.
BOLIVIA
1. La Paz, Rıo Solacama [ca. 168169S,678249W; 2300 m (Paynter, 1992)]. Typelocality of dorothea. Thomas (1911: 517)misspelled the type locality ‘‘Solocame’’and stated that the type was collected byP.O. Simons on 26 January 1901. The co-ordinates given above are those of Choca-chaca on the Rıo Tamampaya, where Si-mons is known to have camped at 1200 mfrom 10 to 29 January 1901 (Chubb, 1919:5). Presumably, Simons was running trap-lines from his base on the Tamampaya upthe valley of the nearby Solacama, but nodocumentary evidence is apparently avail-able to support this conjecture.
2. La Paz, Saynani hydroelectric plant(168079S, 688059W; 2500 m) on southeastside of the Zongo Valley. Type locality ofcreightoni.
3. La Paz, Rıo Unduavi, Pitiguaya, La Florida[ca. 168219S, 678479W; 1785 m (Anderson,1997)]. Type locality of yungasensis.
4. Santa Cruz, Buenavista [178279S, 638409W;ca. 400 m (Paynter, 1992)]. Type localityof ocellatus.
BRAZIL
5. Amapa, Rio Amapari, Serra do Navio[08599N, 528039W; ca. 100 m (Paynter andTraylor, 1991)]. Type locality of pinheiroi.
6. Amazonas, Huitanaa [78409S, 658469W; ca.50 m (Paynter and Traylor, 1991)]. Typelocality of purui, misspelled ‘‘Hyutana-ham’’ by Miller (1913).
7. Bahia, Lamarao [118459S, 388559W; 291 m(Paynter and Traylor, 1991)]. Type localityof bahiensis.
8. Mato Grosso, Aripuanan [78359S, 608269W;no elevation (Paynter and Traylor, 1991)].Type locality of stollei. The coordinates arethose at the confluence of the Rio Aripuanaand the Rio Roosevelt, where the collectorof the holotype (E. Stolle) is known to haveworked (Paynter and Traylor, 1991: 37).
9. Mato Grosso, 264 km N (by road) Xav-antina, Serra do Roncador (128519S,518469W; ca. 533 m). Type locality of bish-opi.
10. Minas Gerais, Rio das Velhas, near LagoaSanta [198389S, 438539W; 760 m (Paynterand Traylor, 1991)]. Type locality of in-canus.
–. Minas Gerais,? Porto Alegre [not located].Type locality of scapulatus. Burmeister(1856) only said that the type was collectedin Minas Gerais, but Matschie (1916) gavethe type locality as Porto Alegre, possiblya fazenda no longer in existence or toosmall to appear in maps and gazetteers.
11. Para, Nova Area Experimental, Utinga,Belem [18279S, 488299W; ca. 10 m (Paynterand Traylor, 1991)]. Type locality of wood-alli.
12. Para, Rio Tapajos, Vila Braga [48259S,568179W; ca. 100 m (Paynter and Traylor,

38 NO. 3466AMERICAN MUSEUM NOVITATES
1991)]. Type locality of collega, misspelled‘‘Villa Braga’’ by Thomas (1920: 281).
13. Rio de Janeiro, Teresopolis [228269S,428599W; 902 m (Paynter and Traylor,1991)]. Type locality of paulensis, spelled‘‘Therezopolis’’ by Tate (1931: 8).
COLOMBIA
14. 14. Antioquia, 9 km S Valdivia ([78069N,758289W; Anderson, 1999], elev. 1400 m).Type locality of handleyi.
15. Cundinamarca, Fusagasuga [48219N,748229W; 1746 m (Paynter, 1997)]. Typelocality of perfuscus.
16. Valle del Cauca, Rıo Cauquita, near Cali([38279N, 768319W; Paynter, 1997], 1000m). Type locality of caucae. Thomas(1900: 222) said the type was collected at‘‘Rio Caqueta, a tributary of the Cauca,near Cali’’, but see Tate (1933: 178).
ECUADOR
17. El Oro, Portovelo ([38439S, 798399W;Paynter, 1993], 610 m). Type locality oforoensis.
18. Loja, Celica ([48079S, 798599W; Paynter,1993], 2103 m). Type locality of celicae.
19. Napo, Coca [5 Puerto Francisco de Orel-lana; 08289S, 768589W, ca. 260 m (Paynter,1993)]. Type locality of politus.
20. Pichincha, Mindo ([08029S, 788489W;Paynter, 1993], 1280 m). Type locality ofsobrinus.
GUYANA
21. Demerara-Mahaica, Hyde Park [68309N,588169W; ca. 100 m (Stephens and Traylor,1985)]. Type locality of parvidens.
PANAMA
22. Darien, Cana ([78479N, 778429W; Fairchildand Handley, 1966], 610 m). Type localityof invictus.
PERU
23. Amazonas, Tambo Carrizal ([ca. 68479S,778529W; Stephens and Traylor, 1983], ca.2743 m). Type locality of leucastrus.
24. Amazonas, Tambo Ventija ([68119S,778339W; Stephens and Traylor, 1983], ca.2743 m), 10 mi E Molinopampa. Type lo-cality of madescens.
25. Cusco, Torontoy ([138109S, 728309W; Ste-phens and Traylor, 1983], 2438 m). Typelocality of albiventris.
26. Junın, Montana de Vitoc, near Chancha-mayo [ca. 118039S, 758199W; no elevation,but presumably .500 m (Stephens andTraylor, 1983)]. Alleged type locality ofimpavidus and noctivagus. AlthoughTschudi (1845) did not state exactly wherehis material was collected, Tate (1933:152–153) inferred that the type of nocti-vagus was collected here based on a pub-lished itinerary of Tschudi’s collecting ac-tivities. Cabrera (1958: 16) assumed thatthe type of impavidus came from the sameplace.
27. Junın, Utcuyacu ([ca. 118129S, 758289W;Stephens and Traylor, 1983], 1463 m).Type locality of juninensis.
28. Loreto, Yurimaguas ([58549S, 768059W;Stephens and Traylor, 1983], 183 m). Typelocality of neglectus.
29. Puno, Inca Mines ([ca. 138519S, 698419W;Stephens and Traylor, 1983], 1829 m).Type locality of keaysi. Allen (1900) statedthat the type was collected at Juliaca, butlater corrected himself (Allen, 1901).
30. San Martın, Yurac Yacu (ca. 762 m), about20 miles WNW of Moyobamba [68039S,768589W; Stephens and Traylor, 1983].Type locality of lugendus.
31. Ucayali, Rıo Ucayali, Lagarto [108409S,738549W; ,500 m (Stephens and Traylor,1983)]. Type locality of ucayalensis.
TRINIDAD AND TOBAGO
32. Trinidad, Caparo [108279N, 618209W; 46 m(Goodwin and Greenhall, 1961)]. Type lo-cality of carri.
VENEZUELA
33. Amazonas, Camp VII (08519N, 658589W;1800 m), Cerro de la Neblina. Type localityof neblina.
34. Falcon, 14 km ENE Mirimire, near La Pas-tora (118129N, 688379W; 150 m). Type lo-cality of cracens.
35. Merida, Rıo Albarregas ([probably nearMerida at 88369N, 718089W; Paynter,1982], 1630 m). Type locality of fuscatus,misspelled ‘‘Abbaregas’’ by Thomas(1896: 314).

2004 39VOSS ET AL.: BOLIVIAN MARMOSOPS
APPENDIX 2
GAZETTEER OF EXAMINED SPECIMENS OF
BOLIVIAN MARMOSOPS
This gazetteer includes all localities from whichwe personally examined Bolivian specimens ofMarmosops. Italicized place names are those ofcurrently recognized departments; boldface typeidentifies collection localities as they appear in thetext of this report. Unless recorded by the collec-tor, geographic coordinates and elevation abovesea level are provided in square brackets with acited secondary source for these data. The speciesname(s), name(s) of the collector(s), and date(s)of collection are separated from each locality en-try by a colon. Numbered localities are plotted infigures 10 and 12.
1. Beni, 1 km E La Embocada, Estancia laCabana [158039S, 668589W; 600 m (Ander-son, 1997)]: Marmosops bishopi and M.noctivagus (coll. G.K. Creighton, 10–13November 1979).
2. Beni, Rurrenabaque (148289S, 678349W;227 m): Marmosops noctivagus (coll. R.H.Pine, 1 September 1980).
3. Chuquisaca, Rıo Limon (198339S,648089W; 1300 m): Marmosops sp. (coll.R.L. Cuellar, 5 August 1990).
4. Cochabamba, Incachaca [178149S,658419W; 2347 m (Anderson, 1997)]: Mar-mosops noctivagus (coll. L.E. Miller andH.S. Boyle, 15 May 1915).
5. Cochabamba, Cordillera de Mosetenes(168149S, 668259W; 1300–1600 m): Mar-mosops bishopi (coll. E. Yensen, 3 Septem-ber 2003) and M. noctivagus (coll. D. Em-bert, F. Guerra, and T. Tarifa, 2–13 Septem-ber 2003).
6. Cochabamba, Tablas Monte, including4.4 km by road N (178049S, 668019W;1833–2100 m) and 9.5 km by road NE(178029S, 658599W; 1500 m): Marmosopsnoctivagus (coll. M.L. Campbell, S.L.Gardner, S.C. Peurach, and C.T. Seaton,13–17 July 1993).
7. La Paz, Iturraldi, Alto Rıo Madidi [5‘‘Moira Camp’’ or ‘‘Moire’’ in Iturraldeprovince; 138359S, 688469W; 270 m (An-derson, 1997)]: Marmosops bishopi (coll.L.H. Emmons, 27 May 1990).
8. La Paz, 20 km NNE Caranavi [158429S,678359W; 610 m (Anderson, 1997)]: Mar-mosops noctivagus (coll. G.K. Creighton, 8May 1978).
9. La Paz, Chijchijpa, 20 km by road fromCoroico (168099S, 678459W; 1114–1400m): Marmosops noctivagus (coll. S. An-
derson, H. Gomez, and R. Tejada, 6–8 July1992).
10. La Paz, 13 km SW Ixiamas (138539S,688159W; 400 m): Marmosops noctivagus(coll. L.H. Emmons, 2 June 1990).
11. La Paz, La Reserva (158439S, 678319W;940 m): Marmosops bishopi (coll. T. Tiu-rina, 22 July 1992) and M. noctivagus(coll. AMNH/MSB party, 28 July 1992).
12. La Paz, Rıo Unduavi, Pitiguaya, La Flor-ida [ca. 168219S, 678479W; 1785 m (An-derson, 1997)]: Marmosops noctivagus(coll. G.H.H. Tate, 9–14 May 1926; G.K.Creighton, 30 November 1979).
13. La Paz, Rıo Solacama [ca. 168169S,678249W; 2300 m (Paynter, 1992)]: Mar-mosops noctivagus (coll. P.O. Simons, 26June 1901). See appendix 1 for commentson this locality.
14. La Paz, Serranıa Bella Vista [158429S,678299W; 1300–1525 m (Anderson,1997)]: Marmosops noctivagus (coll. S.Anderson, 24 July 1992).
15. La Paz, Ticunhuaya [158289S, 688189W;1540 m (Anderson, 1997)]: Marmosopsnoctivagus (coll. G.H.H. Tate, 23 April1926).
16. La Paz, [Valle de Zongo,] Cuticucho ([5Cuti Khuchu at 168089S, 688079W; IGM,1967]; ‘‘2967 m’’): Marmosops creightoni(coll. G.K. Creighton, 29 September–1 Oc-tober 1979). There is some question aboutexactly where Creighton camped at Cuti-cucho. His field notes say 2967 m, but thisis oddly similar to the posted elevation onroad signs that now give the elevation ofCuticucho as 2697, so Creighton’s figuremight have resulted from an accidentaltransposition of two numerals. Modern to-pographic maps (IGM, 1967) give the el-evation at the road adjacent to the Cuticu-cho power plant as 2800 m.
17. La Paz, [Valle de Zongo,] El Vertigo(168059S, 688029W, 1800 m): Marmosopsnoctivagus (coll. F. Guerra, 15 August1993).
18. La Paz, [Valle de Zongo,] Kahua (ca.168029S, 678599W, 1150 m): Marmosopsnoctivagus (coll. T. Tarifa and M. Tarifa,23 January 1995). An alternative spellingon modern topographic maps is ‘‘Khaua’’.
19. La Paz, [Valle de Zongo,] Saynani(168079S, 688059W; 2500 m): Marmosopscreightoni (coll. E. Yensen, 5 August1999).
20. La Paz, [Valle de Zongo,] Cement Mine

40 NO. 3466AMERICAN MUSEUM NOVITATES
([?ca. 168059S, 688039W], 2000 m): Mar-mosops creightoni (coll. G.K. Creighton,4–11 October 1979).
21. Pando, Palmira (118429S, 678569W; 170m): Marmosops impavidus and M. nocti-vagus (coll. L.A. Ruedas, 4 August 1986).
22. Pando, Santa Rosa (128139S, 688249W;180 m): Marmosops noctivagus (coll. L.A.Ruedas, 1 August 1986).
23. Santa Cruz, 6 km by road W Ascencion(158439S, 638099W; 240 m): Marmosopsocellatus (coll. I.D. Mercado T., August1985).
24. Santa Cruz, Aserradero Pontons(178059S, 598349W; no elevation): Mar-mosops ocellatus (coll. H. Aranibar and J.Vargas, 19 April 1999).
25. Santa Cruz, Ayacucho [178519S, 638209W(Anderson, 1997); no elevation]: Marmo-sops ocellatus (coll. F. Bercerra and W.K.Kerr, 24 August 1966).
26. Santa Cruz, Buenavista ([178279S,638409W; Paynter, 1992], 450–500 m):Marmosops ocellatus (coll. J. Steinbach, 6August 1924).
27. Santa Cruz, 4.5 km N and 1.5 km E Cer-ro Amboro, Rıo Pitasama (178449S,638399W; 620 m): Marmosops ocellatus(coll. B.R. Riddle, 15 August 1985).
28. Santa Cruz, El Refugio (148469S, 618029W;ca. 200 m), Parque Nacional Noel KempffMercado: Marmosops bishopi (coll. L.H.Emmons, 6–15 November 1998) and M.ocellatus (coll. L.H. Emmons; 31 August–7 September 1999, 13 September 2000).
29. Santa Cruz, 3.5 km W Estacion El Pailon(178399S, 62845W; 300 m): Marmosopsocellatus (coll. S. Anderson, 22 September1984).
30. Santa Cruz, Hacienda El Pelicano, 3 kmN Zanja Honda (188169S, 638119W; 500m): Marmosops ocellatus (coll. M. Camp-bell, 4 July 1991).
31. Santa Cruz, 7 km E and 3 km N Ingeni-ero Mora ([188109S, 638169W; Anderson,1997] 488–579 m): Marmosops ocellatus(coll. P.A. Remsik, 10 August 1980).
32. Santa Cruz, Lago Caiman [138369S,608559W; ca. 200 m (Killeen and Schulen-berg, 1998)], Parque Nacional Noel
Kempff Mercado: Marmosops ocellatus(coll. E. Cuellar, 27 September 1995).
33. Santa Cruz, 2 km SW Las Cruces(178479S, 638229W; 480 m): Marmosopsocellatus (coll. S. Anderson, 5 July 1990).
34. Santa Cruz, Mangabalito [138559S,608309W; ca. 200 m (DMA, 1992)], ParqueNacional Noel Kempff Mercado: Marmo-sops ocellatus (coll. M.J. Swarner, 4 Oc-tober 2000). Spelled ‘‘Mangabalsinho’’ inDMA (1992); mapped by Killeen andSchulenberg (1998: fig. 1) closer to138509S.
35. Santa Cruz, 3 km SE Montero, 1 km NVilla Copacabana (178239S, 638159W; 250m): Marmosops ocellatus (coll. AMNH/MSB party, 25 June 1991).
36. Santa Cruz, Palmar [178519S, 638099W; noelevation (Anderson, 1997)]: Marmosopsocellatus (coll. D.R. Hadden, 13 August1966).
37. Santa Cruz, Rıo Ariruma, 7 km by roadSE Ariruma (188209S, 648139W; 1750 m):Marmosops sp. (coll. J.A. Cook, 5 June1991).
38. Santa Cruz, San Miguel Rincon (178239S,638329W; 300 m): Marmosops ocellatus(coll. N. Olds, 12 August 1984).
39. Santa Cruz, San Rafael de Amboro(178369S, 638369W; 400 m): Marmosopsbishopi (coll. B.R. Riddle, 31 July 1985).
40. Santa Cruz, 10 km N San Ramon(168369S, 628429W; 250 m): Marmosopsocellatus (coll. S. Anderson and N. Olds,7 August 1985).
41. Santa Cruz, 15 km S Santa Cruz(178539S, 638079W; 400 m): Marmosopsocellatus (coll. MSB parties, 22–25 July1987 and 16 June 1988; AMNH/MSB par-ty, 16 June 1991).
42. Santa Cruz, 27 km SE Santa Cruz(178589S, 638039W; 365 m): Marmosopsocellatus (coll. MSB party, 28 May 1988).
43. Santa Cruz, Santa Rosita [178319S,638119W; no elev. (Anderson, 1997)], 3 kmS Warnes: Marmosops ocellatus (coll.M.L. Kuns, 29 July 1965).
44. Santa Cruz, Tita (188259S, 628109W;300m): Marmosops ocellatus (coll.AMNH/MSB party, 19 October 1984).