an iron–sulfur cluster in the polymerase domain of yeast ... · an iron–sulfur cluster in the...
TRANSCRIPT

Communication
Rinku Jain1, E
0022-2836/$ - see front m
An Iron–Sulfur Cluster in the PolymeraseDomain of Yeast DNA Polymerase ε
va S. Vanamee1, Boris G.
Dzikovski 2, Angeliki Buku1,Robert E. Johnson3, Louise Prakash3, Satya Prakash3 and Aneel K. Aggarwal 11 - Department of Structural and Chemical Biology, Mount Sinai School of Medicine, Box 1677, 1425 Madison Avenue, New York,NY 10029, USA2 - Department of Chemistry and Chemical Biology, Cornell University, Ithaca, NY 14853, USA3 - Department of Biochemistry and Molecular Biology, University of Texas Medical Branch, 301 University Boulevard, Galveston,TX 77755-1061, USA
Correspondence to Aneel K. Aggarwal: Fax: +1 212 849 2456. [email protected]://dx.doi.org/10.1016/j.jmb.2013.10.015Edited by J. Berger
Abstract
DNA polymerase ε (Polε) is a multi-subunit polymerase that contributes to genomic stability via its roles inleading strand replication and the repair of damaged DNA. Polε from Saccharomyces cerevisiae is composedof four subunits—Pol2, Dpb2, Dpb3, and Dpb4. Here, we report the presence of a [Fe-S] cluster directly withinthe active polymerase domain of Pol2 (residues 1–1187). We show that binding of the [Fe-S] cluster ismediated by cysteines in an insertion (Pol2ins) that is conserved in Pol2 orthologs but is absent in thepolymerase domains of Polα, Polδ, and Polζ. We also show that the [Fe-S] cluster is required for Pol2polymerase activity but not for its exonuclease activity. Collectively, our work suggests that Polε is perhapsmore sensitive than other DNA polymerases to changes in oxidative stress in eukaryotic cells.
© 2013 Elsevier Ltd. All rights reserved.
The survival of all organisms depends on the abilityto faithfully replicate the genome. In eukaryotes, withgenomes ranging in size from millions to billions ofbase pairs, the bulk of DNA synthesis is carried out bythree polymerases [1,2]: Polα, Polδ, and Polε. Polαprimes the Okazaki fragments on the lagging strand,which are then elongated by Polδ. Polε is believed tobe the leading strand polymerase and, like Polδ,achieves fidelity via both accurate DNA polymeriza-tion and 3′ → 5′ proofreading exonuclease activities.Polα, Polδ, and Polε belong to the B-family of DNApolymerases and are conserved in all eukaryotes.Eukaryotes also possess a lower fidelity B-familypolymerase, Polζ, which promotes synthesis throughDNA lesions that block replication [3].The Saccharomyces cerevisiae Polα (Pol1, Pol12,
Pri1, Pri2), Polε (Pol2, Dpb2, Dpb3, Dpb4), Polδ (Pol3,Pol31,Pol32), andPolζ (Rev3,Rev7, Pol31, Pol32) aremulti-subunit polymeraseswith catalytic and regulatorysubunits [1,2,4]. The catalytic subunits Pol1, Pol2,Pol3, and Rev3 are modular with a large N-terminalexonuclease–polymerase (exo–pol) catalytic core
atter © 2013 Elsevier Ltd. All rights reserve
followed by a small metal binding domain at theC-terminus (CTD; Fig. 1a). In Pol2, an additionalinactive exo–pol module is observed between theN-terminus active exo–pol module and the CTD [5].Recent studies on eukaryotic B-family Pols haveestablished the presence of a [4Fe-4S] cluster in theCTDs of Pol3 and Rev3 [6–8]; though for the Pol1 andPol2 CTDs, there is some degree of uncertainty [6,8].An [Fe-S] cluster has also been found in the Pri2subunit of Polα [9–11]. Taken together, the observationof Fe-S clusters in eukaryotic DNA polymerases is partof emerging evidence on their importance in essentialcomponents of the nucleic acid processing machi-neries [12]. We show here that Polε contains a [Fe-S]cluster directly within its first active exo–pol catalyticcore and that this [Fe-S] cluster is necessary for itspolymerase activity.The first clue that the Polε catalytic core contained
a [Fe-S] cluster came during protein purification.We observed that samples of Pol2 containing onlythe exo–pol catalytic core (Pol2ΔCTD; residues 1–1187), purified from yeast or Escherichia coli cells,
d. J. Mol. Biol. (2014) 426, 301–308

302 Fe-S Cluster in the Polymerase Domain of Yeast DNA Polymerase ε
were yellowish-brown in color (Fig. 1b) and that thecolor was concentration dependent. UV-Vis spec-trum of Pol2ΔCTD exhibited a broad maximacentered at ~400 nm (Fig. 1c), suggesting thepresence of a [Fe-S] cluster. Biochemical analysiswith an iron-specific indicator (bathophenanthroline),which turned pink in the presence of Pol2ΔCTD butnot a control buffer, also suggested the presenceof non-heme iron in Pol2ΔCTD. The assay yielded astoichiometric molar ratio of 2:1 iron:protein for theE. coli expressed Pol2ΔCTD.Next, we analyzed the Fe-S cluster in Pol2ΔCTD
by extended X-ray absorption fine structure (EXAFS)spectroscopy, using synchrotron radiation at Broo-khaven National Laboratory (beamline X3B). EXAFSis a powerful technique for characterizing an elementand its coordination in a protein sample [13]. The
Fig. 1. Catalytic subunits of eukaryotic B-family polymerorganization in the catalytic subunit of eukaryotic B-family ppolymerase (exo–pol) domain is followed by ametal bindingC-tebetween its active N-terminal exo–pol domain and the CTD.polymerase domain of Pol2 is indicated in yellow. (b) Purified Pcolorless at the same concentration. The S. cerevisiae Pol2YRP654 (MATα ura3-52 trp1Δ leu2Δ1 his3-Δ200 pep4::HIS318 h. Both proteins were purified by affinity chromatography on gS-transferase tag by cleavage with PreScission protease, andcolumn (GE Healthcare). For purification from E. coli cells, yeastag at the N-terminus and purified by affinity chromatographyexchange (HiTrapQ; GE Healthcare) or affinity chromatographGE Healthcare) chromatography. (c) UV-Vis absorption spectrshoulder centered at 400 nm that is characteristic of Fe-S clustePol2ΔCTDmut. UV-Vis spectra were recorded under aerobicSpectrophotometer (Thermo Scientific).
EXAFS data clearly showed the presence of Fe inPol2ΔCTD. Figure 2a shows the Fourier transformEXAFS (FT-EXAFS) data and the best fit. The firstshell peak in the FT-EXAFS data corresponds toFe-S backscattering, and the second shell peak isthe result of Fe-Fe backscattering. Subsequentscans showed a reduction in the size of the Fe-Fepeak, which indicates oxidation. For this reason, weused only the first scan of each spot, merging a totalof three scans for data analysis. The data can bebest fit to a [4Fe-4S] cluster with an Fe-S distance of2.29 Å and a Fe-Fe distance of 2.72 Å. The fittingdata are summarized in Supplementary Table 1. Thefitting results are similar to those obtained for other[4Fe-4S] clusters with a reduced iron center [18]. Fitsto a [2Fe-2S] cluster gave consistently worse results,though the presence of a [2Fe-2S] cluster cannot
ase and characterization of Pol2 variants. (a) Domainolymerases. In each case, an N-terminal exonuclease–rminal domain (CTD). Pol2 has an inactive exo–polmoduleApproximate location of the cysteine-rich motif within theol2ΔCTD is yellowish-brown in color while Pol2ΔCTDmut isΔCTD and Pol2ΔCTDmut were expressed in yeast strainprb1Δ1.6R can1 GAL) by induction with 2% galactose forlutathione Sepharose 4B beads, removal of the glutathionefurther purification by size exclusion on a Superdex 200t Pol2ΔCTD and Pol2ΔCTDmut were expressed with a His6on a Ni-NTA agarose column (Qiagen), followed by ion
y on a Heparin column and size-exclusion (Superdex 200;a of Pol2ΔCTD and Pol2ΔCTDmut. Inset shows the broadr in the Pol2ΔCTD spectrum andmissing in the spectrum ofconditions at room temperature on a Nanodrop 2000c

303Fe-S Cluster in the Polymerase Domain of Yeast DNA Polymerase ε
entirely be ruled out based on the EXAFS dataalone.EPR analysis on Pol2ΔCTD [19] also suggested
the presence of [Fe-S] cluster (Fig. 2b). The EPRsignal of purified Pol2ΔCTD increased considerablyupon reduction with sodium dithionite. The dominantg-values of 1.98 and 2.023 observed in the EPRspectrum are consistent with the presence of either[4Fe-4S] or [2Fe-2S] cluster in Pol2ΔCTD (Fig. 2b).The spectrum was best observed at 12 K, as theintensity decreased with increasing temperature anddisappeared completely at 35 K. The temperaturedependence of the Pol2ΔCTD EPR spectrum ischaracteristic of [Fe-S] clusters, [4Fe-4S] in particular.We further confirmed the presence of iron in
Pol2ΔCTD by inductively coupled plasma (ICP)emission. ICP analysis indicated the presence of2.4 moles of iron per mole of protein and nosignificant amounts of zinc, cobalt, nickel, manga-nese, and magnesium (Table 1). The stoichiometryof less than 4 observed by ICP for Pol2ΔCTD hasalso been observed previously for other [4Fe-4S]proteins [11,20] and can be attributed to loss of ironduring aerobic purification and concentration.Together, our biochemical, EXAFS, EPR, and ICPanalyses provide strong evidence for the existenceof a [Fe-S] cluster in the first active polymerasedomain of Pol2.Pol2 is phylogenetically related to Pol1, Pol3, and
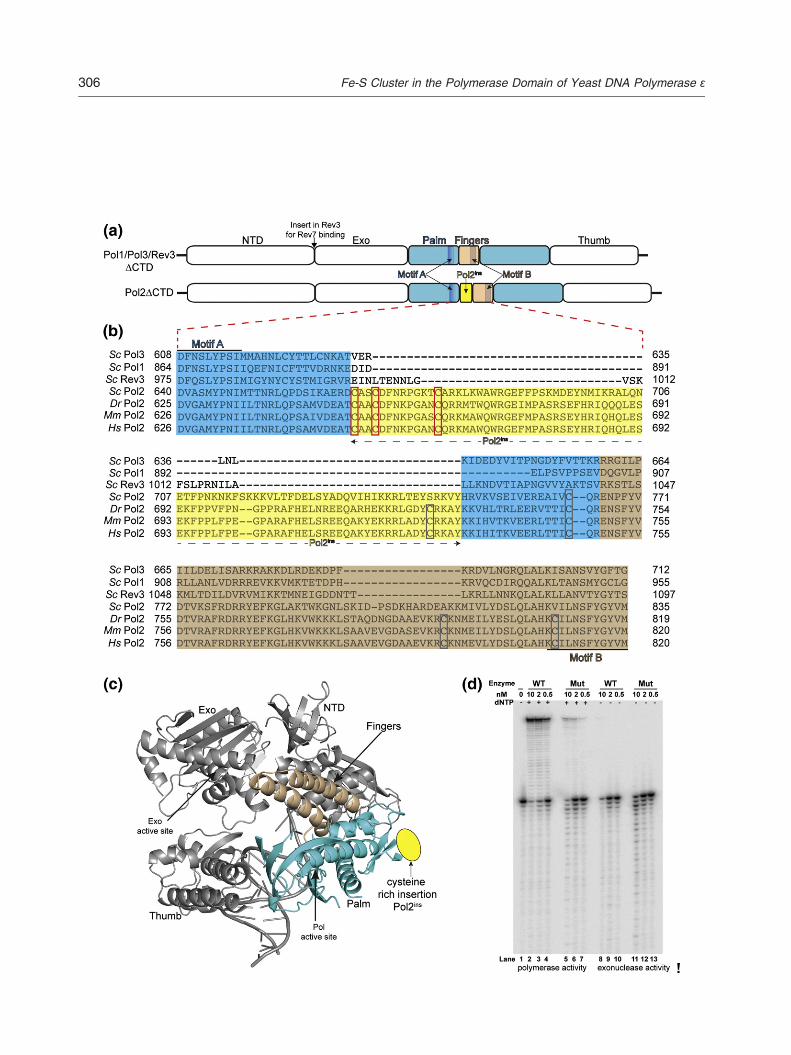
Rev3, which belong to the B-family of DNA poly-merases (Fig. 3a). Despite this homology, samplesof Pol1ΔCTD, Pol3ΔCTD, and Rev3ΔCTD purifiedover the years in our laboratory are colorless (datanot shown), suggesting that the Fe-S cluster bindingis specific to the polymerase domain of Pol2. Toidentify putative metal binding features in Pol2ΔCTDthat are absent from the other homologs, we under-took primary sequence alignment of the catalyticsubunits of eukaryotic B-family polymerases.Sequence alignment revealed a cysteine-rich inser-tion (Pol2ins) between motif A (in the palm domain)and motif B (in the fingers domain) that is conservedin Pol2 from different species (Fig. 3b) but is absentin Pol1, Pol3, and Rev3. Computational analysis withMetalDetector [22] for predicting metal binding sitessuggested four cysteines in the S. cerevisiaePol2ΔCTD (Cys665, Cys668, Cys677, andCys763) as putative metal binding ligands. To testthis prediction, we generated a Pol2ΔCTD mutantwith three of the putative Fe-S ligands (Cys665,Cys668, and Cys677) mutated to serine. Comparedto the wild-type Pol2ΔCTD, the Pol2ΔCTD triplemutant (Pol2ΔCTDmut) was colorless (Fig. 1b) andits UV-Vis spectrum lacked the broad peak centeredat 400 nm (Fig. 1c) that is indicative of a [Fe-S]cluster. Moreover, no significant metal content wasdetected by ICP analysis of Pol2ΔCTDmut at thesame concentration as the wild-type protein(Table 1). Together, these observations suggest
that cysteine residues in Pol2ins are involved in[Fe-S] cluster binding, and mutations to serine resultin impaired cluster incorporation. Also, the mutanthad a slightly higher tendency to aggregate than thewild-type protein, suggesting a role for the [Fe-S]cluster in Pol2 folding.To further examine the role of [Fe-S] cluster in
Pol2ΔCTD folding and enzymatic activity, we testedboth the wild type and the mutant protein for theirability to synthesize primers on a primer-templateduplex (Fig. 3d). As expected, Pol2ΔCTD wascatalytically efficient and able to insert nucleotideand extend the primer. In contrast to the wild-typePol2ΔCTD, Pol2ΔCTDmut was significantly deficientin DNA synthesis activity and unable to extend theprimer to the end of the template strand (Fig. 3d).However, it exhibited a slightly higher exonucleaseactivity, presumably due to the absence of any DNAsynthesis activity. These results suggest that the[Fe-S] cluster is important for the DNA synthesisactivity of Pol2 but has no effect on its exonucleaseactivity.Fe-S clusters are ancient cofactors associated
most commonly with enzymes that mediate electrontransfer during energy conversion [23,24]. Surpris-ingly, [Fe-S] clusters have also now been identifiedin nucleic acid processing enzymes such asglycosylases, helicases, and polymerases, whichhave no obvious roles in electron transfer [12]. The[Fe-S] clusters appear to play a structural role inthese enzymes, though there is evidence that someof these enzymes also use the redox properties ofthe Fe-S cluster to execute DNA transactions, frombinding and unwinding to the sensing of lesions [12].The [4Fe-4S] cluster in the CTD of eukaryotic Polδand Polζ helps to maintain the structural integrity ofthe second metal binding motif (CysB) for interac-tions with the regulatory B-subunit (Pol31) [6–8]. InPolα and Polε, the situation is more complex, withevidence for the presence of Fe or Zn in the CysB oftheir CTDs [6,8,25]. We show here that Polε containsan Fe-S cluster directly within its active N-terminalexo–pol catalytic core. The Fe-S cluster is coordi-nated by cysteines on an “insertion” that is con-served in Pol2 orthologs but is absent from thecatalytic subunits of Polα, Polδ, and Polζ.Although a crystal structure of the Pol2 catalytic core
remains to be determined, a reasonable model can beconstructed based on the crystal structure of the Pol3catalytic domain [21], composed of palm, fingers,thumb, and N-terminal domains for polymerase activityand an exonuclease domain for proofreading activity(Fig. 3a). From our model, the Fe-S cluster in Pol2maps to the “hinge” between the palm and fingersdomain, not too far from the polymerase active site tohave a direct effect on its polymerization activity(Fig. 3c). Interestingly, the [Fe-S] cluster is on theopposite side of the catalytic core as the exonucleasedomain (N60 Å away), which may explain why the loss

304 Fe-S Cluster in the Polymerase Domain of Yeast DNA Polymerase ε
of cluster binding correlates with loss of polymeraseactivity but has no effect on the exonuclease activity(Fig. 3d).Fe-S clusters do not assemble spontaneously but
require a set of proteins in the mitochondria [26].After synthesis in the mitochondria, some of the[Fe-S] clusters are exported to the cytoplasm, wherethe MMS19 protein has recently been shown to bepart of the complex that transfers [Fe-S] clusters totarget proteins [27,28]. Defects in both mitochondriaand MMS19 have been directly linked to nucleargenomic instability [27–29], and based on ourresults, a significant portion of this instability mayarise from the loss of the [Fe-S] cluster in the Polεcatalytic core. Polε contributes to genomic stabilityvia its role not only in leading strand replication butalso in the repair of damaged DNA [30]. We showhere that an [Fe-S] cluster stabilizes the catalyticcore of Polε for DNA synthesis. Also, the relativelysolvent-exposed position of the [Fe-S] cluster in the
hinge between the palm and fingers domain mayallow for interactions with other components of theeukaryotic replication machinery. Importantly, thesensitivity of the [Fe-S] cluster to oxidation mayprovide a means to couple DNA synthesis and repairby Polε to oxidative stress in eukaryotic cells.In conclusion, the finding of a [Fe-S] cluster in the
catalytic core of Pol2 changes our view of Polε. Itsuggests a polymerase that is perhaps moresensitive than others to mitochondrial dysfunction,defects in [Fe-S] biogenesis, and changes inoxidative stress in eukaryotic cells. Altogether, the[Fe-S] cluster in Pol2 adds to the growing evidenceon the importance of [Fe-S] clusters in nucleic acidprocessing machineries and provides a newimpetus for experiments on the correlation betweenDNA metabolism and the redox environment of acell.

Table 1. Element concentration by ICP emissionspectroscopy
Element [Fe]:[protein]Pol2ΔCTD
[Fe]:[protein]Pol2ΔCTDmut
Ca 0.03 0.04Co — 0.00Cu 0.02 0.01Fe 2.35 0.01Mg 0.01 0.00
305Fe-S Cluster in the Polymerase Domain of Yeast DNA Polymerase ε
Acknowledgements
We thank the staff at Brookhaven NationalLaboratory (beamline X3B) for help with EXAFSdata collection. The work was partially supported bygrants CA138546 and CA107650 from the U.S.National Institutes of Health. The EPR studies weresupported by National Institutes of Health grantP41GM103521.
Mn 0.00 0.00Mo — —Ni 0.03 0.13Zn 0.07 0.06
Metal content of Pol2ΔCTD and Pol2ΔCTDmut was determined byICP at the University of Georgia Chemical Analysis Laboratory.Proteins were concentrated to 16 mg/ml in buffer containing25 mM Tris–HCl (pH 8.0), 2 mM tris(2-carboxyethyl)phosphine,5% glycerol, and 400 mM NaCl, and the filtrate from theconcentration step was used as a blank. The concentrations of20 different elements were determined simultaneously. Ratios forsome metals that are commonly found in metalloenzymes areshown.
Appendix A. Supplementary data
Supplementary data to this article can be foundonline at http://dx.doi.org/10.1016/j.jmb.2013.10.015
Received 6 September 2013;Accepted 8 October 2013
Available online 19 October 2013
Keywords:DNA replication;
DNA polymerase;DNA repair;Fe-S cluster;
oxidative stress
Fig. 2. Spectroscopic analysis of Pol2ΔCTD. (a) The FT-EXA(red line). Pol2ΔCTD samples were concentrated to a final concesamples were loaded into a 0.5-mm-thick copper sample holderone side with an Fe-free Kapton tape. Each hole was filled with ~data were collected at the National Synchrotron Light Source ofocused Si(111) double crystal monochromator. A Ni mirror at ancontamination. All experiments were carried out at 15 K in a ccollected with 0.05 Å−1 step sizes in k space starting at 25 eV phrespect to k so that 2 s per point was used at k = 1 and 7 s per pocounting at a specific energy for 1 s and incrementing the energyedge, then in 0.3 eV steps up to 25 eV above the edge with 1 s pdegradation, the X-ray beam was moved on the sample holder,sample holes. Data were taken in the range 180- to 300-mA beamthe incident beam using a 31-element, energy-resolving Ge dete30,000/s per channel to avoid saturation effects in the detector. A cspectrum of a Fe foil simultaneously with all sample spectra. Tcalibration of the sample spectra; the K-edge inflection point assigEXAFS program suite Demeter (v. 0.9.9) that contains theIBM-compatible machine running under Microsoft Windows XPpolynomial spline background subtraction, wavevector cubed wATHENA. The theoretical data were generated using the ab initiextracted fromknowncrystal structures deposited in theProteinDsecond shell [4Fe-4S] contribution from PDB ID 1EYT and the firThe theoretical data were fit to the unfiltered experimental k3 weiiFEFFIT implemented in ARTEMIS by fixing the coordination numalso performs error analysis and calculates goodness-of-fit pabacktransform and fitting. (b) EPR spectra of Pol2ΔCTD after treindicated. For EPR spectrum acquisition, Pol2ΔCTD purified adithionite. Spectra were recorded on BRUKER ELEXSYS E5000.63 mW, and modulation amplitude of 6G. Data were acquiredInstruments liquid helium cryostat. The field sweeps were calibratfrequency was monitored with a frequency counter. Data acquis
Abbreviations used:EXAFS, extended X-ray absorption fine structure;
FT-EXAFS, Fourier transform EXAFS; ICP, inductivelycoupled plasma.
FS data of Pol2ΔCTD (blue line) superimposed on the best fitntration of 370 μM as judged by absorbance at 280 nm. Thecontaining three 4.5 mm × 12 mm holes that were sealed on50 μl of sample. Fe K-edge X-ray absorption spectroscopic
n beamline X3B. The beamline is equipped with a sagittallyangle of 4.5 mrad was used to reject higher-order harmoniclosed cycle He cryostat under vacuum. EXAFS data wereotoelectron energy. The signal averaging was weighted withint was used at k = 15. Below 25 eV, data were collected byby 10 eV from 200 eV below the iron edge to 10 eV below theer point signal averaging. To reduce the possibility of sampleand 3–5 scans were collected on each of the three differentcurrent. Fe K-α fluorescencewas detected at 90° angle fromctor [14]. The internal count rates were kept at or less thanalibration channel behind the samplewas set up to detect thehis calibration channel provided a reference for the energyned as 7112 eV. EXAFS data analysis was performed by theprograms ATHENA, ARTEMIS [15,16], and FEFF on an. Data manipulation with use of a linear pre-edge fit, cubiceighting, and Fourier transformation were performed usingo code FEFF (v.6) [17]. The coordinates of iron models wereataBank using a 6-Å radius around theFe(II) ion. The first andst and second shell [2Fe-2S] contribution from PDB ID 1F37.ghted χ data using the non-linear, least-squares method singber (N) and letting all other parameters to float. The programrameters. An R window of 1–3 was used for the Fourieratment with sodium dithionite at 12 K and 35 K; g-values areerobically was reduced by the addition of 10 mM sodiumEPR spectrophotometer at 9.32 GHz, microwave power offrom frozen glasses at 12 K or 30 K using ESR910 Oxforded with a BRUKERER 035Gauss meter, and the microwaveition and manipulation were performed with Xepr software.

307Fe-S Cluster in the Polymerase Domain of Yeast DNA Polymerase ε
References
[1] Johnson A, O'Donnell M. Cellular DNA replicases: compo-nents and dynamics at the replication fork. Annu RevBiochem 2005;74:283–315.
[2] Johansson E, Macneill SA. The eukaryotic replicative DNApolymerases take shape. Trends Biochem Sci 2010;35:339–47.
[3] Prakash S, Johnson RE, Prakash L. Eukaryotic translesionsynthesis DNA polymerases: specificity of structure andfunction. Annu Rev Biochem 2005;74:317–53.
[4] Johnson RE, Prakash L, Prakash S. Pol31 and Pol32subunits of yeast DNA polymerase delta are also essentialsubunits of DNA polymerase zeta. Proc Natl Acad Sci U S A2012;109:12455–60.
[5] Tahirov TH, Makarova KS, Rogozin IB, Pavlov YI, KooninEV. Evolution of DNA polymerases: an inactivated polymer-ase-exonuclease module in Pol epsilon and a chimeric originof eukaryotic polymerases from two classes of archaealancestors. Biol Direct 2009;4:11.
[6] Baranovskiy AG, Lada AG, Siebler HM, Zhang Y, Pavlov YI,Tahirov TH. DNA polymerase delta and zeta switch bysharing accessory subunits of DNA polymerase delta. J BiolChem 2012;287:17281–7.
[7] Makarova AV, Stodola JL, Burgers PM. A four-subunit DNApolymerase zeta complex containing Pol delta accessorysubunits is essential for PCNA-mediated mutagenesis.Nucleic Acids Res 2012;40:11618–26.
[8] Netz DJ, Stith CM, Stumpfig M, Kopf G, Vogel D, Genau HM,et al. Eukaryotic DNA polymerases require an iron-sulfurcluster for the formation of active complexes. Nat Chem Biol2012;8:125–32.
[9] Klinge S, Hirst J, Maman JD, Krude T, Pellegrini L. An iron-sulfur domain of the eukaryotic primase is essential for RNAprimer synthesis. Nat Struct Mol Biol 2007;14:875–7.
[10] Sauguet L, Klinge S, Perera RL, Maman JD, Pellegrini L.Shared active site architecture between the large subunit ofeukaryotic primase and DNA photolyase. PLoS One 2010;5:e10083.
[11] Weiner BE, Huang H, Dattilo BM, Nilges MJ, Fanning E,Chazin WJ. An iron-sulfur cluster in the C-terminal domain
Fig. 3. A cysteine-rich insertion in Pol2. (a) Schematic of the epolymerases. The catalytic core consists of the N-terminal, palmexonuclease domain for proofreading activity. Approximate locdomains, respectively, is indicated. A cysteine-rich insertion (Ponot withinPol1, Pol3, andRev3. (b) Sequence alignment of the reand Rev3 with Pol2 orthologs. Sc, S. cerevisiae; Dr, Danio reriregions betweenmotifs A and B are shown. Palm domain aminothe fingers domain, in wheat. Cysteines mutated to serine in thisboxes. Other semi-conserved cysteines are highlighted in gray.using the program PROMALS3D, and the alignments were visugenerated by using the structure of Pol3ΔCTD (PDB ID 3IAY) aandwheat, respectively. Approximate locationPol2ins in the hingeyellow oval. (d) Pol2ΔCTDmut (C665S, C668S, and C677S) lacDNA template-primer substrate only. DNA synthesis activity of Ppresence of all four dNTPs. Exonuclease activity of Pol2ΔCTDThe DNA polymerase assay (5 μl) contained 25 mM Tris–HCl (albumin, 10 nMDNA substrate, and 10 μMeach of dGTP, dATPvarious concentrations of DNA polymerase and were carried oaddition of 30 μl of 95% formamide containing 0.03% each of bdenatured by heating to 95 °C for 3 min and resolved by separawere dried and reaction products were visualized and quantifiedIQTL (version 8.0; GE Life Sciences) software.
of the p58 subunit of human DNA primase. J Biol Chem2007;282:33444–51.
[12] White MF, Dillingham MS. Iron-sulphur clusters in nucleic acidprocessing enzymes. Curr Opin Struct Biol 2012;22:94–100.
[13] Strange RW, Feiters MC. Biological X-ray absorptionspectroscopy (BioXAS): a valuable tool for the study oftrace elements in the life sciences. Curr Opin Struct Biol2008;18:609–16.
[14] Cramer SP, Chen E, George SJ, van Elp J, Moore J, TeschO. Soft X-ray spectroscopy of metalloproteins using fluores-cence detection. Nucl Instrum Methods Phys Res Sect A1992;319:289–95.
[15] Newville M. IFEFFIT: interactive XAFS analysis and FEFFfitting. J Synchrotron Radiat 2001;8:322–4.
[16] Ravel B, Newville M. ATHENA, ARTEMIS, HEPHAESTUS:data analysis for X-ray absorption spectroscopy usingIFEFFIT. J Synchrotron Radiat 2005;12:537–41.
[17] Rehr JJ, Mustre de Leon J, Zabinsky SI, Albers RC.Theoretical X-ray absorption fine structure standards. J AmChem Soc 1991;113:5135–40.
[18] Mitra D, George SJ, Guo Y, Kamali S, Keable S, Peters JW,et al. Characterization of [4Fe-4S] cluster vibrations andstructure in nitrogenase Fe protein at three oxidation levelsvia combined NRVS, EXAFS, and DFT analyses. J AmChem Soc 2013;135:2530–43.
[19] Prisner T, Rohrer M, MacMillan F. Pulsed EPR spectro-scopy: biological applications. Annu Rev Phys Chem2001;52:279–313.
[20] Hinks JA, Evans MC, DeMiguel Y, Sartori AA, Jiricny J, PearlLH. An iron-sulfur cluster in the family 4 uracil-DNAglycosylases. J Biol Chem 2002;277:16936–40.
[21] SwanMK, Johnson RE, Prakash L, Prakash S, Aggarwal AK.Structural basis of high-fidelity DNA synthesis by yeast DNApolymerase delta. Nat Struct Mol Biol 2009;16:979–86.
[22] Lippi M, Passerini A, Punta M, Rost B, Frasconi P.MetalDetector: a Web server for predicting metal-bindingsites and disulfide bridges in proteins from sequence.Bioinformatics 2008;24:2094–5.
[23] Brzoska K, Meczynska S, Kruszewski M. Iron-sulfur clusterproteins: electron transfer and beyond. Acta Biochim Pol2006;53:685–91.
xo–pol module in the catalytic subunit of eukaryotic B-family, fingers, and thumb domains for polymerase activity; and anation of conserved motifs A and B in the palm and fingersl2ins) is present between motifs A and B of Pol2 (yellow) butgion betweenmotif A andmotif B forS. cerevisiaePol1, Pol3,o; Mm, Mus musculus; Hs, Homo sapiens. For clarity, onlyacids are highlighted in cyan; Pol2ins residues, in yellow; andstudy and conserved in Pol2 orthologs are highlighted in redAmino acid sequences of Pol2 and homologs were alignedalized and formatted with JalView. (c) Model of Pol2ΔCTDs template [21]. Palm and fingers domain are shown in cyanregion between the palmand fingers domains is shownby a
king the Fe-S cluster is deficient in DNA synthesis. Lane 1:ol2ΔCTD (lanes 2–4) and Pol2ΔCTDmut (lanes 5–7) in the
(lanes 8–10) and Pol2ΔCTDmut (lanes 11–13), respectively.pH 7.5), 10% glycerol, 1 mM DTT, 0.1 mg/ml bovine serum, dTTP, and dCTP. Reactionswere initiated by the addition ofut for 10 min at 30 °C. Reactions were terminated by theromphenol blue and xylene cyanol. Reaction products weretion on 12% polyacrylamide gels containing 8 M urea. Gelson a Storm 860 phosphorsimager (Molecular Dynamics) and

308 Fe-S Cluster in the Polymerase Domain of Yeast DNA Polymerase ε
[24] Johnson DC, Dean DR, Smith AD, Johnson MK. Structure,function, and formation of biological iron-sulfur clusters. AnnuRev Biochem 2005;74:247–81.
[25] Klinge S, Nunez-Ramirez R, Llorca O, Pellegrini L.3D architecture of DNA Pol alpha reveals the functionalcore of multi-subunit replicative polymerases. EMBO J2009;28:1978–87.
[26] Lill R, Muhlenhoff U. Maturation of iron-sulfur proteins ineukaryotes: mechanisms, connected processes, and dis-eases. Annu Rev Biochem 2008;77:669–700.
[27] Stehling O, Vashisht AA, Mascarenhas J, Jonsson ZO,Sharma T, Netz DJ, et al. MMS19 assembles iron-sulfur
proteins required for DNA metabolism and genomic integrity.Science 2012;337:195–9.
[28] Gari K, Leon Ortiz AM, Borel V, Flynn H, Skehel JM, BoultonSJ. MMS19 links cytoplasmic iron-sulfur cluster assembly toDNA metabolism. Science 2012;337:243–5.
[29] Veatch JR, McMurray MA, Nelson ZW, Gottschling DE.Mitochondrial dysfunction leads to nuclear genomeinstabi l i ty via an iron-sulfur cluster defect. Cell2009;137:1247–58.
[30] Pursell ZF, Kunkel TA. DNA polymerase epsilon: a poly-merase of unusual size (and complexity). Prog Nucleic AcidRes Mol Biol 2008;82:101–45.