and aug. vol. isolation regulatory mutants aspartic ...aac.asm.org/content/4/2/73.full.pdf ·...
TRANSCRIPT

ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Aug. 1973, p. 73-79Copyright © 1973 American Society for Microbiology
Vol. 4, No. 2Printed in U.S.A.
Isolation of Regulatory Mutants of the Asparticand Pyruvic Acid Families and Their Effect on
Antibiotic Production in Streptomyces lipmaniiOTIS W. GODFREY
The Lilly Research Laboratories, Eli Lilly and Company, Indianapolis, Indiana 46206
Received for publication 26 February 1973
Streptomyces lipmanii L100 produces two g-lactam ring-containing antibiot-ics: penicillin N and 7-(5-amino-5-carboxy valeramido)-7-methoxy-cephalospo-ranic acid. These antibiotics are modified tripeptides derived from a-aminoadipicacid, cysteine, and valine. Mutants defective either in the biosynthesis or in thecontrol of amino acids belonging to the aspartic and pyruvic acid families wereisolated and characterized with regard to their effect on antibiotic synthesis. In anumber of these mutants antibiotic synthesis was either completely inhibited orincreased up to threefold over wild-type levels.
Since the discovery of the ,B-lactam antibiot-ics, penicillin and cephalosporin C (1, 5), thechemistry, biological activity, and biosynthesisof these compounds have been investigatedextensively. Nagarajan et al. (13) have reportedthe occurrence of three new p-lactam antibioticsfrom two species of Streptomyces. Structureelucidation studies have shown these com-pounds to be chemically related to cephalo-sporin C.Streptomyces lipmanii L100 produces two
,B-lactam antibiotics, penicillin N and 7-(5-amino-5-carboxy valeramido)-7-methoxy-cepha-losporanic acid (8, 13). These antibiotics aremodified tripeptides derived from a-aminoadipicacid, cysteine, and valine (2, 3, 16). In a prokary-otic microorganism such as a Streptomycete, a-aminoadipic and cysteine are directly related tothe biosynthesis of the amino acids present inthe aspartic acid family of amino acids, whereasvaline is derived from pyruvate. Cysteine reactswi,th succinyl homoserine to produce cystathio-nine for methionine biosynthesis (10) and a-aminoadipic acid in this organism is likely aproduct of lysine catabolism (12).
Since all three precursors of the antibiotic arein families that have been amenable to geneticanalysis, regulatory mutants in this pathwaywere isolated and examined for their effect(s) onantibiotic production. Amino acid starvation ofauxotrophs, prototrophic revertants, and ana-logue resistant mutants were utilized in thisstudy.The aim of this communication is to describe
a large array of regulatory mutants and auxo-trophs in an effort to determine whether any
73
particular class or type will have any profoundeffects on antibiotic biosynthesis.
MATERIALS AND METHODSCultures. The wild-type strain of S. lipmanii
(NRRL 3584) was isolated from a South American soilsample by a selective isolation procedure (8). Thisculture is prototrophic and shall be referred to in thisreport as L100. The two major antibiotics produced bythis strain are penicillin N and 7-(5-amino-5-carboxyvaleramido)-7-methoxy-cephalosporanic acid (13).
Other strains derived from L100 used in the courseof this study are described in a previous publication(6). As an aid for mutant classification each strainwas designated by two letters followed by a number.The first letter L refers to the Lilly Laboratories andthe second letter refers to the type of mutant, i.e., LCis a strain that overproduces a primary metabolite,LM is a strain that overproduces a secondary metabo-lite, LA is an auxotroph, and LE is a revertant of anauxotroph that overproduces a primary metabolite. Inmicrobiological assays for antibiotic activity, Lillyculture X142, a strain of Salmonella gallinarum, wasused. A strain of Pseudomonas solanacearum (60-RE-3B) Lilly culture X185 was used to preparebioautograms.
Media. The JS7 minimal medium contained, perliter of distilled deionized water: K2HPO4, 1 g;(NH4) 2SO4, 5 g; NaCI, 1 g; MgSO4, 1 g; FeSO4 7H20,1 mg; MnCl24H20, 1 mg; ZnSO4.7H20, 1 mg;CaCo3, 4 g; maltose, 4 g; glucose, 1 g; glycerol, 0.2 g;L-glutamic acid, 1 g; glycine, 1 g; and agar at 15 g forsolid media. The concentrations of maltose, glucose,and glycerol were increased fivefold for liquid mini-mal medium. The pH value of this medium wasadjusted to 6.8 prior to sterilization. Salts, carbohy-drates, amino acids, and agar were autoclaved sepa-rately and combined aseptically. The fragmentationmedium contained identical salts and carbohydrates
on May 16, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

ANTIMICROB. AG. CHEMOTHER.
as the JS7 medium. The amino acids were replacedby: Difco Casamino Acids, 6 g; yeast extract, 0.5 g;adenine, 5 mg; guanine, cytosine, and thymine, 10 mgeach; and tyrosine, phenylalanine, tryptophan, andanthranilic acid at 100 mg each.The vegetative medium contained, per liter of
distilled deionized water: tryptone (Difco), 5 g;yeast extract, 5 g; glucose, 5 g; dextrin 700, 10 g;MgSO4, 2 g; and glycerol, 1 ml. Sporulation mediumcontained per liter: Contadina tomato paste, 20 g;Gerber baby oatmeal, 20 g; and agar (Difco), 20 g.The pH was adjusted to 7.2 prior to sterilization asdescribed by R. L. Hamill et al. (Abst. Intersci. Conf.Antimicrob. Ag. Chemother., 11th, p. 7, 1971).
The defined fermentation medium contained, perliter of distilled deionized water: NaCl, 1 g; CaCO3, 4g; KH2PO4, 0.2 g; MgSO4, 0.4 g; FeSO4-7H 0, 1 mg;MnCl2-4H20, 1 mg; ZnSO4-7H20, 1 mg; maltose, 20g; glucose, 10 g; glycerol, 1 g; DL-methionine, 0.5 g;L-tryptophan, 1 g; DL-lysine, 4 g; L-valine, 1 g. The pHwas adjusted to 6.8 prior to sterilization with aminoacids and carbohydrates sterilized separately (6).
Preservation of cultures. All cultures were main-tained in liquid nitrogen (vapor phase) and as lyQphi-lized ampoules stored at 5 C. For liquid nitrogenpreservation, a 2-ml sample of an overnight culturegrown in the vegetative medium was added to anequal volume of the suspending agent (20% glyceroland 10% lactose) and immediately placed in liquidnitrogen. When required for use, these cultures werequick-thawed at 45 C (personal communication C.Higgens).
Fermentation. All cultures were grown in wide-mouth 250-ml Erlenmeyer flasks containing 50 ml offermentation medium and incubated at 25 C for 96 hon a gyrotory shaker with a 5-cm stroke at 250 rpm.
Antibiotic assays. After fermentation, myceliawere removed from flasks containing S. lipmanii bycentrifugation. The antibiotic titer of the supernatantbroth was determined by the standard disk-platetechnique with X142 as the assay organism. Antibi-otic activity was recorded as units per milliliter ofculture broth. One unit is equal to 0.33 pg of purifiedantibiotic. Activity of the two antibiotics could bemeasured simultaneously or differentially by selec-tively destroying penicillin N with penicillinase(Riker Laboratories, Northridge, Calif.).
Isolation of analogue resistant mutants. A brothculture of L100 was grown to mid-log phase invegetative medium supplemented with 100 pg ofcytidine per ml. The pH of the medium was thenadjusted to 7.8 with 4N NaOH, and nitrosoguanidine(NTG) was added to a final concentration of 180pAg/ml. The culture was incubated for 20 min at 26 Con a rotary shaker (5-cm stroke) at 250 rpm. Theculture was centrifuged and resuspended in freshvegetative medium and incubated under similar con-ditions for an additional 16 h (6). An alternatetechnique was used when dimethyl sulfate (DMS) wasused as a mutagen. A log-phase culture was exposedto a 10% concentration (vol/vol) of the mutagen for 60min at 26 C with shaking after which the culture waswashed and resuspended in fresh vegetative mediumand incubated an additional 16 h.
After allowing for segregation, the mutated culture(20 ml) was centrifuged and resuspended in 5 ml ofJS7 minimal salts (pH 6.6), and 0.2-ml samples wereplated in minimal medium (JS7) containing thedesired amino acid analogue. The concentrations ofanalogues are given in Table 2.Paper chromatography and bioautograms. The
two major antibiotic factors in the S. lipmanii fermen-tation were resolved by descending chromatographyon Whatman no. 1 paper. The chromatograms weredeveloped in propanol-pyridine-acetic acid CHsCN-water (45:30:9:40:36 vol/vol) for 16 h at 25 C.Bioautograms were prepared by applying developedchromatograms to the surface of agar plates seededwith culture X185.
Quantitation of excreted amino acids. The cul-tures were centrifuged 3,000 x g for 10 min, and 2 mlof the supernatant fluid was collected and dried invacuo. The dried samples were dissolved in buffer (2%thiodiglycol, 0.2 N Na+ citrate, pH 2.2) and analyzedon a Beckman model 120C amino acid analyzerequipped with A BioCal model BC 501 automaticsample injector. Amino acid separation was achievedon a column of Durrum DC-1A resin (0.9- by 52-cm)at 56.5 C, with a 70 iml/h flow of buffer (0.5%thiodiglycol, 0.2 N Na+ citrate, pH 3.23). A ninhydrinflow of 35 ml/h was used.
Chemicals. NTG and dimethyl sulfate were ob-tained from Aldrich Chemical Co., Inc. The aminoacids used in this study were shown to be chromato-graphically pure by two dimensional chromatographyon thin-layer chromatography (TLC) plates. All othercompounds were of reagent grade.
RESULTS
Amino acid limitation of auxotrophs. Rep-resentative strains of each auxotroph weregrown in the defined fermentation mediumsupplemented with 100 to 1,000 gg of therequired amino acid per ml. After four days ofincubation the cultures were assayed both forantibiotic potency and growth. These data aresummarized in Table 1.With the lys, cys, and ilv mutants antibiotic
synthesis could be separated from primary me-tabolism by limiting the levels of the supp4e-mented amino acids. For example, LA 423 (1 is)grew normally (7%) mycelial volume) but failedto produce detectable antibiotic when the L-lysine supplementation was 200 jig/ml. Increas-ing the exogenous levels of lysine to 1 mg/mlresulted in a concomitant increase in antibioticyield.When LA 441 (ilv) was supplemented with
500 ,g of valine per ml and 240 jg of L-isoleucineper ml, the metabolism was shifted towards thepreferential synthesis of the cephalosporins.Both factors were produced when the valineconcentration was increased to 1 mg/ml. Themet mutant (LA 442) failed to synthesize anti-
74 GODFREY
on May 16, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

ANTIBIOTIC PRODUCTION IN STREPTOMYCES
TABLE 1. Amino acid limitation of auxotrophs
L-Amino acid Fermentation D-Amino acidAuxotroph Strain no. suppi. supplied Growthd
(mg/ml) Growtha Potencyb Chroma- (mg/ml)
lys LA423 1.0 8 100 AB 1.0 0.50.2 7 NA 0.2 0.5
L100 1.0 9 100 AB 1.0 50.2 8 50 AB 0.2 4
ilv LA467 1.Oe 11 100 AB 1.0 10.2 12 NAf 0.2
LA441 1.0 12 100 AB 1.0 10.5 11 100 A 0.5
L100 1.0 12 100 AB 1.0 120.2 12 100 AB 0.2 10
cys LA425 1.0 12 100 AB 1.0 120.1 8 NA 0.1 9
L100 1.0 11 100 AB 1.0 120.1 12 130 AB 0.1 10
met LA442 1.0 13 NA 1.0 12L100 1.0 12 100 AB 1.0 12
leu LA473 1.0 14 100 AB 1.0 120.5 11 290 AB 0.5 10
L100 1.0 10 100 AB 1.0 100.5 10 100 AB 0.5 10
a Growth refers to percentage of packed mycelial volume.b Potency refers to the concentration of methoxycephalosporin C measured as units per milliliter on penase
plates seeded with X142.c Chromatography: A, methoxycephalosporin C; B, penicillin N.d Growth refers to the percentage of packed mycelial volume obtained when the L-amino acid was replaced by
the respective D-isomer.e L-isoleucine concentration was kept constant at 240 mg/ml in all instances.f NA, no activity.
biotic at any level of methionine supplementa-tion. However, weak antibiotic activity could bedemonstrated when the medium was supple-mented with a combination of methionine andcysteine.
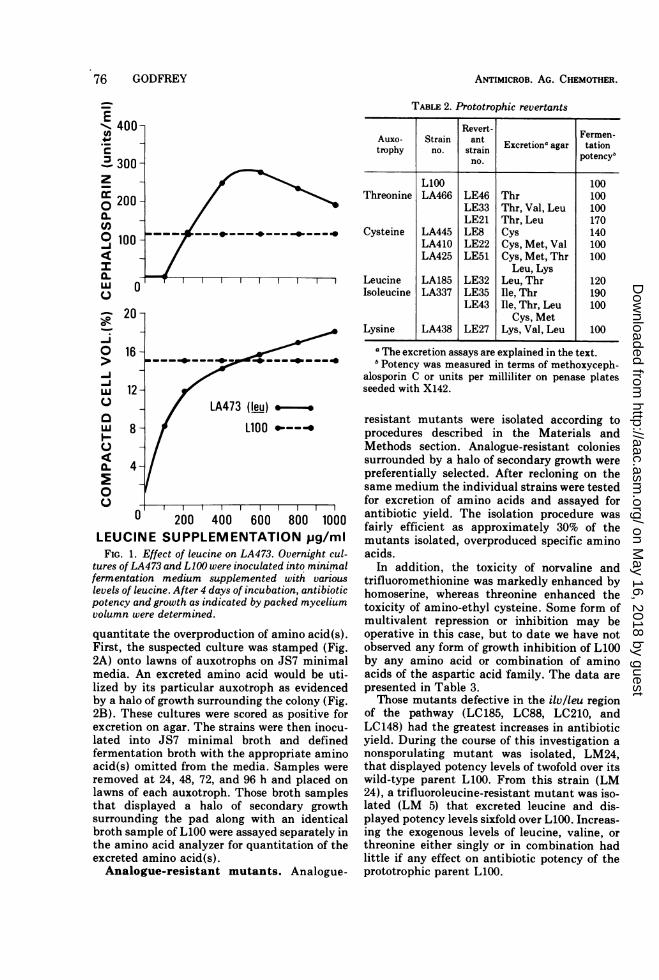
In the case of the leucine mutant, LA 473, thesituation was somewhat different. Growth atlimiting levels of leucine supplementation pro-duced a marked stimulation in antibiotic po-tency as shown in Fig. 1. Although derepressionof the ilv-leu pathway would explain this obser-vation, it was not possible to demonstrateincreased levels of isoleucine or valine in thefree amino acid pools.
In Escherichia coli both cys and met mutantsare capable of using, respectively, D-cysteineand D-methionine for growth. All other aminoacid mutants normally require the appropriateL form (11). However, with S. lipmanii the cys,met, and leu mutants are capable of usingeither isomer for growth, whereas the lys and ilvmutants require the appropriate L-form (Table3). Since aAAA is in the D form in bothantibiotics produced by L100 (13) an effort was
made to obtain mutants of LA423 that coulduse the D isomer. However, of the 40 nmutantsisolated that could grow on D-lysine as a substi-tute for the L isomer all were prototrophic.
Prototrophic revertants. In a previous paper(6) the procedures for the isolation and subse-quent reversion of auxotrophs of S. lipmaniiwere described. Prototrophic colonies thus ob-tained were examined after 3 days of incubationat 26 C. In a few cases colonies were foundsurrounded by a halo of secondary growth of theparental auxotrophs. Such colonies werestreaked on JS7 minimal medium, isolated, andtested both for antibiotic production and excre-tion of amino acids. The data are presented inTable 2.
In addition to increases in antibiotic yield, anumber of the strains excreted amino acids, thelevel of which varied greatly during the fermen-tation. Since the possibility existed that theproducts were being utilized by the cell, it wasnecessary to trap the excreted amino acid(s).
Detection of excreted amino acids. Threesequential steps were employed to detect and
75VOL. 4, 1973
on May 16, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
ANTIMICROB. AG. CHEMOTHER.
E
E 400._
300-z
O 200-
I
0~
CL T I -Wi 0U
20-
16
12-CD _ LA473 (I eu) * --
ui - L100 *---_o
0. 4-
0u
0 200 400 600 800 1000LEUCINE SUPPLEMENTATION pg/miFIG. 1. Effect of leucine on LA473. Overnight cul-
tures ofLA473 and L100 were inoculated into minimalfermentation medium supplemented with variouslevels of leucine. After 4 days of incubation, antibioticpotency and growth as indicated by packed myceliumvolumn were determined.
quantitate the overproduction of amino acid(s).First, the suspected culture was stamped (Fig.2A) onto lawns of auxotrophs on JS7 minimalmedia. An excreted amino acid would be uti-lized by its particular auxotroph as evidencedby a halo of growth surrounding the colony (Fig.2B). These cultures were scored as positive forexcretion on agar. The strains were then inocu-lated into JS7 minimal broth and definedfermentation broth with the appropriate aminoacid(s) omitted from the media. Samples wereremoved at 24, 48, 72, and 96 h and placed onlawns of each auxotroph. Those broth samplesthat displayed a halo of secondary growthsurrounding the pad along with an identicalbroth sample of L100 were assayed separately inthe amino acid analyzer for quantitation of theexcreted amino acid(s).Analogue-resistant mutants. Analogue-
TABLE 2. Prototrophic revertants
Revert- Fermen-Auxo- Strain ant Excretiona agar tationtrophy no. strain potency"
no.
L100 100Threonine LA466 LE46 Thr 100
LE33 Thr, Val, Leu 100LE21 Thr, Leu 170
Cysteine LA445 LE8 Cys 140LA410 LE22 Cys, Met, Val 100LA425 LE51 Cys, Met, Thr 100
Leu, LysLeucine LA185 LE32 Leu, Thr 120Isoleucine LA337 LE35 Ile, Thr 190
LE43 Ile, Thr, Leu 100Cys, Met
Lysine LA438 LE27 Lys, Val, Leu 100
a The excretion assays are explained in the text.Potency was measured in terms of methoxyceph-
alosporin C or units per milliliter on penase platesseeded with X142.
resistant mutants were isolated according toprocedures described in the Materials andMethods section. Analogue-resistant coloniessurrounded by a halo of secondary growth werepreferentially selected. After recloning on thesame medium the individual strains were testedfor excretion of amino acids and assayed forantibiotic yield. The isolation procedure wasfairly efficient as approximately 30% of themutants isolated, overproduced specific aminoacids.
In addition, the toxicity of norvaline andtrifluoromethionine was markedly enhanced byhomoserine, whereas threonine enhanced thetoxicity of amino-ethyl cysteine. Some form ofmultivalent repression or inhibition may beoperative in this case, but to date we have notobserved any form of growth inhibition of L100by any amino acid or combination of aminoacids of the aspartic acid family. The data arepresented in Table 3.
Those mutants defective in the ilv/leu regionof the pathway (LC185, LC88, LC210, andLC148) had the greatest increases in antibioticyield. During the course of this investigation anonsporulating mutant was isolated, LM24,that displayed potency levels of twofold over itswild-type parent L100. From this strain (LM24), a trifluoroleucine-resistant mutant was iso-lated (LM 5) that excreted leucine and dis-played potency levels sixfold over L100. Increas-ing the exogenous levels of leucine, valine, orthreonine either singly or in combination hadlittle if any effect on antibiotic potency of theprototrophic parent L100.
76 GODFREY
on May 16, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

ANTIBIOTIC PRODUCTION IN STREPTOMYCES
FIG. 2. Detection of excreted amino acids on agar.A, One-half milliliter of an overnight culture was
placed into each cup. The cultures were then stampedonto six plates of minimal medium JS7, each contain-ing a lawn composed of a different auxotroph (LA429,LA415, LA351, LA427, LA398, and LA426). The platecontaining LA415 (ilv) was also supplemented with500 pg of isoleucine per ml. B, After 2 and 3 days ofincubation the plates were observed. Those coloniessurrounded by a halo of secondary auxotrophic growthwere scored as positive for excretion on agar (arrow).
All the penicillin resistant mutants had a
mucoid colony appearance. Of the 20 resistantcolonies isolated, only 4 could be recloned. Oneof these, P3, produced antibiotic. This strainexcreted valine, leucine, and alanine. StrainLC194 also excreted alanine but in additionproduced neither antibiotics nor spores. WhenLC194 was incubated with a number of otherstrains their ability to form spores was alsoinhibited. This is similar to the diffusable
compound(s) involved in inhibition of fertilityin S. colicolor (17).
DISCUSSIONS. Iipmanii is an ideal organism in which to
study antibiotic synthesis because all threeantibiotic precursors (aAAA, cys, and val) aredirectly related to the biosynthesis of the aminoacids present in either the aspartic acid familyor pyruvic family of amino acids. In addition,stable mutants can be obtained at high frequen-cies (6), and multiple amino-acid-analogue-resistant mutants have been isolated that pro-duce at least 1 mg of purified antibiotic(s) perml in a defined fermentation medium.
Since all three antibiotic precursors are ineither the aspartic acid family or pyruvic acidfamily, it was decided to isolate regulatorymutants of these pathways. For this studyphysical derepression by amino acid starvation,prototrophic revertants, and analogue resistantmutants were employed. The purpose of thisarticle is to describe the techniques employed toisolate controlless mutants and the ultimateeffect of such mutants on antibiotic potency. Itis hoped that future work in this area will leadto techniques that can be employed to directlyregulate these genes and resultant enzymesconcerned with antibiotic synthesis. Increasedantibiotic synthesis of derepressed strains hasbeen noted previously by Elander et al. (4) forpyrrolnitrin synthesis.By regulating the levels of exogenous amino
acid supplementation of auxotrophs (Table 1) itwas possible to either prevent antibiotic synthe-sis or direct the synthesis to the production ofone factor only (LA441). Apparently, the aminoacids used in antibiotic synthesis are usedpreferentially for protein biosynthesis, becom-ing available for secondary metabolism onlywhen present in excess. Thus, antibiotic pro-duction can be readily divorced from primarymetabolism. This is true in the case of LA425(cys), LA467 (ilv), and LA423 (lys) but not withLA473. In this last instance growth-limitinglevels of leucine supplementation (Fig. 1) mark-edly increased the antibiotic potency of LA473,whereas the wildtype (L 100) was not in-fluenced by any level of leucine supplementa-tion. From the data it would appear that leucinelimitation resulted either in the accumulationof valine or some other intermediate in leucinesynthesis. However, in no instance was thereany correlation between pool levels of valine inLA473 and exogenous leucine supplementation.
Strain LE8 appears to be the most interestingof the revertants, not only because cysteine wasexcreted, but also because its parenteral auxo-
VOL. 4, 1973 77
on May 16, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

ANTIMICROB. AG. CHEMOTHER.
TABLE 3. Analogue-resistant mutants of L100
Analogue ~Inhibitionb Excretion Fermen-Analogue ~~~Strain ItationeCompound concn No.a n. Mutc 1 Broth tec(mg/mi) - + agar (Mumoles/mld) poec
Trifluoromethionine
DL-4-Aza-leucine
DL-Allyl glycine
DL-Norleucinea-Amino-DL butyric
acid
Phenyl mercuricacetate
Seleno-DL-cystine
DL-Norvaline
Penicillin G
Trifluoroleucine
D-Ethionine
2-Amino-ethyl-L-cysteine
9
7
10
215
10
Leu
MetValLeuPheTyrTrp
HomValMet
3 Leu IHomg
4
15
12
26
Leu
MetPheIleValLeuTyrTrp
Thr
LC230
LC227
LC73
LC124LC210LC254LC197
LC194
LC235LC237L100
LC144
LC185
P3
LC88
LC120
LC127
LC171
LC128
LC148
NTG
DMS
NTG
NTGDMSDMSNTG
NTG
DMSNTG
NTG
NTG
NTG
NTG
NTG
NTG
NTG
NTG
NTG
ValIleValIle
Leu
LeuLeuLeuVal
ThrValValLeu
CysMetValLeuThr
ValThr
ThrLeuLeuValVal
LeuThrLysVal
ThrLeu
Cys, 0.109/0.014Val, 0.790/0.018Ala, 1.89/0.009Cys, 0.031/-Leu, 0.184/0.006Leu, 0.508/0.023Ser, 0.204/0.004Lys, 0.242/0.014
Val, 0.138/0.036Leu, 0.127/-
Val, 0.213/trAla, 1.63/0.08
Ile, 0.205/0.008
Cys, 0.166/-'Thr, 0.143/0.058Leu, 0.326/0.026Lsy, > 1.5/0.04Thr, 0.271/-Ser, 0.290/0.023Ala, 0.283/0.035Val, > 1.5/0.018Met, > 1.5/0.007Val, 0.142/0.018Leu, 0.190/0.006Ala, 0.463/0.035Thr, 0.203/-Ser, 0.205/-Ala, 0.403/0.009Leu, 0.549/0.006Thr, 0.065/trSer, 0.499/0.008
Ser, 0.131/0.012
Lys, 0.04/-Thr, 0.154/-Ser, 0.142/0.012
100
100
100
100170100100
NA
100100100
50
120
40
180
NA
100
100
100
130
aNo, number of resistant strains isolated.b Minus sign, reverses inhibition of analogue; plus sign, enhances inhibition of analogue.c Mut, mutagen used.d-, Indicates 1 ml.ePotency refers to the concentration of methoxycephalosporin C measured as units per milliliter on penase
plates seeded with X142.'0.166/, LC 144 had 0.166 umol of cysteine/ml of its broth, whereas L100 under identical growth conditions
possessed no detectable levels of cysteine.g Hom, homoserine.
20
2
20
7010
40.2 x 10-3
0.5
20
0.4
1.5
30
1.9
78 GODFREY
on May 16, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

ANTIBIOTIC PRODUCTION IN STREPTOMYCES
troph LA445 possessed a unique auxanographicpattern. This strain could grow on either homo-serine or cysteine. Work is currently underwayto characterize LA445.
Similar to previous findings (15) homoserineand threonine were found to increase the tox-icity of a number of analogues. In addition, highconcentrations of the analogues were employedto avert permeability mutants. Of the 140mutants isolated during the course of thisstudy, approximately 30% overproduced variousamino acids.As far as antibiotic yield is concerned, strains
lacking control in the ilv-leu region were supe-rior. This in conjunction with loss of feedbackcontrol of LA473 (leu) suggests that valinesynthesis is the rate-limiting step of antibioticproduction in L100. Goulden and Chattaway (7)demonstrated that acetohydroxy acid synthe-tase, of a high yielding (penicillin) strain ofPenicillium chrysogenum was less sensitive tovaline and had a higher specific activity thanthe strain from which it was derived. This,combined with the above, indicates the possi-bility that valine biosynthesis may be ratelimiting in a number of other f-lactam produc-ing cultures.The programmed synthesis of different types
of actinomycins can be accomplished by sup-plying different precursor amino acids throughthe substratum (9). In the case of S. lipmaniithe greatest potential appears to be in removingcontrol from various regions of the aspartic acidfamily. Analogue-resistant mutants are not onlya valuable tool for increasing potency of specificantibiotics but also are an easy marker to use ingene transfer experiments. In addition, thepleiotrophic nature of specific analogue-resist-ance markers will be a great aid in inter andintra species matings. Recombinants from suchmatings hopefully may synthesize new antibiot-ics. Work is currently in progress in obtainingstrains resiistant to multiple combinations ofanalogues either by mutation or recombination.
ACKNOWLEDGMENTS
I thank M. McNeely for her excellent technical assistance.In addition, I acknowledge the assistance of J. Westhead
with the antibiotic assays and thank N. Davis for hisassistance with the chromatography section.
LITERATURE CITED
1. Abraham, E. P., and G. G. F. Newton. 1961. Thestructure of cephalosporin C. Biochem. J. 79:377-393.
2. Caltrider, P. G., and H. F. Niss. 1966. Role of methioninein cephalosporin synthesis. Appl. Microbiol.14:746-753.
3. Demain, A. L., and J. F. Newkirk. 1962. Biosynthesis ofcephalosporin C. Appl. Microbiol. 10:321-325.
4. Elander, R. P., J. A. Mabe, R. L. Hamill, and M.Gorman. 1971. Biosynthesis of pyrrolnitrins by ana-logue-resistant mutants of Pseudomonas fluorescens.Folia Microbiol. 16:156-165.
5. Florey, H. W., E. B. Chain, N. G. Heatley, M. A.Jennings, A. G. Sanders, E. P. Abraham, and M. E.Florey. 1949. Antibiotics, vol. 2. Oxford UniversityPRess, London.
6. Godfrey, 0. W., and J. R. Kirkpatrick. 1972. Theisolation and characterization of auxotrophs of theaspartic acid family from Streptomyces lipmanii. FoliaMicrobiol. 18(2):90.
7. Goulden, S. A., and F. W. Chattaway. 1969. End-productcontrol of acetohydroxy acid synthetase by valine inPenicillium chrysogenum A176 and a high penicillin-yielding mutant. J. Gen. Microbiol. 59:111-118.
8. Higgens, C. E., and R. E. Kastner. 1971. Description ofStreptomyces clavuligerus, a ,-Lactam antibiotic pro-ducing Streptomycete. Int. J. Syst. Bacteriol. 21:326-331.
9. Jayaraman, K., and G. Rangaswam. 1969. Biosynthesis ofantibiotics. J. Sci. Ind. Res. 28:501-504.
10. Kaplan, M. M., and M. Flavin. 1965. Enzymatic synthe-sis of L-cystathionine from the succinic ester of aa-homoserine. Biochim. Biophys. Acta 104:390-394.
11. Kuhn, J., and R. L. Somerville. 1971. Mutant strains ofEscherichia coli K12 that use D-amino acids. Proc.Nat. Acad. Sci. U.S.A. 68:2481-2487.
12. Miller, D. L., and F. Rodwell. 1971. Metabolism of basicamino acids in Pseudomonas putida. J. Biol. Chem.246:2758-2764.
13. Nagarajan, R., L. D. Boeck, M. Gorman, R. L. Hamill, C.E. Higgens, M. M. Hoehn, W. M. Stark, and J. G.Whitney. 1971. ,3-Lactam antibiotics from strep-tomycetes. J. Amer. Chem. Soc. 93:2308-2310.
14. Shijo, I., and S. Nakamori. 1970. Microbial production ofL-threonine. Agr. Biol. Chem. 34:448-455.
15. Trown, P., M. Sharp, and E. P. Abraham. 1963. Biosyn-thesis of Cephalosporin C from amino acids. Biochem.J. 86:284-291.
16. Vivian, A. 1971. Genetic control of fertility in Strep-tomyces coelicolor A3(2): plasmid involvement in theinterconversion of UF and IF strains. J. Gen. Microbiol.69:353-364.
17. Whitney, J. G., D. R. Brannon, J. A. Mabe, and K. J.Wicker. 1972. Incorporation of labeled precursors intoA16886B, a novel 3-lactam antibiotic produced byStreptomyces clavuligerus. Antimicrob. Ag. Chemo-ther. 1:247-251.
VOL. 4, 1973 79
on May 16, 2018 by guest
http://aac.asm.org/
Dow
nloaded from