and polyol dehydrogenases in patella vulgata and · grupos de organismos. ... o segundo maior filo...
TRANSCRIPT

Aromatic alcoholand polyoldehydrogenases inPatella vulgata andother gastropods(Mollusca)Diogo Ribeiro Amaral de CarvalhoMestrado em BioquímicaDepartamento de Química e Bioquímica
2015/2016
Orientador
Alexandre Lobo da Cunha, Professor Catedrático, Instituto de
Ciências Biomédicas Abel Salazar
Coorientador
Vitor Costa, Professor Associado, Instituto de Ciências Biomédicas
Abel Salazar


Todas as correções determinadas
pelo júri, e só essas, foram efetuadas.
O Presidente do Júri,
Porto, /_ /


FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
I
Agradecimentos
Aos meus orientadores: ao Professor Doutor Alexandre Lobo-da-Cunha, pelo
apoio, interesse e auxílio prestados na orientação deste trabalho ao longo destes quase
cinco anos de investigação conjunta; e ao Professor Doutor Vitor Costa, por ter aceite o
desafio de auxiliar a investigação de um organismo tão estranho e muito pouco
conhecido pela comunidade científica em geral.
À equipa técnica do laboratório de Biologia Celular do departamento de
Microscopia do ICBAS, com especial destaque para a Ana Paula Sousa, pelas
infindáveis e martirizantes quantidades de tubos de ensaio lavados em prol deste
trabalho.
À equipa técnica do departamento de Biologia Molecular do ICBAS,
nomeadamente à Carla Oliveira e à Marisa Castro, pela cedência de equipamento e
apoio prestado na realização de alguns procedimentos laboratoriais.
À equipa do grupo de investigação UP3 do IBMC/i3S, nomeadamente ao Doutor
Frederico Silva à Fátima Fonseca, pelo apoio prestado na realização dos procedimentos
cromatográficos.
Ao Doutor Hugo Osório do IPATIMUP/i3S, pelo apoio prestado na identificação e
sequenciação das proteínas das bandas dos géis.
Aos meus, sempre presentes, colegas de mestrado, que individualmente foram
partilhando comigo as suas vivências, familiarizando-me com o que significa ser
bioquímico; e não esquecendo a origem de cada um, aceitam agora um novo conceito,
o de Bioquímico do Meio Aquático.
To you, Lucie, because you were a very important part of my life in this biochemical
journey, and in your six months’ adventure in Portugal, we were still able to share some
unforgettable moments together.
Ao pessoal do Centro Português de fotografia, à equipa de seguranças e às
minhas colegas, nomeadamente à Maria da Luz, a que desejo rápidas melhoras, e mais

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
II
recentemente à Mónica; um grande obrigado por tudo aquilo que me ensinaram nestes
três meses e meio de experiência como trabalhador-estudante.
And to you, my dear Alex, that like a tornado that comes from the sea to make
changes in land, so your overseas arrival testified the end of this chapter of my life, and
brought a new one alive; hopefully for many long years…

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
III
Resumo
As desidrogenases alcoólicas são enzimas bem conhecidas, estudadas em muitos
grupos de organismos. No entanto, pouco se sabe sobre estas enzimas em moluscos,
o segundo maior filo de animais multicelulares. Em estudos preliminares com várias
espécies dos cinco principais clados de gastrópodes, nomeadamente
Patellogastropoda, Neritimorpha, Vetigastropoda, Caenogastropoda e Heterobranchia,
foi possível detetar desidrogenases de álcoois aromáticos (usando álcool cinamílico
como substrato) e poliálcoois (usando manitol e sorbitol como substrato) em
homogeneizados totais de glândula digestiva. Usando a fração citosólica da glândula
digestiva de Patella vulgata, Phorcus lineatus, Littorina littorea e Siphonaria pectinata, a
atividade destas enzimas foi detetadas in situ após eletroforese em gel nativo de
poliacrilamida, verificando-se um perfil enzimático semelhante entre as várias espécies.
Este trabalho tinha como objetivo o isolamento e identificação destas enzimas, da
glândula digestiva da lapa Patella vulgata, uma espécie abundante na zona intertidal
portuguesa. Frações enriquecidas nas enzimas de interesse foram obtidas por
cromatografia de afinidade e de troca iónica. Estudos de proteómica revelaram a
presença de várias desidrogenases, onde se incluem uma álcool desidrogenase de
classe III e duas desidrogenases de poliálcoois (sorbitol e arabinitol desidrogenases) A
clonagem dos genes que codificam para estas enzimas e a expressão heteróloga destas
proteínas, irá permitir a sua caracterização, incluindo a atividade das desidrogenases
para diversos substratos alcoólicos.
Palavras-Chave
Desidrogenases alcoólicas, moluscos, gastrópodes, álcoois aromáticos,
poliálcoois, glândula digestiva, arabinitol desidrogenase, sorbitol desidrogenase.

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
IV
Abstract
Alcohol dehydrogenases are well known enzymes, studied in many types of
organisms. However, little is known about these enzymes in molluscs, the second largest
multicellular animal phylum. Preliminary studies with several species from the five major
gastropod clades, namely Patellogastropoda, Neritimorpha, Vetigastropoda,
Caenogastropoda e Heterobranchia, allowed the detection of aromatic alcohol
dehydrogenases (using cinnamyl alcohol as substrate) and polyalcohol dehydrogenases
(using mannitol and sorbitol as substrate) in digestive gland total homogenates. Using
the digestive gland cytosolic fraction of Patella vulgata, Phorcus lineatus, Littorina littorea
e Siphonaria pectinata, the activity of these enzymes was also detected in situ after
native polyacrylamide gel electrophoresis which resulted in a similar overall profile in all
studied species. This work aimed at the isolation and identification of these enzymes
from the digestive gland of the limpet Patella vulgata, an abundant species in the
Portuguese intertidal zone. Fractions enriched in the enzymes of interest were obtained
by affinity and ion exchange chromatography. Proteomic studies revealed the presence
of several dehydrogenases, which are included one class III alcohol dehydrogenase and
two dehydrogenases (sorbitol and arabinitol dehydrogenase). Cloning of the genes
encoding for these enzymes and the heterologous expression of these enzymes will
allow their characterization, including the dehydrogenase activity for diverse alcoholic
substrates.
.
Keywords
Alcohol dehydrogenases, molluscs, gastropod, aromatic alcohol, polyalcohol,
digestive gland, arabinitol dehydrogenase, sorbitol dehydrogenase.

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
V
Index
List of figures and tables .............................................................................................. VI
List of abbreviations ..................................................................................................... IX
1. Introduction ............................................................................................................ 10
1.1. Gastropod taxonomy and phylogeny ............................................................. 10
1.2. Digestive Gland ............................................................................................. 14
1.3. Oxidation of polyols and aromatic alcohols in gastropods ............................. 16
1.4. Alcohol dehydrogenases ............................................................................... 18
1.4.1. MDR - Type I ADHs ............................................................................. 18
1.4.2. SDR - Type II ADHs ............................................................................. 21
1.4.3. LDR - Type III ADHs ............................................................................ 22
1.4.4. Polyol Dehydrogenases ....................................................................... 23
1.5. Alcohol and polyol dehydrogenases in molluscs............................................ 26
1.6. Aims of this work ........................................................................................... 27
2. Materials and Methods ........................................................................................... 28
2.1. Species and sampling sites ........................................................................... 28
2.2. Preparation of protein extracts ...................................................................... 28
2.3. Enzyme activities .......................................................................................... 30
2.4. Affinity chromatography ................................................................................. 31
2.5. Ion exchange chromatography ...................................................................... 31
2.6. SDS-polyacrylamide gel electrophoresis ....................................................... 31
2.7. Protein sequencing ....................................................................................... 32
3. Results ................................................................................................................... 33
3.1. Enzymatic activities ....................................................................................... 33
3.2. Protein isolation and identification ................................................................. 36
4. Discussion .............................................................................................................. 45
5. Conclusions and perspectives ................................................................................ 49
6. References ............................................................................................................. 50

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
VI
List of figures and tables
Figure 1 - Major gastropod clades, according to morphological and molecular data ... 11
Figure 2 - Pictures of some gastropod species included in this study. A – Patella vulgata,
B – Phorcus lineatus, C – Nucella lapillus, D – Nassarius reticulatus, E – Siphonaria
pectinata, F – Armina maculata, G – Bulla striata, H – Aglaja tricolorata (provided
by A. Lobo-da-Cunha, University of Porto). .......................................................... 13
Figure 3 - Schematic representation of the digestive gland cells in gastropods. mv –
microvilli, ev – endocytic vesicle, ly – lysosome, li – lipid droplets, nu – nucleus, sv
– secretory vesicles, cc – calcium concretions (provided by A. Lobo-da-Cunha,
University of Porto). ............................................................................................. 15
Figure 4 - [A] Ultrastructure of the basophilic cells of the digestive of A. depilans, showing
tubular structures in the interior of rough endoplasmic reticulum cisternae (*) [12].
[B] Frozen section of the digestive gland of A. depilans showing stained basophilic
cells (arrows) after the detection of mannitol oxidase activity [12]. ....................... 16
Figure 5 - Scheme of the polyol pathway of the glucose metabolism. Conversion of
glucose to sorbitol by aldose reductase, and sorbitol to fructose by sorbitol
dehydrogenase [98]. ............................................................................................ 24
Figure 6 - Structure of sheep sorbitol dehydrogenase
(http://www.rcsb.org/pdb/explore/explore.do?structureId=3QE3). ........................ 24
Figure 7 - Native electrophoresis gel sections with in situ detection of the activity of
dehydrogenases of several alcoholic substrates, in four marine herbivorous species.
............................................................................................................................ 35
Figure 8 - Ion exchange chromatogram of 10 mM NAD+ fraction from affinity
chromatography with HiTrap DEAE Sepharose FF column. Proteins eluted with an
ascending and continuous NaCl gradient (0.05 M to 1 M), at a flow rate of 1 ml per
sec. (blue) protein detection (abs 280 nm); (green) high salt solution concentration;
(red) conductivity. ................................................................................................ 38
Figure 9 - SDS-polyacrylamide gel showing the enzymatic profile of some ion exchange
chromatography fractions obtained with HiTrap DEAE Sepharose FF column

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
VII
(Figure 8). Proteins identified by MS/MS using a database of Patella vulgata: 1 –
glutathione-S-transferase mu class; 2 – no statistical significant identification; 3 – D-
arabinitol dehydrogenase 1; 4 – cytosolic malate dehydrogenase; 5 – sorbitol
dehydrogenase-like protein (Table 7). .................................................................. 39
Figure 10 - Ion exchange chromatogram of 10 mM NAD+ fraction from affinity
chromatography with Mono Q 4.6/100 PE column Proteins eluted with an ascending
and continuous NaCl gradient (0.05 M to 1 M), at a flow rate of 1 ml per sec. (green)
protein quantification (abs 280 nm); (black) high salt solution concentration; (red)
conductivity. ......................................................................................................... 42
Figure 11 - Characterization of the fractions obtained by ion exchange chromatography
(Mono Q 4.6/100 PE). Top: analysis of polyol dehydrogenase activity. Bottom:
analysis of proteins by SDS-PAGE. Proteins were stained with Coomassie blue.
Molecular weight markers are shown on the left and right. Spots 1-7 were identified
by MS/MS: 1 & 2 alcohol dehydrogenase class III; 3 – universal stress protein; 4 &
5 - D-arabinitol dehydrogenase and Glyceraldehyde-3-Phosphate dehydrogenase
1; 6 & 7 – sorbitol dehydrogenase-like protein (Table 8). ..................................... 43
Table 1 - Compatible substrates for human liver sorbitol dehydrogenase [105] ........... 26
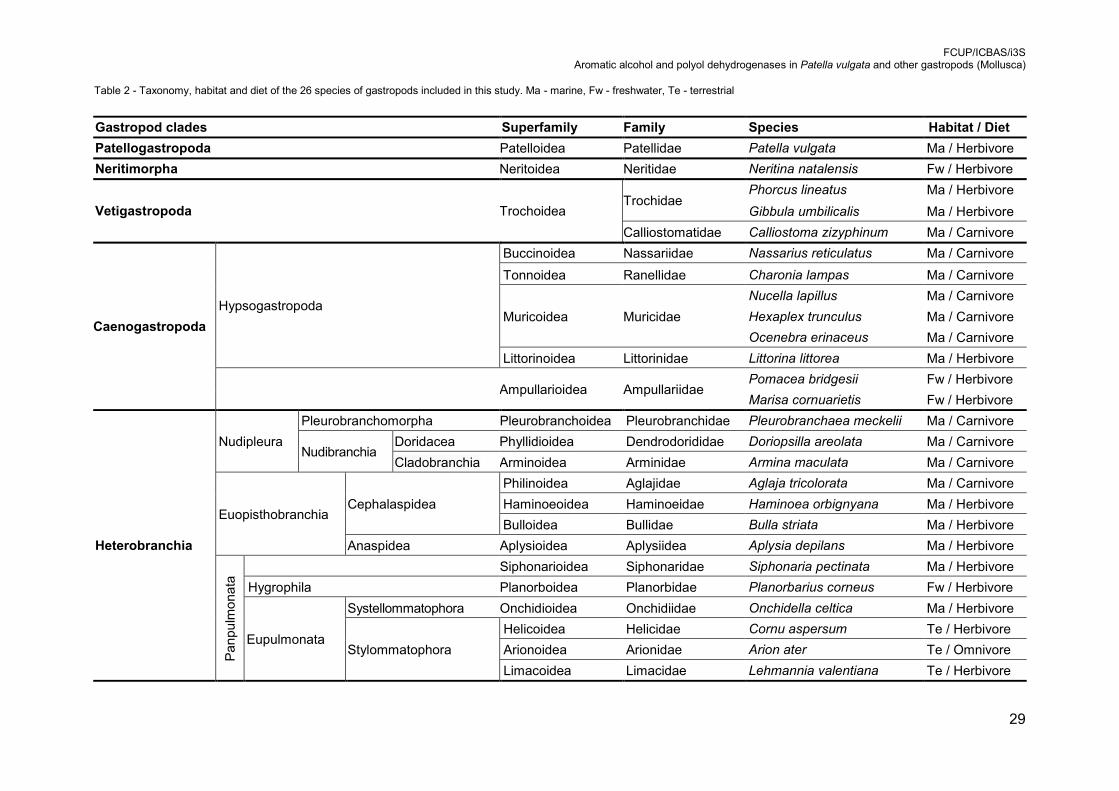
Table 2 - Taxonomy, habitat and diet of the 26 species of gastropods included in this
study. Ma - marine, Fw - freshwater, Te - terrestrial ............................................. 29
Table 3 - Activity of dehydrogenase in the digestive gland of gastropods. Values are
mean ± SD of at least 5 independent experiments (nmol.min-1. g-1 ± SD). Shaded
lines correspond to carnivorous species. ............................................................. 34
Table 4 - The cytosolic fraction of the digestive gland of Patella vulgata was loaded on a
Blue Sepharose CL-6B column, the flow-through was recovered and the column
washed with buffer. Proteins were eluted with NAD+ solutions of increasing
concentrations (5 mM, 10 mM and 20 mM). The column was then washed with
regeneration buffers. Dehydrogenase activities for polyols (D-mannitol and D-
sorbitol) and cinnamyl alcohol (aromatic) in the fractions were assayed by
spectrophotometry. A/ml – activity per ml of fraction; A/mg – activity per mg of
protein (specific activity); A total – activity of total fraction volume. ...................... 37

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
VIII
Table 5 - The cytosolic fraction of the digestive gland of Patella vulgata was loaded on a
Blue Sepharose CL-6B column, the flow-through was recovered and the column
washed with buffer. Proteins were eluted with two NAD+ solutions of different
concentrations (10 mM and 20 mM). The column was then washed with
regeneration buffers. Dehydrogenase activities for polyols (D-mannitol and D-
sorbitol) and cinnamyl alcohol (aromatic) in the fractions were assayed by
spectrophotometry. A/ml – activity per ml of fraction; A/mg – activity per mg of
protein (specific activity); A total – activity of total fraction volume. ...................... 37
Table 6 - 10 mM NAD+ fraction from affinity chromatography loaded on a HiTrap DEAE
Sepharose FF column. Proteins eluted with an ascending and continuous NaCl
gradient (0.05 M to 1 M), at a flow rate of 1 ml per sec. (Figure 8). Distribution of
dehydrogenase activities for polyols (D-mannitol and D-sorbitol) and cinnamyl
alcohol (aromatic) in some fractions were assayed by spectrophotometry. A/ml –
activity per ml of fraction; A/mg – activity per mg of protein (specific activity); A total
– activity of total fraction volume. Fractions: no.4 – flow-through representative, no.
22, 25 and 27 – protein peaks representatives. ................................................... 37
Table 7 - Proteins of interest (Figure 9) were analysed by mass spectrometry (MS/MS)
and identified using a Patella vulgata database. Blastp of the identified proteins was
performed at the NCBI database. Mascot search result scores greater than 65 are
significant (p<0.05). ............................................................................................. 40
Table 8 - Proteins from ion exchange chromatography fraction with Mono Q 4.6/100 PE
column (Figure 11) analysed by mass spectrometry (MS/MS) and identified with a
Patella vulgata database. Mascot search result scores greater than 65 are
significant (p<0.05). Data shows the highest score identifications for each sample.
............................................................................................................................ 44
Table 9 - Peptide ions from ion exchange chromatography fraction with Mono Q 4.6/100
PE column (Figure 11) analysed by mass spectrometry (MS/MS) and identified with
UniProt (Metazoa) database. Mascot search result scores greater than 82 are
significant (p<0.05); Data shows the highest score identifications for each sample.
............................................................................................................................ 44
Table S1 - Oxidase activities in the digestive gland of gastropods (nmol.min-1. g-1 ± SD).
Shaded lines correspond to carnivorous species (data not published). ................ 58
FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
IX
List of abbreviations
RNA Ribonucleic Acid
NAD(H) Nicotinamide Adenine Dinucleotide
FAD Flavin Adenine Dinucleotide
ADH Alcohol Dehydrogenase
NADP(H) Nicotinamide Adenine Dinucleotide Phosphate (reduced form)
MDR Medium-chain Dehydrogenase/Reductase
DNA Deoxyribonucleic Acid
SDR Short-chain Dehydrogenase/Reductase
PGDH 15-Hydroxyprostalandin Dehydrogenase
LDR Long-chain Dehydrogenase/Reductase
HOT Hydroxyacid-Oxoacid Transhydrogenase
SDH Sorbitol Dehydrogenase
ALR Aldose Reductase
EDTA Ethylenediaminetetraacetic Acid
PMSF Phenylmethylsulfonyl Fluoride
NBT Nitro Blue Tetrazolium Chloride
TCA Trichloroacetic Acid
SDS Sodium Dodecyl Sulfate
MS Mass Spectrometry
MALDI Matrix-Assisted Laser Desorption Ionization
GAPDH Glyceraldehyde 3-phosphate Dehydrogenase
ARDH Arabinitol Dehydrogenase

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
10
1. Introduction
1.1. Gastropod taxonomy and phylogeny
Gastropoda is a class of molluscs that includes terrestrial, marine and freshwater
snails and slugs, and it was traditionally divided in three subclasses: Prosobranchia,
Opisthobranchia e Pulmonata [1]. However, morphological and molecular studies
showed that these were not monophyletic groups.
In the last decades, many studies have been dedicated to gastropod phylogeny
[2]. These studies rely mostly in molecular methods to establish phylogenetic trees and
verify group monophyly. Nevertheless, despite the increasing use of rRNA and
mitochondrial genomes, there are still a lot of doubts regarding phylogenetic
relationships between gastropod clades. Thus, gastropods are now divided in the clades:
Patellogastropoda, Neritimorpha, Cocculiniformia, Neomphalina, Vetigastropoda,
Caenogastropoda e Heterobranchia (Figure 1) [2-4].
The clade Patellogastropoda is composed by marine gastropods, known as true or
patellid limpets, which can be found attached to almost any rocky surface, coralline
algae, sunken vessels, whale bone and even to the shell of other molluscs. Their
distribution knows no borders, from the intertidal rocky shores to the deep-sea waters,
from polar to tropical region, and they play very important roles in littoral ecosystems,
having been largely used in ecological studies [5]. These are nocturnal grazing molluscs
that, by scraping the rock surface with their radulae ingest diatoms, settling stages of
algae and invertebrates, filamentous algae, cyanobacteria and detritus. This way, large
rocky areas are maintained in a primary stage of succession [6].
Among all Patellogastropoda families, Patellidae is the most studied one with a
total of 38 species in 4 genera., based in morphological studies. The genus Patella
comprises nine species, that occur in the north-eastern Atlantic area. This genus has
been used for many genetic studies, although very few species were analysed or they
represent a small region [5]. A very common limpet in the Portuguese coast is the species
Patella vulgata (Figure 2A). On the other hand, the family Lottiidae is the most diverse
within Patellogastropoda, comprising 130 species in at least 8 genera, with a worldwide
distribution of temperate and tropical areas. It includes the first mollusc species with full
sequenced genome (Lottia gigantea), being important as a representative of gastropod
basal lineages [5].

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
11
Figure 1 - Major gastropod clades, according to morphological and molecular data
Neritimorpha is a group of large morphological variety, such as the shell shape.
Besides the typical snail appearance, some species resemble limpets or slugs. They are
adapted to some habitats of harsh conditions, like deep-sea hydrothermal vents and
submarine caves, but also distributed in intertidal and subtidal areas of tropical regions,
freshwater habitats and arboreal terrestrial environments [7]. Most neritimorphs are
grazers on algal spores, diatoms and detritus [8]
Regarding recent molecular studies, the groups Neophalina and Cocculinoidea are
now considered two deep-water sister clades. Cocculinids is a group of small, white-
shelled deep-sea limpets, found in decaying squid beaks and sunken-bone/wood, while
neomphalines share features with both cocculinids and neritimorphs and are found
exclusively in hydrothermal vents, hydrocarbon seeps and sunken-wood [3]
The Vetigastropoda clade is constituted by marine snails, largely distributed across
all oceans, from deep waters to the shallow intertidal areas. They display a large variety
of shell morphologies, such as conical, limpet and trochiform types (Figure 2B), that can
also contain holes, slits and fissures [3]. They feed on encrusting invertebrates, however
Heterobranchia
Caenogastropoda
Vetigastropoda
Neritimorpha
Patellogastropoda
“Basal” Heterobranchia
Neomphalina
Cocculiniformia
Euthyneura Nudipleura
Tectipleura Euopisthobranchia
Panpulmonata

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
12
some groups, such as Trochoidea, can feed on algae and plant material such as marine
angiosperms [9].
Caenogastropoda is the largest and most diverse group of living snails, with about
136 existing families. It is a group of ecological and commercial important marine
families, but also contains some terrestrial and freshwater representatives. They present
a wide array of shell morphologies (coiled, uncoiled, globose, elongate, limpet-shaped)
(Figure 2C,D), however, some species present a reduced shell or more rarely do not
contain a shell. Caenogastropoda are found in a wide range of habitats, such as pelagic
drifters or active swimmers, benthic epifaunal or burrowers, herbivores, grazers, active
carnivores, detritus or sedentary suspension feeders, ectoparasites or shell-less
endoparasites [10].
Heterobranchia is a large clade formed by marine, freshwater and terrestrial snails
and slugs, which taxonomy have been changed mainly due to molecular data. This clade
is characterized by several apomorphic features [4], comprising several groups which
systematic position is not yet clear. Those groups presenting more ancestral features
are included in the informal group of “Basal heterobranchs”, which are clearly
paraphyletic [11]. The clade Euthyneura is characterized by a typical non/crossed
nervous system and innervated rhinophores. It is divided in Nudipleura, that occupy a
basal position, suggesting some plesiomorphic features despite the derived ones (Figure
2F), and Tectipleura that is characterized by a monaulic genital system. This last group
is divided in Euopisthobranchia, that share a cuticularized oesophagus (Figure 2G,H),
and Panpulmonata that presents a double-root procerebrum (Figure 2E). Overall, the
antique Euthyneura clade is the only who will survive to the molecular revolution of
gastropod systematics [4]. Regarding their dietary habits, heterobranchs can be
herbivorous, carnivorous, omnivorous or detritivorous, playing an active role hunting
pursuing their pray, or more sedentary one, scavenging or grazing the nearby territory
[11].

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
13
Figure 2 - Pictures of some gastropod species included in this study. A – Patella vulgata, B – Phorcus lineatus, C – Nucella
lapillus, D – Nassarius reticulatus, E – Siphonaria pectinata, F – Armina maculata, G – Bulla striata, H – Aglaja tricolorata
(provided by A. Lobo-da-Cunha, University of Porto).

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
14
1.2. Digestive Gland
The digestive gland is a major organ of the molluscs’ digestive system, being
composed by a vast number of tubules with a dead end, connected to the stomach via a
system of branched ducts. This organ is involved in extracellular and intracellular
digestion of food, nutrient absorption, lipid, glycogen and minerals storage, also having
an important role in detoxification [12-14].
The digestive tubules, also called digestive diverticula, have a one-cell layer
epithelium, surrounded by the basal lamina and connective tissue with muscular cells.
The pulsatile contractions of the muscular cells, combined with the stomach contractions,
move the food materials into the digestive gland. On the other hand, the beating of the
ciliary apparatus of the epithelial cells drives the materials away from the digestive gland,
toward the stomach and intestine. These seem to be the two systems, of opposite
directions that carry the ingested material through the ducts. Inside the ducts, fine
particles and soluble materials are taken up rapidly by endocytosis, however larger
particles are moved directly from the stomach to the intestine, being therefore excluded
from the digestive gland ducts [14].
The detoxification role of the digestive gland is inferred due to its capacity to
accumulate passively ingested metals, correlated with the occurrence of metal binding
proteins. The accumulation of metals can result in severe damage of the digestive gland
cells, when compared with other regions of the digestive tract [14].
In gastropods, the digestive gland epithelium is mainly formed by digestive and
basophilic cells (Figure 3), but in some species it may contain additional cell types [15,
16]. Digestive cells, also known as absorptive or secretion-reabsorption cells, have a
columnar shape and are the most abundant in the digestive gland of molluscs. They
present an intense endocytic activity, having many heterolysosomes where the
intracellular digestion of food particles occurs. They present lipid droplets and glycogen
granules in the cytoplasm, which can also be found in smaller amounts in basophilic cells
(Figure 3) [13, 14]. Basophilic cells, also known as secretory cells, calcium cells or crypt
cells, typically present a pyramidal shape and contain large amounts of rough
endoplasmic reticulum, a well-developed Golgi apparatus and secretory vesicles, being
considered responsible for secretion of enzymes involved in extracellular digestion
(Figure 3). They are responsible for the metal accumulation related with detoxification
[12, 14], but can also accumulate calcium in concentric structures known as calcium
concretions or spherites [17]. Excretory cells are considered a final stage of digestive
cells, exhibiting a large central vacuole and basal nucleus, and are responsible for the

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
15
excretion of residues into the tubule lumen. Thin columnar undifferentiated cells were
also reported in some species, which may give rise to digestive or basophilic cells [14].
Figure 3 - Schematic representation of the digestive gland cells in gastropods. mv – microvilli, ev – endocytic vesicle, ly –
lysosome, li – lipid droplets, nu – nucleus, sv – secretory vesicles, cc – calcium concretions (provided by A. Lobo-da-
Cunha, University of Porto).
Digestive gland cells are rich in peroxisomes. However, they present some
variability when both basophilic and digestive cells are compared. In basophilic cells,
these organelles are abundant in the middle and basal zones and present a nucleoid
with a variable shape between species. In digestive cells, peroxisomes are also found in
great number in the cytoplasm around vacuoles, typically with smaller size.
Morphological differences between digestive and basophilic cells peroxisomes were
reported in some gastropod species, thus suggesting functional differences [12, 13, 18].
Additionally, it was demonstrated that chemical pollutants can lead to morphological and
biochemical changes in peroxisomes, therefore being considered a valuable bioindicator
of pollution [19].
Despite much being known about the digestive system of gastropods, still many
doubts remain to be clarified. For example, how these cells and systems are regulated
accordingly to the animal’s activity and metabolic demands, is still poorly understood.
Another issue, is the fact that most field collected species are carried and maintained in
laboratory captivity in assumingly good conditions. However, some species of
gastropods are very sensitive to environmental conditions that can lead to drastic
changes of their metabolism, namely the digestive system [14]. Therefore, further
research is required for our better understanding of the metabolic regulation in these
organisms.

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
16
1.3. Oxidation of polyols and aromatic alcohols in gastropods
Ultrastructural analysis of the digestive gland of the sea slug Aplysia depilans
revealed many tubular structures inside the rough endoplasmic reticulum cisternae of
basophilic cells (Figure 4) [12]. Similar structures were also described in terrestrial
gastropods, such as the snail Helix pomatia [20] and the slug Arion empiricorum [21],
and in the digestive cells of the marine species Alderia modesta [22]. These structures
were called “mannosomes” since the enzyme mannitol oxidase, which catalyses the
oxidation of D-mannitol to produce the sugar mannose, releasing hydrogen peroxide
[23], is present in these structures, as shown by histochemical and biochemical
localization [24], and by purification methods [23, 25-28]. These studies confirmed the
capacity to metabolize D-mannitol in terrestrial [24-26] and marine [12] species.
Mannitol is a polyol present in plants, algae and fungi. Therefore, it was suggested
that herbivorous gastropods would get nutritional benefits from the ingestion of polyols,
present in plants [29] and algae [30] of their diet, due to the enzymes that turn these
compounds into sugars [24].
Figure 4 - [A] Ultrastructure of the basophilic cells of the digestive of A. depilans, showing tubular structures in the interior
of rough endoplasmic reticulum cisternae (*) [12]. [B] Frozen section of the digestive gland of A. depilans showing stained
basophilic cells (arrows) after the detection of mannitol oxidase activity [12].
More recently, the presence of mannitol oxidase in the digestive gland of several
herbivorous gastropods (Table S1), including terrestrial, marine and freshwater species,
was confirmed by activity assays [31]. In Patellogastropoda, Neritimorpha and
Vetigastropoda the activity of mannitol oxidase was not detected. However, mannitol
oxidase activity was detected in all herbivorous species belonging to Caenogastropoda

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
17
and Heterobranchia. The tubular structures inside rough endoplasmic reticulum
cisternae, typically associated with mannitol oxidase activity [12, 26], were found in
Aplysia depilans, while in Bulla striata, Cornu aspersum and Lehmannia valentiana, they
were found in smooth membrane cisternae. It was also reported that some of the species
expressing mannitol oxidase activity, such as Siphonaria pectinata and Onchidella
celtica, do not exhibit the typical tubular structures [31]. So, in what concerns marine
gastropods, mannitol oxidase is not always associated with intracisternal tubules even
in species with a high activity, such as O. celtica [31].
Another polyol with both nutritional and osmoregulation effects is the mannitol
isomer, sorbitol [32]. In previous studies (Table S1), sorbitol oxidation was only detected
in herbivorous species. In Caenogastropoda only Littorina littorea does not express
sorbitol oxidase activity. In Heterobranchia this activity is restricted to the Eupulmonata
clade. This data supported the hypothesis that mannitol oxidase can also use sorbitol as
substrate (data not published).
Another interesting finding about alcohol oxidases in the digestive gland of
terrestrial gastropods is their capacity to use aromatic alcohols as substrate [33, 34]. This
type of activity was first reported in the culture fluid of the lignolytic white-rot fungus
Polystictus versicolor [35]. Later, several groups have reported enzymes with the same
activity in a variety of other lignolytic fungi [36, 37]. Fungal aromatic alcohol oxidase
activity correlates with lignin degradation. In gastropods, aromatic alcohol oxidase show
some similarities in terms of pH optimum and substrate preferences [34]. Moreover,
herbivorous gastropods species can consume plant matter that has undergone some
lignification, or contains lignin precursors. Thus, it is likely that these species also have
this oxidase activity. Studies on aromatic alcohol oxidase in gastropods showed that this
enzyme has the highest activity towards cinnamyl alcohol, among several substrates
tested [34]. Surprisingly, our data (Table S1) show that all analysed gastropod species,
except O. celtica, present oxidase activity for cinnamyl alcohol (data not published).
In addition to polyols and aromatic alcohols, low molecular weight alcohols were
also tested. In some species, oxidation of ethanol was detected, however none of them
showed oxidation of methanol. Researchers frequently discuss whether the natural
intake of ethanol, in significant concentrations, originated an adaptation in the organisms
that can metabolize it, or if its capacity to oxidize ethanol is just a non-specific and
incidental response [38-40]. Studies using gastropods revealed that this activity is
present in Patella vulgata, in the carnivore caenogastropod Charonia lampas, and in
Heterobranchia, in the freshwater species P. corneus and in all terrestrial species, the
Stylommatophora (Table S1). However, with this data only it was not possible do
conclude if this capacity is an adaptation or an incidental feature (data not published).

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
18
1.4. Alcohol dehydrogenases
Oxidoreductases catalyse oxidation and reduction reactions where an electron is
transferred from a molecule to another. They can be divided into six categories:
oxygenases, reductases, peroxidases, oxidases, hydrolases and dehydrogenases. This
large group of oxidoreductases shows their importance and functional diversity in the
different organisms [41].
The roles of this group of enzymes are quite wide and comprises many
intermediary metabolic functions such as utilization and detoxification of ethanol and
xenobiotics in general, regulation of hormones and signalling molecules or just sensing
the redox status of the organism, thereby regulating vital cellular processes.
The oxidation of a substrate via dehydrogenase is characterized by the
transference of a hydrogen to an electron acceptor, such as NAD+ or FAD, in which the
substrate is losing electrons [42].
Alcohol dehydrogenases (ADH) are widely distributed among organisms of all
phyla and are responsible for the oxidation of alcohols into aldehydes or ketones [43].
There are three non-homologous NAD(P)+ -dependent alcohol dehydrogenase families
in animals. All of them evolved independently and have different structures and reaction
mechanisms [44, 45].
1.4.1. MDR - Type I ADHs
Type I ADH or “medium-chain” dehydrogenase/reductase (MDR) is the most
studied ADH superfamily in animals. Its functions include the metabolism of ethanol,
hydroxysteroids, lipid peroxidation products, formaldehyde, ω-hydroxy fatty acids and
norepinephrine, nitric oxide homeostasis, and retinoid transformation These are NAD+
and zinc-dependent enzymes, with one catalytic and one structural zinc atom, which
have between 350 and 375 amino acid residues. The region around and after the
structural zinc atom is the most divergent region in all ADH proteins [38, 46].
Based on sequence homology, catalytic activity and gene expression, these
enzymes have been sorted in eight different classes (I-VIII) in animals. In invertebrates,
there are only class III ADHs, in fish there are class I and III, in amphibian there are nine
or ten isozymes distributed by four or six classes, and in mammals there are six different
classes, but primates lack class VI ADH [38]. This multiplicity of ADH’s suggests that,
beyond their metabolic function and organism regulation, these enzymes play a role in

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
19
cellular defence against toxic alcohols and aldehydes, both endogenous and exogenous.
They appear to work together with cytochrome P450 and other oxidative systems, acting
without generation of potentially dangerous reactive oxygen species [47].
Class I Zn-dependent alcohol dehydrogenases
This class of enzymes is the most studied regarding the theme of ethanol-
metabolizing enzymes in different organisms, mainly due to its relevance for the alcoholic
tolerance [48]. They were originated by tandem duplication of class III ADH, 450 million
years ago, back in the origin of bony fish [49]. Since then, multiple independent gene
duplication of these enzymes has occurred in the different animal groups. The more
antique groups of vertebrates, fish and amphibians, have 3-5 and 3-6 class I ADH genes,
respectively. In mammals like cow, dog, mouse, and rat there is only on gene, but in
humans and other primates, there are three gene copies [50]. Additionally, a study to
trace overall patters within mammalian ADH genome, which was performed using data
from 15 genomes, revealed that this class of ADH is divided into three isoforms, ADH I
A, ADH I B and ADH I C [51].
In fish and amphibians, the appearance of these ADH isozymes is not related with
ethanol exposure, since they do not have any type of fruit in their diet. So these animals
have what is called a class I ADH ethanol availability-independent gene multiplicity [38].
In mammals, however, there is a broad array of studies on gene expression induced by
ethanol enriched diets [52], influence of alleles on ethanol-metabolizing processes and
its relation with the tolerance of the organism [48] and even how social-economic factors,
such as alcoholic beverages consumption [53] or pre/pro-industrial societies [54], may
be related with these alcohol-metabolizing enzymes occurrences [55].
Despite being highly known for its ethanol-metabolizing action, the list of substrates
for class I ADH can be quite extensive, from short to long aliphatic alcohols, retinol,
hydroxysteroids, bile acids (ADH I C), and hydroxytryptophol (serotonin metabolism). In
mammals, these enzymes are inhibited by pyrazole which has been effectively used to
distinguish class I ADH activity from other activities [51].
Class II Zn-dependent alcohol dehydrogenases
Traced in amphibians 360 million years ago [49], class II ADH is strongly related
with natural exposure of ethanol. Interestingly, in rodents it shows low affinity for ethanol
when compared with other alcohol dehydrogenases [56], but much higher catalytic

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
20
activity with retinol. This is a good evidence of the non-specific behaviour of this group
of enzymes, regarding its substrates [38].
In human, this class of ADH was isolated as pyrazole insensitive enzyme and is
active towards primary alcohols. In the oxidative pathway, the enzyme has been shown
to be the best hepatic retinol dehydrogenase [57]. In the reductive pathway class II ADH
has low activity for benzoquinones, is active towards lipid peroxidation products and has
a suggested redox-specific function in norepinephrine metabolism. However, the main
function for class II ADH is still an open question [51]
Class III Zn-dependent alcohol dehydrogenases
This is the only class of ADHs present in all eukaryotic organisms, and some
bacteria, being the most conserved ADH from all classes. It is considered the ancestral
form of zinc-containing ADHs from which the other ADHs have evolved. It has been
found in all tissues of animals who have their genome completely sequenced [43].
Compared to the other classes, ADH III has two additional activities. It is also
known as glutathione-dependent formaldehyde dehydrogenase and S-
nitrosoglutathione reductase, being expressed in all species containing glutathione [58,
59]. Regarding mammals, this ADH is the only one that has been traced in the central
nervous system [60, 61].
ADH III has very low affinity for ethanol as substrate [62], being suggested to be
involved in the first-pass metabolism of ethanol [63]. Since ethanol is present at low
concentrations in human diet under natural conditions, mainly as ingested ripe fruits, the
role of class III ADH in ethanol metabolism can be considered as incidental and not
adaptive from an evolutionary perspective [38]. However, class III ADH has high activity
for different ω-hydroxyfatty acid and is considered a formaldehyde scavenger in the
pathway where S-hydroxymethyl glutathione is the actual substrate of this enzyme [64].
Class IV Zn-dependent alcohol dehydrogenases
Described as gastric ADH and non-hepatic ADH [65], the origin of class IV ADHs
can be traced to marsupials, 176 million years ago [49]. This is another class of ADHs
that originated before the dietary ethanol being available by means of fruits fermentation
and, therefore, is non-ethanol metabolizing enzyme class [38].
Regarding the human gastric mucosa, this class of enzymes is responsible for the
maintenance of retinoic acid supply [66] and its malfunction is correlated with progression
of inflammation, atrophy and even intestinal metaplasia [67].

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
21
In several species, class IV ADH has not been identified since the region where
the gene encoding for this enzyme is located is poorly analysed in numerous genomes.
On other hand, in opossum for example, there are several isoforms of these enzymes
[51].
Class V and VI Zn-dependent alcohol dehydrogenases
Class VI ADHs also have origin in marsupial, 176 million years ago, and originated
class V ADHs, 76 million year later, by means of tandem duplication. Class V ADHs
appear to be restricted to placental mammals [49]. These functional equivalent enzymes
are known only in rat, which express two isoforms of ADH VI (A and B), and cow.
Primates lack class VI and mouse and dog lack class V ADHs, being the last one a
possible pseudogene in mouse [51]. However, neither human class V nor rat class VI
have been isolated as active enzymes, which is why their physiological role is still
unknown [68, 69].
Overall, these two classes of ADHs are exceptions and have been identified only
at DNA and mRNA levels. At the transcriptional level, different sizes of mRNA have been
identified for class V ADH, but no stable protein has been isolated [70].
Birds Class VII and amphibians class VIII Zn-dependent alcohol dehydrogenases
Class VII ADH has been found only in chicken with higher activity towards all-trans-
retinol and epiandrosterone. Amphibians class VIII ADH displays a strong preference for
NADP+ instead of NAD+ and higher activity with all-trans-retinaldehyde. Thus, it seems
that birds class VII and amphibians class VIII ADH do not play a role in the ethanol
metabolism [38].
1.4.2. SDR - Type II ADHs
Most short-chain dehydrogenases/reductases (SDH) have less than 250 amino
acid residues. However, one family, in a total of five, presents an extension until 350
residues. It was shown that about ¼ of the total described dehydrogenases are SDR.
These are present in insects, such as the genus Drosophila, but about ¾ of the total
SDR’s are of bacterial origin [42].

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
22
The amino acid sequence of the fruit fly Drosophila melanogaster ADH was
obtained, 36 years ago [71], and is clearly different from the amino acid sequences of
mammalians and yeast ADHs [72, 73]. At that time, several authors proposed the
existence of two different ADHs superfamilies [74], one being the short-chain ADHs from
insects, and the others long-chain ADHs, nowadays called medium-chain ADHs.
The type II ADHs have been extensively studied in the genus Drosophila and do
not share any structural relationship with zinc-dependent ADHs, except for the
coenzyme-binding domain, also known as Rossmann-fold [42]. All-common type I class
III ADHs are also expressed in this genus, in addition to type II. This last one is
responsible for most ethanol dehydrogenase activity in Drosophila [75], being considered
the unique fly enzyme involved in ethanol metabolism [76].
The analysis of ADHs in different families of flies, showed that the best substrates
for Drosophila ADH are secondary alcohols instead the primary ones [77]. Contrarily,
analysis of ADH1 and ADH2 from Ceratitis capitata and Anastrepha fraterculus showed
that ethanol and primary alcohols are better substrates than secondary alcohols [78, 79].
This again confirms the variability that these families of enzymes possess towards their
substrates. The origin and diversification of flies with different short-chain ADH genes
occurred shortly after the origin of angiosperms with fleshy fruits from the Lower
Cretaceous [38],which took place about 130-140 million years ago [80].
In vertebrates, the enzyme that shows more similarity to the type II ADH is the 15-
hydroxyprostaglandin dehydrogenase (PGDH), which also shows a broad phyletic
distribution in all animal groups, from the simplest ones, such as Cnidaria, to the most
complex, like the vertebrates. So, it can be assumed that short-chain ADH from insects
are derived by duplication from PGDHs, since these are of older origin [38].
1.4.3. LDR - Type III ADHs
Long-chain dehydrogenases/reductases (LDH) have more than 375 amino acid
residues and are iron dependent molecules that were initially identified in
microorganisms as ethanol oxidizing enzymes [38]. Later, new members of this
superfamily were found displaying different activities, such as: lactaldehyde reductase
and glycerol dehydrogenase in Escherichia coli [81, 82], butanol dehydrogenase in
Clostridium acetobutylicum [83], ethanolamine utilization protein in Salmonella
typhimurium [84], and 1,3-propanediol dehydrogenase in Klebsiella pneumoniae [85].

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
23
These enzymes conserve both an NAD(P)+ binding site and an iron-binding motif.
This last one has only two of the three conserved histidine residues, which make it unable
to determine the specific iron requirement for each enzyme [86].
In rats and humans, the gene encoding for the LDR ADHFe1 is expressed in adult
liver, kidney, thymus, spleen, hypothalamus and adipocytes [87]. ADHFe1 displays
activity as hydroxyacid-oxoacid transhydrogenase (HOT), which catalyses the
conversion of -hydroxybutyrate into succinic semialdehyde with subsequent reduction
of α-cetoglutarate [86]. -Hydroxybutyrate is an energy regulator that promotes the
release of growth hormone [88], and was used illegally by athletes as a performance-
enhancing drug [89]. This data identifies ADHFe1 as an athletic-performance gene that
was positively selected during 400 years in Thoroughbred horses [90].
1.4.4. Polyol Dehydrogenases
Polyols or alditols are acyclic polyfunctional alcohols, also known as sugar
alcohols, that derive from aldoses or ketoses by reduction of the carbonyl group. They
are specially abundant in algae, fungi and plants [91]. Because of their physico-chemical
properties, these alcohols can be seen as simple carbohydrates. In animals, polyols are
important reserves of metabolites that regulate the density and viscosity of some
biological fluids such as blood, lymph and intercellular fluid [92]. These metabolic by-
products have also been associated with an increase of osmotic pressure that provides
a decrease of fluidity of plasma membranes by glucose derivatives, increasing the
cellular resistance to oxidative stress and some toxic agents [93]. The osmotic regulation
by these compounds was also reported in plants and algae [29, 30].
With a broad array of applications, polyols and their derivatives play an important
role in the production of everyday-products. In health industry, compounds like sorbitol
or xylitol are used as sugar substitutes in patient’s nutrition, allowing a better control of
problems regarding oral health and even diabetes control. Also, some polyol esters are
used as emulsifying agents, explosives and pharmaceuticals. Therefore, numerous
dehydrogenases with different substrates specificities have been characterized along the
years, like those involved in the interconversion of D-mannitol/D-fructose or the in vivo
oxidation of sorbitol to fructose by sorbitol dehydrogenase (SDH) [91].
Described as an additional pathway in glucose metabolism [94], the polyol/sorbitol
pathway (Figure 5) involves aldose reduction (ALR2) that convert aldoses into their
corresponding ketoses, and then SDH [95]. This pathway functions as a bypass to

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
24
glycolysis and the pentose phosphate pathway that was used as therapeutic tool in the
control of hyperglycaemia of some diseases, such as diabetes [96, 97].
Figure 5 - Scheme of the polyol pathway of the glucose metabolism. Conversion of glucose to sorbitol by aldose reductase,
and sorbitol to fructose by sorbitol dehydrogenase [98].
Sorbitol dehydrogenase (Figure 6), or L-iditol-2-dehydrogenase, is a 155 kDa
tetrameric protein, with each subunit having around 38 kDa. It has a total of 356 amino
acid residues and a single zinc atom, therefore belonging to the type I medium-chain,
zinc-dependent dehydrogenase/reductase family, but not fitting in any of the eight ADH
classes early described in animals [91]. It is homologous to the alcohol dehydrogenase,
which has a conserved zinc ligand motif that contains two cysteine residues, one
histidine residue and a hydrophobic substrate-binding pocket. The main difference
between ADH and SDH is the replacement of the third Zn ligand in ADH, a cysteine, by
a glutamine residue in SDH [98].
Figure 6 - Structure of sheep sorbitol dehydrogenase (http://www.rcsb.org/pdb/explore/explore.do?structureId=3QE3).
ALR2 SDH
Glucose Sorbitol Fructose
NADPH NAD+ NADP+ NADH

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
25
Sorbitol dehydrogenase is the second enzyme of the polyol pathway (Figure 5) and
catalyses de conversion of sorbitol to fructose. In the first step of this pathway, glucose
is converted to sorbitol by ALR2, a NADPH-dependent aldose reductase. SDH is widely
distributed in almost all mammalian tissues [99-101]. The coenzyme NADH was found
to bind more tightly to SDH than NAD+, at both high and low pH values [102].
The increased flux of glucose through the polyol pathway, by the action of ALR2,
in a hyperglycaemic state, decreases the NADPH/NADP+ ratio. Thereby, the reduction
of glutathione disulphide is impaired leading an oxidative stress. Additionally, the
increase of sorbitol flux through this pathway, because of the increased SDH activity,
may decrease the cytosolic NAD+/NADH ratio resulting in the block of the glycolytic
pathway at the level of glyceraldeyde-3-phosphate dehydrogenase (GAPDH). The
resulting imbalances of these redox states will further affect several other metabolic
pathways [98].
Several studies highlight the importance of these enzymes in the organism
homeostasis, or even as tool for diseases treatment. Nevertheless, other biochemical
imbalances can lead to its overstimulation. Such is the case of the deleterious effect of
sorbitol accumulation on various tissues, because of an overstimulated ALR2 activity
[103]. Stimulators of this pathway include haloalcohols and detergents that will alter the
catalytic site, resulting in a more flexible configuration, aiding the release of NADH from
the coenzyme binding site, and therefore originating a faster rate-limiting dissociation
process of SDH, when in the presence of high concentrations of sorbitol [104].
Similar maximum velocities with various polyol substrates have been previously
reported for human [105] and rat [106] liver SDH, and human [101], rat and bovine [107]
brain SDH. For many years, there was a lack of consensus in the literature regarding de
substrate specificity of SDH. In a first approach, it was suggested that polyol substrates
of SDH have the D-cis-2,4-dihydroxy (2S,4R) configuration, such is the case of sorbitol
and a primary hydroxyl group adjacent to the oxidation site at C2 [91]. Other studies [91]
have shown that SDH catalyses de reversible NAD-linked oxidation of most polyols,
including D-mannitol and galactitol, the 2- and 4-epimers of sorbitol, which do not
possess that specific configuration, into their corresponding ketones. Therefore, the
supposed D-cis-2,4-dihydroxy requirement for the polyol substrates of SDH is not
absolute [108]. Table 1 shows a list of possible substrates for human liver SDH.

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
26
Table 1 - Compatible substrates for human liver sorbitol dehydrogenase [105]
substrate relative rate
sorbitol 100
glycerol 6
erythritol 4
L-threitol 110
ribitol 85
xylitol 103
L-arabinitol 29
D-mannitol 8
galactitol 11
Some reports show that mammalian liver SDH can also use aromatic secondary
alcohols as substrate [109]. Overall, SDH does not bind substrates with C2 configuration
of a polyol and the length of the carbon chain determines the degree of enzymatic
discrimination. The C2 hydroxyl group is coordinated to the Zn atom while the primary
hydroxyl group and the substituent sits in a polar region and in a hydrophobic area,
respectively [91].
1.5. Alcohol and polyol dehydrogenases in molluscs
Molluscs are the second major phylum of multicellular animals. However, that fact
is not reflected in the number of studies of alcohol and polyol dehydrogenases in these
organisms.
A study with cephalopods molluscs (octopus, squid and cuttlefish) showed the
presence of only class III alcohol dehydrogenase, with a wide tissue distribution. This
enzyme behaves like the other class III alcohol dehydrogenases identified in other
organisms, with very low affinity for ethanol and high glutathione-dependent
formaldehyde dehydrogenase activity, proving that multicellular organisms can survive
without ethanol active alcohol dehydrogenases. No other class of alcohol
dehydrogenase enzyme was found in the cephalopod species or organ tissues tested.
In fact, cephalopods were the first multicellular organisms known without significant
amounts of alcohol dehydrogenase forms capable of metabolizing low molecular weight
alcohols, such as ethanol [110].
Biomphalaria glabrata is a gastropod that has been extensively studied in the
matter of public health. To develop mechanisms for the suppression of natural

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
27
populations of this species, several strategies of genetic manipulation have been used.
Some techniques only produce effect after several generations, so it is important to
understand the polymorphisms and inheritance pattern of certain enzymes. Therefore,
there are some studies regarding alcohol dehydrogenases in these species [111].
Additionally, due to the fact of being a subject of extensive investigation, this freshwater
snail is one of the few molluscs with its genome fully sequenced.
Class III alcohol dehydrogenase of the marine species Nucella lapillus was studied
in the context of environmental contamination with endocrine disrupting chemicals.
Chemicals of high persistence and bioaccumulation are used in agriculture, industry and
antifouling paints for boats and fishing nets, and can affect the endocrine system of
several organisms, leading to deregulation of important physiological functions. Studies
with ascidians showed that class III ADH gene expression was strongly downregulated
upon tributyltin exposure. The disruptive consequences of this pollutant in
caenogastropods, such is the case of N. lapillus, is the imposex phenomenon by altering
retinoid X receptors activation. That leads to the development of male secondary sexual
organs in females. Therefore, these animals play a role of major importance as
bioindicators of water contamination. However, in this study, although tributyltin induced
imposex after two-month exposure, class III ADH expression of N. lapillus was not
significantly altered. On the other hand, retinol significantly downregulated class III ADH
levels in female gonads [112].
Regarding organism homeostasis, sorbitol dehydrogenase plays a key role in the
regulation of sorbitol concentration and therefore the viscosity of biological fluids in the
freshwater snail Viviparus viviparus. This way, the organism can adjust some physical
parameters of its liquid phase, such as decreasing the freezing temperature. This stress-
induced cryoprotecting feature is particularly useful for poikilothermic animals allowing
them to resist to condition of low temperature [32],
1.6. Aims of this work
At this point it is quite evident the lack of knowledge about the alcoholic
dehydrogenases in molluscs. Therefore, the objective of this work was to study how
aromatic alcohol and polyol dehydrogenases are distributed among the different clades
of gastropods and how that can be related with their dietary habits. This work also aimed
to isolate and identify the enzyme(s) responsible for the metabolization of specific
substrates in the true limpet Patella vulgata.

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
28
2. Materials and Methods
2.1. Species and sampling sites
Most of the marine species used for this study were collected in the North coastal
region of Portugal: Patella vulgata, Phorcus lineatus (=Monodonta lineata), Gibbula
umbilicalis, Calliostoma zizyphinum, Nassarius reticulatus, Charonia lampas, Nucella
lapillus, Aplysia depilans, Ocenebra erinaceus and Siphonaria pectinata. From Ria de
Aveiro, a coastal lagoon connected to the Atlantic Ocean on the central coastline of
Portugal, was obtained the common periwinkle Littorina littorea. The sea slugs Aglaja
tricolorata, Pleurobranchaea meckelii and Armina maculata were collected in Troia
beach near Setubal. The marine species Hexaplex trunculus, Doriopsilla areolata, Bulla
striata, Haminoea orbignyana and Onchidella celtica were obtained on the South coast
of Portugal (Algarve). The freshwater snails Neritina natalensis, Pomacea bridgesii, and
Marisa cornuarietis were purchased in aquarium shops. Planorbarius corneus
exemplars, also a freshwater snail, were offered by Centro de Educação Ambiental das
Ribeiras de Gaia (CEAR). The terrestrial species Cornu aspersum (=Helix aspersa),
Lehmannia valentiana and Arion ater were collected in the North of Portugal. The
taxonomic position, habitat and diet of these species are included in Table 2.
2.2. Preparation of protein extracts
The digestive gland of each animal was removed and homogenized at 1000 rpm
in a Potter-Elvehjem homogenizer at 4 ºC, using an homogenizing medium for marine
invertebrates (50 mM Tris-HCl buffer pH 7.4, 500 mM sucrose, 150 mM KCl, 1 mM EDTA
and 1 mM PMSF), in an average concentration 0,06 g/ml of digestive gland (adapted
from [113, 114]). The homogenates were then sonicated in three cycles, 15 s each, in
ice-water bath (Bandelin – Sonorex RK 100H sonicator) and centrifuged at 1000 g for 5
min. at 4 ºC. The supernatants were used to determine enzyme activities.
FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
29
Table 2 - Taxonomy, habitat and diet of the 26 species of gastropods included in this study. Ma - marine, Fw - freshwater, Te - terrestrial
Gastropod clades Superfamily Family Species Habitat / Diet
Patellogastropoda Patelloidea Patellidae Patella vulgata Ma / Herbivore
Neritimorpha Neritoidea Neritidae Neritina natalensis Fw / Herbivore
Vetigastropoda Trochoidea Trochidae
Phorcus lineatus Ma / Herbivore
Gibbula umbilicalis Ma / Herbivore
Calliostomatidae Calliostoma zizyphinum Ma / Carnivore
Caenogastropoda
Hypsogastropoda
Buccinoidea Nassariidae Nassarius reticulatus Ma / Carnivore
Tonnoidea Ranellidae Charonia lampas Ma / Carnivore
Muricoidea Muricidae
Nucella lapillus Ma / Carnivore
Hexaplex trunculus Ma / Carnivore
Ocenebra erinaceus Ma / Carnivore
Littorinoidea Littorinidae Littorina littorea Ma / Herbivore
Ampullarioidea Ampullariidae Pomacea bridgesii Fw / Herbivore
Marisa cornuarietis Fw / Herbivore
Heterobranchia
Nudipleura
Pleurobranchomorpha Pleurobranchoidea Pleurobranchidae Pleurobranchaea meckelii Ma / Carnivore
Nudibranchia Doridacea Phyllidioidea Dendrodorididae Doriopsilla areolata Ma / Carnivore
Cladobranchia Arminoidea Arminidae Armina maculata Ma / Carnivore
Euopisthobranchia Cephalaspidea
Philinoidea Aglajidae Aglaja tricolorata Ma / Carnivore
Haminoeoidea Haminoeidae Haminoea orbignyana Ma / Herbivore
Bulloidea Bullidae Bulla striata Ma / Herbivore
Anaspidea Aplysioidea Aplysiidea Aplysia depilans Ma / Herbivore
Pan
pu
lmo
nat
a Siphonarioidea Siphonaridae Siphonaria pectinata Ma / Herbivore
Hygrophila Planorboidea Planorbidae Planorbarius corneus Fw / Herbivore
Eupulmonata
Systellommatophora Onchidioidea Onchidiidae Onchidella celtica Ma / Herbivore
Stylommatophora
Helicoidea Helicidae Cornu aspersum Te / Herbivore
Arionoidea Arionidae Arion ater Te / Omnivore
Limacoidea Limacidae Lehmannia valentiana Te / Herbivore

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
30
For preparation of cytosolic fractions of the digestive gland of Patella vulgata, total
homogenates with 10% digestive gland tissue (4 g of digestive gland in a 40-ml
homogenate), were obtained using a salt-free homogenizing medium (50 mM Tris-HCl
buffer pH 7.4, 500 mM sucrose, 1 mM EDTA and 1 mM PMSF). Samples were
centrifuged at 15 000 g for 15 min. at 4 ºC and then at 100 000 g for 1 h, at 4 ºC (Thermo
Scientific; Sorvall Lynx 6000, T29 - 8x50 ml rotor).
The Bradford method with bovine serum albumin standards was used for protein
quantification [115].
2.3. Enzyme activities
The specific activity of dehydrogenases was determined by using
spectrophotometric assays.
Twenty µl of cytosolic sample were added to 980 µl incubation medium (50 mM
glycine-NaOH buffer pH 10, 5 mM NAD+) supplemented with 50 mM D-sorbitol, 50 mM
D-mannitol or 25 mM cinnamyl alcohol (adapted from [110]). The formation of NADH
(ɛ340 nm = 6.22 mM-1 cm-1) [29] was followed by measuring O.D.340 at 25 ºC, during 200 to
300 sec. A reference cuvette with the incubation medium without sample was used. The
presented values of activity in the total homogenates are mean of at least 5 independent
experiments, and were calculated by using the following equation: Activity (nmol-1ml-
1min-1) = (∆ O.D.340 min. / 0.00622 µM-1) × 50. These assays were done using a double-
beam spectrophotometer (Jenway 68000 UV/Vis.), coupled to a water recirculating
system for temperature regulation in the cuvette.
The activity of specific dehydrogenases was also studied in situ, after separation
of the cytosolic fraction proteins (60-100 µg) by native gel electrophoresis, using 7.5%
non-denaturant polyacrylamide gels. After electrophoresis, the gels were the incubated
in a medium containing 50 mM Tris-HCl buffer pH 8.0, 0.01 mg/ml phenazine
methosulphate, 5 mM NAD+, 0.10 mg/ml NBT (nitro blue tetrazolium chloride) and the
alcoholic substrate (above mentioned concentrations), until the appearance of the
bands. In the control gel the same process was executed but without the alcoholic
substrate (adapted from [116]).

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
31
2.4. Affinity chromatography
The sample (one column volume; 5 ml) was loaded on a Blue Sepharose CL-6B
column and the flow through was recovered. The column was washed with five volumes
(25 ml) of 0.1 M Tris-HCl pH 7.4 buffer. The first 2 volumes were recovered for the control
of activity lost at this step. Then, proteins bound to the column were eluted with one
column volume of increasing concentrations of NAD+ solution (0.05 M, 0.1 M, 0.2 M).
Each of these fractions was recovered independently. Column regeneration was
performed by washing with 2 column volumes, 3 to 5 times, of alternating high (0.1 M
Tris-HCl pH 8.5, 0.5 M NaCl) and low (0.1 M Sodium Acetate pH 4.5, 0.5 M NaCl) buffers.
The NAD+ eluted fractions were concentrated in a Vivaspin 500 Protein Concentrator (6
ml, MWCO 10 000, GE Healthcare Life Sciences) to a final volume around 2.5 ml, due
to its high dilution.
2.5. Ion exchange chromatography
For cation exchange chromatography two columns were used: HiTrap SP HP, 1
ml, controlled by ÄKTAprime plus, a low pressure chromatographic system (GE
Healthcare Life Sciences) and Mono Q 4.6/100 PE controlled by BioLogic DuoFlow™, a
medium pressure chromatographic system (Bio-Rad).
Prior to sample injection, the columns were washed with volumes of alternating low
salt (20 mM Tris-HCl pH 7.4), high salt (20 mM Tris-HCl pH 7.4 1 M NaCl) and low salt
buffers. Concentrated 10 mM NAD+ fractions obtained by affinity chromatography were
diluted with 20 mM Tris-HCl pH 7.4 buffer, for conductivity adjustment, before being
injected in these systems with the aid of a peristaltic bomb. After injection, the column
was washed one more time with low salt buffer for non-bound protein removal. The
columns were then eluted with an ascending and continuous NaCl gradient from 0.05 M
to 1 M, at a flow rate of 1 ml per sec.
2.6. SDS-polyacrylamide gel electrophoresis
To study the protein profile of the samples resulting from the chromatographic
processes, SDS-polyacrylamide precast gels were used (Bio-Rad 12% Mini-

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
32
PROTEAN® TGX™ Precast Protein Gels). Two hundred µl of sample were mixed with
25 µl of 100% TCA, incubated on ice for 5 min., and centrifuged at 10 000 g for 5 min.
The pellet was washed abundantly with acetone to remove the acid, dissolved in 20 µl
of sample buffer (0.25 M Tris-HCl pH 6.8, 8% SDS, 40% glycerol, 20% 2-
mercaptoethanol) and heated at 90 ºC for 5 min. The samples and molecular weight
standards solution (GE Healthcare Life Sciences LMW-SDS Marker Kit: Phosphorylase
b, 97 kDa; Albumin, 66 kDa; Ovalbumin, 45 kDa; carbonic anhydrase, 30 kDa; trypsin
inhibitor, 20.1 kDa; -lactalbumin, 14.4 kDa) were then separated by electrophoresis and
the proteins were stained for 2 hours, using 0,025 M Coomassie Brilliant Blue R-250,
40% methanol, 7% acetic acid). The gel was destained using a 50% methanol, 10%
acetic acid solution.
2.7. Protein sequencing
Coomassie-blue stained protein bands were excised from a SDS-polyacrylamide
gel and analysed by mass spectrometry (MS). Proteins were in gel digested with trypsin
(Promega, USA) and the peptides were extracted using a 2.5% trifluoroacetic acid
solution. Protein digests were desalted and concentrated using C18 ZipTips (Millipore,
USA) and crystallized onto a MALDI plate using α-Cyano-4-hydroxycinnamic acid as a
matrix. Samples were analysed using the 4800 Plus MALDI-TOF/TOF (SCIEX, USA)
[117]. Protein identification was performed by Peptide Mass Fingerprint with Mascot
software (Matrix Science, UK). Some peptides were further characterized by MS/MS
sequencing. The identification of peptides from acquired MS+MS/MS spectra was
performed using a Patella vulgata database (available at:
https://ora.ox.ac.uk/objects/uuid:6471e7d4-dd34-4eb3-883f-a5c438f11741). A Blastp of
the identified proteins was performed at the NCBI database.

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
33
3. Results
3.1. Enzymatic activities
To analyse the distribution of polyol and aromatic alcohol dehydrogenases in five
major clades of gastropods, the activity of these enzymes was quantified using total
homogenates of the digestive gland of 26 species of gastropods.
Regarding the dehydrogenase for the polyols D-mannitol and D-sorbitol (Table 3),
almost all species presented activity for both substrates, with a few exceptions. The
species B. striata and P. meckelii only presented activity for sorbitol whereas the
carnivore vetigastropod C. zizyphinum, the caenogastropods N. reticulatus and H.
trunculus, and the Heterobranchia A. maculata, did not exhibit dehydrogenase activity
for the tested polyols.
Among the species with dehydrogenase activity for both polyols (Table 3), P.
vulgata presented high activity values for both substrates. However, sorbitol
dehydrogenase activity in M. cornuarietis was more than two-fold higher comparing to
that observed in P. vulgata. It was observed that only P. vulgata, P. lineatus and G.
umbilicalis did not present higher dehydrogenase activity for sorbitol than mannitol.
Overall, there was a tendency for higher sorbitol dehydrogenase activities than mannitol
dehydrogenase activities, and the carnivores presented lower dehydrogenase activities
for these substrates than the herbivores.
The dehydrogenase activity for cinnamyl alcohol, representing the aromatic
alcohols, was present in all species, without exceptions (Table 3), with the marine
herbivore Haminoea orbignyana from Heterobranchia exhibiting higher levels. However,
the carnivore N. lapillus also presented a high dehydrogenase activity for cinnamyl
alcohol. Overall, the four studied freshwater species presented low activity values.
Regarding dehydrogenase activities towards the low molecular weight alcohols,
ethanol dehydrogenase activity was only detected, at low levels, in two herbivorous
Eupulmonata species, the marine O. celtica and the terrestrial C. aspersum (Table 3).
No species presented methanol dehydrogenase activity.

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
34
Table 3 - Activity of dehydrogenase in the digestive gland of gastropods. Values are mean ± SD of at least 5 independent experiments (nmol.min-1. g-1 ± SD). Shaded lines correspond to carnivorous
species.
Species Mannitol Sorbitol Cinnamyl alcohol Ethanol
Patellogastropoda Patella vulgata 1,836.1 ± 368.6 1,644.3 ± 723.0 527.6 ± 154.8 -
Neritimorpha Neritina natalensis 870.2 ± 455.9 900.5 ± 356.4 215.4 ± 67.5 -
Vetigastropoda
Phorcus lineatus 1,214.4 ± 340.8 536.6 ± 211.2 1,205.8 ± 424.3 -
Gibbula umbilicalis 1,322.9 ± 244.5 818.9 ± 320.0 560.7 ± 125.2 -
Calliostoma zizyphinum - - 643.6 ± 197.7 -
Caenogastropoda
Nassarius reticulatus - - 1,385.0 ± 392.8 -
Charonia lampas 161.9 ± 60.4 418.4 ± 115.0 824.8 ± 240.5 -
Nucella lapillus 96.2 ± 97.1 381.4 ± 164.8 2,048.5 ± 670.5 -
Hexaplex trunculus - - 894.5 ± 208.9 -
Ocenebra erinaceus 109.1 ± 122.5 254.1 ± 332.5 1,007.4 ± 515.7 -
Littorina littorea 510.2 ± 157.5 944.9 ± 293.8 932.2 ± 432.7 -
Pomacea bridgesii 379.6 ± 65.3 741.0 ± 191.8 181.9 ± 54.5 -
Marisa cornuarietis 1,428.9 ± 232.1 4,095.5 ± 603.3 503.9 ± 75.2 -
Heterobranchia
Pleurobranchaea meckelii - 60.7 ± 68.0 498.0 ± 89.3 -
Doriopsilla areolata 202.0 ± 111.0 327.3 ± 141.9 117.2 ± 39.8 -
Armina maculata - - 1,109.5 ± 279.6 -
Aglaja tricolorata 133.6 ± 71.5 222.3 ± 89.4 1,482.8 ± 205.9 -
Haminoea orbignyana 224.0 ± 32.7 521.3 ± 139.5 2,643.5 ± 996.5 -
Bulla striata - 192.7 ± 82.6 2,017.4 ± 702.2 -
Aplysia depilans 131.5 ± 53.5 530.2 ± 150.1 727.2 ± 248.7 -
Siphonaria pectinata 333.8 ± 116.3 460.2 ± 79.7 1,013.7 ± 372.0 -
Planorbarius corneus 281.3 ± 140.7 847.9 ± 288.6 209.2 ± 60.6 -
Onchidella celtica 623.8 ± 235.5 865.7 ± 157.4 1,604.7 ± 389.0 307.9 ± 186.9
Cornu aspersum 546.2 ± 207.9 1,171.1 ± 335.8 1,329.6 ± 282.5 300.8 ± 137.2
Arion ater 651.6 ± 309.6 1,058.7 ± 501.1 1,264.5 ± 417.6 -
Lehmannia valentiana 437.4 ± 103.3 1,408.9 ± 319.2 978.1 ± 231.0 -

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
35
The activity of dehydrogenases for the polyols D-mannitol and D-sorbitol and for
the aromatic cinnamyl alcohol was also assessed in situ, after native gel electrophoresis
(Figure 7), using four marine herbivorous species: P. vulgata, P. lineatus, L. littorea and
S. pectinata. Regarding the polyols dehydrogenases, a similar overall profile was
observed in all species. For P. vulgata, L. littorea and S. pectinata, one main band of
enzyme activity was detected. Interestingly, sorbitol dehydrogenase activity was higher
when comparing with mannitol dehydrogenase. In L. littorea a second well defined band
appeared for each polyol, again with higher intensity for sorbitol. In P. lineatus, a main
band of sorbitol dehydrogenase activity was detected, whereas for mannitol
dehydrogenase two bands appeared, one of them with lower relative migration distance.
Figure 7 - Native electrophoresis gel sections with in situ detection of the activity of dehydrogenases of several alcoholic
substrates, in four marine herbivorous species.
Patella vulgata Phorcus lineatus
Littorina littorea Siphonaria pectinata
Mannitol Sorbitol Cinnamyl A. Mannitol Sorbitol Cinnamyl A.
Mannitol Sorbitol Cinnamyl A. Mannitol Sorbitol Cinnamyl A.

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
36
The profiles for cinnamyl alcohol dehydrogenase (Figure 7) revealed only one band
of enzyme activity in P. vulgata, while in the other species more bands were detected.
The faster migration band had a higher intensity, except for P. lineatus, in which the band
with lower relative migration distance exhibited a slightly higher intensity. L. littorea was
the species that presented more bands associated with cinnamyl alcohol dehydrogenase
activity (three).
3.2. Protein isolation and identification
Due to the significant amounts of alcoholic dehydrogenase activities, especially for
polyols, observed in previous studies (Table 3), the facility in obtaining good amount of
digestive gland per animal for sample preparation, and the fact of being an all-year
abundant species of easy collection, the species P. vulgata was used for the isolation
and identification of dehydrogenases.
First, proteins (cytosolic fraction) of this species were separated by affinity
chromatography, using Blue Sepharose CL-6B, which binds most enzymes requiring
adenyl-containing cofactors including NAD+. Proteins were eluted with NAD+ (5-20 mM)
and dehydrogenase activities were determined in the different fractions obtained (Table
4). The results show that the polyols dehydrogenases were more concentrated in the
washing steps after the sample being loaded in the column, and in the fractions eluted
with 5 and 10 mM NAD+. On the other hand, the enzymes responsible for cinnamyl
alcohol dehydrogenase were eluted with 20 mM NAD+. Thus, to obtain a fraction
enriched in polyol dehydrogenases for subsequent chromatographic tests, the Blue
Sepharose CL-6B column was eluted with 10 mM NAD+ (Table 5).
After Vivaspin concentration, the 10 mM NAD+ affinity chromatography fraction
was loaded in the HiTrap SP HP chromatography column and proteins were eluted using
NaCl gradient (Figure 8). The chromatogram shows 2 major protein peaks around the
fraction 22 and 25 (blue line). The activity for the alcoholic substrates was measured in
fractions 4 (flow through), 22, 25 and 27 (Table 6). Since high specific activities for
polyols dehydrogenase were identified in the fractions 22 and 25, samples 22 to 26 were
analysed by SDS-PAGE (Figure 9).
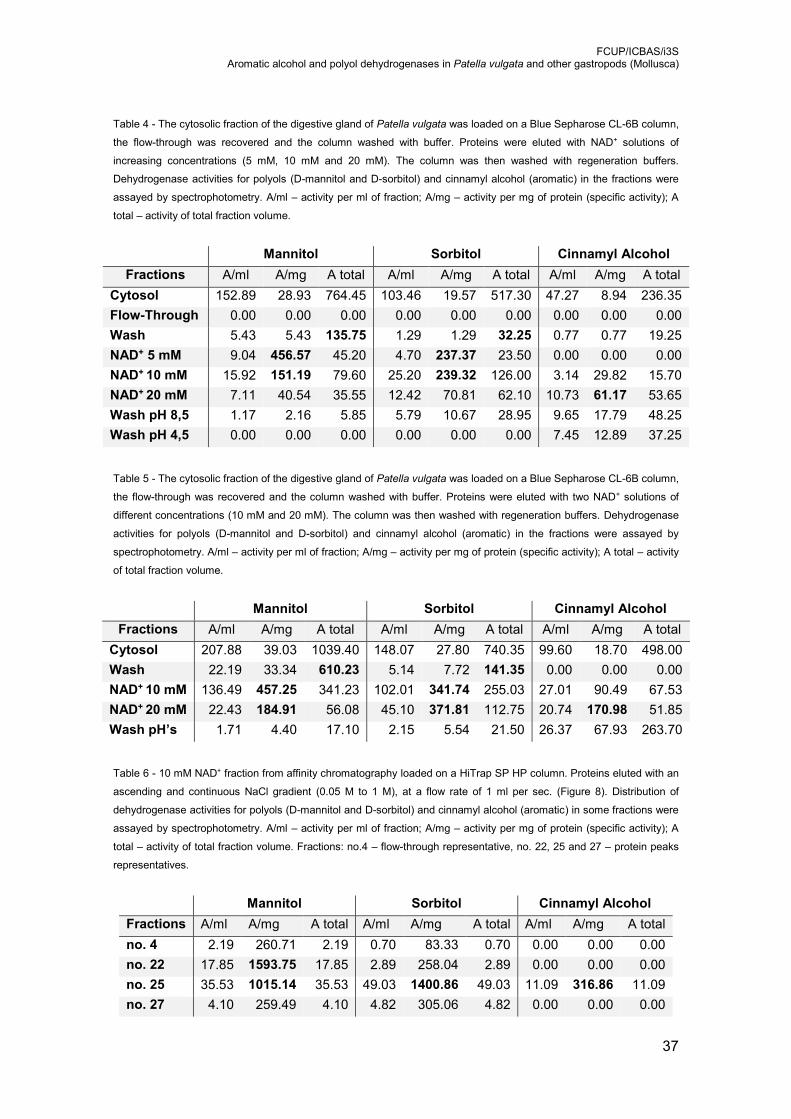
FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
37
Table 4 - The cytosolic fraction of the digestive gland of Patella vulgata was loaded on a Blue Sepharose CL-6B column,
the flow-through was recovered and the column washed with buffer. Proteins were eluted with NAD+ solutions of
increasing concentrations (5 mM, 10 mM and 20 mM). The column was then washed with regeneration buffers.
Dehydrogenase activities for polyols (D-mannitol and D-sorbitol) and cinnamyl alcohol (aromatic) in the fractions were
assayed by spectrophotometry. A/ml – activity per ml of fraction; A/mg – activity per mg of protein (specific activity); A
total – activity of total fraction volume.
Mannitol Sorbitol Cinnamyl Alcohol
Fractions A/ml A/mg A total A/ml A/mg A total A/ml A/mg A total
Cytosol 152.89 28.93 764.45 103.46 19.57 517.30 47.27 8.94 236.35
Flow-Through 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00
Wash 5.43 5.43 135.75 1.29 1.29 32.25 0.77 0.77 19.25
NAD+ 5 mM 9.04 456.57 45.20 4.70 237.37 23.50 0.00 0.00 0.00
NAD+ 10 mM 15.92 151.19 79.60 25.20 239.32 126.00 3.14 29.82 15.70
NAD+ 20 mM 7.11 40.54 35.55 12.42 70.81 62.10 10.73 61.17 53.65
Wash pH 8,5 1.17 2.16 5.85 5.79 10.67 28.95 9.65 17.79 48.25
Wash pH 4,5 0.00 0.00 0.00 0.00 0.00 0.00 7.45 12.89 37.25
Table 5 - The cytosolic fraction of the digestive gland of Patella vulgata was loaded on a Blue Sepharose CL-6B column,
the flow-through was recovered and the column washed with buffer. Proteins were eluted with two NAD+ solutions of
different concentrations (10 mM and 20 mM). The column was then washed with regeneration buffers. Dehydrogenase
activities for polyols (D-mannitol and D-sorbitol) and cinnamyl alcohol (aromatic) in the fractions were assayed by
spectrophotometry. A/ml – activity per ml of fraction; A/mg – activity per mg of protein (specific activity); A total – activity
of total fraction volume.
Mannitol Sorbitol Cinnamyl Alcohol
Fractions A/ml A/mg A total A/ml A/mg A total A/ml A/mg A total
Cytosol 207.88 39.03 1039.40 148.07 27.80 740.35 99.60 18.70 498.00
Wash 22.19 33.34 610.23 5.14 7.72 141.35 0.00 0.00 0.00
NAD+ 10 mM 136.49 457.25 341.23 102.01 341.74 255.03 27.01 90.49 67.53
NAD+ 20 mM 22.43 184.91 56.08 45.10 371.81 112.75 20.74 170.98 51.85
Wash pH’s 1.71 4.40 17.10 2.15 5.54 21.50 26.37 67.93 263.70
Table 6 - 10 mM NAD+ fraction from affinity chromatography loaded on a HiTrap SP HP column. Proteins eluted with an
ascending and continuous NaCl gradient (0.05 M to 1 M), at a flow rate of 1 ml per sec. (Figure 8). Distribution of
dehydrogenase activities for polyols (D-mannitol and D-sorbitol) and cinnamyl alcohol (aromatic) in some fractions were
assayed by spectrophotometry. A/ml – activity per ml of fraction; A/mg – activity per mg of protein (specific activity); A
total – activity of total fraction volume. Fractions: no.4 – flow-through representative, no. 22, 25 and 27 – protein peaks
representatives.
Mannitol Sorbitol Cinnamyl Alcohol
Fractions A/ml A/mg A total A/ml A/mg A total A/ml A/mg A total
no. 4 2.19 260.71 2.19 0.70 83.33 0.70 0.00 0.00 0.00
no. 22 17.85 1593.75 17.85 2.89 258.04 2.89 0.00 0.00 0.00
no. 25 35.53 1015.14 35.53 49.03 1400.86 49.03 11.09 316.86 11.09
no. 27 4.10 259.49 4.10 4.82 305.06 4.82 0.00 0.00 0.00

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
38
Figure 8 - Ion exchange chromatogram of 10 mM NAD+ fraction from affinity chromatography with HiTrap SP HP column. Proteins eluted with an ascending and continuous NaCl gradient (0.05 M to 1
M), at a flow rate of 1 ml per sec. (blue) protein detection (abs 280 nm); (green) high salt solution concentration; (red) conductivity.

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
39
Figure 9 - SDS-polyacrylamide gel showing the enzymatic profile of some ion exchange chromatography fractions
obtained with HiTrap SP HP column (Figure 8). Proteins identified by MS/MS using a database of Patella vulgata: 1 –
glutathione-S-transferase mu class; 2 – no statistical significant identification; 3 – D-arabinitol dehydrogenase 1; 4 –
cytosolic malate dehydrogenase; 5 – sorbitol dehydrogenase-like protein (Table 7).
Some bands of interest were excised from the gel and proteins were analysed by
mass spectrometry (MS/MS) and identified recurring to a Patella vulgata database (Table
7). The following proteins were identified in fraction 22, where the specific activity for
mannitol was much higher than for sorbitol: glutathione-S-transferase and D-arabinitol
dehydrogenase. The analysis of another band around 40 kDa did not give a statistically
valid identification. In fraction 25, where the specific activity is high for both substrates
but higher for sorbitol (Table 6), a cytosolic malate dehydrogenase and a sorbitol
dehydrogenase-like protein were identified (Table 7). Also, the specific activity for
cinnamyl alcohol was higher in fraction 25.
1
2
3
4
5
14
20
30
45
66
97
LMW
kDa 22 23 24 25 26
Fractions
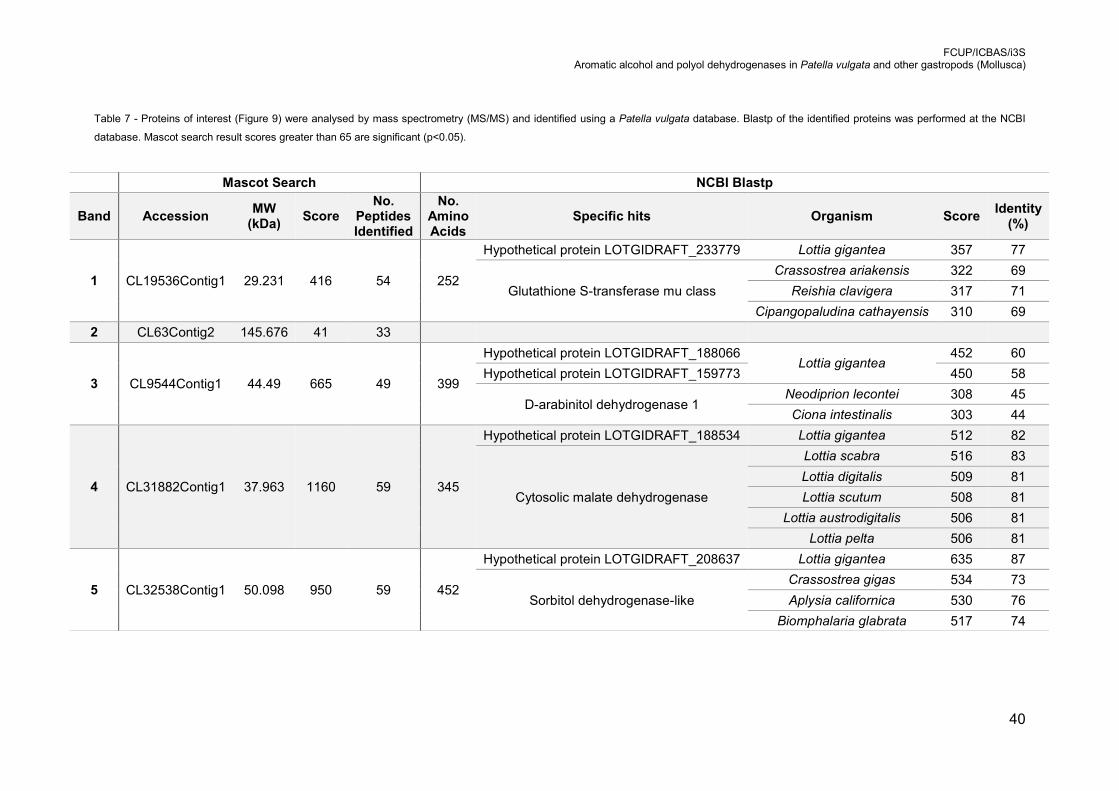
FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
40
Table 7 - Proteins of interest (Figure 9) were analysed by mass spectrometry (MS/MS) and identified using a Patella vulgata database. Blastp of the identified proteins was performed at the NCBI
database. Mascot search result scores greater than 65 are significant (p<0.05).
Mascot Search NCBI Blastp
Band Accession MW
(kDa) Score
No. Peptides Identified
No. Amino Acids
Specific hits Organism Score Identity
(%)
1 CL19536Contig1 29.231 416 54 252
Hypothetical protein LOTGIDRAFT_233779 Lottia gigantea 357 77
Glutathione S-transferase mu class
Crassostrea ariakensis 322 69
Reishia clavigera 317 71
Cipangopaludina cathayensis 310 69
2 CL63Contig2 145.676 41 33
3 CL9544Contig1 44.49 665 49 399
Hypothetical protein LOTGIDRAFT_188066 Lottia gigantea
452 60
Hypothetical protein LOTGIDRAFT_159773 450 58
D-arabinitol dehydrogenase 1 Neodiprion lecontei 308 45
Ciona intestinalis 303 44
4 CL31882Contig1 37.963 1160 59 345
Hypothetical protein LOTGIDRAFT_188534 Lottia gigantea 512 82
Cytosolic malate dehydrogenase
Lottia scabra 516 83
Lottia digitalis 509 81
Lottia scutum 508 81
Lottia austrodigitalis 506 81
Lottia pelta 506 81
5 CL32538Contig1 50.098 950 59 452
Hypothetical protein LOTGIDRAFT_208637 Lottia gigantea 635 87
Sorbitol dehydrogenase-like
Crassostrea gigas 534 73
Aplysia californica 530 76
Biomphalaria glabrata 517 74

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
41
The same affinity chromatography fraction (10 mM NAD+) was then processed by
ion exchange chromatography, using a higher performance column (Mono Q 4.6/100
PE). In this analysis, several protein peaks were observed between fractions 11 and 25
(Figure 10). Therefore, the activity of polyol dehydrogenases was quantified in these
fractions and the protein profile analysed by SDS-PAGE (Figure 11). Once again, two
peaks of activity were detected, the first one with slightly higher activity for mannitol than
sorbitol around fraction 15 and 16, a second peak with an activity for sorbitol two-fold
higher than mannitol around fraction 22.
Seven protein bands of interest, detected after SDS-PAGE of these fractions, were
analysed by MS/MS and further identified (Table 8), using a Patella vulgata database
and by performing a Blastp of the protein at the NCBI database. Bands 1 and 2, from
fractions 16 and 20, respectively, were both identified as being a class III alcohol
dehydrogenase. Band 3 from fraction 22 was identified as a universal stress protein.
Bands 4 and 5, from fraction 15, were both identified as D-arabinitol dehydrogenase 1.
However, this last identified protein was present in low quantity in the samples.
Therefore, 25 of the most abundant peptide ions of each sample were analysed and
glyceraldehyde-3-phosphate dehydrogenase was identified with statistical significance.
Finally, bands 6 and 7, from fraction 22, were identified as sorbitol dehydrogenase-like
protein.

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
42
Figure 10 - Ion exchange chromatogram of 10 mM NAD+ fraction from affinity chromatography with Mono Q 4.6/100 PE column Proteins eluted with an ascending and continuous NaCl gradient (0.05
M to 1 M), at a flow rate of 1 ml per sec. (green) protein quantification (abs 280 nm); (black) high salt solution concentration; (red) conductivity.
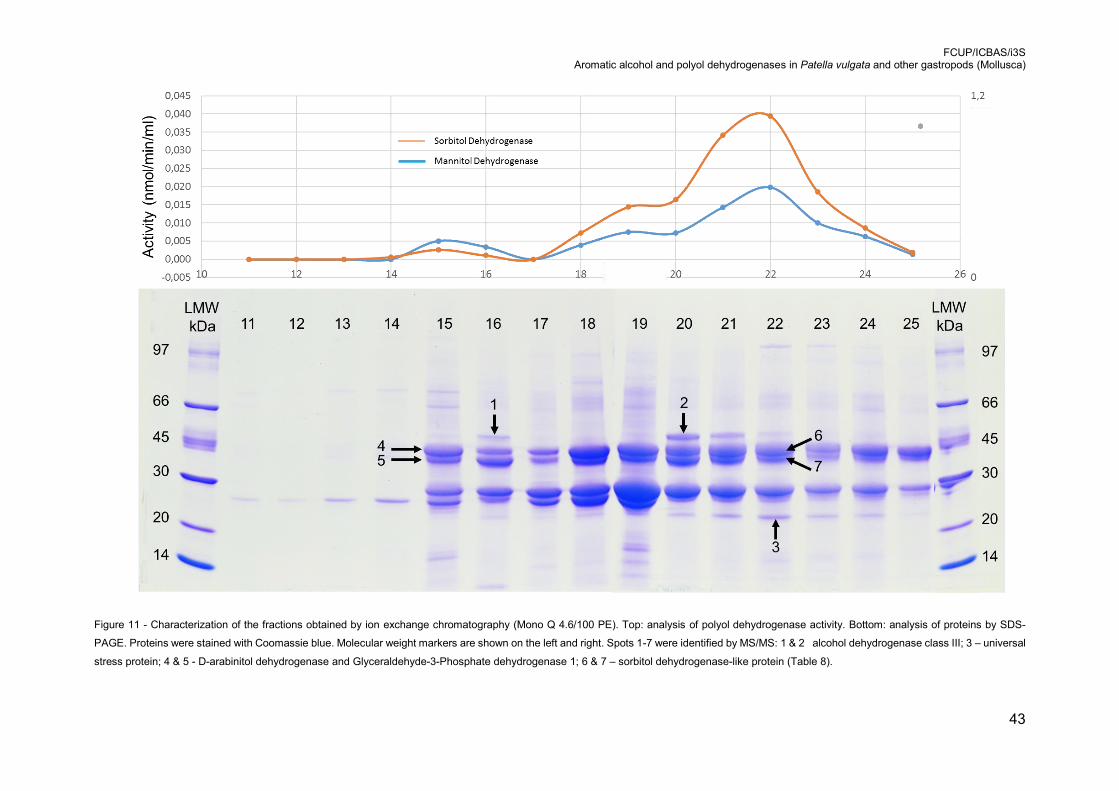
FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
43
Figure 11 - Characterization of the fractions obtained by ion exchange chromatography (Mono Q 4.6/100 PE). Top: analysis of polyol dehydrogenase activity. Bottom: analysis of proteins by SDS-
PAGE. Proteins were stained with Coomassie blue. Molecular weight markers are shown on the left and right. Spots 1-7 were identified by MS/MS: 1 & 2 alcohol dehydrogenase class III; 3 – universal
stress protein; 4 & 5 - D-arabinitol dehydrogenase and Glyceraldehyde-3-Phosphate dehydrogenase 1; 6 & 7 – sorbitol dehydrogenase-like protein (Table 8).

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
44
Table 8 - Proteins from ion exchange chromatography fraction with Mono Q 4.6/100 PE column (Figure 11) analysed by mass spectrometry (MS/MS) and identified with a Patella vulgata database.
Mascot search result scores greater than 65 are significant (p<0.05). Data shows the highest score identifications for each sample.
Mascot Search NCBI Blastp
Band Accession MW
(kDa) Score
No. Peptides Identified
No. Amino Acids
Specific hits Organism Score Identity
(%)
1
CL24363Contig1 48.891
169 35
390
Hypothetical protein LOTGIDRAFT_228995 Lottia gigantea 690 89
Alcohol dehydrogenase class III
Crassostrea gigas 636 81
2 207 41 Aplysia californica 624 80
Biomphalaria glabrata 622 80
3 CL27250Contig1 44.983 88 19 180
Hypothetical protein LOTGIDRAFT_203658 Lottia gigantea 277 81
Universal stress protein Biomphalaria glabrata 230 70
Octopus bimaculoides 221 60
4
CL9544Contig1 44.491
176 29
399
Hypothetical protein LOTGIDRAFT_188066
Lottia gigantea
452 60
Hypothetical protein LOTGIDRAFT_159773 450 58
5 153 29
Hypothetical protein LOTGIDRAFT_122580 362 52
D-arabinitol dehydrogenase 1 Neodiprion lecontei 308 45
Ciona intestinalis 303 44
6
CL32538Contig1 50.098
110 17
452
Hypothetical protein LOTGIDRAFT_208637 Lottia gigantea 635 87
Sorbitol dehydrogenase-like
Crassostrea gigas 534 73
7 88 14 Aplysia californica 530 76
Biomphalaria glabrata 517 74
Table 9 - Peptide ions from ion exchange chromatography fraction with Mono Q 4.6/100 PE column (Figure 11) analysed by mass spectrometry (MS/MS) and identified with UniProt (Metazoa) database.
Mascot search result scores greater than 82 are significant (p<0.05); Data shows the highest score identifications for each sample.
Mascot Search
Band Accession MW (kDa) Score Description
4 C7SP37 20.658 228 Glyceraldehyde-3-Phosphate dehydrogenase (Fragment)
5 G9BBW6 16.599 102 Glyceraldehyde-3-Phosphate dehydrogenase (Fragment)

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
45
4. Discussion
The capacity to metabolize polyols seems to be an important factor that allows
organisms to deal with stress-inducing conditions through osmoregulation [32, 118], but
also to use them as carbon and energy sources after food intake [24]. Our results using
total homogenates of different gastropod species, showed that almost every species
present dehydrogenase activity for both mannitol and sorbitol, suggesting that this
metabolic capacity is generalized throughout all the gastropod clades. In contrast, the
oxidase activity towards polyols was only detected in herbivorous species of
Caenogastropoda and Heterobranchia clades (data not published).
When correlating polyol dehydrogenase with their diet, it was observed that all
herbivores species possess this activity at least for one of the substrates. On the other
hand, half of the analysed carnivorous species did not show polyol dehydrogenase
activity, and those who did presented dehydrogenase activities to these substrates, were
quite lower levels than the observed in herbivores. These data suggest that the
herbivorous species, due their higher polyol intake with the vegetal matter of the diet,
would have an extra need to metabolize these compounds and therefore higher activity
levels. However, this capacity does not seem to be exclusive for herbivores, since some
carnivores also have it. It is unclear whether this is a conserved feature in these animals,
or an adaptation for those whose diet includes polyols and, therefore, use these
compounds for energy intake and survival.
Following the principle that the metabolic activities detected in gastropods
digestive gland would be tightly correlated with the compounds mainly ingested in the
diet [24], the metabolic capacity of the digestive gland for cinnamyl alcohol was also
studied. This compound is representative of the aromatic alcohols [33, 34]. Since it was
already described its correlation with lignin degradation and its oxidative process being,
in part, similar between fungi and gastropods (physico-chemical parameters) [34],
significant activities for this substrate were expected at least in herbivorous species that
might ingest some lignified plant matter.
Interestingly, a previous study reported cinnamyl alcohol oxidase activity in all the
studied species, herbivores and carnivores, except in O. celtica (data not published). In
this work, the dehydrogenase activity for cinnamyl alcohol was detected in all the 26-
studied species, and for the first time in animals. These results suggest that cinnamyl
alcohol metabolism in gastropod digestive gland in generalized among all the studied
species, without any significant correlation with taxonomy and dietary habits. However,

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
46
a slight correlation could be established regarding the species habitat, since the studied
freshwater species presented lower cinnamyl alcohol dehydrogenase activity values in
an overall comparison. Therefore, due to the presence of aromatic alcohol metabolism
in carnivores but also in herbivores that do not feed mainly on lignified plants, the
correlation of this metabolism and the consumption of lignified plant matter does not
seem valid anymore. Another hypothesis for this generalized metabolic capacity through
all the analysed species could be some correlation with detoxification processes.
At this point, data suggest that the polyols metabolism is much more correlated
with dietary habits, and perhaps the taxonomy, in the studied species of gastropods,
while aromatic alcohols metabolisms seems to be a much more generalized feature of
these animals.
There is an ongoing discussion about whereas the natural exposure to ethanol is
responsible for the diversity of oxidoreductases [38]. Studies with cephalopods
dehydrogenases showed that only class III ADH is present in these animals [110]. This
data does not support the hypothesis of ethanol metabolism, typically translated by the
presence of class I ADH, in molluscs. In previous studies, the oxidase activity for ethanol
was evaluated in several gastropod species, where only five of the 26-studied species
presented detectable activity for this compound (data not published). Additionally, in the
present study, ethanol dehydrogenase activity was only detected in two herbivorous
eupulmunates. In both studies, the ethanol-related activities detected presented low
overall values when compared with activity values of other substrates. Therefore, this
data suggests that gastropods digestive gland does not play a major role in ethanol
metabolism (via oxidase or dehydrogenase). It is uncertain if other pathways of ethanol
metabolism exist in molluscs, in the digestive gland or other organs.
Several studies have previously shown that alcohol dehydrogenases [119] and
polyol dehydrogenases [91, 105] are capable of metabolizing, not only the main
substrate responsible for its nomination, but also several other similar substrates. In fact,
they are generally seen as quite unspecific enzymes [38]. In some cases, even higher
activities are found for other substrates [105]. Our results also suggest this unspecific
behaviour of these enzymes. This is supported by the results of in situ activity detection,
where each analysed species presented similar dehydrogenase activity profiles towards
various substrates (polyols and aromatic alcohol). Interestingly, with exception of P.
vulgata, all the other species may have more than one enzyme with dehydrogenase
activity for these substrates. However, it cannot be ruled out the hypothesis that the
second band observed in the analysis of these enzymes corresponds to the same protein
post-translationally modified. Additionally, the well-defined band observed in these gels,

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
47
might be a complex mixture of proteins in which only some of them possess
dehydrogenase activity for these substrates.
The comparison of the relative migration distance for both polyols and aromatic
alcohol dehydrogenases after in situ activity detection suggests that proteins with high
polyols dehydrogenase activity are different from those with high aromatic alcohol
dehydrogenase activity. However, being already known the unspecific behaviour of the
oxidoreductase enzyme family [38, 41, 109], a feature already suggested in this study, it
is not possible to exclude the hypothesis that polyol and aromatic alcohol metabolism
enzymes (oxidases and dehydrogenases) might non-specifically use the substrates of
each other, but with lower efficiency.
Aiming to isolate and identify the proteins with mannitol or sorbitol dehydrogenase
activity, we used cytosolic fractions from Patella vulgata since this species exhibited high
activity values, it is easier to obtain high amounts of tissue due to a big proportion of
digestive gland present in this animal, and this species can be easily collected all year.
The separation of proteins by affinity chromatography allowed the obtaining of
fractions significantly enriched (around ten-fold) in polyols or cinnamyl alcohol
dehydrogenases. Regarding the protein behaviour in this column, this first
chromatography also supported the hypothesis of the significant biochemical
differences, between the proteins responsible for polyols and aromatic alcohol
dehydrogenase activities. Proteins with polyol dehydrogenase activity were eluted from
the column in the initial washing steps, but mainly in the lower NAD+ concentration steps,
and proteins with aromatic alcohol dehydrogenase activity were eluted in the higher
NAD+ concentration steps and in the final washing steps. This protein behaviour
suggests that proteins with polyols dehydrogenase activity present much less affinity for
this column than proteins with aromatic alcohol dehydrogenase activity. In this work, we
focused our attention in the fraction enriched for polyols dehydrogenase activity. Future
studies will be performed to identify cinnamyl alcohol dehydrogenases.
Overall the results show that the Blue Sepharose CL-6B column was quite effective
in the separation of dehydrogenase enzymes, since the characterization of the fractions
enriched in dehydrogenases led to the identification of two dehydrogenases (cytosolic
malate dehydrogenase and GAPDH), one class III alcohol dehydrogenase, and two
polyol dehydrogenases (D-arabinitol dehydrogenase 1 and sorbitol dehydrogenase).
The analysis of dehydrogenase activity in the fractions obtained by ion exchange
chromatography revealed 2 peaks, a first one early eluted from the column that had
mannitol dehydrogenase activity higher than sorbitol dehydrogenase activity, and a
second peak where the sorbitol dehydrogenase activity was higher than mannitol
dehydrogenase activity. These results suggest that different proteins are associated with

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
48
these dehydrogenases (Figure 11). Consistent with the fact that these type of enzymes
can be quite unspecific [91], they can metabolize both D-mannitol and D-sorbitol, it was
identified a D-arabinitol dehydrogenase (ARDH) in the first peak, with higher
dehydrogenase activity for mannitol (band 3 of Figure 9 and band 4 and 5 of Figure 11),
and a sorbitol dehydrogenase-like protein, with higher dehydrogenase activity for sorbitol
(band 5 of Figure 9 and bands 6 and 7 of Figure 11).
D- and L-arabinitol dehydrogenases are enzymes mostly studied in fungi [120,
121]. Interestingly, despite having an apparent classification as a polyol dehydrogenase,
there are some evidences that this enzyme has functions similar to that of medium-chain
dehydrogenases [122] or even short- and long-chain dehydrogenases [123, 124]. Thus,
our Blastp results identified the founded protein as from MDR superfamily and higher
scores were obtained for three uncharacterized sequences of Lottia gigantea, the insect
Neodiprion lecontei and cordate Ciona intestinalis. This data reinforced the idea that little
is known about arabinitol dehydrogenases in other organisms than fungi. However, this
might be one of the first evidences of this enzyme in molluscs.
Regarding the overall data of this work, especially the results related with polyols
dehydrogenases, were obtained the first clues on which enzyme may be responsible for
each dehydrogenase activity. However, more work must be done for the purification of
these enzymes, in order to characterize the enzymes with polyols dehydrogenase
activity. Additionally, more work must be focused in the proteins with aromatic
dehydrogenase activity.

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
49
5. Conclusions and perspectives
With this study, it was revealed for the first time the presence of dehydrogenase of
mannitol and aromatic alcohols in the digestive gland of gastropods. It was reinforced
the biochemical differences between proteins of polyol and aromatic alcohols
metabolisms. Also, this study supports the absence of enzymes with significant ethanol
dehydrogenase activity in molluscs, having the gastropods digestive gland by example.
For a better idea of which specific ADH classes are present in these organisms,
biochemical assays, regarding that ADH class specificity, could be done in the future
studies.
The first steps for isolation of aromatic alcohol and polyalcohol dehydrogenases in
the marine gastropod Patella vulgata, resulted in the identification of at least two
enzymes with polyol dehydrogenase activity (sorbitol and arabinitol dehydrogenases). A
similar line of research regarding the proteins with aromatic alcohols dehydrogenase
activity, namely cinnamyl alcohol, is yet to be developed in future studies. More studies
in the future could be focused in arabinitol dehydrogenase because of the lack of
knowledge about this enzyme in organisms other than fungi.
Thus, with these new knowledge’s, there is a changing of perspectives about the
metabolisms of gastropod`s digestive gland, but also a comparison point for this type of
enzymes with other organisms.
Finally, future work aims to identify the genes codifying these proteins, for their
subsequent heterologous production and to study their metabolic profiles recurring to
other alcoholic substrates.

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
50
6. References
1. Thiele, J., Handbuch der systematischen Weichtierkunde. Vol. 1. 1931, G. Fischer: Jena.
2. Zapata, F., Wilson, N.G., Howison, M., Andrade, S., Jörger, K., Schrödl, M., Goetz, F., Giribet, G., and Dunn, C., Phylogenomic analyses of deep gastropod relationships reject Orthogastropoda. Proc R Soc Lond B Biol Sci, 2014. 281(1794): 20141739.
3. Aktipis, S.W. and Giribet, G., Testing relationships among the vetigastropod taxa: a molecular approach. J Molluscan Stud, 2012. 78(1): 12-27.
4. Wägele, H., Klussmann-Kolb, A., Verbeek, E., and Schrödl, M., Flashback and foreshadowing—a review of the taxon Opisthobranchia. Org Divers Evol, 2014. 14(1): 133-149.
5. Nakano, T. and Sasaki, T., Recent advances in molecular phylogeny, systematics and evolution of patellogastropod limpets. J Molluscan Stud, 2011. 77(3): 203-217.
6. Evans, M.R. and Williams, G.A., Time partitioning of foraging in the limpet Patella vulgata. J Anim Ecol, 1991: 563-575.
7. Uribe, J.E., Colgan, D., Castro, L.R., Kano, Y., and Zardoya, R., Phylogenetic relationships among superfamilies of Neritimorpha (Mollusca: Gastropoda). Mol Phylogenet Evol, 2016. 104: 21-31.
8. Lindberg, D.R., Patellogastropoda, Neritimorpha and Cocculinoidea. The low-diversity gastropod clades. In Phylogeny and Evolution of the Mollusca, Ponder, W.F. and Lindberg, D.R., Editors. 2008, University of California Press: Berkeley, CA. 271-296.
9. Hickman, C.S., Archaeogastropod evolution, phylogeny and systematics: a re-evaluation. Malacol Rev Suppl, 1988. 4: 17-34.
10. Colgan, D., Ponder, W., Beacham, E., and Macaranas, J., Molecular phylogenetics of Caenogastropoda (Gastropoda: Mollusca). Mol Phylogenet Evol, 2007. 42(3): 717-737.
11. Dinapoli, A. and Klussmann-Kolb, A., The long way to diversity–phylogeny and evolution of the Heterobranchia (Mollusca: Gastropoda). Mol Phylogenet Evol, 2010. 55(1): 60-76.
12. Lobo-da-Cunha, A., Ultrastructural and cytochemical aspects of the basophilic cells in the hepatopancreas of Aplysia depilans (Mollusca, Opisthobranchia). Tissue Cell, 1999. 31(1): 8-16.
13. Lobo-da-Cunha, A., The digestive cells of the hepatopancreas in Aplysia depilans (Mollusca, Opisthobranchia): ultrastructural and cytochemical study. Tissue Cell, 2000. 32(1): 49-57.
14. Luchtel, D.L., Martin, A.W., Deyrup-Olsen, I., and Boer, H., Gastropoda: pulmonata. In Microscopic Anatomy of Invertebrates, Harrison, F.W. and Kohn, A.J., Editors. 1997, Wiley-Liss: New York. 459-718.
15. Dimitriadis, V. and Andrews, E., Ultrastructural and cytochemical study of the digestive gland cells of the marine prosobranch mollusc Nucella lapillus (L.) in relation to function. Malacologia, 2000. 42(1-2): 103-112.
16. Franchini, A. and Ottaviani, E., Histochemical and ultrastructural study of the digestive gland of the freshwater snail Panorbarius corneus (L.)(Gastropoda, Pulmonata). Anim Biol, 1993. 2: 191-198.
17. Lobo-da-Cunha, A., Oliveira, E., Alves, Â., Calado, G., and Guimarães, F., Ultrastructural Detection of Calcium with the Pyroantimonate Method in Cells of Marine Gastropods. Microsc Microanal, 2015. 21(S5): 67-68.
18. Lobo-da-Cunha, A., Batista, C., and Oliveira, E., The peroxisomes of the hepatopancreas in marine gastropods. Biol Cell, 1994. 82(1): 67-74.

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
51
19. Cajaraville, M.P., Cancio, I., Ibabe, A., and Orbea, A., Peroxisome proliferation as a biomarker in environmental pollution assessment. Microsc Res Tech, 2003. 61(2): 191-202.
20. David, H. and Götze, J., Elektronenmikroskopische Befunde an der Mitteldarmdrüse von Schnecken. Z mikr-anat Forsch, 1963. 70: 252-272.
21. Moya, J. and Rallo, A.M., Intracisternal polycylinders: a cytoplasmic structure in cells of the terrestrial slug Arion empiricorum Férussac (Pulmonata, Stylommatophora). Cell Tissue Res, 1975. 159(3): 423-433.
22. McLean, N., Unusual aggregations of tubules associated with endoplasmic reticulum in digestive cells of Alderia modesta (Mollusca: Gastropoda: Sacoglossa). Cell Tissue Res, 1978. 194(1): 179-182.
23. Vorhaben, J.E., Scott, J.F., and Campbell, J.W., D-Mannitol oxidation in the land snail, Helix aspersa. J Biol Chem, 1980. 255(5): 1950-1955.
24. Vorhaben, J.E., Smith, D.D., and Campbell, J.W., Histochemical and biochemical localization of mannitol oxidase in the snail Helix aspersa. J Exp Zool, 1984. 231(1): 157-160.
25. Vorhaben, J.E., Smith, D.D., and Campbell, J.W., Mannitol oxidase: partial purification and characterization of the membrane-bound enzyme from the snail Helix aspersa. Int J Biochem, 1986. 18(4): 337-344.
26. Large, A., Jones, C.J., and Connock, M., The association of mannitol oxidase with a distinct organelle in the digestive gland of the terrestrial slug Arion ater. Protoplasma, 1993. 175(3-4): 93-101.
27. Grewal, N., Parveen, Z., Large, A., Perry, C., and Connock, M., Gastropod mollusc aliphatic alcohol oxidase: subcellular localisation and properties. Comp Biochem Physiol B: Biochem Mol Biol, 2000. 125(4): 543-554.
28. Large, A.T. and Connock, M.J., Centrifugal evidence for association of mannitol oxidase with distinct organelles (“mannosomes”) in the digestive gland of several species of terrestrial gastropod mollusc. Comp Biochem Physiol A Physiol, 1994. 107(4): 621-629.
29. Stoop, J.M., Williamson, J.D., and Pharr, D.M., Mannitol metabolism in plants: a method for coping with stress. Trends Plant Sci, 1996. 1(5): 139-144.
30. Iwamoto, K. and Shiraiwa, Y., Salt-regulated mannitol metabolism in algae. Mar Biotechnol, 2005. 7(5): 407-415.
31. Lobo-da-Cunha, A., Amaral-de-Carvalho, D., Oliveira, E., Alves, Â., and Calado, G., Histochemical Detection of Mannitol Oxidase in the Digestive Gland of Gastropods. Microsc Microanal, 2016. 22: 14-15.
32. Tsvetkov, I. and Konichev, A., Stress-induced changes in accumulation of sorbitol and in activities of concomitant enzymes in digestive gland of freshwater snail. Biochemistry (Mosc), 2009. 74(11): 1260-1265.
33. Large, A.T. and Connock, M.J., Aromatic alcohol oxidase and aldehyde oxidase activities in the digestive gland of three species of terrestrial gastropod (Helix aspersa, Arion ater, Limax flavus). Comp Biochem Physiol A Physiol, 1993. 104(3): 489-491.
34. Mann, V., Large, A., Khan, S., Malik, Z., and Connock, M., Aromatic alcohol oxidase: A new membrane‐bound H2O2‐generating enzyme in alimentary tissues of the slug Arion ater. J Exp Zool, 1989. 251(3): 265-274.
35. Farmer, V., Henderson, M.E., and Russell, J., Aromatic-alcohol-oxidase activity in the growth medium of Polystictus versicolor. Biochem J, 1960. 74(2): 257-262.
36. Guillén, F., Martínez, A., Martínez, M., and Evans, C., Hydrogen-peroxide-producing system of Pleurotus eryngii involving the extracellular enzyme aryl-alcohol oxidase. Appl Microbiol Biotechnol, 1994. 41(4): 465-470.
37. Guillén, F., Martínez, A.T., and Martínez, M.J., Substrate specificity and properties of the aryl‐alcohol oxidase from the ligninolytic fungus Pleurotus eryngii. Eur J Biochem, 1992. 209(2): 603-611.

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
52
38. Hernández-Tobías, A., Julián-Sánchez, A., Pina, E., and Riveros-Rosas, H., Natural alcohol exposure: Is ethanol the main substrate for alcohol dehydrogenases in animals? Chem-Biol Interact, 2011. 191(1): 14-25.
39. Lieber, C., Gentry, R., and Baraona, E., First pass metabolism of ethanol. Alcohol Alcohol Suppl, 1993. 2: 163-169.
40. Lieber, C.S., Alcohol, liver, and nutrition. J Am Coll Nutr, 1991. 10(6): 602-632. 41. Jörnvall, H., Höög, J.-O., and Persson, B., SDR and MDR: completed genome
sequences show these protein families to be large, of old origin, and of complex nature. FEBS Lett, 1999. 445(2-3): 261-264.
42. Kavanagh, K., Jörnvall, H., Persson, B., and Oppermann, U., The SDR superfamily: functional and structural diversity within a family of metabolic and regulatory enzymes. Cell Mol Life Sci, 2008. 65: 3895-3906.
43. Riveros‐Rosas, H., Julián‐Sánchez, A., Villalobos‐Molina, R., Pardo, J.P., and Piña, E., Diversity, taxonomy and evolution of medium‐chain dehydrogenase/reductase superfamily. Eur J Biochem, 2003. 270(16): 3309-3334.
44. Jörnvall, H., Hedlund, J., Bergman, T., Oppermann, U., and Persson, B., Superfamilies SDR and MDR: from early ancestry to present forms. Emergence of three lines, a Zn-metalloenzyme, and distinct variabilities. Biochem Biophys Res Commun, 2010. 396(1): 125-130.
45. Reid, M.F. and Fewson, C.A., Molecular characterization of microbial alcohol dehydrogenases. Crit Rev Microbiol, 1994. 20(1): 13-56.
46. Persson, B., Krook, M., and Jörnvall, H., Characteristics of short‐chain alcohol dehydrogenases and related enzymes. Eur J Biochem, 1991. 200(2): 537-543.
47. Jörnvall, H., Nordling, E., and Persson, B., Multiplicity of eukaryotic ADH and other MDR forms. Chem-Biol Interact, 2003. 143: 255-261.
48. Lieber, C.S., The discovery of the microsomal ethanol oxidizing system and its physiologic and pathologic role. Drug Metab Rev, 2004. 36(3-4): 511-529.
49. Kumar, S. and Hedges, S.B., A molecular timescale for vertebrate evolution. Nature, 1998. 392(6679): 917-920.
50. Gonzalez-Duarte, R. and Albalat, R., Merging protein, gene and genomic data: the evolution of the MDR-ADH family. Heredity, 2005. 95(3): 184-197.
51. Höög, J.-O. and Östberg, L.J., Mammalian alcohol dehydrogenases–a comparative investigation at gene and protein levels. Chem-Biol Interact, 2011. 191(1): 2-7.
52. Lieber, C.S., Metabolism of alcohol. Clin Liver Dis, 2005. 9(1): 1-35. 53. Anderson, P. and Baumberg, B., Alcohol in Europe. 2006, London: Institute of
Alcohol Studies. 54. Singer, M., Toward a political-economy of alcoholism: the missing link in the
anthropology of drinking. Soc Sci Med, 1986. 23(2): 113-130. 55. Steinmetz, C.G., Xie, P., Weiner, H., and Hurley, T.D., Structure of mitochondrial
aldehyde dehydrogenase: the genetic component of ethanol aversion. Structure, 1997. 5(5): 701-711.
56. Svensson, S., Strömberg, P., and Höög, J.-O., A novel subtype of class II alcohol dehydrogenase in rodents unique Pro47 and Ser182 modulates hydride transfer in the mouse enzyme. J Biol Chem, 1999. 274(42): 29712-29719.
57. Hellgren, M., Strömberg, P., Gallego, O., Martras, S., Farrés, J., Persson, B., Parés, X., and Höög, J.-O., Alcohol dehydrogenase 2 is a major hepatic enzyme for human retinol metabolism. Cell Mol Life Sci, 2007. 64(4): 498-505.
58. Staab, C., Hellgren, M., and Höög, J.-O., Medium-and short-chain dehydrogenase/reductase gene and protein families. Cell Mol Life Sci, 2008. 65(24): 3950-3960.
59. Jensen, D.E., Belka, G.K., and Du Bois, G.C., S-Nitrosoglutathione is a substrate for rat alcohol dehydrogenase class III isoenzyme. Biochem J, 1998. 331(2): 659-668.

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
53
60. Estonius, M., Danielsson, O., Karlsson, C., Persson, H., Jörnvall, H., and Höög, J.O., Distribution of alcohol and sorbitol dehydrogenases. Eur J Biochem, 1993. 215(2): 497-503.
61. Estonius, M., Svensson, S., and Höög, J.-O., Alcohol dehydrogenase in human tissues: localisation of transcripts coding for five classes of the enzyme. FEBS Lett, 1996. 397(2-3): 338-342.
62. Halsted, C.H., Robles, E.A., and Mezey, E., Distribution of ethanol in the human gastrointestinal tract. Am J Clin Nutr, 1973. 26(8): 831-834.
63. Lee, S.-L., Wang, M.-F., Lee, A.-I., and Yin, S.-J., The metabolic role of human ADH3 functioning as ethanol dehydrogenase. FEBS Lett, 2003. 544(1-3): 143-147.
64. Hedberg, J.J., Höög, J.-O., Nilsson, J.A., Xi, Z., Elfwing, Å., and Grafström, R.C., Expression of alcohol dehydrogenase 3 in tissue and cultured cells from human oral mucosa. Am J Pathol, 2000. 157(5): 1745-1755.
65. Pares, X., Cederlund, E., Moreno, A., Hjelmqvist, L., Farres, J., and Jörnvall, H., Mammalian class IV alcohol dehydrogenase (stomach alcohol dehydrogenase): structure, origin, and correlation with enzymology. Proc Natl Acad Sci USA, 1994. 91(5): 1893-1897.
66. Chou, C.-F., Lai, C.-L., Chang, Y.-C., Duester, G., and Yin, S.-J., Kinetic mechanism of human class IV alcohol dehydrogenase functioning as retinol dehydrogenase. J Biol Chem, 2002. 277(28): 25209-25216.
67. Matsumoto, M., Yokoyama, H., Suzuki, H., Shiraishi-Yokoyama, H., and Hibi, T., Retinoic acid formation from retinol in the human gastric mucosa: role of class IV alcohol dehydrogenase and its relevance to morphological changes. Am J Physiol Gastrointest Liver Physiol, 2005. 289(3): 429-433.
68. Höög, J.-O., Hedberg, J.J., Strömberg, P., and Svensson, S., Mammalian alcohol dehydrogenase–functional and structural implications. J Biomed Sci, 2001. 8(1): 71-76.
69. Höög, J.-O., Strömberg, P., Hedberg, J.J., and Griffiths, W.J., The mammalian alcohol dehydrogenases interact in several metabolic pathways. Chem-Biol Interact, 2003. 143: 175-181.
70. Strömberg, P. and Höög, J.-O., Human class V alcohol dehydrogenase (ADH5): A complex transcription unit generates C-terminal multiplicity. Biochem Biophys Res Commun, 2000. 278(3): 544-549.
71. Thatcher, D.R., The complete amino acid sequence of three alcohol dehydrogenase alleloenzymes (AdhN-11, AdhS and AdhUF) from the fruitfly Drosophila melanogaster. Biochem J, 1980. 187(3): 875-883.
72. Jörnvall, H., Horse liver alcohol dehydrogenase. Eur J Biochem, 1970. 16(1): 25-40.
73. Jörnvall, H., The primary structure of yeast alcohol dehydrogenase. Eur J Biochem, 1977. 72(3): 425-442.
74. Jörnvall, H., Persson, M., and Jeffery, J., Alcohol and polyol dehydrogenases are both divided into two protein types, and structural properties cross-relate the different enzyme activities within each type. Proc Natl Acad Sci USA, 1981. 78(7): 4226-4230.
75. Danielsson, O., Atrian, S., Luque, T., Hjelmqvist, L., Gonzalez-Duarte, R., and Jörnvall, H., Fundamental molecular differences between alcohol dehydrogenase classes. Proc Natl Acad Sci USA, 1994. 91(11): 4980-4984.
76. Geer, B.W., Langevin, M.L., and McKechnie, S.W., Dietary ethanol and lipid synthesis in Drosophila melanogaster. Biochem Genet, 1985. 23(7-8): 607-622.
77. Benach, J., Atrian, S.l., Gonzàlez-Duarte, R., and Ladenstein, R., The catalytic reaction and inhibition mechanism of Drosophila alcohol dehydrogenase: observation of an enzyme-bound NAD-ketone adduct at 1.4 Å resolution by X-ray crystallography. J Mol Biol, 1999. 289(2): 335-355.

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
54
78. Nascimento, J.C. and Oliveira, A.K., Ontogenetic development of Anastrepha fraterculus (Diptera: Tephritidae): isoenzyme patterns of isocitrate and alcohol dehydrogenases. Comp Biochem Physiol A Comp Physiol, 1997. 118(3): 847-854.
79. Gasperi, G., Baruffi, L., Malacrida, A., and Robinson, A., A biochemical genetic study of alcohol dehydrogenase isozymes of the medfly, Ceratitis capitata Wied. Biochem Genet, 1992. 30(5-6): 289-304.
80. Magallón, S., Using fossils to break long branches in molecular dating: a comparison of relaxed clocks applied to the origin of angiosperms. Syst Biol, 2010. 59(4): 384-399.
81. Conway, T. and Ingram, L., Similarity of Escherichia coli propanediol oxidoreductase (fucO product) and an unusual alcohol dehydrogenase from Zymomonas mobilis and Saccharomyces cerevisiae. J Bacteriol, 1989. 171(7): 3754-3759.
82. Truniger, V. and Boos, W., Mapping and cloning of gldA, the structural gene of the Escherichia coli glycerol dehydrogenase. J Bacteriol, 1994. 176(6): 1796-1800.
83. Walter, K.A., Bennett, G., and Papoutsakis, E.T., Molecular characterization of two Clostridium acetobutylicum ATCC 824 butanol dehydrogenase isozyme genes. J Bacteriol, 1992. 174(22): 7149-7158.
84. Stojiljkovic, I., Bäumler, A.J., and Heffron, F., Ethanolamine utilization in Salmonella typhimurium: nucleotide sequence, protein expression, and mutational analysis of the cchA cchB eutE eutJ eutG eutH gene cluster. J Bacteriol, 1995. 177(5): 1357-1366.
85. Yuanyuan, Z., Yang, C., and Baishan, F., Cloning and sequence analysis of the dhaT gene of the 1, 3-propanediol regulon from Klebsiella pneumoniae. Biotechnol Lett, 2004. 26(3): 251-255.
86. Lyon, R.C., Johnston, S.M., Panopoulos, A., Alzeer, S., McGarvie, G., and Ellis, E.M., Enzymes involved in the metabolism of -hydroxybutyrate in SH-SY5Y cells: identification of an iron-dependent alcohol dehydrogenase ADHFe1. Chem-Biol Interact, 2009. 178(1): 283-287.
87. Deng, Y., Wang, Z., Gu, S., Ji, C., Ying, K., Xie, Y., and Mao, Y., Cloning and characterization of a novel human alcohol dehydrogenase gene (ADHFe1). DNA Sequence, 2002. 13(5): 301-306.
88. Van Cauter, E., Plat, L., Scharf, M.B., Leproult, R., Cespedes, S., L'Hermite-Baleriaux, M., and Copinschi, G., Simultaneous stimulation of slow-wave sleep and growth hormone secretion by gamma-hydroxybutyrate in normal young Men. J Clin Invest, 1997. 100(3): 745-753.
89. Koch, J.J., Performance-enhancing: substances and their use among adolescent athletes. Pediatr Rev, 2002. 23(9): 310-317.
90. Gu, J., Orr, N., Park, S.D., Katz, L.M., Sulimova, G., MacHugh, D.E., and Hill, E.W., A genome scan for positive selection in thoroughbred horses. PLoS One, 2009. 4(6): e5767.
91. Lindstad, I.R., Peter, K., and McKinley-McKee, J.S., Substrate specificity of sheep liver sorbitol dehydrogenase. Biochem J, 1998. 330(1): 479-487.
92. Driedzic, W.R. and Ewart, K.V., Control of glycerol production by rainbow smelt (Osmerus mordax) to provide freeze resistance and allow foraging at low winter temperatures. Comp Biochem Physiol B: Biochem Mol Biol, 2004. 139(3): 347-357.
93. Alves-Araújo, C., Pacheco, A., Almeida, M., Spencer-Martins, I., Leão, C., and Sousa, M., Sugar utilization patterns and respiro-fermentative metabolism in the baker's yeast Torulaspora delbrueckii. Microbiology, 2007. 153(3): 898-904.
94. Hers, H., The mechanism of the formation of seminal fructose and fetal fructose. Biochim Biophys Acta, 1960. 37: 127-138.

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
55
95. Jeffery, J. and Jörnvall, H., Enzyme relationships in a sorbitol pathway that bypasses glycolysis and pentose phosphates in glucose metabolism. Proc Natl Acad Sci USA, 1983. 80(4): 901-905.
96. Gabbay, K.H., The sorbitol pathway and the complications of diabetes. New Engl J Med, 1973. 288(16): 831-836.
97. Lindstad, R.I. and McKinley-McKee, J.S., Reversible inhibition of sheep liver sorbitol dehydrogenase by the antidiabetogenic drug 2‐hydroxymethyl‐4‐(4‐N, N‐dimethylaminosulfonyl‐1‐piperazino) pyrimidine. FEBS Lett, 1997. 408(1): 57-61.
98. El-Kabbani, O., Darmanin, C., and Chung, R.-T., Sorbitol dehydrogenase: structure, function and ligand design. Curr Med Chem, 2004. 11(4): 465-476.
99. Barretto, O.C. and Nonoyama, K., Biochemical characteristics of erythrocyte sorbitol dehydrogenase from selected mammals. Comp Biochem Physiol B, 1984. 77(2): 387-389.
100. Jedziniak, J.A., Chylack, L., Cheng, H.-M., Gillis, M.K., Kalustian, A.A., and Tung, W.H., The sorbitol pathway in the human lens: aldose reductase and polyol dehydrogenase. Invest Ophthalmol Visual Sci, 1981. 20(3): 314-326.
101. O'Brien, M.M., Schofield, P.J., and Edwards, M.R., Polyol-pathway enzymes of human brain. Partial purification and properties of sorbitol dehydrogenase. Biochem J, 1983. 211(1): 81-90.
102. Lindstad, R.I., Hermansen, L.F., and McKinley‐McKee, J.S., The kinetic mechanism of sheep liver sorbitol dehydrogenase. Eur J Biochem, 1992. 210(2): 641-647.
103. Nishikawa, T., Edelstein, D., Du, X.L., Yamagishi, S.-i., Matsumura, T., Kaneda, Y., Yorek, M.A., Beebe, D., Oates, P.J., and Hammes, H.-P., Normalizing mitochondrial superoxide production blocks three pathways of hyperglycaemic damage. Nature, 2000. 404(6779): 787-790.
104. Lindstad, R.I., Hermansen, L.F., and McKINLEY‐McKEE, J.S., Inhibition and activation studies on sheep liver sorbitol dehydrogenase. Eur J Biochem, 1994. 221(2): 847-854.
105. Maret, W. and Auld, D.S., Purification and characterization of human liver sorbitol dehydrogenase. Biochemistry, 1988. 27(5): 1622-1628.
106. Leissing, N. and McGuinness, E., Sorbitol dehydrogenase from rat liver. Methods Enzymol, 1982. 89: 136-140.
107. Wiesinger, H. and Hamprecht, B., Purification and characterization of sorbitol dehydrogenase from bovine brain. J Neurochem, 1989. 52(2): 342-348.
108. Karlsson, C., Jörnvall, H., and Höög, J.O., Sorbitol dehydrogenase: cDNA coding for the rat enzyme. Eur J Biochem, 1991. 198(3): 761-765.
109. Lindstad, R.I. and Mckinley‐Mckee, J.S., Reversible inhibition of sheep liver sorbitol dehydrogenase by thiol compounds. Eur J Biochem, 1996. 241(1): 142-148.
110. Fernández, M.R., Jörnvall, H., Moreno, A., Kaiser, R., and Parés, X., Cephalopod alcohol dehydrogenase: purification and enzymatic characterization. FEBS Lett, 1993. 328(3): 235-238.
111. Narang, S. and Narang, N., Alcohol dehydrogenase of Biomphalaria glabrata (Mollusca: Pulmonata): Polymorphism, genetic analysis, and interspecific variations. Biochem Genet, 1976. 14(1-2): 75-86.
112. Coelho, I., Lima, D., André, A., Melo, C., Ruivo, R., Reis-Henriques, M.A., Santos, M.M., and Castro, L.F.C., Molecular characterization of Adh3 from the mollusc Nucella lapillus: tissue gene expression after tributyltin and retinol exposure. J Molluscan Stud, 2012: 1-6.
113. Moyes, C.D., Suarez, R.K., Brown, G.S., and Hochachka, P.W., Peroxisomal ‐oxidation: Insights from comparative biochemistry. J Exp Zool, 1991. 260(2): 267-273.

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
56
114. Stewart, J.M., Carlin, R., MacDonald, J., and Van Iderstine, S., Fatty acid binding proteins and fatty acid catabolism in marine invertebrates: peroxisomal -oxidation. Invertebr Reprod Dev, 1994. 25(1): 73-82.
115. Bradford, M.M., A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem, 1976. 72(1-2): 248-254.
116. Lewis, N. and Gibson, J., Variation in amount of enzyme protein in natural populations. Biochem Genet, 1978. 16(3-4): 159-170.
117. Gomes, C., Almeida, A., Ferreira, J.A., Silva, L., Santos-Sousa, H., Pinto-de-Sousa, J.o., Santos, L.c.L., Amado, F., Schwientek, T., and Levery, S.B., Glycoproteomic analysis of serum from patients with gastric precancerous lesions. J Proteome Res, 2013. 12(3): 1454-1466.
118. Knigge, T., Mann, N., Parveen, Z., Perry, C., Gernhöfer, M., Triebskorn, R., Köhler, H., and Connock, M., Mannosomes: a molluscan intracellular tubular membrane system related to heavy metal stress? Comp Biochem Physiol, C: Toxicol Pharmacol, 2002. 131(3): 259-269.
119. Arslanian, M.J., Pascoe, E., and Reinhold, J.G., Rat liver alcohol dehydrogenase. Purification and properties. Biochem J, 1971. 125(4): 1039-1047.
120. Bae, B., Sullivan, R.P., Zhao, H., and Nair, S.K., Structure and engineering of L-arabinitol 4-dehydrogenase from Neurospora crassa. J Mol Biol, 2010. 402(1): 230-240.
121. Murray, J.S., Wong, M.L., Miyada, C.G., Switchenko, A.C., Goodman, T.C., and Wong, B., Isolation, characterization and expression of the gene that encodes D-arabinitol dehydrogenase in Candida tropicalis. Gene, 1995. 155(1): 123-128.
122. Adachi, O., Fujii, Y., Ghaly, M.F., Toyama, H., Shinagawa, E., and Matsushita, K., Membrane-bound quinoprotein D-arabitol dehydrogenase of Gluconobacter suboxydans IFO 3257: a versatile enzyme for the oxidative fermentation of various ketoses. Biosci Biotechnol Biochem, 2001. 65(12): 2755-2762.
123. Hallborn, J., Walfridsson, M., Penttila, M., Keranen, S., and Hahn-Hagerdal, B., A short-chain dehydrogenase gene from Pichia stipitis having D-arabinitol dehydrogenase activity. Yeast, 1995. 11(9): 839-847.
124. Kavanagh, K.L., Klimacek, M., Nidetzky, B., and Wilson, D.K., Crystal structure of Pseudomonas fluorescens mannitol 2-dehydrogenase: evidence for a very divergent long-chain dehydrogenase family. Chem Biol Interact, 2003. 143-144: 551-558.

FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
57
Annex
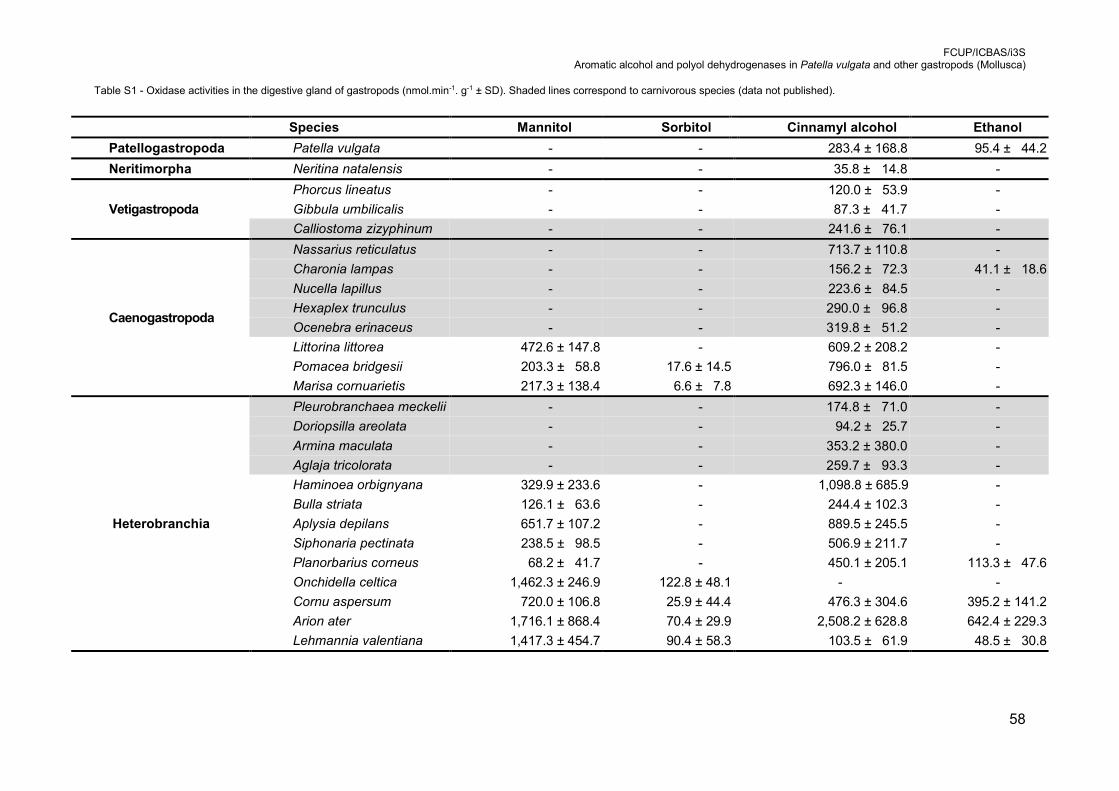
FCUP/ICBAS/i3S Aromatic alcohol and polyol dehydrogenases in Patella vulgata and other gastropods (Mollusca)
58
Table S1 - Oxidase activities in the digestive gland of gastropods (nmol.min-1. g-1 ± SD). Shaded lines correspond to carnivorous species (data not published).
Species Mannitol Sorbitol Cinnamyl alcohol Ethanol
Patellogastropoda Patella vulgata - - 283.4 ± 168.8 95.4 ± 44.2
Neritimorpha Neritina natalensis - - 35.8 ± 14.8 -
Vetigastropoda
Phorcus lineatus - - 120.0 ± 53.9 -
Gibbula umbilicalis - - 87.3 ± 41.7 -
Calliostoma zizyphinum - - 241.6 ± 76.1 -
Caenogastropoda
Nassarius reticulatus - - 713.7 ± 110.8 -
Charonia lampas - - 156.2 ± 72.3 41.1 ± 18.6
Nucella lapillus - - 223.6 ± 84.5 -
Hexaplex trunculus - - 290.0 ± 96.8 -
Ocenebra erinaceus - - 319.8 ± 51.2 -
Littorina littorea 472.6 ± 147.8 - 609.2 ± 208.2 -
Pomacea bridgesii 203.3 ± 58.8 17.6 ± 14.5 796.0 ± 81.5 -
Marisa cornuarietis 217.3 ± 138.4 6.6 ± 7.8 692.3 ± 146.0 -
Heterobranchia
Pleurobranchaea meckelii - - 174.8 ± 71.0 -
Doriopsilla areolata - - 94.2 ± 25.7 -
Armina maculata - - 353.2 ± 380.0 -
Aglaja tricolorata - - 259.7 ± 93.3 -
Haminoea orbignyana 329.9 ± 233.6 - 1,098.8 ± 685.9 -
Bulla striata 126.1 ± 63.6 - 244.4 ± 102.3 -
Aplysia depilans 651.7 ± 107.2 - 889.5 ± 245.5 -
Siphonaria pectinata 238.5 ± 98.5 - 506.9 ± 211.7 -
Planorbarius corneus 68.2 ± 41.7 - 450.1 ± 205.1 113.3 ± 47.6
Onchidella celtica 1,462.3 ± 246.9 122.8 ± 48.1 - -
Cornu aspersum 720.0 ± 106.8 25.9 ± 44.4 476.3 ± 304.6 395.2 ± 141.2
Arion ater 1,716.1 ± 868.4 70.4 ± 29.9 2,508.2 ± 628.8 642.4 ± 229.3
Lehmannia valentiana 1,417.3 ± 454.7 90.4 ± 58.3 103.5 ± 61.9 48.5 ± 30.8