angptl3 blockade with a human monoclonal antibody reduces ... · 1310 journal of lipid research...
TRANSCRIPT

1308 Journal of Lipid Research Volume 56, 2015
Copyright © 2015 by the American Society for Biochemistry and Molecular Biology, Inc.
This article is available online at http://www.jlr.org
LPL plays a central role in the maintenance of normal lipid levels in the blood. LPL is located at the luminal sur-face of the capillary endothelium and is the key enzyme for hydrolysis of core TGs in plasma chylomicron and VLDL particles, and release of fatty acids to adjacent tis-sues for storage and energy production ( 1, 2 ). The activity of LPL is regulated at the transcriptional and posttran-scriptional level in a tissue-specifi c manner ( 2 ). One of the posttranslational regulators of LPL activity is angiopoietin-like protein 3 (ANGPTL3), which belongs to a family of eight secreted proteins ( 3 ).
ANGPTL3 is secreted from the liver ( 4 ). Because the adult liver expresses little to no LPL, it is presumed that ANGPTL3 functions as a circulating inhibitor of LPL. AN-GPTL3 inhibits LPL activity in vitro and in vivo, and mice defi cient in Angptl3 have increased LPL activity and low plasma TG levels ( 5, 6 ). ANGPTL3 inhibits LPL activity by inducing a conformational change in LPL, resulting in in-creased susceptibility to cleavage by proprotein conver-tases, dissociation of LPL from the cell surface, and inhibition of its catalytic activity ( 7 ). In addition to inhibiting LPL, ANGPTL3 also inhibits the activity of endothelial lipase (EL), which hydrolyzes HDL phospholipids ( 8, 9 ).
Genetic studies have shown that humans with sequence variations in ANGPTL3 have reduced plasma lipid levels ( 10–15 ). In particular, individuals who have mutations in both ANGPTL3 alleles have pan-hypolipidemia with low
Abstract Angiopoietin-like protein 3 (ANGPTL3) is a cir-culating protein synthesized exclusively in the liver that inhibits LPL and endothelial lipase (EL), enzymes that hy-drolyze TGs and phospholipids in plasma lipoproteins. Here we describe the development and testing of a fully human monoclonal antibody (REGN1500) that binds ANGPTL3 with high affi nity. REGN1500 reversed ANGPTL3-induced inhibition of LPL activity in vitro. Intravenous admin-istration of REGN1500 to normolipidemic C57Bl/6 mice increased LPL activity and decreased plasma TG levels by � 50%. Chronic administration of REGN1500 to dyslipidemic C57Bl/6 mice for 8 weeks reduced circulating plasma levels of TG, LDL-cholesterol (LDL-C), and HDL-cholesterol (HDL-C) without any changes in liver, adipose, or heart TG contents. Studies in EL knockout mice revealed that REGN1500 reduced serum HDL-C through an EL-dependent mecha-nism. Finally, administration of a single dose of REGN1500 to dyslipidemic cynomolgus monkeys caused a rapid and pronounced decrease in plasma TG, nonHDL-C, and HDL-C. REGN1500 normalized plasma TG levels even in mon-keys with a baseline plasma TG greater than 400 mg/dl. Collectively, these data demonstrate that neutralization of ANGPTL3 using REGN1500 reduces plasma lipids in dys-lipidemic mice and monkeys, and thus provides a potential therapeutic agent for treatment of patients with hyperlipid-emia. —Gusarova, V., C. A. Alexa, Y. Wang, A. Rafi que, J. H. Kim, D. Buckler, I. J. Mintah, L. M. Shihanian, J. C. Cohen, H. H. Hobbs, Y. Xin, D. M. Valenzuela, A. J. Murphy, G. D. Yancopoulos, and J. Gromada. ANGPTL3 blockade with a human monoclonal antibody reduces plasma lipids in dys-lipidemic mice and monkeys. J. Lipid Res. 2015. 56: 1308–1317.
Supplementary key words lipoprotein lipase • endothelial lipase • tri-glycerides • cholesterol • hyperlipidemia • dyslipidemia • angiopoietin-like protein 3
This work was supported by National Institutes of Health Grant PO1 HL20948.
Manuscript received 16 September 2014 and in revised form 29 April 2015.
Published, JLR Papers in Press, May 11, 2015 DOI 10.1194/jlr.M054890
ANGPTL3 blockade with a human monoclonal antibody reduces plasma lipids in dyslipidemic mice and monkeys 1
Viktoria Gusarova , 2, * Corey A. Alexa , * Yan Wang , † Ashique Rafi que , * Jee Hae Kim , * David Buckler , * Ivory J. Mintah , * Lisa M. Shihanian , * Jonathan C. Cohen , § Helen H. Hobbs , † Yurong Xin , * David M. Valenzuela , * Andrew J. Murphy , * George D. Yancopoulos , * and Jesper Gromada *
Regeneron Pharmaceuticals, Inc. ,* Tarrytown, NY 10591; and Howard Hughes Medical Institute and Departments of Molecular Genetics † and Internal Medicine, § University of Texas Southwestern Medical Center , Dallas, TX 75390
Abbreviations: ALT, alanine aminotransferase; ANGPTL3, angio-poietin-like protein 3; AST, aspartate aminotransferase; EL, endo-thelial lipase; HDL-C, HDL-cholesterol; HL, hepatic lipase; LDL-C, LDL-cholesterol; mmH, myc-myc-His6; RU, resonance unit; TC, total cholesterol .
1 See referenced companion article, J. Lipid Res . 2015, 56: 1296–1307. 2 To whom correspondence should be addressed. e-mail: [email protected]
The online version of this article (available at http://www.jlr.org) contains supplementary data in the form of nine fi gures, one table, and text.
by guest, on April 29, 2019
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2015/05/11/jlr.M054890.DC1Supplemental Material can be found at:

ANGPTL3 antibody reduces plasma lipids 1309
running buffer and injected in duplicate over various recombi-nant ANGPTL3 protein captured surfaces for 3 min at 50 � l/min followed by 5 min of dissociation. All capture surfaces were re-generated with one 15 s pulse of 10 mM glycine-HCl (pH 1.5). This format used a saturating concentration of REGN1500 for binding. Dissociation was monitored by a double referencing procedure ( 19 ) ( Table 1 ). The data were processed and kinetic analyses performed using T200 Evaluation (version 1.0, Biacore). The equilibrium dissociation constant ( K D ) was calculated from the ratio of the dissociation rate constant divided by the associa-tion rate constant ( K D = k d / k a ).
Studies in mice Male C57Bl/6 mice (8 weeks of age) were purchased from
Jackson Laboratory or Taconic Farms. Lipg � / � mice were gener-ated on 75% C578Bl/6Tac and 25% 129S6/SvEvTac background using VelociGene® technology ( 20 ). VelociGene® allele identi-fi cation number is VG12135. All procedures were conducted in compliance with protocols approved by the Regeneron Phar-maceuticals Institutional Animal Care and Use Committee. REGN1500 and isotype control antibody were diluted with sterile PBS for sc injection into mice.
Single administration studies. Mice were maintained on a regu-lar chow diet (5001, LabDiet). To establish a baseline for serum chemistry parameters, serum samples were collected 7 days prior to antibody administration after a 4 h fast. On study day 0, mice were sorted into treatment groups based on their serum TG lev-els. Mice (n = 5–6 per group) were then administered a single sc injection of REGN1500 or isotype control antibody at the indi-cated doses. Subsequent serum samples were collected after a 4 h fast over the duration of the experiment and analyzed for serum chemistry parameters and human Fc levels. Postheparin plasma LPL activity was measured as described previously ( 21 ). A lipid tolerance test was performed 4 days after a single dose of REGN1500 (10 mg/kg). Mice were fasted for 2 h following intra-peritoneal administration of 20% intralipid (Baxter Healthcare) at 10 � l/g body weight. TG level was evaluated in blood collected from the tail at subsequent time points.
Multiple administration studies. Eight days prior to antibody administration (day � 8), baseline serum chemistries were mea-sured after a 4 h fast. On day � 3, mice were placed on a high-fat high-cholesterol diet [21% fat, 0.21% cholesterol (wt/wt); cata-log #12079B, Research Diets]. On study day 0, mice were sorted into treatment groups based on their serum TG levels (n = 10 per group). Mice were injected with REGN1500 or isotype control antibody once a week for 8 weeks. Body weights were measured every week. Mice were euthanized 1 week after the last injec-tion and livers, epididymal white adipose tissue, and hearts were collected, weighed, and frozen for subsequent TG con-tent measurements .
Serum chemistry analyses Circulating TG, total cholesterol (TC), LDL-C, HDL-C, NEFA,
alanine aminotransferase (ALT), and aspartate aminotransferase (AST) levels were determined in serum using an ADVIA ® 1800 blood chemistry analyzer (Bayer, Leverkusen, Germany). Non-HDL-C levels were calculated by subtracting HDL-C from TC values.
Lipoprotein separation using HPLC Lipoprotein particles were separated from serum samples by
dual detection HPLC system (Waters Corp.) using a tandem Su-perose 6 HR 10/300GL column ( 22 ). Cholesterol and TG lipo-protein concentrations were calculated as the area under the curve using VLDL, LDL, HDL (Biomedical Technologies), and
plasma TG, LDL-cholesterol (LDL-C), and HDL-choles-terol (HDL-C) levels and increased plasma LPL activity ( 16 ). These fi ndings confi rm the importance of ANGPTL3 in human lipoprotein metabolism and make blocking AN-GPTL3 with a monoclonal antibody a potential therapy to treat hyperlipidemia. In this study, we describe the fully human monoclonal antibody, REGN1500, that binds with high affi nity to ANGPTL3 and effectively inhibits its activ-ity in vivo, leading to robust lowering of plasma lipids in dyslipidemic mice and nonhuman primates.
MATERIALS AND METHODS
Antibodies and protein reagents REGN1500 was derived using Regeneron’s Velocimmune®
technology platform ( 17 ) and is a fully human monoclonal an-tibody with high affi nity to ANGPTL3 from multiple species (mouse, rat, monkey, and human). REGN1500 has a human IgG4 constant region with a stabilizing mutation in the hinge re-gion (serine to proline in position 108 in GenBank #P01864) to minimize half-antibody formation, which is known to occur for the natural IgG4 isotype ( 18 ). An isotype-matched antibody with irrelevant specifi city was used as control.
The following proteins were obtained from R&D Systems, where “HisN” indicates a C-terminal oligohistidine tag (N is the number of His residues): hANGPTL3 (S17-E460)-His10 and mANGPTL3 (S17-T455)-His10. Additional recombinant epitope-tagged proteins were produced in Chinese hamster ovarian cells after stable transfection using vectors that substituted nonnative for endogenous signal peptides. Chinese hamster ovarian-expressed proteins were purifi ed using immobilized metal affi nity chroma-tography and dialyzed into Tris-buffered saline (pH 7.5) or PBS containing 5% glycerol (pH 7.4). These proteins included hANGPTL3 (S17-K170)-His6, MfANGPTL3 (S17-K170)-myc-myc-His6 (mmH) (at the C terminus), rANGPTL3 (S17-D240)-mmH, and mANGPTL3 (S17-T455)-His6 .
Surface plasmon resonance-Biacore Surface plasmon resonance experiments were performed on a
Biacore T200 instrument using a dextran-coated (CM4) chip at 25°C. The running buffer was fi ltered HBS-T [10 mM HEPES, 150 mM NaCl, 3.4 mM EDTA, and 0.05% polysorbate 20 (pH 7.4)]. A capture sensor surface was prepared by covalently immo-bilizing � -histidine antibody (Qiagen) to the chip surface using (1-ethyl-3-[3-dimethylaminopropyl]carbodiimide hydrochloride)/ N -hydroxysuccinimide (EDC/NHS) coupling chemistry. Follow-ing surface activation, � -histidine antibody in coupling buffer [0.1 M acetate buffer (pH 4.5)] was injected over the activated chip surface until a resonance unit (RU) signal of about 4,000 RU ( � -penta histidine mouse monoclonal antibody; Qiagen) was reached. The activated coupled chip surfaces were then washed and treated with 10 mM glycine-HCl (pH 1.5) to remove uncou-pled residual proteins. Recombinant proteins derived from hu-man and nonhuman species ANGPTL3 were diluted into the running buffer and captured through the histidine tag on the coupled � -histidine antibody chip surface at low density (6–28 RU). One fl ow cell of the biosensor chip was left without cap-tured protein to provide a reference surface for the kinetic ex-periment. The capture protocol was designed to yield capture levels of mAb that resulted in an R max no greater than 80 RU. Following the capture step, � -ANGPTL3 antibody was serially diluted 2-fold at concentration (50 nM to 0.39 nM) in the
by guest, on April 29, 2019
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2015/05/11/jlr.M054890.DC1Supplemental Material can be found at:

1310 Journal of Lipid Research Volume 56, 2015
system (Beckman Coulter Inc.). The serum chemistry methods were validated ( 25 ).
Data analyses All data are mean ± SEM. Statistical analyses were performed
utilizing GraphPad software Prism 6.0. LPL and hepatic lipase (HL) activities in REGN1500- and control antibody-treated mice were compared by Student’s t -test. All other parameters were ana-lyzed by two-way ANOVA; a threshold of P < 0.05 was considered statistically signifi cant. If a signifi cant F ratio was obtained with two-way ANOVA, post hoc analysis was conducted between groups with Bonferroni posttests. In the monkey study, the aver-age of each parameter on days � 15, � 7, � 2, and 0 was used as the baseline value. Mean values and standard errors of each pa-rameter in each group at a specifi c date were calculated. In addition, the percentage change of the mean value of each pa-rameter in each experimental group compared with the baseline value was calculated. Student’s t -tests (two tails, paired) were per-formed in each group to compare the differences of the value at a specifi c date and the baseline value.
RESULTS
In vitro characterization of anti-ANGPTL3 antibody REGN1500
The relative affi nity of REGN1500 to human, mouse, rat, and monkey ANGPTL3 was compared using surface plas-mon resonance. REGN1500 bound ANGPTL3 from the four species with comparable affi nities ( K D = 0.26–1.28 nM)
TG (Teco Diagnostics) standards. The cholesterol or TG concen-trations in the standards were determined using enzymatic meth-ods (Teco Diagnostics).
TG tissue content Lipids were extracted from tissues as described ( 23 ). Lipids
were solubilized as outlined earlier ( 24 ). The levels of TG were measured using enzymatic assays (Infi nity; Thermo Fisher Scien-tifi c) and normalized to wet tissue weight.
Study in cynomolgus monkeys The study was performed by Crown Bioscience (Taicang,
Jiangsu, China). Seventeen spontaneous hypertriglyceridemic monkeys were selected for the study based on their overnight fasted serum TG, nonHDL-C, and HDL-C profi le (supplemen-tary Table 1). The monkeys were individually housed, had free access to water and were fed twice daily with a complete nutri-tionally balanced diet (Shanghai Shilin Biotechnology Inc., Shanghai, China), which was enriched with seasonal fruits and vegetables. All animal procedures were approved by the Crown Bioscience Institutional Animal Care and Use Committee and performed according to guidelines approved by the Associa-tion for Assessment and Accreditation of Laboratory Animal Care.
The seventeen animals were divided into three groups. On day 0, the monkeys were given either vehicle or REGN1500 (3 mg/kg or 10 mg/kg). Blood (4 ml) was collected following a 16 h fast with 1 to 5 day intervals up to day 55 via venipuncture into BD sterile venous blood collection tubes for serum preparation and blood glucose measurements (ACCU-CHEK Active, Roche). Se-rum TG, TC, LDL-C, HDL-C, NEFA, ALT, AST, and insulin were assayed in a Beckman Coulter UniCelDxC 800 Synchron clinical
TABLE 1. Summary of kinetic binding parameters for the interaction of REGN1500 with human, monkey, rat, and mouse ANGPTL3 proteins
Protein
Kinetic Binding Parameters
k a (M � 1 s � 1 ) k d (s � 1 ) K D (M) T 1/2 (min)
hANGPTL3 (S17-K170)-His6 1.09 × 10 6 1.39 × 10 � 3 1.28 × 10 � 9 8.3hANGPTL3 (S17-E460)-His10 1.01 × 10 6 9.22 × 10 � 4 9.15 × 10 � 10 12.5MfANGPTL3 (S17-K170)-mmH 2.30 × 10 6 5.99 × 10 � 4 2.61 × 10 � 10 19.3rANGPTL3 (S17-D240)-mmH 2.00 × 10 6 7.71 × 10 � 4 3.85 × 10 � 10 15.0mANGPTL3 (S17-T455)-His6 1.83 × 10 6 6.29 × 10 � 4 3.44 × 10 � 10 18.4
hANGPTL3 (S17-K170)-His6, N-terminal domain human ANGPTL3, amino acids 17-170, with a C-terminal hexahistidine tag; hANGPTL3 (S17-E460)-His10, full-length human ANGPTL3 with a C-terminal decahistidine tag; MfANGPTL3 (S17-K170)-mmH, N-terminal domain of monkey ( Macaca fascicularis ) ANGPTL3, amino acids 17-170, with a C-terminal myc-myc-hexahistidine tag; rANGPTL3 (S17-D240)-mmH, N-terminal domain human ANGPTL3, amino acids 17-240, with a C-terminal myc-myc-hexahistidine tag; mANGPTL3 (S17-T455)-His6, full-length mouse ANGPTL3 with a C-terminal hexahistidine tag; mANGPTL3 (S17-T455)-His6, full-length mouse ANGPTL3 with a C-terminal hexahistidine tag. ANGPTL3 proteins were immobilized onto the sensor surface by anti-penta-histidine capture. REGN1500 (12.5–0.39 nM) was injected across the captured protein chip surface and antibody-dependent changes in RUs were monitored.
TABLE 2. Summary of IC 50 values for REGN1500 blockade of ANGPTL3-mediated LPL inhibition
Parameter
Protein
hANGPTL3 (S17-E460)-His10
hANGPTL3 (S17-K170)-His6
MfANGPTL3 (S17-K170)-mmH
rANGPTL3 (S17-D240)-mmH
mANGPTL3 (S17-T455)-His10
REGN1500, IC 50 (nM) a 9.6 2.9 10.4 1.0 13.6Control Ab, IC 50 (nM) b NB NB NB NB NB
hANGPTL3, human ANGPTL3; MfANGPTL3, M. fascicularis ANGPTL3; rANGPTL3, rat ANGPTL3; mANGPTL3, mouse ANGPTL3; His10, decahistidine tag; His6, hexahistidine tag; mmH, myc-myc-hexahistidine tag; NB, no detectable blocking under the assay conditions used.
a IC 50 values were determined using constant concentrations of ANGPTL proteins that were within 2.5-fold of the measured EC 50 values for the ANGPTL protein inhibiting LPL lipase activity.
b Control Ab, hIgG4 isotype control antibody.
by guest, on April 29, 2019
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2015/05/11/jlr.M054890.DC1Supplemental Material can be found at:
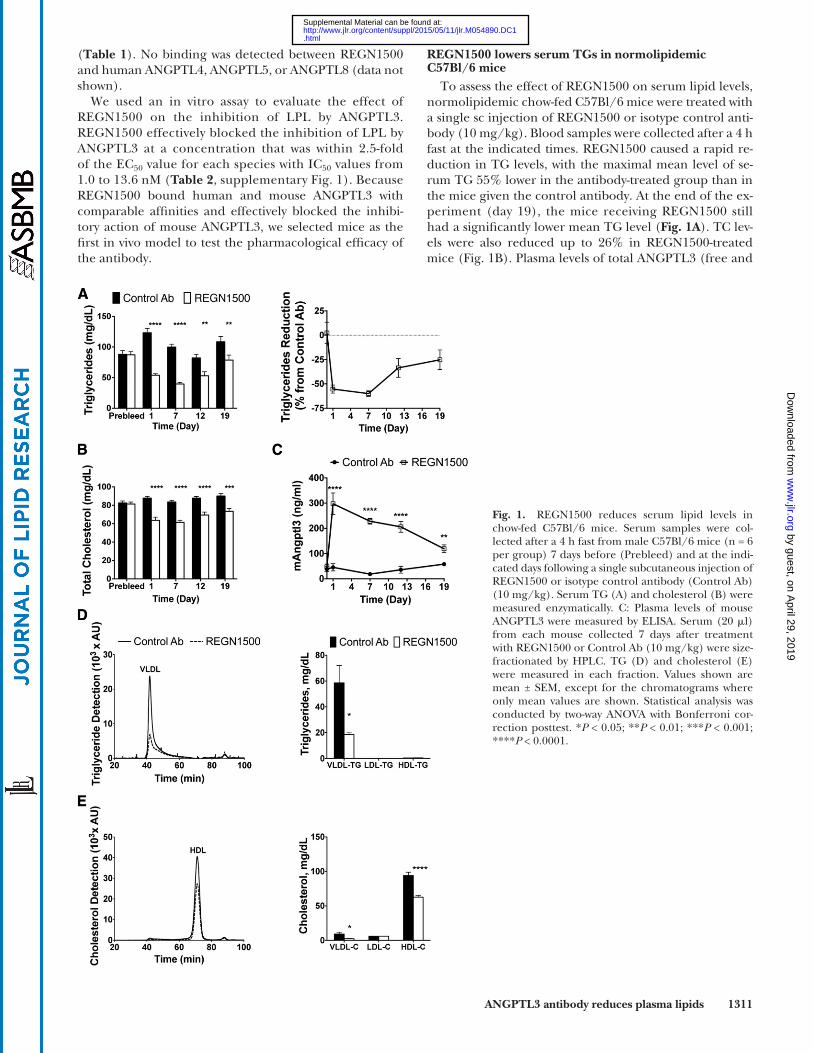
ANGPTL3 antibody reduces plasma lipids 1311
Fig. 1. REGN1500 reduces serum lipid levels in chow-fed C57Bl/6 mice. Serum samples were col-lected after a 4 h fast from male C57Bl/6 mice (n = 6 per group) 7 days before (Prebleed) and at the indi-cated days following a single subcutaneous injection of REGN1500 or isotype control antibody (Control Ab) (10 mg/kg). Serum TG (A) and cholesterol (B) were measured enzymatically. C: Plasma levels of mouse ANGPTL3 were measured by ELISA. Serum (20 � l) from each mouse collected 7 days after treatment with REGN1500 or Control Ab (10 mg/kg) were size-fractionated by HPLC. TG (D) and cholesterol (E) were measured in each fraction. Values shown are mean ± SEM, except for the chromatograms where only mean values are shown. Statistical analysis was conducted by two-way ANOVA with Bonferroni cor-rection posttest. * P < 0.05; ** P < 0.01; *** P < 0.001; **** P < 0.0001.
( Table 1 ). No binding was detected between REGN1500 and human ANGPTL4, ANGPTL5, or ANGPTL8 (data not shown).
We used an in vitro assay to evaluate the effect of REGN1500 on the inhibition of LPL by ANGPTL3. REGN1500 effectively blocked the inhibition of LPL by ANGPTL3 at a concentration that was within 2.5-fold of the EC 50 value for each species with IC 50 values from 1.0 to 13.6 nM ( Table 2 , supplementary Fig. 1). Because REGN1500 bound human and mouse ANGPTL3 with comparable affi nities and effectively blocked the inhibi-tory action of mouse ANGPTL3, we selected mice as the fi rst in vivo model to test the pharmacological effi cacy of the antibody.
REGN1500 lowers serum TGs in normolipidemic C57Bl/6 mice
To assess the effect of REGN1500 on serum lipid levels, normolipidemic chow-fed C57Bl/6 mice were treated with a single sc injection of REGN1500 or isotype control anti-body (10 mg/kg). Blood samples were collected after a 4 h fast at the indicated times. REGN1500 caused a rapid re-duction in TG levels, with the maximal mean level of se-rum TG 55% lower in the antibody-treated group than in the mice given the control antibody. At the end of the ex-periment (day 19), the mice receiving REGN1500 still had a signifi cantly lower mean TG level ( Fig. 1A ). TC lev-els were also reduced up to 26% in REGN1500-treated mice ( Fig. 1B ). Plasma levels of total ANGPTL3 (free and
by guest, on April 29, 2019
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2015/05/11/jlr.M054890.DC1Supplemental Material can be found at:

1312 Journal of Lipid Research Volume 56, 2015
REGN1500 treatment did not change body weight, glu-cose, or insulin levels in the db / db mice (supplementary Fig. 4F–H). Thus, REGN1500 reduces circulating TG by upregulating LPL activity in vivo through blocking of ANGPTL3.
The effect of ANGPTL3 inhibition with REGN1500 on TG clearance was evaluated by acute fat loading. C57Bl/6 mice were treated with either REGN1500 or control antibody (10 mg/kg). Four days later, they were injected intraperitoneally with an intralipid emulsion (20%) after a 2 h fast. Plasma TG levels were signifi -cantly lower at baseline and increased only minimally in response to fat challenge in mice treated with REGN1500 ( Fig. 2B ).
REGN1500 lowers serum TG and cholesterol levels in dyslipidemic C57Bl/6 mice
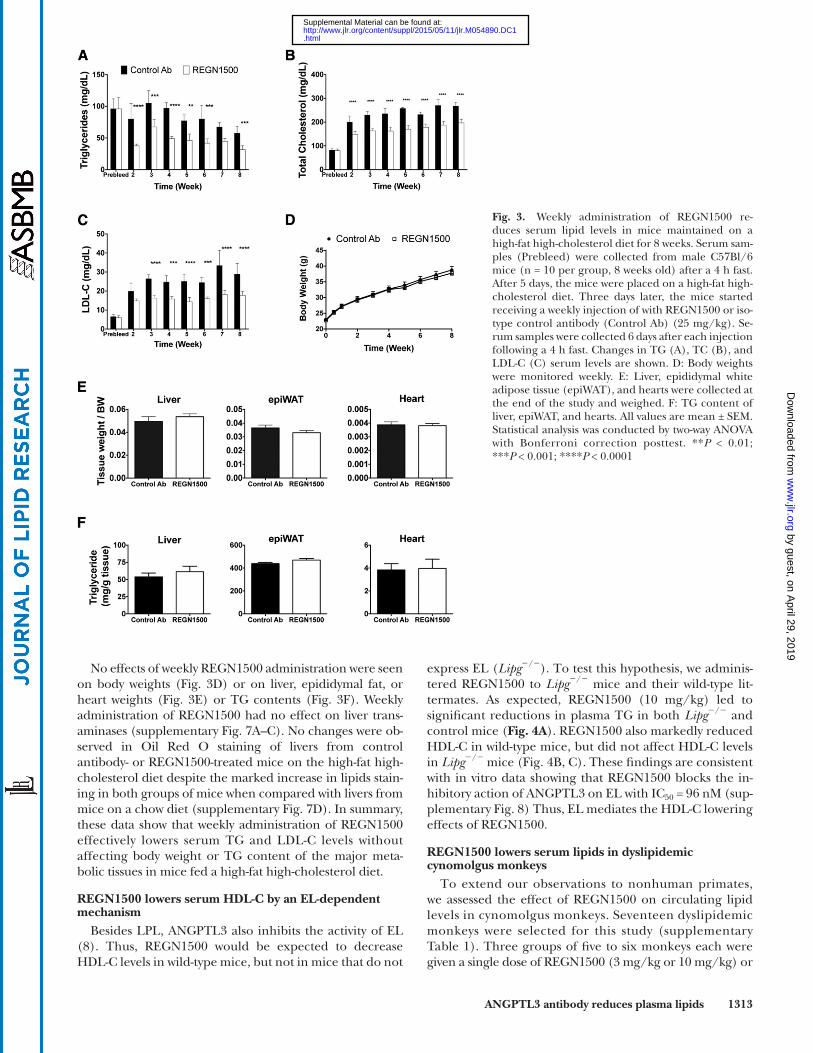
C57Bl/6 mice were placed on a high-fat high-cholesterol diet 3 days prior to initiating treatment with REGN1500. REGN1500 or the control antibody were administered once weekly (25 mg/kg) for 8 weeks. Serum samples were collected after a 4 h fast, 6 days after each injection. A marked and sustained reduction in circulating TG levels (up to � 53%) was seen in the mice given REGN1500 compared with the control antibody ( Fig. 3A ). Hepatic TG secretion rate, assessed following administration of Triton WR1339, was reduced by 58% in mice receiving REGN1500 compared with control antibody (supple-mentary Fig. 5). The reduction in hepatic TG secretion rate was not produced by changes in de novo lipogenesis or fatty acid oxidation, as no differences were found in the gene expression of the major enzymes of these pathways in livers from mice administrated REGN1500 or control antibody (supplementary Fig. 6). Signifi cant reductions in TC (up to � 35%; Fig. 3B ) and LDL-C (up to � 45%; Fig. 3C ) were also observed, showing that REGN1500 can reduce cholesterol levels originally elevated by a high-fat high-cholesterol diet. The reductions in circulating lipids were sustained throughout the 8 weeks of the study.
bound) increased within 1 day of treatment with REGN1500 and then progressively fell over the course of the experi-ment ( Fig. 1C ). Angptl3 liver expression did not change in mice administered REGN1500 (supplementary Fig. 2). Se-rum was separated on a Superose 6 column using HPLC to determine TG ( Fig. 1D ) and cholesterol ( Fig. 1E ) distribu-tion among lipoproteins. Quantifi cation of the lipopro-tein distributions revealed that REGN1500 signifi cantly reduced VLDL-TG, VLDL-cholesterol, and HDL-C levels in normolipidemic C57Bl/6 mice ( Fig. 1D, E ).
A dose-response study was performed in chow-fed C57Bl/6 mice. Administration of a single dose of REGN1500 at 1 mg/kg was not associated with signifi cant changes in plasma TG levels. However, doses of REGN1500 at 5, 10, and 25 mg/kg elicited a progressive dose-dependent re-duction in circulating TG levels with maximal effects ob-served at day 7 (supplementary Fig. 3A). The greatest reduction of TG ( � 66%) occurred with the highest anti-body dose (25 mg/kg) (supplementary Fig. 3B). No change in LDL-C was observed at any dose (data not shown). The corresponding human Fc levels in serum are shown in supplementary Fig. 3C, indicating that a serum REGN1500 concentration of between 5 and 15 � g/ml is required to induce signifi cant pharmacological effects on circulating TG levels in normolipidemic C57Bl/6 mice.
REGN1500 increases postheparin plasma LPL activity First, we confi rmed that inhibition of ANGPTL3 by
treatment with REGN1500 was associated with an increase in postheparin plasma LPL activity in both C57Bl/6 mice ( Fig. 2A ) and in dyslipidemic db / db mice (supplementary Fig. 4A). The postheparin LPL activity was increased by 1.8-fold in C57Bl/6 mice and 2.5-fold in the db / db mice with REGN1500 treatment, whereas HL activity remained unchanged ( Fig. 2A , supplementary Fig. 4B). In the db / db mice, REGN1500 signifi cantly reduced serum TG levels ( � 48%; P < 0.0001), TC levels ( � 37%; P < 0.01), and NE-FAs ( � 35%; P < 0.01) (supplementary Fig. 4C–E).
Fig. 2. REGN1500 increases postheparin plasma LPL and improves lipid tolerance. A: Postheparin plasma LPL and HL activity of chow-fed C57Bl/6 mice treated with REGN1500 or control antibody (Control Ab) (10 mg/kg, n = 5 per group). Postheparin plasma was pooled and fractionated on a heparin column to sepa-rate HL and LPL, and TG hydrolase activity was measured as described in the Materials and Methods. B: Effect of REGN1500 treatment on plasma TG levels following a lipid tolerance test. Male C57Bl/6 mice were treated with a single dose of REGN1500 or control antibody (10 mg/kg, n = 5 per group) 4 days prior to the intraperitoneal administration of Intralipid (10 � l/g of 20% Intralipid). Values are mean ± SEM. Statistical analysis was conducted by Student’s t -test comparing the groups at each time point. * P < 0.05; ** P < 0.01.
by guest, on April 29, 2019
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2015/05/11/jlr.M054890.DC1Supplemental Material can be found at:
ANGPTL3 antibody reduces plasma lipids 1313
express EL ( Lipg � / � ). To test this hypothesis, we adminis-tered REGN1500 to Lipg � / � mice and their wild-type lit-termates. As expected, REGN1500 (10 mg/kg) led to signifi cant reductions in plasma TG in both Lipg � / � and control mice ( Fig. 4A ). REGN1500 also markedly reduced HDL-C in wild-type mice, but did not affect HDL-C levels in Lipg � / � mice ( Fig. 4B, C ). These fi ndings are consistent with in vitro data showing that REGN1500 blocks the in-hibitory action of ANGPTL3 on EL with IC 50 = 96 nM (sup-plementary Fig. 8) Thus, EL mediates the HDL-C lowering effects of REGN1500.
REGN1500 lowers serum lipids in dyslipidemic cynomolgus monkeys
To extend our observations to nonhuman primates, we assessed the effect of REGN1500 on circulating lipid levels in cynomolgus monkeys. Seventeen dyslipidemic monkeys were selected for this study (supplementary Table 1). Three groups of fi ve to six monkeys each were given a single dose of REGN1500 (3 mg/kg or 10 mg/kg) or
Fig. 3. Weekly administration of REGN1500 re-duces serum lipid levels in mice maintained on a high-fat high-cholesterol diet for 8 weeks. Serum sam-ples (Prebleed) were collected from male C57Bl/6 mice (n = 10 per group, 8 weeks old) after a 4 h fast. After 5 days, the mice were placed on a high-fat high-cholesterol diet. Three days later, the mice started receiving a weekly injection of with REGN1500 or iso-type control antibody (Control Ab) (25 mg/kg). Se-rum samples were collected 6 days after each injection following a 4 h fast. Changes in TG (A), TC (B), and LDL-C (C) serum levels are shown. D: Body weights were monitored weekly. E: Liver, epididymal white adipose tissue (epiWAT), and hearts were collected at the end of the study and weighed. F: TG content of liver, epiWAT, and hearts. All values are mean ± SEM. Statistical analysis was conducted by two-way ANOVA with Bonferroni correction posttest. ** P < 0.01; *** P < 0.001; **** P < 0.0001
No effects of weekly REGN1500 administration were seen on body weights ( Fig. 3D ) or on liver, epididymal fat, or heart weights ( Fig. 3E ) or TG contents ( Fig. 3F ). Weekly administration of REGN1500 had no effect on liver trans-aminases (supplementary Fig. 7A–C). No changes were ob-served in Oil Red O staining of livers from control antibody- or REGN1500-treated mice on the high-fat high-cholesterol diet despite the marked increase in lipids stain-ing in both groups of mice when compared with livers from mice on a chow diet (supplementary Fig. 7D). In summary, these data show that weekly administration of REGN1500 effectively lowers serum TG and LDL-C levels without affecting body weight or TG content of the major meta-bolic tissues in mice fed a high-fat high-cholesterol diet.
REGN1500 lowers serum HDL-C by an EL-dependent mechanism
Besides LPL, ANGPTL3 also inhibits the activity of EL ( 8 ). Thus, REGN1500 would be expected to decrease HDL-C levels in wild-type mice, but not in mice that do not
by guest, on April 29, 2019
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2015/05/11/jlr.M054890.DC1Supplemental Material can be found at:

1314 Journal of Lipid Research Volume 56, 2015
vehicle. REGN1500 produced a robust and sustained dose-dependent reduction in circulating TG that reached a maximum within 1 to 2 days following the treatment ( Fig. 5A ). The 3 mg/kg dose of REGN1500 treatment re-sulted in a 48% reduction in mean plasma TG levels (from 320 ± 7 mg/dl to 165 ± 19 mg/dl for days 1 and 2; P < 0.05; n = 6), whereas the 10 mg/kg dose lowered plasma TG by 89% (from 411 ± 42 mg/dl to 46 ± 16 mg/dl for days 1 and 2; P < 0.05; n = 5). The higher dose of REGN1500 lowered serum TG to levels that were similar to the normolipid-emic cynomolgus monkeys (58 ± 3 mg/dl; n = 60, unpub-lished observations). The reductions in plasma TG were maintained for 33 days before returning to baseline levels ( Fig. 5A ). The absolute reduction in plasma TG levels is provided for the higher dose treatment group ( Fig. 5B ). NonHDL-C was reduced by 44% and reached a nadir at day 3 ( Fig. 5C ). REGN1500 did not alter the plasma levels of LDL-C ( Fig. 5D ), perhaps because plasma LDL-C levels were low at baseline (58 ± 7 mg/dl; n = 17) and similar to those of normolipidemic monkeys (54 ± 4 mg/dl; n = 60). At the higher doses of REGN1500, the plasma HDL-C lev-els fell by � 30% at days 10 and 14 ( Fig. 5E ). No changes were observed in liver enzyme levels (ALT and AST, sup-plementary Fig. 9) or fasting glucose or insulin (data not shown).
DISCUSSION
In this paper we show that administration of a high affi nity antibody to ANGPTL3 rapidly and effectively re-duces serum TG and cholesterol levels in normolipid-emic mice, as well as in dyslipidemic mice and monkeys. The phenotype of the treated animals resembles that seen in humans with genetic defi ciency of ANGPTL3 ( 16, 26–31 ) and suggests that inhibition of ANGPTL3 may be benefi cial for treatment of patients with mixed dyslipidemia.
A mouse antibody to ANGPTL3 was shown previously to lower circulating levels of plasma TG and TC in normo-lipidemic and hyperlipidemic mice ( 32, 33 ). Here we de-scribe the development of a new fully human monoclonal antibody that binds human, monkey, mouse, and rat ANG-PTL3 with high affi nity. REGN1500 treatment protected LPL activity from ANGPTL3 inhibition in vitro and in vivo after the antibody was injected into the circulation of mice. The increase in LPL activity in mice treated with REGN1500 resembles that seen in Angptl3 knockout mice ( 5, 6 ) and could account for the rapid clearance of plasma TG following a lipid challenge. A single dose of REGN1500 (5 mg/kg or greater) effectively reduced circulating TG levels in both normolipidemic and hyperlipidemic mice,
Fig. 4. REGN1500 reduces HDL-C in WT but not in EL-defi cient ( Lipg � / � ) mice. Serum samples were col-lected after a 4 h fast from male chow-fed Lipg � / � mice and their WT littermates (n = 7 per group) before (Prebleed; 7 days) and 4 days after a single administration of REGN1500 or isotype control antibody (Control Ab) at 10 mg/kg. Serum was ana-lyzed for TG (A) and HDL-C (B). C: Chromatogram showing cholesterol distribution among lipoprotein subclasses after REGN1500 or control antibody ad-ministration in WT and Lipg � / � mice. All values are mean ± SEM, except chromatograms where only mean values are shown. Statistical analysis was by two-way ANOVA with Bonferroni correction posttest. * P < 0.05; ** P < 0.01.
by guest, on April 29, 2019
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2015/05/11/jlr.M054890.DC1Supplemental Material can be found at:

ANGPTL3 antibody reduces plasma lipids 1315
We also evaluated the effect of chronic treatment with REGN1500. Weekly administration of REGN1500 in C57Bl/6 mice fed a high-fat high-cholesterol diet for 8 weeks caused a sustained reduction in circulating TG,
and in dyslipidemic cynomolgus monkeys. These fi nd-ings indicate that REGN1500 reduces circulating TG lev-els by upregulating LPL activity through inhibition of ANGPTL3.
Fig. 5. REGN1500 reduces circulating TG, nonHDL-C, and HDL-C levels in dyslipidemic cynomolgus monkeys. Baseline serum samples were collected at days � 15, � 7, � 2, and 0 following a 16 h fast. Seventeen monkeys were divided into three groups and were administered vehicle or REGN1500 (3 or 10 mg/kg) on day 0 of the study. Serum samples were collected on multiple days and analyzed for TG (A), nonHDL-C (C), LDL-C (D), and HDL-C (E), and are represented as a percent change from baseline. All values are mean ± SEM. Panel (B) shows aver-age changes in absolute TG values before and following treatment with REGN1500 (10 mg/kg). For each parameter, Student’s t -test (two tails, paired) was performed in each group comparing the mean value at each time point to the baseline value. * P < 0.05; ** P < 0.01.
by guest, on April 29, 2019
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2015/05/11/jlr.M054890.DC1Supplemental Material can be found at:

1316 Journal of Lipid Research Volume 56, 2015
TC, and LDL-C levels without affecting body weight, liver enzymes, or hepatic, adipose, or heart TG content. The reduction in serum TG levels can in part be attributed to reduced hepatic TG secretion rate. We show that the abil-ity of REGN1500 to lower serum lipid levels is indepen-dent of changes in genes involved in de novo lipogenesis or in fatty acid oxidation in the liver.
ANGPTL3 inactivation with REGN1500 reduced LDL-C levels only in animals fed a high-fat high-cholesterol diet, but not in normolipidemic mice or cynomolgus monkeys that have normally low baseline LDL-C levels. The mecha-nism responsible for the LDL-lowering effect of ANGPTL3 is currently not clear, but appears not to involve the major canonical pathways of clearance of ApoB-containing lipo-proteins ( 34 ) .
The administration of REGN1500 to normolipidemic and hyperlipidemic mice also caused a reduction in plasma levels of HDL-C. Humans who are homozygous for inactivating mutations in ANGPTL3 have lower plasma HDL-C levels ( 26–30 ). It has been proposed that the re-duction in plasma HDL-C levels in ANGPTL3-defi cient individuals is due to the inhibitory effects of ANGPTL3 on the phospholipase activity of EL ( 8 ). Plasma phospholi-pase activity is higher in Angptl3 � / � mice than in wild-type mice and the EL activity can be inhibited by recombinant human ANGPTL3 in vitro ( 8 ). In this study, administra-tion of REGN1500 to mice lacking EL resulted in a reduc-tion in circulating TG levels, but not in plasma HDL-C levels, thus confi rming that HDL-C reduction results from the relief of EL inhibition by ANGPTL3.
HDL-C plays a key role in the process of reverse cho-lesterol transport by promoting the effl ux of excess cho-lesterol from peripheral tissues to the liver for biliary excretion ( 35 ). The possible therapeutic benefi t of ANG-PTL3 blockade using REGN1500 to lower both TG and LDL-C is possibly tempered by concern about its ability to decrease HDL-C. However, recent genetic and pharmaco-logical fi ndings have questioned the importance of HDL-C as a marker for cardiovascular risk prediction ( 36–38 ), particularly when refl ecting changes due to EL action . Consistent with this scenario, it was recently shown that humans with a common inactivating mutation in LIPG and higher plasma levels of HDL-C are not protected from coronary heart disease ( 36 ). Measurement of reverse cho-lesterol transport and assessment of HDL functionality may help to better understand the potential consequences of reduced plasma HDL-C in humans.
Although very limited analysis is available on the preva-lence of coronary heart disease in carriers of homozygous ANGPTL3 mutations, Pisciotta et al. ( 28 ) reported no clinical evidence of atherosclerosis in their patients, attrib-uting these fi ndings to markedly reduced levels of pro-atherogenic apoB-containing lipoproteins (VLDL, VLDL remnants, and LDL) in the mutation carriers.
In summary, biochemical characterization and in vivo stud-ies in normolipidemic and dyslipidemic mice and monkeys demonstrate that inhibition of ANGPTL3 by REGN1500 lowers the circulating levels of both cholesterol and TG sig-nifi cantly. These data suggest that REGN1500 may provide
a new therapeutic approach for lipid-lowering, especially in subjects with combined hyperlipidemia.
REFERENCES
1 . Merkel , M. , R. H. Eckel , and I. J. Goldberg . 2002 . Lipoprotein lipase: genetics, lipid uptake, and regulation. J. Lipid Res. 43 : 1997 – 2006 .
2 . Wang , H. , and R. H. Eckel . 2009 . Lipoprotein lipase: from gene to obesity. Am. J. Physiol. Endocrinol. Metab. 297 : E271 – E288 .
3 . Shimizugawa , T. , M. Ono , M. Shimamura , K. Yoshida , Y. Ando , R. Koishi , K. Ueda , T. Inaba , H. Minekura , T. Kohama , et al . 2002 . ANGPTL3 decreases very low density lipoprotein triglyceride clearance by inhibition of lipoprotein lipase. J. Biol. Chem. 277 : 33742 – 33748 .
4 . Ono , M. , T. Shimizugawa , M. Shimamura , K. Yoshida , C. Noji-Sakikawa , Y. Ando , R. Koishi , and H. Furukawa . 2003 . Protein region important for regulation of lipid metabolism in angiopoi-etin-like 3 (ANGPTL3): ANGPTL3 is cleaved and activated in vivo. J. Biol. Chem. 278 : 41804 – 41809 .
5 . Köster , A. , Y. B. Chao , M. Mosior , A. Ford , P. A. Gonzalez-DeWhitt , J. E. Hale , D. Li , Y. Qiu , C. C. Fraser , D. D. Yang , et al . 2005 . Transgenic angiopoietin-like (angptl)4 overexpression and tar-geted disruption of angptl4 and angptl3: regulation of triglyceride metabolism. Endocrinology . 146 : 4943 – 4950 .
6 . Fujimoto , K. , R. Koishi , T. Shimizugawa , and Y. Ando . 2006 . Angptl3-null mice show low plasma lipid concentrations by en-hanced lipoprotein lipase activity. Exp. Anim. 55 : 27 – 34 .
7 . Liu , J. , H. Afroza , D. J. Rader , and W. Jin . 2010 . Angiopoietin-like protein 3 inhibits lipoprotein lipase activity through enhanc-ing its cleavage by proprotein convertases. J. Biol. Chem. 285 : 27561 – 27570 .
8 . Shimamura , M. , M. Matsuda , H. Yasumo , M. Okazaki , K. Fujimoto , K. Kono , T. Shimizugawa , Y. Ando , R. Koishi , T. Kohama , et al . 2007 . Angiopoietin-like protein3 regulates plasma HDL choles-terol through suppression of endothelial lipase. Arterioscler. Thromb. Vasc. Biol. 27 : 366 – 372 .
9 . Jin , W. , X. Wang , J. S. Millar , T. Quertermous , G. H. Rothblat , J. M. Glick , and D. J. Rader . 2007 . Hepatic proprotein convertases modulate HDL metabolism. Cell Metab. 6 : 129 – 136 .
10 . Willer , C. J. , S. Sanna , A. U. Jackson , A. Scuteri , L. L. Bonnycastle , R. Clarke , S. C. Heath , N. J. Timpson , S. S. Najjar , H. M. Stringham , et al . 2008 . Newly identifi ed loci that infl uence lipid concentrations and risk of coronary artery disease. Nat. Genet. 40 : 161 – 169 .
11 . Kathiresan , S. , C. J. Willer , G. M. Peloso , S. Demissie , K. Musunuru , E. E. Schadt , L. Kaplan , D. Bennett , Y. Li , T. Tanaka , et al . 2009 . Common variants at 30 loci contribute to polygenic dyslipidemia. Nat. Genet. 41 : 56 – 65 .
12 . Kathiresan , S. , O. Melander , C. Guiducci , A. Surti , N. P. Burtt , M. J. Rieder , G. M. Cooper , C. Roos , B. F. Voight , A. S. Havulinna , et al . 2008 . Six new loci associated with blood low-density lipopro-tein cholesterol, high-density lipoprotein cholesterol or triglycer-ides in humans. Nat. Genet. 40 : 189 – 197 .
13 . Hegele , R. A. , M. R. Ban , N. Hsueh , B. A. Kennedy , H. Cao , G. Y. Zou , S. Anand , S. Yusuf , M. W. Huff , and J. Wang . 2009 . A polygenic basis for four classical Fredrickson hyperlipoproteinemia pheno-types that are characterized by hypertriglyceridemia. Hum. Mol. Genet. 18 : 4189 – 4194 .
14 . Teslovich , T. M. , K. Musunuru , A. V. Smith , A. C. Edmondson , I. M. Stylianou , M. Koseki , J. P. Pirruccello , S. Ripatti , D. I. Chasman , C. J. Willer , et al . 2010 . Biological, clinical and population relevance of 95 loci for blood lipids. Nature . 466 : 707 – 713 .
15 . Romeo , S. , W. Yin , J. Kozlitina , L. A. Pennacchio , E. Boerwinkle , H. H. Hobbs , and J. C. Cohen . 2009 . Rare loss-of-function mutations in ANGPTL family members contribute to plasma triglyceride lev-els in humans. J. Clin. Invest. 119 : 70 – 79 .
16 . Robciuc , M. R. , M. Maranghi , A. Lahikainen , D. Rader , A. Bensadoun , K. Oorni , J. Metso , I. Minicocci , E. Ciociola , F. Ceci , et al . 2013 . Angptl3 defi ciency is associated with increased insulin sensitivity, lipoprotein lipase activity, and decreased serum free fatty acids. Arterioscler. Thromb. Vasc. Biol. 33 : 1706 – 1713 . [Erratum. 2013. Arterioscler. Thromb. Vasc. Biol. 33: e124.]
17 . Murphy , A. J. , L. E. Macdonald , S. Stevens , M. Karow , A. T. Dore , K. Pobursky , T. T. Huang , W. T. Poueymirou , L. Esau , M. Meola , et al . 2014 . Mice with megabase humanization of their immunoglobulin
by guest, on April 29, 2019
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2015/05/11/jlr.M054890.DC1Supplemental Material can be found at:

ANGPTL3 antibody reduces plasma lipids 1317
genes generate antibodies as effi ciently as normal mice. Proc. Natl. Acad. Sci. USA . 111 : 5153 – 5158 .
18 . Labrijn , A. F. , A. O. Buijsse , E. T. van den Bremer , A. Y. Verwilligen , W. K. Bleeker , S. J. Thorpe , J. Killestein , C. H. Polman , R. C. Aalberse , J. Schuurman , et al . 2009 . Therapeutic IgG4 antibod-ies engage in Fab-arm exchange with endogenous human IgG4 in vivo. Nat. Biotechnol. 27 : 767 – 771 .
19 . Myszka , D. G. 1999 . Improving biosensor analysis. J. Mol. Recognit. 12 : 279 – 284 .
20 . Valenzuela , D. M. , A. J. Murphy , D. Frendewey , N. W. Gale , A. N. Economides , W. Auerbach , W. T. Poueymirou , N. C. Adams , J. Rojas , J. Yasenchak , et al . 2003 . High-throughput engineering of the mouse genome coupled with high-resolution expression analy-sis. Nat. Biotechnol. 21 : 652 – 659 .
21 . Wang , Y. , F. Quagliarini , V. Gusarova , J. Gromada , D. M. Valenzuela , J. C. Cohen , and H. H. Hobbs . 2013 . Mice lacking ANGPTL8 (Betatrophin) manifest disrupted triglyceride metabolism with-out impaired glucose homeostasis. Proc. Natl. Acad. Sci. USA . 110 : 16109 – 16114 .
22 . Usui , S. , Y. Hara , S. Hosaki , and M. Okazaki . 2002 . A new on-line dual enzymatic method for simultaneous quantifi cation of choles-terol and triglycerides in lipoproteins by HPLC. J. Lipid Res. 43 : 805 – 814 .
23 . Folch , J. , M. Lees , and G. H. Sloane Stanley . 1957 . A simple method for the isolation and purifi cation of total lipides from animal tis-sues. J. Biol. Chem. 226 : 497 – 509 .
24 . Carr , T. P. , C. J. Andresen , and L. L. Rudel . 1993 . Enzymatic deter-mination of triglyceride, free cholesterol, and total cholesterol in tissue lipid extracts. Clin. Biochem. 26 : 39 – 42 .
25 . Han , S. , A. M. Flattery , D. McLaren , R. Raubertas , S. H. Lee , V. Mendoza , R. Rosa , N. Geoghagen , J. M. Castro-Perez , T. P. Roddy , et al . 2012 . Comparison of lipoprotein separation and lipid analysis methodologies for human and cynomolgus monkey plasma sam-ples. J. Cardiovasc. Transl. Res. 5 : 75 – 83 .
26 . Musunuru , K. , J. P. Pirruccello , R. Do , G. M. Peloso , C. Guiducci , C. Sougnez , K. V. Garimella , S. Fisher , J. Abreu , A. J. Barry , et al . 2010 . Exome sequencing, ANGPTL3 mutations, and familial combined hypolipidemia. N. Engl. J. Med. 363 : 2220 – 2227 .
27 . Martín-Campos , J. M. , R. Roig , C. Mayoral , S. Martinez , G. Marti , J. A. Arroyo , J. Julve , and F. Blanco-Vaca . 2012 . Identifi cation of a novel mutation in the ANGPTL3 gene in two families diagnosed of familial hypobetalipoproteinemia without APOB mutation. Clin. Chim. Acta . 413 : 552 – 555 .
28 . Pisciotta , L. , E. Favari , L. Magnolo , S. Simonelli , M. P. Adorni , R. Sallo , T. Fancello , I. Zavaroni , D. Ardigo , F. Bernini , et al . 2012 . Characterization of three kindreds with familial combined
hypolipidemia caused by loss-of-function mutations of ANGPTL3. Circ Cardiovasc Genet . 5 : 42 – 50 .
29 . Noto , D. , A. B. Cefalu , V. Valenti , F. Fayer , E. Pinotti , M. Ditta , R. Spina , G. Vigna , P. Yue , S. Kathiresan , et al . 2012 . Prevalence of ANGPTL3 and APOB gene mutations in subjects with combined hypolipidemia. Arterioscler. Thromb. Vasc. Biol. 32 : 805 – 809 .
30 . Minicocci , I. , A. Montali , M. R. Robciuc , F. Quagliarini , V. Censi , G. Labbadia , C. Gabiati , G. Pigna , M. L. Sepe , F. Pannozzo , et al . 2012 . Mutations in the ANGPTL3 gene and familial combined hy-polipidemia: a clinical and biochemical characterization. J. Clin. Endocrinol. Metab. 97 : E1266 – E1275 .
31 . Minicocci , I. , S. Santini , V. Cantisani , N. Stitziel , S. Kathiresan , J. A. Arroyo , G. Marti , L. Pisciotta , D. Noto , A. B. Cefalu , et al . 2013 . Clinical characteristics and plasma lipids in subjects with famil-ial combined hypolipidemia: a pooled analysis. J. Lipid Res. 54 : 3481 – 3490 .
32 . Lee , E. C. , U. Desai , G. Gololobov , S. Hong , X. Feng , X. C. Yu , J. Gay , N. Wilganowski , C. Gao , L. L. Du , et al . 2009 . Identifi cation of a new functional domain in angiopoietin-like 3 (ANGPTL3) and angiopoietin-like 4 (ANGPTL4) involved in binding and inhibition of lipoprotein lipase (LPL). J. Biol. Chem. 284 : 13735 – 13745 .
33 . Sonnenburg , W. K. , D. Yu , E. C. Lee , W. Xiong , G. Gololobov , B. Key , J. Gay , N. Wilganowski , Y. Hu , S. Zhao , et al . 2009 . GPIHBP1 stabilizes lipoprotein lipase and prevents its inhibition by angiopoi-etin-like 3 and angiopoietin-like 4. J. Lipid Res. 50 : 2421 – 2429 .
34 . Wang, Y., V. Gusarova, S. Banfi , J. Gromada, J. C. Cohen, and H. H. Hobbs. 2015. Inactivation of ANGPTL3 reduces hepatic VLDL-triglyceride secretion. J. Lipid Res. 56: 1296 – 1307 .
35 . Rosenson , R. S. , H. B. Brewer , Jr ., W. S. Davidson , Z. A. Fayad , V. Fuster , J. Goldstein , M. Hellerstein , X. C. Jiang , M. C. Phillips , D. J. Rader , et al . 2012 . Cholesterol effl ux and atheroprotection: ad-vancing the concept of reverse cholesterol transport. Circulation . 125 : 1905 – 1919 .
36 . Voight , B. F. , G. M. Peloso , M. Orho-Melander , R. Frikke-Schmidt , M. Barbalic , M. K. Jensen , G. Hindy , H. Holm , E. L. Ding , T. Johnson , et al . 2012 . Plasma HDL cholesterol and risk of myo-cardial infarction: a mendelian randomisation study. Lancet . 380 : 572 – 580 .
37 . Barter , P. J. , M. Caulfi eld , M. Eriksson , S. M. Grundy , J. J. Kastelein , M. Komajda , J. Lopez-Sendon , L. Mosca , J. C. Tardif , D. D. Waters , et al . 2007 . Effects of torcetrapib in patients at high risk for coro-nary events. N. Engl. J. Med. 357 : 2109 – 2122 .
38 . Boden , W. E. , J. L. Probstfi eld , T. Anderson , B. R. Chaitman , P. Desvignes-Nickens , K. Koprowicz , R. McBride , K. Teo , and W. Weintraub . 2011 . Niacin in patients with low HDL cholesterol levels receiving intensive statin therapy. N. Engl. J. Med. 365 : 2255 – 2267 .
by guest, on April 29, 2019
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2015/05/11/jlr.M054890.DC1Supplemental Material can be found at: