announcements 1. first lab report deadline extended by one week: x-linked cross lab report due 11/...
TRANSCRIPT

Announcements
1. First lab report deadline extended by one week: X-linked cross lab report due 11/ 5,6.
2. Get lab overview for PCR lab - quiz in lab this week
3. If your transformation did not work, look at plates in lab refrigerator, both groups “A” and “B”; make counts from these plates. Lab assignment due 10/29, 30 (extra week).
4. Group B presentations in lab 10/29, 30 - get sources approved
5. Monk papers will be handed back after lecture in the lab - avg. 67/75 = 89%.
6. Currently: reading chapters 12, 13 this week.

Review of Last Lecture
I. Bacterial DNA replication - semiconservative
II. Many complex issues to resolve during DNA replication- DNA helix must be unwound, RNA primer needed- Synthesis is both continuous and discontinuous- Proofreading is critical

Outline of Lecture 22
I. Eukaryotic DNA replication is more complex
II. The “end” problem and telomerase
III. DNA Recombination
IV. The Genetic Code
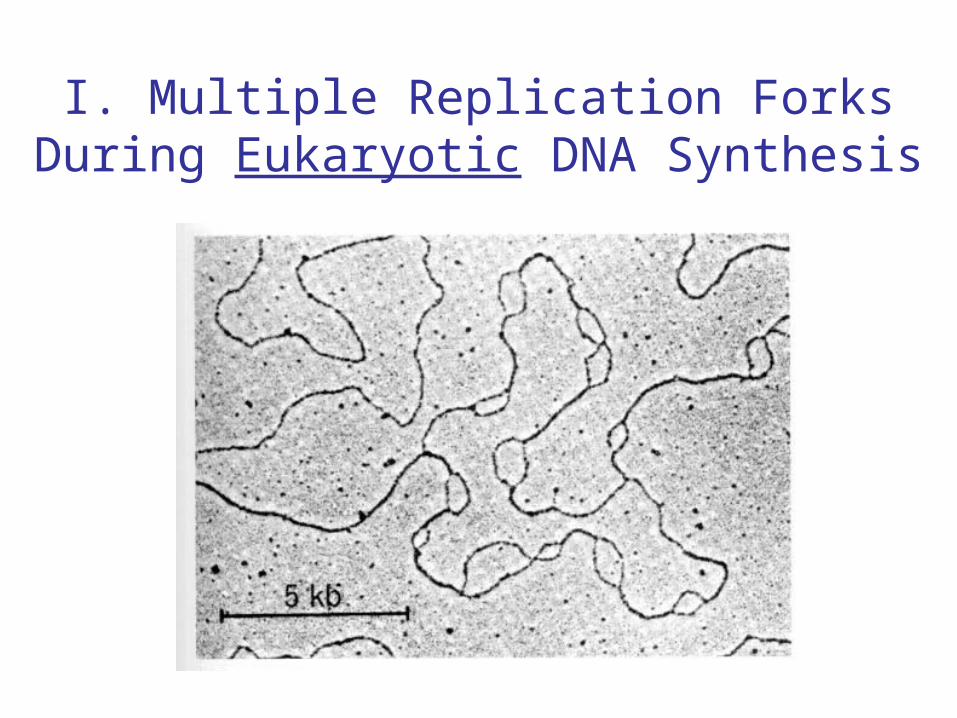
I. Multiple Replication Forks During Eukaryotic DNA Synthesis

When, during the cell cycle, can new replication origins be formed?
G1 S G2Pre-replication replication post replication
ARSs complex DNA synthesis DNA synthesisInto ORCs completed
Pre-RCs can form No new pre-RCs No new pre-RCs
ARS = autonomously replicating sequence = originORC = origin recognition complexPre-RC = pre-replication complex

Eukaryotic DNA PolymerasesEnzyme Location Function• Pol (alpha) Nucleus DNA replication
– includes RNA primase activity, starts DNA strand
• Pol (gamma) Nucleus DNA replication– replaces Pol to extend DNA strand, proofreads
• Pol (epsilon) Nucleus DNA replication– similar to Pol , shown to be required by yeast mutants
• Pol (beta) Nucleus DNA repair• Pol (zeta) Nucleus DNA repair• Pol (gamma) Mitochondria DNA replication
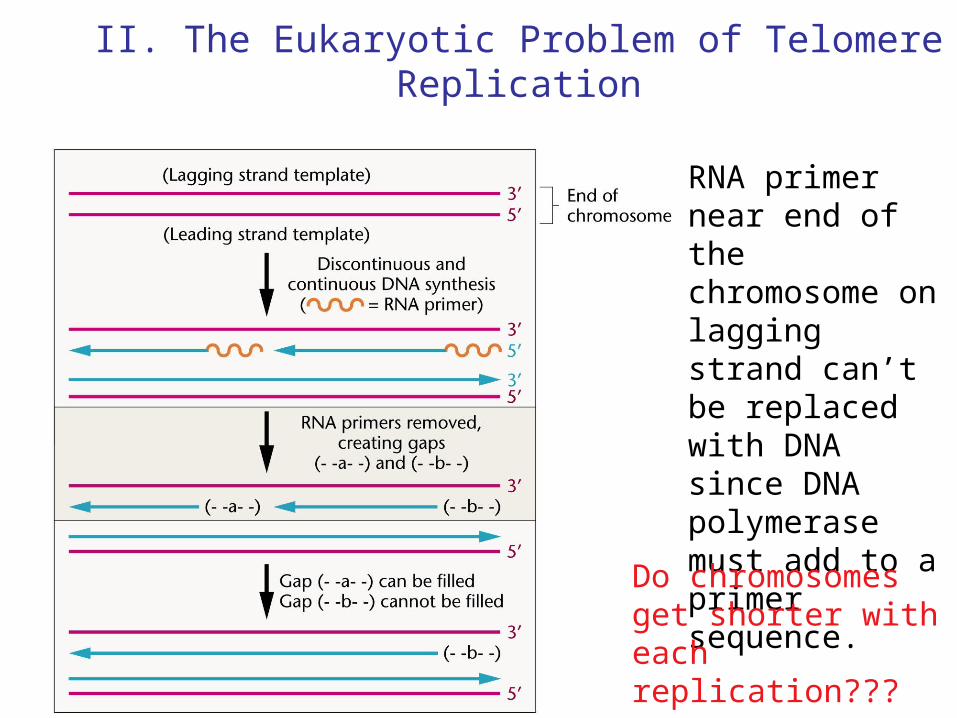
II. The Eukaryotic Problem of Telomere Replication
RNA primer near end of the chromosome on lagging strand can’t be replaced with DNA since DNA polymerase must add to a primer sequence.
Do chromosomes get shorter with each replication???
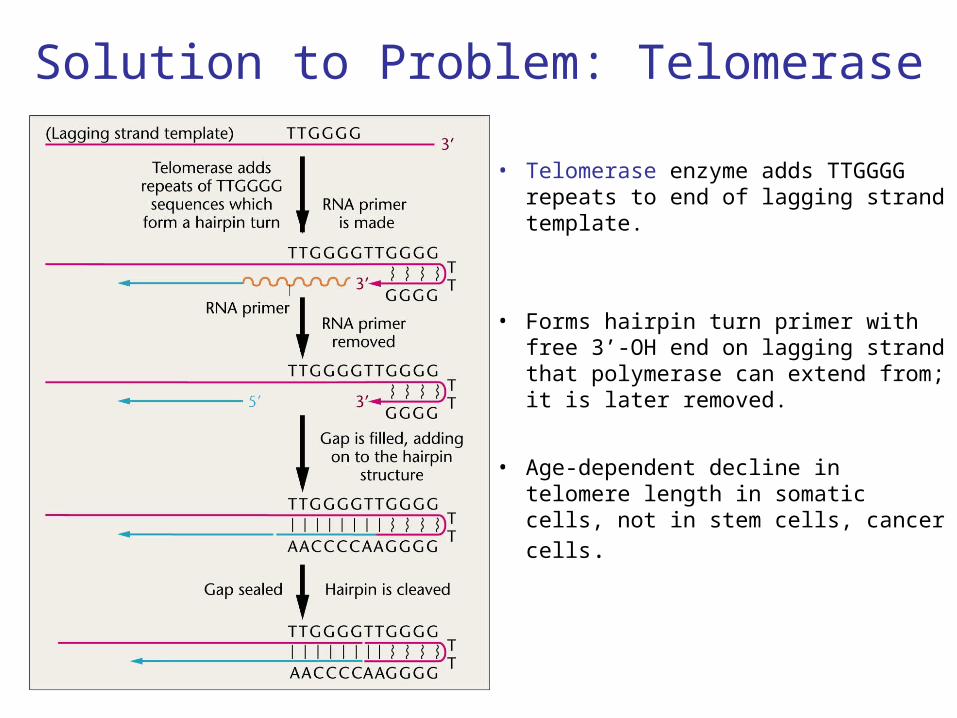
Solution to Problem: Telomerase
• Telomerase enzyme adds TTGGGG repeats to end of lagging strand template.
• Forms hairpin turn primer with free 3’-OH end on lagging strand that polymerase can extend from; it is later removed.
• Age-dependent decline in telomere length in somatic cells, not in stem
cells, cancer cells.

III. Recombination at the Molecular Level
• Breakage and joining also directed by enzymes.
• Homologous recombination occurs during synapsis in meiosis I, general recombination in bacteria, and viral genetic exchange.
• Molecular mechanism proposed by Holliday and Whitehouse (1964).
• Depends on complementary base pairing.
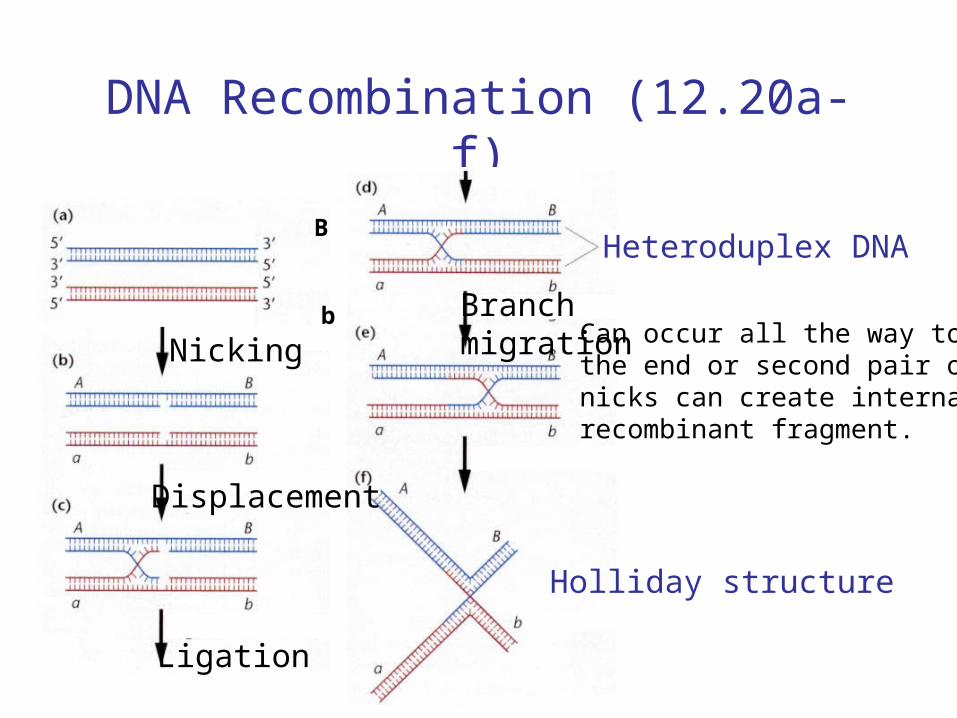
DNA Recombination (12.20a-f)
Heteroduplex DNA
Holliday structure
Can occur all the way tothe end or second pair ofnicks can create internalrecombinant fragment.
Nicking
Displacement
Ligation
Branchmigration
A B
a b
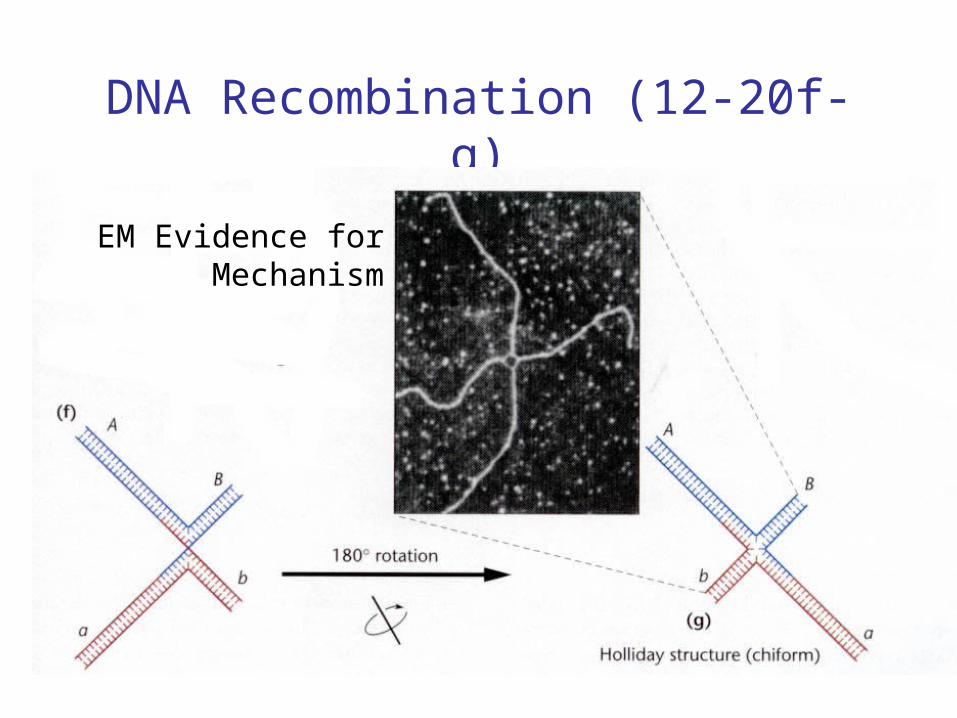
DNA Recombination (12-20f-g)
EM Evidence forMechanism
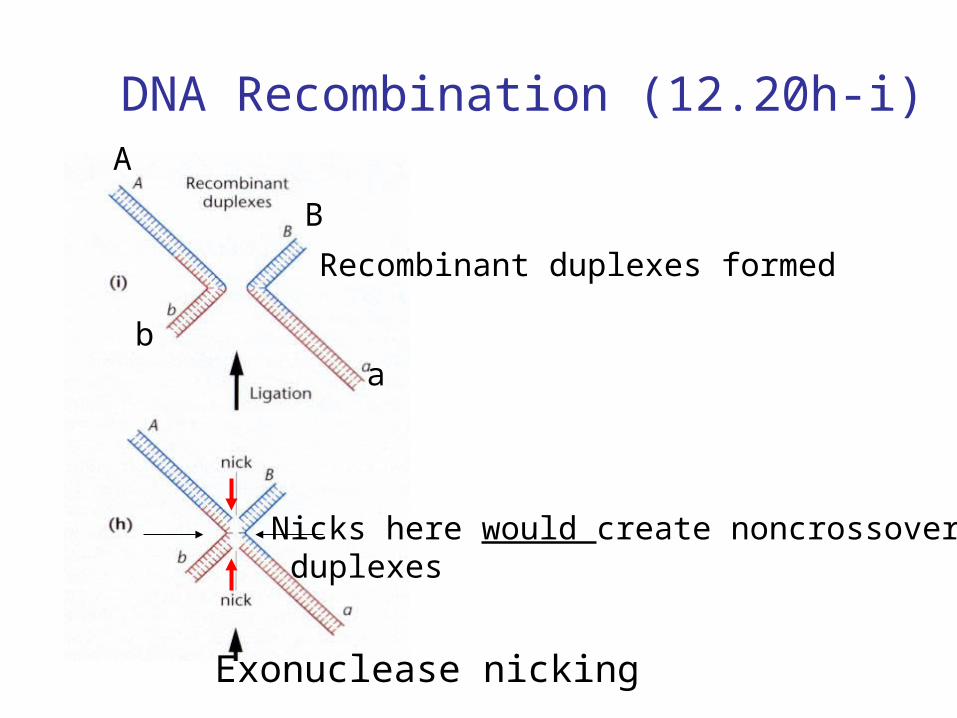
DNA Recombination (12.20h-i)
Exonuclease nicking
Nicks here would create noncrossover duplexes
Recombinant duplexes formed
A
b
B
a

IV. Early Evidence for the Genetic Code
• 1940’s: Beadle and Tatum noted correlation between gene mutation and nonfunctional enzyme
• First direct evidence: sickle-cell hemoglobin– single nucleotide change > change in amino acid
• 1961: Jacob and Monod proposed that mRNA is an unstable intermediate between DNA and protein
• How could four letters (A, T, G, C) spell out 20 words (the amino acids)?

Theoretical Evidence
• Sidney Brenner (early 1960’s) argued that code must be triplet theoretically.
• If a two letter code, how many amino acid “words” could be made from A, U, G, C? 42 = 16
• If a three letter code, how many “words” could be made? 43 = 64, more than enough for the 20 amino acids.

Genetic Evidence: Frameshift Mutations
• 1961: Francis Crick, Barnett, Brenner, and Watts-Tobin
• Created insertion and deletion mutants in cistron B of rII locus of phage T4– A cistron codes for a single polypeptide chain within a gene
• Proflavin (a DNA dye) was used as a mutagen.
• Proflavin caused insertion or deletion of one or more nucleotides in the cistron, usually causing a frameshift of the putative genetic code.
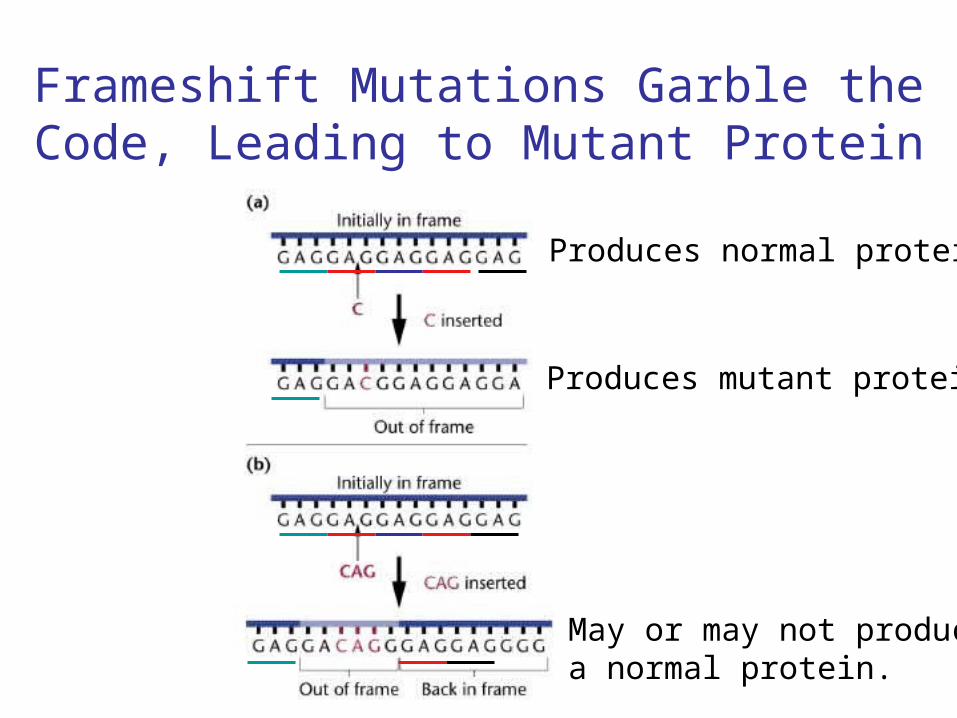
Frameshift Mutations Garble the Code, Leading to Mutant Protein
Produces mutant protein
Produces normal protein
May or may not producea normal protein.

5’ UGC GAA AAC ACA AGA GCA UUA U 3’ WT C E N T R A L
Functional Site 5’ UGC GAA AAC GAC AAG AGC AUU AU 3’ + MUT C E N N K S I
A5’ UGC GAA AACCAA GAG CAU UAU 3’ - MUT C E N Q Q H Y
Wildtype, Single Insertion and Deletion

Insertion/Deletion, Triple Deletion, Triple Insertion
A5’ UGC GAA AAC GCA AGA GCA UUA U 3’ +/- WT C E N A R A L
GAA5’ UGC AAC ACA AGA GCA UUA U 3’ -/-/- WT C N T R A L
5’ UGC GAA GAA AAC ACA AGA GCA UUA U 3’ +/+/+ WT C E E N T R A L

Biochemical Evidence
• 1961: Nirenberg, Matthaei used synthetic mRNAs and an in vitro translation system to decipher the code.
• Polynucleotide Phosphorylase enzyme links NTPs to make RNA without a template
• Homopolymers:– poly(U) codes for Phe-Phe-Phe-Phe-…– poly(A) codes for Lys-Lys-Lys-Lys-…– poly(C) codes for Pro-Pro-Pro-Pro-...

Repeating Copolymers
• Khorana, early 1960’s
• UGUGUGUGUGUGUGUGU...– Cys-Val-Cys-Val-Cys-Val-...– Therefore GUG or UGU codes for either Cys or Val
• UUCUUCUUCUUCUUC…– Phe-Phe-Phe-Phe-... or– Ser-Ser-Ser-Ser-… or– Leu-Leu-Leu-Leu-...
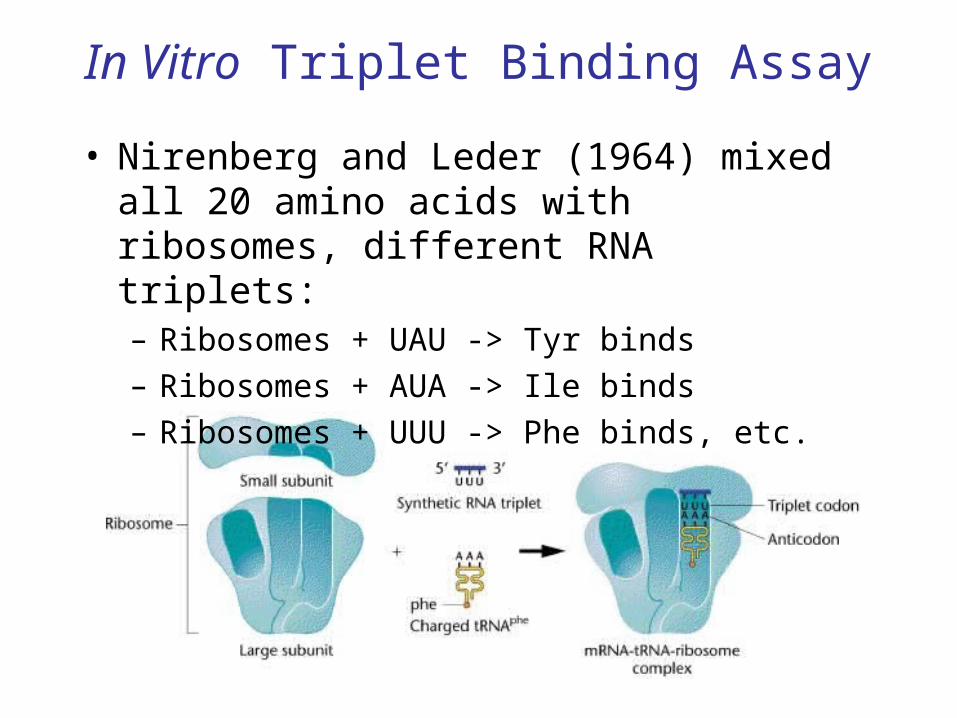
In Vitro Triplet Binding Assay
• Nirenberg and Leder (1964) mixed all 20 amino acids with ribosomes, different RNA triplets:– Ribosomes + UAU -> Tyr binds– Ribosomes + AUA -> Ile binds– Ribosomes + UUU -> Phe binds, etc.
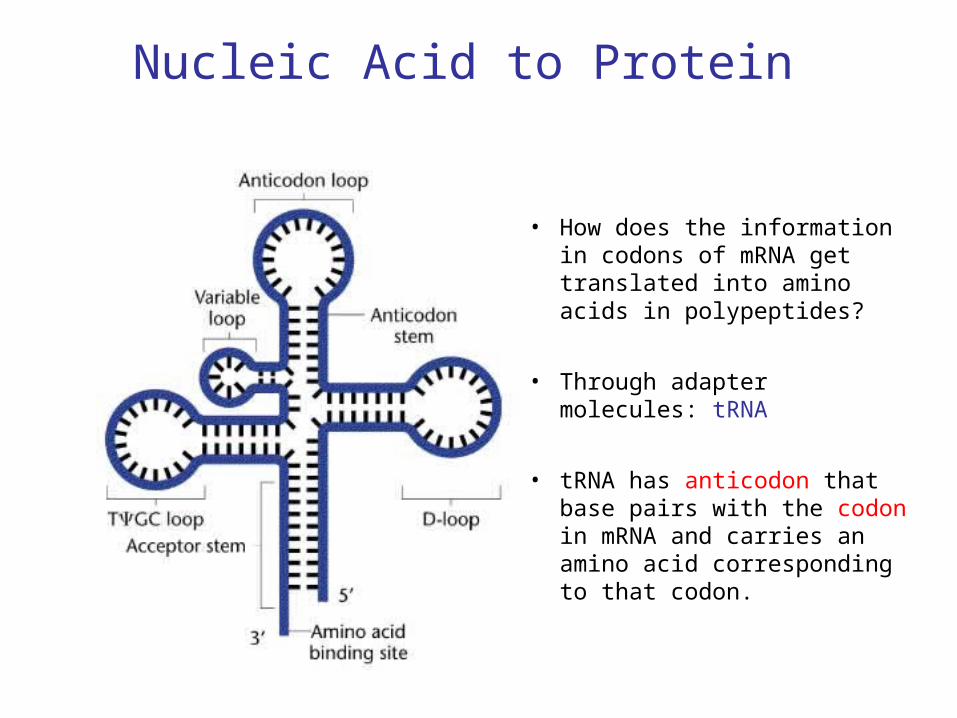
Nucleic Acid to Protein
• How does the information in codons of mRNA get translated into amino acids in polypeptides?
• Through adapter molecules: tRNA
• tRNA has anticodon that base pairs with the codon in mRNA and carries an amino acid corresponding to that codon.
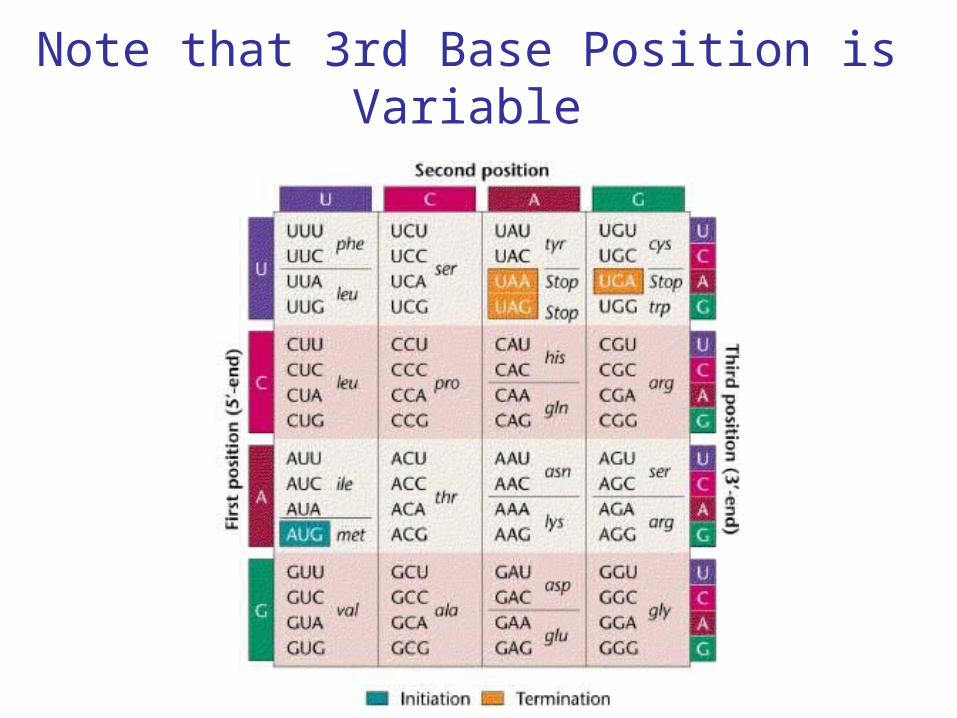
Note that 3rd Base Position is Variable
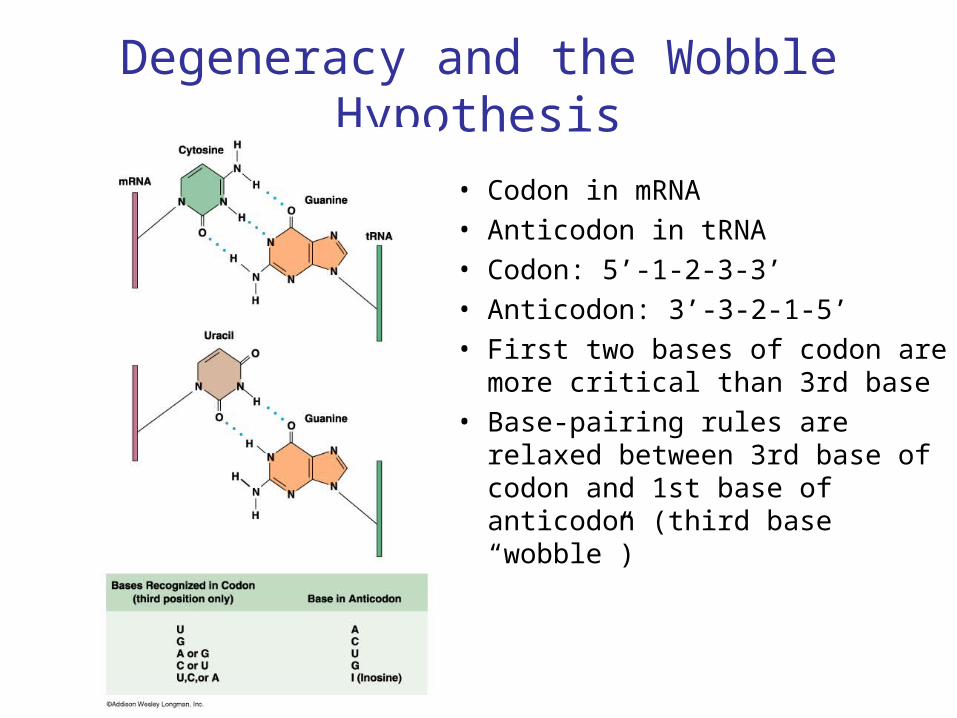
Degeneracy and the Wobble Hypothesis
• Codon in mRNA• Anticodon in tRNA• Codon: 5’-1-2-3-3’• Anticodon: 3’-3-2-1-5’• First two bases of codon are
more critical than 3rd base• Base-pairing rules are relaxed
between 3rd base of codon and 1st base of anticodon (third base “wobble”)
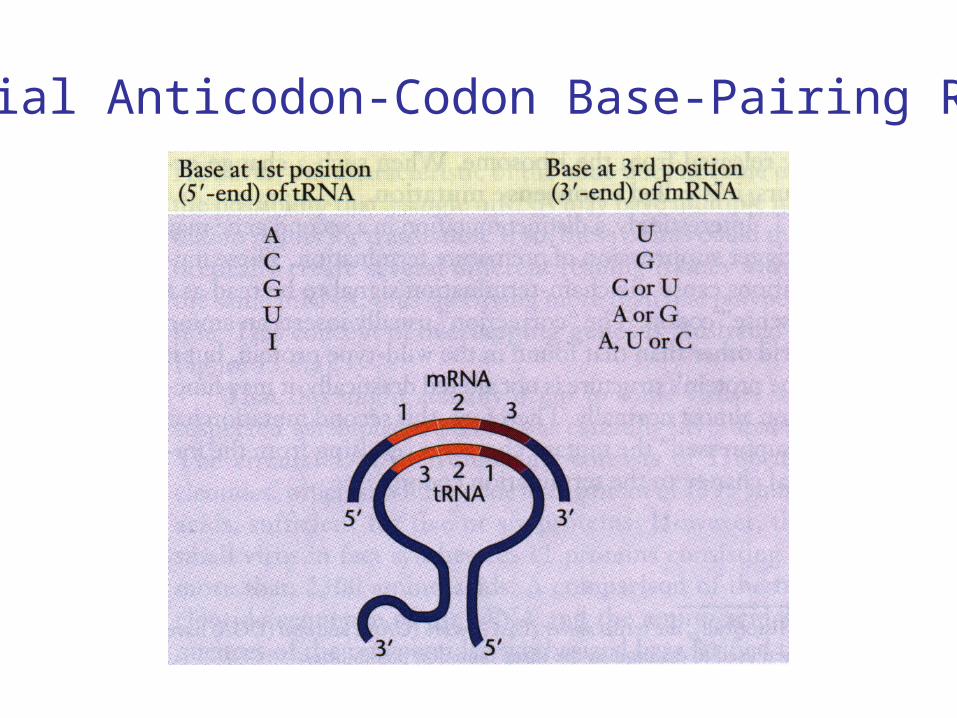
Special Anticodon-Codon Base-Pairing Rules