antibodies in cancer - biomarkers and immune activation
DESCRIPTION
Cancer and Antibodies with BiomarketsTRANSCRIPT

577
Review
www.expert-reviews.com ISSN 1478-9450© 2011 Expert Reviews Ltd10.1586/EPR.11.48
IntroductionCurrent cancer biomarkers & the role of autoantibodies for cancer detection The development of blood-based biomarkers for the early detection and prognosis of cancer remains an elusive target. Multiple labora tories have used genomic and proteomic discovery techniques to identify molecular changes in tumor-specific methylation [1], mRNAs [2], glycosylation [3], proteins [4], autoantibodies (AAbs) [5,6] and circulating tumor cells [7] related to cancer. However, despite these efforts, the vast majority of newly discovered protein bio-markers for the early detection of cancer have not progressed beyond blinded Phase III validation studies into clinical practice [8]. There are sev-eral reasons for this, including natural variation of the human proteome, subtle differences in sample collection techniques that introduce bias and limited resources to perform early biomarker discovery projects in the prediagnostic samples that represent the very population that would ultimately be screened.
Despite these limitations, the ideal protein biomarker would reflect true biologic hetero-geneity in tumor protein expression. The bio-marker may be a differentially expressed tumor antigen, or the corresponding AAb with affinity to the tumor antigen. AAb may be detectable both at early onset of the disease and in higher
concentrations as compared with the tumor anti-gen itself, representing an in vivo amplification of signal detection. Many of these antibodies are being developed as diagnostic biomarkers for early detection [9,10] and have been extensively reviewed. In comparison, their utility as prog-nostic biomarkers for disease outcome is less well understood but critical to understanding their function in the immune surveillance of cancer. For clinical utility, the association of these mark-ers must be independent of established clinico-pathologic criteria, such as tumor stage, grade, subtype, mutation status or treatment.
The protein structure of immunogenic B-cell epitopesComputer algorithms are available for predict-ing B-cell immunogenicity [11], but in general the protein structural features that induce AAbs in cancer patients (or healthy individuals) are poorly understood. One explanation is that AAbs are generated from B cells that have escaped the self-tolerance check points of the immune sys-tem [12,13]. In addition, antigen mutation [14], overexpression [15], altered antigen folding [16], aberrant degradation [17] and aberrant glycosy-lation [18] are all associated with AAb formation. The epitopes of the tumor antigens can be linear stretches of amino acids, conformational epitopes of discontinuous amino acids that develop 3D conformations, or cryptic epitopes, embedded
Kerstin Järås1 and Karen Anderson†2
1Barnett Institute of Chemical and Biological Analysis, Northeastern University, Boston, MA 02115, USA 2Biodesign Institute, Arizona State University, PO Box 876401, Tempe, AZ 85287, USA †Author for correspondence:Tel.: +1 480 965 6982 Fax: +1 480 965 3051 [email protected]
The development of proteomic technologies that display a wide variety of antigenic structures has led to the identification of autoantibodies to cancer-derived tumor antigens. These autoantibodies have been detected in sera from patients with multiple cancer types, and are being evaluated as biomarkers for early cancer detection. It is not known whether these antibodies also contribute to active immune surveillance or even tumorigenicity of developing tumors. Here, we review which tumor antigen-specific antibodies are prognostic biomarkers of cancer outcome, and emerging proteomic methods for the isolation and cloning of these antibodies for potential molecular diagnostics and therapeutics.
Keywords: autoantibodies • cancer • clinical outcome • function • human monoclonal antibodies • prognosis • prognostic • recurrence • survival • therapeutic antibodies • tumor antigens
Autoantibodies in cancer: prognostic biomarkers and immune activationExpert Rev. Proteomics 8(5), 577–589 (2011)
For reprint orders, please contact [email protected]

Expert Rev. Proteomics 8(5), (2011)578
Review
within the protein [6]. As proteomic tools emerge for proteome-wide immune monitoring, improved prediction of likely AAb epitopes will enhance our ability to identify these biomarkers.
Potential of prognostic AAbsAs biomarkers, AAbs are highly specific, easily purified from serum and easily identified with high-quality secondary reagents. Many AAbs have been correlated with clinical prognosis [19–34]. Because these AAbs are indirect measurements of changes in tumor antigenic structure and/or expression, they could represent nonfunctional surrogate markers of antigen overexpression; active B-cell immunity, which functions to limit both tumor growth and dissemination; or active B-cell immunity that is protumori-genic. Here, we review the proteomic technologies that are used for the detection of AAbs and their corresponding tumor anti-gens, the evidence of the association of certain AAbs with cancer prognosis and methods to determine the mechanisms of AAb function. In addition, AAbs associated with improved prognosis may represent partially effective cancer immunosurveillance, and may be candidates for immunotherapy development.
Proteomic technologies for tumor AAb discovery & validationTechnologies used to detect AAbs to tumor antigens can be divided into methods for discovery of novel tumor antigens generating AAb responses and methods for validation of selected tumor antigens. The discovery methods include serological ana lysis of tumor antigens by recombinant cDNA expression cloning (SEREX) [35], sero logical proteome analysis (SERPA) [36], also called PROTEOMEX by Seliger et al. [37,38], multiple affinity protein profiling (MAPPing) [39] and high-density protein microarrays [40–42]. These approaches are labor and cost intensive, and are used to screen large numbers of antigens with small numbers of sera.
Once potential tumor antigens are discovered, confirmation of their association with clinical parameters requires highly reproduc-ible, low-cost assays for validation using large numbers of sera. These validation assays include recombinant protein ELISA [22,30,43–45], protein microarrays [46–50] and bead-based immunoassays [51].
SEREXSerological ana lysis of tumor antigens by recombinant cDNA expression cloning, initially developed in the Pfreundschuh and Old laboratories [35,52], uses phage-expression cDNA libraries con-structed from tumor tissue or cell lines to display tumor antigens. Expression products are transferred to membranes and probed with patient sera. SEREX has been used to identify thousands of tumor antigens in multiple tumor types [53–56]. SEREX has limitations, including restricted detection to denatured epitopes and a bias toward gene products with high RNA content. SEREX has been combined with solution-based phage-display technol-ogy [57] or printed as phage-displayed protein microarrays [42]. Solution-based phage display uses serial rounds of biopanning to enrich for antigens of interest [58,59]. Yeast display systems, which can incorporate eukaryotic post-translational modifications, have also been investigated [60–62].
SERPA & reversed-phase microarraysAn alternative discovery technology is SERPA [36], based on a classical proteomics strategy using 2D gel electrophoresis fol-lowed by differential immunoblotting with patient and control sera. In-gel digestion is used to identify tumor antigens by mass spectroscopy analysis [36,43,63–66]. In contrast to phage- and yeast-based display methods, the SERPA technology allows for the identification of immunogenic post-translational modifications and isoforms. Limitations of the technique include preferential detection of highly abundant proteins, limited identification of transmembrane tumor antigens and denaturation of proteins by immunoblotting.
Fractions from chromatographically separated tumor cell lysates can also be arrayed onto slides. These reversed-phase microarrays are then probed with sera to detect AAbs [67–69]. As with SERPA, the arrayed fraction contains a complex mixture of proteins, requiring further separation techniques and mass spectrometry to identify the tumor antigens [68].
MAPPMultiple affinity protein profile is a 2D immunoaffinity separation of cell lysate and subsequent MS/MS analysis for the identification of immunogenic tumor antigens [39]. The first chromato graphic column contains IgG from healthy control serum, to deplete the cell lysate from antigens that will bind to antibodies found in nor-mal sera. The second immunoaffinity column contains the IgG of patient serum. Several columns can be used in parallel, each corresponding to different patients. Tumor antigens captured in the second dimension are eluted and digested for identification by nano-liquid chromatography MS/MS. MAPPing maintains tumor antigen in solution, allowing for the potential identifica-tion of structural epitopes. However, immunoprecipitation using affinity columns limits detection of the tumor antigen within complex protein solutions, such as cell lysates. The identification of tumor antigen is often restricted to antibody interactions with low dissociation rate constant (k
off).
Protein microarraysProtein microarrays constitute a quick and convenient techno-logy for characterizing humoral immune response in serum in a high-throughput manner. Spotted protein microarrays are now commercially available, with proteins generally expressed using phage [42] or baculovirus [70]. Tagged proteins can be expressed and captured in situ using programmable protein expression arrays (NAPPA) [71–73]. NAPPA arrays circumvent the challenges of pro-tein purification and stability during storage, but require rigorous standardization for reproducibility of protein expression.
Validation techniquesThe primary challenge with tumor antigen discovery using pro-teomic techniques is that validation of serum biomarkers requires highly reproducible, quantitative and cost-effective assays for screening large numbers of sera on small numbers of antigens. To evaluate the prognostic effect of a specific humoral response, immunoassays, such as ELISA and protein microarrays, are used
Järås & Anderson

www.expert-reviews.com 579
Review
to measure the serum AAb levels in patients with known clinical history. These AAb levels are then correlated with clinical out-come. Protein microarrays have been used for validation of AAb from cancer patients and tumor-bearing mice [46–50], but single-antigen ELISA is more frequently used to validate antibody detec-tion in both cancer and autoimmunity [22,30,43–45]. Protein micro-arrays are excellent discovery tools, but currently lack the speed, precision, quantitation and cost efficiency needed for expanded clinical use. Bead-based arrays, such as Luminex™ arrays, are highly versatile arrays that allow for limited multi plexing, con-servation of limited patient sera and high -throughput serologic screening. These are increasingly used as high -throughput assays to detect viral-specific and tumor antigen-specific antibodies in sera [51,74,75]. Emerging microfluidic technologies have the poten-tial as cost-effective, point-of-care devices that could be adapted for AAb detection [76]. Overall, the lack of high-precision clinical-grade assays, which require reproducible (and expensive) gen-eration of tumor antigen and internal standards, has limited the application of AAb biomarkers to clinical practice.
A general limitation of the application of AAb biomarkers to clinical use has been the overall low sensitivity of detection of these AAbs in cancer patient sera. This could be due to true bio-logic hetero geneity (i.e., only a few tumors overexpress the tumor antigen) or technical limitations of detection of specific AAbs. Validation assays for AAb can be optimized to markedly improve the sensitivity of detection while maintaining specificity for the antigen [77]. Such optimizations include micro- and nano-porous silicon surfaces for increased capture molecule deposition, and sig-nal amplification with europium-incorporated nano particles, which allows for quantification over a wide concentration range [78,79]. Other signal-enhancement methods include catalyzed signal ampli-fication with colorimetric readout [80], proximity ligation assays [81] and rolling circle amplification [82]. Solution-based ELISAs, such as luciferase immunoprecipitation, may also increase the sensitivity of detection [83].
Ideal protein biomarkers reflect consistent, biologically rel-evant changes in the tumor microenvironment. AAb validation includes determination of the tumor antigen overexpression in primary tumor tissue by immunohistochemistry, real-time PCR or immuno blot [84]. In general, the detection of AAb correlates with tumor antigen overexpression. Snyder and coworkers identi-fied potential ovarian cancer AAbs, and confirmed tumor antigen expression using tumor tissue microarrays [70]. In this approach, the AAbs were used to identify novel aberrantly expressed proteins. There are several other examples of validation of tumor antigen expression associated with humoral immune response. Mutations in the tumor protein p53 result in intracellular accumu lation of p53 protein and are strongly associated with AAb generation [85–88]. In a study by Goodell et al., 82% of breast cancers that strongly over-expressed the HER2/neu protein had AAb specific to HER2/neu in their sera as compared with none of the cases with weak expres-sion [89]. For the tumor antigen NY-ESO-1, up to 83% of patients with antigen-expressing tumors also had AAb to NY-ESO-1, while patients with tumors lacking NY-ESO-1 expression had low (17%) [90] or absent [91,92] NY-ESO-1 AAb in their sera.
However, unknown factors impact AAb production. First, strong protein over expression does not invariably induce AAb; strong over expression of the tumor antigens p16 and melan A are not associated with AAb detection [92,93]. Conversely, AAbs have been detected in patients with antigen-negative tumors [90,94]. This may reflect loss of antigen expression due to immunoselection (selection pressure by the immune system towards tumor antigen loss) of the tumor cells or tumor heterogeneity of antigen expression. In addi-tion, there are examples of AAb present in healthy individuals to epithelial mucin 1 (MUC1) [95], a glycosylated protein that is both a serum tumor marker and a cellular tumor antigen. Therefore, tumor-specific antigen overexpression, while ideal, is not a scientific prerequisite for the development of an AAb biomarker.
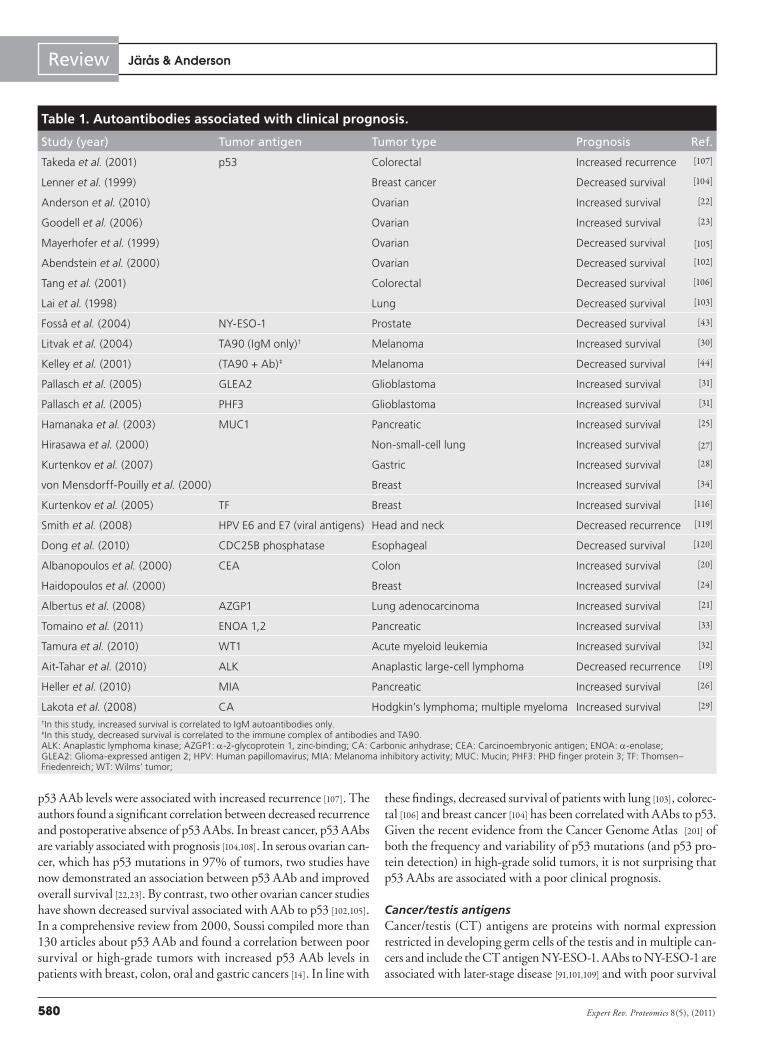
AAbs associated with clinical prognosisPrognostic biomarkers that predict cancer recurrence and/or sur-vival are crucial for targeting therapies to high-risk populations. Clinical parameters, such as tumor–node–metastasis staging and tumor grade/differ entiation, are routinely used for risk stratifica-tion in clinical practice. Emerging molecular prognostic markers that measure gene expression within primary tumor specimens have broad applications across multiple cancer types, and have revealed fundamental differences in tumor biology between can-cers with similar histologies [96]. While tumor antigen-specific AAbs have been detected prior to cancer development [10,97–100], few have been assessed as prognostic biomarkers of cancer recurrence and/or survival.
There are several challenges associated with the evaluation of prognostic AAb biomarkers. First, clinical samples must be derived prior to treatment from well-defined populations who received uniform therapy with long-term clinical follow-up [91,101]. Second, prognostic significance must be independent of recog-nized clinical variables, such as stage, grade and molecular sub-type. Most importantly, unlike target antigens, AAbs are indi-rect measures of overexpression or specific mutation of the target antigen and depend on host variability of immune competence, such as polymorphisms in FcG receptors. At this time, it is not known if any prognostic AAbs are simply highly sensitive serum bio markers of tumor antigen expression/mutation (such as specific p53 mutations) or reflect underlying functional immuno reactivity against the tumor. A summary of published studies of the asso-ciation of AAbs with clinical prognosis in cancer is shown in Table 1. The most frequently studied tumor antigens from Table 1 are discussed in the following section.
Tumor-suppressor antigen p53One of the earliest identified AAbs is specific for the tumor sup-pressor antigen p53, which is mutated in many cancers. Anti-p53 antibodies in sera of workers occupationally exposed to vinyl chlo-ride have been detected prior to the clinical diagnosis of angio-sarcoma of the liver [10] and in patients with chronic obstructive pulmonary disease prior to lung cancer diagnosis [99].
Several studies have addressed the correlation between p53 AAb and survival [22,23,102–106]. In a study of 40 patients who had undergone operations for colorectal cancer, elevated postoperative
Autoantibodies in cancer: prognostic biomarkers & immune activation
Expert Rev. Proteomics 8(5), (2011)580
Review
p53 AAb levels were associated with increased recurrence [107]. The authors found a significant correlation between decreased recurrence and postoperative absence of p53 AAbs. In breast cancer, p53 AAbs are variably associated with prognosis [104,108]. In serous ovarian can-cer, which has p53 mutations in 97% of tumors, two studies have now demonstrated an association between p53 AAb and improved overall survival [22,23]. By contrast, two other ovarian cancer studies have shown decreased survival associated with AAb to p53 [102,105]. In a comprehensive review from 2000, Soussi compiled more than 130 articles about p53 AAb and found a correlation between poor survival or high-grade tumors with increased p53 AAb levels in patients with breast, colon, oral and gastric cancers [14]. In line with
these findings, decreased survival of patients with lung [103], colorec-tal [106] and breast cancer [104] has been correlated with AAbs to p53. Given the recent evidence from the Cancer Genome Atlas [201] of both the frequency and variability of p53 mutations (and p53 pro-tein detection) in high-grade solid tumors, it is not surprising that p53 AAbs are associated with a poor clinical prognosis.
Cancer/testis antigensCancer/testis (CT) antigens are proteins with normal expression restricted in developing germ cells of the testis and in multiple can-cers and include the CT antigen NY-ESO-1. AAbs to NY-ESO-1 are associated with later-stage disease [91,101,109] and with poor survival
Table 1. Autoantibodies associated with clinical prognosis.
Study (year) Tumor antigen Tumor type Prognosis Ref.
Takeda et al. (2001) p53 Colorectal Increased recurrence [107]
Lenner et al. (1999) Breast cancer Decreased survival [104]
Anderson et al. (2010) Ovarian Increased survival [22]
Goodell et al. (2006) Ovarian Increased survival [23]
Mayerhofer et al. (1999) Ovarian Decreased survival [105]
Abendstein et al. (2000) Ovarian Decreased survival [102]
Tang et al. (2001) Colorectal Decreased survival [106]
Lai et al. (1998) Lung Decreased survival [103]
Fosså et al. (2004) NY-ESO-1 Prostate Decreased survival [43]
Litvak et al. (2004) TA90 (IgM only)† Melanoma Increased survival [30]
Kelley et al. (2001) (TA90 + Ab)‡ Melanoma Decreased survival [44]
Pallasch et al. (2005) GLEA2 Glioblastoma Increased survival [31]
Pallasch et al. (2005) PHF3 Glioblastoma Increased survival [31]
Hamanaka et al. (2003) MUC1 Pancreatic Increased survival [25]
Hirasawa et al. (2000) Non-small-cell lung Increased survival [27]
Kurtenkov et al. (2007) Gastric Increased survival [28]
von Mensdorff-Pouilly et al. (2000) Breast Increased survival [34]
Kurtenkov et al. (2005) TF Breast Increased survival [116]
Smith et al. (2008) HPV E6 and E7 (viral antigens) Head and neck Decreased recurrence [119]
Dong et al. (2010) CDC25B phosphatase Esophageal Decreased survival [120]
Albanopoulos et al. (2000) CEA Colon Increased survival [20]
Haidopoulos et al. (2000) Breast Increased survival [24]
Albertus et al. (2008) AZGP1 Lung adenocarcinoma Increased survival [21]
Tomaino et al. (2011) ENOA 1,2 Pancreatic Increased survival [33]
Tamura et al. (2010) WT1 Acute myeloid leukemia Increased survival [32]
Ait-Tahar et al. (2010) ALK Anaplastic large-cell lymphoma Decreased recurrence [19]
Heller et al. (2010) MIA Pancreatic Increased survival [26]
Lakota et al. (2008) CA Hodgkin’s lymphoma; multiple myeloma Increased survival [29]
†In this study, increased survival is correlated to IgM autoantibodies only.‡In this study, decreased survival is correlated to the immune complex of antibodies and TA90.ALK: Anaplastic lymphoma kinase; AZGP1: a-2-glycoprotein 1, zinc-binding; CA: Carbonic anhydrase; CEA: Carcinoembryonic antigen; ENOA: a-enolase; GLEA2: Glioma-expressed antigen 2; HPV: Human papillomavirus; MIA: Melanoma inhibitory activity; MUC: Mucin; PHF3: PHD finger protein 3; TF: Thomsen–Friedenreich; WT: Wilms’ tumor;
Järås & Anderson

www.expert-reviews.com 581
Review
in hormone-refractory prostate cancer [43]. A more recently identi-fied member of the CT antigen family is sperm-associated antigen 9, which is associated with high risk of breast cancer recurrence [110].
Cell surface antigensThe tumor antigen HER-2/neu is a transmembrane tyrosine kinase and is the target of the therapeutic antibody trastu-zumab, used for the treatment of HER2/neu-overexpressing breast cancers [111]. AAbs specific for HER2/neu have been reported [95,112,113]. HER2/neu-targed vaccination can induce HER2-specific AAbs that modulate the HER2/neu signaling cascade [114] and are associated with improved survival [111,115]. Whether naturally occurring HER2/neu AAbs develop similar functional effects on the HER2/neu molecule is not known.
Mucin 1 is a high-molecular-weight transmembrane glyco-protein, overexpressed in multiple epithelial cancers, including pancreatic, ovarian, lung and breast cancers. AAbs to MUC1 have been detected in multiple MUC1-positive cancers [25,27,28,34]. Studies have demonstrated an association between anti-MUC1 AAbs and improved survival in pancreatic [25], gastric [28], breast [34] and non-small-cell lung cancers [27]. AAbs to the MUC1-related Thomsen–Friedenreich epitope are associated with improved survival of stage II breast cancer patients [116].
Viral antigensViral proteins induce viral-specific antibodies that may mimic the effects of AAbs. Human papillomavirus (HPV) is associated with cervical cancer and the majority of oropharyngeal cancers [117]. HPV-associated oropharyngeal cancers have an improved clinical prognosis compared with HPV-negative cancers when matched for stage [118,119]. In a study of 156 serum samples from patients with HPV16+ head and neck cancer, patients with antibodies against the HPV16 viral proteins E6 and/or E7 had improved clinical prognosis (p = 0.003) [119].
Overall, the results from Table 1 correlating AAbs and clinical prognosis are mixed. Certain AAbs, such those as for TA90 [44] and CDC25B [120], are associated with an unfavorable prognosis, but others are associated with an improved prognosis. It is likely that as our methods for measuring the genetic, epigenetic and protein over-expression and intratumoral variability early in cancer development improve, clear correlations with clinical prognosis will be detected.
Functional effects of AAbs in cancerIn parallel to biomarker studies assessing the association between AAbs and clinical prognosis, significant progress has been made in our understanding of the role of the antibody-producing B cells in cancer progression. Animal models have demonstrated that combinations of cellular, antibody, cytokine and innate immunity can function paradoxically both in tumor control via immunosurveillance, as well as tumor progression via inflamma-tion and tissue invasion [121]. In humans, the presence of tumor-infiltrating lymphocytes correlates with improved survival for multiple types of cancers, including follicular lymphoma [122], and colorectal [123,124], breast [125] and ovarian cancers [126]. Further characterization of immune subsets has identified an association
with improved survival for CD8+ cytotoxic T cells [127] and for CD45RO+-activated memory T cells [128].
In addition to T lymphocyte-based immune regulation, emerg-ing evidence suggests that CD20+ B cells and antibody-secreting plasma cells are also associated with active tumor surveillance [129–131]. Immunohistochemistry of primary non-small-cell lung can-cers has demonstrated intratumoral tertiary lymphoid structures that resemble the B-cell follicles found in secondary lymphoid organs such as lymph nodes [132]. These structures are aggregates of immune cells in nonlymphoid tissue, which are thought to facilitate rapid and robust local immune responses [133,134], and may indicate ongoing B-cell maturation, somatic mutation and isotype switch-ing. The function of these tertiary lymphoid structures in cancer development are not known, but gene-expression analysis of the stroma surrounding breast tumors showed that high expression of a B-cell/plasma cell metagene carries a favorable prognosis [135]. Overall, these data suggest that tumor antigen-specific B cells and their corresponding secreted antibodies may actively function in immune control at the tumor site.
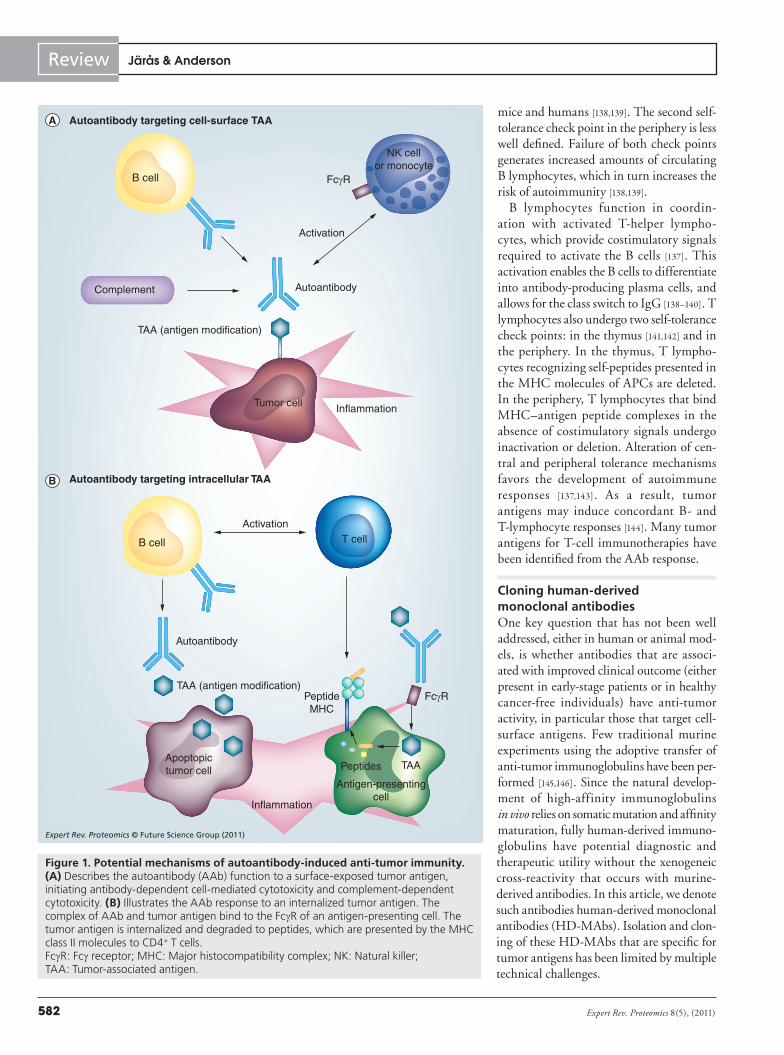
As one arm of the adaptive immune response, antibodies serve multiple functions to prevent pathogenic infections. AAbs that bind self-antigens have escaped self-tolerance, but the function of these antibodies is generally unknown. Upon binding to cell surface molecules, the Fc portion induces antibody-dependent cell medi-ated cytotoxicity (ADCC) and complement-dependent cytotoxicity (CDC) (Figure 1a). AAbs also enhance antigen cross-presentation and subsequent activation of T lymphocytes (Figure 1b). AAb–tumor antigen complexes bind the Fcg receptor of antigen-presenting cells (APCs), with internalization and degradation of tumor antigen to peptides that are presented to CD4+ T cells by the MHC class II molecules of the APC. AAbs can interfere with receptor/ligand interactions; HER2/neu-specific AAbs can be induced that interfere with signal transduction and HER2 phosphorylation [114], although the impact of this effect on clinical outcome is not yet known.
In autoimmune diseases, as well as cancer immunity, many of the identified autoantigens are intracellular proteins found in com-plexes with nucleic acids involved in protein synthesis [136]. These antigens might be immunogenic due to overexpression, apoptosis, reduced degradation or similarity to cross-reacting foreign antigens (mimicry). Whether these AAbs to intra cellular antigens are inert, functionally active in immunosurveillance (either active induc-tion of localized immune responses), simply biomarkers of CD4+ T-lymphocyte immunity or contribute to tumorigenic inflamma-tion is not known. Figure 1 is a schematic of the tumor microenviron-ment, where inflammation and antigen modification are driving forces for AAb production. The B lymphocytes are usually con-trolled by two self-tolerance check points: one in the bone marrow, where they are generated by hemato poietic stem cells, and the other in the periphery, where they undergo matur ation. The self-toler-ance check point in the bone marrow engages the B-cell receptor complex, consisting of the membrane-bound immunoglobulin of immature B cells as well as accessory signaling molecules [137–139]. If the B-cell receptor complex binds self -antigens, B-cell matura-tion is hindered and the receptor edited. Ineffective receptor edit-ing is believed to play an important role in auto immunity in both
Autoantibodies in cancer: prognostic biomarkers & immune activation
Expert Rev. Proteomics 8(5), (2011)582
Review
mice and humans [138,139]. The second self-tolerance check point in the periphery is less well defined. Failure of both check points generates increased amounts of circulating B lymphocytes, which in turn increases the risk of autoimmunity [138,139].
B lymphocytes function in coordin-ation with activated T-helper lympho-cytes, which provide costimulatory signals required to activate the B cells [137]. This activation enables the B cells to differentiate into antibody-producing plasma cells, and allows for the class switch to IgG [138–140]. T lymphocytes also undergo two self-tolerance check points: in the thymus [141,142] and in the periphery. In the thymus, T lympho-cytes recognizing self-peptides presented in the MHC molecules of APCs are deleted. In the periphery, T lymphocytes that bind MHC–antigen peptide complexes in the absence of costimulatory signals undergo inactivation or deletion. Alteration of cen-tral and peripheral tolerance mechanisms favors the development of auto immune responses [137,143]. As a result, tumor antigens may induce concordant B- and T-lymphocyte responses [144]. Many tumor antigens for T-cell immunotherapies have been identified from the AAb response.
Cloning human-derived monoclonal antibodiesOne key question that has not been well addressed, either in human or animal mod-els, is whether antibodies that are associ-ated with improved clinical outcome (either present in early-stage patients or in healthy cancer-free individuals) have anti-tumor activity, in particular those that target cell-surface antigens. Few traditional murine experiments using the adoptive transfer of anti-tumor immunoglobulins have been per-formed [145,146]. Since the natural develop-ment of high-affinity immunoglobulins in vivo relies on somatic mutation and affinity maturation, fully human-derived immuno-globulins have potential diagnostic and therapeutic utility without the xeno geneic cross-reactivity that occurs with murine-derived anti bodies. In this article, we denote such anti bodies human-derived monoclonal antibodies (HD-MAbs). Isolation and clon-ing of these HD-MAbs that are specific for tumor antigens has been limited by multiple technical challenges.
Autoantibody targeting cell-surface TAA
Autoantibody targeting intracellular TAA
Complement
Activation
B cell
Autoantibody
Autoantibody
InflammationTumor cell
TAA (antigen modification)
TAA (antigen modification)
B cell
ActivationT cell
Inflammation
Apoptopictumor cell
FcγR
NK cellor monocyte
PeptideMHC
FcγR
Peptides
Antigen-presentingcell
TAA
Expert Rev. Proteomics © Future Science Group (2011)
Figure 1. Potential mechanisms of autoantibody-induced anti-tumor immunity. (A) Describes the autoantibody (AAb) function to a surface-exposed tumor antigen, initiating antibody-dependent cell-mediated cytotoxicity and complement-dependent cytotoxicity. (B) Illustrates the AAb response to an internalized tumor antigen. The complex of AAb and tumor antigen bind to the FcgR of an antigen-presenting cell. The tumor antigen is internalized and degraded to peptides, which are presented by the MHC class II molecules to CD4+ T cells. FcgR: Fcg receptor; MHC: Major histocompatibility complex; NK: Natural killer; TAA: Tumor-associated antigen.
Järås & Anderson
www.expert-reviews.com 583
Review
HD-MAb from PBMC or B-cell follicles
Tumor antigen
Autoantibody
Autoantibodies
PBMC
Fusion partner
Hybridoma cells
producing antibodies
B-cell follicles fromtumor microenvironment
HD-MAb from a single-cell RT-PCR
HD-MAb from phage display
B-cell follicles fromtumor microenvironment
B-cell mRNA fromtumor microenvironment
Single-cell RT-PCRof B-cell immunoglobulin
Molecular cloning
Production of fully human antibodies
Molecular cloning
cDNA libray construction
TAAMagnetic
bead
Selection of scFv
Fusion partner
Production of fully human antibodies
Antibodycharacterization
Hybridoma cells
producing antibodies
Phage with scFv
Expert Rev. Proteomics © Future Science Group (2011)
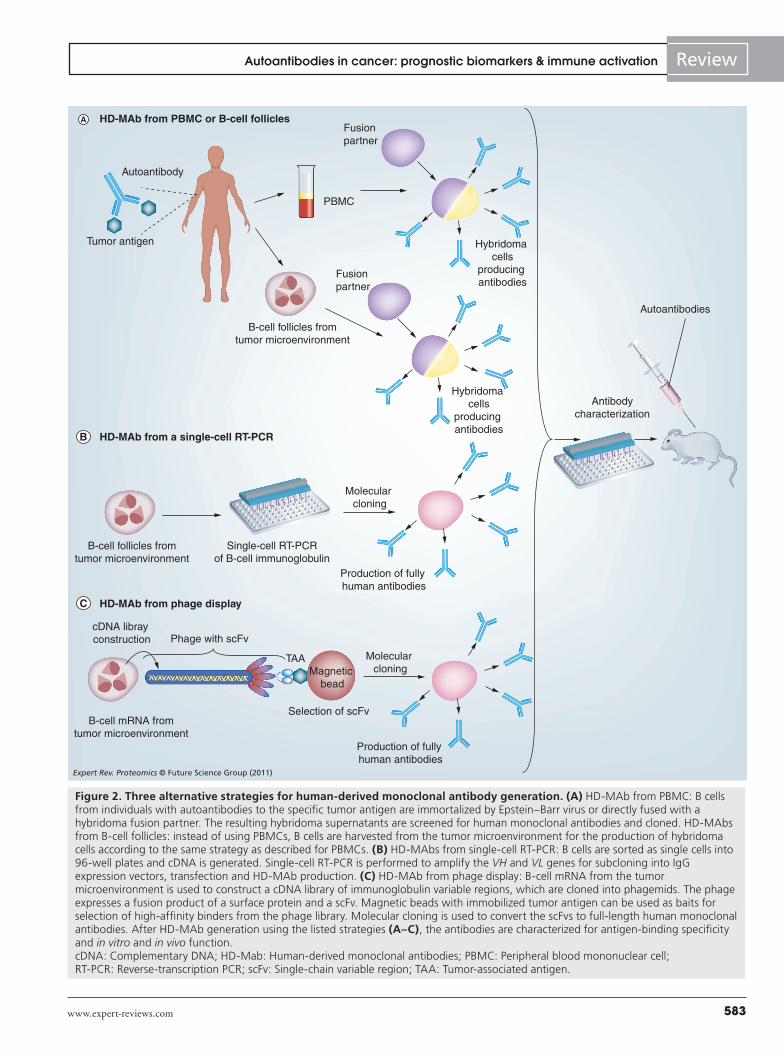
Figure 2. Three alternative strategies for human-derived monoclonal antibody generation. (A) HD-MAb from PBMC: B cells from individuals with autoantibodies to the specific tumor antigen are immortalized by Epstein–Barr virus or directly fused with a hybridoma fusion partner. The resulting hybridoma supernatants are screened for human monoclonal antibodies and cloned. HD-MAbs from B-cell follicles: instead of using PBMCs, B cells are harvested from the tumor microenvironment for the production of hybridoma cells according to the same strategy as described for PBMCs. (B) HD-MAbs from single-cell RT-PCR: B cells are sorted as single cells into 96-well plates and cDNA is generated. Single-cell RT-PCR is performed to amplify the VH and VL genes for subcloning into IgG expression vectors, transfection and HD-MAb production. (C) HD-MAb from phage display: B-cell mRNA from the tumor microenvironment is used to construct a cDNA library of immunoglobulin variable regions, which are cloned into phagemids. The phage expresses a fusion product of a surface protein and a scFv. Magnetic beads with immobilized tumor antigen can be used as baits for selection of high-affinity binders from the phage library. Molecular cloning is used to convert the scFvs to full-length human monoclonal antibodies. After HD-MAb generation using the listed strategies (A–C), the antibodies are characterized for antigen-binding specificity and in vitro and in vivo function. cDNA: Complementary DNA; HD-Mab: Human-derived monoclonal antibodies; PBMC: Peripheral blood mononuclear cell; RT-PCR: Reverse-transcription PCR; scFv: Single-chain variable region; TAA: Tumor-associated antigen.
Autoantibodies in cancer: prognostic biomarkers & immune activation

Expert Rev. Proteomics 8(5), (2011)584
Review
Human hybridoma fusionsThe classical method for isolating monoclonal antibodies is by performing cellular fusions of antibody-producing B cells with a nonsecreting, immortalized hybridoma fusion partner cell. While this is well established in murine systems, performing human hybridoma fusions is technically very difficult. Peripheral blood mononuclear cells or B cells sorted by flow cytometry directly from the tumor microenvironment can be generated by direct fusion to a fusion partner, or first transformed with Epstein–Barr virus to immortalize the enriched B cells [147,148] and then fused to the hybridoma partner to improve the stability and increase the amounts of secreted antibody. In vitro screenings of the antibody supernatants using standard ELISA, or, when generating multiple antibody targets, by bead-array ELISA [51] or protein microarrays, can then be performed. Therapeutic function of the generated antibodies can finally be tested in vivo by administration of the antibodies into murine cancer models (Figure 2a).
Molecular cloning of HD-AbsAlternative ways to produce fully human antibodies are to use single-cell RT-PCR (Figure 2b) [149] or phage-display techno-logy (Figure 2C) [150]. In molecular cloning, single-cell reverse-tran-scription PCR is performed to amplify the rearranged VH and VL genes for subcloning into IgG expression vectors. The drawbacks of this strategy are those linked to single-cell PCR – that is, contami-nations, amplification failure and allele dropouts. Alternatively, a recombinant library of diverse variable regions of immunoglobulin genes isolated as a pool either from tumor -infiltrating lymphocytes or from peripheral blood mononuclear cells is expressed as fusion proteins with the gene III surface protein of bacteriophage. By using protein display of tumor antigen panels as baits, the high-affinity binders are selected from the phage library.
Expert commentary & five-year viewThe identification of AAbs in cancer has multiple potential downstream clinical applications.
Early diagnostic biomarkersThe identification of these antibodies in the sera of patients with early-stage cancer has led to their development as potential
biomarkers for early cancer diagnosis [6,72]. The utility of these AAb panels for early diagnosis awaits confirmation in large, multicenter clinical studies involving blinded prediagnostic samples.
Prognostic biomarkersThe search for prognostic signatures for cancer has led to the iden-tification of multiple molecular biomarkers to predict the risk of systemic disease recurrence. The majority of these markers are based on gene-expression analysis of primary tumor specimens, and the best markers correlate with key biologic changes that impact tumorigenesis, such as cellular proliferation, stem-cell features and invasion. Many of these expression signatures are now available for clinical use, such as Oncotype DX™ and Mammaprint®. These markers identify low-risk patients who are unlikely to benefit from adjuvant chemotherapy. These prog nostic biomarkers have the added potential for tailoring adjuvant therapy trials for high-risk patients. AAbs have advantages as potential independent prog-nostic biomarkers for multiple cancer types, as they depend on blood-based assays and reflect both changes in the primary tumor as well as immuno surveillance. However, the development of these markers depends on retro spective, blinded analysis of large patient cohorts that have defined clinical end points of disease recurrence.
Anti-tumor immunotherapyOne key application of AAb biomarkers is the development of target antigens for T cell-based vaccine development. The development of a robust antibody response to a target antigen is often associated with effective T-cell immunity [151]. For example, NY-ESO-1 was initially identified as an AAb target [84] and subsequently, T-cell epitopes were identified and developed as peptide-based vaccines for multiple cancer types. HER2/neu, which induces AAb in a subset of HER2+ breast and ovarian cancer patients, is associated with multiple T cell-specific epitopes. Melanoma-inhibitor of apoptosis is associated with both B- and T-cell immunity in melanoma [152].
The identification of AAbs that are prognostic for improved or variable clinical outcome, such as p53, MUC1 and carcino-embryonic antigen, suggests that these antibodies are involved in active immunosurveillance. Since naturally occurring antibodies undergo affinity maturation, these antibodies have the potential for use as immunotherapeutics (therapeutic antibodies), either due to
Key issues
• Certain autoantibodies (AAbs) may be prognostic biomarkers for clinical outcome of cancer, but need to be evaluated independently of tumor antigen expression.
• Key technologies for the discovery of tumor antigens and their cognate AAbs include serological analysis of tumor antigens by recombinant complementary DNA expression cloning, serological proteome analysis, multiple affinity protein profiling and protein microarrays.
• Emerging data from functional assays in particular, but also RNA-expression microarrays and in situ histochemistry, which shows evidence of overexpressed tumor antigens associated with the presence of AAbs in serum, suggest that AAbs may have direct and/or indirect functions in cancer immunosurveillance.
• Most studies have evaluated AAbs as diagnostic, but not prognostic, biomarkers. Well-established validation assays and uniform sample cohorts linked to clinical outcome are needed to evaluate the prognostic significance of these AAbs.
• Technologies for the generation of human-derived monoclonal antibodies include hybridoma cell production from B cell-rich material, such as peripheral blood mononuclear cell or peritumoral B-cell follicles, phage display and single-cell reverse-transcription PCR followed by molecular cloning.
• B-cell immunity to tumor antigens may function as part of a coordinated immune response to limit both tumor growth and dissemination.
Järås & Anderson

www.expert-reviews.com 585
Review
direct anti-tumor effects or via immune recruitment and ADCC. The development of these HD-MAbs will require high-throughput molecular cloning and demonstration of anti-tumor efficacy in animal models. While antibodies targeting cell-surface antigens are those that are most likely to be of clinical utility, antibodies targeting intracellular antigens, such as p53, may have efficacy by focusing locoregional immune responses in areas of apoptotic tumor beds (Figure 1b).
Financial & competing interests disclosureKaren Anderson has received travel funding from Luminex and has a patent pending for breast cancer autoantibodies. Kerstin Järås has received Contribution No. 995 from the Barnett Institute. The authors have no other relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript apart from those disclosed.
No writing assistance was utilized in the production of this manuscript.
Autoantibodies in cancer: prognostic biomarkers & immune activation
ReferencesPapers of special note have been highlighted as:• of interest•• of considerable interest
1 Lee JS, Lo PK, Fackler MJ et al. A comparative study of Korean with Caucasian breast cancer reveals frequency of methylation in multiple genes correlates with breast cancer in young, ER, PR-negative breast cancer in Korean women. Cancer Biol. Ther. 6(7), 1114–1120 (2007).
2 Resnick KE, Alder H, Hagan JP, Richardson DL, Croce CM, Cohn DE. The detection of differentially expressed microRNAs from the serum of ovarian cancer patients using a novel real-time PCR platform. Gynecol. Oncol. 112(1), 55–59 (2009).
3 Jacob F, Goldstein DR, Bovin NV et al. Serum anti-glycan antibody detection of non-mucinous ovarian cancers by using a printed glycan array. Int. J. Cancer DOI: 10.1002/ijc.26002 (2011) (Epub ahead of print).
4 Miller JC, Zhou H, Kwekel J et al. Antibody microarray profiling of human prostate cancer sera: antibody screening and identification of potential biomarkers. Proteomics 3(1), 56–63 (2003).
5 Desmetz C, Maudelonde T, Mange A, Solassol J. Identifying autoantibody signatures in cancer: a promising challenge. Expert Rev. Proteomics 6(4), 377–386 (2009).
• Describesthediscoveryandthediagnosticutilityoftumorantigensandtheircognateautoantibodies(AAbs).
6 Tabernero MD, Lv LL, Anderson KS. Autoantibody profiles as biomarkers of breast cancer. Cancer Biomark. 6(5–6), 247–256 (2009).
7 Nagrath S, Sequist LV, Maheswaran S et al. Isolation of rare circulating tumour cells in cancer patients by microchip technology. Nature 450(7173), 1235–1239 (2007).
8 Cramer DW, Bast RC Jr, Berg CD et al. Ovarian cancer biomarker performance in prostate, lung, colorectal, and ovarian cancer screening trial specimens. Cancer Prev. Res. 4(3), 365–374 (2011).
9 Chapman CJ, Thorpe AJ, Murray A et al. Immunobiomarkers in small cell lung cancer: potential early cancer signals. Clin. Cancer Res. 17(6), 1474–1480 (2010).
10 Trivers GE, Cawley HL, DeBenedetti VM et al. Anti-p53 antibodies in sera of workers occupationally exposed to vinyl chloride. J. Natl Cancer Inst. 87(18), 1400–1407 (1995).
11 Rubinstein ND, Mayrose I, Martz E, Pupko T. Epitopia: a web-server for predicting B-cell epitopes. BMC Bioinformatics 10, 287 (2009).
12 Gaudin E, Hao Y, Rosado MM, Chaby R, Girard R, Freitas AA. Positive selection of B cells expressing low densities of self-reactive BCRs. J. Exp. Med. 199(6), 843–853 (2004).
13 Wardemann H, Yurasov S, Schaefer A, Young JW, Meffre E, Nussenzweig MC. Predominant autoantibody production by early human B cell precursors. Science 301(5638), 1374–1377 (2003).
14 Soussi T. p53 antibodies in the sera of patients with various types of cancer: a review. Cancer Res. 60(7), 1777–1788 (2000).
15 Chen YT, Scanlan MJ, Sahin U et al. A testicular antigen aberrantly expressed in human cancers detected by autologous antibody screening. Proc. Natl Acad. Sci. USA 94(5), 1914–1918 (1997).
16 Schubert U, Anton LC, Gibbs J, Norbury CC, Yewdell JW, Bennink JR. Rapid degradation of a large fraction of newly synthesized proteins by proteasomes. Nature 404(6779), 770–774 (2000).
17 Ulanet DB, Torbenson M, Dang CV, Casciola-Rosen L, Rosen A. Unique conformation of cancer autoantigen B23 in hepatoma: a mechanism for specificity in the autoimmune response. Proc. Natl Acad. Sci. USA 100(21), 12361–12366 (2003).
18 von Mensdorff-Pouilly S, Petrakou E, Kenemans P et al. Reactivity of natural and induced human antibodies to MUC1 mucin with MUC1 peptides and N-acetylgalactosamine (GalNAc) peptides. Int. J. Cancer 86(5), 702–712 (2000).
19 Ait-Tahar K, Damm-Welk C, Burkhardt B et al. Correlation of the autoantibody response to the ALK oncoantigen in pediatric anaplastic lymphoma kinase-positive anaplastic large cell lymphoma with tumor dissemination and relapse risk. Blood 115(16), 3314–3319 (2010).
• Demonstratesthefirstclinicalevidencethatincreasedpre-existinglevelsofAAbstoanaplasticlymphomakinaseiscorrelatedtoinhibitionoflymphomadisseminationanddecreasedriskofrelapseinanaplasticlarge-celllymphoma.
20 Albanopoulos K, Armakolas A, Konstadoulakis MM et al. Prognostic significance of circulating antibodies against carcinoembryonic antigen (anti-CEA) in patients with colon cancer. Am. J. Gastroenterol. 95(4), 1056–1061 (2000).
21 Albertus DL, Seder CW, Chen G et al. AZGP1 autoantibody predicts survival and histone deacetylase inhibitors increase expression in lung adenocarcinoma. J. Thorac. Oncol. 3(11), 1236–1244 (2008).
22 Anderson KS, Wong J, Vitonis A et al. p53 autoantibodies as potential detection and prognostic biomarkers in serous ovarian cancer. Cancer Epidemiol. Biomarkers Prev. 19(3), 859–868 (2010).
• Recentstudydescribingthecorrelationbetweenelevatedlevelsofp53AAbandimprovedsurvivalinserousovariancancer.
23 Goodell V, Salazar LG, Urban N et al. Antibody immunity to the p53 oncogenic protein is a prognostic indicator in ovarian cancer. J. Clin. Oncol. 24(5), 762–768 (2006).
24 Haidopoulos D, Konstadoulakis MM, Antonakis PT et al. Circulating anti-CEA antibodies in the sera of patients with breast cancer. Eur. J. Surg. Oncol. 26(8), 742–746 (2000).
25 Hamanaka Y, Suehiro Y, Fukui M, Shikichi K, Imai K, Hinoda Y. Circulating anti-MUC1 IgG antibodies as a favorable prognostic factor for pancreatic cancer. Int. J. Cancer 103(1), 97–100 (2003).

Expert Rev. Proteomics 8(5), (2011)586
Review Järås & Anderson
26 Heller A, Zornig I, Muller T et al. Immunogenicity of SEREX-identified antigens and disease outcome in pancreatic cancer. Cancer Immunol. Immunother. 59(9), 1389–1400 (2010).
27 Hirasawa Y, Kohno N, Yokoyama A, Kondo K, Hiwada K, Miyake M. Natural autoantibody to MUC1 is a prognostic indicator for non-small cell lung cancer. Am. J. Respir. Crit. Care Med. 161(2 Pt 1), 589–594 (2000).
28 Kurtenkov O, Klaamas K, Mensdorff-Pouilly S, Miljukhina L, Shljapnikova L, Chuzmarov V. Humoral immune response to MUC1 and to the Thomsen–Friedenreich (TF) glycotope in patients with gastric cancer: relation to survival. Acta Oncol. 46(3), 316–323 (2007).
29 Lakota J, Skultety L, Dubrovcakova M, Altaner C. Presence of serum carbonic anhydrase autoantibodies in patients relapsed after autologous stem cell transplantation indicates an improved prognosis. Neoplasma 55(6), 488–492 (2008).
30 Litvak DA, Gupta RK, Yee R, Wanek LA, Ye W, Morton DL. Endogenous immune response to early- and intermediate-stage melanoma is correlated with outcomes and is independent of locoregional relapse and standard prognostic factors. J. Am. Coll. Surg. 198(1), 27–35 (2004).
31 Pallasch CP, Struss AK, Munnia A et al. Autoantibodies against GLEA2 and PHF3 in glioblastoma: tumor-associated autoantibodies correlated with prolonged survival. Int. J. Cancer 117(3), 456–459 (2005).
32 Tamura H, Dan K, Yokose N et al. Prognostic significance of WT1 mRNA and anti-WT1 antibody levels in peripheral blood in patients with myelodysplastic syndromes. Leuk. Res. 34(8), 986–990 (2010).
33 Tomaino B, Cappello P, Capello M et al. Circulating autoantibodies to phosphorylated a-enolase are a hallmark of pancreatic cancer. J. Proteome Res. 10(1), 105–112 (2011).
34 von Mensdorff-Pouilly S, Verstraeten AA, Kenemans P et al. Survival in early breast cancer patients is favorably influenced by a natural humoral immune response to polymorphic epithelial mucin. J. Clin. Oncol. 18(3), 574–583 (2000).
35 Sahin U, Tureci O, Schmitt H et al. Human neoplasms elicit multiple specific immune responses in the autologous host. Proc. Natl Acad. Sci. USA 92(25), 11810–11813 (1995).
36 Klade CS, Voss T, Krystek E et al. Identification of tumor antigens in renal cell carcinoma by serological proteome analysis. Proteomics 1(7), 890–898 (2001).
37 Seliger B, Kellner R. Design of proteome-based studies in combination with serology for the identification of biomarkers and novel targets. Proteomics 2(12), 1641–1651 (2002).
38 Seliger B, Lichtenfels R, Kellner R. Detection of renal cell carcinoma-associated markers via proteome- and other ‘ome’-based analyses. Brief Funct. Genomic Proteomic 2(3), 194–212 (2003).
39 Hardouin J, Lasserre JP, Canelle L et al. Usefulness of autoantigens depletion to detect autoantibody signatures by multiple affinity protein profiling. J. Sep. Sci. 30(3), 352–358 (2007).
40 Ramachandran N, Hainsworth E, Bhullar B et al. Self-assembling protein microarrays. Science 305(5680), 86–90 (2004).
41 Ramachandran N, Raphael JV, Hainsworth E et al. Next-generation high-density self-assembling functional protein arrays. Nat. Methods 5(6), 535–538 (2008).
42 Wang X, Yu J, Sreekumar A et al. Autoantibody signatures in prostate cancer. N. Engl. J. Med. 353(12), 1224–1235 (2005).
• Describesasuccessfuldiagnosticapproachfordetectionof22AAbsdirectedtopeptidesderivedfromprostatecancertumortissueanddisplayedbyphagesinamicroarrayformat.
43 Fosså A, Berner A, Fosså SD, Hernes E, Gaudernack G, Smeland EB. NY-ESO-1 protein expression and humoral immune responses in prostate cancer. Prostate 59(4), 440–447 (2004).
44 Kelley MC, Gupta RK, Hsueh EC, Yee R, Stern S, Morton DL. Tumor-associated antigen TA90 immune complex assay predicts recurrence and survival after surgical treatment of stage I–III melanoma. J. Clin. Oncol. 19(4), 1176–1182 (2001).
45 Regele S, Vogl FD, Kohler T, Kreienberg R, Runnebaum IB. p53 autoantibodies can be indicative of the development of breast cancer relapse. AntiCancer Res. 23(1B), 761–764 (2003).
46 Ehrlich JR, Caiazzo RJ, Jr., Qiu W et al. A native antigen ‘reverse capture’ microarray platform for autoantibody profiling of prostate cancer sera. Proteomics Clin. Appl. 1(5), 476–485 (2007).
47 Li L, Chen SH, Yu CH, Li YM, Wang SQ. Identification of hepatocellular-carcinoma-associated antigens and autoantibodies by serological proteome analysis combined with protein microarray. J. Proteome Res. 7(2), 611–620 (2008).
48 Merbl Y, Itzchak R, Vider-Shalit T et al. A systems immunology approach to the host–tumor interaction: large-scale patterns of natural autoantibodies distinguish healthy and tumor-bearing mice. PLoS One 4(6), e6053 (2009).
49 Qin S, Qiu W, Ehrlich JR et al. Development of a ‘reverse capture’ autoantibody microarray for studies of antigen-autoantibody profiling. Proteomics 6(10), 3199–3209 (2006).
50 Stempfer R, Syed P, Vierlinger K et al. Tumour auto-antibody screening: performance of protein microarrays using SEREX derived antigens. BMC Cancer 10, 627 (2010).
51 Wong J, Sibani S, Lokko NN, LaBaer J, Anderson KS. Rapid detection of antibodies in sera using multiplexed self-assembling bead arrays. J. Immunol. Methods 350(1–2), 171–182 (2009).
52 Sahin U, Tureci O, Pfreundschuh M. Serological identification of human tumor antigens. Curr. Opin. Immunol. 9(5), 709–716 (1997).
53 Gure AO, Altorki NK, Stockert E, Scanlan MJ, Old LJ, Chen YT. Human lung cancer antigens recognized by autologous antibodies: definition of a novel cDNA derived from the tumor suppressor gene locus on chromosome 3p21.3. Cancer Res. 58(5), 1034–1041 (1998).
54 Gure AO, Stockert E, Scanlan MJ et al. Serological identification of embryonic neural proteins as highly immunogenic tumor antigens in small cell lung cancer. Proc. Natl Acad. Sci. USA 97(8), 4198–4203 (2000).
55 Jager D, Stockert E, Gure AO et al. Identification of a tissue-specific putative transcription factor in breast tissue by serological screening of a breast cancer library. Cancer Res. 61(5), 2055–2061 (2001).
56 Jager D, Unkelbach M, Frei C et al. Identification of tumor-restricted antigens NY-BR-1, SCP-1, and a new cancer/testis-like antigen NW-BR-3 by serological screening of a testicular library with breast cancer serum. Cancer Immun. 2, 5 (2002).
57 Minenkova O, Pucci A, Pavoni E et al. Identification of tumor-associated antigens by screening phage-displayed human

www.expert-reviews.com 587
ReviewAutoantibodies in cancer: prognostic biomarkers & immune activation
cDNA libraries with sera from tumor patients. Int. J. Cancer 106(4), 534–544 (2003).
58 Chatterjee M, Mohapatra S, Ionan A et al. Diagnostic markers of ovarian cancer by high-throughput antigen cloning and detection on arrays. Cancer Res. 66(2), 1181–1190 (2006).
59 Chen G, Wang X, Yu J et al. Autoantibody profiles reveal ubiquilin 1 as a humoral immune response target in lung adenocarcinoma. Cancer Res. 67(7), 3461–3467 (2007).
60 Boder ET, Wittrup KD. Yeast surface display for screening combinatorial polypeptide libraries. Nat. Biotechnol. 15(6), 553–557 (1997).
61 Kim MS, Choi HY, Choi YS, Kim J, Kim YS. Optimized serological isolation of lung-cancer-associated antigens from a yeast surface-expressed cDNA library. J. Microbiol. Biotechnol. 17(6), 993–1001 (2007).
62 Wadle A, Mischo A, Imig J et al. Serological identification of breast cancer-related antigens from a Saccharomyces cerevisiae surface display library. Int. J. Cancer 117(1), 104–113 (2005).
63 Li Q, Lv Y, Li C et al. Vitiligo autoantigen VIT75 is identified as lamin A in vitiligo by serological proteome analysis based on mass spectrometry. J. Invest. Dermatol. 131(3), 727–734 (2011).
64 Fujita Y, Nakanishi T, Hiramatsu M et al. Proteomics-based approach identifying autoantibody against peroxiredoxin VI as a novel serum marker in esophageal squamous cell carcinoma. Clin. Cancer Res. 12(21), 6415–6420 (2006).
65 Park S, Lim Y, Lee D et al. Identification and characterization of a novel cancer/testis antigen gene CAGE-1. Biochim. Biophys. Acta 1625(2), 173–182 (2003).
66 Yang F, Xiao ZQ, Zhang XZ et al. Identification of tumor antigens in human lung squamous carcinoma by serological proteome analysis. J. Proteome Res. 6(2), 751–758 (2007).
67 Bouwman K, Qiu J, Zhou H et al. Microarrays of tumor cell derived proteins uncover a distinct pattern of prostate cancer serum immunoreactivity. Proteomics 3(11), 2200–2207 (2003).
68 Nam MJ, Madoz-Gurpide J, Wang H et al. Molecular profiling of the immune response in colon cancer using protein microarrays: occurrence of autoantibodies to ubiquitin C-terminal hydrolase L3. Proteomics 3(11), 2108–2115 (2003).
69 Qiu J, Madoz-Gurpide J, Misek DE et al. Development of natural protein microarrays for diagnosing cancer based on an antibody response to tumor antigens. J. Proteome Res. 3(2), 261–267 (2004).
70 Hudson ME, Pozdnyakova I, Haines K, Mor G, Snyder M. Identification of differentially expressed proteins in ovarian cancer using high-density protein microarrays. Proc. Natl Acad. Sci. USA 104(44), 17494–17499 (2007).
71 Anderson KS, Ramachandran N, Wong J et al. Application of protein microarrays for multiplexed detection of antibodies to tumor antigens in breast cancer. J. Proteome Res. 7(4), 1490–1499 (2008).
72 Anderson KS, Sibani S, Wallstrom G et al. Protein microarray signature of autoantibody biomarkers for the early detection of breast cancer. J. Proteome Res. 10(1), 85–96 (2011).
73 Ramachandran N, Anderson KS, Raphael JV et al. Tracking humoral responses using self assembling protein microarrays. Proteomics Clin. Appl. 2(10–11), 1518–1527 (2008).
74 Anderson KS. Multiplexed detection of antibodies using programmable bead arrays. Methods Mol. Biol. 723, 227–238 (2011).
75 Waterboer T, Sehr P, Michael KM et al. Multiplex human papillomavirus serology based on in situ-purified glutathione s-transferase fusion proteins. Clin. Chem. 51(10), 1845–1853 (2005).
76 Lenshof A, Ahmad-Tajudin A, Jaras K et al. Acoustic whole blood plasmapheresis chip for prostate specific antigen microarray diagnostics. Anal. Chem. 81(15), 6030–6037 (2009).
77 Tiberti C, Verrienti A, Fiore B et al. IA-2 combined epitope assay: a new, highly sensitive approach to evaluate IA-2 humoral autoimmunity in Type 1 diabetes. Clin. Immunol. 115(3), 260–267 (2005).
78 Jaras K, Ressine A, Nilsson E et al. Reverse-phase versus sandwich antibody microarray, technical comparison from a clinical perspective. Anal. Chem. 79(15), 5817–5825 (2007).
79 Jaras K, Tajudin AA, Ressine A et al. ENSAM: europium nanoparticles for signal enhancement of antibody microarrays on nanoporous silicon. J. Proteome Res. 7(3), 1308–1314 (2008).
80 Nishizuka S, Charboneau L, Young L et al. Proteomic profiling of the NCI-60 cancer cell lines using new high-density reverse-phase lysate microarrays. Proc. Natl Acad. Sci. USA 100(24), 14229–14234 (2003).
81 Gustafsdottir SM, Schlingemann J, Rada-Iglesias A et al. In vitro analysis of DNA–protein interactions by proximity ligation. Proc. Natl Acad. Sci. USA 104(9), 3067–3072 (2007).
82 Zhou H, Bouwman K, Schotanus M et al. Two-color, rolling-circle amplification on antibody microarrays for sensitive, multiplexed serum-protein measurements. Genome Biol. 5(4), R28 (2004).
83 Burbelo PD, Bren KE, Ching KH et al. LIPS arrays for simultaneous detection of antibodies against partial and whole proteomes of HCV, HIV and EBV. Mol. Biosyst. 7(5), 1453–1462 (2011).
84 Gnjatic S, Nishikawa H, Jungbluth AA et al. NY-ESO-1: review of an immunogenic tumor antigen. Adv. Cancer Res. 95, 1–30 (2006).
85 Davidoff AM, Iglehart JD, Marks JR. Immune response to p53 is dependent upon p53/HSP70 complexes in breast cancers. Proc. Natl Acad. Sci. USA 89(8),3439–3442 (1992).
86 Green JA, Mudenda B, Jenkins J et al. Serum p53 auto-antibodies: incidence in familial breast cancer. Eur. J. Cancer 30A(5), 580–584 (1994).
87 Lechpammer M, Lukac J, Lechpammer S, Kovacevic D, Loda M, Kusic Z. Humoral immune response to p53 correlates with clinical course in colorectal cancer patients during adjuvant chemotherapy. Int. J. Colorectal Dis. 19(2), 114–120 (2004).
88 Warnakulasuriya S, Soussi T, Maher R, Johnson N, Tavassoli M. Expression of p53 in oral squamous cell carcinoma is associated with the presence of IgG and IgA p53 autoantibodies in sera and saliva of the patients. J. Pathol. 192(1), 52–57 (2000).
89 Goodell V, Waisman J, Salazar LG et al. Level of HER-2/neu protein expression in breast cancer may affect the development of endogenous HER-2/neu-specific immunity. Mol. Cancer Ther. 7(3), 449–454 (2008).
90 Maio M, Coral S, Sigalotti L et al. Analysis of cancer/testis antigens in sporadic medullary thyroid carcinoma: expression and humoral response to NY-ESO-1. J. Clin. Endocrinol. Metab. 88(2), 748–754 (2003).
91 Jager E, Stockert E, Zidianakis Z et al. Humoral immune responses of cancer patients against ‘cancer-testis’ antigen NY-ESO-1: correlation with clinical events. Int. J. Cancer 84(5), 506–510 (1999).

Expert Rev. Proteomics 8(5), (2011)588
Review Järås & Anderson
92 Stockert E, Jager E, Chen YT et al. A survey of the humoral immune response of cancer patients to a panel of human tumor antigens. J. Exp. Med. 187(8), 1349–1354 (1998).
93 Reuschenbach M, Waterboer T, Wallin KL et al. Characterization of humoral immune responses against p16, p53, HPV16 E6 and HPV16 E7 in patients with HPV-associated cancers. Int. J. Cancer 123(11), 2626–2631 (2008).
94 Akcakanat A, Kanda T, Koyama Y et al. NY-ESO-1 expression and its serum immunoreactivity in esophageal cancer. Cancer Chemother. Pharmacol. 54(1), 95–100 (2004).
95 Reuschenbach M, von Knebel Doeberitz M, Wentzensen N. A systematic review of humoral immune responses against tumor antigens. Cancer Immunol. Immunother. 58(10), 1535–1544 (2009).
96 Sotiriou C, Pusztai L. Gene-expression signatures in breast cancer. N. Engl. J. Med. 360(8), 790–800 (2009).
97 Qiu J, Choi G, Li L et al. Occurrence of autoantibodies to annexin I, 14–13–3 q and LAMR1 in prediagnostic lung cancer sera. J. Clin. Oncol. 26(31), 5060–5066 (2008).
98 Suzuki H, Graziano DF, McKolanis J, Finn OJ. T cell-dependent antibody responses against aberrantly expressed cyclin B1 protein in patients with cancer and premalignant disease. Clin. Cancer Res. 11(4), 1521–1526 (2005).
99 Trivers GE, De Benedetti VM, Cawley HL et al. Anti-p53 antibodies in sera from patients with chronic obstructive pulmonary disease can predate a diagnosis of cancer. Clin. Cancer Res. 2(10), 1767–1775 (1996).
100 Jimenez LG, Aguilar MC, Monroy OL et al. Detection of autoantibodies to survivin in cervical mucus from patients with human papillomavirus-associated cervical cancer and precursor lesions. Autoimmunity 40(1), 66–72 (2007).
101 Tureci O, Mack U, Luxemburger U et al. Humoral immune responses of lung cancer patients against tumor antigen NY-ESO-1. Cancer Lett. 236(1), 64–71 (2006).
102 Abendstein B, Marth C, Muller-Holzner E, Widschwendter M, Daxenbichler G, Zeimet AG. Clinical significance of serum and ascitic p53 autoantibodies in epithelial ovarian carcinoma. Cancer 88(6), 1432–1437 (2000).
103 Lai CL, Tsai CM, Tsai TT et al. Presence of serum anti-p53 antibodies is associated with pleural effusion and poor prognosis in
lung cancer patients. Clin. Cancer Res. 4(12), 3025–3030 (1998).
104 Lenner P, Wiklund F, Emdin SO et al. Serum antibodies against p53 in relation to cancer risk and prognosis in breast cancer: a population-based epidemiological study. Br. J. Cancer 79(5–6), 927–932 (1999).
105 Mayerhofer K, Tempfer C, Kucera E et al. Humoral p53 antibody response is a prognostic parameter in ovarian cancer. Anti Cancer Res. 19(1B), 875–878 (1999).
106 Tang R, Ko MC, Wang JY et al. Humoral response to p53 in human colorectal tumors: a prospective study of 1,209 patients. Int. J. Cancer 94(6), 859–863 (2001).
107 Takeda A, Shimada H, Nakajima K et al. Monitoring of p53 autoantibodies after resection of colorectal cancer: relationship to operative curability. Eur. J. Surg. 167(1), 50–53 (2001).
108 Metcalfe S, Wheeler TK, Picken S, Negus S, Jo Milner A. P53 autoantibodies in 1006 patients followed up for breast cancer. Breast Cancer Res. 2(6), 438–443 (2000).
109 Rodolfo M, Luksch R, Stockert E et al. Antigen-specific immunity in neuroblastoma patients: antibody and T-cell recognition of NY-ESO-1 tumor antigen. Cancer Res. 63(20), 6948–6955 (2003).
110 Kanojia D, Garg M, Gupta S, Gupta A, Suri A. Sperm-associated antigen 9, a novel biomarker for early detection of breast cancer. Cancer Epidemiol. Biomarkers Prev. 18(2), 630–639 (2009).
111 Jager D, Knuth A. Antibodies and vaccines – hope or illusion? Breast 14(6), 631–635 (2005).
112 Disis ML, Knutson KL, Schiffman K, Rinn K, McNeel DG. Pre-existent immunity to the HER-2/neu oncogenic protein in patients with HER-2/neu overexpressing breast and ovarian cancer. Breast Cancer Res. Treat.62(3), 245–252 (2000).
113 Disis ML, Pupa SM, Gralow JR, Dittadi R, Menard S, Cheever MA. High-titer HER-2/neu protein-specific antibody can be detected in patients with early-stage breast cancer. J. Clin. Oncol. 15(11), 3363–3367 (1997).
114 Montgomery RB, Makary E, Schiffman K, Goodell V, Disis ML. Endogenous anti-HER2 antibodies block HER2 phosphorylation and signaling through extracellular signal-regulated kinase. Cancer Res. 65(2), 650–656 (2005).
115 Disis ML, Schiffman K. Cancer vaccines targeting the HER2/neu oncogenic protein. Semin. Oncol. 28(6 Suppl. 18), 12–20 (2001).
116 Kurtenkov O, Klaamas K, Rittenhouse-Olson K et al. IgG immune response to tumor-associated carbohydrate antigens (TF, Tn, aGal) in patients with breast cancer: impact of neoadjuvant chemotherapy and relation to the survival. Exp. Oncol. 27(2), 136–140 (2005).
117 Anderson KS, Wong J, D’Souza G et al. Serum antibodies to the HPV16 proteome as biomarkers for head and neck cancer. Br. J. Cancer 104(12), 1896–1905 (2011).
118 Lowy DR, Munger K. Prognostic implications of HPV in oropharyngeal cancer. N. Engl. J. Med. 363(1), 82–84 (2010).
119 Smith EM, Rubenstein LM, Ritchie JM et al. Does pretreatment seropositivity to human papillomavirus have prognostic significance for head and neck cancers? Cancer Epidemiol. Biomarkers Prev. 17(8), 2087–2096 (2008).
120 Dong J, Zeng BH, Xu LH et al. Anti-CDC25B autoantibody predicts poor prognosis in patients with advanced esophageal squamous cell carcinoma. J. Transl. Med. 8, 81 (2010).
121 Bei R, Masuelli L, Palumbo C, Modesti M, Modesti A. A common repertoire of autoantibodies is shared by cancer and autoimmune disease patients: inflammation in their induction and impact on tumor growth. Cancer Lett. 281(1), 8–23 (2009).
• Describestheinteractionbetweeninflammationandtumorprogression.
122 Dave SS, Wright G, Tan B et al. Prediction of survival in follicular lymphoma based on molecular features of tumor-infiltrating immune cells. N. Engl. J. Med. 351(21), 2159–2169 (2004).
123 Galon J, Costes A, Sanchez-Cabo F et al. Type, density, and location of immune cells within human colorectal tumors predict clinical outcome. Science 313(5795), 1960–1964 (2006).
124 Pages F, Berger A, Camus M et al. Effector memory T cells, early metastasis, and survival in colorectal cancer. N. Engl. J. Med. 353(25), 2654–2666 (2005).
125 Schmidt M, Bohm D, von Torne C et al. The humoral immune system has a key prognostic impact in node-negative breast cancer. Cancer Res. 68(13), 5405–5413 (2008).

www.expert-reviews.com 589
ReviewAutoantibodies in cancer: prognostic biomarkers & immune activation
126 Zhang L, Conejo-Garcia JR, Katsaros D et al. Intratumoral T cells, recurrence, and survival in epithelial ovarian cancer. N. Engl. J. Med. 348(3), 203–213 (2003).
127 Pages F, Galon J, Dieu-Nosjean MC, Tartour E, Sautes-Fridman C, Fridman WH. Immune infiltration in human tumors: a prognostic factor that should not be ignored. Oncogene 29(8), 1093–1102 (2010).
128 Rauser S, Langer R, Tschernitz S et al. High number of CD45RO+ tumor infiltrating lymphocytes is an independent prognostic factor in non-metastasized (stage I–IIA) esophageal adenocarcinoma. BMC Cancer 10, 608 (2010).
129 Milne K, Kobel M, Kalloger SE et al. Systematic analysis of immune infiltrates in high-grade serous ovarian cancer reveals CD20, FoxP3 and TIA-1 as positive prognostic factors. PLoS One 4(7), e6412 (2009).
130 Nelson BH. CD20+ B cells: the other tumor-infiltrating lymphocytes. J. Immunol. 185(9), 4977–4982 (2010).
131 Willis SN, Mallozzi SS, Rodig SJ et al. The microenvironment of germ cell tumors harbors a prominent antigen-driven humoral response. J. Immunol. 182(5), 3310–3317 (2009).
132 Dieu-Nosjean MC, Antoine M, Danel C et al. Long-term survival for patients with non-small-cell lung cancer with intratumoral lymphoid structures. J. Clin. Oncol. 26(27), 4410–4417 (2008).
133 Carragher DM, Rangel-Moreno J, Randall TD. Ectopic lymphoid tissues and local immunity. Semin. Immunol. 20(1), 26–42 (2008).
134 Drayton DL, Liao S, Mounzer RH, Ruddle NH. Lymphoid organ development: from ontogeny to neogenesis. Nat. Immunol. 7(4), 344–353 (2006).
135 Bianchini G, Qi Y, Alvarez RH et al. Molecular anatomy of breast cancer stroma and its prognostic value in estrogen receptor-positive and -negative cancers. J. Clin. Oncol. 28(28), 4316–4323 (2010).
136 Routsias JG, Tzioufas AG, Moutsopoulos HM. The clinical value of intracellular autoantigens B-cell epitopes in systemic rheumatic diseases. Clin. Chim. Acta 340(1–2), 1–25 (2004).
137 Goodnow CC, Sprent J, Fazekas de St Groth B, Vinuesa CG. Cellular and genetic mechanisms of self tolerance and autoimmunity. Nature 435(7042), 590–597 (2005).
138 Wardemann H, Nussenzweig MC. B-cell self-tolerance in humans. Adv. Immunol. 95, 83–110 (2007).
139 Yurasov S, Nussenzweig MC. Regulation of autoreactive antibodies. Curr. Opin. Rheumatol. 19(5), 421–426 (2007).
140 Bei R, Masuelli L, Moriconi E et al. Immune responses to all ErbB family receptors detectable in serum of cancer patients. Oncogene 18(6), 1267–1275 (1999).
141 Hogquist KA, Baldwin TA, Jameson SC. Central tolerance: learning self-control in the thymus. Nat. Rev. Immunol. 5(10), 772–782 (2005).
142 Kyewski B, Klein L. A central role for central tolerance. Annu. Rev. Immunol. 24, 571–606 (2006).
143 Plotz PH. The autoantibody repertoire: searching for order. Nat. Rev. Immunol. 3(1), 73–78 (2003).
144 Anderson KS, LaBaer J. The sentinel within: exploiting the immune system for cancer biomarkers. J. Proteome Res. 4(4), 1123–1133 (2005).
145 Hardy B, Indjiia L, Rodionov G, Raiter A, Inbal A. Treatment with BAT monoclonal antibody decreases tumor burden in a murine model of leukemia/lymphoma. Int. J. Oncol. 19(5), 897–902 (2001).
146 Liebman MA, Roche MI, Williams BR, Kim J, Pageau SC, Sharon J. Antibody treatment of human tumor xenografts elicits active anti-tumor immunity in nude mice. Immunol. Lett. 114(1), 16–22 (2007).
147 Dejnirattisai W, Jumnainsong A, Onsirisakul N et al. Cross-reacting antibodies enhance dengue virus infection in humans. Science 328(5979), 745–748 (2010).
148 Traggiai E, Becker S, Subbarao K et al. An efficient method to make human monoclonal antibodies from memory B cells: potent neutralization of SARS coronavirus. Nat. Med. 10(8), 871–875 (2004).
149 Tiller T, Meffre E, Yurasov S, Tsuiji M, Nussenzweig MC, Wardemann H. Efficient generation of monoclonal antibodies from single human B cells by single cell RT-PCR and expression vector cloning. J. Immunol. Methods 329(1–2), 112–124 (2008).
• Describesthegenerationofhumanmonoclonalantibodiesbysingle-cellreverse-transcriptionPCRandRNAfromBcells.
150 Gruel N, Kotlan B, Beuzard M, Teillaud JL. Generation of scFv from a phage display mini-library derived from tumor-infiltrating B-cells. Methods Mol. Biol. 193, 281–300 (2002).
151 Philip R, Murthy S, Krakover J et al. Shared immunoproteome for ovarian cancer diagnostics and immunotherapy: potential theranostic approach to cancer. J. Proteome Res. 6(7), 2509–2517 (2007).
152 Schmollinger JC, Vonderheide RH, Hoar KM et al. Melanoma inhibitor of apoptosis protein (ML-IAP) is a target for immune-mediated tumor destruction. Proc. Natl Acad. Sci. USA 100(6), 3398–3403 (2003).
Website
201 The Cancer Genome Atlas http://cancergenome.nih.gov

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.