antigen-specific suppressor factors produced by t cell hybridomas for delayed-type hypersensitivity
TRANSCRIPT

572 J. Hewitt and F. Y. Liew Eur. J. Immunol. 1979. 9: 572-575
Jane Hewitt and Foo Y. Liew
Department of Experimental
Antigen-specific suppressor factors produced by T cell hybridomas for delayed-type hypersensitivity
Immunobiology, The Wellcome Research Laboratories, Beckenham
This study describes the generation of T hybridoma lines which secrete factors specifi- cally suppressing delayed-type hypersensitivity (DTH) to sheep red blood cells (SRBC). AKR strain-derived T lymphoma BW 5147 cells were fused with spleen cells from mice primed with SRBC and containing antigen-specific T suppressor cells for DTH. Supernatants from the derived hybridomas were tested for suppression of either expression or induction of DTH to SRBC. Six lines produced specific suppres- sor activity for the expression of DTH; 4 lines produced suppressor activity for the induction of DTH of which only one line was antigen-specific. These lines were passaged in normal AKR mice, and the serum obtained had activity up to dilution. The factor was effective across the H-2 barrier.
1 Introduction
The role of T cell subpopulations and their secreted products in the regulation of the immune response is the subject of much investigation. Application of the cell hybridization technique [l, 21 to establish cell lines of functional, differenti- ated T cells could advance such studies by providing large quantities of homogeneous material. Fusion of lymphocytes with the BW 5147 [3-51 or EL4 T cell [6] murine lymphoma lines resulting in T cell hybridomas has already been achieved. Recently,'T cell hybridomas or their products which specifi- cally suppress antibody responses in vitro [7] or in vivo [6, 81 have been reported. We have obtained seven hybridoma lines secreting antigen-specific factors which suppress either the induction or the expression of delayed-type hypersensitivity (DTH) to sheep red blood cells (SRBC) in vivo. These factors share characteristics with T cell suppressor factors effective in other systems [9-141 and may be similar, if not identical, to the factor 1151 produced by DTH suppressor cells [16].
2 Materials and methods
2.1 Cells
The AKR-derived T cell line BW 5147 (hypoxanthine guanine phosphoribosyl transferase-negative), derived by Dr. R. Hymann and obtained from Dr. C. Milstein, was used. These cells cannot grow in selective HAT medium [17]. CBA mice, at least 8 weeks old, were injected with 1 X lo9 SRBC intravenously (i.v.), and the spleen cells harvested 3 days later.
2.2 Cell fusion
Cell fusion was performed essentially as described previously using polyethylene glycol (PEG) IS]. Spleen lymphocytes and
[I 23951
Correspondence: Jane Hewitt, Department of Experimental Immunobiulogy, The Wellcome Research Laboratories, Beckenham, Kent. GB
Abbreviations: DTH: Delayed-type hypersensitivity SRBC: Sheep red blood cells CRBC: Chicken red blood cells PEG: Polyethylene glycol DMEM: Dulbecco's modified Eagle's medium HAT medium: DMEM supplemented with hypoxanthine, aminopterin and thymidine BSS: Balanced salt solution
BW 5147 cells mixed in a 10: 1 ratio were washed twice in serum-free balanced salt solution (BSS) and pelleted together at 4OOxg. In two fusion experiments designated N and 0, 3 X lo8 or 1 X lo8 spleen lymphocytes were used, respectively. The cell pellets were gently resuspended in PEG solution (BDH, mol. wt. 1500, 41.7% in Dulbecco's modified Eagle's medium (DMEM) containing 15% dimethyl sulfoxide) using 2 m1/108 lymphocytes. After 1 min at 37 "C, the suspension was slowly diluted by dropwise addition of 30 ml BSS. Then the cells were pelleted again and resuspended in 5 ml of HAT medium and distributed into 96 wells of tissue culture plates (Costar 3524) which contained 1.5 ml of prewarmed HAT medium. The plates were then incubated at 37 "C. Only hyb- ridoma growth can occur in HAT selective medium, and growth was observed in all (Fusion N) or half (Fusion 0) of the wells 10-14 days later.
2.3 Passage of hybridomas in mice
Hybridoma lines were passaged as solid or ascitic tumors by injecting 5 x lo6 cells subcutaneously (s.c.) or intraperitone- ally (i.p.) into AKR mice. (Both the CBA lymphocytes and BW 5147 cells are of H-2k haplotype.) Growth of S.C. or ascitic tumors, respectively, was observed 10-20 days later, at which time cells and sera were harvested.
2.4 Measurement of DTH and suppressor activity
Mice from CBA, BALBic or C 57 BL/6 strains, and at least 8 weeks old, were used. The mice were sensitized by S.C. injec- tion of 1 X 10' SRBC or chicken red blood cells (CRBC). Five days later, the mice were challenged by injecting 1 X 10' SRBC or CRBC into the right hind footpad. Footpad increase was measured 24 h later with a dial-caliper (Mitutoyo, No. 7309, Mitutoyo Co., Japan). The level of DTH was expressed as percentage of footpad increase.
The culture media from the hybridoma lines (0.5 ml from 105-106 cells/m1/2-3 days) were tested for suppressor activity by injecting i.v. into either (a) sensitized mice (suppression of expression of DTH) or (b) normal mice which were challenged immediately with 1 x 10' SRBC or CRBC S.C. (suppression of induction of DTH). The degree of suppression caused by any of the hybridomas' culture media was calculated as follows: % Suppression = [(DTKontrol - DTH,,,,)/DTH,o,,,o, J x 100. DTHcontrol was the DTH of mice given HAT medium.
0014-2980/79/0707-0572$02.50/0 0 Verlag Chemie, GmbH, D-6940 Weinheim, 1979
Eur. J. Immunol. 1979. 9: 572-575
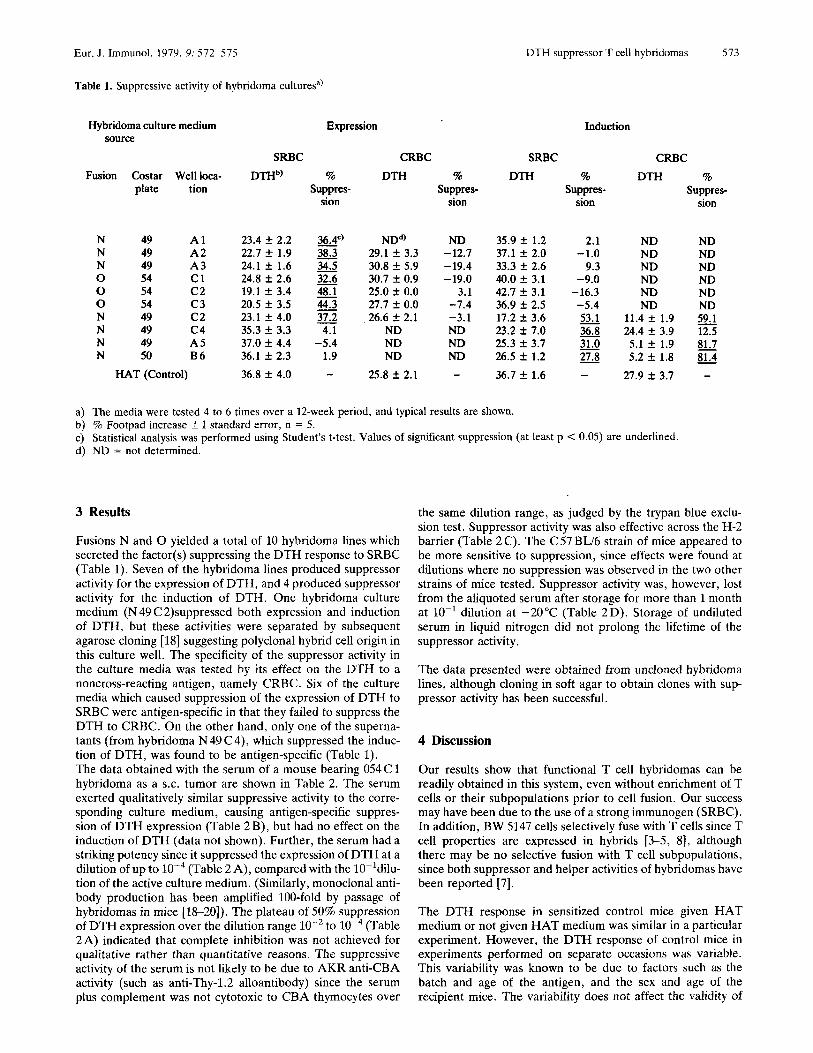
Table 1. Suppressive activity of hybridoma culturesa)
Hybridoma culture medium Expression
DTH suppressor T cell hybridomas 573
Induction source
SRBC CRBC SRBC Fusion Costar Well loca- DTHb) % DTH % DTH %
plate tion Suppres- Suppres- Suppres- sion sion sion
N 49 A 1 23.4 f 2.2 36.4’) N 49 A 2 22.7 f 1.9 38.3 N 49 A 3 24.1 f 1.6 34.5 0 54 c1 24.8 f 2.6 32.6 0 54 c 2 19.1 f 3.4 48.1 0 54 c 3 20.5 f 3.5 44.3 N 49 c 2 23.1 f 4.0 37.2 N 49 c 4 35.3 f 3.3 4.1
N 50 B6 36.1 f 2.3 1.9 N 49 A5 37.0 f 4.4 -5.4
ND*) 29.1 ? 3.3 30.8 f 5.9 30.7 f 0.9 25.0 f 0.0 27.7 f 0.0 26.6 f 2.1
ND ND ND
ND 35.9 f 1.2 -12.7 37.1 f 2.0 -19.4 33.3 f 2.6 -19.0 40.0 f 3.1
3.1 42.7 f 3.1 -7.4 36.9 f 2.5 -3.1 17.2 f 3.6 ND 23.2 f 7.0 ND 25.3 f 3.7 m 26.5 f 1.2
2.1 -1.0
9.3 -9.0
-16.3 -5.4 53.1
31.0 - 27.8
- - 36.8 -
HAT (Control) 36.8 f 4.0 - 25.8 f 2.1 - 36.7 f 1.6 -
CRBC DTH
ND ND ND ND ND ND
11.4 f 1.9 24.4 f 3.9
5.1 f 1.9 5.2 f 1.8
27.9 f 3.7
% Suppres-
sion
ND ND ND ND ND ND 59.1 12.5 - 81.7 81.4
-
- -
a) The media were tested 4 to 6 times over a 12-week period, and typical results are shown. b) % Footpad increase f 1 standard error, n = 5. c) Statistical analysis was performed using Student’s t-test. Values of significant suppression (at least p < 0.05) are underlined. d) ND = not determined.
3 Results
Fusions N and 0 yielded a total of 10 hybridoma lines which secreted the factor(s) suppressing the DTH response to SRBC (Table 1). Seven of the hybridoma lines produced suppressor activity for the expression of DTH, and 4 produced suppressor activity for the induction of DTH. One hybridoma culture medium (N 49 C 2)suppressed both expression and induction of DTH, but these activities were separated by subsequent agarose cloning [18] suggesting polyclonal hybrid cell origin in this culture well. The specificity of the suppressor activity in the culture media was tested by its effect on the DTH to a noncross-reacting antigen, namely CRBC. Six of the culture media which caused suppression of the expression of DTH to SRBC were antigen-specific in that they failed to suppress the DTH to CRBC. On the other hand, only one of the superna- tants (from hybridoma N 49 C4), which suppressed the induc- tion of DTH, was found to be antigen-specific (Table 1). The data obtained with the serum of a mouse bearing 054 C 1 hybridoma as a S.C. tumor are shown in Table 2. The serum exerted qualitatively similar suppressive activity to the corre- sponding culture medium, causing antigen-specific suppres- sion of DTH expression (Table 2 B), but had no effect on the induction of DTH (data not shown). Further, the serum had a striking potency since it suppressed the expression of DTH at a dilution of up to (Table 2A), compared with the IO-ldilu- tion of the active culture medium. (Similarly, monoclonal anti- body production has been amplified 100-fold by passage of hybridomas in mice [18-201). The plateau of 50% suppression of DTH expression over the dilution range lo-’ to W4 (Table 2 A) indicated that complete inhibition was not achieved for qualitative rather than quantitative reasons. The suppressive activity of the serum is not likely to be due to AKR anti-CBA activity (such as anti-Thy-1.2 alloantibody) since the serum plus complement was not cytotoxic to CBA thymocytes over
the same dilution range, as judged by the trypan blue exclu- sion test. Suppressor activity was also effective across the H-2 barrier (Table 2C). The C57BL/6 strain of mice appeared to be more sensitive to suppression, since effects were found at dilutions where no suppression was observed in the two other strains of mice tested. Suppressor activity was, however, lost from the aliquoted serum after storage for more than 1 month at lo-’ dilution at -20°C (Table 2D). Storage of undiluted serum in liquid nitrogen did not prolong the lifetime of the suppressor activity.
The data presented were obtained from uncloned hybridoma lines, although cloning in soft agar to obtain clones with sup- pressor activity has been successful.
4 Discussion
Our results show that functional T cell hybridomas can be readily obtained in this system, even without enrichment of T cells or their subpopulations prior to cell fusion. Our success may have been due to the use of a strong immunogen (SRBC). In addition, BW 5147 cells selectively fuse with T cells since T cell properties are expressed in hybrids [3-5, 81, although there may be no selective fusion with T cell subpopulations, since both suppressor and helper activities of hybridomas have been reported [7].
The DTH response in sensitized control mice given HAT medium or not given HAT medium was similar in a particular experiment. However, the DTH response of control mice in experiments performed on separate occasions was variable. This variability was known to be due to factors such as the batch and age of the antigen, and the sex and age of the recipient mice. The variability does not affect the validity of

574 J. Hewitt and F. Y . Liew
Table 2. Assay of serum from a T hybridoma tumor (054 C 1)-bearing mouse
Immuno- Recipient strain Storage of Serum”) gen (haplotype)
(A) Potency SRBC CBA
(B) Specificity SRBC CBA
CRBC
(C) Different SRBC CBA (H-2’) recipients
BALBk (H-2d)
C 57 BU6 ( H-2b)
(D) Storage SRBC CBA
serum (days)
12
12
20
5
7
30
37
dilution or HAT
HAT 10-2 10-3 1 0 - ~ 10-5 10-6 HAT 10-9 HAT
HAT
HAT
HAT
HAT
HAT
HAT 10-2
HAT
10-3
10-3
10-3
10-3
10-3
10-3
10-4
10-3
the suppressive effect caused by the hybridoma products, which was tested in 4-6 separate experiments, but could affect the extent of suppression computed in an individual experi- ment.
Preliminary immunochemical analysis of the factor suppres- sive for the expression of DTH has been made by analyzing the culture media after absorption or elution from immunosor- bent columns. Experiments so far have established that the factor is devoid of immunoglobulin or heterologous las or I-Js determinants, but bears the homologous Iak and I-Jk determin- ants. Furthermore, concanavalin A columns have absorbed out the factor’s activity suggesting the presence of carbohy- drate determinants. Also, separation of the active diluted serum, described in Table 2, by Amicon membrane filtration indicated that the active component has a molecular weight of less than 50000 (data to be published elsewhere). These prop- erties were found for other suppressor factors for DTH [15], and other T cell factors [9-141.
Further immunochemical, structural and functional analysis of the suppressor factors in the sera of tumor-bearing mice is under study. Should the factor prove to specifically suppress DTH across the species barrier, as has been shown in another system [21], then the generation of suppressor factors to other antigens by the hybridization technique may well be applicable to situations where specific suppression of a cell-mediated response is required.
Eur. J. Immunol. 1979. 9: 572-575
DTH % sup- pression”)
18.6 f 1.4 - 54.4 39.3 50.6
15.2 f 2.8 18.3 17.2 f 2.7 7.6 18.6 f 1.4 - 11.3 f 4.3 -39.3 - 21.5 f 2.5 - 24.7 f 1.9 -14.8 43.7 f 3.2 -
22.0 55.0 f 4.4 -
27.8 28.4 f 2.5 -
52.8 f 5.0 - 35.6 f 3.0 - 32.6 54.4 i 3.4 -
59.2 32.4 f 4.4 -
32.4 28.7 f 3.0 11.4 20.8 f 1.7 24.8 f 3.2 -19.2 HATat -20°C.
8.5 f 4.2 - 11.3 f 4.3 -
- 9.2 f 4.1
34.1 f 1.9 -
39.7 f 8.2 -
13.6 f 1.9 47.8
22.2 f 4.6 -
21.9 k 4.2 - - a) Serum was stored at 10-1 dilution
b) See Table 1, c).
We thank Mr. D. Y. Sia for performing the immunoabsorption experi- ments and Ms. B . Pearce for technical assistance.
Received January 19, 1979
5 References
1 2
3
4
5 6 7
8
9 10 11
Kohler, G. and Milstein, C., Nature 1975. 256: 495. Hammerling, G. J., Reth, M., Lemke, H., Hewitt, J., Melchers, I. and Rajewsky, K., in Peeters, H. (Ed.), XXV Colloquium - Pro- tides of the Biological Fluids, Pergamon Press, London 1978, p. 551. Goldsby, R. A, , Osborne, B. A , , Simpson, E. and Herzenberg, L. A., Nature 1977. 267: 707. Kohler, G . , Lefkovits, I., Elliott, B. and Coutinho, A , , Eur. J . Immunol. 1977. 7: 758. Hammerling, G . J., Eur. J . Immunol. 1977. 7: 743. Taniguchi, M. and Miller, J . F. A. P., J . Exp. Med. 1978.148: 373. Kontianen, S. , Simpson, E., Bohrer, E., Beverley, P. C. L., Her- zenberg, L. A., Fitzpatrick, W. C., Vogt, P., Torano, A., McKen- zie, I. F. C . and Feldmann, M., Nature 1978. 274: 477. Watanabe, T., Kimoto, M., Maruyama, S. , Kishimoto, T. and Yarnarnura, Y . , J . Immunol. 1978. 121: 2113. Feldmann, M., J . Exp. Med. 1972. 136: 731. Munro, A. and Taussig, M. J . , Nature 1975. 256: 103. Tada, T., Taniguchi, M. and Tokushisa, T., in McDevitt, H. 0. (Ed.), Zr Genes and la Antigens, Academic Press, New York 1978, p. 517.

Eur. J. Immunol. 1979.9: 575-578 Lectin affinity fractionation of cytotoxic T lymphocytes 575
12 Takemori, T. and Tada, T., J. Exp. Med. 1975.142: 1241. 13 Kapp, J. A , , Pierce, C. W., Dela Croix, F. and Benacerraf, B., J.
14 Kontianen, S. and Feldmann, M., Eur. J. Immunol. 1977. 7: 417. 15 Liew, F. Y. and Chan-Liew, W. L., Eur. J . Immunol. 1978.8: 168. 16 Liew, F. Y., Eur. J. Immunof. 1977. 7: 714. 17 Littlefield, J. W., Science 1964. 145: 709.
18 Cotton, R. G. H., Secher, D. S. and Milstein, C., Eur. J.
19 Kohler, G. and Milstein, C., Eur. J. Immunol. 1976. 6: 511. 20 Pearson, T., Galfrt, G., Ziegler, A. and Milstein, C., Eur. J.
21 Luzzati, A. L., Taussig, M. J., Meo, T. and Pernis, B., J. Exp.
Immunol. 1973. 3: 135. Immunol. 1976. 116: 305.
Imnunol. 1977. 7: 684.
Med. 1976. 144: 573.
Arthur Kimura, Anders Om, Goran Holmquist, Hans Wigzell and Bo Ersson+
Department of Immunology, University of Uppsala Biomedical Center and Separation Central, University of Uppsala Biomedical Center’, Uppsala
Unique lectin-binding characteristics of cytotoxic T lymphocytes allowing their distinction from natural killer cells and “K” cells*
Cytotoxic T lymphocytes (CTL) can be selectively depleted from in vitro or in vivo alloactivated populations of T cells on Vicia villosa lectin adsorbents through the lectin-specific interaction with the CTLassociated surface glycoprotein T 145 (Kim- ura, A, Wigzell, H. and Holmquist, G., J . Exp. Med. 1979. 149: 473). Results from these and other experiments have demonstrated the general applicability of this frac- tionation procedure in which no constraints related to antigenic specificitiy of the CTL have been observed. When this fractionation procedure was applied to other compartments of cytolytic cells (natural killer cells and “ K ’ cells), no detectable impact could be seen. This differential lectin binding would appear to offer a means of dissecting the activities of CTL from other compartments of cytolytic lymphoid cells.
1 Introduction
The existence of at least three major tissue-destructive compart- ments of lymphoid cells has been extensively documented for a variety of species (see for review [l]). Cytotoxic T lymphocytes (CTL), natural killer cells (NK) and antibody-dependent “K” cells seemingly all contribute towards the maintenance of a versatile cellular immune effector mechanism. Although a great deal of experimental evidence has been reported to distinguish the cytolytic activities of these three categories of cell types as separate entities, the lineage(s) of both NK and K cells have not been firmly established [l]. Considerable effort has been put forth to identify cell surface antigenic systems which would clearly dissociate these three categories of cell types. Unfortu- nately, approaches in which a particular cell type is selected for by the use of antibody and complement elimination pose
[I 23861
* This work was supported by an NIH contract NOI-CB-64033, NIH grant AI-CA-13485-03 and the Swedish Cancer Society.
Correspondence: Arthur Kimura, Department of Immunology, Uni- versity of Uppsala Biomedical Center, Box 582, S-751 23 Uppsala, Sweden
Abbreviations: Con A: Concanavalin A ADCC: Antibody-dependent cellular cytotoxicity CTL: Cytotoxic T lymphocyte(s) D-G~NAc: N- acetyl-D-galactosamine FCS: Fetal calf serum K cell: Naturally occur- ring Fc receptor-positive cytotoxic cells mediating contactual lysis against IgG-coated target cells NK cell: Natural killer cell NP40 Nonidet-P 40 PBS: Phosphate-buffered saline MLC: Mixed lympho- cyte culture
0 Verlag Chemie, GmbH, D-6940 Weinheim, 1979
further problems in the creation of soluble immune complexes which can be shown to alter the activity of cells participating in antibody-dependent cellular cytotoxicity (ADCC) [2].
We have previously described a CTL-associated cell surface glycoprotein (T 145) whose expression and distribution among activated T lymphocytes is restricted to the Ly-1- 2’ popula- tion of alloactivated T blasts, being undetectable on their pre- cursors and other populations of activated T blasts lacking cytolytic function [3]. During the course of purification proce- dures for T 145, a panel of over 20 lectins were analyzed for their quantitative reactivity against various Nonidet-P 40-sol- ubilized, CTL-derived membrane glycoproteins as a first step towards purification. Unexpectedly, one blood group A-reac- tive lectin derived from the seeds of Vicia villosa was found to display an unusually selective binding to T 145 from among the total pool of surface-labeled glycoproteins [4]. When subse- quently applied to cell fractionation studies, Vicia villosa affin- ity adsorbents could be shown to selectively retain those T cells capable of expressing cytolytic activity [4]. In this study, we present further evidence for the discriminatory power of Vicia villosa by the comparison of its reactivity for other cellu- lar compartments of cytolytic activity.
2 Materials and methods
2.1 Animals
Male and female C B N H mice (6-8 weeks of age) used in these studies were raised and maintained in our breeding facilities, Department of Immunology, Uppsala.
0014-2980/79/0707-0575$02.50/0