apoptosis and apoptosis regulating proteins and factors in...
TRANSCRIPT

APOPTOSIS AND APOPTOSIS REGULATING PROTEINS AND FACTORS IN SMALL AND LARGE CELL LUNG CARCINOMA
ANNA-KAISAEEROLA
Department of Pathology
OULU 1999

OULUN YLIOPISTO, OULU 1999
APOPTOSIS AND APOPTOSIS REGULATING PROTEINS AND FACTORS IN SMALL AND LARGE CELL LUNG CARCINOMA
ANNA-KAISA EEROLA
Academic Dissertation to be presented with the assent of the Faculty of Medicine, University of Oulu, for public discussion in the Auditorium of the Department of Pharmacology and Toxicology, on November 5th, 1999, at 12 noon.

Copyright © 1999Oulu University Library, 1999
OULU UNIVERSITY LIBRARYOULU 1999
ALSO AVAILABLE IN PRINTED FORMAT
Manuscript received 23 Sebtember 1999Accepted 30 Sebtember 1999
Communicated by Docent Pekka Klemi Docent Pertti Lipponen
ISBN 951-42-5406-6(URL: http://herkules.oulu.fi/isbn9514254066/)
ISBN 951-42-5405-8ISSN 0355-3221 (URL: http://herkules.oulu.fi/issn03553221/)

To my Mother and Father

This is the way the world endsNot with a bang but a whimper.
T.S Elliot THE HOLLOW MEN, 1925


Eerola, Anna-Kaisa, Apoptosis and apoptosis regulating proteins and factors insmall and large cell lung carcinomaDepartment of Pathology, University of Oulu, FIN-90401, Oulu, Finland1999Oulu, Finland(Manuscript received 23 September 1999)
Abstract
Aptosis denotes a biochemically and morphologically distinct chain of events leading to self-destruction of cell. It is pivotal in the maintenance of tissue homeostasis and also plays a role inneoplasm. In this work, the extent of apoptosis and apoptosis regulating proteins and factors wasstudied in a total of 94 patients operated for lung carcinoma, including 56 small cell lungcarcinomas (SCLC) and 38 large cell lung carcinomas (LCLC). The extent of apoptosis wasdetermined by detecting and counting the relative and absolute numbers of apoptotic cells andbodies using 3 ´- end labelling of the apoptotic DNA. The extent of apoptosis in SCLC wascompared with the cell proliferation activity as determined by Ki-67 immunohistochemistry, withthe volume density of necrosis and with the occurrence of immunohistochemically detectable p53and bcl-2 proteins. In order to test the hypothesis that increased apoptotic activity is connected withneuroendocrine differentiation and with low differentiation degree in LCLC and that it is regulatedby bcl-2 family proteins, the extent of apoptosis and tumour necrosis was analysed in relation to theexpression of bcl-2 family proteins bcl-2, mcl-1, bax and bak. Apoptosis, tumour infiltratinglymphocytes (TILs), and angiogenesis are important factors that contribute to tumour growth. In thepresent study immunohistochemical methods were used to investigate the relationships of thesefactors and their role in the prognosis of the patients with LCLC and SCLC.
A remarkably high apoptotic activity was detected in both SCLC and LCLC. The meanapoptotic index in SCLC was 2.70 % and in LCLC 2.49 %. Exceptionally high proliferation activityand high percentage of tumour necrosis was seen in SCLC. 58 % of SCLC showed more than 40 %of Ki-67 positive nuclei, and tumour necrosis was seen in 83 % of the cases. P53 proteinaccumulation was detected in 38 % and bcl-2 expression in 50 % of SCLC. The extent of apoptosisin SCLC was inversely related to tumour necrosis and p53 protein accumulation. In LCLC, bcl-2expression was detected in 40 % of the cases. It was associated with neuroendocrine differentiationand predicted favourable prognosis of the patients. A high number of T cells and macrophages witha small number of B cells was detected in both SCLC and LCLC. The occurrence of intratumouralcytotoxic CD8 cells was associated with the occurrence of apoptotic bodies in SCLC. The increasednumber of intratumoural T cells, CD8-positive cells and macrophages predicted favourableprognosis of the patients with SCLC. In LCLC, an increased number of B cells and macrophages,but not T cells, was associated with better survival.
In addition to tumour cells, numerous apoptotic bodies could also be found within alveolarmacrophages within and close to tumour tissue. In order to test whether such cells could be found insputum smears and if their presence could be utilised as a marker of malignancy in tumourdiagnosis, the occurrence of alveolar macrophages with apoptotic bodies (AMWABs) was analysedin 84 sputum samples and 13 broncho-alveolar lavage (BAL) specimens from patients with andwithout lung carcinoma. AMWABs could be found in cytological samples of the patients with lungcarcinoma. In sputum specimens, enhanced apoptosis, as measured by an increased number ofAMWABs reflected and was indicative of malignancy. This was also true for cytological specimensof the patients even when the actual malignant cells were not found. Therefore the AMWABsserved as a marker of pulmonary malignancy.
Keywords: small cell lung carcinoma (SCLC), large cell lung carcinoma (LCLC), tumourinfil trating lymphocytes (TILs), alveolar macrophages with apoptotic bodies (AMWABs)


ulu,the
ty toind
y fortingts ofseed to
n thisast
my
eirnnaithErjaul to
inaheing
my
hisloveeathear
rk.
Acknowledgements
The present thesis was carried out at the Department of Pathology, University of Oduring the years 1994-1999. I wish to thank Professor Veli-Pekka Lehto, Head ofDepartment of Pathology and Professor Frej Stenbäck for giving me the opportuniwork at their department. I am most grateful to Professor Veli-Pekka Lehto for his kand supporting attitude towards my work and for revision of this manuscript.
I owe my deepest gratitude to Docent Paavo Pääkkö, the supervisor of this studintroducing me to the field of pulmonary pathology and apoptosis. His everlasoptimism and firm guidance throughout this work has carried me through the momendespair towards new solutions and the delight of discovery. Due to his ability to forethe importance of the concept of apoptosis in tumour pathology I have been privilegetake part in apoptosis research and to witness the enormous progress made ifascinating field of study. I am also grateful to Docent Ylermi Soini for sharing his vknowledge and ideas, for his true interest in apoptosis research and for reviewingpapers and this manuscript.
Docents Pertti Lipponen and Pekka Klemi are gratefully acknowledged for thvaluable remarks and criticism which have improved this thesis. I want to thank AVuolteenaho for correcting my English and Risto Bloigu for his kind and expert help wthe statistics. I am sincerely grateful for the technical assistance of Mirja Vahera,Tomperi, Marja Tolppanen, Tapio Leinonen and Hannu Wäänänen. I am also gratefHilkka Penttilä and Kati Hietala for their help in secretarial matters.
My warmest thanks belong to my co-workers Ulla Näpäkangas, Merja Vakkala, NTurunen and Henni Ruokolainen for all their help during this work. Outside of tdepartment, I want to thank my dearest friend Elina Markkula for her long-lastfriendship, which goes back to our early childhood and her encouragement duringyears of study in Oulu.
I want to thank my life-companion Enzy Tamme for sharing the last five years oflife with me. Many obstacles during this work have been overcome because of yourand understanding. Our many nightly philosophical discussions about life and d(apoptosis) have helped me to widen my scope of life itself. I also want to thank my dsister Elina Armstrong and her family in USA for their love and support during this wo

andou
. I amand
, thetinese
rs
I owe my deepest gratitude to my dear mother and father, Docent Mirja EerolaProfessor Risto Eerola for their love and caring throughout my life. I want to thank yfor being an excellent example as medical doctors and scientists as well as parentsalso most grateful for your everlasting encouragement during my medical studiesespecially during this work. To you I dedicate this work.
This work was supported by the Cancer Societies of Finland and Northern FinlandFinnish Anti-Tuberculosis Association, Finnish Medical Foundation, Ida MonFoundation, Emil Aaltonen Foundation and Finnish Medical Society Duodecim. All thare gratefully acknowledged.
The permission of John Wiley & Sons Limited (I) and Kluwer Academic Publishe(V) to reprint the original articles is acknowledged.
Oulu, September 1999 Anna-Kaisa Eerola

Abbreviations
AC Atypical carcinoid
AMWABs Alveolar macrophage with apoptotic bodies
Apaf-1 Apoptosis protease activating factor 1
ApoI Apoptotic index
Bak Bcl-2 homologous antagonist / killer
BAL Broncho-alveolar lavage
Bax Bcl-2 homologous antagonist x
BCGF B cell growth factor
Bcl-2 B-cell leukaemia 2
BH Bcl-2 homology
bp base pair
CT Carcinoid tumour
DAB Diaminobenzidine
DNA Deoxyribonucleic acid
FADD Fas associating death domain
FasL Fas ligand
FasR Fas receptor
FLICE Fas ligand interacting cell effector
HE Haematoxylin-eosin
HPF High power field
INFλ Interferon-gamma
ISEL In situ end labelling
Kbp Kilo base pair

LCLC Large cell lung carcinoma
LCNEC Large cell neuroendocrine carcinoma
LCNNEC Large cell non-neuroendocrine carcinoma
LT Lymphotoxin
Mcl-1 Myeloid cell leukeamia 1
MD Microvessel density
MGG May-Grünwald-Giemsa
NE Neuroendocrine
NK Natural killer
NSCLC Non-small cell lung carcinoma
NSE Neuron specific enolase
NT Nick translation
PBS Phosphate buffered saline
PCNA Proliferating cell nuclear antigen
PBS Phosphate buffered saline
PS Phosphatidylserine
RIP Receptor interacting protein
SCLC Small cell lung carcinoma
TC Typical carcinoid
TILs Tumour infiltrating lymphocytes
TNF Tumour necrosis factor
TUNEL Terminal dUTP nick end labelling
VEGF Vascular endothelial growth factor

heir
htois
72-
dtion
ncol
lung
nlung
lar6.
List of original papers
This thesis is based mainly on following articles that are referred to in the text by tRoman numerals:
I Eerola A-K, Törmänen U, Rainio P, Sormunen R, Bloigu R, Vähäkangas K, LeV-P, Soini Y & Pääkkö P (1997) Apoptosis in operated small cell lung carcinomainversely related to tumour necrosis and p53 immunoreactivity. J Pathol 181:1177.
II Eerola A-K, Ruokolainen H, Soini Y, Raunio H & Pääkkö P (1999) Accelerateapoptosis and low bcl-2 expression associated with neuroendocrine differentiapredict shortened survival in operated large cell carcinoma of the lung. Pathol ORes. In press.
III Eerola A-K, Soini Y & Pääkkö P (1999) A high number of tumour infiltratinglymphocytes is associated with a favourable prognosis in operated small cellcarcinoma. Submitted.
IV Eerola A-K, Soini Y & Pääkkö P (1999) Tumour infiltrating lymphocytes in relatioto tumour angiogenesis, apoptosis and prognosis in patients with large cellcarcinoma. Lung Cancer. In press.
V Eerola A-K, Soini Y, Lehto V-P & Pääkkö P (1998) Increased numbers of alveomacrophages with apoptotic bodies predict lung carcinoma. Apoptosis 3:261-26


921211123
232345
27782829299303131132323334
3435
35
Contents
AbstractAcknowledgementsAbbreviationsList of original papers1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12. Review of the literature . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
2.1. Apoptosis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .2.1.1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22.1.2. History . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22.1.3. Molecular mechanisms of apoptosis . . . . . . . . . . . . . . . . . . . . . . . . . . .
2.1.3.1. CED genes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .2.1.3.2. Caspases . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .2.1.3.3. Bcl-2 family proteins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22.1.3.4. p53 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
2.1.4. Morphology of apoptosis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .2.1.4.1. Electron microscopy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22.1.4.2. Light microscopy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
2.1.5. Apoptosis and necrosis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .2.1.6. Biochemical features of apoptosis . . . . . . . . . . . . . . . . . . . . . . . . . . . . .2.1.7. Histochemical detection of apoptosis . . . . . . . . . . . . . . . . . . . . . . . . . .
2.1.7.1. In situ end labelling (ISEL) . . . . . . . . . . . . . . . . . . . . . . . . . . . 22.1.8. Histological evaluation of apoptosis . . . . . . . . . . . . . . . . . . . . . . . . . . .
2.2. Lung carcinoma . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .2.2.1. Epidemiology of lung carcinoma . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .2.2.2. Etiology of lung carcinoma . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
2.2.2.1. Tobacco smoking and lung carcinoma . . . . . . . . . . . . . . . . . . .2.2.2.2. Asbestos and lung carcinoma . . . . . . . . . . . . . . . . . . . . . . . . . .
2.2.3. Histological typing of lung tumours . . . . . . . . . . . . . . . . . . . . . . . . . . . 32.2.4. Pulmonary cytology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
2.3. Neuroendocrine lung tumours . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .2.3.1. Small cell lung carcinoma (SCLC) . . . . . . . . . . . . . . . . . . . . . . . . . . . .2.3.2. Large cell neuroendocrine carcinoma (LCNEC) . . . . . . . . . . . . . . . . . .2.3.3. Bronchopulmonary carcinoid tumour (CT) . . . . . . . . . . . . . . . . . . . . . .

66363677378389404041411422433344
4445
55
747484848
949490051515151
253
2.4. Markers of cell proliferation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32.4.1. Ki-67 and PCNA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
2.5. Apoptosis in tumours . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .2.5.1. Apoptosis in lung carcinoma . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
2.6. Tumour immunity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32.6.1. Tumour infiltrating lymphocytes (TILs) . . . . . . . . . . . . . . . . . . . . . . . . 32.6.2. Apoptosis and the immune system . . . . . . . . . . . . . . . . . . . . . . . . . . . .
2.6.2.1. Fas(CD95/APO-1) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32.7. Tumour angiogenesis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
3. Aims of the study . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 34. Materials and methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
4.1. Tumour samples and identification of neuroendocrine differentiation (I-IV) . .4.2. Cytological samples (V) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
4.2.1. Sputum smears . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .4.2.2. BAL samples . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
4.3. Detection of apoptosis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .4.3.1. In situ 3´- end labelling (ISEL) (I-V) . . . . . . . . . . . . . . . . . . . . . . . . . . . 44.3.2. Detection of apoptotic bodies within alveolar macrophages (V) . . . . .4.3.3. Controls of apoptosis (I-IV) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44.3.4. Evaluation of apoptosis in tumours (I-IV) . . . . . . . . . . . . . . . . . . . . . . . 44.3.5. Evaluation of apoptotic bodies within alveolar macrophages (V) . . . . .
4.4. Evaluation of tumour necrosis (I-II) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .4.5. Immunohistochemistry (I-IV) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
4.5.1. p53 protein (I) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44.5.2. Ki-67 (I) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44.5.3. Bcl-2 and related proteins Mcl-1, Bax and Bak (I-II) . . . . . . . . . . . . . . 44.5.4. Tumour infiltrating lymphocytes (TILs) (III-IV) . . . . . . . . . . . . . . . . . 464.5.5. Factor VIII (III-IV) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46
4.6. Western Blotting (II) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44.7. Statistical methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
5. Results . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .5.1. Apoptosis in tumour samples (I-IV) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
5.1.1. Morphological features of apoptosis . . . . . . . . . . . . . . . . . . . . . . . . . . .5.1.2. Extent of apoptosis in SCLC (I, III) and LCLC ( II, IV) . . . . . . . . . . . . 485.1.3. Apoptosis in relation to clinical data (I-II) . . . . . . . . . . . . . . . . . . . . . . 4
5.2. Apoptosis regulating proteins (I-II) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .5.2.1. p53 protein accumulation in SCLC (I) . . . . . . . . . . . . . . . . . . . . . . . . .5.2.2. Bcl-2 protein expression in SCLC and LCLC (I- II) . . . . . . . . . . . . . . . 55.2.3. Bax, Bak and Mcl-1 proteins in LCLC (II) . . . . . . . . . . . . . . . . . . . . . . 5
5.3. Western Blot analysis (II) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .5.4. Tumour necrosis and cell proliferation (I) . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
5.4.1. Ki-67 expression in SCLC . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .5.4.2. Tumour necrosis in SCLC . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
5.5. Tumour infiltrating lymphocytes (TILs) in SCLC and in LCLC (III-IV) . . . . . 525.6. Microvessel density (MD) in SCLC and LCLC (III-IV) . . . . . . . . . . . . . . . . . . 55.7. Parameters as prognostic markers (I-IV) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .

53. 5454
5555
57788595959
6061626236365
656769
5.7.1. Extent of apoptosis as a prognostic marker (II) . . . . . . . . . . . . . . . . . . .5.7.2. Tumour diameter and postoperative treatment as prognostic markers5.7.3. Bcl-2 protein expression as a prognostic marker . . . . . . . . . . . . . . . . . .5.7.4. TILs as prognostic markers (III-IV) . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
5.8. Apoptosis in cytological samples (V) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .5.8.1. AMWABs in sputum samples . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 55.8.2. Specific demonstration of the presence of apoptotic DNA in alveolar
macrophages . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .5.8.3. AMWABs and inflammation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 55.8.4. AMWABs in BAL samples . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 55.8.5. AMWABs in relation to clinical data . . . . . . . . . . . . . . . . . . . . . . . . . . 5
6. Discussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .6.1. Detection of apoptosis in tumours . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .6.2. Extent of apoptosis in malignant tumours . . . . . . . . . . . . . . . . . . . . . . . . . . . . .6.3. Apoptosis in relation to prognosis, cell proliferation, necrosis and age of
the patients . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .6.4. p53 protein accumulation in SCLC . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .6.5. Expression of bcl-2 family proteins in lung carcinoma . . . . . . . . . . . . . . . . . . .
6.5.1. Bcl-2 expression in relation to survival . . . . . . . . . . . . . . . . . . . . . . . . .6.6. TILs in lung carcinoma . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6
6.6.1. The association between TILs and prognosis . . . . . . . . . . . . . . . . . . . .6.7. Tumour angiogenesis in lung carcinoma . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .6.8. Apoptosis in cytological samples . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
7. Summary and Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .8. References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .


both
ourand
rtanttionwthand
imits
self-ofof
rld,994).
vivals torvival987,C)urs.less
ying
iscal
1. Introduction
Apoptosis is a genetically regulated active process abolishing cell populations inphysiological and pathological processes (Kerret al. 1972, reviewed by Wyllie 1997a).Impaired apoptosis, either due to expression of oncogenes or mutations of tumsuppressor genes, leads to an uncontrolled accumulation of malignant cellseventually to formation of cancer (Kerret al. 1994, Carcon & Ribeiro 1993). The growthrate is a principal determinant of the aggressiveness of a tumour and an impoprognostic factor. It has become increasingly clear, however, that not only proliferabut also death of tumour cells by apoptosis is an important determinant of tumour grorate. Tumour angiogenesis has been shown to be essential for tumour growthmetastasis (Folkman 1990) and there is evidence that inhibition of angiogenesis ltumour growth by promoting apoptosis (Holmgrenet al. 1995). Apoptosis also has afundamental role in immune response, such as deletion of immune cells recognisingantigens and also in cytotoxic killing in the tumour microenvironment. The importanceapoptosis is now widely recognised in many fields of medicine rasing a lotexpectations as to a future use in tumour diagnosis and therapy.
Lung carcinoma is the most frequently diagnosed malignancy throughout the wocausing more cancer deaths than all the other types of cancer combined (Hammar 1Compared to many other cancers, there has been relatively little change in surdespite intensive investigation into early detection and trials of new approachesurgical, radiation and adjuvant therapies, as well as chemotherapy. Five-year surate for all cases of lung cancer was 6 % in 1950 to 1954 and 13 % in 1981 to 1showing only a slight improvement (Beckett 1993). Small cell lung carcinoma (SCLand large cell lung carcinoma (LCLC) are the most aggressive types of lung tumoThey are highly associated with cigarette smoking and have a five-year survival ofthan 10 % (Hammar 1994, Traviset al. 1998). Lung cancer mortality in the next centurwill depend primarily on the effectiveness of current efforts to reduce smokprevalence. For improvement of survival of the patients with lung carcinoma itessential to find early detection tools that pick up lung cancer prior to its clinidetection.

20
iationandin
r of
The present study was undertaken to study the extent of apoptosis and its assocto apoptosis regulating proteins, tumour infiltrating lymphocytes (TILs), angiogenesissurvival in surgically resected highly malignant lung tumours, namely SCLC andLCLC. Additionally, the detection of enhanced apoptosis was applied as a markemalignancy in diagnostic respiratory cytology.

essself-rrine-cifictheb).
tivetheding
ed bysed tooxiases:
hasee cell
toticrallyr a
2. Review of the literature
2.1. Apoptosis
2.1.1. Introduction
Apoptosis is an evolutionarily conserved, genetically regulated, active proccharacterised by profound and distinct changes in cellular architecture leading todestruction of cells (Kerret al. 1994, reviewed by White 1996 and Yang & Korsmeye1996). Apoptosis has a pivotal role in sculpting tissues during development, endocdependent atrophy, normal cell turnover in many tissues, the selection of speimmunologically competent subpopulation in both T and B cell lineages duringresponse to antigen, and cytotoxic T lymphocyte killing (for a review, see Wyllie 1997Apoptosis limits the accumulation of potentially harmful cells, such as self-reaclymphocytes, virus-infected cells and tumour cells (reviewed by Reed 1995). Onother hand, extravagant apoptosis contributes to a wide variety of diseases, inclucancer, AIDS, stroke, myopathies and various neurodegenerative disorders (reviewThompson 1995). Furthermore, apoptosis occurs in normal and tumour tissues expolow or moderate doses of chemotherapeutic agents, ionising radiation and hyp(reviewed by Wyllie 1997b). As a process, apoptosis can be divided into three phainduction, effector and degradation (for a review, see Kroemeret al. 1995). In theinduction phase the cell receives apoptosis-triggering signals. During the effector pthese signals activate the apoptotic machinery, and in the degradation phase thpasses the point of no return and finally disperses.
2.1.2. History
Although it is thought that regression of ductus arteriosus after birth, a typical apopphenomenon, was already known to Aristotle, research work concerning natuoccurring cell death did not commence until the beginning of the 19th century (foreview, see Clarke & Clarke 1996 and Cummingset al. 1997). At that time, although

22
ringncept
byewed
thearlyand42,
oads.rend
of&as
d alets”.cell.epteds. Iniew
toers
ong
athdiesed ittheell
in
deen.oo &idd a84)ical
wereNA
ledentshas
ce ofttest
our
known much earlier, the question of the regression of foetal and larval structures dudevelopment was clearly recognised and formulated as a topic of research. The coof cell death became feasible especially due to the formulation of the cell theorySchleiden and Schwann and the advent of microscopes and fixation methods (reviby Clarke & Clarke 1996). In the 19th century, ontogenesis and phylogenesis weremajor progressive fields in medical research and therefore, not surprisingly, epublications of naturally occurring cell death dealt with metamorphosis of insectsamphibia. The first discovery of developmental cell death was made by Vogt in 18when he reported cell death of notochord and adjacent cartilage of metamorphic tLater Vogt abandoned his early interest in the field of cell death setting, however, a tthat was followed by many eminent scientists. The first morphological descriptionnaturally occurring cell death was that of Flemming in 1885 (reviewed by ClarkeClarke 1996). Flemming described dying granulosa cells of rabbit Graafian follicleshaving an ”ill-defined nucleus containing several small, heavily stained lumps, anpale, homogeneous cytoplasm containing what appeared to be fine fat dropFlemming was the first to argue that cell death involved chemical changes within theHe coined the word chromatolysis at the end of the 19th century which became accas a distinct form of cell death corresponding to the currently used term apoptosi1914 German anatomist Ludvig Gräper published a paper ”A new point of vregarding the elimination of cells” with the idea that mechanisms must existcounterbalance mitosis, referring to Flemming ´s chromatolysis. Although Gräppublication was overlooked, the original concept of chromatolysis survived amembryologists like Glücksmann (for a review, see Manjo & Joris 1995).
First evidence of the existence of two morphologically distinct types of cell decame from an Australian pathologist, John Kerr. In 1965, as a result of his Ph. D. stuconcerning hepatocyte atrophy, Kerr recognised a distinct form of cell death and namshrinkage necrosis. In 1972 John Kerr, Alastair Currie and Andrew Wyllie introducedterm apoptosis for morphologically distinctive, active, inherently controlled form of cdeath, complementary to mitosis in the regulation of animal cell populations bothphysiological and pathological conditions (Kerret al. 1972). The word apoptosis wassuggested by James Cormac, professor of classical Greek at the University of AberIt means ”falling off” or ”dropping off” (apó = from, ptósis = a fall), previously used tdescribe the dropping off of the leaves from trees in the autumn (reviewed by MajJoris 1995 and Cummingset al. 1997). In 1976 and 1981, studies on irradiated lymphotissues led to the notion that chromatin broke down into fragments that producetypical, ladder-like pattern. This phenomenon was linked to apoptosis by Wyllie (19and it came to serve as a specific biochemical marker for the distinctive morphologchanges of apoptotic cells. During apoptosis DNA was fragmented into pieces thatmultiples of nucleosomes seen in polyacrylamide gel electrophoresis analysis of D(Yamadaet al. 1981). Since then, exploitation of molecular biological techniques hasto rapid advances in knowledge about apoptosis, one of the most exciting developmin modern biology. Discovery of cellular oncogenes and tumour suppressor geneshelped to focus research on cell proliferation and growth suppression. The importanapoptosis is now widely recognised in many fields of medicine and it is one of the hofields in biomedical research rasing a lot of expectations as to a future use in tumdiagnosis and therapy.

23
eticg theEllis
itivevitzf C.
n theighlyals.C.
tingand
andld
cellhlyion ofns
istosisof
ptosis
based
ticals can
duced0 Kdctive
ins,A-gicalure of
2.1.3. Molecular mechanisms of apoptosis
2.1.3.1. CED genes
Much of the current knowledge about apoptosis regulation comes from the genstudies of nematode Caenorhabditis elegans. Of the 1090 somatic cells formed durindevelopment of an adult hermaphrodite, 131 die through apoptosis (for a review, seeet al. 1991). The most important of the 11 identified genes of C. elegans are posregulators of apoptosis CED3, CED4 and antiapoptotic CED9 (Hengartner & Hor1994). The death-suppressing activity of CED9 is essential for the development oelegans and, in the case of CED9 mutation and inactivation, the animal dies early idevelopment. The apoptotic program delineated in C. elegans seems to be hconserved in evolution, and homologous genes have been identified in mammMammalian proto-oncogene bcl-2 shows striking functional and structural similarity toelegans CED9. Furthermore, bcl-2 can partially substitute for CED9 in prevenapoptosis in C. elegans (Herngartner & Horvitz 1994). The activities of both CED3CED4 are essential for cell death occurring in the development C. elegans. Mutationinactivation of either CED3 or CED4 leads to the survival of all 131 cells that wounormally die. If both CED3 and CED9 are inactivated, neither the normal nor extradeaths occur (Hengartner & Horviz 1994). CED3 has been shown to be highomologous to human cystein proteases caspases, which are crucial in the executapoptosis (Zouet al. 1997). The genetic studies indicate that CED4 functiodownstream of CED9 but upstream of CED3 (Shaham & Horviz 1996). Thereaccumulating evidence suggesting that CED4 might be homologous to human apopactivating factor 1 (Apaf-1), which participates in cytocrome c dependent activationcaspase-3 (Zouet al.1997).
2.1.3.2. Caspases
The caspase family of cystein proteases functions as fundamental effectors in apo(for a review, see Thornberry & Lazebnik 1998 and Nuñezet al. 1998). At present, 13caspases have been identified in mammals, which can be divided into two classeson the lengths of their N-terminal prodomains (Humkeet al. 1998). Caspases normallyexist in cells as inactive proenzymes and need to be activated through proteolyprocessing in order to trigger apoptosis (Hengartner 1998). The activation of caspasebe achieved by at least two mechanisms, i.e. by caspase cascade or by proximity-inmechanism (Herngartner 1998). This activation cleaves the molecule to 10 and 2subunits, which heterodimerize and associate into tetramers that constitute the aenzyme (Harveyet al.1997). Caspase activity results in cleavage of cytoskeletal protedisruption of the nuclear membrane, disruption of cell-cell contact and DNfragmentation. These irreversible proteolytic events are responsible for the morpholochanges of apoptotic cells and the DNA agarose gel ladder pattern that is the signatapoptosis.

24
ialses inandypesactornti-toticdad,
l-2ch
ally
sts
thebaxion
nesh as
clearof
1) isase-3on ofsticat istive
heasecell
ssalso
beto
cl-2st and
n-
2.1.3.3. Bcl-2 family proteins
The bcl-2 family proteins, of which 16 are currently known in humans, are crucregulators of apoptosis (for a review, see Reed 1998). They act upstream of caspathe apoptosis pathway, either inhibiting or promoting cell death in a largely cell-typestimulus-dependent manner (reviewed by Chao & Korsmeyer 1998). In some cell tapoptosis triggered by Fas/CD95 and certain members of the tumour necrosis f(TNF) family of death receptors is not blocked by over-expression of bcl-2 or other aapoptotic members of the family (for a review, see Reed 1998). Human anti-apopproteins of the bcl-2 family include bcl-2, bcl-xL, mcl-1, bcl-w, A1, Boo anproapoptotic members bax, bak, bok/mtd, bcl-xS, bik/nbk, hrk/dp5, bim/bod, blk, bbid (reviewed by Reed 1998, Yang & Korsmeyer 1996, Kroemer 1997, O´Connoret al.1998, Songet al. 1999). The bcl-2 family proteins possess variable numbers of bchomology (BH) regions (BH1-BH4), which determine their capacity to interact with eaother or with other unrelated proteins (Oltvaiet al. 1993). The proportions ofhomodimers and heterodimers that bcl-2 family proteins can form with each other findetermine the fate of a cell to either survive or die (Oltvaiet al.1993).
The ability of bcl-2 family proteins to form channels in artificial membranes suggethat they may regulate apoptosis by influencing ion or protein transport (Liuet al. 1996).Bcl-2 group proteins resides on the outer mitochondrial membrane oriented towardcytosol, while bax protein resides in the cytosol. Upon resieving apoptotic signalproteins migrate and bind to the mitochondrial membrane inducing loss of selectivepermeability. Channels formed by bcl-2 family proteins in mitochondrial membracontribute to calcium fluxes and the release of apoptosis-promoting agents succytochrome c and apoptotis inducing factor (AIF) (Liuet al. 1996, Yanget al.1997). AIFmoves directly to the nucleus, where it produces chromtin condensation and nufragmentation, while cytosolic cytochrome c sets in motion the terminal eventsapoptosis. Binding of cytochrome c to apoptosis protease activating factor-1 (Apaf-necessary for the activation of procaspase-9, which in turn induces procasepresponsible for the cytological characteristic changes of apoptosis. Altered expressibcl-2 family proteins is common in a variety of human cancers, contributing to neoplacell accumulation by suppressing apoptosis and extending tumour cell life span. Whmore, the ultimate effect of most anticancer drugs is profoundly influenced by the relalevels and state of activation of bcl-2 family members (reviewed by Reed 1998).
The first member of this family, proto-oncogene bcl-2, was originally found at tbreakpoints of t(14;18) chromosomal translocation in low-grade B cell lymphom(Tsujimotoet al. 1984, Bakhshiet al. 1985). The bcl-2 gene was thought to be a uniquoncogene that contributed to cell expansion through failed cell death rather than rapiddivision (Tsujimotoet al. 1984). However, it is now known that many cancers exprehigh levels of bcl-2 without evident gene alterations, suggesting that other pathwayscontribute to bcl-2 expression (Packham 1998). The bcl-2 protein seems tomultifunctional since it is able to heterodimerize its pro-apoptotic relative bax, to bindnon-homologous proteins and to form ion-channels (reviewed by Reed 1998). Bprotein expression has been associated with an enhanced growth rate in lung, breaurinary bladder tumours, probably due to a blocked cell death (Joensuuet al. 1994, Jianget al. 1996, Kinget al. 1996). In lung carcinoma, bcl-2 is expressed in 8 to 30 % of no

25
ns. ALC
tosis
of aratingx andleevel in
and
axvarybax-
omalyy cell
marilyas a53wn inver,
gLC
ofisms:
small cell lung carcinoma (NSCLC) and up to 90 % in SCLC (Ikegakiet al. 1994,Higashiyamaet al. 1995, Fleminget al. 1998). A high level of bcl-2 expression haspreviously been detected in SCLC cell lines (Ikegakiet al. 1994). Moreover, higher bcl-2protein expression has been detected in SCLC compared to that of NSCLCin vivo(Pezzellaet al. 1993, Wanget al. 1998), suggesting that the bcl-2 protein may play aimportant role in the development and progression of neuroendocrine lung tumourhigh level of bcl-2 protein is suggested to be linked to multidrug resistance in SCpatients (Wanget al. 1998).
Bax (Bcl-2 homologous antagonist x) is a bcl-2 related protein that promotes apopand acts as a tumour suppressor (Yinet al. 1997). Bax is thought to be a downstreamtranscription target of p53 and may thus play a part in p53 apoptotic pathway (Yinet al.1997). There is evidence that the ratio of bcl-2 to bax determines the susceptibilitycell to apoptosis. When overexpressed, bax forms homodimers, thereby acceleapoptosis. In contrast, when bcl-2 is expressed in excess, it heterodimerises with baapoptosis is suppressed (Oltvaiet al. 1993). Consistently with its tumour suppressor roin human cancer, bax has been shown to be mutated or expressed at a reduced lseveral human cancers, including colon and (Rampinoet al. 1997) breast cancer(Krajewski et al. 1995a) as well as haematopoietic malignancies (Brimmelet al. 1998).The expression of bax is predominant in large cell neuroendocrine lung carcinomainversely associated with bcl-2 expression (Brambillaet al. 1996).
Mcl-1 (Myeloid cell leukaemia 1) is an anti-apoptotic protein which can bind to band suppress bax-induced cytotoxicity. Transfection of mcl-1 into Chinese hamster ocells partially blocks myc-induced apoptosis, and mcl-1 has been shown to blockmediated apoptosis in a yeast two-hybrid system (Reynoldset al. 1994, Satoet al. 1994).In solid tumours, mcl-1 has been expressed widely in high grade prostate carcin(Krajewskaet al. 1996). Bak (bcl-2 homologous antagonist/killer), like bax, primaripromotes apoptosis, and it has been suggested that its function may be mediated bdeath inhibitory factors, particularly in cell types with a long life span (Chittendenet al.1995, Kieferet al. 1995).
2.1.3.4. p53
The p53 tumour suppressor gene encodes a 53 kDa phosphoprotein that resides priin the nucleus, binds to specific DNA sequences, and functions at least partlytranscriptional regulator (for a review, see Vogelstein & Kinzler 1992). Loss of ptumour suppressor gene function represents the most common genetic lesion knohuman cancer, occurring in 50-55 % of all tumours including colon, lung, breast, libone, brain and haematological malignancies (reviewed by Hollsteinet al. 1991,Vogelstein & Kinzler 1992, Greenblattet al. 1994). The mutational spectra of p53 in luncancers differ among histological types, with highest prevalence (70 %) in SC(D´Amico et al. 1992, Lohmannet al. 1993). In 47 % of NSCLC p53 mutations havebeen detected, the ratios being 65 % squamous, 60 % large cell, and 33 %adenocarcinomas. P53 contributes to tumour suppression by at least two mechan

26
ell
h as
age
en
olled
Thisp53-
ple,e
esed
-givethe
in-
own-
y bein
c and,this
ucedve no
elyild-
by
f-lifeation
p53
through arrest of cell proliferation (reviewed by Hartwell 1992) and induction of cdeath through apoptosis (Yonish-Rouachet al. 1991, Shawet al. 1992). For thesefunctions p53 is known as ”guardian of the genome” (Lane 1992).
P53 plays a role in triggering apoptosis under several physiological conditions, suchypoxia (Graeberet al. 1996), and in response to UV irradiation (Ziegleret al. 1994).Many pathological conditions induce p53 dependent apoptosis, including DNA dam(Liebermannet al. 1995), asbestos exposure in vitro (Pääkköet al. 1996), adenovirusE1A expression (Debbas & White 1993), withdrawal of growth factors (Gottlieb & Or1994) and expression of mitogenic oncogenes such as c-myc (Wagneret al. 1994).Depending on cell type and external signals, c- myc is able to induce either uncontrproliferation or to activate p53 to promote apoptosis (Wagneret al. 1994). Myc and othermitogenic oncogenes activate p53 through another tumour suppressor, p19/ARF.pathway is disabled in most human cancers, implying that an oncogene-activateddependent apoptosis pathway contributes to tumour suppression (Kamijoet al. 1997,Bateset al. 1998). However, not all apoptotic events are p53-mediated. For examthymocytes from p53 -/- mice die of apoptosis when exposed to glucocorticoids (Lowetal. 1993).
How activated p53 promotes apoptosis is still not known, but it probably involvdownregulation of bcl-2 and upregulation of its family member bax (Miyashita & Re1995), p53-inducing genes known as PIGs (Polyaket al. 1997), or signalling through fas-related pathways (Owen-Schaubet al. 1995, Bennettet al. 1998). P53 acts as a tumoursuppressor arresting cells in the G0/G1 phase whenever DNA is damaged in order tomore time for the cell´s DNA repair mechanism to function. This takes place throughinduction of CIP/WAF1/p21, a protein which prevents phosphorylation of cycldependent kinases (El-Deiryet al. 1993, Harperet al. 1993). If the DNA repair isunsuccessful, p53 triggers apoptosis by upregulating the apoptosis-inducer bax and dregulating the anti-apoptotic bcl-2, leading cells to apoptotic death (Miyashitaet al. 1994,Miyashita & Reed 1995). Also caspases, downstream executors of apoptosis, mainvolved in p53 mediated apoptosis. It seems that immediate effectors of p53apoptosis, such as bax, target the mitochondria, leading to the release of cytochromeactivation of caspase-9 (Casp9) (Soengaset al. 1999). In the presence of cytochrome cCasp9 associates with a specific adaptor molecule, Apaf-1. Oligomerization ofcomplex can activate a caspase cascade leading to apoptotic cell death (Zouet al. 1997).It has been shown, however, that caspase inhibitors do not prevent myc- or bax -indapoptosis, implying that caspases act too late in these death programmes and hasubstantial effect on long-term survival (McCarthyet al. 1997).
Normally, the p53 protein is kept at a low concentration in a cell due to its relativshort half-life, which is about 20 minutes (for a review, see Levine 1991). Unstable wtype p53 protein does not accumulate in nuclei and thus it is not detectableimmunohistochemistry (Iggo et al. 1990). It becomes detectable byimmunohistochemistry in tumour cells as a consequence of lengthening of the haland accumulation which is due to mutational events in the p53 gene or due inactivthrough formation of complex with cellular or viral oncoproteins (Iggoet al. 1990,Momandet al. 1992). It has been shown that more than 80 % of the mutations in thegene produce a stabilised protein (Harris & Hollstein 1993).

27
of
3
in53
d in
lungitive
a
n ofe (fores incell
ch asCell
and
ofwithces toelles
ed byell aspeartiontosising
Immunohistochemical studies have shown p53 protein accumulation in 40-88 %SCLCs and 40-60 % of NSCLC (Nuorvaet al. 1994, Brambillaet al. 1996, Nishioet al.1996, Konishiet al. 1997, Wanget al. 1998). In neuroendocrine lung tumours, p5protein accumulation has been linked to a more malignant phenotype (Prqygodzkiet al.1996, Wanget al. 1998). A low level of 53 expression (up to 21 %) has been detectedtypical carcinoid tumours, while SCLCs express generally a high level of pimmunoreactivity (up to 88 %) (Prqygodzkiet al.1996, Lohmannet al.1993, Wanget al.1998). In SCLC, a higher level of p53 protein accumulation has been detecteintermediate cell type of SCLC compared to oat cell type SCLC (Korkolopoulouet al.1993). The relationship between p53 protein accumulation and prognosis incarcinoma is controversial. In NSCLC some authors have reported that p53 posimmunoreactivity in tumour cells is a predictor of poor prognosis (Quinlandet al. 1992,Fujino et al. 1995, Törmänenet al. 1995, Ishidaet al. 1997), while others have reportedthat no correlation exists between p53 protein expression and prognosis (McLarenet al.1992, Brambillaet al. 1996). Leeet al. (1995) have reported that high p53 expression isfavourable prognostic factor in NSCLC.
2.1.4. Morphology of apoptosis
2.1.4.1. Electron microscopy
The earliest detectable ultrastructural change in apoptosis is the condensatiochromatin to form uniformly dense, crescentic masses that abut the nuclear envelopa review, see Cummingset al. 1997). This is followed by nuclear changes including thdispersal of peripheral nuclear chromatin to form aggregates of osmiophilic granulethe centre of the nucleus. Simultaneously with the nuclear changes the apoptoticseparates from its neighbours, usually with loss of special membrane structures, sumicrovilli and desmosomes, and undergoes a period of budding and contortion.volume decreases, cell density increases, cytoplasmic organelles compact,convolution of the cellular and nuclear outline is evident (Kerret al. 1994, Cummingsetal. 1997). At the same time, cytoplasmic changes occur, including aggregationcytoskeletal filaments and clumping of ribosomal particles. Closely associatednuclear and cytoplasmic changes, cells form extensive surface blebs and protuberanproduce membrane-bound apoptotic bodies with well-preserved cytoplasmic organ(Kerr et al. 1994, reviewed by Cummingset al. 1997 and Wyllieet al. 1997b). Apoptoticbodies are dense particles which represent apoptotic remants of cell phagocytostheir viable neighbours or by specialist phagocytes. In tumours, macrophages as wtumour cells themselves are involved in this phagocytosis. The dying cells disaprapidly from the tissue without any inflammatory reaction. The absence of inflammais a crucial feature, since it permits cell death without damage to adjacent cells. Apopis completed within a few hours at most, the majority of which is spent undergodegradation within the phagocytic cell (Kerret al. 1994, reviewed by Wyllie 1997b andCummingset al. 1997).

28
icalcell
icibleeren are
thoutll asnes
is in
ich is
ereasiablelicmatinage,
omalcell
ellsinand
that
cing). Inareas,
omecatedn be
2.1.4.2. Light microscopy
The detection of apoptotic cells by using light microscopy is based on morphologfeatures including condensation of chromatin and cytoplasm, fragmentation of theand apoptotic body formation (Kerret al. 1972). The budding phenomenon of apoptotcells lasts only a few minutes, but the formation of apoptotic bodies remains vishistologically for 1 to 2 hours (Barres 1992, Coles 1993). Thus, in tissues whapoptosis affects scattered cells, apoptotic bodies of various stages of degeneratiodetected. Histologically, apoptotic bodies appear as round or oval masses with or wibasophilic nuclear material. They vary considerably in size; apoptotic bodies as sma0.5 µm in diameter can be readily detected by light microscopy, while the smallest oremain undetectable (reviewed by Cummingset al. 1997). Short half-life and small sizeof apoptotic bodies indicates that even a small increase in the proportion of apoptostissues can represent considerable cumulative cell loss (Howieet al. 1994).
2.1.5. Apoptosis and necrosis
In contrast to apoptosis, necrosis is considered a passive event, the nature of whlargely dependent of the type of the external injurious agent (Kerret al. 1994). Intumours, necrosis leads to destruction of a large group of cells in the same area, whindividual cells undergoing apoptosis are observed scattered throughout the vtumour tissue (Kerret al. 1994). In contrast to apoptosis, necrotic cells swell, cytosoas well as nuclear structures alter, but the general disposition of hetero- and euchrois maintained (reviewed by Wyllie 1997b). In necrosis, because of membrane damcytosolic material leaks into the extracellular space, leading to a release of lysosenzymes and to inflammation, whereas in apoptosis the outer membrane of theremains intact and no inflammation reaction is provoked, leaving the neighbouring cand tissues unharmed (Kerret al. 1994). Moreover, the distinct morphological featuresapoptosis, i.e. chromatin margination, nuclear fragmentation, cellular shrinkingformation of apoptotic bodies do not take place in necrosis (Kerret al. 1994).
Although previously considered unrelated, there is now accumulating evidenceapoptosis and necrosis are related phenomena.In vivo, certain stimuli can induce bothapoptosis and necrosis (Leist & Nicotera 1997). Furthermore, certain apoptosis-industimuli can bring about necrosis when present in high doses (Leist & Nicotera 1997tumours, an increased number of apoptotic cells are seen adjacent to necroticsuggesting that apoptosis and necrosis are related phenomena alsoin vivo (Arai &Katayama 1997). Not only some of the death-inducing mechanisms but also ssignalling pathways seem to be shared, namely, caspases 8 and 10, which are loupstream of the apoptotic signalling pathway. They can provoke necrosis and this cainhibited by anti-apoptotic bcl-2 (Leist & Nicotera 1997).

29
clearof
llieKbpquent
o be
en, lossfor aaat ther cell
leteele bygespointnt of
ingleakselled
nto
or
he
theours
2.1.6. Biochemical features of apoptosis
The characteristic biochemical feature of apoptosis is double-strand cleavage of nuDNA at the linker regions between nucleosomes, leading to a productionoligonucleosomal fragments which are multiples of units comprising 180-200 bp (Wy1980). Internucleosomal cleavage is preceded by cleavage of DNA into 300- or 50fragments, which occurrs in at least some cases where there is no subsedevelopment of oligonucleosomes (Oberhammeret al. 1993). However, the cleavage mayoccasionally be delayed or absent in cell death that by other criteria appears tapoptotic (Cohenet al. 1992, Oberhammeret al. 1993).
Viable cells maintain an asymmetric distribution of different phospholipids betwethe inner and outer leaflets of the plasma membrane. In apoptosis, on the other handof this plasma membrane asymmetry is an early event independent of the cell type (review, see van Engelandet al. 1998). In the presence of calcium, loss of plasmmembrane asymmetry results in the exposure of phosphatidylserine (PS) residuesouter plasma membrane leaflet. At present, the molecular machinery responsible fosurface exposure of PS remains unidentified.
2.1.7. Histochemical detection of apoptosis
The defining characteristic of apoptosis is DNA fragmentation associated with a compchange in cellular morphology (Kerret al. 1972). The morphological changes in cells armanifested in routinely stained sections, such as in tumours, and are thus detectablight and electron microscopy. However, the short duration of morphological chanduring apoptosis and low frequency of apoptotic cells present in tissues at one timehas led to the development of more sensitive techniques for the measuremeapoptosis.
2.1.7.1. In situ end labelling (ISEL)
In situ end labelling technique (ISEL) enables detection of apoptosis in tissues at scell level making use of the newly generated free ends of DNA. The DNA strand breare detected either enzymatically, using terminal deoxytransferase to add labnucleotides to the DNA 3´- OH termini (TUNEL) (Gavrieliet al. 1992), or by using E.coli polymerase I or its Klenow fragment to incorporate labelled nucleotides ifragmented DNA by nick translation (NT) (Wijsmanet al. 1993). Nucleotides that bind toDNA fragments can be labelled radioactively and detected by autoradiographynonradioactively and detected by using the appropriate chromogen reaction.
The maximal intensity of labelling of apoptotic cells is generally higher with tTUNEL method than in NT, TUNEL being thus more sensitive (Gorczycaet al. 1993).Furthermore, digoxigenin-labelled nucleotide (dUTP) incorporation is more rapid inTUNEL method; a 30-min incubation being adequate, compared with several h

30
ith
omage
totic
nd-I).ourten
s insamemay
ssueiencythen ofore,
odiesr ofttenthehen
y beship
e ofugh
g ofplaint 20
tionsand
required for the NT assay to maximise labelling (Gorczycaet al. 1993). Thein situ endlabelling (ISEL) technique detects practically all DNA strand breaks, i.e. also cells wDNA damage, autolytic and necrotic cells are labelled (Wijsmanet al. 1993, Grasil-Krauppet al. 1995). However, necrotic cells can be separated by their morphology frapoptotic cells which occur sporadically and lie scattered in tissues. The DNA daminduced during fixation and embedding of the tissue can lead to labelling of non-apopcells (Wijsmanet al.1993).
2.1.8. Histological evaluation of apoptosis
In histological tumour material, the extent of apoptosis is detected by using the DNA elabelling technique or by plain morphology, and given as apoptotic index (ApoApoptotic index can be defined as percentage of apoptotic cells and bodies of all tumcells per high power field (HPF), or as a number of apoptotic cells and bodies perHPFs (Törmänenet al. 1995, Lipponenet al. 1994a). Even though apoptotic index isused widely, there are no uniform criteria for measuring the extent of apoptositumours, and the results between different authors vary a great deal even amongtumour type. There are a number of technical and methodological factors thatexplain these differences.
The duration of the apoptotic process is largely unknown and it is dependent on tiand cell type (Potten 1996). The extent of apoptosis depends essentially on the efficwith which small fragments can be detected, i.e. the sensitivity and specificity ofdetection procedure which depends on pretreatment, the type and concentratiolabelling enzyme, method of tissue processing and the incubation times. Furthermbecause a dying cell breaks up into several apoptotic bodies, a score of apoptotic bdoes not equal a cell death score. It is therefore more reliable to count a clusteapoptotic bodies likely originating from the same dying cell as one death event (Po1996). Other factors that largely affect apoptotic indices in different tissues aremechanisms and efficiency of removal of the apoptotic fragments. Furthermore, wdetecting apoptosis in tumours, apoptotic macrophages and lymphocytes mamisinterpreted as neoplastic apoptotic cells, giving a distorted view of the relationbetween apoptosis and tumour growth (reviewed by Soiniet al. 1998a).
There are great differences in the rate of cell proliferation as well as in the ratapoptosis at different locations within one tumour. It is thus essential to include enofields and to use a high power lens in the analysis (reviewed by Soiniet al. 1998a). Thereare only few methodological studies that evaluate sources of error in the countinapoptotic cells in tissue sections. When detecting apoptotic cells was based onmorhology a good reproducibility was found when counting apoptotic cells in at leasmicroscopic fields at 630 x magnification (Wijsmanet al. 1993, van de Schepopet al.1996). The most reliable technique in detecting the extent of apoptosis in tissue secwould most likely be combining the two techniques based on apoptotic morphologyspecific labelling of apoptotic DNA.

31
gistry94).mostThe
d 440f thebut
sameased
rlier,ease
omall cell
vivals tote forram981nalwill
oreherherc andintot and
2.2. Lung carcinoma
2.2.1. Epidemiology of lung carcinoma
Lung carcinoma is the leading cause of cancer death in Finland (Finnish Cancer Re1995) and it is the most common type of tumour throughout the world (Hammar 19In 1995 lung carcinoma was the second most common cancer in males and the fifthcommon cancer in females, responsible for 20 % of all cancer deaths in Finland.number of new lung cancer cases diagnosed in Finland in 1997 was 1390 male anfemale cases (Finnish Cancer Registry, unpublished data 1999). From the middle o1970s the incidence of lung cancer in Finland has declined moderately in malesincreased continuously in females. According to two prospective studies done in thegeographical area in northern Finland, the incidence of lung cancer has decresignificantly among males, (from 87 to 63 per 100 000) compared with 20 years eabut increased among females (from 4.1 to 9.5 per 100 000), mainly owing to an incrin lung adenocarcinoma (Huhtiet al. 1980, Mäkitaroet al. 1999). The main histologicaltypes of lung cancer diagnosed in Finland in 1985-1994 were squamous cell carcin(males 33 %, females 17 %), adenocarcinoma (males 16 %, females 28 %) and smacarcinoma (males 16 %, females 16 %) (Dickmanet al. 1999).
Compared to many other cancers, there has been relatively little change in surdespite intensive investigation into early detection and trials of new approachesurgical, radiation, and adjuvant therapies, and chemotherapy. The 5-year survival raall cases of lung cancer in the Surveillance, Epidemiology and End Results prog(SEER) of the National Cancer Institute was 6 % from 1950 to 1954 and 13 % from 1to 1987, showing some improvement in lung cancer survival. Even with additiointerventions to reduce the smoking rate, it is suggested that lung cancer mortalitycontinue to rise through the end of the century.
2.2.2. Etiology of lung carcinoma
The malignant transformation of bronchial cells requires the occurrence of two or mcellular events which include disruptions of intranuclear genetic material or otelements of the cell regulatory system by ionising radiation, viral, chemical, or otphysical factors. Conditions associated with these causative events include genetienvironmental risk factors. The environmental factors can be further subdividedthose encountered through smoking, occupation, domestic and outdoor environmendiet.

32
g for
al tocer- to
oker2- to
ithex-nderossses can
king-p53
mayas
ahas
estosestos-ss
of
osure
f allenceound
2.2.2.1. Tobacco smoking and lung carcinoma
Tobacco smoking is the leading preventable cause of cancer mortality, accountin40 % of cancer deaths in men and 20 % in women (Greenblattet al. 1994). Severalepidemiological studies have confirmed a dose-response relationship proportionduration and amount of smoking (Guyatt & Newhouse 1985). The risk of lung candeclines from an increased risk of approximately 15-fold for current smokers to a 1.54-fold risk for lung cancer 15 years after stopping smoking compared to a nonsm(reviewed by Garfinkel & Silverberg 1991). Passive smoking has shown to cause 1.1.5-fold increased risk of lung carcinoma (reviewed by Fielding & Phenow 1988).
Different histological types of lung carcinoma have varying associations wsmoking. Namely, more than 95 per cent of patients with SCLC are current orsmokers, compared to 80 per cent of patients with adenocarcinoma (Morabia & Wy1991, McDuffieet al. 1990). The histologic changes due to cigarette smoking include lof cilia, cellular atypia, and proliferation of the bronchial epithelium. In addition to thechanges and hypoxia the dispersed neuroendocrine cells and neuroepithelial bodieincrease in number and may contribute to the origin of SCLC (Gosneyet al. 1988).Mutations of p53 tumour suppressor have been shown to be very common in smoassociated lung carcinoma (Harris & Hollstein 1993). G:C to T:A transversions of thetumour suppressor gene occur more frequently in smokers than in non-smokers andbe result of specific carcinogenic agents present in tobacco smoke, suchbenzo(a)pyrene (Greenblattet al. 1994, Rämetet al. 1995, Denissenkoet al. 1996).
2.2.2.2. Asbestos and lung carcinoma
The most frequent occupational cause of cancer is past exposure to asbestos (Okset al.1997, Steenlandet al. 1996). Furthermore, interaction between smoking and asbestosproduced a very high risk for lung cancer in those with both heavy smoke and asbexposures, estimated to be 50 times greater than that of the non-smoking, non-asbexposed male population (Steenlandet al. 1996). Asbestos can cause chromosome loand deletion which can lead to accumulation of proto-oncogenes or inactivationtumour suppressor genes (Barrettet al. 1989). In lung carcinoma, frequency of p53protein accumulation has been associated with clinical and histological asbestos expas well as with heavy tobacco smoking (Nuorvaet al. 1994). Lung cancer mortality ofFinnish men with diagnosed asbestosis in 1980 was 35 %, compared to 10.8 % odeaths among Finnish men (Huuskonen in 1995). It is estimated that the peak incidof asbestos-induced cancer will increase for 15-20 years, reaching its maximum ar2010 in Finland (Huuskonenet al. 1995).

33
asonrld.nentare
) onant, 1c)mall, 3b)olid
3f)ith4d)5)
sisiblechialder
s ofand
cer
yieldromthethat
hodowingf lung
theults,ng
2.2.3. Histological typing of lung tumours
The third edition of WHO's Histological Typing of Lung and Pleural Tumours wpublished in 1999 (Traviset al. 1999). The definitions of the tumour types are basedlight microscopy in order to achieve the widest application throughout the woCommon lung neoplasms are classified according to the best-differentiated compoand graded by the most poorly differentiated component. Malignant lung tumoursclassified into two groups, namely SCLC and non-small cell lung carcinoma (NSCLCthe basis of their histological features. The five major histological types of malignlung neoplasms are 1) squamous cell carcinoma with its 1a) papillary, 1b) clear cellsmall cell and 1d) basaloid variants, 2) small cell carcinoma with its 2a) combined scell variant, 3) adenocarcinoma with its subtypes of 3a) acinar adenocarcinomapapillary adenocarcinoma, 3c) bronchiolo-alveolar adenocarcinoma, 3d) sadenocarcinoma with mucin, 3e) adenocarcinoma with mixed subtypes andadenocarcinoma variants, 4) large cell carcinoma with its variants 4a) LCNEC wcombined subtype, 4b) basaloid carcinoma, 4c) lymphoepithelioma-like carcinoma,clear cell carcinoma, 4e) large cell carcinoma with rhabdoid phenotype, andadenosquamous carcinoma.
2.2.4. Pulmonary cytology
Currently, flexible fiberoptic bronchoscopy has an important role in both the diagnoand staging of lung carcinoma. However, the diagnosis of non-endoscopically visperipheral lesions requires a combination of multiple techniques such as transbronbiopsy, bronchial washing, brushing or transbronchial needle aspiration unfluoroscopic guidance to achieve a high diagnostic yield (Popovichet al. 1982, Shure &Fedullo 1982).
At present, broncho-alveolar lavage (BAL) plays an important role in the diagnosimany pulmonary diseases such as pulmonary infection, pulmonary haemorrhagealveolar proteinosis. The reports on the value of BAL in diagnosing primary lung canshow variable diagnostic yields from 14 to 68.6 % (Wongsurakiatet al. 1998). In thestudy of Wongsurakiatet al. (1998), the diagnostic yield of BAL was found to beinfluenced by the size and the segmental location of the lesion, since the diagnosticof BAL increased from 14.3 % to 80 % when the diameter of the tumour increased f3.5 cm to over 7 cm, respectively. Furthermore, the diagnostic yield of the BAL indiagnosis of the middle and the lingula lobe lesions was much higher (88.9 %) thanof the apical or posterior segments of the upper lobe lesions (12.5 %).
Cytologic analysis of sputum is a simple, economical and non-invasive metcurrently used to screen for, or to diagnose lung cancer. There are some reports shthat three adequate sputum specimens will give enough evidence for a diagnosis ocarcinoma in about 80 % of central lesions and 50 % of perhipheral lesions (Tanakaet al.1985). However, there are several limitations in this technique. The major problem isinability to collect adequate sputum in most of the patients. According to previous resadding sputum cytology to chest X-ray did not improve the early diagnosis of lu

34
opic
rtainTheadeaderesing,
CLC
inoided
and
ticcific
egralsuchhalin
ithidlylingf
ivecavatedg´s
o 3 %haverm
carcinoma (Mulshine & Scott 1995). Also the results of immediate postbronchoscsputum cytology reported by Wongsurakiet al. (1998) were disappointing, with adiagnostic yield of only 7.7 %.
2.3. Neuroendocrine lung tumours
Neuroendocrine tumours of the lung are a distinct subset of tumours which share cemorphologic, ultrastructural, immunohistochemical and molecular characteristics.major categories of morphologically identifiable neuroendocrine tumours are low-grtypical carcinoid (TC), intermediate-grade atypical carcinoid (AC) and the high grSCLC and LCNEC (Traviset al. 1999). All these tumours share neuroendocrine featuof varying degrees detectable by light microscopy including organoid nesting, palisada trabecular pattern and rosette-like structures (Traviset al. 1999). Histogenesis ofpulmonary neuroendocrine tumours is poorly understood and precursor lesions to Shave not been recognised (for a review, see Colbyet al. 1998). However, it is suggestedthat neuroendocrine cell hyperplasia may be a precursor to a peripheral typical carctumour (reviewed by Colbyet al. 1998). Neuroendocrine lung tumours are distinguishultrastructurally by the presence of variable numbers of dense core granules,immunohistochemically by the production of small polypeptides (Traviset al 1991).Immunohistochemical markers of neuroendocrine differentiation include glycolyenzyme neuron specific enolase (NSE) and the neuroprotein PGP 9.5. Specomponents of neurosecretory granules include the matrix protein chromogranin, intmembrane glycoprotein synaptophysin and biologically active amines and peptidesas serotonin, bombesin, vasoactive intestinal polypeptide, gastrin and leu-enkep(Hammar 1994).
2.3.1. Small cell lung carcinoma (SCLC)
SCLC accounts for 25 to 30 % of all lung tumours and it is strongly associated wcigarette smoking (Hammar 1994, Glisson & Hong 1997). SCLC is the most rapprogressing form of lung cancer with the highest growth fraction and shortest doubtime (Brighamet al. 1978). SCLC is also most clinically distinctive of the major types olung cancer for its biologic characteristics and responsiveness to chemotherapy.
At the time of diagnosis patients with SCLC commonly suffer from postobstructpneumonitis, haemoptysis, atelectasis, vocal cord paralysis, or superior venasydrome (Ihdeet al. 1984). Certain paraneoplastic sydromes are distinctively associawith SCLC, namely inappropriate secretion of antidiuretic hormone, ectopic Cushinsyndrome and the Eaton-Lambert syndrome (Morriset al. 1992). Approximately 70 % ofSCLCs have metastasised at the time of diagnosis (Carter 1983). Because only 1 % tof these patients are potential candidates for surgery, chemotherapy and radiationplayed the most significant role in the treatment of SCLC (Carter 1983). Long-te

35
0 %.p to
t isnd
es
ithn 10
lsar tor 10andis
ize,ther
ion of
d totcrineC
d ofquent
lackaly
ity
ACrosis,998).ie of
survival rate, even for patients presenting with clinically limited disease is less than 1However, with combined chemotherapy and radiation, a 5- year survival rate of u23 % has recently been achieved (Turrisiet al. 1999).
2.3.2. Large cell neuroendocrine carcinoma (LCNEC)
LCNEC of the lung is a poorly differentiated high-grade neuroendocrine tumour thamorphologically and biologically intermediate-type between atypical carcinoid (AC) aSCLC (Traviset al. 1999). Together with atypical and typical carcinoids, LCNEC makup only 2-3 % of neuroendocrine lung tumours (Traviset al. 1999). Of theneuroendocrine lung tumours, LCNEC together with SCLC is highly associated wcigarette smoking and they have the poorest prognosis with 5-year survival less tha% (Downeyet al. 1989, Traviset al. 1999).
According to Traviset al. (1991), LCNEC is a tumour composed of large celcharacterised by a light microscopic neuroendocrine appearance with a low nuclecytoplasmic ratio, frequent nucoleoli, a high mitotic rate (greater than 10 mitoses pehigh power fields), and abundant necrosis. The main criterion for separating LCNECSCLC from AC is a mitotic count of eleven or more mitoses per ten HPFs. LCNECseparated from SCLC using a constellation of criteria, which include larger cell sabundant cytoplasm, prominent nucleoli, vesicular or coarse chromatin, polygonal rathan fusiform shape, less prominent nuclear moulding and less conspicuous deposithaematoxylin-stained material (DNA) in blood vessel walls (Traviset al. 1999).
2.3.3. Bronchopulmonary carcinoid tumour (CT)
CT account for 1 to 2 % of all primary lung tumours and are not necessarily relatesmoking or environmental pollution (Brensteinet al. 1989). These potentially malignantumours occur equally in men and women with a mean age of 55 years. Neuroendocell hyperplasia with or without tumourlets is relatively frequent in CT but not in LCNEor SCLC (Traviset al. 1999).
TC is the best differentiated pulmonary neuroendocrine tumour that is composeuniform cells containing numerous dense-core neuroendocrine granules and frelysosomes (Hammar 1994). TC have less than two mitoses per ten HPFs andnecrosis (Traviset al. 1999). At the time of diagnosis, 5 to 20 % of TC have regionlymph node involvement (Brensteinet al. 1989). Although this rarely happens, TC mametastasize to distant sites, usually many years after the initial diagnosis (Brensteinet al.1989). The most important criterion for separating TC from AC is lower mitotic activand lack of tumour necrosis in TC.
AC represent 11 to 24 % of pulmonary CT (Hammar 1994). Compared to TC,have a larger tumour size, more frequent mitoses (2 to 10 per ten HPFs), tumour neca higher rate of metastases, and a significantly reduced survival rate (Travis et al. 1The mortality ranges between 27 to 47 %, and the mean survival for patients who dAC is slightly over 2 years, ranging up to 10 years.

36
rtantlearty of
in
nis,
sue
s a
A
re of
inournceheasedrent
nd is
ungor aby
tein
2.4. Markers of cell proliferation
2.4.1. Ki-67 and PCNA
Growth rate is a principal determinant of the aggressiveness of a tumour and an impoprognostic factor. The proliferation associated antigen Ki-67 and proliferating cell nucantigen (PCNA) have been used as proliferation and prognostic markers in a variemalignancies, including lung tumours (Korkolopoulouet al. 1993, Kawaiet al. 1994).Ki-67 is a DNA-binding nuclear protein expressed throughout the cell cycleproliferating cells but not in quiescent (G0) cells (Gerdeset al. 1984). Due to theinstability of the Ki-67 epitope in formalin fixation, analysis with Ki-67 has beepreviously restricted to frozen sections. MIB-1, an antibody against Ki-67 antigenhowever, able to detect proliferating cells in formalin-fixed, paraffin-embedded tissections after microwave antigen retrieval (Cattorettiet al. 1992).
Proliferating cell nuclear antigen (PCNA) is a 36 kD nuclear protein acting acofactor for DNA polymerase delta (Mathewset al. 1984). Synthesis of PCNA is reportedto correlate directly with DNA replication and cell proliferation. The expression of PCNincreases in late G1, reaches its maximum in S1 and then declines (Hallet al. 1990). Theexpression of PCNA in tumours is, however, considered to be an unreliable measuproliferative activity (Hallet al. 1990).
2.5. Apoptosis in tumours
Apoptosis has a pivotal role in limiting the population expansion of tumour cells earlythe process of tumour growth, at the stage in which angiogenesis is limiting tumprogression (Naiket al. 1996). Disturbances in the homeostatic mechanisms that balacell proliferation and cell death can contribute to the growth rate of a tumour. Taccumulation of neoplastic cells is not, however, consistently associated with decrerate of apoptosis. On the contrary, high apoptotic activity seems to be appaparticularly in aggressive tumours, such as urinary bladder carcinoma (Kinget al. 1996,Lipponenet al. 1994a). In general, apoptosis seems to be accelerated in cancer ahighly dependent on tumour type (for a review, see Soiniet al. 1998a).
2.5.1. Apoptosis in lung carcinoma
There is great variability in the extent of apoptosis detected in different types of ltumours as well as in same type of lung tumours reported by different authors (freview, see Soiniet al. 1998a). In NSCLC, the extent of apoptosis as detectedapoptotic index (%) varies between 0.37 to 1.00 (Stammler & Volm 1996, Kargiet al.1997) and in SCLC between 2.65 to 10.9 (Brambillaet al. 1996, Gaffneyet al. 1995).Paradoxically, despite a higher apoptotic activity, also higher anti-apoptotic bcl-2 proexpression has been detected in SCLC compared to NSCLC (Stauntonet al. 1995,

37
ourlow
andenentsds a
T
ar aune
muchTheble
r
ryand). B
in
nse,ux
tort &
Törmänenet al. 1995). One may assume that enhanced apoptosis will retard tumgrowth and hence indicate a favourable prognosis compared with tumours showing aapoptotic activity. However, according to the few studies available on apoptosisprognosis for lung carcinomas, the opposite seems to be true. According to Törmänetal. (1995), a high apoptotic activity is associated with a shortened survival of the patiwith non-small cell lung carcinoma. Stammel and Volm (1996) reported a trend towarsimilar association.
2.6. Tumour immunity
2.6.1. Tumour infiltrating lymphocytes (TILs)
Tumour infiltrating lymphocytes (TILs) include helper and suppressor/cytotoxiclymphocytes, natural killer (NK) cells, B lymphocytes and macrophages (Balchet al.1990). In the middle of the 1970s it was for the first time proposed that TILs may bedirect relationship to improved prognosis and may represent an expression of immsurveillance in human tumours (Underwood 1974). Since then, this idea has gainedsupport but even today, the role of TILs in solid tumours has remained controversial.infiltration of immunologic cells in tumour tissue has been associated with a favouraprognosis in melanoma (Poppemaet al. 1983), carcinoma of the urinary bladde(Lipponenet al. 1994a), cervix (Nakanoet al.1992), prostate (Vesalainenet al. 1994) andcolorectum (Naitoet al. 1998). However, in many cases TILs are viewed as inflammatocells mediating nonspecific rather than tumour specific interactions. T lymphocytesmacrophages are the main component of TILs in solid tumours (Underwood 1974cells are rarely found, with the exception of melanoma (Ruiteret al. 1982), and NK arepresent in relatively small numbers (Balchet al. 1990). In lung carcinoma, TILs havebeen associated with histological type and degree of differentiation (Watanabeet al.1983, Kerret al. 1998). Previously, a high percentage of B cells (Kerret al. 1998) andmacrophages (Watanabeet al. 1983) has been associated with a better survivalNSCLC.
2.6.2. Apoptosis and the immune system
Apoptosis plays a key role in the immune system controlling the immune respodeleting immune cells recognising self-antigens, and cytotoxic killing. (Ekert & Va1997). Defects in the regulation of apoptosis in the immune system may leadautoimmune disease, immunodeficiency as well as to progression of cancer (EkeVaux 1997).

38
aceg of
cell.tionaseracthecell
oticbe
llstes,
mour
ptoticand
oblastncee p53
r the
2.6.2.1. Fas(CD95/APO-1)
Fas(CD95/APO-1) belongs to the tumour necrosis factor family (TNF) of cell surfreceptors (Ekert & Vaux 1997) which is expressed in wide range of cell types. Bindinligand (FasL) or anti-Fas antibodies to Fas results in apoptosis of the Fas bearingThe cytoplasmic domain of Fas bears a motif called ”death domain”, which, after ligaof FasL, allows it to bind the death domain of cytoplasmic proteins FADD/Mort-1 (Fassociating death domain) and RIP (Receptor interacting protein). These, in turn, intwith FLICE/Mach-1 (Fas ligand interacting cell effector), leading to the activation of tcaspases and to apoptotic cell death (Ekert & Vaux 1997). There is evidence that Tfunction in tumour micro-environment is modulated through the Fas/FasL apoptpathway (Cardiet al. 1998). Furthermore, functional FasL has been shown toexpressed in several tumour types including lung carcinoma (Strandet al. 1996, Hellquistet al. 1997, Niehanset al. 1997). It is suggested that FasL expressing tumour ceactively protect themselves by destroying Fas bearing tumour infiltrating lymphocythus contributing to tumour growth (Ekert & Vaux 1997).
2.7. Tumour angiogenesis
There are several studies indicating that angiogenesis has an important role in tudevelopment, progression and formation of metastases (Folkmanet al. 1990). In theabsence of an adequate vasculature tumour cells become necrotic and/or apo(Holmgrenet al. 1995). Angiogenesis can be regulated by the balance of inducersinhibitors of endothelial cell proliferation (Hanahanet al. 1996). Most commonlyexpressed inducers of angiogenesis in human and animal cancers are basic fibrgrowth factor (bFGF) and vascular endothelial growth factor (VEGF). There is evidethat some of these inhibitors and inducers of angiogenesis are under the control of thtumour-suppressor gene (Dameronet al. 1994, Fontaniniet al. 1997). In NSCLC, tumourvascularity has been found to be an independent prognostic factor and important fodevelopment of haematogenous metastases (Macchiariniet al. 1992, Matsuyamaet al.1998, Yamazakiet al. 1994).

3. Aims of the study
The specific aims of this study were:1. To investigate the extent of apoptosis and its association to apoptosis regulating
proteins in SCLC (I) and in LCLC (II).2. To elucidate the rate of the apoptosis and apoptosis regulating proteins as a prognostic
marker in LCLC (II) and SCLC (unpublished data).3. To evaluate the occurrence of tumour infiltrating lymphocytes in relation to tumour
angiogenesis, apoptosis and prognosis in SCLC (III) and LCLC (IV).4. To apply the detection of enhanced apoptosis as a marker of lung carcinoma in
diagnostic respiratory cytology (V).

Cs3120
ingLCfiletients,ll and98.56
asesIII-themetientno
onlyf thetionrapy
to-n in-coreindilylungor
4. Materials and methods
4.1. Tumour samples and identification of neuroendocrinedifferentiation (I-IV)
A total of 94 patients operated for lung carcinoma including 56 SCLCs and 38 LCLwere included in the original papers as follows: 40 cases of SCLC includingintermediate cell type and 9 oat cell type SCLCs (I); 35 cases of LCLC includingLCNECs and 15 LCNNECs (II); 56 cases of SCLC (III) and 38 cases of LCLC includ20 LCNECs and 18 LCNNECs (IV). The original number of the patients in the SCgroup increased by 16 (III) and in the LCLC group by three (IV) due to expandingsearch and the new operated patients that were added to the material. The paincluding 84 males and 10 females, represented all the cases operated for large cesmall cell lung carcinoma at Oulu University Hospital during the years 1975-19Clinical follow-up data of the patients were collected from the hospital records. Of theSCLC cases, there were 37 stage I-II and 19 stage III-IV tumours. Of the 38 LCLC cthere were 24 stage I-II and 11 stage III-IV tumours. Of all LCLCs, most of the stageIV tumours were LCNECs (8 out of 11). 64 % of the patients with SCLC and 46 % ofpatients with LCLC died of the disease during the 5-year follow-up (mean follow-up ti42 months). The postoperative treatment of the patients was collected from the parecords and it was compared to the survival. Of the patients with SCLC 14 hadpostoperative treatment, 13 were treated by radiation only, 12 with chemotherapyand 14 patients were treated with the combination of chemotherapy and radiation. Opatients with LCLC 19 had no postoperative treatment, 16 were treated with radiaonly, one patient with chemotherapy only, and two patients with combined chemotheand radiation.
Neuroendocrine (NE) differentiation was identified on the basis of immunohischemically detectable NE markers synaptophysin or chromogranin. Chromogranilung tumours has been shown to correlate with the number of cytoplasmic densegranules (Saidet al. 1985). Synaptophysin is an integral membrane glycoproteoriginally isolated from presynaptic vesicles of bovine neurons that can be reademonstrated in most neuroendocrine neoplasms including NE carcinomas of(Gould et al. 1987). 20 out of 38 LCLCs expressed either chromogranin

41
avisall
g of
78sityps: 1)
ungs in
ostncyinence,. Theand0 =
inedereupst2)
lia)acute
synaptophysin, and by using this information, along with the criteria presented by Tret al (1991), they were classified as NE tumours. NE differentiation was detected inSCLCs and the diagnosis was based on the WHO International Histological TypinLung Tumours (WHO 1981).
4.2. Cytological samples (V)
4.2.1. Sputum smears
Cytological material included 84 Papanicolau-stained sputum smear samples frompatients collected from the files of the Department of Pathology of the Oulu UniverHospital between the years 1986-1995. Comparisons were made between three grousamples from patients without malignancy (n = 31), 2) samples from patients with lcarcinoma (n = 53) and 3) samples from patients with SCLC without malignant cellsputum smear (n = 11). Cytological material is presented in detail in Table 1. The mcommon indications of collecting sputum smears from the patients without malignawere inflammatory or fibrotic alterations in chest X-ray, haemoptysis, difficultybreathing and weight loss. All samples were collected for diagnostic purposes, and hthe patients had not been subjected to any chemo or radiation therapy regimensextent of inflammation in each sputum sample was evaluated by light microscopydivided into four groups according to the number of neutrophils present as follows:no neutrophils, 1 = a few, 2 = moderate, 3 = abundant number of neutrophils.
4.2.2. BAL samples
For control purposes, cytological samples of 13 May-Grünwald-Giemsa (MGG) stabroncho-alveolar lavage (BAL) specimens from patients without lung carcinoma walso included in the study (see Table 1). BAL specimens were divided into two groaccording to the findings in cell differential count, 1) normal finding or slighabnormalities in cell differential count and cytological analysis (n = 5) andabnormalities in cell differential count (lymphocytosis, neutrophilia and eosinophiassociated with different diseases such as sarcoidosis, other chronic infections,infections and asbestos exposure (n = 8).
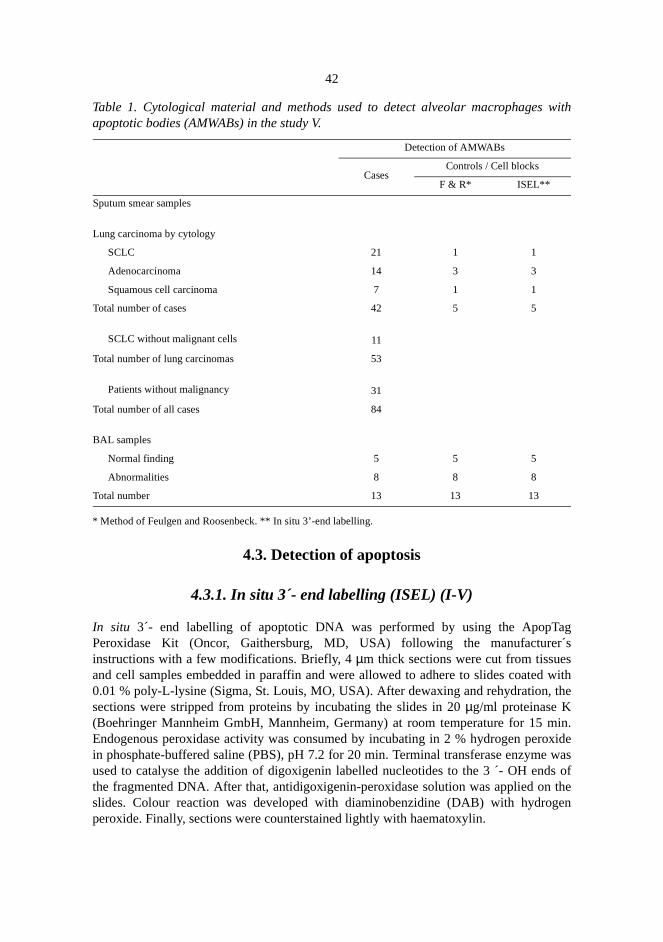
42
with
agr´s
withhe
in.xidewass oftheen
Table 1. Cytological material and methods used to detect alveolar macrophagesapoptotic bodies (AMWABs) in the study V.
* Method of Feulgen and Roosenbeck. ** In situ 3’-end labelling.
4.3. Detection of apoptosis
4.3.1. In situ 3´- end labelling (ISEL) (I-V)
In situ 3´- end labelling of apoptotic DNA was performed by using the ApopTPeroxidase Kit (Oncor, Gaithersburg, MD, USA) following the manufactureinstructions with a few modifications. Briefly, 4µm thick sections were cut from tissuesand cell samples embedded in paraffin and were allowed to adhere to slides coated0.01 % poly-L-lysine (Sigma, St. Louis, MO, USA). After dewaxing and rehydration, tsections were stripped from proteins by incubating the slides in 20µg/ml proteinase K(Boehringer Mannheim GmbH, Mannheim, Germany) at room temperature for 15 mEndogenous peroxidase activity was consumed by incubating in 2 % hydrogen peroin phosphate-buffered saline (PBS), pH 7.2 for 20 min. Terminal transferase enzymeused to catalyse the addition of digoxigenin labelled nucleotides to the 3 ´- OH endthe fragmented DNA. After that, antidigoxigenin-peroxidase solution was applied onslides. Colour reaction was developed with diaminobenzidine (DAB) with hydrogperoxide. Finally, sections were counterstained lightly with haematoxylin.
Detection of AMWABs
CasesControls / Cell blocks
F & R* ISEL**
Sputum smear samples
Lung carcinoma by cytology
SCLC 21 1 1
Adenocarcinoma 14 3 3
Squamous cell carcinoma 7 1 1
Total number of cases 42 5 5
SCLC without malignant cells 11
Total number of lung carcinomas 53
Patients without malignancy 31
Total number of all cases 84
BAL samples
Normal finding 5 5 5
Abnormalities 8 8 8
Total number 13 13 13

43
usingy, 2)and
orlin,
olarlight
g andardacid
wereurpleNA
ontrolcyteala,e at,sticm,lls ofer ofAs adex
and
and0 xes
4.3.2. Detection of apoptotic bodies within alveolar macrophages (V)
In selected cases, apoptotic bodies within alveolar macrophages were analysed bythree different techniques, i.e. 1) Papanicolaou staining and routine light microscopin situ 3 ´- end labelling of apoptotic DNA and 3) the standard method of FeulgenRoosenbeck (Vaccaet al. 1985) (see Table 1). For these techniques, cells from sputumBAL samples were harvested by centrifugation, fixed in 10 % buffered formaimmersed in 2 % agar, and embedded in paraffin.The in situ3´- end labelling methodwas used as previously described (see 4.3.1). Apoptotic bodies within alvemacrophages of Papanicolaou stained sputum smear samples were identified bymicroscopy based on the characteristic morphology of apoptotic bodies, includinround and regular and condensed appearance and a dark blue staining. The stamethod of Feulgen and Roosenbeck was used for demonstrating deoxyribonucleic(DNA) (Vaccaet al. 1985). Acid hydrolysis, employing 1 M HCl at 60° C, was used tobreak the purine-deoxyribose bond, and the resulting reactive aldehydesdemonstrated by the use of Schiff´s reagent. Elements containing DNA gave a red pcolour. The ribose-purine bond remains unaffected by the hydrolysis, leaving Runstained.
4.3.3. Controls of apoptosis (I-IV)
Peripheral blood lymphocytes treated with dexamethasone were used as a positive cfor apoptosis (I). Lymphocytes were separated from heparinised blood using lymphoseparation medium (Ficoll-Paque, Pharmacia LKB Biotechnology Inc., UppsSweden). After 48 hours´ culture, lymphocytes were treated with dexamethazonconcentrations of 1, 5 and 10µM for 24 hours. Lymphocytes were harvested, fixedembedded in 2 % agar, and after solidification, embedded in paraffin. Non-neoplacontrol tissues included lung tissue with uninvolved bronchial respiratory epitheliuserous and mucous glands of the bronchial wall, alveolar macrophages, interstitial celung parenchyma. A hyperplastic lymph node sample, showing an increased numbapoptotic cells within germinal centres, was used as a positive control for apoptosis.negative control, a tumour sample previously shown to exhibit a high apoptotic inwhere TdT-enzyme was omitted was used.
4.3.4. Evaluation of apoptosis in tumours (I-IV)
The extent of apoptosis in tumour samples was evaluated by the labelling patternmorphology of apoptotic cells and bodies detected byin situ 3 ´- end labelling. Twoindices were used to indicate the extent of apoptosis; the number of apoptotic cellsbodies per high power field (HPF) as counted from a minimum of ten HPFs (4objective, diameter of the field 400µm) and a percentage of apoptotic cells and bodi(% index) of all the cells within an HPF.

44
tive.erage,ence oflogywasverL
seen
cles,ereining
thodd,ith aume
ed toons
wereed ined by
utes.in
darywiththe
tlyy).
4.3.5. Evaluation of apoptotic bodies within alveolar macrophages (V)
The cytological samples were analysed by using light microscopy with a 40 x objecSpecifically, the presence of apoptotic bodies in macrophages was recorded. On av500 consecutive alveolar macrophages were analysed in each sample, and the presapoptotic bodies within alveolar macrophages was identified on the basis of morphoand staining pattern. The number of alveolar macrophages with apoptotic bodiescounted originally by two persons (A.-K.E. and P.P.). In order to test the interobsevariation AMWABs was counted also by a third person (Y.S.) in a set of 13 BAsamples stained by the method of Feulgen and Roosenbeck.
Usually one, but in a few cases also more than one apoptotic body could bewithin one macrophage. Apoptotic bodies as small as 0.5µm in diameter were discernedby using the 40 x objective. Contrary to foreign bodies, such as anthracotic partiwhich showed an angular and asymmetric morphology, apoptotic bodies wdistinguished on the basis of their round and regular appearance and dark, blue stanuclei. These were often surrounded by a halo.
4.4. Evaluation of tumour necrosis (I-II)
The volume density of tumour necrosis was determined by a morphometric me(Pääkkö et al. 1989). Briefly, a Nikon Labophot light microscope was employeconnected via a video camera (Panasonic F10) to a TV monitor (Sony) equipped wnet of 100 points adherent to the screen. The total magnification for counting the voldensity was x 100 and the total number of counts was 2000 in each case.
4.5. Immunohistochemistry (I-IV)
For all immunostainings, a 4 µm thick section was cut from each specimen and placthe slides coated with poly-L-lysine solution (Sigma Chemicals, St. Louis, MO). Sectifor all other immunostainings but for p53 were dried at 37° C overnight to facilitate thesticking of the sections to the slides. Sections selected for p53 immunostainingallowed to dry overnight at room temperature. After that the specimens were dewaxxylene and rehydrated in graded alcohol. The endogenous peroxidase was consumimmersing the sections in 0.1 % hydrogen peroxide in absolute methanol for 20 minNon-specific binding was blocked by incubating the slides in 20 % foetal calf serumphosphate-buffered saline (PBS) for 20 minutes. Specific primary and seconantibodies were used for each immunostaining as described below. After incubationsecondary antibody, the avidin-biotin-complex method was used (Hsu 1990) andcolour was developed with diaminobenzidine (DAB). Finally, the sections were lighconterstained with haematoxylin and mounted with Eukitt (Kindler, Freiburg, German

45
ntlyred.
arytraith
f the
as
anti-by
ark).tric
the%,
at aiesforndith6.0)
ive1-24of
s 0 =
All the immunohistochemical stainings were evaluated by two persons independe(A.-K.E. and P.P.) and consensus was made when the opinions diffeImmunohistochemical methods is presented in Table 2.
4.5.1. p53 protein (I)
For p53 immunostaining, sections were incubated overnight at 4º C with primpolyclonal rabbit anti-human p53 antibody CM-1 (dilution 1:1000; NovocasLaboratories, Newcastle upon Tyne, United Kingdom) followed by incubation wbiotinylated anti-rabbit antibody (Dakopatts, Copenhagen, Denmark).
The p53 positivity was evaluated semiquantitatively and graded on the basis onumber of stained nuclei as follows: - = negative, + =≤ 5 %, ++ = 6-10 %, +++ =11-40%, and ++++ = > 40 % of the nuclei positive. The proportion of positive cells wcounted in ten HPFs (objective, x 40; diameter of the field 400µm).
4.5.2. Ki-67 (I)
For Ki-67 immunostaining, the sections were incubated with a monoclonal mousehuman MIB-1 antibody (Immunotech, Marseille, France, at a dilution 1:25) followedincubation with biotinylated anti-mouse antibody (Dakopatts, Copenhagen, DenmPrior to incubation with the MIB-1 antibody, the sections were heated in 10 mM ciacid monohydrate (pH 6.0) in a microwave oven.
Ki-67 positivity was evaluated semiquantitatively and graded on the basis ofnumber of stained nuclei as follows: - = negative, + = 1-5 %, ++ = 6-20 %, +++ = 30++++ = 40-50 % and +++++≥ 60 % of the nuclei positive. The proportion of positivecells was counted in ten HPFs (objective, x 40; diameter of the field 400µm).
4.5.3. Bcl-2 and related proteins Mcl-1, Bax and Bak (I-II)
Monoclonal mouse anti-human bcl-2 antibody (clone 124, Dako, Glostrup, Denmark,dilution of 1:50) and polyclonal rabbit anti-human bax, bak and mcl-1 antibod(Pharmingen, San Diego, CA, USA, all at a dilution of 1:1000) were usedimmunostainings followed by incubation with biotinylated anti-mouse (for bcl-2) aanti-rabbit (for bax, bak and mcl-1) antibodies, respectively. Prior to incubation wprimary antibodies, the sections were heated in 10 mM citric acid monohydrate (pHin a microwave oven.
For evaluation of bcl-2, bax, bak and mcl-1 positivity, the percentage of posittumour cells was analysed semiquantitatively and graded as follows: 0 = none, 1 =%, 2 = 25-50 %, and 3 = 51-100 % of the tumour cells positive (I, II). The proportionpositive cells was counted in ten HPFs (objective, x 40; diameter of the field 400µm),and the intensity of staining was evaluated in the whole tumour area and graded a

46
bakwas
26,stlells
toxicjoritytion
(III)xute
Binally,
withhewashtly
utivePF isdianlow
negative, 1 = weak, 2 = moderate, 3 = strong. Finally, a staining index for bcl-2, bax,and mcl-1 was given in which the sum of the semiquantitative and qualitative scorescalculated for each tumour specimen (II).
4.5.4. Tumour infiltrating lymphocytes (TILs) (III-IV)
For immunophenotypic analysis of TILs, monoclonal mouse antibodies UCHL1, LPGM-1 (DAKO, Glostrup, Denmark) and CD8 (Novocastra Laboratories, Newcaupon Tyne, United Kingdom) were used. UCHL1 (anti-CD45) labels resting T cewithin both the CD4 and CD8 subsets and mature activated T cells. CD8 labels cytosuppressor T cells. L26 (anti-CD20) is directed against an antigen present in the maof B cells while PGM-1 stains macrophages. All these antibodies were used at a diluof 1:50.
An area containing on average 500-700 tumour cells, range 100 – 1200 for SCLCand 100-1100 for LCLC (IV) in a minimum of five high power fields (HPF; 40objective, diameter of the field: 400µm) was evaluated in each case and the absolnumber of intratumoural T lymphocytes, CD8 positive suppressor T cells,lymphocytes, and macrophages was calculated per HPFs avoiding necrotic areas. Fthe average number of each inflammatory cell type per HPF was given.
4.5.5. Factor VIII (III-IV)
Microvessels in the tumour tissue were detected by immunohistochemical staininganti-factor VIII related antigen polyclonal antibody (dilution 1:200) A0082 that labels tvascular endothelium (Dako, Glostrup, Denmark). The avidin-biotin staining methodused, with diaminobenzidine (DAB) as a chromogen. Finally, the sections were ligcounterstained with haematoxylin.
The absolute number of microvessels in tumours was evaluated from five consecHPFs avoiding necrotic areas. Finally, the average number of microvessels per Hgiven. Microvessel density (MD) was considered a continuous variable, and a mevalue of nine (III) and 6.6 (IV) microvessels per HPF was used as a limit betweenand high MD for SCLC and LCLC, respectively.

47
zenlesfor
rmedto
raturekieswithweredish-een
een-ng to
kagenge.
riatethe
ith
n
Table 2. Immunohistochemical methods used in studies I-IV.
4.6. Western Blotting (II)
In order to test the specificity of the antibodies for bcl-2, bax, bak and mcl-1, fresh-frotissue from 3 LCLCs (one LCNEC and two LCNNEC) was used (II). The tissue sampwere homogenised in PBS, centrifuged at 10,000 g for 15 min, and sampleselectrophoresis were prepared from the supernatants. SDS-PAGE was perfoessentially according to Laemmli (1970). Resolved proteins were transferrednitrocellulose membranes. The membranes were incubated overnight at room tempein TBS (50 mM Tris-HCl/200 mM NaCl, pH 7.4) containing 5 % dried milk to blocnon-specific binding sites. Anti-bcl-2, anti-bax, anti-bak and anti-mcl-1 antibod(1:250, 1:2,000, 1:2,000 and 1:2,000 dilutions in TBS, respectively) were incubatedthe membranes for 60 min at room temperature. Biotinylated secondary antibodiesused, after which the membranes were incubated with streptavidin-horseraperoxidase complex (ECL Western blotting kit, Amersham, Amersham, UK). Betweach step, the membranes were washed extensively with TBS containing 0.05 % Tw20. Finally, chemiluminescence-based detection of proteins was performed accordithe manufacturer’s protocol (Amersham).
4.7. Statistical methods
The statistical analyses were performed with the SPSS for Windows program pac(Chicago, IL. USA). Data on the apoptotic indices are presented as means with raThe significances of the associations were determined usingχ2 test, Fisher’s exactprobability test, Pearson´s test, the two-tailed t-test and Mann-Whitney U-test. Univaanalyses of the survival data were performed using survival curves and applyingKaplan-Meyer method with log rank analysis. The probability values are two-tailed, wa probability of p < 0.05 considered as statistically significant.
Antigen Antibody T* Dilution Specificity SourceKi-67 MIB-1 + 1:25 All proliferating cells Immunotech, Marseille, FranceBcl-2 Bcl-2 Ab
(clone124)+
1:50 Bcl-2 protein Dako, Glostrup, Denmark
Bax Bax Ab + 1:1000 Bax protein Pharmingen, San Diego, CA, USABak Bak Ab + 1:1000 Bak protein Pharmingen, San Diego, CA, USAMcl-1 Mcl-1 Ab + 1:1000 Mcl-1 protein Pharmingen, San Diego, CA, USAP53 CM-1 - 1:1000 All forms of p53 Novocastra Laboratories, Newcastle upo
Tyne, UK
Factor VIII A0082 - 1:200 Endothelial cells Dako, Glostrup, DenmarkCD45 UCHL1 - 1:50 T cell Dako, Glostrup, DenmarkCD20 L26 - 1.50 B cells Dako, Glostrup, DenmarkCD68 PGM-1 - 1:50 Macrophages Dako, Glostrup, DenmarkCD8 NCL-CD8 - 1:50 Cytotoxic/suppressor
T cellsNovocastra LaboratoriesNewcastle upon Tyne, UK
*T = pretreatment, + = microwave, citrate buffer, - = none

s ofuclearcted.e
ting
its(I),
res ofr ofto the2 %).ialthe
CLCp =heNond
5. Results
5.1. Apoptosis in tumour samples (I-IV)
5.1.1. Morphological features of apoptosis
By light microscopy, the sections labelled for apoptotic DNA revealed different stageapoptosis. At the onset of apoptosis, labelled nuclear chromatin was seen at the nenvelope. At later stages, more intense and uniform nuclear labelling was deteApoptotic bodies, as small as 0.5µm, representing final stages of apoptosis could breadily detected by using a 40 x objective. Clusters of apoptotic bodies likely originafrom the same dying cell were counted as one apoptotic cell.
5.1.2. Extent of apoptosis in SCLC (I, III) and LCLC ( II, IV)
The apoptotic indices in SCLC (mean 2.70 %) and in LCLC (mean 2.49 %) withsubtypes LCNEC and LCNNEC are presented in Fig. 1 (unpublished data). In SCLCthe apoptotic indices (mean 2.33 %) were in the same range as in the germinal centhyperplastic lymph nodes (mean 1.92 %). Interestingly, in SCLC a high numbeapoptotic bodies was detected within alveolar macrophages (mean 1.99 %), closeextent of apoptosis seen in germinal centres of hyperplastic lymph nodes (mean 1.9Low apoptotic activity (mean 0.00-0.34 %) was detected in uninvolved bronchepithelium, serous and mucous glands, interstitial cells and lymphocytes outsidegerminal centres of hyperplastic lymph nodes (I). When all the studied cases of Sand LCLC (I-IV) were analysed, higher apoptotic indices were detected in SCLC (0.05 by Mann-Whitney U-test, unpublished data). Both in SCLC and in LCLC tnumber of apoptotic bodies was roughly twice as high as that of apoptotic cells.significant differences could be found in apoptotic indices between LCNEC aLCNNEC.

49
Fig. 1. Apoptotic indices (%) of the whole cell population small cell lung carcinoma (SCLC) andin large cell lung carcinoma (LCLC) with its subtypes of large cell neuroendocrine carcinoma(LCNEC) and large cell non-neuroendocrine carcinoma LCNNEC.
5.1.3. Apoptosis in relation to clinical data (I-II)
High apoptotic activity was associated with high age and long smoking history of thepatients with SCLC (p = 0.01 and p = 0.001) (I). In contrast, in LCNEC, patients whowere younger than 60 years had tumours with significantly higher apoptotic indices (p =0.01) (II). No association was found between apoptosis and tumour stage, tumourdiameter, or metastases in either of the tumour types.
5.2. Apoptosis regulating proteins (I-II)
5.2.1. p53 protein accumulation in SCLC (I)
38 % of SCLCs showed positive p53 immunoreactivity (Table 3). The intermediate celltype SCLC showed more than 5 % of p53-positive nuclei more frequently than the oatcell type (p = 0.05). Low apoptotic activity was associated with p53 protein expression, inother words, p53-postive tumours showed low apoptotic indices (≤ 2 %) significantlymore often than p53-negative tumours (p = 0.008, Table 4).
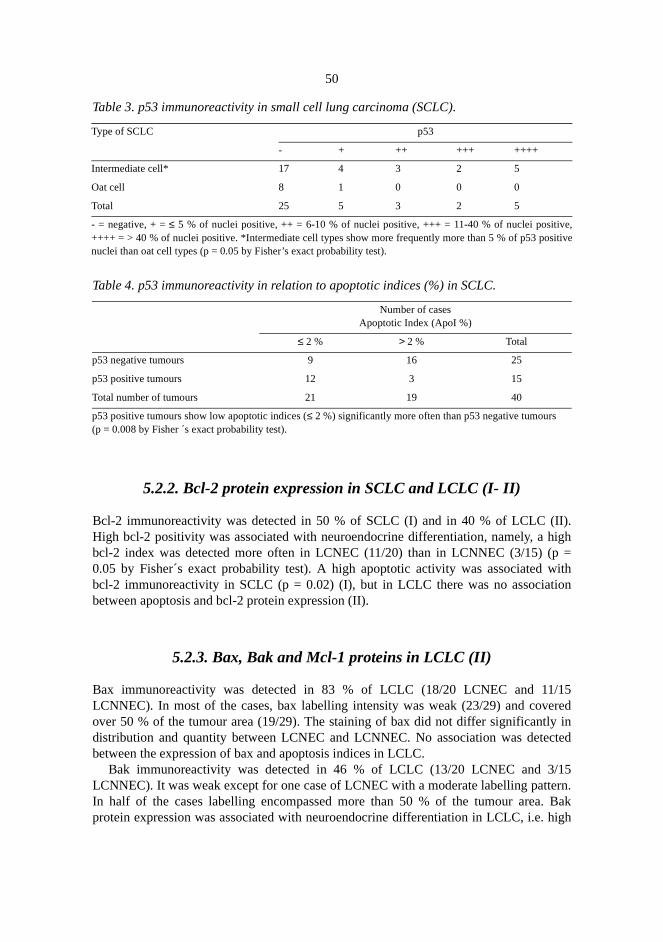
50
).igh=ith
on
15redin
cted
15ern.
Bakhigh
e,itive
Table 3. p53 immunoreactivity in small cell lung carcinoma (SCLC).
Table 4. p53 immunoreactivity in relation to apoptotic indices (%) in SCLC.
5.2.2. Bcl-2 protein expression in SCLC and LCLC (I- II)
Bcl-2 immunoreactivity was detected in 50 % of SCLC (I) and in 40 % of LCLC (IIHigh bcl-2 positivity was associated with neuroendocrine differentiation, namely, a hbcl-2 index was detected more often in LCNEC (11/20) than in LCNNEC (3/15) (p0.05 by Fisher´s exact probability test). A high apoptotic activity was associated wbcl-2 immunoreactivity in SCLC (p = 0.02) (I), but in LCLC there was no associatibetween apoptosis and bcl-2 protein expression (II).
5.2.3. Bax, Bak and Mcl-1 proteins in LCLC (II)
Bax immunoreactivity was detected in 83 % of LCLC (18/20 LCNEC and 11/LCNNEC). In most of the cases, bax labelling intensity was weak (23/29) and coveover 50 % of the tumour area (19/29). The staining of bax did not differ significantlydistribution and quantity between LCNEC and LCNNEC. No association was detebetween the expression of bax and apoptosis indices in LCLC.
Bak immunoreactivity was detected in 46 % of LCLC (13/20 LCNEC and 3/LCNNEC). It was weak except for one case of LCNEC with a moderate labelling pattIn half of the cases labelling encompassed more than 50 % of the tumour area.protein expression was associated with neuroendocrine differentiation in LCLC, i.e.
Type of SCLC p53
- + ++ +++ ++++
Intermediate cell* 17 4 3 2 5
Oat cell 8 1 0 0 0
Total 25 5 3 2 5
- = negative, + =≤ 5 % of nuclei positive, ++ = 6-10 % of nuclei positive, +++ = 11-40 % of nuclei positiv++++ = > 40 % of nuclei positive. *Intermediate cell types show more frequently more than 5 % of p53 posnuclei than oat cell types (p = 0.05 by Fisher’s exact probability test).
Number of casesApoptotic Index (ApoI %)
≤ 2 % > 2 % Total
p53 negative tumours 9 16 25
p53 positive tumours 12 3 15
Total number of tumours 21 19 40
p53 positive tumours show low apoptotic indices (≤ 2 %) significantly more often than p53 negative tumours(p = 0.008 by Fisher ´s exact probability test).

51
.02n and
/15tivecl-1byeneent in
onel 3ding
kDa)kDa
ity.oatLCivity
LC,ivity;ices05).ere
bak indices were detected significantly more often in LCNEC than in LCNNEC (p = 0by Fisher´s exact probability test). No association was found between bak expressioapoptotic indices in LCLC.
Mcl-1 immunoreactivity was detected in 80 % of the cases (19/20 LCNEC and 9LCNNEC). Mcl-1 labelling was considered weak in 74 % of the cases and mcl-1 positumour cells covered over 50 % of tumour area in 17 out of 26 cases. Strong mlabelling intensity was associated with neuroendocrine differentiation (p = 0.03Fisher´s exact probability test), while quantity was not significantly different betweLCNEC and LCNNEC. There was a statistically significant positive association betwmcl-1 expression and apoptotic index in LCNNEC (p = 0.04 by Fisher´s test) but noLCNEC.
5.3. Western Blot analysis (II)
Western blot analysis was performed from freshly frozen tissue samples fromLCNEC and two LCNNEC to validate the specificity of the antibodies used. In alsamples, anti-bax and anti-bak antibodies specifically detected their corresponantigens (22 and 24 kDa). In contrast, anti-mcl-1 antibody detected its antigen (40in only one out of the three samples. The anti-bcl-2 antibody detected a faint 26-band corresponding to the known molecular weight of the bcl-2 protein.
5.4. Tumour necrosis and cell proliferation (I)
5.4.1. Ki-67 expression in SCLC
SCLC expressed high proliferative activity as determined by Ki-67 immunoreactiv58 % of SCLC showed more than 40 % the nuclei as being positive for Ki-67. Thecell type showed a higher proliferative activity than the intermediate cell type of SC(p = 0.02). The extent of apoptosis showed no association with the proliferative actas determined by Ki-67 immunoreactivity.
5.4.2. Tumour necrosis in SCLC
Tumour necrosis was seen in 83 % of SCLC (mean 25 % of tumour volume). In SCtumour necrosis was associated with p53 protein accumulation and low apoptotic actin cases with necrosis exceeding 20 % of relative tumour volume, low apoptotic indwere detected significantly more often than in less necrotic tumours (p = 0.Moreover, SCLCs containing necrotic tissue exceeding 20 % of their volume, wp53-positive significantly more often than less necrotic tumours (p = 0.02).
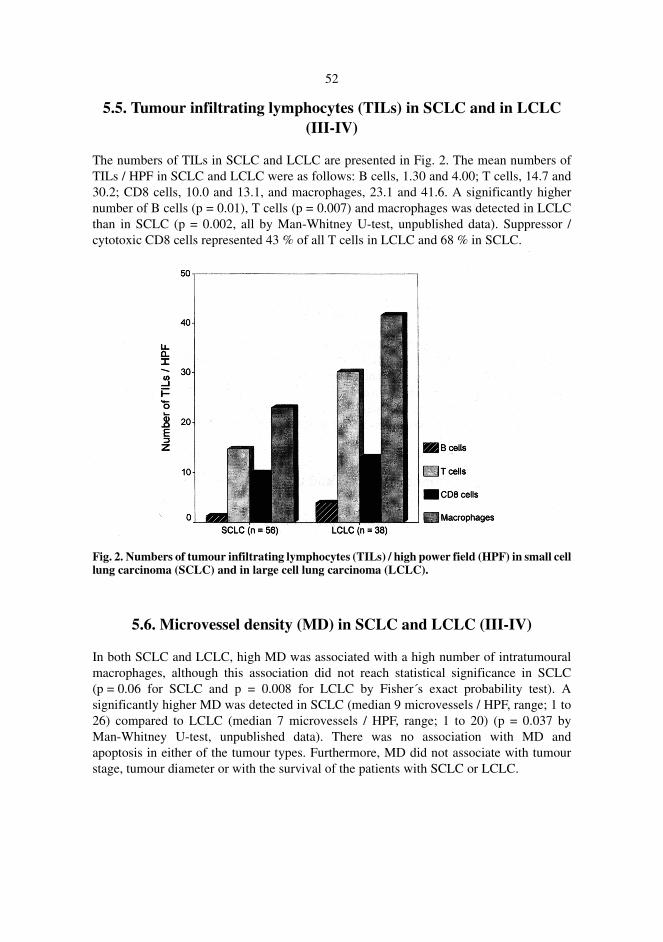
52
5.5. Tumour infiltrating lymphocytes (TILs) in SCLC and in LCLC(III-IV)
The numbers of TILs in SCLC and LCLC are presented in Fig. 2. The mean numbers ofTILs / HPF in SCLC and LCLC were as follows: B cells, 1.30 and 4.00; T cells, 14.7 and30.2; CD8 cells, 10.0 and 13.1, and macrophages, 23.1 and 41.6. A significantly highernumber of B cells (p = 0.01), T cells (p = 0.007) and macrophages was detected in LCLCthan in SCLC (p = 0.002, all by Man-Whitney U-test, unpublished data). Suppressor /cytotoxic CD8 cells represented 43 % of all T cells in LCLC and 68 % in SCLC.
Fig. 2. Numbers of tumour infiltrating lymphocytes (TILs) / high power field (HPF) in small celllung carcinoma (SCLC) and in large cell lung carcinoma (LCLC).
5.6. Microvessel density (MD) in SCLC and LCLC (III-IV)
In both SCLC and LCLC, high MD was associated with a high number of intratumouralmacrophages, although this association did not reach statistical significance in SCLC(p = 0.06 for SCLC and p = 0.008 for LCLC by Fisher´s exact probability test). Asignificantly higher MD was detected in SCLC (median 9 microvessels / HPF, range; 1 to26) compared to LCLC (median 7 microvessels / HPF, range; 1 to 20) (p = 0.037 byMan-Whitney U-test, unpublished data). There was no association with MD andapoptosis in either of the tumour types. Furthermore, MD did not associate with tumourstage, tumour diameter or with the survival of the patients with SCLC or LCLC.
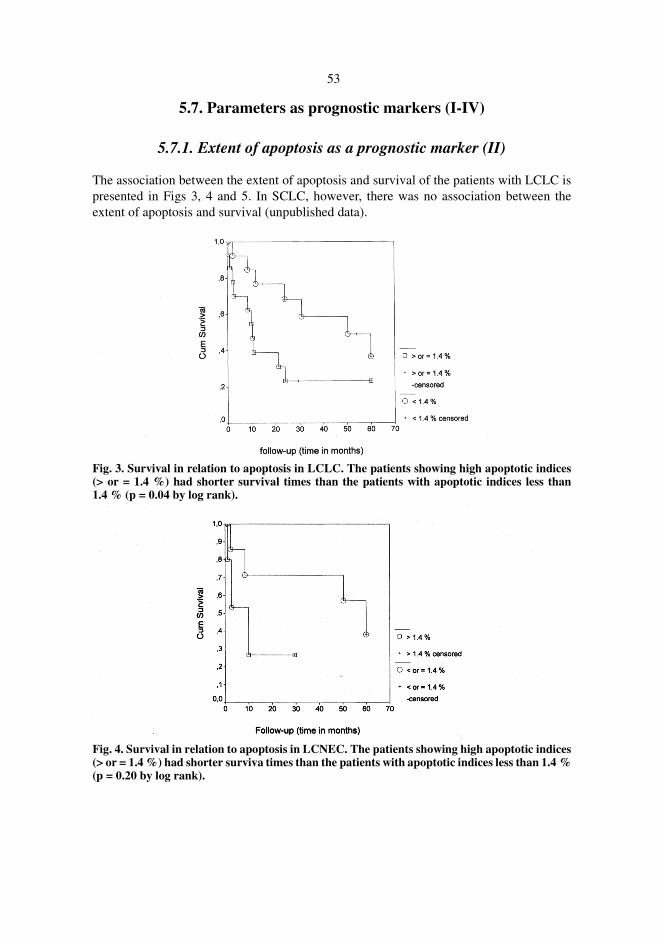
53
5.7. Parameters as prognostic markers (I-IV)
5.7.1. Extent of apoptosis as a prognostic marker (II)
The association between the extent of apoptosis and survival of the patients with LCLC ispresented in Figs 3, 4 and 5. In SCLC, however, there was no association between theextent of apoptosis and survival (unpublished data).
Fig. 3. Survival in relation to apoptosis in LCLC. The patients showing high apoptotic indices(> or = 1.4 %) had shorter survival times than the patients with apoptotic indices less than1.4 % (p = 0.04 by log rank).
Fig. 4. Survival in relation to apoptosis in LCNEC. The patients showing high apoptotic indices(> or = 1.4 %) had shorter surviva times than the patients with apoptotic indices less than 1.4 %(p = 0.20 by log rank).

54
Fig. 5. Survival in relation to apoptosis in LCNNEC. The patients showing high apoptoticindices (> or = 1.4 %) had shorter survival times than the patients with apoptotic indices lessthan 1.4 % (p = 0.03 by log rank).
5.7.2. Tumour diameter and postoperative treatment as prognosticmarkers
The patients with LCLC whose tumour diameter exceeded 4 cm had a significantlyshorter survival time than those with a smaller tumour (p = 0.05 by log rank) (II). Theextent of tumour necrosis or diameter of the tumour was not associated with survival ofthe patients with SCLC (unpublished data). None of the treatment modalities (radiationand/or chemotherapy) were associated with better survival of the patients (unpublisheddata).
5.7.3. Bcl-2 protein expression as a prognostic marker
The association between bcl-2 protein expression and survival of the patients withLCNEC is presented in Fig. 6. However, there was no association between bcl-2 andsurvival in patients with SCLC (unpublished data).

55
Fig. 6. Survival in relation to bcl-2 protein expression in LCNEC. The patients with a low bcl-2index (0-2) in their lung tumours had a significantly shorter survival time than those with a highbcl-2 index (3-6) (p = 0.04 by log rank).
5.7.4. TILs as prognostic markers (III-IV)
A high total number of TILs was a prognostic factor predicting favourable prognosis ofthe patients with SCLC (III). When analysed separately, patients with a high number ofintratumoural T cells (≥ 7.8 / HPF) and CD8 cells (≥ 5.8 / HPF) had a significantly longersurvival time than those with a low number of T cells (< 7.8 / HPF) or CD8 cells (< 5.8 /HPF) (p = 0.007 and p = 0.02 by log rank). A high number of intratumoural macrophages(≥ 19.0 / HPF) was also associated with favourable survival time of the patients withSCLC (p = 0.05 by log rank).
In LCLC a high number of intratumoural B cells (≥ 1.40 per HPF) and a high numberof intratumoural macrophages (≥ 30 per HPF) was a prognostic factor predicting afavourable prognosis of the patients (p = 0.05 and p = 0.02 by log rank) (IV). WhenLCNNECs and LCNECs were analysed separately, a high number of macrophages wasalso associated with a favourable survival of the patients with LCNNEC (p = 0.02), butnot of those with LCNEC (p = 0.16). Occurrence of intratumoural T cells was notassociated with survival in LCLC.
5.8. Apoptosis in cytological samples (V)
5.8.1. AMWABs in sputum samples
The differential cell count in Papanicolaou-stained sputum samples from patients withoutmalignancy showed a low percentage of AMWABs (Figs 7 and 8). In patients with lungcarcinoma, on the other hand, a striking feature was the high percentage of AMWABs
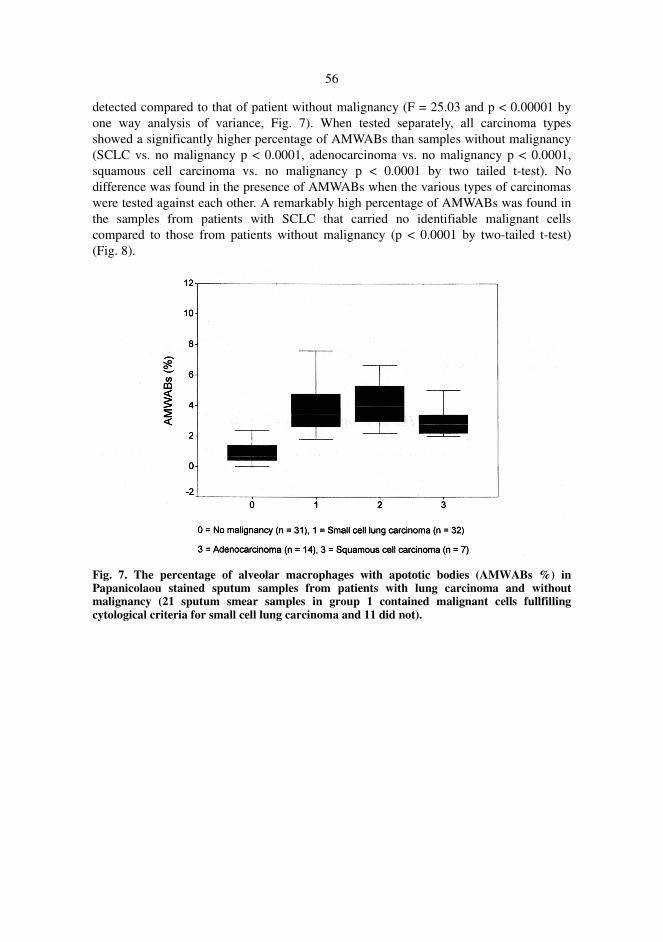
56
detected compared to that of patient without malignancy (F = 25.03 and p < 0.00001 byone way analysis of variance, Fig. 7). When tested separately, all carcinoma typesshowed a significantly higher percentage of AMWABs than samples without malignancy(SCLC vs. no malignancy p < 0.0001, adenocarcinoma vs. no malignancy p < 0.0001,squamous cell carcinoma vs. no malignancy p < 0.0001 by two tailed t-test). Nodifference was found in the presence of AMWABs when the various types of carcinomaswere tested against each other. A remarkably high percentage of AMWABs was found inthe samples from patients with SCLC that carried no identifiable malignant cellscompared to those from patients without malignancy (p < 0.0001 by two-tailed t-test)(Fig. 8).
Fig. 7. The percentage of alveolar macrophages with apototic bodies (AMWABs %) inPapanicolaou stained sputum samples from patients with lung carcinoma and withoutmalignancy (21 sputum smear samples in group 1 contained malignant cells fullfillingcytological criteria for small cell lung carcinoma and 11 did not).

57
Fig. 8. The percentage of alveolar macrophages with apoptotic bodies (AMWABs %) inPapanicolaou stained sputum samples from patients without malignancy and with SCLCwithout malignant cells.
5.8.2. Specific demonstration of the presence of apoptotic DNA inalveolar macrophages
Application of in situ 3 ´- end labelling of DNA and staining by the Feulgen andRoosenbeck (Vacca 1985) techniques yielded results highly concordant with the analysisbased on Papanicolaou staining. Labelling of DNA showed a distinct staining of theapoptotic bodies within alveolar macrophages, verifying that they did indeed representapoptotic bodies. In lung carcinoma AMWABs were detected in the same range offrequence by the in situ 3 ´ - end labelling technique (mean 3.13 %, range 1.25 - 6.25 %)as by Papanicolaou staining (mean 4.56 %, range 3.50 - 6.67 %). There was highlysignificant correspondence between these two techniques in revealing the percentage ofAMWABs (p < 0.0001).
5.8.3. AMWABs and inflammation
There was a significant correlation between the relative abundance of AMWABs and theextent of acute inflammation in the samples from the patients with lung carcinoma (F =5.08 and p = 0.005 by one way analysis of variance) with the highest association in thesamples with SCLC (p < 0.0001 by two-tailed t-test, F = 4.4 and p = 0.01 by one way

58
philshan
theutumtagein
out.25of
ut not.61 -theas0.5
alellthodsn
byhoutroupnts
and
analysis of variance). SCLC samples with a moderate or abundant number of neutroshowed approximately a four times higher percentage of AMWABs (mean 6.31 %) tthose without malignancy (mean 1.51 %) when the samples were equalized forneutrophils present in the samples. The difference was even more clear in the spsamples with a few or without any inflammatory cells; in such cases the mean percenof AMWABs was over nine times higher in cases with SCLC (mean 3.41 %) thanthose without malignancy (mean 0.36 %).
5.8.4. AMWABs in BAL samples
Light microscopical analysis of MGG stained BAL specimens from patients withcarcinoma showed a fairly low percentage of AMWABs (mean 0.51 %, range 0.0 - 1%). When usingin situ 3´- end labelling technique and a standard staining methodFeulgen and Roosenbeck, the corresponding percentages were slightly higher, bsignificantly different (mean 0.74 %, range 0.0 - 2.0 %, and mean 1.00 %, range 02.50 %, respectively). A fairly good interobserver reproducibility was obtained whennumber of AMWABs detected by a third person (Y.S.) in a set of 13 BAL samples wcompared to the original data counted by two other persons (A.-K.E. and P.P.) (R =by Pearson´s test).
For further analysis, BAL specimens were divided into two subgroups 1) normfinding or slight abnormalities in cell differential count 2) distinct abnormalities in cdifferential count. A significantly higher percentage of AMWABs in the sample widistinct abnormalities in cell differential count was seen with all three detection meth(MGG-staining, p = 0.004,in situ 3 ´- end labelling, p = 0.01 and the method of Feulgeand Roosenbeck, p = 0.05).
5.8.5. AMWABs in relation to clinical data
There was a significant increase in the percentage of AMWABs with age (p = 0.003linear regression analysis). However, when samples from patients with and witmalignancy were tested separately, a significant correlation was found only in the gof patients with malignancy (p = 0.05 by linear regression analysis) but not in patiewithout malignancy. There was no correlation between the occurrence of AMWABstotal smoking time.

ofand
n of
es ofnted
sue
aptosist in%)
eention.ostw
6. Discussion
6.1. Detection of apoptosis in tumours
The detection of apoptotic cells and bodies by using light microscopic analysisapoptosis is based on morphological features including condensation of chromatincytoplasm, fragmentation of the cell and apoptotic body formation (Kerret al.1972). Thebudding phenomenon of apoptotic cells lasts only a few minutes, but the formatioapoptotic bodies remains histologically visible for 1 to 2 hours (Barreset al. 1992). Thus,in tissues, where apoptosis affects scattered cells, apoptotic bodies of various stagdegeneration are detected. Visualisation of apoptotic cells by labelling of the fragmeDNA allows the identification of apoptotic bodies as small as 0.5µm in diameter, whichis clearly superior to the light microscopic investigation of conventionally stained tissections allowing positive identification of apoptotic bodies of 2µm diameter (Aiharaetal. 1994).
6.2. Extent of apoptosis in malignant tumours
Apoptosis, along with cell proliferation, has an important role in the growth ofneoplasm. Since the development of the techniques for measuring the extent of apoin vivo, an increasing number of papers have been published reporting its extendifferent tumours and in non-neoplastic tissues. Low apoptotic indices (less than 1have previously been reported in hepatocellular carcinoma (Soiniet al. 1996), pancreaticcarcinoma (Virkajärviet al. 1997) and in malignant salivary gland tumours (Soiniet al.1998), while moderate to high rates of apoptosis (1.20-8.10 %) have previously breported in tumours sharing neuroendocrine features and low degree of differentiaNeuroendocrine differentiation has been associated with high apoptotic activity mdistinctively in SCLC (Staunton & Gaffney 1995), while a high apoptotic index and lodifferentiation degree has been demonstrated in breast carcinoma (Mustonenet al. 1997),urogenital carcinoma (Kinget al.1996) and thyroid carcinoma (Basoloet al.1997).

60
LCherdy/:1
erateded
ofin
ce toticsibleytosed
perryofs ofharaof anHPFes in
thellingissue
e, bevity.me ofith a
s in
is inoursw
ries
In the present study, high apoptotic activity was detected in both SCLC and LC(mean apoptotic index 2.70 % and 2.49 %, respectively). In LCLC, somewhat higapoptotic activity was detected in LCNECs than in LCNNECs. The apoptotic boapoptotic cell ratio in SCLC and in LCLC was roughly 2:1, in contrast to a ratio of 1seen in non-neoplastic control tissue and cells. This suggests that there is accelapoptosis both in SCLC and in LCLC compared with the surrounding uninvolvepithelial tissues. A high apoptotic index was also seen in germinal centreshyperplastic lymph nodes. This is due to an elevated apoptotic activity in B cellsgerminal centres, while the T cells around the germinal centres showed resistanapoptosis (Yoshinoet al. 1994). Interestingly, there was a high number of apoptobodies within alveolar macrophages in the tumour-bearing lungs. It is, however, posthat these apoptotic bodies are released from the tumours and have become phagocby macrophages.
Usually, the extent of apoptosis is indicated simply as a number of apoptotic cellsHPF or per mm2 of tissue. On the other hand, it is well known that the cell sizes vafrom one tumour type to another. It is thus difficult to compare directly the levelapoptosis found in SCLC and in LCLC with the numbers presented for other typetumour, such as bladder (Lipponen & Aaltomaa 1994a) and prostatic carcinoma (Aiet al. 1994). Some theoretical calculations can be done, however, based on the sizeHPF and assuming that in SCLC there are approximately 4-5 times more cells perthan in bladder and prostatic carcinoma. Such calculations show that apoptotic indicSCLC are clearly higher than in prostatic carcinoma (Aiharaet al. 1994) and high-gradebladder cancer (Lipponen & Aaltomaa 1994a). It is possible, however, that part ofdifference could be assigned to a supposedly higher sensitivity of the in 3 ´- end labemethod when compared to the determination of apoptosis in conventionally stained tsections.
6.3. Apoptosis in relation to prognosis, cell proliferation, necrosis andage of the patients
One may assume that enhanced apoptosis will retard tumour growth and, hencindicative of a favourable prognosis compared to tumours showing low apoptotic actiThere are a few studies available on apoptosis and the prognosis for carcinomas, sowhich are contradictory. Previously, a high rate of apoptosis has been associated wshortened survival of the patients with NSCLC (Törmänenet al. 1995, Stammler & Volm1996). High apoptotic activity also seems to be connected with poor prognosihormone-dependent tumours, such as breast carcinoma (Silvestriniet al. 1994, Vakkalaetal. 1999), and in prostate carcinoma (Aiharaet al. 1994). Also Lipponen and Aaltomaa(1994) have shown that a high apoptotic index is associated with a poor prognostransitional cell carcinoma of the urinary bladder. In hepatocellular carcinoma, tumwith a high growth index, i.e. tumours showing a high proliferation activity and a lodegree of apoptosis and necrosis, were associated with a shortened survival (Soiniet al.1996). As is known for the rate of cell proliferation, also the rate of apoptosis va

61
velop
poora
rs.ty asigh
ourt the
in
menaeenthere
such
ntsosenced
ndingl other
ofheips asnd
half-. Onomes993,
blyn thatis
considerably in different tumours (Kerret al.1972, Kerr & Lamb 1984). Thus, in order touse the rate of cell death as a reliable prognostic marker, it would be essential to demethods to determine the duration of apoptosis.
In the present study, we found an association between enhanced apoptosis andprognosis in LCLC but not in SCLC. It is well known that both LCLC and SCLC showhigh mitotic activity, and hence proliferate rapidly (Brighamet al. 1978, Travis 1991).High apoptotic activity could reflect a generally high cell turnover in these tumouHowever, in the present study there was no association between proliferation actividetermined by Ki-67 immunoreactivity and apoptosis in SCLC. It is possible that the hrate of apoptosis detected in SCLC would be protracted, ineffective to limit tumgrowth and thus not associated with survival. Further studies are needed to teshypothesis that high apoptotic activity is connected with increased cell proliferationLCLC.
There is accumulating evidence that apoptosis and necrosis are related pheno(Leist & Nicotera 1997). In tumours, increased number of apoptotic cells are sadjacent to necrotic areas (Arai & Katayama 1997). In the present study, however,was an inverse association between tumour necrosis and apoptosis in SCLC, but noassociation was found in LCLC.
In SCLC significantly higher apoptotic activity was detected in tumours from patieover 65 years of age and with a long smoking history. However, all the patients whdata were included in the study were current or ex-smokers, and thus, the enhaapoptosis is rather due to tobacco smoking than to age. It is possible that a long-stasmoking habit, besides being able to cause p53 mutations, is able to cause severagene mutations, which in turn are able to modulate the extent of apoptosis.
6.4. p53 protein accumulation in SCLC
We detected abnormal p53 protein accumulation in 38 % of SCLC and in 45 %intermediate cell type and in 11 % of oat cell type of SCLC, which is in line with tresult presented by Korkolopoulouet al. (1993). There was a distinct inverse relationshbetween p53 immunohistochemistry and the extent of apoptosis in SCLC. This iexpected, in view of the well known differences in the capacity of the wild-type amutated p53 proteins to induce apoptosis; the wild type p53 which, due to its shortlife, remains immunohistochemically undetectable, is a potent inducer of apoptosisthe other hand, mutated p53, which due to its extended half-life accumulates and becdetectable by anti-p53 antibodies, is no longer capable of inducing apoptosis (Lowe 1Bardeesyet al. 1995). The inability of the mutated p53 to induce apoptosis is probadue to its decreased ability to transactivate the Bax gene, which encodes a proteibinds to bcl-2 (Miyashita & Reed 1995), a well known inhibitor of apoptos(Hockenberyet al.1990, reviewed by Reed 1994).

62
s, a
y inction
cl-2 by
BH4tion
due
s ofinwith
ith ae
thehighalrEshiftlliteurvaluate
tosiscl-2ours.ation
ansion
T
withl-2
6.5. Expression of bcl-2 family proteins in lung carcinoma
In the present study, positive bcl-2 immunoreactivity was detected in half of the SCLCfrequency similar to that reported by Ikegakiet al. (1994). Surprisingly, there was apositive association between the extent of apoptosis and bcl-2 immunoreactivitSCLC, which is contrary to what was expected based on the generally accepted funof bcl-2 as a suppressor of apoptosis. Recent evidence suggests that cleavage of bcaspase-3 may either activate or inactivate essential functions of bcl-2 (Chenget al.1997). Cleavage of bcl-2 by caspases has beem shown to result in loss of thehomology domain that is required for its antiapoptotic activity. Furthermore, the deleof this domain has been shown to release a potent proapoptotic fragment (Chenget al.1997). It is possible that also in SCLC bcl-2 had lost its ability to suppress apoptosisto the cleavage of caspase-3.
In the present study, bcl-2 immunoreactivity was detected in 40 % of all caseLCLC. It was detected significantly more often in LCNECs than in LCNNEC, which isline with some other studies suggesting that bcl-2 expression is associatedneuroendocrine differentiation (Jianget al. 1996, Reedet al. 1991, Segalet al. 1994).Bcl-2 overexpression and Bax down-regulation has previously been associated whigh apoptotic index in LCNEC (Brambillaet al.1996). However, in the present study wwere not able to find any association between either bcl-2 or bax expression andextent of apoptosis in LCLC. Instead, mcl-1 showed an interesting association withapoptotic activity in LCNNEC but not in LCNEC. This is contrary to the generexpectation that mcl-1 expression is associated with low apoptotic activity (Kroemeetal. 1997, Reynoldset al. 1994). Bax was widely expressed in LCLC regardless of Ndifferentiation. It has recently been demonstrated that certain somatic framemutations in the bax gene take place in colon cancer displaying microsateheterogeneity (Rampinoet al. 1997), suggesting that bax may be an important tumosuppressor gene in human cancer on a wider scale. Further research is needed to ebcl-2 family genes as potential tumour suppressors in lung carcinoma as well.
Based on the results of the present study, bcl-2 is not involved in suppressing apopin SCLC or in LCLC. To explain this, it can be assumed that there are also bindependent pathways that are involved in the regulation of apoptosis in these tumOne of these could be mediated by the FAS/FAS-ligand system and caspase activ(Longthorneet al.1997). In accordance with this idea, Hellquistet al. (1997) were able todemonstrate the presence of FAS receptor in squamous cell lung carcinomas. Niehetal. (1997) pointed to the presence of FAS ligand in lung carcinoma cell lines. Activatof this pathway would presuppose the presence of intratumoural cytotoxiclymphocytes.
6.5.1. Bcl-2 expression in relation to survival
There are a few studies indicating that bcl-2 expression in tumours is connectedsurvival, most of which report its association with a favourable prognosis. Bcexpression has been linked with a better survival in NSCLC (Fontaniniet al. 1995) and

63
ateg no
dtedave
ucells,
r cells
en
TILss
ofeorly
LC.n inaree
ly inwing
t thema
host
theget
with better short term survival for breast carcinoma (Silvestriniet al. 1994). In contrast,Brambillaet al. (1996) reported that bcl-2 expression correlates with a lower survival rin cases of LCNEC and CTs. However, there are some studies demonstratinassociation between bcl-2 expression and survival in lung carcinoma (Fleminget al.1998, Higashiyamaet al.1995, Törmänenet al.1995). In contrast to the results presenteby Brambilla et al. (1996), in the present study, a low bcl-2 expression was associawith a shortened survival in LCNEC. This suggests that bcl-2 expression may hprognostic value in lung carcinoma, at least in tumours showing NE differentiation.
6.6. TILs in lung carcinoma
TILs are known to have diverse functions in tumour microenvironment. They prodsoluble cytokines that regulate proliferation and metastatic activity of tumour cepromote angiogenesis and participate in host defence mechanisms against tumou(Ekert & Vaux 1997, Matsuyamaet al. 1998, Kan-Mitchellet al. 1985). Functionaldeficiency of TILs, i.e. diminished cytotoxic capacity and lowered tumour-antigrecognition (Moyet al. 1985) or immune escape of TILs by apoptosis (O´Mahoniet al.1993) are mechanisms that can lead to tumour growth.
In the present study, T cells and macrophages were the most commonly detectedin SCLC and in LCLC with a minority of B cells. This is in line with previous studiepresented by Kerret al. (1998) and Watanabeet al. (1983). Previously, the number ofTILs in lung carcinoma has been associated with tumour type and degreedifferentiation (Watanabeet al. 1983, Kerret al. 1998). Namely, the highest levels havbeen found in well differentiated squamous cell lung carcinoma and the lowest in podifferentiated SCLC (Watanabeet al. 1983, Kerret al. 1998). In the present study, theoverall number of TILs detected in SCLC was less than half of that detected in LCMoreover, CD8 cells represented a much larger proportion of T cells in SCLC thaLCLC. In some previous studies only relative numbers of TILs in lung carcinomagiven (Watanabeet al. 1983, Kerret al. 1998), and thus cannot be compared with thresults presented in this work.
6.6.1. The association between TILs and prognosis
A high number of TILs has been associated with a favourable prognosis particularfast growing tumours such as bladder cancer (Lipponen & Aaltomaa 1994a), fast grobreast tumours (Aaltomaaet al. 1992) and in melanoma (Clementeet al. 1996).According to a few studies available, there seems to be a general trend thaoccurrence of TILs is associated with a better survival of patients with lung carcino(Kerr et al. 1998, Riemannet al.1997).
According to the results presented in this study there seems to be an activedefence mechanism present in SCLC. Indeed, in line with Kerret al. (1998), it is shownhere that intratumoural T cells and cytotoxic CD8 T cells predict better survival ofpatients with SCLC. Cytotoxic T lymphocyte can specifically induce death in the tar

64
ulesgand
hastioncyteasR)andfrom
itiveasL
ungweeng atDue
hattherroml toILs in
ith ad bysticthers B
d Titing
ofourby
wn
heirtheng artant
andthat
only
cell by at least two mechanisms. The first is mediated by direct exocytosis of granthat contain perforin and granzymes, and the second involves signalling by Fas li(FasL) (Ekert & Vaux 1997). Co-operation between secreted granzymes and perforinbeen shown to cause typical cytotoxic lymphocyte induced lysis and DNA fragmenta(Henkart 1994). In the Fas mediated pathway, engagement of cytotoxic lymphomembrane ligand (FasL) with an apoptosis-inducing target cell surface receptor (Ftriggers apoptosis of the target cell (Berke 1995). The functional expression of Fas lig(FasL) on tumour cells is a mechanism proposed by several groups of tumour escapeimmunological detection of TILs (Strandet al. 1996, Bennettet al. 1998, Cardiet al.1998). Also lung carcinoma has been shown to express FasL bothin vitro and in vivo(Niehanset al. 1997). This enables lung carcinoma cells to counterattack Fas-sensTILs and thus contribute to tumour growth. On the other hand, impaired FasR/Fapoptotic pathway may contribute to immunologically nonreactive TILs detected in lcarcinoma (Holmes 1985). In the present study, there was an association betcytotoxic CD8 T cells and the occurrence of apoptotic bodies in SCLC, suggestinleast some involvement of active FasR/FasL or perforin mediated apoptotic pathway.to the positive association of apoptotic bodies and cytotoxic T cells it is likely tapoptotic bodies represent dying tumour cells attacked by cytotoxic T cells. Anopossibility would be that apoptotic bodies are remains of activated T cells resulting fthe down-regulation of the immune response. To verify this, it would be essentiaevaluate the expression of FasL and FasR and their association to apoptosis and Tthese tumours.
In the present study, an increased number of B cells in tumours was associated wbetter survival of the patients with LCLC, which is supported by the results presenteRiemannet al. (1997) also suggesting that B cells are a more significant prognomarker in NSCLC than T cells. T cells secrete several cytokines, which activate omononuclear cells. Among these are B cell growth factor (BCGF), which stimulatecell differentiation and proliferation, interferon-λ (INFλ) which activates macrophagesand promotes cytotoxic functions, and IL-2 which stimulates proliferation of activatecells (Herberman 1997). There is evidence that tumour cells are capable of inhibimmune response by producing factors that downregulate IL-2 and INFλ production(Guentheret al. 1993). This downregulation could explain why a fairly large numberintratumoural T cells was not associated with the survival of the patients or with tumsize in LCLC. B cells participate actively in the tumour mediated immune responsesecreting lymphokines. One of these lymphokines is lymphotoxin (LT), which is knoto kill target tumour or virus infected cells (Quinet al. 1995). According to animalexperiments, LT is a potent cytokine against tumour growth and metastasisin vivo(Funahashiet al. 1991). Activated B cells also produce IL-12, which in turn induces tproduction of INFλ from natural killer cells, thus activating them and enhancing thecytotoxic activity against tumour target cells (Trinchieri 1995). In the present study,occurrence of a high number of intratumoural B cells was a prognostic marker showibetter survival time, suggesting that lymphokines secreted by B cells might be impomediators for the regulation of the tumour growth in LCLC.
Macrophages are pivotal cells in both humoral and cell-mediated immunityparticipate in host defence against cancer (Herberman 1997). However, it is possiblepart of intratumoural macrophages in lung carcinoma accumulate in tumour tissue

65
moureolar
with
ighofand/
and a
tionl
is insis inngment
esselntated
d toeen
earstumentsble
ion ofonarycells
n ofis of
arlyof
ate
secondary to inflammation or represent alveolar macrophages entrapped by the tuand serve as phagocytes of foreign particles. Previously, human pulmonary alvmacrophages have been shown to be cytotoxic towards tumour cells (Kan-Mitchellet al.1985) and to produce growth inhibitory factors (Graveset al. 1989). In the present study ahigh number of macrophages was associated with better survival of the patientsSCLC and LCLC, which is in accordance with the results presented by Kerret al. (1998).A high microvessel density (MD) in both SCLC and LCLC was associated with a hnumber of intratumoural macrophages, which is well in line with the knowledgemacrophage function to promote angiogenesis by secreting proangiogenic cytokinesor enzymes that degrade the extracellular matrix (Lewiset al. 1995). Also Leeket al.(1996) detected a significant association between an increased macrophage indexhigh vascular grade in breast carcinoma.
6.7. Tumour angiogenesis in lung carcinoma
Despite the undoubtedly important role of angiogenesis in tumour growth and formaof metastases (Graveset al. 1989, Folkmanet al. 1990), in the present study microvessedensity (MD) was not associated with metastatic status, tumour size or prognosSCLC or in LCLC. Previously, microvessel count has been associated with metastaNSCLC (Graveset al. 1989, Macchiarini 1998). It is suggested, however, that lucarcinomas and especially their peripheral parts use preserved alveolar basemembranes for their local spread and are thus not dependent on new microvformation (Pääkköet al. 1990). Fontaniniet al. (1997) reported that angiogenesis iNSCLC is under p53 tumour-suppressor gene control. Their results suggest that mup53 protein promotes angiogenesis through VEGF expression. This would leaabrogation of p53 mediated apoptosis. However, we found no association betwapoptosis and MD in SCLC or in LCLC.
6.8. Apoptosis in cytological samples
In the present study it is shown for the first time that macrophages in sputum smcontain apoptotic bodies and evidence is provided that, in conventional clinical spusmear and BAL specimens, an elevated number of AMWABs can be found in patiwith lung cancer, irrespective of the presence or absence of cytologically verifiatumour cells in the sample. Thus, the results of the present study suggest that detectapoptotic macrophages can be used as an adjunct in the diagnosis of pulmcarcinoma and that it might be especially valuable in those cases were carcinomacannot be detected. This is of great importance, since currently positive identificatiolung cancer can be reached in only 42 to 63 % of the peripheral lesions on the bassputum smears (Nasiellet al 1967, Dahlgren & Lind 1972, Tanakaet al. 1985). It is alsoknown that combining sputum cytology and chest X-ray has not improved the edetection of lung cancer (Mulshine & Scott 1995). Thus adding determinationAMWABs into routine sputum analysis has the potential of improving the detection r

66
h ass withicaletrict of
velyfled by
inctingis
xtraacute
ver,ry
l of
Bsientsheto thetotic, this
and
foundtumotheralso
of lung carcinomas, e.g. in patients with an increased risk of lung cancer, sucsmokers. Whether the concept of the occurrence of apoptotic bodies or macrophageapoptotic bodies as an indicator of malignancy is valid also in other types of cytologsamples, such as pleural and peritoneal fluid, remains to be studied. Flow cytomtechniques, for example, could provide possibilities to reveal and quantify the extenapoptosis rapidly (Ormerod 1994, Perryet al. 1994).
Apoptotic bodies were readily detectable by conventional staining and were positiidentified as apoptotic bodies by usingin situ 3 ´ - end labelling and standard method oFeulgen and Roosenbeck (Vacca 1985). The percentage of apoptotic bodies reveathese two specific methods was not substantially different from that detectedPapanicolaou-stained samples. Moreover, the interobserver reproducibility of deteAMWABs was fairly good. Thus, sufficiently accurate detection of apoptotic bodiespossible in conventional cytological sputum smear and BAL specimens without any eprocedures. In the present study, there was a weak correlation between the extent ofinflammation and the occurrence of AMWABs, especially in cases of SCLC. Howethe percentage of AMWABs was distinctively higher in SCLC even when iflammatocells were not present. Thus, inflammation does not significantly affect the leveAMWABs in lung carcinoma.
In this study, there was a significant correlation between the percentage of AMWAand the age of the patient. A much lower correlation was seen, however, in patwithout malignancy, indicating that the occurrence of AMWABs is mostly due to tpresence of cancer and not to ageing. Age as such may have a contributing roleprevalence of tumour associated apoptosis, since in SCLC significantly higher apopindices were detected in the tumours of the patients over 65 years of age. Obviouslycould also be a reflection of differences in the biological nature of SCLC of youngold age.
Because an increased number of apoptotic bodies in alveolar macrophages wasaround tumour tissue in SCLC, it is most likely that apoptotic bodies seen in spusmears and BAL samples originate from tumour cells undergoing apoptosis. On thehad, it is evident that the apoptotic bodies seen in association with inflammationresult from inflammatory cells undergoing apoptosis.

s ofs and
theelywas
edooris inbewaste ofourtrary
oftiontein
val ininguman
ofand
Theand
asR/th inal inwasouse tond if
7. Summary and Conclusions
Impaired apoptosis due to the expression of bcl-2 family oncogenes or mutationtumour suppressor genes leads to the uncontrolled accumulation of malignant celleventually to formation of cancer. In the present study, apoptosis and its role inprognosis of the patients was investigated in highly malignant lung tumours, namsmall and large cell lung carcinoma. Finally, the detection of enhanced apoptosisapplied as a marker of lung carcinoma in diagnostic respiratory cytology.
Surprisingly, both highly malignant lung tumours, SCLC and LCLC showremarkably high apoptotic indices. High apoptotic activity was associated with pprognosis in LCLC, but no association was found between apoptosis and prognosSCLC. High apoptotic activity could reflect a generally high turnover rate orprotracted and ineffective to limit growth in these tumours. P53 protein accumulationdetected in nearly half of both tumour types and was associated with decreased raapoptosis in SCLC. This supports the previous results indicating that p53 tumsuppressor has an important role in the genesis of lung carcinoma. In this study, conto the generally accepted function of bcl-2 it was not involved in the suppressonapoptosis in SCLC nor in LCLC. On the contrary, there was a positive associabetween bcl-2 protein expression and apoptosis in SCLC. Furthermore, bcl-2 proexpression was associated with neuroendocrine differentation and shortened surviLCLC. Bax was widely expressed in LCLC regardless of NE differentation, supportsome previous ideas that bax might be an important tumour suppressor gene in hcancer on a wider scale. In conclusion, the role of bcl-2 family proteins in regulationapoptosis in lung carcinoma is complicated and needs to be studied further. T cellsmacrophages were the most commonly detected TILs in SCLC and LCLC.occurrence of intratumoural cytotoxic CD8 cells was associated with apoptotic bodiespredicted favourable prognosis in the patients with SCLC. This indicates that the FFasL apoptotic pathway may be a key mechanism in the regulation of tumour growSCLC. On the other hand, there was no association between T cells and survivLCLC. Instead, the high number of intratumoural B cells and macrophagesassociated with better survival of the patients with LCLC. In this study, numerapoptotic bodies could also be found within alveolar macrophages within and clostumour tissue. In order to test whether such cells could be found in sputum smears, a

68
, thefromns,
wasntsfore,than
their presence could be used as a marker of malignancy in tumour diagnosisoccurrence of AMWABs was analysed in a set of sputum smear and BAL samplespatients with and without a pulmonary malignancy. In sputum and BAL specimeenhanced apoptosis, as measured by increased number of AMWABs reflected andthus indicative of malignancy. This was also true for cytological specimens of patiewith lung carcinoma, even when the actual malignant cells were not present. Therethe occurrence of AMWABs served as a more sensitive marker for lung carcinomathe malignant cells per se.

2)-864.of
: 797-
-cell
hol
.
P &lysis
90)200-
ilm´s
ell
ts of
ion541.ks
5.
ace
Theith
8. References
Aaltomaa S, Lipponen P, Eskelinen M, Kosma V-M, Marin S, Alhava E & Syrjänen K (199Lymphocyte infiltrates as a prognostic variable in female breast cancer. Eur J Cancer 28:859
Aihara M, Truong LD, Dunn JK, Wheeler TM, Scardino PT & Thompson TC (1994) Frequencyapoptotic bodies positively correlates with Gleason grade in prostate cancer. Hum Pathol 25801.
Arai E & Katayama I (1997) Role of apoptosis in spontaneous regression of peripheral Tlymphoma arising in the skin subcutis. Hum Pathol 28: 472-477.
Arends MJ, Morris RG & Wyllie AH (1990) Apoptosis. The role of endonuclease (1990) Am J Pat136: 593-608.
Arrigoni MG, Woolner LB, Bernatz PE & Minn R (1972) Atypical carcinoid tumors of the lungThorac Cardiovasc Surg 64: 413-421.
Bakhshi A, Wright JJ, Graninger W, Seto M, Owens J, Cossman J, Jensen JP, GoldmanKorsmeyer SJ (1987) Mechanism of the t(14;18) chromosomal translocation: structural anaof both derivative 14 and 18 reciprocal partners. Proc Natl Acad Sci USA 84: 2396-2400.
Balch CM, Riley LB, Bae YJ, Salmeron MA, Plautsoucas CD, von Eschenbach A & Itoh K (19Patterns of human tumour-infiltrating lymphocytes in 120 human cancers. Arch Surg 125:205.
Bardeesy N, Beckwith JB & Pelletier J (1995) Clonal expansion and attenuated apoptosis in Wtumours are associated wtih p53 gene mutations. Cancer Res 55: 215-219.
Barres BA, Hart IK, Coles HS, Burne JF, Voyvodic JT, Richardson WE & Raff MC (1992) Cdeath and control of cell survival in the oligodendrocyte lineage. Cell 70: 31-46.
Barrett JC, Lamb PW & Wiseman RV (1989) Multiple mechanisms for the carcinogenetic effecasbestos and other mineral fibres. Environ Health Perspect 81: 81-89.
Basolo F, Pollina L, Fontanini G, Fiore L, Pacini F & Baldanzi A (1997) Apoptosis and proliferatin thyroid carcinoma: correlation with bcl-2 and p53 protein expression. Br J Cancer 75: 537-
Bates S, Phillips AC, Clark PA, Stott F, Peters G, Ludwig RL & Vousden KH (1998) p14ARF linthe tumour suppressors RB and p53. Nature 395: 124-125.
Beckett WS (1993) Epidemiology and etiology of lung cancer (review). Clin Chest Med 14: 1-1Berke G (1995) The CTL ´s kiss of death. Cell 81: 9-12.Bennett M, Macdonald K, Chan SW, Luzio JP, Simari R & Weissberg P (1998) Cell surf
trafficking of fas: a rapid mechanism of p53 mediated apoptosis. Science 282: 290-293.Bennett MW, O´Connell J, O´Sullivan GC, Brady C, Roche D, Collins JK & Shanahan F (1998)
fas counterattack in vivo: apoptotic depletion of tumor-infiltrating lymphocytes associated wfas ligand expression by human esophageal carcinoma. J Immunol 160: 5669-5675.

70
6)149:
93-
of
esis and
e of39.
or-Res
nalells
nol
7)
5)
and
lema.
res286:
nat
ney
8-
racic
y
D,p53
1B.
Brambilla E, Negoescu A, Gazzeri S, Lantuejoul S, Moro D, Brambilla C & Coll JL (199Apoptosis-related factors p53, bcl-2, and bax in neuroendocrine lung tumors. Am J Pathol1941-1952.
Brenstein C, McGoey J & Lertzman M (1989) Recurrent bronchial carcinoid tumor. Chest 95: 6694.
Brigham BA, Bunn PA, Minna JD, Cohen MH, Ihde DC & Shackney SE (1978) Growth ratessmall cell bronchogenic carcinomas. Cancer 42: 2880-2886.
Brimmell M, Mendiola R, Mangion J & Packham G (1998) Bax frameshift mutations in cell linderived from human haemopoietic malignancies are associated with resistance to apoptosmicrosatellite instability. Oncogene 16: 1803-1812.
Brown DG, Sun XM & Cohen GM (1993) Dexamethasone-induced apoptosis involves cleavagDNA to large fragments prior to internucleosomal fragmentation. J Biol Chem 268: 3037-30
Carcon DA & Ribeiro JM (1993) Apoptosis and disease. Lancet 341: 1251-1254.Cardi G, Heaney JA, Schned AR & Ernstoff MS (1998) Expression of Fas(APO-1/CD95) in tum
infiltrating and peripheral blood lymphocytes in patients with renal cell carcinoma. Cancer58: 2078-2080.
Carter D (1983) Small-cell carcinoma of the lung. Am J Surg Pathol 7: 787-795.Cattoretti G, Becker MH, Key G, Duchrow M, Schluter C, Galle J & Gerdes J (1992) Monoclo
antibodies agains recombinant part of Ki-67 antigen (MIB1 and MIB3) detect proliferating cin microwave-processed formalin-fixed paraffin sections. J Pahtol 168: 357-363.
Chao DT & Korsmeyer SJ (1998) Bcl-2 family: regulators of cell death (review). Annu Rev Immu16: 395-419.
Cheng EHY, Kirsch DG, Clem RJ, Ravi R, Kastan MB, Bedi A, Ueno K & Hardwick JM (199Conversion of bcl-2 to a bax-like death effector by caspases. Science 278: 1966- 1968.
Chittenden T, Harrington EA, O´Connor R, Flemington C, Lutz RJ, Evan GI & Guild BC (199Induction of apoptosis by the bcl-2 homologue bak. Nature 374: 733-736.
Clarke PG & Clarke S (1996) Nineteenth century research on naturally occurring cell deathrelated phenomena (review). Anat Embryol 193: 81-99.
Clemente CG, Mihm MCJ, Bufalino R, Zurrida S, Collini P & Cascinelli N (1996) Prognostic vauof tumor infiltrating lymphocytes in the vertical growth phase of primary cutaneus melanoCancer 77: 1303-1310.
Cohen GM, Sun XM, Snowden RT, Dinsdale D & Skilleter DN (1992) Key morphological featuof apoptosis may occur in the absence of internucleosomal DNA fragmentation. Biochem J331-334.
Colby TV, Wistuba II & Gazdar A (1998) Percursors to pulmonary neoplasia (review). Adv APathol 5: 205-215
Coles HS, Burne JF & Raff MC (1993) Large-scale normal cell death in the developing rat kidand its reduction by epidermal growth factor. Development 118: 777-784.
Cummings MC, Winterford CM & Walker NI (1997) Apoptosis (review). Am J Surg Pathol 21: 8101.
Dahlgren SE & Lind B (1972) Comparison between diagnostic results obtained by transthoneedle biopsy and sputum cytology. Acta Cytol 16: 53-58.
Dameron KM, Volpert OV, Tainsky MA & Buck N (1994) Control of angiogenesis in fibroblasts bp53 regulation of thrombospondin-1. Science 265: 1582-1584.
D´Amico D, Carbone D, Mitsudomi T, Nau M, Fedorko J, Russell E, Johnson B, BuchhagenBodner S, Phelps R, Gazdar A & Minna JD (1992) High frequency of somatically acquiredmutations in small cell lung cancer cell lines and tumors. Oncogene 7: 339-346.
Debbas M & White E (1993) Wild-type p53 mediates apoptotsis by E1A, which is inhibited by EGenes Dev 7: 546-554.

71
99)
ene
ive
er17-
ev
ed
rltered
laarcl-athol
er Inst
cer.
)gical
titheir. Br
a Hwthther
)457-
t
past
ctive
hill
Dickman PW, Hakulinen T, Luostarinen T, Pukkala E, Sankila R, Söderman B & Teppo L (19Survival of cancer patients in Finland 1955-1994. Acta Oncol 38: 43.
Denissenko MF, Pao A, Tang M & Pfeifer GP (1996) Preferential formation of benzy(a)pyradducts at lung cancer mutational hotspots in p53. Science 274: 430-432.
Downey RS, Sewell W & Mansour KA (1989) Large cell carcinoma of the lung. a highly aggresstumour with dismal prognosis. Ann Thorac Surg 47: 806-808.
Ekert PG & Vaux DL (1997) Apoptosis and immune system. Br Med Bulletin 53: 591-603.El-Deiry WS, Tokino T, Velculescu VE, Levy DB, Parsons R, Trent JM, Lin D, Mercer WE, Kinzl
KW & Vogelstein B (1993) WAF1, a potential mediator of p53 tumor suppression. Cell 75: 8825.
Ellis RE, Yuan JY & Horvitz HR (1991) Mechanisms and functions of cell death (review). Annu RCell Biol 7: 663-698.
Fielding JE & Phenow KJ (1988) Health effects of involuntary smoking (review). New Engl J M319: 1452-1460.
Finlay CA, Hinds PW, Tan TH, Eliyahu D, Oren M & Levin AJ (1988) Activating mutations fotransformation by p53 produce a gene product that forms a hsc-70-p53 compex with an ahalf life. Mol Cell Biol 8: 531-539.
Fleming MV, Guinee DG, Chu WS, Freedman AN, Caporaso NE, Bennett WP, Colby TV, TazeH, Abbondanzo SL, Jett J, Pairolero P, Trastek V, Liotta LA, Harris CC & Travis WD (1998) B2 immunohistochemistry in a surgical series of non-small cell lung cancer patients. Hum P29: 60-64.
Folkman J (1990) What is the evidence that tumors are angiogenesis dependent ? J Natl Canc82: 4-6.
Fontanini G, Vignati S, Bigini D, Mussi A, Lucchi M, Angeletti CA, Basolo F & Bevilacqua G(1995) Bcl-2 protein: a prognostic factor inversely correlated to p53 in non-small cell lung canBr J Cancer 71: 1003-1007.
Fontanini G, Vignati S, Bigini D, Mussi A, Lucchi M, Chiné S, Angeletti C & Bevilacqua G (1996Recurrence and death in non-small cell lung carcinomas: a prognostic model using patholoparameters, microvessel count and gene protein products. Clin Cancer Res 2: 1067-1075.
Fontanini G, Vignati S, Lucchi M, Mussi A, Calcinai A, Boldrini L, Chine S, Silvestri V, AngeletCA, Basolo F & Bevilacqua G (1997) Neoangiogenesis and p53 protein in lung cancer:prognostic role and their relation with vascular endothelial growth factor (VEGF) expressionJ Cancer 75: 1295-1301.
Funahashi I, Kawatsu M, Kajikawa T, Takeo K, Asahi T, Kakutani T, Yamashita T, Kawaharad& Watanabe K (1991) Usefulness of glycosylated recombinant human lymphotoxin for groinhibition of human and murine solid tumors and experimental metastasis in mice. J Immuno10: 28-38.
Fujino M, Dosaka-Akita H, Harada M, Hiroumi H, Kinoshita I, Akie K & Kawakami Y (1995Prognostic significance of p53 and p21 expression in nonsmall cell lung cancer. Cancer 76: 22463.
Gaffney EF, O´Neill AJ & Staunton MJ (1995)In situ end-labelling, light microscopic assessmenand ultrastructure of apoptosis in lung carcinoma. J Clin Pathol 48: 1017-1021.
Garfinkel L & Silverberg E (1991) Lung cancer and smoking trends in the United States over the25 years (review). Cancer J Clin 41: 137-145.
Gavrieli Y, Sherman Y & Ben-Sasson SA (1992) Identification of programmed cell deathin situ viaspecific labeling of nuclear DNA fragmentation. J Cell Biol 119: 493-501.
Gerdes J, Schwab U, Lemke H & Stein H (1984) Production of a mouse monoclonal antibody reawith a human nuclear antigen associated with cell proliferation. Int J Cancer 31: 13-20.
Glisson BS & Hong WK (1997) Survival after treatment of small cell lung cancer: an endless upbattle. J Natl Cancer Inst 89: 1745-1747.

72
alys.
man
7-
JA,as
996)ture
Rng
tic
ssor
ing
,genof
logy.
uring
ein
. N
ility
arly
ssedd. Br J
onal
tor
l-2
Gorczyca W, Gong J & Darzynkiewicz Z (1993) Detection of DNA strand breaks in individuapoptotic cells by thein situ terminal deoxynucleotidyl transferase and nick translation assaCancer Res 53: 1945-1951.
Gosney JR, Sissons MJ & Allibone RO (1988) Neuroendocrine cell populations in normal hulungs: a quantitative study. Thorax 43: 878-882.
Gottlieb TM & Oren M (1996) p53 in growth control and neoplasia. Bioch Biophys Acta 1287: 7102.
Gould VE, Wiedenmann B, Lee I, Schwechheimer K, Dockhorn-Dworniczak B, RadosevichMoll R & Franke WW (1987) Synaptophysin expression in neuroendocrine neoplasmsdetermined by immunohistochemistry. Am J Pathol 126: 243-257.
Graeber TG, Osmanian C, Jacks T, Housman, DE, Koch CJ, Lowe SW & Giaccia AJ (1Hypoxia-mediated selection of cells with diminished apoptotic potential in solid tumours. Na379: 88-91.
Grasil-Kraupp B, Ruttkay-Nedecky B, Koudelka H, Bukowska K, Bursch W & Schulte-Hermann(1995) In situ detection of fragmentated DNA (TUNEL assay) fails to discriminate amoapoptosis, necrosis, and autolytic cell death: a cautionary note. Hepatology 21: 1465-1468.
Graves DT, Jiang YL, Williamson MJ & Valente AJ (1989) Identification of monocyte chemotacactivity produced by malignant cells. Science 245: 1490-1493.
Greenblatt MS, Bennett WP, Hollstein M & Harris CC (1994) Mutations in the p53 tumor suppregene: clues to cancer etiology and molecular pathogenesis. Cancer Res 54: 4855-4878.
Guyatt GH & Newhouse MT (1985) Are active and passive smoking harmful ? Determincausation. Chest 88: 445-451.
Hall PA, Levison DA, Woods AI, Yu CCW, Kellock B, Watkins JA, Barnes DM, Gillett CECamplejohn R, Dover R, Waseem NH & Lane DP (1990) Proliferating cell nuclear anti(PCNA) immunolocalization in paraffin sections: an index of cell proliferation with evidencederegulated expression in some neoplasms. J Pathol 162: 285-294.
Hammar S (1994) Common neoplasms. In : Dail DH & Hammar SP (eds) Pulmonary PathoSpringer-Verlag, New York, p. 1123-1278.
Hanahan D & Folkman J (1996) Patterns and emerging mechanisms of the angiogenic switch dtumorigenesis. Cell 86: 353-364.
Harper JW, Adami GR, Wei N, Keyomarsi K & Elledge SJ (1993) The p21 Cdk-interacting protCip1 is a potent inhibitor of G1 cyclin-dependent kinases. Cell 75: 805-816.
Harris CC & Hollstein M (1993) Clinical implications of the p53 tumor suppressor gene (review)Engl J Med 329: 1318-1327.
Hartwell L (1992) Defects in a cell cycle checkpoint may be responsible for the genomic instabof cancer cells (review) Cell 71: 543-546.
Harvey NL, Butt AJ & Kumar S (1997) Functional activation of Nedd2/ICH-1 (caspase-2) is an eprocess in apoptosis. J Biol Chem 272: 13134-13139.
Hellquist HB, Olejnicka B, Jadner M, Andersson T & Sederholm C (1997) Fas receptor is exprein human lung squamous cell carcinomas, whereas bcl-2 and apoptosis are not pronounceCancer 76: 175-179.
Hengartner M (1998) Apoptosis. Death by crowd control. Science 281: 1298-1299.Hengartner MO & Horvitz HR (1994) C. Elegans cell survival gene ced-9 encodes a functi
homolog of the mammalian proto-oncogene bcl-2. Cell 76: 665-676.Henkart PA (1994) Lymphocyte-mediated cytotoxicity: two pathways and multiple effec
molecules. Immunity 1: 343-346.Herberman RB (1997) Cellular Immunity. In: Crystal RG & West JB (eds) The Lung. 2nd edn.
Lippincott-Raven Publishers, Philadelphia. p. 511-533.Higashiyama M, Doi O, Kodama K, Yokouchi H & Tateishi R (1995) High prevalence of bc
oncoprotein expression in small cell lung cancer. Anticaner Res 15: 503-505.

73
er
s in: 369-
ed-153.bylin
ed
3:
land.
p53
728.g
ndncer
sion837-
al in
use
&Res
53ncer
4)
cer
e-
in
ry
Hockenbery D, Nuñez G, Milliman C, Schreider RD & Korsmeyer S (1990) bcl-2 is an innmitochnodrial membrane protein that blocks programmed cell death. Nature 348: 334-336.
Hollstein M, Soussi T, Thomas G, vonBrevern MC & Bartsch 2nd (1991) p53 gne alterationhuman tumors: perspectives for cancer control (review). Recent Results Cancer Res 143389.
Holmes EC (1985) Immunology of tumor infiltrating lymphocytes. Ann Surg 201: 158-163.Holmgren L, O´Reilly MS & Folkman J (1995) Dormancy of micrometastases: balanc
proliferation and apoptosis in the presence of angiogenesis suppression. Nature Med 1: 149Howie SE, Sommerfield AJ, Gray E & Harrison DJ (1994) Peripheral T lymphocyte depletion
apoptosis after CD4 ligation in vivo: selective loss of CD44- and activating memory T cells. CExp Immunol 95: 195-200.
Hsu SM (1990) Immunohistochemistry. Methods Enzymol 184: 357-363.Huhti E, Sutinen S, Reinilä A, Poukkula A & Saloheimo M (1980) Lung cancer in a defin
geographical area: history and histological types. Thorax 35: 660-667.Humke EW, Ni J & Dixit VM (1998) ERICE, a novel FLICE-activated caspase. J Biol Chem 27
15702-15707.Huuskonen MS, Karjalainen A, Tossavainen A & Rantanen J (1995) Asbestos and cancer in Fin
Med Lav 86: 426-434.Iggo R, Gatter K, Bartek J, Lane D & Harris AL (1990) Increased expression of mutant forms of
oncogene in primary lung cancer. Lancet 335: 675-679.Ihde DC (1984) Current status of therapy for small cell carcinoma of the lung. Cancer 54: 2722- 2Ikegaki N, Katsumata M, Minna J & Tsujimoto Y (1994) Expression of bcl-2 in small cell lun
carcinoma cells. Cancer Res 54: 6-8.Ishida H, Irie K, Itoh T, Furukawa T & Tokunaga O (1997) The prognostic significance of p53 a
bcl-2 expression in lung adenocaricnoma and its correlation with Ki-67 growth fraction. Ca80: 1034-1045.
Jiang SX, Kameya T, Sato Y, Yanase N, Yoshimura H & Kodama T (1996) Bcl-2 protein expresin lung cancer and close correlation with neuroendocrine differentation. Am J Pathol 148:846.
Joensuu H, Pylkkänen L & Toikkanen S (1994) Bcl-2 protein expression and long-term survivbreast cancer. Am J Pathol 145: 1191-1198.
Kamijo T, Zindy F, Rousse MF, Quelle DE & Downing JR (1997) Tumor suppression at the moINK4a locus mediated by the alternative reading frame product p19ARF. Cell 91: 649-659.
Kan-Mitchell J, Hengst JCD, Kempf RA, Rothbart RK, Simons SM, Brooker AS, Kortes VLMitchell MS (1985) Cytotoxic activity of human pulmonary alveolar macrophages. Cancer45: 453-458.
Kargi HA, Aktas S, Sagol O, Ermete S, Akpinar O & Akkoclu A (1997) Apoptosis, bcl-2 and pexpresion and their relation to tumour stage in non-small cell lung carcinomas (NSCLC). CaLett 116: 185-189.
Kawai T, Suzuki M, Kono S, Shinomiya N, Rokutanda M, Takagi K, Ogata T & Tamai S (199Proliferating cell nuclear antigen and Ki-67 in lung carcinoma. Cancer 74: 2468-2475.
Kerr JF, Winterford CM & Harmon BV (1994) Apoptosis. Its significance in cancer and cantherapy. Cancer 73: 2013-2026.
Kerr JF, Wyllie AH & Currie AR (1972) Apoptosis: a basic biological phenomenon with widranging implications in tissue kinetics. Br J Cancer 26: 239-257.
Kerr KM, Johnson SK, Kling G, Kennedy MM, Weir J & Jeffrey R (1998) Partial regressionprimary carcinoma of the lung: does it occur ? Histopathology 33: 55-63.
Kerr KM & Lamb D (1984) Actual growth rate and tumour cell proliferation in human pulmonaneoplasms. Br J Cancer 50: 343-349.

74
ion
andsion.
ge
small93-
996). Am
&sponcereast
5b)319.614-
of
hage
5)r. J
ofancer
236:
ast
sis:
ostic
ed to
gancer
ee
Kiefer MC, Brauer MJ, Powers VC, Wu JJ, Umansky SR, Tomei LD & Barr PJ (1995) Modulatof apoptosis by the widely distributed bcl-2 homologue Bak. Nature 374: 736-739.
King ED, Matteson J, Jacobs SC & Kyprianou N (1996) Incidence of apoptosis, cell proliferationbcl-2 expression in transitional cell carcinoma of the bladder: association with tumor progresJ Urol 155: 316-320.
Konishi T, Lin Z, Fujino S, Kato H & Mori A (1997) Association of p53 protein expression in staI lung adenocarcinoma with reference to cytologicl subtypes. Hum Pathol 28: 544-548.
Korkolopoulou P, Oates J, Crocker J & Edwards J (1993) p53 expression in oat and non-oatcell lung carcinomas: correlations with proliferating cell nuclear antigen. J Clin Pathol 46: 101096.
Krajewska M, Krajewski S, Epstein JI, Shabaik A, Sauvageot J, Song K, Kitada S & Reed JC (1Immunohistochemical analysis of bcl-2, bax, bcl-X, and mcl-1 expression in prostate cancersJ Pathol 148: 1567-1576.
Krajewski S, Blomqvuist C, Franssila K, Krajewska M, Wasenius VM, Niskanen E, Nordling SReed JC (1995a) Reduced expression of proapoptotic gene bax is associated with poor rerates to combination chemotherapy and shorter survival in women with metastatic badenocarcinoma. Cancer Res 55: 4471-4478.
Krajewski S, Bodrug S, Krajewska M, Shabaik A, Gascoyne R, Berean K & Reed JC (199Immunohistochemical analysis of mcl-1 protein in human tissues. Am J Pathol 146: 1309- 1
Kroemer G (1997) The proto-oncogene bcl-2 and its role in regulation apoptosis. Nature Med 3:620.
Kroemer G, Petit P, Zamzami N, Vayssière JL & Mignotte B (1995) The biochemistryprogrammed cell death (review). FASEB J 9: 1277-1287.
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriopY4. Nature 227: 680-685.
Lane DP (1992) p53, guardian of the genome. Nature 358: 15-16.Lee JS, Yoon A, Kalapurakal SK, Ro JY, Lee JJ, Tu N, Hittelman WN & Hong WK (199
Expression of p53 oncoprotein in non-small cell lung cancer: a favourable prognostic factoClin Oncology 13: 1893-1903.
Leek RD, Lewis CE, Whitehouse R, Greenall M, Clarke J & Harris AL (1996) Associationmacrophage infiltration with angiogenesis and prognosis in invasive breast carcinoma. CRes 56: 4625-4629.
Leist M & Nicotera P (1997) The shape of cell death (review). Biochem Biophys Res Commun1-9.
Levine AJ, Momand J & Finlay CA (1991) The tumour suppressor gene. Nature 315: 453-456.Lewis CE, Leek R, Harris A & McGee JO (1995) Cytokine regulation of angiogenesis in bre
cancer: the role of tumor-associated macrophages. J Leukocyte Biol 57: 747-751.Liebermann D, Hoffman B & Steinman R (1995) Molecular controls of growth arrest and apopto
p53-dependent and independent pathways. Oncogene 11: 199-210.Lipponen PK & Aaltomaa S (1994a) Apoptosis in bladder cancer as related to standard progn
factors and prognosis. J Pathol 173: 333-339.Lipponen P, Aaltomaa S, Kosma V-M & Syrjänen K (1994b) Apoptosis in breast cancer as relat
histopathological characteristics and prognosis. Eur J Cancer 30: 2068-2073.Lipponen PK, Eskelinen MJ, Jauhiainen K, Harju E & Terho R (1992) Tumour infiltratin
lymphocytes as an independent prognostic factor in transitional cell bladder cancer. Eur J C29: 69-75.
Liu X, Kim CN, Yang J, Jemmerson R & Wang X (1996) Induction of apoptotic program in cell- frextracts: requirement for dATP and cytochrome c. Cell 86: 147-157.

75
p53athol
ted
on-
f
thol
ear
esis409.esak. J
cer
3
dnd in
the
duct37-
04:
ofathol
in
ols.
iated-354.g
J 13:
esis:v 10:
lsr Res
Lohmann D, Pütz B, Reich U, Böhm J, Präuer H & Höfler H (1993) Mutational spectrum of thegene in human small-cell lung cancer and relationship to clinicopathological data. Am J P142: 907-915.
Longthorne VL & Williams GT (1997) Caspase activity is required for commitment to Fas- mediaapoptosis. EMBO J 16: 3805-3812.
Lowe SW, Scmhitt EM, Smith SW, Osborne BA & Jacks T (1993) p53 is required for radiatiinduced apoptosis in mouse thymocyctes. Nature 326: 847-849.
Macchiarini P, Fontanini G, Hardin MJ, Squartini F & Angeletti CA (1992) Relation oneovascularisation to metastasis of non-small cell lung cancer. Lancet 340: 145-146.
Manjo G & Joris I (1995) Apoptosis, oncosis and necrosis. an overview of cell death. Am J Pa146: 3-15.
Mathews MB, Bernstein RM, Franza BR & Garrels JI (1984) Identity of the proliferating cell nuclantigen and cyclin. Nature 303: 374-376.
Matsuyama K, Chiba Y, Sasaki M, Tanaka H, Muraoka R & Tanigawa N (1998) Tumor angiogenas a prognostic marker in operable non-small cell lung cancer. Ann Thorac Surg 65: 1405-1
McCarthy NJ, Whyte MK, Gilbert CS & Evan GI (1997) Inhibition of Ced-3/ICE-related proteasdoes not prevent cell death induced by oncogenes, DNA damage, or the bcl-2 homologue bCell Biol 136: 215-227.
McDuffie HH, Klaassen DJ & Dosman JA (1990) Determinants of cell type in patients with canof the lung. Chest 98: 1187-1193.
McLaren R, Kuzu I, Dunnill M, Harris A, Lane D & Gatter KC (1992) The relationship of p5immunostaining to survival in carcinoma of the lung. Br J Cancer 66: 735-738.
Miyashita T, Krajewski S, Krajewska M Wang HG, Lin HK, Liebermann Da, Hoffman B & ReeJC (1994) Tumor suppressor p53 is a regulator of bcl-2 and bax gene expression in vitro avivo. Oncogene 9: 1799-1805.
Miyashita T & Reed JC (1995) Tumor suppressor p53 is a direct transcriptional activator ofhuman bax gene. Cell 80: 293-299.
Momand J, Zambetti GP, Olson DC, George D & Levine AJ (1992) The mdm-2 oncogene proforms a complex with the p53 protein and inhibits p53-mediated transactivation. Cell 69: 121245.
Morabia A & Wynder EL (1992) Relation of bronchioloalveolar carcinoma to tobacco. Br Med J 3541-543.
Morris CS, Esiri MM, Marx A & Newsom-Davis J (1992) Immunocytochemical caracteristicssmall cell lung carcinoma associated with the Lambert-Eaton myasthenic syndrome Am J P140: 839-845.
Moy PM, Holmes EC & Golub SH (1985) Depression of natural killer cytotoxic activitylymphocytes infiltrating human pulmonary tumors. Cancer Res 45: 57-60.
Mulshine JL & Scott F (1995) Molecular markers in early cancer detection: new screening toChest 107: 280S-286S.
Mustonen M, Raunio H, Pääkkö P & Soini Y (1997) The extent of apoptosis is inversely assocwith bcl-2 expression in premalignant and malignant breast lesions. Histopathology 31: 347
Mäkitaro R, Pääkkö P, Huhti E, Bloigu R & Kinnula VL (1999) An epidemiological study of luncancer: history and histological types in general population in northern Finland. Eur Respir436-440.
Naik P, Karrim J & Hanahan D (1996) The rise and fall of apoptosis during multistage tumorigendown-modulation contributes to tumor progression from angiogenic progenitors. Genes De2105-2116.
Naito Y, Saito K, Shiiba K, Ohuchi A, Saigenji K, Nagura H & Ohtani H (1998) CD8+ T celinfiltrated within cancer cell nests as a prognostic factor in human colorectal cancer. Cance58: 3491-3494.

76
Tncer.
thod
RA
T,p53
hway
53e. Am
kants
im:
andd Med
ed
eal and
erothant
se
on
is.
stictral
nghe
1748.ossrs.
Nakano T, Oka K, Takahashi T, Morita S & Arai T (1992) Roles of Langerhans´cells andlymphocytes infiltrating cancer tissues in patients treated by radiation therapy for cervical caCancer 70: 2839-2844.
Nasiell M (1967) Diagnosis of lung cancer by aspiration biopsy and comparison between this meand exfoliative cytology. Acta Cytol 11: 114-119.
Niehans GA, Brunner T, Frizelle SP, Liston JC, Salerno CT, Knapp DJ, Green DR & Kratzke(1997) Human lung carcinomas express fas ligand. Cancer Res 57: 1007-1012.
Nishio M, Koshikawa T, Kuroishi T, Suyama M, Uchida K, Takagi Y, Washimi O, SugiuraAriyoshi Y, Takahashi T, Ueda R & Takahashi T (1996) Prognostic significance of abnormalaccumulation in primary, resected non-small cell lung cancers. J Clin Oncol 14: 497-502.
Nuñez G, Benedict MA, Hu Y & Inohara N (1998) Caspases: the proteases of the apoptotic pat(review). Oncogene 17: 3237-3245.
Nuorva K, Mäkitaro R, Huhti E, Kamel D, Vähäkangas K, Bloigu R, Soini Y & Pääkkö P (1994) pprotein accumulation in lung carcinomas of patients exposed to asbestos and tobacco smokJ Respir Crit Care Med 150: 528-533.
Oberhammer F, Wilson JW, Dive C, Morris ID, Hickman JA, Wakeling AE, Walker PR & SikorsM (1993) Apoptosis death in epithelial cells: cleavage of DNA to 300 and/or 50 kb fragmeprior to or in the absence of internucleosomal fragmentation. EMBO J 12: 3679-3684.
O´Connor L, Strasser A, O´Reilly LA, Hausmann G, Adamas JM, Cory S & Huang DC (1998) Ba novel member of the bcl-2 family that promotes apoptosis. EMBO J 17: 384-395.
Oksa P, Pukkala E, Karjalainen A, Ojajärvi A & Huuskonen MS (1997) Cancer incidencemortality among Finnish asbestos sprayers and in asbestosis and silicosis patients. Am J In31: 693-698.
Oltvai ZN, Milliman CL & Korsmeyer SJ (1993) Bcl-2 heterodimerizes in vivo with a conservhomolog, bax, that accelerates programmed cell death. Cell 74: 609-619.
O´Mahony AM, O ´Sullivan GC, O ´Connell J, Cotter TG & Collins JK (1993) An immunsuppressive factor derived from esophageal squamous carcinoma induces apoptosis in normtransformed cells of lymphoid lineage. J Immunol 15: 4847-4856.
Ormerod MG (1994) Flow cytometric studies of apoptosis. CMB 1: 35-43.Owen-Schaub LB, Zhang W, Cusack JC, Angelo LS, Santee SM, Fujiwara T, Roth JA, Deiss
AB, Zhang WW & Kruzel E (1995) Wild-type human p53 and temperature-sensitive mutinduce Fas/APO-1 expression. Mol Cel Biol 151: 3032-3040.
Packham G (1998) Mutation of bcl-2 family proteins in cancer. Apoptosis 3: 75-82.Perry A, Duenzl ML & Ansari Q (1994) Flow cytometric terminal deoxynucleotidyltransfera
analysis. Arch Pathol Lab Med 118: 1119-1122.Pezzella F, Turley H, Kuzu I, Tungekar MF, Dunnill MS, Pierce CB, Harris A, Gatter KC & Mas
DY (1993) Bcl-2 protein in non-small cell lung carcinoma. N Engl J Med 329: 690-694.Polyak K, Xia Y, Zweier JL, Kinzler KW & Vogelstein B (1997) A model of p53-induced apoptos
Nature 389: 300-305.Popovich J Jr, Kvale PA, Eichenhorn MS, Radke JR, Ohorondnik JM & Fine G (1982) Diagno
accuracy of multiple biopsies from flexible fiberoptic bronchoscopy: a comparison of cenversus peripheral carcinoma. Am Rev Respir Dis 125: 521-523.
Poppema S, Brocker EB, DELeji L, Terbrack D, Visscher T, TerHaar A, Macher E, The TH & SoC (1983)In situ analysis of mononuclear cell infiltrate in primary malignant melanoma Of tskin. Clin Exp Immunol 51: 77-82.
Potten CS (1996) What is an apoptotic index measuring. A commentary. Br J Cancer 74: 1743-Przygodzki RM, Finkelstein SD, Langer JC, Swaltsky PA, Fishback N, Bakker A, Guinee DG, K
M & Travis WD (1996) Analysis of p53, k-ras-2, c-raf-1 in pulmonary neuroendocrine tumoCorrelation with histological subtype and clinical outcome. Am J Pathol 148: 1531-1541.

77
ung
n Alung
ntse-
f B
53
tictype.
6.and
tialeural
, amster
97)rgy
nsnd
&cts in
cell
85)s of
angtwo-
the
type
rome: 21-
Pääkkö P, Risteli J, Risteli L & Autio-Harmainen H (1990) Immunohistochemical evidence that lcarcinomas grow on alveolar basement membranes. Am J Surg Pahtol 14: 464-473.
Pääkkö P, Rämet M, Vähäkangas K, Korpela N, Soini Y, Turunen S, Jaworska M & Gillisse(1996) Crocidolite asbestos causes an induction of p53 and apoptosis in cultured A-549carcinoma cells. Apoptosis 3: 203-212.
Pääkkö P, Sormunen R, Risteli L, Risteli J, Ala-Kokko L & Ryhänen L (1989) Malotilate preveaccumulation of type IIIpN-collagen, type IV collagen and laminin in carbon tetrachloridinduced pulmonary fibrosis in rats. Am Rev Respir Dis 139: 1105-1111.
Quin Z & Blankenstein T (1995) Tumor growth inhibition mediated by lymphotoxin: evidence olymphocyte involvement in the antitumor responce. Cancer Res 55: 4747-4751.
Quinland DC, Davidson AG, Summers CL, Warden HE & Doshi HM (1992) Accumulation of pprotein correlates with a poor prognosis in human lung cancer. Cancer Res 52: 4828-4831.
Rampino N, Yamamoto H, Ionov Y, Li Y, Sawai H, Reed JC & Perucho M (1997) Somaframeshift mutations in the bax gene in colon cancers of the microsatellite mutator phenoScinece 275: 967-969.
Reed JC (1994) Bcl-2 and the regulation of programmed cell death (review). J Cell Biol 124: 1-Reed JC (1995) Regulation of apoptosis by bcl-2 family proteins and its role in cancer
chemoresistance (review). Curr Opin Oncol 7:541-546.Reed JC (1998) Bcl-2 family proteins (review). Oncogene 17: 3225-3236.Reed JC, Meister L, Tanaka S, Cuddy M, Yum S, Geyer C & Pleasure D (1991) Differen
expression of bcl-2 protooncogene in neuroblastoma and other human tumour cell lines of norigin. Cancer Res 15: 6529-6538.
Reynolds JE, Yang T, Qian L, Jenkinson JD, Zhou P, Eastman A & Craig RW (1994) Mcl-1member of bcl-2 family, delays apoptosis induced by c-myc overexpresssion in Chinese haovary cells. Cancer Res 54: 6348-6352.
Riemann D, Wenzel K, Schultz T, Hofmann S, Neef H, Lautenschläger C & Langner J (19Phenotypic analysis of T lymphocytes isolated from non-small cell lung cancer. Int Arch AlleImmunol 114: 38-45.
Ruiter DJ, Bhan AK, Harrist TJ, Sober AJ & Mihm MC Jr (1982) Major histocompatibility antigeand mononuclear inflammatory infiltrate in beningn nevomelanocytic proliferations amalignant melanoma. J Immunol 129: 2808-2815.
Rämet M, Castrén K, Järvinen K, Pekkala K, Turpeenniemi-Hujanen T, Soini Y, Pääkkö PVähäkangas K (1995) p53 protein expression is correlated with benzo(a)pyrene-DNA adducarcinoma cell lines. Carcinogenesis 16: 2117-2124.
Shaham S & Horviz HR (1996) An alternatively spliced C. Elegans ced-4 RNA encodes a noveldeath inhibitor. Cell 86: 201-208.
Said JW, Vimadalal S, Nash G, Shintaku IP, Heusser RC, Sassoon AF & Lloyd RV (19Immunoreactive neuron-specific enolase, bombesin and chromogranin as markerneuroendocrine lung tumors. Hum Pathol 16: 236-240.
Sato T, Hanada M, Bodrug S, Irie S, Iwama N, Boise LH, Thompson CB, Colemis E, Fong L & WHG (1994) Interactions among members of the bcl-2 protein family analyzed with a yeasthybrid system. Proc Natl Acad Sci USA 91: 9238-9242.
Segal NH, Cohen RJ, Haffejee Z & Savage N (1994) Bcl-2 proto-oncogene expression inprostatic neuroendocrine cell. Arch Pathol Lab Med 118: 616-618.
Shaw P, Bovey R, Tardy S, Sahali R, Sordat B & Costa J (1992) Induction of apoptosis by wild-p53 in human colon tumor-derived cell line. Proc Natl Acad Sci USA 89: 4495-4499.
Shepherd FA, Laskey J, Evans WK, Goss PE, Johansen E & Khamsi F (1992) Cushing´s syndassociated with ectopic corticotropin production and small-cell lung cancer. J Clin Oncol 1027.

78
espir
&n in
f-1
m J
bax
ghtened
th,
nce
Am
esis.
ersing
er Gation
5-37.)able
e 267:
316.d
&cell
isnd
nsRev
e097-
Shure D & Fedullo PF (1982) Transbronchial needle aspiration of peripheral masses. Am Rev RDis 128: 1090-1092.
Silvestrini R, Veneroni S, Daidone MG, Benini E, Boracchi P, Mezzetti M, DiFronzo G, Rilke FVeronesi U (1994) The bcl-2 protein: a prognostic indicator strongly related to p53 proteilymph node-negative breast cancer patients. J Natl Cancer Inst 86: 499-504.
Soengas MS, Alarcón RM, Yoshida H, Giaccia AJ, Hakem R, Mak TW & Lowe SW (1999) ApaAnd caspase-9 in p53-dependent apoptosis and tumor inhibition. Science 284: 156-159.
Soini Y, Pääkkö P & Lehto V-P (1998a) Histopathological evaluation of apoptosis (review). APathol 153: 1041-1053.
Soini Y, Törmänen U & Pääkkö P (1998b) Apoptosis is inversely related to bcl-2 but not toexpresion in salivary gland tumours. Histopathology 32: 28-34.
Soini Y, Virkajärvi N, Lehto VP & Pääkkö P (1996) Hepatocellular carcinomas with a hiproliferation index and low degree of apoptosis and necrosis are associated with a shorsurvival. Br J Cancer 73: 1025-1030.
Song Q, Kuang Y, Dixit VM, Vincenz C (1999) Boo, a novel negative regulator of cell deainteracts with Apaf-1. EMBO J 18: 167-178.
Stammler G & Volm M (1996) Apoptosis in non-small cell lung cancer as related to drug resistaand prognosis. Apoptosis 1: 95-99.
Staunton MJ & Gaffney EF (1995) Tumour type is a determinant of susceptibility to apoptosis.J Clin Pathol 103: 300-307.
Steenland K, Loomis D, Shy C & Simonsen N (1996) Review of occupational lung carcinogenAm J Ind Med 29: 474-490.
Strand S, Hoffman WJ, Hug H, Müller M, Otto G, Strand D, Mariani SM, Stremmel W, KrammPH & Galle PR (1996) Lymphocyte apoptosis induced by CD95(APO-1/Fas) ligand-exprestumor cells - a mechanism of immune evasion ? Nature Med 2: 1361-1366.
Susin SA, Zamzami N, Castedo M, Daugas E, Wang HG, Geley S, Fassy F, Reed JC & Kroem(1997) The central executioner of apoptosis: multiple connections between protease activand mitochondria in Fas/APO-1/CD95 - and ceramide-induced apoptosis. J Exp Med 186:2
Tanaka T, Yamamoto M, Tamura T, Moritani Y, Miyai M, Hiraki S, Ohnoshi T & Kimura I (1985Cytologic and histologic correlation in primary lung cancer. A study of 154 cases with resecttumors. Acta Cytol 29: 49-56
Thompson CB (1995) Apoptosis in the pathogenesis and treatment of disease (review). Scienc1456-1462.
Thornberry NA & Lazebnik Y (1998) Caspases: enemies within (review). Science 281: 1312-1Travis WD, Colby TV, Corrin B, Shimosato Y & Brambilla E in collaboration with Sobin LH an
pathologists from 14 countries (1999) Histological typing of lung an pleural tumours. 3rd edn.Springer-Verlag Berlin Heidelberg. Germany.
Travis WD, Linnoila I, Tsokos MG, Hitchcock CL, Cutler GB, Nieman L, Chrousos G, Pass HDoppman J (1991) Neuroendocrine tumors of the lung with proposed criteria for largeneuroendocrine carcinoma. Am J Surg Pathol 15: 529-553.
Travis WD, Rush W, Flieder DB, Falk R, Fleming MV, Gal AA & Koss MN (1998) Survival analysof 200 pulmonary neuroendocrine tumors with clarification of criteria for atypical carcinoid aits separation from typical carcinoid. Am J Surg Pathol 22: 934-944.
Trinchieri G (1995) Interleukin-12: a proinflammatory cytokine with immunoregulatory functiothat bridge innatet resistance and antigen-specific adaptive immunity (review) AnnualImmunol 13: 251-276.
Tsujimoto Y, Finger LR, Yunis J, Nowell PC & Croce CM (1984) Cloning of the chromosombreakpoint of neoplastic B cells with the t(14;18) chromosome translocation. Science 226; 11099.

79
nng
&lung
ical
73-
ast
exinosure
s: a ¨
on,tatic
ion,-772.
onus
n aDev
ng
ith89.
993)
ion
a J,jamgy in
ase
atin
Turrisi AT, Kim K, Blum R, Sause WT, Livingston RB, Komaki R, Wagner H, Aisner S & JohnsoDH (1999) Twice daily compared with once daily thoracic radiotherapy in limited small-cell lucancer treated concurrently with cisplatin and etoposide. New Engl J Med 340 265-271.
Törmänen U, Eerola A-K, Rainio P, Vähäkangas K, Soini Y, Sormunen R, Bloigu R, Lehto V-PPääkkö P (1995) Enhanced apoptosis predicts shortened survival in non-small cellcarcinoma. Cancer Res 55: 5595-5602.
Underwood JCE (1974) Lymphoreticular infiltration in human tumours: prognostic and biologimplications (review). Br J Cancer 30: 537-548.
Vacca LL (1985) Laboratory Manual of Histochemistry. New York, NY, USA. Raven Press, p. 1175.
Vakkala M, Lähteenmäki K, Raunio H, Pääkkö P & Soini Y (1999) Apoptosis during brecarcinoma progression. Clin Cancer Res 5: 319-324.
Van Engeland M, Nieland LJ, Ramaekers FCS, Schutte B & Reutelingsperger CPM (1998) AnnV-affinity assay: a review on an apoptotis detection system based on phospatidylserine exp(review). Cytometry 31: 1-9.
Van de Schepop AM, de Jong JS, van Diest PJ & Baak JPA (1996) Counting of apoptotic cellMethodological study in invasive breast cancer. J Clin Pathol: Mol Pathol 49: 214-217.
Vesalainen S, Lipponen P, Talja M & Syrjänen K (1994) Histological grade, perineural infiltratitumor-infiltrating lymphocytes and apoptosis as determinants of long-term prognosis in prosadenocarcinoma. Eur J Cancer 30: 1797-1803.
Virkajärvi N, Pääkkö P & Soini Y (1997) Association between p53 overexpression, cell proliferattumour necrosis and extent of apoptosis in operated pancreatic carcinoma. APMIS 105: 765
Vogelstein B & Kinzler KW (1992) p53 function and dysfuction (review). Cell 70: 523-526.Vose BM, Vánky F & Klein E (1977) Human tumour-lymphocyte interaction in vitro. Comparis
of the reactivity of tumour-infiltrating, blood and lymph-node lymphocytes with autologotumour cells. Int J Cancer 20: 895-902.
Wagner AJ, Kokontis JM & Hay N (1994) Myc-mediated apoptosis requires wild-type p53 imanner independent of cell cycle arrest and ability of p53 to induce p21WAF/CIP1. Genes8: 2817-2830.
Wang DG, Johnston CF, Sloan JM & Buchanan KD (1998) Expression of bcl-2 in luneuroendocrine tumours: comparison with p53. J Pathol 184: 247-251.
Watanabe S, Sato Y, Kodama T & Shimosato Y (1983) Immunohistochemical study wmonoclonal antibodies on immune response in human lung cancer. Cancer Res 3: 5883-58
White E (1996) Life, death, and the pursuit of apoptosis (review). Genes Dev 10: 1-15.Wijsman JH, Jonker RR, Keijzer R, van de Velde CJ, Cornelisse CJ & van Dierendonch JH (1
A new method to detect apoptosis in paraffin sections:in situend-labeling of fragmentated DNA.J Histochem Cytochem 41: 7-12.
World Health Organization. (1981) Histological typing of lung tumours. International classificatof Tumours. No. 1.2nd edn. Geneva: World Health Organization.
Wongsurakiat P, Wongbunnate S, Dejsomritrutai W, Charoenratanakul S, TscheikunYoungchaiyud P, Pushpakom R, Maranetra N, Nana A, Chierarkul N, Sakiyalak U & RuengC (1998) Diagnostic value of bronchoalevolar lavage and postbronchoscopic sputum cytoloperipheral lung cancer. Respirology 3: 131-137.
Wyllie AH (1980) Glucocorticoid-induced thymocyte apoptosis is associated with endonucleactivation. Nature 284: 555-556.
Wyllie AH (1984) Chromatin cleavage in apoptosis: association with condensed chrommorphology and dependence on macromolecular synthesis. J Pathol 142: 67-77.
Wyllie AH (1997a) Apoptosis and carcinogenesis (review). Eur J Cell Biol 73: 189-197.Wyllie AH (1997b) Apoptosis: an overview (review). Br Med Bull 53: 451-465.

80
eta-
age
noue250.eath
)ence
and
n
5345-
seated
s T
C.-413.
Xiang J, Chao DT & Korsmeyer SJ (1996) Bax-induced cell death may not require interleukin 1 bconverting enzyme-like proteases. Proc Natl Acad Sci USA 93: 14559-1463.
Yamada T, Ohyama H, Kinjo Y & Watanabe M (1981) Evidence for the internucleosomal breakof chromatin in rat thymocytes irradiated in vitro. Radiat Res 85: 544-553.
Yamazaki K, Shosaku A, Takekawa H, Sukoh N, Watanabe N, Ogura S, Nakajima I, Isobe H, IK & Kawakami Y (1994) Tumor angiogenesis in human adenocarcinoma. Cancer 74: 2245-2
Yang E & Korsmeyer SJ (1996) Molecular thanatopsis: a discourse on the bcl-2 family and cell d(review). Blood 88: 386-401.
Yang J, Liu X, Bhalla K, Kim CN, Ibrado AM, Cai J, Peng TI, Jones DP & Wang X (1997Prevention of apoptosis by bcl-2: release of cytochrome c from mitochondria blocked. Sci275: 1129-1132.
Yin C, Knudson CM, Korsmwyer SJ & Van Dyke T (1997) Bax suppresses tumorigenesisstimulates apoptosis in vivo. Nature 385: 637-640.
Yin XM, Oltvai ZN & Korsmeyer S (1994) BH1 and BH2 domains of bcl-2 are required for Inhibitioof apoptosis and heterodimerization with bax. Nature 369: 321-322.
Yonish-Rouach E, Resnitzky D, Lotem J, Sachs L, Kimchi A & Oren M (1991) Wild-type pinduces apoptosis of myeloid leukaemic cells that is inhibited by interleukin-6. Nature 352: 3347.
Yoshino T, Kondo E, Cao L, Takahashi K, Hayashi K, Nomura S & Akagi T (1994) Inverexpression of bcl-2 and Fas antigen in lymphoblas in peripheral lymph nodes and activperipheral blood T and B lymphocytes. Blood 7: 1856-1861.
Ziegler A, Johanson AS, Leffell DJ, Simon JA, Sharma HW, Kimmelman J, Remington L, Jack& Brash DE (1994) Sunburn and p53 in the onset of skin cancer. Nature 372: 773-776.
Zou H, Henzel WJ, Liu X, Lutschg A & Wang X (1997) Apaf-1, a human protein homologous toElegans CED-4 participates in cytochrome c-dependent activation of caspase-3. Cell 90: 405

Original papers
