arbovirus infection dynamics in the mosquito …
TRANSCRIPT

The Pennsylvania State University
The Graduate School
ARBOVIRUS INFECTION DYNAMICS IN THE MOSQUITO AEDES AEGYPTI
A Dissertation in
Entomology
by
Mario De Jesus Novelo Canto
© 2021 Mario De Jesus Novelo Canto
Submitted in Partial Fulfillment
of the Requirements
for the Degree of
Doctor of Philosophy
August 2021

ii
The dissertation of Mario De Jesus Novelo Canto was reviewed and approved by the
following:
Elizabeth A. McGraw
Professor and Huck Scholar in Entomology
Head of the Department of Biology
Dissertation Adviser
Chair of Committee
Jason L. Rasgon
Professor of Entomology and Disease Epidemiology
Margarita López-Uribe
Lorenzo L. Langstroth Early Career Professor
Assistant Professor of Entomology
David Kennedy
Assistant Professor of Biology
Gary W. Felton
Professor of Entomology
Head of the Department of Entomology

iii
ABSTRACT
Arboviral diseases account for more than 17% of all infectious diseases globally,
among the most prevalent are dengue fever and chikungunya, with over half of world’s
population at risk of infection. Although mortality is generally low, the morbidity caused
by these pathogens is associated with a substantial socioeconomic burden. Occasionally,
infection with dengue virus (DENV) can lead to death due to dengue shock syndrome in
all ages and chikungunya virus (CHIKV) infection can be severe for infants, with nearly
half of infected newborns experiencing encephalopathy and multiple organ dysfunction.
Both DENV and CHIKV are RNA viruses transmitted through the bite of the mosquito
Aedes aegypti. Arboviral infection in the mosquito is a complex process, as the dynamics
of infection can be influenced by the genetic variation of the virus altering the infection
rates and transmission potential in mosquitoes. Competition between strains or serotypes
can also affect viral population dynamics within the vector, thus modulating the
transmission potential. This happens in nature when mosquitoes take multiple blood meals
from several different hosts, each infected with a different or multiple virus. How different
viruses interact and compete inside the mosquito during the stepwise process of infection
or how the vector immune response changes in response to these different viruses is an
area of vital research.
Due to the lack of an effective vaccine and antiviral drugs, vector control is the
most efficient strategy in reducing arbovirus transmission. Therefore, understanding the
basic processes shaping vector-virus interactions is necessary for both implementation of
more effective control strategies and the development of genetic modifications (GMO) and

iv
biocontrol in mosquitoes to limit transmission. Current example of such technologies
includes clustered regularly interspaced short palindromic repeats (CRISPR) and the use
of the endosymbiont Wolbachia. The three major aims of my dissertation are to investigate
the 1) infection dynamics of DENV in the mosquito Ae. aegypti; 2) competitive interactions
between co-infecting DENV serotypes in both wild type and Wolbachia – infected
mosquitoes and 3) genetic basis of Ae. aegypti immune response to DENV and CHIKV
infection.
In chapter 1 1 I reviewed the history and role of Ae. aegypti as a disease vector and
the importance of my research by studying the process of infection with different
arboviruses inside the vector. In chapter 2 I examined the infection dynamics of DENV
in the mosquito Ae. aegypti, specifically, the effect of infectious dose and serotype on the
growth kinetics of virus in the tissues define the stepwise process of infection in the
mosquito body. In chapter 3 I studied the competitive interactions between co-infecting
DENV serotypes both in wild type mosquitoes and in Wolbachia-infected Ae. aegypti.
Chapter 4 examines the comparative genetic basis of Ae. aegypti response to DENV and
CHIKV infection using a family-breeding design. Finally, chapter 5 synthesizes the
findings of the dissertation, including future directions, as well as discussing unanswered
questions in this research area.

v
TABLE OF CONTENTS
LIST OF FIGURES ................................................................................................................. viii
LIST OF TABLES ................................................................................................................... ix
ACKNOWLEDGEMENTS ..................................................................................................... x
Chapter 1 Introduction ............................................................................................................ 1
1.1 Aedes aegypti ............................................................................................................ 1 1.2 Dengue and chikungunya .......................................................................................... 4 1.3 Vector – virus dynamics ............................................................................................ 5 1.4 Vector control ............................................................................................................ 8 1.5 References ................................................................................................................. 12
Chapter 2 Intra-host growth kinetics of dengue virus in the mosquito Aedes aegypti ............. 23
2.1 Abstract ..................................................................................................................... 23 2.2 Author summary ........................................................................................................ 24 2.3 Introduction ............................................................................................................... 25 2.4 Methods ...................................................................................................................... 28
2.4.1 Ae. aegypti rearing and virus preparation ........................................................ 28
2.4.2 Mosquito infections ......................................................................................... 29
2.4.3 Mosquito dissection and RNA extraction ....................................................... 29
2.4.4 DENV absolute quantification via RT-qPCR ................................................. 30
2.4.5 Survival assay .................................................................................................. 31
2.4.6 Data analysis ................................................................................................... 31
2.5 Results ....................................................................................................................... 32
2.5.1 The proportion of DENV – infected mosquitoes is affected by both
infectious dose and DPI .................................................................................... 33
2.5.2 EIP is affected by the serotype strain and initial infectious dose .................... 34
2.5.3 Intra-host DENV kinetics are influenced by initial infectious dose and
strain ................................................................................................................. 37
2.5.4 Some treatments do not show substantial DENV replication and others
exhibit subpopulation structure of DENV load in infected mosquitoes ........... 37
2.5.5 DENV load is always higher in the midgut than other tissues for high
infectious dose .................................................................................................. 42
2.5.6 Tissue-specific DENV kinetics vary by strain in the high infectious dose .... 43
2.5.7 The DENV-2 strain has a higher maximum DENV load than the DENV-1
strain in the low infectious dose ....................................................................... 44
2.6 Discussion ................................................................................................................. 46
2.7 Acknowledgments ...................................................................................................... 50
2.8 References ................................................................................................................. 51
Chapter 3 The effects of DENV serotype competition and co-infection on viral kinetics in
Wolbachia-infected and uninfected Aedes aegypti mosquitoes ............................................... 57

vi
3.1 Abstract ..................................................................................................................... 57
3.2 Introduction ............................................................................................................... 59
3.3 Methods ..................................................................................................................... 62
3.3.1 Mosquito lines and rearing ............................................................................. 62
3.3.2 Virus culture and titration .............................................................................. 63
3.3.3 Mosquito infections ......................................................................................... 64
3.3.4 DENV absolute quantification via RT-qPCR ................................................ 65
3.3.5 Statistical analysis .......................................................................................... 66
3.4 Results ....................................................................................................................... 67
3.4.1 DENV serotype competition in co-infection ................................................... 67
3.4.2 Co-infection alters infection dynamics of DENV serotypes in wild type
mosquitoes ........................................................................................................ 72
3.4.3 Tissue specific differences in viral dynamics .................................................. 72
3.5 Discussion ................................................................................................................. 74
3.6 Conclusion ................................................................................................................. 78
3.7 Acknowledgements .................................................................................................... 79
3.8 References ................................................................................................................. 80
Chapter 4 Family level variation in the mosquito Aedes aegypti susceptibility and
immune response to dengue and chikungunya viral infection ........................................ 85
4.1 Abstract ..................................................................................................................... 85
4.2 Introduction ............................................................................................................... 86
4.3 Methods ...................................................................................................................... 91
4.3.1 Mosquito line and stock practices ................................................................... 91
4.3.2 Breeding design .............................................................................................. 92
4.3.3 Virus culture ................................................................................................... 92
4.3.4 Mosquito infections ......................................................................................... 93
4.3.5 Viral quantification ......................................................................................... 93
4.3.6 Selection of immune genes for expression analysis ....................................... 94
4.3.7 Immune gene expression ................................................................................. 95
4.3.8 Analysis of genetic variance ........................................................................... 96
4.4 Results ....................................................................................................................... 97
4.4.1 Family level variation in the heritability of DENV and CHIKV viral loads ... 97
4.4.2 Genetic correlation between DENV and CHIKV across mosquito families ... 99
4.4.3 Phenotypic extremes of viral loads for both DENV and CHIKV infection
in the mosquito families ................................................................................... 99
4.4.4 Immune genes relative expression .................................................................. 101
4.5 Discussion ................................................................................................................. 105
4.6 References ................................................................................................................. 112
Chapter 5 Synthesis and General Discussion .......................................................................... 120
5.1 References .................................................................................................................. 125
Appendices .............................................................................................................................. 127
Appendix A Supplementary materials from Chapter 2 .................................................... 127

vii
Appendix B Supplementary materials from Chapter 3 .................................................... 135
Appendix C Supplementary materials from Chapter 4 .................................................... 136

viii
LIST OF FIGURES
Figure 2.1: Susceptibility of DENV strains by DPI, tissue and infectious dose ...................... 35
Figure 2.2: DENV kinetics by strain, tissue and infectious dose ............................................. 39
Figure 2.3: “Trickledown” or declining viral load with tissue progression ............................. 43
Figure 3.1: Mosquito susceptibility to DENV in co-infections by DPI, tissue, and line ......... 69
Figure 3.2: DENV load in co-infection by DPI, tissue, and mosquito line .............................. 70
Figure 3.3: Co-infection serotype ratio by DPI and tissue ....................................................... 71
Figure 3.4: WT line susceptibility to DENV in co- and mono-infections by DPI, tissue,
and serotype ..................................................................................................................... 73
Figure 3.5: WT line DENV viral RNA load in co- and mono-infection by DPI, tissue, and
serotype ............................................................................................................................ 74
Figure 4.1:Ranked families by viral load ................................................................................ 97
Figure 4.2: Scatter plot showing the relationship between mean DENV and CHIKV loads
from individual families ........................................................................................................... 98
Figure 4.3:Whole mosquito viral loads per family representing the extremes of the
distribution ....................................................................................................................... 99
Figure 4.4: AGO2 and DCR2 expression relative to the housekeeping gene RPS17 in
family groups classified as High and Low Viral loads .................................................... 102
Figure 4.5: TEP3 and CECH expression relative to the housekeeping gene RPS17 in
family groups classified as High and Low Viral loads .................................................... 103

ix
LIST OF TABLES
Table 2.1: The DENV strains used in this study ...................................................................... 29
Table 2.2: Susceptibility by Strain, Tissue, Dose and DPI ...................................................... 36
Table 2.3: DRC parameters for successful infections by tissue, infectious dose and strain .... 40
Table 2.4: DRC parameter contrasts for successful infections between strains, infectious
dose and subpopulations ................................................................................................. 41
Table 2.5: Linear regression model for SG ............................................................................ 45
Table 2.6: Serotype contrasts for growth rate in SG ................................................................ 45
Table 3.1: Dengue serotypes and strains use ........................................................................... 64
Table 3.2: Viral titers ............................................................................................................... 64
Table 4.1: Primers and probes for qPCR gene expression and viral quantification ................ 95

x
ACKNOWLEDGEMENTS
I would like to express my sincere gratitude to my Adviser, Professor Elizabeth A.
McGraw, for giving me the opportunity to be part of her lab, for her guidance, advice and
most of importantly, for her unwavering support in making me a better scientist. And also,
if I’m honest, for having the patience to deal with countless manuscript drafts filled with
grammatical errors that I send her way. I would also like to thank the members of my
committee, Professor Jason L. Rasgon, Assistant Professors Margarita López -Uribe and
David Kennedy for their insights and advice.
I want to acknowledge the past and present McGraw lab members and friends from
Melbourne and State College, I will be forever grateful for their company, generosity, for
the laughs, drinks and the good times. Cass, Gerard, Michelle, Suzie, Jim, Helena, Tent,
Heather, Toby and Louise, miss you guys. A very special thanks to Emily Kerton from our
time in Monash, for being a superstar, and helping me carry out two of my three research
projects. I want to thank my PSU lab mates for making me feel welcome in a new place:
Fhallon, Leah, Heverton, Austin and Matt.
To my family and friends from back home, for their love and support. To my
parents, Mario and Carolina, and my sisters Marinha, Cari and Paz for their unconditional
love and strength. Como decia Jorge Negrete: “México lindo y querido, si muero lejos de
ti que digan que estoy dormido y que me traigan aquí”.

xi
Finally, to my loving partner Staci Cibotti, for being the most awesome human
being. To be honest I don’t think you contribute at all to me finishing my PhD, but you
sure make my life happier. To Jack the cat.
This work was conducted under the Institutional Biosafety Committee Protocol
47902, with funding from the Australian Research Council (DP160100588), the Australian
Research Council Laureate Fellowship (FL170100022), the National Health and Medical
Research Council Grant (APP1103804), the National Institute of Allergy and Infectious
Diseases (1R01 AI143758), start-up funds from the PSU College of Agriculture and the
Department of Entomology.
The findings and conclusions in this dissertation do not reflect the view of the
funding agencies.

xii
“Try not. Do or do not, there is no try” – Master Yoda.

1
Chapter 1
Introduction
1.1 Aedes aegypti
The yellow fever mosquito, Ae. aegypti, is one of the most feared and important
dipterans around the world (McGregor & Connelly, 2020). The reason behind its notoriety
is because no other mosquito species has had such a dramatic impact on human history
(Jeffrey R. Powell & Tabachnick, 2013). Often called, “the most dangerous animal in the
planet”, Ae. aegypti alone is responsible for the transmission of at least half a dozen
medically important pathogens to humans (Jeffrey R. Powell, 2016), with the earliest
recorded mosquito-borne virus outbreak of yellow fever occurring in the XV century
(Staples & Monath, 2008). Several of the pathogens transmitted by Ae. aegypti are
responsible for substantial global socioeconomic losses. Chiefly among them are: yellow
fever (YFV), Zika (ZIKV), chikungunya (CHIKV) and dengue (DENV) viruses (Leta et
al., 2018). Globally, DENV is the most important vector-borne viral disease, with estimates
of 3.6 billion people living in areas of high transmission risk, and up to $20 billion in
economic losses linked to DENV infection . Additionally, newly emerging pathogens like
CHIKV have caused several major epidemics in the past 20 years, and although mortality
remains low, CHIKV infection can be severe in the young and cause ongoing arthritis like
complications. Congenital infection rates are upwards of 50% and nearly half of infected

2
newborns will experience central nervous system disease (encephalopathy) (Morrison,
2014).
The Ae. aegypti life cycle occurs in four defined stages: 1) female mosquitoes lay
eggs on wet walls of water containers above the waterline, preferably in artificial containers
around human habitation, 2) larvae hatch once the water rises enough to cover the eggs, 3)
pupae develop until the adult mosquito is ready to emerge and, 4) after emergence, male
mosquitoes feed on nectar and females feed almost exclusively on human blood to produce
eggs (Christophers, 1960). Because of this, the natural history of Ae. aegypti is deeply
intertwined with human history (McNeill, 2010). Native and once confined to the African
continent, where ancestral populations exhibit sylvatic cycles, Ae. aegypti territorial
expansion around the globe coincides with the transatlantic slave trade by European
colonialists (Lounibos, 1981; Tabachnick, 1991). “Domestication” of the mosquito is
thought to have occurred in two ways; 1) with the transition from nomadic to sedentary
lifestyles, humans began living in villages in close proximity and storing water year around,
creating an ideal niche for the mosquito to exploit, and 2) during slave or trade voyages,
where ships had optimal breeding conditions such as crowding and long-term water
storages (Jeffrey R. Powell & Tabachnick, 2013; Wilke et al., 2020). Regardless of the true
domestication origin of Ae. aegypti, its current preference for both human habitat and
blood, made this species and efficient disease vector (J. R. Powell, 2018; Scott A Ritchie,
2014).
Since the first confirmed outbreak of YFV in Yucatán, México, in 1614 (Staples &
Monath, 2008) and the first time Ae. aegypti was identified as an arboviral vector in Cuba

3
in 1906 (Bancroft, 1906; Reed & Carroll, 1901), there has been a search for mosquito
abatement methods, eventually leading to the current concept of vector control. From an
epidemiological point of view, vector control methods can only be effective if they are
contrasted with a mosquito efficacy in transmitting a pathogen (Roiz et al., 2018; David L.
Smith et al., 2012). This efficacy has been described as the vectorial capacity (VC), a
mathematical link between the mosquito’s natural history and pathogen transmission
(Macdonald, 1957). VC defines, within a vector population, the pathogen transmission
potential by using mosquito metrics such as: daily survival rate, biting rate and the extrinsic
incubation period (EIP) (Macdonald, 1957; David L. Smith et al., 2012; D. L. Smith et al.,
2014; Souza-Neto, Powell, & Bonizzoni, 2019). The EIP has been defined as the time it
takes from pathogen acquisition to transmission by a vector, to a susceptible vertebrate
host, and it represents one of the most influential parameters in modeling mosquito-borne
pathogen transmission (Ohm et al., 2018; Claudia Rückert & Ebel, 2018; Tjaden, Thomas,
Fischer, & Beierkuhnlein, 2013). Several factors can influence the length of the EIP, such
as temperature, vector microbiome, initial pathogen dose, and both the mosquito and viral
genotype (Armstrong & Rico-Hesse, 2003; Carrington & Simmons, 2014; Fontaine et al.,
2018; A. Gloria-Soria, Armstrong, Powell, & Turner, 2017; Gould & Higgs, 2009;
Kauffman & Kramer, 2017; L. Lambrechts et al., 2009; Le Flohic, Porphyre, Barbazan, &
Gonzalez, 2013). The interactions between any of these factors in shaping the transmission
dynamics of a pathogen represents vital research area. Specifically, the vector - pathogen
interactions are the main focus of this dissertation, using Ae. aegypti and DENV and
CHIKV viruses.

4
1.2 Dengue and chikungunya
Dengue fever is one of the most prevalent arboviral diseases globally (Murray,
Quam, & Wilder-Smith, 2013), with estimates of more than 390 million infections per year
(Bhatt et al., 2013) and over half of the world’s population at risk (Brady et al., 2012).
Although mortality is generally low, the morbidity caused by dengue fever is associated
with a substantial socioeconomic burden (Duane J. Gubler, 2011). Dengue fever is caused
by four different serotypes of virus (DENV 1–4), belonging to the Flaviviridae family,
(Deen et al., 2006) that are 65–70% similar at the DNA sequence level across their ~11-kb
genomes (Perera & Kuhn, 2008), while also exhibiting a high-average within-serotype
diversity of ~3% at the amino acid level (Azhar et al., 2015).
Compared to DENV, CHIKV was relatively unknown until it started disseminating
across the globe in 2004, causing large outbreaks in Latin America, Asia and Africa
(Petersen & Powers, 2016). Chikungunya fever is caused by a single CHIKV serotype that
belongs to the Alphavirus genus and it has an ~11 kb single stranded positive-sense RNA
genome (Tanabe et al., 2018). The more substantial impact of the disease, is in long term
morbidity (Silva & Dermody, 2017). While adults tend to experience a syndrome involving
rash, fever and arthritis, many go on to have long-term complications and up to 50% of
infected newborns will develop encephalopathy. Additionally, recent studies have
determined that 48% of infected individuals develop chronic arthritis within 20 months of
infection (Yactayo, Staples, Millot, Cibrelus, & Ramon-Pardo, 2016).

5
1.3 Vector-virus dynamics
Arboviral infection in the mosquito is a complex process (Lequime, Fontaine, Ar
Gouilh, Moltini-Conclois, & Lambrechts, 2016). Viruses must infect several tissues,
including the midgut and salivary glands, in a stepwise fashion (Franz, Kantor, Passarelli,
& Clem, 2015). Following ingestion, the virus enters the midgut epithelial cells, where it
replicates. Subsequently, it disseminates and infects secondary tissues, ultimately reaching
the salivary glands (Khoo, Doty, Held, Olson, & Franz, 2013). The midgut is thought to
represent a key barrier in the infection process, preventing many mosquitoes from reaching
a disseminated infection stage (Cox, Brown, & Rico-Hesse, 2011). Once the mosquito is
infected with a virus, the infection can persist in multiple tissues and the vector has the
potential of constantly spreading the pathogen throughout its life (Miller, Mitchell, &
Ballinger, 1989). A key determinant in the kinetics of infection is the antiviral immunity
of the mosquito, that ultimately affects the EIP and the mosquito VC. Several innate
immune pathways have been reported to control or modulate viral infection inside the
mosquito in both systemic and tissue specific ways. Some of the main pathways are, RNAi,
Toll, Janus Kinase - signal transduction and activators of transcription (JAK-STAT), the
Immune deficiency factor (IMD) and antimicrobial peptides (AMP’s) (Bartholomay et al.,
2010; Blair & Olson, 2015; Cheng, Liu, Wang, & Xiao, 2016; R. Zhang et al., 2017).
The kinetics of infection are equally influenced by genetic variation in the virus.
For DENV, comparisons between strains within single serotypes, have revealed variation
in both infection rates and EIP in mosquitoes (Anderson & Rico-Hesse, 2006; Armstrong
& Rico-Hesse, 2003), potentially due to differences in viral replication rates (Cox et al.,

6
2011). In humans, genetic variation within and between serotypes also determines relative
viral fitness (replication rate) (Louis Lambrechts et al., 2012; Ty Hang et al., 2010), with
particular strains linked to more severe disease outcome. The intra-host/vector diversity of
DENV can also play a role in transmission, such as variants with a replicative advantage
can spread more rapidly overall, eventually displacing those lower replication rates.
(Guzman & Harris, 2015). For example, an uncharacteristically large outbreak of dengue
in Cairns, Australia in 2008/2009 was attributed to the very short EIP of the DENV-3 strain
in the mosquito (S. A. Ritchie et al., 2013). DENV-2 strains from the American and South
East Asian genotypes differ in their EIP lengths, with the South East Asian genotypes
having shorter EIPs. This shorter EIP was thought, in part, to explain the displacement of
the American DENV strains in South America by the Asian lineage (Anderson & Rico-
Hesse, 2006). Surprisingly, little is known about DENV kinetics in the mosquito and how
the virus interacts with individual tissues during infection. Given that the virus moves in a
stepwise fashion, selective pressures in initial tissues might have cumulative effects on the
viral kinetics in downstream tissues (Hall, Bento, & Ebert, 2017).
Competition between strains or serotypes can also affect viral population dynamics
within the vector thus modulating the transmission potential (Louis Lambrechts et al.,
2012; Vogels et al., 2019). This happens in nature when mosquitoes take multiple blood
meals from several different hosts, each infected with a different or multiple DENV
serotypes (Shrivastava et al., 2018). In controlled laboratory experiments, using field
derived mosquito populations, there are no differences in dissemination and transmission
rates between DENV-1 and DENV-4 mono-infections in Ae. aegypti, but during co-

7
infection, DENV-4 has a much higher dissemination rate, leading to the exclusive presence
of DENV-4 in the saliva (Vazeille, Gaborit, Mousson, Girod, & Failloux, 2016).
Furthermore, differential replication between DENV-2 and DENV-3 have been shown,
with DENV-2 exhibiting a much higher replication efficiency in both in vitro and in vivo
during co-infection (Quintero-Gil, Ospina, Osorio-Benitez, & Martinez-Gutierrez, 2014).
Additionally, the impact of co-infection with different families of arboviruses in vector
competence has just been recently studied. For example, mosquitoes exposed to double or
triple CHIKV, ZIKV and DENV viruses were capable of transmitting all pathogens
concurrently, without noticeable changes to mosquito infection and dissemination rates (C.
Rückert et al., 2017). Co-infection studies may shed light on the outcome of competitive
processes in field mosquitoes, even if rare, but also importantly allow for direct
comparisons of the transmissibility between viruses.
Compared to other viruses, CHIKV is much less studied. This is in part due to its
very recent re-emergence (Wahid, Ali, Rafique, & Idrees, 2017), but also the requirement
that all in vitro and in vivo CHIKV research be carried out under BSL-3 conditions. To
date, there are only a handful of studies in the literature that examine the basis of the
mosquito’s antiviral response to CHIKV infection (Chowdhury et al., 2020; McFarlane et
al., 2014; Zhao, Alto, Jiang, Yu, & Zhang, 2019). Interestingly, the traditional mosquito
innate immune pathways (Toll, JAK/STAT, RNAi) play little or no role in limiting CHIKV
infection (McFarlane et al., 2014; Shrinet, Srivastava, & Sunil, 2017). Not surprisingly
then, the transcriptional response of mosquitoes infected with CHIKV and DENV also
share little overlap (Shrinet et al., 2017). Additionally, there have been several recent

8
discoveries of novel non-canonical antiviral genes in Drosophila (Kounatidis et al., 2017;
L. Zhang et al., 2020) and in Ae. aegypti (Sigle & McGraw, 2019). The antiviral action of
each of these genes was revealed only by examining population level genetic variation in
the antiviral response. Quantitative genetic studies have also revealed differences in the
transmission potential between arboviruses (Sanchez-Vargas, Olson, & Black, 2021) and
helped understand the genetic basis of Wolbachia mediated pathogen blocking in in Ae.
aegypti (Terradas, Allen, Chenoweth, & McGraw, 2017).
1.4 Vector control
One of the most important outcomes of researching mosquito-pathogen interactions
is the development and optimization of vector control methods. Ae. aegypti natural history
has allowed this species to encroach and establish in new geographic areas (McGregor &
Connelly, 2020). Classic vector control methods, used to this day, include the removal of
breeding containers and areas of stagnant water (D. J. Gubler & Clark, 1996; Jones, Ant,
Cameron, & Logan, 2021; Lobo, Achee, Greico, & Collins, 2018). However, because of
the mosquito’s preference for small and often cryptic breeding grounds, complete removal
its challenging, even for the most prepare abatement programs particularly in areas of high
annual rainfall (Cheong, 1967; McGregor & Connelly, 2020). Additionally, Ae. aegypti
has been shown to breed in septic tanks, sewage plants, cesspits and stormwater drains,
making the task of monitoring mosquito populations even more cumbersome (Arana-
Guardia et al., 2014; Babu, Panicker, & Das, 1983; Barrera et al., 2008). Insecticide-based
approaches offer the next line of suppression against mosquito populations. This method
can target both larvae and adult mosquitoes and it has been responsible for the eradication

9
of diseases like typhus, malaria, yellow fever and even dengue fever in many countries
(Achee et al., 2015; Keatinge, 1949; Rogan & Chen, 2005). The two main problems with
the use of insecticides are that 1) they can cause widespread negative health effects in non-
targets, such as humans, animals or beneficial insects and 2) target insects can develop
resistance, driving the use of higher insecticides concentrations and increasing the negative
effects (Aktar, Sengupta, & Chowdhury, 2009; Calvo-Agudo et al., 2019; Rivero, Vézilier,
Weill, Read, & Gandon, 2010). For Ae. aegypti, many studies have shown low efficiency
of insecticides in wild populations (Maciel-de-Freitas et al., 2014; Marcombe et al., 2011;
Moyes et al., 2017; Plernsub et al., 2016) particularly due to point mutations in the voltage
sodium channel, the main target of several classes of insecticides (Brito et al., 2013; Moyes
et al., 2017).
Novel biotechnological vector control strategies that could work in tandem with the
classic methods are currently being developed (Barrera et al., 2008; Jones et al., 2021; Roiz
et al., 2018; Wilson et al., 2020). Some of the new approaches for reducing arthropod borne
diseases include the use of gene editing technologies like RNA interference (RNAi) and
CRISPR to induce pathogen resistant phenotypes or to kill mosquitoes at early
developmental stages (Chaverra-Rodriguez et al., 2018; Macias, Ohm, & Rasgon, 2017).
Other strategies are the use of sterile insect technique (SIT) involving mass rearing of the
targeted mosquito, followed by sterilization and release into the wild to suppress the
population reproductive potential (Alphey et al., 2010; Kandul et al., 2019; Zheng et al.,
2019). Similarly, the release of insects carrying a dominant lethal (RIDL) uses dominant
lethal transgenes that are suppress during mass laboratory rearing, but once the mosquitoes

10
are release in the wild their offspring will die due to the lack of lethal transgene suppression
in the field (Lin & Wang, 2015; Phuc et al., 2007; Watkinson-Powell & Alphey, 2017).
Finally, the bacteria Wolbachia pipientis is currently being trialed and used as a vector
control method (John H. Werren, Baldo, & Clark, 2008; P.-S. Yen & Failloux, 2020).
Wolbachia is an intracellular Gram-negative bacterium that is maternally transmitted and
naturally occurs in more than 60% of insect species but not in Ae. aegypti (Andrea Gloria-
Soria, Chiodo, & Powell, 2018; Hertig & Wolbach, 1924; Hilgenboecker, Hammerstein,
Schlattmann, Telschow, & Werren, 2008; J. H. Werren, 1997). Wolbachia’s ability to
manipulate the host reproductive biology has been shown to cause a several phenotypes in
its host, including sex ratio distortions such as male-killing, feminization, parthenogenesis
and cytoplasmic incompatibility (CI). In the case of CI, Wolbachia-infected males mate
with uninfected females and prevent them from producing viable offspring, whereas all
other possible crosses are successful. Because Wolbachia is vertically-inherited, infected
females have a reproductive advantage by default, and the infection tends to spread in
populations (Asgharian, Chang, Mazzoglio, & Negri, 2014; McGraw & O'Neill, 1999;
Weeks & Breeuwer, 2001; J. H. Yen & Barr, 1971; Zabalou et al., 2004). After Wolbachia
was introduced into Ae. aegypti, where it formed stably inherited infections, it was also
shown to limit the replication of several pathogens during coinfection in the mosquito
including; ZIKV, YFV, CHIKV and DENV (Hoffmann et al., 2014; Hurk et al., 2012;
Moreira et al.; Walker et al., 2011). Subsequently, Wolbachia has been tested in large scale
field release programs in numerous locations around the globe, where the bacterium has
spread to near fixation and lead to overall reductions in human disease incidence,

11
measuring up to 77% for dengue virus and 60% for chikungunya virus (Durovni et al.,
2019; Indriani et al., 2020; Pinto et al., 2021).
Regardless of the vector control strategy, the effectiveness of these methods can be
shaped by vector-pathogen interactions. Therefore, any insights into vector x virus
interactions will inform the development and successful implementation of control
practices. My research will provide a better understanding of virus-vector dynamics, which
will assist in developing intervention points for genetic modification. My findings will
help expand our model of vectorial capacity by adding components such as dose and
serotype, and our understanding of DENV epidemiology, such as the effect of serotype
competition in both transmission and Wolbachia mediated pathogen blocking.
Additionally, my work will reveal mosquito genes involved with mosquito immunity to
CHIKV, that is less well studied than DENV, offering new potential targets for control.

12
1.5 References
Achee, N. L., Gould, F., Perkins, T. A., Reiner, R. C., Jr., Morrison, A. C., Ritchie, S. A.,
Gubler, D. J., Teyssou, R., & Scott, T. W. (2015). A critical assessment of vector control for
dengue prevention. PLoS Neglected Tropical Diseases, 9(5), e0003655-e0003655.
doi:10.1371/journal.pntd.0003655
Aktar, M. W., Sengupta, D., & Chowdhury, A. (2009). Impact of pesticides use in
agriculture: their benefits and hazards. Interdisciplinary toxicology, 2(1), 1-12.
doi:10.2478/v10102-009-0001-7
Alphey, L., Benedict, M., Bellini, R., Clark, G. G., Dame, D. A., Service, M. W., &
Dobson, S. L. (2010). Sterile-insect methods for control of mosquito-borne diseases: an analysis.
Vector borne and zoonotic diseases (Larchmont, N.Y.), 10(3), 295-311.
doi:10.1089/vbz.2009.0014
Anderson, J. R., & Rico-Hesse, R. (2006). Aedes aegypti vectorial capacity is determined
by the infecting genotype of dengue virus. American Journal of Tropical Medicine and Hygiene,
75(5), 886-892.
Arana-Guardia, R., Baak-Baak, C. M., Loroño-Pino, M. A., Machain-Williams, C.,
Beaty, B. J., Eisen, L., & García-Rejón, J. E. (2014). Stormwater drains and catch basins as
sources for production of Aedes aegypti and Culex quinquefasciatus. Acta Tropicaica, 134, 33-42.
doi:10.1016/j.actatropica.2014.01.011
Armstrong, P. M., & Rico-Hesse, R. (2003). Efficiency of dengue serotype 2 virus strains
to infect and disseminate in Aedes aegypti. The American Society of Tropical Medicine and
Hygiene, 68.
Asgharian, H., Chang, P. L., Mazzoglio, P. J., & Negri, I. (2014). Wolbachia is not all
about sex: male-feminizing Wolbachia alters the leafhopper Zyginidia pullula transcriptome in a
mainly sex-independent manner. Frontiers in microbiology, 5, 430-430. Do i:1 0.3 389 /fmi cb.2
014.00430
Azhar, E. I., Hashem, A. M., El-Kafrawy, S. A., Abol-Ela, S., Abd-Alla, A. M., &
Sohrab, S. S. (2015). Complete genome sequencing and phylogenetic analysis of dengue type 1
virus isolated from Jeddah, Saudi Arabia. Virology Journal, 12. doi:10.1186/s12985-014-0235-7
Babu, C. J., Panicker, K. N., & Das, P. K. (1983). Breeding of Aedes aegypti in closed
septic tanks. Indian Journal of Medical Research, 77, 637.
Bancroft, T. L. (1906). On the aetiology of dengue fever. Australian Medical Gazette.,
25, 17-18.
Barrera, R., Amador, M., Diaz, A., Smith, J., Munoz-Jordan, J. L., & Rosario, Y. (2008).
Unusual productivity of Aedes aegypti in septic tanks and its implications for dengue control.
Medical and Veterinary Entomology, 22(1), 62-69. doi:10.1111/j.1365-2915.2008.00720.x

13
Bartholomay, L. C., Waterhouse, R. M., Mayhew, G. F., Campbell, C. L., Michel, K.,
Zou, Z., Ramirez, J. L., Das, S., Alvarez, K., Arensburger, P., Bryant, B., Chapman, S. B., Dong,
Y., Erickson, S. M., Karunaratne, S. H. P. P., Kokoza, V., Kodira, C. D., Pignatelli, P., Shin, S.
W., Vanlandingham, D. L., Atkinson, P. W., Birren, B., Christophides, G. K., Clem, R. J.,
Hemingway, J., Higgs, S., Megy, K., Ranson, H., Zdobnov, E. M., Raikhel, A. S., Christensen, B.
M., Dimopoulos, G., & Muskavitch, M. A. T. (2010). Pathogenomics of Culex quinquefasciatus
and meta-analysis of infection responses to diverse pathogens. Science (New York, N.Y.),
330(6000), 88-90. doi:10.1126/science.1193162
Bhatt, S., Gething, P. W., Brady, O. J., Messina, J. P., Farlow, A. W., Moyes, C. L.,
Drake, J. M., Brownstein, J. S., Hoen, A. G., Sankoh, O., Myers, M. F., George, D. B., Jaenisch,
T., Wint, G. R. W., Simmons, C. P., Scott, T. W., Farrar, J. J., & Hay, S. I. (2013). The global
distribution and burden of dengue. Nature, 496(7446), 504-507. doi:10.1038/nature12060 http:
//www.nature.com/nature/journal/v496/n7446/abs/nature12060.
Blair, C. D., & Olson, K. E. (2015). The role of RNA interference (RNAi) in arbovirus-
vector interactions. Viruses, 7(2), 820-843. doi:10.3390/v7020820
Brady, O. J., Gething, P. W., Bhatt, S., Messina, J. P., Brownstein, J. S., Hoen, A. G.,
Moyes, C. L., Farlow, A. W., Scott, T. W., & Hay, S. I. (2012). Refining the global spatial limits
of dengue virus transmission by evidence-based consensus. PLoS Neglected Tropical Diseases,
6(8), e1760. doi:10.1371/journal.pntd.0001760
Brito, L. P., Linss, J. G. B., Lima-Camara, T. N., Belinato, T. A., Peixoto, A. A., Lima, J.
B. P., Valle, D., & Martins, A. J. (2013). Assessing the effects of Aedes aegypti kdr mutations on
pyrethroid resistance and its fitness cost. PLoS ONE, 8(4), e60878. doi:10 .137 1/jour nal.p one.0
060878
Calvo-Agudo, M., González-Cabrera, J., Picó, Y., Calatayud-Vernich, P., Urbaneja, A.,
Dicke, M., & Tena, A. (2019). Neonicotinoids in excretion product of phloem-feeding insects kill
beneficial insects. Proceedings of the National Academy of Sciences, 116(34), 16817-16822.
doi:10.1073/pnas.1904298116
Carrington, L. B., & Simmons, C. P. (2014). Human to mosquito transmission of dengue
viruses. Frontiers in Immunology, 5, 290. doi:10.3389/fimmu.2014.00290
Chaverra-Rodriguez, D., Macias, V. M., Hughes, G. L., Pujhari, S., Suzuki, Y., Peterson,
D. R., Kim, D., McKeand, S., & Rasgon, J. L. (2018). Targeted delivery of CRISPR-Cas9
ribonucleoprotein into arthropod ovaries for heritable germline gene editing. Nature
Communications, 9(1), 3008. doi:10.1038/s41467-018-05425-9
Cheng, G., Liu, Y., Wang, P., & Xiao, X. (2016). Mosquito defense strategies against
viral infection. Trends in Parasitology, 32(3), 177-186. doi:10.1016/j.pt.2015.09.009
Cheong, W. H. (1967). Preferred Aedes aegypti larval habitats in urban areas. Bulletin of
the World Health Organization, 36(4), 586-589.

14
Chowdhury, A., Modahl, C. M., Tan, S. T., Wong Wei Xiang, B., Missé, D., Vial, T.,
Kini, R. M., & Pompon, J. F. (2020). JNK pathway restricts DENV2, ZIKV and CHIKV
infection by activating complement and apoptosis in mosquito salivary glands. PLoS Pathogens,
16(8), e1008754. doi:10.1371/journal.ppat.1008754
Christophers, S. (1960). Aedes aegypti (L.) the yellow fever mosquito: its life history,
bionomics and structure: London: The Syndics of the Cambridge University Press, Bentley
House, 200, Euston Road, N.W.I.
Cox, J., Brown, H. E., & Rico-Hesse, R. (2011). Variation in vector competence for
dengue viruses Does not depend on mosquito midgut binding affinity. PLoS Neglected Tropical
Diseases, 5(5), e1172. doi:10.1371/journal.pntd.0001172
Deen, J. L., Harris, E., Wills, B., Balmaseda, A., Hammond, S. N., Rocha, C., Dung, N.
M., Hung, N. T., Hien, T. T., & Farrar, J. J. (2006). The WHO dengue classification and case
definitions: time for a reassessment. The Lancet, 368(9530), 170-173. doi:10.1016/S0140-
6736(06)69006-5
Durovni, B., Saraceni, V., Eppinghaus, A., Riback, T. I. S., Moreira, L. A., Jewell, N. P.,
Dufault, S. M., O'Neill, S. L., Simmons, C. P., Tanamas, S. K., & Anders, K. L. (2019). The
impact of large-scale deployment of Wolbachia mosquitoes on dengue and other Aedes-borne
diseases in Rio de Janeiro and Niterói, Brazil: study protocol for a controlled interrupted time
series analysis using routine disease surveillance data. F1000Research, 8, 1328.
doi:10.12688/f1000research.19859.2
Fontaine, A., Lequime, S., Moltini-Conclois, I., Jiolle, D., Leparc-Goffart, I., Reiner, R.
C., Jr., & Lambrechts, L. (2018). Epidemiological significance of dengue virus genetic variation
in mosquito infection dynamics. PLoS Pathogens, 14(7), e1007187.
doi:10.1371/journal.ppat.1007187
Franz, A. W. E., Kantor, A. M., Passarelli, A. L., & Clem, R. J. (2015). Tissue Barriers to
Arbovirus Infection in Mosquitoes. Viruses, 7(7), 3741-3767. doi:10.3390/v7072795
Gloria-Soria, A., Armstrong, P. M., Powell, J. R., & Turner, P. E. (2017). Infection rate
of Aedes aegypti mosquitoes with dengue virus depends on the interaction between temperature
and mosquito genotype. Proceedings of the Royal Society B: Biological Sciences, 284(1864).
doi:10.1098/rspb.2017.1506
Gloria-Soria, A., Chiodo, T. G., & Powell, J. R. (2018). Lack of evidence for natural
Wolbachia infections in Aedes aegypti (Diptera: Culicidae). Journal of Medical Entomology,
55(5), 1354-1356. doi:10.1093/jme/tjy084
Gould, E. A., & Higgs, S. (2009). Impact of climate change and other factors on
emerging arbovirus diseases. Transactions of The Royal Society of Tropical Medicine and
Hygiene, 103(2), 109-121. doi:10.1016/j.trstmh.2008.07.025
Gubler, D. J. (2011). Dengue, urbanization and globalization: The unholy trinity of the
21(st) century. Tropical Medicine and Health, 39(4 Suppl), 3-11. doi:10.2149/tmh.2011-S05

15
Gubler, D. J., & Clark, G. G. (1996). Community involvement in the control of Aedes
aegypti. Acta Tropica, 61(2), 169-179. doi:10.1016/0001-706x(95)00103-l
Guzman, M. G., & Harris, E. (2015). Dengue. The Lancet, 385(9966), 453-465.
doi:https://doi.org/10.1016/S0140-6736(14)60572-9
Hall, M. D., Bento, G., & Ebert, D. (2017). The Evolutionary consequences of stepwise
infection processes. Trends in Ecology & Evolution, 32(8), 612-623.
doi:10.1016/j.tree.2017.05.009
Hertig, M., & Wolbach, S. B. (1924). Studies on rickettsia-like micro-organisms in
insects. The Journal of medical research, 44(3), 329-374.327.
Hilgenboecker, K., Hammerstein, P., Schlattmann, P., Telschow, A., & Werren, J. H.
(2008). How many species are infected with Wolbachia?--A statistical analysis of current data.
FEMS microbiology letters, 281(2), 215-220. doi:10.1111/j.1574-6968.2008.01110.x
Hoffmann, A. A., Iturbe-Ormaetxe, I., Callahan, A. G., Phillips, B., Billington, K., &
Axford, J. K. (2014). Stability of the wMel Wolbachia infection following invasion into Aedes
aegypti populations. PLoS Neglected Tropical Diseases, 8. doi:10.1371/journal.pntd.0003115
Hurk, A. F., Hall-Mendelin, S., Pyke, A. T., Frentiu, F. D., McElroy, K., & Day, A.
(2012). Impact of Wolbachia on infection with chikungunya and yellow fever viruses in the
mosquito vector Aedes aegypti. PLoS Neglected Tropical Diseases, 6. doi:1 0.1371/ journal.
.0001892
Indriani, C., Tantowijoyo, W., Rancès, E., Andari, B., Prabowo, E., Yusdi, D., Ansari, M.
R., Wardana, D. S., Supriyati, E., Nurhayati, I., Ernesia, I., Setyawan, S., Fitriana, I., Arguni, E.,
Amelia, Y., Ahmad, R. A., Jewell, N. P., Dufault, S. M., Ryan, P. A., Green, B. R., McAdam, T.
F., O'Neill, S. L., Tanamas, S. K., Simmons, C. P., Anders, K. L., & Utarini, A. (2020). Reduced
dengue incidence following deployments of Wolbachia-infected Aedes aegypti in Yogyakarta,
Indonesia: a quasi-experimental trial using controlled interrupted time series analysis. Gates open
research, 4, 50-50. doi:10.12688/gatesopenres.13122.1
Jones, R. T., Ant, T. H., Cameron, M. M., & Logan, J. G. (2021). Novel control strategies
for mosquito-borne diseases. Philosophical Transactions of the Royal Society B: Biological
Sciences, 376(1818), 20190802. doi:10.1098/rstb.2019.0802
Kandul, N. P., Liu, J., Sanchez C, H. M., Wu, S. L., Marshall, J. M., & Akbari, O. S.
(2019). Transforming insect population control with precision guided sterile males with
demonstration in flies. Nature Communications, 10(1), 84. doi:10.1038/s41467-018-07964-7
Kauffman, E. B., & Kramer, L. D. (2017). Zika virus mosquito vectors: competence,
biology, and vector control. The Journal of Infectious Diseases, 216(suppl_10), S976-s990.
doi:10.1093/infdis/jix405
Keatinge, A. F. (1949). A hundred years of insecticides and repellents in the army; a
historical summary. Journal of the Royal Army Medical Corps, 92(6), 290-312.

16
Khoo, C. C. H., Doty, J. B., Held, N. L., Olson, K. E., & Franz, A. W. E. (2013).
Isolation of midgut escape mutants of two American genotype dengue 2 viruses from. Virology
Journal, 10(1), 257. doi:10.1186/1743-422X-10-257
Kounatidis, I., Chtarbanova, S., Cao, Y., Hayne, M., Jayanth, D., Ganetzky, B., &
Ligoxygakis, P. (2017). NF-κB Immunity in the brain determines fly lifespan in healthy aging
and age-related neurodegeneration. Cell Reports, 19(4), 836-848. Do i:10. 1016 /j.celre p.2017
.04.007
Lambrechts, L., Chevillon, C., Albright, R. G., Thaisomboonsuk, B., Richardson, J. H.,
Jarman, R. G., & Scott, T. W. (2009). Genetic specificity and potential for local adaptation
between dengue viruses and mosquito vectors. BMC Ecology and Evolution, 9, 160.
doi:10.1186/1471-2148-9-160
Lambrechts, L., Fansiri, T., Pongsiri, A., Thaisomboonsuk, B., Klungthong, C.,
Richardson, J. H., Ponlawat, A., Jarman, R. G., & Scott, T. W. (2012). Dengue-1 virus clade
replacement in Thailand associated with enhanced mosquito transmission. Journal of Virology,
86(3), 1853-1861. doi:10.1128/jvi.06458-11
Le Flohic, G., Porphyre, V., Barbazan, P., & Gonzalez, J. P. (2013). Review of climate,
landscape, and viral genetics as drivers of the Japanese encephalitis virus ecology. PLoS
Neglected Tropical Diseases, 7(9), e2208. doi:10.1371/journal.pntd.0002208
Lequime, S., Fontaine, A., Ar Gouilh, M., Moltini-Conclois, I., & Lambrechts, L. (2016).
Genetic drift, purifying selection and vector genotype shape dengue virus intra-host genetic
diversity in mosquitoes. PLoS Genetics, 12(6), e1006111. doi:10.1371/journal.pgen.1006111
Leta, S., Beyene, T. J., De Clercq, E. M., Amenu, K., Kraemer, M. U. G., & Revie, C. W.
(2018). Global risk mapping for major diseases transmitted by Aedes aegypti and Aedes
albopictus. International Journal of Infectious Diseases, 67, 25-35. doi: htt ps: //do i.or g/1 0.101
6/j.ijid.2017.11.026
Lin, X., & Wang, G. (2015). Development of a RNAi-based release of insects carrying a
dominant lethal (RIDL) system in Drosophila melanogaster. Science Bulletin, 60(3), 356-362.
doi:https://doi.org/10.1007/s11434-014-0667-x
Lobo, N. F., Achee, N. L., Greico, J., & Collins, F. H. (2018). Modern vector control.
Cold Spring Harbor perspectives in medicine, 8(1), a025643. doi:10.1101/cshperspect.a025643
Lounibos, L. P. (1981). Habitat segregation among African treehole mosquitoes.
Ecological Entomology, 6(2), 129-154. doi:https://doi.org/10.1111/j.1365-2311.1981.tb00601.x
Macdonald, G. (1957). The epidemiology and control of malaria. Amen House, Warwick
Square, London E.C.4.: Oxford University Press.
Macias, V. M., Ohm, J. R., & Rasgon, J. L. (2017). Gene drive for mosquito control:
where did it come from and where are we headed? International Journal of Environmental
Research and Public Health, 14(9). doi:10.3390/ijerph14091006

17
Maciel-de-Freitas, R., Avendanho, F. C., Santos, R., Sylvestre, G., Araújo, S. C., Lima, J.
B. P., Martins, A. J., Coelho, G. E., & Valle, D. (2014). Undesirable consequences of insecticide
resistance following Aedes aegypti control activities due to a dengue outbreak. PLoS ONE, 9(3),
e92424. doi:10.1371/journal.pone.0092424
Marcombe, S., Darriet, F., Tolosa, M., Agnew, P., Duchon, S., Etienne, M., Yp Tcha, M.
M., Chandre, F., Corbel, V., & Yébakima, A. (2011). Pyrethroid resistance reduces the efficacy
of space sprays for dengue control on the island of Martinique (Caribbean). PLoS Neglected
Tropical Diseases, 5(6), e1202. doi:10.1371/journal.pntd.0001202
McFarlane, M., Arias-Goeta, C., Martin, E., O'Hara, Z., Lulla, A., Mousson, L., Rainey,
S. M., Misbah, S., Schnettler, E., Donald, C. L., Merits, A., Kohl, A., & Failloux, A.-B. (2014).
Characterization of Aedes aegypti innate-immune pathways that limit chikungunya virus
replication. PLoS Neglected Tropical Diseases, 8(7), e2994. doi:10.1371/journal.pntd.0002994
McGraw, E. A., & O'Neill, S. L. (1999). Evolution of Wolbachia pipientis transmission
dynamics in insects. Trends in Microbiology, 7(7), 297-302. doi:10.1016/s0966-842x(99)01531-0
McGregor, B. L., & Connelly, C. R. (2020). A review of the control of Aedes aegypti
(Diptera: Culicidae) in the continental United States. Journal of Medical Entomology, 58(1), 10-
25. doi:10.1093/jme/tjaa157
McNeill, J. R. (2010). Mosquito empires: Ecology and war in the greater Caribbean,
1620–1914. Cambridge: Cambridge University Press.
Miller, B. R., Mitchell, C. J., & Ballinger, M. E. (1989). Replication, tissue tropisms and
transmission of yellow fever virus in Aedes albopictus. Transactions of The Royal Society of
Tropical Medicine and Hygiene, 83(2), 252-255. doi:10.1016/0035-9203(89)90667-6
Moreira, L. A., Iturbe-Ormaetxe, I., Jeffery, J. A., Lu, G., Pyke, A. T., Hedges, L. M.,
Rocha, B. C., Hall-Mendelin, S., Day, A., Riegler, M., Hugo, L. E., Johnson, K. N., Kay, B. H.,
McGraw, E. A., van den Hurk, A. F., Ryan, P. A., & O'Neill, S. L. A Wolbachia symbiont in
Aedes aegypti limits infection with dengue, chikungunya, and plasmodium. Cell, 139(7), 1268-
1278. doi:10.1016/j.cell.2009.11.042
Morrison, T. E. (2014). Reemergence of chikungunya virus. Journal of Virology, 88(20),
11644-11647. doi:10.1128/jvi.01432-14
Moyes, C. L., Vontas, J., Martins, A. J., Ng, L. C., Koou, S. Y., Dusfour, I.,
Raghavendra, K., Pinto, J., Corbel, V., David, J.-P., & Weetman, D. (2017). Contemporary status
of insecticide resistance in the major Aedes vectors of arboviruses infecting humans. PLoS
Neglected Tropical Diseases, 11(7), e0005625. doi:10.1371/journal.pntd.0005625
Murray, N. E., Quam, M. B., & Wilder-Smith, A. (2013). Epidemiology of dengue: past,
present and future prospects. Clinical Epidemiology, 5.

18
Ohm, J. R., Baldini, F., Barreaux, P., Lefevre, T., Lynch, P. A., Suh, E., Whitehead, S.
A., & Thomas, M. B. (2018). Rethinking the extrinsic incubation period of malaria parasites.
Parasites & Vectors, 11(1), 178. doi:10.1186/s13071-018-2761-4
Perera, R., & Kuhn, R. J. (2008). Structural proteomics of dengue virus. Current opinion
in microbiology, 11(4), 369-377. doi:10.1016/j.mib.2008.06.004
Petersen, L. R., & Powers, A. M. (2016). Chikungunya: epidemiology. F1000Research,
5, F1000 Faculty Rev-1082. doi:10.12688/f1000research.7171.1
Phuc, H. K., Andreasen, M. H., Burton, R. S., Vass, C., Epton, M. J., Pape, G., Fu, G.,
Condon, K. C., Scaife, S., Donnelly, C. A., Coleman, P. G., White-Cooper, H., & Alphey, L.
(2007). Late-acting dominant lethal genetic systems and mosquito control. BMC Biology, 5(1),
11. doi:10.1186/1741-7007-5-11
Pinto, S. B., Riback, T. I. S., Sylvestre, G., Costa, G., Peixoto, J., Dias, F. B. S.,
Tanamas, S. K., Simmons, C. P., Dufault, S. M., Ryan, P. A., Neill, S. L., Muzzi, F. C., Kutcher,
S., Montgomery, J., Green, B. R., Smithyman, R., Eppinghaus, A., Saraceni, V., Anders, K. L.,
Durovni, B., & Moreira, L. A. (2021). Effectiveness of Wolbachia infected mosquito
deployments in reducing the incidence of dengue and chikungunya in Niterói, Brazil: a quasi-
experimental study. medRxiv, 2021.2001.2031.21250726. doi:10.1101/2021.01.31.21250726
Plernsub, S., Saingamsook, J., Yanola, J., Lumjuan, N., Tippawangkosol, P., Walton, C.,
& Somboon, P. (2016). Temporal frequency of knockdown resistance mutations, F1534C and
V1016G, in Aedes aegypti in Chiang Mai city, Thailand and the impact of the mutations on the
efficiency of thermal fogging spray with pyrethroids. Acta Tropica, 162, 125-132.
doi:https://doi.org/10.1016/j.actatropica.2016.06.019
Powell, J. R. (2016). New contender for most lethal animal. Nature, 540(7634), 525-525.
doi:10.1038/540525c
Powell, J. R. (2018). Mosquito-Borne Human Viral Diseases: Why Aedes aegypti? The
American Society of Tropical Medicine and Hygiene, 98(6), 1563-1565. doi:10.4269/ajtmh.17-
0866
Powell, J. R., & Tabachnick, W. J. (2013). History of domestication and spread of Aedes
aegypti - A Review. Memórias do Instituto Oswaldo Cruz, 108(Suppl 1), 11-17.
doi:10.1590/0074-0276130395
Quintero-Gil, D. C., Ospina, M., Osorio-Benitez, J. E., & Martinez-Gutierrez, M. (2014).
Differential replication of dengue virus serotypes 2 and 3 in coinfections of C6/36 cells and Aedes
aegypti mosquitoes. The Journal of Infection in Developing Countries, 8(7), 876-884.
doi:10.3855/jidc.3978
Reed, W., & Carroll, J. (1901). The Prevention of yellow fever. Public health papers and
reports, 27, 113-129.

19
Ritchie, S. A. (2014). 24 Dengue Vector Bionomics: Why Aedes aegypti is such a good
vector: CABI International Oxford, UK.
Ritchie, S. A., Pyke, A. T., Hall-Mendelin, S., Day, A., Mores, C. N., Christofferson, R.
C., Gubler, D. J., Bennett, S. N., & van den Hurk, A. F. (2013). An explosive epidemic of DENV-
3 in Cairns, Australia. PLoS ONE, 8(7), e68137. doi:10.1371/journal.pone.0068137
Rivero, A., Vézilier, J., Weill, M., Read, A. F., & Gandon, S. (2010). Insecticide control
of vector-borne diseases: When is insecticide resistance a problem? PLoS Pathogens, 6(8),
e1001000. doi:10.1371/journal.ppat.1001000
Rogan, W. J., & Chen, A. (2005). Health risks and benefits of bis(4-chlorophenyl)-1,1,1-
trichloroethane (DDT). The Lancet, 366(9487), 763-773. doi:10.1016/S0140-6736(05)67182-6
Roiz, D., Wilson, A. L., Scott, T. W., Fonseca, D. M., Jourdain, F., Müller, P.,
Velayudhan, R., & Corbel, V. (2018). Integrated Aedes management for the control of Aedes-
borne diseases. PLoS Neglected Tropical Diseases, 12(12), e0006845-e0006845.
doi:10.1371/journal.pntd.0006845
Rückert, C., & Ebel, G. D. (2018). How do virus-mosquito interactions lead to viral
emergence? Trends in Parasitology, 34(4), 310-321. doi:10.1016/j.pt.2017.12.004
Rückert, C., Weger-Lucarelli, J., Garcia-Luna, S. M., Young, M. C., Byas, A. D.,
Murrieta, R. A., Fauver, J. R., & Ebel, G. D. (2017). Impact of simultaneous exposure to
arboviruses on infection and transmission by Aedes aegypti mosquitoes. Nature Communications,
8, 15412. doi:10.1038/ncomms15412
Sanchez-Vargas, I., Olson, K. E., & Black, W. C. t. (2021). The Genetic basis for salivary
gland barriers to arboviral transmission. Insects, 12(1). doi:10.3390/insects12010073
Shrinet, J., Srivastava, P., & Sunil, S. (2017). Transcriptome analysis of Aedes aegypti in
response to mono-infections and co-infections of dengue virus-2 and chikungunya virus.
Biochemical and Biophysical Research Communications, 492(4), 617-623. doi:10.1 0 16/ j.b brc
.2017.01.162
Shrivastava, S., Tiraki, D., Diwan, A., Lalwani, S. K., Modak, M., Mishra, A. C., &
Arankalle, V. A. (2018). Co-circulation of all the four dengue virus serotypes and detection of a
novel clade of DENV-4 (genotype I) virus in Pune, India during 2016 season. PLoS ONE, 13(2),
e0192672. doi:10.1371/journal.pone.0192672
Sigle, L. T., & McGraw, E. A. (2019). Expanding the canon: Non-classical mosquito
genes at the interface of arboviral infection. Insect Biochemistry and Molecular Biology, 109, 72-
80. doi:10.1016/j.ibmb.2019.04.004
Silva, L. A., & Dermody, T. S. (2017). Chikungunya virus: epidemiology, replication,
disease mechanisms, and prospective intervention strategies. Journal of Clinical Investigation,
127(3), 737-749. doi:10.1172/jci84417

20
Smith, D. L., Battle, K. E., Hay, S. I., Barker, C. M., Scott, T. W., & McKenzie, F. E.
(2012). Ross, Macdonald, and a theory for the dynamics and control of mosquito-transmitted
pathogens. PLoS Pathogens, 8(4), e1002588. doi:10.1371/journal.ppat.1002588
Smith, D. L., Perkins, T. A., Reiner, R. C., Jr., Barker, C. M., Niu, T., Chaves, L. F.,
Ellis, A. M., George, D. B., Le Menach, A., Pulliam, J. R., Bisanzio, D., Buckee, C., Chiyaka, C.,
Cummings, D. A., Garcia, A. J., Gatton, M. L., Gething, P. W., Hartley, D. M., Johnston, G.,
Klein, E. Y., Michael, E., Lloyd, A. L., Pigott, D. M., Reisen, W. K., Ruktanonchai, N., Singh, B.
K., Stoller, J., Tatem, A. J., Kitron, U., Godfray, H. C., Cohen, J. M., Hay, S. I., & Scott, T. W.
(2014). Recasting the theory of mosquito-borne pathogen transmission dynamics and control.
Transactions of The Royal Society of Tropical Medicine and Hygiene, 108(4), 185-197.
doi:10.1093/trstmh/tru026
Souza-Neto, J. A., Powell, J. R., & Bonizzoni, M. (2019). Aedes aegypti vector
competence studies: A review. Infection, Genetics and Evolution 67, 191-209.
doi:https://doi.org/10.1016/j.meegid.2018.11.009
Staples, J. E., & Monath, T. P. (2008). Yellow fever: 100 Years of discovery. JAMA,
300(8), 960-962. doi:10.1001/jama.300.8.960
Tabachnick, W. J. (1991). Evolutionary genetics and arthropod-borne disease: The
yellow fever mosquito. American Entomologist, 37(1), 14-26. doi:10.1093/ae/37.1.14
Tanabe, I. S. B., Tanabe, E. L. L., Santos, E. C., Martins, W. V., Araújo, I. M. T. C.,
Cavalcante, M. C. A., Lima, A. R. V., Câmara, N. O. S., Anderson, L., Yunusov, D., & Bassi, Ê.
J. (2018). Cellular and molecular immune response to chikungunya virus infection. Frontiers in
cellular and infection microbiology, 8, 345-345. doi:10.3389/fcimb.2018.00345
Terradas, G., Allen, S. L., Chenoweth, S. F., & McGraw, E. A. (2017). Family level
variation in Wolbachia-mediated dengue virus blocking in Aedes aegypti. Parasites & Vectors,
10(1), 622-622. doi:10.1186/s13071-017-2589-3
Tjaden, N. B., Thomas, S. M., Fischer, D., & Beierkuhnlein, C. (2013). Extrinsic
incubation period of dengue: Knowledge, backlog, and applications of temperature dependence.
PLoS Neglected Tropical Diseases, 7(6), e2207. doi:10.1371/journal.pntd.0002207
Ty Hang, V. T., Holmes, E. C., Veasna, D., Quy, N. T., Tinh Hien, T., Quail, M.,
Churcher, C., Parkhill, J., Cardosa, J., Farrar, J., Wills, B., Lennon, N. J., Birren, B. W., Buchy,
P., Henn, M. R., & Simmons, C. P. (2010). Emergence of the Asian 1 genotype of dengue virus
serotype 2 in Viet Nam: In vivo fitness advantage and lineage replacement in South-East Asia.
PLoS Neglected Tropical Diseases, 4(7), e757. doi:10.1371/journal.pntd.0000757
Vazeille, M., Gaborit, P., Mousson, L., Girod, R., & Failloux, A. B. (2016). Competitive
advantage of a dengue 4 virus when co-infecting the mosquito Aedes aegypti with a dengue 1
virus. BMC Infectious Diseases, 16, 318. doi:10.1186/s12879-016-1666-0

21
Vogels, C. B. F., Rückert, C., Cavany, S. M., Perkins, T. A., Ebel, G. D., & Grubaugh, N.
D. (2019). Arbovirus coinfection and co-transmission: A neglected public health concern? PLoS
Biology, 17(1), e3000130. doi:10.1371/journal.pbio.3000130
Wahid, B., Ali, A., Rafique, S., & Idrees, M. (2017). Global expansion of chikungunya
virus: mapping the 64-year history. Int The Journal of Infectious Diseases, 58, 69-76.
doi:10.1016/j.ijid.2017.03.006
Walker, T., Johnson, P. H., Moreira, L. A., Iturbe-Ormaetxe, I., Frentiu, F. D.,
McMeniman, C. J., Leong, Y. S., Dong, Y., Axford, J., Kriesner, P., Lloyd, A. L., Ritchie, S. A.,
O/'Neill, S. L., & Hoffmann, A. A. (2011). The wMel Wolbachia strain blocks dengue and
invades caged Aedes aegypti populations. Nature, 476(7361), 450-453. doi:http: //www.natur e . c
om/nature/journal/v476/n7361/abs/nature10355
Watkinson-Powell, B., & Alphey, N. (2017). Resistance to genetic insect control:
Modelling the effects of space. Journal of Theoretical Biology, 413, 72-85. doi:https://doi.org/
10.1016/j.jtbi.2016.10.014
Weeks, A. R., & Breeuwer, J. A. (2001). Wolbachia-induced parthenogenesis in a genus
of phytophagous mites. Proceedings. Biological sciences, 268(1482), 2245-2251.
doi:10.1098/rspb.2001.1797
Werren, J. H. (1997). Biology of Wolbachia. The Annual Review of Entomology, 42, 587-
609. doi:10.1146/annurev.ento.42.1.587
Werren, J. H., Baldo, L., & Clark, M. E. (2008). Wolbachia: master manipulators of
invertebrate biology. Nature Reviews Microbiology, 6(10), 741-751. doi:10.1038/nrmicro1969
Wilke, A. B. B., Vasquez, C., Carvajal, A., Medina, J., Chase, C., Cardenas, G., Mutebi,
J.-P., Petrie, W. D., & Beier, J. C. (2020). Proliferation of Aedes aegypti in urban environments
mediated by the availability of key aquatic habitats. Scientific Reports, 10(1), 12925.
doi:10.1038/s41598-020-69759-5
Wilson, A. L., Courtenay, O., Kelly-Hope, L. A., Scott, T. W., Takken, W., Torr, S. J., &
Lindsay, S. W. (2020). The importance of vector control for the control and elimination of vector-
borne diseases. PLoS Neglected Tropical Diseases, 14(1), e0007831. doi:10.1371/journal.pntd.
0007831
Yactayo, S., Staples, J. E., Millot, V., Cibrelus, L., & Ramon-Pardo, P. (2016).
Epidemiology of chikungunya in the Americas. The Journal of Infectious Diseases, 214(suppl 5),
S441-s445. doi:10.1093/infdis/jiw390
Yen, J. H., & Barr, A. R. (1971). New hypothesis of the cause of cytoplasmic
incompatibility in Culex pipiens L. Nature, 232(5313), 657-658. doi:10.1038/232657a0
Yen, P.-S., & Failloux, A.-B. (2020). A Review: Wolbachia-based population
replacement for mosquito control shares common points with genetically modified control
approaches. Pathogens (Basel, Switzerland), 9(5), 404. doi:10.3390/pathogens9050404

22
Zabalou, S., Riegler, M., Theodorakopoulou, M., Stauffer, C., Savakis, C., & Bourtzis, K.
(2004). Wolbachia induced cytoplasmic incompatibility as a means for insect pest population
control. Proceedings of the National Academy of Sciences, 101(42), 15042-15045.
doi:10.1073/pnas.0403853101
Zhang, L., Xu, W., Gao, X., Li, W., Qi, S., Guo, D., Ajayi, O. E., Ding, S.-W., & Wu, Q.
(2020). lncRNA Sensing of a Viral Suppressor of RNAi Activates non-canonical innate immune
signaling in Drosophila. Cell Host & Microbe, 27(1), 115-128.e118.
doi:https://doi.org/10.1016/j.chom.2019.12.006
Zhang, R., Zhu, Y., Pang, X., Xiao, X., Zhang, R., & Cheng, G. (2017). Regulation of
antimicrobial peptides in Aedes aegypti Aag2 Cells. Frontiers in Cellular and Infection
Microbiology, 7, 22. doi:10.3389/fcimb.2017.00022
Zhao, L., Alto, B. W., Jiang, Y., Yu, F., & Zhang, Y. (2019). Transcriptomic analysis of
Aedes aegypti innate immune system in response to ingestion of chikungunya virus. International
Journal of Molecular Sciences, 20(13), 3133. doi:10.3390/ijms20133133
Zheng, X., Zhang, D., Li, Y., Yang, C., Wu, Y., Liang, X., Liang, Y., Pan, X., Hu, L.,
Sun, Q., Wang, X., Wei, Y., Zhu, J., Qian, W., Yan, Z., Parker, A. G., Gilles, J. R. L., Bourtzis,
K., Bouyer, J., Tang, M., Zheng, B., Yu, J., Liu, J., Zhuang, J., Hu, Z., Zhang, M., Gong, J.-T.,
Hong, X.-Y., Zhang, Z., Lin, L., Liu, Q., Hu, Z., Wu, Z., Baton, L. A., Hoffmann, A. A., & Xi, Z.
(2019). Incompatible and sterile insect techniques combined eliminate mosquitoes. Nature,
572(7767), 56-61. doi:10.1038/s41586-019-1407-9

23
Chapter 2
Intra-host growth kinetics of dengue virus in the mosquito Aedes aegypti
Citation: Novelo M, Hall MD, Pak D, Young PR, Holmes EC, McGraw EA. Intra-host
growth kinetics of dengue virus in the mosquito Aedes aegypti. PLoS Pathogens.
2019;15(12):e1008218
2.1 Abstract
Dengue virus (DENV) transmission by mosquitoes is a time-dependent process that
begins with the consumption of an infectious blood-meal. DENV infection then proceeds
stepwise through the mosquito from the midgut to the carcass, and ultimately to the salivary
glands, where it is secreted into saliva and then transmitted anew on a subsequent bite. We
examined viral kinetics in tissues of the Ae. aegypti mosquito over a finely graded time
course, and as per previous studies, found that initial viral dose and serotype strain diversity
control infectivity. We also found that a threshold level of virus is required to establish
body-wide infections and that replication kinetics in the early and intermediate tissues do
not predict those of the salivary glands. Our findings have implications for mosquito GMO
design, modeling the contribution of transmission to vector competence and the role of
mosquito kinetics in the overall DENV epidemiological landscape.

24
2.2 Author summary
DENV infection in the mosquito is a complex and dynamic process. Following
ingestion of an infected blood meal, DENV enters the mosquito midgut epithelial cells,
where it replicates. Subsequently, the virus disseminates and infects other tissues, including
hemocytes, fat body and reproductive organs, ultimately reaching the salivary glands. The
kinetics of infection are influenced by genetic variation in the virus. Comparisons between
strains within single serotypes, have revealed variation in infection rates in mosquitoes. To
explore the role of infectious dose, serotype and tissue in viral infection kinetics we
sampled DENV loads in populations of infected mosquitoes over numerous, sequential
time-points. We reveal that the kinetics of DENV infection in the midgut, carcass and
salivary glands of the mosquito Ae. aegypti are strikingly different among the strains
selected for this study, and that these differences are also driven by the initial infectious
dose.
Keywords: mosquito; Aedes aegypti; DENV; infection dynamics; within-vector
infection; EIP, intra-host; viral kinetics; dengue virus.

25
2.3 Introduction
Dengue is the most prevalent arboviral disease globally (Murray, Quam, & Wilder-
Smith, 2013), with estimates of more than 390 million infections per year (Bhatt et al.,
2013) and over half of the world’s population at risk (Brady et al., 2012). Although
mortality is generally low, the morbidity caused by dengue disease is associated with a
substantial socioeconomic burden (Duane J. Gubler, 2011). Dengue virus (DENV),
transmitted to humans through the bite of the Ae. aegypti mosquito (Simmons , Farrar , van
Vinh Chau , & Wills 2012), is spreading in large part due to the expanding geographic
range of the vector (Powell & Tabachnick, 2013). Previously confined to Africa, Ae.
aegypti is now present in tropical and temperate regions worldwide, and its spread is
assisted by climate change, globalization and ineffective vector control programs (Colón-
González, Fezzi, Lake, & Hunter, 2013).
DENV infection in the mosquito is a complex and dynamic process (Lequime,
Fontaine, Ar Gouilh, Moltini-Conclois, & Lambrechts, 2016). The virus must circumvent
multiple tissue barriers, including the midgut and salivary glands, and infect a range of
intermediate tissues in a stepwise fashion (Franz, Kantor, Passarelli, & Clem, 2015).
Following ingestion, DENV enters the midgut epithelial cells, where it replicates.
Subsequently, the virus disseminates and infects secondary tissues, including hemocytes,
fat body and reproductive tissue, ultimately reaching the salivary glands (Khoo, Doty,
Held, Olson, & Franz, 2013). The midgut is thought to represent the primary barrier to the
process of infection (Franz et al., 2015), capable of preventing many mosquitoes from
reaching the stage of disseminated infections (Cox, Brown, & Rico-Hesse, 2011). The rate

26
of this progression dictates the extrinsic incubation period (EIP), or the delay before a
mosquito can infect another human on a subsequent bite (Black Iv et al., 2002). The EIP
plays an important role in shaping transmission rates (Macdonald, 1957), with longer time
windows reducing the number of opportunities for pathogen transmission over a
mosquito’s lifetime.
The kinetics of infection are equally influenced by genetic variation in the virus.
Dengue fever is caused by four different serotypes of virus (DENV 1–4) (Deen et al., 2006)
that are 65–70% similar at the DNA sequence level across their ~11-kb genomes (Perera
& Kuhn, 2008), while also exhibiting a high-average within-serotype diversity of ~3% at
the amino acid level (Azhar et al., 2015). Comparisons between strains within single
serotypes, have revealed variation in both infection rates and EIP in mosquitoes (Anderson
& Rico-Hesse, 2006; Armstrong & Rico-Hesse, 2003), most likely due to differences in
viral replication rates (Cox et al., 2011). In humans, genetic variation within and between
serotypes also determines relative viral fitness (replication rate), epidemic potential and
virulence (Lambrechts et al., 2012; Ty Hang et al., 2010), with particular strains linked to
more severe clinical manifestations. The intra-host/vector diversity of DENV can also play
a role in transmission, such as variants with a replicative advantage can spread more rapidly
overall, eventually displacing those with lower fitness (Guzman & Harris, 2015). For
example, an uncharacteristically large outbreak of dengue in Cairns, Australia in
2008/2009 was attributed to the very short EIP of the DENV-3 strain in the mosquito
(Ritchie et al., 2013).

27
Surprisingly, little is known about DENV kinetics in the mosquito and how the
virus interacts with individual tissues during infection. Mosquitoes have evolved both
systemic and tissue specific antiviral mechanisms to limit viral replication (Cheng, Liu,
Wang, & Xiao, 2016); thus, viral kinetics may differ between tissues. Given that the virus
moves in a stepwise fashion, selective pressures in an initial tissue might therefore have
cascading effects on the viral kinetics in downstream tissues (Hall, Bento, & Ebert, 2017).
Additionally, different tissue types may offer a diversity of cell types and cellular niches
that may vary in their capacity to support DENV replication. For example, the midgut
epithelium is a dynamic niche, with cells being shed frequently (Zieler, Garon, Fischer, &
Shahabuddin, 2000), whereas other cell types in intermediate tissues may be more stable
sites of virus production.
A better understanding of intra vector kinetics will assist with developing optimal
intervention points for genetic modification, improve our model of vectorial capacity
(Macdonald, 1957) by adding components of dose and serotype, etc. (Christofferson &
Mores, 2011) and, expand our understanding of DENV epidemiology, and vector-virus
interactions such as peaks in viral load and latency periods in the host. Herein we compared
midgut, carcass and salivary gland loads for 4 strains of DENV representing each of the 4
serotypes, fed at either high or low infectious doses, daily over a period of 3 weeks. In so
doing, we were able to assess how the factors of dose and serotype strain diversity define
susceptibility, EIP and transmissibility but also how tissue specific differences and the inter
relationships between tissue kinetics drive transmissibility.

28
2.4 Methods
2.4.1 Ae. aegypti rearing and virus preparation
Approximately 8000 eggs of an Ae. aegypti inbred line collected in Townsville,
Australia 13 generations previously were vacuum hatched and divided into individual 30
× 40 × 8-cm plastic trays containing 1 liter of reverse osmosis (RO) autoclaved water.
Larvae (~150) were placed in individual trays containing 3 liters of RO autoclaved water,
supplemented with common fish food (Tetramin, Melle, Germany). Pupae were collected
and placed into breeding cages, containing approximately 450 mosquitoes each. Adults
were provided 10% sucrose ad libitum. All mosquitoes were reared in a controlled
environment at 26°C, 75% relative humidity and a 12-hr light/dark cycle.
The DENV serotypes/strains used for this experiment are listed in Table 2.1. The
virus was propagated in cell culture, as described previously (Frentiu, Robinson, Young,
McGraw, & O'Neill, 2010). Ae. albopictus C6/36 cells were grown at 26°C in RPMI 1640
medium (Invitrogen, Carlsbad, CA) supplemented with 10% fetal bovine serum (FBS), 1×
Glutamax (Invitrogen) and HEPES buffer. Cells were first allowed to form monolayers of
around 60–80% confluence in T175 flasks (Sigma Aldrich, St. Louis, MO), and then
inoculated with DENV and maintained in RPMI medium supplemented with 2% FBS.
After 7 days post-inoculation, live virus was harvested, titrated via absolute quantification
PCR and adjusted to two final viral loads of 105 and 108 DENV copies per ml. Single-use
aliquots were stored at −80°C for subsequent plaque assay titration.

29
2.4.2 Mosquito infections
Prior to infection, female mosquitoes were sorted and placed in 1-liter plastic cups
with a density of ~150 individuals. Sucrose was removed from the mosquitoes 24 hrs, prior
to oral infection. Double-chamber glass feeders were covered with pig intestine previously
immersed in a 10% sucrose solution. Water heated to 37°C was circulated in the outer
chamber of the feeders, and a 1:1 mix of defibrinated sheep blood and the previously
titrated DENV virus was placed inside the feeder. The mosquitoes were split into 2 feeding
groups, each allowed to feed for ~2 hrs. One group was fed with a DENV infectious blood
meal concentration of 108 and the other with 105 DENV copies/ml. After 24 hrs, blood fed
mosquitoes were identified by visual inspection and separated into cups of 10 individuals.
2.4.3 Mosquito dissection and RNA extraction
Blood fed Ae. aegypti mosquitoes were collected daily, 10 per feeding group, over
20 days. Mosquitoes were anesthetized by chilling and dissected on a chill plate in a drop
of sterile phosphate buffer saline (PBS). Midgut, salivary glands and carcass tissues were
Table 2.1. The DENV strains used in this study
Serotype Strain Passage Accession
number
Place of origin Collection date
DENV-1 FR-50 10 FJ432734.1 Vietnam 2007
DENV-2 ET-300 11 EF440433.1 East Timor 2000
DENV-3 Cairns/08/09 9 JN406515.1 Australia 2008
DENV-4 33-188 12 None Vietnam NA
NA: not available

30
placed in separated 1.5-ml microcentrifuge tubes (Sarstedt, Nümbrecht, Germany)
containing 200 µl of TRIzol reagent (Invitrogen, Carlsbad, CA, USA) and 2-mm glass
beads. Samples were homogenized and frozen at −80°C. RNA extraction was performed
using the TRIzol RNA extraction protocol according to the manufacturer’s instructions,
and RNA was eluted in 25 µl of nucleic acid-free water. Samples were DNase treated (Life
Technologies, Carlsbad, CA, USA) according to the manufacturer instructions (Amuzu,
Simmons, & McGraw, 2015). Total RNA was determined with a NanoDropTM lite
spectrophotometer (Thermo Scientific, Waltham, MA).
2.4.4 DENV absolute quantification via RT-qPCR
Quantitative real-time PCR (RT-qPCR) reactions for DENV detections were
performed with 4× TaqMan® fast virus 1-step master mix (Roche Applied Science,
Switzerland), PCR-grade water, 10 µM of DENV primers and probe and 2.5 µl of RNA in
a final volume of 10 µl. Reactions were run in microseal 96 microplates (Life Technologies
Life, Technologies, Carlsbad, CA) covered with optically clear film. LightCycler 480
(Roche Applied Science, Switzerland) thermal cycling conditions were 50°C for 5 min for
reverse transcription, 95°C for 10 s for RT inactivation/denaturation followed by 50
amplification cycles of 95°C for 3 s, 60°C 30 s and 72°C for 1 s.
Standard curves were generated from triplicate samples on each plate spanning the
range of 10 to 108 copies/reaction of DENV fragment copies. DENV standards were
constructed as described elsewhere (Ye et al., 2015), and they contained the 3’UTR of the
DENV genome. The limit of detection was set at 10 copies in this experiment; water was

31
used as a negative control and standards were run in triplicates. The concentration of
DENV genome copies in each sample was extrapolated from the standard curve as DENV
copies per nanogram of total RNA.
The primer sequences used for the detection of DENV, as previously used (Ye et
al., 2014), were F: 5′-AAGGACTAGAGGTTAGAGGAGACCC-3′, R: 5′CGTTC TG TG
CCTGGAATGATG-3′ and P: 5′-HEX- AACAG CATATTGA CGCTGGGAGAG ACC
AGA-BHQ1-3′.
2.4.5 Survival assay
A second set of Townsville mosquitoes were reared in the same conditions as
above. Mosquitoes were blood fed with the two viral concentrations as above (108 and 105
DENV copies/ml). A total of 80 fed mosquitoes per concentration were separated into clear
200-ml cups, each containing 10 individuals. They were monitored daily to assess death
until all had expired. Neither DENV serotypes nor infectious doses had an effect on
survival compared to a blood-only fed control (Appendix A).
2.4.6 Data analysis
Data analysis was carried out in R v 3.6.0 (http://www.r-project.org/). For
statistical analysis of the infection frequency, quasibinomial GLM was used to correct
for overdispersion of data, and Tukey for contrasts was used for post hoc comparisons.
For the kinetics of DENV virus, first we fit a 3-parameter logistic DRC model to DENV
loads in each tissue, dose and serotype combination, which modeled the upper asymptote

32
or the maximum DENV load, the slope or growth rate at the midpoint, and the midpoint or
infection age which is halfway between the lower and upper asymptotes (i.e. ED50). We
then estimated separate trends for each treatment combination. From the candidate models,
the best fitting model for each treatment combination was selected using Akaike’s
information criterion (AIC). For MG and CA tissues the best fitting model was the 3-
parameter logistic model, while for the SG tissue we identified a linear relationship
between DPI and DENV load. DRC parameters and pairwise comparisons were estimated
with the DRC package (Ritz, Baty, Streibig, & Gerhard, 2016). The best model was
selected based on the AIC (yielding the lowest) values. In general, for the salivary
glands, the best fit was a linear model; thus, they were subsequently analyzed in this
fashion. For the average DENV load at DPI 20, significant differences were based on
Tukey post hoc comparison following ANOVAs. All DENV loads were reported on a log
scale given the value range.
2.5 Results
We orally challenged inbred wild type Ae. aegypti mosquitoes with two infectious
doses (108 and 105 DENV copies/ml), representative of plasma viremia ranges in humans
(Duong et al., 2015), of the 4 DENV serotypes. In each vector competence experiment,
mosquitoes were blood fed with a strain representing each of the 4 DENV serotypes at the
2 infectious doses. Ten individuals were then collected daily for 20 days, starting from the
first day post-infection (DPI), to measure DENV infection status and infection kinetics in
3 key tissues - midgut (MG), carcass (CA) and salivary glands (SG) - per mosquito.

33
2.5.1 The proportion of DENV-infected mosquitoes is affected by both infectious dose
and DPI
Infection status was determined by analyzing the proportion of individuals positive
for DENV in each tissue at each DPI, based on the presence of a positive qRT-PCR.
Infection prevalence (Figure 2.1) in the MG, CA and SG in all the serotypes was
significantly influenced by the infectious dose and DPI (Table 2.2). In only one case was
there a significant interaction between these 2 factors (Table 2.2, DENV-3 SG, GLM,
F=5.23, p=0.02), indicating that, in general, dose and age of infection act independently to
shape infection status.
Mosquito populations fed with higher doses had higher infection rates across
tissues and strains (Figure 2.1). The DENV-1 and -2 strains exhibited a higher infection
rate in all tissues and at both infectious doses compared to the DENV-3 and -4 strains.
DENV-4 exhibited especially low infection rates in all tissues at the low infectious dose.
The significance of DPI demonstrates that infection status is heavily time-dependent. For
the DENV-1 and -2 strains, in the high infectious dose, infection rates were constantly high
throughout 20 DPI in contrast to the low infectious dose, where there is noticeable
difference between the infection rates between early and late DPI. For the DENV-3 strain,
both infectious doses peaked in infection rate at ~12 days in all tissues. In the DENV-4
strain, infection levels are weaker than the other strains, which may contribute to the greater
variation in the effect of time across tissues and doses.

34
2.5.2 EIP is affected by the serotype strain and initial infectious dose
If we take SG detection of viral load as a proxy for presence of virus in the saliva,
the susceptibility data (Figure 2.1) can also be used to shed light on EIP. EIP can be
measured in multiple ways, including the day of first arrival of virus in the tissue/saliva or
the point at which 50% of the population has infected tissue/saliva. For all strains and
infectious doses, viral load can be detected in the SG already by day 2. In many cases,
especially for low doses, viral load appears in the SG and then detection is lost for several
days before returning and becoming stable (Figure 2.1). If we calculate an EIP50 for
populations where infection continues for more than two days, we see that infectious dose
and serotype strain diversity are predictors; for the strains representing DENV 1-4 at low-
dose, they are 5, 6, 9 and 15 days, respectively, while for the high-dose they are 1, 3, 7 and
7 days.

35
Figure 2.1: Susceptibility of DENV strains by DPI, tissue and infectious dose. The percentage
of DENV infectious mosquitoes calculated daily out of 10 individuals. Low infectious dose in red
and high infectious dose in blue. Stars represent EIP50 at the salivary gland for both infectious
doses. Statistical analysis is presented in Table 2.3.

36
Table 2.2. Susceptibility by Strain, Tissue, Dose and DPI
Serotype Tissue Factor F df p-value Sig.
DENV-1
MIDGUT
DOSE 29.79 38 <0.001 ***
DPI 19.38 37 <0.001 ***
DOSE: DPI 0.4 36 0.50 ns
CARCASS
DOSE 49.33 38 <0.001 ***
DPI 30.68 37 <0.001 **
DOSE: DPI 0.31 36 0.57 ns
SALIVARY
GLANDS
DOSE 31.78 38 <0.001 ***
DPI 12.09 37 0.001 **
DOSE: DPI 0.40 36 0.52 ns
DENV-2
MIDGUT
DOSE 16.76 38 <0.001 ***
DPI 4.67 37 0.03 *
DOSE: DPI 0.30 36 0.58 ns
CARCASS
DOSE 15.86 38 <0.001 ***
DPI 7.49 37 0.001 **
DOSE: DPI 0.12 36 0.72 ns
SALIVARY
GLANDS
DOSE 5.21 38 0.02 *
DPI 30.71 37 <0.001 ***
DOSE: DPI 0.25 36 0.61 ns
DENV-3
MIDGUT
DOSE 16.28 38 <0.001 ***
DPI 18.52 37 <0.001 ***
DOSE: DPI 0.08 36 0.77 ns
CARCASS
DOSE 15.48 35 <0.001 ***
DPI 6.24 34 0.01 *
DOSE: DPI 1.04 33 0.31 ns
SALIVARY
GLANDS
DOSE 6.95 38 0.01 *
DPI 13.72 37 <0.001 ***
DOSE: DPI 5.23 36 0.02 *
DENV-4
MIDGUT
DOSE 30.69 37 <0.001 ***
DPI 17.35 36 <0.001 ***
DOSE: DPI 1.47 35 0.23 ns
CARCASS
DOSE 20.78 33 <0.001 ***
DPI 14.01 32 <0.001 ***
DOSE: DPI 0.66 34 0.42 ns
SALIVARY
GLANDS
DOSE 10.62 38 0.002 **
DPI 18.07 37 <0.001 ***
DOSE: DPI 0.12 36 0.73 ns
*P<0.05, **P<0.01, ***P<0.001. ns. not significant.

37
2.5.3 Intra-host DENV kinetics are influenced by initial infectious dose and strain
To quantify the kinetics of DENV infection, we assessed whether the cumulative
change in DENV load in each tissue followed a trend that was either a sigmoidal or linear
over time. Thus, we first fit a 3-parameter logistic model to DENV loads in each tissue,
dose and serotype combination, which modeled the upper asymptote or the maximum
DENV load, the slope or growth rate at the midpoint, and the midpoint or infection age
which is halfway between the lower and upper asymptotes (i.e., ED50). We then estimated
separate trends for each treatment combination. From the candidate models, the best fitting
model for each treatment combination was selected using Akaike’s information criterion
(Appendix A). For MG and CA tissues the best fitting model was the 3-parameter logistic
model, while for the SG tissue we identified a linear relationship between DPI and DENV
load. We used the slope and maximum DENV load parameters to compare the behavior of
the different serotypes at different infectious doses and across tissues and times post-
infection (Figure 2.2).
2.5.4 Some treatments do not show substantial DENV replication and others exhibit
subpopulation structure of DENV load in infected mosquitoes
For a number of treatments DENV loads did not rise to very high levels (Figure
2.2). For example, in the low-dose categories neither the DENV-3 nor -4 strains reached a
load higher than log 103 in any tissue as measured by population mean. In these cases,
although we were able to fit a dose response curve (DRC) to the data, the parameters of
max load and growth rate are not informative (Appendix A). Therefore, the subsequent

38
discussion of growth parameter estimates focuses only on those treatments with substantial
DENV loads and replication (Tables 2.3 and 2.4). For the remaining treatments we aimed
to fit a single DRC to the different treatment combinations (infectious dose, tissue and
strain). In several treatments, however, we identified potential subpopulation structure,
with a proportion of the mosquitoes exhibiting high loads and the rest exhibiting very low
loads. Those treatments were the low infectious dose in the MG and CA for strains of
serotype DENV-1 and -2 and the high infectious dose in the MG and CA for strains of
serotypes DENV-3 and -4 (Figure 2.2). After noticing subpopulation structure in our data,
we applied Hartigan’s dip test for all datasets. For any with a significant result, we then
used a bimodality coefficient test to partition the data. We found that using a coefficient of
bimodality of >0.75 effectively split our datasets into two clear subpopulations (Appendix
A) using the following criteria: (1) for the y-axis, we selected the lowest load in the
histogram located between the two highest peaks (Appendix A); and (2) for the x-axis, we
identified the DPI where the two populations began to diverge.
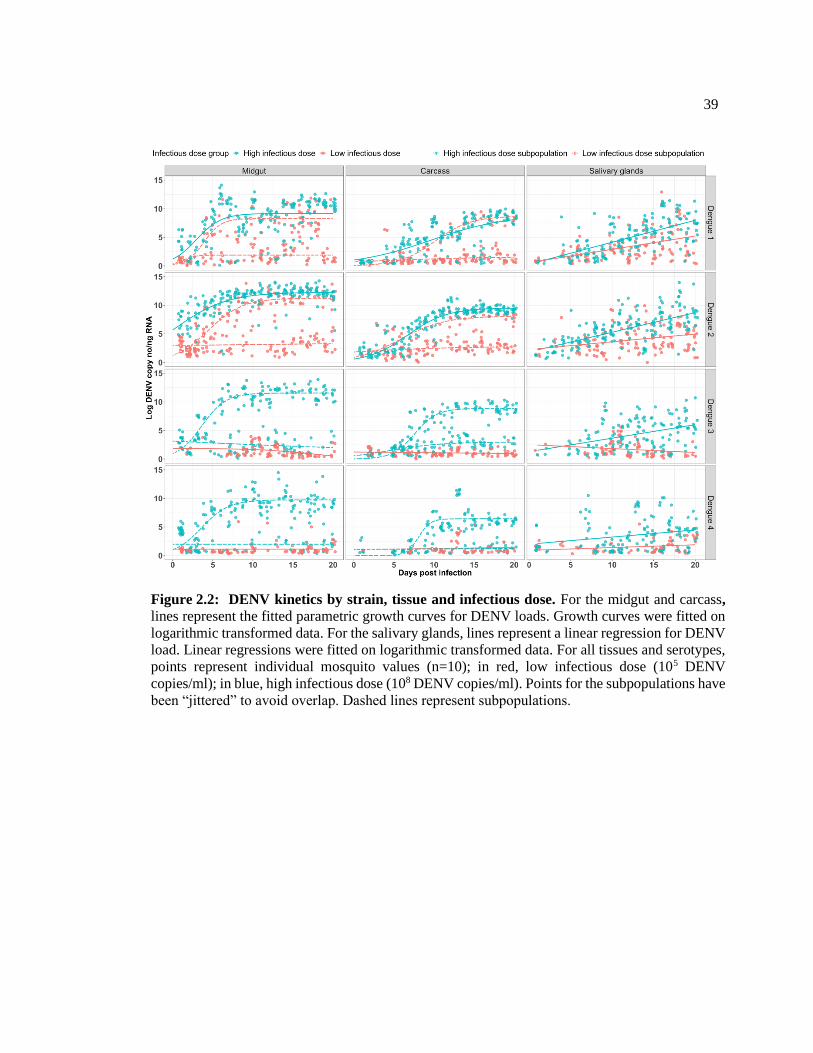
39
Figure 2.2: DENV kinetics by strain, tissue and infectious dose. For the midgut and carcass,
lines represent the fitted parametric growth curves for DENV loads. Growth curves were fitted on
logarithmic transformed data. For the salivary glands, lines represent a linear regression for DENV
load. Linear regressions were fitted on logarithmic transformed data. For all tissues and serotypes,
points represent individual mosquito values (n=10); in red, low infectious dose (105 DENV
copies/ml); in blue, high infectious dose (108 DENV copies/ml). Points for the subpopulations have
been “jittered” to avoid overlap. Dashed lines represent subpopulations.

40
Table 2.3 DRC parameters for successful infections by tissue, infectious dose and strain
Infectious
dose Serotype Tissue Parameter
Parameter
estimate
Standard
error p-value Sig.
HIGH
DENV-3
MIDGUT
Growth rate
0.66 0.07 <0.001 ***
DENV-1 0.64 0.21 0.003 **
DENV-4 0.59 0.11 <0.001 ***
DENV-2 0.31 0.04 <0.001 ***
DENV-4
CARCASS
1.36 0.48 0.005 **
DENV-3 0.73 0.1 <0.001 ***
DENV-2 0.4 0.03 <0.001 ***
DENV-1 0.21 0.05 <0.001 ***
DENV-2
MIDGUT
Max DENV
load
12.2 0.2 <0.001 ***
DENV-3 11.5 0.17 <0.001 ***
DENV-4 9.74 0.26 <0.001 ***
DENV-1 9.11 0.27 <0.001 ***
DENV-2
CARCASS
9.53 0.16 <0.001 ***
DENV-1 8.99 1.24 <0.001 ***
DENV-3 8.85 0.19 <0.001 ***
DENV-4 6.44 0.21 <0.001 ***
DENV-1
MIDGUT
ED50
3.6 0.38 <0.001 ***
DENV-2 3.6 0.43 0.28
DENV-3 2.94 0.19 <0.001 ***
DENV-4 0.46 0.31 <0.001 ***
DENV-1
CARCASS
9.61 1.72 <0.001 ***
DENV-4 8.06 0.32 <0.001 ***
DENV-3 7.56 0.18 <0.001 ***
DENV-2 6.69 0.19 <0.001 ***
LOW
DENV-1 MIDGUT
Growth rate
0.92 0.27 0.001 ***
DENV-2 0.46 0.05 <0.001 ***
DENV-1 CARCASS
0.45 0.07 <0.001 ***
DENV-2 0.36 0.08 <0.001 ***
DENV-2 MIDGUT
Max DENV
load
11.24 0.27 <0.001 ***
DENV-1 8.28 0.27 <0.001 ***
DENV-1 CARCASS
8.7 0.44 <0.001 ***
DENV-2 8.1 0.37 <0.001 ***
DENV-2 MIDGUT
ED50
4.44 0.26 <0.001 ***
DENV-1 3.88 0.33 <0.001 ***
DENV-1 CARCASS
10.1 0.53 <0.001 ***
DENV-2 5.77 0.69 <0.001 ***
*P<0.05, **P<0.01, ***P<0.001. p-value indicates if parameter estimate is different than zero.
Standard error of parameter estimates. Treatments ranked by parameter estimate.

41
Table 2.4 DRC parameter contrasts for successful infections between strains, infectious
dose and subpopulations
Infectious
dose Serotype contrast Tissue
Parameter
estimate
Standard
error p-value Sig.
HIGH
DENV-1 vs DENV 2
MIDGUT
Growth rate 0.18 0.08 ns
Max DENV
load 0.36 <0.001 ***
CARCASS
Growth rate 0.05 0.001 **
Max DENV
load 1.02 0.58 ns
LOW
MIDGUT
Growth rate 0.25 0.07 ns
Max DENV
load 0.39 <0.001 ***
CARCASS
Growth rate 0.11 0.42 ns
Max DENV
load 0.59 0.31 ns
HIGH DENV-1 vs DENV-3
MIDGUT
Growth rate 0.20 0.94 ns
Max DENV
load 0.34 <0.001 ***
CARCASS
Growth rate 0.13 <0.001 ***
Max DENV
load 1.00 0.88 ns
HIGH DENV-1 vs DENV-4
MIDGUT
Growth rate 0.20 0.78 ns
Max DENV
load 0.33 0.08 ns
CARCASS
Growth rate 0.46 0.01 **
Max DENV
load 0.99 0.01 **
HIGH DENV-2 vs DENV-3
MIDGUT
Growth rate 0.13 0.01 **
Max DENV
load 0.33 0.05 ns
CARCASS
Growth rate 0.13 0.01 **
Max DENV
load 0.37 0.04 **
HIGH DENV-2 vs DENV-4
MIDGUT
Growth rate 0.13 0.03 **
Max DENV
load 0.30 <0.001 ***
CARCASS
Growth rate 0.46 0.04 ***
Max DENV
load 0.30 <0.001 ***
HIGH DENV-3 vs DENV-4
MIDGUT
Growth rate 0.14 0.63 ns
Max DENV
load 0.32 <0.001 ***
CARCASS
Growth rate 0.43 0.15 ns
Max DENV
load 0.29 <0.001 ***
*P<0.05, **P<0.01, ***P<0.001; ns, not significant.

42
2.5.5 DENV load is always higher in the midgut than other tissues for high infectious
dose
In all serotypes, maximum DENV loads were consistently higher in the MG than
the CA. Because we fit linear models to the SG data, we examined the average DENV load
at the last DPI (20 days) in lieu of a maximum load (Appendix A). For all serotypes and
high infectious doses, the SG load at this point was lower than the CA loads. This strong
pattern suggests a “trickledown” effect for DENV load across the sequentially infected
tissues (Figure 2.3). In contrast, low infectious doses resulted in low DENV loads in all
tissues and for all strains, and DENV-3 and -4 could not be evaluated due to their lack of
successful infection and failure of the selected model to run. Additionally, we ran
generalized linearized models on DENV load in the MG and CA (for high dose only) as a
predictor for the SG DENV load. We found no significant predictive ability regardless of
the serotype and tissue (Appendix A).

43
2.5.6 Tissue-specific DENV kinetics vary by strain in the high infectious dose
In the MG, the maximum loads of DENV-2 (12.2 log c/ng) and -3 (11.5 log c/ng)
are similarly high and greater than the strains representing serotypes 1 (9.11 log c/ng) and
4 (9.74 log c/ng) (Table 2.3). In the CA, the DENV-1 (8.99 log c/ng) and -2 (9.53 log c/ng)
strains have the highest loads and are both significantly different from DENV-3 (8.85 log
c/ng) and -4 (6.44 log c/ng) strains (Table 2.4). In the SG, the average final load (DPI 20)
for the DENV-2 strain is not different from the DENV-1 and -3 strains but is greater than
the DENV-4 (Appendix A). Although we have examined only one representative strain
from each serotype, these patterns suggest that DENV-2 may be better at replicating in
Figure 2.3: “Trickledown” or declining viral load with tissue progression. Average 20 DPI
DENV load for the 4 dengue serotypes across infectious dose and tissues. Box plots show median
values (horizontal line in the box), 25–75% interquartile range (upper–lower limits of the box),
95% range of data (error lines).

44
mosquitoes and that the other serotypes do not show consistent patterns of dominance over
one another across tissues. For growth rate, no one strain appeared to dominate over the
others across multiple tissues (Tables 2.4 and 2.5). For example, the DENV-3 strain has
the highest growth rate (change in viral copies/day) in the MG (0.66 log DENV copies per
DPI), whereas DENV-2 strain has the lowest (0.31 log DENV copies per DPI). In the CA,
DENV-4 has the highest growth rate (1.36 log DENV copies per DPI), whereas DENV-1
has the lowest (0.21 log DENV copies per DPI). Interestingly, in the SG all strains have
distinctly different growth rates, ranked in the following order: DENV-1, 2, 3, and 4 (Table
2.5). This suggests that the growth rates in early process/upstream tissues are not predictive
of rates in the final tissue controlling transmissibility.
2.5.7 The DENV-2 strain has a higher maximum DENV load than the DENV-1 strain
in the low infectious dose
For the low infectious dose, we only considered mosquitoes infected with either
DENV-1 or -2 strains, as the remaining strains did not exhibit “successful” infections
meaning that even though infection rate was high, the actual DENV load was substantially
low. Here, we saw that the DENV-2 strain had a significantly higher max DENV load in
the MG and a higher average final load in the SG than the DENV-1 strain. For growth rate,
there were no differences between the DENV-1 and DENV-2 strains in the MG or CA, but
in SG, the DENV-1 strain was significantly higher (Table 2.6).

45
Table 2.5 Linear regression model for SG
Infectious
dose Serotype Term Slope
Standard
error F p-value Sig
LOW
DENV-1 DPI 0.23 0.03 6.12 <0.01 **
DENV-2 DPI 0.13 0.03 3.51 <0.01 **
DENV-3 DPI −0.06 0.03 -1.81 0.073 ns
DENV-4 DPI 0.04 0.02 2.31 0.023 *
HIGH
DENV-1 DPI 0.38 0.03 10.98 <0.01 **
DENV-2 DPI 0.34 0.03 11.38 <0.01 **
DENV-3 DPI 0.23 0.04 5.24 <0.01 **
DENV-4 DPI 0.12 0.04 2.87 <0.01 **
*P<0.05, **P<0.01, ***P<0.001. Tukey for contrasts. ns, not significant.
Table 2.6 Serotype contrasts for growth rate in SG
Infectiou
s dose Serotype contrast
Standard
error df p-value Sig
LOW
DENV-1 – DENV-2 0.16 1026 0.24 ns
DENV-1 – DENV-3 1.81 1026 <.0001 ***
DENV-1 – DENV-4 2.14 1026 <.0001 ***
DENV-2 – DENV-3 2.95 1026 <.0001 ***
DENV-2 – DENV-4 3.49 1026 <.0001 ***
DENV-3 – DENV-4 0.40 1026 0.97 ns
HIGH
DENV-1 – DENV-2 0.08 1026 0.0001 ***
DENV-1 – DENV-3 0.58 1026 0.004 **
DENV-1 – DENV-4 1.15 1026 <.0001 ***
DENV-2 – DENV-3 1.64 1026 <.0001 ***
DENV-2 – DENV-4 3.27 1026 <.0001 ***
DENV-3 – DENV-4 0.51 1026 0.054 ns
*P<0.05, **P<0.01, ***P<0.001. Tukey for contrasts. ns, not significant.

46
2.6 Discussion
To explore the role of infectious dose, serotype and tissue in viral infection kinetics
we sampled DENV loads in populations of infected mosquitoes over numerous, sequential
time-points of the mosquito’s lifespan relevant to transmission in the field. We reveal that
the kinetics of DENV infection in the midgut, carcass and salivary glands of the mosquito
Ae. aegypti are strikingly different among the strains selected for this study, and that these
differences are also driven by the initial infectious dose. Specifically, we showed that (1)
initial infectious dose dictates infection frequency, with the DENV-1 and -2 strains
analyzed here infecting a greater proportion of mosquitoes than the DENV-3 and -4 strains;
(2) for DENV-3 and -4 strains only, the high infectious dose led to strong infection, with
lower doses producing lower viral replication; (3) viral growth rates in particular tissues
did not predict the total DENV load in that same tissue, with some strains having higher
growth rate but lower total DENV load; (4) in the salivary glands all strains have
independent growth rates not predicted by growth rates in previous tissues; and (5) for
every tissue and infectious dose, the DENV-2 strain reached a higher viral load according
to both the average final load at 20 DPI and max DENV load.
The initial infectious dose has been shown to influence the likelihood of a mosquito
becoming infected following an infectious blood meal and whether there is ultimately
dissemination to the salivary glands after a blood meal (D. J. Gubler, Nalim, Tan, Saipan,
& Sulianti Saroso, 1979; Rosen, Roseboom, Gubler, Lien, & Chaniotis, 1985). The amount
of virus in human blood required to infect ~50% of a mosquito population varies between
serotypes, with DENV-1 and -2 requiring ~10-fold less than DENV-3 and -4. This same

47
rank order is observed here. However, strain variation within each serotype is substantial
(Fontaine et al., 2018), so further comparisons across multiple strains within each serotype
are clearly needed to draw any conclusions about whether the strains we have selected are
predictive of the entire serotype.
We were also able to identify a potential threshold of virus needed in the blood
meal to initiate successful infections and assure likely transmissibility via the salivary
glands. A similar threshold effect has been observed in Drosophila melanogaster
populations challenged with systemic bacterial infections (Duneau et al. 2017). The study
identified two possible outcomes of infection; (1) where hosts died with high bacterial
burden or (2) where hosts survived with chronic infection and low pathogen load. Bacterial
load and the time it took the host to establish an immune response were found to determine
the trajectory of infection. Similarly, we saw either high infection (leading to
transmissibility) or low/loss of infection. Interestingly, for our study, the threshold effect
was driven by the initial infectious dose and by serotype. DENV-1 and -2 strains required
less virus to establish an infection, whereas, DENV-3 and -4 strains required substantially
more. The failure to initiate a successful infection could be due to poor replication in the
midgut or the midgut preventing dissemination (Cox et al., 2011; Khoo et al., 2013;
Taracena et al., 2018). This pattern is not unique to the midgut, however, with low and
inefficient replication also in disseminated tissues, such as the carcass. The data may
simply be the result of low population sizes of virus in combination with the large number
of vacant cellular niches. During infection DENV selectively annexes and manipulates host
metabolism and machinery to increase viral translation and replication (Chotiwan et al.,

48
2018; Choy, Sessions, Gubler, & Ooi, 2015; Reid et al., 2018), while also enhancing
replication of mosquito cells to increase niche availability in the mosquito (Helt & Harris,
2005). The more virions that infect a given cell, the more efficient this process becomes
and, thus, more viruses are produced. Initial viral load combined with inherent replicative
differences between strains is likely to produce the pattern we see in our data, that the
DENV-1 and 2 strains require a lower dose to produce successful infections.
Interestingly, higher viral growth rates in tissues did not necessarily lead to higher
DENV loads. For example, for the high infectious dose of DENV-1, the midgut had the
highest max DENV load compared to the carcass, however its growth rate was smaller.
This relationship was seen for the other treatments as well. It is possible that the migration
of viruses between tissues via the tracheal system or hemolymph (Forrester, Guerbois,
Seymour, Spratt, & Weaver, 2012) or directly from nearby tissues is impacting the total
viral counts. In support of tracheal or hemolymph travel is the initial spike in infection rate
in salivary glands followed by a reduction to zero and then a second front of infection
several days later. It is possible that the latter wave represents the larger migration of virus
from bodily tissues that then establishes permanent infection. Given how the circulatory
system of the mosquito works, viral particles could be carried from tissues that are not in
immediate proximity to the salivary glands. This process could directly affect growth rate
in upstream tissues. The salivary glands also represent a special case of the entry and exit
balance, with virus being excreted into the saliva and hence lost from tissue measures.
Under a more natural setting additional factors would likely affect kinetics including,
exposure to non-infectious blood meals prior to consumption of an infectious (Zhu et al.,

49
2017), consumption of sequential dengue infectious meals (Muturi, Buckner, & Bara,
2017) or interactions between DENV and other viruses present in blood meals or the
mosquito body (Le Coupanec et al., 2017).
The disconnect between kinetics in intermediate tissues and those of the salivary
glands suggests that there are either stochastic processes driving kinetics in early tissues
that do not predict how many viruses make it to the salivary glands, or that factors affecting
replication rate in the salivary glands are somewhat independent (Cheng et al., 2016). For
example, previous research has demonstrated that the strength and efficacy of immunity
varies across tissues (Cheng et al., 2016) and in response to different serotypes (Smartt,
Shin, & Alto, 2017). It is also possible that DENV is modulating the vector immune
response in a strain- or tissue-dependent manner (Sim & Dimopoulos, 2010). Differences
between the salivary glands and other tissues may also be due to the number of cellular
niches, and/or unique replication rates in those niches. For example, in contrast to the
midgut epithelium, DENV tropism in the salivary glands appears to be heterogenous within
the tissue; this might be due to the distribution of cellular receptors (Cao-Lormeau, 2009),
with a possible higher concentration of viral entry receptors in the lateral distal lobes of the
tissue (Salazar, Richardson, Sánchez-Vargas, Olson, & Beaty, 2007). In future studies, it
would be useful to quantify the production rate of virus using negative strand strain-specific
PCR methods (Raquin & Lambrechts, 2017) in the midgut and in the salivary glands
relative to the availability of cellular niches.
The differences in viral kinetics between the DENV strains analyzed here,
particularly in the salivary glands, might explain disparities in transmission potential in the

50
field. A range of studies have linked different genotypes to overall dominance in terms of
circulation in human populations both at local and global scales (Dumre et al., 2017;
Liebman et al., 2012; Sasmono et al., 2018; Shrivastava et al., 2018). Additionally, DENV
variants with a replicative advantage in both the host and vector can spread more rapidly,
eventually displacing those with lower fitness (Guzman & Harris, 2015). Our findings
indicate that 2 of the 4 DENV strains studied here show greater capacity to replicate in the
mosquito. In some cases, these differences may have consequences for public health, as
some specific variants of DENV have been linked to more severe clinical manifestations
than others (Balmaseda et al., 2006; Halsey et al., 2012; Nisalak et al., 2000; OhAinle et
al., 2011). These findings also have implications for the mosquito’s contribution to the
DENV epidemiological landscape, with the understanding that aspects of human
immunity, including cross-reactivity and non-specific serotype responses also shape the
nature of circulating variants (Reich et al., 2013). Lastly, our work provides better insight
into ideal design of vector competence experiments and the selection of gene candidates
for the engineering of virus refractory mosquitoes. Specifically, our findings support a
greater focus on salivary gland or saliva-based measures of infection and genes that either
completely reduce viral loads in the midgut or that control viral replication in the endpoint
tissue, salivary glands.
2.7 Acknowledgments
The authors would like to thank Emily Kerton, for her help with insect rearing and
sample preparation.

51
2.8 References
Amuzu, H. E., Simmons, C. P., & McGraw, E. A. (2015). Effect of repeat human blood
feeding on Wolbachia density and dengue virus infection in Aedes aegypti. Parasites & Vectors,
8, 246. doi:10.1186/s13071-015-0853-y
Anderson, J. R., & Rico-Hesse, R. (2006). Aedes aegypti vectorial capacity is determined
by the infecting genotype of dengue virus. The American Society of Tropical Medicine and
Hygiene, 75.
Armstrong, P. M., & Rico-Hesse, R. (2003). Efficiency of dengue serotype 2 virus strains
to infect and disseminate in Aedes aegypti. The American Society of Tropical Medicine and
Hygiene, 68.
Azhar, E. I., Hashem, A. M., El-Kafrawy, S. A., Abol-Ela, S., Abd-Alla, A. M., &
Sohrab, S. S. (2015). Complete genome sequencing and phylogenetic analysis of dengue type 1
virus isolated from Jeddah, Saudi Arabia. Virology Journal, 12. doi:10.1186/s12985-014-0235-7
Balmaseda, A., Hammond, S. N., Perez, L., Tellez, Y., Saborio, S. I., & Mercado, J. C.
(2006). Serotype-specific differences in clinical manifestations of dengue. The American Society
of Tropical Medicine and Hygiene, 74.
Bhatt, S., Gething, P. W., Brady, O. J., Messina, J. P., Farlow, A. W., Moyes, C. L.,
Drake, J. M., Brownstein, J. S., Hoen, A. G., Sankoh, O., Myers, M. F., George, D. B., Jaenisch,
T., Wint, G. R. W., Simmons, C. P., Scott, T. W., Farrar, J. J., & Hay, S. I. (2013). The global
distribution and burden of dengue. Nature, 496(7446), 504-507.Doi:10.1038/nature12060
Black Iv, W. C., Bennett, K. E., Gorrochótegui-Escalante, N., Barillas-Mury, C. V.,
Fernández-Salas, I., de Lourdes Muñoz, M. a., Farfán-Alé, J. A., Olson, K. E., & Beaty, B. J.
(2002). Flavivirus susceptibility in Aedes aegypti. Archives of Medical Research, 33(4), 379-388.
doi:http://dx.doi.org/10.1016/S0188-4409(02)00373-9
Brady, O. J., Gething, P. W., Bhatt, S., Messina, J. P., Brownstein, J. S., Hoen, A. G.,
Moyes, C. L., Farlow, A. W., Scott, T. W., & Hay, S. I. (2012). Refining the global spatial limits
of dengue virus transmission by evidence-based consensus. PLoS Neglected Tropical Diseases,
6(8), e1760. doi:10.1371/journal.pntd.0001760
Cao-Lormeau, V.-M. (2009). Dengue viruses binding proteins from Aedes aegypti and
Aedes polynesiensis salivary glands. Virology Journal, 6(1), 35. doi:10.1186/1743-422x-6-35
Cheng, G., Liu, Y., Wang, P., & Xiao, X. (2016). Mosquito defense strategies against
viral infection. Trends in Parasitology, 32(3), 177-186. doi:10.1016/j.pt.2015.09.009
Chotiwan, N., Andre, B. G., Sanchez-Vargas, I., Islam, M. N., Grabowski, J. M., Hopf-
Jannasch, A., Gough, E., Nakayasu, E., Blair, C. D., Belisle, J. T., Hill, C. A., Kuhn, R. J., &
Perera, R. (2018). Dynamic remodeling of lipids coincides with dengue virus replication in the

52
midgut of Aedes aegypti mosquitoes. PLoS Pathogens, 14(2), e1006853.
doi:10.1371/journal.ppat.1006853
Choy, M. M., Sessions, O. M., Gubler, D. J., & Ooi, E. E. (2015). Production of
infectious dengue virus in Aedes aegypti is dependent on the ubiquitin proteasome pathway. PLoS
Neglected Tropical Diseases, 9(11), e0004227-e0004227. doi:10.1371/journal.pntd.0004227
Christofferson, R. C., & Mores, C. N. (2011). Estimating the magnitude and direction of
altered arbovirus transmission due to viral phenotype. PLoS ONE, 6(1), e16298.
doi:10.1371/journal.pone.0016298
Colón-González, F. J., Fezzi, C., Lake, I. R., & Hunter, P. R. (2013). The effects of
weather and climate change on dengue. PLoS Neglected Tropical Diseases, 7(11), e2503.
doi:10.1371/journal.pntd.0002503
Cox, J., Brown, H. E., & Rico-Hesse, R. (2011). Variation in vector competence for
dengue viruses does not depend on mosquito midgut binding affinity. PLoS Neglected Tropical
Diseases, 5(5), e1172. doi:10.1371/journal.pntd.0001172
Deen, J. L., Harris, E., Wills, B., Balmaseda, A., Hammond, S. N., Rocha, C., Dung, N.
M., Hung, N. T., Hien, T. T., & Farrar, J. J. (2006). The WHO dengue classification and case
definitions: time for a reassessment. The Lancet, 368(9530), 170-173. doi:10.1016/S0140-
6736(06)69006-5
Dumre, S. P., Bhandari, R., Shakya, G., Shrestha, S. K., Cherif, M. S., Ghimire, P.,
Klungthong, C., Yoon, I.-K., Hirayama, K., Na-Bangchang, K., & Fernandez, S. (2017). Dengue
virus serotypes 1 and 2 responsible for major dengue outbreaks in Nepal: Clinical, laboratory, and
epidemiological features. The American Society of Tropical Medicine and Hygiene, 97(4), 1062-
1069. doi:10.4269/ajtmh.17-0221
Duneau, D., Ferdy, J. B., Revah, J., Kondolf, H., Ortiz, G. A., Lazzaro, B. P., & Buchon,
N. (2017). Stochastic variation in the initial phase of bacterial infection predicts the probability of
survival in D. melanogaster. eLife, 6. doi:10.7554/eLife.28298
Duong, V., Lambrechts, L., Paul, R. E., Ly, S., Lay, R. S., Long, K. C., Huy, R.,
Tarantola, A., Scott, T. W., Sakuntabhai, A., & Buchy, P. (2015). Asymptomatic humans transmit
dengue virus to mosquitoes. Proceedings of the National Academy, 112(47), 14688-14693.
doi:10.1073/pnas.1508114112
Fontaine, A., Lequime, S., Moltini-Conclois, I., Jiolle, D., Leparc-Goffart, I., Reiner, R.
C., Jr., & Lambrechts, L. (2018). Epidemiological significance of dengue virus genetic variation
in mosquito infection dynamics. PLoS Pathogens, 14(7), e1007187.
doi:10.1371/journal.ppat.1007187
Forrester, N. L., Guerbois, M., Seymour, R. L., Spratt, H., & Weaver, S. C. (2012).
vector-borne transmission imposes a severe bottleneck on an RNA virus population. PLoS
Pathogens, 8(9), e1002897. doi:10.1371/journal.ppat.1002897

53
Franz, A. W. E., Kantor, A. M., Passarelli, A. L., & Clem, R. J. (2015). Tissue Barriers to
Arbovirus infection in mosquitoes. Viruses, 7(7), 3741-3767. doi:10.3390/v7072795
Frentiu, F. D., Robinson, J., Young, P. R., McGraw, E. A., & O'Neill, S. L. (2010).
Wolbachia-mediated resistance to dengue virus infection and death at the cellular level. PLoS
ONE, 5(10), e13398. doi:10.1371/journal.pone.0013398
Gubler, D. J. (2011). Dengue, urbanization and globalization: The unholy trinity of the
21(st) century. Tropical Medicine and Health, 39(4 Suppl), 3-11. doi:10.2149/tmh.2011-S05
Gubler, D. J., Nalim, S., Tan, R., Saipan, H., & Sulianti Saroso, J. (1979). Variation in
susceptibility to oral infection with dengue viruses among geographic strains of Aedes aegypti.
The American Society of Tropical Medicine and Hygiene, 28(6), 1045-1052.
Guzman, M. G., & Harris, E. (2015). Dengue. The Lancet, 385(9966), 453-465.
doi:https://doi.org/10.1016/S0140-6736(14)60572-9
Hall, M. D., Bento, G., & Ebert, D. (2017). The evolutionary consequences of stepwise
infection processes. Trends in Ecology & Evolution, 32(8), 612-623.
doi:10.1016/j.tree.2017.05.009
Halsey, E. S., Marks, M. A., Gotuzzo, E., Fiestas, V., Suarez, L., Vargas, J., Aguayo, N.,
Madrid, C., Vimos, C., Kochel, T. J., & Laguna-Torres, V. A. (2012). Correlation of serotype-
specific dengue virus infection with clinical manifestations. PLoS Neglected Tropical Diseases,
6(5), e1638-e1638. doi:10.1371/journal.pntd.0001638
Helt, A.-M., & Harris, E. (2005). S-phase-dependent enhancement of dengue virus 2
replication in mosquito cells, but not in human cells. Journal of Virology, 79(21), 13218-13230.
doi:10.1128/jvi.79.21.13218-13230.2005
Khoo, C. C., Doty, J. B., Held, N. L., Olson, K. E., & Franz, A. W. (2013). Isolation of
midgut escape mutants of two American genotype dengue 2 viruses from Aedes aegypti. Virology
Journal, 10(1), 1-11. doi:10.1186/1743-422x-10-257
Lambrechts, L., Fansiri, T., Pongsiri, A., Thaisomboonsuk, B., Klungthong, C.,
Richardson, J. H., Ponlawat, A., Jarman, R. G., & Scott, T. W. (2012). Dengue-1 virus clade
replacement in Thailand associated with enhanced mosquito transmission. Journal of Virology,
86(3), 1853-1861. doi:10.1128/jvi.06458-11
Le Coupanec, A., Tchankouo-Nguetcheu, S., Roux, P., Khun, H., Huerre, M., Morales-
Vargas, R., Enguehard, M., Lavillette, D., Misse, D., & Choumet, V. (2017). Co-infection of
mosquitoes with chikungunya and dengue viruses reveals modulation of the replication of both
viruses in midguts and salivary glands of Aedes aegypti mosquitoes. International Journal of
Molecular Sciences, 18(8). doi:10.3390/ijms18081708
Lequime, S., Fontaine, A., Ar Gouilh, M., Moltini-Conclois, I., & Lambrechts, L. (2016).
Genetic drift, purifying selection and vector genotype shape dengue virus intra-host genetic
diversity in mosquitoes. PLoS Genetics, 12(6), e1006111. doi:10.1371/journal.pgen.1006111

54
Liebman, K. A., Stoddard, S. T., Morrison, A. C., Rocha, C., Minnick, S., Sihuincha, M.,
Russell, K. L., Olson, J. G., Blair, P. J., Watts, D. M., Kochel, T., & Scott, T. W. (2012). Spatial
dimensions of dengue virus transmission across interepidemic and epidemic periods in Iquitos,
Peru (1999-2003). PLoS Neglected Tropical Diseases, 6(2), e1472.doi:10.1371/journal.pntd.
0001472
Macdonald, G. (1957). The Epidemiology and Control of Malaria. Amen House,
Warwick Square, London E.C.4.: Oxford University Press.
Murray, N. E., Quam, M. B., & Wilder-Smith, A. (2013). Epidemiology of dengue: past,
present and future prospects. Clinical Epidemiology, 5.
Muturi, E. J., Buckner, E., & Bara, J. (2017). Superinfection interference between
dengue-2 and dengue-4 viruses in Aedes aegypti mosquitoes. Tropical Medicine & International
Health, 22(4), 399-406. doi:10.1111/tmi.12846
Nisalak, A., Raengsakulrach, B., Endy, T. P., Vaughn, D. W., Kalayanarooj, S.,
Nimmannitya, S., Suntayakorn, S., Rothman, A. L., Ennis, F. A., Green, S., & Innis, B. L. (2000).
Dengue Viremia Titer, Antibody Response Pattern, and Virus Serotype Correlate with Disease
Severity. The Journal of Infectious Diseases, 181(1), 2-9. doi:10.1086/315215
OhAinle, M., Balmaseda, A., Macalalad, A. R., Tellez, Y., Zody, M. C., Saborio, S.,
Nunez, A., Lennon, N. J., Birren, B. W., Gordon, A., Henn, M. R., & Harris, E. (2011). Dynamics
of dengue disease severity determined by the interplay between viral genetics and serotype-
specific immunity. Science Translational Medicine, 3(114), 114ra128.
doi:10.1126/scitranslmed.3003084
Perera, R., & Kuhn, R. J. (2008). Structural proteomics of dengue virus. Current opinion
in microbiology, 11(4), 369-377. doi:10.1016/j.mib.2008.06.004
Powell, J. R., & Tabachnick, W. J. (2013). History of domestication and spread of Aedes
aegypti - A review. Memórias do Instituto Oswaldo Cruz, 108(Suppl 1), 11-17.
doi:10.1590/0074-0276130395
Raquin, V., & Lambrechts, L. (2017). Dengue virus replicates and accumulates in Aedes
aegypti salivary glands. Virology, 507, 75-81. doi:https://doi.org/10.1016/j.virol.2017.04.009
Reich, N. G., Shrestha, S., King, A. A., Rohani, P., Lessler, J., Kalayanarooj, S., Yoon,
I.-K., Gibbons, R. V., Burke, D. S., & Cummings, D. A. T. (2013). Interactions between
serotypes of dengue highlight epidemiological impact of cross-immunity. Journal of The Royal
Society Interface, 10(86). doi:10.1098/rsif.2013.0414
Reid, D. W., Campos, R. K., Child, J. R., Zheng, T., Chan, K. W. K., Bradrick, S. S.,
Vasudevan, S. G., Garcia-Blanco, M. A., & Nicchitta, C. V. (2018). Dengue virus selectively
annexes endoplasmic reticulum-associated translation machinery as a strategy for co-opting host
cell protein synthesis. Journal of Virology, 92(7), e01766-01717. doi:10.1128/jvi.01766-17

55
Ritchie, S. A., Pyke, A. T., Hall-Mendelin, S., Day, A., Mores, C. N., Christofferson, R.
C., Gubler, D. J., Bennett, S. N., & van den Hurk, A. F. (2013). An explosive epidemic of DENV-
3 in Cairns, Australia. PLoS ONE, 8(7), e68137. doi:10.1371/journal.pone.0068137
Ritz, C., Baty, F., Streibig, J. C., & Gerhard, D. (2016). Dose-response analysis using R.
PLoS ONE, 10(12), e0146021. doi:10.1371/journal.pone.0146021
Rosen, L., Roseboom, L. E., Gubler, D. J., Lien, J. C., & Chaniotis, B. N. (1985).
Comparative susceptibility of mosquito species and strains to oral and parenteral infection with
dengue and Japanese encephalitis viruses. The American Society of Tropical Medicine and
Hygiene, 34(3), 603-615.
Salazar, M. I., Richardson, J. H., Sánchez-Vargas, I., Olson, K. E., & Beaty, B. J. (2007).
Dengue virus type 2: replication and tropisms in orally infected Aedes aegypti mosquitoes. BMC
Microbiology, 7, 9-9. doi:10.1186/1471-2180-7-9
Sasmono, R. T., Taurel, A. F., Prayitno, A., Sitompul, H., Yohan, B., Hayati, R. F.,
Bouckenooghe, A., Hadinegoro, S. R., & Nealon, J. (2018). Dengue virus serotype distribution
based on serological evidence in pediatric urban population in Indonesia. PLoS Neglected
Tropical Diseases, 12(6), e0006616. doi:10.1371/journal.pntd.0006616
Shrivastava, S., Tiraki, D., Diwan, A., Lalwani, S. K., Modak, M., Mishra, A. C., &
Arankalle, V. A. (2018). Co-circulation of all the four dengue virus serotypes and detection of a
novel clade of DENV-4 (genotype I) virus in Pune, India during 2016 season. PLoS ONE, 13(2),
e0192672. doi:10.1371/journal.pone.0192672
Sim, S., & Dimopoulos, G. (2010). Dengue virus inhibits immune responses in Aedes
aegypti cells. PLoS ONE, 5(5), e10678. doi:10.1371/journal.pone.0010678
Simmons , C. P., Farrar , J. J., van Vinh Chau , N., & Wills , B. (2012). Dengue. New
England Journal of Medicine, 366(15), 1423-1432. doi:doi:10.1056/NEJMra1110265
Smartt, C. T., Shin, D., & Alto, B. W. (2017). Dengue serotype-specific immune
response in Aedes aegypti and Aedes albopictus. Memórias do Instituto Oswaldo Cruz, 112(12),
829-837. doi:10.1590/0074-02760170182
Taracena, M. L., Bottino-Rojas, V., Talyuli, O. A. C., Walter-Nuno, A. B., Oliveira, J. H.
M., Angleró-Rodriguez, Y. I., Wells, M. B., Dimopoulos, G., Oliveira, P. L., & Paiva-Silva, G.
O. (2018). Regulation of midgut cell proliferation impacts Aedes aegypti susceptibility to dengue
virus. PLoS Neglected Tropical Diseases, 12(5), e0006498. doi:10.1371/journal.pntd.0006498
Ty Hang, V. T., Holmes, E. C., Veasna, D., Quy, N. T., Tinh Hien, T., Quail, M.,
Churcher, C., Parkhill, J., Cardosa, J., Farrar, J., Wills, B., Lennon, N. J., Birren, B. W., Buchy,
P., Henn, M. R., & Simmons, C. P. (2010). Emergence of the Asian 1 genotype of dengue virus
serotype 2 in Viet Nam: In vivo fitness advantage and lineage replacement in South-East Asia.
PLoS Neglected Tropical Diseases, 4(7), e757. doi:10.1371/journal.pntd.0000757

56
Ye, Y. H., Carrasco, A. M., Frentiu, F. D., Chenoweth, S. F., Beebe, N. W., van den
Hurk, A. F., Simmons, C. P., O'Neill, S. L., & McGraw, E. A. (2015). Wolbachia reduces the
transmission potential of dengue-infected Aedes aegypti. PLoS Neglected Tropical Diseases, 9(6),
e0003894. doi:10.1371/journal.pntd.0003894
Ye, Y. H., Ng, T. S., Frentiu, F. D., Walker, T., van den Hurk, A. F., O'Neill, S. L.,
Beebe, N. W., & McGraw, E. A. (2014). Comparative susceptibility of mosquito populations in
North Queensland, Australia to oral infection with dengue virus. The American Society of
Tropical Medicine and Hygiene, 90(3), 422-430. doi:10.4269/ajtmh.13-0186
Zhu, Y., Zhang, R., Zhang, B., Zhao, T., Wang, P., Liang, G., & Cheng, G. (2017). Blood
meal acquisition enhances arbovirus replication in mosquitoes through activation of the
GABAergic system. Nature Communications, 8(1), 1262. doi:10.1038/s41467-017-01244-6
Zieler, H., Garon, C. F., Fischer, E. R., & Shahabuddin, M. (2000). A tubular network
associated with the brush-border surface of the Aedes aegypti midgut: implications for pathogen
transmission by mosquitoes. Journal of Experimental Biology, 203(Pt 10), 1599-1611.

57
Chapter 3
The effects of DENV serotype competition and co-infection on viral kinetics
in Wolbachia-infected and uninfected Aedes aegypti mosquitoes
Citation: Novelo M, Audsley M, McGraw EA. The impact of DENV serotype competition
and co-infection viral kinetics in Wolbachia-infected and uninfected Aedes aegypti
mosquitoes. Parasites & Vectors. 2021. In press.
3.1 Abstract
Background: The Ae. aegypti mosquito is responsible for the transmission of
several medically important arthropod-borne viruses, including multiple serotypes of
dengue virus (DENV-1, -2, -3, and -4). Competition within the mosquito between DENV
serotypes can affect viral infection dynamics, modulating the transmission potential of the
pathogen. Vector control remains the main method for limiting dengue fever. The insect
endosymbiont Wolbachia pipientis is currently being trialed in field releases globally as a
means of biological control because it reduces virus replication inside the mosquito. It is
not clear how co-infection between DENV serotypes in the same mosquito might alter the
pathogen-blocking phenotype elicited by Wolbachia in Ae. aegypti.
Methods: Five- to 7-day-old female Ae. aegypti from two lines, namely, with
(wMel) and without Wolbachia infection (WT), were fed virus-laden blood through an
artificial membrane with either a mix of DENV-2 and -3 or the same DENV serotypes

58
singly. Mosquitoes were subsequently incubated inside environmental chambers and
collected on the following days post-infection: 3, 4, 5, 7, 8, 9, 11, 12, and 13. Midgut,
carcass, and salivary glands were collected from each mosquito at each timepoint and
individually analyzed to determine the percentage of DENV infection and viral RNA load
via RT-qPCR.
Results: We saw that for WT mosquitoes DENV-3 grew to higher viral infection
rates across multiple tissues when co-infected with DENV-2 than when it was in a mono-
infection. Additionally, we saw a strong pathogen-blocking phenotype in wMel mosquitoes
independent of co-infection status.
Conclusion: In this study, we demonstrated that the wMel mosquito line is capable
of blocking DENV serotype co-infection in a systemic way across the mosquito body.
Moreover, we showed that for WT mosquitoes, serotype co-infection can affect infection
frequency in a tissue- and time-specific manner and that both viruses have the potential of
being transmitted simultaneously. Our findings suggest that the long-term efficacy of
Wolbachia pathogen blocking is not compromised by arthropod-borne virus co-infection.
Key words: dengue; co-infection, Aedes aegypti; serotype; Wolbachia; infection
dynamics

59
3.2 Introduction
Dengue viruses (DENVs) are medically important arthropod-borne viruses
(arboviruses) responsible for up to 300 million cases of dengue fever a year, and it can be
caused by any of the four related but antigenically distinct DENV serotypes (DENV-1 to -
4) (Murray, Quam, & Wilder-Smith, 2013). In regions with endemic transmission of all
four serotypes of DENV, varying predominance of certain serotypes has been observed
between seasons (Vasilakis & Weaver, 2008). Differences in the DENV infection kinetics
and transmission potential are influenced by the genetic diversity of the four different
DENV serotypes. The overall genome sequence-level differences between serotypes are
estimated at 20-30% (Holmes & Twiddy, 2003; Rico-Hesse, 2003).
Substantial evidence indicates that variation in the DENV genome of serotypes and
strains can have epidemiological significance by altering the extrinsic incubation period
(EIP) (Fontaine et al., 2018; Ko, Salem, Chang, & Chao, 2020; L. Lambrechts et al., 2009),
defined as the time it takes for the pathogen to be transmitted by the vector (Macdonald,
1957), and therefore has a powerful effect on the scale and speed of epidemics. DENV-2
strains from the American and Southeast Asian genotypes differ in their EIP lengths, with
the Southeast Asian genotypes having shorter EIPs. This shorter EIP was thought, in part,
to explain the displacement of the American DENV strains in South America by the Asian
lineage (Anderson & Rico-Hesse, 2006). Additionally, different DENV serotypes exhibit
various degrees of infectivity across the same mosquito populations (C. V. F. Carrington,
Foster, Pybus, Bennett, & Holmes, 2005; Ekwudu et al., 2020; Gubler, Nalim, Tan, Saipan,
& Sulianti Saroso, 1979; Novelo et al., 2019; Weaver & Vasilakis, 2009). Moreover, oral

60
susceptibility to DENV-1 was shown to be up to four times higher than that of DENV-3 in
Ae. aegypti from Senegal, with DENV-1 having higher infection and dissemination rates
(Gaye et al., 2014). Finally, systematic analyses of DENV replication kinetics of all four
DENV serotypes found significant differences in the infection rate and EIP between
serotypes (Ekwudu et al., 2020; Novelo et al., 2019).
Competition between DENV strains and serotypes can also affect viral population
dynamics within the vector, thus modulating the transmission potential (Louis Lambrechts
et al., 2012; Vogels et al., 2019). This happens in nature when mosquitoes take multiple
blood meals from several different hosts that are each infected with a different or multiple
DENV serotypes (Shrivastava et al., 2018). In controlled laboratory experiments, using
field-derived mosquito populations, there were no differences in dissemination and
transmission rates between DENV-1 and DENV-4 mono-infections in Ae. aegypti, but
during co-infection, DENV-4 had a much higher dissemination rate, leading to the
exclusive presence of DENV-4 in the saliva for this particular experimental mosquito
population (M. Vazeille, Gaborit, Mousson, Girod, & Failloux, 2016). Furthermore,
differential replication between DENV-2 and DENV-3 has been shown, with DENV-2
exhibiting a much higher replication efficiency both in vitro and in vivo during co-infection
(Quintero-Gil, Ospina, Osorio-Benitez, & Martinez-Gutierrez, 2014). Additionally, the
effect of co-infection with different families of arboviruses on vector competence has just
been recently studied. For example, mosquitoes exposed to double or triple infections with
chikungunya virus (CHIKV; Togaviridae), Zika virus (ZIKV; Flaviviridae), and DENV
(Flaviviridae) were capable of transmitting all pathogens concurrently, without noticeable

61
changes to mosquito infection and dissemination rates (Rückert et al., 2017). Co-infection
studies may shed light on the outcome of competitive processes in field mosquitoes, even
if rare, but also importantly allow for direct comparisons of the transmissibility between
viruses.
Given the ease of rearing Ae. aegypti in the laboratory, vector competence
experiments are an important tool with which to study the effect and interaction between
DENV and mosquito genotypes in the transmission potential of the virus. However, one of
the main issues with artificial vector competence experiments is that there is too much
heterogeneity between experiments that results from environmental variation and its
interaction with genetic variation in both the mosquitoes and viruses (Souza-Neto, Powell,
& Bonizzoni, 2019). Individual vector competence experiments using single DENV
serotypes or strains often give varying results in both infection and transmission rates,
making pairwise comparisons difficult to interpret. Moreover, limited data are available on
co-infections with different serotypes, with some experiments suggesting competitive
disadvantage or superinfection interference between DENV serotypes (Muturi, Buckner,
& Bara, 2017). Additionally, it is not clear how these viral dynamics and interactions may
be altered in the presence of Wolbachia infection, which is currently being trialed in global
releases as a means of reducing virus transmission to humans (Dorigatti, McCormack,
Nedjati-Gilani, & Ferguson, 2018) and that is known to reduce viral replication in serotype-
specific ways (Bell, Katzelnick, & Bedford, 2019; L. B. Carrington et al., 2018; Ferguson
et al., 2015). To the best of our knowledge, only one study has looked at the effects of

62
arboviral co-infection in Wolbachia-infected mosquitoes using ZIKV and DENV (Dutra et
al., 2016).
To assess the effect of DENV serotype co-infection on transmissibility and any
corresponding interactions with Wolbachia infection, we used two interdependent
approaches. First, we challenged two Ae. aegypti mosquito lines that were either
Wolbachia infected (wMel) or Wolbachia uninfected (WT) in both mono- and co-infection
vector competence experiments with DENV-2 and DENV-3. We collected midgut,
carcass, and salivary glands at nine timepoints post-infection. We then used the infection
rate and viral RNA load data to assess the effects of competing serotypes and evaluate their
performance in individual mosquitoes and between wMel and WT lines. Second, we
determined whether serotype co-infection altered viral infection dynamics relative to the
mono-infected state by comparing viral RNA load and infection rate between the two
vector competence experiments.
3.3 Methods
3.3.1 Mosquito lines and rearing
The mosquito lines used for this work have been described previously (Terradas,
Allen, Chenoweth, & McGraw, 2017; Walker et al., 2011). The wMel line was collected
from the Wolbachia release zone in Cairns, Australia, as part of the Eliminate Dengue
Program, whereas the WT line, naturally free from Wolbachia, was collected outside the
Wolbachia release zone. Both lines were identified morphologically and with genetic
markers as well as screened for the presence/absence of pathogens before being used in

63
our study (31). WT and wMel larvae were fed Tetramin® fish food (Melle, Germany), and
adults were maintained on 10% sucrose. All mosquitoes were reared in a controlled
environment at 26°C, 75% relative humidity, and a 12-hr light/dark cycle.
3.3.2 Virus culture and titration
The DENV serotypes/strains used for this experiment are listed in Table 3.1. The
virus was propagated in cell culture, as described previously (Frentiu, Robinson, Young,
McGraw, & O'Neill, 2010). Briefly, Ae. albopictus C6/36 cells were grown at 26°C in
RPMI 1640 medium (Invitrogen, Carlsbad, CA) supplemented with 10% fetal bovine
serum (FBS), 1× Glutamax (Invitrogen), and HEPES buffer. Cells were first allowed to
form monolayers of around 60–80% confluence in T175 flasks (Sigma Aldrich, St. Louis,
MO), and then, they were inoculated with DENV and maintained in RPMI medium
supplemented with 2% FBS. After 7 days post-inoculation, live virus was harvested,
titrated via absolute quantification PCR (qPCR) and plaque forming unit assay (as per
below), and adjusted to a final concentration of ~4 × 105 plaque-forming units (PFU)/ml
for both serotypes (Table 3.2) prior to mixing with blood. Live virus was used for all vector
competence experiments.
Prior to the above steps, we isolated the viruses at different timepoints from C6/36
cells and assessed their viral RNA loads by qPCR and plaque assay to select the most
appropriate day to harvest virus for the vector competence experiments. These pilot
experiments also revealed the relationship between viral RNA load estimates by qPCR and
live virus estimates by plaque assay. In general, we saw that higher viral RNA loads

64
correlated with higher plaque assay titers (Appendix B) with viral RNA loads ranging from
~10- to 2000-fold higher than live virus titers, as expected (Choy, Ellis, Ellis, & Gubler,
2013; Richardson, Molina-Cruz, Salazar, & Black, 2006). The average ratio of viral RNA
copies relative to PFU/ml was larger for DENV-3 (1300-fold) than for DENV-2 (270-fold),
indicating that RT-qPCR was more sensitive for DENV-3. This effect was consistent
across the two tested collection timepoints.
3.3.3 Mosquito infections
The methods for mosquito oral infections have been described previously (Novelo et al.,
2019; Terradas et al., 2017). Briefly, prior to oral DENV infections, 6- to 7-day-old adult
female mosquitoes were sorted and starved for 24 hrs. For mono-infections, a 1:1 mix of
Table 3.1 Dengue serotypes and strains use
Serotype Strain Passage Accession number Place of origin Collection
date
DENV-2 ET-300 11 EF440433.1 East Timor 2000
DENV-3 Cairns 2008 9 JN406515.1 Australia 2008
Table 3.2. Viral titers
Serotype
Undiluted RT-
qPCR titer of
infected
supernatant
(copies/ml)
Final plaque
assay titer for
the blood meal
(PFU/ml)
Fold change
difference
between RNA
copies/mL and
PFU/mL
Average ratio of
viral RNA
copies/ml relative
to PFU/ml
DENV-2 4 × 107 ~3 × 105 146 277.1
DENV-3 8 × 108 ~4 × 105 2046 1382.1

65
virus culture and defibrinated sheep blood was prepared. For co-infections, 1 ml of each
DENV serotype was combined, and from that blend, 1 ml was combined with 1 ml of
defibrinated sheep blood. Glass feeders with double chambers were covered with pig
intestine, and water heated to 37°C was circulated in the outer chamber of the feeders. The
mosquitoes were allowed to feed for ~2 hrs. Immediately after blood feeding, mosquitoes
were knocked down and sorted on ice. Unfed females were discarded. The remaining
mosquitoes were returned to 70-ml plastic cups and maintained on 10% sucrose. At days
post-infection (DPI) 3, 4, 5, 7, 8, 9, 11, 12, and 13, mosquitoes were anesthetized and
dissected in sterile phosphate-buffered saline (PBS). We collected midgut, carcass, and
salivary glands from individual mosquitoes. For our vector competence experiments, the
mosquito carcass was the collection of tissues that remained after dissecting the midgut
and salivary glands. We used the carcass as a proxy for viral dissemination from the
mosquito midgut. All tissue collections were conducted on live mosquitoes. Individual
tissues were collected in 1.5-ml microcentrifuge tubes (Sarstedt, Nümbrecht, Germany)
containing 200 µl of Trizol reagent (Invitrogen) and 2-mm glass beads. Samples were
homogenized and frozen at −80°C until RNA extraction.
3.3.4 DENV absolute quantification via RT-qPCR
The RT-qPCR mixture contained 2.5 μl of LC480 master mix (Roche); 4.25 μl of
PCR-grade water; 0.25 μl of 10 μM forward (F) primer, reverse (R) primer, and probe (P)
specific to each DENV serotype; and 2 μl of RNA. Reactions were run in the LightCycler
480 instrument (Roche), and the thermal cycling conditions were 95°C for 3 minutes, 30
cycles of 95°C for 30 seconds, 50°C for 1 min, and 68°C for 1 min, finalizing with 68°C

66
for 5 min. Standard curves were generated by triplicate on each plate by analyzing 100 to
107 copies/reaction of DENV fragment copies with a limit of detection of set at 100 copies.
DENV genome copies were extrapolated from the standard curve as DENV copies per
tissue. The standards for both serotypes were generated using a ~100-bp conserved region
of the NS5 protein of the DENV genome. The detection threshold for both DENV-2 and
DENV-3 was set at ~35 Ct. The primers and probes used for the detection of specific
DENV serotypes were as follows:
DENV-2-ET300: F primer (primer 9873681; TCCATACACGCCACACATGAG),
R primer (primer 98736818; GGGATTTCCTCCCATGATTC) and Probe-FAM (probe
98084286; 56-fam/AGGGTGTGGATTCGAGAAAACCCATGG/3BHQ_1).
DENV-3-Cairns08/09: F primer (primer 98644632; TTTCTG CTCCCAC CAC
TTTC), R primer (primer 98451787; CCATCCYGCTCCCTGAGA) and Probe-LC640
(probe9845178; 5LtC640N /AAGAA AGTTGGT AGTCCCC TGCAGACC TCA/3IAb R
QSp).
3.3.5 Statistical analysis
All statistical analysis used R v3.6.0 (http://www.r-project.org/). For the analysis
of the infection frequency, a one-way ANOVA was fitted, and Tukey for contrasts was
used for post hoc comparisons. Viral RNA load analysis was carried out using Kruskal-
Wallis by rank test for the non-parametric data. All DENV RNA loads were reported on a
log10 scale given the value range.

67
3.4 Results
In this study, we challenged both WT and wMel Ae. aegypti mosquitoes with two
serotypes in the co- and mono-infected states. Specifically, we investigated (1) the relative
infection dynamics of two DENV serotypes inside the mosquito by competing them
directly and (2) how individual serotypes behave when they are in a co-infection relative
to mono-infection. Mosquitoes were blood fed with DENV-2 and DENV-3, both separately
and together. In the case of mono-infections, a 1:1 mix of each DENV serotype and blood
was used. For co-infections, the two serotypes were first combined, and from that blend, a
1:1 mix of blood and the two serotypes was used. Overall, the viral titers for both
experiments were equivalent. We used RT-qPCR with serotype-specific TaqMan probes
to quantify DENV RNA load in midgut, carcass, and salivary gland tissues to assess
dissemination and infectiousness at nine timepoints post-infection.
3.4.1 DENV serotype competition in co-infection
By competing the serotypes in same mosquitoes, we were able to powerfully
compare their performance, controlling some of the substantial variation that can occur
across vector competence experiments. Infection rates in the WT mosquito line indicate
that DENV-3 was a better competitor than DENV-2 but that the magnitude of this
difference changed depending on the tissue and DPI. In the co-infection experiments, we
classified the infection rates as uninfected (no viral RNA load detectable for either
serotype), only DENV-2 infected (DENV-2/alone), only DENV-3 infected (DENV-
3/alone), or co-infected (both serotypes detected). We saw significant variation in the

68
percentage of infected WT mosquitoes between serotypes (df=1, F=22.2, P < 0.0001). In
the midgut, most mosquitoes were co-infected at all DPIs (infection rate: 25% to 100%).
Mosquitoes were DENV-3/alone at 6 timepoints with an infection rate between 10 and
50% and DENV-2/alone at DPI 4 and 9 with an infection rate of ~15% (Figure 3.1, midgut).
In the carcass, most WT mosquitoes were either co-infected or DENV-3/alone, but
we saw no DENV-2/alone mosquitoes at any timepoint (Figure 3.1, carcass). For the
salivary glands, most WT mosquitoes were similarly either co-infected or DENV-3/alone,
and there was only one timepoint at which we observed DENV-2/alone mosquitoes, with
an infection frequency of ~10% at DPI 3 (Figure 3.1, salivary glands). For wMel
mosquitoes, no pairwise comparison between serotypes was possible, as there were too few
infected mosquitoes to perform statistical analysis, due to the action of Wolbachia-
mediated pathogen blocking. Overall, our results indicate that DENV-3 is better at
replicating and disseminating in mosquitoes, regardless of tissue or timepoint, than DENV-
2.

69
We also compared viral RNA loads between serotypes across tissues and DPI.
Overall, we saw no significant differences between DENV-2 and DENV-3 (Figure 3.2, Chi
square = 1.66, P = 0.19, df =1). The viral RNA load for both serotypes in the WT
mosquitoes ranged from ~103 to ~108 in the midgut, ~102 to ~107 in the carcass, and ~102
to ~106 log10 DENV copies per tissue in the salivary glands. In contrast, the viral RNA
load in the wMel mosquitoes was diminished and only observed in 10 timepoints across all
Figure 3.1. Mosquito susceptibility to DENV in co-infections by DPI, tissue, and line. Ae.
aegypti were orally challenged with DENV-2 (orange) and DENV-3 (navy) simultaneously,
whereby mosquitoes were fed a blood meal containing both viruses at ~3 × 105 DENV genome
copies/ml. Mosquito tissues (midgut, carcass, and salivary glands) were collected at nine
timepoints post-infection. DENV RNA load was determined via RT-qPCR using serotype-specific
probes. Each bar represents the proportion of mosquitoes positive for either (orange or navy) or
both (blue) serotypes for each day post-infection. Total number of mosquitoes screened per day
was n = 7 for the wild type line and n = 30 for wMel.

70
tissues, ranging from ~102 to ~105 in the midgut, ~102 to ~108 in the carcass, and ~102
log10 DENV copies per tissue in the salivary glands. Additionally, in the wMel
mosquitoes, viral RNA loads from both serotypes were observed at only one timepoint
(Figure 3.1, wMel, carcass, DPI 5).
Figure 3.2. DENV load in co-infection by DPI, tissue, and mosquito line. Ae. aegypti were
orally challenged with DENV-2 (orange circles) and DENV-3 (navy diamonds) simultaneously,
and both viruses were fed to mosquitoes at ~3 × 105 DENV genome copies/ml. Mosquito tissues
(midgut, carcass, and salivary glands) were collected at nine days post-infection, and DENV
RNA load was determined via RT-qPCR using serotype-specific probes. Mosquitoes with
undetectable viral RNA load are not represented in this graph. Black bars represent treatment
medians. Each symbol represents a single mosquito sample. Total number of mosquitoes
screened per day was n=7 for the wild type line and n=30 for wMel.
Viral RNA loads from both serotypes were expressed as DENV-2/DENV-3 ratio
(Figure 3.3). Although for all tissues there was a general trend of co-infected samples

71
having higher levels of DENV-3 than DENV-2, they were not significantly different
from one another. For the wMel mosquitoes, only DPI 5 had sufficient data with which
to calculate a ratio due to the action of Wolbachia pathogen blocking; in this case,
DENV-3 appeared higher than DENV-2, but with so few data points, statistical
comparisons were not possible.
Figure 3.3. Co-infection serotype ratio by DPI and tissue. Graphs depict the log10 DENV-
2/DENV-3 ratio for all wild type (grey) and wMel (red) samples found to be positive for infection
with both viruses via RT-qPCR for each tissue. Ratios greater than zero indicate that DENV-2
levels were higher than DENV-3 levels for that sample. Color lines represent mean DENV-
2/DENV-3 ratio and the standard error estimate of the mean ratio.

72
3.4.2 Co-infection alters infection dynamics of DENV serotypes in wild type
mosquitoes
After examining the competition dynamics between DENV-2 and DENV-3, we
then sought to determine whether or not co-infection altered viral infection dynamics
relative to the mono-infected state. For the wMel mosquitoes, no pairwise comparisons
between mono- and co-infection for each serotype were possible due to the action of
Wolbachia-mediated viral blocking. In the WT line, we saw higher infection rates when
DENV-3 was in co-infection relative to mono-infection (Figure 3.4) but that the magnitude
of this difference varied by DPI and tissue (df=1, F=7.5, P < 0.005). In the salivary glands,
infection was only observed at four timepoints in the mono-infected state, whereas it was
present in eight out of the nine timepoints in the co-infected mosquitoes (Figure 3.4). For
viral RNA load, we saw no significant difference between DENV-3 mono- and DENV-3
co-infection (Figure 3.5, Chi square = 1.54, P = 0.21, df =1). For DENV-2, there were no
differences in either infection frequency (Figure 3.4, df= 1, F=0.97, P=0.33) or viral RNA
load (Figure 3.5, Chi square = 0.01, P = 0.91, df =1) between mono- and co-infection,
indicating that co-infection did not alter dynamics.
3.4.3 Tissue specific differences in viral dynamics
Overall, for DENV-2, the infection rates were highest in the midgut and declined
as the virus moved into the carcass and salivary glands (Figure 3.4). DENV-3, in contrast,
demonstrated the highest infection rates in the midgut, followed by the salivary glands
(Figure 3.4). Viral RNA loads for the two serotypes were highest in the midgut and

73
decreased in the carcass and then the salivary glands (Figure 3.5, DENV-2; Chi square =
57.48, P < 0.001, df =2; DENV-3; Chi square = 57.48, P < 0.001, df =2). The carcass was
the tissue most likely to have either serotype present despite the action of wMel-mediated
pathogen blocking. Dissemination rates of each of the two serotypes in the presence of
wMel were similar across all three tissues (Figure 3.1).
Figure 3.4. WT line susceptibility to DENV in co- and mono-infections by DPI, tissue, and
serotype. Ae. aegypti mosquitoes were orally challenged with DENV-2 and DENV-3 in both
mono- and co-infection experiments. Each bar represents the proportion of mosquitoes positive
for either serotype in the mono- (grey) and co-infection (blue) experiments for each day post-
infection. Mosquito tissues (midgut, carcass, and salivary glands) were collected at nine days
post-infection, and DENV RNA load was determined via RT-qPCR using serotype-specific
probes. Sample size n=7 per day.

74
3.5 Discussion
We sought to explore the role of DENV serotype co-infection and competition in
both Wolbachia-mediated blocking and within-vector infection dynamics. Particularly, we
examined DENV RNA loads and infection rates in DENV-2/DENV-3 co-infected Ae.
aegypti midguts, carcasses, and salivary glands of two mosquito lines with (wMel) and
without Wolbachia infection (WT). We identified that in WT mosquitoes, there was a
Figure 3.5. WT line DENV viral RNA load in co- and mono-infection by DPI, tissue, and
serotype. Ae. aegypti mosquitoes were orally challenged with DENV-2 (orange) and DENV-3
(navy) in both co-infection (square) and mono-infection (triangle) experiments, with viruses
offered at equivalent titers in both cases. Mosquito tissues (midgut, carcass, and salivary glands)
were collected at nine days post-infection, and DENV RNA load was determined via RT-qPCR
using serotype-specific probes. Mosquitoes with undetectable viral RNA load are not represented
in this graph. Black bars represent treatment medians. Each symbol represents a single mosquito
sample. Data were log transformed. Sample size n=7.

75
competitive advantage for DENV-3 when co-infected with DENV-2 across multiple tissues
compared to a mono-infection. Additionally, we saw a strong pathogen-blocking
phenotype in wMel mosquitoes independent of co-infection status, tissue, and DPI.
Our data showed that for the WT mosquitoes, DENV serotype co-infection altered
the infection frequency of each serotype in a tissue-specific manner, with DENV-3 having
a competitive advantage over DENV-2. This advantage was clearer at the dissemination
stage of infection once the virus reached the hemocoel from the midgut and ultimately at
the transmission level when the virus arrived at the salivary glands. Moreover, the DENV-
3 competitive advantage was confirmed when we compared each serotype in co-infected
mosquitoes vs mono-infected mosquitoes, and we identified that DENV-3 produced higher
infection rates in all tissues when mosquitoes were co-infected. Conversely, DENV-2
infection dynamics did not change significantly when mosquitoes were co-infected
compared to mono-infected. Only one other study has looked at the infection dynamics of
co-infection with DENV-2 and -3 both in vitro and in vivo (Quintero-Gil et al., 2014);
contrary to our findings, they showed an increase in replication efficiency of 1000-fold for
DENV-2. These contradictory results may be due to the use of different DENV and
mosquito genotypes.
When the replication capacities of the two DENV serotypes were assessed in WT
mosquitoes, no significant differences in viral RNA load between DENV-2 and DENV-3
in either mono- and co-infection were found. Differences in viral replication rates without
affecting the viral RNA load in each tissue have been previously reported (Novelo et al.,
2019), in which DENV serotypes that had high growth rates did not necessarily achieve

76
high viral RNA loads or high infection frequencies in experimental mosquito populations.
The disconnect between viral RNA load and infection frequency for each virus suggests
that stochastic processes are potentially taking place or/and genotype-by-genotype
interactions are affecting the virus infection dynamics. For example, previous research has
shown that the strength of the mosquito immune response can be tissue and serotype
dependent (Cheng, Liu, Wang, & Xiao, 2016; Smartt, Shin, & Alto, 2017; Taracena et al.,
2018) and could lead to scenarios in which mosquitoes are more susceptible to dengue
infection with a particular serotype (i.e., higher infection frequency) but can also mount a
relatively strong immune response in particular tissues (i.e., low viral RNA load).
Arboviral co-infection is not limited to DENV serotypes; although most reported
cases are with DENV, co-circulation of DENV with CHIKV and/or ZIKV is increasing
around the globe (Bisanzio et al., 2018; Rodriguez-Morales, Villamil-Gómez, & Franco-
Paredes, 2016). This co-circulation represents a major challenge for many national and
international public health organizations, particularly because there is little information
about the potential clinical and biological consequences of these interactions. Individual
case reports of arboviral co-infection in humans suggest enhanced disease severity. Co-
infection with ZIKV and CHIKV has been associated with severe meningoencephalitis in
a male patient, and co-infection with DENV and CHIKV was linked to severe metrorrhagia
in a female patient (Brito, Azevedo, Cordeiro, Marques, & Franca, 2017; Schilling,
Emmerich, Günther, & Schmidt-Chanasit, 2009). Increased disease severity may occur
when both viruses interfere with different parts of the same immune pathways. For
example, interferon signaling is a major part of the human antiviral response, and it is

77
mediated by the signal transducer and activator of transcription one and two (STAT-1 and
STAT-2) (Murira & Lamarre, 2016). DENV has been shown to block STAT-1, and
CHIKV can potentially interfere with STAT-2 (Hollidge, Weiss, & Soldan, 2011),
therefore blocking the activation cascade of interferon and potentially increasing disease
severity.
The wMel mosquitoes challenged in DENV serotype co-infection were far less
susceptible than the WT line, indicating that the pathogen-blocking phenotype caused by
Wolbachia infection is not affected by concomitant DENV serotypes. Additionally, the
effect of Wolbachia blocking was seen in all three tissues and was stable across nine
timepoints, from 3 to 13 DPI, encompassing days of the mosquito’s lifespan relevant to
viral transmission in the field. Pathogen blocking by Wolbachia-infected mosquitoes
challenged with co-infecting arboviruses has only been shown once before (Caragata et al.,
2019). Co-infection was performed using DENV/ZIKV challenges but not with multiple
DENV serotypes at the same time, and it was limited to three timepoints and only one
mosquito tissue. Although co-circulation of novel emerging arboviruses like ZIKV or
CHIKV coupled with DENV has been reported (Carrillo-Hernández, Ruiz-Saenz,
Villamizar, Gómez-Rangel, & Martínez-Gutierrez, 2018; dos Santos S. Marinho et al.,
2020; Villamil-Gómez et al., 2016), most countries where DENV is endemic have reported
co-circulation of all 4 DENV serotypes, resulting in hyperendemicity for the virus (Bastos
Mde et al., 2012; Villabona-Arenas et al., 2014). Specifically, for DENV-2 and DENV-3
infections, one study showed that from 303 human serum samples, up to 21% were infected
with both viruses (Senaratne, Murugananthan, Sirisena, Carr, & Noordeen, 2020).This

78
phenomenon has been linked to an increased frequency of severe dengue cases and an
overall increase of virulence (Guzman & Harris, 2015).
Another effect of co-transmission can be epidemiological, where co-infection
occurring from a single biting event can significantly increase disease burden.
Mathematical modeling using in silico data in Ae. aegypti has shown that co-transmission
events can potentially lead to an increased number of cases for both viruses (Vogels et al.,
2019). An additional unanswered question in co-infection in Ae. aegypti is how sequential
viral infections might affect pathogen transmission dynamics, a scenario that can also occur
in nature (Nuckols et al., 2015; Marie Vazeille, Mousson, Martin, & Failloux, 2010).
Whether or not viruses can potentially interact after sequential acquisition by the mosquito
has yet to be determined. Last, in our co-infection experiments, we used half as much of
each virus but the same overall viral RNA load as mono-infections; however, no
statistically significant differences were observed in viral RNA load between mono-
infection and co-infection for either serotype. This still begs the question of the potential
outcome of infecting the mosquitoes with twice the amount of each serotype and twice the
overall amount of virus in co-infection.
3.6 Conclusion
Here, we present the first examination of DENV serotype co-infection and its effect
on Wolbachia-mediated pathogen blocking. We demonstrated that the wMel mosquito
strain is capable of blocking DENV serotype co-infection in a systemic way across the
mosquito body. Moreover, we showed that for WT mosquitoes, serotype co-infection can

79
affect infection frequency in a tissue- and time-specific manner and that both viruses have
the potential of being transmitted simultaneously.
3.7 Acknowledgements
We would like to thank E. Kerton, H. Amuzu, G. Terradas, C. Koh, L. Jimenez, S,
Ford, and C. Hammer for technical support throughout the experiment. Additionally, we
would like to thank Ana Ramírez for sharing and letting us use Ae. aegypti illustrations.

80
3.8 References
Anderson, J. R., & Rico-Hesse, R. (2006). Aedes aegypti vectorial capacity is determined
by the infecting genotype of dengue virus. The American Society of Tropical Medicine and
Hygiene, 75.
Bastos Mde, S., Figueiredo, R. M., Ramasawmy, R., Itapirema, E., Gimaque, J. B.,
Santos, L. O., Figueiredo, L. T., & Mourão, M. P. (2012). Simultaneous circulation of all four
dengue serotypes in Manaus, State of Amazonas, Brazil in 2011. Revista da Sociedade Brasileira
de Medicina Tropical, 45(3), 393-394. doi:10.1590/s0037-86822012000300022
Bell, S. M., Katzelnick, L., & Bedford, T. (2019). Dengue genetic divergence generates
within-serotype antigenic variation, but serotypes dominate evolutionary dynamics. eLife, 8,
e42496. doi:10.7554/eLife.42496
Bisanzio, D., Dzul-Manzanilla, F., Gomez-Dantés, H., Pavia-Ruz, N., Hladish, T. J.,
Lenhart, A., Palacio-Vargas, J., González Roldan, J. F., Correa-Morales, F., Sánchez-Tejeda, G.,
Kuri Morales, P., Manrique-Saide, P., Longini, I. M., Halloran, M. E., & Vazquez-Prokopec, G.
M. (2018). Spatio-temporal coherence of dengue, chikungunya and Zika outbreaks in Merida,
Mexico. PLoS Neglected Tropical Diseases, 12(3), e0006298. doi:10.1371/journal.pntd.0006298
Brito, C. A. A., Azevedo, F., Cordeiro, M. T., Marques, E. T. A., Jr., & Franca, R. F. O.
(2017). Central and peripheral nervous system involvement caused by Zika and chikungunya
coinfection. PLoS Neglected Tropical Diseases, 11(7), e0005583.
doi:10.1371/journal.pntd.0005583
Caragata, E. P., Rocha, M. N., Pereira, T. N., Mansur, S. B., Dutra, H. L. C., & Moreira,
L. A. (2019). Pathogen blocking in Wolbachia-infected Aedes aegypti is not affected by Zika and
dengue virus co-infection. PLoS Neglected Tropical Diseases, 13(5), e0007443-e0007443.
doi:10.1371/journal.pntd.0007443
Carrillo-Hernández, M. Y., Ruiz-Saenz, J., Villamizar, L. J., Gómez-Rangel, S. Y., &
Martínez-Gutierrez, M. (2018). Co-circulation and simultaneous co-infection of dengue,
chikungunya, and Zika viruses in patients with febrile syndrome at the Colombian-Venezuelan
border. BMC Infectious Diseases, 18(1), 61. doi:10.1186/s12879-018-2976-1
Carrington, C. V. F., Foster, J. E., Pybus, O. G., Bennett, S. N., & Holmes, E. C. (2005).
Invasion and maintenance of dengue virus type 2 and type 4 in the Americas. Journal of Virology,
79(23), 14680. doi:10.1128/JVI.79.23.14680-14687.2005
Carrington, L. B., Tran, B. C. N., Le, N. T. H., Luong, T. T. H., Nguyen, T. T., Nguyen,
P. T., Nguyen, C. V. V., Nguyen, H. T. C., Vu, T. T., Vo, L. T., Le, D. T., Vu, N. T., Nguyen, G.
T., Luu, H. Q., Dang, A. D., Hurst, T. P., O’Neill, S. L., Tran, V. T., Kien, D. T. H., Nguyen, N.
M., Wolbers, M., Wills, B., & Simmons, C. P. (2018). Field and clinically derived estimates of
Wolbachia mediated blocking of dengue virus transmission potential in Aedes aegypti
mosquitoes. Proceeds of the National Academy of Science, 115(2), 361.
doi:10.1073/pnas.1715788115

81
Cheng, G., Liu, Y., Wang, P., & Xiao, X. (2016). Mosquito defense strategies against
viral infection. Trends in Parasitology, 32(3), 177-186. doi:10.1016/j.pt.2015.09.009
Choy, M. M., Ellis, B. R., Ellis, E. M., & Gubler, D. J. (2013). Comparison of the
mosquito inoculation technique and quantitative real time polymerase chain reaction to measure
dengue virus concentration. The American Society of Tropical Medicine and Hygiene, 89(5),
1001-1005. doi:10.4269/ajtmh.13-0100
Dorigatti, I., McCormack, C., Nedjati-Gilani, G., & Ferguson, N. M. (2018). Using
Wolbachia for dengue control: insights from modelling. Trends in Parasitology, 34(2), 102-113.
doi:10.1016/j.pt.2017.11.002
dos Santos S. Marinho, R., Sanz Duro, R. L., Santos, G. L., Hunter, J., da Aparecida
Rodrigues Teles, M., Brustulin, R., de Padua Milagres, F. A., Sabino, E. C., Diaz, R. S., &
Komninakis, S. V. (2020). Detection of coinfection with chikungunya virus and dengue virus
serotype 2 in serum samples of patients in state of Tocantins, Brazil. Journal of Infection and
Public Health, 13(5), 724-729. doi:https://doi.org/10.1016/j.jiph.2020.02.034
Dutra, Heverton Leandro C., Rocha, Marcele N., Dias, Fernando Braga S., Mansur,
Simone B., Caragata, Eric P., & Moreira, Luciano A. (2016). Wolbachia blocks currently
circulating Zika virus isolates in Brazilian Aedes aegypti mosquitoes. Cell Host & Microbe,
19(6), 771-774. doi:https://doi.org/10.1016/j.chom.2016.04.021
Ekwudu, O. m., Marquart, L., Webb, L., Lowry, K. S., Devine, G. J., Hugo, L. E., &
Frentiu, F. D. (2020). Effect of serotype and strain diversity on dengue virus replication in
Australian mosquito vectors. Pathogens (Basel, Switzerland), 9(8), 668.
doi:10.3390/pathogens9080668
Ferguson, N. M., Kien, D. T. H., Clapham, H., Aguas, R., Tuan Trung, V. T., & Chau, T.
N. B. (2015). Wolbachia-mediated interference of dengue virus infection in Aedes aegypti and its
projected epidemiological impact on transmission. Science Translational Medicine, 7.
doi:10.1126/scitranslmed.3010370
Fontaine, A., Lequime, S., Moltini-Conclois, I., Jiolle, D., Leparc-Goffart, I., Reiner, R.
C., Jr., & Lambrechts, L. (2018). Epidemiological significance of dengue virus genetic variation
in mosquito infection dynamics. PLoS Pathogens, 14(7), e1007187.
doi:10.1371/journal.ppat.1007187
Frentiu, F. D., Robinson, J., Young, P. R., McGraw, E. A., & O'Neill, S. L. (2010).
Wolbachia-mediated resistance to dengue virus Infection and death at the cellular level. PLoS
ONE, 5(10), e13398. doi:10.1371/journal.pone.0013398
Gaye, A., Faye, O., Diagne, C. T., Faye, O., Diallo, D., Weaver, S. C., Sall, A. A., &
Diallo, M. (2014). Oral susceptibility of Aedes aegypti (Diptera: Culicidae) from Senegal for
dengue serotypes 1 and 3 viruses. Tropical Medicine & International Health, 19(11), 1355-1359.
doi:10.1111/tmi.12373

82
Gubler, D. J., Nalim, S., Tan, R., Saipan, H., & Sulianti Saroso, J. (1979). Variation in
susceptibility to oral infection with dengue viruses among geographic strains of Aedes aegypti.
The American Society of Tropical Medicine and Hygiene, 28(6), 1045-1052.
Guzman, M. G., & Harris, E. (2015). Dengue. The Lancet, 385(9966), 453-465.
doi:https://doi.org/10.1016/S0140-6736(14)60572-9
Hollidge, B. S., Weiss, S. R., & Soldan, S. S. (2011). The role of interferon antagonist,
non-structural proteins in the pathogenesis and emergence of arboviruses. Viruses, 3(6), 629-658.
doi:10.3390/v3060629
Holmes, E. C., & Twiddy, S. S. (2003). The origin, emergence and evolutionary genetics
of dengue virus. Infection, genetics and evolution : journal of molecular epidemiology and
evolutionary genetics in infectious diseases, 3(1), 19-28. doi:https://doi.org/10.1016/S1567-
1348(03)00004-2
Ko, H.-Y., Salem, G. M., Chang, G.-J. J., & Chao, D.-Y. (2020). Application of next-
generation sequencing to reveal how evolutionary dynamics of viral population shape dengue
epidemiology. Frontiers in Microbiol, 11(1371). doi:10.3389/fmicb.2020.01371
Lambrechts, L., Chevillon, C., Albright, R. G., Thaisomboonsuk, B., Richardson, J. H.,
Jarman, R. G., & Scott, T. W. (2009). Genetic specificity and potential for local adaptation
between dengue viruses and mosquito vectors. BMC Ecology and Evolution, 9, 160.
doi:10.1186/1471-2148-9-160
Lambrechts, L., Fansiri, T., Pongsiri, A., Thaisomboonsuk, B., Klungthong, C.,
Richardson, J. H., Ponlawat, A., Jarman, R. G., & Scott, T. W. (2012). Dengue-1 virus clade
replacement in Thailand associated with enhanced mosquito transmission. Journal of Virology,
86(3), 1853-1861. doi:10.1128/jvi.06458-11
Macdonald, G. (1957). The epidemiology and control of malaria. Amen House, Warwick
Square, London E.C.4.: Oxford University Press.
Murira, A., & Lamarre, A. (2016). Type-I interferon responses: from friend to foe in the
battle against chronic viral infection. Frontiers in Immunology, 7(609).
doi:10.3389/fimmu.2016.00609
Murray, N. E., Quam, M. B., & Wilder-Smith, A. (2013). Epidemiology of dengue: past,
present and future prospects. Clinical Epidemiology, 5.
Muturi, E. J., Buckner, E., & Bara, J. (2017). Superinfection interference between
dengue-2 and dengue-4 viruses in Aedes aegypti mosquitoes. Tropical Medicine & International
Health, 22(4), 399-406. doi:10.1111/tmi.12846
Novelo, M., Hall, M. D., Pak, D., Young, P. R., Holmes, E. C., & McGraw, E. A. (2019).
Intra-host growth kinetics of dengue virus in the mosquito Aedes aegypti. PLoS Pathogens,
15(12), e1008218. doi:10.1371/journal.ppat.1008218

83
Nuckols, J. T., Huang, Y. J., Higgs, S., Miller, A. L., Pyles, R. B., Spratt, H. M., Horne,
K. M., & Vanlandingham, D. L. (2015). Evaluation of simultaneous transmission of chikungunya
virus and dengue virus type 2 in infected Aedes aegypti and Aedes albopictus (Diptera:
Culicidae). The Journal of Medical Entomology, 52(3), 447-451. doi:10.1093/jme/tjv017
Quintero-Gil, D. C., Ospina, M., Osorio-Benitez, J. E., & Martinez-Gutierrez, M. (2014).
Differential replication of dengue virus serotypes 2 and 3 in coinfections of C6/36 cells and Aedes
aegypti mosquitoes. The Journal of Infection in Developing Countries, 8(7), 876-884.
doi:10.3855/jidc.3978
Richardson, J., Molina-Cruz, A., Salazar, M. I., & Black, W. t. (2006). Quantitative
analysis of dengue-2 virus RNA during the extrinsic incubation period in individual Aedes
aegypti. The American Society of Tropical Medicine and Hygiene, 74(1), 132-141.
Rico-Hesse, R. (2003). Microevolution and virulence of dengue viruses. Advances in
Virus Research, 59, 315-341. doi:10.1016/s0065-3527(03)59009-1
Rodriguez-Morales, A. J., Villamil-Gómez, W. E., & Franco-Paredes, C. (2016). The
arboviral burden of disease caused by co-circulation and co-infection of dengue, chikungunya and
Zika in the Americas. Travel Medicine and Infectious Disease, 14(3), 177-179.
doi:https://doi.org/10.1016/j.tmaid.2016.05.004
Rückert, C., Weger-Lucarelli, J., Garcia-Luna, S. M., Young, M. C., Byas, A. D.,
Murrieta, R. A., Fauver, J. R., & Ebel, G. D. (2017). Impact of simultaneous exposure to
arboviruses on infection and transmission by Aedes aegypti mosquitoes. Nature Communications,
8, 15412. doi:10.1038/ncomms15412
Schilling, S., Emmerich, P., Günther, S., & Schmidt-Chanasit, J. (2009). Dengue and
chikungunya virus co-infection in a German traveller. Journal of Clinical Virology, 45(2), 163-
164. doi:10.1016/j.jcv.2009.04.001
Senaratne, U. T. N., Murugananthan, K., Sirisena, P. D. N. N., Carr, J. M., & Noordeen,
F. (2020). Dengue virus co-infections with multiple serotypes do not result in a different clinical
outcome compared to mono-infections. Epidemiology & Infection, 148, e119-e119.
doi:10.1017/S0950268820000229
Shrivastava, S., Tiraki, D., Diwan, A., Lalwani, S. K., Modak, M., Mishra, A. C., &
Arankalle, V. A. (2018). Co-circulation of all the four dengue virus serotypes and detection of a
novel clade of DENV-4 (genotype I) virus in Pune, India during 2016 season. PLoS ONE, 13(2),
e0192672. doi:10.1371/journal.pone.0192672
Smartt, C. T., Shin, D., & Alto, B. W. (2017). Dengue serotype-specific immune
response in Aedes aegypti and Aedes albopictus. Memórias do Instituto Oswaldo Cruz, 112(12),
829-837. doi:10.1590/0074-02760170182
Souza-Neto, J. A., Powell, J. R., & Bonizzoni, M. (2019). Aedes aegypti vector
competence studies: A review. Infection, Genetics and Evolution 67, 191-209. doi:http s : / /
doi.org /10.1016/ j. meegid.2018.11.009

84
Taracena, M. L., Bottino-Rojas, V., Talyuli, O. A. C., Walter-Nuno, A. B., Oliveira, J. H.
M., Angleró-Rodriguez, Y. I., Wells, M. B., Dimopoulos, G., Oliveira, P. L., & Paiva-Silva, G.
O. (2018). Regulation of midgut cell proliferation impacts Aedes aegypti susceptibility to dengue
virus. PLoS Neglected Tropical Diseases, 12(5), e0006498. doi:10.1371/journal.pntd.0006498
Terradas, G., Allen, S. L., Chenoweth, S. F., & McGraw, E. A. (2017). Family level
variation in Wolbachia-mediated dengue virus blocking in Aedes aegypti. Parasites & Vectors,
10(1), 622-622. doi:10.1186/s13071-017-2589-3
Vasilakis, N., & Weaver, S. C. (2008). The history and evolution of human dengue
emergence. Advances in Virus Research, 72, 1-76. doi:10.1016/s0065-3527(08)00401-6
Vazeille, M., Gaborit, P., Mousson, L., Girod, R., & Failloux, A. B. (2016). Competitive
advantage of a dengue 4 virus when co-infecting the mosquito Aedes aegypti with a dengue 1
virus. BMC Infectious Diseases, 16, 318. doi:10.1186/s12879-016-1666-0
Vazeille, M., Mousson, L., Martin, E., & Failloux, A.-B. (2010). Orally co-infected
Aedes albopictus from la Reunion island, Indian ocean, can deliver both dengue and chikungunya
infectious viral particles in their saliva. PLoS Neglected Tropical Diseases, 4(6), e706.
doi:10.1371/journal.pntd.0000706
Villabona-Arenas, C. J., de Oliveira, J. L., Capra, C. d. S., Balarini, K., Loureiro, M.,
Fonseca, C. R. T. P., Passos, S. D., & Zanotto, P. M. d. A. (2014). Detection of four dengue
serotypes suggests rise in hyperendemicity in urban centers of Brazil. PLoS Neglected Tropical
Diseases, 8(2), e2620-e2620. doi:10.1371/journal.pntd.0002620
Villamil-Gómez, W. E., Rodríguez-Morales, A. J., Uribe-García, A. M., González-
Arismendy, E., Castellanos, J. E., Calvo, E. P., Álvarez-Mon, M., & Musso, D. (2016). Zika,
dengue, and chikungunya co-infection in a pregnant woman from Colombia. International
Journal of Infectious Diseases, 51, 135-138. doi:10.1016/j.ijid.2016.07.017
Vogels, C. B. F., Rückert, C., Cavany, S. M., Perkins, T. A., Ebel, G. D., & Grubaugh, N.
D. (2019). Arbovirus coinfection and co-transmission: A neglected public health concern? PLoS
Biol, 17(1), e3000130. doi:10.1371/journal.pbio.3000130
Walker, T., Johnson, P. H., Moreira, L. A., Iturbe-Ormaetxe, I., Frentiu, F. D., &
McMeniman, C. J. (2011). The wMel Wolbachia strain blocks dengue and invades caged Aedes
aegypti populations. Nature, 476. doi:10.1038/nature10355
Weaver, S. C., & Vasilakis, N. (2009). Molecular evolution of dengue viruses:
contributions of phylogenetics to understanding the history and epidemiology of the preeminent
arboviral disease. Infection, genetics and evolution : journal of molecular epidemiology and
evolutionary genetics in infectious diseases, 9(4), 523-540. doi:10.1016/j.meegid.2009.02.003

85
Chapter 4
Family level variation in the mosquito Aedes aegypti susceptibility and
immune response to dengue and chikungunya viral infection.
4.1 Abstract
Aedes aegypti is the main vector of the arboviruses dengue (DENV) and
chikungunya (CHIKV). Viral susceptibility and transmissibility in mosquito populations
depends on the GxG interactions between the virus and the vector. Both genotypes can also
have a great impact in the evolutionary potential of the vector:virus association. Here, we
used a modified full-sibling design to estimate the contribution of mosquito genetic
variation to the susceptibility and immune response to DENV and CHIKV. We
demonstrated substantial genetic variation in the susceptibility to both viruses.
Heritabilities were significant, but much higher for DENV susceptibility than to CHIKV,
at 40 and 18%, respectively. The differences are largely driven by greater within family
variation for CHIKV. Interestingly, we showed lack of genetic correlation between DENV
and CHIKV susceptibility between siblings of the same families. These data suggest that
the mosquito is much better at controlling DENV infection, and that the mosquito responds
to the viruses with two very different mechanisms. These disparities may result from
different coevolutionary histories, with CHIKV only recently emerging and acting as a
selective agent in mosquito populations. Last, in we tested for correlations between viral

86
susceptibility and the expression levels of several immune genes, identifying two genes
that may underpin the insect’s response to CHIKV (TEP3 and CECH).
4.2 Introduction
Arboviral diseases account for more than 17% of all infectious diseases globally
(Kading, Brault, & Beckham, 2020), among the most prevalent are dengue and
Chikungunya, with over half of the world’s population at risk and no effective vaccines or
treatments (Wilder-Smith et al., 2017). Although mortality is generally low, the morbidity
caused by these pathogens is associated with a substantial socioeconomic burden. In some
cases, infection with any of the four dengue virus serotypes (DENV-1, 2, 3, 4) can lead to
dengue shock syndrome in all ages (Rajapakse, 2011), and chikungunya virus (CHIKV)
infection can be severe for infants, with nearly half of infected newborns experiencing
encephalopathy and multiple organ dysfunction (Mehta et al., 2018). Both DENV and
CHIKV are single stranded RNA viruses, belonging to the Flaviridae and Togaviridae
viral families respectively, and are transmitted primarily through the bite of the mosquito
Ae. aegypti (Huang, Higgs, Horne, & Vanlandingham, 2014; Lim, Lee, Madzokere, &
Herrero, 2018). Unlike DENV, CHIKV is less well studied, largely as it was a relatively
unknown pathogen causing small outbreaks around the Indian ocean until recently
(Rougeron et al., 2015). In 2013 the virus was first reported in the Americas, and by 2018
there have been more than 3 million cases in 45 countries (Yactayo, Staples, Millot,
Cibrelus, & Ramon-Pardo, 2016; Zeller, Van Bortel, & Sudre, 2016).

87
Arboviral infection in the mosquito is a complex and dynamic process, as the
kinetics of infection can be influenced by the genetic variation of both the mosquito and
the virus (Novelo et al., 2019). Natural Ae. aegypti populations have been shown to vary
in their susceptibility to multiple arboviruses, including DENV and CHIKV (Gaye et al.,
2014; Gokhale, Paingankar, Sudeep, & Parashar, 2015; Gubler, Nalim, Tan, Saipan, &
Sulianti Saroso, 1979; Reiskind, Pesko, Westbrook, & Mores, 2008; Y. H. Ye et al., 2014),
however, studies have shown that diverse mosquito populations from the Americas, India
and the Pacific region have, are highly susceptible to CHIKV infection (Alto et al., 2017;
Chen et al., 2015; Gokhale et al., 2015; Tesh, Gubler, & Rosen, 1976; Vega-Rúa, Zouache,
Girod, Failloux, & Lourenço-de-Oliveira, 2014). Transmission efficiency in Ae. aegypti
populations from America have been shown to be up to 83% for multiple CHIKV
genotypes (Vega-Rúa et al., 2014). Moreover, CHIKV infectious particles have been
detected in Ae. aegypti salivary glands as early as 2 days post infection (DPI), indicating
that the dissemination of the virus within the mosquito is rapid and concomitant with
infection of the salivary glands (Dubrulle, Mousson, Moutailler, Vazeille, & Failloux,
2009). Comparatively, infectious DENV particles have been detected in the mosquito
salivary glands 4 DPI (Salazar, Richardson, Sánchez-Vargas, Olson, & Beaty, 2007).
Overall, the differences in susceptibility between CHIKV and other arboviruses, such as
DENV, have been linked to viral replication inside the mosquito, where genetic variation
and specific pathogen infection mechanisms can have an important role in determining
susceptibility and transmission.

88
Upon ingestion of a viral infected blood meal, the molecular interactions between
the mosquito cells and the pathogen starts with the viral particle attachment to cellular
receptors, facilitating viral endocytosis or activating signaling pathways to gain
intracellular access (Alonso-Palomares, Moreno-García, Lanz-Mendoza, & Salazar, 2018).
For DENV, the specific identity of mosquito cellular receptors remains elusive, although
several putative receptors have been suggested (Cruz-Oliveira et al., 2015; Smith, 2012).
To date, only one DENV cell expressed receptor has been characterized, Prohibitin, a
ubiquitously expressed conserved protein shown to interact with DENV-2 (Kuadkitkan,
Wikan, Fongsaran, & Smith, 2010). CHIKV cellular internalization is thought to be
mediated through clathrin-mediated endocytosis (CME), a key process in vesicular
trafficking (Dong, Balaraman, et al., 2017; Hoornweg et al., 2016; Lee et al., 2013). Upon
entry, both viruses are uncoated through intracellular specific conditions, such as pH, and
replicate in the cell cytoplasm (Rodriguez, Muñoz, Segura, Rangel, & Bello, 2019).
Cellular tropism for both DENV and CHIKV appears widespread to multiple mosquito
organs, including midgut, fat body tissue, nervous, tracheal and reproductive systems and
salivary glands (Kantor, Grant, Balaraman, White, & Franz, 2018; Matusali et al., 2019;
Salazar et al., 2007; Vega-Rúa, Schmitt, Bonne, Krijnse Locker, & Failloux, 2015; Wong,
Chan, Sam, Sulaiman, & Vythilingam, 2016; Zhang et al., 2010; Ziegler et al., 2011).
Interestingly, variation in the replication rates between viruses have been reported inside
Ae. aegypti. Specifically, one study measuring viral RNA copies of each virus (Le
Coupanec et al., 2017), showed that in the mosquito midguts, DENV had a lower copy
number than CHIKV at 3 DPI, similarly, in the salivary glands, CHIKV was detected as
early as 4 DPI whereas DENV was detected no earlier than 10 DPI. Moreover, co-infection

89
with DENV and CHIKV has shown to increased overall mosquito infection rates up to
100% for both viruses, while also enhancing CHIKV and DENV viral replication in the
midgut and salivary glands, compared to each virus alone (Le Coupanec et al., 2017).
Viral infection triggers cellular and humoral immune responses inside the
mosquito. The core pathways of the insect immune response are the Janus Kinase-Signal
Transducer Activator of transcription (JAK/STAT), Toll, Immune Deficiency (IMD) and
RNA interference (RNAi) (Alonso-Palomares et al., 2018). Viral components also interact
with pattern recognition receptors, triggering processes, such as melanization and the
production of anti-microbial peptides (AMP), including acting, defensin and cecropin
among others (Agaisse & Perrimon, 2004; Antonova, Alvarez, Kim, Kokoza, & Raikhel,
2009; Sim & Dimopoulos, 2010). Additionally, complement related proteins in the
mosquito hemolymph, such as thioester containing proteins (TEP), can recognize
arboviruses and trigger further AMP production (Mukherjee, Das, Begum, Mal, & Ray,
2019). Interestingly, some of the traditional innate immune pathways (Toll,
JAK/STAT,IMD) that been shown to contribute to the reduction of DENV replication both
in vitro and in vivo, play little role in limiting CHIKV infection. Specifically, exogenous
activation of the JAK-STAT pathway has been shown to modulate DENV infection, but
resulted in no enhanced resistance to CHIKV (Jupatanakul et al., 2017). Moreover, CHIKV
infection has also been shown to significantly repress the Toll pathway stimulation, and
neither the IMD pathway has been found to mediate viral activities (McFarlane et al.,
2014). Of the mayor insect immune pathways known to control infection of arboviruses
such as DENV, only the RNAi pathway has been shown to play a vital role in limiting

90
CHIKV replication. Knockdown of Argonaute2 (AGO2), an RNAi effector molecule,
resulted in significant increase of viral RNA replication and titers (McFarlane et al., 2014).
Transcriptomic profiles of DENV and CHIKV infected mosquitoes have shown to
be distinct, with little overlap between responsive genes (Shrinet, Srivastava, & Sunil,
2017). Expression profiles of CHIKV-infected mosquitoes have revealed upregulation of
immune genes encoding AMPs, such as defensin and cecropin (CECH), and TEP (Dong,
Behura, & Franz, 2017). Both TEP and CECH have been shown to be highly upregulated
upon CHIKV infection up 10 DPI. In the same study, TEP3 expression was shown to be
highly induced with up to 16-fold greater expression than other genes (Zhao, Alto, Jiang,
Yu, & Zhang, 2019). Another study in malaria infected Anopheles gambie mosquitoes,
revealed that TEP molecules can function as positive immune regulators, reducing
pathogen intensity (Clayton, Dong, & Dimopoulos, 2014). Further reports have shown
increased expression of the CECH gene upon pathogen infection (Antonova et al., 2009;
Kokoza et al., 2010; Muturi, Blackshear, & Montgomery, 2012), including with Sindbis
virus (SINV) infection, a member of the Togaviridae family (Kim & Muturi, 2013).
Overall, the current data indicates that these genes might be important for CHIKV infection
response inside the vector.
There have been several recent discoveries of novel non-canonical antiviral genes
in the fly (Magwire et al., 2012; Piontkivska et al., 2016) and in Ae. aegypti (Ford et al.,
2019) revealed by examining population level genetic variation in the antiviral response.
Studying the natural genetic variation in Ae. aegypti could lead to insights in the basis of
the vector’s own natural immune response to CHIKV and DENV, and may offer a means

91
to identify candidate antiviral genes to target with emerging insect genetic tools, such as
CRISPR-cas9, (Terradas, Allen, Chenoweth, & McGraw, 2017; Terradas & McGraw,
2019). Here, we used a modified full-sib breeding design to assess the family level
variation in CHIKV and DENV susceptibility, using a U.S. population of Ae. aegypti. By
examining the susceptibility trait for both viruses in the same population, we can show the
contribution of the genetic variation to the viral load variance. We then used the families
showing extreme phenotypes in viral load to screen candidate genes relate to infection
susceptibility. The set of candidate genes selected, were either common responders to viral
infection of both CHIKV and DENV, or were shown to have a specific effect only in
CHIKV-infection based on the literature.
4.3. Methods
4.3.1 Mosquito line and stock practices
An F3 Ae. aegypti mosquito line originating from Monterrey, Mexico, was reared in the
laboratory for 3 generations to expand and adapt the colony to laboratory feeding regimes.
Mosquitoes were reared under standard conditions; 26°C, 65% relative humidity and a 12
h light/dark cycle. Larvae were maintained on fish food (Tetramin, Tetra). Adults were fed
with 10% sucrose solution ad libitum. Mosquitoes were fed human blood (BioIVT,
Hicksville, NY, USA) for egg collection using a Hemotek system (Hemotek).

92
4.3.2. Breeding design
A modified full – sib breeding design (William G. Hill & Mackay, 2004; Terradas
et al., 2017; Yixin H. Ye et al., 2016) was carried out on the mosquitoes in combination
with DENV and CHIKV infection. After expanding the mosquito line over 3 generations,
F6 eggs were hatched in synchrony and reared at low density (~150 larvae per 3 ltr of RO
water). After pupation, males and females were separated and transferred to 30 x 30 x 30
cm cages at a density of ~250 individuals per cage. Six to seven- day old virgin adult
females were blood-fed and then 250 virgin males were then added for mating. A total of
600 families were created by placing 600 blood-fed and mated females in small individual
housings containing moist filter paper. Egg papers were collected and dried 3-4 days later,
for short term storage. We selected female lineages that had produced a minimum of 60
eggs. For each family, eggs were hatched separately and after pupation, females were
separated and split into two cups with a minimum of 8 individuals per cup. Mosquitoes
were maintained on sucrose until vector competence experiments.
4.3.3. Virus culture
All experiments were carried out using the CHIKV strain 20235-St Martin 2013
(BEI Resources), and the DENV-2 strain 429557-Mex 2005 (BEI Resources), both from
Latin American outbreaks. Virus was cultured in C6/36 cells as previously reported
(Frentiu, Robinson, Young, McGraw, & O'Neill, 2010; Novelo et al., 2019; Terradas et al.,
2017; Yixin H. Ye et al., 2016). Briefly, C6/36 Ae albopictus cells were grown in RPMI
1640 media (Life Technologies, Carlsband, CA, USA) and supplemented with 10% heat-

93
inactivated fetal bovine serum (FBS, Life Technologies), 1% Glutamax (Life
Technologies) and 25 mM HEPES (Sigma-Aldrich, St. Louis, MO, USA). Cells were
grown to a confluency of 80% and then independently infected with DENV-2 and CHIKV.
Infected culture flasks were incubated at 27 oC. For DENV-2, after 7 days post-infection,
supernatant was harvested at a titer of 1.0 x 105 focus forming units per ml (FFU/ml). For
CHKV, supernatant was harvested at 2 days post infection at a titer of 8.7 x 105 and
adjusted to a final concentration of 1.0 x 105 FFA/ml. Both viruses were store at -80oC in
1 ml single use aliquots for vector competence experiments.
4.3.4. Mosquito infections
Methods for oral mosquito infections have been fully described previously
(Amuzu, Simmons, & McGraw, 2015; Novelo et al., 2019; Terradas et al., 2017). Prior to
infections, mosquitoes were starved for 24 hrs. Half of the 6–7-day old females of each
family were challenged with DENV-2 (1st cup) and the other half (2nd cup) with CHIKV
at equal viral titers using a 1:1 mix of the frozen titrated aliquots and human blood.
Mosquitoes were allowed to feed for 30 minutes and shortly after they were anesthetized
with CO2 and unfed individuals were removed and discarded. All CHIKV work was carried
out at the Penn State Eva J. Pell ABSL-3 laboratory.
4.3.5. Viral quantification
After 7 days post infection, whole mosquitoes were collected, homogenized and
stored in trizol (Invitrogen, Carlsbad, CA, USA) reagent at -80 oC for downstream
molecular analysis. RNA was extracted according to the manufacturer’s protocol. RNA

94
was eluted in 25 ul of DNA/RNA free water and DNAse treated (Life Technologies,
Carlsbad, CA, USA). DENV and CHIKV was quantified using 4× TaqMan fast virus 1-
step master mix (Applied biosystems, Foster city, CA, USA) in individual 10 ul reactions
containing virus specific primers and probes (Table 4.1). The RT-qPCR reactions were run
in a LightCycler 480 instrument (Roche Applied Science, Switzerland). The thermal
cycling conditions were 50°C for 5 min for reverse transcription, 95°C for 10s for RT
inactivation/denaturation followed by 50 amplification cycles of 95°C for 3s, 60°C for 30s
and 72°C for 1s. Standard curves for both DENV and CHIKV were generated as described
elsewhere (Rückert et al., 2017; Yixin H. Ye et al., 2016) and contained a ~100 bp fragment
of the 3’UTR of the DENV genome and a ~140 bp fragment of the CHIKV genome,
respectively. The standard curve spanned from 107 to 10 copies/reaction with a limit of
detection of 100 copies for both viruses. The viral load in each sample was extrapolated
from the standard curve as copies per mosquito.
4.3.6. Selection of immune genes for expression analysis
Both viral load and infection intensity have been shown to be modulated by the
mosquito immune response (Blair & Olson, 2014). To assess the potential role of specific
immune genes in viral infection, we used families with extreme viral loads to test the for
associations with gene expression. The null genetic correlation between DENV and
CHIKV infected mosquitoes suggests a genetic decoupling in the mechanisms that drive
the susceptibility to either virus in mosquitoes. Therefore, we aimed to test immune genes
that represent mosquito immune pathways that have 1) shown to be of great importance in
modulating infection for both viruses, and 2) that have might be important in the immune

95
response against CHIKV. We examined whether virus type and viral load group altered the
expression level of several immune related genes. We compared the expression levels of
four genes associated with viral infection in Ae. aegypti. For the first group, we focused in
two genes, AGO2 and DCR2, that are part of the mosquito RNA RNAi pathway which
have been shown to modulate both CHIKV and DENV. For the second group, we tested
the antimicrobial peptide (AMP), cecropin (CECH), a TOLL pathway-regulated immune
effector (Mukherjee et al., 2019; Ramirez & Dimopoulos, 2010; Xi et al., 2008) and the
complement - related immune protein TEP3 (Blandin & Levashina, 2004; Mukherjee et
al., 2019) which have been shown to be upregulate in CHIKV infection but not DENV.
4.3.7. Immune gene expression
Retrotranscription of RNA to cDNA and gene expression analysis was carried out
on a LightCycler 480 instrument (Roche) using the Script One-step SYBR Green qRT-
PCR (Quantabio, Beverly, MA, USA) according to the manufacturer’s protocol. All CT
values were normalized to the housekeeping Ae. aegypti gene RpS17. Gene expression
ratios were obtained using the ∆∆Ct method (Livak & Schmittgen, 2001). PCR
amplification cycled 45 times at 95° C for 3 sec and 60° C for 30 sec, and the final cycle
was followed by a melting curve analysis. All primers for candidate genes and viral
quantification are listed in table 4.1.

96
4.3.8. Analysis of genetic variance
We tested for genetic variation and broad-sense heritability (H2) for DENV and
CHIKV loads across our family design. We estimated the parameters using a modified full-
sib breeding design and the random effect linear model previously described (Terradas et
al., 2017; Yixin H. Ye et al., 2016):
zij = fi + εij (1)
Zij is the trait value for the jth female from the ith family, f i is the random effect of
the ith family and ε ij is the unexplained error. To test if the genetic variation was
significant, the family term was fitted as a random effect and the model (1) was compared
to a reduce model without the family term. Loglikelihoods of both models were compared
and twice the difference was tested against a chi-squared distribution with a single degree
Table 4.1. Primers and probes for qPCR gene expression and viral quantification.
Gene ID Target Direction Sequence (5’-3’)
AY713296 DICER 2 Forward GAATTCCTCGGCGATGCGGTATTA
Reverse GATGCCGACTCTGCCAGGATGT
AAEL008607 TEP-3 Forward AGTGTCCGTTGAGTCTCCTG
Reverse TCTACCGATCCCTTGCCATC
AAEL004175 RPS17 Forward TCCGTGGTATCTCCATCAAGCT
Reverse CACTTCCGGCACGTAGTTGTC
NC_001474.2 DENV
Forward AAGGACTAGAGGTTAGAGGAGACCC
Reverse CGTTCTGTGCCTGGAATGATG
Probe
FAM-
AACAGCATATTGACGCTGGGAGAGACCAG
A-BHQ1
MT228632 CHIKV
Forward CACCCGAAGTAGCCCTGAATG
Reverse TCCGAACATCTTTCCTCCCG
Probe 5CY5-GAGAATAGCCCGCTGTCTAGATCCAC-
3BHQ2

97
of freedom. Broad sense heritability was estimated as twice the family variance component
(σ family) divided by the total phenotypic variance (σ family + σ error), (Saxton, 2004). All
models were constructed in SAS studio version 3.8 (SAS Institute, Cary, NC, USA).
Genetic correlations between DENV and CHIKV infected siblings was estimated using a
bivariate version of model (1) and fitting unrestrictive covariance correlations at the family
level (TYPE=UNR option) suing SAS Proc MIXED command. Significance of the genetic
correlation was tested by loglikelihood ratio tests.
4.4 Results
4.4.1. Family level variation in the heritability of DENV and CHIKV viral loads
To test for differences in the heritability across DENV and CHIKV viral loads we
assayed a total of 800 DENV and CHIKV fed individual mosquitoes from 50 families in a
modified full-sib design. Overall, we obtained 37 families for DENV and 38 for CHIKV
that had enough infected mosquitoes for further analysis. Infected mosquitoes were tested
for both viruses at day 7 post infection. After RNA extraction, we quantified DENV and
CHIKV via RT-qPCR in 3-5 individuals per family. The broad sense heritability (H2) of
DENV load was estimated to be 0.40 (Figure 4.1A. LRT: X2=24.8, P=>0.001) and for
CHIKV load, H2 was 0.18 (Figure 4.1B. LRT: X2= 5.9, P=0.015) across all the families.
Both heritability values were significantly greater than zero, indicating a role for multi-
genetic interactions in determining the viral load trait across the mosquito families for
DENV and CHIKV. The range of viral load was extremely variable across the families,

98
spanning from hundreds to millions of copies; 102-107 per body for DENV and 102-108 per
body for CHIKV.
Figure 4.1. Ranked families by viral load. A) DENV load ranked families and B) CHIKV load
ranked families. Each point represents a single infected mosquito. Box plots show mean viral load
± SEM of each distinct Ae. aegypti family. Whole mosquito viral load was log10 transformed. N=3-
5 individuals per family.

99
4.4.2. Genetic correlation between DENV and CHIKV across mosquito families
We found no correlation between DENV and CHIKV loads between siblings of the
same family based on 37 families (Figure 4.2. Pearson’s r = 0.0018, p = 0.99). Furthermore,
the correlation was not significantly greater than zero (LRT: X2= >0.001, P= 1.00). The
decoupling between susceptibility traits for siblings indicates a lack of shared control
mechanisms for the two viruses.
Figure 4.2. Scatter plot showing the relationship between mean DENV and CHIKV loads
from individual families. Gray area represents the 95% confidence interval of the regression line.
N = 37 mosquito families.
4.4.3 Phenotypic extremes of viral loads for both DENV and CHIKV infection in the
mosquito families
We selected the five most extreme mosquito families with respect to mean viral
load for both viruses. We then tested for differences between the viral loads between these
groups and found a statistically significant difference between our family extremes for both
DENV (Figure 4.3A, w=256, df=1, p= <0.0001) and CHIKV (Figure 4.3B, w=393, df=1,
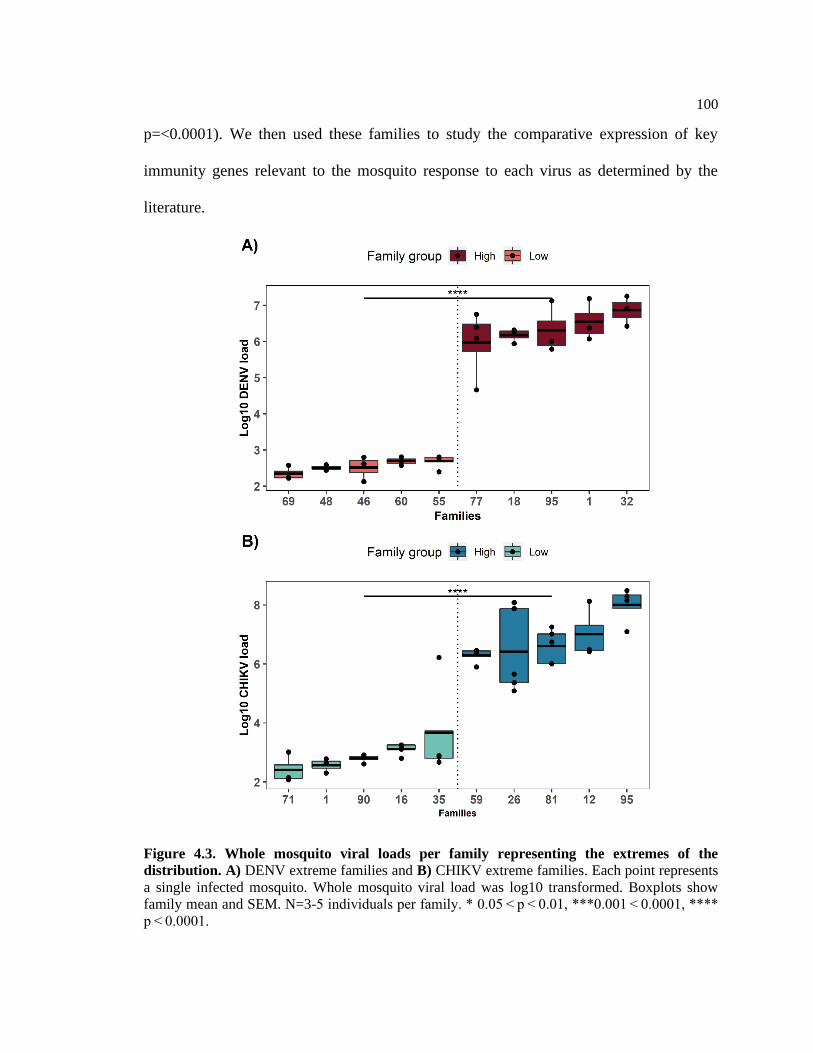
100
p=<0.0001). We then used these families to study the comparative expression of key
immunity genes relevant to the mosquito response to each virus as determined by the
literature.
Figure 4.3. Whole mosquito viral loads per family representing the extremes of the
distribution. A) DENV extreme families and B) CHIKV extreme families. Each point represents
a single infected mosquito. Whole mosquito viral load was log10 transformed. Boxplots show
family mean and SEM. N=3-5 individuals per family. * 0.05 < p < 0.01, ***0.001 < 0.0001, ****
p < 0.0001.

101
4.4.4. Immune genes relative expression
We evaluate the relative expression of two genes known to be involved in viral
control of both CHIKV and DENV, DCR2 (AAEL006794) and AGO2 (AAEL017251),
part of the RNAi pathway. For DENV infected mosquitoes, there was a significant
difference in relative AGO2 expression between family groups, with higher expression of
the gene in the High viral load group (Figure 4.4A, w=236, df=1, p=0.001). The average
gene expression fold change of was 1.49 between High/Low viral load groups. For DCR2
expression, there was no significant difference between extreme family groups (Fig 4.4C,
w= 116, df=1, = 0.48) with a average fold change in gene expression of 1.22 between
High/Low viral load groups. For CHIKV infected mosquitoes, we saw no significant
difference in relative AGO2 gene expression between extreme family groups (Fig 4.4B,
w= 288, df=1, p= 0.08). The average gene expression fold change of High/Low viral load
groups was 1.39. However, there was a significant difference in DCR2 expression between
family groups were the High viral load group had higher expression (Fig 4.4D, w=352,
df=1, p=<0.001) with an average gene expression fold change of 2.98 over the Low viral
load group.
The second set of genes we evaluate are two effector molecules that have been
shown to be important in the overall control of CHIKV but not DENV infection in Ae.
aegypti, CECH (AAEL017211) and TEP3 (AAEL008607). For DENV infected
mosquitoes we identify a significant difference between groups (Fig 4.5A, w=67, df=1,
p=0.01), with a higher expression of the TEP3 in the Low viral load group. The average
gene expression fold change between High/Low viral load groups was 0.17. However, we

102
saw no significant difference in CECH gene expression between the High and Low viral
load groups (Fig 4.5C, w=115, df=1, p=0.31) with an average gene expression fold change
of 0.83. For CHIKV infected mosquitoes we saw a significant difference in both TEP3 (Fig
4.5B, w=342, df=1, p=<0.0001) and CECH (Fig 4.5D, w=394, df=1, p=<0.0001) and, with
higher gene expression in the High CHIKV group. The average gene expression fold
change between High/Low viral load groups was 3.84 for TEP3 and 2.98 for CECH.
Additionally, we looked at the relationship between gene expression and viral load
regardless of the family groups (i.e., High and Low) for both DENV and CHIKV infected
mosquitoes. This was done to take into account the variation in gene expression in families
that might not correspond to their viral load groups. For DENV infected mosquitoes the
only significant interaction was for AGO2 expression, that was positively correlated to viral
load (Appendix C, Spearman’s r = 0.63, p= .05). For CHIKV infected mosquitoes all genes
were positively correlated to viral load (Appendix C). In general, the correlation trends
corresponded to the viral load family groupings previously observed.

103
Figure 4.4. AGO2 and DCR2 expression relative to the housekeeping gene RPS17 in family
groups classified as High and Low Viral loads. A) AGO2 expression in DENV infected families;
B) AGO2 expression in CHIKV infected families and C) DCR2 expression in DENV infected
families; D) DCR2 expression in CHIKV infected families. Boxplots show family mean with SEM.
N=3-5 individuals per family. * 0.05 < p < 0.01, ***0.001 < 0.0001, **** p < 0.0001. The average
fold change of High viral load /Low viral load groups is A) 1.49, B) 1.36, C) 1.22, D) 2.98.

104
Figure 4.5. TEP3 and CECH expression relative to the housekeeping gene RPS17 in family
groups classified as High and Low Viral loads. A) TEP3 expression in DENV infected families;
B) TEP3 expression in CHIKV infected families and C) CECH expression in DENV infected
families; D) CECH expression in CHIKV infected families. Boxplots show family mean with SEM.
N=3-5 individuals per family. * 0.05 < p < 0.01, ***0.001 < 0.0001, **** p < 0.0001. The average
fold change of High viral load /Low viral load groups is A) 0.17, B) 3.84, C) 0.83, D) 2.98.

105
4.5. Discussion
We performed a modified full-sib breeding design to study the contribution of the
mosquito’s genetic variation, while limiting environmental variation, to susceptibility to
each virus. By exploring susceptibility to the two viruses in sisters of the same family, we
were also able to measure the genetic correlation between the individual susceptibilities.
We used the distribution of susceptibilities to select families representing the phenotypic
extremes for each virus. In these extreme families we tested for differences in expression
for key genes previously shown to represent the mosquito immune response to each virus.
Our experiments demonstrate the role of multi-genic interactions in determining the viral
load trait across mosquito families for both DENV and CHIKV. Additionally, the lack of
genetic correlation between DENV and CHIKV viral loads across siblings, demonstrated
independence of the genetic response to these two viruses. Finally, our gene expression
analysis on the extreme families showed that, although some viral control mechanisms,
such as the RNAi pathway, play a universal role in modulating viral replication, the
mosquitos’ response to infection appears markedly different depending on the infecting
virus.
The heritability values of the viral susceptibility for both DENV and CHIKV were
significant but fell within different ranges. For DENV, the H2 was 40%, this is in
accordance with previous studies examining susceptibility in Ae. aegypti using multiple
tissues as proxy. Both Bosio et al. (Bosio, Beaty, & Black, 1998) and Ye et al. (Yixin H.
Ye et al., 2016) reported heritabilities of ~40% in the midgut, head and saliva of DENV
infected mosquitoes. This suggests, that for DENV infection susceptibility, the mosquito

106
genotype differences contribute to vector competence almost equally as the viral genotype
or environmental effects do. Heritability of CHIKV susceptibility was estimated at 18%,
almost half that estimated for siblings in the same family. The lower heritability for CHIKV
suggests that environmental or stochastic factors as well as the viral genotype have a
greater role than the mosquito’s own genetic factors in determining the susceptibility to
infection (Charmantier & Garant, 2005; William G. Hill, Goddard, & Visscher, 2008;
William G. Hill & Mackay, 2004). CHIKV infected mosquitoes also exhibited a wider
range in in viral loads than DENV and reached higher viral loads than DENV infected
mosquitos. Overall, the wider range of viral loads in CHIKV is a key contributor to the
overall low heritability for this trait. The genetic correlation between siblings for the DENV
and CHIKV susceptibility trait was not significantly different than zero. This shows
independence between the genes that control the susceptibility trait between viruses.
Additionally, a value less than one indicates the presence of genotype x environment
interactions (William G. Hill & Mackay, 2004). In general, the primary source of genetic
correlation is the pleiotropy of genes, though linkage disequilibrium can be a contributing
factor (Sgrò & Hoffmann, 2004; Slatkin, 2008). Our results could suggest that genes
controlling this trait are present in different loci among the mosquito population or that
they lack pleiotropic effect, meaning that different genes control susceptibility to infection
depending on the virus (W. G. Hill, 2013).
Phylogenetic studies on Ae. aegypti and DENV have revealed a coevolutionary
process between the virus and the co-indigenous mosquito populations (Alonso-Palomares
et al., 2018; Barón, Ursic-Bedoya, Lowenberger, & Ocampo, 2010; Sim & Dimopoulos,
2010) which has in turn shape the susceptibility to the virus. Since CHIKV is a re-emerging

107
disease, the lack of the coevolutionary process with naïve, such as American, Ae. aegypti
populations, could be primary source of the differences between the mechanisms that
control infection inside the vector (Alonso-Palomares et al., 2018). The global spread of
CHIKV and DENV was preceded by the global expansion of Ae. aegypti and Ae.
albopictus. While the expansion of Ae. aegypti started over 400 years ago, the geographic
expansion of Ae. albopictus, the secondary vector of DENV and CHIKV started since the
1960’s, previously being limited to Southeast Asia (Louis Lambrechts, Scott, & Gubler,
2010). Among the evolutionary forces that drive the evolution of arboviruses, vector
interactions may play an important role in selecting viral genotypes with low fitness costs
and high transmissibility rates (L. Lambrechts et al., 2009). CHIKV emergence in the
Indian Ocean was linked to a single mutation that enhanced transmission by Ae. albopictus
over Ae. aegypti (Tsetsarkin, Vanlandingham, McGee, & Higgs, 2007). It has been
hypothesized that the emergence of arboviruses is partially caused by adaptations to native
vector populations. Invasive viral genotypes, that outcompete local genotypes tend to have
enhanced viral replication in the vector (Chevillon & Failloux, 2003).
Using the natural genetic variation in a mosquito population, we were able to select
family extremes with respect to DENV and CHIKV viral loads. Using these families, we
then quantify the expression levels of four genes associated with the mosquito immunity
after viral infection with DENV and CHIKV. We tested two genes involve in the immune
response for both viruses and two immune genes that have been recently shown to be of
importance upon CHIKV infection. We decided for this this approach since very little is
known about Ae. aegypti immune response to CHIKV compared to DENV. Additionally,

108
the two genes of importance for CHIKV that we chose, have been shown to be upregulated
in transcriptomic studies of CHIKV infected mosquitoes (Zhao et al., 2019).
The first set of genes, AGO2 and DCR2, are part of the RNAi-induced silencing
complex (RISC) (Pratt & MacRae, 2009) and been shown to play a vital role in limiting
CHIKV and DENV replication (Franz et al., 2006; Liu, Swevers, Kolliopoulou, &
Smagghe, 2019; McFarlane et al., 2014; Mukherjee et al., 2019). Interestingly, the
expression pattern of both genes was different depending on the infecting virus. AGO2
expression was positively correlated to higher viral loads in DENV infected families but
not so for CHIKV. In contrast, DCR2 expression was positively correlated with load in
CHIKV infected families, but no correlation was seen for DENV loads. Induction of RISC
starts with DCR2 recognition and cleavage of exogenous viral double stranded RNA
(dsRNA) into small interfering RNAs (siRNAs). The siRNAs are loaded into RISC and
cleaved by AGO2 leading to further viral degradation (Blair, 2011). Arbovirus infection
studies in Ae. aegypti have indicate that, given the ability of viruses to replicate despite
RNAi response, that arboviruses must possess a mechanism of immune evasion or
suppression (Blair & Olson, 2014). Studies with Flock House Virus (FHV) infected
Drosophila have shown that some viral genome siRNAs are inefficiently loaded into RISC,
potentially reducing the efficacy of RNAi downstream of DCR2 and allowing the virus to
maintain replication (Flynt, Liu, Martin, & Lai, 2009). For DENV-2, it has been shown
that its genome contains an overrepresentation of DCR2 target sites suggesting a decoy
mechanism for RNAi evasion (Sánchez-Vargas et al., 2009). These hotspots have also been
shown to be a poor target for AGO2 cleavage potentially due to secondary intra-strand
structures (Siu et al., 2011). Additionally, RNAi targeting has been shown to produce viral

109
genotype diversification, leading to the proliferation of escape mutants within the same
viral population that are not easily recognized by the effectors in this pathway (Brackney,
Beane, & Ebel, 2009). Whether either virus genotype actively promotes these evasion
strategies, that result in differential RISC viral modulation, remains unknown.
The second set of genes, CECH and TEP3, are a TOLL pathway-regulated
(Mukherjee et al., 2019; Ramirez & Dimopoulos, 2010; Xi, Ramirez, & Dimopoulos, 2008)
antimicrobial peptide (AMP), and a complement - related immune protein (Blandin &
Levashina, 2004; Mukherjee et al., 2019), respectively. The expression pattern for both
genes was also different depending on the virus. TEP3 expression was negatively
correlated with DENV viral load while it was positively correlated to CHIKV viral load.
No expression pattern was seen for CECH in DENV infected families but it was positively
correlated to CHIKV viral load. The expression pattern of both genes in is in line with other
studies suggesting their role in CHIKV immune modulation (Zhao et al., 2019).
Interestingly, the inverse correlation between TEP3 expression and CHIKV and DENV
viral loads suggest a differential interaction with the viruses. For CHIKV, the TEP3
expression pattern suggest that the immune response mediated through the immune protein
is reactive to the pathogen while for DENV, it suggests viral suppression. The mechanisms
by which each virus might affect the expression of these genes remains unknown.
Our experimental design has some caveats that may limit the scope of its overall
interpretation. Immune effector expression can vary depending on the time post infection
and its possible that the expression levels of specific genes might peak immediately after
blood feeding and were missed from our sampling regime (Erler, Popp, & Lattorff, 2011;
Sim & Dimopoulos, 2010). Given the scale of the family-based design we could only

110
sample one single time point. The experimental conditions differ from past transcriptomic
and gene expression studies used to identify our gene candidates. The expression results
shown here do not exclude genes that might be relevant in different tissues and timepoints.
Finally, the immune response of Ae. aegypti is complex and dynamic and several questions
regarding the mosquitos’ immunity remain outstanding, since most of what is known about
the general immunity of arthropods has been elucidated in the Drosophila model system
(Wang et al., 2015). Specifically, the precise molecular details of Ae. aegypti antiviral
immune systems remains undescribed (Cheng, Liu, Wang, & Xiao, 2016).
From an evolutionary perspective, the differences in quantitative genetics shown
here indicate that the selective pressures that lead to DENV infected mosquitoes having a
higher heritability and more homogeneous viral loads are stronger than those for CHIKV,
suggesting that the selection for higher or lower viral loads within a mosquito population
would occur faster for DENV than CHIKV infection (William G. Hill & Mackay, 2004).
This would have implication in our ability to predict the strength and impact of new
outbreaks and the overall infection dynamics between Ae. aegypti and CHIKV. These
changes could also explain CHIKV re-emergence and explosion in the American continent
in naïve Aedes populations. Additionally, the results shown here suggest that the infection
dynamics and some of the mechanisms that control viral susceptibility in mosquito
populations are markedly different between DENV and CHIKV. From an epidemiological
perspective, if the genes that control viral susceptibility to multiple viruses are different
and are located at different loci in the mosquito genome, this could introduce a challenge
to vector control methods aiming to target and modify universal loci that could be used
against multiple arbovirus. In this case, more individualized gene editing approaches would

111
be necessary depending on the predominant virus present (Scott & Zhang, 2017). Our gene
expression results also provide with some candidate genes that are not traditionally
associated with antiviral activity, that should be further examined as potential targets to
make refractory mosquito populations through gene editing techniques.

112
4.6. References
Agaisse, H., & Perrimon, N. (2004). The roles of JAK/STAT signaling in Drosophila
immune responses. Immunological Reviews, 198(1), 72-82. doi:https://doi.org/10.1111/j.0105-
2896.2004.0133.x
Alonso-Palomares, L. A., Moreno-García, M., Lanz-Mendoza, H., & Salazar, M. I.
(2018). Molecular basis for arbovirus transmission by Aedes aegypti mosquitoes. Intervirology,
61(6), 255-264. doi:10.1159/000499128
Alto, B. W., Wiggins, K., Eastmond, B., Velez, D., Lounibos, L. P., & Lord, C. C.
(2017). Transmission risk of two chikungunya lineages by invasive mosquito vectors from
Florida and the Dominican Republic. PLoS Neglected Tropical Diseases, 11(7), e0005724.
doi:10.1371/journal.pntd.0005724
Amuzu, H. E., Simmons, C. P., & McGraw, E. A. (2015). Effect of repeat human blood
feeding on Wolbachia density and dengue virus infection in Aedes aegypti. Parasites & Vectors,
8, 246. doi:10.1186/s13071-015-0853-y
Antonova, Y., Alvarez, K. S., Kim, Y. J., Kokoza, V., & Raikhel, A. S. (2009). The role
of NF-kappaB factor REL2 in the Aedes aegypti immune response. Insect biochemistry and
molecular biology, 39(4), 303-314. doi:10.1016/j.ibmb.2009.01.007
Barón, O. L., Ursic-Bedoya, R. J., Lowenberger, C. A., & Ocampo, C. B. (2010).
Differential gene expression from midguts of refractory and susceptible lines of the mosquito,
Aedes aegypti, infected with Dengue-2 virus. Journal of Insect Science, 10(1).
doi:10.1673/031.010.4101
Blair, C. D. (2011). Mosquito RNAi is the major innate immune pathway controlling
arbovirus infection and transmission. Future Microbiology, 6(3), 265-277. doi:10.2217/fmb.11.11
Blair, C. D., & Olson, K. E. (2014). Mosquito immune responses to arbovirus infections.
Current opinion in insect science, 3, 22-29. doi:10.1016/j.cois.2014.07.005
Blandin, S., & Levashina, E. A. (2004). Thioester-containing proteins and insect
immunity. Molecular Immunology, 40(12), 903-908. doi:https://doi.org/10.101 6/j.molim
m.2003.10.010
Bosio, C. F., Beaty, B. J., & Black, W. C. t. (1998). Quantitative genetics of vector
competence for dengue-2 virus in Aedes aegypti. The American Society of Tropical Medicine and
Hygiene, 59(6), 965-970.
Brackney, D. E., Beane, J. E., & Ebel, G. D. (2009). RNAi targeting of West Nile virus in
mosquito midguts promotes virus diversification. PLoS Pathogens, 5(7), e1000502.
doi:10.1371/journal.ppat.1000502
Charmantier, A., & Garant, D. (2005). Environmental quality and evolutionary potential:
lessons from wild populations. Proceedings. Biological sciences, 272(1571), 1415-1425.
doi:10.1098/rspb.2005.3117

113
Chen, T.-H., Jian, S.-W., Wang, C.-Y., Lin, C., Wang, P.-F., Su, C.-L., Teng, H.-J., Shu,
P.-Y., & Wu, H.-S. (2015). Susceptibility of Aedes albopictus and Aedes aegypti to three
imported chikungunya virus strains, including the E1/226V variant in Taiwan. Journal of the
Formosan Medical Association, 114(6), 546-552. doi:https://doi.org/10.1016/j.jfma.2014.12.002
Cheng, G., Liu, Y., Wang, P., & Xiao, X. (2016). Mosquito defense strategies against
viral infection. Trends in Parasitology, 32(3), 177-186. doi:10.1016/j.pt.2015.09.009
Chevillon, C., & Failloux, A. B. (2003). Questions on viral population biology to
complete dengue puzzle. Trends in Microbiology, 11(9), 415-421. doi:10.1016/s0966-
842x(03)00206-3
Clayton, A. M., Dong, Y., & Dimopoulos, G. (2014). The Anopheles innate immune
system in the defense against malaria infection. The Journal of Innate Immunity, 6(2), 169-181.
doi:10.1159/000353602
Cruz-Oliveira, C., Freire, J. M., Conceição, T. M., Higa, L. M., Castanho, M. A. R. B., &
Da Poian, A. T. (2015). Receptors and routes of dengue virus entry into the host cells. FEMS
Microbiology Reviews, 39(2), 155-170. doi:10.1093/femsre/fuu004
Dong, S., Balaraman, V., Kantor, A. M., Lin, J., Grant, D. G., Held, N. L., & Franz, A.
W. E. (2017). Chikungunya virus dissemination from the midgut of Aedes aegypti is associated
with temporal basal lamina degradation during bloodmeal digestion. PLoS Neglected Tropical
Diseases, 11(9), e0005976-e0005976. doi:10.1371/journal.pntd.0005976
Dong, S., Behura, S. K., & Franz, A. W. E. (2017). The midgut transcriptome of Aedes
aegypti fed with saline or protein meals containing chikungunya virus reveals genes potentially
involved in viral midgut escape. BMC Genomics, 18(1), 382-382. doi:10.1186/s12864-017-3775-
6
Dubrulle, M., Mousson, L., Moutailler, S., Vazeille, M., & Failloux, A.-B. (2009).
Chikungunya virus and Aedes mosquitoes: Saliva is infectious as soon as two days after oral
infection. PLoS ONE, 4(6), e5895. doi:10.1371/journal.pone.0005895
Erler, S., Popp, M., & Lattorff, H. M. G. (2011). Dynamics of immune system gene
expression upon bacterial challenge and wounding in a social insect (Bombus terrestris). PLoS
ONE, 6(3), e18126. doi:10.1371/journal.pone.0018126
Flynt, A., Liu, N., Martin, R., & Lai, E. C. (2009). Dicing of viral replication
intermediates during silencing of latent Drosophila viruses. Proceedings of the National Academy
of Sciences, 106(13), 5270-5275. doi:10.1073/pnas.0813412106
Ford, S. A., Allen, S. L., Ohm, J. R., Sigle, L. T., Sebastian, A., Albert, I., Chenoweth, S.
F., & McGraw, E. A. (2019). Selection on Aedes aegypti alters Wolbachia-mediated dengue virus
blocking and fitness. Nature Microbiology, 4(11), 1832-1839. doi:10.1038/s41564-019-0533-3
Franz, A. W. E., Sanchez-Vargas, I., Adelman, Z. N., Blair, C. D., Beaty, B. J., James, A.
A., & Olson, K. E. (2006). Engineering RNA interference-based resistance to dengue virus type 2
in genetically modified Aedes aegypti. Proceeds of the National Academy of Science USA, 103.
doi:10.1073/pnas.0600479103

114
Frentiu, F. D., Robinson, J., Young, P. R., McGraw, E. A., & O'Neill, S. L. (2010).
Wolbachia-mediated resistance to dengue virus Infection and death at the cellular level. PLoS
ONE, 5(10), e13398. doi:10.1371/journal.pone.0013398
Gaye, A., Faye, O., Diagne, C. T., Faye, O., Diallo, D., Weaver, S. C., Sall, A. A., &
Diallo, M. (2014). Oral susceptibility of Aedes aegypti (Diptera: Culicidae) from Senegal for
dengue serotypes 1 and 3 viruses. Tropical Medicine & International Health, 19(11), 1355-1359.
doi:10.1111/tmi.12373
Gokhale, M. D., Paingankar, M. S., Sudeep, A. B., & Parashar, D. (2015). Chikungunya
virus susceptibility & variation in populations of Aedes aegypti (Diptera: Culicidae) mosquito
from India. The Indian journal of medical research, 142 Suppl(Suppl 1), S33-S43.
doi:10.4103/0971-5916.176614
Gubler, D. J., Nalim, S., Tan, R., Saipan, H., & Sulianti Saroso, J. (1979). Variation in
susceptibility to oral infection with dengue viruses among geographic strains of Aedes aegypti.
The American Society of Tropical Medicine and Hygiene, 28(6), 1045-1052.
Hill, W. G. (2013). Genetic Correlation. In S. Maloy & K. Hughes (Eds.), Brenner's
Encyclopedia of Genetics (Second Edition) (pp. 237-239). San Diego: Academic Press.
Hill, W. G., Goddard, M. E., & Visscher, P. M. (2008). Data and theory point to mainly
additive genetic variance for complex traits. PLoS Genetics, 4(2), e1000008.
doi:10.1371/journal.pgen.1000008
Hill, W. G., & Mackay, T. F. C. (2004). D. S. Falconer and introduction to quantitative
genetics. Genetics, 167(4), 1529-1536.
Hoornweg, T. E., van Duijl-Richter, M. K. S., Ayala Nuñez, N. V., Albulescu, I. C., van
Hemert, M. J., & Smit, J. M. (2016). Dynamics of chikungunya virus cell entry unraveled by
single-virus tracking in living cells. Journal of Virology, 90(9), 4745-4756.
doi:10.1128/jvi.03184-15
Huang, Y.-J. S., Higgs, S., Horne, K. M., & Vanlandingham, D. L. (2014). Flavivirus-
mosquito interactions. Viruses, 6(11), 4703-4730. doi:10.3390/v6114703
Jupatanakul, N., Sim, S., Angleró-Rodríguez, Y. I., Souza-Neto, J., Das, S., Poti, K. E.,
Rossi, S. L., Bergren, N., Vasilakis, N., & Dimopoulos, G. (2017). Engineered Aedes aegypti
JAK/STAT pathway-mediated immunity to dengue virus. PLoS Neglected Tropical Diseases,
11(1), e0005187. doi:10.1371/journal.pntd.0005187
Kading, R. C., Brault, A. C., & Beckham, J. D. (2020). Global perspectives on arbovirus
outbreaks: A 2020 snapshot. Tropical medicine and infectious disease, 5(3), 142.
doi:10.3390/tropicalmed5030142
Kantor, A. M., Grant, D. G., Balaraman, V., White, T. A., & Franz, A. W. E. (2018).
Ultrastructural analysis of chikungunya virus dissemination from the midgut of the yellow fever
mosquito, Aedes aegypti. Viruses, 10(10), 571. doi:10.3390/v10100571

115
Kim, C.-H., & Muturi, E. J. (2013). Effect of larval density and Sindbis virus infection on
immune responses in Aedes aegypti. Journal of Insect Physiology, 59(6), 604-610.
doi:https://doi.org/10.1016/j.jinsphys.2013.03.010
Kokoza, V., Ahmed, A., Woon Shin, S., Okafor, N., Zou, Z., & Raikhel, A. S. (2010).
Blocking of plasmodium transmission by cooperative action of cecropin A and defensin A in
transgenic Aedes aegypti mosquitoes. Proceeds of the National Academy of Science U S A,
107(18), 8111-8116. doi:10.1073/pnas.1003056107
Kuadkitkan, A., Wikan, N., Fongsaran, C., & Smith, D. R. (2010). Identification and
characterization of prohibitin as a receptor protein mediating DENV-2 entry into insect cells.
Virology, 406(1), 149-161. doi:https://doi.org/10.1016/j.virol.2010.07.015
Lambrechts, L., Chevillon, C., Albright, R. G., Thaisomboonsuk, B., Richardson, J. H.,
Jarman, R. G., & Scott, T. W. (2009). Genetic specificity and potential for local adaptation
between dengue viruses and mosquito vectors. BMC Ecology and Evolution, 9, 160.
doi:10.1186/1471-2148-9-160
Lambrechts, L., Scott, T. W., & Gubler, D. J. (2010). Consequences of the expanding
global distribution of Aedes albopictus for dengue virus transmission. PLoS Neglected Tropical
Diseases, 4(5), e646. doi:10.1371/journal.pntd.0000646
Le Coupanec, A., Tchankouo-Nguetcheu, S., Roux, P., Khun, H., Huerre, M., Morales-
Vargas, R., Enguehard, M., Lavillette, D., Misse, D., & Choumet, V. (2017). Co-Infection of
mosquitoes with chikungunya and dengue viruses reveals modulation of the replication of both
viruses in midguts and salivary glands of Aedes aegypti mosquitoes. International Journal of
Molecular Sciences, 18(8). doi:10.3390/ijms18081708
Lee, R. C. H., Hapuarachchi, H. C., Chen, K. C., Hussain, K. M., Chen, H., Low, S. L.,
Ng, L. C., Lin, R., Ng, M. M.-L., & Chu, J. J. H. (2013). Mosquito cellular factors and functions
in mediating the infectious entry of chikungunya virus. PLoS Neglected Tropical Diseases, 7(2),
e2050-e2050. doi:10.1371/journal.pntd.0002050
Lim, E. X. Y., Lee, W. S., Madzokere, E. T., & Herrero, L. J. (2018). Mosquitoes as
suitable vectors for Alphaviruses. Viruses, 10(2). doi:10.3390/v10020084
Liu, J., Swevers, L., Kolliopoulou, A., & Smagghe, G. (2019). Arboviruses and the
challenge to establish systemic and persistent infections in competent mosquito vectors: The
interaction with the RNAi mechanism. Frontiers in Physiology, 10, 890-890.
doi:10.3389/fphys.2019.00890
Livak, K. J., & Schmittgen, T. D. (2001). Analysis of relative gene expression data using
real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods, 25(4), 402-408.
doi:10.1006/meth.2001.1262
Magwire, M. M., Fabian, D. K., Schweyen, H., Cao, C., Longdon, B., Bayer, F., &
Jiggins, F. M. (2012). Genome-wide association studies reveal a simple genetic basis of
resistance to naturally coevolving viruses in Drosophila melanogaster. PLoS Genetics, 8(11),
e1003057. doi:10.1371/journal.pgen.1003057

116
Matusali, G., Colavita, F., Bordi, L., Lalle, E., Ippolito, G., Capobianchi, M. R., &
Castilletti, C. (2019). Tropism of the chikungunya virus. Viruses, 11(2). doi:10.3390/v11020175
McFarlane, M., Arias-Goeta, C., Martin, E., O'Hara, Z., Lulla, A., Mousson, L., Rainey,
S. M., Misbah, S., Schnettler, E., Donald, C. L., Merits, A., Kohl, A., & Failloux, A.-B. (2014).
Characterization of Aedes aegypti innate-immune pathways that limit chikungunya virus
replication. PLoS Neglected Tropical Diseases, 8(7), e2994. doi:10.1371/journal.pntd.0002994
Mehta, R., Gerardin, P., de Brito, C. A. A., Soares, C. N., Ferreira, M. L. B., & Solomon,
T. (2018). The neurological complications of chikungunya virus: A systematic review. Reviews in
medical virology, 28(3), e1978-e1978. doi:10.1002/rmv.1978
Mukherjee, D., Das, S., Begum, F., Mal, S., & Ray, U. (2019). The Mosquito immune
system and the life of dengue virus: What we know and do not know. Pathogens (Basel,
Switzerland), 8(2). doi:10.3390/pathogens8020077
Muturi, E. J., Blackshear, M., Jr., & Montgomery, A. (2012). Temperature and density-
dependent effects of larval environment on Aedes aegypti competence for an alphavirus. Journal
of vector ecology : journal of the Society for Vector Ecology, 37(1), 154-161. doi:10.1111/j.1948-
7134.2012.00212.x
Novelo, M., Hall, M. D., Pak, D., Young, P. R., Holmes, E. C., & McGraw, E. A. (2019).
Intra-host growth kinetics of dengue virus in the mosquito Aedes aegypti. PLoS Pathogens,
15(12), e1008218. doi:10.1371/journal.ppat.1008218
Piontkivska, H., Matos, L. F., Paul, S., Scharfenberg, B., Farmerie, W. G., Miyamoto, M.
M., & Wayne, M. L. (2016). Role of host-driven mutagenesis in determining genome evolution of
sigma virus (DMelSV; Rhabdoviridae) in Drosophila melanogaster. Genome Biology and
Evolution, 8(9), 2952-2963. doi:10.1093/gbe/evw212
Pratt, A. J., & MacRae, I. J. (2009). The RNA-induced silencing complex: a versatile
gene-silencing machine. The Journal of biological chemistry, 284(27), 17897-17901.
doi:10.1074/jbc.R900012200
Rajapakse, S. (2011). Dengue shock. Journal of Emergencies, Trauma and Shock, 4(1),
120-127. doi:10.4103/0974-2700.76835
Ramirez, J. L., & Dimopoulos, G. (2010). The Toll immune signaling pathway control
conserved anti-dengue defenses across diverse Ae. aegypti strains and against multiple dengue
virus serotypes. Developmental & Comparative Immunology, 34. doi:10.1016/j.dci.2010.01.006
Reiskind, M. H., Pesko, K., Westbrook, C. J., & Mores, C. N. (2008). Susceptibility of
Florida mosquitoes to infection with chikungunya virus. The American Society of Tropical
Medicine and Hygiene, 78(3), 422-425.
Rodriguez, A. K., Muñoz, A. L., Segura, N. A., Rangel, H. R., & Bello, F. (2019).
Molecular characteristics and replication mechanism of dengue, zika and chikungunya
arboviruses, and their treatments with natural extracts from plants: An updated review. EXCLI
Journal, 18, 988-1006. doi:10.17179/excli2019-1825

117
Rougeron, V., Sam, I. C., Caron, M., Nkoghe, D., Leroy, E., & Roques, P. (2015).
Chikungunya, a paradigm of neglected tropical disease that emerged to be a new health global
risk. Journal of Clinical Virology, 64, 144-152. doi:10.1016/j.jcv.2014.08.032
Rückert, C., Weger-Lucarelli, J., Garcia-Luna, S. M., Young, M. C., Byas, A. D.,
Murrieta, R. A., Fauver, J. R., & Ebel, G. D. (2017). Impact of simultaneous exposure to
arboviruses on infection and transmission by Aedes aegypti mosquitoes. Nature Communications,
8, 15412. doi:10.1038/ncomms15412
Salazar, M. I., Richardson, J. H., Sánchez-Vargas, I., Olson, K. E., & Beaty, B. J. (2007).
Dengue virus type 2: replication and tropisms in orally infected Aedes aegypti mosquitoes. BMC
Microbiology, 7, 9-9. doi:10.1186/1471-2180-7-9
Sánchez-Vargas, I., Scott, J. C., Poole-Smith, B. K., Franz, A. W. E., Barbosa-Solomieu,
V., Wilusz, J., Olson, K. E., & Blair, C. D. (2009). Dengue virus type 2 infections of Aedes
aegypti are modulated by the mosquito's RNA interference pathway. PLoS Pathogens, 5(2),
e1000299. doi:10.1371/journal.ppat.1000299
Saxton, A. M. (2004). Genetic analysis of complex traits using SAS. Cary, N.C: SAS
Institute.
Scott, D. A., & Zhang, F. (2017). Implications of human genetic variation in CRISPR-
based therapeutic genome editing. Nature medicine, 23(9), 1095-1101. doi:10.1038/nm.4377
Sgrò, C. M., & Hoffmann, A. A. (2004). Genetic correlations, tradeoffs and
environmental variation. Heredity, 93(3), 241-248. doi:10.1038/sj.hdy.6800532
Shrinet, J., Srivastava, P., & Sunil, S. (2017). Transcriptome analysis of Aedes aegypti in
response to mono-infections and co-infections of dengue virus-2 and chikungunya virus.
Biochemical and Biophysical Research Communications, 492(4), 617-623.
doi:10.1016/j.bbrc.2017.01.162
Sim, S., & Dimopoulos, G. (2010). Dengue Virus Inhibits Immune Responses in Aedes
aegypti Cells. PLoS ONE, 5(5), e10678. doi:10.1371/journal.pone.0010678
Siu, R. W. C., Fragkoudis, R., Simmonds, P., Donald, C. L., Chase-Topping, M. E.,
Barry, G., Attarzadeh-Yazdi, G., Rodriguez-Andres, J., Nash, A. A., Merits, A., Fazakerley, J. K.,
& Kohl, A. (2011). Antiviral RNA interference responses induced by Semliki forest virus
infection of mosquito cells: Characterization, origin, and frequency-dependent functions of virus-
derived small interfering RNAs. Journal of Virology, 85(6), 2907-2917. doi:10.1128/jvi.02052-10
Slatkin, M. (2008). Linkage disequilibrium — understanding the evolutionary past and
mapping the medical future. Nature Reviews Genetics, 9(6), 477-485. doi:10.1038/nrg2361
Smith, D. R. (2012). An update on mosquito cell expressed dengue virus receptor
proteins. Insect Molecular Biology, 21(1), 1-7. doi:https://doi.org/10.1111/j.1365-
2583.2011.01098.x

118
Terradas, G., Allen, S. L., Chenoweth, S. F., & McGraw, E. A. (2017). Family level
variation in Wolbachia-mediated dengue virus blocking in Aedes aegypti. Parasites & Vectors,
10(1), 622-622. doi:10.1186/s13071-017-2589-3
Terradas, G., & McGraw, E. A. (2019). Using genetic variation in Aedes aegypti to
identify candidate anti-dengue virus genes. BMC Infectious Diseases, 19(1), 580-580.
doi:10.1186/s12879-019-4212-z
Tesh, R. B., Gubler, D. J., & Rosen, L. (1976). Variation among goegraphic strains of
Aedes albopictus in susceptibility to infection with chikungunya virus. The American Society of
Tropical Medicine and Hygiene, 25(2), 326-335. doi:10.4269/ajtmh.1976.25.326
Tsetsarkin, K. A., Vanlandingham, D. L., McGee, C. E., & Higgs, S. (2007). A single
mutation in chikungunya virus affects vector specificity and epidemic potential. PLoS Pathogens,
3(12), e201. doi:10.1371/journal.ppat.0030201
Vega-Rúa, A., Schmitt, C., Bonne, I., Krijnse Locker, J., & Failloux, A.-B. (2015).
Chikungunya virus replication in salivary glands of the mosquito Aedes albopictus. Viruses,
7(11), 5902-5907. doi:10.3390/v7112917
Vega-Rúa, A., Zouache, K., Girod, R., Failloux, A.-B., & Lourenço-de-Oliveira, R.
(2014). High level of vector competence of Aedes aegypti and Aedes albopictus from ten
American countries as a crucial factor in the spread of chikungunya virus. Journal of Virology,
88(11), 6294-6306. doi:10.1128/jvi.00370-14
Wang, Z., Wu, D., Liu, Y., Xia, X., Gong, W., Qiu, Y., Yang, J., Zheng, Y., Li, J., Wang,
Y.-F., Xiang, Y., Hu, Y., & Zhou, X. (2015). Drosophila Dicer-2 has an RNA interference–
independent function that modulates Toll immune signaling. Science Advances, 1(9), e1500228.
doi:10.1126/sciadv.1500228
Wilder-Smith, A., Gubler, D. J., Weaver, S. C., Monath, T. P., Heymann, D. L., & Scott,
T. W. (2017). Epidemic arboviral diseases: priorities for research and public health. The Lancet
Infectious Diseases, 17(3), e101-e106. doi:10.1016/S1473-3099(16)30518-7
Wong, H. V., Chan, Y. F., Sam, I. C., Sulaiman, W. Y., & Vythilingam, I. (2016).
Chikungunya virus infection of Aedes Mosquitoes. Methods in Molecular Biology, 1426, 119-
128. doi:10.1007/978-1-4939-3618-2_11
Xi, Z., Ramirez, J. L., & Dimopoulos, G. (2008). The Aedes aegypti Toll pathway
controls dengue virus infection. PLoS Pathogens, 4(7), e1000098.
doi:10.1371/journal.ppat.1000098
Yactayo, S., Staples, J. E., Millot, V., Cibrelus, L., & Ramon-Pardo, P. (2016).
Epidemiology of chikungunya in the Americas. The Journal of Infectious Diseases, 214(suppl 5),
S441-s445. doi:10.1093/infdis/jiw390
Ye, Y. H., Chenoweth, S. F., Carrasco, A. M., Allen, S. L., Frentiu, F. D., van den Hurk,
A. F., Beebe, N. W., & McGraw, E. A. (2016). Evolutionary potential of the extrinsic incubation
period of dengue virus in Aedes aegypti. Evolution. doi:10.1111/evo.13039

119
Ye, Y. H., Ng, T. S., Frentiu, F. D., Walker, T., van den Hurk, A. F., O'Neill, S. L.,
Beebe, N. W., & McGraw, E. A. (2014). Comparative susceptibility of mosquito populations in
North Queensland, Australia to oral infection with dengue virus. The American Society of
Tropical Medicine and Hygiene, 90(3), 422-430. doi:10.4269/ajtmh.13-0186
Zeller, H., Van Bortel, W., & Sudre, B. (2016). Chikungunya: Its history in Africa and
Asia and its spread to new regions in 2013–2014. The Journal of Infectious Diseases,
214(suppl_5), S436-S440. doi:10.1093/infdis/jiw391
Zhang, M., Zheng, X., Wu, Y., Gan, M., He, A., Li, Z., Liu, J., & Zhan, X. (2010).
Quantitative analysis of replication and tropisms of Dengue virus type 2 in Aedes albopictus. The
American Society of Tropical Medicine and Hygiene, 83(3), 700-707. doi:10.4269/ajtmh.2010.10-
0193
Zhao, L., Alto, B. W., Jiang, Y., Yu, F., & Zhang, Y. (2019). Transcriptomic analysis of
Aedes aegypti innate immune system in response to ingestion of chikungunya virus. International
Journal of Molecular Sciences, 20(13), 3133. doi:10.3390/ijms20133133
Ziegler, S. A., Nuckols, J., McGee, C. E., Huang, Y.-J. S., Vanlandingham, D. L., Tesh,
R. B., & Higgs, S. (2011). In vivo imaging of chikungunya virus in mice and Aedes mosquitoes
using a Renilla luciferase clone. Vector borne and zoonotic diseases (Larchmont, N.Y.), 11(11),
1471-1477. doi:10.1089/vbz.2011.0648

120
Chapter 5
Synthesis and General Discussion
Aedes aegypti remains as one of the deadliest animals in the world (McGregor &
Connelly, 2020). Because of this, the development of vector control methods remains as
the primary tool to fight the spread of vector borne diseases. A key part of development of
optimal and effective vector control strategies depends on a deep understanding of the
vector-virus interactions that take place when a mosquito takes pathogen-laden blood meal.
The viral infection process inside the mosquito is a complex and dynamic process that is
poorly understood. (Guzman & Harris, 2015). Additionally, the strength and efficacy of
the mosquito immunity varies across tissues and in response to different serotypes and
viruses. These vector - virus interactions can ultimately shape the transmission potential of
multiple arboviruses (Bennett et al., 2002; Gubler, Nalim, Tan, Saipan, & Sulianti Saroso,
1979).
Chapter 2 focused on comparing tissue - specific mosquito responses to DENV
infection using four DENV serotypes. Chapter 3 further expanded this work by examining
the effect of DENV serotype co-infection in tissue-specific responses to infection, for both
a Wolbachia infected and uninfected mosquito lines. Finally, in Chapter 4, we explored
the mosquito infection dynamics from a population perspective and between different

121
arboviral families. We tested the role of the Ae. aegypti own genetic variation in
determining susceptibility to DENV and CHIKV.
Differences between individual infections of viruses of the same family, in our case,
of DENV serotypes, had markedly different dynamics in our vector competence assays in
Chapter 2, DENV-1 and 2 outperform DENV-3 and 4, reaching higher viral and infection
rates. This can potentially explain the disparities in the transmission potential in the field
between the different DENV serotypes, since DENV variants with replicative advantages
can move more rapidly in susceptible vector or host population. If individual infections of
DENV serotypes have different dynamics inside the mosquito, how do they behave when
they are co-infecting the vector? moreover, how can this scenario affect the development
and deployment of vector control methods? In Chapter 3 we try to answered these
questions by analyzing the co-infection process of DENV-2 and DENV-3 inside a
Wolbachia - infected Ae. aegypti mosquito line. We saw that DENV co-infection has little
effect in Wolbachia’s ability to block pathogen replication and that Wolbachia’s blocking
phenotype was systemic across the mosquito body. From a vector control point of view,
this seems promising, since it suggests that the long-term efficacy of Wolbachia won’t be
compromised by DENV-coinfections. In Wolbachia-free mosquitoes, DENV co-infection
had a significant effect in the dynamics of each DENV serotype. While in Chapter 2,
DENV-2 ha greater capacity to replicate in the mosquito than DENV-3, when in co-
infection, DENV-3 had a competitive advantage over DENV-2, suggesting either
enhancement of DENV-3 or competitive inhibition of DENV-2, although the mechanisms
by which this occur remain unknown. These chapters support the notions that the infection

122
process inside the vector, that ultimately affects transmissibility, is variable and dynamic
and can be depended on the infecting virus.
If the infection dynamics of DENV differ across (chapter 2) and between serotypes
(chapter 3) at an individual level, how do the differences in the infectious process translate
to a populations level? And how much the mosquito population’s own genetic variation
can impact this process, when infection is caused by an arbovirus from a different viral
family? In chapter 4, we demonstrate not only that the heritability to the susceptibility to
DENV and CHIKV is different within a population but that some of the immune
mechanisms that control it are different. We showed that indeed, the mosquitoes immunity
responds differently to each virus and that although some viral control mechanisms, such
as the RNAi pathway, play a universal role in modulating viral replication, the mosquitos’
response to infection appears markedly different depending on the infecting virus.
While these experiments shed light on the G x G interaction on the susceptibility
and viral transmission potential, I was only able to test individual viral and mosquito
strains, thus extrapolating the results for other genotypes is not without limitations. Further
expedients could use our framework with a diverse arrange of mosquito and viral genotypes
incorporating different variables such as more initial infectious doses, different modes of
co-infection and different immune genes (Souza-Neto, Powell, & Bonizzoni, 2019).
Presently, my findings from chapter 2 have been further expanded not only by testing
more DENV strains per serotype but also doing with Ae. albopictus, in the work done by
Ekwudu et al. (2020). Additionally, the tissue specific infection dynamics and the presence
of a viral subpopulation structure caused by a dose-dependent effect was reported in

123
CHIKV infected mosquitoes by Merwaiss et al. (2021). Similarly, chapter’s 3 results are
in agreement with the work done by Caragata et al. (2019) showing that co-infection does
not adversely affect pathogen blocking caused by Wolbachia, currently being trialed
worldwide.
The selection for viral genotypes that have the potential for rapid and explosive
spread is actively driven by withing vector selection (Chevillon & Failloux, 2003). Single
adaptive mutations within the viral genotype have been linked to the re-emergence and
spread of viruses like CHIKV. Viruses with such selective advantages can displace local
and more mild viral genotypes of the same or different viral families (Tsetsarkin,
Vanlandingham, McGee, & Higgs, 2007). Outcompeted genotypes tend to be milder
clinical manifestations and slow disease transmission within mosquito populations. How
much does the viral populations reflect the genetic structure of their vectors? Worldwide,
Ae. aegypti populations consists of connected islands of genetically different populations
(Apostol, Black, Reiter, & Miller, 1996). The vector competence within a population is, up
to some degree, determine by the G x G interactions, therefore viral genotype structure is
a result from adaptations to the vector (Lambrechts et al., 2009). The transmission potential
of a virus is therefore dependent on the vector-virus compatibility, i.e., the combination of
initial viral load, host and pathogen genotypes.
Through my work, I demonstrate the importance of the viral genetic diversity,
initial viral dose and tissues - specific response in dictating infectivity. We showed that a
threshold level of virus is required to stablish body-wide infection and that this threshold
might be different between DENV serotypes. We also showed that the pathogen blocking

124
effect in Wolbachia-infected mosquitoes is not affected by DENV serotype co-infection.
Finally, we showcase the contribution of mosquito’s genetic variation in the differential
susceptibility to infection between DENV and CHIKV. Taken together, my dissertation
addresses different areas of the complex vector-virus interactions and their effect on
pathogen transmissibility. While many unanswered questions remained, my research
provides valuable insight into the infection dynamics, immunity and feasibility of vector
control strategies.

125
5.1 References
Apostol, B. L., Black, W. C., Reiter, P., & Miller, B. R. (1996). Population
genetics with RAPD-PCR markers: the breeding structure of Aedes aegypti in Puerto
Rico. Heredity, 76(4), 325-334. doi:10.1038/hdy.1996.50
Bennett, K. E., Olson, K. E., Munoz Mde, L., Fernandez-Salas, I., Farfan-Ale, J.
A., Higgs, S., Black, W. C., & Beaty, B. J. (2002). Variation in vector competence for
dengue 2 virus among 24 collections of Aedes aegypti from Mexico and the United
States. American Journal of Tropical Medicine and Hygiene, 67.
Caragata, E. P., Rocha, M. N., Pereira, T. N., Mansur, S. B., Dutra, H. L. C., &
Moreira, L. A. (2019). Pathogen blocking in Wolbachia-infected Aedes aegypti is not
affected by Zika and dengue virus co-infection. PLOS Neglected Tropical Diseases,
13(5), e0007443-e0007443. doi:10.1371/journal.pntd.0007443
Chevillon, C., & Failloux, A. B. (2003). Questions on viral population biology to
complete dengue puzzle. Trends in Microbiology, 11(9), 415-421. doi:10.1016/s0966-
842x(03)00206-3
Ekwudu, O. m., Marquart, L., Webb, L., Lowry, K. S., Devine, G. J., Hugo, L. E.,
& Frentiu, F. D. (2020). Effect of serotype and strain diversity on dengue virus
replication in Australian mosquito vectors. Pathogens (Basel, Switzerland), 9(8), 668.
doi:10.3390/pathogens9080668
Gubler, D. J., Nalim, S., Tan, R., Saipan, H., & Sulianti Saroso, J. (1979).
Variation in susceptibility to oral infection with dengue viruses among geographic strains
of Aedes aegypti. American Journal of Tropical Medicine and Hygiene, 28(6), 1045-
1052.
Guzman, M. G., & Harris, E. (2015). Dengue. The Lancet, 385(9966), 453-465.
doi:https://doi.org/10.1016/S0140-6736(14)60572-9
Lambrechts, L., Chevillon, C., Albright, R. G., Thaisomboonsuk, B., Richardson,
J. H., Jarman, R. G., & Scott, T. W. (2009). Genetic specificity and potential for local
adaptation between dengue viruses and mosquito vectors. BMC Ecology and Evolution,
9, 160. doi:10.1186/1471-2148-9-160
McGregor, B. L., & Connelly, C. R. (2020). A Review of the Control of Aedes
aegypti (Diptera: Culicidae) in the Continental United States. Journal of Medical
Entomology, 58(1), 10-25. doi:10.1093/jme/tjaa157
Merwaiss, F., Filomatori, C. V., Susuki, Y., Bardossy, E. S., Alvarez, D. E., &
Saleh, M. C. (2021). Chikungunya virus replication rate determines the capacity of

126
crossing tissue barriers in mosquitoes. Journal of Virology, 95(3). doi:10.1128/jvi.01956-
20
Souza-Neto, J. A., Powell, J. R., & Bonizzoni, M. (2019). Aedes aegypti vector
competence studies: A review. Infection, Genetics and Evolution, 67, 191-209.
doi:https://doi.org/10.1016/j.meegid.2018.11.009
Tsetsarkin, K. A., Vanlandingham, D. L., McGee, C. E., & Higgs, S. (2007). A
single mutation in chikungunya virus affects vector specificity and epidemic potential.
PLoS Pathogens, 3(12), e201. doi:10.1371/journal.ppat.0030201

127
Appendices
Appendix A: Supplementary materials from Chapter 2
Appendix A: DRC estimates for unsuccessful infections by tissue, infectious dose and DENV
strain
Infectious
dose Serotype Tissue
Subpopulation
threshold Parameter Estimate SE
p-value Sig
LOW
DENV-1
MIDGUT
Unsuccessful
Growth
rate 1.25 0.81
0.12
Max
DENV
load
1.96 0.14
<0.001 ***
ED50 1.14 0.69 0.1
CARCASS
Growth
rate 0.13 0.15
0.38
Max
DENV
load
1.64 0.58
0.001
ED50 1.91 5.16 0.71
DENV-2
MIDGUT
Growth
rate 0 0.03
0.79
Max
DENV
load
6.56 8.63
0.81
ED50 1.14 0.68 1.67
CARCASS
Growth
rate 0.04 NA
NA
Max
DENV
load
4.04 NA
NA
ED50 0 NA NA
HIGH DENV-3
MIDGUT
Growth
rate -0.03 0.03
0.93
Max
DENV
load
6.22 9.66
0.64
ED50 0 78.7 1
LOW DENV-3 NTA
Growth
rate -0.3 0.17
0.08
Max
DENV
load
1.85 0.26
<0.001 ***
ED50 17.1 1.84 <0.001 ***

128
HIGH DENV-3
CARCASS
Unsuccessful
Growth
rate 0.31 0.13
0.02
Max
DENV
load
2.96 0.28
<0.001 ***
ED50 4.56 1.09 <0.001 ***
LOW DENV-3 NTA
Growth
rate -0.02 0.03
0.49
Max
DENV
load
2.44 4.99
0.62
ED50 0 90.7 1
HIGH DENV-4
MIDGUT
Unsuccessful
Growth
rate -0.04 .03
0.1552
Max
DENV
load
4.93 0.19
<0.001 ***
ED50 0 0.3 1
LOW DENV-4 NTA
Growth
rate 0.15 0.3
0.6
Max
DENV
load
1.38 0.52
0.01 *
ED50 0 .55 1
HIGH DENV-4
CARCASS
Unsuccessful
Growth
rate 0.02 0.06
0.74
Max
DENV
load
3.63 20.4
0.85
ED50 17.12 745 0.9
LOW DENV-4 NTA
Growth
rate 0.64 0.43
0.14
Max
DENV
load
1.31 0.16
<0.001 ***
ED50 7.25 2.17 0.001 **
*NA, not available due to poor fitting to the log logistic model. *NTA no threshold applied. *P<0.05,
**P<0.01, ***P<0.001. SE of the estimate, p-value indicates if estimate is different than zero.

129
Appendix A: Multimodal distribution analysis
Treatments Hartigan’s dip test Bimodality coefficient
DENV-1 CA LOW <2.2e-16 0.81
DENV-1 MG LOW 2.60E-05 0.86
DENV-2 CA LOW 0.001386 0.72
DENV-2 MG LOW 4.24E-05 0.79
DENV-3 CA HIGH 7.32E-06 0.73
DENV-3 MG HIGH <2.2e-16 0.75
DENV-4 CA HIGH 5.18E-06 0.89
DENV-4 MG HIGH <2.2e-16 0.93
DENV-1 CA HIGH 7.83E-06 0.67
DENV-1 MG HIGH 0.5921 0.63
DENV-2 CA HIGH 0.0001161 0.75
DENV-2 MG HIGH 0.9904 0.63
If the p value is >0.05, it suggests bimodality.
Appendix A: Survival curve statistical analysis
Treatment N Observed Expected (O-E)^2/E (O-E)^2/V
Control 80 80 88.5 0.82 1.06
DENV-1 High 80 80 98.7 3.52 4.93
DENV-1 Low 80 80 75.2 0.31 0.39
DENV-2 High 80 80 64.7 3.62 4.44
DENV-2 Low 80 80 86.4 0.47 0.60
DENV-3 High 80 80 81.1 0.02 0.03
DENV-3 Low 80 80 82.1 0.05 0.07
DENV-4 High 80 80 68.7 1.85 2.29
DENV-4 Low 80 80 74.3 0.43 0.54
Chisq = 12.9 on 8 degrees of freedom, p= 0.01.

130
Appendix A: AIC values for the DRC model
Serotype Dose Tissue Subpopulation threshold AIC
DENV-1 High MG NA 967
DENV-1 Low MG Successful 318
DENV-1 Low MG Unsuccessful 341
DENV-2 High MG NA 775
DENV-2 Low MG Successful 386
DENV-2 Low MG Unsuccessful 360
DENV-3 High MG Successful 392
DENV-3 High MG Unsuccessful 306
DENV-3 Low MG NA 263
DENV-4 High MG Successful 605
DENV-4 High MG Unsuccessful 228
DENV-4 Low MG NA 282
DENV-1 High CA NA 786
DENV-1 Low CA Successful 181
DENV-1 Low CA Unsuccessful 300
DENV-2 High CA NA 621
DENV-2 Low CA Successful 199
DENV-2 Low CA Unsuccessful 340
DENV-3 High CA Successful 340
DENV-3 High CA Unsuccessful 301
DENV-3 Low CA NA 160
DENV-4 High CA Successful 362
DENV-4 High MG Unsuccessful 163
DENV-4 Low MG NA 189
Appendix A: Results of the GLM fitted to salivary glands DENV load
Estimate SE t-value P=value
Intercept 1.06 2.36 0.44 0.66
DENGUE-2 41.94 75.43 0.55 0.58
DENGUE-3 0.14 2.41 0.05 0.95
DENGUE-4 -29.65 20.55 -1.44 0.16
Carcass load 0.13 0.29 0.44 0.66
Midgut load 0.05 0.24 0.21 0.83
DENGUE-2:Carcass load -4.69 8.34 -0.56 0.58
DENGUE-3:Carcass load -0.03 0.30 -0.13 0.89
DENGUE-4:Carcass load 6.15 =4.24 1.44 0.16
DENGUE-2:Midgut load -3.50 6.34 -0.55 0.58
DENGUE-3:Midgut load -0.01 0.24 -0.07 0.94
DENGUE-4:Midgut load 3.02 2.09 1.44 0.16
Carcass load:Midgut load -0.01 0.03 -0.33 0.73
DENGUE-2:Carcass load:Midgut load 0.39 0.70 0.56 0.58
DENGUE-3:Carcass load:Midgut load 0.008 0.03 0.27 0.78
DENGUE-4:Carcass load:Midgut load -0.62 0.42 -1.45 0.16
131
Appendix A: Density plots. Dotted line indicates data split point
DENV-1 MG LOW DENV-2 MG LOW
DENV-1 CA LOW DENV-2 CA LOW
DENV-3 MG HIGH DENV-4 MG HIGH

132
DENV-3 CA HIGH DENV-4 CA HIGH
DENV-1 CA HIGH DENV-2 CA HIGH
DENV-1 MG HIGH DENV-2 MG HIGH

133
Appendix A: Average salivary gland DENV load at the last DPI (20 days).
Average DENV load at DPI 20 for all 4 DENV serotypes at 2 infectious doses (High: 1 × 108, low:
1 × 105 DENV copies/ml). All box plots show median and interquartile ranges (n = 10 per
treatment). Significant differences are based on Tukey post hoc comparison following ANOVAs
on log-transformed data. Only significant differences are shown. *p<0.05.

134
Appendix A: Survival curves for all treatments
Survival curve analysis of control and the 4 DENV serotypes at 2 infectious doses (High: 1 × 108,
low: 1 × 105 DENV copies/ml). There was no significant difference between treatments (χ2 = 12.9,
df = 8, p = 0.1). Dotted outside lines represent 95% confidence intervals. Each line at the top
represents individual treatments.
Treatment

135
Appendix B: Supplementary materials from Chapter 3
Appendix B: Supplementary figure 1. Relationship between PFU and viral RNA copies
for both DENV-2 and DENV-3
Relationship between PFU and viral RNA copies for both DENV-2 and DENV-3. Viruses were
harvested from C6/36 cells at DPI 5 and 7 and on the day of the experiment (DPI7-Exp). A) PFU/ml
for both DENV-2 and DENV-3 for DPI5 (red), DPI7 (blue), and on the day of the experiment DPI7
(black). B) Log10 viral RNA copies/ml for both DENV-2 and DENV-3 for DPI5 (red), DPI7 (blue),
and on the day of the experiment DPI7 (black). All assays were done using live virus.

136
Appendix C: Supplementary materials from Chapter 4
Appendix C: Supplementary figure 1. Relationship between DENV viral load and gene
expression levels.
Relationship between DENV viral load and gene expression levels in extreme DENV load
mosquito families. DENV viral load was correlated with expression levels of A) AGO2 (Spearmans
r = 0.63, p = 0.05), B) DCR2 (Spearmans r = 0.0022, p = 0.95), C) TEP3 (Spearmans r = -0.46, p
= 0.18) and D) CECH (Spearmans r = -0.35, p = 0.32). Each point represents a mosquito family.
N=10 families.

137
Appendix C: Supplementary figure 3. Relationship between DENV viral load and gene
expression levels.
Relationship between CHIKV viral load and gene expression levels in extreme CHIKV load
mosquito families. CHIKV viral load was correlated with expression levels of A) AGO2
(Spearmans r = 0.7, p = 0.016), B) DCR2 (Spearmans r = 0.64, p = 0.035), C) TEP3 (Spearmans r
= 0.76, p = 0.0061) and D) CECH (Spearmans r = 0.62, p = 0.043). Each point represents a mosquito
family. N=10 families.

VITA
Mario de Jesús Novelo Canto
Education
Publications
• Novelo M, Hall MD, Pak D, Young PR, Holmes EC, McGraw EA. Intra-host
kinetics of dengue virus in the mosquito Aedes aegypti. PLOS Pathogens.
2019;15(12): e1008218. Novelo M, Audsley M, McGraw EA.
• Novelo M, Audsley M, McGraw EA. The impact of DENV serotype competition
and co-infection viral kinetics in the mosquito Aedes aegypti. Parasites & Vectors.
2021. In press.
Awards
• Jose de la Tore Endowment Opportunity for Graduate students from Latin America
and the College of Agricultural Sciences Graduate Student Travel Award - Penn
State (USA).
• Dean's Postgraduate Research Scholarship and Deans International Postgraduate
Research Scholarship - Monash University (AUS).
• National Council of Science and Technology Scholarship - CONACyT (MEX)
Teaching experience
• Teaching assistant: ENT202 The Insect Connection (Spring 2018)
• Teaching assistant: ENT216 Plagues Through the Ages (Spring 2020)
2018 – 2021 PhD in Entomology
Penn State University, University Park, PA, USA
2014 – 2016 Master of Science in Molecular Medicine (Distinction)
University of Sheffield, United Kingdom
2009 – 2014 Bachelor of Science in Biology
Universidad Autónoma de Yucatán, México