arbuscular in tibet
TRANSCRIPT

7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 17
MycorrhizaDOI 101007s00572-005-0031-8
ORIGINAL PAPER
J P Gai X B Cai G Feng P Christie X L Li
Arbuscular mycorrhizal fungi associated with sedgeson the Tibetan plateau
Received 4 May 2005 Accepted 28 October 2005 Springer-Verlag 2005
Abstract The arbuscular mycorrhizal (AM) status of ninedominant sedge species and the diversity of AM fungi inTibetan grassland were surveyed in the autumn of 2003 and
2004 Most of the sedge species and ecotypes examinedwere mycorrhizal but Carex moorcroftii and Kobresia pusilla were of doubtful AM status and Kobresia humiliswas facultatively mycorrhizal This is the first report of themycorrhizal status of eight of the nine sedge speciesexamined Intraradical vesicles and aseptate hyphae werethe structures most frequently observed Appressoria coilsand arbuscules were found in the roots of a few sedgespecies A strong negative correlation was found betweensoil organic matter content and the extent of mycorrhizalcolonization Using trap cultures 26 species of AM fungi belonging to six genera Glomus Acaulospora Paraglomus Archaeospora Pacispora and Scutellospora were isolated
from the soil samples collected The frequency of occur-rence of different taxa of AM fungi varied greatly Glomusand Acaulospora were the dominant genera and Acaulospora scrobiculata was the most frequent and abundant species The species richness of AM fungi was 273 in the study areaSpecies richness and diversity index differed among thesedge species but were not correlated with soil factors such as pH available P or organic matter content
Keywords Sedges Arbuscular mycorrhiza Fungalstructure Grassland Species diversity Tibet
Introduction
About 80 of terrestrial plant species in natural ecosys-tems are mycorrhizal and the associations may improvethe fitness of the fungal and plant partners (Smith andRead 1997 ) Ecologists have taken increasing interest in theincidence and ecological role of arbuscular mycorrhizal(AM) fungi in natural plant communities in recent years(Bever 2002 Bever et al 2001 Burrows and Pfleger 2002 )Despite the occurrence of mycorrhiza in the great majorityof modern plant taxa and in almost all ecosystems there arestill some plant families such as Cyperaceae Brassicaceae
Caryophyllaceae Juncaceae and Amaranthaceae presumedto be nonmycorrhizal or rarely mycorrhizal (Newman andReddell 1987 Peterson and Bradbury 1995 Hirsch andKapulnik 1998 Regvar et al 2003 Fuchs and Haselwandter 2004 ) The Cyperaceae have become particularly promi-nent in recent years for the many conflicting reports ontheir mycorrhizal status
Sedges often dominate wetlands and arctic and alpinevegetation in which the mycorrhizal inoculum is often lowor absent and were therefore often designated nonmycor-rhizal (Harley and Harley 1987 Tester et al 1987 ) How-ever Muthukumar et al ( 2004 ) recently reviewed thecurrent information on mycorrhizal associations in sedges
and showed that the family can no longer be considerednonmycorrhizal Indeed the mycorrhizal status of itsmembers seems to be greatly influenced by environmentalconditions Since 1987 information has become availablefor 221 sedge species indicating that 88 (40) are mycor-rhizal 109 (49) are nonmycorrhizal and 24 (11) arefacultatively mycorrhizal There are still many sedge spe-cies that remain to be investigated and detailed informa-tion on the AM fungal composition and communitydiversity in sedges is still unavailable Further functionalaspects of the association in the extreme environments that sedges often inhabit remain unclear
J P Gai G Feng P Christie X L Li ( )Department of Plant Nutrition China Agricultural University2 Yuanmingyuan RoadBeijing 94 Chinae-mail lixlcaueducnTel +86-10-62731325Fax +86-10-62731016
X B CaiCollege of Agricultural and Animal HusbandryUniversity of TibetLinzhi 860000 China
P ChristieAgricultural and Environmental Science DepartmentQueens University Belfast Newforge Lane Belfast BT9 5PX UK
7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 27
Tibet (26deg44 primendash 36deg32 prime N 78deg25 primendash 99deg06 prime E) is the worldslargest and highest plateau The average altitude is about 4500 m above sea level and the weather differs sharplyduring day and night Because of the different geographicalconditions contrasting temperatures are found in the northand south The north has a continental climate and thesouth is relatively warm and rainy Grassland is veryimportant because of the harsh geographical and climaticconditions occupying 8207 million hectares of the Tibetan plateau The main types of grassland are alpine steppesalpine meadow grasslands alpine meadows and montanescrub Poaceae Cyperaceae and Asteraceae are the threemost important plant families in terms of abundanceherbage quality productivity and distribution for pastoralagriculture Most of the sedges especially Kobresia occur in the high quality pasture and information is required onthe mycorrhizal status and ecological role of AM as-sociations in sedges However there are no reports on themycorrhizal status of sedges on the Tibetan plateau
The objectives of the present study were to determine themycorrhizal status of the dominant sedge species Kobresiahumilis Kobresia prainii Kobresia pygmaea and Carexmoorcroftii and to assess the diversity of AM fungi in therhizosphere of sedges in Tibetan grassland The relation-ships between the extent of AM colonization and soil
properties (soil pH organic matter content and available P)were also investigated
Materials and methods
Sampling area and procedures
The investigation was conducted in the prefectures of Lhasa Rikaze Shannan and Naqu (29deg19 primendash 32deg52 prime N 88deg57 primendash 92deg20 prime E) Soils and roots were sampled in the grass-lands at altitudes ranging from 3798 to 5220 m above sealevel The climate of this region from north to south variesfrom alpine plateau to a semiarid type of temperate climateThe mean annual temperature is minus 12 to 8degC and annual precipitation is about 300 ndash 450 mm
Vegetation types are mainly montane scrub alpinesteppe alpine meadow grassland and alpine meadow Thedominant sedge species in the different types of grasslandwere sampled in this study In the autumn of 2003 and2004 22 soil samples were collected from the rhizosphereof nine sedge species in the main grasslands The sedgespecies selected are often dominant and scattered amongdense grasses in the sampling areas Three or more plantswere dug out together from each sampling point and
Table 1 Sedges examined vegetation type altitude soil characteristics and AM status of Tibetan sedges
Host plants Landtype
Altitude(m)
Soiltexture
Organicmatter (gkg)
Olsen-P(mgkg)
pH AM fungalstructures
Colonization()
Sporedensity(per 20 g)
Carex capillacea var linzensis MS 3980 Loam 547 523 674 ar v h 303plusmn85 66
C capillacea varlinzensis
LM 3977 Loam 151 1035 647 v h 318plusmn102 10
C moorcroftii AMG 5220 Sandy loam 182 1499 667 v h 227plusmn59 30C moorcroftii AT 4510 Gravel soil 368 1708 819 v h 221plusmn41 79C tibetica MS 3798 Sand 009 6216 755 v h c 309plusmn123 17C compressus AM 4827 Loam 360 665 603 ar v h 253plusmn67 10C compressus AM 4827 Loam 546 525 601 ar v h 177plusmn20 120C compressus AM 4394 Loam 211 636 626 ar v h 513plusmn157 48C compressus AM 4857 Loam 829 844 602 ar v h 201plusmn32 23C compressus AM 4827 Loam 836 515 61 ar v h 199plusmn19 42 K humilis AM 4385 Loam 251 152 703 ar v h 342plusmn101 58 K humilis AMG 5138 Sand 258 967 686 v h 379plusmn38 16 K humilis AM 4789 Loam 1156 1245 697 ND 0 14 K humilis AS 4559 Loam 209 1817 675 ap v h 427plusmn147 72 K humilis AM 4455 Loam 357 1145 637 v h 352plusmn89 24 K prainii AMG 4897 Loam 813 964 715 ap ar v h 650plusmn164 16 K prainii AS 4543 Gravel soil 082 178 739 ap v h 300plusmn29 70 K pusilla AMG 4717 Loam 778 1102 792 v h 61plusmn14 87 K pusilla AT 4837 Loam 306 749 661 v h 481plusmn129 24 K pygmaea AMG 5006 Gravel soil 036 294 818 ap h 106plusmn19 189 K pygmaea AS 4603 Silty soil 328 514 848 ar v h 153plusmn32 8 K tibetica AMG 4738 Sand 176 925 811 ar v h 560plusmn27 16
AM Arbuscular mycorrhizal Ap appressoria ar arbuscules h intercellular aseptate hyphae v vesicles c coils ND not detected AMG alpinemeadow grassland AS alpine steppe AM alpine meadow MS montane scrub LM lowland meadow
7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 37
approximately 2 kg soil was collected from the rootingzone of the sedges to a depth of 20 cm Care was taken toavoid contamination from other plants The soil sampleswere taken back to the laboratory and soil texture and soil
pH were determined immediately After air-drying the soil
samples were passed through a 2-mm sieve and stored at room temperature Soil available P and organic matter content were determined using the methods described byLu (2000 )
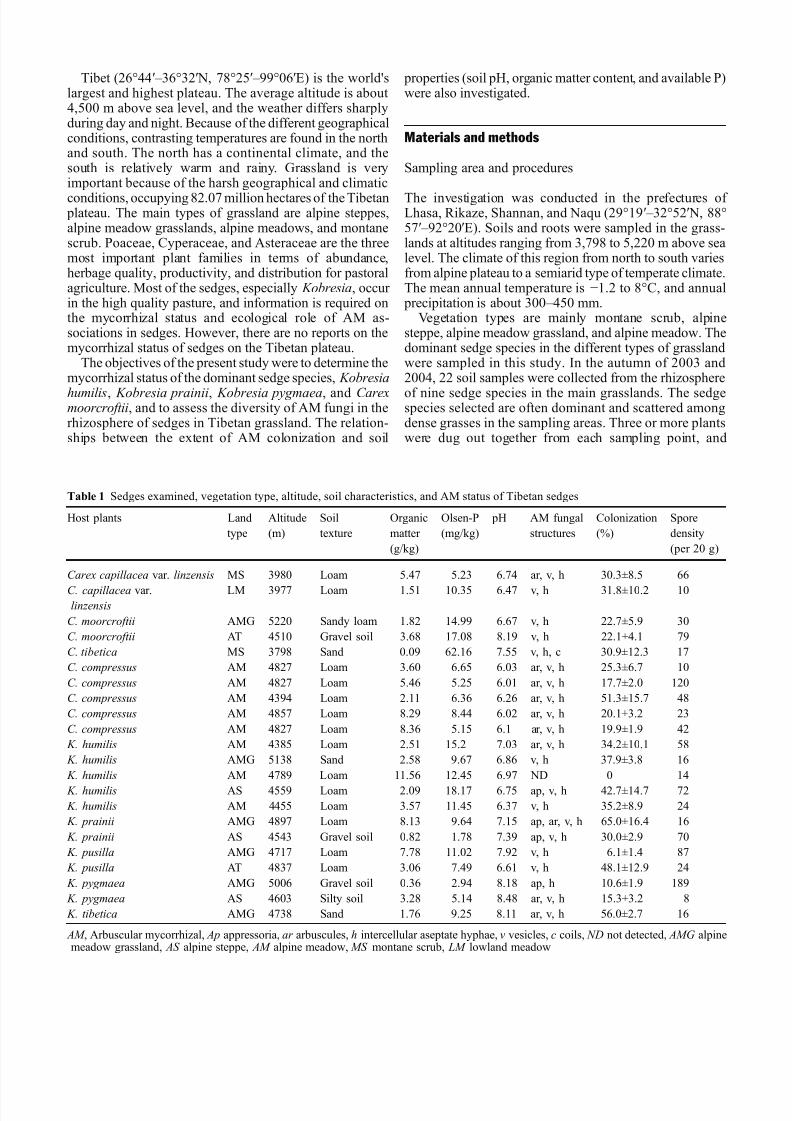
Fig 1 Fungal colonization inTibetan grassland sedgesa Arbuscule (a) intraradicalhyphae and vesicle (v) in root of K prainii b Intraradicalhyphae and vesicle (v) in cellsof C moorcroftii c Vesicle (v)and intraradical hyphae in cellsof K humilis d Extramatricalspores (s) attached to K pygmaea root e Intraradicalhyphae and vesicle (v) in root of Carex capillacea varlinzensis f Hyphal coil (hc)formation on C tibetica rootg Intraradical hyphae (h) andvesicle in root of C compressus h Appressorium (ap) formationin K pygmaea Bar=50 μ m(a d h ) bar=100 μ m (b c f ) bar=150 μ m (e g)
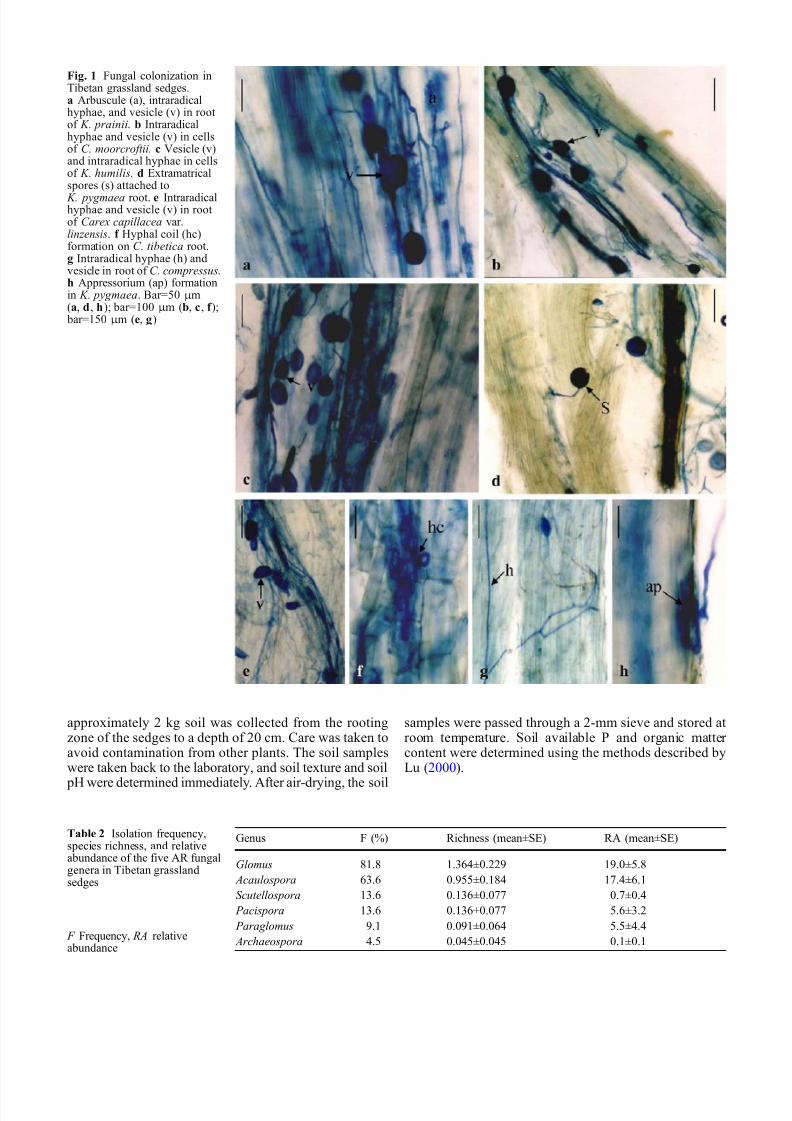
Table 2 Isolation frequencyspecies richness and relativeabundance of the five AR fungalgenera in Tibetan grasslandsedges
F Frequency RA relativeabundance
Genus F () Richness (meanplusmnSE) RA (meanplusmnSE)
Glomus 818 1364plusmn0229 190plusmn58 Acaulospora 636 0955plusmn0184 174plusmn61Scutellospora 136 0136plusmn0077 07plusmn04 Pacispora 136 0136plusmn0077 56plusmn32 Paraglomus 91 0091plusmn0064 55plusmn44 Archaeospora 45 0045plusmn0045 01plusmn01
7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 47
Root staining
Roots were gently washed with water to remove adheringsoil and carefully separated according to their morpholog-ical characteristics to avoid misinterpretation Washed root samples were cleared in 10 (wv) KOH for 30 min at 90degC acidified in 2 (vv) lactic acid for 10 min andstained for 30 min at 90degC with 005 (wv) Trypan blue(Phillips and Hayman 1970 ) Roots that remained dark after clearing were bleached in alkaline H 2 O2 prior toacidification with lactic acid Fifty 05- to 1-cm root fragments from each sample were examined under acompound microscope (100 ndash 400times) for AM fungal struc-tures The presence of arbuscules was used to designateAM associations (Brundrett 1991 ) and the presence of anAM association was considered doubtful when arbusculeswere absent The proportion of root length colonized wasestimated by the root slide technique (Brundrett et al 1996 )
Spore extraction and counting
Spores were isolated from air-dried soil using the methodof Daniels and Skipper ( 1982 ) Twenty grams of soil takenfrom each sample was suspended in 1 l water and left tostand for 20 min The suspensions were passed throughnested 500- and 45- μ m sieves and wet sieved followed bysucrose density gradient centrifugation The AM fungalspores were counted on a grid-patterned dish under a binocular stereomicroscope
Culture establishment
Trap cultures were established from fresh soil samplesmixed with autoclaved sand in a ratio of 21 Clover (Trifolium repens L) and sorghum ( Sorghum vulgare Pers)were used as host plants Cultures were grown in agreenhouse at China Agricultural University for 5 monthswith a temperature regime of 28degC (day) and 15degC (night)and a 14-h photoperiod at a light intensity of 250 μ mol m
minus2
sminus2
provided by supplementary illumination
Identification of AM fungi
Spores of AM fungi isolated from the field soils and trap
cultures were mounted on glass slides in polyvinyl ndash
lacto ndash
glycerol (PVLG) or PVLG + Melzers reagent (11 vv)The spores were examined microscopically and identifiedaccording to current taxonomic criteria (Schenck and Peacutereacutez1990 ) and using the Internet information from the INVAMwebsite ( http invamcafwvuedu )
Data analysis
AM fungal diversity was evaluated by spore density rel-ative abundance isolation frequency (F) and species
richness Spore density was expressed as numbers of AMfungal spores per 20 g dried field soil Relative abundancewas defined as the percentage of numbers of spores of the particular species or genera in the field soil Data from fieldsoil and trap cultures were combined to determine speciescomposition Isolation frequency was calculated as the percentage of samples in which the particular genus or species was present Species richness was defined asnumbers of AM fungal species per soil sample Speciesdiversity was measured by the Shannon ndash Weiner index asfollows
Shannon Weiner index frac14 X Pi ln Pifrac12 eth THORN
where
Pi frac14 ni =N
and n i = number of spores in species i and N = total sporenumber of all species
Results
Mycorrhizal status of sedges
AM fungal structures were observed in all the sedge rootsexamined except for one ecotype of K humilis (Table 1Fig 1) At species level six species in our survey weremycorrhizal K humilis was facultatively mycorrhizal andC moorcroftii and Kobresia pusilla were of doubtfulmycorrhizal status The colonization pattern varied amongthe plant species and even among the ecotypes of individual
species Vesicles and intercellular aseptate hyphae were themost frequently observed structures present in the sedgeroots examined Vesicles were observed in 20 root samples(90) whereas arbuscules were observed in only 10 sam- ples (46) Appressoria were present in K humilis sampledfrom alpine steppe K prainii from both alpine meadowgrassland and alpine steppe and K pygmaea from alpinemeadow grassland Intracellular coils were found only inCarex tibetica No AM fungal structures were observed inthe root sample of K humilis from alpine meadow
Colonization levels in sedges varied from 0 to 65spore density from 8 to 189 per 20 g dried soil and bothvaried greatly with plant species and ecotype (Table 1)
Table 3 Isolation frequency ( ge 15) and relative abundance of arbuscular mycorrhizal fungal species
Species F () RA (meanplusmnSE)
A laevis 227 35plusmn16 A scrobiculata 318 58plusmn22G claroideum 182 53plusmn42G intraradices 227 24plusmn15G mosseae 273 28plusmn15
F Isolation frequency RA relative abundance
7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 57
Further Pearsons correlation analysis of colonization lev-els and soil factors (soil pH organic matter and availableP) revealed a strong negative correlation between organicmatter and colonization rate ( r = minus 0733 P lt000 n=22)
AM fungal diversity
Twenty-six taxa of AM fungi were isolated from the soilsamples belonging to Glomus Acaulospora Paraglomus Archaeospora Pacispora and Scutellospora (Table 2)Species richness in the survey areas was 273 Glomus and Acaulospora were the dominant genera both in frequencyand relative abundance The five most commonly observedspecies were Acaulospora scrobiculata Glomus mosseae Glomus intraradices Acaulospora laevis and Glomusclaroideum Of these A scrobiculata was the most fre-quent and abundant species (Table 3)
The species richness and fungal diversity of nine sedge
species differed substantially (Table 4) Species richnessvaried from 10 ( C tibetica ) to 45 ( K prainii ) and diversityfrom 0070 ( Kobresia tibetica ) to 0252 ( K pygmaea ) Norelationship existed between AM fungal diversity (diversityindex spore density) and soil factors (soil pH organicmatter and available P)
Discussion
Here we report for the first time the mycorrhizal status of eight of the nine sedge species investigated Only Cyperuscompressus has previously been reported as mycorrhizal
(Allsopp and Stock 1993 Muthukumar and Udaiyan 2000 )and was defined as facultatively mycorrhizal whenMuthukumar et al ( 2004 ) summarized the published in-formation on the mycorrhizal status of sedges C compres- sus was mycorrhizal in all ecotypes collected in our studyOnly intercellular aseptate hyphae and vesicles were foundin C moorcroftii and K pusilla It has been reported that vesicles and hyphae are found in nonmycorrhizal hosts suchas Cyperus iria and Cyperus rotundus (Koske et al 1992 Giovannetti and Sbrana 1998 ) These plants can therefore bereferred to as of doubtful AM status K humilis was found to be facultatively mycorrhizal in our survey The incidence of
mycorrhizal species in the present study was much higher (67) than the average level of 40 reported Tibetansedges therefore appear to be readily colonized by AMfungi
Vesicles and intercellular aseptate hyphae were the most frequent structures present in the sedge roots and thisaccords with earlier reports (Muthukumar et al 2003 Muthukumar and Udaiyan 2000 ) The incidence of ar- buscules was 46 not a very low value compared withother surveys (Muthukumar et al 2003 Muthukumar andUdaiyan 2000 ) However the percentage of root lengthcolonized by AM fungi was very low in most root samplesand hyphal coils and appressoria were found in only a fewspecimens The potential nutritional benefit of the mycor-rhizal association in sedges therefore has yet to be as-certained More detailed studies are necessary to ascertainthe exact degree of mycorrhizal dependency of Cyperaceaeand the functional aspects of the association in theseTibetan grassland communities
The rarely colonized or nonmycorrhizal status of sedgesis often attributed to the small amount or absence of mycorrhizal propagules in the soils that sedges inhabit (Peat and Fitter 1993 ) However many sedges tend to benonmycorrhizal even in the presence of AM fungal prop-agules (Van der Heijden et al 1998 ) indicating that absence of AM fungal inoculum may not always explainthe low incidence of AM in these plants Spore densities inthe rhizosphere of sedges were determined in our studyand there was no correlation between spore density andcolonization rate Other important factors likely to influ-ence root colonization are soil properties A strong negativecorrelation was found between soil organic matter content and root colonization rate This is in agreement with theanalysis of Muthukumar et al ( 2004 ) However root
colonization was not related to important factors such assoil pH or soil available P which are shown to influence themycorrhizal status of sedges (Lovera and Cuenca 1996 Muthukumar et al 2004 ) One likely explanation could bethe narrow data range in a particular habitat of this studyThe mechanisms by which soil properties and other factorsinfluence mycorrhizal associations still remain unclear andrequire more detailed research
AM fungi are known to exhibit ecological specificity(McGonigle and Fitter 1990 ) There are no published reportson the AM fungal community associated with sedges under the climatic conditions prevailing on the Tibet plateau Onlya few studies have addressed the ability of AM fungi to
survive winter freezing or drought stress (van der Heijden et al 1998 Addy et al 1998 ) The predominance of Glomusand Acaulospora is in accordance with Klironomos et al(2001 ) who studied interspecific differences in AM fungaltolerance to freezing and dry conditions These authorsfound that Glomus and Acaulospora were more frequentlyisolated in the field and Glomus species were the least affected by freezing in pot experiment conditions whiledrying gave more variable responses in colonization by AMfungi Soil properties are important factors influencing theAM fungal community (Bever et al 2001) Acaulosporaspecies are often associated with acid soils Most of the soils
Table 4 Arbuscular mycorrhizal species richness and diversity indifferent sedge species
Host plants Species richness Diversity
C capillacea var linzensis 35plusmn20 0143plusmn0135C moorcroftii 35plusmn10 0201plusmn0120C tibetica 10plusmn00 0067plusmn0000C compressus 24plusmn06 0169plusmn0062
K humilis 20plusmn04 0137plusmn0042 K prainii 45plusmn21 0177plusmn0150 K prattii 30plusmn00 0195plusmn0123 K pygmaea 30plusmn14 0252plusmn0252 K tibetica 20plusmn00 0070plusmn0000
7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 67
in our study were acidic and this could be one explanationfor our frequent detection of Acaulospora
The colonization pattern varied among plant species andeven among ecotypes of particular plant species K humiliswas mycorrhizal or nonmycorrhizal which demonstratesthat mycorrhizal fungi and their hosts occupy different positions on the ldquo mutualism ndash parasitism continuum rdquo under different environmental conditions (Johnson et al 1997 )The AM fungal community composition of particular sedge species varied with habitat type similar to the resultsof Brundrett et al ( 1999 ) G mosseae and G intraradiceswere also found by Klironomos et al ( 2001 ) in similar environmental conditions This indicates that some specieshave higher specific ecological adaptability
Host species is an important factor influencing AMfungal species diversity (Brundrett 1991 Bever et al 2001)Species richness and fungal diversity differed among thenine sedge species in our study It has been shown that AMfungal diversity has a strong effect on the plant communityon plant productivity and on succession (Allen 1991 Bever et al 2001) The available data suggest an important ecological role for AM fungi in the Tibetan sedge commu-nity However full elucidation of the effects of AM fungi onthe sedges still remains unclear and the ecology andimportance of AM associations require further detailedresearch
Acknowledgements We thank the National Natural Science Foun-dation of China (Projects 30370818 30260055 and 30470341) andthe Royal Society (China Exchanges Project 15360) for financialsupport
Appendix
AM fungal species and their frequency of isolation from rhizospheresoils of sedges
Species Frequency Acaulospora appendicula Spain Sieverding amp Schenck 45 Acaulospora dilatata Morton 91 Acaulospora laevis Gerd amp Trappe 227 Acaulospora mellea Spain amp Schenck 45 Acaulospora scrobiculata Trappe 318 Acaulospora spinosa Walker amp Trappe 91 Acaulospora sp 1 91 Acaulospora sp 2 45 Archaeospora gerdemanni (Rose Daniels amp Trappe)
Morton amp Redecker 45Glomus claroideum Schenck amp Sm emend Walker ampVestberg
182
Glomus convolutum Gerd amp Trappe 45Glomus clarum Nicolson amp Schenck 45Glomus diaphanum Morton amp Walker 91Glomus etunicatum Becker amp Gerdemann 91Glomus geosporum (Nicol amp Gerd) Walker 91Glomus glomerulatum Sieverd 45Glomus intraradices Schenck amp Smith 227
Species FrequencyGlomus luteum Kennedy Stutz et Morton 45Glomus manihotis Howeler Sieverd amp Schenck 45Glomus mosseae (Nicol amp Gerd) Gerd amp Trappe 273Glomus verruculosum Błaszk 91Glomus sp 1 91 Pacispora scintillans (Rose amp Trappe) Oehl amp Sieverd 136
Paraglomus occultum (Walker) Morton amp Redecker 91Scutellospora calospora (Nicol amp Gerd) Walker 91Scutellospora verrucosa (Koske amp C Walker)Walker amp Sanders
45
References
Addy HD Boswell EP Koide RT (1998) Low temperatureacclimation and freezing resistance of extraradical VA mycor-rhizal hyphae Mycol Res 102582 ndash 586
Allen ME (1991) The ecology of mycorrhiza Cambridge University
Press New York Allsopp N Stock WD (1993) Mycorrhizal status of plants growingin the Cape Floristic Region South Africa Bothalia 2391 ndash 104
Bever JD (2002) Host-specificity of AM fungal population growthrates can generate feedback on plant growth Plant Soil244281 ndash 290
Bever JD Schultz PA Pringle A Morton JB (2001) Arbuscular mycorrhizal fungi more diverse than meets the eye and theecological tale of why Bioscience 51923 ndash 931
Brundrett M (1991) Mycorrhizas in natural ecosystems Adv EcolRes 21171 ndash 313
Brundrett M Bougher N Dell B Grove T Malajczuk N (1996)Working with mycorrhizas in forestry and agriculture ACIAR Monograph 32 Australian Centre for International AgriculturalResearch Canberra
Brundrett MC Jasper DA Ashwath N (1999) Glomalean mycor-rhizal fungi from tropical Australia II The effect of nutrient levels and host species on the isolation of fungi Mycorrhiza8315 ndash 321
Burrows RL Pfleger FL (2002) Arbuscular mycorrhizal fungirespond to increasing plant diversity Can J Bot 80120 ndash 130
Daniels BA Skipper HD (1982) Methods for the recovery andquantitative estimation of propagules from soil In Schenck NC(ed) Methods and principles of mycorrhizal research AmericanPhytopathologic Society St Paul MN pp 29 ndash 35
Fuchs B Haselwandter K (2004) Red list plants colonization byarbuscular mycorrhizal fungi and dark septate endophytesMycorrhiza 14277 ndash 281
Giovannetti M Sbrana C (1998) Meeting a non-host the behaviour of AM fungi Mycorrhiza 8123 ndash 130
Harley JL Harley EL (1987) A checklist of mycorrhiza in theBritish flora New Phytol 105 (Suppl)1 ndash 102
Hirsch AM Kapulnik Y (1998) Signal transduction pathways inmycorrhizal associations comparisons with the Rhizobium -legume symbiosis Fungal Genet Biol 23205 ndash 212
Johnson NC Graham JH Smith FA (1997) Functioning of mycorrhizal associations along the mutualism ndash parasitism con-tinuum New Phytol 135575 ndash 586
Klironomos JN Hart MM Gurney JE Moutoglis P (2001)Interspecific differences in the tolerance of arbuscular mycor-rhizal fungi to freezing and drying Can J Bot 791161 ndash 1166
Koske SE Gemma JN Flynn T (1992) Mycorrhizae in Haiwaiianangiosperms a survey with implications for the origin of thenative flora Am J Bot 79853 ndash 862
Lovera M Cuenca G (1996) Arbuscular mycorrhizal infection inCyperaceae and Gramineae from natural disturbed and restoredsavannas in La Gran Sabana Venezuela Mycorrhiza 6111 ndash 118
7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 77
Lu RK (2000) Methods of soil and agricultural chemistry analyses(in Chinese) China Agricultural Science and Technology PressBeijing
McGonigle TP Fitter AH (1990) Ecological specificity of vesicular ndash
arbuscular mycorrhizal associations Mycol Res 94120 ndash 122Muthukumar T Udaiyan K (2000) Arbuscular mycorrhizas of plants
growing in the Western Ghats region Southern India Mycor-rhiza 9297 ndash 313
Muthukumar T Sha LQ Yang XD Cao M Tang JW Zheng Z(2003) Mycorrhiza of plants in different vegetation types intropical ecosystems of Xishuangbanna southwest ChinaMycorrhiza 13289 ndash 297
Muthukumar T Udaiyan K Shanmughavel P (2004) Mycorrhiza insedges an overview Mycorrhiza 1465 ndash 77
Newman EI Reddell P (1987) The distribution of mycorrhizasamong families of vascular plants New Phytol 106745 ndash 751
Peat HJ Fitter AH (1993) The distribution of arbuscular mycor-rhizas in the British flora New Phytol 125845 ndash 854
Peterson RL Bradbury SM (1995) Use of plant mutants intraspe-cific variants and non-hosts in studying mycorrhiza formationand function In Varma A Hock B (eds) Mycorrhiza SpringerBerlin Heidelberg New York pp 521 ndash 560
Phillips JM Hayman DS (1970) Improved procedures for cleaningand staining parasitic and vesicular arbuscular mycorrhizalfungi for rapid assessment of infection Trans Br Mycol Soc55158 ndash 160
Regvar M Vogel K Irgel N Wraber T Hildebrandt U Wilde PBothe H (2003) Colonization of pennycresses ( Thlaspi spp) of the Brassicaceae by arbuscular mycorrhizal fungi J Plant Physiol 160615 ndash 626
Schenck NC Peacutereacutez Y (1990) Manual for the identification of vesicular ndash arbuscular mycorrhizal fungi INVAM University of Florida Gainesville
Smith SE Read DJ (1997) Mycorrhizal symbiosis 2nd ednAcademic Press London
Tester M Smith SE Smith FA (1987) The phenomenon of lsquo nonmycorrhizal rsquo plants Can J Bot 65419 ndash 431
van der Heijden MGA Klironomos JN Ursic M Moutoglis PStreitwolf-Engel R Boller T Wiemken A Sanders IR (1998)Mycorrhizal fungal diversity determines plant biodiversityecosystem variability and productivity Nature 39669 ndash 72
7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 27
Tibet (26deg44 primendash 36deg32 prime N 78deg25 primendash 99deg06 prime E) is the worldslargest and highest plateau The average altitude is about 4500 m above sea level and the weather differs sharplyduring day and night Because of the different geographicalconditions contrasting temperatures are found in the northand south The north has a continental climate and thesouth is relatively warm and rainy Grassland is veryimportant because of the harsh geographical and climaticconditions occupying 8207 million hectares of the Tibetan plateau The main types of grassland are alpine steppesalpine meadow grasslands alpine meadows and montanescrub Poaceae Cyperaceae and Asteraceae are the threemost important plant families in terms of abundanceherbage quality productivity and distribution for pastoralagriculture Most of the sedges especially Kobresia occur in the high quality pasture and information is required onthe mycorrhizal status and ecological role of AM as-sociations in sedges However there are no reports on themycorrhizal status of sedges on the Tibetan plateau
The objectives of the present study were to determine themycorrhizal status of the dominant sedge species Kobresiahumilis Kobresia prainii Kobresia pygmaea and Carexmoorcroftii and to assess the diversity of AM fungi in therhizosphere of sedges in Tibetan grassland The relation-ships between the extent of AM colonization and soil
properties (soil pH organic matter content and available P)were also investigated
Materials and methods
Sampling area and procedures
The investigation was conducted in the prefectures of Lhasa Rikaze Shannan and Naqu (29deg19 primendash 32deg52 prime N 88deg57 primendash 92deg20 prime E) Soils and roots were sampled in the grass-lands at altitudes ranging from 3798 to 5220 m above sealevel The climate of this region from north to south variesfrom alpine plateau to a semiarid type of temperate climateThe mean annual temperature is minus 12 to 8degC and annual precipitation is about 300 ndash 450 mm
Vegetation types are mainly montane scrub alpinesteppe alpine meadow grassland and alpine meadow Thedominant sedge species in the different types of grasslandwere sampled in this study In the autumn of 2003 and2004 22 soil samples were collected from the rhizosphereof nine sedge species in the main grasslands The sedgespecies selected are often dominant and scattered amongdense grasses in the sampling areas Three or more plantswere dug out together from each sampling point and
Table 1 Sedges examined vegetation type altitude soil characteristics and AM status of Tibetan sedges
Host plants Landtype
Altitude(m)
Soiltexture
Organicmatter (gkg)
Olsen-P(mgkg)
pH AM fungalstructures
Colonization()
Sporedensity(per 20 g)
Carex capillacea var linzensis MS 3980 Loam 547 523 674 ar v h 303plusmn85 66
C capillacea varlinzensis
LM 3977 Loam 151 1035 647 v h 318plusmn102 10
C moorcroftii AMG 5220 Sandy loam 182 1499 667 v h 227plusmn59 30C moorcroftii AT 4510 Gravel soil 368 1708 819 v h 221plusmn41 79C tibetica MS 3798 Sand 009 6216 755 v h c 309plusmn123 17C compressus AM 4827 Loam 360 665 603 ar v h 253plusmn67 10C compressus AM 4827 Loam 546 525 601 ar v h 177plusmn20 120C compressus AM 4394 Loam 211 636 626 ar v h 513plusmn157 48C compressus AM 4857 Loam 829 844 602 ar v h 201plusmn32 23C compressus AM 4827 Loam 836 515 61 ar v h 199plusmn19 42 K humilis AM 4385 Loam 251 152 703 ar v h 342plusmn101 58 K humilis AMG 5138 Sand 258 967 686 v h 379plusmn38 16 K humilis AM 4789 Loam 1156 1245 697 ND 0 14 K humilis AS 4559 Loam 209 1817 675 ap v h 427plusmn147 72 K humilis AM 4455 Loam 357 1145 637 v h 352plusmn89 24 K prainii AMG 4897 Loam 813 964 715 ap ar v h 650plusmn164 16 K prainii AS 4543 Gravel soil 082 178 739 ap v h 300plusmn29 70 K pusilla AMG 4717 Loam 778 1102 792 v h 61plusmn14 87 K pusilla AT 4837 Loam 306 749 661 v h 481plusmn129 24 K pygmaea AMG 5006 Gravel soil 036 294 818 ap h 106plusmn19 189 K pygmaea AS 4603 Silty soil 328 514 848 ar v h 153plusmn32 8 K tibetica AMG 4738 Sand 176 925 811 ar v h 560plusmn27 16
AM Arbuscular mycorrhizal Ap appressoria ar arbuscules h intercellular aseptate hyphae v vesicles c coils ND not detected AMG alpinemeadow grassland AS alpine steppe AM alpine meadow MS montane scrub LM lowland meadow
7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 37
approximately 2 kg soil was collected from the rootingzone of the sedges to a depth of 20 cm Care was taken toavoid contamination from other plants The soil sampleswere taken back to the laboratory and soil texture and soil
pH were determined immediately After air-drying the soil
samples were passed through a 2-mm sieve and stored at room temperature Soil available P and organic matter content were determined using the methods described byLu (2000 )
Fig 1 Fungal colonization inTibetan grassland sedgesa Arbuscule (a) intraradicalhyphae and vesicle (v) in root of K prainii b Intraradicalhyphae and vesicle (v) in cellsof C moorcroftii c Vesicle (v)and intraradical hyphae in cellsof K humilis d Extramatricalspores (s) attached to K pygmaea root e Intraradicalhyphae and vesicle (v) in root of Carex capillacea varlinzensis f Hyphal coil (hc)formation on C tibetica rootg Intraradical hyphae (h) andvesicle in root of C compressus h Appressorium (ap) formationin K pygmaea Bar=50 μ m(a d h ) bar=100 μ m (b c f ) bar=150 μ m (e g)
Table 2 Isolation frequencyspecies richness and relativeabundance of the five AR fungalgenera in Tibetan grasslandsedges
F Frequency RA relativeabundance
Genus F () Richness (meanplusmnSE) RA (meanplusmnSE)
Glomus 818 1364plusmn0229 190plusmn58 Acaulospora 636 0955plusmn0184 174plusmn61Scutellospora 136 0136plusmn0077 07plusmn04 Pacispora 136 0136plusmn0077 56plusmn32 Paraglomus 91 0091plusmn0064 55plusmn44 Archaeospora 45 0045plusmn0045 01plusmn01
7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 47
Root staining
Roots were gently washed with water to remove adheringsoil and carefully separated according to their morpholog-ical characteristics to avoid misinterpretation Washed root samples were cleared in 10 (wv) KOH for 30 min at 90degC acidified in 2 (vv) lactic acid for 10 min andstained for 30 min at 90degC with 005 (wv) Trypan blue(Phillips and Hayman 1970 ) Roots that remained dark after clearing were bleached in alkaline H 2 O2 prior toacidification with lactic acid Fifty 05- to 1-cm root fragments from each sample were examined under acompound microscope (100 ndash 400times) for AM fungal struc-tures The presence of arbuscules was used to designateAM associations (Brundrett 1991 ) and the presence of anAM association was considered doubtful when arbusculeswere absent The proportion of root length colonized wasestimated by the root slide technique (Brundrett et al 1996 )
Spore extraction and counting
Spores were isolated from air-dried soil using the methodof Daniels and Skipper ( 1982 ) Twenty grams of soil takenfrom each sample was suspended in 1 l water and left tostand for 20 min The suspensions were passed throughnested 500- and 45- μ m sieves and wet sieved followed bysucrose density gradient centrifugation The AM fungalspores were counted on a grid-patterned dish under a binocular stereomicroscope
Culture establishment
Trap cultures were established from fresh soil samplesmixed with autoclaved sand in a ratio of 21 Clover (Trifolium repens L) and sorghum ( Sorghum vulgare Pers)were used as host plants Cultures were grown in agreenhouse at China Agricultural University for 5 monthswith a temperature regime of 28degC (day) and 15degC (night)and a 14-h photoperiod at a light intensity of 250 μ mol m
minus2
sminus2
provided by supplementary illumination
Identification of AM fungi
Spores of AM fungi isolated from the field soils and trap
cultures were mounted on glass slides in polyvinyl ndash
lacto ndash
glycerol (PVLG) or PVLG + Melzers reagent (11 vv)The spores were examined microscopically and identifiedaccording to current taxonomic criteria (Schenck and Peacutereacutez1990 ) and using the Internet information from the INVAMwebsite ( http invamcafwvuedu )
Data analysis
AM fungal diversity was evaluated by spore density rel-ative abundance isolation frequency (F) and species
richness Spore density was expressed as numbers of AMfungal spores per 20 g dried field soil Relative abundancewas defined as the percentage of numbers of spores of the particular species or genera in the field soil Data from fieldsoil and trap cultures were combined to determine speciescomposition Isolation frequency was calculated as the percentage of samples in which the particular genus or species was present Species richness was defined asnumbers of AM fungal species per soil sample Speciesdiversity was measured by the Shannon ndash Weiner index asfollows
Shannon Weiner index frac14 X Pi ln Pifrac12 eth THORN
where
Pi frac14 ni =N
and n i = number of spores in species i and N = total sporenumber of all species
Results
Mycorrhizal status of sedges
AM fungal structures were observed in all the sedge rootsexamined except for one ecotype of K humilis (Table 1Fig 1) At species level six species in our survey weremycorrhizal K humilis was facultatively mycorrhizal andC moorcroftii and Kobresia pusilla were of doubtfulmycorrhizal status The colonization pattern varied amongthe plant species and even among the ecotypes of individual
species Vesicles and intercellular aseptate hyphae were themost frequently observed structures present in the sedgeroots examined Vesicles were observed in 20 root samples(90) whereas arbuscules were observed in only 10 sam- ples (46) Appressoria were present in K humilis sampledfrom alpine steppe K prainii from both alpine meadowgrassland and alpine steppe and K pygmaea from alpinemeadow grassland Intracellular coils were found only inCarex tibetica No AM fungal structures were observed inthe root sample of K humilis from alpine meadow
Colonization levels in sedges varied from 0 to 65spore density from 8 to 189 per 20 g dried soil and bothvaried greatly with plant species and ecotype (Table 1)
Table 3 Isolation frequency ( ge 15) and relative abundance of arbuscular mycorrhizal fungal species
Species F () RA (meanplusmnSE)
A laevis 227 35plusmn16 A scrobiculata 318 58plusmn22G claroideum 182 53plusmn42G intraradices 227 24plusmn15G mosseae 273 28plusmn15
F Isolation frequency RA relative abundance
7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 57
Further Pearsons correlation analysis of colonization lev-els and soil factors (soil pH organic matter and availableP) revealed a strong negative correlation between organicmatter and colonization rate ( r = minus 0733 P lt000 n=22)
AM fungal diversity
Twenty-six taxa of AM fungi were isolated from the soilsamples belonging to Glomus Acaulospora Paraglomus Archaeospora Pacispora and Scutellospora (Table 2)Species richness in the survey areas was 273 Glomus and Acaulospora were the dominant genera both in frequencyand relative abundance The five most commonly observedspecies were Acaulospora scrobiculata Glomus mosseae Glomus intraradices Acaulospora laevis and Glomusclaroideum Of these A scrobiculata was the most fre-quent and abundant species (Table 3)
The species richness and fungal diversity of nine sedge
species differed substantially (Table 4) Species richnessvaried from 10 ( C tibetica ) to 45 ( K prainii ) and diversityfrom 0070 ( Kobresia tibetica ) to 0252 ( K pygmaea ) Norelationship existed between AM fungal diversity (diversityindex spore density) and soil factors (soil pH organicmatter and available P)
Discussion
Here we report for the first time the mycorrhizal status of eight of the nine sedge species investigated Only Cyperuscompressus has previously been reported as mycorrhizal
(Allsopp and Stock 1993 Muthukumar and Udaiyan 2000 )and was defined as facultatively mycorrhizal whenMuthukumar et al ( 2004 ) summarized the published in-formation on the mycorrhizal status of sedges C compres- sus was mycorrhizal in all ecotypes collected in our studyOnly intercellular aseptate hyphae and vesicles were foundin C moorcroftii and K pusilla It has been reported that vesicles and hyphae are found in nonmycorrhizal hosts suchas Cyperus iria and Cyperus rotundus (Koske et al 1992 Giovannetti and Sbrana 1998 ) These plants can therefore bereferred to as of doubtful AM status K humilis was found to be facultatively mycorrhizal in our survey The incidence of
mycorrhizal species in the present study was much higher (67) than the average level of 40 reported Tibetansedges therefore appear to be readily colonized by AMfungi
Vesicles and intercellular aseptate hyphae were the most frequent structures present in the sedge roots and thisaccords with earlier reports (Muthukumar et al 2003 Muthukumar and Udaiyan 2000 ) The incidence of ar- buscules was 46 not a very low value compared withother surveys (Muthukumar et al 2003 Muthukumar andUdaiyan 2000 ) However the percentage of root lengthcolonized by AM fungi was very low in most root samplesand hyphal coils and appressoria were found in only a fewspecimens The potential nutritional benefit of the mycor-rhizal association in sedges therefore has yet to be as-certained More detailed studies are necessary to ascertainthe exact degree of mycorrhizal dependency of Cyperaceaeand the functional aspects of the association in theseTibetan grassland communities
The rarely colonized or nonmycorrhizal status of sedgesis often attributed to the small amount or absence of mycorrhizal propagules in the soils that sedges inhabit (Peat and Fitter 1993 ) However many sedges tend to benonmycorrhizal even in the presence of AM fungal prop-agules (Van der Heijden et al 1998 ) indicating that absence of AM fungal inoculum may not always explainthe low incidence of AM in these plants Spore densities inthe rhizosphere of sedges were determined in our studyand there was no correlation between spore density andcolonization rate Other important factors likely to influ-ence root colonization are soil properties A strong negativecorrelation was found between soil organic matter content and root colonization rate This is in agreement with theanalysis of Muthukumar et al ( 2004 ) However root
colonization was not related to important factors such assoil pH or soil available P which are shown to influence themycorrhizal status of sedges (Lovera and Cuenca 1996 Muthukumar et al 2004 ) One likely explanation could bethe narrow data range in a particular habitat of this studyThe mechanisms by which soil properties and other factorsinfluence mycorrhizal associations still remain unclear andrequire more detailed research
AM fungi are known to exhibit ecological specificity(McGonigle and Fitter 1990 ) There are no published reportson the AM fungal community associated with sedges under the climatic conditions prevailing on the Tibet plateau Onlya few studies have addressed the ability of AM fungi to
survive winter freezing or drought stress (van der Heijden et al 1998 Addy et al 1998 ) The predominance of Glomusand Acaulospora is in accordance with Klironomos et al(2001 ) who studied interspecific differences in AM fungaltolerance to freezing and dry conditions These authorsfound that Glomus and Acaulospora were more frequentlyisolated in the field and Glomus species were the least affected by freezing in pot experiment conditions whiledrying gave more variable responses in colonization by AMfungi Soil properties are important factors influencing theAM fungal community (Bever et al 2001) Acaulosporaspecies are often associated with acid soils Most of the soils
Table 4 Arbuscular mycorrhizal species richness and diversity indifferent sedge species
Host plants Species richness Diversity
C capillacea var linzensis 35plusmn20 0143plusmn0135C moorcroftii 35plusmn10 0201plusmn0120C tibetica 10plusmn00 0067plusmn0000C compressus 24plusmn06 0169plusmn0062
K humilis 20plusmn04 0137plusmn0042 K prainii 45plusmn21 0177plusmn0150 K prattii 30plusmn00 0195plusmn0123 K pygmaea 30plusmn14 0252plusmn0252 K tibetica 20plusmn00 0070plusmn0000
7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 67
in our study were acidic and this could be one explanationfor our frequent detection of Acaulospora
The colonization pattern varied among plant species andeven among ecotypes of particular plant species K humiliswas mycorrhizal or nonmycorrhizal which demonstratesthat mycorrhizal fungi and their hosts occupy different positions on the ldquo mutualism ndash parasitism continuum rdquo under different environmental conditions (Johnson et al 1997 )The AM fungal community composition of particular sedge species varied with habitat type similar to the resultsof Brundrett et al ( 1999 ) G mosseae and G intraradiceswere also found by Klironomos et al ( 2001 ) in similar environmental conditions This indicates that some specieshave higher specific ecological adaptability
Host species is an important factor influencing AMfungal species diversity (Brundrett 1991 Bever et al 2001)Species richness and fungal diversity differed among thenine sedge species in our study It has been shown that AMfungal diversity has a strong effect on the plant communityon plant productivity and on succession (Allen 1991 Bever et al 2001) The available data suggest an important ecological role for AM fungi in the Tibetan sedge commu-nity However full elucidation of the effects of AM fungi onthe sedges still remains unclear and the ecology andimportance of AM associations require further detailedresearch
Acknowledgements We thank the National Natural Science Foun-dation of China (Projects 30370818 30260055 and 30470341) andthe Royal Society (China Exchanges Project 15360) for financialsupport
Appendix
AM fungal species and their frequency of isolation from rhizospheresoils of sedges
Species Frequency Acaulospora appendicula Spain Sieverding amp Schenck 45 Acaulospora dilatata Morton 91 Acaulospora laevis Gerd amp Trappe 227 Acaulospora mellea Spain amp Schenck 45 Acaulospora scrobiculata Trappe 318 Acaulospora spinosa Walker amp Trappe 91 Acaulospora sp 1 91 Acaulospora sp 2 45 Archaeospora gerdemanni (Rose Daniels amp Trappe)
Morton amp Redecker 45Glomus claroideum Schenck amp Sm emend Walker ampVestberg
182
Glomus convolutum Gerd amp Trappe 45Glomus clarum Nicolson amp Schenck 45Glomus diaphanum Morton amp Walker 91Glomus etunicatum Becker amp Gerdemann 91Glomus geosporum (Nicol amp Gerd) Walker 91Glomus glomerulatum Sieverd 45Glomus intraradices Schenck amp Smith 227
Species FrequencyGlomus luteum Kennedy Stutz et Morton 45Glomus manihotis Howeler Sieverd amp Schenck 45Glomus mosseae (Nicol amp Gerd) Gerd amp Trappe 273Glomus verruculosum Błaszk 91Glomus sp 1 91 Pacispora scintillans (Rose amp Trappe) Oehl amp Sieverd 136
Paraglomus occultum (Walker) Morton amp Redecker 91Scutellospora calospora (Nicol amp Gerd) Walker 91Scutellospora verrucosa (Koske amp C Walker)Walker amp Sanders
45
References
Addy HD Boswell EP Koide RT (1998) Low temperatureacclimation and freezing resistance of extraradical VA mycor-rhizal hyphae Mycol Res 102582 ndash 586
Allen ME (1991) The ecology of mycorrhiza Cambridge University
Press New York Allsopp N Stock WD (1993) Mycorrhizal status of plants growingin the Cape Floristic Region South Africa Bothalia 2391 ndash 104
Bever JD (2002) Host-specificity of AM fungal population growthrates can generate feedback on plant growth Plant Soil244281 ndash 290
Bever JD Schultz PA Pringle A Morton JB (2001) Arbuscular mycorrhizal fungi more diverse than meets the eye and theecological tale of why Bioscience 51923 ndash 931
Brundrett M (1991) Mycorrhizas in natural ecosystems Adv EcolRes 21171 ndash 313
Brundrett M Bougher N Dell B Grove T Malajczuk N (1996)Working with mycorrhizas in forestry and agriculture ACIAR Monograph 32 Australian Centre for International AgriculturalResearch Canberra
Brundrett MC Jasper DA Ashwath N (1999) Glomalean mycor-rhizal fungi from tropical Australia II The effect of nutrient levels and host species on the isolation of fungi Mycorrhiza8315 ndash 321
Burrows RL Pfleger FL (2002) Arbuscular mycorrhizal fungirespond to increasing plant diversity Can J Bot 80120 ndash 130
Daniels BA Skipper HD (1982) Methods for the recovery andquantitative estimation of propagules from soil In Schenck NC(ed) Methods and principles of mycorrhizal research AmericanPhytopathologic Society St Paul MN pp 29 ndash 35
Fuchs B Haselwandter K (2004) Red list plants colonization byarbuscular mycorrhizal fungi and dark septate endophytesMycorrhiza 14277 ndash 281
Giovannetti M Sbrana C (1998) Meeting a non-host the behaviour of AM fungi Mycorrhiza 8123 ndash 130
Harley JL Harley EL (1987) A checklist of mycorrhiza in theBritish flora New Phytol 105 (Suppl)1 ndash 102
Hirsch AM Kapulnik Y (1998) Signal transduction pathways inmycorrhizal associations comparisons with the Rhizobium -legume symbiosis Fungal Genet Biol 23205 ndash 212
Johnson NC Graham JH Smith FA (1997) Functioning of mycorrhizal associations along the mutualism ndash parasitism con-tinuum New Phytol 135575 ndash 586
Klironomos JN Hart MM Gurney JE Moutoglis P (2001)Interspecific differences in the tolerance of arbuscular mycor-rhizal fungi to freezing and drying Can J Bot 791161 ndash 1166
Koske SE Gemma JN Flynn T (1992) Mycorrhizae in Haiwaiianangiosperms a survey with implications for the origin of thenative flora Am J Bot 79853 ndash 862
Lovera M Cuenca G (1996) Arbuscular mycorrhizal infection inCyperaceae and Gramineae from natural disturbed and restoredsavannas in La Gran Sabana Venezuela Mycorrhiza 6111 ndash 118
7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 77
Lu RK (2000) Methods of soil and agricultural chemistry analyses(in Chinese) China Agricultural Science and Technology PressBeijing
McGonigle TP Fitter AH (1990) Ecological specificity of vesicular ndash
arbuscular mycorrhizal associations Mycol Res 94120 ndash 122Muthukumar T Udaiyan K (2000) Arbuscular mycorrhizas of plants
growing in the Western Ghats region Southern India Mycor-rhiza 9297 ndash 313
Muthukumar T Sha LQ Yang XD Cao M Tang JW Zheng Z(2003) Mycorrhiza of plants in different vegetation types intropical ecosystems of Xishuangbanna southwest ChinaMycorrhiza 13289 ndash 297
Muthukumar T Udaiyan K Shanmughavel P (2004) Mycorrhiza insedges an overview Mycorrhiza 1465 ndash 77
Newman EI Reddell P (1987) The distribution of mycorrhizasamong families of vascular plants New Phytol 106745 ndash 751
Peat HJ Fitter AH (1993) The distribution of arbuscular mycor-rhizas in the British flora New Phytol 125845 ndash 854
Peterson RL Bradbury SM (1995) Use of plant mutants intraspe-cific variants and non-hosts in studying mycorrhiza formationand function In Varma A Hock B (eds) Mycorrhiza SpringerBerlin Heidelberg New York pp 521 ndash 560
Phillips JM Hayman DS (1970) Improved procedures for cleaningand staining parasitic and vesicular arbuscular mycorrhizalfungi for rapid assessment of infection Trans Br Mycol Soc55158 ndash 160
Regvar M Vogel K Irgel N Wraber T Hildebrandt U Wilde PBothe H (2003) Colonization of pennycresses ( Thlaspi spp) of the Brassicaceae by arbuscular mycorrhizal fungi J Plant Physiol 160615 ndash 626
Schenck NC Peacutereacutez Y (1990) Manual for the identification of vesicular ndash arbuscular mycorrhizal fungi INVAM University of Florida Gainesville
Smith SE Read DJ (1997) Mycorrhizal symbiosis 2nd ednAcademic Press London
Tester M Smith SE Smith FA (1987) The phenomenon of lsquo nonmycorrhizal rsquo plants Can J Bot 65419 ndash 431
van der Heijden MGA Klironomos JN Ursic M Moutoglis PStreitwolf-Engel R Boller T Wiemken A Sanders IR (1998)Mycorrhizal fungal diversity determines plant biodiversityecosystem variability and productivity Nature 39669 ndash 72
7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 37
approximately 2 kg soil was collected from the rootingzone of the sedges to a depth of 20 cm Care was taken toavoid contamination from other plants The soil sampleswere taken back to the laboratory and soil texture and soil
pH were determined immediately After air-drying the soil
samples were passed through a 2-mm sieve and stored at room temperature Soil available P and organic matter content were determined using the methods described byLu (2000 )
Fig 1 Fungal colonization inTibetan grassland sedgesa Arbuscule (a) intraradicalhyphae and vesicle (v) in root of K prainii b Intraradicalhyphae and vesicle (v) in cellsof C moorcroftii c Vesicle (v)and intraradical hyphae in cellsof K humilis d Extramatricalspores (s) attached to K pygmaea root e Intraradicalhyphae and vesicle (v) in root of Carex capillacea varlinzensis f Hyphal coil (hc)formation on C tibetica rootg Intraradical hyphae (h) andvesicle in root of C compressus h Appressorium (ap) formationin K pygmaea Bar=50 μ m(a d h ) bar=100 μ m (b c f ) bar=150 μ m (e g)
Table 2 Isolation frequencyspecies richness and relativeabundance of the five AR fungalgenera in Tibetan grasslandsedges
F Frequency RA relativeabundance
Genus F () Richness (meanplusmnSE) RA (meanplusmnSE)
Glomus 818 1364plusmn0229 190plusmn58 Acaulospora 636 0955plusmn0184 174plusmn61Scutellospora 136 0136plusmn0077 07plusmn04 Pacispora 136 0136plusmn0077 56plusmn32 Paraglomus 91 0091plusmn0064 55plusmn44 Archaeospora 45 0045plusmn0045 01plusmn01
7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 47
Root staining
Roots were gently washed with water to remove adheringsoil and carefully separated according to their morpholog-ical characteristics to avoid misinterpretation Washed root samples were cleared in 10 (wv) KOH for 30 min at 90degC acidified in 2 (vv) lactic acid for 10 min andstained for 30 min at 90degC with 005 (wv) Trypan blue(Phillips and Hayman 1970 ) Roots that remained dark after clearing were bleached in alkaline H 2 O2 prior toacidification with lactic acid Fifty 05- to 1-cm root fragments from each sample were examined under acompound microscope (100 ndash 400times) for AM fungal struc-tures The presence of arbuscules was used to designateAM associations (Brundrett 1991 ) and the presence of anAM association was considered doubtful when arbusculeswere absent The proportion of root length colonized wasestimated by the root slide technique (Brundrett et al 1996 )
Spore extraction and counting
Spores were isolated from air-dried soil using the methodof Daniels and Skipper ( 1982 ) Twenty grams of soil takenfrom each sample was suspended in 1 l water and left tostand for 20 min The suspensions were passed throughnested 500- and 45- μ m sieves and wet sieved followed bysucrose density gradient centrifugation The AM fungalspores were counted on a grid-patterned dish under a binocular stereomicroscope
Culture establishment
Trap cultures were established from fresh soil samplesmixed with autoclaved sand in a ratio of 21 Clover (Trifolium repens L) and sorghum ( Sorghum vulgare Pers)were used as host plants Cultures were grown in agreenhouse at China Agricultural University for 5 monthswith a temperature regime of 28degC (day) and 15degC (night)and a 14-h photoperiod at a light intensity of 250 μ mol m
minus2
sminus2
provided by supplementary illumination
Identification of AM fungi
Spores of AM fungi isolated from the field soils and trap
cultures were mounted on glass slides in polyvinyl ndash
lacto ndash
glycerol (PVLG) or PVLG + Melzers reagent (11 vv)The spores were examined microscopically and identifiedaccording to current taxonomic criteria (Schenck and Peacutereacutez1990 ) and using the Internet information from the INVAMwebsite ( http invamcafwvuedu )
Data analysis
AM fungal diversity was evaluated by spore density rel-ative abundance isolation frequency (F) and species
richness Spore density was expressed as numbers of AMfungal spores per 20 g dried field soil Relative abundancewas defined as the percentage of numbers of spores of the particular species or genera in the field soil Data from fieldsoil and trap cultures were combined to determine speciescomposition Isolation frequency was calculated as the percentage of samples in which the particular genus or species was present Species richness was defined asnumbers of AM fungal species per soil sample Speciesdiversity was measured by the Shannon ndash Weiner index asfollows
Shannon Weiner index frac14 X Pi ln Pifrac12 eth THORN
where
Pi frac14 ni =N
and n i = number of spores in species i and N = total sporenumber of all species
Results
Mycorrhizal status of sedges
AM fungal structures were observed in all the sedge rootsexamined except for one ecotype of K humilis (Table 1Fig 1) At species level six species in our survey weremycorrhizal K humilis was facultatively mycorrhizal andC moorcroftii and Kobresia pusilla were of doubtfulmycorrhizal status The colonization pattern varied amongthe plant species and even among the ecotypes of individual
species Vesicles and intercellular aseptate hyphae were themost frequently observed structures present in the sedgeroots examined Vesicles were observed in 20 root samples(90) whereas arbuscules were observed in only 10 sam- ples (46) Appressoria were present in K humilis sampledfrom alpine steppe K prainii from both alpine meadowgrassland and alpine steppe and K pygmaea from alpinemeadow grassland Intracellular coils were found only inCarex tibetica No AM fungal structures were observed inthe root sample of K humilis from alpine meadow
Colonization levels in sedges varied from 0 to 65spore density from 8 to 189 per 20 g dried soil and bothvaried greatly with plant species and ecotype (Table 1)
Table 3 Isolation frequency ( ge 15) and relative abundance of arbuscular mycorrhizal fungal species
Species F () RA (meanplusmnSE)
A laevis 227 35plusmn16 A scrobiculata 318 58plusmn22G claroideum 182 53plusmn42G intraradices 227 24plusmn15G mosseae 273 28plusmn15
F Isolation frequency RA relative abundance
7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 57
Further Pearsons correlation analysis of colonization lev-els and soil factors (soil pH organic matter and availableP) revealed a strong negative correlation between organicmatter and colonization rate ( r = minus 0733 P lt000 n=22)
AM fungal diversity
Twenty-six taxa of AM fungi were isolated from the soilsamples belonging to Glomus Acaulospora Paraglomus Archaeospora Pacispora and Scutellospora (Table 2)Species richness in the survey areas was 273 Glomus and Acaulospora were the dominant genera both in frequencyand relative abundance The five most commonly observedspecies were Acaulospora scrobiculata Glomus mosseae Glomus intraradices Acaulospora laevis and Glomusclaroideum Of these A scrobiculata was the most fre-quent and abundant species (Table 3)
The species richness and fungal diversity of nine sedge
species differed substantially (Table 4) Species richnessvaried from 10 ( C tibetica ) to 45 ( K prainii ) and diversityfrom 0070 ( Kobresia tibetica ) to 0252 ( K pygmaea ) Norelationship existed between AM fungal diversity (diversityindex spore density) and soil factors (soil pH organicmatter and available P)
Discussion
Here we report for the first time the mycorrhizal status of eight of the nine sedge species investigated Only Cyperuscompressus has previously been reported as mycorrhizal
(Allsopp and Stock 1993 Muthukumar and Udaiyan 2000 )and was defined as facultatively mycorrhizal whenMuthukumar et al ( 2004 ) summarized the published in-formation on the mycorrhizal status of sedges C compres- sus was mycorrhizal in all ecotypes collected in our studyOnly intercellular aseptate hyphae and vesicles were foundin C moorcroftii and K pusilla It has been reported that vesicles and hyphae are found in nonmycorrhizal hosts suchas Cyperus iria and Cyperus rotundus (Koske et al 1992 Giovannetti and Sbrana 1998 ) These plants can therefore bereferred to as of doubtful AM status K humilis was found to be facultatively mycorrhizal in our survey The incidence of
mycorrhizal species in the present study was much higher (67) than the average level of 40 reported Tibetansedges therefore appear to be readily colonized by AMfungi
Vesicles and intercellular aseptate hyphae were the most frequent structures present in the sedge roots and thisaccords with earlier reports (Muthukumar et al 2003 Muthukumar and Udaiyan 2000 ) The incidence of ar- buscules was 46 not a very low value compared withother surveys (Muthukumar et al 2003 Muthukumar andUdaiyan 2000 ) However the percentage of root lengthcolonized by AM fungi was very low in most root samplesand hyphal coils and appressoria were found in only a fewspecimens The potential nutritional benefit of the mycor-rhizal association in sedges therefore has yet to be as-certained More detailed studies are necessary to ascertainthe exact degree of mycorrhizal dependency of Cyperaceaeand the functional aspects of the association in theseTibetan grassland communities
The rarely colonized or nonmycorrhizal status of sedgesis often attributed to the small amount or absence of mycorrhizal propagules in the soils that sedges inhabit (Peat and Fitter 1993 ) However many sedges tend to benonmycorrhizal even in the presence of AM fungal prop-agules (Van der Heijden et al 1998 ) indicating that absence of AM fungal inoculum may not always explainthe low incidence of AM in these plants Spore densities inthe rhizosphere of sedges were determined in our studyand there was no correlation between spore density andcolonization rate Other important factors likely to influ-ence root colonization are soil properties A strong negativecorrelation was found between soil organic matter content and root colonization rate This is in agreement with theanalysis of Muthukumar et al ( 2004 ) However root
colonization was not related to important factors such assoil pH or soil available P which are shown to influence themycorrhizal status of sedges (Lovera and Cuenca 1996 Muthukumar et al 2004 ) One likely explanation could bethe narrow data range in a particular habitat of this studyThe mechanisms by which soil properties and other factorsinfluence mycorrhizal associations still remain unclear andrequire more detailed research
AM fungi are known to exhibit ecological specificity(McGonigle and Fitter 1990 ) There are no published reportson the AM fungal community associated with sedges under the climatic conditions prevailing on the Tibet plateau Onlya few studies have addressed the ability of AM fungi to
survive winter freezing or drought stress (van der Heijden et al 1998 Addy et al 1998 ) The predominance of Glomusand Acaulospora is in accordance with Klironomos et al(2001 ) who studied interspecific differences in AM fungaltolerance to freezing and dry conditions These authorsfound that Glomus and Acaulospora were more frequentlyisolated in the field and Glomus species were the least affected by freezing in pot experiment conditions whiledrying gave more variable responses in colonization by AMfungi Soil properties are important factors influencing theAM fungal community (Bever et al 2001) Acaulosporaspecies are often associated with acid soils Most of the soils
Table 4 Arbuscular mycorrhizal species richness and diversity indifferent sedge species
Host plants Species richness Diversity
C capillacea var linzensis 35plusmn20 0143plusmn0135C moorcroftii 35plusmn10 0201plusmn0120C tibetica 10plusmn00 0067plusmn0000C compressus 24plusmn06 0169plusmn0062
K humilis 20plusmn04 0137plusmn0042 K prainii 45plusmn21 0177plusmn0150 K prattii 30plusmn00 0195plusmn0123 K pygmaea 30plusmn14 0252plusmn0252 K tibetica 20plusmn00 0070plusmn0000
7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 67
in our study were acidic and this could be one explanationfor our frequent detection of Acaulospora
The colonization pattern varied among plant species andeven among ecotypes of particular plant species K humiliswas mycorrhizal or nonmycorrhizal which demonstratesthat mycorrhizal fungi and their hosts occupy different positions on the ldquo mutualism ndash parasitism continuum rdquo under different environmental conditions (Johnson et al 1997 )The AM fungal community composition of particular sedge species varied with habitat type similar to the resultsof Brundrett et al ( 1999 ) G mosseae and G intraradiceswere also found by Klironomos et al ( 2001 ) in similar environmental conditions This indicates that some specieshave higher specific ecological adaptability
Host species is an important factor influencing AMfungal species diversity (Brundrett 1991 Bever et al 2001)Species richness and fungal diversity differed among thenine sedge species in our study It has been shown that AMfungal diversity has a strong effect on the plant communityon plant productivity and on succession (Allen 1991 Bever et al 2001) The available data suggest an important ecological role for AM fungi in the Tibetan sedge commu-nity However full elucidation of the effects of AM fungi onthe sedges still remains unclear and the ecology andimportance of AM associations require further detailedresearch
Acknowledgements We thank the National Natural Science Foun-dation of China (Projects 30370818 30260055 and 30470341) andthe Royal Society (China Exchanges Project 15360) for financialsupport
Appendix
AM fungal species and their frequency of isolation from rhizospheresoils of sedges
Species Frequency Acaulospora appendicula Spain Sieverding amp Schenck 45 Acaulospora dilatata Morton 91 Acaulospora laevis Gerd amp Trappe 227 Acaulospora mellea Spain amp Schenck 45 Acaulospora scrobiculata Trappe 318 Acaulospora spinosa Walker amp Trappe 91 Acaulospora sp 1 91 Acaulospora sp 2 45 Archaeospora gerdemanni (Rose Daniels amp Trappe)
Morton amp Redecker 45Glomus claroideum Schenck amp Sm emend Walker ampVestberg
182
Glomus convolutum Gerd amp Trappe 45Glomus clarum Nicolson amp Schenck 45Glomus diaphanum Morton amp Walker 91Glomus etunicatum Becker amp Gerdemann 91Glomus geosporum (Nicol amp Gerd) Walker 91Glomus glomerulatum Sieverd 45Glomus intraradices Schenck amp Smith 227
Species FrequencyGlomus luteum Kennedy Stutz et Morton 45Glomus manihotis Howeler Sieverd amp Schenck 45Glomus mosseae (Nicol amp Gerd) Gerd amp Trappe 273Glomus verruculosum Błaszk 91Glomus sp 1 91 Pacispora scintillans (Rose amp Trappe) Oehl amp Sieverd 136
Paraglomus occultum (Walker) Morton amp Redecker 91Scutellospora calospora (Nicol amp Gerd) Walker 91Scutellospora verrucosa (Koske amp C Walker)Walker amp Sanders
45
References
Addy HD Boswell EP Koide RT (1998) Low temperatureacclimation and freezing resistance of extraradical VA mycor-rhizal hyphae Mycol Res 102582 ndash 586
Allen ME (1991) The ecology of mycorrhiza Cambridge University
Press New York Allsopp N Stock WD (1993) Mycorrhizal status of plants growingin the Cape Floristic Region South Africa Bothalia 2391 ndash 104
Bever JD (2002) Host-specificity of AM fungal population growthrates can generate feedback on plant growth Plant Soil244281 ndash 290
Bever JD Schultz PA Pringle A Morton JB (2001) Arbuscular mycorrhizal fungi more diverse than meets the eye and theecological tale of why Bioscience 51923 ndash 931
Brundrett M (1991) Mycorrhizas in natural ecosystems Adv EcolRes 21171 ndash 313
Brundrett M Bougher N Dell B Grove T Malajczuk N (1996)Working with mycorrhizas in forestry and agriculture ACIAR Monograph 32 Australian Centre for International AgriculturalResearch Canberra
Brundrett MC Jasper DA Ashwath N (1999) Glomalean mycor-rhizal fungi from tropical Australia II The effect of nutrient levels and host species on the isolation of fungi Mycorrhiza8315 ndash 321
Burrows RL Pfleger FL (2002) Arbuscular mycorrhizal fungirespond to increasing plant diversity Can J Bot 80120 ndash 130
Daniels BA Skipper HD (1982) Methods for the recovery andquantitative estimation of propagules from soil In Schenck NC(ed) Methods and principles of mycorrhizal research AmericanPhytopathologic Society St Paul MN pp 29 ndash 35
Fuchs B Haselwandter K (2004) Red list plants colonization byarbuscular mycorrhizal fungi and dark septate endophytesMycorrhiza 14277 ndash 281
Giovannetti M Sbrana C (1998) Meeting a non-host the behaviour of AM fungi Mycorrhiza 8123 ndash 130
Harley JL Harley EL (1987) A checklist of mycorrhiza in theBritish flora New Phytol 105 (Suppl)1 ndash 102
Hirsch AM Kapulnik Y (1998) Signal transduction pathways inmycorrhizal associations comparisons with the Rhizobium -legume symbiosis Fungal Genet Biol 23205 ndash 212
Johnson NC Graham JH Smith FA (1997) Functioning of mycorrhizal associations along the mutualism ndash parasitism con-tinuum New Phytol 135575 ndash 586
Klironomos JN Hart MM Gurney JE Moutoglis P (2001)Interspecific differences in the tolerance of arbuscular mycor-rhizal fungi to freezing and drying Can J Bot 791161 ndash 1166
Koske SE Gemma JN Flynn T (1992) Mycorrhizae in Haiwaiianangiosperms a survey with implications for the origin of thenative flora Am J Bot 79853 ndash 862
Lovera M Cuenca G (1996) Arbuscular mycorrhizal infection inCyperaceae and Gramineae from natural disturbed and restoredsavannas in La Gran Sabana Venezuela Mycorrhiza 6111 ndash 118
7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 77
Lu RK (2000) Methods of soil and agricultural chemistry analyses(in Chinese) China Agricultural Science and Technology PressBeijing
McGonigle TP Fitter AH (1990) Ecological specificity of vesicular ndash
arbuscular mycorrhizal associations Mycol Res 94120 ndash 122Muthukumar T Udaiyan K (2000) Arbuscular mycorrhizas of plants
growing in the Western Ghats region Southern India Mycor-rhiza 9297 ndash 313
Muthukumar T Sha LQ Yang XD Cao M Tang JW Zheng Z(2003) Mycorrhiza of plants in different vegetation types intropical ecosystems of Xishuangbanna southwest ChinaMycorrhiza 13289 ndash 297
Muthukumar T Udaiyan K Shanmughavel P (2004) Mycorrhiza insedges an overview Mycorrhiza 1465 ndash 77
Newman EI Reddell P (1987) The distribution of mycorrhizasamong families of vascular plants New Phytol 106745 ndash 751
Peat HJ Fitter AH (1993) The distribution of arbuscular mycor-rhizas in the British flora New Phytol 125845 ndash 854
Peterson RL Bradbury SM (1995) Use of plant mutants intraspe-cific variants and non-hosts in studying mycorrhiza formationand function In Varma A Hock B (eds) Mycorrhiza SpringerBerlin Heidelberg New York pp 521 ndash 560
Phillips JM Hayman DS (1970) Improved procedures for cleaningand staining parasitic and vesicular arbuscular mycorrhizalfungi for rapid assessment of infection Trans Br Mycol Soc55158 ndash 160
Regvar M Vogel K Irgel N Wraber T Hildebrandt U Wilde PBothe H (2003) Colonization of pennycresses ( Thlaspi spp) of the Brassicaceae by arbuscular mycorrhizal fungi J Plant Physiol 160615 ndash 626
Schenck NC Peacutereacutez Y (1990) Manual for the identification of vesicular ndash arbuscular mycorrhizal fungi INVAM University of Florida Gainesville
Smith SE Read DJ (1997) Mycorrhizal symbiosis 2nd ednAcademic Press London
Tester M Smith SE Smith FA (1987) The phenomenon of lsquo nonmycorrhizal rsquo plants Can J Bot 65419 ndash 431
van der Heijden MGA Klironomos JN Ursic M Moutoglis PStreitwolf-Engel R Boller T Wiemken A Sanders IR (1998)Mycorrhizal fungal diversity determines plant biodiversityecosystem variability and productivity Nature 39669 ndash 72

7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 47
Root staining
Roots were gently washed with water to remove adheringsoil and carefully separated according to their morpholog-ical characteristics to avoid misinterpretation Washed root samples were cleared in 10 (wv) KOH for 30 min at 90degC acidified in 2 (vv) lactic acid for 10 min andstained for 30 min at 90degC with 005 (wv) Trypan blue(Phillips and Hayman 1970 ) Roots that remained dark after clearing were bleached in alkaline H 2 O2 prior toacidification with lactic acid Fifty 05- to 1-cm root fragments from each sample were examined under acompound microscope (100 ndash 400times) for AM fungal struc-tures The presence of arbuscules was used to designateAM associations (Brundrett 1991 ) and the presence of anAM association was considered doubtful when arbusculeswere absent The proportion of root length colonized wasestimated by the root slide technique (Brundrett et al 1996 )
Spore extraction and counting
Spores were isolated from air-dried soil using the methodof Daniels and Skipper ( 1982 ) Twenty grams of soil takenfrom each sample was suspended in 1 l water and left tostand for 20 min The suspensions were passed throughnested 500- and 45- μ m sieves and wet sieved followed bysucrose density gradient centrifugation The AM fungalspores were counted on a grid-patterned dish under a binocular stereomicroscope
Culture establishment
Trap cultures were established from fresh soil samplesmixed with autoclaved sand in a ratio of 21 Clover (Trifolium repens L) and sorghum ( Sorghum vulgare Pers)were used as host plants Cultures were grown in agreenhouse at China Agricultural University for 5 monthswith a temperature regime of 28degC (day) and 15degC (night)and a 14-h photoperiod at a light intensity of 250 μ mol m
minus2
sminus2
provided by supplementary illumination
Identification of AM fungi
Spores of AM fungi isolated from the field soils and trap
cultures were mounted on glass slides in polyvinyl ndash
lacto ndash
glycerol (PVLG) or PVLG + Melzers reagent (11 vv)The spores were examined microscopically and identifiedaccording to current taxonomic criteria (Schenck and Peacutereacutez1990 ) and using the Internet information from the INVAMwebsite ( http invamcafwvuedu )
Data analysis
AM fungal diversity was evaluated by spore density rel-ative abundance isolation frequency (F) and species
richness Spore density was expressed as numbers of AMfungal spores per 20 g dried field soil Relative abundancewas defined as the percentage of numbers of spores of the particular species or genera in the field soil Data from fieldsoil and trap cultures were combined to determine speciescomposition Isolation frequency was calculated as the percentage of samples in which the particular genus or species was present Species richness was defined asnumbers of AM fungal species per soil sample Speciesdiversity was measured by the Shannon ndash Weiner index asfollows
Shannon Weiner index frac14 X Pi ln Pifrac12 eth THORN
where
Pi frac14 ni =N
and n i = number of spores in species i and N = total sporenumber of all species
Results
Mycorrhizal status of sedges
AM fungal structures were observed in all the sedge rootsexamined except for one ecotype of K humilis (Table 1Fig 1) At species level six species in our survey weremycorrhizal K humilis was facultatively mycorrhizal andC moorcroftii and Kobresia pusilla were of doubtfulmycorrhizal status The colonization pattern varied amongthe plant species and even among the ecotypes of individual
species Vesicles and intercellular aseptate hyphae were themost frequently observed structures present in the sedgeroots examined Vesicles were observed in 20 root samples(90) whereas arbuscules were observed in only 10 sam- ples (46) Appressoria were present in K humilis sampledfrom alpine steppe K prainii from both alpine meadowgrassland and alpine steppe and K pygmaea from alpinemeadow grassland Intracellular coils were found only inCarex tibetica No AM fungal structures were observed inthe root sample of K humilis from alpine meadow
Colonization levels in sedges varied from 0 to 65spore density from 8 to 189 per 20 g dried soil and bothvaried greatly with plant species and ecotype (Table 1)
Table 3 Isolation frequency ( ge 15) and relative abundance of arbuscular mycorrhizal fungal species
Species F () RA (meanplusmnSE)
A laevis 227 35plusmn16 A scrobiculata 318 58plusmn22G claroideum 182 53plusmn42G intraradices 227 24plusmn15G mosseae 273 28plusmn15
F Isolation frequency RA relative abundance
7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 57
Further Pearsons correlation analysis of colonization lev-els and soil factors (soil pH organic matter and availableP) revealed a strong negative correlation between organicmatter and colonization rate ( r = minus 0733 P lt000 n=22)
AM fungal diversity
Twenty-six taxa of AM fungi were isolated from the soilsamples belonging to Glomus Acaulospora Paraglomus Archaeospora Pacispora and Scutellospora (Table 2)Species richness in the survey areas was 273 Glomus and Acaulospora were the dominant genera both in frequencyand relative abundance The five most commonly observedspecies were Acaulospora scrobiculata Glomus mosseae Glomus intraradices Acaulospora laevis and Glomusclaroideum Of these A scrobiculata was the most fre-quent and abundant species (Table 3)
The species richness and fungal diversity of nine sedge
species differed substantially (Table 4) Species richnessvaried from 10 ( C tibetica ) to 45 ( K prainii ) and diversityfrom 0070 ( Kobresia tibetica ) to 0252 ( K pygmaea ) Norelationship existed between AM fungal diversity (diversityindex spore density) and soil factors (soil pH organicmatter and available P)
Discussion
Here we report for the first time the mycorrhizal status of eight of the nine sedge species investigated Only Cyperuscompressus has previously been reported as mycorrhizal
(Allsopp and Stock 1993 Muthukumar and Udaiyan 2000 )and was defined as facultatively mycorrhizal whenMuthukumar et al ( 2004 ) summarized the published in-formation on the mycorrhizal status of sedges C compres- sus was mycorrhizal in all ecotypes collected in our studyOnly intercellular aseptate hyphae and vesicles were foundin C moorcroftii and K pusilla It has been reported that vesicles and hyphae are found in nonmycorrhizal hosts suchas Cyperus iria and Cyperus rotundus (Koske et al 1992 Giovannetti and Sbrana 1998 ) These plants can therefore bereferred to as of doubtful AM status K humilis was found to be facultatively mycorrhizal in our survey The incidence of
mycorrhizal species in the present study was much higher (67) than the average level of 40 reported Tibetansedges therefore appear to be readily colonized by AMfungi
Vesicles and intercellular aseptate hyphae were the most frequent structures present in the sedge roots and thisaccords with earlier reports (Muthukumar et al 2003 Muthukumar and Udaiyan 2000 ) The incidence of ar- buscules was 46 not a very low value compared withother surveys (Muthukumar et al 2003 Muthukumar andUdaiyan 2000 ) However the percentage of root lengthcolonized by AM fungi was very low in most root samplesand hyphal coils and appressoria were found in only a fewspecimens The potential nutritional benefit of the mycor-rhizal association in sedges therefore has yet to be as-certained More detailed studies are necessary to ascertainthe exact degree of mycorrhizal dependency of Cyperaceaeand the functional aspects of the association in theseTibetan grassland communities
The rarely colonized or nonmycorrhizal status of sedgesis often attributed to the small amount or absence of mycorrhizal propagules in the soils that sedges inhabit (Peat and Fitter 1993 ) However many sedges tend to benonmycorrhizal even in the presence of AM fungal prop-agules (Van der Heijden et al 1998 ) indicating that absence of AM fungal inoculum may not always explainthe low incidence of AM in these plants Spore densities inthe rhizosphere of sedges were determined in our studyand there was no correlation between spore density andcolonization rate Other important factors likely to influ-ence root colonization are soil properties A strong negativecorrelation was found between soil organic matter content and root colonization rate This is in agreement with theanalysis of Muthukumar et al ( 2004 ) However root
colonization was not related to important factors such assoil pH or soil available P which are shown to influence themycorrhizal status of sedges (Lovera and Cuenca 1996 Muthukumar et al 2004 ) One likely explanation could bethe narrow data range in a particular habitat of this studyThe mechanisms by which soil properties and other factorsinfluence mycorrhizal associations still remain unclear andrequire more detailed research
AM fungi are known to exhibit ecological specificity(McGonigle and Fitter 1990 ) There are no published reportson the AM fungal community associated with sedges under the climatic conditions prevailing on the Tibet plateau Onlya few studies have addressed the ability of AM fungi to
survive winter freezing or drought stress (van der Heijden et al 1998 Addy et al 1998 ) The predominance of Glomusand Acaulospora is in accordance with Klironomos et al(2001 ) who studied interspecific differences in AM fungaltolerance to freezing and dry conditions These authorsfound that Glomus and Acaulospora were more frequentlyisolated in the field and Glomus species were the least affected by freezing in pot experiment conditions whiledrying gave more variable responses in colonization by AMfungi Soil properties are important factors influencing theAM fungal community (Bever et al 2001) Acaulosporaspecies are often associated with acid soils Most of the soils
Table 4 Arbuscular mycorrhizal species richness and diversity indifferent sedge species
Host plants Species richness Diversity
C capillacea var linzensis 35plusmn20 0143plusmn0135C moorcroftii 35plusmn10 0201plusmn0120C tibetica 10plusmn00 0067plusmn0000C compressus 24plusmn06 0169plusmn0062
K humilis 20plusmn04 0137plusmn0042 K prainii 45plusmn21 0177plusmn0150 K prattii 30plusmn00 0195plusmn0123 K pygmaea 30plusmn14 0252plusmn0252 K tibetica 20plusmn00 0070plusmn0000
7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 67
in our study were acidic and this could be one explanationfor our frequent detection of Acaulospora
The colonization pattern varied among plant species andeven among ecotypes of particular plant species K humiliswas mycorrhizal or nonmycorrhizal which demonstratesthat mycorrhizal fungi and their hosts occupy different positions on the ldquo mutualism ndash parasitism continuum rdquo under different environmental conditions (Johnson et al 1997 )The AM fungal community composition of particular sedge species varied with habitat type similar to the resultsof Brundrett et al ( 1999 ) G mosseae and G intraradiceswere also found by Klironomos et al ( 2001 ) in similar environmental conditions This indicates that some specieshave higher specific ecological adaptability
Host species is an important factor influencing AMfungal species diversity (Brundrett 1991 Bever et al 2001)Species richness and fungal diversity differed among thenine sedge species in our study It has been shown that AMfungal diversity has a strong effect on the plant communityon plant productivity and on succession (Allen 1991 Bever et al 2001) The available data suggest an important ecological role for AM fungi in the Tibetan sedge commu-nity However full elucidation of the effects of AM fungi onthe sedges still remains unclear and the ecology andimportance of AM associations require further detailedresearch
Acknowledgements We thank the National Natural Science Foun-dation of China (Projects 30370818 30260055 and 30470341) andthe Royal Society (China Exchanges Project 15360) for financialsupport
Appendix
AM fungal species and their frequency of isolation from rhizospheresoils of sedges
Species Frequency Acaulospora appendicula Spain Sieverding amp Schenck 45 Acaulospora dilatata Morton 91 Acaulospora laevis Gerd amp Trappe 227 Acaulospora mellea Spain amp Schenck 45 Acaulospora scrobiculata Trappe 318 Acaulospora spinosa Walker amp Trappe 91 Acaulospora sp 1 91 Acaulospora sp 2 45 Archaeospora gerdemanni (Rose Daniels amp Trappe)
Morton amp Redecker 45Glomus claroideum Schenck amp Sm emend Walker ampVestberg
182
Glomus convolutum Gerd amp Trappe 45Glomus clarum Nicolson amp Schenck 45Glomus diaphanum Morton amp Walker 91Glomus etunicatum Becker amp Gerdemann 91Glomus geosporum (Nicol amp Gerd) Walker 91Glomus glomerulatum Sieverd 45Glomus intraradices Schenck amp Smith 227
Species FrequencyGlomus luteum Kennedy Stutz et Morton 45Glomus manihotis Howeler Sieverd amp Schenck 45Glomus mosseae (Nicol amp Gerd) Gerd amp Trappe 273Glomus verruculosum Błaszk 91Glomus sp 1 91 Pacispora scintillans (Rose amp Trappe) Oehl amp Sieverd 136
Paraglomus occultum (Walker) Morton amp Redecker 91Scutellospora calospora (Nicol amp Gerd) Walker 91Scutellospora verrucosa (Koske amp C Walker)Walker amp Sanders
45
References
Addy HD Boswell EP Koide RT (1998) Low temperatureacclimation and freezing resistance of extraradical VA mycor-rhizal hyphae Mycol Res 102582 ndash 586
Allen ME (1991) The ecology of mycorrhiza Cambridge University
Press New York Allsopp N Stock WD (1993) Mycorrhizal status of plants growingin the Cape Floristic Region South Africa Bothalia 2391 ndash 104
Bever JD (2002) Host-specificity of AM fungal population growthrates can generate feedback on plant growth Plant Soil244281 ndash 290
Bever JD Schultz PA Pringle A Morton JB (2001) Arbuscular mycorrhizal fungi more diverse than meets the eye and theecological tale of why Bioscience 51923 ndash 931
Brundrett M (1991) Mycorrhizas in natural ecosystems Adv EcolRes 21171 ndash 313
Brundrett M Bougher N Dell B Grove T Malajczuk N (1996)Working with mycorrhizas in forestry and agriculture ACIAR Monograph 32 Australian Centre for International AgriculturalResearch Canberra
Brundrett MC Jasper DA Ashwath N (1999) Glomalean mycor-rhizal fungi from tropical Australia II The effect of nutrient levels and host species on the isolation of fungi Mycorrhiza8315 ndash 321
Burrows RL Pfleger FL (2002) Arbuscular mycorrhizal fungirespond to increasing plant diversity Can J Bot 80120 ndash 130
Daniels BA Skipper HD (1982) Methods for the recovery andquantitative estimation of propagules from soil In Schenck NC(ed) Methods and principles of mycorrhizal research AmericanPhytopathologic Society St Paul MN pp 29 ndash 35
Fuchs B Haselwandter K (2004) Red list plants colonization byarbuscular mycorrhizal fungi and dark septate endophytesMycorrhiza 14277 ndash 281
Giovannetti M Sbrana C (1998) Meeting a non-host the behaviour of AM fungi Mycorrhiza 8123 ndash 130
Harley JL Harley EL (1987) A checklist of mycorrhiza in theBritish flora New Phytol 105 (Suppl)1 ndash 102
Hirsch AM Kapulnik Y (1998) Signal transduction pathways inmycorrhizal associations comparisons with the Rhizobium -legume symbiosis Fungal Genet Biol 23205 ndash 212
Johnson NC Graham JH Smith FA (1997) Functioning of mycorrhizal associations along the mutualism ndash parasitism con-tinuum New Phytol 135575 ndash 586
Klironomos JN Hart MM Gurney JE Moutoglis P (2001)Interspecific differences in the tolerance of arbuscular mycor-rhizal fungi to freezing and drying Can J Bot 791161 ndash 1166
Koske SE Gemma JN Flynn T (1992) Mycorrhizae in Haiwaiianangiosperms a survey with implications for the origin of thenative flora Am J Bot 79853 ndash 862
Lovera M Cuenca G (1996) Arbuscular mycorrhizal infection inCyperaceae and Gramineae from natural disturbed and restoredsavannas in La Gran Sabana Venezuela Mycorrhiza 6111 ndash 118
7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 77
Lu RK (2000) Methods of soil and agricultural chemistry analyses(in Chinese) China Agricultural Science and Technology PressBeijing
McGonigle TP Fitter AH (1990) Ecological specificity of vesicular ndash
arbuscular mycorrhizal associations Mycol Res 94120 ndash 122Muthukumar T Udaiyan K (2000) Arbuscular mycorrhizas of plants
growing in the Western Ghats region Southern India Mycor-rhiza 9297 ndash 313
Muthukumar T Sha LQ Yang XD Cao M Tang JW Zheng Z(2003) Mycorrhiza of plants in different vegetation types intropical ecosystems of Xishuangbanna southwest ChinaMycorrhiza 13289 ndash 297
Muthukumar T Udaiyan K Shanmughavel P (2004) Mycorrhiza insedges an overview Mycorrhiza 1465 ndash 77
Newman EI Reddell P (1987) The distribution of mycorrhizasamong families of vascular plants New Phytol 106745 ndash 751
Peat HJ Fitter AH (1993) The distribution of arbuscular mycor-rhizas in the British flora New Phytol 125845 ndash 854
Peterson RL Bradbury SM (1995) Use of plant mutants intraspe-cific variants and non-hosts in studying mycorrhiza formationand function In Varma A Hock B (eds) Mycorrhiza SpringerBerlin Heidelberg New York pp 521 ndash 560
Phillips JM Hayman DS (1970) Improved procedures for cleaningand staining parasitic and vesicular arbuscular mycorrhizalfungi for rapid assessment of infection Trans Br Mycol Soc55158 ndash 160
Regvar M Vogel K Irgel N Wraber T Hildebrandt U Wilde PBothe H (2003) Colonization of pennycresses ( Thlaspi spp) of the Brassicaceae by arbuscular mycorrhizal fungi J Plant Physiol 160615 ndash 626
Schenck NC Peacutereacutez Y (1990) Manual for the identification of vesicular ndash arbuscular mycorrhizal fungi INVAM University of Florida Gainesville
Smith SE Read DJ (1997) Mycorrhizal symbiosis 2nd ednAcademic Press London
Tester M Smith SE Smith FA (1987) The phenomenon of lsquo nonmycorrhizal rsquo plants Can J Bot 65419 ndash 431
van der Heijden MGA Klironomos JN Ursic M Moutoglis PStreitwolf-Engel R Boller T Wiemken A Sanders IR (1998)Mycorrhizal fungal diversity determines plant biodiversityecosystem variability and productivity Nature 39669 ndash 72

7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 57
Further Pearsons correlation analysis of colonization lev-els and soil factors (soil pH organic matter and availableP) revealed a strong negative correlation between organicmatter and colonization rate ( r = minus 0733 P lt000 n=22)
AM fungal diversity
Twenty-six taxa of AM fungi were isolated from the soilsamples belonging to Glomus Acaulospora Paraglomus Archaeospora Pacispora and Scutellospora (Table 2)Species richness in the survey areas was 273 Glomus and Acaulospora were the dominant genera both in frequencyand relative abundance The five most commonly observedspecies were Acaulospora scrobiculata Glomus mosseae Glomus intraradices Acaulospora laevis and Glomusclaroideum Of these A scrobiculata was the most fre-quent and abundant species (Table 3)
The species richness and fungal diversity of nine sedge
species differed substantially (Table 4) Species richnessvaried from 10 ( C tibetica ) to 45 ( K prainii ) and diversityfrom 0070 ( Kobresia tibetica ) to 0252 ( K pygmaea ) Norelationship existed between AM fungal diversity (diversityindex spore density) and soil factors (soil pH organicmatter and available P)
Discussion
Here we report for the first time the mycorrhizal status of eight of the nine sedge species investigated Only Cyperuscompressus has previously been reported as mycorrhizal
(Allsopp and Stock 1993 Muthukumar and Udaiyan 2000 )and was defined as facultatively mycorrhizal whenMuthukumar et al ( 2004 ) summarized the published in-formation on the mycorrhizal status of sedges C compres- sus was mycorrhizal in all ecotypes collected in our studyOnly intercellular aseptate hyphae and vesicles were foundin C moorcroftii and K pusilla It has been reported that vesicles and hyphae are found in nonmycorrhizal hosts suchas Cyperus iria and Cyperus rotundus (Koske et al 1992 Giovannetti and Sbrana 1998 ) These plants can therefore bereferred to as of doubtful AM status K humilis was found to be facultatively mycorrhizal in our survey The incidence of
mycorrhizal species in the present study was much higher (67) than the average level of 40 reported Tibetansedges therefore appear to be readily colonized by AMfungi
Vesicles and intercellular aseptate hyphae were the most frequent structures present in the sedge roots and thisaccords with earlier reports (Muthukumar et al 2003 Muthukumar and Udaiyan 2000 ) The incidence of ar- buscules was 46 not a very low value compared withother surveys (Muthukumar et al 2003 Muthukumar andUdaiyan 2000 ) However the percentage of root lengthcolonized by AM fungi was very low in most root samplesand hyphal coils and appressoria were found in only a fewspecimens The potential nutritional benefit of the mycor-rhizal association in sedges therefore has yet to be as-certained More detailed studies are necessary to ascertainthe exact degree of mycorrhizal dependency of Cyperaceaeand the functional aspects of the association in theseTibetan grassland communities
The rarely colonized or nonmycorrhizal status of sedgesis often attributed to the small amount or absence of mycorrhizal propagules in the soils that sedges inhabit (Peat and Fitter 1993 ) However many sedges tend to benonmycorrhizal even in the presence of AM fungal prop-agules (Van der Heijden et al 1998 ) indicating that absence of AM fungal inoculum may not always explainthe low incidence of AM in these plants Spore densities inthe rhizosphere of sedges were determined in our studyand there was no correlation between spore density andcolonization rate Other important factors likely to influ-ence root colonization are soil properties A strong negativecorrelation was found between soil organic matter content and root colonization rate This is in agreement with theanalysis of Muthukumar et al ( 2004 ) However root
colonization was not related to important factors such assoil pH or soil available P which are shown to influence themycorrhizal status of sedges (Lovera and Cuenca 1996 Muthukumar et al 2004 ) One likely explanation could bethe narrow data range in a particular habitat of this studyThe mechanisms by which soil properties and other factorsinfluence mycorrhizal associations still remain unclear andrequire more detailed research
AM fungi are known to exhibit ecological specificity(McGonigle and Fitter 1990 ) There are no published reportson the AM fungal community associated with sedges under the climatic conditions prevailing on the Tibet plateau Onlya few studies have addressed the ability of AM fungi to
survive winter freezing or drought stress (van der Heijden et al 1998 Addy et al 1998 ) The predominance of Glomusand Acaulospora is in accordance with Klironomos et al(2001 ) who studied interspecific differences in AM fungaltolerance to freezing and dry conditions These authorsfound that Glomus and Acaulospora were more frequentlyisolated in the field and Glomus species were the least affected by freezing in pot experiment conditions whiledrying gave more variable responses in colonization by AMfungi Soil properties are important factors influencing theAM fungal community (Bever et al 2001) Acaulosporaspecies are often associated with acid soils Most of the soils
Table 4 Arbuscular mycorrhizal species richness and diversity indifferent sedge species
Host plants Species richness Diversity
C capillacea var linzensis 35plusmn20 0143plusmn0135C moorcroftii 35plusmn10 0201plusmn0120C tibetica 10plusmn00 0067plusmn0000C compressus 24plusmn06 0169plusmn0062
K humilis 20plusmn04 0137plusmn0042 K prainii 45plusmn21 0177plusmn0150 K prattii 30plusmn00 0195plusmn0123 K pygmaea 30plusmn14 0252plusmn0252 K tibetica 20plusmn00 0070plusmn0000
7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 67
in our study were acidic and this could be one explanationfor our frequent detection of Acaulospora
The colonization pattern varied among plant species andeven among ecotypes of particular plant species K humiliswas mycorrhizal or nonmycorrhizal which demonstratesthat mycorrhizal fungi and their hosts occupy different positions on the ldquo mutualism ndash parasitism continuum rdquo under different environmental conditions (Johnson et al 1997 )The AM fungal community composition of particular sedge species varied with habitat type similar to the resultsof Brundrett et al ( 1999 ) G mosseae and G intraradiceswere also found by Klironomos et al ( 2001 ) in similar environmental conditions This indicates that some specieshave higher specific ecological adaptability
Host species is an important factor influencing AMfungal species diversity (Brundrett 1991 Bever et al 2001)Species richness and fungal diversity differed among thenine sedge species in our study It has been shown that AMfungal diversity has a strong effect on the plant communityon plant productivity and on succession (Allen 1991 Bever et al 2001) The available data suggest an important ecological role for AM fungi in the Tibetan sedge commu-nity However full elucidation of the effects of AM fungi onthe sedges still remains unclear and the ecology andimportance of AM associations require further detailedresearch
Acknowledgements We thank the National Natural Science Foun-dation of China (Projects 30370818 30260055 and 30470341) andthe Royal Society (China Exchanges Project 15360) for financialsupport
Appendix
AM fungal species and their frequency of isolation from rhizospheresoils of sedges
Species Frequency Acaulospora appendicula Spain Sieverding amp Schenck 45 Acaulospora dilatata Morton 91 Acaulospora laevis Gerd amp Trappe 227 Acaulospora mellea Spain amp Schenck 45 Acaulospora scrobiculata Trappe 318 Acaulospora spinosa Walker amp Trappe 91 Acaulospora sp 1 91 Acaulospora sp 2 45 Archaeospora gerdemanni (Rose Daniels amp Trappe)
Morton amp Redecker 45Glomus claroideum Schenck amp Sm emend Walker ampVestberg
182
Glomus convolutum Gerd amp Trappe 45Glomus clarum Nicolson amp Schenck 45Glomus diaphanum Morton amp Walker 91Glomus etunicatum Becker amp Gerdemann 91Glomus geosporum (Nicol amp Gerd) Walker 91Glomus glomerulatum Sieverd 45Glomus intraradices Schenck amp Smith 227
Species FrequencyGlomus luteum Kennedy Stutz et Morton 45Glomus manihotis Howeler Sieverd amp Schenck 45Glomus mosseae (Nicol amp Gerd) Gerd amp Trappe 273Glomus verruculosum Błaszk 91Glomus sp 1 91 Pacispora scintillans (Rose amp Trappe) Oehl amp Sieverd 136
Paraglomus occultum (Walker) Morton amp Redecker 91Scutellospora calospora (Nicol amp Gerd) Walker 91Scutellospora verrucosa (Koske amp C Walker)Walker amp Sanders
45
References
Addy HD Boswell EP Koide RT (1998) Low temperatureacclimation and freezing resistance of extraradical VA mycor-rhizal hyphae Mycol Res 102582 ndash 586
Allen ME (1991) The ecology of mycorrhiza Cambridge University
Press New York Allsopp N Stock WD (1993) Mycorrhizal status of plants growingin the Cape Floristic Region South Africa Bothalia 2391 ndash 104
Bever JD (2002) Host-specificity of AM fungal population growthrates can generate feedback on plant growth Plant Soil244281 ndash 290
Bever JD Schultz PA Pringle A Morton JB (2001) Arbuscular mycorrhizal fungi more diverse than meets the eye and theecological tale of why Bioscience 51923 ndash 931
Brundrett M (1991) Mycorrhizas in natural ecosystems Adv EcolRes 21171 ndash 313
Brundrett M Bougher N Dell B Grove T Malajczuk N (1996)Working with mycorrhizas in forestry and agriculture ACIAR Monograph 32 Australian Centre for International AgriculturalResearch Canberra
Brundrett MC Jasper DA Ashwath N (1999) Glomalean mycor-rhizal fungi from tropical Australia II The effect of nutrient levels and host species on the isolation of fungi Mycorrhiza8315 ndash 321
Burrows RL Pfleger FL (2002) Arbuscular mycorrhizal fungirespond to increasing plant diversity Can J Bot 80120 ndash 130
Daniels BA Skipper HD (1982) Methods for the recovery andquantitative estimation of propagules from soil In Schenck NC(ed) Methods and principles of mycorrhizal research AmericanPhytopathologic Society St Paul MN pp 29 ndash 35
Fuchs B Haselwandter K (2004) Red list plants colonization byarbuscular mycorrhizal fungi and dark septate endophytesMycorrhiza 14277 ndash 281
Giovannetti M Sbrana C (1998) Meeting a non-host the behaviour of AM fungi Mycorrhiza 8123 ndash 130
Harley JL Harley EL (1987) A checklist of mycorrhiza in theBritish flora New Phytol 105 (Suppl)1 ndash 102
Hirsch AM Kapulnik Y (1998) Signal transduction pathways inmycorrhizal associations comparisons with the Rhizobium -legume symbiosis Fungal Genet Biol 23205 ndash 212
Johnson NC Graham JH Smith FA (1997) Functioning of mycorrhizal associations along the mutualism ndash parasitism con-tinuum New Phytol 135575 ndash 586
Klironomos JN Hart MM Gurney JE Moutoglis P (2001)Interspecific differences in the tolerance of arbuscular mycor-rhizal fungi to freezing and drying Can J Bot 791161 ndash 1166
Koske SE Gemma JN Flynn T (1992) Mycorrhizae in Haiwaiianangiosperms a survey with implications for the origin of thenative flora Am J Bot 79853 ndash 862
Lovera M Cuenca G (1996) Arbuscular mycorrhizal infection inCyperaceae and Gramineae from natural disturbed and restoredsavannas in La Gran Sabana Venezuela Mycorrhiza 6111 ndash 118
7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 77
Lu RK (2000) Methods of soil and agricultural chemistry analyses(in Chinese) China Agricultural Science and Technology PressBeijing
McGonigle TP Fitter AH (1990) Ecological specificity of vesicular ndash
arbuscular mycorrhizal associations Mycol Res 94120 ndash 122Muthukumar T Udaiyan K (2000) Arbuscular mycorrhizas of plants
growing in the Western Ghats region Southern India Mycor-rhiza 9297 ndash 313
Muthukumar T Sha LQ Yang XD Cao M Tang JW Zheng Z(2003) Mycorrhiza of plants in different vegetation types intropical ecosystems of Xishuangbanna southwest ChinaMycorrhiza 13289 ndash 297
Muthukumar T Udaiyan K Shanmughavel P (2004) Mycorrhiza insedges an overview Mycorrhiza 1465 ndash 77
Newman EI Reddell P (1987) The distribution of mycorrhizasamong families of vascular plants New Phytol 106745 ndash 751
Peat HJ Fitter AH (1993) The distribution of arbuscular mycor-rhizas in the British flora New Phytol 125845 ndash 854
Peterson RL Bradbury SM (1995) Use of plant mutants intraspe-cific variants and non-hosts in studying mycorrhiza formationand function In Varma A Hock B (eds) Mycorrhiza SpringerBerlin Heidelberg New York pp 521 ndash 560
Phillips JM Hayman DS (1970) Improved procedures for cleaningand staining parasitic and vesicular arbuscular mycorrhizalfungi for rapid assessment of infection Trans Br Mycol Soc55158 ndash 160
Regvar M Vogel K Irgel N Wraber T Hildebrandt U Wilde PBothe H (2003) Colonization of pennycresses ( Thlaspi spp) of the Brassicaceae by arbuscular mycorrhizal fungi J Plant Physiol 160615 ndash 626
Schenck NC Peacutereacutez Y (1990) Manual for the identification of vesicular ndash arbuscular mycorrhizal fungi INVAM University of Florida Gainesville
Smith SE Read DJ (1997) Mycorrhizal symbiosis 2nd ednAcademic Press London
Tester M Smith SE Smith FA (1987) The phenomenon of lsquo nonmycorrhizal rsquo plants Can J Bot 65419 ndash 431
van der Heijden MGA Klironomos JN Ursic M Moutoglis PStreitwolf-Engel R Boller T Wiemken A Sanders IR (1998)Mycorrhizal fungal diversity determines plant biodiversityecosystem variability and productivity Nature 39669 ndash 72

7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 67
in our study were acidic and this could be one explanationfor our frequent detection of Acaulospora
The colonization pattern varied among plant species andeven among ecotypes of particular plant species K humiliswas mycorrhizal or nonmycorrhizal which demonstratesthat mycorrhizal fungi and their hosts occupy different positions on the ldquo mutualism ndash parasitism continuum rdquo under different environmental conditions (Johnson et al 1997 )The AM fungal community composition of particular sedge species varied with habitat type similar to the resultsof Brundrett et al ( 1999 ) G mosseae and G intraradiceswere also found by Klironomos et al ( 2001 ) in similar environmental conditions This indicates that some specieshave higher specific ecological adaptability
Host species is an important factor influencing AMfungal species diversity (Brundrett 1991 Bever et al 2001)Species richness and fungal diversity differed among thenine sedge species in our study It has been shown that AMfungal diversity has a strong effect on the plant communityon plant productivity and on succession (Allen 1991 Bever et al 2001) The available data suggest an important ecological role for AM fungi in the Tibetan sedge commu-nity However full elucidation of the effects of AM fungi onthe sedges still remains unclear and the ecology andimportance of AM associations require further detailedresearch
Acknowledgements We thank the National Natural Science Foun-dation of China (Projects 30370818 30260055 and 30470341) andthe Royal Society (China Exchanges Project 15360) for financialsupport
Appendix
AM fungal species and their frequency of isolation from rhizospheresoils of sedges
Species Frequency Acaulospora appendicula Spain Sieverding amp Schenck 45 Acaulospora dilatata Morton 91 Acaulospora laevis Gerd amp Trappe 227 Acaulospora mellea Spain amp Schenck 45 Acaulospora scrobiculata Trappe 318 Acaulospora spinosa Walker amp Trappe 91 Acaulospora sp 1 91 Acaulospora sp 2 45 Archaeospora gerdemanni (Rose Daniels amp Trappe)
Morton amp Redecker 45Glomus claroideum Schenck amp Sm emend Walker ampVestberg
182
Glomus convolutum Gerd amp Trappe 45Glomus clarum Nicolson amp Schenck 45Glomus diaphanum Morton amp Walker 91Glomus etunicatum Becker amp Gerdemann 91Glomus geosporum (Nicol amp Gerd) Walker 91Glomus glomerulatum Sieverd 45Glomus intraradices Schenck amp Smith 227
Species FrequencyGlomus luteum Kennedy Stutz et Morton 45Glomus manihotis Howeler Sieverd amp Schenck 45Glomus mosseae (Nicol amp Gerd) Gerd amp Trappe 273Glomus verruculosum Błaszk 91Glomus sp 1 91 Pacispora scintillans (Rose amp Trappe) Oehl amp Sieverd 136
Paraglomus occultum (Walker) Morton amp Redecker 91Scutellospora calospora (Nicol amp Gerd) Walker 91Scutellospora verrucosa (Koske amp C Walker)Walker amp Sanders
45
References
Addy HD Boswell EP Koide RT (1998) Low temperatureacclimation and freezing resistance of extraradical VA mycor-rhizal hyphae Mycol Res 102582 ndash 586
Allen ME (1991) The ecology of mycorrhiza Cambridge University
Press New York Allsopp N Stock WD (1993) Mycorrhizal status of plants growingin the Cape Floristic Region South Africa Bothalia 2391 ndash 104
Bever JD (2002) Host-specificity of AM fungal population growthrates can generate feedback on plant growth Plant Soil244281 ndash 290
Bever JD Schultz PA Pringle A Morton JB (2001) Arbuscular mycorrhizal fungi more diverse than meets the eye and theecological tale of why Bioscience 51923 ndash 931
Brundrett M (1991) Mycorrhizas in natural ecosystems Adv EcolRes 21171 ndash 313
Brundrett M Bougher N Dell B Grove T Malajczuk N (1996)Working with mycorrhizas in forestry and agriculture ACIAR Monograph 32 Australian Centre for International AgriculturalResearch Canberra
Brundrett MC Jasper DA Ashwath N (1999) Glomalean mycor-rhizal fungi from tropical Australia II The effect of nutrient levels and host species on the isolation of fungi Mycorrhiza8315 ndash 321
Burrows RL Pfleger FL (2002) Arbuscular mycorrhizal fungirespond to increasing plant diversity Can J Bot 80120 ndash 130
Daniels BA Skipper HD (1982) Methods for the recovery andquantitative estimation of propagules from soil In Schenck NC(ed) Methods and principles of mycorrhizal research AmericanPhytopathologic Society St Paul MN pp 29 ndash 35
Fuchs B Haselwandter K (2004) Red list plants colonization byarbuscular mycorrhizal fungi and dark septate endophytesMycorrhiza 14277 ndash 281
Giovannetti M Sbrana C (1998) Meeting a non-host the behaviour of AM fungi Mycorrhiza 8123 ndash 130
Harley JL Harley EL (1987) A checklist of mycorrhiza in theBritish flora New Phytol 105 (Suppl)1 ndash 102
Hirsch AM Kapulnik Y (1998) Signal transduction pathways inmycorrhizal associations comparisons with the Rhizobium -legume symbiosis Fungal Genet Biol 23205 ndash 212
Johnson NC Graham JH Smith FA (1997) Functioning of mycorrhizal associations along the mutualism ndash parasitism con-tinuum New Phytol 135575 ndash 586
Klironomos JN Hart MM Gurney JE Moutoglis P (2001)Interspecific differences in the tolerance of arbuscular mycor-rhizal fungi to freezing and drying Can J Bot 791161 ndash 1166
Koske SE Gemma JN Flynn T (1992) Mycorrhizae in Haiwaiianangiosperms a survey with implications for the origin of thenative flora Am J Bot 79853 ndash 862
Lovera M Cuenca G (1996) Arbuscular mycorrhizal infection inCyperaceae and Gramineae from natural disturbed and restoredsavannas in La Gran Sabana Venezuela Mycorrhiza 6111 ndash 118
7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 77
Lu RK (2000) Methods of soil and agricultural chemistry analyses(in Chinese) China Agricultural Science and Technology PressBeijing
McGonigle TP Fitter AH (1990) Ecological specificity of vesicular ndash
arbuscular mycorrhizal associations Mycol Res 94120 ndash 122Muthukumar T Udaiyan K (2000) Arbuscular mycorrhizas of plants
growing in the Western Ghats region Southern India Mycor-rhiza 9297 ndash 313
Muthukumar T Sha LQ Yang XD Cao M Tang JW Zheng Z(2003) Mycorrhiza of plants in different vegetation types intropical ecosystems of Xishuangbanna southwest ChinaMycorrhiza 13289 ndash 297
Muthukumar T Udaiyan K Shanmughavel P (2004) Mycorrhiza insedges an overview Mycorrhiza 1465 ndash 77
Newman EI Reddell P (1987) The distribution of mycorrhizasamong families of vascular plants New Phytol 106745 ndash 751
Peat HJ Fitter AH (1993) The distribution of arbuscular mycor-rhizas in the British flora New Phytol 125845 ndash 854
Peterson RL Bradbury SM (1995) Use of plant mutants intraspe-cific variants and non-hosts in studying mycorrhiza formationand function In Varma A Hock B (eds) Mycorrhiza SpringerBerlin Heidelberg New York pp 521 ndash 560
Phillips JM Hayman DS (1970) Improved procedures for cleaningand staining parasitic and vesicular arbuscular mycorrhizalfungi for rapid assessment of infection Trans Br Mycol Soc55158 ndash 160
Regvar M Vogel K Irgel N Wraber T Hildebrandt U Wilde PBothe H (2003) Colonization of pennycresses ( Thlaspi spp) of the Brassicaceae by arbuscular mycorrhizal fungi J Plant Physiol 160615 ndash 626
Schenck NC Peacutereacutez Y (1990) Manual for the identification of vesicular ndash arbuscular mycorrhizal fungi INVAM University of Florida Gainesville
Smith SE Read DJ (1997) Mycorrhizal symbiosis 2nd ednAcademic Press London
Tester M Smith SE Smith FA (1987) The phenomenon of lsquo nonmycorrhizal rsquo plants Can J Bot 65419 ndash 431
van der Heijden MGA Klironomos JN Ursic M Moutoglis PStreitwolf-Engel R Boller T Wiemken A Sanders IR (1998)Mycorrhizal fungal diversity determines plant biodiversityecosystem variability and productivity Nature 39669 ndash 72

7252019 Arbuscular in Tibet
httpslidepdfcomreaderfullarbuscular-in-tibet 77
Lu RK (2000) Methods of soil and agricultural chemistry analyses(in Chinese) China Agricultural Science and Technology PressBeijing
McGonigle TP Fitter AH (1990) Ecological specificity of vesicular ndash
arbuscular mycorrhizal associations Mycol Res 94120 ndash 122Muthukumar T Udaiyan K (2000) Arbuscular mycorrhizas of plants
growing in the Western Ghats region Southern India Mycor-rhiza 9297 ndash 313
Muthukumar T Sha LQ Yang XD Cao M Tang JW Zheng Z(2003) Mycorrhiza of plants in different vegetation types intropical ecosystems of Xishuangbanna southwest ChinaMycorrhiza 13289 ndash 297
Muthukumar T Udaiyan K Shanmughavel P (2004) Mycorrhiza insedges an overview Mycorrhiza 1465 ndash 77
Newman EI Reddell P (1987) The distribution of mycorrhizasamong families of vascular plants New Phytol 106745 ndash 751
Peat HJ Fitter AH (1993) The distribution of arbuscular mycor-rhizas in the British flora New Phytol 125845 ndash 854
Peterson RL Bradbury SM (1995) Use of plant mutants intraspe-cific variants and non-hosts in studying mycorrhiza formationand function In Varma A Hock B (eds) Mycorrhiza SpringerBerlin Heidelberg New York pp 521 ndash 560
Phillips JM Hayman DS (1970) Improved procedures for cleaningand staining parasitic and vesicular arbuscular mycorrhizalfungi for rapid assessment of infection Trans Br Mycol Soc55158 ndash 160
Regvar M Vogel K Irgel N Wraber T Hildebrandt U Wilde PBothe H (2003) Colonization of pennycresses ( Thlaspi spp) of the Brassicaceae by arbuscular mycorrhizal fungi J Plant Physiol 160615 ndash 626
Schenck NC Peacutereacutez Y (1990) Manual for the identification of vesicular ndash arbuscular mycorrhizal fungi INVAM University of Florida Gainesville
Smith SE Read DJ (1997) Mycorrhizal symbiosis 2nd ednAcademic Press London
Tester M Smith SE Smith FA (1987) The phenomenon of lsquo nonmycorrhizal rsquo plants Can J Bot 65419 ndash 431
van der Heijden MGA Klironomos JN Ursic M Moutoglis PStreitwolf-Engel R Boller T Wiemken A Sanders IR (1998)Mycorrhizal fungal diversity determines plant biodiversityecosystem variability and productivity Nature 39669 ndash 72