arbuscular mycorrhizal associations in lycopodiaceae...
TRANSCRIPT

790
www.newphytologist.org
Research
Blackwell Publishing Ltd
Arbuscular mycorrhizal associations in Lycopodiaceae
Jennifer L. Winther and William E. Friedman
University of Colorado, Department of Ecology and Evolutionary Biology, Boulder, Colorado 80309-0334, USA
Summary
• This study characterizes the molecular and phylogenetic identity of fungi involvedin arbuscular mycorrhizal (AM) associations in extant
Huperzia
and
Lycopodium
(Lycopodiaceae).
Huperzia
and
Lycopodium
are characterized by a life cycle withlong-lived autotrophic sporophytes and long-lived mycoheterotrophic (obtain allorganic carbon from fungal symbionts) gametophytes.• 18S ribosomal DNA was isolated and sequenced from
Glomus
symbionts inautotrophic sporophytes of seven species of
Huperzia
and
Lycopodium
andmycoheterotrophic
Huperzia
gametophytes collected from the Páramos of Ecuador.• Phylogenetic analyses recovered four
Glomus A
phylotypes in a single clade(MH3) that form AM associations with
Huperzia
and
Lycopodium
. In addition,phylogenetic analyses of
Glomus
symbionts from other nonphotosynthetic plantsdemonstrate that most AM fungi that form mycoheterotrophic associations belongto at least four specific clades of
Glomus A
.• These results suggest that most mycoheterotrophic plants that form AMassociations do so with restricted clades of
Glomus A
. Moreover, the correspondenceof identity of AM symbionts in
Huperzia
sporophytes and gametophytes raises thepossibility that photosynthetic sporophytes are a source of carbon to conspecificmycoheterotrophic gametophytes via shared fungal networks.
Key words:
arbuscular mycorrhizal (AM) associations,
Glomus
,
Huperzia
,Lycopodiaceae,
Lycopodium
, mycoheterotrophy.
New Phytologist
(2008)
177
: 790–801
© The Authors (2007). Journal compilation ©
New Phytologist
(2007)
doi
: 10.1111/j.1469-8137.2007.02276.x
Author for correspondence:
William E. FriedmanTel:
+
1 303 4923082Fax:
+
1 303 4928699Email: [email protected]
Received:
7 August 2007
Accepted:
18 September 2007
Introduction
Most land plants, from liverworts to angiosperms, formmutualistic arbuscular mycorrhizal (AM) symbioses withfungi in the Glomeromycota (glomalean fungi) where theplant host gains essential mineral nutrients from the fungalsymbiont in exchange for fixed carbon (Smith & Read, 1997;Schussler
et al
., 2001). It is hypothesized that the evolution ofAM symbioses was critical to the successful establishment andlater diversification of land plants (Pirozynski & Malloch,1975; Simon
et al
., 1993a; Remy
et al
., 1994; Simon, 1995;Redecker
et al
., 2000). However, current understanding ofthe diversity, physiology, ecology, and evolution of glomaleanfungi symbioses is mostly limited to AM associations inphotosynthetic angiosperms (Smith & Read, 1997; Brundrett,2002, 2004). Notably, most studies have overlooked AM
associations in ferns and lycophytes, ancient lineages thatcommonly have mycoheterotrophic life cycle phases that gainall of their organic carbon through their glomalean symbionts(Bierhorst, 1971; Leake, 1993, 2004; Wang & Qiu, 2006).In order to address the broader evolutionary and ecologicalhistory of AM associations across the almost 500 million yrhistory of land plants, glomalean symbioses from a broadersampling of plant diversity need to be characterized.
The focus of this study is on AM associations in extantLycopodiaceae, a family in the ancient and diverse lycophyteclade that is sister to all other extant vascular plants (Kenrick& Crane, 1997). Members of this family (
Huperzia
,
Lycopodium
,
Lycopodiella
and
Phylloglossum
) are characterized by a life cyclewith independent sporophyte and long-lived (several years)gametophyte phases (Bierhorst, 1971; Ollgaard, 1992; Kenrick& Crane, 1997). AM associations can be found throughout

© The Authors (2007). Journal compilation ©
New Phytologist
(2007)
www.newphytologist.org
New Phytologist
(2008)
177
: 790–801
Research 791
the life cycle of these four genera: from the gametophyte, tothe young sporophyte that is still attached to the gametophyte,to the mature sporophyte (Bierhorst, 1971).
It is assumed that Lycopodiaceae sporophytes have typicalmutualistic AM associations (Smith & Read, 1997). However,a review of the literature reveals that there is little consensusconcerning AM associations in the sporophytes of
Huperzia
,
Lycopodium
,
Lycopodiella
and
Phylloglossum
. Surveys of AMassociations in Lycopodiaceae report that AM associations areonly sometimes found within individuals of a single speciesand that some species have never been found to have AMassociations (Boullard, 1957; Harley & Harley, 1987; Gemma
et al
., 1992; Treu
et al
., 1996; Zhao, 2000; Fuchs & Hasel-wandter, 2004). Furthermore, other types of mycorrhizalassociations (i.e. ericoid, orchid, microsclerotia) with septate,nonglomalean fungi (Ascomycota and Basidiomycota) havebeen reported in the roots of multiple
Lycopodium
speciesinstead of, and in addition to, AM associations (Spessard,1917; Freeberg, 1962; Bruce, 1979a; Treu
et al
., 1996).The long-lived gametophytes of Lycopodiaceae fall into
two general categories: photosynthetic surface dwelling, andnonphotosynthetic subterranean (Bierhorst, 1971). Extensivemicroscopy since the late 1800 s (Treub, 1884, 1886, 1888a,b,1890a,b; Bruchmann, 1885, 1898; Holloway, 1910, 1916,1935; Chamberlain, 1917; Spessard, 1917; Bruce, 1979a,b;Schmid & Oberwinkler, 1993) shows that both above-groundand below-ground gametophytes have fungal symbionts thatform AM associations.
In nature, all
Huperzia
,
Lycopodium
,
Lycopodiella
and
Phylloglossum
gametophytes, whether photosynthetic or non-photosynthetic, appear to require mycorrhizal symbionts forgrowth and the production of gametes (Treub, 1884, 1886,1888a,b, 1890a,b; Bruchmann, 1885, 1898; Bruce, 1979a,b;Schmid & Oberwinkler, 1993; Whittier
et al
., 2005). Sporesgrown axenically require sugars in order to develop intogametophytes without a fungal symbiont (Whittier & Webster,1986; Whittier & Braggins, 1992; Whittier, 1998). Thus, bothsurface and subterranean gametophytes are consideredobligate mycorrhizal symbionts (Kenrick & Crane, 1997).
Members of the Lycopodiaceae are found throughout theworld, from wet alpine meadows to tropical rainforests, andtheir habitats range from terrestrial to epiphytic (Bierhorst,1971; Kenrick & Crane, 1997). We chose to collect from thehigh-altitude tropical mountain habitats (Páramos) ofEcuador, a center of Lycopodiaceae diversity (Ollgaard, 1992).Plant material from seven species (
H. hypogeae
,
H. urbani
,
H. tetragona
,
H. crassa
var.
crassa
,
H. affinis
,
H. hypogeae
,
L.calvatum
ssp.
clavatum
and
L. clavatum
ssp.
contiguum
) in twogenera,
Huperzia
and
Lycopodium
, was collected throughoutthis region (Fig. 1, Table 1).
Huperzia
and
Lycopodium
gametophytes, in particular subterranean gametophytes, arenotoriously hard to find (Holloway, 1916; Bruce & Bietel,1979; Whittier, 1998). Although gametophytes were searchedfor at all sites, only two subterranean gametophytes were
found, both near
H. hypogeae
sporophytes. Each gametophytewas later confirmed as
H. hypogeae
based on DNA sequences.Our broad sampling of
Lycopodium
and
Huperzia
speciesacross the Páramos of Ecuador, and the fortuitous recovery ofunderground gametophytes, enabled us to address three
Fig. 1 Map of Ecuador with collecting sites (modified from Ecuador physical map at http://oddens.geog.uu.nl/result.php?limit_result=0&zoek=ecuador&themecountry=0&category=0&submitButtonName=Search). Numbers correspond to collecting sites in Table 1.
Table 1 Huperzia and Lycopodium species sampled from each site and the Glomus 18S phylotype isolated from each species
Collection site and species sampledGlomus 18S phylotype isolated
Site 1Lycopodium clavatum ssp. contiguum CHuperzia tetragona B
Site 2Huperzia crassa var. crassa CHuperzia tetragona C, D
Site 3Huperzia affinis C, D
Site 4Huperzia hypogeae A, C, D
Site 5Lycopodium clavatum ssp. clavatum D
Site 6Huperzia urbanii C, D
Site number corresponds to those in Fig. 1; phylotype letters correspond to those in Fig. 4.

New Phytologist
(2008)
177
: 790–801
www.newphytologist.org
© The Authors (2007). Journal compilation ©
New Phytologist
(2007)
Research792
questions concerning the ecology and evolution of glomaleansymbionts that form AM associations in this ancient lineageof vascular land plants:• What are the molecular/phylogenetic identities of AMsymbionts in the mycoheterotrophic gametophyte andphotosynthetic sporophyte of
H. hypogeae
?• What is the distribution and phylogenetic identity of AMsymbionts across a range of seven species of
Huperzia
and
Lycopodium
?• How are Lycopodiaceae AM symbionts phylogeneticallyrelated to other Glomeromycota taxa, especially those knownto engage in AM associations with other mycoheterotrophicplant species?
Materials and Methods
Collection
In May 2004, seven
Lycopodium
and
Huperzia
species (
H.hypogeae
,
H. urbanii
,
H. tetragona
,
H. crassa
var.
crassa
,
H.affinis
,
L. calvatum
ssp.
clavatum
and
L. clavatum
ssp.
contiguum
)were collected from six sites in the Páramos of Ecuador(Table 1, Fig. 1). The collections were predominately fromgrass Páramos habitats dominated by tussock grasslandsusually composed of species of
Calamagrostis
and/or Festuca(Luteyn, 1999). Voucher specimens for each of the sevenspecies collected were deposited at the University of ColoradoHerbarium.
For each species, roots were collected throughout the axis(roots are shoot-borne) from a single sporophyte. Some rootswere surface-sterilized in a 5% bleach solution, rinsed threetimes with sterile water and placed in silica gel for futureextraction of DNA. The remaining roots were fixed in formalinacetic acid (FAA) for anatomical study. The gametophyteswere isolated by sifting soil collected next to the H. hypogeaesporophyte using USA standard sieves no. 80 and no. 20.Surface-sterilized gametophytes were cut in half. Half of eachgametophyte was frozen for DNA extraction, amplification,and sequencing, and the other half was chemically fixed formicroscopy.
Light microscopy
Tissues were fixed in the field in FAA. The plant materialwas dehydrated in an ethanol series to 95%, and infiltratedand embedded in glycol methacrylate ( JB-4 embedding kit,Polysciences, Warrington, PA, USA). Five-micrometer serialsections of plastic-embedded specimens were made using aLeica rotary microtome (Leica Microsystems, Bannockburn,IL, USA) and glass knives. Serial sections were mounted onslides and stained with 0.1% toluidine blue. Slides showingimportant features were digitally photographed using a ZeissAxiophot microscope equipped with a Zeiss Axiocam digitalcamera (Zeiss MicroImaging, Thornwood, NY, USA).
Sequence amplification
DNA was extracted from surface-sterilized silica dried rootmaterial or frozen gametophyte tissue by grinding tissue inCTAB buffer and using the phenol-chloroform extractionmethod (DeSalle et al., 2002). The nuclear small subunitribosomal DNA (18S) was amplified using GEOA2, GEO11(Schwarzott & Schussler, 2001), and SS1492 (Simon et al.,1993b) primers previously used in the amplification of alllineages of Glomeromycota fungi from plant roots andisolated spores. It should be noted that GEOA2 and GEO11primers have previously only been successful in nested PCRreactions (Schwarzott & Schussler, 2001), and recent studieshave developed additional PCR primers and protocols thatreliably amplify AM 18S and ITS sequences (Hijri et al.,2006). The nuclear ribosomal internal transcribed spacerregion (ITS), including ITS1, 5.8S and ITS2, was amplifiedusing fungal specific primer ITS1F (Redecker et al., 1997)and ITS4 (White et al., 1990). The PCR program was 95°Cfor 1 min 30 s, 35 cycles of 94°C for 45 s, 56.5°C for 55 s,72°C for 2 min 30 s + 1 s per cycle, hold 72°C for 10 min.
Sequencing
All amplified DNA samples (25 18S, 17 ITS) were clonedusing TOPO TA cloning kits (Invitrogen, Carlsbad, CA, USA).A minimum of 12, and usually 20, colonies were chosen fromeach reaction for sequencing. Colonies were isolated andgrown in TSB broth at 35°C for 18 h. Plasmid DNA wasextracted using the Mini Wizard Prep Plasmid Kit (Promega,Madison, WI, USA). Extracted plasmid DNA was useddirectly in a cycle sequence reaction using the DYEnamic ETTerminator Cycle Sequencing Kit (Amersham Biosciences,Piscataway, NJ, USA). The cycle sequence reaction mixturewas 2 µl of sequencing reagent premix, 2 µl of template,0.33 µl of primer, 7 µl of water, 0.75 µl of 5 × sequencingbuffer. The cycle sequence reaction was 96°C for 10 s, 50°Cfor 5 s, 60°C for 4 min, repeated 25 times. The cycle sequencereactions were cleaned using Sephadex Column System(Applied Biosystems, Foster City, CA, USA). Samples wererun at the University of Colorado Sequencing Facility in theDepartment of Ecology and Evolutionary Biology on an MJResearch Base Station 51. Additional cloned PCR productswere cleaned with shrimp alkaline phosphatase (USB, Cleveland,OH, USA) and sent to Macrogen (Rockville, MD, USA) forcycle sequencing and gel runs. 18S sequences (EF558776–EF558800) and ITS sequences (EF558801–EF55837)have been submitted to GenBank (www.ncbi.nlm.nih.gov/GenBank/).
Phylogenetic analyses
Each 18S and ITS sequence was run through a BLASTN search(http://www.ncbi.nlm.nih.gov/blast/) on GenBank to confirm

© The Authors (2007). Journal compilation © New Phytologist (2007) www.newphytologist.org New Phytologist (2008) 177: 790–801
Research 793
it as glomalean. Sequences were aligned and edited in ClustalX 1.83 (Thompson et al., 1997), Sequencher version 4.2 (GeneCodes Corporation, Ann Arbor, MI, USA), and MacCladeversion 4.6 OS X (Sinauer Associates, Sunderland, MA, USA)(see Appendix S1, Supplementary Material, for sequencealignment). Neighbor-joining (NJ), maximum-parsimony(MP) and Bayesian analyses were conducted in PAUP v.4.2.b(Swofford, 2002) and MrBayes (Huelsenbeck & Ronquist,2001). Bayesian analyses were carried out using models ofevolution determined by Modeltest (Posada & Crandall, 1998).Published 18S Glomaceae, Acaulosporaceae, Gigasporaceae,Archaeosporaceae, Geosiphonaceae and Paraglomaceae sequencesused in the present phylogenetic analyses are from Schwarzottet al. (2001). We included previously recovered 18S Glomussymbionts from field-collected mycoheterotrophic andautotrophic life cycle stages of Botrychium crenulatum andBotrychium lanceolatum (Winther & Friedman, 2007) and 18Ssequences recovered from the mycoheterotrophic angiosperms,Voyria, Voyriella, and Arachnitis in our phylogenetic analyses(Bidartondo et al., 2002) (see Appendix S2 for accessionnumbers).
Results
Light microscopy
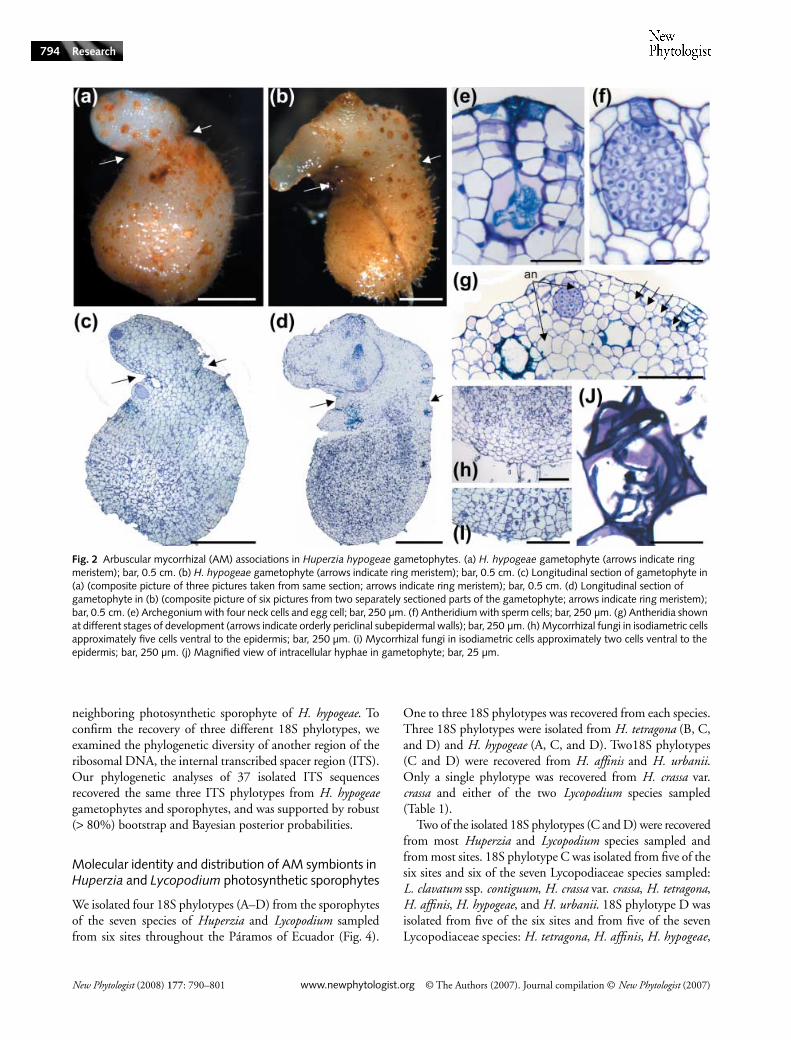
The H. hypogeae gametophytes are orientated with gametangiaon the top of the gametophyte, as has been reported in fieldand germination studies (Treub, 1884, 1886, 1888a,b, 1890a,b;Bruchmann, 1885; Holloway, 1910, 1916; Chamberlain,1917; Spessard, 1917; Bruce, 1979a,b; Whittier & Webster,1986; Whittier, 1998). H. hypogeae gametophytes have abulbous reddish-brown basal region that comprises most ofthe body of the gametophyte. A narrow region (indicatedby arrows in Fig. 2a–d) corresponds to the ring meristem(Bierhorst, 1971) that demarcates the boundary between theupper whitish oval region and the larger basal region (Fig. 2a–d).
Antheridia and archegonia are found predominately alongthe perimeter of the upper portion of the bisexual gametophyteand are sunken ventrally (Fig. 2e–g). The antheridia andarchegonia in the two gametophytes are in various states ofdevelopment from initiation to senescence (Fig. 2g).
Arbuscular–mycorrhizal fungi in the gametophytes arerestricted to the basal bulbous portion of the gametophyte(Fig. 2c,d,h,i) below the antheridia and archegonia. Hyphaeare aseptate and typically intracellular, and are found inisodiametric cells similar to those reported by Whittier &Webster (1986) for the gametophytes of Lycopodium lucidulum(Fig. 2c,d,h–j). Most of the mycorrhizal fungi are concentratedin the centers of the gametophytes, two to five cells internalto the epidermis (Fig. 2h,i).
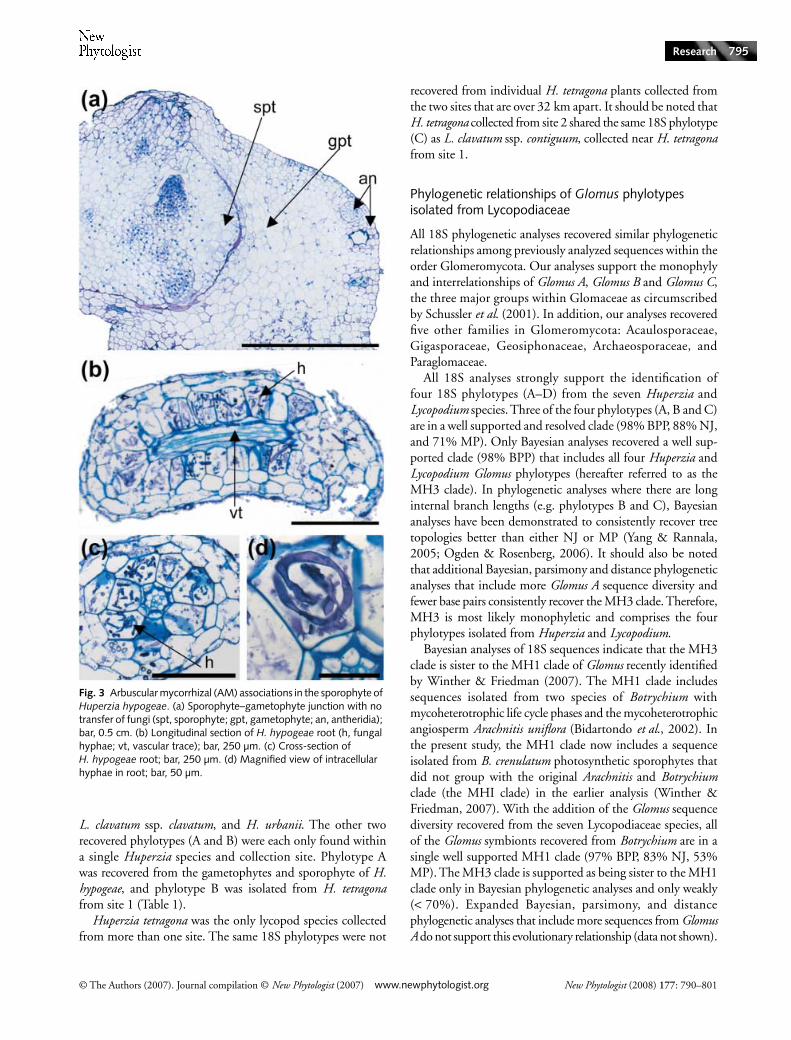
A single young sporophyte was observed growing laterallyfrom one of the H. hypogeae gametophytes (Figs 2b,d, 3a). Atthe interface between the gametophyte and sporophyte tissue
there are no fungal hyphae. No mycorrhizal symbionts werefound in the primary root of the young sporophyte (data notshown).
The AM fungi in the roots of Huperzia and Lycopodiumsporophytes are found predominately intracellularly withinthe large cortical parenchyma cells that surround the vasculartissue (Fig. 3b–d). The AM fungi found in sporophytes areaseptate and have a coil-like appearance (Fig. 3d). Not all of theroots sampled from each plant had AM associations. Typically,the smaller-diameter roots sectioned from the seven Huperziaand Lycopodium species had AM associations in the cortex.
Molecular identity of isolated 18S sequences
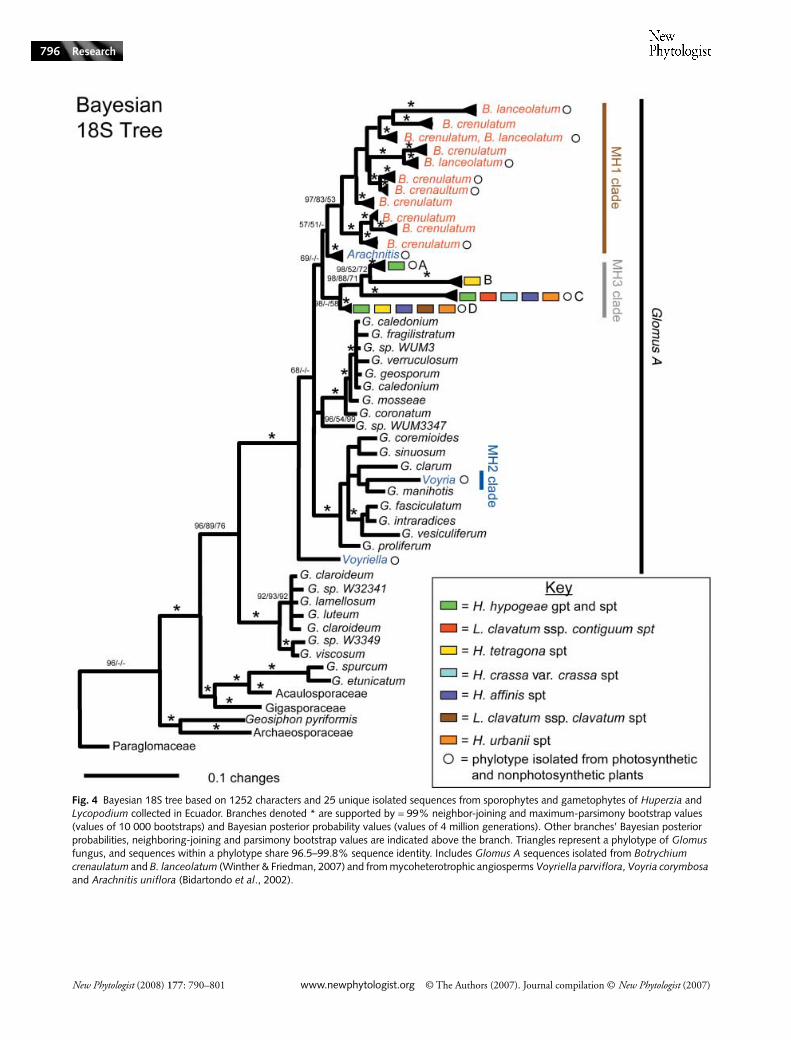
The 18S Bayesian phylogeny (Fig. 4), based on 1252 charactersand 25 unique fungal sequences isolated from seven speciesof Lycopodiaceae, is fairly well resolved. All 25 isolatedsequences are in the family Glomaceae and in Glomus groupA as circumscribed by Schussler et al. (2001). The recoveredsequences represent previously unknown glomalean sequencediversity based on both phylogenetic analyses and GenBanksearches.
The diversity of glomalean symbionts within a plant can becharacterized phylogenetically via the number of recoveredphylotypes, for example, clades of closely related sequenceswith high statistical support. Previous studies have identifiedGlomus phylotypes with bootstrap values from 77 to 100%and sequence identity within the phylotypes that ranges from96.5 to 100% (Vandenkoornhuyse et al., 2002; Opik et al.,2003; Gollotte et al., 2004; Rosendahl & Stukenbrock, 2004;Winther & Friedman, 2007). Our phylogenetic analysespartition the 25 18S sequences isolated from Huperzia andLycopodium into four phylotypes supported by > 95% NJ andMP bootstrap values and > 95% Bayesian posterior probability(BPP) values. Sequence identity within the phylotypes rangesfrom 99.3 to 99.8%. These four phylotypes (A–D) cannot befurther divided into statistically robust groupings.
We are not equating the four 18S glomalean phylotypeswith glomalean species. Glomalean fungi are asexual, multinu-cleate organisms, and 18S and ITS sequences isolated fromsingle spores have been shown to vary 0.5–3% and 6–18%,respectively (Clapp et al., 1995; Antoniolli et al., 2000;Kjoller & Rosendahl, 2001; Jansa et al., 2002). As a result,circumscription of glomalean species based on moleculardata can be difficult. The isolated 18S phylotypes representdiscrete evolutionary units that are statistically robustphylogenetic groupings of genotypes that share an evolutionaryhistory (Winther & Friedman, 2007).
Molecular identity of AM symbionts in gametophytes and sporophyte roots of H. hypogeae
Three 18S phylotypes (A, C, and D) were isolated from eachof the two underground gametophytes and roots of the
New Phytologist (2008) 177: 790–801 www.newphytologist.org © The Authors (2007). Journal compilation © New Phytologist (2007)
Research794
neighboring photosynthetic sporophyte of H. hypogeae. Toconfirm the recovery of three different 18S phylotypes, weexamined the phylogenetic diversity of another region of theribosomal DNA, the internal transcribed spacer region (ITS).Our phylogenetic analyses of 37 isolated ITS sequencesrecovered the same three ITS phylotypes from H. hypogeaegametophytes and sporophytes, and was supported by robust(> 80%) bootstrap and Bayesian posterior probabilities.
Molecular identity and distribution of AM symbionts in Huperzia and Lycopodium photosynthetic sporophytes
We isolated four 18S phylotypes (A–D) from the sporophytesof the seven species of Huperzia and Lycopodium sampledfrom six sites throughout the Páramos of Ecuador (Fig. 4).
One to three 18S phylotypes was recovered from each species.Three 18S phylotypes were isolated from H. tetragona (B, C,and D) and H. hypogeae (A, C, and D). Two18S phylotypes(C and D) were recovered from H. affinis and H. urbanii.Only a single phylotype was recovered from H. crassa var.crassa and either of the two Lycopodium species sampled(Table 1).
Two of the isolated 18S phylotypes (C and D) were recoveredfrom most Huperzia and Lycopodium species sampled andfrom most sites. 18S phylotype C was isolated from five of thesix sites and six of the seven Lycopodiaceae species sampled:L. clavatum ssp. contiguum, H. crassa var. crassa, H. tetragona,H. affinis, H. hypogeae, and H. urbanii. 18S phylotype D wasisolated from five of the six sites and from five of the sevenLycopodiaceae species: H. tetragona, H. affinis, H. hypogeae,
Fig. 2 Arbuscular mycorrhizal (AM) associations in Huperzia hypogeae gametophytes. (a) H. hypogeae gametophyte (arrows indicate ring meristem); bar, 0.5 cm. (b) H. hypogeae gametophyte (arrows indicate ring meristem); bar, 0.5 cm. (c) Longitudinal section of gametophyte in (a) (composite picture of three pictures taken from same section; arrows indicate ring meristem); bar, 0.5 cm. (d) Longitudinal section of gametophyte in (b) (composite picture of six pictures from two separately sectioned parts of the gametophyte; arrows indicate ring meristem); bar, 0.5 cm. (e) Archegonium with four neck cells and egg cell; bar, 250 µm. (f) Antheridium with sperm cells; bar, 250 µm. (g) Antheridia shown at different stages of development (arrows indicate orderly periclinal subepidermal walls); bar, 250 µm. (h) Mycorrhizal fungi in isodiametric cells approximately five cells ventral to the epidermis; bar, 250 µm. (i) Mycorrhizal fungi in isodiametric cells approximately two cells ventral to the epidermis; bar, 250 µm. (j) Magnified view of intracellular hyphae in gametophyte; bar, 25 µm.
© The Authors (2007). Journal compilation © New Phytologist (2007) www.newphytologist.org New Phytologist (2008) 177: 790–801
Research 795
L. clavatum ssp. clavatum, and H. urbanii. The other tworecovered phylotypes (A and B) were each only found withina single Huperzia species and collection site. Phylotype Awas recovered from the gametophytes and sporophyte of H.hypogeae, and phylotype B was isolated from H. tetragonafrom site 1 (Table 1).
Huperzia tetragona was the only lycopod species collectedfrom more than one site. The same 18S phylotypes were not
recovered from individual H. tetragona plants collected fromthe two sites that are over 32 km apart. It should be noted thatH. tetragona collected from site 2 shared the same 18S phylotype(C) as L. clavatum ssp. contiguum, collected near H. tetragonafrom site 1.
Phylogenetic relationships of Glomus phylotypes isolated from Lycopodiaceae
All 18S phylogenetic analyses recovered similar phylogeneticrelationships among previously analyzed sequences within theorder Glomeromycota. Our analyses support the monophylyand interrelationships of Glomus A, Glomus B and Glomus C,the three major groups within Glomaceae as circumscribedby Schussler et al. (2001). In addition, our analyses recoveredfive other families in Glomeromycota: Acaulosporaceae,Gigasporaceae, Geosiphonaceae, Archaeosporaceae, andParaglomaceae.
All 18S analyses strongly support the identification offour 18S phylotypes (A–D) from the seven Huperzia andLycopodium species. Three of the four phylotypes (A, B and C)are in a well supported and resolved clade (98% BPP, 88% NJ,and 71% MP). Only Bayesian analyses recovered a well sup-ported clade (98% BPP) that includes all four Huperzia andLycopodium Glomus phylotypes (hereafter referred to as theMH3 clade). In phylogenetic analyses where there are longinternal branch lengths (e.g. phylotypes B and C), Bayesiananalyses have been demonstrated to consistently recover treetopologies better than either NJ or MP (Yang & Rannala,2005; Ogden & Rosenberg, 2006). It should also be notedthat additional Bayesian, parsimony and distance phylogeneticanalyses that include more Glomus A sequence diversity andfewer base pairs consistently recover the MH3 clade. Therefore,MH3 is most likely monophyletic and comprises the fourphylotypes isolated from Huperzia and Lycopodium.
Bayesian analyses of 18S sequences indicate that the MH3clade is sister to the MH1 clade of Glomus recently identifiedby Winther & Friedman (2007). The MH1 clade includessequences isolated from two species of Botrychium withmycoheterotrophic life cycle phases and the mycoheterotrophicangiosperm Arachnitis uniflora (Bidartondo et al., 2002). Inthe present study, the MH1 clade now includes a sequenceisolated from B. crenulatum photosynthetic sporophytes thatdid not group with the original Arachnitis and Botrychiumclade (the MHI clade) in the earlier analysis (Winther &Friedman, 2007). With the addition of the Glomus sequencediversity recovered from the seven Lycopodiaceae species, allof the Glomus symbionts recovered from Botrychium are in asingle well supported MH1 clade (97% BPP, 83% NJ, 53%MP). The MH3 clade is supported as being sister to the MH1clade only in Bayesian phylogenetic analyses and only weakly(< 70%). Expanded Bayesian, parsimony, and distancephylogenetic analyses that include more sequences from GlomusA do not support this evolutionary relationship (data not shown).
Fig. 3 Arbuscular mycorrhizal (AM) associations in the sporophyte of Huperzia hypogeae. (a) Sporophyte–gametophyte junction with no transfer of fungi (spt, sporophyte; gpt, gametophyte; an, antheridia); bar, 0.5 cm. (b) Longitudinal section of H. hypogeae root (h, fungal hyphae; vt, vascular trace); bar, 250 µm. (c) Cross-section of H. hypogeae root; bar, 250 µm. (d) Magnified view of intracellular hyphae in root; bar, 50 µm.
New Phytologist (2008) 177: 790–801 www.newphytologist.org © The Authors (2007). Journal compilation © New Phytologist (2007)
Research796
Fig. 4 Bayesian 18S tree based on 1252 characters and 25 unique isolated sequences from sporophytes and gametophytes of Huperzia and Lycopodium collected in Ecuador. Branches denoted * are supported by = 99% neighbor-joining and maximum-parsimony bootstrap values (values of 10 000 bootstraps) and Bayesian posterior probability values (values of 4 million generations). Other branches’ Bayesian posterior probabilities, neighboring-joining and parsimony bootstrap values are indicated above the branch. Triangles represent a phylotype of Glomus fungus, and sequences within a phylotype share 96.5–99.8% sequence identity. Includes Glomus A sequences isolated from Botrychium crenaulatum and B. lanceolatum (Winther & Friedman, 2007) and from mycoheterotrophic angiosperms Voyriella parviflora, Voyria corymbosa and Arachnitis uniflora (Bidartondo et al., 2002).

© The Authors (2007). Journal compilation © New Phytologist (2007) www.newphytologist.org New Phytologist (2008) 177: 790–801
Research 797
The phylogenetic relationships of the MH3 clade to otherGlomus sequences recovered from mycoheterotrophic vascularplants are ambiguous. All 18S Glomus sequences recoveredfrom mycoheterotrophic life cycle stages of Lycopodium andHuperzia (MH3), B. crenulatum (MH1), B. crenulatum (MH1),Arachnitis (MH1), Voyria (MH2), and Voyriella are in thewell supported Glomus group A (sensu Schussler et al., 2001).Although relationships among clades within Glomus A are notresolved, it is clear that the MH3 clade represents Glomusdiversity that is different from other Glomus sequences previouslyrecovered from other nonphotosynthetic vascular land plantsand isolated Glomus spores.
Discussion
This study of AM associations in Lycopodiaceae, an ancientlineage of vascular land plants, builds upon our previousresearch on the Glomus symbionts found throughout the lifecycle of two field-collected species of the fern Botrychium(Winther & Friedman, 2007). Botrychium, like Huperzia andLycopodium, is characterized by a life cycle with independentmycoheterotrophic gametophytes that are dependent on AMassociations for a source of fixed carbon. Although the presenceof fungal symbionts in the gametophytes and sporophytes ofLycopodiaceae had been documented (Bruchmann, 1885;Treub, 1890a; Holloway, 1910; Bruce, 1979a; Schmid &Oberwinkler, 1993), nothing was known of the phylogeneticaffinities of the fungal partners, within and between thedifferent life cycle stages of Huperzia and Lycopodium. OurDNA data and phylogenetic analyses provide an emergingpicture of the ecological and evolutionary relationships betweennonseed vascular plants and their fungal partners.
AM associations in life cycle of H. hypogeae
The AM fungi in H. hypogeae above-ground sporophytes andunderground gametophytes are aseptate and have coil-likearbuscules (Figs 2, 3) similar to Paris-type AM associationsthat have also been reported in the sporophytes andsubterranean gametophytes of many mycoheterotrophicland plants, including Lycopodiaceae (Treub, 1884, 1886,1888a,b, 1890a,b; Bruchmann, 1885, 1898; Bruce, 1979a,b;Whittier & Webster, 1986), Ophioglossales (Campbell, 1911;Schmid & Oberwinkler, 1994), Psilotales (Holloway, 1939;Bierhorst, 1953, 1954, 1971; Peterson et al., 1981; Duckett& Ligrone, 2005), and nonphotosynthetic angiosperms(Imhof & Weber, 1997; Imhof, 1999a,b, 2004; Dominguezet al., 2005). Our data provide further evidence that ancientlineages of nonseed vascular plants form Paris-type AMassociations (Smith & Smith, 1997).
Three 18S and ITS phylotypes were isolated from H. hypogeaesporophyte roots and gametophytes. Both H. hypogeaegametophytes had the same three 18S and ITS Glomusphylotypes (Fig. 4, Table 1) as their neighboring photosynthetic
H. hypogeae sporophyte. Our sequence data provide compelling,but circumstantial, evidence that underground H. hypogeaegametophytes obtain carbon via ‘epiparasitism’ from conspecificneighboring sporophytes through shared glomalean fungalsymbionts. However, it is important to note that we did notexamine the AM symbionts in neighboring heterospecificplants. In other plant taxa with AM mycoheterotrophicassociations such as Botrychium (Winther & Friedman, 2007),Arachnitis, Voyria and Voyriella (Bidartondo et al., 2002),neighboring heterospecific plants are likely to contributeorganic carbon through shared glomalean networks and it ispossible that this also occurs in H. hypogeae.
Distribution of AM symbionts in seven species of Lycopodium and Huperzia
Previous studies have reported nonglomalean mycorrhizalassociations in Lycopodiaceae sporophytes (Spessard, 1917;Freeberg, 1962; Bruce, 1979a,b; Treu et al., 1996). It ispossible Lycopodium, Huperzia, Lycopodiella, and Phylloglossumspecies form plant–fungal associations with a diversity of fungallineages. However, we found no evidence of nonglomaleanAM associations in the seven species of Huperzia andLycopodium with either microscopy or DNA data (Figs 2–4).
Four Glomus phylotypes that form AM associations wererecovered from the seven Huperzia and Lycopodium photosyn-thetic sporophytes sampled (Fig. 4, Table 1). The handful ofpublished accounts of AM associations (based on microscopy)in roots of Lycopodium, Huperzia, Phylloglossum, and Lycopodiellareport that AM associations are only sometimes found withinindividuals of a single species and that some species have neverbeen found to have AM associations (Boullard, 1957; Harley& Harley, 1987; Gemma et al., 1992; Treu et al., 1996; Zhoa,2000; Fuchs & Haselwandter, 2004). It is possible that someLycopodiaceae sporophytes are facultatively mycorrhizal, butour data do not support this. Each plant we surveyed had AMfungi, but not all roots from a single plant contained AMfungi. Previous studies may have suffered from insufficientsampling.
Two of the four Glomus phylotypes (C and D) were recoveredfrom most sites and most Huperzia and Lycopodium speciessampled (Table 1). This suggests that these phylotypes arerelatively common mycorrhizal associates of Lycopodiaceaespecies in the Páramos of Ecuador. These two phylotypes werealso recovered from the subterranean gametophytes of H.hypogeae and might form AM associations with the obligatemycorrhizal gametophytes of the other Lycopodium andHuperzia species sampled.
Two of the four Glomus phylotypes (A and B) were eachonly recovered from a single species of Huperzia (H. hypogeaeand H. tetragona, respectively). This could be because of ourlimited sample sizes of photosynthetic sporophytes andmycoheterotrophic gametophytes and/or to PCR bias (unequalamplification of certain sequences over others when the

New Phytologist (2008) 177: 790–801 www.newphytologist.org © The Authors (2007). Journal compilation © New Phytologist (2007)
Research798
template is a mixture homologous genes (Kanagawa, 2003)).If this result is not an artifact of sampling, then the AMsymbionts found in a given Huperzia or Lycopodium speciesmay, in certain cases, be restricted by the geographic distributionof Glomus symbionts and/or the taxonomic identity of theplant host.
Phylogenetic relationships of AM symbionts in Huperzia and Lycopodium
The phylogenetic relationships of the isolated 18S phylotypesprovide compelling evidence for the discovery of a majorclade of Glomus, the MH3 clade, capable of forming AMassociations with Huperzia and Lycopodium sporophytes andmycoheterotrophic gametophytes. This is consistent withprevious studies that have demonstrated that plants withmycoheterotrophic life cycle phases appear to specialize on‘narrow’ clades of fungi (Bidartondo et al., 2002; Winther &Friedman, 2007). By contrast, diverse clades of glomaleanfungi are routinely recovered from environmental samplingof AM symbionts from autotrophic plant species (recentexamples include Vandenkoornhuyse et al., 2002; Bergemann& Garbelotto, 2006; reviewed in Johnson et al., 2005; Opiket al., 2006). Given the broad fungal diversity that is consistentlyreported from environmental sampling of soils and plantsfrom sites all over the world (Anderson & Cairney, 2004;Mangan et al., 2004; Johnson et al., 2005; Borstler et al.,2006), it is likely that the MH3 clade represents a smallfraction of the total AM diversity available to the sampledspecies of Huperzia and Lycopodium.
It is not surprising that mycoheterotrophic plants are limitedin their AM partners, but it is striking that photosyntheticsporophytes of Huperzia and Lycopodium are restricted to fungalsymbionts in the same clade (MH3) as recovered from the twoH. hypogeae gametophytes. This is consistent with what wasreported in two field-collected species of Botrychium, wherea single clade of Glomus (MH1) was recovered from bothphotosynthetic sporophytes and mycoheterotrophic life cyclephases (Winther & Friedman, 2007). It may be that photo-synthetic sporophytes in plant lineages with an alternationof generations characterized by free-living photosyntheticsporophytes and mycoheterotrophic gametophytes are con-strained to form AM associations with small clades of GlomusA. This could be the result of coevolution of mycoheterotrophicplant lineages with a certain subset of Glomus that can engagein mycoheterotrophic associations and/or the fact that thesporophyte is potentially the ultimate source of fixed carbonfor the gametophytes via shared fungal hyphae.
As we begin to amass data on the phylogenetic identity ofAM symbionts from diverse vascular plant lineages, a pictureof the ecological and evolutionary biology of AM associationsis emerging. Most fungal symbionts recovered from plantswith mycoheterotrophic life cycle stages (including lycophytes,ferns and angiosperms) are members of the Glomus A clade
(Bidartondo et al., 2002; Franke et al., 2006; Winther &Friedman, 2007), one of three major lineages within theGlomaceae (Schussler et al., 2001). Moreover, analyses ofphylogenetic relationships among members of the Glomus Aclade suggest that the majority of mycoheterotrophic plantsstudied to date form AM associations with restricted cladesof Glomus A (Fig. 4). It is possible there have been manyindependent origins of clades of Glomus A that can formmycoheterotrophic AM associations. Alternatively, it may bethat with further sampling of AM fungal symbionts frommycoheterotrophic plant species, all members of Glomus Awill be found to be able to form mycoheterotrophic AMassociations with specific clades of plants. If true, Glomus A maybe viewed as a fungal clade with the ability to form complexlabile AM networks where the fungus may receive or transfercarbon to the plant host. It is yet unknown whether geographyand/or plant identity are the primary determinants of thespecific pattern of association that we have uncovered inlycopods and ferns.
The seven Huperzia and Lycopodium species we sampled areextant members of two Lycopodiaceae genera whose ancestorsdiverged approx. 350 million yr (Wikstrom, 2001; Wikstrom& Kenrick, 2001). Our data show that both genera form AMassociations with the same narrow Glomus A clade, MH3, thatforms Paris-type AM associations with photosynthetic and myco-heterotrophic life cycle phases of Huperzia and Lycopodium.In addition, the distribution of AM symbionts in H. hypogeaeprovides circumstantial evidence that photosyntheticsporophytes are a source of carbon to its mycoheterotrophicgametophytes via shared fungal networks. Fossil evidenceindicates that the ancestors of lycophytes had photosyntheticgametophytes and sporophytes with intracellular mycorrhizalassociations that resemble AM associations (Taylor et al.,2004; Helgason & Fitter, 2005; Krings et al., 2005; Taylor &Krings, 2005). Thus, we are looking at a potentially veryancient association between a subset of Glomus A fungi andHuperzia and Lycopodium sporophytes and gametophytes.
Concluding remarks
Molecular identities of the fungal symbionts in mycoheter-otrophic AM plant–fungal associations are now known fromland plant lineages that span approx. 390 million yr of landplant evolution (Wikstrom & Kenrick, 2001). Our phylogeneticanalyses suggest that most mycoheterotrophic lineages formAM associations with restricted clades of Glomus A. We wouldexpect to recover additional lineages of Glomus A from furthermolecular identification of Glomus symbionts of the approx.1000 fern and lycopod mycoheterotrophic plant species thatrepresent multiple independent evolutions of mycoheterotrophicAM symbioses (Wang & Qiu, 2006). Moreover, additionalsampling will establish if all fungal taxa in Glomus A arecapable of forming mycoheterotrophic associations and/or towhat extent there is plant-fungal specificity in mycoheterotrophic

© The Authors (2007). Journal compilation © New Phytologist (2007) www.newphytologist.org New Phytologist (2008) 177: 790–801
Research 799
AM associations. Without question, future molecularidentification of Glomus symbionts in mycoheterotrophicplants from a broader ecological and phylogenetic breadthshould yield a fascinating picture of the evolutionary historyand the complex biological relationships of mycoheterotrophsand their fungi symbionts.
Acknowledgements
The authors thank Devin O’Conner for collecting Lycopodiumand Huperzia species. We thank Benjamin Ollgaard for helpin the identification of the different Lycopodium and Huperziaspecies. We thank Kirsten Ryerson for help with sectioning.This work was supported by a NASA Astrobiology grant toWEF.
References
Anderson IC, Cairney JWG. 2004. Diversity and ecology of soil fungal communities: increased understanding through the application of molecular techniques. Environmental Microbiology 6: 769–779.
Antoniolli ZI, Schachtman DP, Ophel-Keller K, Smith SE. 2000. Variation in rDNA ITS sequences in Glomus mosseae and Gigaspora margarita spores from a permanent pasture. Mycological Research 104: 708–715.
Bergemann SE, Garbelotto M. 2006. High diversity of fungi recovered from the roots of mature tanoak (Lithocarpus densiflorus) in northern California. Canadian Journal of Botany 84: 1380–1394.
Bidartondo MI, Redecker D, Hijri I, Wiemken A, Bruns TD, Dominguez T, Sersic A, Leake JR, Read DJ. 2002. Epiparasitic plants specialized on arbuscular mycorrhizal fungi. Nature 419: 389–392.
Bierhorst DW. 1953. Structure and development of the gametophyte of Psilotum-nudum. American Journal of Botany 40: 649–658.
Bierhorst DW. 1954. The gametangia and embryo of Psilotum-nudum. American Journal of Botany 41: 274–281.
Bierhorst DW. 1971. Morphology of vascular plants. New York, NY, USA: The Macmillan Co.
Borstler B, Renker C, Kahmen A, Buscot F. 2006. Species composition of arbuscular mycorrhizal fungi in two mountain meadows with differing management types and levels of plant biodiversity. Biology of Fertility and Soils 42: 286–298.
Boullard B. 1957. Bulletin for the Society of Mycology Fr 23: 225–244.Bruce JG. 1979a. Gametophyte of Lycopodium-Digitatum. American Journal
of Botany 66: 1138–1150.Bruce JG. 1979b. Gametophyte and young sporophyte of
Lycopodium-Carolinianum. American Journal of Botany 66: 1156–1163.Bruce JG, Bietel JM. 1979. A community of Lycopodium gametophytes in
Michigan. American Fern Journal 69: 33–41.Bruchmann H. 1885. Das prothallium von Lycopodium. Botanisches
Zentralblatt 21: 23–28.Bruchmann H. 1898. Uber die Prothallien und die Keimpflanzen
mehrerer europäischer Lycopodien, und zwar über die von Lycopodium clavatum, L. annotinum, L. complanatum und L. Selago. Gotha, Germany: Perthes.
Brundrett MC. 2002. Coevolution of roots and mycorrhizas of land plants. New Phytologist 154: 275–304.
Brundrett M. 2004. Diversity and classification of mycorrhizal associations. Biological Reviews 79: 473–495.
Campbell DH. 1911. The eusporangiate: the comparative morphology of the Ophioglossaceae and the Marattiaceae. Washington, DC, USA: Carnegie Institution of Washington.
Chamberlain CJ. 1917. Prothallia and sporelings of three New Zealand species of Lycopodium. Botanical Gazette 63: 51–65.
Clapp JP, Young JPW, Merryweather JW, Fitter AH. 1995. Diversity of fungal symbionts in arbuscular mycorrhizas from a natural community. New Phytologist 130: 259–265.
DeSalle R, Giribet G, Wheeler W. 2002. Techniques in molecular systematics and evolution. Basel, Germany: Birkhauser.
Dominguez L, Sersic A, Melville L, Peterson RL. 2005. ‘Prepackaged symbioses’: propagules on roots of the myco-heterotrophic plant Arachnitis uniflora. New Phytologist 169: 191–198.
Duckett JG, Ligrone R. 2005. A comparative cytological analysis of fungal endophytes in the sporophyte rhizomes and vascularized gametophytes of Tmesipteris and Psilotum. Canadian Journal of Botany 83: 1443–1456.
Franke T, Beenken L, Doring M, Kocyan A, Agerer R. 2006. Arbuscular mycorrhizal fungi of the Glomus-group A lineage (Glomerales; Glomeromycota) detected in myco-heterotrophic plants from tropical Africa. Mycological Progress 5: 24–31.
Freeberg JA. 1962. Lycopodium prothalli and their endophytic fungi as studies in vitro. American Journal of Botany 49: 530–535.
Fuchs B, Haselwandter K. 2004. Red list plants; colonization by arbuscular mycorrhizal fungi and dark septate endophytes. Mycorrhiza 14: 277–281.
Gemma JN, Koske RE, Flynn T. 1992. Mycorrhizae in Hawaiian Pteridophytes: occurrence and evolutionary significance. American Journal of Botany 79: 843–852.
Gollotte A, van Tuinen D, Atkinson D. 2004. Diversity of arbuscular mycorrhizal fungi colonising roots of the grass species Agrostis capillaris and Lolium perenne in a field experiment. Mycorrhiza 14: 111–117.
Harley JL, Harley EL. 1987. A check-list of mycorrhiza in British flora. New Phytologist 105: 1–102.
Helgason T, Fitter AH. 2005. The ecology and evolution of the arbuscular mycorrhizal fungi. Mycologist 19: 96–101.
Hijri I, Sykorova Z, Oehl F, Ineichen K, Mader P, Wiemken A, Redecker D. 2006. Communities of arbuscular mycorrhizal fungi in arable soils are not necessarily low in diversity. Molecular Ecology 15: 2277–2289.
Holloway JE. 1910. A comparative study of the anatomy of six New Zealand species of Lycopodium. Transactions and Proceedings of the New Zealand Institute 42: 356–370.
Holloway JE. 1916. Studies in the New Zealand species of the genus Lycopodium. Part 1. Transactions and Proceedings of the New Zealand Institute 48: 253–303.
Holloway JE. 1935. The gametophyte of Phylloglossum drummondii. Annals of Botany 49: 513–519.
Holloway JE. 1939. The gametophyte, embryo, and young rhizome of Psilotum Triquetrum Swartz. Annals of Botany 53: 313–319.
Huelsenbeck JP, Ronquist F. 2001. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 17: 754–755.
Imhof S. 1999a. Subterranean structures and mycorrhiza of the achlorophyllous Burmannia tenella (Burmanniaceae). Canadian Journal of Botany 77: 637–643.
Imhof S. 1999b. Root morphology, anatomy and mycotrophy of the achlorophyllous Voyria aphylla ( Jacq.) Pers. (Gentianaceae). Mycorrhiza 9: 33–39.
Imhof S. 2004. Morphology and development of the subterranean organs of the achlorophyllous Sciaphila polygyna (Triuridaceae). Botany Journal of the Linnean Society 146: 295–301.
Imhof S, Weber HC. 1997. Root anatomy and mycotrophy (AM) of the achlorophyllous Voyria truncata (Standley) Standley & Steyermark (Gentianaceae). Botanica Acta 110: 127–134.
Jansa J, Mozafar A, Banke S, Mcdonald BA, Frossard E. 2002. Intra- and intersporal diversity of ITS rDNA sequences in Glomus intraradices assessed by cloning and sequencing, and by SSCP analysis. Mycological Research 106: 670–681.

New Phytologist (2008) 177: 790–801 www.newphytologist.org © The Authors (2007). Journal compilation © New Phytologist (2007)
Research800
Johnson D, Ijdo M, Genney DR, Anderson IC, Alexander IJ. 2005. How do plants regulate the function, community structure, and diversity of mycorrhizal fungi? Journal of Experimental Botany 56: 1751–1760.
Kanagawa et al. 2003. Bias and artifact in multitemplate polymerase chain reactions (PCR). Journal of Bioscience and Bioengineering 96: 317–323.
Kenrick P, Crane PR. 1997. The origin and early evolution of plants on land. Nature 389: 33–39.
Kjoller R, Rosendahl S. 2001. Molecular diversity of glomalean (arbuscular mycorrhizal) fungi determined as distinct Glomus specific DNA sequences from roots of field grown peas. Mycological Research 105: 1027–1032.
Krings M, Grewing A, Taylor TN, Kerp H, Galtier J. 2005. Lageniastrum macrosporae (fossil Volvocales, Lageniastraceae nov fam.), an endophyte in megaspores from the Carboniferous of the French Massif Central. Geobios 38: 451–465.
Leake JR. 1993. The biology of myco-heterotrophic (‘saprophytic’) plants. New Phytologist 127: 171–216.
Leake JR. 2004. Myco-heterotroph/epiparasitic plant interactions with ectomycorrhizal and arbuscular mycorrhizal fungi. Current Opinion in Biology 7: 422–428.
Luteyn. 1999. Páramos: checklist of plant diversity, geographical distribution and botanical literature. New York, NY, USA: Botanical Garden Press.
Mangan SA, Eom AH, Adler GH, Yavitt JB, Herre EA. 2004. Diversity of arbuscular mycorrhizal fungi across a fragmented forest in Panama: insular spore communities differ from mainland communities. Oecologia 141: 687–700.
Ogden TH, Rosenberg MS. 2006. Multiple sequence alignment accuracy and phylogenetic inference. Systematic Biology 55: 314–328.
Ollgaard B. 1992. Neotropical Lycopodiaceae – an overview. Annals of the Missouri Botanical Garden 79: 687–717.
Opik M, Moora M, Liira J, Koljalg U, Zobel M, Sen R. 2003. Divergent arbuscular mycorrhizal fungal communities colonize roots of Pulsatilla spp. in boreal Scots pine forest and grassland soils. New Phytologist 160: 581–593.
Opik M, Moora M, Liira J, Zobel M. 2006. Composition of root-colonizing arbuscular mycorrhizal fungal communities in different ecosystems around the globe. Journal of Ecology 94: 778–790.
Peterson RL, Howarth MJ, Whittier DP. 1981. Interactions between a fungal endophyte and gametophyte cells in Psilotum–Nudum. Canadian Journal of Botany-Revue Canadienne de Botanique 59: 711–720.
Pirozynski KA, Malloch DW. 1975. The origin of land plants: a matter of mycotrophism. Biosystems 6: 153–164.
Posada D, Crandall KA. 1998. Modeltest: testing the model of DNA substitution. Bioinformatics 9: 817–818.
Redecker D, Kodner R, Graham LE. 2000. Glomalean fungi from the Ordovician. Science 289: 1920–1921.
Redecker D, Thierfelder H, Walker C, Werner D. 1997. Restriction analysis of PCR-amplified internal transcribed spacers of ribosomal DNA as a tool for species identification in different genera of the order glomales. Applied and Environmental Microbiology 63: 1756–1761.
Remy W, Thomas TN, Hass H, Kerp H. 1994. Four hundred-million-year-old vesicular arbuscular mycorrhizae. Proceedings of the National Academy of Sciences, USA 91: 11841–11843.
Rosendahl D, Stukenbrock EH. 2004. Community structure of arbuscular mycorrhizal fungi in undisturbed vegetation revealed by analyses of LSU rDNA sequences. Molecular Ecology 13: 3179–3186.
Schmid E, Oberwinkler F. 1993. Mycorrhiza-like interaction between the achlorophyllous gametophyte of Lycopodium-Clavatum L. and its fungal endophyte studied by light and electron-microscopy. New Phytologist 124: 69–81.
Schmid E, Oberwinkler F. 1994. Light and electron microscopy of the host–fungus interaction in the achlorophyllous gaetophyte of Botrychium lunaria. Canadian Journal of Botany 72: 182–188.
Schussler A, Schwarzott D, Walker C. 2001. A new fungal phylum, the Glomeromycota: phylogeny and evolution. Mycological Research 105: 1413–1421.
Schwarzott D, Schussler A. 2001. A simple and reliable method for SSU rRNA gene DNA extraction, amplification, and cloning from single AM fungal spores. Mycorrhiza 10: 203–207.
Schwarzott D, Walker C, Schussler A. 2001. Glomus, the largest genus of the arbuscular mycorrhizal fungi (Glomales), is nonmonophyletic. Molecular Phylogenetics and Evolution 21: 190–197.
Simon L. 1995. Phylogeny of the glomales: deciphering the past to understand the present. New Phytologist 133: 95–101.
Simon L, Bousquet J, Levesque RC, Lalonde M. 1993a. Origin and diversification of endomycorrhizal fungi and coincidence with vascular land plants. Nature 363: 67–69.
Simon L, Levesque RC, Lalonde M. 1993b. Identification of endomycorrhizal fungi colonizing roots by fluorescent single-strand conformation polymorphism polymerase chain-reaction. Applied and Environmental Microbiology 59: 4211–4215.
Smith SE, Read DJ. 1997. Mycorrhizal symbiosis. San Diego, CA, USA: Academic Press.
Smith FA, Smith SE. 1997. Structural diversity in (vesicular)-arbuscular mycorrhizal symbioses. New Phytologist 137: 373–388.
Smith. 1938. Cryptogamic botany, Vol. II. Bryophytes and Pteridophytes. New York, NY, USA: McGraw-Hill.
Spessard EA. 1917. Prothallia of Lycopodium in America. Botanical Gazette 63: 66–76.
Swofford DL. 2002. PAUP*: Phylogenetic Analysis Using Parsimony. Sunderland, MA, USA: Sinauer.
Taylor TN, Hass H, Krings M, Klavins SD, Kerp H. 2004. Fungi in the Rhynie chert: a view from the dark side. Transactions of the Royal Society of Edinburgh, Earth Science 94: 455–471.
Taylor TN, Krings M. 2005. Fossil microorganisms and land plants: Associations and interactions. Symbiosis 40: 119–135.
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. 1997. The ClustalX windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Research 24: 4876–4882.
Treu R, Laursen GA, Stephenson SL, Landolt JC, Densmore R. 1996. Mycorrhizae from Denali National Park and Perserve, Alaska. Mycorrhiza 6: 21–29.
Treub M. 1884. Etude sur les lycopodiacees. I. Le prothalle du Lycopodium cernuum. Annales du Jardin Botanique de Buitenzorg 4: 107–135.
Treub M. 1886. Etude sur les lycopodiacees II. Le prothalle du Lycopodium phlegmaria. Annales du Jardin Botanique de Buitenzorg 5: 87–114.
Treub M. 1888a. Etudes sur les lycopodiacees V. Les prothalle des Lycopodium carinatum. nummularifolium et. Hippuris. Annales du Jardin Botanique de Buitenzorg 7: 146–150.
Treub M. 1888b. Etudes sur les lycopodiacees IV Le prothalle du Lycopodium salakense. Annales du Jardin Botanique de Buitenzorg 7: 141–148.
Treub M. 1890a. Etudes sur les Lycopodiacees. VIII. Considerations theorique. Annales du Jardin Botanique de Buitenzorg 8: 23–27.
Treub M. 1890b. Etudes sur les lycopodiacees VI. L’embron et la plantule du Lycopodium cernuum. Etudes sur les lycopodiacea VIII. les tubercules radicaux du Lycopodium cernuum. Annales du Jardin Botanique de Buitenzorg 8: 1–36.
Vandenkoornhuyse P, Husband R, Daniell TJ, Watson IJ, Duck JM, Fitter AH, Young JPW. 2002. Arbuscular mycorrhizal community composition associated with two plant species in a grassland ecosystem. Molecular Ecology 11: 1555–1564.
Wang B, Qiu YL. 2006. Pyhlogenetic distribution and evolution of mycorrhizas in land plants. Mycorrhiza 16: 299–363.
White TJ, Bruns T, Lee S, Taylor JW. 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics.

© The Authors (2007). Journal compilation © New Phytologist (2007) www.newphytologist.org New Phytologist (2008) 177: 790–801
Research 801
In: Innis MA, Gelfand DH, Sninsky JJ, White. TJ, eds. PCR protocols: a guide to methods and applications. New York, NY, USA: Academic Press, 315–322.
Whittier DP, Braggins JE. 1992. The young gametophyte of Phylloglossum (Lycopodiaceae). Annals of Missouri Botanic Garden 79: 730–736.
Whittier DP. 1998. Germination of spores of the Lycopodiaceae in axenic culture. American Fern Journal 88: 106–113.
Whittier DP, Pintaud JC, Braggins JE. 2005. The gametophyte of Lycopodium deuterodensum – type II or I. American Fern Journal 95: 22–29.
Whittier DP, Webster TR. 1986. Gametophytes of Lycopodium-Lucidulum from axenic culture. American Fern Journal 76: 48–55.
Wikstrom N. 2001. Diversification and relationships of extant homosporous lycopods. American Fern Journal 91: 150–165.
Wikstrom N, Kenrick P. 2001. Evolution of Lycopodiaceae (Lycopsida): estimating divergence times from rbcL gene sequences by use of nonparametric rate smoothing. Molecular Phylogenetics and Evolution 19: 177–186.
Winther JL, Friedman WF. 2007. Arbuscular mycorrhizal symbionts in Botrychium (Ophioglossaceae). American Journal of Botany 94: 1248–1255.
Yang Z, Rannala B. 2005. Branch-length prior influences Bayesian posterior probability of phylogeny. Systematic Biology 54: 455–470.
Zhao Z. 2000. The arbuscular mycorrhizas of pteridophytes in Yunnan, southwest China: evolutionary interpretations. Mycorrhiza 10: 145–149.
Supplementary Material
The following supplementary material is available for thisarticle online:
Appendix S1 Alignment of 18S Glomus sequences isolatedfrom Huperzia and Lycopodium including previously publishedsequences.
Appendix S2 Accession numbers for previously published18S and accession numbers for Glomus sequences previouslyisolated from mycoheterotrophic plants used in phylogeneticanalyses.
This material is available as part of the online article from:http://www.blackwell-synergy.com/doi/abs/10.1111/j.1469-8137.2007.02276.x(This link will take you to the article abstract).
Please note: Blackwell Publishing are not responsible for thecontent or functionality of any supplementary materials suppliedby the authors. Any queries (other than missing material) shouldbe directed to the journal at New Phytologist Central Office.
About New Phytologist
• New Phytologist is owned by a non-profit-making charitable trust dedicated to the promotion of plant science, facilitating projectsfrom symposia to open access for our Tansley reviews. Complete information is available at www.newphytologist.org.
• Regular papers, Letters, Research reviews, Rapid reports and both Modelling/Theory and Methods papers are encouraged.We are committed to rapid processing, from online submission through to publication ‘as-ready’ via OnlineEarly – our averagesubmission to decision time is just 28 days. Online-only colour is free, and essential print colour costs will be met if necessary. Wealso provide 25 offprints as well as a PDF for each article.
• For online summaries and ToC alerts, go to the website and click on ‘Journal online’. You can take out a personal subscription tothe journal for a fraction of the institutional price. Rates start at £135 in Europe/$251 in the USA & Canada for the online edition(click on ‘Subscribe’ at the website).
• If you have any questions, do get in touch with Central Office ([email protected]; tel +44 1524 594691) or, for a localcontact in North America, the US Office ([email protected]; tel +1 865 576 5261).