asms 2009 poster
TRANSCRIPT
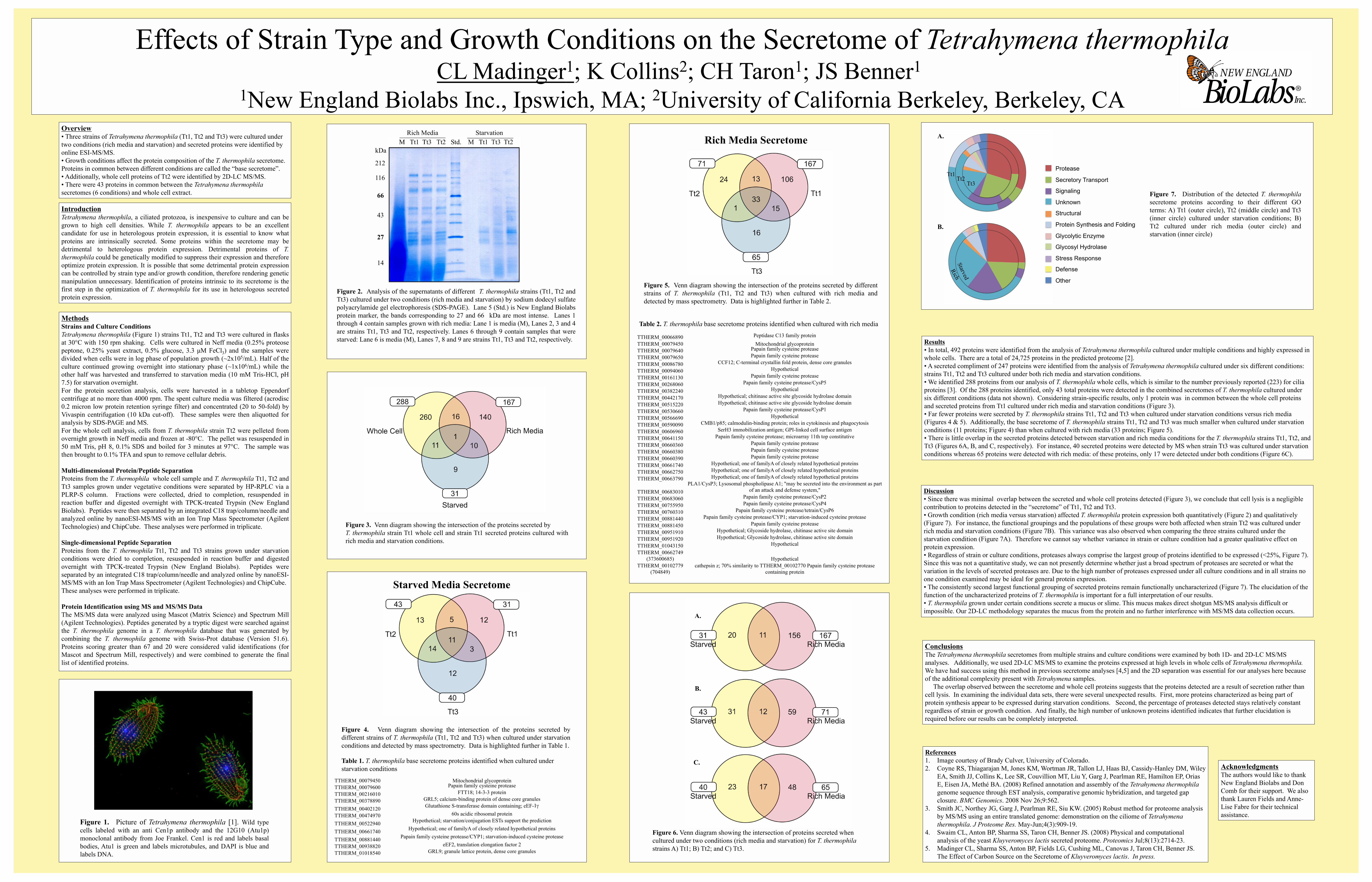
Figure 4. Venn diagram showing the intersection of the proteins secreted by
different strains of T. thermophila (Tt1, Tt2 and Tt3) when cultured under starvation
conditions and detected by mass spectrometry. Data is highlighted further in Table 1.
Effects of Strain Type and Growth Conditions on the Secretome of Tetrahymena thermophila CL Madinger1; K Collins2; CH Taron1; JS Benner1
1New England Biolabs Inc., Ipswich, MA; 2University of California Berkeley, Berkeley, CA
Introduction Tetrahymena thermophila, a ciliated protozoa, is inexpensive to culture and can be
grown to high cell densities. While T. thermophila appears to be an excellent
candidate for use in heterologous protein expression, it is essential to know what
proteins are intrinsically secreted. Some proteins within the secretome may be
detrimental to heterologous protein expression. Detrimental proteins of T.
thermophila could be genetically modified to suppress their expression and therefore
optimize protein expression. It is possible that some detrimental protein expression
can be controlled by strain type and/or growth condition, therefore rendering genetic
manipulation unnecessary. Identification of proteins intrinsic to its secretome is the
first step in the optimization of T. thermophila for its use in heterologous secreted
protein expression.
Methods Strains and Culture Conditions
Tetrahymena thermophila (Figure 1) strains Tt1, Tt2 and Tt3 were cultured in flasks
at 30°C with 150 rpm shaking. Cells were cultured in Neff media (0.25% proteose
peptone, 0.25% yeast extract, 0.5% glucose, 3.3 μM FeCl3) and the samples were
divided when cells were in log phase of population growth (~2x105/mL). Half of the
culture continued growing overnight into stationary phase (~1x106/mL) while the
other half was harvested and transferred to starvation media (10 mM Tris-HCl, pH
7.5) for starvation overnight.
For the protein secretion analysis, cells were harvested in a tabletop Eppendorf
centrifuge at no more than 4000 rpm. The spent culture media was filtered (acrodisc
0.2 micron low protein retention syringe filter) and concentrated (20 to 50-fold) by
Vivaspin centrifugation (10 kDa cut-off). These samples were then aliquotted for
analysis by SDS-PAGE and MS.
For the whole cell analysis, cells from T. thermophila strain Tt2 were pelleted from
overnight growth in Neff media and frozen at -80°C. The pellet was resuspended in
50 mM Tris, pH 8, 0.1% SDS and boiled for 3 minutes at 97°C. The sample was
then brought to 0.1% TFA and spun to remove cellular debris.
Multi-dimensional Protein/Peptide Separation
Proteins from the T. thermophila whole cell sample and T. thermophila Tt1, Tt2 and
Tt3 samples grown under vegetative conditions were separated by HP-RPLC via a
PLRP-S column. Fractions were collected, dried to completion, resuspended in
reaction buffer and digested overnight with TPCK-treated Trypsin (New England
Biolabs). Peptides were then separated by an integrated C18 trap/column/needle and
analyzed online by nanoESI-MS/MS with an Ion Trap Mass Spectrometer (Agilent
Technologies) and ChipCube. These analyses were performed in triplicate.
Single-dimensional Peptide Separation
Proteins from the T. thermophila Tt1, Tt2 and Tt3 strains grown under starvation
conditions were dried to completion, resuspended in reaction buffer and digested
overnight with TPCK-treated Trypsin (New England Biolabs). Peptides were
separated by an integrated C18 trap/column/needle and analyzed online by nanoESI-
MS/MS with an Ion Trap Mass Spectrometer (Agilent Technologies) and ChipCube.
These analyses were performed in triplicate.
Protein Identification using MS and MS/MS Data
The MS/MS data were analyzed using Mascot (Matrix Science) and Spectrum Mill
(Agilent Technologies). Peptides generated by a tryptic digest were searched against
the T. thermophila genome in a T. thermophila database that was generated by
combining the T. thermophila genome with Swiss-Prot database (Version 51.6).
Proteins scoring greater than 67 and 20 were considered valid identifications (for
Mascot and Spectrum Mill, respectively) and were combined to generate the final
list of identified proteins.
Overview • Three strains of Tetrahymena thermophila (Tt1, Tt2 and Tt3) were cultured under
two conditions (rich media and starvation) and secreted proteins were identified by
online ESI-MS/MS.
• Growth conditions affect the protein composition of the T. thermophila secretome.
Proteins in common between different conditions are called the “base secretome”.
• Additionally, whole cell proteins of Tt2 were identified by 2D-LC MS/MS.
• There were 43 proteins in common between the Tetrahymena thermophila
secretomes (6 conditions) and whole cell extract.
Results
• In total, 492 proteins were identified from the analysis of Tetrahymena thermophila cultured under multiple conditions and highly expressed in
whole cells. There are a total of 24,725 proteins in the predicted proteome [2].
• A secreted compliment of 247 proteins were identified from the analysis of Tetrahymena thermophila cultured under six different conditions:
strains Tt1, Tt2 and Tt3 cultured under both rich media and starvation conditions.
• We identified 288 proteins from our analysis of T. thermophila whole cells, which is similar to the number previously reported (223) for cilia
proteins [3]. Of the 288 proteins identified, only 43 total proteins were detected in the combined secretomes of T. thermophila cultured under
six different conditions (data not shown). Considering strain-specific results, only 1 protein was in common between the whole cell proteins
and secreted proteins from Tt1 cultured under rich media and starvation conditions (Figure 3).
• Far fewer proteins were secreted by T. thermophila strains Tt1, Tt2 and Tt3 when cultured under starvation conditions versus rich media
(Figures 4 & 5). Additionally, the base secretome of T. thermophila strains Tt1, Tt2 and Tt3 was much smaller when cultured under starvation
conditions (11 proteins; Figure 4) than when cultured with rich media (33 proteins; Figure 5).
• There is little overlap in the secreted proteins detected between starvation and rich media conditions for the T. thermophila strains Tt1, Tt2, and
Tt3 (Figures 6A, B, and C, respectively). For instance, 40 secreted proteins were detected by MS when strain Tt3 was cultured under starvation
conditions whereas 65 proteins were detected with rich media: of these proteins, only 17 were detected under both conditions (Figure 6C).
Conclusions The Tetrahymena thermophila secretomes from multiple strains and culture conditions were examined by both 1D- and 2D-LC MS/MS
analyses. Additionally, we used 2D-LC MS/MS to examine the proteins expressed at high levels in whole cells of Tetrahymena thermophila.
We have had success using this method in previous secretome analyses [4,5] and the 2D separation was essential for our analyses here because
of the additional complexity present with Tetrahymena samples.
The overlap observed between the secretome and whole cell proteins suggests that the proteins detected are a result of secretion rather than
cell lysis. In examining the individual data sets, there were several unexpected results. First, more proteins characterized as being part of
protein synthesis appear to be expressed during starvation conditions. Second, the percentage of proteases detected stays relatively constant
regardless of strain or growth condition. And finally, the high number of unknown proteins identified indicates that further elucidation is
required before our results can be completely interpreted.
References
1. Image courtesy of Brady Culver, University of Colorado.
2. Coyne RS, Thiagarajan M, Jones KM, Wortman JR, Tallon LJ, Haas BJ, Cassidy-Hanley DM, Wiley
EA, Smith JJ, Collins K, Lee SR, Couvillion MT, Liu Y, Garg J, Pearlman RE, Hamilton EP, Orias
E, Eisen JA, Methé BA. (2008) Refined annotation and assembly of the Tetrahymena thermophila
genome sequence through EST analysis, comparative genomic hybridization, and targeted gap
closure. BMC Genomics. 2008 Nov 26;9:562.
3. Smith JC, Northey JG, Garg J, Pearlman RE, Siu KW. (2005) Robust method for proteome analysis
by MS/MS using an entire translated genome: demonstration on the ciliome of Tetrahymena
thermophila. J Proteome Res. May-Jun;4(3):909-19.
4. Swaim CL, Anton BP, Sharma SS, Taron CH, Benner JS. (2008) Physical and computational
analysis of the yeast Kluyveromyces lactis secreted proteome. Proteomics Jul;8(13):2714-23.
5. Madinger CL, Sharma SS, Anton BP, Fields LG, Cushing ML, Canovas J, Taron CH, Benner JS.
The Effect of Carbon Source on the Secretome of Kluyveromyces lactis. In press.
Figure 2. Analysis of the supernatants of different T. thermophila strains (Tt1, Tt2 and
Tt3) cultured under two conditions (rich media and starvation) by sodium dodecyl sulfate
polyacrylamide gel electrophoresis (SDS-PAGE). Lane 5 (Std.) is New England Biolabs
protein marker, the bands corresponding to 27 and 66 kDa are most intense. Lanes 1
through 4 contain samples grown with rich media: Lane 1 is media (M), Lanes 2, 3 and 4
are strains Tt1, Tt3 and Tt2, respectively. Lanes 6 through 9 contain samples that were
starved: Lane 6 is media (M), Lanes 7, 8 and 9 are strains Tt1, Tt3 and Tt2, respectively.
Figure 5. Venn diagram showing the intersection of the proteins secreted by different
strains of T. thermophila (Tt1, Tt2 and Tt3) when cultured with rich media and
detected by mass spectrometry. Data is highlighted further in Table 2.
Table 1. T. thermophila base secretome proteins identified when cultured under
starvation conditions
Figure 1. Picture of Tetrahymena thermophila [1]. Wild type
cells labeled with an anti Cen1p antibody and the 12G10 (Atu1p)
monoclonal antibody from Joe Frankel. Cen1 is red and labels basal
bodies, Atu1 is green and labels microtubules, and DAPI is blue and
labels DNA.
TTHERM_00079450 Mitochondrial glycoprotein
TTHERM_00079600 Papain family cysteine protease
TTHERM_00216010 FTT18; 14-3-3 protein
TTHERM_00378890 GRL5; calcium-binding protein of dense core granules
TTHERM_00402120 Glutathione S-transferase domain containing; eEF-1
TTHERM_00474970 60s acidic ribosomal protein
TTHERM_00522940 Hypothetical; starvation/conjugation ESTs support the prediction
TTHERM_00661740 Hypothetical; one of familyA of closely related hypothetical proteins
TTHERM_00881440 Papain family cysteine protease/CYP1; starvation-induced cysteine protease
TTHERM_00938820 eEF2, translation elongation factor 2
TTHERM_01018540 GRL9; granule lattice protein, dense core granules
TTHERM_00066890 Peptidase C13 family protein
TTHERM_00079450 Mitochondrial glycoprotein
TTHERM_00079640 Papain family cysteine protease
TTHERM_00079650 Papain family cysteine protease
TTHERM_00086780 CCF12; C-terminal crystallin fold protein, dense core granules
TTHERM_00094060 Hypothetical
TTHERM_00161130 Papain family cysteine protease
TTHERM_00268060 Papain family cysteine protease/CysP5
TTHERM_00382240 Hypothetical
TTHERM_00442170 Hypothetical; chitinase active site glycoside hydrolase domain
TTHERM_00515220 Hypothetical; chitinase active site glycoside hydrolase domain
TTHERM_00530660 Papain family cysteine protease/CysP1
TTHERM_00566690 Hypothetical
TTHERM_00590090 CMB1/p85; calmodulin-binding protein; roles in cytokinesis and phagocytosis
TTHERM_00606960 SerH3 immobilization antigen; GPI-linked cell surface antigen
TTHERM_00641150 Papain family cysteine protease; microarray 11th top constitutive
TTHERM_00660360 Papain family cysteine protease
TTHERM_00660380 Papain family cysteine protease
TTHERM_00660390 Papain family cysteine protease
TTHERM_00661740 Hypothetical; one of familyA of closely related hypothetical proteins
TTHERM_00662750 Hypothetical; one of familyA of closely related hypothetical proteins
TTHERM_00663790 Hypothetical; one of familyA of closely related hypothetical proteins
TTHERM_00683010
PLA1/CysP3; Lysosomal phospholipase A1; "may be secreted into the environment as part
of an attack and defense system,"
TTHERM_00683060 Papain family cysteine protease/CysP2
TTHERM_00755950 Papain family cysteine protease/CysP4
TTHERM_00760310 Papain family cysteine protease/tetrain/CysP6
TTHERM_00881440 Papain family cysteine protease/CYP1; starvation-induced cysteine protease
TTHERM_00881450 Papain family cysteine protease
TTHERM_00951910 Hypothetical; Glycoside hydrolase, chitinase active site domain
TTHERM_00951920 Hypothetical; Glycoside hydrolase, chitinase active site domain
TTHERM_01043150 Hypothetical
TTHERM_00662749
(373600685) Hypothetical
TTHERM_00102779
(704849)
cathepsin z; 70% similarity to TTHERM_00102770 Papain family cysteine protease
containing protein
Table 2. T. thermophila base secretome proteins identified when cultured with rich media
Discussion
• Since there was minimal overlap between the secreted and whole cell proteins detected (Figure 3), we conclude that cell lysis is a negligible
contribution to proteins detected in the “secretome” of Tt1, Tt2 and Tt3.
• Growth condition (rich media versus starvation) affected T. thermophila protein expression both quantitatively (Figure 2) and qualitatively
(Figure 7). For instance, the functional groupings and the populations of these groups were both affected when strain Tt2 was cultured under
rich media and starvation conditions (Figure 7B). This variance was also observed when comparing the three strains cultured under the
starvation condition (Figure 7A). Therefore we cannot say whether variance in strain or culture condition had a greater qualitative effect on
protein expression.
• Regardless of strain or culture conditions, proteases always comprise the largest group of proteins identified to be expressed (<25%, Figure 7).
Since this was not a quantitative study, we can not presently determine whether just a broad spectrum of proteases are secreted or what the
variation in the levels of secreted proteases are. Due to the high number of proteases expressed under all culture conditions and in all strains no
one condition examined may be ideal for general protein expression.
• The consistently second largest functional grouping of secreted proteins remain functionally uncharacterized (Figure 7). The elucidation of the
function of the uncharacterized proteins of T. thermophila is important for a full interpretation of our results.
• T. thermophila grown under certain conditions secrete a mucus or slime. This mucus makes direct shotgun MS/MS analysis difficult or
impossible. Our 2D-LC methodology separates the mucus from the protein and no further interference with MS/MS data collection occurs.
Figure 3. Venn diagram showing the intersection of the proteins secreted by
T. thermophila strain Tt1 whole cell and strain Tt1 secreted proteins cultured with
rich media and starvation conditions.
Acknowledgments The authors would like to thank
New England Biolabs and Don
Comb for their support. We also
thank Lauren Fields and Anne-
Lise Fabre for their technical
assistance.
Figure 6. Venn diagram showing the intersection of proteins secreted when
cultured under two conditions (rich media and starvation) for T. thermophila
strains A) Tt1; B) Tt2; and C) Tt3.
A.
B.
C.
M Tt1 Tt3 Tt2 Std. M Tt1 Tt3 Tt2
kDa
212
116
66
43
27
14
Rich Media Starvation
Protease
Secretory Transport
Signaling
Unknown
Structural
Protein Synthesis and Folding
Glycolytic Enzyme
Glycosyl Hydrolase
Stress Response
Defense
Other
Figure 7. Distribution of the detected T. thermophila
secretome proteins according to their different GO
terms: A) Tt1 (outer circle), Tt2 (middle circle) and Tt3
(inner circle) cultured under starvation conditions; B)
Tt2 cultured under rich media (outer circle) and
starvation (inner circle) B.
A.
Tt1 Tt2
Tt3
Rich Media Secretome
Starved Media Secretome