bacterial associations with the hydromedusa nemopsis bachei and scyphomedusa aurelia aurita from the...
TRANSCRIPT

Full Terms & Conditions of access and use can be found athttp://www.tandfonline.com/action/journalInformation?journalCode=smar20
Download by: [North Shore Med Ctr] Date: 22 November 2016, At: 08:50
Marine Biology Research
ISSN: 1745-1000 (Print) 1745-1019 (Online) Journal homepage: http://www.tandfonline.com/loi/smar20
Bacterial associations with the hydromedusaNemopsis bachei and scyphomedusa Aureliaaurita from the North Atlantic Ocean
Meaghan C. Daley, Juanita Urban-Rich & Pia H. Moisander
To cite this article: Meaghan C. Daley, Juanita Urban-Rich & Pia H. Moisander (2016): Bacterialassociations with the hydromedusa Nemopsis bachei and scyphomedusa Aurelia aurita fromthe North Atlantic Ocean, Marine Biology Research, DOI: 10.1080/17451000.2016.1228974
To link to this article: http://dx.doi.org/10.1080/17451000.2016.1228974
View supplementary material
Published online: 07 Nov 2016.
Submit your article to this journal
Article views: 17
View related articles
View Crossmark data

ORIGINAL ARTICLE
Bacterial associations with the hydromedusa Nemopsis bachei andscyphomedusa Aurelia aurita from the North Atlantic OceanMeaghan C. Daleya,b, Juanita Urban-Richa and Pia H. Moisanderb
aSchool for the Environment, University of Massachusetts Boston, Boston, MA, USA; bDepartment of Biology, University of MassachusettsDartmouth, North Dartmouth, MA, USA
ABSTRACTRecent studies suggest that jellyfish influence the community composition of marinemicroorganisms, but few studies have been conducted contrasting communities amongdifferent jellyfish taxa. In this study microbial communities were compared between twocnidarians found in North Atlantic Ocean coastal waters during the spring–summer months.Microbial communities associated with the invasive hydrozoan Nemopsis bachei and thecosmopolitan scyphozoan Aurelia aurita (Cnidaria) were characterized based on the 16S rRNAgene sequence. The bacterial communities associated with the jellyfish were significantlydifferent from the communities in seawater, and N. bachei and A. aurita hosted taxon-specificbacterial groups. Gammaproteobacteria, Bacteroidetes and Alphaproteobacteria dominatedbacterial sequences on N. bachei, the dominant orders including Vibrionales, Flavobacteriales,Rhizobiales and Rickettsiales. Vibrio spp. and Photobacterium spp. were abundant in N. bachei,and Tenacibaculum sp. (Bacteroidetes) had a host-specific association with N. bachei.Mycoplasmatales was a prominent, unique, and potentially host-promoted association inA. aurita, and overall Cyanobacteria, Tenericutes and unclassified bacteria dominated thesequences in A. aurita. This is the first description of the microbial community composition inN. bachei, which has been reported as an invasive species in eastern North Atlantic waters.Overall these results suggest that different cnidarians in North Atlantic coastal waterspromote growth of distinct microbial communities. Jellyfish could thus differentially influencemicrobially mediated biogeochemical cycles and food webs in regions where they proliferate.
ARTICLE HISTORYReceived 9 March 2016Accepted 13 July 2016
RESPONSIBLE EDITORHongyue Dang
KEY WORDS16S rRNA; Hydrozoa; jellyfish;microbial diversity;Mycoplasma; Vibrio
Introduction
Marine zooplankton from protozoans to mesozoo-plankton are known to host a range of microbialassociates (Grossart et al. 2013). Although the natureof such associations in zooplankton is not well under-stood, they may range from pathogenic interactionsto commensal partnerships and symbioses thatprovide important benefits to one or both of thecounterparts (Grossart et al. 2010; Tang et al. 2010).Bacteria also form associations with cnidarian jellyfish.Jellyfish-produced nutrients and dissolved organicmatter (DOM) have been shown to influence sur-rounding microbial communities (Pitt et al. 2009;Tinta et al. 2012). Investigation of the influence of jel-lyfish on marine microbial communities has drawnattention recently because increases in jellyfish abun-dances in some regions has been reported (Brotzet al. 2012), this trend potentially being linked tointer-decadal oscillations caused by large-scale
climate-driven forcing (Brotz et al. 2012; Condonet al. 2013). Jellyfish compete for food with mesozoo-plankton, thus potentially disrupting marine foodwebs (Condon et al. 2011). They withstand eutrophi-cation-linked low oxygen conditions relatively well(Breitburg et al. 2001; Purcell et al. 2001; Boeroet al. 2008), and along with climate fluctuations, over-fishing, eutrophication, aquaculture, and coastaldevelopment may all have contributed to thereported variability (Purcell et al. 2007; Richardsonet al. 2009).
Although jellyfish vary widely in size and ecology,potentially leading to significant differences in theirrelationship with microbial communities, few studiesthus far have compared the microbial communitiesassociated with different species of jellyfish collectedfrom the same geographic area. In this study thecomposition of microbial communities were com-pared in two distantly related jellyfish, Nemopsisbachei L. Agassiz, 1849 and Aurelia aurita (Linnaeus,
© 2016 Informa UK Limited, trading as Taylor & Francis Group
CONTACT Pia H. Moisander [email protected] Department of Biology, University of Massachusetts Dartmouth, 285 Old Westport Road,North Dartmouth, MA 02747, USA
The supplementary material for this article (Table SI; Figure S1) is available at http://doi:10.1080/17451000.2016.1228974
MARINE BIOLOGY RESEARCH, 2016http://dx.doi.org/10.1080/17451000.2016.1228974

1758), of the Cnidarian classes Hydrozoa and Scypho-zoa, respectively, both collected from Western NorthAtlantic coastal waters. Nemopsis bachei is a colonial,relatively small species (up to 11 mm bell height),with the polyp phase found in the Northern latitudesbetween approximately 30°N and 60°N in both theAtlantic and Pacific Oceans (Mendoza-Becerril &Marques 2013), and the range extending to the Atlan-tic Seashore and Gulf of Mexico (Marshalonis & Pinck-ney 2007; Johnson & Allen 2012). Nemopsis bachei isreported as native to the east coast of the UnitedStates and the Gulf of Mexico (Moore 1958), and asan invasive species in Northern European coastalwaters (Frost et al. 2010; Laakmann & Holst 2014). InChesapeake Bay in the Mid-Atlantic coast of theUnited States, N. bachei has been noted as the mostimportant gelatinous predator of micro- and meso-zooplankton during summer months (Purcell &Nemazie 1992), and it was the most abundant hydro-medusa in a South Carolina estuary (Marshalonis &Pinckney 2007).
In contrast to the more geographically restricted,potentially invasive N. bachei, A. aurita is cosmopolitan.Aurelia aurita is larger than N. bachei (50–300 mm),present in coastal waters and enclosed bays duringwarm summer months, and feeds on micro- and meso-zooplankton (Uye 2011). High abundances of A. auritahave been shown to lead to decreased abundancesof herbivorous zooplankton and increased abundancesof phytoplankton and protozoa (Möller 1980; Feigen-baum & Kelly 1984).
Cnidarians can have highly taxon-specific microbialcommunities, reflecting a close interdependence ofthe microbiome and the host in at least some species(Bosch et al. 2015). In addition, environmental prefer-ences and differences in food sources could promotea distinct microbiome composition among invertebratehost species (Tang et al. 2009). Differences have alsobeen reported even among subpopulations withinspecies, as well as among life stages and body parts(Weiland-Bräuer et al. 2015). The goal of this studywas to characterize and contrast bacterial communitiesassociated with N. bachei and A. aurita, both of whichare a common part of the ecosystem in the temperatecoastal waters of the Atlantic Ocean, but could havedistinct microbial signatures, differentially influencingthe jellyfish ecophysiology or the ecosystem function.The microbial communities in these jellyfish couldpotentially reflect symbioses, an induced communityresponse, feeding interactions, or some combinationof these. Jellyfish could also potentially serve asvectors or hosts of human or animal pathogens. Thismay be the first published investigation of microbial
community composition in association with N. bachei,and the first description of communities in A. auritain waters of the North-western Atlantic Ocean.
Materials and methods
Individuals of Nemopsis bachei (n = 15) were collectedfrom Mayflower Beach in Dennis, Massachusetts, USA(41.744°N, 70.219°W) in 2–4 August 2012 (hereafter’the Cape Cod site’). Specimens were collected at adepth of 1 m by towing a hand net (34 cm in diameter,2.5 mm mesh size) parallel to shore in 5 m transects.Specimens collected from 10 tows were placed in con-tainers holding 3 l of seawater, and seawater sampleswere collected (n = 3) in acid-washed and combustedIPEX or Wheaton bottles (Wheaton, Millville, NJ).Samples were transported to the University of Massa-chusetts Boston (UMB) within 3 h from collection.Aurelia aurita samples (n = 7) were collected fromBoston Harbor on 12 and 22 May 2013. Specimenswere collected using buckets off two docks (42.311°N,71.040°W and 42.293°N, 71.039°W) and placed in 3 lof seawater. Samples for parallel seawater analysis forA. aurita (n = 2) samples were collected similarly tothose for N. bachei. Boston Harbor samples ofA. aurita and parallel seawater were transported toUMB within 1 h of collection.
In the lab the jellyfish individuals were transferredfrom the seawater into 3 l of 0.2-µm filtered and auto-claved artificial seawater (ASW) at local salinities (32and 29 ppt for Cape Cod and Boston Harbor, respect-ively). ASW was made using the manufacturer’s proto-col (Instant Ocean, Blacksburg, VA). The jellyfishremained in the ASW for approximately 2–3 h inorder for the animals to evacuate their guts, afterwhich they were then rinsed three times with theASW. Nemopsis bachei individuals were transferredinto autoclaved 2 ml bead beater tubes, then flashfrozen in liquid nitrogen. Aurelia aurita were dissectedusing sterile scalpels and tweezers to select symmetri-cal portions containing parts of the bell, cilia and tenta-cles. The different parts from each individual werecombined into the same tube, then flash frozen asabove. All containers for collection and holdinganimals were acid washed. Seawater (150 ml) fromeach site was filtered through 0.2 μm Supor membranefilters (Pall-Gelman, Port Washington, NY), which werethen placed in autoclaved 2 ml bead beater tubesand flash frozen. All samples were stored at −80°Cuntil further processing.
DNA from seawater samples was extracted using aMO BIO UltraClean Tissue & Cells DNA Isolation Kit(Carlsbad, CA) using the manufacturer’s protocol.
2 M. C. DALEY ET AL.

Jellyfish samples were extracted using the same proto-col, but the samples were homogenized before usingthe kit protocol. The jellyfish samples were manuallyground for one minute using a sterile plastic pestle, fol-lowed by the DNA extraction directly or after removalof the largest jellyfish tissue particles. The large tissuepieces were removed by filtration through a 2 µm poly-carbonate filter (GE Osmonics, Greenville, SC) with20 ml of nuclease free water, and the filtrate wasthen collected and filtered on a 0.2 µm Supor filter,which was used for DNA extraction. The microbial16S rRNA gene was amplified from DNA extractedfrom jellyfish and seawater samples using polymerasechain reaction (PCR) primers 8F (5’-AGRGTTYGA-TYMTGGCTCAG-3’) (Morris et al. 2004) and 519R (5’-GWATTACCGCGGCKGCTG-3’) (Turner et al. 1999). PCRreactions consisted of 1 μl each of the primers at25 μM stock concentration, 5 μl 10× buffer, 2 μl 50-mM MgCl2, 1 μl 10-mM dNTPs, 1 U (0.2 μl) PlatinumTaq (Invitrogen, Life Technologies, Carlsbad, CA), and2 μl of template DNA. The reactions were adjusted to50 μl with nuclease free water. The PCR consisted ofan initial denaturation at 95°C for 3 min, followed by30 cycles of 94°C for 1 min, 55°C for 1 min, 72°C for 1min and extension at 72°C for 7 min. Products wereseparated on a 1.2% TAE agarose gel and visualizedwith ethidium bromide. Bands were excised and gelpurified, then ligated into the pGEM-T vector, trans-formed into competent E. coli JM109, and grown onLB plates under ampicillin selection using the
manufacturer’s protocol (Promega, Madison, WI).Sequencing was performed at the MassachusettsGeneral Hospital DNA Core Facility at Harvard Univer-sity (Cambridge, MA), the sequences were trimmedusing CLC Main Workbench 6 (CLC Bio, Cambridge,MA), and a chimera check was conducted using DECI-PHER (Wright et al. 2012). Sequences were identifiedusing BLASTn and the RDP Classifier (Cole et al. 2009),the latter based on an 80% threshold for phylum identi-fication. The sequences were aligned using the onlineSINA aligner (Pruesse et al. 2012) and imported intothe 16S rRNA SILVA database release 115 (Quast et al.2013) in ARB (Ludwig et al. 2004) for creating neigh-bour-joining phylogenetic trees. Bootstrapping wasperformed in MEGA (version 5.2.2) with 1000 replicates.The 16S rRNA sequences from this study are inGenBank under accession numbers KP198297–KP198503 (Table S1). Nemopsis bachei identificationwas confirmed by conducting PCR targeting the mito-chondrial cytochrome oxidase gene as previouslydescribed (Folmer et al. 1994; Bucklin et al. 1999; Shoe-maker & Moisander 2015). The COI sequence GenBankaccession numbers are KX265102–KX265109.
Statistical comparisons between bacterial commu-nities were conducted (redundancy analysis and analy-sis of variance) using R Studio. The sequenceabundances in different OTUs were standardized toproportion of all sequences. The Euclidian distancewas used to compare operational taxonomic unit(OTU, 97% DNA identity) presence and abundancebased on Hellinger transformed per cent compositionOTU data (Legendre & Gallagher 2001). An ordinationplot was created using a redundancy analysis (RDA),with the constrained eigenvalues representing thesample type (jellyfish or seawater) and location(Boston Harbor or Cape Cod). Analysis of variance(ANOVA) was performed on the RDA to test for differ-ences in bacterial communities among the samples.The ANOVA tests were run with 999 permutationsand P < 0.05 was considered significant.
Results
In the initial PCR reactions to amplify microbial 16SrRNA, the high abundance of jellyfish DNA in thesamples resulted in strong amplification of the 18SrRNA gene from the jellyfish. In subsequent DNAextractions, the majority of the homogenized jellyfishtissue was removed through a 2 µm filter and the fil-trate used for DNA extraction and PCR (see Methods).In most subsequent PCR reactions, bands for both18S rRNA and 16S rRNA were observed, but using posi-tive controls for each, the two bands could be
Figure 1. Per cent composition of bacterial phyla associatedwith Cape Cod seawater (three samples, total of 61 sequences),Nemopsis bachei (15 individuals, total of 92 sequences), Bostonseawater (two samples, total of 41 sequences) and Aureliaaurita (seven individuals, total of 51 sequences) based onRDP classifier results on 90% similarity.
MARINE BIOLOGY RESEARCH 3

distinguished in gel electrophoresis and 16S rRNAbands selectively excised, cloned and sequenced(Figure S1, supplementary material). A total of 245sequences were recovered, of which 207 wereunique. A total of 74 and 43 unique sequences wererecovered from Nemopsis bachei and Aurelia aurita,respectively, and 50 and 40 unique sequences wererecovered from the seawater collected in parallel withN. bachei (Cape Cod seawater) and A. aurita (BostonHarbor), respectively. All of the COI sequences fromthe individuals morphologically identified as N. bacheihad a 99% nucleotide identity with N. bachei(sampled from the North Sea, GenBank accessionKC440112.1).
Microbial community in Nemopsis bachei andthe Cape Cod seawater
Gammaproteobacterial sequences formed a total of45% and 7% of Nemopsis bachei and Cape Cod sea-water sequences, respectively (Figure 1). Vibrionaleswere the most abundant Gammaproteobacteria inN. bachei, forming 35% of all sequences. Sequencesfrom Oceanospirillales, Enterobacteriales, Alteromona-dales and Vibrionales were all found in associationwith N. bachei but not in Cape Cod seawater (Figure2). The majority of the Vibrionales sequences fromN. bachei clustered with Vibrio spp., and one clusterof sequences within Vibrionales was closely relatedwith Photobacterium swingsii (HE584802, 99% identity).A cluster of Enterobacteriaceae sequences fromN. bachei was closely related with Shigella flexneri(ADUV01000046, 97–99% identity).
Sequences from both N. bachei and the surroundingseawater contained Alphaproteobacteria (20% ofN. bachei sequences and 10% of Cape Cod seawatersequences) but were from different groups in N. bacheiand in the seawater. The majority of alphaproteobacter-ial sequences from N. bachei grouped with Rickettsiales(AF206298 Ehrlichia sp. 91–92% identity and U12457Neorickettsia helminthoeca 90–91% identity) (Figure 2),while one sequence grouped with Rhizobiales(NR_117855, Devoisa glacialis 99% identity). In contrast,Rhodospirillaceae and Rhodobacteraceae (Alphaproteo-bacteria) sequences were recovered from the Cape Codseawater samples but not from N. bachei. Betaproteo-bacteria formed 7% of Cape Cod seawater sequencesbut were not found in N. bachei (Figure 1).
Bacteroidetes represented 8% of the sequencesfrom N. bachei but they were not obtained from theparallel seawater (Figures 1 and 3). Of the total of 92sequences from N. bachei, three sequences had a94% identity with Polaribacter sp. (HM010403), one
had a 99% identity with Tenacibaculum mesophilum(AB681059), and one had a 96% identity with Psychro-serpens damuponensis (NR_109097), all withinFlavobacteriales.
Sequences from cyanobacteria or chloroplastsformed 52% of all sequences from the Cape Cod sea-water and 24% of sequences from N. bachei (Figures1 and 3). The majority of them were within the cladeGpIIa that encompasses the picocyanobacteria Syn-echococcus and Prochlorococcus (HM057705 andGU170756, 98–100% identity). One of the phylotypes,closely related to Synechococcus WH 8102, waspresent in both N. bachei and the parallel seawater,while two of the Synechococcus phylotypes (Synecho-coccus CC9311 and CC9902) were present in the sea-water only. An additional cyanobacterial sequencewas recovered from N. bachei clustering with thenon-heterocystous, filamentous cyanobacterium Lepto-lyngbya sp. (JX481735, 97% identity).
Within Firmicutes, two sequences were recoveredfrom N. bachei but none were obtained from anyother sample types; these sequences had a high iden-tity with Streptococcus salivarius (AB680535; 99% iden-tity), falling in the Lactobacillales order (Figure 4).Planctomycetes, Actinobacteria and Verrucomicrobiarepresented 16%, 3% and 2% of the sequences in theCape Cod seawater (Figure 1) but were not found inN. bachei. One N. bachei sequence had a 90%identity with an uncultured Epsilonproteobacterium(AB189341, isolated from a cold-seep). Of thesequences from Cape Cod seawater, 3% were unclassi-fied, with 98% identity to an uncultured marine bacter-ium (HM225808) (Figure 1).
Microbial community in Aurelia aurita and theBoston Harbor seawater
As in Nemopsis bachei, microbial communities fromAurelia aurita and the parallel seawater were domi-nated by different groups (Figure 1). Gammaproteo-bacteria formed 6% and 2% of the sequences fromA. aurita and the corresponding seawater, respectively.One gammaproteobacterial phylotype was recoveredfrom A. aurita, clustering with Psychrobacter immobilis(U85880, 96–97% identity). In the Boston Harbor sea-water, another gammaproteobacterial sequence type,closely related with Vibrio sp. S3791 (FJ457558, 99%identity), was present that was identical to sequencesfrom N. bachei sampled from the Cape Cod watersfurther south (Figure 2). Alphaproteobacteria formed22% of the community in the Boston Harbor seawater,and a smaller proportion (4%) of sequences fromA. aurita (Figure 1). An alphaproteobacterial sequence
4 M. C. DALEY ET AL.

type within Rickettsiales was recovered from A. aurita(Figure 2). Alphaproteobacterial sequences that werenot detected in any other sample type were recovered
from Boston Harbor seawater, including a sequencefrom the SAR11 cluster, and sequences clusteringwith Rhodobacteria.
Figure 2. Neighbour-joining tree of 16S rRNA sequences clustering with (a) Alphaproteobacteria and (b) Gammaproteobacteria.Bootstrap values are shown where the value was 50% or higher based on 1000 permutations.
MARINE BIOLOGY RESEARCH 5

Flavobacteriales within Bacteroidetes were highlyrepresented in the Boston Harbor seawater. However,Bacteroidetes formed only 2% of sequences inA. aurita (99% identity with an uncultured Bacteroi-detes/Chlorobi HQ242389 from the Northeast Pacificseawater) (Figure 3). Cyanobacteria were detected inboth A. aurita and Boston Harbor seawater. All cyano-bacterial sequences from A. aurita had the greatestidentity with cyanobacteria similar to the
Synechococcus CC9902 related phylotype obtainedfrom the Cape Cod seawater (Figure 3). The sequencesof this phylotype made up a large portion of all of thesequences from A. aurita (35%). Chloroplast sequencesfrom Boston Harbor seawater accounted for 15% oftotal sequences from this sample type and clusteredclosely with uncultured Streptophyta (98% identity,JF703621). Sequences clustering within the phylumTenericute of the class Mollicutes and order
Figure 3. Neighbour-joining tree of 16S rRNA sequences clustering with (a) Bacteroidetes and (b) Cyanobacteria. Bootstrap valuesare shown where the value was 50% or higher based on 1000 permutations.
6 M. C. DALEY ET AL.

Mycoplasmatales were found at a high proportionamong A. aurita sequences but not in any othersample type (Figure 4). Sequences distantly related toMycoplasma represented as much as 20% of thesequences from A. aurita (84% identity with Myco-plasma mobile strain NR_074620 and 85% identitywith an uncultured Mollicutes bacterium KC511181)(Figure 4). Thirty-one per cent of the sequences inA. aurita were distant (< 79% identity) to any classifiedbacteria.
Microbial community in cnidarians vs. seawater
Rarefaction curves for Nemopsis bachei and Cape Codseawater had a similar slope, suggesting that thelevel of richness recovered through sampling was
closely similar (Figure 5). Both jellyfish had lower diver-sity than that of the surrounding seawater at thephylum level, but at the OTU level, N. bachei diversitywas higher than that in the parallel seawater (Table I).The microbial community composition of N. bacheiand that of Cape Cod seawater was significantly differ-ent based on a one-way ANOVA on a redundancyanalysis (RDA) whether the analysis was conductedby grouping the sequences by phylum (P = 0.048) orOTU (P = 0.009) (Figure 6).
Rarefaction curves suggested that Aurelia aurita hadthe lowest OTU richness of the four sample types(Figure 5). The richness in Boston Harbor and CapeCod seawater was relatively similar, and similar toN. bachei when comparing the curves at same samplesize as the seawater samples. Microbial communitiesin A. aurita and Boston Harbor seawater were statisti-cally different based on ANOVA and RDA analysiswhether the sequences were discriminated byphylum (P = 0.045) or OTU (P = 0.049) (Figure 6). The
Figure 4. Neighbour-joining tree of 16S rRNA sequences clustering with Tenericutes, Firmicutes, Betaproteobacteria, Actinobac-teria, Verrucomicrobia and Planctomycetes. Bootstrap values are shown where the value was 50% or higher based on 1000permutations.
Figure 5. Rarefaction curves for the sequences recovered fromthe different sample types based on 97% similarity. Number ofOTUs shown observed as a function of sample size in eachsample type (# of sequences). 1, Cape Cod seawater; 2,Boston Harbor seawater; 3, Nemopsis bachei; 4, Aurelia aurita.
Table I. Shannon H diversity. Diversity calculated on thenumber of sequences identified at OTU and phylum level(OTU defined as 97% similarity).
#samples
#sequences
Shannon H(OTU)
Shannon H(phylum)
Nemopsisbachei
15 92 3.66 1.46
Cape Codseawater
3 61 3.30 1.57
Aurelia aurita 7 51 2.92 1.57Boston Harborseawater
2 41 3.20 1.64
MARINE BIOLOGY RESEARCH 7
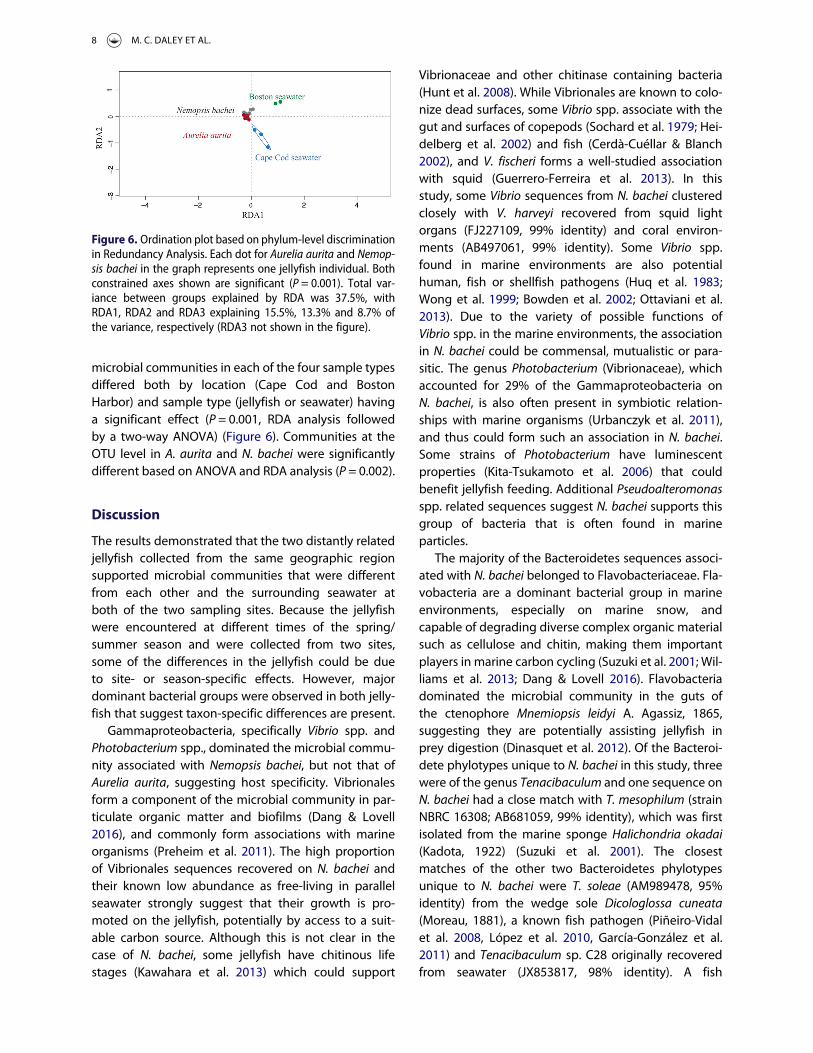
microbial communities in each of the four sample typesdiffered both by location (Cape Cod and BostonHarbor) and sample type (jellyfish or seawater) havinga significant effect (P = 0.001, RDA analysis followedby a two-way ANOVA) (Figure 6). Communities at theOTU level in A. aurita and N. bachei were significantlydifferent based on ANOVA and RDA analysis (P = 0.002).
Discussion
The results demonstrated that the two distantly relatedjellyfish collected from the same geographic regionsupported microbial communities that were differentfrom each other and the surrounding seawater atboth of the two sampling sites. Because the jellyfishwere encountered at different times of the spring/summer season and were collected from two sites,some of the differences in the jellyfish could be dueto site- or season-specific effects. However, majordominant bacterial groups were observed in both jelly-fish that suggest taxon-specific differences are present.
Gammaproteobacteria, specifically Vibrio spp. andPhotobacterium spp., dominated the microbial commu-nity associated with Nemopsis bachei, but not that ofAurelia aurita, suggesting host specificity. Vibrionalesform a component of the microbial community in par-ticulate organic matter and biofilms (Dang & Lovell2016), and commonly form associations with marineorganisms (Preheim et al. 2011). The high proportionof Vibrionales sequences recovered on N. bachei andtheir known low abundance as free-living in parallelseawater strongly suggest that their growth is pro-moted on the jellyfish, potentially by access to a suit-able carbon source. Although this is not clear in thecase of N. bachei, some jellyfish have chitinous lifestages (Kawahara et al. 2013) which could support
Vibrionaceae and other chitinase containing bacteria(Hunt et al. 2008). While Vibrionales are known to colo-nize dead surfaces, some Vibrio spp. associate with thegut and surfaces of copepods (Sochard et al. 1979; Hei-delberg et al. 2002) and fish (Cerdà-Cuéllar & Blanch2002), and V. fischeri forms a well-studied associationwith squid (Guerrero-Ferreira et al. 2013). In thisstudy, some Vibrio sequences from N. bachei clusteredclosely with V. harveyi recovered from squid lightorgans (FJ227109, 99% identity) and coral environ-ments (AB497061, 99% identity). Some Vibrio spp.found in marine environments are also potentialhuman, fish or shellfish pathogens (Huq et al. 1983;Wong et al. 1999; Bowden et al. 2002; Ottaviani et al.2013). Due to the variety of possible functions ofVibrio spp. in the marine environments, the associationin N. bachei could be commensal, mutualistic or para-sitic. The genus Photobacterium (Vibrionaceae), whichaccounted for 29% of the Gammaproteobacteria onN. bachei, is also often present in symbiotic relation-ships with marine organisms (Urbanczyk et al. 2011),and thus could form such an association in N. bachei.Some strains of Photobacterium have luminescentproperties (Kita-Tsukamoto et al. 2006) that couldbenefit jellyfish feeding. Additional Pseudoalteromonasspp. related sequences suggest N. bachei supports thisgroup of bacteria that is often found in marineparticles.
The majority of the Bacteroidetes sequences associ-ated with N. bachei belonged to Flavobacteriaceae. Fla-vobacteria are a dominant bacterial group in marineenvironments, especially on marine snow, andcapable of degrading diverse complex organic materialsuch as cellulose and chitin, making them importantplayers in marine carbon cycling (Suzuki et al. 2001; Wil-liams et al. 2013; Dang & Lovell 2016). Flavobacteriadominated the microbial community in the guts ofthe ctenophore Mnemiopsis leidyi A. Agassiz, 1865,suggesting they are potentially assisting jellyfish inprey digestion (Dinasquet et al. 2012). Of the Bacteroi-dete phylotypes unique to N. bachei in this study, threewere of the genus Tenacibaculum and one sequence onN. bachei had a close match with T. mesophilum (strainNBRC 16308; AB681059, 99% identity), which was firstisolated from the marine sponge Halichondria okadai(Kadota, 1922) (Suzuki et al. 2001). The closestmatches of the other two Bacteroidetes phylotypesunique to N. bachei were T. soleae (AM989478, 95%identity) from the wedge sole Dicologlossa cuneata(Moreau, 1881), a known fish pathogen (Piñeiro-Vidalet al. 2008, López et al. 2010, García-González et al.2011) and Tenacibaculum sp. C28 originally recoveredfrom seawater (JX853817, 98% identity). A fish
Figure 6. Ordination plot based on phylum-level discriminationin Redundancy Analysis. Each dot for Aurelia aurita and Nemop-sis bachei in the graph represents one jellyfish individual. Bothconstrained axes shown are significant (P = 0.001). Total var-iance between groups explained by RDA was 37.5%, withRDA1, RDA2 and RDA3 explaining 15.5%, 13.3% and 8.7% ofthe variance, respectively (RDA3 not shown in the figure).
8 M. C. DALEY ET AL.

pathogen T. maritimum was previously reported in thehydromedusa Phialella quadrata (Forbes, 1848) whereit may influence digestion of food particles (Fergusonet al. 2010). Based on this and previous studies, jellyfish,including N. bachei, may serve as vectors for severalTenacibaculum spp. (Avendaño-Herrera et al. 2006, Fer-guson et al. 2010, Delannoy et al. 2011).
Lactobacillales in Firmicutes was also specific toN. bachei compared with the surrounding seawater.Additionally, Rickettsiales within Alphaproteobacteriawere present in N. bachei but not in the Cape Cod sea-water, Rickettsiales related to Neorickettsia accountingfor 20% of Alphaproteobacterial sequences onN. bachei. Related but different sequences found inA. aurita suggest importance of this group for jellyfish.Rickettsiales include many endosymbionts or patho-gens (Walker 1996) as well as the important marinefree-living SAR11 cluster of Alphaproteobacteria thatwere distant to the sequences recovered fromN. bachei. The Neorickettsia sequences from N. bacheiwere most closely similar to Ehrlichia sp. (AF206298,91–92% similarity) and Neorickettsia helminthoeca(U12457, 90–91% identity).
The results suggest that bacteria within Gamma-and Alphaproteobacteria, Bacteroidetes, Firmicutes,and possibly Cyanobacteria may associate withN. bachei. These are typical seawater phyla whoserepresentatives also associate with other marineanimals such as corals (Harder et al. 2003), sponges(Schmitt et al. 2012), copepods (Tang et al. 2010) andctenophores (Daniels & Breitbart 2012). Notably, acommon marine snow colonizer group Roseobacterwas not detected on the jellyfish. Although we canonly speculate on the function of the associations,the distinct communities found in the cnidarians andseawater suggest some of them are host-promotedand taxon-specific.
Bacteria suggestive of both mutualistic and parasiticcapabilities were found also on A. aurita. Mycoplasmawithin Tenericutes was specific to A. aurita comparedwith all sample types, and represented a large pro-portion of the sequences recovered from A. aurita,suggesting the host promoted the association.Results from this study and a previous study on cteno-phores (Daniels & Breitbart 2012), suggest that Myco-plasmatales are common associates in marinejellyfish; however, Mycoplasmatales were not detectedin N. bachei, suggesting these associations are jellyfishtaxon-specific. Mycoplasmatales have been found inthe marine invertebrate bryozoanWatersipora cucullata(Busk, 1854) (Zimmer & Woollacott 1983; Rottem 2003)as well as in corals (Penn et al. 2006; Kellogg et al. 2009;Neulinger et al. 2009). Some Mycoplasmatales
parasitize plants and animals to obtain nutrients, andcause human illnesses such as pneumonia and infec-tions (Rottem 2003; Pitcher & Nicholas 2005). One ofthe strains clustering closely with sequences fromA. aurita was Mycoplasma mobile, first isolated from afish. Its genome analysis showed that M. mobile isable to transport and metabolize several sugars (Jaffeet al. 2004) which could aid in obtaining nutrientsfrom hosts such as jellyfish. Mycoplasma could beobtaining nutrients from A. aurita, thus forming a com-mensal, mutualistic or parasitic association. A likelyendosymbiont in the Mycoplasma genus was recentlyreported for A. aurita in the Eastern North Atlanticpopulations (Weiland-Bräuer et al. 2015). Our studyprovides support to the idea that Mycoplasma spp.are common endosymbionts in various subpopu-lations/clades of A. aurita, although the mechanismsof this association are unknown.
The Gammaproteobacteria Psychrobacter spp. (clus-tering closely with P. immobilis; Figure 2), found to bespecific to A. aurita compared with the surroundingseawater, contain species that have been isolatedfrom a wide range of habitats that vary in temperature,salinity and oxygen levels (Bowman 2006). Thus, it isdifficult to speculate on the nature of the associationof A. aurita with this phylotype.
The high proportion of picocyanobacteria Synecho-coccus spp. of the microbial community in A. auritawas somewhat unexpected, since the jellyfish weretreated to empty their gut contents before preserving.While some of the sequences could be remnants of thegut contents, they could alternatively indicate morestable epi- or endobiotic associations, or the cyanobac-teria could have been entangled in the outer mucus ofthe jellyfish. The mucus of A. aurita and other jellyfishspecies is a rich source of labile carbon and can effec-tively entrap nanoparticles (Patwa et al. 2015) whichcould also lead to concentration of surrounding bac-teria. While dominant free-living cyanobacteria incoastal waters, Synechococcus spp. are also found inmutualistic associations with marine animals such assome sponges, where they provide organic carbon tothe sponges via photosynthesis, while the spongereturns metabolic waste products useful to the bacter-ium (Taylor et al. 2007). Clade GpIIa cyanobacteria havebeen found in association with sponges (White et al.2012) and the cyanobacteria Synechoccocus (strainPCC7943) was found in association with corals(Rohwer et al. 2002), suggesting that photosyntheticnutrients and byproducts may play an important rolebetween the host and bacterium. Synechococcus sp.strain CC9902 (clade IV), that grouped most closelywith sequences obtained from A. aurita (CP000097,
MARINE BIOLOGY RESEARCH 9

99% identity), is a common component in coastalsurface waters of Southern California (Palenik et al.2009), suggesting the sequences on A. aurita mayhave been from either feeding or entanglement onthe jellyfish mucus (Patwa et al. 2015).
The clone libraries for different sample types arecomposed of pooled libraries from several individualsamong which there was variability in sequences recov-ered, hence the data suggest variability is present incommunities among jellyfish individuals. This may bea real trend, but sequence coverage depth availablefor this study is a limitation for drawing conclusionsabout the community stability among the jellyfish indi-viduals. However, the RDA analysis incorporates thisvariability and even at this relatively small sample sizeallowed separation of the communities in differentsample types. Deeper sequencing would allow amore detailed assessment of these communities. Allprimers have certain biases and it is possible suchbiases influenced our data as well; however, the 16SrRNA primers used in this study were designed as uni-versal and have been used in comparative studies ofmarine bacterioplankton communities (Morris et al.2004; Treusch et al. 2009). Jellyfish blooms are patchyand seasonal in the waters where sampling was con-ducted. The two species typically occur at differenttimes of the year and were sampled in the spring(A. aurita) and summer (N. bachei), which may haveinfluenced the results due to changing temperatures,food availability, and other environmental factors. Theinfluence of seasonality on the microbiomes of thesejellyfish merits further study. The degree of anthropo-genic influence at the sampling sites may also haveplayed a role in structuring the communities observed,as Boston Harbor is influenced by pollution from amajor urban city, and the Cape Cod Bay experiencesat least some effects from elevated nutrients fromcoastal activities.
The taxon-specific associations in the hydromedusaN. bachei and scyphomedusa A. aurita demonstratedby this and other studies overall suggest that the rela-tive fitness of different jellyfish taxa plays a role inshaping microbial communities in coastal ecosystems.Low oxygen tolerant N. bachei may be promoted inthe zooplankton community in coastal and estuarinewaters under periodic hypoxia (Marshalonis & Pinckney2007), a condition enhanced by eutrophication. Thereported microbial communities dominated by Vibriospp. are notable in the case of N. bachei that currentlyappears to be expanding its range to European waters(Frost et al. 2010). The observation of Mycoplasma spp.in A. aurita in Western North Atlantic waters in thisstudy suggests this association is a stable symbiosis
found in several subpopulations of the species(Weiland-Bräuer et al. 2015) in geographically distantlocations.
Acknowledgements
We thank Eugene Gallagher and Scott Morey (University ofMassachusetts Boston) for help with statistical analyses. Com-ments from the anonymous reviewers and the AssociateEditor improved the manuscript.
Disclosure statement
No potential conflict of interest was reported by the authors.
Funding
The work was supported by the University of MassachusettsSchool of Marine Science to Pia Moisander and JuanitaUrban-Rich, and a National Science Foundation award[grant number OCE-1130495] to Pia Moisander.
ORCID
Pia H. Moisander http://orcid.org/0000-0002-3262-3662
References
Avendaño-Herrera R, Toranzo AE, Magariños B. 2006.Tenacibaculosis infection in marine fish caused byTenacibaculum maritimum: a review. Diseases of AquaticOrganisms 71:255–66. doi:10.3354/dao071255
Boero F, Bouillon J, Gravili C, Miglietta MP, Parsons T, Piraino S.2008. Gelatinous plankton: irregularities rule the world(sometimes). Marine Ecology Progress Series 356:299–310. doi:10.3354/meps07368
Bosch TCG, Grasis JA, Lachnit T. 2015. Microbial ecology inHydra: why viruses matter. Journal of Microbiology53:193–200. doi:10.1007/s12275-015-4695-2
Bowden TJ, Menoyo-Luque D, Bricknell IR, Wergeland H. 2002.Efficacy of different administration routes for vaccinationagainst Vibrio anguillarum in Atlantic halibut(Hippoglossus hippoglossus L.). Fish & ShellfishImmunology 12:283–85. doi:10.1006/fsim.2001.0386
Bowman JP. 2006. The genus Psychrobacter. In: Dworkin M,Falkow S, Rosenberg E, Schleifer K-H, Stackebrandt E,editors. The Prokaryotes. Volume 6. New York: Springer, p920–30.
Breitburg DL, Pihl L, Kolesar SE. 2001. Effects of low dissolvedoxygen on the behavior, ecology and harvest of fishes: acomparison of the Chesapeake Bay and Baltic-Kattegatsystems. In: Rabalais NN, editor. Coastal Hypoxia:Consequences for Living Resources and Ecosystems.Coastal and Estuarine Sciences, Volume 58. Washington,DC: American Geophysical Union, p 241–67.
Brotz L, Cheung WWL, Kleisner K, Pakhomov E, Pauly D. 2012.Increasing jellyfish populations: trends in large marine eco-systems. Hydrobiologia 690:3–20. doi:10.1007/s10750-012-1039-7
10 M. C. DALEY ET AL.

Bucklin A, Guarnieri M, Hill RS, Bentley AM, Kaartvedt S. 1999.Taxonomic and systematic assessment of planktonic cope-pods using mitochondrial COI sequence variation andcompetitive, species-specific PCR. Hydrobiologia 401:239–54. doi:10.1023/A:1003790411424
Cerdà-Cuéllar M, Blanch AR. 2002. Detection and identifi-cation of Vibrio scophthalmi in the intestinal microbiotaof fish and evaluation of host specificity. Journal ofApplied Microbiology 93:261–68. doi:10.1046/j.1365-2672.2002.01697.x
Cole JR, Wang Q, Cardenas E, Fish J, Chai B, Farris RJ, et al.2009. The ribosomal database project: improved align-ments and new tools for rRNA analysis. Nucleic AcidsResearch 37(Suppl 1):D141–45. doi:10.1093/nar/gkn879
Condon RH, Steinberg DK, del Giorgio PA, Bouvier TC, BronkDA, Graham WM, Ducklow HW. 2011. Jellyfish bloomsresult in a major microbial respiratory sink of carbon inmarine systems. Proceedings of the National Academy ofSciences 108:10225–30. doi:10.1073/pnas.1015782108
Condon RH, Duarte CM, Pitt KA, Robinson KL, Lucas CH,Sutherland KR, et al. 2013. Recurrent jellyfish blooms area consequence of global oscillations. Proceedings of theNational Academy of Sciences 110:1000–05. doi:10.1073/pnas.1210920110
Dang H, Lovell CR. 2016. Microbial surface colonization andbiofilm development in marine environments.Microbiology and Molecular Biology Reviews 80:91–138.doi:10.1128/MMBR.00037-15
Daniels C, Breitbart M. 2012. Bacterial communities associatedwith the ctenophores Mnemiopsis leidyi and Beroe ovata.FEMS Microbiology Ecology 82:90–101. doi:10.1111/j.1574-6941.2012.01409.x
Delannoy CMJ, Houghton JDR, Fleming NEC, Ferguson HW.2011.Mauve stingers (Pelagia noctiluca) as carriers of thebac-terial fish pathogen Tenacibaculum maritimum. Aquaculture311:255–57. doi:10.1016/j.aquaculture.2010.11.033
Dinasquet J, Granhag L, Riemann L. 2012. Stimulated bacter-ioplankton growth and selection for certain bacterial taxain the vicinity of the ctenophore Mnemiopsis leidyi.Frontiers in Microbiology 3:1–8. doi:10.3389/fmicb.2012.00302
Feigenbaum D, Kelly M. 1984. Changes in the lowerChesapeake Bay food-chain in presence of the sea nettleChrysaora quinquecirrha (Scyphomedusa). Marine EcologyProgress Series 19:39–47. doi:10.3354/meps019039
Ferguson HW, Delannoy CMJ, Hay S, Nicolson J, Sutherland D,Crumlish M. 2010. Jellyfish as vectors of bacterial diseasefor farmed salmon (Salmo salar). Journal of VeterinaryDiagnostic Investigation 22:376–82. doi:10.1177/104063871002200305
Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R. 1994. DNAprimers for amplification of mitochondrial cytochrome coxidase subunit I from diverse metazoan invertebrates.Molecular Marine Biology and Biotechnology 3:294–99.
Frost JR, Jacoby CA, Youngbluth MJ. 2010. Behavior ofNemopsis bachei L. Agassiz, 1849 medusae in the presenceof physical gradients and biological thin layers.Hydrobiologia 645:97–111. doi:10.1007/s10750-010-0213-z
García-González P, García-Lamas N, Edfuf CF, Santos Y. 2011.Development of a PCR method for the specific identificationof the marine fish pathogen Tenacibaculum soleae.Aquaculture 319:1–4. doi:10.1016/j.aquaculture.2011.06.013
Grossart HP, Dziallas C, Leunert F, Tang KW. 2010. Bacteria dis-persal by hitchhiking on zooplankton. Proceedings of theNational Academy of Sciences 107:11959–64. doi:10.1073/pnas.1000668107
Grossart HP, Riemann L, Tang KW. 2013. Molecular and func-tional ecology of aquatic microbial symbionts. Frontiers inMicrobiology 4:1–2. doi:10.3389/fmicb.2013.00059
Guerrero-Ferreira R, Gorman C, Chavez AA, Willie S, NishiguchiMK. 2013. Characterization of the bacterial diversity in Indo-West Pacific loliginid and sepiolid squid light organs.Microbial Ecology 65:214–26. doi:10.1007/s00248-012-0099-6
Harder T, Lau SCK, Dobretsov S, Fang TK, Qian PY. 2003. A dis-tinctive epibiotic bacterial community on the soft coralDendronephthya sp. and antibacterial activity of coraltissue extracts suggest a chemical mechanism against bac-terial epibiosis. FEMS Microbiology Ecology 43:337–47.doi:10.1111/j.1574-6941.2003.tb01074.x
Heidelberg JF, Heidelberg KB, Colwell RR. 2002. Bacteria of theγ-subclass Proteobacteria associated with zooplankton inChesapeake Bay. Applied and Environmental Microbiology68:5498–507. doi:10.1128/AEM.68.11.5498-5507.2002
Hunt DE, Gevers D, Vahora NM, Polz MF. 2008. Conservationof the chitin utilization pathway in the Vibrionaceae.Applied and Environmental Microbiology 74:44–51.doi:10.1128/AEM.01412-07
Huq A, Small EB, West PA, Huq MI, Rahman R, Colwell RR.1983. Ecological relationships between Vibrio choleraeand planktonic crustacean copepods. Applied andEnvironmental Microbiology 45:275–83.
Jaffe JD, Stange-Thomann N, Smith C, DeCaprio D, Fisher S,Butler J, et al. 2004. The complete genome and proteomeof Mycoplasma mobile. Genome Research 14:1447–61.doi:10.1101/gr.2674004
Johnson WS, Allen DM. 2012. Zooplankton of the Atlantic andGulf Coasts – A Guide to their Identification and Ecology.Baltimore, MD: The Johns Hopkins University Press. 472pages.
Kawahara M, Ohtsu K, Uye SI. 2013. Bloom or non-bloom inthe giant jellyfish Nemopilema nomurai (Scyphozoa:Rhizostomeae): roles of dormant podocysts. Journal ofPlankton Research 35:213–17. doi:10.1093/plankt/fbs074
Kellogg CA, Lisle JT, Galkiewicz JP. 2009. Culture-independentcharacterization of bacterial communities associated withthe cold-water coral Lophelia pertusa in the NortheasternGulf of Mexico. Applied and Environmental Microbiology75:2294–303. doi:10.1128/AEM.02357-08
Kita-Tsukamoto K, Yao K, Kamiya A, Yoshizawa S, Uchiyama N,Kogure K, Wada M. 2006. Rapid identification of marinebioluminescent bacteria by amplified 16S ribosomal RNAgene restriction analysis. FEMS Microbiology Letters256:298–303. doi:10.1111/j.1574-6968.2006.00129.x
Laakmann S, Holst S. 2014. Emphasizing the diversity of NorthSea hydromedusae by combined morphological and mol-ecular methods. Journal of Plankton Research 36:64–76.doi:10.1093/plankt/fbt078
Legendre P, Gallagher ED. 2001. Ecologically meaningfultransformations for ordination of species data. Oecologia129:271–80. doi:10.1007/s004420100716
López JR, Piñeiro-Vidal M, García-Lamas N, de la Herran R,Navas JI, Hachero-Cruzado I, Santos Y. 2010. First iso-lation of Tenacibaculum soleae from diseased culturedwedge sole, Dicologoglossa cuneata (Moreau), and brill,
MARINE BIOLOGY RESEARCH 11

Scophthalmus rhombus (L.). Journal of Fish Diseases33:273–78. doi:10.1111/j.1365-2761.2009.01105.x
Ludwig W, Strunk O, Westram R, Richter L, Meier H,Yadhukumar, et al. 2004. ARB: a software environmentfor sequence data. Nucleic Acids Research 32:1363–71.doi:10.1093/nar/gkh293
Marshalonis D, Pinckney JL. 2007. Respiration rates of domi-nant hydromedusae in the North Inlet tidal estuaryduring winter and summer. Journal of Plankton Research29:1031–40. doi:10.1093/plankt/fbm077
Mendoza-Becerril MA, Marques AC. 2013. Synopsis on theknowledge and distribution of the family Bougainvilliidae(Hydrozoa, Hydroidolina). Latin American Journal of AquaticResearch 41:908–24. doi:10.3856/vol41-issue5-fulltext-11
Möller H. 1980. Scyphomedusae as predators and food com-petitors of larval fish. Meeresforschung 28:90–100.
Moore DR. 1958. Occurrence and distribution of Nemopsisbachei Agassiz (Hydrozoa) in the Northern Gulf ofMexico. Bulletin of Marine Science of the Gulf andCaribbean 12:399–402.
Morris RM, Rappe MS, Urbach E, Connon SA, Giovannoni SJ.2004. Prevalence of the Chloroflexi-related SAR202 bacter-ioplankton cluster throughout the mesopelagic zone anddeep ocean. Applied and Environmental Microbiology70:2836–42. doi:10.1128/AEM.70.5.2836-2842.2004
Neulinger SC, Gärtner A, Järnegren J, Ludvigsen M, Lochte K,Dullo WC. 2009. Tissue-associated “CandidatusMycoplasma corallicola” and filamentous bacteria on thecold-water coral Lophelia pertusa (Scleractinia). Appliedand Environmental Microbiology 75:1437–44. doi:10.1128/AEM.01781-08
Ottaviani D, Leoni F, Rocchegiani E, Mioni R, Costa A, Virgilio S,et al. 2013. An extensive investigation into the prevalenceand the genetic and serological diversity of toxigenic Vibrioparahaemolyticus in Italian marine coastal waters.Environmental Microbiology 15:1377–86. doi:10.1111/j.1462-2920.2012.02839.x
Palenik B, Ren Q, Tai V, Paulsen IT. 2009. CoastalSynechococcus metagenome reveals major roles for hori-zontal gene transfer and plasmids in population diversity.Environmental Microbiology 11:349–59. doi:10.1111/j.1462-2920.2008.01772.x
Patwa A, Thiéry A, Lombard F, Lilley MKS, Boisset C, BranardJ-F, et al. 2015. Accumulation of nanoparticles in “jelly-fish” mucus: a bio-inspired route to decontamination ofnano-waste. Scientific Reports 5:11387. 8 pages. doi:10.1038/srep11387
Penn K, Wu DY, Eisen JA, Ward N. 2006. Characterization ofbacterial communities associated with deep-sea corals onGulf of Alaska seamounts. Applied and EnvironmentalMicrobiology 72:1680–83. doi:10.1128/AEM.72.2.1680-1683.2006
Piñeiro-Vidal M, Carballas CG, Gómez-Barreiro O, Riaza A,Santos Y. 2008. Tenacibaculum soleae sp nov., isolatedfrom diseased sole (Solea senegalensis Kaup).International Journal of Systematic and EvolutionaryMicrobiology 58:881–85. doi:10.1099/ijs.0.65539-0
Pitcher DG, Nicholas RAJ. 2005. Mycoplasma host specificity:fact or fiction? The Veterinary Journal 170:300–06. doi:10.1016/j.tvjl.2004.08.011
Pitt KA, Welsh DT, Condon RH. 2009. Influence of jellyfishblooms on carbon, nitrogen and phosphorus cycling and
plankton production. Hydrobiologia 616:133–49. doi:10.1007/s10750-008-9584-9
Preheim SP, Boucher Y, Wildschutte H, David LA, Veneziano D,Alm EJ, Polz MF 2011. Metapopulation structure ofVibrionaceae among coastal marine invertebrates.Environmental Microbiology 13:265–75. doi:10.1111/j.1462-2920.2010.02328.x
Pruesse E, Peplies J, Glöckner FO. 2012. SINA: accurate high-throughput multiple sequence alignment of ribosomalRNA genes. Bioinformatics 28:1823–29. doi:10.1093/bioinformatics/bts252
Purcell JE, Nemazie DA. 1992. Quantitative feeding ecology ofthe hydromedusan Nemopsis-bachei in Chesapeake Bay.Marine Biology 113:305–11.
Purcell JE, Breitburg DL, Decker MB, GrahamWM, YoungbluthMJ, Raskoff KA. 2001. Pelagic cnidarians and ctenophoresin low dissolved oxygen environments: a review. In:Rabalais NN, editor. Coastal Hypoxia: Consequences forLiving Resources and Ecosystems. Coastal and EstuarineSciences, Volume 58. Washington, DC: AmericanGeophysical Union, p 77–100.
Purcell JE, Uye S, Lo WT. 2007. Anthropogenic causes of jelly-fish blooms and their direct consequences for humans: areview. Marine Ecology Progress Series 350:153–74.doi:10.3354/meps07093
Quast C, Pruesse E, Yilmaz P, Gerken J, Schweer T, Yarza P,et al. 2013. The SILVA ribosomal RNA gene databaseproject: improved data processing and web-based tools.Nucleic Acids Research 41:D590–96. doi:10.1093/nar/gks1219
Richardson AJ, Bakun A, Hays GC, Gibbons MJ. 2009. The jelly-fish joyride: causes, consequences and managementresponses to a more gelatinous future. Trends in Ecology& Evolution 24:312–22. doi:10.1016/j.tree.2009.01.010
Rohwer F, SeguritanV, AzamF, KnowltonN. 2002.Diversity anddistribution of coral-associated bacteria. Marine EcologyProgress Series 243:1–10. doi:10.3354/meps243001
Rottem S. 2003. Interaction of mycoplasmas with host cells.Physiological Reviews 83:417–32. doi:10.1152/physrev.00030.2002
Schmitt S, Tsai P, Bell J, Fromont J, IlanM, LindquistN, et al. 2012.Assessing the complex sponge microbiota: core, variableand species-specific bacterial communities in marinesponges. ISME Journal 6:564–76. doi:10.1038/ismej.2011.116
Shoemaker KM, Moisander PH. 2015. Microbial diversityassociated with copepods in the North Atlantic subtropicalgyre. FEMS Microbiology Ecology 91(7):fiv064. 11 pages.doi:10.1093/femsec/fiv064
Sochard MR, Wilson DF, Austin B, Colwell RR. 1979. Bacteriaassociated with the surface and gut of marine copepods.Applied and Environmental Microbiology 37:750–59.
Suzuki M, Nakagawa Y, Harayama S, Yamamoto S. 2001.Phylogenetic analysis and taxonomic study of marineCytophaga-like bacteria: proposal for Tenacibaculum gen.nov. with Tenacibaculum maritimum comb. nov. andTenacibaculum ovolyticum comb. nov., and description ofTenacibaculum mesophilum sp. nov. and Tenacibaculumamylolyticum sp. nov. International Journal of Systematicand Evolutionary Microbiology 51:1639–52. doi:10.1099/00207713-51-5-1639
Tang K, Dziallas C, Hutalle-Schmelzer K, Grossart H-P. 2009.Effects of food on bacterial community composition
12 M. C. DALEY ET AL.

associated with the copepod Acartia tonsa Dana. BiologyLetters 5:549–53. doi:10.1098/rsbl.2009.0076
Tang KW, Turk V, Grossart HP. 2010. Linkage between crus-tacean zooplankton and aquatic bacteria. AquaticMicrobial Ecology 61:261–77. doi:10.3354/ame01424
Taylor MW, Radax R, Steger D, Wagner M. 2007. Sponge-associated microorganisms: evolution, ecology, and bio-technological potential. Microbiology and MolecularBiology Reviews 71:295–347. doi:10.1128/MMBR.00040-06
Tinta T, Kogovšek T, Malej A, Turk V. 2012. Jellyfish modulatebacterial dynamic and community structure. PLoS One 7(6):e39274. 11 pages. doi:10.1371/journal.pone.0039274
Treusch AH, Vergin KL, Finlay LA, Donatz MG, Burton RM,Carlson CA, Giovannoni SJ. 2009. Seasonality and verticalstructure of microbial communities in an ocean gyre.ISME Journal 3:1148–63. doi:10.1038/ismej.2009.60
Turner S, Pryer KM, Miao VPW, Palmer JD. 1999. Investigatingdeep phylogenetic relationships among cyanobacteria andplastids by small submit rRNA sequence analysis. Journal ofEukaryotic Microbiology 46:327–38. doi:10.1111/j.1550-7408.1999.tb04612.x
Urbanczyk H, Ast JC, Dunlap PV. 2011. Phylogeny, genomics,and symbiosis of Photobacterium. FEMS MicrobiologyReviews 35:324–42. doi:10.1111/j.1574-6976.2010.00250.x
Uye S. 2011. Human forcing of the copepod-fish-jellyfish tri-angular trophic relationship. Hydrobiologia 666:71–83.doi:10.1007/s10750-010-0208-9
Walker DH. 1996. Rickettsiae. In: Baron S, editor. MedicalMicrobiology. Galveston, TX: University of Texas MedicalBranch at Galveston, p 487–501.
Weiland-Bräuer N, Neulinger SC, Pinnow N, Künzel S, BainesJF, Schmitz RA. 2015. Composition of bacterial commu-nities associated with Aurelia aurita changes with compart-ment, life stage and population. Applied andEnvironmental Microbiology 81:6038–52. doi:10.1128/AEM.01601-15
White JR, Patel J, Ottesen A, Arce G, Blackwelder P, Lopez JV.2012. Pyrosequencing of bacterial symbionts withinAxinella corrugata sponges: diversity and seasonal variabil-ity. PLoS One 7:e38204. 12 pages. doi:10.1371/journal.pone.0038204
Williams TJ, Wilkins D, Long E, Evans F, DeMaere MZ, RafteryMJ, Cavicchioli R. 2013. The role of planktonic Flavobacteriain processing algal organic matter in coastal EastAntarctica revealed using metagenomics and metaproteo-mics. Environmental Microbiology 15:1302–17. doi:10.1111/1462-2920.12017
Wong HC, Liu CC, Pan TM, Wang TK, Lee CL, Shih YC. 1999.Molecular typing of Vibrio parahaemolyticus isolates,obtained from patients involved in food poisoning out-breaks in Taiwan, by random amplified polymorphic DNAanalysis. Journal of Clinical Microbiology 37:1809–12.
Wright ES, Yilmaz LS, Noguera DR. 2012. DECIPHER, a search-based approach to chimera identification for 16S rRNAsequences. Applied and Environmental Microbiology78:717–25. doi:10.1128/AEM.06516-11
Zimmer RL, Woollacott RM. 1983. Mycoplasma-like organisms:occurrence with the larvae and adults of a marinebryozoan. Science 220:208–10. doi:10.1126/science.220.4593.208
MARINE BIOLOGY RESEARCH 13