balancing the dumbbell: summer habitats need protection in addition to winter dens for northern...
TRANSCRIPT

Habitat Relations
Balancing the Dumbbell: Summer HabitatsNeed Protection in Addition to Winter Densfor Northern Snake Communities
LAURA E. GARDINER, Biology Department, University of Regina, 3737 Wascana Parkway, Regina, SK, Canada S4S 0A2
CHRISTOPHER M. SOMERS, Biology Department, University of Regina, 3737 Wascana Parkway, Regina, SK, Canada S4S 0A2
JESSICA A. MARTINO, Biology Department, University of Regina, 3737 Wascana Parkway, Regina, SK, Canada S4S 0A2
DENNILYN L. PARKER, Western College of Veterinary Medicine, University of Saskatchewan, Saskatoon, SK, Canada S7N 5B4
RAY G. POULIN,1 Royal Saskatchewan Museum, 2340 Albert Street, Regina, SK, Canada S4P 2V7
ABSTRACT The grasslands of southwestern Saskatchewan, Canada are home to several snake species ofconservation concern at the northern extreme of their geographic range. To aid conservation assessment andmanagement planning for these snakes, we used radio-telemetry, a geographic information system, andmultivariate modeling to identify and compare macrohabitat selection by eastern yellow-bellied racers(n ¼ 33; Coluber constrictor flaviventris), bullsnakes (n ¼ 16; Pituophis catenifer sayi), and prairie rattlesnakes(n ¼ 23; Crotalus viridis). All 3 species shared communal hibernacula in the inactive winter season, butdispersed into different macrohabitats across the landscape during the spring and summer. Their homeranges varied in size by species and were dumbbell-shaped with activity centers near hibernacula and in well-defined summer grounds; activity centers were connected by narrow movement corridors. Racers stronglyselected for riparian areas, bullsnakes selected for valley grassland habitats, and rattlesnakes selected for areasassociated with prairie dog colonies. Some rattlesnakes traveled great distances (over 11 km) from the denscompared to the other species (bullsnake max. ¼ 4 km; racer max. ¼ 5 km), which may be a result of theirselected macrohabitat being more patchily distributed in the landscape. Our results indicate thatmanagement plans for these snakes must consider the den area, corridors, and separate summering grounds,as well as differences in home range size and movement patterns for each species. � 2013 The WildlifeSociety.
KEYWORDS bullsnake, Coluber constrictor flaviventris, Crotalus viridis, eastern yellow-bellied racer, grassland snakes,habitat partitioning, Pituophis catenifer sayi, prairie rattlesnake.
Multi-species studies provide insight into habitat partition-ing and may have important conservation implications.Studies on habitat selection of 1 species reveal little abouthow their selection differs from, and interacts with, otherspecies in the community. Differences in habitat selectionbetween similar species reduce interspecific competitionallowing ecological equivalents to coexist stably (Schoener1974), and these differences should be accounted for inconservation plans. Studies on the community ecology ofsnakes are rare in general, though some notable exceptionsexist (Hirth et al. 1966, Fitch and Shirer 1971, Keller andHeske 2000). Data on habitat partitioning in snakes areespecially scarce and lag behind that of other taxonomicgroups such as lizards, birds, and small mammals(Schoener 1974). Keller and Heske (2000) documentedhabitat partitioning in a more southern snake community inIllinois, USA, but no previous studies, to our knowledge,
have documented habitat partitioning in peripheral pop-ulations. Northern snake communities are of particularinterest because many species hibernate communally in alimited number of dens, and therefore share winter habitatfollowed by dispersal into summer habitat (Hirth et al. 1966,Fitch and Shirer 1971, Brown and Parker 1976, Prior andWeatherhead 1996, Keller and Heske 2000). Conservationplans for hibernating snake communities usually includebuffers around known den locations, but these are likelynot large enough to encompass summer habitats (Martinoet al. 2012, Williams et al. 2012). Habitat partitioning ismost likely to occur during the active season when speciesmay compete for resources such as food or nesting sites(Keller and Heske 2000), and these habitat differences willneed to be included in conservation plans by identifyingspecies-specific summer buffer zones in addition to denprotection.Southwestern Saskatchewan contains a unique (for
Canada) snake community where eastern yellow-belliedracers (Coluber constrictor flaviventris), prairie rattlesnakes(Crotalus viridis), bullsnakes (Pituophis catenifer sayi), westernhognose (Heterodon nasicus), and plains (Thamnophis radix)
Received: 4 November 2012; Accepted: 12 February 2013Published: 14 June 2013
1E-mail: [email protected]
The Journal of Wildlife Management 77(5):975–982; 2013; DOI: 10.1002/jwmg.555
Gardiner et al. � Summer Habitat Use by Snakes 975

and terrestrial (T. elegans) garter snakes coexist. Easternyellow-bellied racers (hereafter racers) are a Threatenedspecies in Canada (Committee on the Status of EndangeredWildlife in Canada [COSEWIC] 2004), with their rangeconfined to a few river valleys in southern Saskatchewan andsoutheastern Alberta (Gardiner et al. 2011). Racers meet thecriteria for an Endangered species in Canada, but arecurrently listed as Threatened because the majority of theirknown range is protected by Grasslands National Park(COSEWIC 2004). Bullsnakes are listed as Data Deficient,because sufficient information does not exist for a properconservation status assessment (COSEWIC 2002). Prairierattlesnakes (hereafter rattlesnakes) have been listed forassessment, and therefore updated information on theecology of this species is in immediate demand (www.cosewic.gc.ca, accessed 27 Oct 2012). These 3 snake speciesare vulnerable to extinction in Canada because of habitat lossand negative interactions with humans; they have specificrequirements for suitable hibernacula, which are limiting inthis portion of their range because of extreme wintertemperatures (Watson and Russell 1997, Ernst andErnst 2003, COSEWIC 2004, Kapfer et al. 2008, Gardinerand Sonmor 2011). Although essential for effectiveconservation planning, little information exists on the spatialecology and habitat selection of these species at northernrange limits (Kapfer et al. 2008, Martino et al. 2012).We compared the movement patterns and macrohabitat
selection of 3 sympatric snake species (racers, bullsnakes, andrattlesnakes) during the active season at the northern extremeof their ranges in the mixed-grass prairie ecoregion of southernCanada. Martino et al. (2012) found that racers and bullsnakestravel up to 10 times farther from hibernacula and occupymuch larger home ranges (3–104 times larger) than theirsouthern counterparts. They proposed that this phenomenonmay be because they use distinct winter and summer habitatslinked by corridors, creating a dumbbell-shaped home range.However, the basic characterization of home ranges usingminimum convex polygon (MCP) methods by Martino et al.(2012) did not provide specific information on home rangeshape or the relative intensity of habitat use in different areas.Correspondingly, the primary objective of our study was todetermine whether home ranges for racers, bullsnakes, andrattlesnakes are actually dumbbell-shaped based on separatewinter and summer activity centers. As part of this process,we also characterized macrohabitat selection and potential nicepartitioning among the 3 snake species.
STUDY AREA
We conducted our research within the Frenchman RiverValley around the west block of Grasslands National Park(4981003700 10782503300W), and a nearby Agri-EnvironmentServices Branch (AESB, formerly PFRA) communitypasture in Saskatchewan, Canada from May to Septemberof 2010 and 2011 (Fig. 1); data included from Martino et al.(2012), however, were collected in 2008 and 2009. Our studyarea is described in detail inMartino et al. (2012). In additionto this description, unstable slump zones along the slopes ofthe river valley are a common location for winter hibernation
sites for snakes in the area (Gardiner and Sonmor 2011).Landslides, or slumps, along unstable slopes create terracescontaining slight depressions or sinkholes, fissures, or smallfaults, and loosely packed soil that attracts burrowingmammals. These factors provide opportunities for snakes tofind refuge below the frost line. Snakes are dependent onsuitable winter hibernacula for persistence at higherlatitudes, and more than 1 species are commonly foundhibernating together at communal hibernation sites; this islikely a response to limited availability of suitable locationsfor overwintering. Hibernacula contain a combination ofracers, bullsnakes, and rattlesnakes, in addition to plainsgarter snakes. The number of snakes varies by site with somesites containing hundreds of individuals (e.g., Gardiner et al.2011), which may indicate hibernacula vary in quality. Wecollected snakes from 3 known dens within GrasslandsNational Park, and 2 known dens within the AESBcommunity pasture.
METHODS
Study SpeciesThe racer is an elusive, non-venomous colubrid with anextremely limited known range on the Canadian prairies(Gardiner et al. 2011). Closely related subspecies of racers to
Figure 1. Ranges of the racer (Coluber constrictor), bullsnake (Pituophiscatenifer), and prairie rattlesnake (Crotalus viridis) in North America, and theregion where their ranges overlap. The location of our study area (2008–2011) in Saskatchewan, Canada, is indicated by the arrow.
976 The Journal of Wildlife Management � 77(5)

those found in Canada have been studied extensively in theUSA (Hirth et al. 1966; Fitch and Shirer 1971; Brown andParker 1976; Klug et al. 2011a, b), but only 1 study exists inCanada (Martino et al. 2012). The range of racers in Canadatends to coincide with river valleys, which is likely because oftheir reliance on hibernacula as well as their selection forriparian areas during the summer season (Martino et al.2012). Bullsnakes are also non-venomous colubrids with alimited known range on the Canadian prairies. Littleinformation is available on the habitat requirements of thissubspecies (Kapfer et al. 2008, Martino et al. 2012), thoughthey are known to select for valley grassland habitats in ourstudy area and remain within closer proximity to the den thanthe racer (Martino et al. 2012). Racers and bullsnakes aredescribed in greater detail in Martino et al. (2012). Prairierattlesnakes are a wide-ranging pit viper capable of reachingover 1.5 m in length with a diet mainly consisting of smallmammals (Hill et al. 2001, Ernst and Ernst 2003). Fewpublished studies exist on the ecology of the prairierattlesnakes at the northern extreme of their range (Gannonand Secoy 1984, 1985; Charland and Gregory 1990), noneof which focused on their spatial ecology. All 3 of thesespecies have extensive ranges throughout the USA, thoughthey are limited in their Canadian range to southernSaskatchewan and Alberta (Pendlebury 1977, Ernst andErnst 2003, Gardiner et al. 2011; Fig. 1).
Snake Capture and TrackingThe active season for snakes in Saskatchewan extends fromMay until October. We caught snakes at 5 different densduring emergence from hibernation using a combination ofdrift fences with snake traps, opportunistic visual encounters,and road surveys. We sexed snakes via cloacal probing(Schaefer 1934) and surgically implanted them with HolohilSystems Ltd. radio-transmitters (model SB-2, mass ¼ 4–5 g; Holohil Systems Ltd., Carp, Ontario, Canada) based onpreviously established methods (Reinert and Cundall 1982).We implanted 14 racers and 23 rattlesnakes, which enabledus to add rattlesnakes to the Martino et al. (2012) multi-species dataset and increase the sample size of racers to 33(from n ¼ 19). We only implanted snakes when transmittermass represented less than or equal to 5% of body mass andthe diameter of the transmitter was less than 50% of thediameter of the snake at the insertion site.We allowed snakesto recover for 48 hours before releasing them at their site ofcapture.Using radio-telemetry, we tracked individual snakes from
their emergence from hibernacula in May until their return
to hibernacula in October. We attempted to relocate allsnakes every 48 hours, but the number of relocations andduration of tracking varied by individual and species(Table 1). We did not directly approach the snakes to avoidinfluencing their movements; we either visually confirmedthe snake site from a distance, or precisely triangulated thesnake’s location from a short distance. We recordedUniversal Transverse Mercator (UTM) coordinates when-ever we located a snake and marked the site. All methodswere approved by the President’s Committee on AnimalCare at the University of Regina.
Movement and Home RangeWe quantified movement patterns using ArcGISDesktop 10(Environmental Systems Research Institute, Inc., Redlands,CA). We determined maximum distances moved with thepoint distance tool. We determined daily movement ratesaccording to previously published methods (Charland andGregory 1995). We first converted snake location points intolines using the points to lines tool. We calculated the totaldistance (i.e., length) of each line and divided the sum by thenumber of days we tracked each individual.We calculated 95% fixed kernels (ha) to estimate home
range size of individuals using the Hawths Tools extensionfor ArcMap 9.2 (Beyer 2004). We only included individualstracked over a minimum of 50 days with at least 15relocations in this analysis. We used the average dailymovement rate calculated for each species as the smoothingfactor to estimate kernel home ranges, with the assumptionthat larger than average movements reflect movementsbetween activity centers (the dispersal kernel; Kot et al.1996). We assigned a weighted value to points representingsnake locations based on the amount of time spent at eachlocation, determined by how many times a snake wasrelocated in the same spot versus a new location. Weconsidered locations to be new when an individual movedgreater than 20 m from the previous location (as in Martinoet al. 2012). We also calculated 100% MCPs (ha) usingArcMap 10 to enable comparisons with previous studies. Weonly included individuals with at least 10 new locations inthis analysis (Himes et al. 2006, Kapfer et al. 2008, Martinoet al. 2012). We re-calculated mean home range size usingthe MCP method for racers from Martino et al. (2012) withthe increased sample size. We analyzed all movement andhome range comparisons between species with a 1-wayanalysis of variance (ANOVA) and Tukey–Kramer post hoctests when data met the assumptions of normality andhomoscedasticity. When data did not meet the assumptions
Table 1. Mean tracking duration (days), number of relocations, daily movement rate (DMR; m/day), and maximum distance moved from the den (max.distance; m) by sympatric eastern yellow-bellied racers (EYBR), prairie rattlesnakes (PRS), and bullsnakes (BS) in Saskatchewan, Canada, 2008–2011.
Species
Tracking duration (days) No. of relocations Movement parameters (mean � SD)
Mean � SD (range) Mean � SD (range) DMR (m/day) Max. distance (m)
EYBR (n ¼ 33) 93 � 45.4 (6–151) 22 � 13.1 (2–51) 65.3 � 5.0 2,463.7 � 1,282.4PRS (n ¼ 23) 70 � 47.5 (13–151) 17 � 10.0 (4–32) 92.0 � 16.2 2,812.5 � 2,819.9BS (n ¼ 16) 84 � 30.2 (30–114) 25 � 13.8 (8–50) 52.4 � 7.0a 1,547.6 � 990.6a
a Data are from Martino et al. (2012).
Gardiner et al. � Summer Habitat Use by Snakes 977
required for parametric tests, we used Kruskal–Wallis andDunn’s post hoc tests. We used Mann–Whitney U tests tocompare home range size between sexes for each species. Weperformed statistical tests using GraphPad Prism 5 with analpha value of 0.05.
Macrohabitat SelectionWe defined macrohabitat as the large-scale environmentincluding broad landscape features that are readily observ-able, such as vegetation and topography (Johnson 1980).Macrohabitats available to the snakes included native upland,lowland pasture, hills and slopes, mudflats, crop areas, prairiedog colonies, irrigation areas, open water, roads, and riparianareas. These macrohabitats are easily differentiated fromeach other in the mixed-grass prairie ecosystem in southernSaskatchewan, and have been used in other grassland snakestudies (e.g., Martino et al. 2012). We determined themacrohabitat used by radio-tracked snakes at each locationby visual observation of the surrounding area, which we laterconfirmed by overlaying the snake locations on satelliteimagery using ArcGIS Desktop 10. We quantified macro-habitat selection by determining the proportion of time spentin each macrohabitat type in comparison to the proportion ofthat macrohabitat available (as in Martino et al. 2012) in abuffer zone around the hibernaculum.We used 5-km buffersaround hibernacula to determine macrohabitats available toracers and bullsnakes (Martino et al. 2012), and 12-kmbuffers for rattlesnakes. We determined buffer size based onthe maximum distances moved (from their den) by anyindividual from each species. We traced macrohabitatswithin the buffers in ArcMap over a 2.5-m resolution SPOTimage (satellite photo; www.flysask.ca, accessed 18 Oct2011). We used the union tool to calculate the area of eachmacrohabitat polygon, and then determined the percentageof each polygon within the buffer. We performed composi-tional analysis to statistically analyze macrohabitat selection(R package, adehabitatHS; R statistics v2.14.2, The RFoundation, Vienna, Austria), because it is well suited tolandscape-level analyses (Aebischer et al. 1993, Marshallet al. 2006). Compositional analysis statistically compareshabitat use by individuals to the proportions of all availablehabitats simultaneously rather than univariately (usingWilkslambda; Aebischer et al. 1993). When macrohabitat use isnon-random (P < 0.05), habitats are ranked in order of use
and analyzed using 2-tailed t-tests to determine whetherrankings differ significantly from each other (P-valuesBonferroni corrected).
RESULTS
Movement and Home RangeDaily movement rate varied among the 3 study species(Table 1; ANOVA, F2, 69 ¼ 3.61, P ¼ 0.032). Post hoctesting revealed that rattlesnakes moved significantly farther(roughly 2 times farther) than bullsnakes on a daily basis,and that racers were intermediate. The maximum distanceracers traveled from their den ranged from 187 m to4,994 m, whereas rattlesnakes ranged from 534 m to11,107 m. Both racers and rattlesnakes appeared to travelfarther from hibernacula than bullsnakes, but these differ-ences were not statistically significant (Table 1; F2, 67 ¼ 2.25,P ¼ 0.113).Regardless of method, racers and rattlesnakes had larger
home range sizes on average than bullsnakes (Table 2).Kernel density analysis revealed racers and rattlesnakes havesimilar home range sizes that are 3 times larger than those ofbullsnakes (F2, 43 ¼ 22.86, P � 0.001). Using the MCPmethod, however, home range size did not differ significantlybetween the 3 species because of the large amount ofvariation (Kruskal–Wallis, H2 ¼ 5.31, P ¼ 0.070). Homerange sizes did not differ by sex usingMann–WhitneyU tests(Table 2). Home range size also did not vary significantlywith total snake length or number of relocations for anyspecies (data not shown). Home ranges were dumbbellshaped for all 3 species with 2 activity centers connected by amovement corridor (Fig. 2). Concentrations in habitat useexisted around the overwintering (i.e., den) sites and inseparate summering grounds. Snakes typically movedbetween winter and summer habitats quickly over the courseof several days, although we found some inter-individualvariation, particularly among rattlesnakes (Table 3). Summeractivity centers were varying distances fromwintering kernelsbased on the maximum distances moved by individuals.Given this structure to home ranges, size estimates usingMCP methods are not as accurate or potentially useful asthose derived from kernel density analysis.
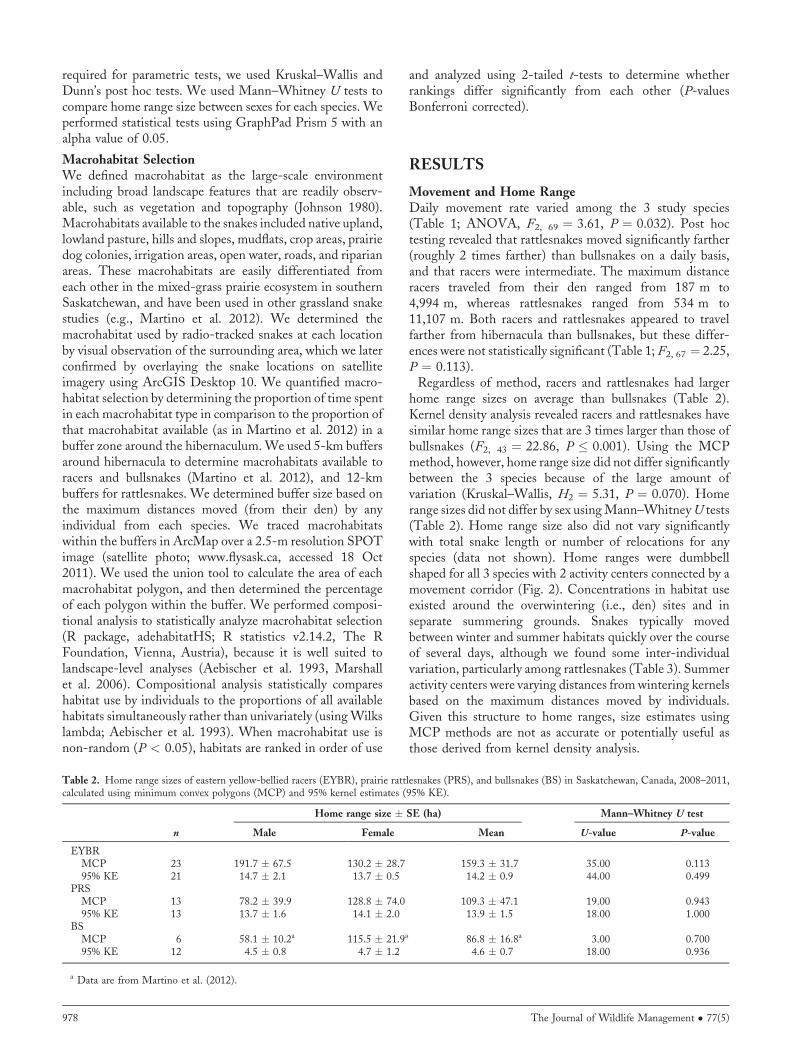
Table 2. Home range sizes of eastern yellow-bellied racers (EYBR), prairie rattlesnakes (PRS), and bullsnakes (BS) in Saskatchewan, Canada, 2008–2011,calculated using minimum convex polygons (MCP) and 95% kernel estimates (95% KE).
n
Home range size � SE (ha) Mann–Whitney U test
Male Female Mean U-value P-value
EYBRMCP 23 191.7 � 67.5 130.2 � 28.7 159.3 � 31.7 35.00 0.11395% KE 21 14.7 � 2.1 13.7 � 0.5 14.2 � 0.9 44.00 0.499
PRSMCP 13 78.2 � 39.9 128.8 � 74.0 109.3 � 47.1 19.00 0.94395% KE 13 13.7 � 1.6 14.1 � 2.0 13.9 � 1.5 18.00 1.000
BSMCP 6 58.1 � 10.2a 115.5 � 21.9a 86.8 � 16.8a 3.00 0.70095% KE 12 4.5 � 0.8 4.7 � 1.2 4.6 � 0.7 18.00 0.936
a Data are from Martino et al. (2012).
978 The Journal of Wildlife Management � 77(5)

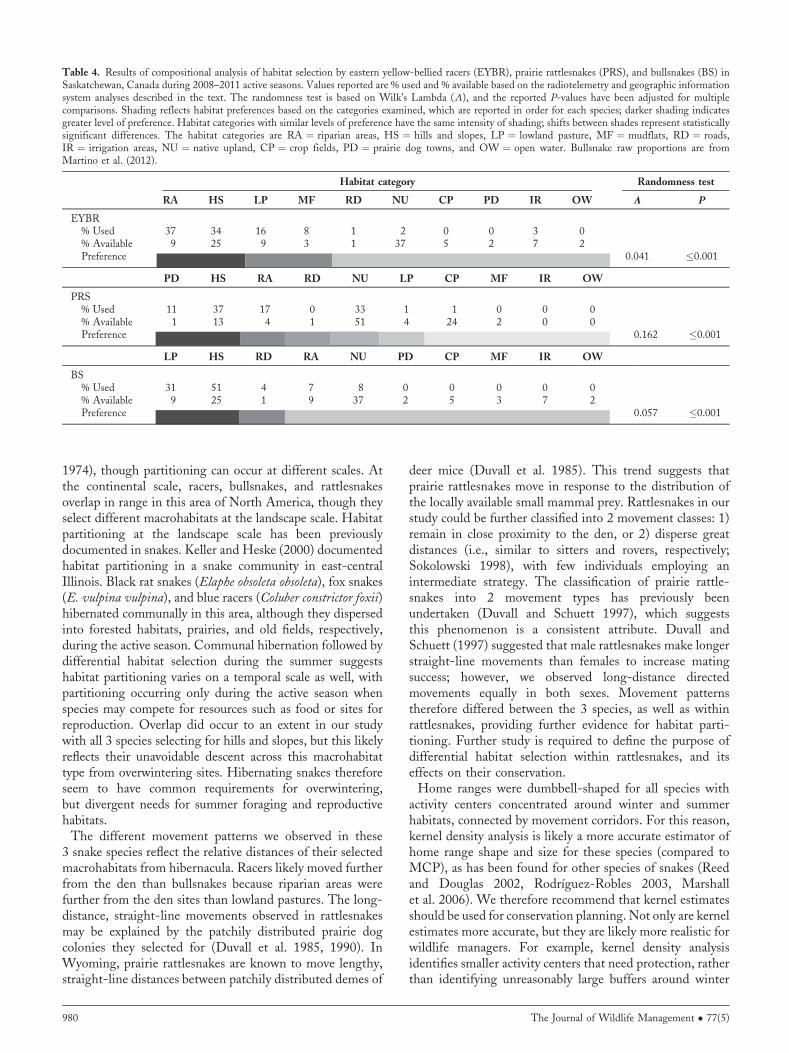
Macrohabitat SelectionRacers, bullsnakes, and rattlesnakes exhibited habitatselection at the landscape scale (Table 4). Compositionalanalysis revealed that racers selected for riparian areas, hillsand slopes, lowland pastures, and mudflats, but riparian areas(t28 ¼ 2.736, P ¼ 0.010) and hills and slopes (t28 ¼ 2.764,P ¼ 0.009) were used more than lowland pastures andmudflats. Rattlesnakes selected for prairie dog colonies, hillsand slopes, and riparian areas, though prairie dog colonies(t22 ¼ 3.450, P ¼ 0.002) and hills and slopes (t22 ¼ 3.628,P ¼ 0.001) were used more than riparian areas. Roads wereused according to their availability in the environment byracers and rattlesnakes. In contrast, compositional analysisrevealed bullsnakes (data fromMartino et al. 2012) select for
lowland pasture, hills and slopes, and roads, though lowlandpasture (t15 ¼ 3.434, P ¼ 0.003) and hills and slopes(t15 ¼ 4.830, P � 0.001) were used more than roads. All3 species avoided native upland, crop areas, irrigated areas,and open water, which were used significantly less than theother available landscape types (Table 4).
DISCUSSION
Habitat partitioning is occurring with little overlap betweenracers, bullsnakes, and rattlesnakes during the active season,despite their communal occurrence at winter hibernacula.Habitat partitioning reduces competition for resourcesallowing the existence of sympatric species (Schoener
Figure 2. Kernel density maps of home range shape of representative individuals of sympatric bullsnakes (a), eastern yellow-bellied racers (b), and prairierattlesnakes (c) in Saskatchewan, Canada, 2008–2011. Home range maps are based on both location and duration data from radio-telemetry; shaded areaswithin home ranges reflecting a greater probability of use are based on relocations of snakes at the same location (points) for many days or weeks.
Table 3. Duration (days) and number of relocations per species during movement between winter hibernacula and summer activity areas by eastern yellow-bellied racers (EYBR), prairie rattlesnakes (PRS), and bullsnakes (BS) in Saskatchewan, Canada, 2008–2011. Locations (points) were identified as part ofthis movement phase if they were between, and not within, winter and summer kernels.
Species
Duration (days) No. of relocations
Mean � SD Range Mean � SD Range
EYBR (n ¼ 21) 8.9 � 4.6 2–18 2.3 � 1.1 0–4PRS (n ¼ 13)a 18.1 � 17.9 2–60 3.8 � 3.5 0–11BS (n ¼ 12) 4.9 � 3.0 2–10 0.1 � 0.4 0–1
a Includes 2 individual rattlesnakes that traveled large distances (over 11 km) from winter hibernacula. Excluding these 2 outliers, meanduration � SD ¼ 12.3 � 11.2 (range: 2–38) and mean number of relocations � SD ¼ 3.4 � 3.5 (range: 0–11).
Gardiner et al. � Summer Habitat Use by Snakes 979
1974), though partitioning can occur at different scales. Atthe continental scale, racers, bullsnakes, and rattlesnakesoverlap in range in this area of North America, though theyselect different macrohabitats at the landscape scale. Habitatpartitioning at the landscape scale has been previouslydocumented in snakes. Keller and Heske (2000) documentedhabitat partitioning in a snake community in east-centralIllinois. Black rat snakes (Elaphe obsoleta obsoleta), fox snakes(E. vulpina vulpina), and blue racers (Coluber constrictor foxii)hibernated communally in this area, although they dispersedinto forested habitats, prairies, and old fields, respectively,during the active season. Communal hibernation followed bydifferential habitat selection during the summer suggestshabitat partitioning varies on a temporal scale as well, withpartitioning occurring only during the active season whenspecies may compete for resources such as food or sites forreproduction. Overlap did occur to an extent in our studywith all 3 species selecting for hills and slopes, but this likelyreflects their unavoidable descent across this macrohabitattype from overwintering sites. Hibernating snakes thereforeseem to have common requirements for overwintering,but divergent needs for summer foraging and reproductivehabitats.The different movement patterns we observed in these
3 snake species reflect the relative distances of their selectedmacrohabitats from hibernacula. Racers likely moved furtherfrom the den than bullsnakes because riparian areas werefurther from the den sites than lowland pastures. The long-distance, straight-line movements observed in rattlesnakesmay be explained by the patchily distributed prairie dogcolonies they selected for (Duvall et al. 1985, 1990). InWyoming, prairie rattlesnakes are known to move lengthy,straight-line distances between patchily distributed demes of
deer mice (Duvall et al. 1985). This trend suggests thatprairie rattlesnakes move in response to the distribution ofthe locally available small mammal prey. Rattlesnakes in ourstudy could be further classified into 2 movement classes: 1)remain in close proximity to the den, or 2) disperse greatdistances (i.e., similar to sitters and rovers, respectively;Sokolowski 1998), with few individuals employing anintermediate strategy. The classification of prairie rattle-snakes into 2 movement types has previously beenundertaken (Duvall and Schuett 1997), which suggeststhis phenomenon is a consistent attribute. Duvall andSchuett (1997) suggested that male rattlesnakes make longerstraight-line movements than females to increase matingsuccess; however, we observed long-distance directedmovements equally in both sexes. Movement patternstherefore differed between the 3 species, as well as withinrattlesnakes, providing further evidence for habitat parti-tioning. Further study is required to define the purpose ofdifferential habitat selection within rattlesnakes, and itseffects on their conservation.Home ranges were dumbbell-shaped for all species with
activity centers concentrated around winter and summerhabitats, connected by movement corridors. For this reason,kernel density analysis is likely a more accurate estimator ofhome range shape and size for these species (compared toMCP), as has been found for other species of snakes (Reedand Douglas 2002, Rodrı́guez-Robles 2003, Marshallet al. 2006). We therefore recommend that kernel estimatesshould be used for conservation planning. Not only are kernelestimates more accurate, but they are likely more realistic forwildlife managers. For example, kernel density analysisidentifies smaller activity centers that need protection, ratherthan identifying unreasonably large buffers around winter
Table 4. Results of compositional analysis of habitat selection by eastern yellow-bellied racers (EYBR), prairie rattlesnakes (PRS), and bullsnakes (BS) inSaskatchewan, Canada during 2008–2011 active seasons. Values reported are % used and % available based on the radiotelemetry and geographic informationsystem analyses described in the text. The randomness test is based on Wilk’s Lambda (L), and the reported P-values have been adjusted for multiplecomparisons. Shading reflects habitat preferences based on the categories examined, which are reported in order for each species; darker shading indicatesgreater level of preference. Habitat categories with similar levels of preference have the same intensity of shading; shifts between shades represent statisticallysignificant differences. The habitat categories are RA ¼ riparian areas, HS ¼ hills and slopes, LP ¼ lowland pasture, MF ¼ mudflats, RD ¼ roads,IR ¼ irrigation areas, NU ¼ native upland, CP ¼ crop fields, PD ¼ prairie dog towns, and OW ¼ open water. Bullsnake raw proportions are fromMartino et al. (2012).
Habitat category Randomness test
RA HS LP MF RD NU CP PD IR OW L P
EYBR% Used 37 34 16 8 1 2 0 0 3 0% Available 9 25 9 3 1 37 5 2 7 2Preference 0.041 �0.001
PD HS RA RD NU LP CP MF IR OW
PRS% Used 11 37 17 0 33 1 1 0 0 0% Available 1 13 4 1 51 4 24 2 0 0Preference 0.162 �0.001
LP HS RD RA NU PD CP MF IR OW
BS% Used 31 51 4 7 8 0 0 0 0 0% Available 9 25 1 9 37 2 5 3 7 2Preference 0.057 �0.001
980 The Journal of Wildlife Management � 77(5)

den sites. Snakes at the northern periphery of theirgeographic range are tied to winter hibernation sites, whichmay or may not be in close proximity to summer habitats.Therefore, the distance between winter and summergrounds, and thus the required activity centers, for snakesmay vary. Although racers did not travel nearly as far fromthe den as rattlesnakes, kernel density analysis revealed theirhome range sizes to be similar. This is because the activitycenters were similar in size, whereas the length of themovement corridor between the winter and summer groundsfor the 2 species differed. The distance between the winterand summer grounds varied more within rattlesnakes becauseof the 2 movement strategies exhibited by this species (i.e.,sitters and rovers). These distinct movement patterns werethe cause of the variation seen in the average daily movementrate, maximum distance moved from the den, and MCPhome range size for rattlesnakes as well. In contrast,bullsnakes had concentrated activity centers closer to the denrelative to the other species; again, likely reflecting theirselection of a macrohabitat within close proximity to the densites. All 3 species therefore vary in the length of theirmovement corridors, further suggesting they select distinctsummer habitats.
MANAGEMENT IMPLICATIONS
Habitat differences between species during the active seasonshould be considered when making management decisionsregarding snake populations that hibernate in communaldens. Though suitable overwintering sites are imperative tothe persistence of northern snake populations, small reservesaround communal hibernacula have been identified asinsufficient to protect snakes during the active season(Martino et al. 2012, Williams et al. 2012); our results showthat identifying summer grounds will likely require species-specific considerations. Martino et al. (2012) suggested thatsimply creating management zones around dens alone wouldlikely not be sufficient to include summer grounds fornorthern snake populations, and they further suggested thatthis can be mitigated by either defining much larger denzones or identifying a corridor system linking smallerprotected areas surrounding winter and summer habitats.Our results show the home ranges of these northerngrassland snakes are indeed dumbbell-shaped, and manage-ment considerations must include the den area, corridors,and the summering grounds. Habitat use during the activeseason differs markedly by snake species, so conservationstrategies must incorporate these differences by identifyingspecies-specific summer habitats for protection. For example,racers strongly avoid prairie dog colonies, though these areasare heavily used by rattlesnakes. So, even though activitycenters were similar in size (14 ha) for these 2 species,activity centers will need to be identified around theirrespective summer habitats (e.g., riparian areas for racers,small mammal concentrations for rattlesnakes). Further-more, corridors connecting winter and summer grounds maynot be the shortest path between the 2. For example, if aprairie dog colony is located between the 2 activity centersfor racers, they appear to take a longer route to avoid this
habitat type. Snakes with more tortuous movement paths,such as racers, may therefore require wider movementcorridors than snakes that travel in straighter paths, such asrattlesnakes. We suggest future studies identify factorsaffecting snake movement corridors. Features of thecorridors to be measured could include topography,vegetation type and cover, physical barriers (natural andanthropogenic), and human disturbance. The results couldthen be used to identify and protect effective movementcorridors for snakes in fragmented habitats.
ACKNOWLEDGMENTS
We thank J. Piwowar and L. Heisler for their statisticalguidance, and the vets and field assistants without whom thisresearch would not have been possible. Funding wasprovided for this study by Saskatchewan EnvironmentFish and Wildlife Development Fund, EnvironmentCanada’s Interdepartmental Recovery Fund, the CanadianMuseum of Nature, the Friends of the Royal SaskatchewanMuseum, Parks Canada Agency—Grasslands NationalPark, Agriculture and Agri-Food Canada-PFRA, and theCanada Research Chairs Program.
LITERATURE CITEDAebischer, N. J., P. A. Robertson, and R. E. Kenward. 1993. Compositionalanalysis of habitat use from animal radiotracking data. Ecology 74:1313–1325.
Beyer, H. L. 2004. Hawth’s analysis tools for ArcGIS. <http://www.spatialecology.com/htools>. Accessed 18 Nov 2011.
Brown, W. S., and W. S. Parker. 1976. Movement ecology of Coluberconstrictor near communal hibernacula. Copeia 1976:225–242.
Charland, M. B., and P. T. Gregory. 1990. The influence of femalereproductive status on thermoregulation in a viviparous snake, Crotalusviridis. Copeia 1990:1089–1098.
Charland, M. B., and P. T. Gregory. 1995. Movements and habitat use ingravid and nongravid female garter snakes (Colubridae, Thamnophis).Journal of Zoology (London) 236:543–561.
Committee On the Status of Endangered Wildlife In Canada[COSEWIC]. 2002. COSEWIC assessment and status report onthe Gophersnake Pituophis catenifer in Canada. COSEWIC, Ottawa,Canada.
Committee On the Status of Endangered Wildlife In Canada[COSEWIC]. 2004. COSEWIC assessment and update status reporton the eastern and western yellow-bellied racers, Coluber constrictorflaviventris and Coluber constrictor mormon in Canada. COSEWIC,Ottawa, Canada.
Duvall, D., and G. W. Schuett. 1997. Straight-line movement andcompetitive mate searching in prairie rattlesnakes, Crotalus viridis viridis.Animal Behaviour 54:329–334.
Duvall, D., M. B. King, and K. J. Gutzwiller. 1985. Behavioral ecologyand ethology of the prairie rattlesnake. National Geographic Research1:80–111.
Duvall, D., M. J. Goode, W. K. Hayes, J. K. Leonhardt, and D. Brown.1990. Prairie rattlesnake vernal migrations: field experimental analyses andsurvival value. National Geographic Research 6:457–469.
Ernst, C. H., and E. M. Ernst. 2003. Snakes of the United States andCanada. Smithsonian Books, Washington, D.C.
Fitch, H. S., and H. W. Shirer. 1971. A radiotelemetric study of spatialrelationships in some common snakes. Copeia 1971:118–128.
Gannon, V. P. J., andD.M. Secoy. 1984. Growth and reproductive rates of anorthern population of the prairie rattlesnake,Crotalus v. viridis. Journal ofHerpetology 18:13–19.
Gannon, V. P. J., andD.M. Secoy. 1985. Seasonal and daily activity patternsin a Canadian population of the prairie rattlesnake, Crotalus viridis viridis.Canadian Journal of Zoology 63:86–91.
Gardiner et al. � Summer Habitat Use by Snakes 981

Gardiner, L. E., and K.W. Sonmor. 2011. Major slump event at GrasslandsNational Park Snake Pit in southwestern Saskatchewan. Blue Jay 69:120–124.
Gardiner, L. E., J. A. Martino, R. G. Poulin, and C. M. Somers. 2011.Eastern yellow-bellied racer populations on the Canadian prairies. Blue Jay69:70–74.
Hill, M. M. A., G. L. Powell, and A. P. Russell. 2001. Diet of the prairierattlesnake, Crotalus viridis viridis, in Southeastern Alberta. CanadianField-Naturalist 115:241–246.
Himes, J. G., L. M. Hardy, D. C. Rudolf, and S. J. Burgdorf. 2006.Movement patterns and habitat selection by native and repatriatedLouisiana pine snakes (Pituophis ruthveni): implications for conservation.Herpetological Natural History 9:103–116.
Hirth, H. F., R. C. Pendleton, A. C. King, and T. R. Downard. 1966.Dispersal of snakes from a hibernaculum in northwestern Utah. Ecology50:332–339.
Johnson, D. H. 1980. The comparison of usage and availability measure-ments for evaluating resource preference. Ecology 61:65–71.
Kapfer, J. M., J. R. Coggins, and R. Hay. 2008. Spatial ecology and habitatselection of bullsnakes (Pituophis catenifer sayi) at the northern periphery oftheir geographic range. Copeia 2008:815–826.
Keller,W. L., and E. J. Heske. 2000.Habitat use by three species of snakes atthe Middle Fork Fish and Wildlife Area, Illinois. Journal of Herpetology34:558–564.
Klug, P. E., J. Fill, and K. A.With. 2011a. Spatial ecology of eastern yellow-bellied racer (Coluber constrictor flaviventris) and great plains rat snake(Pantherophis emoryi) in a contiguous tallgrass prairie landscape.Herpetologica 67:428–439.
Klug, P. E., S. M. Wisely, and K. A. With. 2011b. Population geneticstructure and landscape connectivity of the eastern yellow-bellied racer(Coluber constrictor flaviventris) in the contiguous tallgrass prairie ofNortheastern Kansas, USA. Landscape Ecology 26:281–294.
Kot, M., M. A. Lewis, and P. van den Driessche. 1996. Dispersal data andthe spread of invading organisms. Ecology 77:2027–2042.
Marshall, J. C. Jr., J. V. Manning, and B. A. Kingsbury. 2006. Movementand macrohabitat selection of the Eastern Massasauga in a fen habitat.Herpetologica 62:141–150.
Martino, J. A., R. G. Poulin, D. L. Parker, and C.M. Somers. 2012. Habitatselection by grassland snakes at northern range limits: implications forconservation. Journal of Wildlife Management 76:759–767.
Pendlebury, G. B. 1977. Distribution and abundance of the prairierattlesnake, Crotalus viridis viridis, in Canada. Canadian Field-Naturalist91:122–129.
Prior, K. A., and P. J.Weatherhead. 1996.Habitat features of black rat snakehibernacula in Ontario. Journal of Herpetology 30:211–218.
Reed, R. N., and M. E. Douglas. 2002. Ecology of the grand canyonrattlesnake (Crotalus viridis abyssus) in the Little Colorado River Canyon,Arizona. Southwestern Naturalist 47:30–39.
Reinert, H. K., and D. Cundall. 1982. An improved surgical implantationmethod for radio-tracking snakes. Copeia 1982:702–705.
Rodrı́guez-Robles, J. A. 2003. Home ranges of gopher snakes (Pituophiscatenifer, Colubridae) in central California. Copeia 2003:391–396.
Schaefer, W. H. 1934. Diagnosis of sex in snakes. Copeia 1934:81.Schoener, T. W. 1974. Resource partitioning in ecological communities.Science 174:27–37.
Sokolowski, M. B. 1998. Genes for normal behaviour variation: recent cluesfrom flies and worms. Neuron 21:1–4.
Watson, S. M., and A. P. Russell. 1997. Status of the prairie rattlesnake(Crotalus viridis viridis) in Alberta. Alberta Environmental Protection,Wildlife, Management Division, Wildlife Status Report No. 6,Edmonton, Alberta, Canada.
Williams, K. E., K. E. Hodges, and C. A. Bishop. 2012. Small reservesaround hibernation sites may not adequately protect mobile snakes: theexample of great basin gophersnakes (Pituophis catenifer deserticola) inBritish Columbia. Canadian Journal of Zoology 90:304–312.
Associate Editor: Paul Beier.
982 The Journal of Wildlife Management � 77(5)