basic research alterations of mast cells and tgf- 1 on · pdf file1142 issn 1007-9327 cn...
TRANSCRIPT

• BASIC RESEARCH •
Alterations of mast cells and TGF-1 on the silymarin treatment
for CCl4-induced hepatic fibrosis
Da-Hee Jeong, Gi-Ppeum Lee, Won-Il Jeong, Sun-Hee Do, Hai-Jie Yang, Dong-Wei Yuan, Ho-Yong Park, Kyu-Jong Kim,
Kyu-Shik Jeong
EL SEVIER
PO Box 2345, Beijing 100023, China World J Gastroenterol 2005;11(8):1141-1148
www.wjgnet.com World Journal of Gastroenterology ISSN 1007-9327
[email protected] © 2005 The WJG Press and Elsevier Inc. All rights reserved.
Da-Hee Jeong, Gi-Ppeum Lee, Won-Il Jeong, Sun-Hee Do,Hai-Jie Yang, Dong-Wei Yuan, Kyu-Shik Jeong, Department ofPathology, College of Veterinary Medicine, Kyungpook NationalUniversity, Daegu 702-701, Republic of KoreaHo-Yong Park, Korea Research Institute of Bioscience andBiotechnology, Daejon, Republic of KoreaKyu-Jong Kim, Dr. Kim Medicos Clinics, JungGu, SamdeokDong3-21, Daegu 702-701, Republic of KoreaSupported by the Brain Korea 21 project in 2004 and the Ministryof Agriculture and Forestry project, No. 202059032WTO11Correspondence to: Professor, Kyu-Shik Jeong, D.V.M., Ph.D.,College of Veterinary Medicine, Kyungpook National University,Daegu 702-701, Republic of Korea. [email protected]: +82-53-950-5975 Fax: +82-53-950-5955Received: 2004-07-26 Accepted: 2004-09-09
Abstract
AIM: Silymarin is a potent antioxidant, antiinflammatoryand anti-fibrogenic agent in the liver, which is mediatedby alteration of hepatic Kupffer cell function, lipidperoxidation, and collagen production. Especially, inhepatic fibrogenesis, mast cells are expressed in chronicinflammatory conditions, and promote fibroblast growthand stimulate production of the extracellular matrix byhepatic stellate cells.
METHODS: We examined the inhibitory mechanism ofsilymarin on CCl4-induced hepatic cirrhosis in rats. At 4, 8,and 12 wk, liver tissues were examined histopathologicallyfor fibrotic changes produced by silymarin treatment.
RESULTS: In the silymarin with CCl4-treated group,increase of hepatic stellate cells and TGF-1 productionwere lower than in the CCl4-treated group at early stages.Additionally, at the late fibrogenic stage, expressions ofTGF-1 were weaker and especially not expressed inhepatocytes located in peripheral areas. Moreover, thenumber of mast cell in portal areas gradually increasedand was dependent on the fibrogenic stage, but those ofCCl4+silymarin-treated group decreased significantly.
CONCLUSION: Anti-fibrotic and antiinflammatory effectsof silymarin were associated with activation of hepaticstellate cells through the expression of TGF-1 andstabilization of mast cells. These results suggest thatsilymarin prevent hepatic fibrosis through suppression ofinflammation and hypoxia in the hepatic fibrogenesis.
© 2005 The WJG Press and Elsevier Inc. All rights reserved.
Key words: Silymarin; TGF-1; Mast cell; Hepatic fibrosis
Jeong DH, Lee GP, Jeong WI, Do SH, Yang HJ, Yuan DW,Park HY, Kim KJ, Jeong KS. Alterations of mast cells andTGF-1 on the silymarin treatment for CCl4-induced hepaticfibrosis. World J Gastroenterol 2005; 11(8): 1141-1148
http://www.wjgnet.com/1007-9327/11/1141.asp
INTRODUCTION
Silymarin, a standardized extract of the milk thistle (Silybummarianum [L.] Gaertner) has a long tradition as an herbalremedy[1]. The flavonoid silymarin was introduced as a“hepatoprotective” agent a few years ago and is usedclinically in Europe and Asia for the treatment of liverdiseases[2]. The protective action of silymarin is explicablein terms of its capacity for trapping free radicals and has astabilizing effect on the cytoplasmic membranes. Inexperimental animals, this flavonoid has a protective actionon the liver, which is particularly vulnerable to poisoning byseveral hepatotoxic substances such as carbon tetrachloride(CCl4), thioacetamide, and D-galactosamine[3]. Silymarin isa potent antioxidant that inhibits lipid peroxide formationin the liver cells[4], and possesses antiinflammatory propertiesmediated by alteration of hepatic Kupffer cell function[5].
Hepatic fibrosis has been noted in chronic liver disease,and is characterized by increased production and depositionof collagen, glycoproteins, and proteoglycans that composethe extracellular matrix (ECM)[6]. Availability of animalmodels is crucial for the study of liver fibrosis and/orcirrhosis. It is well known that hepatic cirrhosis animalmodels for chronic liver damage induced by CCl4 in ratsproduce liver fibrosis and biochemical and histologicalpatterns that resemble human liver cirrhosis[7]. Thus, therat model of liver cirrhosis has been useful in studying theeffects of hepatoprotective drugs with therapeutic potentialto be used in humans[8]. In hepatic fibrogenesis,myofibroblasts (MFBs) such as hepatic stellate cells (HSCs)are the major source of increased ECM[6]. When they areexposed to soluble factors from damaged hepatocytes andfrom activated Kupffer cells, MFBs will lose Vitamin Aand their lipid contents and undergo activation. The activatedMFBs migrate and proliferate at the site of liver injury,playing a pivotal role in the formation of fibrous tissue[9].Among the various cytokines, TGF-1 plays an importantrole as a profibrogenic factor in chronic liver disease,triggering the expression of procollagen-I and tissue inhibitorof metalloproteinases-1 (TIMP-1), key effectors of

1142 ISSN 1007-9327 CN 14-1219/ R World J Gastroenterol February 28, 2005 Volume 11 Number 8
fibrogenesis. TGF-1 is also the most potent mast cellchemo-attractant so far identified and induces mast cellmigration at femtomolar (fM) concentrations. Various othercell types, e.g., monocytes, neutrophils, and fibroblasts alsomigrate towards TGF-1[10].
Mast cells, which are derived from hematopoieticprogenitors, leave the bone marrow and migrate to areasof inflammation. A number of factors responsible for thisdirectional migration and tissue maturation of mast cellshave been identified. These include the CXC family ofchemokines, stem cell factor (also known as kit-ligand, steelfactor, and mast cell growth factor), and TGF-1[11]. Thus,the activation of mast cells and the subsequent exocytosisof granules are followed by production and secretion ofcytokines and other factors that lead to leukocyte infiltrationand local inflammation. Mast cell hyperplasia in the liverhas also been observed in a variety of experimental modelsof rat liver fibrosis, such as that induced by CCl4,diethylnitrosamine, radiation, porcine serum, and bile ductresection[12]. In addition, silymarin acts to stabilize hepatocytemembranes and block receptor binding of various toxinsand drugs. Antioxidant activity is also hepatoprotectivein vivo and in vitro studies, showing that silymarin has freeradical scavenging activity and enhances superoxidedismutase action in erythrocytes and lymphocytes[13].Silymarin also protects against glutathione depletion andincreases protein synthesis by hepatocytes when there isdamage to parenchymatous tissue[14].
In this study, we examined the inhibitory mechanism ofsilymarin on CCl4-induced hepatic cirrhosis in rats. Theobjective of the present study was to observe the alterationof MFBs, TGF-1, and mast cells histopathologically onthe silymarin treatment and to elucidate the correlationbetween these changes and the antifibrotic effect ofsilymarin.
MATERIALS AND METHODS
Animals and treatmentsStudies were performed on thirty male Wistar rats weighing130-150 g. They were housed in a room at 22±2 ℃ and a12-h light-dark cycle. Feed (PMI Nutrition International,USA) and water were supplied ad libitum.
Hepatic fibrosis was induced experimentally byintraperitoneal injection (IP) of 1.0 mL/kg body weight of10% CCl4 (Sigma, USA) dissolved in olive oil (Sigma, USA),three times a week for 12 wk. Two groups of fifteen animalseach were used. These groups are schematically shown inTable 1. The first (CCl4) group received CCl4, 1.0 mL/kgIP three times a week and 0.25% carboxymethylcellulose(CMC, Sigma, USA), 1.0 mL/kg per oral, 5 d a weekfor 12 wk. The second (CCl4+Sily) group received CCl4,1.0 mg/kg IP and a daily oral dose of 50 mg/kg silymarin5 times a week. Silymarin was given as a suspension in0.25% CMC. Five rats of each group were sacrificed at wk4, 8, and 12 respectively.
Serum biochemical measurementsSerum was collected and assayed for alanine aminotransferase(ALT) and aspartate aminotransferase (AST) using standard
enzymatic assay kits. Each assay is a colorimetric assay withdetection of a highly colored end product measured at490-520 nm using autoanalyzer - UV/V is spectrophotometer(Hitachi 736-10, Hitachi, Japan). The absorbance of eachend product is proportional to the enzyme’s activity.
Determination of hepatic hydroxyproline contentHydroxyproline (HYP) was determinated colorimetricallyin duplicates from 0.2 g of liver tissue using a modifiedmethod of Jamall et al[15]. Briefly, the frozen tissue washomogenized in 4 mL of 6N HCl and hydrolyzed at 110 ℃for 16 h. The hydrolysate was filtered, and then 30 Laliquot of these samples was evaporated under vacuum.The sediment was dissolved in 1.2 mL of isopropanol andincubated with 0.2 mL of 0.84% chloramines-T in acetate-citrate buffer (pH 6.0) for 10 min at room temperature.Then, 1.0 mL of Ehrlich’s reagent was added and themixture was incubated at 60 ℃ for 25 min. The absorbanceof the sample solution was measured at 560 nm wavelength(Hitachi 736-10, Hitachi, Japan). Next, the hydroxyprolinecontent in 100 mg of liver was calculated from the standardcurve of 4-hydroxy-L-proline (Sigma, USA) (g/100 mgliver weight).
Histopathological analysisLiver tissues from each rat were rapidly removed, fixed in10% neutral-buffered formalin, and processed routinely.Paraffin-embedded sections were cut into 4 m thicksections. The sections were stained with hematoxylin andeosin (HE) and with special Azan stain, for collagen fibers.In there experiments, the degree of fibrosis in each sectionof liver was classified as a grade 0-4[16].
ImmunohistochemistryLiver sections were deparaffinized in xylene, dehydrated ingraded alcohol series, and for the block of endogenousperoxidase, sections were incubated in a solution of 3%hydrogen peroxide (H2O2) in methanol for 10 min. Tissuesections were washed with PBS containing 0.03% non-fatmilk and 0.01% Tween 20, and then immunostained withprimary antibodies for alpha-smooth muscle actin (alpha-SMA) and TGF-1. The antigen-antibody complex wasvisualized by a labeled streptavidin-biotin method using aHistostatin-PLUS Bulk Kit (Zymed Laboratories Inc., USA)and followed by diaminobenzidine (DAB) as a chromogen.After washing, slides were counter-stained with Meyer’shematoxylin and washed with tap water. The primaryantibodies used were monoclonal anti -SMA at a dilutionof 1:800 (clone 1A4, Sigma, USA) and polygonal rabbitLC[1-30] antibody to mature TGF-1 (kindly provided byDr. Seong-Jin Kim) at a dilution of 1:100 respectively. Non-
Table 1 Experimental designs used in this study
Group ID Number Treatment Sacrificed time of animal
CCl4 15 CCl4, 1.0 mg/(kg·d), IP 3 times a wk 4, 8, 12 wk
0.25% CMC, 1 mL/(kg·d), PO 5 times a wk
CCl4 + Sily 15 CCl4, 1.0 mg/(kg·d), IP 3 times a wk 4, 8, 12 wk
Silymarin, 50 mg/(kg·d), PO 5 times a wk

Jeong DH et al. Alterations of mast cell and TGF-1 by silymarin 1143
immunized goat sera, which were used instead of theprimary antibody, served as the negative control.
Number of mast cells in liver tissueToluidine blue staining for mast cells was performed byimmersion of liver sections in 0.1% toluidine blue (Sigma,USA) for 1 min at room temperature. The number of mastcells was quantified in 25 randomly selected, non-overlappingfields and expressed as the number of mast cells/mm2.
Statistical analysisAll results were expressed as mean and standard deviation(SD). Statistical analysis of the data was done using InStatprogram (GraphPad Software Inc.). A Mann-WhitneyU-test was conducted and the data that were considered tobe significantly different were reported at probability levelsof P<0.05 or P<0.005, as indicated.
RESULTS
Serum biochemistrySerum ALT and AST levels of all CCl4-tested animalsincreased in a time-dependent fashion during the whole ofexperimental period (Figure 1). These changes indicated thatliver damage and inflammation were induced successfullyby injection of CCl4 and were similar to previous results.However, in the CCl4+silymarin treatment group, increasesof serum ALT and AST levels were lower than those ofthe CCl4, control group. The reduced ALT and AST activitiesof the silymarin-treated group were 30%, 72% and 33%compared to increases of 8%, 55% and 34% in the CCl4-treated groups, at 4, 8, and 12 wk.
Figure 1 Serum ALT and AST levels in the CCl4- or CCl4 with silymarin-treated groups. A: Serum ALT levels were detected at 4, 8, and 12 wk;B: Serum AST levels were detected at 4, 8, and 12 wk. In the CCl4
with silymarin-treated group (black bar), increase of serum ALT andAST levels were lower than those of CCl4-treated group (open bar).
Collagen accumulationCollagen contents of liver tissue were quantified bydetermination of the hydroxyproline content (Figure 2).In the CCl4-treated group, hydroxyproline contents at4, 8, and 12 wk increased 21.81±3.37, 24.31±0.91, and43.26±1.41 g/100 mg liver weight respectively.However, in the CCl4+silymarin-treated group, the HYPcontents at 4, 8, and 12 wk were 18.05±1.89, 20.5±1.92,and 27.5±0.69 g/100 mg liver weight respectively. At 4,8, and 12 wk, these HYP contents of CCl4+silymarin-treatedgroup were reduced by 17%, 16%, and 36% respectivelycompared to those of control group. Reduced HYP contentsof 8 and 12 wk in the CCl4+silymarin-treated group werestatistically significant, P<0.05 and <0.005 respectively.
Figure 2 Hydroxyproline content determinated in the livers of CCl4-or CCl4 with silymarin-treated groups. At 4, 8 and 12 wk, HYP con-tents of CCl4 with silymarin-treated group (black bar) reduced com-pared to those of control group (open bar). Data are expressed asthe mean±SD (n = 5). aP<0.05 means significant compared to that ofCCl4-treated groups. cP<0.005 means significant compared to thatof CCl4-treated groups.
Histopathological observationsDuring the process of cirrhosis, the grade of hepatic fibrosischanged from grade 0 to grade 4 (Table 2). At 4 wk,centrilobular necrosis and moderate fatty change of liverwere found in the CCl4-only-treated group. In this group,there was collagen accumulation around the blood vessels(Figure 3A, G). At 8 wk, fatty change was rarely observed,but collagen fibers were more abundant in the centrilobulararea and neighboring central veins were bridged by fibroussepta. Then, pseudolobuli were formed by thin fibrous septa(Figure 3B, H). At 12 wk, pseudolobuli were formed andmacrovesicular lipid droplets were detected. The collagenoussepta were much thicker, and pseudolobuli were subdividedinto smaller lobuli (Figure 3C, I).
In the silymarin with CCl4 injection group, at 4 wk, thepresence of connective tissue was almost normal aroundthe central veins (Figure 3D, J). At wk 8, moderate to severefatty change was detected around the periportal area andcentral vein. A slight accumulation and spread of collagenfibers around central veins was observed too (Figure 3E, K).At wk 12, large lipid accumulation was detected more than
CCl4 Sily CCl4 Sily CCl4 Sily
1 500
1 000
500
0
4 8 12 (wk)
ALT
(IU
/L)
CCl4 Sily CCl4 Sily CCl4 Sily
1 500
1 000
500
0
AST (
IU/L
)
A
B
CCl4 Sily CCl4 Sily CCl4 Sily
40
30
20
10
0
a
Hyd
roxy
pro
line c
once
ntr
ation
(g/1
00 m
g liv
er
weig
ht)
c
4 8 12 (wk)
4 8 12 (wk)
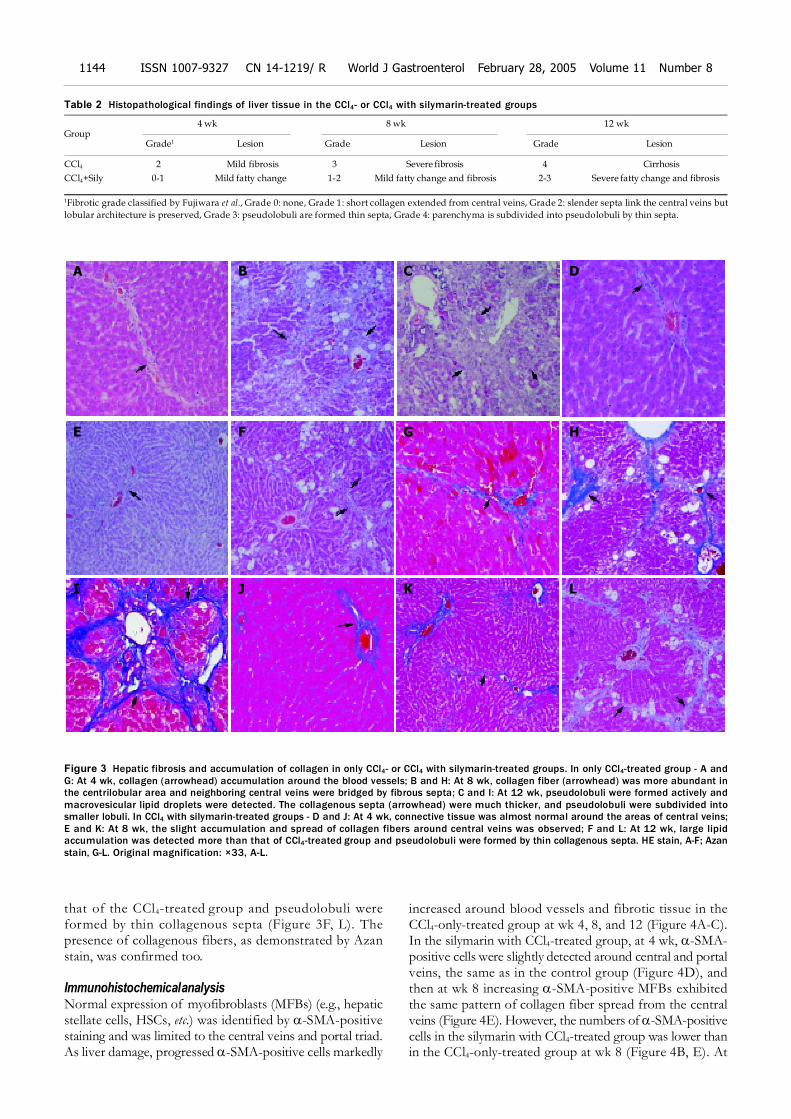
that of the CCl4-treated group and pseudolobuli wereformed by thin collagenous septa (Figure 3F, L). Thepresence of collagenous fibers, as demonstrated by Azanstain, was confirmed too.
Immunohistochemical analysisNormal expression of myofibroblasts (MFBs) (e.g., hepaticstellate cells, HSCs, etc.) was identified by -SMA-positivestaining and was limited to the central veins and portal triad.As liver damage, progressed -SMA-positive cells markedly
increased around blood vessels and fibrotic tissue in theCCl4-only-treated group at wk 4, 8, and 12 (Figure 4A-C).In the silymarin with CCl4-treated group, at 4 wk, -SMA-positive cells were slightly detected around central and portalveins, the same as in the control group (Figure 4D), andthen at wk 8 increasing -SMA-positive MFBs exhibitedthe same pattern of collagen fiber spread from the centralveins (Figure 4E). However, the numbers of -SMA-positivecells in the silymarin with CCl4-treated group was lower thanin the CCl4-only-treated group at wk 8 (Figure 4B, E). At
Table 2 Histopathological findings of liver tissue in the CCl4- or CCl4 with silymarin-treated groups
4 wk 8 wk 12 wkGroup
Grade1 Lesion Grade Lesion Grade Lesion
CCl4 2 Mild fibrosis 3 Severe fibrosis 4 Cirrhosis
CCl4+Sily 0-1 Mild fatty change 1-2 Mild fatty change and fibrosis 2-3 Severe fatty change and fibrosis
1Fibrotic grade classified by Fujiwara et al., Grade 0: none, Grade 1: short collagen extended from central veins, Grade 2: slender septa link the central veins but
lobular architecture is preserved, Grade 3: pseudolobuli are formed thin septa, Grade 4: parenchyma is subdivided into pseudolobuli by thin septa.
Figure 3 Hepatic fibrosis and accumulation of collagen in only CCl4- or CCl4 with silymarin-treated groups. In only CCl4-treated group - A andG: At 4 wk, collagen (arrowhead) accumulation around the blood vessels; B and H: At 8 wk, collagen fiber (arrowhead) was more abundant inthe centrilobular area and neighboring central veins were bridged by fibrous septa; C and I: At 12 wk, pseudolobuli were formed actively andmacrovesicular lipid droplets were detected. The collagenous septa (arrowhead) were much thicker, and pseudolobuli were subdivided intosmaller lobuli. In CCl4 with silymarin-treated groups - D and J: At 4 wk, connective tissue was almost normal around the areas of central veins;E and K: At 8 wk, the slight accumulation and spread of collagen fibers around central veins was observed; F and L: At 12 wk, large lipidaccumulation was detected more than that of CCl4-treated group and pseudolobuli were formed by thin collagenous septa. HE stain, A-F; Azanstain, G-L. Original magnification: ×33, A-L.
A B C D
E F G H
I J K L
1144 ISSN 1007-9327 CN 14-1219/ R World J Gastroenterol February 28, 2005 Volume 11 Number 8
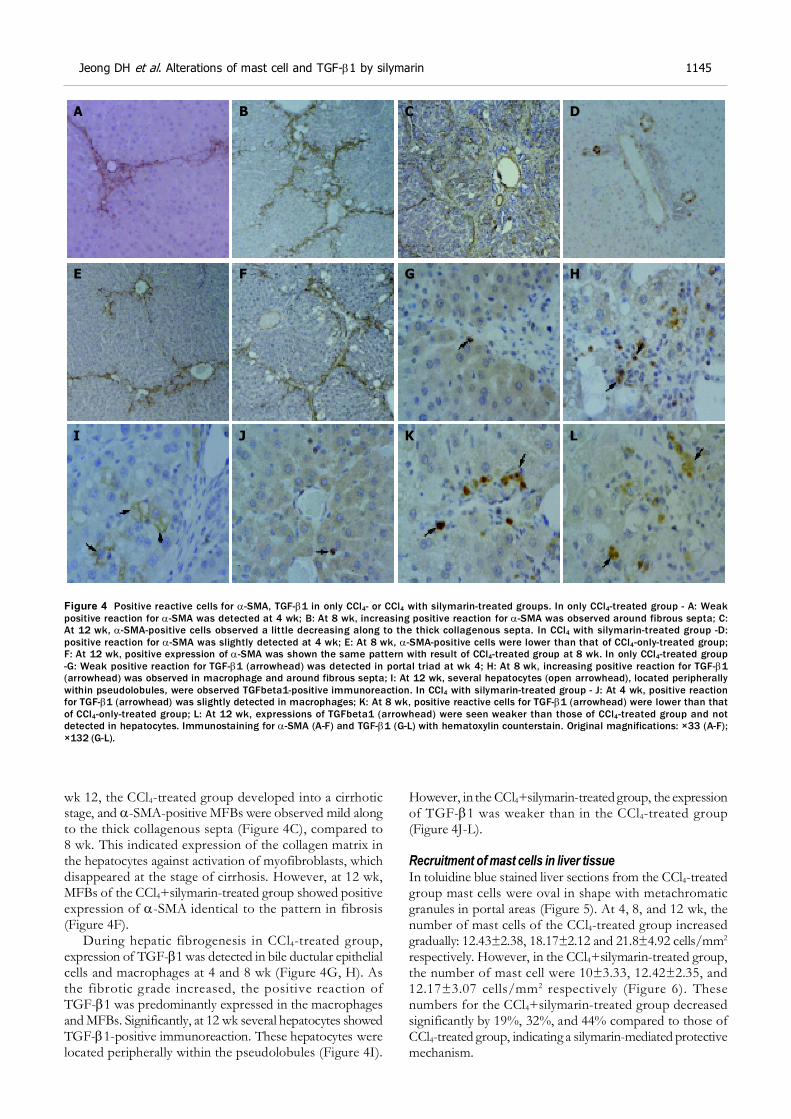
wk 12, the CCl4-treated group developed into a cirrhoticstage, and -SMA-positive MFBs were observed mild alongto the thick collagenous septa (Figure 4C), compared to8 wk. This indicated expression of the collagen matrix inthe hepatocytes against activation of myofibroblasts, whichdisappeared at the stage of cirrhosis. However, at 12 wk,MFBs of the CCl4+silymarin-treated group showed positiveexpression of -SMA identical to the pattern in fibrosis(Figure 4F).
During hepatic fibrogenesis in CCl4-treated group,expression of TGF-1 was detected in bile ductular epithelialcells and macrophages at 4 and 8 wk (Figure 4G, H). Asthe fibrotic grade increased, the positive reaction ofTGF-1 was predominantly expressed in the macrophagesand MFBs. Significantly, at 12 wk several hepatocytes showedTGF-1-positive immunoreaction. These hepatocytes werelocated peripherally within the pseudolobules (Figure 4I).
However, in the CCl4+silymarin-treated group, the expressionof TGF-1 was weaker than in the CCl4-treated group(Figure 4J-L).
Recruitment of mast cells in liver tissueIn toluidine blue stained liver sections from the CCl4-treatedgroup mast cells were oval in shape with metachromaticgranules in portal areas (Figure 5). At 4, 8, and 12 wk, thenumber of mast cells of the CCl4-treated group increasedgradually: 12.43±2.38, 18.17±2.12 and 21.8±4.92 cells/mm2
respectively. However, in the CCl4+silymarin-treated group,the number of mast cell were 10±3.33, 12.42±2.35, and12.17±3.07 cells/mm2 respectively (Figure 6). Thesenumbers for the CCl4+silymarin-treated group decreasedsignificantly by 19%, 32%, and 44% compared to those ofCCl4-treated group, indicating a silymarin-mediated protectivemechanism.
Figure 4 Positive reactive cells for -SMA, TGF-1 in only CCl4- or CCl4 with silymarin-treated groups. In only CCl4-treated group - A: Weakpositive reaction for -SMA was detected at 4 wk; B: At 8 wk, increasing positive reaction for -SMA was observed around fibrous septa; C:At 12 wk, -SMA-positive cells observed a little decreasing along to the thick collagenous septa. In CCl4 with silymarin-treated group -D:positive reaction for -SMA was slightly detected at 4 wk; E: At 8 wk, -SMA-positive cells were lower than that of CCl4-only-treated group;F: At 12 wk, positive expression of -SMA was shown the same pattern with result of CCl4-treated group at 8 wk. In only CCl4-treated group-G: Weak positive reaction for TGF-1 (arrowhead) was detected in portal triad at wk 4; H: At 8 wk, increasing positive reaction for TGF-1(arrowhead) was observed in macrophage and around fibrous septa; I: At 12 wk, several hepatocytes (open arrowhead), located peripherallywithin pseudolobules, were observed TGFbeta1-positive immunoreaction. In CCl4 with silymarin-treated group - J: At 4 wk, positive reactionfor TGF-1 (arrowhead) was slightly detected in macrophages; K: At 8 wk, positive reactive cells for TGF-1 (arrowhead) were lower than thatof CCl4-only-treated group; L: At 12 wk, expressions of TGFbeta1 (arrowhead) were seen weaker than those of CCl4-treated group and notdetected in hepatocytes. Immunostaining for -SMA (A-F) and TGF-1 (G-L) with hematoxylin counterstain. Original magnifications: ×33 (A-F);×132 (G-L).
A B C D
E F G H
LKJI
Jeong DH et al. Alterations of mast cell and TGF-1 by silymarin 1145

DISCUSSION
Silymarin is well known to be a protective agent againstvarious hepatotoxins, such as acetaminophen, alcohol,carbon tetrachloride, tetrachloromethane, and toluene[17].Pretreating rats and mice with silymarin before exposure tothese chemical hepatotoxins significantly reduced lipidperoxidation and hepatotoxicity[3]. Additionally, thepharmacological effects of silymarin include regulation ofcell membrane permeability, leukotriene inhibition, reactiveoxygen species scavenging, and suppression of NF-kappaBDNA binding activity[18].
In animal studies, silymarin is as effective as colchicinein reversing hepatic fibrosis due to CCl4-induced damage[1].A CCl4-induced hepatic cirrhosis rat model has been useful
in studying the effects of hepatoprotective drugs withtherapeutic potential to be used in humans[8]. In the currentstudy, hepatic fibrosis/cirrhosis was successfully inducedby CCl4 injection in rats and flavorous results were obtainedsimilar to previous study. Total hepatic collagen contentsdetermined by hydroxyproline content increased graduallyduring experimental period and histopathological findingsof fibrosis/cirrhosis were observed in H&E and the Azan-stained section. In the present study, the hepatoprotectiveeffect of silymarin was determined in CCl4-induced livercirrhosis of rats. In the CCl4 with silymarin-treated group,total hepatic collagen contents were significantly lower thanin the CCl4-treated group especially at 8 and 12 wk.Additionally histopathological findings of fibrosis/cirrhosiswere revealed a significant reduction in the CCl4-treatedgroup. Similar experiments by Favari et al reported thereduction of lipid peroxidation, Na+, K+, and Ca2+-ATPaselevels and increases of collagen content[3,19].
In hepatic fibrogenesis, myofibroblasts (MFBs) are themajor source of increased ECM. The activated MFBsmigrate and proliferate at the site of liver injury and play apivotal role in the formation of fibrous tissue. Therefore,activated MFBs are considered the major cellular target toprevent the progression of liver fibrosis during the newdrug development[9]. In addition, transforming growth factorbeta (TGF-) is a potent fibrogenic cytokine produced byKupffer cells and HSCs. There is a prolonged increase ofTGF-1 expression during hepatic fibrosis in CCl4- anddiethylnitrosamine-induced models[20] and in patients withcirrhosis induced by alcohol or viral hepatitis[21].
In the current study, as liver damage progressed in theCCl4-only-treated group, -SMA-positive cells markedlyincreased around fibrous septa. The number of these cellsincreased from wk 4 to 8, but slightly decreased along the
Figure 5 Toluidine blue-stained liver tissue of only CCl4- or CCl4 with silymarin-treated groups. In only CCl4-treated group - A: At 4 wk, specifictoluidine blue-stained mast cells (arrowhead) were detected in periportal region; B: At 8 wk, the number of mast cell increased; C: At 12 wk,mast cells increased along the thick collagenous septa. In CCl4 with silymarin-treated group - D: Positive reaction for toluidine blue(arrowhead) was slightly detected at 4 wk; E: At 8 wk, -SMA-positive cells were lower than that of CCl4-only-treated group; F: At 12 wk,positive expression of -SMA was shown in the same pattern as with the result of CCl4-treated group at 8 wk. Toluidine blue stain, A-F. Originalmagnification: ×132, A-F.
Figure 6 Recruitment of mast cell numbers in only CCl4- or CCl4 withsilymarin-treated groups. At 4, 8 and 12 wk, mast cell numbers ofCCl4 with silymarin-treated group (black bar) significantly reducedcompared to those of control group (open bar). Data are expressedas the mean±SD. aP<0.05 means significant compared to that ofCCl4-treated groups. cP<0.005 smeans significant compared to thatof CCl4-treated groups.
A B C
D E F
CCl4 Sily CCl4 Sily CCl4 Sily
30
20
10
0
Num
bers
of
mast
cells
(cel
ls/m
m2 s
ect
ion)
1146 ISSN 1007-9327 CN 14-1219/ R World J Gastroenterol February 28, 2005 Volume 11 Number 8
4 8 12 (wk)
c a c

thick collagenous septa at 12 wk, developed into the cirrhoticstage. Characteristically, in immunohistochemical analysisfor TGF-1, positive reactions were mainly expressed byHSC and macrophages around the portal region, at early(4 wk) and middle (8 wk) stages of the fibrotic processes,but they were predominantly observed in hepatocytes locatedin pseudolobules peripherally, at the cirrhosis-occurred stage(12 wk). The mechanism of TGF-1 expression inhepatocytes has been studied by several researchers, but isstill unclear. Furthermore, these alterations of TGF-1expression were already reported by our laboratory and wesuggested that hypoxia might be associated with fibrogenesisin the liver[22]. However, in the silymarin+CCl4-treated groupof our study, there was increase of -SMA-positive cellssuch as MFBs, lower than that of CCl4-only-treated group,at 4, 8, and 12 wk. Additionally, expressions of TGF-1were weaker than those of CCl4-treated group, during allexperimental periods, especially at 12 wk not expressed inhepatocyte located in peripheral areas of pseudolobules.Thus, based on the results of the current study, it is concludedthat silymarin has protective effect of proliferation andTGF-1 production in MFBs.
Fuchs et al[23] reported on the basis of in vitro studiesthat the potential antifibrotic properties of silymarin mightbe the inhibition of hepatic stellate cell proliferation andtransformation. Jia et al[24] observed that silymarin suppressesexpression of profibrogenic procollagen alpha1 (I) andTIMP-1 most likely via down-regulation of TGF-1 mRNAin rats with biliary fibrosis. However, in vivo studies of theanti-fibrotic activities of silymarin have not yet elucidatedhistopathologically the preventive mechanism of activationor proliferation of MFBs by silymarin during the CCl4-induced hepatic fibrogenesis. These results suggest thatalterations of the numbers of MFBs and TGF-1 expressionin the liver may be involved in the hepatoprotective effectsof silymarin observed in other studies. Another study ofsilymarin explained the antifibrotic action through the effectson TGF-1 expression[24]. Silymarin has been noted toregenerate cells and enhance RNA synthesis in the rat liver[25].
In liver fibrosis and/or cirrhosis, several studies havereported a relationship between mast cell density,hepatocellular damage, mRNA encoding TGF-1, hepaticstellate cell activation, and collagen levels[26-28]. Mast cellshave been implicated in chronic inflammatory conditionsresulting in fibrosis, such as Crohn’s disease, and have beenidentified in human liver. The number of mast cells arereported to increase in chronic liver diseases associated withfibrosis[12]. Armbrust et al[29] demonstrated that in the latestage of liver fibrogenesis, mast cells may be involved bydisplaying protease inhibitory activity in the fibrotic septa.In our previous study, the chronic injection of CCl4 inducedrat liver cirrhosis concomitant with a marked increase ofmast cells[30]. In this study, the number of mast cells inportal areas gradually increased in the fibrogenic stage, butthe number of mast cells in the CCl4+silymarin-treated groupdecreased significantly compared to those of CCl4-treatedgroup. In the study of Fantozzi et al, there was inhibitionof neutrophil-mediated histamine release dose-dependently.These results further stress the concept of a neutrophil-mast cell interaction, which may be involved in inflammatory
processes[31]. Moreover, mast cells secrete various mediators,which promote fibroblast growth, stimulate production ofthe extracellular matrix by fibroblasts of hepatic stellatecells, and produce components of the extramedullary matrixthemselves[32,33]. However, it is unclear whether they play acentral role in its development.
The anti-inflammatory effects of silymarin are also basedon multiple activities including mast cell stabilization,inhibition of neutrophil migration, Kupffer cell inhibition,inhibition of leukotrienes, and prostaglandin formation.However, results of earlier studies have not histopathologicallyshown expression of mast cells in fibrotic liver tissue aftersilymarin treatment. Thus, silymarin has been histopathologicallyshown to have significant antiinflammatory effect on hepatictissue, including mast cell stabilization. In addition, it is likelythat the hepatoprotective effect of silymarin is related toprevention of hypoxia in hepatic fibrogenesis.
In conclusion, the anti-fibrotic and antiinflammatoryeffects of silymarin were histopathologically observed inthe hepatic fibrogenesis of chronic liver damage inducedby CCl4 treatment. Furthermore, these effects wereassociated with activation of MFBs, expression of TGF-1,and stabilization of mast cells. These results suggest thatsilymarin prevents hepatic fibrosis through the suppressionof inflammation and hypoxia in CCl4-induced rat livercirrhosis.
REFERENCES
1 Valenzuela A, Garrido A. Biochemical bases of the pharma-cological action of the flavonoid silymarin and of its struc-
tural isomer silibinin. Biol Res 1994; 27: 105-112
2 Down WH, Chasseaud LF, Grundy RK. Effect of silybin onthe hepaticmicrosomal drug metabolizing enzyme system in
the rat. Arzneim-Forsch. Drug Res 1974; 24: 1986-1988
3 Mourelle M, Muriel P, Favari L, Franco T. Prevention ofCCL4-induced liver cirrhosis by silymarin. Fundam Clin
Pharmacol 1989; 3: 183-191
4 Muriel P, Garcia-pina T, Perez-Alvarez V, Mourelle M.Si lymarin protects against paracetamol-induced l ipid
peroxidation and liver damage. J Appl Toxicol 1992; 12: 439-
4425 Dehmlow C, Erhard J, de Groot H. Inhibition of Kupffer cell
functions as an explanation for the hepatoprotective proper-
ties of silibinin. Hepatology 1996; 23: 749-7546 Akiyoshi H, Terada T. Mast cell, myofibroblast and nerve
terminal complexes in carbon tetrachloride-induced cirrhotic
rat livers. J Hepatol 1998; 29: 112-1197 Perez-Tamayo R. Is cirrhosis of the liver experimentally pro-
duced by CCl4 and adequate model of human cirrhosis?Hepatology 1983; 3: 112-120
8 Paquet KJ, Kamphausen U. The carbon-tetrachloride-hepa-totoxicity as a model of liver damage. First report: Long-timebiochemical changes. Acta Hepatogastroenterol (Stuttg) 1975;22 : 84-88
9 Wu J, Zern MA. Hepatic stellate cells: a target for the treat-ment of liver fibrosis. J Gastroenterol 2000; 35: 665-672
1 0 Olsson N, Piek E, ten Dijke P, Nilsson G. Human mast cellmigration in response to members of the transforming growth
factor-beta family. J Leukoc Biol 2000; 67: 350-3561 1 Jones SE, Kelly DJ, Cox AJ, Zhang Y, Gow RM, Gilbert RE.
Mast cell infiltration and chemokine expression in progressiverenal disease. Kidney Int 2003; 64: 906-913
1 2 Sugihara A, Tsujimura T, Fujita Y, Nakata Y, Terada N.Evaluation of role of mast cells in the development of liverfibrosis using mast cell-deficient rats and mice. J Hepatol 1999;30 : 859-867
Jeong DH et al. Alterations of mast cell and TGF-1 by silymarin 1147

1 3 Altorjay I, Dalmi L, Sari B, Imre S, Balla G. The effect of
silibinin (Legalon) on the the free radical scavenger mecha-nisms of human erythrocytes in vitro. Acta Physiol Hung 1992;
80 : 375-380
1 4 Galisteo M, Rissel M, Sergent O, Chevanne M, Cillard J,Guillouzo A, Lagadic-Gossmann D. Hepatotoxicity of tacrine:
occurrence of membrane fluidity alterations without involve-
ment of lipid peroxidation. J Pharmacol Exp Ther 2000; 294:160-167
1 5 Jamall IS, Finelli VN, Que Hee SS. A simple method to deter-
mine nanogram levels of 4-hydroxyproline in biological tissues.Anal Biochem 1981; 112: 70-75
1 6 Fujiwara K, Ogata I, Ohta Y, Hayashi S, Mishiro S, Takatsuki
K, Sato Y, Yamada S, Hirata K, Oka H. Decreased collagenaccumulation by a prolyl hydroxylase inhibitor in pig serum-
induced fibrotic rat liver. Hepatology 1988; 8: 804-807
1 7 Saller R, Meier R, Brignoli R. The use of silymarin in thetreatment of liver diseases. Drugs 2001; 61: 2035-2063
1 8 Schumann J, Prockl J, Kiemer AK, Vollmar AM, Bang R,
Tiegs G. Silibinin protects mice from T cell-dependent liverinjury. J Hepatol 2003; 39: 333-340
1 9 Favari L, Perez-Alvarez V. Comparative effects of colchicine
and silymarin on CCl4-chronic liver damage in rats. Arch MedRes 1997; 28: 11-17
2 0 Nakatsukasa H, Nagy P, Evarts RP, Hsia CC, Marsden E,
Thorgeirsson SS. Cellular distribution of transforming growthfactor-beta 1 and procollagen types I, III, and IV transcripts
in carbon tetrachloride-induced rat liver fibrosis. J Clin Invest
1990; 85 : 1833-18432 1 Annoni G, Weiner FR, Zern MA. Increased transforming
growth factor-beta 1 gene expression in human liver disease.
J Hepatol 1992; 14: 259-2642 2 Jeong WI, Do SH, Yun HS, Song BJ, Kim SJ, Kwak WJ, Yoo
SE, Park HY, Jeong KS. Hypoxia potentiates transforming
growth factor-beta expression of hepatocyte during the cir-rhotic condition in rat liver. Liver Int 2004; 24: 658-668
2 3 Fuchs EC, Weyhenmeyer R, Weiner OH. Effects of silibinin
and of a synthetic analogue on isolated rat hepatic stellate
cells and myofibroblasts. Arzneimittelforschung 1997; 47 :1383-1387
2 4 Jia JD, Bauer M, Cho JJ, Ruehl M, Milani S, Boigk G, Riecken
EO, Schuppan D. Antifibrotic effect of silymarin in rat second-ary biliary fibrosis is mediated by downregulation of procollagen
alpha1(I) and TIMP-1. J Hepatol 2001; 35: 392-398
2 5 Sonnenbichler J, Zetl I. Mechanism of action of silibinin. V.Effect of silibinin on the synthesis of ribosomal RNA, mRNA
and tRNA in rat liver in vivo. Hoppe Seylers Z Physiol Chem
1984; 365 : 555-5662 6 Grizzi F, Franceschini B, Gagliano N, Moscheni C, Annoni G,
Vergani C, Hermonat PL, Chiriva-Internati M, Dioguardi N.
Mast cell density, hepatic stellate cell activation and TGF-beta1 transcripts in the aging Sprague-Dawley rat during
early acute liver injury. Toxicol Pathol 2003; 31: 173-1782 7 Li CY, Baek JY. Mastocytosis and fibrosis: role of cytokines.
Int Arch Allergy Immunol 2002; 127: 123-1262 8 Zheng M, Ruan Y, Wu Z. Correlation study of TGF beta
expression in diethylnitrosamine-induced rat liver cancer andmast cells in its vicinity. Zhonghua Zhongliu Zazhi 2000; 22:463-465
2 9 Armbrust T, Batusic D, Ringe B, Ramadori G. Mast cellsdistribution in human liver disease and experimental rat liverfibrosis. Indications for mast cell participation in develop-ment of liver fibrosis. J Hepatol 1997; 26: 1042-1054
3 0 Jeong WI, Lee CS, Park SJ, Chung JY, Jeong KS. Kinetics ofmacrophages, myofibroblasts and mast cells in carbon tetra-chloride-induced rat liver cirrhosis. Anticancer Res 2002; 22:869-877
3 1 Fantozzi R, Brunelleschi S, Rubino A, Tarli S, Masini E,Mannaioni PF. FMLP-activated neutrophils evoke histaminerelease from mast cells. Agents Actions 1986; 18: 155-158
3 2 Hatamochi A, Fujiwara K, Ueki H. Effects of histamine oncollagen synthesis by cultured fibroblasts derived from guineapig skin. Arch Dermatol Res 1985; 277: 60-64
3 3 Weber S, Kruger-Krasagakes S, Grabbe J, Zuberbier T,Czarnetzki BM. Mast cells. Int J Dermatol 1995; 34: 1-10
1148 ISSN 1007-9327 CN 14-1219/ R World J Gastroenterol February 28, 2005 Volume 11 Number 8