basis of rise in intracellular sodium in airway hyperresponsiveness and asthma
TRANSCRIPT

Basis of Rise in Intracellular Sodium in Airway
Hyperresponsiveness and Asthma
Anurag Agrawal,1,3 Krishna P. Agrawal,1,2 Arjun Ram,3 Anita Sondhi,1 SunilK. Chhabra,1 Sharad V. Gangal,3 and Dolly Mehta2*
1Department of Cardio-respiratory Physiology, Vallabhbhai Patel Chest Institute, University of Delhi;2Department of Physiology, Institute of Nuclear Medicine and Allied Sciences, Lucknow RoadA;3Allergy and Immunology Section, Institute of Genomics and Integrative Biology, Mall Road, Delhi,
India
Abstract. The aim of this study was to investigate the basis of disturbances insodium transport in asthma and in airway hyperresponsiveness without symp-toms of asthma (asymptomatic AHR). We measured the intracellular sodium(Nai); activity of Na
+/K+-ATPase in unstimulated cells (resting activity) and incell homogenate under optimal conditions (maximal activity); and sodium in-flux, in mixed leukocytes of 15 normal subjects, 12 subjects with asymptomaticAHR, and 26 asthmatics with or without active symptoms. Resting Na+/K+-ATPase activity was the same as sodium influx, consistent with homeostasis.Compared with normal subjects, those with asymptomatic AHR or asthma withcontrolled symptoms had a twofold increase in sodium influx and Nai. Symp-tomatic asthmatics also had a twofold increase in sodium influx but a fourfoldelevation of Nai. Maximal Na
+/K+-ATPase activity was reduced by half insymptomatic asthmatics compared with normal subjects. The reduction ofmaximal Na+/K+-ATPase activity was associated with a significant decrease inATP turnover per Na+/K+-ATPase molecule but not number of Na+/K+-ATPase molecules per cell. In summary, airway hyperresponsiveness with orwithout asthma is associated with increased sodium influx and Na in leukocytes.Resting activity of Na+/K+-ATPase is also increased as a compensatory re-sponse to the increased sodium influx, but it is achieved at the expense of higherNai. Symptomatic asthma is additionally associated with reduction in maximalactivity of Na+/K+-ATPase, resulting in reduced capacity to handle the in-crease in sodium influx and consequent severe elevations in Nai.
*Present address:Department of Pharmacology, University of Chicago, Chicago, IL 60612, USA
Correspondence to: Anurag Agrawal, Department of Physiology, Vallabhbhai Patel Chest
Institute, Delhi University, Delhi 110007, India; email: [email protected]
Lung (2005) 183:375–387
DOI: 10.1007/s00408-005-2549-0

Key words: Asthma—Airway hyperresponsiveness–Airway hyperreactivi-ty—Na+/K+-ATPase—Sodium.
Introduction
Cellular sodium homeostasis is known to be disturbed in asthma [1, 21, 25, 31,34]. However, there has been disagreement on whether Na+/K+-ATPaseactivity increases or decreases, and very limited data are available on otherparameters.Na+/K+-ATPase activity has been reported to be decreased in leukocytes
and platelets of asthmatic patients [21, 31]. These studies used cell homogen-ates under optimal conditions, i.e., when both Na+ and K+ sites of the Na+/K+-ATPase were fully occupied, resulting in maximal enzyme activation.Therefore, while the results of these studies show decreased maximal Na+/K+-ATPase activity, they did not address the activity of this enzyme in intactcells.Orlov et al. [25] reported increased 86Rb uptake in intact erythrocytes of
asthmatic patients, indicating increased resting Na+/K+-ATPase activity. Theyalso observed an increase in intracellular sodium (Nai) in these cells. Tribe et al.[34] found increased sodium influx in leukocytes of patients with asthma.Increase in Nai directly effects Ca
2+regulation via Na+/Ca2+ exchange.Other than its well-described role in calcium efflux, Na+/Ca2+ exchange is amajor mechanism of Ca2+ influx in many cells including lymphocytes, mastcells, and airway smooth muscle that have important roles in asthma [2, 4, 8,27, 29]. Therefore, an increase in Nai possibly results in increased calciuminflux through Na+/Ca2+ exchange, increased activation of leukocytes, andbronchoconstriction. Sodium channel blockers reduce smooth muscle contrac-tion in vitro [20]. Of these, mexiletine and lidocaine have been shown to inhibitbronchoconstriction in asthmatic patients in vivo, and inhaled lidocaine hasbeen used to treat asthma, although the to exact mechanism of action is notclear [10].At any given Nai, a certain fraction of the maximal Na
+/K+-ATPaseenzyme activity is activated [32]. We hypothesized that the resting Na+/K+-ATPase activity, as measured by Rb uptake in intact cells, and the maximalNa+/K+-ATPase activity, as measured in cell homogenates, are related to eachother by Nai and that combined analysis of these three parameters and sodiuminflux would unify the disparate findings previously published. We furtherhypothesized that asymptomatic subjects with airway hyperresponsivenes(AHR) may also have abnormalities in their sodium homeostasis. Separateanalyses of cellular sodium homeostasis in normal subjects, asymptomaticsubjects with AHR, symptomatic asthma patients, and treated asymptomaticasthma patients may importantly explain the relationship of these abnormali-ties to AHR and asthma.
376 A. Agrawal et al.

Materials and Methods
Study Subjects and Design
A total of 53 subjects were recruited for this study from the outpatient asthma clinic of Vallabhbhai
Patel Chest Institute, New Delhi, and from volunteers. Informed consent was obtained from all
participants. Twenty-six had clinically and established asthma, showing AHR and reversible airway
obstruction. AHR was established when the concentration of histamine to produce a fall of 35% in
specific airway conductance (SGaw) (PD35hist) was less than 5 mg/mL [6]. We also recruited 12 subjects
with asymptomatic AHR and 15 normal subjects with normal airway responsiveness to histamine.
Asthma specialists examined each subject with asymptomatic AHR to exclude a clinical diagnosis of
asthma. This included absence of airway obstruction during body plethysmography, absence of cough
or nocturnal awakening, normal physical activity without shortness of breath, and normal health.
All subjects were tested for their SGaw. Subjects with clinically diagnosed asthma were subjected
to bronchoprovocation studies if AHR had not been previously documented. Asthmatic subjects were
divided into symptomatic and asymptomatic groups (asymptomatic was defined as no nocturnal
awakenings, two or less episodes of shortness of breath requiring use of rescue medication in the prior
4 weeks, no oral steroids, and no persisent cough or sputum).
After acceptance into the study, venous blood was collected and study parameters were measured
as described below. The overlap between the personnel responsible for subject enrollment and those
measuring study parameters did not permit adequate blinding. The sample size was based on achieving
a minimum of five different measurements for every study parameter for every group.
As stated in the Introduction, our study parameters were sodium influx, sodium efflux, resting
Na+/K+-ATPase activity in intact cells, and maximal Na+/K+-ATPase activity. Measured values
were compared to a simple mathematical model that we derived from existing literature [32]:
1. Na influx = Na efflux (homeostasis in intact cells)
2. Na influx = 3 · Resting Na+/K+-ATPase activity (since the primary driving of sodium efflux isNa+/K+-ATPase: 3Na+ pumped out per ATP hydrolyzed)
3. Resting Na+/K+-ATPase activity = Maximal Na+/K+-ATPase activity · Nai · k (since the
activated fraction (resting/maximal) of total Na+/K+-ATPase is constantly related to Nai by a
factor k)
4. Nai ¼ ðNa influx=Maximal Naþ Kþ �ATPase activityÞ x K ðsubstituting andrearranging between 2 and 3 such that K ¼ l/3 kÞ
This predicts that either an increase in sodium influx or a decrease in maximal Na+/K+-ATPase
activity will lead to an increase in Nai. We therefore also measured the two components of maximal
Na+/K+-ATPase activity, i.e. molecule number and molecule activity.
Measurement of Specific Airway Conductance (SGaw) and AirwayHyperresponsiveness
SGaw was measured in a constant-volume body plethysmograph by the quiet breathing technique [12].
Briefly, the patient was seated in the plethysmograph, and SGaw was calculated from oscilloscope
tracings of airflow and box-pressure signal. Airway responsiveness to histamine was measured by serial
inhalations of increasing concentrations of nebulized histamine, followed by SGaw measurement.
PD35hist was calculated by interpolation [6].
Isolation of Leukocytes
Mixed peripheral leukocytes were separated by the method of Boyum [5]. Thirty milliliters of blood
were collected by venipuncture and mixed with acid citrate dextrose anticoagulant solution containing
Abnormal Sodium Homeostasis in Leukocytes of Asthmatics 377

(9:1). The mixture was centrifuged at 5000 rpm at 4�C in a refrigerated centrifuge for 10 min. Afterseparation of plasma, 6% dextran reagent (1/3 v/v) was added for sedimentation of red blood cells
(RBC). The leukocyte-rich supernatant was then separated and centrifuged at 2000 rpm for 20 min.
Pelleted leukocytes were washed with 0.8% NH4CI and kept at room temperature for 5 min to lyse
contaminating RBC. They were then centrifuged at 2000 rpm and resuspended in respective buffers to
estimate resting or maximal ATPase activity, 3H-ouabain binding, sodium influx, Or Nai. The viability
of the cells was verified by trypan blue exclusion test to be more than 99%. The leukocyte content of the
cell suspension was greater than 95% as found by simple light microscopy.
Measurement of Maximal Na+/K+-ATPase Activity
ATPase activity was determined as described by Schmalzing and Kushera [28]. Leukocyte suspensions
were sonicated and an aliquot of homogenate corresponding to 100–150 lg of protein was incubated at37�C in an Eppendorf tube containing a total volume of 500:1 reaction mixture. For estimation ofMg2+–(Na+/K+)-ATPase activity, the reaction mixture contained 100 mM Tris-HCl (pH 7.4), 5 mM
MgCl2, 100 mM NaCl, 20 mM KCl, and 2 mM EGTA. For estimation of Mg2+-ATPase activity, the
reaction mixture contained 100 mM Tris-HCl (pH 7.4), 5 mM MgCl2, 100 mM NaCl, 2 mM EGTA
plus 100 lM ouabain. Reactions were started by addition of ATP (2 mM final concentration). After 30min, reactions were terminated by addition of 5% trichloroacetic acid and the contents of the tubes
were centrifuged at 4�C. For reaction controls, the homogenate was added after termination of thereaction. ATPase activity was determined by measuring the amount of inorganic phosphorus liberated
in each tube following the method of Le Bel et al. [16]. Activity of Na+/K+-ATPase was calculated
from the difference between quantity of Pi liberated in Mg2+–(Na+/k+)-ATPase and Mg2+-ATPase
activity. The mean interassay coefficient of variation of Na+/K+-ATPase activity on three similar
normal subjects determined at five different times was within 7%. The mean intra-assay coefficient of
variation of Na+/K+-ATPase activity in cells from the same subject was within 2%. The ouabain-
sensitive ATPase activity was considered equal to Na+/K+-ATPase activity. Since this assay is per-
formed under conditions optimal for ATPase activity, this reflects maximal ATPase capacity of the
Na+/K+-ATPase, expressed as nmol ATP hydrolyzed /106 cells/30 min at 37�C.
Measurement of Resting Na+/K+-ATPase Activity (86RbUptake)
Ouabain-sensitive 86Rb uptake was measured as described by Oh et al. [23]. After isolation, intact
leukocytes were washed and suspended in freshly prepared potassium-containing Ringer solution
(composition in mmol/L: 142 Na+, 1.25 Mg2+, 4.48 K+, 1.34 Ca2+, 127 Cl, 17 PO43), and 5.6
glucose). Cells were incubated at 37�C in Ringer buffer containing 8.3 lM 86Rb (final concentration)
with or without 1 mM ouabain to determine total uptake and ouabain-resistant uptake, respectively.
After 1 h incubation, cell-containing suspension was centrifuged, pellet was washed multiple times with
potassium-free Ringer solution at 4�C, and the tubes were drained of excess water. Distiller water(200 lL) was added to each tube and Cerenkov radiation was counted for 10min in a liquid scintil-lation counter. Ouabain-sensitive 86Rb uptake was calculated by subtracting ouabain-resistant uptake
from total uptake. The coefficient of variation in ouabain-sensitive 86Rb uptake in the same sample was
not more than 5%. Ouabain-sensitive uptake equals the K+ uptake activity of Na+/K+-ATPase. Since
two potassium ions are internalized per ATP consumed by Na+/K+-ATPase, the measured value was
divided by two to convert it to ATPase activity.
Measurement of Specific 3H–Ouabain Binding
Specific 3H–ouabain binding in the intact leukocytes was estimated as described by Oh et al. [23]. After
isolation, intact leukocytes were washed once and then resuspended at a density of 2 · 106 cells/mL in a
378 A. Agrawal et al.

freshly prepared potassium-free Ringer solution (composition in mM: 142 Na+, 1.25Mg2+, 1.34 Ca2+,
127 Cl), 17 PO43) and 5.6 glucose). Cells were incubated with 25 nM [3H]–ouabain for 2 h. Incubation
was stopped by centrifugation at 8000 rpm for 3 min. Cell pellet was washed four times to remove
extracellular bound label, after which retained radioactivity was counted in a liquid scintillation
counter. Specific ouabain binding sites were calculated by subtracting non specific 3H–ouabain binding
sites obtained in the presence of 5 mM digoxin from total 3H–ouabain binding sites (in the absence of
digoxin). Ouabain binding sites were expressed in fmol/106 cells. Each sample was run in triplicate. The
coefficient of variation in ouabain binding sites in the same sample was in the range of 4–7%.
Measurement of Sodium Influx
Sodium influx was measured using the method of Simcowitz et al. [30]. After isolation, leukocytes were
suspended in 5 mM Tris buffer (pH 7.4) containing (in mM) 140 NaCl 5 KCl, 1 CaCl2, 0.5 MgCl2, 5.6
glucose, and 1mg/mL bovine serum albumin at a concentration of 106/1.8 mL. They were pre-equili-
brated for 1 h at 37�Cbefore addition of 22NaCl (4.5 lCi/mL). Following 30min of incubation, a 0.5mLaliquot was removed and layered on 0.7 mL silicone oil and centrifuged at 10,000 rpm. The aqueous and
oil phases were carefully aspirated after which radioactivity retained in the leukocyte pellet was counted
in the gamma counter. In parallel, controls were run in each experiment in which leukocytes were pelleted
immediately after the addition of the radioactivity (for uptake at 0 min). Na+ uptake was calculated by
subtracting the influx at 0 min from that which occurred at 30 min. Na+ uptake by leukocytes was
expressed as pmol/min/106 cells.
Measurement of Nai
Nai was determined as described byHartoonium et al. [11]. In brief, leukocytes were isolated as described
above. The leukocyte pellet was resuspended in potassium-containing Ringer�s solution and incubatedwith the dye SBFI-AM (Sodium Binding Fluorescent Indicator, Sigma Chemicals) per manufacturer�sprotocol at 37�C. The cells were then washed and the fluorescence was measured in triplicate. The intra-assay coefficient of variation of mean Nai in cells from the same subject was within 5%.
Analysis
The parameters measured above were grouped as per the protocol. The four groups, i.e., normore-
active normals (N), hyperreactive normals (HN), asymptomatic asthmatic patients (AP), and symp-
tomatic asthmatic patients (SP) were analyzed as described below for differences in each of the
parameters under study. Because of non-normal, nonhomogenous subpopulations, nonparametric
analysis based on ranks was carried out. Kruskal–Wallis (K–W) statistic was determined for all groups
and, if found significant (p< 0.05), the Mann–Whitney test was applied to compare any two groups.
Differences were considered significant if p £ 0.0086, to provide an effective a value of about 0.05(Bonferroni�s correction for multiple comparisons). No analysis was performed for parameters indi-rectly calculated from the measured means.
Results
General
A total of 53 subjects were recruited. Of these 15 were normal subjects with nohyperresponsiveness (N), 12 were normal subjects with airway hyperresponsive-
Abnormal Sodium Homeostasis in Leukocytes of Asthmatics 379

ness (HN), 15 were patients with symptomatic asthma (SP), and 11 wereasymptomatic asthmatic patients (AP). The populations were similar for age andgender. All subjects were otherwise healthy. Patients with symptomatic asthmahad airway obstruction as measured by a reduced airway conductance(SGaw = 0.07 ± 0.02), different from all other groups, which were comparable(Table 1). It was not possible to measure all parameters in every subject, butmeasurements in triplicate in at least five different subjects were made for everyparameter other than directly measured sodium influx. Sodium influx was notdirectly measured in more subjects because of the duplicate nature of the infor-mation. Since resting Na+/K+-ATPase activity determines sodium efflux that istheoretically the same as sodium influx, assuming equilibrium (see Methods), wedid not perform any further direct measurements of sodium influx after con-firming this quantitatively in three subjects.
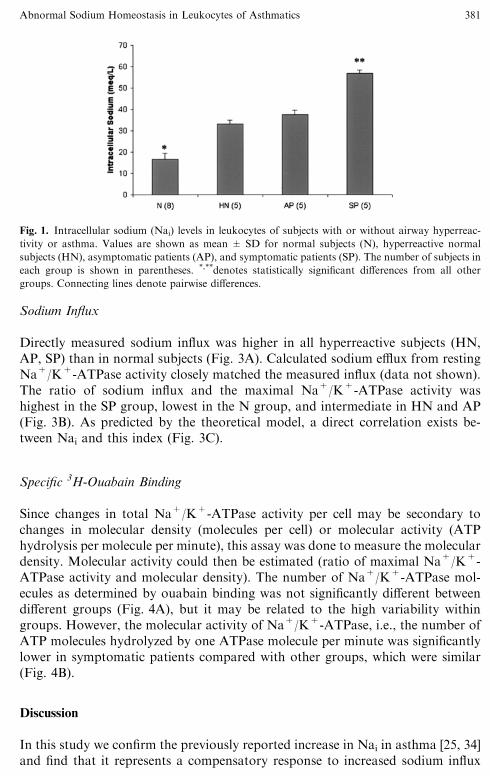
Intracellular Sodium
The distribution of Nai in each patient group is shown in Figure 1. All hyperre-active subjects (HN, AP, SP) had significantly elevated Nai (mmol/L,mean ± SD) when compared with normals (N, 17 ± 3). Among them, symp-tomatic patients had significantly higher levels than others (SP, 56 ± 2). Inasymptomatic patients and hyperreactive normals, the levels were similar (AP,37 ± 2; HN, 33 ± 2).
Resting and Maximal Na+/K+-ATPase Activity
The resting Na+/K+-ATPase activity was measured as an ionic flux and ex-pressed as ATPase activity for comparison (see Methods). All hyperreactivesubjects (HN, AP, SP) had similarly elevated resting Na+/K+-ATPase activity(Fig. 2A), indicating higher sodium efflux. Maximal activity of Na+/K+-ATPasewas similar in all groups except SP, in which it was reduced by about half(Fig. 2B). The activated fraction of Na+/K+-ATPase was calculated as the ratioof resting and maximal activity (percent activation). It was highest in symptom-atic patients (SP), lowest in normal subjects (N), and similar in hyperreactivenormals (HN) and asymptomatic patients (AP) (Fig. 2C). This pattern was sim-ilar to that of Nai.
Table 1. Comparison of age and specific airway conductance (SGaw) between groups
Normal Hyperreactive normal Asymptomatic patients Symptomatic patients
Age 26 ± 2 25 ± 3 26 ± 7 22 ± 5
SGaw 0.21 ± 0.03 0.20 ± 0.07 0.15 ± 0.02 0.07 ± 0.02*
*Statistically significant differences (p < 0.009). All values are shown as mean ± SD.
380 A. Agrawal et al.
Sodium Influx
Directly measured sodium influx was higher in all hyperreactive subjects (HN,AP, SP) than in normal subjects (Fig. 3A). Calculated sodium efflux from restingNa+/K+-ATPase activity closely matched the measured influx (data not shown).The ratio of sodium influx and the maximal Na+/K+-ATPase activity washighest in the SP group, lowest in the N group, and intermediate in HN and AP(Fig. 3B). As predicted by the theoretical model, a direct correlation exists be-tween Nai and this index (Fig. 3C).
Specific 3H-Ouabain Binding
Since changes in total Na+/K+-ATPase activity per cell may be secondary tochanges in molecular density (molecules per cell) or molecular activity (ATPhydrolysis per molecule per minute), this assay was done to measure the moleculardensity. Molecular activity could then be estimated (ratio of maximal Na+/K+-ATPase activity and molecular density). The number of Na+/K+-ATPase mol-ecules as determined by ouabain binding was not significantly different betweendifferent groups (Fig. 4A), but it may be related to the high variability withingroups. However, the molecular activity of Na+/K+-ATPase, i.e., the number ofATP molecules hydrolyzed by one ATPase molecule per minute was significantlylower in symptomatic patients compared with other groups, which were similar(Fig. 4B).
Discussion
In this study we confirm the previously reported increase in Nai in asthma [25, 34]and find that it represents a compensatory response to increased sodium influx
Fig. 1. Intracellular sodium (Nai) levels in leukocytes of subjects with or without airway hyperreac-
tivity or asthma. Values are shown as mean ± SD for normal subjects (N), hyperreactive normal
subjects (HN), asymptomatic patients (AP), and symptomatic patients (SP). The number of subjects in
each group is shown in parentheses. *,**denotes statistically significant differences from all other
groups. Connecting lines denote pairwise differences.
Abnormal Sodium Homeostasis in Leukocytes of Asthmatics 381

and decreased maximal ATPase activity of Na+/K+-ATPase. While our resultswere obtained in a mixed leukocyte population, which is predominantlyneutrophils, other investigators have described similar disturbances in sodiumhomeostasis in other cell types [9, 35]. In our study we found that Nai is related tothe ratio of sodium influx and maximal Na+/K+-ATPase activity, suggesting thatthis ratio is a useful index of the functional sodium loading of a cell (see Eq. 4 instudy design, and Fig. 3C).The disturbances in cellular sodium homeostasis found in asthma are also
found to a lesser degree in subjects with asymptomatic AHR. Patients withasymptomatic asthma had parameters of sodium homeostasis identical to those
Fig. 2. Relationship of resting and maximal capacities of Na+/K+-ATPase. Resting Na/K-ATPase
activity (A); maximal activity (B); and percent activation of Na/K-ATPase, i.e., resting activity as
percent of maximal activity (C) in leukocytes of the subject groups as described in Figure 1.
382 A. Agrawal et al.

with asymptomatic AHR. In our study, increased sodium influx and Nai is seen inAHR irrespective of symptoms, while a reduction in maximal Na+/K+-ATPaseactivity is associated with symptoms of asthma, presumably through potentiatingthe increase in Nai. It is intriguing that biochemical changes in normal subjectswith AHR and patients with asthma are similar. Laprise and Boulet [14] reportedthat 4 of 28 normal subjects with AHR developed asthma over a three-yearperiod. Asymptomatic AHR was also associated with airway inflammation andremodeling [15]. Asymptomatic family members of asthma patients have both ahigher likelihood of AHR and of developing asthma [13]. Combining these data
Fig. 3. Relationship of sodium influx and calculated efflux capacity to Nai. Sodium influx (A); ratio of
measured influx to calculated maximal efflux capacity (B); and XY plot of mean Nai to the ratio of
mean sodium influx and calculated efflux capacity (C) in leukocytes of the subject groups as described
in Figure 1. Statistical tests of significance were not performed on extrapolated measurements (B, C).
Abnormal Sodium Homeostasis in Leukocytes of Asthmatics 383

with our biochemical findings, it is likely that asymptomatic AHR is a pre-asthmastate.The reason for asthma to be associated with disturbances in sodium
homeostasis is unknown. However, Na+/Ca2+ exchange has been known to bean important regulator of Ca2+ influx during secretion by neutrophils and lym-phocytes. Thus, an increase in Nai reduces the transmembrane Na
+ gradient,enhances Ca2+ influx, and stimulates secretion [1, 24]. Mast cell secretion hasbeen shown to similarly depend on Na+/Ca2+ exchange [27]. It is, therefore,likely that increased Nai results in activation of leukocytes, with increased releaseof cytokines and other inflammatory mediators. Feedback effects of these prod-ucts on sodium influx could initiate a self-sustaining cycle (see below).The molecular mechanisms of increase in sodium influx or reduction in Na+/
K+-ATPase activity are not clear. Tribe et al. [34] have reported that serum fromsubjects showing AHR causes increased sodium influx and rise in Nai inleukocytes from normal subjects. The nature of this serum-borne factor remainsunknown. It is expected that since increased influx occurs in the presence ofincreased Nai and, therefore, a decreased Na
+ gradient across the cell membrane,it must be a result of increased membrane permeability to Na+. Lys-ophosphatidylcholine (LPC), which consistently has been found to be elevated inthe serum of asthmatic subjects [7, 19] and is also generated in their airwaysduring antigen challenge [9], is known to increase cell membrane permeability in
Fig. 4. Functional characteristics of Na+/K+-ATPase activity. Molecular number (A) and molecular
activity (B) of Na+/K+-ATPase in leukocytes of various subject groups, as described in Figure 1.
384 A. Agrawal et al.

various tissues [3, 17, 22] and is positively correlated with AHR [19, 21]. Recently,LPC has been reported to open nonselective cation channels in myocytes that aremore permeable to Na+ than Ca2+ [18]. LPC also decreases the affinity of Na+
binding site of Na+/K+-ATPase for Na+ [24], resulting in reduced activity ofNa+/K+-ATPase for any given concentration of Na+, possibly accounting forour observation of reduced maximal Na+/K+-ATPase activity in symptomaticasthmatic patients. Interleukin-13, known to be important in asthma [35], in-creases cytosolic phospholipase A2 (cPLA2) expression [26] that may furtherregulate Na+ channel activity [33].There were some limitations to this study. The majority of asymptomatic
asthmatic subjects in our study were receiving anti-asthma medications other thanoral glucocorticoids. Withholding their medications was not appropriate and wetherefore did not obtain measurements off medications. Also, we did not furtherdefine our nonasthmatic asymptomatic subjects with AHR. Such patients havealready been studied in detail regarding incidence of asthma, atopy, rhinitis, andother similar parameters [13–15].We conclude that AHR with or without asthma is associated with increased
sodium influx and Nai in leukocytes. Symptomatic asthma is additionally asso-ciated with reduction in maximal activity of Na+/K+-ATPase, resulting in re-duced capacity to handle the increase in sodium influx and severe elevations inNai. Previous reports of increased activity of Na
+/K+-ATPase in asthma areattributable to the resting activity of Na+/K+-ATPase being measured. That is acompensatory response to the increased sodium influx achieved at the expense ofhigher Nai and reflects failure rather than efficiency of cellular sodium homeo-stasis.
Acknowledgments. This work was funded by the Council of Scientific and Industrial Research, India.
The authors thank Ramveer Singh for technical assistance.
References
1. Agrawal KP, Mehta D, Gupta S, Chhabra SK (1986) Metabolic basis for asthma and rhinitis: an
integrated approach. Lung 164(5):279–291
2. BalasubramanyamM, Rohowsky–Kochan C, Reeves JP, Gardner JP (1994) Na+/Ca2+ exchange-
mediated calcium entry in human lymphocytes. J Clin Invest 94(5):2002–2008
3. Bierbaum TJ, Bouma SR, Huestis WH (1979) A mechanism of erythrocyte lysis by lys-
ophosphatidylcholine. Biochim Biophys Acta 5551:102–110
4. Blaustein MP, Lederer WJ (1999) Sodium/calcium exchange: its physiological implications.
Physiol Rev 79(3):763–854
5. Boyum A (1968) Separation of leukocytes from blood and bone marrow. Introduction. Scand. J
Clin Lab Invest (Suppl) 97:7
6. Chai H, Farr RS, Froehlich LA, et al. (1975) Standardization of bronchial inhalation challenge
procedures. J Allergy Clin Immunol 564:323–327
7. Chhabra SK, Khanduja A, Jain D (1999) Increased intracellular calcium and decreased activities
of leucocyte Na+/K+-ATPase and Ca2+-ATPase in asthma. Clin Sci (Lond) 975:595–601
8. Chideckel EW, Frost JL, Mike P, Fedan JS (1987) The effect of ouabain on tension in isolated
respiratory tract smooth muscle of humans and other species. Br J Pharmacol 923:609–614
Abnormal Sodium Homeostasis in Leukocytes of Asthmatics 385

9. Chilton FH, Averill FJ, Hubbard WC, et al. (1996) Antigen-induced generation of lyso-phos-
pholipids in human airways. J Exp Med 1835:2235–2245
10. Groeben H, Foster WM, Brown RH (1996) Intravenous lidocaine and oral mexiletine block reflex
bronchoconstriction in asthmatic subjects. Am J Respir Grit Care Med 1544 Pt 1:885–888
11. Harootunian AT, Kao JP, Eckert BK, Tsien RY (1989) Fluorescence ratio imaging of cytosolic
free Na+ in individual fibroblasts and lymphocytes. J Biol Chem 26432:19458–19467
12. Krell WS, Agrawal KP, Hyatt RE (1984) Quiet-breathing vs. panting methods for determination
of specific airway conductance. J Appl Physiol 576:1917–1922
13. Laprise C, Boulet LP (1996) Airway responsiveness and atopy in families of patients with asthma.
Clin Invest Med 196:461–469
14. Laprise C, Boulet LP (1997) Asymptomatic airway hyperresponsiveness: a three-year follow-up.
Am J Respir Crit Care Med 1562 Pt 1:403–409
15. Laprise C, Laviolette M, Boutet M, Boulet LP (1999) Asymptomatic airway hyperresponsiveness:
relationships with airway inflammation and remodelling. Eur Respir J 141:63–73
16. LeBel D, Poirier GG, Beaudoin AR (1978) A convenient method for the ATPase assay. Anal
Biochem 851:86–89
17. Lindahl M, Hede AR, Tagesson C (1986) Lysophosphatidylcholine increases airway and capillary
permeability in the isolated perfused rat lung. Exp Lung Res 111:1–12
18. Magishi K, Kimura J, Kubo Y, Ablko Y (1996) Exogenous lysophosphatidylcholine increases
non-selective cation current in guinea-pig ventricular myocytes. Pflugers Arch 4322:345–350
19. Mehta D, Gupta S, Gaur SN, Gangal SV, Agrawal KP (1990) Increased leukocyte phospholipase
A2 activity and plasma lysophosphatidyicholine levels in asthma and rhinitis and their relationship
to airway sensitivity to histamine. Am Rev Respir Dis 1421:157–161
20. Nakashima Y, Sugiyama S, Shindoh J, et al. (1990) Effects of sodium channel blockers on elec-
trical field stimulation-induced guinea-pig tracheal smooth muscle contraction. Arch Int Phar-
macodyn Ther 306:130–138
21. Nath P, Joshi AP, Agrawal KP (1983) Biochemical correlates of airway hyperreactivity in guinea
pigs: role of lysophosphatidyl choline. J Allergy Clin Immunol 72(4):351–358
22. Niewoehner DE, Rice K, Sinha AA, Wangensteen D (1987) Injurious effects of lys-
ophosphatidylcholine on barrier properties of alveolar epithelium. J Appl Physiol 635:1979–
1986
23. Oh VM, Taylor EA, Ding JL, et al. (1987) Enhancement of specific [3H]ouabain binding and
ouabain sensitive 86rubidium influx in intact human lymphocytes by a dialysable factor in human
and fetal calf serum. Clin Sci (Lond) 721:71–79
24. Oishi K, Zheng B, Kuo JF (1990) Inhibition of Na, K-ATPase and sodium pump by protein kinase
C regulators sphingosine, lysophosphatidylcholine, and oleic acid. J Biol Chem 2651:70–75
25. Orlov SN, Baranov IA, Pokudin NI, Kubatiev AA, Chuehalin AC (1991) The transport of
monovalent ions and calcium in the erythrocytes of patients with bronchial asthma. Vestn Akad
Med Nauk SSSR 3:43–49
26. Rey A, M�Rini C, Sozzani P, et al. (1998) IL-13 increases the cPLA2 gene and protein expressionand the mobilization of arachidonic acid during an inflammatory process in mouse peritoneal
macrophages. Biochim Biophys Acta 13932–3:244–252
27. Rumpel E, Pilatus U, Mayer A, Pecht I (2000) Na(+)-dependent Ca(2+) transport modulates the
secretory response to the Fcepsilon receptor stimulus of mast cells. Biophys 796:2975–2986
28. Schmalzing G, Kutschera P (1982) Modulation of ATPase activities of human erythrocyte
membranes by free fatty acids or phospholipase A2. J Membr Biol 691:65–76
29. Simchowitz L, Cragoe EJ Jr (1998) Na+–Ca2+ exchange in human neutrophils. Am J Physiol 2541
Pt 1:C150–C164
30. Simchowitz L, Spilberg I, De Weer P (1982) Sodium and potassium fluxes and membrane potential
of human neutrophils: evidence for an electrogenic sodium pump. J Gen Physiol 79(3):453–479
31. Skoner DP, Gentile D, Evans R (1990) Decreased activity of the platelet Na+/K(+)-adenosine
triphosphatase enzyme in allergic subjects. J Lab Clin Med 115(5):535–540
32. Skou JC (1988) The Na, K-pump. Methods Enzymol 156:1–25
33. Smith PR, Fuller CM, Bubien JS, Bones DJ (2001) Focus on ‘‘contrasting effects of cPLA2 on
epithelial Na+ transport’’. Am J Physiol Cell Physiol 2811:C12–C14
386 A. Agrawal et al.

34. Tribe RM, Barton JR, Poston L, Burney PC (1994) Dietary sodium intake, airway responsiveness,
and cellular sodium transport. Am J Respir crit Care Med 149(6):1426–1433
35. Wills–Karp M, Luyimbazi J, Xu X, et al. (1998) Interfeukin-13: central mediator of allergic
asthma. Science 2825397:2258–2261
Accepted for publication: 12 May 2005
Abnormal Sodium Homeostasis in Leukocytes of Asthmatics 387