benthic marine ecosystems of great britain and the north-east
TRANSCRIPT

6 Benthic marine ecosystems in the north-eastAtlantic6.1 IntroductionDescriptive marine ecological studies undertaken inoffshore areas and in inshore areas of countries otherthan Great Britain in the north-east Atlantic (Figure 1)provide important information to:
◆ assist with the classification of marine habitats andcommunities of inshore areas in Great Britain, and
◆ compare against MNCR findings for Great Britain so asto provide a geographical context to the occurrence of
27
Hiscock: Introduction and Atlantic-European perspective
ESTUARINE HABITATS
Mud Sand Rock Zostera marina
Benthic FishesAnguilla anguillaNerophis lumbriciformisPholis gunnellusAgonus cataphractusPomotoschistus micropsPlatichthys flesusPleuronectes platessa
Epibenthic FishesSalmo truttaSalmo salarPollachius pllachiusTrisopterus minutusGasterosteus aculeatusMullus surmuletus
Benthic FishesAnguilla anguillaAgonus cataphractusPomatoschistus micropsPomatoschistus pictusPomatoschistus minutusPlatichthys flesusPleuronectes platessa
Epibenthic FishesSalmo truttaSalmo salarOsmerus eperlanusPollachius pollachiusTrisopterus minutusGasterosteus aculeatusDicentrachus labraxMullus surmuletusPholis gunnellus
Benthic FishesNeropis lumbriciformisSyngnathus acusSyngnathus typhleCyclopterus lumpusGobius nigerGobius paganellus
Epibenthic FishesLepadogaster lepadogasterPollachius pollachius (juv.)Spinachia spinachiaTaurulus bubalisLiparis liparisLiparis montaguiDicentrarchus labraxCentrolabrus exoletus (juv.)Crenilabrus melops (juv.)Ctenolabrus rupestris (juv.)Labrus bergylta (juv.)Gobiusculus flavescens
Benthic FishesLepadogaster lepadogasterSyngnathus acusSyngnathus typhleLiparis montagui
Epibenthic FishesGasterosteus aculeatusSpinachia spinachiaTaurulus bubalisCentrolabrus exoletus (juv.)Crenilabrus melops (juv.)Ctenolabrus rupestris (juv.)Labrus bergylta (juv.)Gobiusculus flavescens
LITTORAL HABITATS
Mud Sand Bedrock & boulders Rockpool & crevices
Benthic FishesAnguilla anguillaNerophis lumbriciformisAgonus cataphractusPomatoschistus micropsPlatichthys flesusPleuronectes platessa
Epibenthic FishesSalmo truttaSalmo salarPollachius pollachiusTrisopterus minutusGasterosteus aculeatusDicentrachus labraxMugilidaePholis gunnellus
Benthic FishesRajidae (juv.)Agonus cataphractusTrachinus viperaPomatoschistus minutusAmmodytidaeCallionymidaePsetta maximaScophthalmus rhombusLimanda limandaPlatichthys flesusPleuronectes platessaSolea solea
Epibenthic FishesOsmerus eperlanuaMullus surmuletusMugilidaeAmmodytidae*
Benthic FishesApletodon microcephalusLepadogaster lepadogasterNerophis lumbriciformisNerophis sp.Syngnathus acusSyngnathus typhleMyococephalus scorpiusTaurulus bubalisCyclopterus lumpusLiparis liparisLiparis montaguiCoryphoblennius galeritaLipophris pholisParablennius gattorugineGobius nigerGobius pagenellusZeugopterus punctatus
Epibenthic FishesSpinachia spinachiaCentrolabrus exoletusCrenilabrus melopsCtenolabrus rupestrisLabrus bergyltaGobiusculus flavescensThorogobius ephippiatus
Benthic FishesApledon microcephalusLepadogaster lepadogasterCiliata mustelaNerophis lumbriciformisNerophis sp.Myoxocephalus scorpiusTaurulus bubalisCyclopterus lumpusCoryphoblennius galeritaLipophrys pholisParablennius gattorugineGobius cobitisGobius nigerGobius paganellusZeugopterus punctatus
Epibenthic FishesCentrolabrus exoletusCrenilabrus melopsCtenolabrus rupestrisLabrus bergyltaGobiusculus flavescens
* Also present in the seabed (cont’d overleaf)
Table 2. Fish assemblages of benthic habitats. Derived from Potts & Swaby (1991) but not including species listed as uncommon. Nofish assemblages are listed for littoral and inshore gravel where very few species are associated with the gravel

habitats, communities and species and therefore oftheir importance at an international scale.
‘Inshore’ areas are taken as those encompassed withinabout 5 to 6 km offshore of low water on the open coastbut may include more extensive areas within shallow(taken as less than 50 m deep) bays or inlets (for instanceCardigan Bay in west Wales, the Bristol Channel insouth-west England) or enclosed by islands (forinstance, The Minch in north-west Scotland). Thehistorical importance of various studies undertakenoutside Great Britain has been referred to in the Section2. In the current section, special attention is given to theidentification of recent sources of data describing marinehabitats and communities comparable to those occurringin Great Britain. Review of the literature for areasoutside of Great Britain has necessarily been lessthorough than for our main study area.
Studies undertaken solely in the Baltic have not beenincluded in detail as its special features makedescriptions of the benthos less relevant for comparisonwith Great Britain.
A final section (6.21) gives a brief description of someMediterranean literature.
6.2 The North SeaStudies of the North Sea benthos in British waters canbe said to have started with the work of Davis (1923,1925) and Stephen (1923, 1933, 1934). Recentdescriptions based on physical conditions andcommunities of species associated with those conditionswere undertaken by Glémarec (1973), Dyer et al. (1983a),Basford & Eleftheriou (1988), Eleftheriou & Basford(1989) and Basford, Eleftheriou & Raffaelli (1989, 1990).Many of the papers describing North Sea benthos areincluded in the review of biological effects of human
activities (Rees & Eleftheriou 1989). Künitzer et al. (1992)combined the results of benthos sampling undertaken in1986 by participants of the Benthos Ecology WorkingGroup of the International Council for the Explorationof the Seas (ICES) with the results of Eleftheriou &Basford (1989) to produce descriptions of the benthicinfauna for the whole of the North Sea proper. There is agreat deal of localised work undertaken in the region ofoil exploration and production areas but usuallydescribed in limited-circulation reports (resultspublished in the scientific literature include those ofAddy et al. (1978), Hartley (1984) and Hartley & Bishop(1986)). The proceedings of an international symposiumon the ecology of the North Sea held in May 1988 werepublished in Volume 25 (parts 1 & 2) of the NetherlandsJournal of Sea Research. Most recently, the series ofAssessment Reports contributing to the North SeaQuality Status Report (QSR) (Anonymous 1993) includessummaries of information on the biology of the NorthSea.
The work of Glémarec (1973) is particularly importantin describing sediment benthos in the North Sea. Thedistribution of infralittoral, coastal and open sea étages(Glémarec 1973) together with the major macrofaunalcommunities indicated by Kingston & Rachor (1982) isillustrated in Figure 17.
The term ‘infralittoral’ in the terminology of Glémarec(1973) can be confused with the ‘infralittoral’ of rockysubstratum zonation where light penetration andconsequent algal coverage determines its extent.Nevertheless, the étages of Glémarec have been widelyused in describing level bottom (sediment) communitiesin the North Sea. The importance of depth and thermalstability is also suggested by later authors. For instance,studying epibenthic species, Dyer et al. (1983a) andFrauenheim et al. (1989) separate the North Sea benthic
28
Marine Nature Conservation Review: benthic marine ecosystems
SUBLITTORAL HABITATS
Inshore mud & sand Inshore bedrock, boulders,artificial substrate and wrecks
Inshore crevices Offshore seabed
Benthic FishesSquatina squatinaRajidaeLophius piscatoriusCepola rubescensTrachinus viperaCallionymidaeGobiidaePomatoschistus minutusLimanda limandaPleuronectes platessa
Epibenthic FishesScyliorhinus caniculaScyliorhinus stellarisGadus morhuaMelanogrammus aeglefinusTrisopterus luscus (juv.)Trispoterus minutus (juv.)TriglidaeMullus surmuletusAmmodytidae*
Benthic FishesConger congerGaidropsarus vulgarisRaniceps raninusMyoxocephalus scorpiusTaurulus bubalisLiparis liparisGobius nigerGobius paganellusThorogobius ephippiatusZeugopterus punctatus
Epibenthic FishesMolva molvaTrisopterus luscusTrisopterus minutusZeus faberCentrolabrus exoletusCrenilabrus melopsCtenolabrus rupestrisLabrus bergyltaLabrus mixtusGobiusculus flavescensBalistes carolinensis
Benthic/Epibenthic FishesConger congerLepadogaster lepadogasterGaidropsarus vulgarisMolva molvaRaniceps raninusTrisopterus luscusTrisopterus minutusLiparis liparisCentrolabrus exoletusCrenilabrus melopsCtenolabrus rupestrisLabrus bergyltaLabrus mixtusBlennius ocellarisParablennius gattorugineAnarhichas lupusChirolophis ascaniiThorogobius ephippiatusZeugopterus punctatusBalistes carolinensis
Benthic FishesRajidaeRaja clavataScophthalmidaePsetta maximaScophthalmus rhombusBothidaePleuronectidaeHippoglossus hippoglossusMicrostomus kittPleuronectes platessaSoleidaeSolea solea
Epibenthic FishesGadidaeGadus morhuaMelanogrammus aeglefinusMerlangius merlangusMolva molvaMerluccius merluccius
* Also present in the seabed
Table 2 (continued)

29
Hiscock: Introduction and Atlantic-European perspective
Figure 17. The distribution of infralittoral, coastal and open sea étages and of major macrofaunal communities in the North Sea.(After Glémarec 1973 and Kingston & Rachor 1982. Re-drawn from Mitchell 1987.)

regions at around 50 m depth. Adams (1987) separates‘offshore northern’, ‘offshore central’ and ‘offshoresouthern’ sectors of the North Sea according toplanktonic communities but with boundariesapproximately corresponding to the 50 m and 100 mdepth contours and therefore closely parallel to those ofthe Glémarec étages. Künitzer et al. (1992) identify aseparation of northern and southern assemblages alongthe 70 m depth contour with further separation at the 30,50 and 100 m depth contours as well as by sedimenttype.
The paper by Basford, Eleftheriou & Raffaelli (1990)summarises their work in the northern North Sea anddiscusses the separation of regions. Using TWINSPANanalysis separately on infauna and epifauna, theydescribe the main groupings including separation ofcoastal and offshore stations for infauna. The
distribution of the major faunistic groupings suggestedby the TWINSPAN analysis is illustrated in Figure 18.The taxa characteristic of the major divisions are shownin Table 3.
Basford, Eleftheriou & Raffaelli (1990) conclude thatthe major factors underlying the distribution andabundance of infauna and epifauna are related to depthand sedimentary characteristics with sediment moreimportant than depth for the infauna and depth ofgreatest importance for the epifauna. Although the tworegions they describe for infauna coincide with the‘north British coastal’ and ‘offshore northern’ sectors ofAdams (1987), similarity of a significant proportion ofthe infauna in those two regions led to the conclusionthat it was not justified to demarcate the northern andcentral parts of the North Sea at the 100 m depthcontour in the manner of Adams (1987) or Glémarec
30
Marine Nature Conservation Review: benthic marine ecosystems
Figure 18. Distribution of the main infaunal TWINSPAN groupings from Basford, Eleftheriou & Raffaelli (1990). Characteristic taxaare in Table 3.

(1973). The analysis of infaunal data for the whole of theNorth Sea undertaken by Künitzer et al. (1992) also usedTWINSPAN and came to somewhat differentconclusions regarding the species assemblages whichcould be identified (Figures 19 & 20).
Krönke (1992) has undertaken a detailed study of theDogger Bank and compared her results with those of workundertaken in the 1950s. Results from 1985 to 1987 revealedthree assemblages of species illustrated in Figure 21.
6.3 The English ChannelFew studies extend to the offshore areas of the EnglishChannel. The most detailed undertaken is that of Holme(1961, 1966), who sampled 311 stations by anchor dredgethroughout the Channel. He differentiated species into:
1. those with a general distribution;
2. western species;
3. west Channel species;
4. Cornubian (warm water) species;
5. Sarnian species (centred on the Channel Isles).
The seven animal communities he identified werebased on the classification of Jones (1950), and aresummarised below.
1. The boreal shallow-sand association. A shallow watercommunity with Arenicola marina, Nephtys sp., Tellina(now Angulus) tenuis, Donax vittatus, etc.
2. The boreal shallow-mud association. Petersen’s‘Macoma’ community.
3. The boreal offshore-sand association. Petersen’s ‘Venus’community. Characteristic molluscs and echinodermsinclude Cardium echinatum, Dosinia lupinus, Venusstriatula, (now Chamelea gallina), Gari fervensis, Abraprismatica, Echinocardium cordatum and Acrochnidabrachiata, with Callista chione, Tellina (now Fabulina)fabula, Mactra corallina (now Mactra stultorum) and Ensissiliqua mainly confined to shallow parts.
31
Hiscock: Introduction and Atlantic-European perspective
Infaunal TWINSPAN analysis
COASTAL STATIONS MAINLY OFFSHORE STATIONS
Ophelina neglectaSphaerosyllis bulbosaEchinocyamus pusillus
Thyasira spp.Prionospio mulibranchiata
Group 1 Group 2 Group 1 Group 2
Pisione remota Nucula tenuis Spiophanes bombyx Eriopisa elongataThyasira spp.Lumbrineris gracilisCeratocephale loveni
Epifaunal TWINSPAN analysis
COASTAL STATIONS MAINLY OFFSHORE STATIONS
PoriferaFlustra foliaceaHyas coarctatusBolocera tuediae
Asterias rubensAstropecten irregularisBrissopis lyrifera
Group 1 Group 2 Group 1 Group 2
Porifera TunicatesSpirontocaris lilljeborgi
Pagurus bernhardusCrangon allmanniSpatangus purpureusColus gracilis
Pennatula phosphorea
Table 3. Characteristic taxa of the major TWINSPAN divisions for infauna (see Figure 18) and epifauna. FromBasford, Eleftheriou & Raffaelli (1990).
Figure 19. Classification of stations from the ICES North SeaBenthos Survey combined with data from Eleftheriou &Basford (1989) by TWINSPAN, incorporating speciesabundances. (From Künitzer et al. 1992.) Indicator species aregiven in Figure 20.

4. Boreal offshore muddy-sand association. Petersen’s‘Echinocardium cordatum-Amphiura filiformis’community and ‘Abra’ community. Typical speciesinclude: Nucula turgida, Cyprina (now Arctica)islandica, Cardium echinatum, Dosinia lupinus, Abra alba,Abra prismatica, Phaxas pellucidus, Ensis ensis, Spisulasubtruncata, Lutraria lutraria, Corbula gibba, Dentalium(now Antalis) entalis, Aporrhais pespelecani, Philinequadripartita, Callianassa subterranea, Ophiura texturata(now Ophiura ophiura), Amphiura filiformis,Echinocardium cordatum, Cucumaria elongata,Leptosynapta inhaerens and Labidoplax digitata.
5. Boreal offshore mud association. Corresponds toPetersen’s ‘Brissopsis lyrifera-Amphiura chiajei’
community which is, however, not present in theChannel. Holme infers localised usually inshoreoccurrence for this community with very fewlocations for it identified. The most characteristicspecies were the echiuroid Maxmuelleria lankesteriwith the bivalve Saxicavella jeffreysi.
6. Boreal offshore gravel association. Characteristicspecies include Nucula hanleyi, Glycymeris glycymeris,Venus (now Circomphalus) casina, Venus (now Timoclea)ovata, Venerupis (now Tapes) rhomboides, Gari tellinella,Ensis arcuatus, Spisula elliptica, Lutraria angustior,Echinocyamus pusillus, Echinocardium flavescens,Spatangus purpureus, Amphioxus (now Branchiostoma)lanceolatus. Often associated with beds of thebrittlestar Ophiothrix fragilis on harder ground.
7. Boreal offshore muddy-gravel association. (Notrecognised by Jones 1950.) Commonest species arecertain burrowing crustacea, particularly Upogebiadeltaura, Upogebia stellata and more rarely Squilla (nowMeiosquilla) desmaresti. Molluscs include Nuculanucleus, Venus verrucosa, Turritella communis andGibbula magus. There are also the sipunculid wormsGolfingia elongata and Golfingia vulgaris, species of theholothurian Thyone and the burrowing sea anemoneMesacmaea mitchelli.
The distribution of the communities described byHolme (1966) is illustrated in Figure 22. Holme alsorecognised a number of faunistic boundaries in theChannel including separation of the Channel Isles faunafrom that of the English side of the Channel and ofabrupt boundaries associated with headlands,particularly Start Point, St Alban’s Head and theCotentin Peninsula with the main faunistic boundaryidentified with the boundary between the summer-stratified waters of the western Channel with theunstratified waters of the eastern Channel. A further
32
Marine Nature Conservation Review: benthic marine ecosystems
Figure 20. TWINSPAN classification incorporating species abundance data. (Adapted from Künitzer et al. 1992.)
Figure 21. Macrofaunal communities in 1985 to 1987 on theDogger Bank. The ten numerically dominant species from eachof three groupings identified by cluster analyses are listed.Based on Krönke 1992.

boundary area is suggested off Looe in Cornwall. Thesamples collected by Holme (1966) were further studiedfor their bryozoan populations by Grant & Hayward(1983), who identified three distinct assemblagescharacterised as ‘shallow’, ‘intermediate’ and ‘deep’.
Cabioch et al. (1977) used the results of dredgesampling to indicate distribution of hard substratumspecies in the English Channel (Figure 23). Offshoreareas of the English Channel were further investigatedusing towed video and still cameras by Holme & Wilson(1985). The area studied, about 37 km south of theDorset coast, was predominantly of hard substrata oftenwith transitory sand cover. The epifaunal assemblagesencountered were separated into:
6.4 The Celtic SeaThe Celtic Sea is the area to the south of Ireland andwest of Cornwall in south-west England. The mostextensive published survey of the benthic fauna of the
Celtic Sea is that undertaken in 1974 and 1975 by theField Studies Council Oil Pollution Research Unit(Hartley & Dicks 1977; Hartley 1979). The fauna at mostsites was typical of a ‘deep Venus community’ asdescribed by Mackie (1990) and included in the nextsection. At the edge of the Celtic Deep, the communitieswere typical of a ‘boreal deep mud association’ andincluded the brittlestars Amphiura chiajei and Amphiurafiliformis, the bivalves Nucula sulcata, Nucula tenuis,Thyasira flexuosa and Abra nitida, and polychaetesMyriochele heeri, Lagis (now Pectinaria) koreni andAmphicteis gunneri. Bryozoan species occurring on hardsubstratum in depths of 159 to 1582 m are recorded byHayward & Ryland (1978).
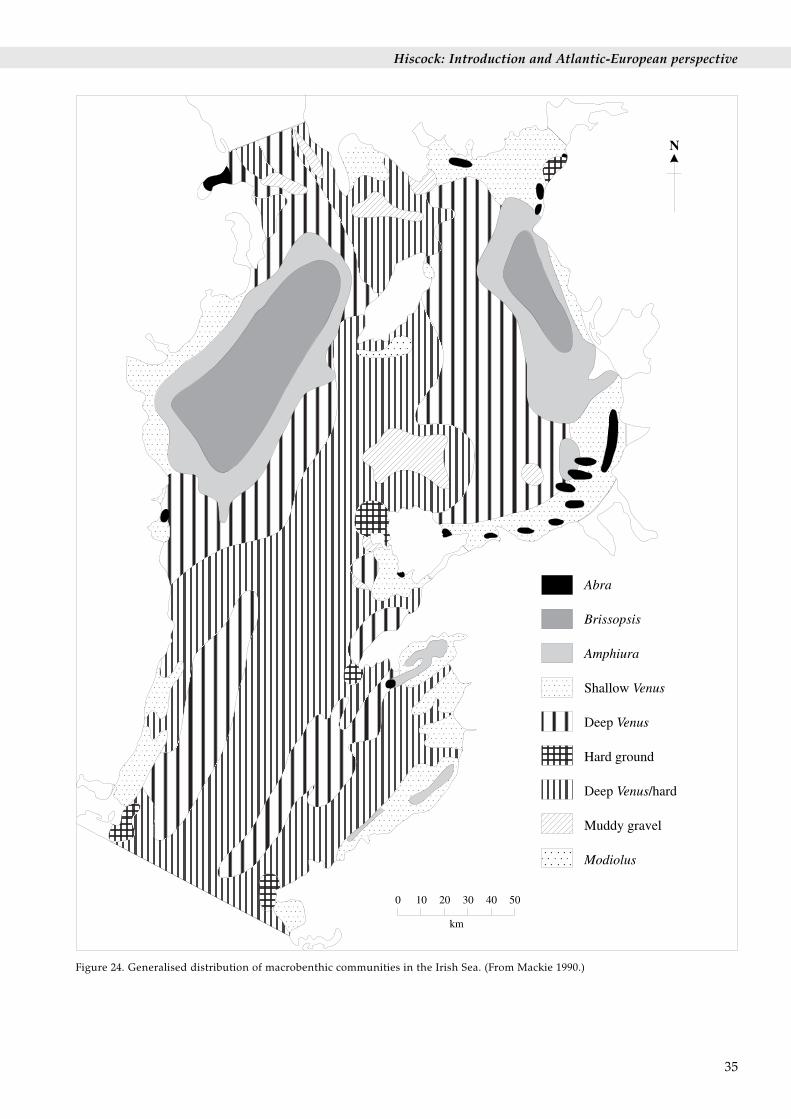
6.5 The Irish SeaPublished and unpublished information on the offshoresediment macrofaunal benthic communities of the IrishSea are brought together and their distribution mappedby Mackie (1990) (Figure 24). Nine separate communitytypes are noted:
1. The Amphiura community [the ‘Boreal offshoremuddy sand association’ of Jones 1950] present inoffshore sandy muds at shallow to moderate depths(15 m to 100 m) and typically including the brittle-starAmphiura filiformis, the urchin Echinocardium cordatumand the tower shell Turritella communis.
33
Hiscock: Introduction and Atlantic-European perspective
Figure 22. Distribution of benthic communities in the English Channel. (Re-drawn from Holme 1966.)
Type A Stable faunal assemblage with diverse spongecover;
Type B B1 Well developed faunal assemblage withPolycarpa violacea
B2 Impoverished Polycarpa violacea – Flustrafoliacea assemblage
B3 Impoverished Balanus – Pomatocerosassemblage;
Type C Cobble floor covered by sand.

2. The Brissopsis community [the ‘Boreal offshore mudassociation’ of Jones 1950] present in offshore mudsat shallow to moderate depths (15 m to 100 m) andtypically including the urchin Brissopsis lyrifera andthe brittle-star Amphiura chiajei.
3. The Abra community [included in the ‘Boreal offshoremuddy sand association’ of Jones 1950] present inshallow (5 m to 30 m) nearshore muddy sands/mudswith rich organic content and typically including thebivalve mollusc Abra alba and the polychaete wormPectinaria koreni.
4. The Shallow Venus community [the ‘Boreal offshoresand association’ of Jones 1950] present in shallow (5m to 40 m) nearshore sands. There are two sub-communities. The ‘Tellina sub-community’ occurs infine sands and typically includes the bivalve Tellina(now Fabulina) fabula and the polychaete Magelonamirabilis. The Spisula sub-community occurs inmedium to coarse sands subject to disturbance andtypically includes the bivalve Spisula elliptica and thepolychaete Nephtys cirrosa.
5. The Deep Venus community [the ‘Boreal offshoregravel association’ of Jones 1950] occurs in coarsesand/gravel/shell sediments at moderate depths (40 mto 100 m) and typically includes the urchin Spatanguspurpureus and the bivalves Glycymeris glycymeris,Astarte sulcata and Venus spp.
6. The muddy-gravel community [referred to in relationto the ‘Boreal offshore muddy-gravel association’ of
Holme (1966)] includes very rich faunas from mixedmuddy gravels.
7. The Modiolus community [part of the ‘Boreal offshoregravel association’ of Jones 1950] occurs on coarsesand/gravel or shell/stone substrata at moderatedepths and typically includes the horse musselModiolus modiolus and the brittle-star Ophiothrixfragilis and the mussel clumps attract a rich fauna.
8. Hard substratum communities.
Mackie, Oliver & Rees (1995) describe the results offurther sampling undertaken in 1989 and 1991. Theyconclude that the southern Irish Sea can be said to be partof the boreal zoogeographical province but with moresouthern lusitanian influences in area of the Celtic Deep atthe southern entrance to the Irish Sea. They identify amosaic of loose overlapping assemblages with three majortypes corresponding to general sediment distribution:
1. Assemblage A occurred in the deeper mud and sandymud regions.
2. Assemblage B was found in the inshore sandy andmuddy sand areas.
3. Assemblage C coincided with the offshore gravelysediments.
The frontal systems in the Irish Sea (Pingree &Griffiths 1978; Figure 7) are important areas for planktonproductivity and for the marine species which
34
Marine Nature Conservation Review: benthic marine ecosystems
Figure 23. Eastern boundaries of epibenthic species in the English Channel (from Cabioch et al. 1977). A = Porella compressa; B =Diphasia pinaster; C = Thuiaria articulata; D = Lafoea dumosa; E = Caryophyllia smithii; F = Sertularella gayi: G = Rhynchozoonbispinosum.
35
Hiscock: Introduction and Atlantic-European perspective
Figure 24. Generalised distribution of macrobenthic communities in the Irish Sea. (From Mackie 1990.)

congregate there to feed and may also affectproductivity of the benthos and biogeography.
6.6 The Faeroe IslandsThe marine algal flora of the Faeroes has been studiedsince the late 18th century. Most importantly for the useof the MNCR, distinct species assemblages weredescribed by Børgesen (1908). Børgesen’s assemblageshave provided a basis for classification and comparisonof algal communities in southern Ireland (Cotton 1912),the Netherlands (den Hartog 1959) and in Great Britain(for example, Tittley, Irvine & Jephson 1976). A series ofpapers (Irvine 1982; Tittley, Farnham & Gray 1982; Price& Farnham 1982) describe the seaweeds of the Faeroesincorporating and comparing the much earlier work ofBørgesen. Another series of papers entitled The zoology ofthe Faeroe Islands published in Copenhagen provides listsof shore and shallow sublittoral species. In the Faeroes,the species present are, with few exceptions, thosefound in Great Britain although the Faeroesecommunities include a lower number of species. Inrecent years, a wide-ranging study of the deeper watercommunities around the Faeroes has been undertaken(the BioFar programme: Nörrevang et al. 1996) and thisis now continuing inshore. The distribution and ecologyof the reef-building coral Lophelia pertusa on the shelfbreak around the Faeroe Islands has been especially wellstudied as a part of the BIOFAR programme. Jensen &Frederiksen (1992) describe the fauna associated withsamples of Lophelia. A total of 298 species was recorded,predominantly Porifera, Polychaeta, Bivalvia, Crustacea,Bryozoa, Echinodermata and Brachiopoda. Theassociated fauna, like the coral itself, was of suspensionfeeders suggesting the importance of water movementto the community.
6.7 NorwayThe coastline of Norway (Figure 25) extends from about58°N to 71°N and, at just north of 60°N, Bergen is on thesame latitude as Lerwick in Shetland. Much of thecoastline is exposed to very strong wave action butfjords and offshore islands create many very extensivewave-sheltered areas. Studies of marine biology inNorway extend back into the early 19th century with thepioneering work of Michael Sars. Much of this historicalwork is referred to in the review given by Gislén (1930)and in Brattström (1967).
Little published descriptive survey work has beenundertaken between Tromsø and Trondheim along thenorthern part of the Norwegian coastline. However,communities recorded from a submarine gully near toTromsø (Gulliksen 1978) are very similar to those in suchgullies in Britain. Other studies by Holte & Gulliksen(1987) on sediment communities and by Jakola &Gulliksen (1987) on hard substratum communities onjetty pilings provide further comparative data from theTromsø area. The species living in sediments are similarto those which would be encountered in similarsituations in Great Britain. However, the most abundantlarge species found on jetty piles were arctic or arcticboreal; for instance, the ascidians Styela rustica, Ascidiacallosa and Halocynthia pyriformis. South of Tromsø, the
characteristics of marine communities change with thedominance of arctic or arctic-boreal species giving wayto species with a more southern distribution which alsooccur in Britain. One key species which reaches itsnorthern limit at about the Lofoten Islands is the limpetPatella vulgata.
Farther south, Borgenfjorden near the head ofTrondheimfjord has been particularly thoroughlystudied for hard substratum sublittoral communities(papers listed in Gulliksen 1980) and for littoralsediments (Strömgren, Lande & Engen 1973) revealingsimilar communities to those present in enclosed areason the west coast of Scotland and Shetland.
The main centre for the study of benthic communitieshas been Bergen with the establishment of laboratoriesat the coast by the Bergen museum. A historical accountof work undertaken there and of habitats and fauna isgiven by Brattström (1967). Jorde (1975) reviews thepapers describing the ecology and distribution of algaein western Norway. The paper by Jorde (1966) whichdescribes 38 algal associations in the coastal area southof Bergen is particularly important for comparison ofcommunities with those found in Britain. Fjords near toBergen have also provided the opportunity to studyeffects of environmental gradients along their lengthincluding, for instance, on rocky shores inHardangerfjord (Jorde & Klavestad 1963, Figure 26;Brattegard 1966). These two papers further providedescriptions of the littoral and shallow sublittoralcommunities occurring in Hardangerfjord. Descriptionsof algal communities are particularly useful forcomparison with those present in Britain and includeaccounts of 25 associations. Many of the features ofspecies distribution along fjords are comparable withthose in sealochs both in terms of decreasing exposureto wave action and where large volumes of freshwaterenter at the head of the inlet. Tunberg (1982) describesthe communities of Raunefjorden near Bergen. The twocommunities (characterised by the molluscs Thraciavillosiuscula and Dosinia borealis and by Lucinoma borealisand Thyasira flexuosa) described from there may be ofparticular interest as they do not correspond to any ofthose described by Thorson (1957). On the open coasthere, marine communities include many more Arcticelements than in British waters at the same latitude. Forexample, the sea urchin Echinus acutus has not yet beenrecorded in British coastal waters but is abundant inshallow depths near Bergen and Strongylocentrotusdroebachiensis is extremely abundant in Norwaycompared with the isolated populations of Shetland.
The fjords of western Norway provide opportunitiesfor the study of deep isolated environments andBrattström (1967) comments that there must be fewplaces where one sails inland to sample the deep-seafauna. The communities present include onescharacterised by the pogonophoran Siboglinum ekmaniwith large foramaniferans present (Brattegard 1967) aswell as communities associated with the reef-formingcoral Lophelia pertusa (Tambs-Lyche 1958). Sognfjord,with a maximum depth of 1,300 m, includes a deep seafauna similar to that of the north-westernMediterranean (Carpine 1970). Although not occurringin inshore waters in Britain, some of these communities
36
Marine Nature Conservation Review: benthic marine ecosystems

37
Hiscock: Introduction and Atlantic-European perspective
LofotenIslands
Figure 25. Norway, Sweden and Denmark, showing locations of places mentioned in the text.

38
Marine Nature Conservation Review: benthic marine ecosystems
Rhodymenia Palmaria palmata(now )
Ceramium rubrum Ceramium nodulosum(now )
Phyllophora membranifiolia Phyllophora pseudoceranoides(now )
Polysiphonia nigrescens (now Polysiphonia fucoides)
Polysiphonia violacea (now Polysiphonia fibrillosa)
Gigartina stellata Mastocarpus stellatus(now )
Himanthalia elongata
Alaria esculenta
Laminaria digitata f. stenophylla
Callophyllis laciniata
Ptilota plumosa
Delesseria sanguinea
Euthora cristata
Laminaria hyperborea
Lometaria clavellosa
Phycodrys rubens
Chylocladia verticillata
Pelvetia canaliculata
Polysiphonia elongata
Rhodomela confervoides
Fucus spiralis
Laminaria digitata
Laminaria saccharina
Ahnfeltia plicata
Sphacelaria plumosa
Polyides rotundus
Fucus serratus
Ascophyllum nodosum
Fucus vesiculosus
Figure 26. The horizontal distribution of selected species in Hardangerfjord and Sörfjord. (From Jorde & Klavestad 1963.)

may occur at considerable depths off the British coasts.Siboglinum ekmani has been found in the westernapproaches to the English Channel (Southward &Southward 1958) and in The Minch (A. McIntyre, pers.comm.) and Lophelia pertusa is known from the RockallBank and Hebrides.
Polls are marine inlets which are partly cut-off fromthe open sea by, for instance, shallow sills and are awidespread and particular feature especially in the areaof Bergen. They have hydrographic characteristics andcommunities similar to larger brackish water obs inScotland. Lindaspollene north of Bergen are an exampleof this type of habitat described by Taasen & Evans(1977) and subsequent papers (for example, Evans 1981,Taasen & Høisaeter 1989).
The abundance of species on rocky shores of widelydifferent exposures was described in the Fensfjord area ofwestern Norway in relation to studies of effects of waveexposure and the production of a biologically definedexposure scale (Dalby et al. 1978). These records help torelate community descriptions developed for Britain withthose in other parts of the north-east Atlantic.
The marine habitats and associated communities ofplants and animals present on the coast of southernNorway often appear to be described incidentally toother studies, particularly those monitoring effects ofpollution. Sediment communities in Oslofjord aredescribed in detail by Mirza & Gray (1981). Theydescribe six site-species groupings and similar trendsand species composition in relation to organicenrichment described in Loch Eil (Pearson 1975). Withinthe same area, sublittoral rock communities aredescribed by Christie (1980). Monitoring studies usingunderwater photogrammetry extend onto the opencoast in the Skaggerak region. The methods used andspecies studied were similar to those of Swedish workersand some of the main results of these studies from 22sites on the Norwegian and Swedish coasts are given byLundälv & Christie (1986).
On the Skagerrak coast of Norway outside Oslofjord,there are habitats ranging from steeply sloping wave-exposed rock to extremely sheltered inlets. Here,sublittoral communities, at least below the haloclinewhere present, are very similar to those in Scottishsealochs. Those above the halocline where water is ofvariable or low salinity are comparable with thoseoccurring in a very small number of locations in Britain;for instance Loch Obisary in the Outer Hebrides.(Dipper, Lumb & Palmer 1987; K. Hiscock pers. obs.). In1991, the MNCR undertook a survey of some of thefjords in southern Norway to compare the benthiccommunities present with those of Scottish sea lochs(Connor 1991). Rocky intertidal communities were veryrestricted in extent due to small tidal range but also lessrich than in Scottish sea loch most likely as a result oflow salinity conditions. There were also considerabledifferences in sublittoral communities with bryozoans,fish, sponges and calcareous tubeworms betterrepresented in the fjords but crustaceans, molluscs,hydroids and burrowing anemones less wellrepresented. In general, the variety of sublittoralhabitats in the fjords was less than in Scottish sea lochsmainly owing to lack of tidal currents and the
predominance of bedrock habitats in the upper 50 m offjords compared with the more varied substrata in lochs.
The distribution of marine species along the coast ofNorway has been recorded and mapped (described inBrattegard & Holthe 1995) by gathering data frommarine laboratories, field workers and taxonomists.
6.8 Sweden – west coastThe Swedish west coast (Figure 25) is affected by theoutflow of water from the Baltic and the salinity ofsurface waters commonly drops to 10‰, althoughsalinity below the halocline at 10 m to 15 m depthremains above 30‰. Rosenberg & Möller (1979) describehow sediment macrofaunal communities below thehalocline are much richer in species and have a higherbiomass than shallower communities subject to reducedand variable salinity. Much of the work undertaken inthis part of Sweden and of relevance to the MNCR hasbeen carried out at the Kristinebergs Marine ZoologyStation at Lysekil. From here, Molander (1928) tested thecommunity hypothesis of Petersen by his studies in theGullmar Fjord. Later, in 1962, Molander published afurther study of the sediment communities in fjordsalong this coast. Some of the earliest descriptions ofrocky sublittoral communities were those undertaken byGislén in the Gullmar Fjord and published in 1930.Much later, a programme aimed at describing dynamicaspects of marine communities in the same area started(Lundälv, Larsson & Axelsson 1986). Svane & Gröndahl(1988) revisited the sites surveyed by Gislén (1930) and,although finding that the downward vertical extent ofmacroalgae had diminished, the deeper animal-dominated fauna were very similar. Rex (1975),describing the algal assemblages in the eutrophicatedByfjorden, also compares them with samples taken byGislén (1930) and obtained substantially differentresults. Other observations by diving which describemarine communities include those of Michanek (1967).
6.9 The BalticAlthough biogeographically in the same region as theNorth Sea, the Baltic has many features which makecommunities there very different from those widelyencountered in the rest of the north-east Atlantic;especially with regard to low salinity and the de-oxygenation of deeper waters. A country-by-countryreview is not therefore undertaken here. The physical,chemical (salinity) and biological characteristics of theBaltic are summarised by Jansson (1978), who notes thatthe salinity of surface waters is about 6‰ to 7‰ withwater flowing into the Baltic from the Kattegat havingan average salinity of 17.5‰ and maintaining thesalinity of deep water at about 11‰. Shallow rockyareas are dominated by filamentous algae and Fucusvesiculosus whereas sediment areas, particularly in thesouth, are extensively covered by seagrass Zosteramarina. Jansson (1978) gives numbers of macroscopicspecies recorded from various parts of the Baltic and theinner limits of marine and brackish water species(summarised from Segerstråle 1957; Figure 27). Eighty-eight species are recorded in the central Baltic comparedwith 1,500 in the east of the Skagerrak. Andersin, Lassig& Sandler (1977) describe the sediment communities in
39
Hiscock: Introduction and Atlantic-European perspective

the Baltic. In deep water, below about 60 m, thesediments switch between aerobic and anaerobic statesand prolonged periods of deoxygenation occur,exacerbated by eutrophication. Inputs of pollutants to asea where residence time of the water is 25 to 40 years isa therefore serious problem. Shallow rocky sublittoralcommunities in the north of the Baltic are described byJansson & Kautsky (1977). Olenin (in press) summarisesinformation on benthic zonation in the Baltic anddescribes the southern Baltic in more detail. Kiel Bayand the adjacent Lübeck Bay in the south-western Balticare described and considered relatively rich in speciescompared with the rest of the Baltic (Rosenberg 1980).Lübeck Bay in the south-west was the location for theuse of an underwater laboratory from which Gulliksen(1977) described the fauna of rocks and boulders. Here,rocks were dominated by the ascidian Dendrodoagrossularia and the polychaete worm Polydora ciliata,both species found in low salinity in Great Britain.
Baltic marine biologists are currently developing amarine habitat classification for HELCOM with abroadly similar structure to the MNCR classification (ECNature 1996).
6.10 DenmarkThe coast of Denmark (Figure 25) is markedly differentfrom west to east. The west coast is predominantlysandy and, south of Esbjerg, is part of the Wadden Sea.To the east, in the Kattegat at the entrance to the Baltic,there are large and small islands virtually blocking the
entrance to the Baltic. The seabed here is predominantlysedimentary and shallow areas are extensively coveredby seagrass Zostera marina. However, areas of hardsubstratum occur in the form of glacial boulder dumps.The Limfjord, which cuts through Denmark from westto east, is a large (1,500 km2) shallow inland seaconnected by narrow entrances to the North Sea andKattegat.
It was off the coast of Denmark that C.G.J. Petersenundertook his classic studies sampling the seabed todescribe the communities present there (introduced inPetersen & Jensen 1911, summarised in Petersen 1915,1918, 1924). Petersen’s initial intention was to obtainsamples to calculate the quantity of fish food available tobottom-living fish but he soon recognised distinctspecies groupings which occurred over large areas. Hiswork was mainly in the Kattegat and detailed maps ofcommunity distribution there were presented. An‘Echinocardium–Filiformis’ community is illustrated inFigure 28.
The communities described by Petersen (1918) forareas off Denmark are listed below.
I The Macoma or Baltic community
II The Abra community
III The Venus community
IV The Echinocardium–Filiformis community
V The Brissopsis–Chiajei community
VI The Brissopsis–Sarsii community
VII The Amphilepis–Pecten community
VIII The Haploops community
[IX The deep Venus community]
Petersen also described the fauna associated with bedsof Zostera marina.
The communities identified by Petersen have beendescribed in a wide range of studies around the coast ofGreat Britain and the Petersen approach continues to beused today in naming sediment communities.
Petersen’s work in Denmark was followed by thatundertaken by Blegvad (1922, 1928, and 1930) for thesouthern part of the North Sea, Limfjord and theKattegat respectively. The stations sampled by Petersenin the Kattegat in 1911–12 were sampled again usingsimilar techniques (Pearson, Josefson & Rosenberg 1985)and significant changes attributed mainly toeutrophication are described (for instance Figure 29).Although predominantly sedimentary, the Kattegatincludes ‘stone reefs’; stones and boulders depositedduring the ice age. These hard substrata are colonised byepibiota and the algae are described for the TønnebergBanke by Nielsen (1991). Her records also compare thespecies present in the late 1980s with those collected atthe same locality by Professor L.K. Rosenvinge 75 to 100years ago. Although there were differences in speciescomposition, species richness was similar and there waslittle effect from eutrophication. The predominantlyremote sampling techniques used in the Kattegat missedthe discovery and description of carbonate-cemented
40
Marine Nature Conservation Review: benthic marine ecosystems
Figure 27. Numbers of macroscopic animals (in circles), salinity(dotted lines) and the innermost limit of some marine speciesin the Baltic. a = Baltic tellin Macoma balthica; b = musselMytilus edulis; c = cod Gadus morhua; d = bladderwrack Fucusvesiculosus; e = moon jellyfish Aurelia aurita; f = plaicePleuronectes platessa; g = mackerel Scomber scombrus; h =common starfish Asterias rubens; i = shore crab Carcinus maenas.(From Jansson 1978.)

sandstone reefs including columns of carbonate rockformed as a result of methane gas seeps (Jensen et al.1992). These columns, up to 4 m high and currently onlyknown in the Kattegat, are islands for rich epifaunalcommunities and also attract fish. Sediments around themare very varied and therefore the range of communities ina small area is great.
The Limfjord includes extensive shallow watercommunities but about 22% of the bottom is influencedby annual oxygen depletion in late summer (Rosenberg1980). Studies of the Limfjord figure greatly in theReports of the Danish Biological Station including aspecial note in Petersen (1918). Communities there werelater described by Blegvad (1928) and by Jørgensen(1980), who notes the presence of a typical Macomacommunity in shallow water and a Syndosmya (nowAbra) community in the soft muds. Farther south, on theNorth Sea coast, Ringkøbing Fjord is a very extensiveshallow area of brackish water behind a coastal landstrip. Its history during this century has been one of
considerable change in saltwater influence (Johansen,Blegvad & Spärck 1933-1936).
The brackish water habitats of the shallow fjords on theeast coast were extensively surveyed by Muus (1967), whoconcluded that they were dominated by a Cardium lamarki(now Cerastoderma glaucum) – Hydrobia ventrosa community.The most recent description of benthic communities there(Spärck 1936) compares the fauna with other brackishnorth European waters. The situation today is doubtlessdifferent again as a new opening to the sea has been madein recent years (H. Christensen pers. comm.).
In the Wadden Sea area of Denmark, studies wereundertaken by Thamdrup (1935) and Smidt (1951) in thebrackish inshore waters. Thamdrup describes a seawardzone with Arenicola marina, Cerastoderma edule andMacoma balthica, and a zone closer inshore with Hydrobiaulvae, Pygospio elegans and Corophium volutator. Smidtalso describes the epifauna of Zostera (presumablyZostera marina), Mytilus edulis, Ostrea edulis and Sabellariaspinulosa as well as mobile fauna.
41
Hiscock: Introduction and Atlantic-European perspective
Figure 28. An ‘Echinocardium – Filiformis’ community present in a 0.25 m2 sample collected from 20 m to 22 m depth in the Kattegat.(From Petersen 1918.)

6.11 The Wadden SeaThe ‘Wadden Sea’ (‘Waddenzee’ in the Netherlands,‘Wattenmeer ’ in Germany and ‘Vadehavet’ in Denmark)extends along the western and northern coasts ofDenmark, Germany and the Netherlands (Figure 30). Itis bounded on the open coast by 17 large barrier islandsas well as many small islands and sand banks. TheRivers Ems, Weser and Elbe empty into the Wadden Seawhich occupies about 10,000 km2 making it the largestestuarine area in Europe (Wolff 1979). Studies of theWadden Sea have mostly been carried out withincountry boundaries but brought together by the WaddenSea Working Group. The work carried out issummarised in Wolff (1983). Accounts of workundertaken solely within the political boundaries of aparticular country are noted in the country descriptions.Volume 1, parts 3 and 4 of the series The ecology of theWadden Sea edited by Wolff (1983) are very usefulsummaries. Hoek et al. (1983) describe seven macrophytecommunities from the Wadden Sea: the upper eulittoralEnteromorpha community; the lower eulittoralEnteromorpha-Ulva community; the lower eulittoralcommunity of Zostera noltii, Zostera marina, Enteromorphaand Ulva; the lower eulittoral community of Fucus
vesiculosus on Mytilus banks; the ‘rich’ lower eulittoralcommunity and upper sublittoral algal community; the‘rich’ lower eulittoral community of Zostera noltii andZostera marina; and the ‘rich’ sublittoral community ofZostera marina. Diatom communities are also described.Dankers & Beukema in the section edited by Dankers,Kühl & Wolff (1983) on invertebrates do not identifydiscrete communities in the Wadden Sea but describethe distribution of species in relation to sediment typesand location. Descriptions of conservation effort andrequirements and of changes in flora and fauna aredescribed in Dankers, Smit & Scholl (1992).
6.12 Germany (North Sea)Near the border between Denmark and Germany, theisland of Sylt has been especially thoroughly studiedand patterns of meiofaunal and macrofaunal speciesdistribution reviewed by Armonies & Hellwig-Armonies(1987). Figure 31 illustrates those recorded fromKonigshafen. Further south, the North Sea coast ofGermany is greatly influenced by the presence of threemajor estuaries: the Elbe, Weser and Ems. Michaelis(1981) describes the intertidal sediment communities ofthe Ems and Weser while van Arkel and Mulder (1982)describe the benthic fauna of the Ems–Dollard estuary.These authors identify the different communities thatoccur along the gradient of decreasing salinity. Thesubtidal fauna at stations along the salinity gradient inthe Weser estuary is also described by Gosselck et al.(1993). Comparison is made with the results of previousstudies and the possible effects of and changes in faunafollowing dredging is discussed. Eighty-nine specieswere recorded with a strong polarisation of samples toouter (high salinity) and inner (brackish) regions of theestuary using principal component analysis. The typicalinhabitants of brackish waters and tidal flats are notedas the hydroid Cordylophora caspia and the wormsStreblospio shrubsoli and Manayunkia aestuarina.Differences in the estuarine fauna between 1967 and1991 are accounted for partly by man-made deepeningof the Weser and by eutrophication but also rapidproliferation of non-native species (for instance, theworm Marenzelleria viridis and the bivalve Ensis directus).
Other parts of the inshore intertidal areas especiallythoroughly studied include the area of Norderney(Dörjes 1992) and a portion of the tidal flats behindbarrier islands on the north coast (Hertweck 1995)where the distribution of eight intertidal assemblagesand distinctive shore types has been surveyed andmapped. Jade Bay, adjacent to Wilhelmshaven, has alsobeen extensively studied for both intertidal assemblages(for instance, Hertweck 1994; Figure 32) and the biota ofsubtidal channels (Dörjes 1992).
Descriptions of the marine communities present offthe North Sea coast of Germany (the German Bight)include the work of Hagmeir (1925), Remane (1940),Stripp (1969a, 1969b), Reineck et al. (1968) andSalzwedel, Rachor & Gerdes (1985) for the level seabed.Reise & Bartsch (1990) characterise epifaunalcommunities for the German Wadden Sea and offshoreareas in the German Bight. These constituted twoseparate regions in terms of species composition, withthe main dominant species in the Wadden Sea being
42
Marine Nature Conservation Review: benthic marine ecosystems
Figure 29. Pictorial representation of the distribution andabundance of the larger macrobenthic fauna in the Skaldervik,southeastern Kattegat in 1912 and 1984. From Pearson, Josefson& Rosenberg (1985).

43
Hiscock: Introduction and Atlantic-European perspective
Wilhelmshaven
GERMANY
0 100
km
N
Hamburg
54˚N54˚N
52˚52˚
10˚10˚
DENMARK
Amsterdam
Esbjerg
NETHERLANDS
BELGIUM
River ElbeRiver Elbe
River WeserRiver Ems
River Rhine
River Meuse
River Scheldt
Ijsselmeer
FRANCE
Amsterdam
Esbjerg
DENMARK
GERMANY
NETHERLANDS
BELGIUM
Sylt
German Bight
Helgoland
Oyster Ground
NORTH SEA
Texel
River Elbe
River WeserRiver Ems
River Rhine
River Meuse
River Scheldt
Delta Region
Lake Grevelingen
Ijsselmeer
Fris ian Is lands
FRANCE
0 100
km
N
Wad
den Sea
Oosterschelde
Westerschelde
54˚N
52˚
4˚E 10˚6˚ 8˚
Hamburg
North
Frisian
Islands
Wadden Sea
Wilhelmshaven
Figure 30. The German Bight, Wadden Sea and the Low Countries, showing locations of places mentioned in the text.
Figure 31. Benthic communities in Konigshafen on the island ofSylt, Wadden Sea. (From Reise 1985.) 1. Pomatoschistus microps,2. Hydrobia ulvae, 3. Pygospio elegans, 4. Macoma balthica, 5.Scoloplos armiger, 6. Cerastoderma edule, 7. Arenicola marina, 8.Carcinus maenas, 9. Mytilus edulis, 10. Littorina littorea, 11.Tubificoides benedeni, 12. Heteromastus filiformis, 13. Mya arenaria,14. Nephtys hombergii, 15. Lanice conchilega, 16. Nereis (nowHediste) diversicolor, 17. Corophium volutator.
Figure 32. Typical assemblage of endobenthos andlebensspuren [architectural patterns in the sediment typical ofcertain species] in the transitional zone between the mudflatand mixed flat in the Jade area near Wilhelmshaven. 1.Heteromastus filiformis; 2. Nereis (now Hediste) diversicolor; 3.Pygospio elegans; 4. Arenicola marina, a) juvenile, b) half adultsize; 5. Scrobicularia plana; 6. Macoma balthica; 7. Mya arenaria,juvenile; 8. Cerastoderma edule. (From Hertweck 1994.)

decapod crustaceans and in the North Sea echinoderms.Five communities are listed for the German Bight inSalzwedel, Rachor & Gerdes (1985) and theirdistribution is illustrated in Figure 33. Changes in thesublittoral zoobenthos of the German Bight over 60years based on published data are described by Rachor(1990). He suggests an overall increase in biomass andchange in species dominance which is interpreted asbeing influenced and driven by eutrophication.
The island of Helgoland is a small area of rockysubstratum in an otherwise sedimentary offshore area.Sandstone, limestone and chalk bedrock is present in thesubtidal. The Biologische Anstalt Helgoland has provideda centre for marine biological studies since itsestablishment on the island in 1892 although studies ofbenthic algae extend back to the earliest part of the 19thcentury. Studies of algae are reviewed by Mollenhauer &Lüning (1988). Papers describing the marine biologicalcommunities on the shores of Helgoland include those ofden Hartog (1959) incorporated in his review ofNetherlands algal communities, Munda & Markham(1982), who describe littoral algal communities andseasonal change, and Janke (1986), who describes littoralanimal populations, and later (Janke 1990), biologicalinteractions on the shores. The sandstone shores ofHelgoland are dominated by the fucoid alga Fucusserratus with lesser amounts of Fucus spiralis and Fucusvesiculosus and a small number of other species. Pelvetiacanaliculata, which is usually abundant on fucoiddominated European shores, is absent on Helgoland.Munda & Markham (1982) note that the winter and early
spring flora of Helgoland shows several resemblances tothe summer flora of Scandinavia. Animal communities onHelgoland shores are notable for the absence of Patellaspecies and for the high abundances of rock-boringpolychaetes Polydora ciliata and Fabricia sabella in the softsandstone rock. The noted absence of some othercommon rocky shore species (the gastropods Monodontalineata, Gibbula umbilicalis and Littorina (now Melarhaphe)neritoides) is to be expected in the cold North Sea. Thedistribution of macroscopic algae is described by Lüning(1970). Laminaria digitata and Laminaria saccharina occurbetween 0.5 m and 1.5 m below mean low water of springtides followed by a Laminaria hyperborea forest to 4 m. Thedeepest algae (encrusting species) were recorded at 15 m.The development of a subtidal hard substratumcommunity is described by Anger (1978). Recently, severalmostly unpublished studies have investigated the faunaof hard substrata. Schultze et al. (1990) describe themacrofauna and macroflora of kelp plants. De Kluijver(1991) describes analysis of samples from 80 stations at 14locations around Helgoland. Nine communities areidentified.
6.13 The NetherlandsThe coast of the Netherlands is long and indented withmany important features for comparison with Britain.The intertidal algal communities on hard substrata(mainly artificial surfaces) are thoroughly described andclassified by den Hartog (1959) and by Nienhuis (1976).Communities colonising brackish waters are alsodescribed, often in relation to major coastal protectionor land claim works. For instance, Kroon, de Jong &Verhoeven (1985) list the macrofauna of ponds, ditchesand drainage channels on the island of Texel identifyingthose which are euryhaline fresh water, ‘true’ brackishwater, euryhaline marine and holeurhaline species witha general distribution over the whole salinity range.
The fauna of tidal flats in the Dutch Wadden Sea isdescribed by Beukema (1976), who found predominantlya Macoma balthica community present. Beukema (1989,1992) describes long-term and recent changes inmacrofaunal abundance on tidal flats in the western partof the Wadden Sea. This provides a valuable insight intothe changes in 29 species over a twenty-year period.One group of 12 species were identified as being sensitiveto low winter temperatures, which caused low densitiesafter cold winters. A further group of 11 species showed anupward trend in biomass over the twenty years, probablyas a consequence of increasing eutrophication. Remainingspecies showed no specific trend. The composition andseasonal variation of Zostera marina and Zostera noltiicommunities on tidal flats in the south-west Netherlands isdescribed by Jacobs, Hegger & Willens (1983).
Farther south and to the Belgian border, is the DeltaRegion where three major European rivers, the Rhine,Meuse and Scheldt (which separates into the Westernand Eastern Scheldt) enter the sea. This was previouslyan extensive estuarine area but, following disastrousfloods in 1953, the Netherlands government decided toclose-off three of the four main estuaries, a decisionwhich would have created huge brackish or freshwaterlakes. However, environmental considerations led toonly a partial implementation of the plan. The Delta
44
Marine Nature Conservation Review: benthic marine ecosystems
Figure 33. The distribution of deeper sublittoral communities inthe German Bight and adjacent areas. (From Salzwedel, Rachor& Gerdes 1985.)

Institute for Hydrobiological Research was founded tostudy the changes which occurred in relation to thebuilding of the various tidal barriers (for example,volume 18 of the Netherlands Journal of Sea Research isdedicated to papers on Lake Grevelingen: from an estuaryto a saline lake (Hummel, Brummelhuis & de Wolf 1986)).More recently, a volume of Hydrobiologia described theOosterschelde area following its closure (Nienhuis &Smaal 1994). Before these changes started, detailedstudies were undertaken and are reported in Wolff(1973), who concentrates on describing and comparingbrackish water faunas. Figure 34 illustrates thedistribution of soft-bottom animal species over thesalinity gradient from the North Sea to the rivers.Changes to the Rhine, Meuse and Scheldt estuaries as aresult of tidal barrier construction were described byHeip (1989), who suggests that only the Western Scheldtremains a true unchanged estuary. He points out that,from the original situation of four comparable estuaries
entering the Delta region, many ecologically verydifferent water bodies have been created.
Offshore areas of the Netherlands were sampled instudies by Creutzberg et al. (1984), who suggested that atidal flow strength below about 0.9 knots (1.8 m s–1)allowed the settlement of suspended organic mattercreating a rich benthic fauna. The ‘front’ established bydecrease in current is static and thus creates a clearboundary. The fauna of the ‘Oyster Ground’ north of theDutch Wadden Sea is described by de Wilde, Berghuis &Kok (1984) and by Cadée (1984). Here, there was asouthern sandy area in which communities resembledthe Venus (now Chamelea) gallina community of Petersenand a northern rich Amphiura filiformis community withspatangids, Chaetopterus variopedatus, callianassids,Arctica islandica and amphiuroids on a muddy seabed indepths of about 30 m to 50 m (Figure 35). Thedistributionn of meiobenthic and macrobenthic speciesand identification of species groupings (using
45
Hiscock: Introduction and Atlantic-European perspective
ANorth Sea
BLower reaches ofestuaries
CMiddle reaches ofestuaries
DUpper reaches ofestuaries
ERivers
Very stable, high salinityExposed
Stable, high salinitySheltered
Unstable, mediumsalinitySheltered
Stable, low stabilitySheltered
Stable, low stabilitySheltered
Restricted to A(types 1, 2)44 species
Restricted to B(type 4)17 species
Restricted to C(type 6)Boccardia ligericaRhithropanopeus harrisiiCyathura carinataLeptocheirus pilosusCorophium multisetosum
Restricted to D(type 8)Perforatella rubiginosaErpobdella monstriataHaemopsis sanguisugaGlossiphonia heteroclita
Restricted to E(type 9)4 species
Restricted to A and B (types 2, 3, 4)104 species
(Talitrus saltator)(Talorchestia deshayesii)
Restricted to A, B and C (type 5)Macoma balthicaMya arenariaEteone longaHeteromastus filiformusCrangon crangonCarcinus maenas
Restricted to D and E (type 9)22 species
Restricted to B and C (types 5, 6)
Nereis succineaPolydora ligniPygospio elegans(Orchestia gammarella)
Assiminea grayanaLimapontia depressaAlderia modestaStreplospio shrubsoliiManayunkia aestuarinaNeomyosis integerSphaeroma rugicaudaGammarus salinusCorophium volutator
Restricted to B, C and D (type 7)Nereis diversicolor
Restricted to C and D (type 7)Gammarus zaddachiPseudamnicola confusa
Restricted to C, D and E (types 7, 8)Potamopyrgus jenkinsiLimnaea peregraTrocheta bykowskii(Orchestia cavimana)
Migrating species (type 10)Eriocheir sinensis
Figure 34. The distribution of sediment benthos species over the salinity gradient from the North Sea to the Rivers Rhine, Meuseand Scheldt. The large number of species characteristic of the North Sea and lower reaches of the estuaries are not shown. (FromWolff 1973.)

TWINSPAN) off the north coast of the Netherlands isdescribed by van Scheppingen & Groenewold (1990) andby Holtmann & Groenewold (1994). Their results aredescribed in terms of the indicator species andgroupings identified by TWINSPAN analyses. Theoffshore macrobenthic assemblages are described as:
north of the 30 m depth line
◆ an Amphiura filifomis, Callianassa subterranea andMysella bidentata assemblage;
◆ a Callianassa subterranea, Cultellus pellucidus,Lumbrinereis latrelli, Magelona papillicornis and Nephtyshombergii assemblage.
south of the 30 m depth line
◆ a Chaetozone setosa, Magelona papillicornis andSpiophanes bombyx assemblage;
◆ a Nepthys cirrhosa and Scoloplos armiger assemblage.
Off the south coast of the Netherlands, in theSouthern Bight of the North Sea, benthic communitieswere sampled by Govaere et al. (1980). They describe anearshore Abra community, a transition zone and anoffshore Venus community.
Extensive studies of macrobenthos continue off theNetherlands; they include a long-term monitoringprogramme described by Holtmann et al. (1995) andpublication of an atlas of zoobenthos of the Dutchcontinental shelf (Holtmann et al. in press). The paper
by Holtmann et al. (1995) suggests, as before, four stationclusters although they are distributed differently.Analysis of data from 1991 to 1994 showed no majorchange in benthos over the years.
6.14 BelgiumThe coast of Belgium is about 65 km long and is sandybeach backed by dunes. Due to the influence of thenearby Scheldt estuary, the eastern section is much moresilty than to the west. The intertidal coast is described byWarmoes, Backeljau & de Bruyn (1988), who studied thelittorinid molluscs found there. Dykes for coastal defenceare constructed along about 40 km of coastline but areonly rarely within the littoral zone. However, largenumbers of breakwaters built at right angles to the coast,together with piers and harbours, provide littoral hardsubstrata. Below low water, the seabed is very graduallysloping sand which only reaches 10 m depth between 3km and 10 km offshore. The seabed off the Belgium coastwas one of the first to be sampled systematically (Gilson1907). The area offshore from the eastern part of the coastwas included in the sampling of benthic communitiesdescribed by Govaere et al. 1980. A few kilometresoffshore, the western half is characterised by the FlemishBanks; sandbanks set obliquely to the shore. Locallycoarse sandy sediments are found with a rich interstitialfauna (Rappé 1978; Vanosmael et al. 1982). A monitoringprogramme sampling sublittoral sediments off the coasthas been under way for several years but results are notyet published (A. Catrysse pers. comm.).
6.15 Channel IslandsThe Channel Islands were especially mentioned byForbes & Godwin-Austen (1859) as peculiar for thepresence of many Lusitanean species not only absentfrom south-western coasts of England but also from thenearby western coast of France. Crisp & Southward(1958) included the Channel Islands in their study of thedistribution of intertidal organisms along the coasts ofthe English Channel. Holme (1966) included samplesites in the region of the Channel Islands in his surveyof benthic communities in the English Channel (Section5.2.5). However, remarkably little information describingmarine biological communities around the islands hasbeen found. The marine algae of Guernsey weredescribed by Lyle (1920) although recent studies (Dr C.Maggs pers. comm.) have revealed a great increase innon-native species, which now dominate some areas.The littoral fish of Guernsey were listed by Wheeler(1970). Surveys of the species living in a wide variety ofhabitats, particularly in the littoral, at Portelet Bay andsome other locations in Jersey were reported by Culleyet al. (1983), Thomas & Culley (1987) and Culley, Thomas& Thorp (1988). Their studies do not includeidentification of community types but descriptions fromhabitats rarely sampled in Great Britain, includingrockpools and crevices, and are useful for comparativepurposes. Crisp & Southward noted the presence of thetopshell Gibbula pennanti and the ormer Haliotistuberculata, which are not found in Great Britain. Thiswas mentioned again by Culley, Thomas & Thorp (1988)along with several species of algae that are rarelyencountered or not found in Great Britain.
46
Marine Nature Conservation Review: benthic marine ecosystems
Figure 35. The community present on the Oyster Ground.(From de Wilde, Berghuis & Kok 1984.) 1. Spatangids (includesEchinocardium cordatum, Echinocardium flavescens, Brissopsislyrifera). 2. Chaetopterus variopedatus. 3. Callianassids (includesCallianassa subterranea, Upogebia deltaura). 4. Arctica islandica. 5.Ophiuroids (includes Amphiura filiformis, Amphiura chiajei). 6.Gattyana cirrosa. 7. Glycera rouxi, Glycera alba. 8. Nereis (nowHediste) and Nephtys spp. 9. Notomastus latericeus. 10. Echiurusechiurus.

6.16 Isle of ManSome of the earliest studies of marine biology in the firsthalf of the 19th century were undertaken by EdwardForbes off the Isle of Man (Figure 36)(for example,Forbes 1835a&b). However, it was following the move ofthe Liverpool Marine Biology Committee’s interests tothe Isle of Man in 1892 and later by the establishment ofthe Liverpool University Marine Biology Laboratory atPort Erin that marine research became firmly establishedthere. Since then, much ecological work has beenundertaken, although this has been almost entirelyaround the southern part of the island. The algae of theIsle of Man are listed in Knight & Parke (1931) and themarine fauna together with descriptions of the maincollecting grounds in Bruce, Colman & Jones (1963).Jones (1951) describes the bottom fauna off the southend of the Isle of Man where he identified fourcommunities in relation to sediment types: ‘the offshoregravel community’; ‘the offshore fine sand community’;‘the offshore muddy sand community’ and ‘the offshoremud community’. Further descriptions of seabedcommunities are given in Eggleston (1963) and Ward(1988), who concentrate on assemblages of Bryozoa.Some of the communities in littoral sediments aredescribed in Southward (1953). One of the early studiesusing self-contained underwater breathing apparatus tosurvey areas of rocky seabed is published in Kain (1960).She sampled algae at intervals of approximately 1 mdepth from low water to the depth to which rock waspresent at ten different locations around the south coast.In addition, mobile hard substrata were sampled at foursites. A study of seasonal change on the breakwater atPort Erin was described in the same paper.
Much of the work undertaken from the Isle of Manhas been experimental rather than descriptive in natureand has added considerably to our knowledge of theecology and dynamics of marine ecosystems. Of specialnote are the studies of limpet grazing (reviewed inSouthward 1953 and in Hartnoll & Hawkins 1985), seaurchin grazing (Jones & Kain 1967) and the ecology ofkelp (reviewed in Kain 1979). There are severalunpublished reports describing rocky sublittoralcommunities and a great deal of local knowledge,mainly about the southern area of the island, is held bymarine station staff. Most recently, Geffen, Hawkins &Fisher (1990) have characterised the shores of the Isle ofMan and mapped the occurrence of marine habitats andcommunities based on those defined by the MNCR. Thebiota is distinctly northern or cold-water with theabsence of several southern species most probablycompounded by the isolation of the Isle of Man frompossible mainland sources of larvae. The southernmostrecords on the west coast of Britain for the red algaOdonthalia dentata occur off the Isle of Man.
6.17 Ireland
6.17.1 IntroductionThe coast of Ireland (Figure 36) is predominantly rockyand encompasses a western seaboard bathed by thewarm waters of the North-East Atlantic Drift around tothe much colder waters of the enclosed Irish Sea.Differences in seawater temperature around Ireland
result in a distinctive geographical distribution ofspecies (for example, of littoral species: Southward &Crisp 1954). Communities on the west coast include arich mixture of lusitanean elements and thosecharacteristic of enclosed inlets within such areas asGalway Bay.
Recorded studies of marine biology in Ireland extendback to the beginning of the 19th century but usefulcomparative information is found in more recentreports. Northern Ireland’s sublittoral and littoral areaswere thoroughly surveyed in exercises similar to theMNCR (Erwin et al. 1986, 1990; Wilkinson et al. 1988).The littoral and inshore sublittoral areas of the Republicof Ireland have been surveyed since 1993 as a part of theEuropean Commission Life-funded BioMar programme(Costello & Mills 1996). MNCR are partners in theprogramme and the BioMar team based in Dublin usedMNCR methods and entered data to the MNCRdatabase.
Papers describing the marine ecology of Ireland andpublished in the Irish Naturalists Journal are listed inKelly et al. (1996).
6.17.2 Republic of IrelandThe Clare Island Survey which was undertaken in theearly part of the 20th century under the auspices of theRoyal Irish Academy was a ‘model’ descriptive surveyencompassing studies of hydrology, sedimentology andthe relationships between various communities andtheir environment. The results of this survey aresummarised in Southern (1915). The descriptions ofalgal communities (Cotton 1912) and animalcommunities (Southern 1915) are easily compared withthe work of the MNCR. Fifteen algal ‘associations’(although some are described by habitat type ratherthan species) are described for exposed rocky littoralareas while ten are described for sheltered rocky shores.Further associations are described for sublittoral regions,littoral and sublittoral sediments, saltmarshes, rivermouths and brackish bays. Southern (1915) provides ahierarchical classification of habitat types andcharacterising animal species and notes 15 sedimenttypes and associated species and 11 hard substratumtypes and associated species for both littoral andsublittoral areas. Petersen (1918) commented onSouthern’s communities and suggested the presence of‘Macoma’ and ‘Venus’ communities and somethingapproaching the ‘Echinocardium – Filiformis’ community.
The series of papers entitled The ecology of Lough Inepublished since 1948 have greatly enhanced ourknowledge of littoral and shallow sublittoral ecology.Lough Ine (now called Lough Hyne) is a deep enclosedmarine basin in south-west Ireland. The lough has beenstudied since the late 1920s (Renouf 1931) and later byJ.A. Kitching and his co-workers (early papers aresummarised in Kitching & Ebling 1967 and updated inKitching 1987). Those studies have included descriptionsof the distribution of marine species in different habitatsand particularly in relation to environmental conditionsincluding wave exposure, tidal current velocity, siltation,light and deoxygenation. Few studies list species interms of communities. Nevertheless, several areparticularly useful for comparative purposes including
47
Hiscock: Introduction and Atlantic-European perspective

the study of boulder faunas in relation to tidal currentvelocity and surfaces aspect (Lilly et al. 1953; Figure 37),the study of species distribution in relation to light in asea cave (Norton, Ebling & Kitching 1971), thedescription of rock pool communities on the open coastthere (Goss-Custard et al. 1979), and the description ofthe open coast Laminaria forest (Norton, Hiscock &Kitching 1977). Hiscock (1976) described the animalcommunities of underwater cliffs on the open coast, justwithin the Lough and in the most sheltered part of theLough (Figure 38). Other studies undertaken in south-western Ireland include the description of rocky shorecommunities in Bantry Bay (Crapp 1973), the survey ofKillary Fjord (Mathers 1975), of Roaringwater Bay on the
south-west tip of Ireland (Hiscock & Hiscock 1980) andof Cape Clear Island and a site on the Dingle Peninsula(Cullinane & Whelan 1983).
Galway Bay and nearby Kilkieran Bay have beenmuch studied in recent years, particularly because of theproximity of University College Galway. One of the keypapers against which we compare results of our studiesin Great Britain is the work of Könnecker (1977) whichdescribed rocky sublittoral marine communities andtheir distribution in relation to temperature stability asfollows:
I. Stenohaline, stenothermal, offshore(a) Tethyopsilla – Tetilla association (below 40 m)
48
Marine Nature Conservation Review: benthic marine ecosystems
Figure 36. Ireland and the Isle of Man, showing locations of places mentioned in the text.

(b) Axinella dissimilis (now A. polypoides) association(25 m to 40 m)
II. Stenohaline, eurythermal, offshore-inshore(a) Upper Laminaria hyperborea association (0 m to
15 m)(b) Lower Laminaria hyperborea association (15 m to
lower limit of Laminaria)III. Euryhaline, eurythermal, inshore
(a) Lithothamnium association (0 m to 20 m)(b) Laminaria saccharina association (0 m to 10 m)(c) Raspailia – Stelligera association (below 10 m)(d) Musculus discors association (no depth limits,
strong currents)
A later paper (Könnecker & Keegan 1983) expandsthese descriptions.
The fauna of extensive beds of maerl in Galway Bayand nearby Kilkieran Bay were described by Keeganmost recently in 1974. Keegan (1974) describes six maerlor predominantly maerl substrata and associatedcommunities providing a basis, with the work ofCabioch (1968) in Brittany, for comparison ofcommunities associated with this particular substratum.Similarly, the work of Maggs (1986) on the algaeattached to maerl in Galway Bay provides a comparison
49
Hiscock: Introduction and Atlantic-European perspective
Figure 37. Studies at Lough Ine (now Lough Hyne) havedemonstrated the relationship between environmental factorsand species distributions. Here, the distribution of three speciesof sea anemone in relation to tidal current velocity is illustratedfor the rapids area. (From Lilly et al. 1953.)
Figure 38. Illustration of the sessile species colonising cliffs at adepth of 15 m on the wave exposed open coast outside ofLough Hyne, the tidal stream exposed area just within thelough and the extremely sheltered area in the western basin ofthe lough. (The key is re-drawn and current names used.)(From Hiscock 1976.)

with the work of Cabioch (1969) in the Bay of Morlaixnear Roscoff and the NCC-commissioned surveys insouth-west Britain. Another notable species present inKilkieran Bay is the large tube-building anemonePachycerianthus multiplicatus, also found in Scottishsealochs. O’Connor et al. (1977) describe a distinctiveassociation of animals living in or on the tube of thisanemone.
A major survey of littoral and nearshore sublittoralbiotopes in the Republic of Ireland has been undertakenas part of the BioMar programme since 1993 (Costello &Mills 1996). MNCR field survey methods and the MNCRdatabase have been used in the work and the resultsfrom Ireland are being incorporated into thedevelopment of the biotopes classification for the BritishIsles.
6.17.3 Northern IrelandMany of the investigations of marine benthiccommunities in Northern Ireland have been undertakenin Strangford Lough; notably in the work of Williams(1954), who described the fauna of the Lough and theneighbouring coasts. Roberts (1975) described the faunaassociated with beds of Modiolus modiolus in the Loughand this was incorporated into a comparison of horsemussel bed faunas undertaken by Hiscock & Mitchell(1980). Mapping surveys of the benthic communities inStrangford Lough have been undertaken using acousticsurvey and photography by Magorrian, Service & Clarke(1995). They describe the distribution and extent ofhorse mussel Modiolus modiolus communities and ofareas of dense burrows of the Norway lobster Nephropsnorvegicus and brittlestar Ophiothrix fragilis beds.
Systematic descriptive surveys of both nearshoresublittoral (Erwin et al. 1986, 1990) and littoral(Wilkinson et al. 1988; Fuller et al. 1991) benthiccommunities were undertaken in Northern Ireland bythe Department of Environment (NI). The sublittoralsurvey describes communities in the following habitats(from Erwin et al. 1986):
PLANT-DOMINATED COMMUNITIES1. Bedrock & boulder: Laminaria hyperborea2. Bedrock & boulders: Laminaria saccharina3. Sand covered rock4. Sand scoured rock5. Mobile cobbles6. Pebble & gravel7. Phymatolithon calcareum (maerl)8. Very sheltered conditions9. Zostera marina
ANIMAL-DOMINATED COMMUNITIES ON HARDSUBSTRATA
1. Bedrock(a) Species composition (= ubiquitous species)(b) Sand scoured rock(c) Sabellaria spinulosa reefs(d) Terraced bedrock(e) Caves & fissures(f) Surge gullies
2. Boulder
3. Mixed boulder, cobble, gravel and sand habitats(a) Unstable substrata(b) Stable substrata
ANIMAL-DOMINATED COMMUNITIES ON SOFTSUBSTRATA
1. Mixed substrata of boulder and cobble withmuddy gravel(a) Ophiothrix fragilis beds(b) Muddy gravel with cobbles & boulder
2. Pebble3. Gravel
(a) Coarse clean shell gravel(b) Clean stone gravel with pebbles(c) Muddy gravel
4. Coarse sand5. Sand
Clean mobile sand with Ammodytes tobianus6. Fine sand
(a) Clean, firm, rippled sand with Echinocardiumcordatum
(b) Clean fine sand with Arenicola marina(c) Muddy fine sand
7. Mud(a) Mud with shell(b) Soft mud
The Stage 1 littoral survey (Wilkinson et al. 1988)identified the location of 12 sediment and 13 rockycommunity types whilst the more detailed Stage 2survey used computer analysis of survey results toillustrate the distribution of different shore types aroundthe coast, and rocky shore types are illustrated in Figure39. Interpretation of the biological results suggested thatgeographical location, wave exposure andsedimentological features were mainly responsible fordetermining the distribution of sites with similar speciesassemblages.
The results of both surveys were summarised and thecommunity descriptions interpreted to the MNCRhabitat/community types for purposes of mapping theirdistribution (Baxter & Boaden 1990); 24 littoral and 19sublittoral habitat/community types in 12 major sitetypes are described.
6.18 Atlantic FranceThe Atlantic coast of France (Figure 40) extends from itsNorth Sea border in the east where conditions aredominated by sedimentary coasts and turbid cold waterspast the mainly rocky coasts of Normandy and Brittanybathed in clearer and warmer waters to the sedimentarycoasts of the Bay of Biscay to the south. The results of awide range of studies undertaken on sedimentary androcky habitats on the French Atlantic coast are availablefor comparison with MNCR work. Those studies havealso been brought together to compile a classification ofbenthic communities in Dauvin (1994), providing aparticularly valuable comparison with the MNCRbiotopes classification.
The widest ranging studies offshore are those for thewhole of the English Channel undertaken by Cabioch etal. (1977) investigating the distribution of species frommobile hard substrata (Figure 23). Descriptions of
50
Marine Nature Conservation Review: benthic marine ecosystems

benthos from a data set of 707 sites sampled by dredgebetween 1971 and 1975 and extending from Belgium toNormandy in France and across the Channel to thesouth-east coast of England will be published inSanvicente-Añorve & Leprêtre (in press.).
The sediment communities along the North Sea andEnglish Channel coast between Belgium and CapeGris-Nez have been extensively sampled and sevengroups identified and mapped (Davoult et al. 1988).These are:
◆ Pebbles with sessile epifauna community
◆ Modiolus modiolus facies
◆ Gravel with Amphioxus (now Branchiostoma) lanceolatuscommunity
◆ Muddy heterogeneous sediment community
◆ Fine to medium clean sand with Ophelia borealiscommunity
◆ Muddy fine sand with Abra alba community
◆ Soft intertidal sediment communities
Farther south along the open coast, Cabioch & Glaçon(1975) describe and map five benthic communities[termed “peuplements” in the French text and translatedhere as “communities”] south of Boulogne to the estuaryof the Somme (Figure 41). Desprez, Ducrotoy & Sylvand
51
Hiscock: Introduction and Atlantic-European perspective
5m
0
5m
0
5m
0
5m
0
5m
0
5m
0
12
3
4 5
Zonation:1. Supralittoral. Very wide zone dominated byVerrucaria maura with littorinids in crevices.2. Verrucaria maura covers rock surface.Porphyra umbilicalis & Enteromorpha spp.also present.3. Dominated by limpet and barnacle species.Tufts of Lichina pygmaea present.4. Encrusting calcareous algae. Corallinaofficinalis, Mastocarpus stellatus & other red algae.5. Alaria esculenta with understory ofencrusting calcareous algae.
1A PORT NA TOBER HEADLAND, NORTH COASTNotes - Very exposed to wave action - Rock types - basalt and hardlimestone - Similar sites found along thenorth coast around Rathlin Island and outer Islandmagee - Subhabitat diversity and species richness low
Horizontal Extent0 200m
Heightabovelow water
Zonation1. Supralittoral Verrucaria maura with littorinids in crevices.2. Verrucaria maura with Porphyra umbilicalis & Enteromorpha spp.3. Mixed zone. Limpet and barnacle species on rock surface & exposed faces of boulders. Fucoids & green algae in shelter between boulders. Mytilus edulis in crevices.4. Red algal turf (Mastocarpus stellatus & Corallina officinalis & Laurencia pinnatifida) with Himanthalia elongata & clumps of Mytilus edulis.5. Laminaria digitata; understory of Corallina officinalis & Mastocarpus stellatus.
1B EAST LIGHTHOUSE PLATFORMS. RATHLIN ISLANDNotes - Wave exposure variesaccording to local shoretopography. - Rock types - basalt & hard limestone. - Example of wave - cut platform which are found mainly along the North Coast.
12
34 5
Zonation1. Patchy Pelvetia canaliculata2. Patchy Fucus spiralis with littorinids.3. Fucus serratus between boulders. Boulder surfaces covered by limpets, barnacles & Nucella lapillus.4. Fucus serratus with mixed red algae (Mastocarpus stellatus, Laurencia spp., Palmaria palmata & Lomentaria articulata)5. Laminaria digitata with an understory of mixed red algae & encrusting corallines.
1C TURNLY'S PORT, EAST ANTRIMNotes - Moderately exposed to wave action. - Similar sites found all along the East Antrim coast. - Shores consisting of boulders and cobbles, more mobile at some sites than others. - Subhabitat diversity and species richness low.
1 23
45
Zonation1. Dense Pelvetia canaliculata with patchy Verrucaria maura and littorinids.2. Dense Fucus spiralis with littorinids.3. Coverage of Ascophyllum nodosum & Fucus vesiculosus with understory of green, brown &red algae. Rock pools & crevices with a highanimal & plant diversity.4. Blanket coverage of Fucus serratus with anunderstory of Corallina officinalis, red algae &encrusting corallines.
2A WALLACE'S ROCKS, NORTH COUNTY DOWN, OPEN COAST
Notes - Sheltered. - Extensive littoral region composed of creviced & fractured sedimentary rock with areas of boulders & rock pools. - Similar sites found along outer Ards Peninsula. - Subhabitat diversity high. Species diversity also high.
1
23
4
Zonation1. Verrucaria maura with Enteromorpha spp. in pools and littorinids in crevices.2. Rock surface dominated by limpet and barnacle species, with patches of Enteromorpha spp. and Porphyra umbilicalis. Rock pools have a high animal & plant diversity. Nucella lapillus common.3. Open rock surface - mixed red algal community. Fucus serratus dominant in shelter.4. Laminaria digitata dominated in shelter with mixed community of red algal understory.
2B WRECK PORT, SOUTH COUNTY DOWN, OPEN COAST
1
2
3
4
Notes - Exposed to moderate wave action - Littoral region composed of creviced & fractured sedimentary rock with areas of boulders & rock pools. - Similar sites found all along southern Co. Down. In some areas boulder shores predominate, in others sedimentary bedrock. - Subhabitat diversity high. Species richness low on open rock surface, but high in crevices and rock pools.
Zonation1. Patchy Pelvetia canaliculata & Verrucaria maura.2. Dense Fucus spiralis.3. Dominated by Ascophyllum nodosum with an understory of brown & red algae. High epifaunal diversity.4. Dominated by Fucus serratus. Understory of filiform & filamentous red & brown algae. High diversity of invertebrates & epifauna beneath boulders.5. Laminaria digitata with Laminaria saccharina & Chorda filum.
2C RINGBURR POINT, STRANGFORD LOUGH
Notes - Very sheltered. - Extensive intertidal region of boulders with wide areas of mixed sediments & wide shallow pools. Similar sites found in Strangford Lough & Carlingford Lough. Conditions of extreme shelter allow heavy growth of epifaunal species on algae. Invertebrate species richness beneath boulders is high. Algal species richness only moderate, but community composition is distinctive.
1
2
34
5
Figure 39. Characterising species for major rocky shore types inNorthern Ireland. (From Fuller et al. 1991.)
Bay of Mont-St-MichelBay of Mont-St-Michel
BoulogneBoulogne
0˚0˚0˚SPAINSPAIN
FRANCEFRANCEFRANCE
ENGLANDENGLAND
2˚2˚2˚ 2˚E2˚E2˚E
GGAASSCCOOGGNNEE4˚4˚4˚
RRiivveerr SSoommmmee
RRiivveerr SSeeiinnee
44˚44˚44˚
46˚46˚46˚
48˚48˚48˚
50˚N50˚N50˚N
N
0 100
km
Dunkirk
Morlaix
Concarneau
Gironde
San SebastianSan Sebastian
BRITTANY
NORMANDY
N
0 100
km
Dunkirk
English Channel
Guernsey
Jersey
St MaloRoscoff
MorlaixBrest
Ushant
Concarneau
Isles Glenans
Gironde
Arcachon
Bay of
Biscay
SPAIN
FRANCE
CHANNEL ISLANDS
ENGLAND
Strait of Dover
San Sebastian
Santander
BRITTANY
NORMANDY
2˚ 2˚E
44˚
46˚
48˚
50˚N
GASCOGNE
´
0˚4˚
Boulogne
River Somme
River Seine
Bay of Mont-St-Michel
Figure 40. Atlantic France and the Channel Isles, showinglocations of places mentioned in the text.
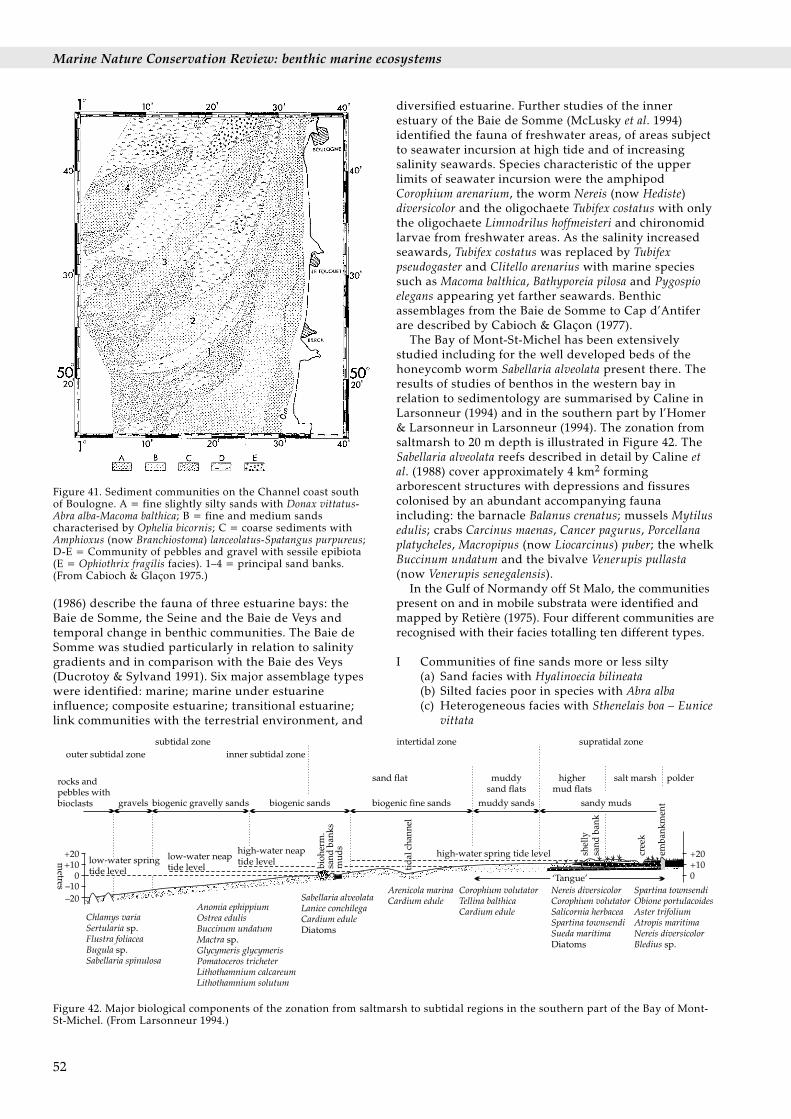
(1986) describe the fauna of three estuarine bays: theBaie de Somme, the Seine and the Baie de Veys andtemporal change in benthic communities. The Baie deSomme was studied particularly in relation to salinitygradients and in comparison with the Baie des Veys(Ducrotoy & Sylvand 1991). Six major assemblage typeswere identified: marine; marine under estuarineinfluence; composite estuarine; transitional estuarine;link communities with the terrestrial environment, and
diversified estuarine. Further studies of the innerestuary of the Baie de Somme (McLusky et al. 1994)identified the fauna of freshwater areas, of areas subjectto seawater incursion at high tide and of increasingsalinity seawards. Species characteristic of the upperlimits of seawater incursion were the amphipodCorophium arenarium, the worm Nereis (now Hediste)diversicolor and the oligochaete Tubifex costatus with onlythe oligochaete Limnodrilus hoffmeisteri and chironomidlarvae from freshwater areas. As the salinity increasedseawards, Tubifex costatus was replaced by Tubifexpseudogaster and Clitello arenarius with marine speciessuch as Macoma balthica, Bathyporeia pilosa and Pygospioelegans appearing yet farther seawards. Benthicassemblages from the Baie de Somme to Cap d’Antiferare described by Cabioch & Glaçon (1977).
The Bay of Mont-St-Michel has been extensivelystudied including for the well developed beds of thehoneycomb worm Sabellaria alveolata present there. Theresults of studies of benthos in the western bay inrelation to sedimentology are summarised by Caline inLarsonneur (1994) and in the southern part by l’Homer& Larsonneur in Larsonneur (1994). The zonation fromsaltmarsh to 20 m depth is illustrated in Figure 42. TheSabellaria alveolata reefs described in detail by Caline etal. (1988) cover approximately 4 km2 formingarborescent structures with depressions and fissurescolonised by an abundant accompanying faunaincluding: the barnacle Balanus crenatus; mussels Mytilusedulis; crabs Carcinus maenas, Cancer pagurus, Porcellanaplatycheles, Macropipus (now Liocarcinus) puber; the whelkBuccinum undatum and the bivalve Venerupis pullasta(now Venerupis senegalensis).
In the Gulf of Normandy off St Malo, the communitiespresent on and in mobile substrata were identified andmapped by Retière (1975). Four different communities arerecognised with their facies totalling ten different types.
I Communities of fine sands more or less silty(a) Sand facies with Hyalinoecia bilineata(b) Silted facies poor in species with Abra alba(c) Heterogeneous facies with Sthenelais boa – Eunice
vittata
52
Marine Nature Conservation Review: benthic marine ecosystems
Figure 41. Sediment communities on the Channel coast southof Boulogne. A = fine slightly silty sands with Donax vittatus-Abra alba-Macoma balthica; B = fine and medium sandscharacterised by Ophelia bicornis; C = coarse sediments withAmphioxus (now Branchiostoma) lanceolatus-Spatangus purpureus;D-E = Community of pebbles and gravel with sessile epibiota(E = Ophiothrix fragilis facies). 1–4 = principal sand banks.(From Cabioch & Glaçon 1975.)
Figure 42. Major biological components of the zonation from saltmarsh to subtidal regions in the southern part of the Bay of Mont-St-Michel. (From Larsonneur 1994.)

(d) Heterogeneous facies silted with Pista cristata(e) (As above), within bottoms of maerl
II Communities of fine clean sands with Armandiapolyophthalma
III Communities of coarse sediments with Amphioxus(now Branchiostoma) lanceolatus(a) (As above)(b) (In maerl)
IV Sessile epibenthic communities of stones andgravels(a) (As above)(b) Facies with Ophiothrix fragilis
The distribution of shore animals along the Atlanticcoast of France from north-west Brittany to the borderwith Spain has been described and mapped by Crisp &Fischer-Piette (1959) while the most abundant speciespresent in the different subzones on rocky shores at sixlocations in the same area are described by Evans (1957).These studies revealed a clear discontinuity in speciesdistribution south of the Gironde estuary, doubtlessaided by the almost entirely sedimentary nature ofshores for over a hundred kilometres. Thus the rockyshores of Brittany are similar to those of south-westBritain whereas those to the south appear to have adifferent biogeographical character. Indeed, the speciesand communities present on the rocky coast around theFrench-Spanish border (the Basque coast) are muchmore southern in character than those farther west andsouth along the Spanish coast. Studying algae, Hoek &Donze (1966) drew together the conclusions of severalstudies to suggest that the Basque coast east of SanSebastian in Spain had a flora intermediate betweenMorocco and north-west Spain.
Castric Fey (in press) describes the sublittoral rockcommunities near to Trébeurden in eastern Brittany inrelation to their exposure to wave action and tidalcurrents. Characteristic species for situations dominatedby wave action and moderate currents where coarsesand occurs on rocks are contrasted with those ofsituations dominated by strong currents. Thesecommunities are similar to those found in south-westEngland.
Studies of marine ecosystems have been carried outover many years by the marine biological station atRoscoff. This work is reflected in part in the series ofpublications which list the marine fauna of the area andthe list of algae (Feldmann 1954).
The seagrass beds of the area of Roscoff have beenextensively studied (many of the papers are included inJacobs 1982). Jacobs & Huisman (1982) describedmacrobenthos associated with Zostera marina and Zosteranoltii.
Off the coast of Brittany, the seabed communitieswere defined and mapped from about 84 km west to 48km east and 60 km north of Roscoff by Cabioch (1968).This study outlined the physical environment of theEnglish Channel, described the distribution of species inrelation to depth, substratum, current and longitude,and described the biocenoses, communities (as“peuplements”) and facies of communities present.Many of the species listed are hard substratum epifauna.
Nineteen assemblages are described and mapped for thenearshore ‘frontolittoral’ zone and a further ten for theoffshore ‘prelittoral’ zone. Part of this map is shown inFigure 43. This very extensive study cannot easily besummarised here but provides a basis for comparisonwith the classification of sublittoral biotopes beingundertaken by the MNCR. A translation of thedescription of regions and assemblages by Cabioch(1968) is given below.
A THE FRONTOLITTORAL REGION [with irregulartopography and extending from tidal regions to theChannel ‘plain’]a. Infralittoral communities:
communities composed of fine sands moreor less silty with Abra alba and Corbula gibba;biocoenosis of maerl;biocoenosis of rock with laminarians.
b. Circalittoral communities:biocoenosis of hard bottoms with Axinelladissimilis.
c. Populations relatively independent of the verticalzones:
biocoenosis of coarse sediments with Venus(now Clausinella) fasciata including facies ofepifauna on Sabellaria spinulosa bound[sediment], particularly at the transitionwith prelittoral populations;facies on rock with Musculus discors.
B THE PRELITTORAL REGION [flat plains, the greatestpart of the western basin of the Channel]
biocoenosis of hard bottoms with Axinelladissimilis;biocoenosis of stones and gravels of thecoastal prelittoral;biocoenoses of stones and gravels in thewider prelittoral;biocoenosis of coarse sediments with Venus(now Clausinella) fasciata, facies withEchinocardium pennatifidum.
(The above include a number of facies.)
The studies undertaken by Cabioch used remotesampling techniques but diving has also been used onthe Brittany coast. Drach undertook a series ofobservations using SCUBA during the late 1940s andearly 1950s on the Atlantic coast of France summarisedin Drach (1952). These were followed by those of Ernst(1955) studying algae.
Algal communities surveyed by diving in the Bay ofMorlaix east of Roscoff are described by L’Hardy-Halos(1972). Earlier studies there had described the flora ofthe extensive beds of maerl occurring on the shallowseabed (Cabioch 1969) providing a basis for comparisonof maerl communities in NCC-commissioned studies insouth-west Britain (for example, Little & Hiscock 1987).At the time of the Amoco Cadiz oil spill in 1978, severalstudies of sediment benthos had been started in the areaof the Bay of Morlaix near Roscoff. These studies rapidlybecame surveys of the effects of oil pollution providingmuch useful information on the response of sedimentbenthos communities. Dauvin (1988) mentions threespecies groupings in the area:
53
Hiscock: Introduction and Atlantic-European perspective

54
Marine Nature Conservation Review: benthic marine ecosystems
Figure 43. Seabed biocoenoses, communities and facies of communities off the Bay of Morlaix. Part of a much larger map of inshore(frontolittoral) region of the north coast of Brittany. (From Cabioch 1968.)

1. community of fine sands with little silt with Abra alba– Hyalinoecia bilineata;
2. community of muddy sands with Abra alba – Melinnapalmata;
3. community of coarse sand with Amphioxus (nowBranchiostoma) lanceolatus – Venus (now Clausinella)fasciata.
There appears to be no published description of hardsubstratum sublittoral communities present in themarine inlets and coast adjacent to Roscoff except for abrief report of an expedition by British divers in 1985(Ackers 1986). This study revealed communities basicallysimilar to those found in south-west Britain but enrichedby many southern species not recorded in British watersincluding, most conspicuously, the sponge Ulosa digitata,the fan worm Sabella spallanzanii, the starfish Echinastersepositus and the ascidian Polysyncraton lacazei. Somespecies, for example the sea fan Eunicella verrucosa, weremuch more abundant in Brittany than off the Britishcoast although some others such as the alga Carpomitracostata, the cup coral Leptopsammia pruvoti, the anemonesAiptasia mutabilis and Parazoanthus axinellae and soft coralAlcyonium glomeratum were much less abundant thanmight have been expected farther south than GreatBritain. Some northern species were not recorded(Nemertesia ramosa and Flustra foliacea) or were present inlow numbers (Alcyonium digitatum, Urticina felina,Asterias rubens, Echinus esculentus).
Studies using diving techniques in the Glénanarchipelago by L’Hardy-Hales et al. (1973) and byCastric-Fey et al. (1973) provide a description of thecomposition and distribution of rocky sublittoralcommunities very similar to those occurring off south-western Britain. Castric-Fey et al. (1973) described fourdistinct zonal communities (Figure 44):
1. Upper infralittoral community characterised byLaminaria digitata extending from 0 m to 6 m onhorizontal surfaces and +0.5 m to 3 m on verticalsurfaces.
2. Lower infralittoral community characterised by denseLaminaria hyperborea with rich algae and animals to18 m followed by more sparse Laminaria hyperborea (to
a maximum depth of 26 m) and a monotonouscommunity predominantly of the algae Delesseriasanguinea, Bonnemaisonia asparagoides and Dictyopterismembranacea.
3. Upper circalittoral community characterised byAxinellidae and brachiopods from 30 m to 55 m.
4. Lower circalittoral community characterised byDendrophyllia cornigera and Swiftia rosea below 55 m.
A later paper (Castric-Fey, Girard-Descatoire &Lafargue 1978) expanded these descriptions byincluding faunal lists and noting species present incaves, fissures and other habitats. The uppermost zoneis described as the infralittoral fringe in the later paper.
Castric-Fey (1988) also undertook a survey of 21sublittoral sites in the Bay of Concarneau revealing thedistribution of species in relation to wave exposure andturbidity. The results of that study were compared withwork in Britain and Ireland, and several similarcommunities were found to occur. Further work(Castric-Fey & Chassé 1991) described rocky sublittoralcommunities and the distribution of species in relationto various environmental factors in the region of Brest.Again, comparisons are made with work undertaken inBritain and broadly similar distributions of species andassemblages in relation to environmental conditionsrevealed. An extensive study of the Bay of Brest usingmainly underwater video by Hily (1989) provided aclassification which defined 17 different habitat types infive major groups with their associated assemblages ofspecies.
Farther south, off the northern Gascony coast,Glémarec (1973) describes 19 sediment communitiespresent from shallow to deep water divided intoinfralittoral, coastal, and open sea étages (Figure 45).Glémarec (1973) also reviews work undertaken to thenorth of Gascony and around Britain. The communitiesare further described and incorporated with ones for theEnglish Channel in Glémarec (1995). Sediment types andcharacterising species are used and the communities canbe matched to those described elsewhere in theliterature and provide a basis for comparison with thosepresent around Great Britain. Infaunal sedimentcommunities off south Gascony are described by Cornetet al. (1983) whilst epifaunal sediment communities aredescribed in Sorbe (1989).
(A significant publication, Dauvin (1997), whichdescribes marine biocoenoses of Atlantic France, wasreceived after completion of this section but is notreviewed here.)
6.19 Atlantic SpainThe north-western and northern coasts of Spain (Figure46) are predominantly rocky and were the subject of adetailed study of littoral communities by Fischer-Piette(1955), who records species present and theirdistribution along the coast. Littoral fish are recorded byIbanez et al. (1989). These studies, together with thework of Hoek & Donze (1966), emphasise theincreasingly southern character of fauna and floraeastwards along the northern coast of Spain. Various
55
Hiscock: Introduction and Atlantic-European perspective
Figure 44. Schematic profile of the seabed showing zonation ofthe main characterising species for rocky substrata in theGlénan archipelago. (From Castric-Fey et al. 1973.)

papers referred to in Hoek & Donze (1966) also point tothe dynamic nature of fucoid cover along the northerncoast in which fucoids had become increasingly rare inthe first half of the 19th century but had increased inabundance since 1950. However, the general sparsity offucoids greatly affects the appearance of shores andHoek & Donze (1966) note that the marine vegetation ofnorth-west Spain is intermediate in appearance betweenthat of north-west Brittany and the Basque coastreferred to above. Papers by López-Cotelo Viéitez &Diaz-Pineda (1982) and Vilas (1986) include descriptionsof sandy beach fauna in north-west Spain. Here, the
amphipod Talitrus saltator dominated the upper shorewith remaining zones characterised by polychaeteworms: Scolelepis squamata and Scoloplos armiger on theupper midshore, Nephtys hombergii and Nereis (nowHediste) diversicolor on the midshore and Euclmeneoerstedii, Pectinaria koreni and Nephtys hombergii lowerdown. Studies of sublittoral communities appear few.López-Cotelo, Viéitez & Diaz-Pineda (1982) include adescription of polychaete and mollusc communities insublittoral sediments for areas offshore of Santander.Here, a boreal-lusitanean Tellina community in whichthe polychaete worm Nephtys cirrosa was most abundant
56
Marine Nature Conservation Review: benthic marine ecosystems
Figure 45. The distribution of sediment communities on the continental shelf of North Gascony. (Adapted from Glémarec 1973.)

and a reduced Macoma balthica community in which thebivalve molluscs Cerastoderma edule and Scrobiculariaplana with the polychaete worm Nephtys hombergii werecharacteristic. Fine sediments were characterised by aheterogeneous community including the worms Glyceraunicornis, Euclymene oerstedi, Polydora ciliata andPectinaria koreni. Within Santander Bay, Lastra et al.(1990) distinguished two major communities: an Abraalba community in the inner area of the bay and a sandycommunity characterised by the hermit crab Diogenespugilator, the cumacean Iphinoe trispinosa and thepolychaete Nephtys cirrosa in the open bay.
The north-west and west-facing coast is rocky andhighly indented including several rias. The sedimentcommunities of those inlets are described in a series ofpapers by López-Jamar (1978, 1981, 1982 and López-Jamar & Mejuto 1985). López-Jamar & Mejuto (1985)describe sediment communities in the ría of Corunnaand found a Tellina community dominant. Donze (1968)records the marine algal vegetation of the Ría de Arosain north-west Spain, and López-Jamar (1982) theinfaunal communities. He records two majorcommunities: a Spiochaetopterus costarum communityinhabiting anoxic sediments with high organic content,and a Sternaspis scutata – Tharyx marioni communityinhabiting muddy sediments that are not anoxic. In theouter part and partially off the northern shore, there aremore sandy bottoms with three further infaunalassemblages identified. The rias are extensively used formariculture and Sanjurjo (1981) describes the fauna ofmussel ropes in the Ría de Arosa whilst Tenore, Corral &González (1985) assess effects of this intense cultivation.Most southerly along the Spanish coast are the rias ofPontevedra and Vigo. Sandy beach communities here aredescribed by Vieitez (1982), who reports the presence of areduced Macoma community and a boreal lusitaneanTellina community (sensu Thorson 1957). The fauna ofsandy littoral and shallow sublittoral substrata in twoareas outside of the Ría de El Barquero is described in adetailed study by Mazé, Laborda & Luis (1990). Theyfound a boreal lusitanean Tellina community on theBancos Arenosus, an offshore bank, and a clear zonationof fauna consisting mostly of species also found on sandybeaches in Great Britain, on the sandy beach fringing thecoast (the Area Longa beach).
6.20 PortugalThe coast of Portugal (Figure 46) is predominantlyexposed to strong wave action with both sandy androcky habitats. Studies of marine communities havebeen mainly undertaken in the sheltered inlets along thecoast. The ‘lagoons’ of western and southern Portugalare shallow inlets of the sea, some of which arelandlocked and some of which have channelsconnecting them to the sea. Quintino, Rodrigues &Gentil (1989) describe the benthic species in the lagoonof Obidos in western Portugal and conclude that speciescharacteristic of both Atlantic and Mediterraneanlagoons were present. The lagoon of Albufeira isessentially marine with a maximum depth exceeding 13m. Dredge and grab samples of fauna were analysed byQuintino et al. (1987) using correspondence analysis toidentify the communities illustrated in Figure 47. They
concluded that the overall faunistic composition can berelated to the Atlantic and Mediterranean fauna ofcoastal or lagoonal systems. They identify their coarseclean sand assemblage (AI) precisely with the ‘biocenosedes sables grossiers et fins graviers sous l’influencehydrodynamique’ described by Pérès & Picard (1964).Their transition group (AII) includes several speciescharacteristic of the biocenosis ‘sables à Amphioxus’ (nowBranchiostoma) (Pérès & Picard 1964; Bellan 1964). Thethird group, which covered about 90% of the lagoonfloor, was considered typical of coastal lagoons both forthe Atlantic and Mediterranean and is related to the‘biocenose lagunaire euryhaline et eurytherme’ of Pérès& Picard (1964) and Bellan (1964). The Ría de Aveiro isdescribed by Moriera et al. (1993). The ‘ria’ extends as theCanal de Mira along which is a salinity gradient from 35to 0‰ providing a typical longitudinal estuarinegradient. Analysis of biological samples identified threemajor groupings representing subtidal stations in theouter channel (separated into three groups), intertidalstations in the middle and outer reaches and innerstations. The numerically dominant species in thesegroups (the polychaetes Hediste diversicolor, Amagesadspersa, Streblospio dekuyzeni, Pygospio elegans, Tharyxmarioni and Heteromastus filiformis, the crustaceansUrothoe brevicornis, Cyathura carinata and Corophiummultisetosum, the prosobranchs Hydrobia ulvae andPotamopyrgus jenkensi, and the bivalves Scrobiculariaplana, Cerastoderma edule and Venerupis pullastra) aremostly species which would characterise similar habitatsin Britain. Studies have also been undertaken in the ‘ria’of Alvor on the southern coast of Portugal (for instance,Rodrigues & Dauvin 1987). Communities are describedaccording to the hydrosedimentary gradient as: amedium clean sand community near the mouth, anintermediate community and a landward community onsandy mud. The fauna was considered typical ofEuropean coastal lagoons.
The lagoon of St Andre is a landlocked system.Benthic and fish communities were described by daFonseca, Costa & Bernardo (1989). There is a markedchange in the fauna through the year in relation tochanges in salinity, and the species (most of which alsooccur in Britain) characteristic of the different salinityregimes are listed. The Ría Formosa is a large area ofenclosed coast with four openings into the Atlantic.Austen, Warwick & Rosado (1989) note that the benthicecology of the area is not very well known. Their workincluded the sampling of macro-and meiofauna along agradient of sewage pollution identifying organismsgenerally to family level. Thus, no comparison withcommunities elsewhere is possible although it is notablethat they conclude digging for shellfish disturbsmacrobenthic communities well beyond any influencefrom sewage.
For the open coast, Dexter (1988, 1990) describes thesandy beach fauna from 60 locations. The zonation shedescribed was of: the talitrid Talitrus saltator and theoniscid Tylos europeus isopods in the supralittoral fringe;cirolanid isopods especially Eurydice affinis in the‘retention’ zone (see McLachlane & Jaramillo 1995 forterminology); Spio filiformis, Nephtys cirrosa, haustoriid,urothoid, oedicerotid and pontoporinid amphipods and
57
Hiscock: Introduction and Atlantic-European perspective

the isopods Spaeroma rugicauda and Spaeroma hookeri inthe resurgence zone and Donax trunculus, Tellina tenuis,Gastrosaccus sanctus and the crab Portumnus latipes in thesaturation zone.
6.21 The MediterraneanThe Mediterranean Sea includes many species andfeatures in common with the Atlantic but also manyspecies and features not found west and north of theStraits of Gibraltar. Similarities with the Atlantic as wellas species richness generally decreases with increasingdistance eastwards. Much marine biological work hasbeen carried out in the Mediterranean. This contributesto our knowledge of the taxonomy and ecology ofmarine species, to the sampling techniques that can beused to record and separate marine communities and tothe identification of biotopes. The absence of anextensive littoral zone and the warmer waters in theMediterranean encouraged the study of sublittoral areasusing diving techniques much earlier than in the north-east Atlantic. A remarkable early study that providesmuch information on the ecology of hard substratumsublittoral species and communities is the work by Riedl(1966) Biologie der Meereshöhlen. However, the manystudies undertaken on specifically Mediterraneansystems are not described here but attention is given towork aimed at classifying benthic marine habitats andcommunities, mainly undertaken on the French andSpanish coasts, which provides important reference forMNCR studies.
58
Marine Nature Conservation Review: benthic marine ecosystems
SPAIN
SantanderLa Coruna
Vigo
MOROCCO
ALGERIA
2˚E2˚E
36˚36˚
San Sebastian
Lisbon
FRANCE
PORTUGAL
N
Strait of Gibraltar
La Coruna SantanderSan Sebastian
Ria de Vigo
Ria dePontevedra
BalearicIslands
Bay of Biscay
Lisbon
Albufeira
FRANCE
PORTUGALSPAIN
MOROCCO
GIBRALTAR
0 200
km
10˚W 6˚ 2˚ 2˚E
42˚N
40˚
38˚
36˚Mediterranean Sea
ALGERIA
MedesIslands
Ria de Barquero
Ria deArosa
Vigo
Ria de Aveiro& Canalde Mira
LagunaSt Andre
Ria Formosa
Figure 46. Spain, Portugal and Mediterranean France, showing locations of places mentioned in the text.
Figure 47. Distribution of macrobenthic communities in thelagoon at Albufeira. AI = marine assemblage of Saccocirruspapillocercus, Nephtys cirrosa and Spisula ovalis; AII = transitionassemblage; AIIa = high richness group; AIIb = medium sandswith Spio martinensis; AIII = lagoon assemblage; AIIIa = sandymud mainly of Abra ovata and Heteromastus filiformis; AIIIb =impoverished muds with Phoronis psammophila. (From Quintinoet al. 1987.)

The work of Pérès & Molinier (1957) provides thebasis for defining zonation of epibenthic communities inthe sublittoral which, although mainly based on theMediterranean coast, is easily converted to work innorth-east Atlantic ecosystems at a broad scale (Hiscock& Mitchell 1980). Pérès & Picard (1964) extended thezonal classification to one for the major assemblages ofspecies in the Mediterranean benthos in their Nouveaumanuel de bionomie benthique de la Méditerranée. Theirconclusions are largely repeated, in English, in Pérès(1967), where the extent and depths of the major zonesare illustrated (Figure 48). Augier (1982) provides aninventory and classification of marine benthicbiocoenoses of the Mediterranean derived greatly fromthe work of Pérès & Picard (1964) and which reviews themajor works. Bellan-Santini, Lacaze & Poizat (1994) givean up-to-date summary of the marine biocoenoses andthreats to them in the Mediterranean. The system ofclassification for benthic communities established nowsince 1957 in the Mediterranean has provided theopportunity to map the extent of distinctive habitats andcommunities as a tool in management for natureconservation. For example, Augier & Boudouresque(1967) map marine benthic biocoenoses in the Port-CrosNational Park (Figure 49). The studies of marine benthichabitats and communities undertaken in the MedesIslands off the north-east coast of Spain (Ros, Olivella &Gili 1984) also followed the classification system firstestablished by Pérès & Picard (1964) and the inventorywith detailed descriptions of species richness and thedistribution of communities thus provided is the basisfor their proposal of protected status for the islands.
It is the high proportion of the species present in theMediterranean which do not occur or do not occur inabundance on the Atlantic coast of Europe (in shallowwaters at least – below a depth of about 200 m, Carpine(1970) records communities with many species similar tothose of muddy substrata in the north-east Atlanticincluding a close similarity between the Mediterraneanbathyal communities and those from the deep water inSognfjord in Norway) and the great differences indominant species of particular habitats which makesdirect comparison or integration of classifications at adetailed level difficult between the Mediterranean andAtlantic coasts of Europe. Nevertheless, the frameworkfor a classification which can be applied across Europeshould be the same for all of the north temperateAtlantic area.
7 AcknowledgementsThis chapter was originally circulated as a MNCROccasional Report (Hiscock 1991). That report benefitedparticularly from an earlier review of marine benthicbiocenoses in the North Sea and Baltic undertaken by DrK. Probert and published in Mitchell (1987) and from thebibliography published by Palmer, Mitchell & Probert(1983). I am particularly grateful for comments on theOccasional Report from Dr M. Costello, Dr R.Hammond, Dr S. Hawkins, Dr S. Hull, Dr S. McGrorty,Professor A.D. McIntyre and Dr G. Rappé.
Dr M. Elliott, Dr P. Kingston and E.I.S. Rees arethanked for their comments on the final draft of thischapter and Dr L.M. Davies is thanked for comments onan early draft.
Much of the information-gathering for this report hasbeen undertaken at the National Marine BiologicalLibrary at Plymouth and I am grateful to the staff thereboth for the excellence of the facilities they provide andfor always giving help when needed.
59
Hiscock: Introduction and Atlantic-European perspective
Figure 48. General scheme of the distribution of the mainbiocoenoses on the French coast of the Mediterranean: 1, rockypoints; 2, alluvial area; 3, high and middle beach (supralittoraland mediolittoral sandy biocoenoses); 4, biocoenoses of the finesands in very shallow waters; 5, biocoenosis of the well sortedsands; 6, biocoenosis of the photophilous algae on rockysubstratum; 7, biocoenosis of the Posidonia meadows; 8,coralligenous biocoenosis; 9, biocoenosis of the coastal detritic;10, biocoenosis of the terrigenous mud; 11, biocoenosis of theshelf-edge detritic; 12, biocoenosis of the bathyal mud; 13,biocoenosis of the deep sea corals. (From Pérès 1967.)
Figure 49. Marine benthic biocoenoses in the Bay of La Palu, Port-Cros National Park, France. (From Augier & Boudouresque 1967.)