binding of c-reactive protein to chromatin and … .journal of biological chemistry vol. 259, no....
TRANSCRIPT

THE . J O U R N A L OF BIOLOGICAL CHEMISTRY Vol. 259, No. 11, Issue of June 10, pp. 7311-7316, 1984 Prrnted in U S.A.
Binding of C-reactive Protein to Chromatin and Nucleosome Core Particles A POSSIBLE PHYSIOLOGICAL ROLE OF C-REACTIVE PROTEIN*
(Received for publication, November 28,1983)
Frank A. Robey, Karen D. Jones, Taeko Tanaka, and Teh-Yung Liu From the Division of Biochemistry and Biophysics, Office of Biologics Research and Review, National Center for Drugs and Biologics, Food and Drug Administration, Bethesda, Maryland 20205
By using a variety of biochemical techniques, chro- matin and chromatin fragments have been identified as probable physiological ligands for C-reactive pro- tein. Studies using 14C-labeled C-reactive protein show that binding to chromatin is saturable with a K, = 8 x
M, a value indicating that the affinity of C-reac- tive protein for chromatin is at least four times its affinity for phosphorylcholine. At saturation, there is approximately one C-reactive protein-binding site for every 160 base pairs of DNA in chromatin. The inter- action of C-reactive protein with chicken erythrocyte nucleosome core particle has been studied. Fifty per cent inhibition of the binding of C-reactive protein to phosphorylcholine is obtained at a core particle con- centration of 1.25 X lo-@ M, indicating that the affinity of C-reactive protein for one of the sites on core par- ticles is at least 2400 times greater than the affinity of C-reactive protein for phosphorylcholine. The possi- bility that C-reactive protein may act as a scavenger for chromatin fragments released from damaged cells is discussed.
C-reactive protein is an acute phase serum protein in most higher order mammals. In humans, serum levels of CRP' may increase by as much as 2000-fold during the inflammatory process or other pathological events (1, 2).
Since CRP was discovered in 1930 by Tillet and Francis (3), many speculations with regard to its function have been offered, but, as yet, none compellingly argue for a unique role for this protein. Several of the biological properties ascribed of CRP resemble those of immunoglobulins. Like the immu- noglobulins, CRP possesses the ability to promote reactions of precipitation (3) and agglutination (4), bacterial capsular swelling (5), phagocytosis (6), and complement fixation (7). However, there is no apparent amino acid sequence homology between CRP and the immunoglobulins (8).
In 1971, Volanakis and Kaplan (9) showed that PC was the most effective inhibitor of the CRP-pneumococcal C-polysac- charide precipitation reaction. This observation led to the development of a simple one-step purification scheme for CRP (10, 11) and the suggestion that CRP might exert its
* The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
' The abbreviations used are: CRP, C-reactive protein; PC, phos- phorylcholine; FITC, fluorescein isothiocyanate; BME, basal Eagle's medium; ELISA, enzyme-linked immunosorbent assay; TLCK, N"- p-tosyl-L-lysine cbloromethyl ketone; BSA-PC, bovine serum albu- min-phosphorylcholine conjugate.
physiological function through Ca2+-dependent binding to PC. In this report, we summarize our finding that CRP binds
to chromatin and chromatin fragments which may be present in the extracellular milieu at sites of tissue damage. That this property may be related to the physiological role of CRP is discussed in relation to the above phenomena involving CRP, especially phagocytosis and complement fixation.
MATERIALS AND METHODS
The CRP used in these studies was purified from acute phase rabbit sera using a PC-Sepharose affinity resin as described by Oliveira et al. (11).
['4C]CRP-['4C]CRP was prepared by reacting 5 mg of rabbit CRP in 2 ml of 0.15 M NaCl, 0.05 M Tris, 0.01 M CaC12, pH 7.4, buffer with 1 mCi of ["CC)CH,I (New England Nuclear; specific activity = 59 mCi/mmol). The reaction was allowed to proceed at room temper- ature for 16 h, and the ['*CJCRP was purified by affinity chromatog- raphy using PC-Sepharose (11). The specific activity of the final product was 0.02 pCi/pg of CRP.
CRP-Sepharose-CRP-Sepharose was prepared by reacting 30 mg of purified rabbit CRP in 0.1 M NaHC03, 0.1 M EDTA, pH 8.0, with 2.0 g of activated Sepbarose CH-4B (Pharmacia Fine Chemicals). After 2 h at ambient temperature, the residual reactive groups on the Sepharose were destroyed by addition of ethanolamine (Sigma) to give a final concentration of 0.1 mM, and the reaction was allowed to proceed for 16 h at 4 "C. The gel was then washed with 500 ml of buffer containing 0.15 M NaCI, 0.05 M Tris, pH 7.4, 0.01 M CaC12.
FZTC-CRP-FITC-CRP was prepared by reacting 10 mg of fluo- rescein isothiocyanate (Aldrich) with 5 mg of CRP in the above Tris, saline, 0.01 M EDTA, pH 7.4, buffer (1.0 ml). The solution was mixed rapidly for 1 min at ambient temperature and then filtered through a Millipore membrane (0.22 pm). After standing a t room temperature for 1 h, the filtrate (1 ml) was added to 1 ml of saturated Na2S0, and the FITC-CRP was precipitated. The FITC-CRP precipitate was pelleted by centrifugation a t 15,000 rpm for 5 min in a Eppendorf centrifuge. The product was dissolved in Tris/saline/Ca2+ buffer (0.5 ml), and the FITC-CRP was precipitated again after the addition of 0.5 ml of saturated Na2S0,. The above precipitation-dissolution pro- cedure was repeated three times more, and the FITC-CRP was dialyzed extensively against either the Tris/saline/Ca2+ buffer or BME. The quantity of FITC-CRP used in the experiments was determined by the PC ELISA (12).
Affinity-purified Goat Anti-CRP Ig-The preparation of affinity- purified goat anti-CRP Ig was accomplished exactly as previously described (13).
Goat Anti-CRP Ig-Sepharose-The goat anti-CRP Ig was immo- bilized on Sepharose by reacting 20 mg of affinity-purified goat anti- rabbit CRP Ig in 0.1 M NaHCO, a t p H 8.0 with 2.0 g of activated Sepharose CH-4B. The rest of the preparation was exactly as de- scribed above for the preparation of CRP-Sepharose.
Preparation of Chromatin-Chromatin was isolated from rabbit liver nuclei using the method of Rizzo and Bustin (14) except a hypotonic buffer of 0.015 M NaC1, 5 mM Tris, 0.3 mM MgC12, 0.3 mM CaCL, and 0.1 mM TLCK, pH 7.4, a t 4 "C was used throughout the preparation of the nuclei, and the nuclei were lysed by homogenization in 0.015 M NaC1, 5 mM Tris, 1 mM EDTA, 0.1 mM TLCK, pH 7.4, in a Type B Dounce homogenizer. The nuclear lysate was layered on a
7311

7312 CRP Reacts with Chromatin 1.7 M sucrose cushion and subjected to centrlfugation to give the chromatin pellet as described in Ref. 14. The quantity of chromatin used in the binding experiments was determined spectroscopically in the presence of 0.1% sodium dodecyl sulfate, 0.1 N NaOH using an extinction coefficient a t 260 nm of 0.05 ml mg”.
Chicken Erythrocyte Nucleosome Core Particles-The crude nu- cleosome core particles were prepared by a limited micrococcal nu- clease digestion of chicken erythrocyte chromatin followed by puri- fication through a sucrose gradient containing 0.5 M NaCl and was supplied to us as a gift from Dr. Robert T. Simpson at National Institutes of Health, Bethesda, MD. The exact procedure is described in Ref. 15. Core particles with an average molecular weight of 200,000 were used for inhibition studies?
Binding of /“C/CRP to Chromatin-In a typical binding experi- ment, 34.5 pg of chromatin in 100 pl of Tris/saline/Ca2+ buffer were added to several 1.5-ml Eppendorf microtest tubes followed by pre- determined varying amounts of buffer and [“CJCRP to give a final volume of 0.5 ml. After centrifugation at 15,000 rpm for 1 min, 100 ~l of the supernatant were withdrawn and added to 10 ml of Aquasol (New England Nuclear) for scintillation counting. Standards were run under identical conditions but the chromatin was omitted. The difference between the total counts in the standard minus the total supernatant counts with chromatin present gives the number of counts of [“CICRP bound to the chromatin.
Inhibition of [“CICRP binding to chromatin by PC or by unlabeled CRP was studied by adding varying amounts of PC or unlabeled CRP to solutions containing 34.5 pg of chromatin, 1 pM [14C]CRP in Tris/ saline/Ca2+, pH 7.4, buffer a t a total volume of 0.5 ml. The suspension was centrifuged, and the amount of [“CICRP in the supernatant was determined as above.
PC ELISA-The inhibition of CRP binding to PC by core particles was studied using the PC ELISA developed in this laboratory without modification (12). Serial dilutions of inhibitor were made in the wells of BSA-PC-coated polyvinyl chloride microtiter plates followed by the addition of 10 ng of purified rabbit CRP/well. The quantity of CRP bound to BSA-PC in the presence of inhibitor was determined by the PC ELISA as described in Ref. 12. Further details are given in the legends to the appropriate figures.
Agarose Diffusion Studies-The reactivity of core particles with CRP was demonstrated on agarose plates (3 X 5 inches) by addition of noted volumes of the reagents in wells set in the agarose. The buffer used for the 0.5% agarose was 0.03 M NaCI, 0.002 M CaCI2, and 0.01 M Tris, pH 7.4. The Tris/saline/Ca2+ buffer used for most experiments described above caused core particles to precipitate, so it was necessary to use this lower ionic strength buffer.
BSA-PC was prepared exactly as described (11). All other materials and methods are described in the figure legends.
RESULTS
CRP Binding to Damaged Rabbit Skin and Lung Fibroblasts in Culture-CRP is known to accumulate at the site of tissue injury. In order to study the possible physiological ligand for CRP, FITC-CRP was synthesized to directly visualize the binding of CRP to damaged cells in culture. Fig. 1 shows rabbit lung fibroblasts treated with either 1.0 pg/ml of bra- dykinin or sublytic amounts of bee venom phospholipase AZ or lo-’ M egg lysolecithin. Bradykinin causes hydrolysis of phospholipid in many cell types through activation of endog- enous phospholipases (17, 18). Treatment of cultured lung fibroblasts with 1 pg/ml of bradykinin for 30 min at 37 “C resulted in approximately 5% cell death as measured by the appearance of lactate dehydrogenase in the media (19). We observed dead cells that had incorporated FITC-CRP as a result of bradykinin treatment. In Fig. 1, a cluster of fluoresc- ing attached cells is shown. The same type of fluorescent pattern seen in Fig. 1 was observed when FITC-CRP was added to cells treated with sublytic quantities of phospholi- pase Az or lysolecithin (lo-’ M). Because these cells allow the passage of CRP through the membrane, the cells are presumed to be dead. Most of the fluorescent material was seen as punctate patterns in the nuclei of cells which do fluoresce.
R. T. Simpson, personal communication.
FIG. 1. Damaged rabbit lung fibroblasts treated with FITC- CRP. Rabbit lung fibroblasts (CCL 193; American Type Culture Collection) were grown to 40-50% confluency in BME on tissue culture chamber slides (No. 4808, Miles Laboratories, Inc.). The fibroblasts were treated for 30 min with 1 pg/ml bradykinin (Sigma) in BME followed by the addition of 100 pg/ml of FITC-CRP. The slide was washed three times in BME and observed under the fluo- rescence microscope (Leitz). The picture shown is an enlarged print of a slide taken at a magnification of X 100. The quantity of FITC- CRP used in the experiment was determined by the PC ELISA (12).
There was no indication of FITC-CRP binding to the plasma or nuclear membrane despite the probable presence of lyso- lecithin in these membranes. FITC-CRP uptake also occurred at 4 “C after bradykinin treatment at 37 “C, indicating passive diffusion of CRP to the nucleus. Isolated nuclei (not shown) which were not stimulated with any agent included FITC- CRP with a similar spotty pattern as seen in the intact damaged cells. By adding a 100-fold excess of unlabeled CRP or 0.1 mM PC, the punctate fluorescent pattern in the cell nuclei or isolated nuclei could be abolished, resulting in diffuse homogeneous fluorescence in the nuclei. The same results were obtained using rabbit skin fibroblasts (CRL 1414, Amer- ican Type Culture Collection). No fluorescence was detected in cell cultures where the plasma membranes of the cells were not damaged by treatment with bradykinin, phospholipase A*, or lysolecithin prior to the addition of FITC-CRP to the cell culture.
Controls using FITC-rabbit serum albumin or FITC alone showed that damaged cells did include these fluorescent sub- stances, but the detailed spots seen with FITC-CRP were not apparent, i.e. with FITC-albumin or FITC alone, nuclear fluorescence appeared totally diffuse.
CRP Reacts with Chromatin 7313
The key point of these experiments is to show that if the plasma membrane of a cell is damaged to the extent that exogenous substances such as CRP can enter the cell, CRP will bind specifically to a major component in the damaged cell's nucleus. That one of these CRP-binding substances is chromatin is shown in the experiments below.
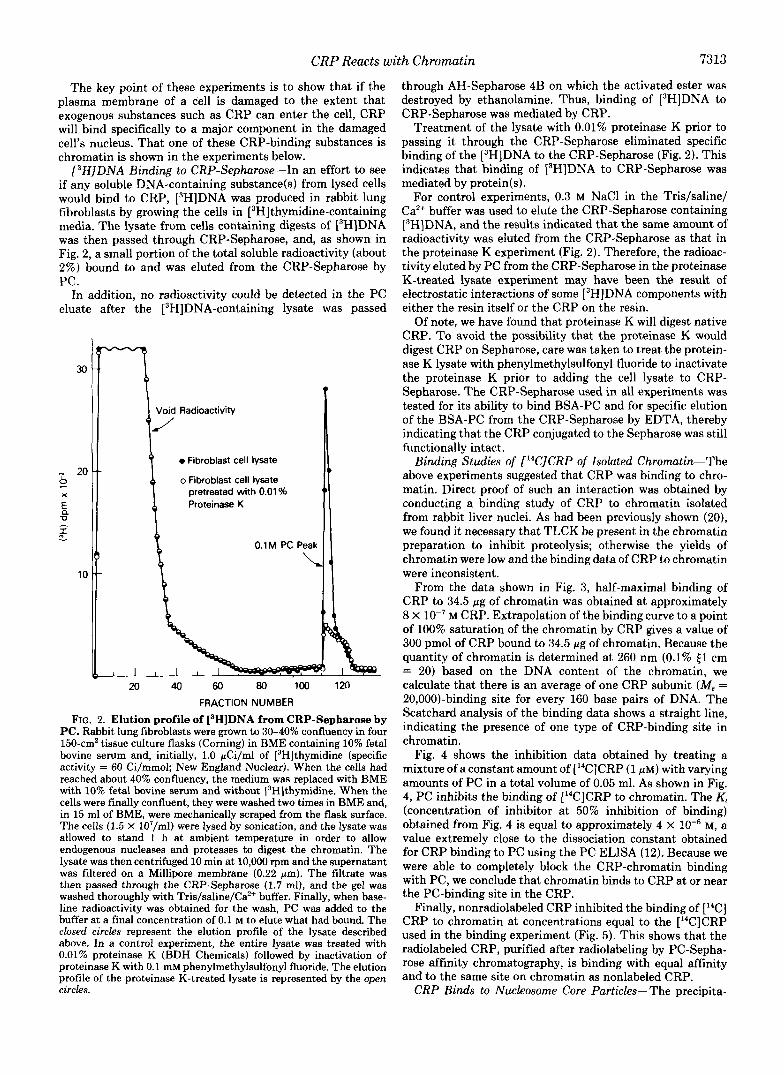
[3H]DNA Binding to CRP-Sepharose-In an effort to see if any soluble DNA-containing substance(s) from lysed cells would bind to CRP, [3H]DNA was produced in rabbit lung fibroblasts by growing the cells in [3H]thymidine-containing media. The lysate from cells containing digests of L3H]DNA was then passed through CRP-Sepharose, and, as shown in Fig. 2, a small portion of the total soluble radioactivity (about 2%) bound to and was eluted from the CRP-Sepharose by PC.
In addition, no radioactivity could be detected in the PC eluate after the [3H]DNA-containing lysate was passed
Void Radioactivity
1 0 Fibroblast cell lysate
o Fibroblast cell lysate n 20 9 X pretreated with 0.01 %
E, Proteinase K 0
I - -
0.1 M PC Peak
10
20 40 60 80 100 120
FRACTION NUMBER
FIG. 2. Elution profile of [3H]DNA from CRP-Sepharose by PC. Rabbit lung fibroblasts were grown to 30-40% confluency in four 150-cmZ tissue culture flasks (Corning) in BME containing 10% fetal bovine serum and, initially, 1.0 pCi/ml of [3H]thymidine (specific activity = 60 Ci/mmol; New England Nuclear). When the cells had reached about 40% confluency, the medium was replaced with BME with 10% fetal bovine serum and without [3H]thymidine. When the cells were finally confluent, they were washed two times in BME and, in 15 ml of BME, were mechanically scraped from the flask surface. The cells (1.5 X 107/ml) were lysed by sonication, and the lysate was allowed to stand 1 h at ambient temperature in order to allow endogenous nucleases and proteases to digest the chromatin. The lysate was then centrifuged 10 min at 10,000 rpm and the supernatant was filtered on a Millipore membrane (0.22 pm). The filtrate was then passed through the CRP-Sepharose (1.7 ml), and the gel was washed thoroughly with Tris/saline/Ca2+ buffer. Finally, when base- line radioactivity was obtained for the wash, PC was added to the buffer at a final concentration of 0.1 M to elute what had bound. The closed circles represent the elution profile of the lysate described above. In a control experiment, the entire lysate was treated with 0.01% proteinase K (BDH Chemicals) followed by inactivation of proteinase K with 0.1 mM phenylmethylsulfonyl fluoride. The elution profile of the proteinase K-treated lysate is represented by the open circles.
through AH-Sepharose 4B on which the activated ester was destroyed by ethanolamine. Thus, binding of [3H]DNA to CRP-Sepharose was mediated by CRP.
Treatment of the lysate with 0.01% proteinase K prior to passing it through the CRP-Sepharose eliminated specific binding of the [3H]DNA to the CRP-Sepharose (Fig. 2). This indicates that binding of i3H]DNA to CRP-Sepharose was mediated by protein(s).
For control experiments, 0.3 M NaCl in the Tris/saline/ Ca2+ buffer was used to elute the CRP-Sepharose containing [3H]DNA, and the results indicated that the same amount of radioactivity was eluted from the CRP-Sepharose as that in the proteinase K experiment (Fig. 2). Therefore, the radioac- tivity eluted by PC from the CRP-Sepharose in the proteinase K-treated lysate experiment may have been the result of electrostatic interactions of some [3H]DNA components with either the resin itself or the CRP on the resin.
Of note, we have found that proteinase K will digest native CRP. To avoid the possibility that the proteinase K would digest CRP on Sepharose, care was taken to treat the protein- ase K lysate with phenylmethylsulfonyl fluoride to inactivate the proteinase K prior to adding the cell lysate to CRP- Sepharose. The CRP-Sepharose used in all experiments was tested for its ability to bind BSA-PC and for specific elution of the BSA-PC from the CRP-Sepharose by EDTA, thereby indicating that the CRP conjugated to the Sepharose was still functionally intact.
Binding Studies of [I4C]CRP of Isolated Chromatin-The above experiments suggested that CRP was binding to chro- matin. Direct proof of such an interaction was obtained by conducting a binding study of CRP to chromatin isolated from rabbit liver nuclei. As had been previously shown (20), we found it necessary that TLCK be present in the chromatin preparation to inhibit proteolysis; otherwise the yields of chromatin were low and the binding data of CRP to chromatin were inconsistent.
From the data shown in Fig. 3, half-maximal binding of CRP to 34.5 pg of chromatin was obtained at approximately 8 X lo-' M CRP. Extrapolation of the binding curve to a point of 100% saturation of the chromatin by CRP gives a value of 300 pmol of CRP bound to 34.5 pg of chromatin. Because the quantity of chromatin is determined at 260 nm (0.1% E1 cm = 20) based on the DNA content of the chromatin, we calculate that there is an average of one CRP subunit (M, = 20,00O)-binding site for every 160 base pairs of DNA. The Scatchard analysis of the binding data shows a straight line, indicating the presence of one type of CRP-binding site in chromatin.
Fig. 4 shows the inhibition data obtained by treating a mixture of a constant amount of [14C]CRP (1 PM) with varying amounts of PC in a total volume of 0.05 ml. As shown in Fig. 4, PC inhibits the binding of [14C]CRP to chromatin. The K, (concentration of inhibitor at 50% inhibition of binding) obtained from Fig. 4 is equal to approximately 4 X M, a value extremely close to the dissociation constant obtained for CRP binding to PC using the PC ELISA (12). Because we were able to completely block the CRP-chromatin binding with PC, we conclude that chromatin binds to CRP at or near the PC-binding site in the CRP.
Finally, nonradiolabeled CRP inhibited the binding of [14C] CRP to chromatin at concentrations equal to the [14C]CRP used in the binding experiment (Fig. 5). This shows that the radiolabeled CRP, purified after radiolabeling by PC-Sepha- rose affinity chromatography, is binding with equal affinity and to the same site on chromatin as nonlabeled CRP.
CRP Binds to Nucleosome Core Particles-The precipita-

7314
1 .o 0.9
0.8
0.7
E 0.6 a 5 0.5 0 m 0.4
0.3
0.2
0.1
W W
CRP Reacts with Chromatin
1 2 3 4
CRP BOUND (xlO”Omol) FIG. 3. Scatchard plot and binding curve of [“CICRP bind-
ing to isolated chromatin. Experimental details are given under “Materials and Methods”.
I d
I 0 I I I 0.1 1 .o 10 100
[PHOSPHORYLCHOLINE], pm
FIG. 4. Inhibition of [“CICRP binding to chromatin by PC. Experimental details are given under “Materials and Methods.”
tion reaction of CRP with core particles is shown in Fig. 6. Also shown is the precipitation of CRP with BSA-PC. The single line of identity seen between the core particles and the BSA-PC indicates that both substances are binding to CRP through the same binding site in the CRP.
To visualize the precipitate formed between CRP and core particles, the sodium dodecyl sulfate-polyacrylamide gel sys- tem described by LeStourgen and Rusch (21) was used. The gels (Fig. 7) clearly show that CRP precipitates with intact core particles and probably requires the protein component as well as the DNA component.
Core particles pretreated with DNase I or proteinase K failed to precipitate with CRP in the presence of Ca2+. How- ever, the DNase I-treated core particles may bind to CRP without forming a precipitate as revealed by the experiment shown in Fig. 2. An estimate of this binding by the PC ELISA
100 0 1 0
I s
0
1 .o 10 100 m CRP.pgm
FIG. 5. Inhibition of [“CICRP binding to chromatin by CRP. Experimental procedure was exactly the same as described for inhibition by PC except incremental amounts of nonradiolabeled CRP were used with 23 pg of [“CICRP in each triplicate experiment.
FIG. 6. Agarose diffusion experiment demonstrating the reactivity of CRP with nucleosome core particles. Plates are 0.5% agarose set in 0.03 M NaCl, 0.01 M Tris, pH 7.4, 0.002 M CaCh, 0.01% NaN3. A, 10 pl of nucleosome core particles (1 mg/ml); B, 10 pl of BSA-PC (2 mg/ml); C, 0.5 mg/ml of purified rabbit CRP (20 4 ) .
was not possible due to interference of the PC ELISA by the DNase I-treated core particles similar to that seen for poly- cations such as poly-L-lysine and histones (12). Proteinase K- treated core particles did not bind to CRP as indicated by the PC ELISA, clearly showing that the protein component of core particles is essential for binding CRP.
Finally, the inhibition of CRP binding to PC in the PC ELISA by core particles is shown in Fig. 8. Using an average molecular weight of 200,000 for the core particles, 50% inhi- bition of CRP binding to core particles occurs at approxi- mately 1.25 X lo-’ M core particles. This affinity is a t least 2,400 times that value obtained for the 50% inhibition of CRP binding to PC by PC in the same assay, i.e. 2.95 X M (12). Although there may be several CRP-binding sites on the core particles there appears to be a single very high affinity binding site; 50% inhibition of CRP binding to PC was occurring a t a core particle concentration which was half the concentration of CRP used in the assay. There must be at least two binding sites for CRP on the core particles otherwise the precipitation reaction between CRP and core particle (Fig. 6) would probably not have been observed.
Calcium-dependent Binding of CRP to Chromatin-All of the binding of CRP to chromatin and chromatin fragments

CRP Reacts with Chromatin 7315
1 2
CRP-
3 4
4 5 K
30 K
2 0 K
1 2 K
FIG. 7. Sodium dodecyl sulfate-polyacrylamide gel electro- phoresis of CRP (lane I ) , core particles (lane 2), and the precipitate formed on mixing CRP with core particles (lane 3). Lane 4 contains molecular weight standards (45K represents, for example, M, = 45,000). The gel was stained for protein with Coomas- sie brilliant blue R-250. The buffer system of LeStourgeon and Rusch (21) was used. The separating gel contained 18% acrylamide, 0.16% bisacrylamide, and the stacking gel contained 3% acrylamide, 0.08% bisacrylamide. The precipitate was formed by mixing 50 pl of core particles (1 mg/ml) in 0.03 M NaCI, 0.001 M Tris, pH 7.4, 0.002 M CaCI2 with 50 pl of purified rabbit CRP (0.25 mg/ml). The suspension was centrifuged, and the pellet was washed three times in the above buffer before running the sodium dodecyl sulfate-polyacrylamide gel electrophoresis.
100
70 /- / 0.1 1 .o 10 100
[NUCLEOSOME CORE PARTICLEI nM
FIG. 8. Inhibition of the binding of CRP to immobilized PC by core particles using the PC ELISA (12). The initial concen- tration of core particle was 0.1 mg/ml, and serial dilutions of this were made to evaluate the Ki.
described in this manuscript is inhibited by EDTA, indicating that calcium is required for this interaction.
DISCUSSION
CRP is the prototypical acute phase reactant, i.e. unlike any other serum protein, the CRP levels in serum rapidly
increase by as much as 2000-fold during the onset of certain diseases (1,2). This fact, plus the finding that CRP is a major blood constituent of primitive animals such as the horseshoe crab Limulus polyphemus (22) and the dogfish shark (13), argue strongly for an important physiological role for this protein.
CRPs exist either as pentamers or hexamers (23). A recent study from this laboratory (24) using spin-labeled derivatives of PC showed that the subunits which comprise the polymers of the respective human, dogfish, or Limulus CRPs bind PC noncooperatively. From a functional point of view, it would seem logical that CRP has such a geometry for the purpose of cross-linking ligands to form aggregates. By virtue of the strong affinity of CRP for PC (9), it has been suggested that CRP may exert its effect by binding to PC in damaged membranes where PC is a major constituent. How the penta- or hexameric configuration of CRP interacts with the PC moiety in the membrane to exert its effect is not clear.
This study shows that, in uitro, CRP forms a calcium- dependent complex with chromatin. If we accept the hypoth- esis that CRP mediates clearance of certain ligands which are exposed as a result of tissue damage, two general mechanisms of clearance are possible. First, a CRP-chromatin complex may be amenable to complement-mediated phagocytosis. This hypothesis receives support from the recent finding of Volan- akis (25) that CRP-C-polysaccharide complexes are solublized by the complement system. Second, the CRP-chromatin com- plexes may travel directly to the liver or spleen where they are degraded. A recent report dealing with the half-life of CRP in blood revealed a noticeably short half-life of CRP in uiuo, i.e. 5.5 h (26), indicating that, in response to inflamma- tion, the liver constantly produces CRP in order to maintain high levels of the protein in serum. CRP synthesized in the liver is delivered to the site of inflammation where it could bind to chromatin fragments and the complex could be se- questered in the liver or spleen. With a K d of a t least lo-’ M for the binding of CRP to core particles, such a mechanism may be reasonable. In Limulus where there are no sophisti- cated immune systems such as immunoglobulins and phago- cytic cells but where CRP is a major serum component (22), this proposal would be particularly attractive.
Neither naked DNA nor histone alone binds to CRP in a PC-inhibitable, Ca2+-dependent manner (12, 27). In the cur- rent series of experiments, we have demonstrated that CRP contains a site capable of interacting with chromatin, a con- jugate of DNA and histones. This site on CRP is either contiguous with or modulated by events in the PC-binding site since the binding with chromatin is calcium-dependent and readily inhibitable with PC. It is conceivable that CRP may be binding to chromatin through the DNA-histone region where a basic group of lysine or arginine from histone is adjacent to a negatively charged phosphate moiety in DNA. Such a binding site on chromatin would be far more favorable and efficient than PC to bind to CRP since the K d for one of the sites on core particles binding to CRP is at least 2400 times that found for PC binding to CRP. However, we have found no single compound which can mimic the DNA-histone- binding region as well as PC.
Although CRP was discovered over fifty years ago (3), its definitive function has been unknown. As we show in this paper, CRP binds to chromatin and chromatin fragments, and we suggest that this may be a suitable starting point from which CRP function can be explored. Presently, the fate of chromatin fragments released from cells as a result of damage or death to the cells is unknown. Likewise, whether or not these chromatin fragments are harmful to the body, possibly

7316 CRP Reacts with Chromatin
as oncogenes, or are totally innocuous is not known. We suggest that CRP may help mediate the removal of chromatin and/or chromatin fragments from the body after cell death provided the barriers separating CRP from chromatin are destroyed.
Acknoruledgments-We are pleased to acknowledge Dr. Michael Bustin’s helpful discussions on the preparation and properties of chromatin and Dr. Robert T. Simpson for the generous gift of core particle. The clerical assistance of Nancy Wyne is appreciated.
REFERENCES 1. Claus, D. R., Osmand, A. P., and Gewurz, H. (1976) J. Lab. Clin.
2. Morley, J. J., and Kushner, I. (1982) Ann. N. Y. Acad. Sci. 389 ,
3. Tillet, W. S., and Francis, T., Jr. (1930) J. Exp. Med. 52, 561-
4. Gal, K., and Miltenyi, M. (1955) Acta Microbiol. Acad. Sci. Hung.
5. Lofstrom, G . (1944) Br. J. Exp. Pathol. 25, 21-26 6. Hokema, Y., Coleman, M. K., and Riley, R. F. (1962) J . Bacteriol.
7. Kaplan, M. H., and Volanakis, J. (1974) J. Zmmunol. 112,2135-
8. Oliveira, E. B., Gotschlich, E. C., and Liu, T. (1979) J. Biol.
9. Volanakis, J. E., and Kaplan, M. H. (1971) Proe. SOC. Exp. Biol.
Med. 87,120-128
406-418
571
3,41-51
83,1017-1024
2147
Chem. 254,489-502
Med. 136.612-614
10. Bach, B. A., Gewurz, H., and Osmand, A. P. (1977) Immunochem-
11. Oliveira, E. B., Gotschlich, E. C., and Liu, T.-Y. (1980) J. Im-
12. Tanaka, T., and Robey, F. A. (1983) J. Immunol. Methods 65,
13. Robey, F. A., Tanaka, T., and Liu, T.-Y. (1983) J. Biol. Chem.
14. Rizzo, W. B., and Bustin, M. (1977) J. Biol. Chem. 252, 7062-
15. Simpson, R. T. (1978) Biochemistry 17, 5524-5531
17. Hong, S. L., and Deykin, D. (1981) J. Bwl. Chem. 256, 5215- 16. Deleted in proof
18. Hong, S. L., and Deykin, D. (1982) J. Biol. Chem. 257, 7151-
19. Stambaugh, R., and Post, D. (1966) J. BioL Chem. 241, 1462-
20. Krueger, R. C. (1983) Biochem. Biophys. Res. Commun 110,216-
21. LeStourgeon, W. M., and Rusch, H. P. (1973) Arch. Biochem.
22. Robey, F. A., and Liu, T.-Y. (1981) J. Biol. Chem. 256,969-975 23. Liu, T.-Y., Robey, F. A., and Wang, C.-M. (1982) Ann. N. Y.
24. Robey, F. A., and Liu, T.-Y. (1983) J. Biol. Chem. 258, 3895-
25. Volanakis, J. E. (1982) J . Immunol. 128,2745-2750 26. Chelladurai, M., Macintyre, S. S., and Kushner, I. (1982) Ann.
27. Dougherty, T., Potempa, L. A. Gewurz, H., Siegel, J. (1982) Ann.
istry 1396-1402
munol. 124 , 1396-1402
333-341
258,3889-3894
7067
5219
7154
1467
219
Biophys. 155, 144-158
Acad. Sci. 389, 151-162
3900
N. Y. Acad. Sci. 389,437-438
N. Y. Acad. Sci. 389,444-445