binding of intermediate density lipoproteins rich or poor in high molecular weight apolipoprotein b...
TRANSCRIPT

Binding of intermediate density lipoproteins rich or poor in high molecular weight apolipoprotein B to rat liver membranes
LOUISE BRISSETTE' AND SIMON-PIERRE NOEL' De'partement de biochimie, Universite' de Montrial, Montrial (Que'bec), Canada H3C 357
Received February 16, 1988
BRISSETTE, L., and NOEL, S.-P. 1988. Binding of intermediate density lipoproteins rich or poor in high molecular weight apolipoprotein B to rat liver membranes. Biochem. Cell Biol. 66: 1238-1247.
Very low density lipoproteins rich or poor in high molecular weight apolipoprotein B (Bh-rich or Bh-poor VLDL, respectively) were prepared from rats fasted for 2 days and animals fasted and then refed for 2 days, respectively. Bh-rich or Bh-pOOr VLDL remnants (IDL) were also prepared by in vitro lipolysis of the corresponding VLDL preparations, and their apolipoprotein (apo) profile and lipid composition determined. Bh-rich IDL are richer in esterified cholesterol than Bh-poor IDL, but poorer in apoC and triglycerides. The binding of 125~-labeled Bh-rich IDL and '25~-labeled Bh-poor IDL to rat liver membranes was assessed by saturation-curve studies. Both types of IDL bound to high- and low-affinity sites on rat liver membranes. There were no significant differences between the binding of IDL produced from Bh-rich or Bh-poor VLDL to either the high- or low-affinity sites. However, by masking the low-affinity binding sites with saturating amounts of human high density lipoproteins 3 (HDL3), we were able to demonstrate that Bh-rich IDL bound to high-affinity binding sites with five times less affinity than Bh-poor IDL. These results show that saturating the low-affinity binding sites of rat liver membranes reveals differences in the binding abilities of lipoproteins to the high-affinity sites. Also, an analysis of apo and lipid compositions of the two types of IDL reveals that the apoBh contribution is likely to be responsible for differences in affinities of IDL for the high-affinity binding sites of rat liver membranes.
BRISSETTE, L., et NOEL, S.-P. 1988. Binding of intermediate density lipoproteins rich or poor in high molecular weight apolipoprotein B to rat liver membranes. Biochem. Cell Biol. 66 : 1238-1247.
Nous avons prepart des 1ipoprotCines de trks faible densit6 (VLDL) riches ou pauvres en apolipoprottine B de haut poids moltculaire (ap0Bh) il partu de plasma de rats mis h jeQn pour une pCriode de 2 jours dans le premier cas ou rnis h jecn et renoums pour des pCriodes de 2 jours dans le second cas. Par lipolyse in vitro, nous avons produit des rCsidus de VLDL (IDL) h partir de ces deux types de preparations et CtudiC leurs compositions prottiques et lipidiques. Nous avons observC que les IDL riches en apoBh contiennent plus de cholestCro1 estCrifiC et moins d'apoC et de triacylglycCrols que les IDL pauvres en apoBh. La liaison de ces lipoprottines aux membranes hCpatiques de rat a CtC CtudiCe par des essais de courbes de saturation. Nous avons 0 b S e ~ C que les IDL riches ou pauvres en apoBh se lient h des sites de liaison de haute et de faible affinitt. Selon ces conditions de liaison, aucune difftrence significative entre la liaison des deux types de IDL n'a pu &tre dtcelke. Toutefois, en saturant les sites de liaison de faible affinitt avec des lipoprottines 3 de trks forte densitt (HDL3), nous avons pu dtmontrer que les IDL riches en ap0Bh se lient aux sites de haute affinitk avec cinq fois moins d'affinitC que les IDL pauvres en apoBh. Ces r6sultats demontrent qu'il est avantageux de saturer les sites de liaison de faible affinitC des membranes de foie de rat pour percevoir des diffkrences au niveau des paramktres de liaison de diverses prCparations de IipoprotCines aux sites de haute affinith. De plus, I'analyse approfondie des compositions protkiques et lipidiques a rCvC1C que la composition en apoBh des deux types de IDL semble &tre responsable des affinitCs observ6es pour la liaison des IDL aux sites de haute affinitt des membranes de foie de rat.
Introduction In the fasted state, serum VLDL is mainly secreted by
the liver (Ockner and Isselbacher 1974). Within the circulation, part of the VLDL triglyceride load is hydrolyzed by lipoprotein lipase (Eisenberg et al. 1973). This process leads to the formation of VLDL remnants, also referred to as IDL. In rats, most IDL are
ABBREVIATIONS: VLDL, very low density lipoprotein; IDL, intermediate density lipoprotein; LDL, low density lipo- protein; apoB, apoE, apoC, apolipoproteins B, E, and C, respectively; apoBh, high molecular weight apoB; apoBl, low molecular weight apoB; HDL3, high density lipoprotein 3.
' ~ u t h o r to whom correspondence should be sent to the following address: Cancer Research Laboratories, Queen's University, Kingston, Ont., Canada K7L 3N6.
'~eceased February 5, 1987.
rapidly taken up by the liver (Eisenberg and Rachmile- witz 1973; Mjos eta l . 1975) and a very small proportion is converted to LDL. Windler et al. (1980) have shown that IDL bind to high- and low-affinity binding sites on rat liver membranes. We have demonstrated (Brissette and Noel 1986; Brissette et al . 1986) that both the LDL and remnant receptors are responsible for the high- affinity binding, whereas currently uncharacterized lipoprotein-binding sites are responsible for the low- affinity binding.
In contrast with human VLDL, which contain only the high molecular weight form of apoB (apoB- loo), rat VLDL contain both the high (Bh) and low (B,) molecular weight apoB species (Wu and Windmueller 1981; Elovson et al. 1981). Although the majority of apoBl and apoBh are removed by the liver at the IDL stage (Mjos et al. 1975), Elovson et al. (1981), Wu and
Bio
chem
. Cel
l Bio
l. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
OF
NO
RT
H C
AR
OL
INA
AT
on
11/1
3/14
For
pers
onal
use
onl
y.

BRISSETTE
Windmueller ( l981) , and Sparks and Marsh (1981) have shown that in the rat, apoBh disappears slower from circulation than apoB1. In addition, the results of Sparks et al. (1981) suggest that the VLDL catabolism cascade is different for the particles containing either apoBh or apoB1, since only the high molecular weight form of apoB remains associated with LDL. It was therefore suggested that apoBh impedes the uptake of lipoproteins (Brown and Goldstein 1983).
The aim of this work was to study the effect of the IDL composition in apoB on the binding of these lipoproteins to rat liver membranes. VLDL rich or poor in apoBh were prepared according to the method of Krishnaiah et al. (1980) and their correspondent Bh-rich or Bh-poor IDL produced by in vitro lipolysis. The binding abilities of these two types of IDL to receptors on rat liver membranes were determined and w e found that Bh-rich IDL bound five times less readily than B,-poor IDL to the high-affinity binding sites of rat liver membranes.
Materials and methods Materials
Carrier-free sodium ['25~] iodide was purchased from Amer- sham Canada Ltd. (Oakville, Ont.) and [9, 10-3H(N)]triolein was bought from New England Nuclear (Boston, MA). Fetal bovine serum was obtained from Gibco Laboratories (Burling- ton, Ont.). Fatty acid-free bovine serum albumin, fraction V , phenylmethylsulfonyl fluoride, and heparin (sodium salt, grade 1) were all purchased from Sigma Chemical Co. (St. Louis, MO). Human blood, 3- to 5-days old, was acquired from the MontrCal Blood Transfusion Centre of the Canadian Red Cross.
Animals Male Sprague-Dawley rats were obtained from Charles
River CanadaInc. (St. Constant, Que.). They were maintained in a 12-h light-darkness cycle and fed Rat Chow (Ralston Purina Co., St. Louis, MO) ad libitum unless otherwise stated. Collection of blood for VLDL preparation and removal of livers for membrane isolation were camed out under diethyl ether anesthesia. For the preparation of postheparin plasma, animals were anesthetized with 60 mglkg body weight of sodium pentobarbital (Centre de distribution de MCdicaments VCtCrinaires, St. Hyacinthe, Que.).
Preparation of lipoproteins All lipoproteins were isolated at 16OC according to the
method described by Hatch and Lees (1968). Phenylmethyl- sulfonyl fluoride (10 pM), EDTA (0.3 mM), and sodium azide (3 rnM) were added to all sera before isolation of lipoproteins. Sodium bromide was used to adjust the density of sera and solutions unless otherwise stated.
All VLDL preparations were isolated from the serum of 300- to 400-g adult male rats. Normal VLDL were prepared from rats fed ad libitum. As outlined in the method of Krishnaiah et al. (1980), Bh-rich VLDL were prepared from rats fasted for 2 days, while Bh-poor VLDL were isolated from rats fasted for 2 days and refed for 2 days. Chylomicrons were removed from normal and Bh-poor VLDL serum prepara-
tions by ultracentrifugation for 30 rnin at 85 000 x g. VLDL were isolated by ultracentrifugation at 100 000 X g for 18 h .
Rat IDL were produced by incubation of VLDL with rat postheparin plasma containing lipoprotein lipase. To minimize possible contamination with normal VLDL, we incubated B,-rich and Bh-poor VLDL with postheparin plasma prepared horn rats [hat had folfowed the fasting-feeding conditions used for their isolation. Each VLDL preparation was incu- bated at 37OC for 50 min with lipase (80 mU/mg VLDL triglycerides), in the presence of 2.5% fatty acid-free bovine serum albumin, as previously described (Brissette and Noel 1984). In this work, 1 m u was defined as the amount of postheparin plasma that produced 1 nmol of fatty acid per minute at 37OC, pH 8.0. The enzymatic activity was measured by using an emulsion of tritiated triolein in glycerol (Nilsson- Ehle and Schotz 1976). Under these lipolytic conditions, 60-70% of VLDL triglycerides were hydrolyzed. Following incubation, the medium was placed on ice and solid sodium chloride was added to raise the density to 1.019 g/mL. IDL were then isolated by ultracentrifugation at 100 000 X g for 20 h at 4°C and washed under the same conditions. IDL were iodinated by a modification (Langer et al. 1972) of the iodine monochloride method of McFarlane (1958), as previously described (Brissette and Noel 1986).
Human HDL3 (density: 1.125-1.21 g/rnL) were prepared as previously described (Brissette and N&l 1986).
Liver membrane-binding assay Liver membranes were prepared from fasted rats weighing
200-300 g, according to the method described by Kovanen et al. (1979).
The binding assay was carried out essentially as described previously (Brissette and N&l 1986). 1251-labeled IDL and membranes were incubated for 60 min at O°C in 80 mM NaCI, 1.0 rnM CaC12, 50 mM Tris-HC1 (pH 7.5), and 20 m g / d bovine serum albumin. Each tube contained 100 pg of membrane protein and the indicated amount of rat 125~-labeled IDL with or without a 200-fold excess of HDL3, in a total volume of 120 p L Nonspecific binding was determined by the addition of a 200-fold excess of unlabeled rat IDL in the incubation medium. Following incubation, a 60-pL aliquot from each assay tube was layered onto 125 p L of fetal bovine serum in a silanized tube (Prosil28, PCR Research Chemicals Inc.) and the tubes were centrifuged at 100 000 x g for 20 min at room temperature in a Beckman Airfuge (rotor A-1001 18). The supernatant was then removed and each pellet was overlaid with 175 pL of fetal bovine serum and recentrifuged for 10 min at 100 000 x g. The supernatant was removed, each tube was sliced with a razor blade, and the bottom of the tube containing the pellet was counted in an LKB gamma counter.
Analysis of binding data Specific binding was calculated by subtracting the binding
of rat lZ51-labeled IDL obtained in the presence of a 200-fold excess of unlabeled IDL (nonspecific binding) from that obtained in the absence of labeled IDL (total binding). According to a modification of the method of Scatchard (1949), the curves generated by the specific binding data were transformed into plots of the ratio membrane-boundlfree lipoprotein versus membrane-bound lipoprotein. Concave- upward curvilinear Scatchard plots were resolved by the
Bio
chem
. Cel
l Bio
l. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
OF
NO
RT
H C
AR
OL
INA
AT
on
11/1
3/14
For
pers
onal
use
onl
y.

1240 BIOCHEM. CELL BIOL. VOL. 66, 1988
graphical method of Rosenthal (1967). The dissociation constants ( K d ) were calculated from the slopes of the resulting linear components. The maximum binding (B,,,) was ob- tained from the intercept of each linear component on the x axis.
Other analyses Protein mass was determined by the method of Lowry et al.
(1951) as adapted by Kashyap et al. (1980) for solutions containing lipoproteins, using bovine serum albumin as a standard. Total lipids were extracted according to the method of Folch etal. (1957) and separated by thin-layerchromatogra- phy on silica gel G (Baker Chemical Co., Phillipsburg, NJ). The migrating solvent contained petroleum ether, diethyl ether, and glacial acetic acid in the ratio 90: 10: 1 (vlv). Phospholipids were eluted with a mixture of chloroform, methanol, and water (75:25:4, vlv), triglycerides with solution of hexane and isopropanol(7:4, vlv), and free and esterified cholesterol with chloroform. The phospholipids, triglycerides, free and esteri- tied cholesterol were recovered from the silica gel plates with an average efficiency of 80, 90, 81, and 87%, respectively. The recovery of all lipids was determined each time and the raw lipid values were corrected to 100%. The phosphate released from phospholipid by perchloric acid digestion was determined by the method of Fiske and SubbaRow (1926) and converted to phospholipids as distearyl phosphatidylcholine. Triglycerides and cholesterol, respectively, were determined by the method of Van Handel (1961) and Franey and Amador (1968).
For polyacrylamide gel electrophoresis, VLDL solutions were dialyzed against water and lyophilized prior to delipida- tion with ethanol and ether (3: 1, vlv) as described by Scanu and Edelstein (1 97 1). The apolipoproteins were separated by slab-gel electrophoresis, essentially as described elsewhere (Brissette and Noel 1986). Apolipoprotein profiles were scanned by a laser densitometer (Ultroscan, LKB) and the relative contribution of each band was calculated using the GelScan Apple IIe software.
Results Rat VLDL rich or poor in apoBh were prepared
according to the nutritional conditions described by Krishnaiah et al. (1980). The polyacrylamide gel electrophoretic patterns of these VLDL preparations were compared with a normal VLDL preparation (ad libitum fed rat). Figure 1 shows that each VLDL preparation contains apoBh, apoB,, apoE, and apoc and that the proportion of apoBh and apoB, differs in each preparation. Several preparations of Bh-rich or Bh-poor VLDL were scanned by densitometry and the relative contribution of each protein band was calculat- ed. Apolipoprotein ratios were also calculated from these data (Table 1). The analysis of these results shows that the contribution of each apolipoprotein differs significantly between the two types of VLDL, and that apoBh/apoBI, apoE/apoB, and apoC/apoB ratios also differ significantly. On a total apoB basis, it appears that Bh-rich VLDL is poorer in apoc and apoE than Bh-poor VLDL. Since apoE1apoC ratios are not significantly dif-
FIG. 1. Polyacrylamide gel electrophoretic patterns of rat normal VLDL, Bh-poor and Bh-rich VLDL. A 50-kg sample of protein was applied on a 4-18% polyacrylamide gradient gel in the presence of sodium dodecyl sulfate and P-mercapto- ethanol. (A) Normal VLDL, (B) Bh-pOOr VLDL, (C) Bh-rich VLDL.
ferent between the two VLDL types, it is apparent that both apolipoproteins are decreased by comparable amounts in the Bh-rich particles. An analysis of the lipid composition of the VLDL preparations shows that the proportional surface lipid content (phospholipids and free cholesterol) of the two types of VLDL are similar, while the core lipid content (triglycerides and esterified cholesterol) is different (Table 2). Indeed, we observe that Bh-rich VLDL contains more esterified cholesterol and less triglycerides than Bh-poor VLDL, which could be due to a smaller intrahepatic triglyceride pool in fasted than in refed rats.
We have also prepared IDL by in vitro incubation of VLDL with postheparin plasma containing lipoprotein lipase. To minimize the risk of contamination with normal VLDL, we avoided the customary use of overnight fasted rats to prepare postheparin plasma. Instead, Bh-rich and Bh-poor VLDL were incubated with postheparin plasma prepared from rats that had followed the same fasting-feeding conditions. The enzymatic activities were determined to be 215 * 14 mU/rnL in fasted rats and 272 + 27 mU/mL (mean + SEM, N = 4) inrefed rats. The lower enzymatic activity in fasted rats may be due to a decline of lipoprotein lipase activity in adipose tissue, as shown previously by Fried et al. (1983). These postheparin plasma prepara- tions were used in a ratio of 80 mUlmg triglycerides to hydrolyze their corresponding VLDL preparations. In this fashion, we were able to achieve the same extent of
Bio
chem
. Cel
l Bio
l. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
OF
NO
RT
H C
AR
OL
INA
AT
on
11/1
3/14
For
pers
onal
use
onl
y.

TABLE 1. Apolipoprotein composition of rat Bh-rich or Bh-pOOr VLDL and IDL
Apolipoprotein content (%)" -
Apolipoproteins Bh-rich VLDL Bh-poor VLDL Bh-rich IDL Bh-poor IDL
Bh 36.224.0 6.920.67~ 34.62 1.8 10.6k1.2~ BI 11.5k1.5 18.7k1.02~ 17.624.2 18.4k3.3' E 24.224.1 ~ 7 2 3 . 7 ~ 31.123.3 30.0+3.6' C 22.123.9 36.62 10.7~ 10.5k2.1 34.6k2.7b
B h / B ~ 3.920.78 0.3920.05~ 2.3-tO.75 0.62k0.1 lb E/B 0.56kO. 12 1.4620.14~ 0.6020.06 1.20k0.35' C/B 0.7920. 18 1.7320.24~ 0.21r0.06 1.2020.11b E/C 1.4820.52 1.01+0.19' 3.20-tO.75 0.90kO. 1 8 ~
N 11 15 3 4
NOTE: Each value represents the mean 5 SEM of the indicated number of experiments (N). "The apolipoproteins of B,-rich and B,-poor VLDL and IDL were separated by a 4-18% polyacrylamide
gel electmphoresis in the presence of sodium dodecyl sulfate and p-mercaptoethanol. Each electrophoretic pattern was scanned by a densitometer (Ultroscan. LKB), and the relative contribution of each band was calculated by the Gelscan Software Apple Ile.
bStatistically different fmm Bh-rich lipoprotein counterpart (p < 0.0125). 'Not statistically significant.
TABLE 2. Lipid composition of rat &-rich or Bh-poor VLDL and IDL
Lipid content (%)"
Lipids Bh-rich VLDL Bh-poor VLDL Bh-rich IDL Bh-poor IDL
NOTE: CL, free cholesterol; CE, esterified cholesterol; TG, uiglycerides; PL, phospholipids. Each value represents the mean f SEM of the number of experiments (N).
"The lipid content was calculated as the relative content of the total lipid mass. bStatistically different from Bh-rich lipoprotein counterpart ( p < 0.01). 'Not statistically significant.
hydrolysis for the two IDL preparations (60-70%). The analysis of the lipid composition (Table 2) shows that the lipolysis produced, as expected, a loss of triglycer- ides and a relative enrichment in the other lipids. The analysis also reveals that Bh-rich lipoproteins are still richer in esterified cholesterol and poorer in triglycer- ides than Bh-poor lipoproteins. Analysis of the apolipo- protein electrophoretic patterns (Table 1) of these IDL preparations, compared with their related VLDL prepa- rations, shows that the expected loss of apoC usually occurring during lipolysis (Eisenberg and Levy 1976) was not observed for IDL derived from Bh-poor VLDL. It is possible that HDL, which are known to be apoC acceptors (Patsch et al. 1978), were not as potent in the latter preparation of postheparin plasma obtained from refed rats, since they might have been saturated with apoC derived from the catabolism of chylomicrons.
Analysis of apolipoprotein ratios of the particks reveals that apoBh/apoBI and apoC/apoB ratios remain signi- ficantly higher and lower, respectively, for &-rich than for Bh-poor lipoproteins. These ratios also reveal that lipolysis creates a statistically significant difference between the apoElapoC ratios of the two types of IDL (Bh-rich IDL having the highest ratio). Also, it appears that the apoE/apoB ratios, as in the VLDL preparations, are lower for Bh-rich than for Bh-poor IDL. However, it appears that this twofold difference is not significantly different (just below the 95% P value) because of the high variability of the results.
The binding of these two types of IDL to rat liver membranes was assessed by a saturation-curve analysis. Figure 2 shows the binding of rat lZ5I-labeled Bh-poor IDL to hepatic membranes. The saturation curve ap- pears in panel A and the Scatchard plot of the specific
Bio
chem
. Cel
l Bio
l. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
OF
NO
RT
H C
AR
OL
INA
AT
on
11/1
3/14
For
pers
onal
use
onl
y.

BIOCHEM. CELL BIOL. VOL. 66, 1988
'2'~-lobeled Bh- poor IDL (yg proteins/mL)
'"1- lobeled Bh- poor IDL bound ( yg proteins/mg membrone proteins)
FIG. 2. The binding of rat '25~-labeled Bh-poor IDL to rat liver membranes. (A) Saturation curve of the binding of rat 125~-labeled Bh-poor IDL to rat liver membranes. Each tube contained 100 k g of membrane proteins and the indicated concentration of '25~-labeled Bh-poor IDL. The specific binding (0) was calculated by subtracting the binding of 125~-labeled Bh-pOOr IDL obtained in the presence of a 200-fold excess of unlabeled Bh-poor IDL (0) from the binding obtained in the absence of unlabeled IDL (0). (B) Scatchard plot of the specific binding data. The boundlfree ratio represents the amount of specifically bound lipoprotein (p,g proteinlmg membrane protein) divided by the concentration of unbound lipoprotein in the reaction mixture (kg protein/mL). Each point represents the mean of duplicate values. These results are typical of four such experiments that gave essentially the same results.
binding data is shown in panel B . The curvilinearity of the Scatchard plot shows that I2'I-labeled Bh-poer IDL bind to multiple binding sites, which can be resolved by the graphical method of Roseatha1 into two binding cornpo- nents that are identified as high- and tow-affinity binding sires. Similar results were obtained with the binding of 12'1-labeled Bh-rich IDL (Fig. 3). An analysis of the binding parameters of each type of IDL to each binding site (Table 3) shows the absence of significant differences between the binding of these IDL to either the high- or the low-affinity binding sites.
The binding of I2%labeled Bh-rich IDL and '251- labeled Bh-poor IDL to the high-affinity sites was also investigated in the presence of a 200-fold excess of HDL3, since previous work (Brissette and Noel 1986) had shown that the addition of such a quantity of HDL3 was able to completely abolish the binding of IDL to the low-affinity sites. Typical results are shown in Figs. 4
and 5. Linear Scatchard plots are obtained, showing that in the presence of HDL3, both IDL preparations interact only with a single binding site or with binding sites of undistinguished affinities. A comparison of the binding parameters deduced from these linear Scatchard plots (Table 4) with the binding parameters calculated from the Rosenthal decomposition of regular saturation-curve studies (Table 3) reveals that under these conditions, Bh-poor and Bh-rich IDL bind only to the high- affinity binding sites, as expected. The analysis of the binding parameters obtained with Bh-rich and Bh-poor IDL also demonstrates that the affinity of the Bh-poor IDL binding to the high-affinity binding sites is higher (fivefold) than the Bh-rich IDL binding.
To verify that the two types of IDL preparations bind to the same high-affinity bindin sites, we studied the B competition of the binding of l2 I-labeled Bh-poor IDL with Bh-poor or Bh-rich IDL in the presence of HDL3.
Bio
chem
. Cel
l Bio
l. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
OF
NO
RT
H C
AR
OL
INA
AT
on
11/1
3/14
For
pers
onal
use
onl
y.

BRISSETTE AND NOEL
''=I - labeled 8,-rich IDL (yg prateins/mLl
Iz51 - labeled Bh- rich IDL bound OJg proteins/mg membrane proteins)
FIG. 3. The binding of rat '251-labeled Bh-rich IDL in rat liver membranes. (A) Saturation curve of the binding of rat lZ5I-labeled Bh-rich IDL to rat liver membranes. Each tube contained 100 kg of membrane proteins and the indicated concentration of lZ51-labeled Bh-rich IDL. The specific binding (0) was calculated by subtracting the binding of 'Z5~-labeled Bh-rich IDL obtained in the presence of a 200-fold excess of unlabeled Bh-rich IDL (0) from the binding obtained in the absence of unlabeled IDL (e). (B) Scatchard plot of the specific binding data. The boundlfree ratio represents the amount of specifically bound lipoprotein (pg proteinlmg membrane protein) divided by the concentration of unbound lipoprotein in the reaction mixture (kg protein/mL). Each point represents the mean of duplicate values. These results are typical of six such experiments that gave essentially the same results.
TABLE 3. Binding parameters of rat Bh-rich or Bh-poor IDL to rat liver membranes
High-affinity component Low-affinity component
Kd, B m , Kd, Bm,, (pg protein/mL) (pg proteinling protein membrane) (pg proteinlml) (pg proteinlmg protein membrane)
Bh-rich IDL (4) 0.45k0.023 0.038+0.005 68.3229.9 3.9k2.1 Bh-p00rIDL(4) 0.38k0.024 0.025+0.005 100.0a36.6 8.224.1
P NS NS NS NS
NOTE: The binding assays were carried out as described under Materials and methods. Kd and B , were determined from the Scatchard plots of the specific binding data using the graphical method of Rosenthal. Each value represents the mean + SEM of the indicated number of experiments in parentheses. The Student's I-test was performed on the two means of each column. NS, not statistically significant.
The result shown in Fig. 6 reveals that Bh-rich IDL slower disappearance of apoBh from the circulation (Wu can compete with the binding of Bh-poor IDL to the and Windmueller 1981; Sparks and Marsh 1981) may be high-affinity sites. due to slower kinetics of lipolysis of apOBh-cOnstituted
VLDL and (or) to a less efficient uptake by the liver of Discussion apoBh-constituted IDL compared with their apoBl-con-
Rat VLDL consist of a mixture of particles that con- stituted lipoproteins counterparts. In this manuscript, tain either high or low molecular weight apoB (Wu and we have investigated the latter possibility. Windmueller 198 1 ; Elovson et al. 198 1). The relatively Bh-rich and Bh-poor VLDL were prepared accord-
Bio
chem
. Cel
l Bio
l. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
OF
NO
RT
H C
AR
OL
INA
AT
on
11/1
3/14
For
pers
onal
use
onl
y.

1244 BIOCHEM. CELL BIOL. VOL. 66, 1988
12'1- labeled Bh- poor IDL (pg proteins/mL)
l e 5 1 - lobeled Bh- poor I D L bound (pg proteins/mg membrane proteins)
FIG. 4. The binding of rat 12s1-labeled &-poor IDL to rat liver membranes in the presence of human HDL3. (A) Saturation c w e of the binding of rat ' ' f~ l&eled Bh-por [DL to rat liver membranes. Each tube contained 100 pg of membrane proteins, the indicated concentration of '25~-labeled &poor IDL, and a 200-fold excess of human HDL3 protein relative to the amount of rat ''5~-labeled Bh-poor 113L protein in each away tube. The specific binding (0) was calculated by subtracting the binding of 1251-labeled Bh-poor 1DL obtained in the presence of a 200-fold excess of unlabeled Bh-poor IDL (0) from the binding obtained in the absence of unlabeled IDL (a). (B) Scatchard plot of the specific binding data. The boundlfree ratio represents the amount of specifically bound lipoprotein (pg proteinlmg membrane protein) divided by the concentration of unbound lipoprotein in the reaction mixture (kg proteinlml). Each point represents the mean of duplicate values. These results are typical of nine such experiments that gave essentially the same results.
'25~-labeled Bh-rich IDL (yg proteins/mL
'"I -labeled Bh-rich IDL bound ( J I ~ proleins/mg membrane proteins1
FIG. 5. The binding of rat 125~-labeled Bh-rich IDL to rat liver membranes in the presence of human HDL3. (A) Saturation curve of the binding of rat lZ51-labeled &,-rich IDL to rat liver membranes. Each tube contained 100 pg of membranes proteins, the indicated concentration of 'Z5~-labeled Bh-rich IDL, and a 200-fold excess of human HDL3 protein relative to the amount of rat lZ51-labeled Bh-rich IDL protein in each assay tube. The specific binding (0) was calculated by subtracting the binding of 'Z51-labeled Bh-rich IDL obtained in the presence of a 200-fold excess of unlabeled Bh-rich IDL (0) from the binding obtained in the absence of unlabeled IDL (@). (B) Scatchard plot of the specific binding data. The boundlfree ratio represents the amount of specifically bound lipoprotein (kg proteinlmg membrane protein) divided by the concentration of unbound lipoprotein in the reaction mixture (pg protein/mL). Each point represents the mean of duplicate values. These results are typical of six such experiments that gave essentially the same results.
Bio
chem
. Cel
l Bio
l. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
OF
NO
RT
H C
AR
OL
INA
AT
on
11/1
3/14
For
pers
onal
use
onl
y.
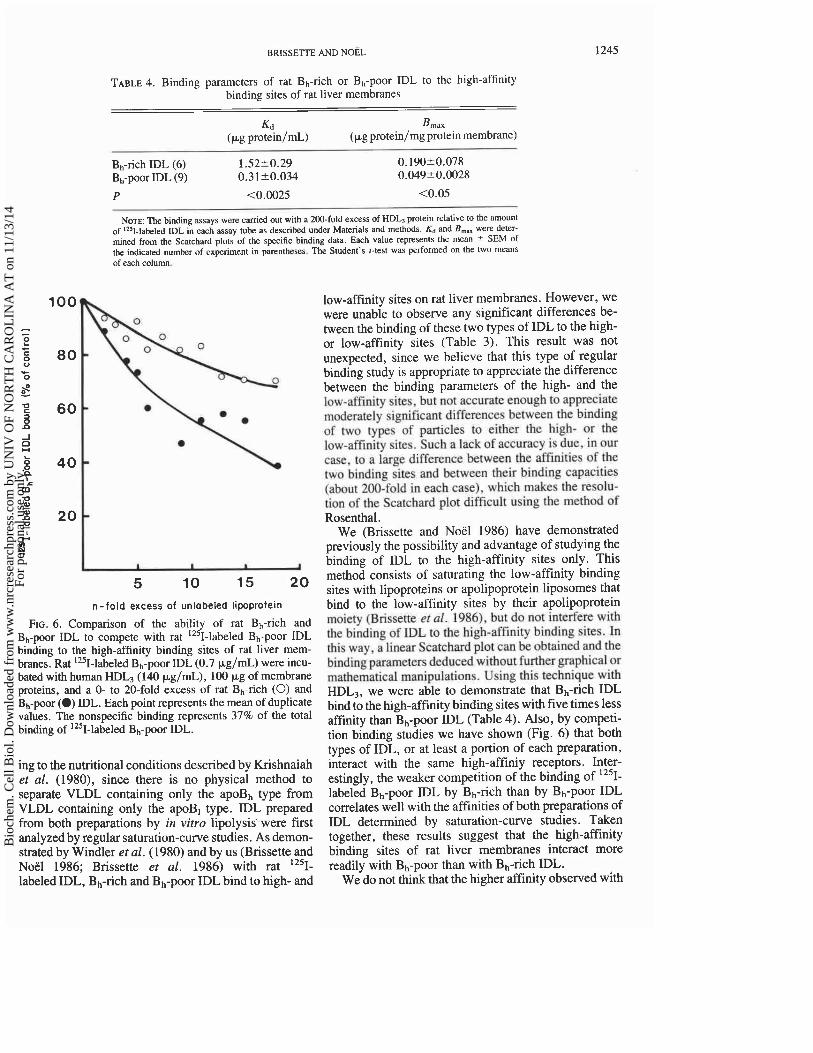
TABLE 4. Binding parameters of rat Bh-rich or Bh-pOOr IDL to the high-affinity binding sites of rat liver membranes
Kd Bmax (kg protein/rnL) (kg protein/mg protein membrane)
Bh-rich IDL (6) 1.5220.29 0.190?0.078 Bh-pOOr IDL (9) 0.31+0.034 0.04920.0028 P <0.0025 cO.05
-
NOTE: The binding assays were carried out with a 200-fold excess of HDL, protein relative to the amounl of 12s1-labeled IDL in each assay tube as described under Materials and methods. K, and B,,. were deter- mined from the Scatchard plots of the specific binding data. Each value represents the mean + SEM of the indicated number of experiment in parentheses. The Student's I-test was performed on the two means of each column.
n-fold excess of unlabeled lipoprotein
FIG. 6. Comparison of the ability of rat Bh-rich and Bh-pOOr IDL to compete with rat '25~-labeled Bh-poor IDL binding to the high-affinity binding sites of rat liver mem- branes. Rat '25~-labeled Bh-poor IDL (0.7 pg/mL) were incu- bated with human HDL, (140 pg/mL), 100 pg of membrane proteins, and a 0- to 20-fold excess of rat Bh-rich (0) and Bh-poor (0) IDL. Each point represents the mean of duplicate values. The nonspecific binding represents 37% of the total binding of '25~-labeled Bh-poor IDL.
ing to the nutritional conditions described by Krishnaiah et al. (1980), since there is no physical method to separate VLDL containing only the apoBh type from VLDL containing only the apoBl type. IDL prepared from both preparations by in vitro lipolysis were first analyzed by regular saturation-curve studies. As demon- strated by Windler et al. (1980) and by us (Brissette and Noel 1986; Brissette et al. 1986) with rat 12'1- labeled IDL, Bh-rich and Bh-poor IDL bind to high- and
low-affinity sites on rat liver membranes. However, we were unable to observe any significant differences be- tween the binding of these two types of IDL to the high- or low-affinity sites (Table 3). This result was not unexpected, since we believe that this type of regular binding study is appropriate to appreciate the difference between the binding parameters of the high- and the low-affinity sites, but not accurate enough to appreciate moderately significant differences between the binding of two types of particles to either the high- or the low-affinity sites. Such a lack af accuracy is due, in our case. to a large difference between the affinities of the two binding sites and between their binding capacities (about 200-fold in each case), which makes the resolu- tion of the Scatchard plot difficult using the method of Rosenthal.
We (Brissette and Noel 1986) have demonstrated previously the possibility and advantage of studying the binding of IDL to the high-affinity sites only. This method consists of saturating the low-affinity binding sites with lipoproteins or apolipoprotein liposomes that bind to the low-affinity sites by their apolipoprotein moiety (Brissette et al. 19861, but do not interfe~ with the binding of IDL to the high-affinity binding sites. In this way, a linear Scatchard plot can be obtained and the binding parameters deduced without further graphical or mathematical manipulations. Using this technique with HDL3, we were able to demonstrate that Bh-rich IDL bind to the high-affinity binding sites with five times less affinity than Bh-poor IDL (Table 4). Also, by competi- tion binding studies we have shown (Fig. 6) that both types of IDL, or at least a portion of each preparation, interact with the same high-affiniy receptors. Inter- estingly, the weaker competition of the binding of 125~-
labeled Bh-pOOr IDL by Bh-rich than by Bh-pOOr IDL correlates well with the affinities of both preparations of IDL determined by saturation-curve studies. Taken together, these results suggest that the high-affinity binding sites of rat liver membranes interact more readily with Bh-poor than with Bh-rich IDL.
We do not think that the higher affinity observed with
Bio
chem
. Cel
l Bio
l. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
OF
NO
RT
H C
AR
OL
INA
AT
on
11/1
3/14
For
pers
onal
use
onl
y.

1246 BIOCHEM. CELL BIOL. VOL. 66. 1988
Bh-poor IDL is due to its higher apoC/apoB or its lower apoE/apoC ratios, since in each case, this would imply that a higher apoC content (Table 4) favors binding. Such a behaviour would be contrary to the many studies suggesting that apoC (particularly apoC-111) is more likely to interfere with binding (Shelburne et al. 1980; Windler and Have1 1985). If this is in fact the case, it is possible that the higher apoC content of Bh-poor IDL used in this study may have resulted in an underestima- tion of the affinity of Bh-poor IDL for the high-affinity binding sites. Also, we cannot rule out the possibility that differences in affinity could reflect the lipid compo- sition of the IDL preparations, since Bh-poor IDL is richer in triglycerides than Bh-rich IDL. A recent study has shown that the immunoreactivity of an apoB epitope involved in the binding of LDL to human LDL receptor (Y. Marcel, personal communication) increases with increasing density of lipoprotein subclasses. In light of that, it appears that if the triglyceride composition is exerting an effect on the binding of our IDL prepara- tions, it would be by decreasing the ability of Bh-poor IDL to bind to the high-affinity sites. This difference between the two IDL preparations cannot explain the differences observed between the affinities of the two lipoprotein preparations, but could result in an underes- timation of Bh-poor IDL affinity. Thus, we propose that the form of apoB (apoBh/apoB1) present in IDL is responsible for the difference between the affinity of different types of IDL for the high-affinity binding sites of rat liver membranes. We also suggest that apoBh interferes with the binding of IDL containing apoBh to the high-affinity sites of rat hepatic membranes since, the affinity of the particle richer in apoBh is lower than that of the particle poor in apoBh.
We have previously demonstrated (Brissette and Noel 1986) that the LDL (apoB, apoE) receptors and the remnant (apoE) receptors both constitute the high- affinity binding sites. It appears unlikely that it is the binding of IDL rich or poor in apoBh to the LDL receptors that is responsible for the difference in affinity observed by us, since apoBh is known to bind to this receptor (Brown and Goldstein 1983) and there is no evidence suggesting that this apolipoprotein interferes with the binding of apoE to the LDL receptor. Thus, it appears that the difference in affinity is attributable to the binding of IDL to the remnant receptor. Our results agree well with the observation that human lipoproteins containing high molecular weight apoB (apoB-100) and apoE disappear less rapidly from the circulation of familial hypercholesterolernic subjects (absence of func- tional LDL receptors) than lipoproteins containing low molecular weight apoB (apoB-48) and apoE (Goldstein and Brown 1977).
In conclusion, by studying the binding of preparations of IDL rich or poor in apoBh to the high-affinity sites of
rat liver membranes, we have shown that the former type of particles binds with less affinity to these sites than to the latter type. These results could explain, at least in part, why apoBh disappears less rapidly than apoBl from rat blood circulation.
Acknowledgments We wish to thank the Montrkal Transfusion Centre of
the Canadian Red Cross and M. Caisse for supplying us with fresh human blood. We gratefully acknowledge the help of L. Charette in typing this manuscript and of C. Ostiguy for the drawing of the figures. The skillful technical collaboration of T. Buy and R. Dupras was also much appreciated. We are indebted to Dr. R. G. Deeley for his useful suggestions in the preparation of this manuscript. This work was supported by grant MA-8056 from the Canadian Medical Research Coun- cil. L.B. was a recipient of a studentship from the Canadian Medical Research Council.
BRISSETTE, L., and NOEL, S. P. 1984. An in vitro model to study lipolysis of rat hepatic very low density lipoprotein using cardiac lipoprotein lipase. Can. J. Biochem. Cell Biol. 62: 1076-1086.
1986. The effects of low and high density lipoproteins on the binding of rat intermediate density lipoproteins to rat liver membranes. J. Biol. Chem. 261: 6847-6852.
BRISSETTE, L., ROACH, P. D., and NOEL, S. P. 1986. The effects of liposome-reconstituted apolipoproteins on the binding of rat intermediate density lipoproteins to rat liver membrane. J. Biol. Chem. 261: 11 631 - 11 638.
BROWN, M. S., and GOLDSTEIN, J. L. 1983. Lipoprotein receptors in the liver. Control signals for plasma cholesterol traffic. J. Clin. Invest. 72: 743-747.
EISENBERG, S . , and LEVY, R. I. 1976. Lipoprotein metabo- lism. Adv. Lipid Res. 13: 1-89.
EISENBERG, S., and RACHMILEWITZ, D. 1973. Metabolism of rat plasma very low density lipoprotein. Fate in circulation of the whole lipoprotein. Biochim. Biophys. Acta, 326: 378-390.
EISENBERG, S., BILHEIMER, D. W., LEVY, R. Y., and LINDGREN, F. T. 1973. On the metabolic conversion of human plasma very low density lipoprotein to low density lipoprotein. Biochim. Biophys. Acta, 326: 361-377.
ELOVSON, J . , HUANG, Y. O., BAKER, N., and KAUNAN, R. 1981. Apolipoprotein B is structurally and metabolically heterogeneous in the rat. Proc. Natl. Acad. Sci. U.S.A. 78: 157-161.
FISKE, C. H., and SUBBAROW, Y. 1926. The calorimetric determination of phosphorous. J. Biol. Chem. 66: 375- 400.
FOLCH, L., LEES, M., and SLOANE-STANLEY, G. H. 1957. A simple method for the isolation and purification of total lipids from animal tissue. J. Biol. Chem. 226: 497-509.
FRANEY, R. J. , and AMADOR, E. 1968. Serum cholesterol measurement based on ethanol extraction and fenic chloride-sulfuric acid. Clin. Chim. Acta, 21: 255-263.
FRIED, S. K., HILL, J. O., NICKEL, M., ~~~DIGIROLIANO, M.
Bio
chem
. Cel
l Bio
l. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
OF
NO
RT
H C
AR
OL
INA
AT
on
11/1
3/14
For
pers
onal
use
onl
y.

1983. Prolonged effects of fasting-refeeding on rat adipose tissue lipoprotein lipase activity. J. Nutr. 113: 1861- 1869.
GOLDSTEIN, J . L., and BROWN, M. S. 1977. The low density lipoprotein pathway and its relation to atherosclerosis. Annu. Rev. Biochem. 46: 897-930.
HATCH, F. T., and LEES, R. S. 1968. Practical methods for plasma lipoprotein analysis. Adv. Lipid Res. 6: 1-68.
KASHYAP, M. L., HYND, B., and ROBERNSON, K. 1980. A rapid and simple method for measurement of total protein in very low density lipoproteins by Lowry assay. J. Lipid Res. 21: 491-495.
KOVANEN, P. T. , BROWN, M. S., GOL GOLD STEIN, J . L. 1979. Increased binding of low density lipoprotein to liver membrane from rats treated with 17 a-ethinyl estradiol. J. Biol. Chem. 254: 1528-1936.
~RISHNAIAH, K. V., WALKER, L. F., BORENSZTAJN, J . C., SCHONFELD, G., and GETZ, G. S. 1980. Apolipoprotein B variant derived from rat intestine. Proc. Natl. Acad. Sci. U.S.A. 77: 3806-3810.
LANGER, T. W., STROKER, W., and LEVY, R. L. 1972. The metabolism of low density lipoprotein in familial type I1 hyperlipoproteinemia. J. Clin. Invest. 51: 1528-1536.
LOWRY, 0. H., ROSEBROUGH, N. J., FARR, A. L., and RANDALL, R. J. 1951. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 193: 265-275.
MCFARLANE, A. S. 1958. Efficient trace-labeling of proteins with iodine. Nature (London), 182: 53-54.
Uros, 0. D., FAERGEMAN, O., HAMILTON, R. L., and HAVEL, R. J. 1975. Characterization of remnants produced during the metabolism of triglyceride-rich lipoproteins of blood plasma and intestinal lymph in the rat. J. Clin. Invest. 56: 603-615.
NILSSON-EHLE, P., and SCHOTZ, M. C. 1976. A stable, radioactive substrate emulsion for assay of lipoprotein lipase. J. Lipid Res. 17: 536-541.
OCKNER, R. K., and ISSELBACHER, K. J. 1974. Recent concepts of intestinal fat absorption. Rev. Physiol. Bio- chem. Pharmacol. 71: 107- 146.
PATSCH, J. R., GOTTO, A. M., OLIVECRONA, T., and EISENBERG, S. 1978. Formation of high density lipoprotein-
like particles during lipolysis of very low density lipopro- teins in vitro. Proc. Natl. Acad. Sci. U.S.A. 75: 4519- 4523.
ROSENTHAL, H. E. 1967. A graphic method for the determina- tion and presentation of binding parameters in a complex system. Anal. Biochem. 20: 525-532.
SCANU, A. M., and EDELSTEIN, C. 1971. Solubility in aqueous solutions of ethanol of the small molecular weight peptides of the serum very low density and high density lipoproteins. Anal. Biochem. 44: 576-588.
SCATCHARD, G. 1949. The attractions of proteins for small molecules and ions. Ann. N.Y. Acad. Sci. 51: 660-672.
SHELBURNE, F., HANKS, J., MEYERS, M., and QUARFORDT, S. 1980. Effects of apoproteins on hepatic uptake of triglyceride emulsions in the rat. J. Clin. Invest. 65: 652-658.
SPARKS, C. E., ED MARSH, J. B. 1981. Metabolic heterogene- ity of apolipoprotein B in the rat. J. Lipid Res. 22: 519-527.
SPARKS, C. E., HNATUIK, O., and MARSH, J. B. 1981. Hepatic and intestinal contribution of two forms of apolipo- protein B to plasma lipoprotein fractions in the rat. Can. J. Biochem. 59: 693-699.
VAN HANDEL, C. 1961. Suggested modification of the micro-detennination of triglycerides. Clin. Chem. 7: 249- 251.
WINDLER, E., and HAVEL, R. J. 1985. Inhibitory effects of C apolipoproteins from rats and humans on the uptake of triglyceride-rich lipoprotein and their remnants by the perfused rat liver. J . Lipid Res. 26: 556-565.
WINDLER, E. E. T., KOVANEN, P. T., CHAO, Y. S., BROWN, M. S., HAVEL, R. J., and GOLDSTEIN, J. L. 1980. The estradiol-stimulated lipoprotein receptor of rat liver. A binding site that mediates the uptake of rat lipoproteins con- taining apoproteins B end E. J. Biol. Chem. 255: 10 464 - 10 471.
Wu, A., and WINDMUELLER, H. G. 1981. Variant forms of plasma apolipoproteins B . Hepatic and intestinal biosynthe- sis and heterogeneous metabolism in the rat. J. Biol. Chem. 256: 3615-3618.
Bio
chem
. Cel
l Bio
l. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
OF
NO
RT
H C
AR
OL
INA
AT
on
11/1
3/14
For
pers
onal
use
onl
y.