bioenergetics-quest for energy - marine biological … … · · 2013-07-12emp pathway now easier...
TRANSCRIPT

Bioenergetics-Quest for energy • All cellular organisms need energy to grow and
survive. • The energy derived from catabolism of growth
substrates is used to fuel anabolism (biosynthesis) and to fuel other cellular functions such as transport and motility.
• Two catabolic modes – Phototrophy- energy from light – Chemotrophy - energy from chemical reactions
• Chemotrophs catalyze thermodynamically favorable (exergonic) reactions and conserve part of the energy released, either as high energy ester bonds (such as ATP) or as an ion-motive force (usually protons).
• Bacteria and Archaea are incredibly versatile at conserving energy– If there’s a buck to be made some bug will do it.....R. Wolfe (penny?)

Worth looking at
Gottschalk, 1985 2 in library Great chapter on fermentations
White, Drummond, and Fuqua 2012 Not in library - $110.67 at Amazon- 2006 Ed. much cheaper Lots of info – not very user friendly
Thauer, R. K., K. Jungermann, and K. Decker. 1977. Energy conservation in chemotrophic anaerobic bacteria. Bacteriol. Rev. 41:100-180. Out of date, with nothing on chemiosmosis, but still a trove of information on fundamentals of anaerobic metabolism and killer tables at end

Free energy of reactions ∆G°' = ∑∆Gf° products - ∑∆Gf° reactants
-917.2 + 6(0) 6(-586.9) + 6(-39.9) glucose + 6O2 6HCO3- + 6H+
-917.2 -3760.8 ∆G°' = -3760.8 - (-917.2 ) = -2843.6 kJ/rxn (-237 kJ/e- pair)
glucose 2lactate- + 2H+
-917.2 2(-517.8) + 2 (-39.9) = -198.2 kJ/mol
4H2 + HCO3- + H+ CH4 + 3H2O
4(0) + (-586.9) +(-39.9) (-50.8) + 3(-237.2) = -135.6 kJ/rxn (-33.2 kJ/e- pair - but actually less)
Std conditions = 25 oC, 1 M solutes, 1 atm gases, aqueous (H20 ≈ 55 M) For ∆Go’, H+ is constant at 10-7 M (pH 7)
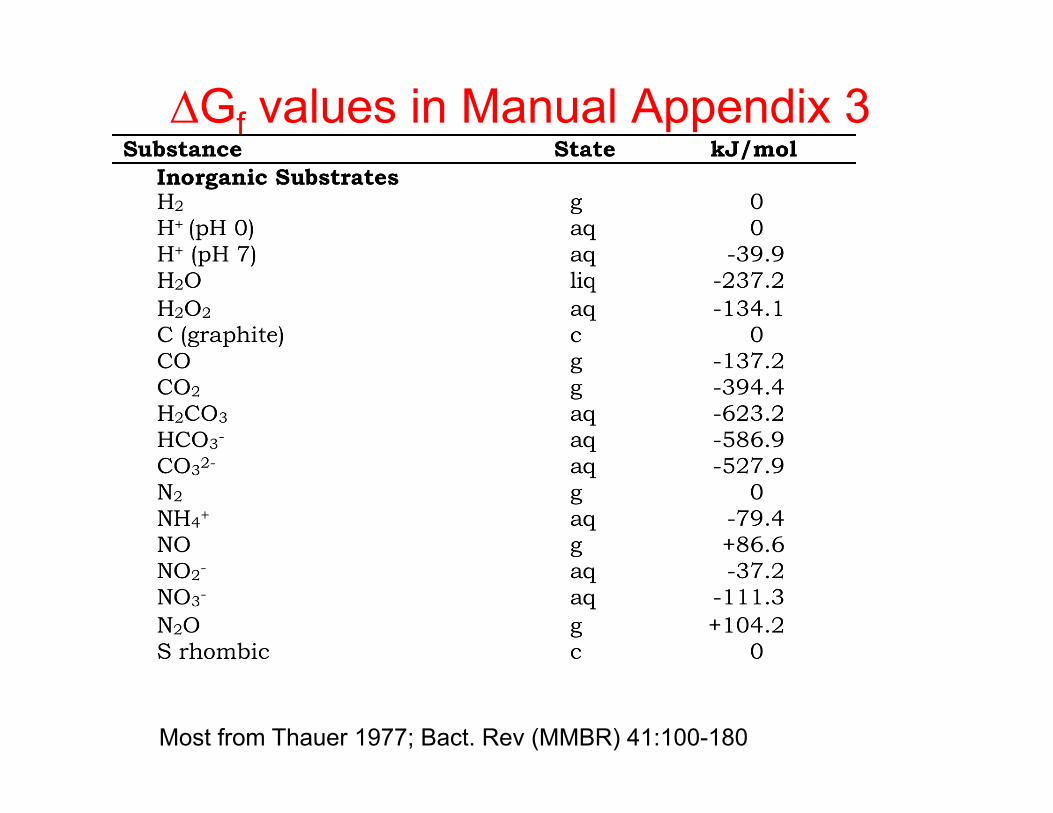
∆Gf values in Manual Appendix 3
Most from Thauer 1977; Bact. Rev (MMBR) 41:100-180

Balancing equations using H2

Example: Aerobic CH4 oxidation Hydrogenation Eqn 27- is backwards
Hydrogenation Eqn 43 – need to balance hydrogens
2O2 +4H2 4H2O -949.0 kJ/rxn
CH4+ 3H2O HCO3- + 4H2 + H+ +135.6 kJ/rxn
CH4+ 2O2 HCO3- + H+ + H2O -813.4 kJ/rxn
Methane + 3H2O HCO3- + 4H2 + H+ +135.6 kJ/rxn
Turn it around:

Concentration affects the free energy of reactions....
€
ΔG'= ΔGo '+RT ln (C)c (D)d
(A)a (B)b
For a reaction: aA + bB cC + dD
....particularly important for anaerobes €
ΔG'= ΔGo '+5.7log (C)c (D)d
(A)a (B)b
“Free energy form of the Nernst Equation”
At 25o C:

Effect of H2 partial pressure on methanogenesis
H2 typically 10-4-10-5 atm in methanogenic habitats
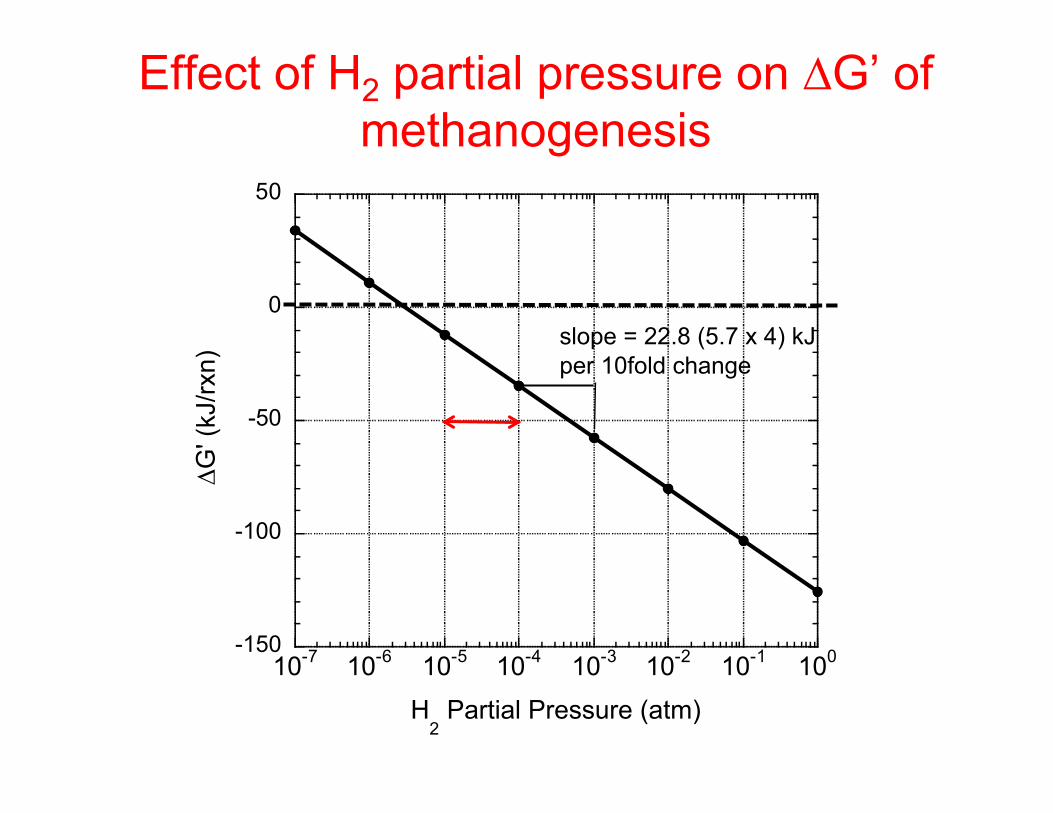
-150
-100
-50
0
50
10-7 10-6 10-5 10-4 10-3 10-2 10-1 100
∆G' (
kJ/rx
n)
H2 Partial Pressure (atm)
Effect of H2 partial pressure on ∆G’ of methanogenesis
slope = 22.8 (5.7 x 4) kJ per 10fold change
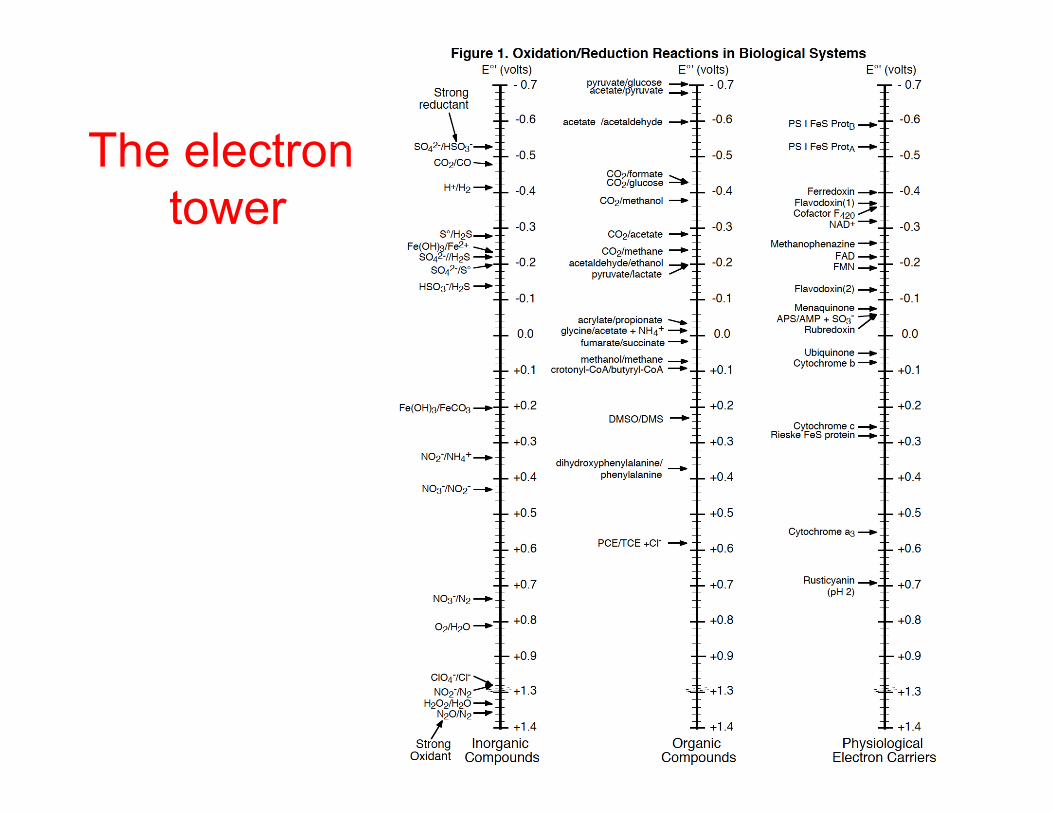
The electron tower

Substrate-level phosphorylation - ATP
∆Go’synthesis = +31.8 kJ/mol from ADP ≈ +40-80 kJ/mol under physiological conditions
Five ATPs are hydrolyzed to ADP to form each amide bond in a protein

R. Thauer MD 2010
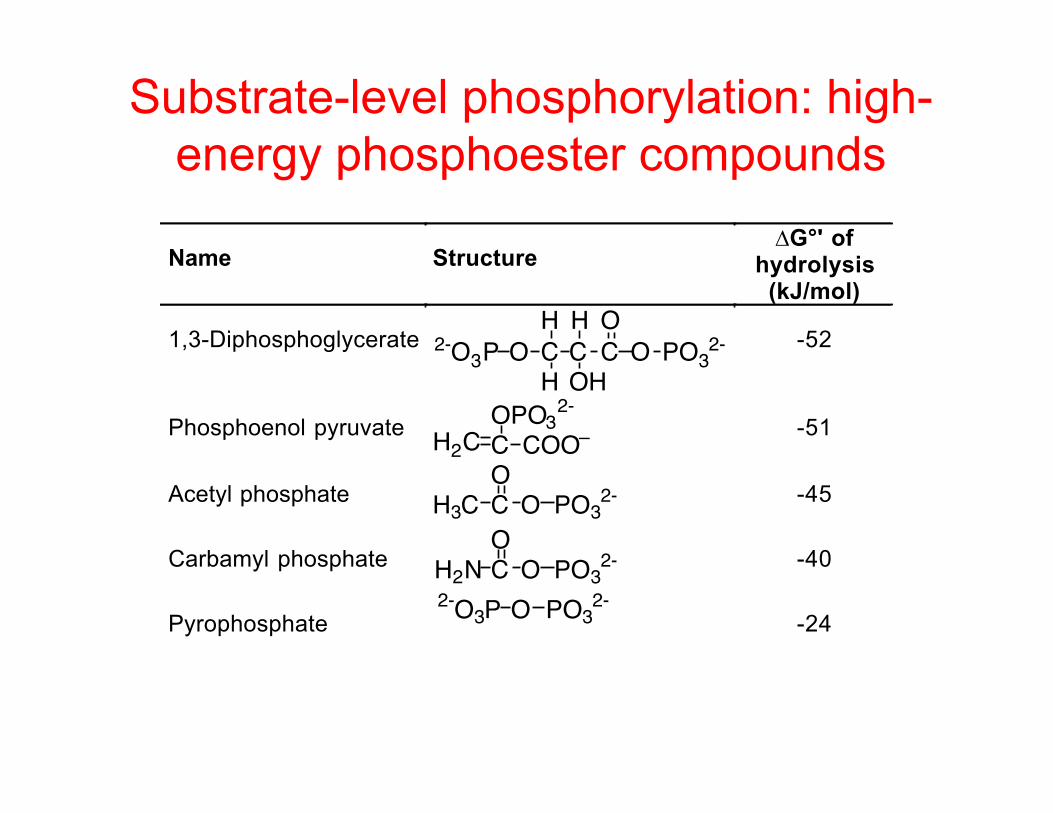
Substrate-level phosphorylation: high-energy phosphoester compounds
Name Structure∆G°' of
hydrolysis(kJ/mol)
1,3-Diphosphoglycerate C C C OOH
O PO32-
H
H OH2-O3P
-52
Phosphoenol pyruvate H2C C COO–OPO32- -51
Acetyl phosphate H3C C OO
PO32- -45
Carbamyl phosphate H2N C OO
PO32- -40
Pyrophosphate2-O3P O PO3
2--24
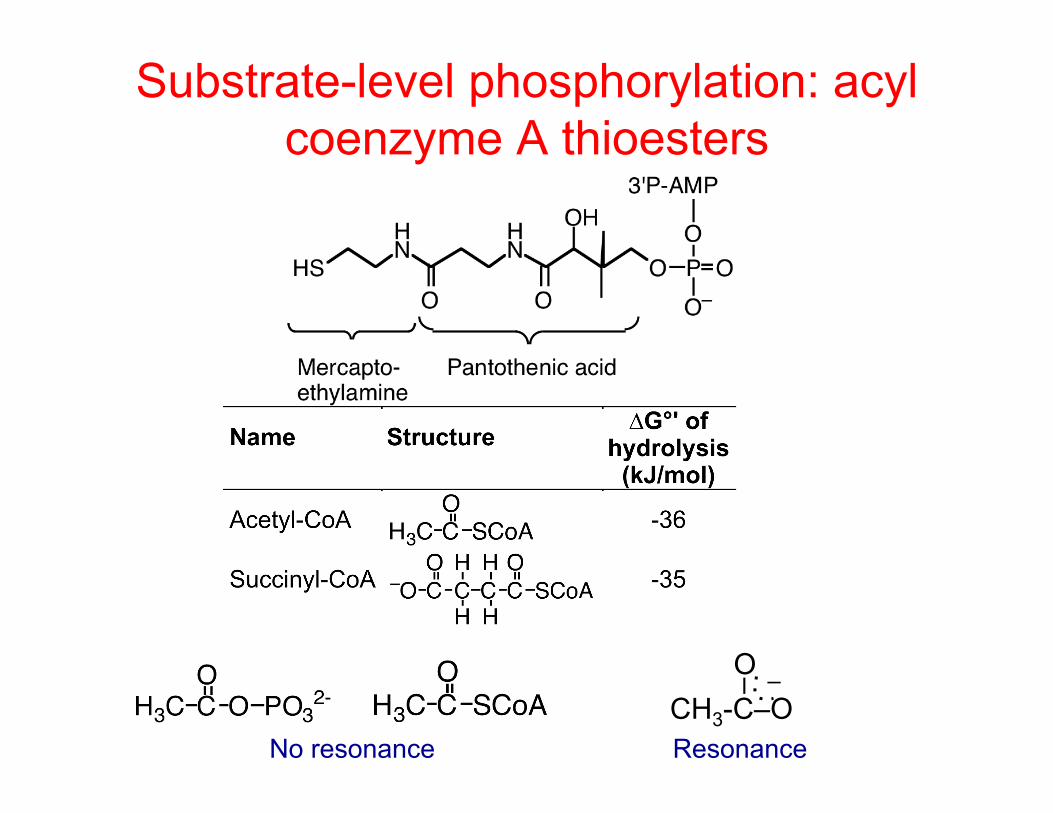
Substrate-level phosphorylation: acyl coenzyme A thioesters
No resonance CH3-C–O
–
O –
Resonance

Acyl coenzyme A thioesters can be cashed in as ATP
Fatty acid or succinate as fermentation product Acyl CoA was probably cashed in as ATP

Fermentation • Latin: "fermentum" - brewing beverages -
connotation of bubbling • Alchemy: a process in which organic chemicals
were transformed – Still used by industrial microbiologists, e.g. the "penicillin
fermentation” • Early 20th century: metabolism of organics in the
absence of oxygen • Brock (13th): Anaerobic catabolism of an organic
compound in which the compound serves both as electron donor and an electron acceptor and in which ATP is usually produced via substrate-level phosphorylation (SLP)

EMP pathway
Now easier to cleave
-ATP -ATP
+2ATP
+2ATP
Aldehyde high energy
+2NADH

Fermentation • Another glycolysis pathway often used by microbes is the Entner
Douderoff (ED) pathway, which only conserves 1 ATP/glucose • There are many variations on these pathways especially in Archaea
Selig et al. Arch Microbiol. 167:217 (1997)

Glycolysis diversity

Fermentation • Net result of EMP is that glucose is converted to pyruvate with the
production of NADH, which needs to be re-oxidized • In aerobes and some anaerobic respirers, the electrons can go
down the electron transport chain to the electron acceptor. • Fermentative organisms don't have that option • Must dispose of electrons from glycolysis • Show three (and a half) solutions
Glucose
2 NADH + 2 H+2 ATP
2 ADP + 2Pi
2 Pyruvic acid
2 NAD+
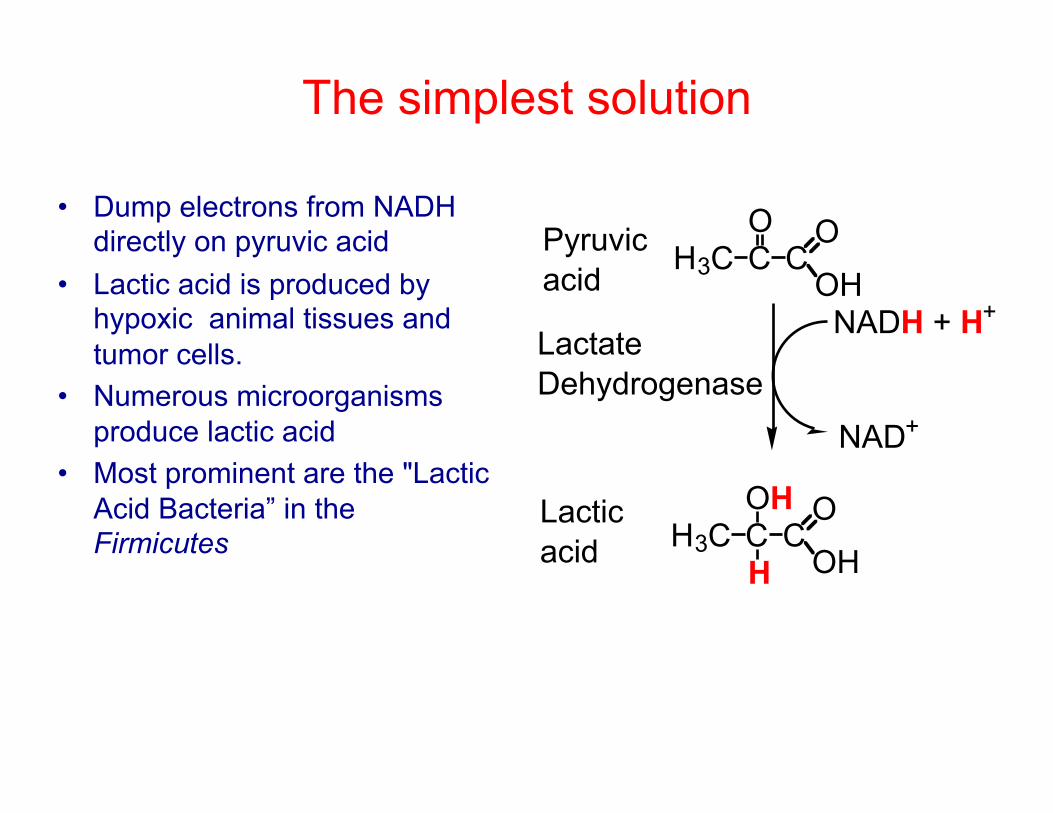
The simplest solution
• Dump electrons from NADH directly on pyruvic acid
• Lactic acid is produced by hypoxic animal tissues and tumor cells.
• Numerous microorganisms produce lactic acid
• Most prominent are the "Lactic Acid Bacteria” in the Firmicutes
H3C CO
CO
OH
H3C COH
CO
OH
Pyruvicacid
Lacticacid
NADH + H+
NAD+
H
LactateDehydrogenase
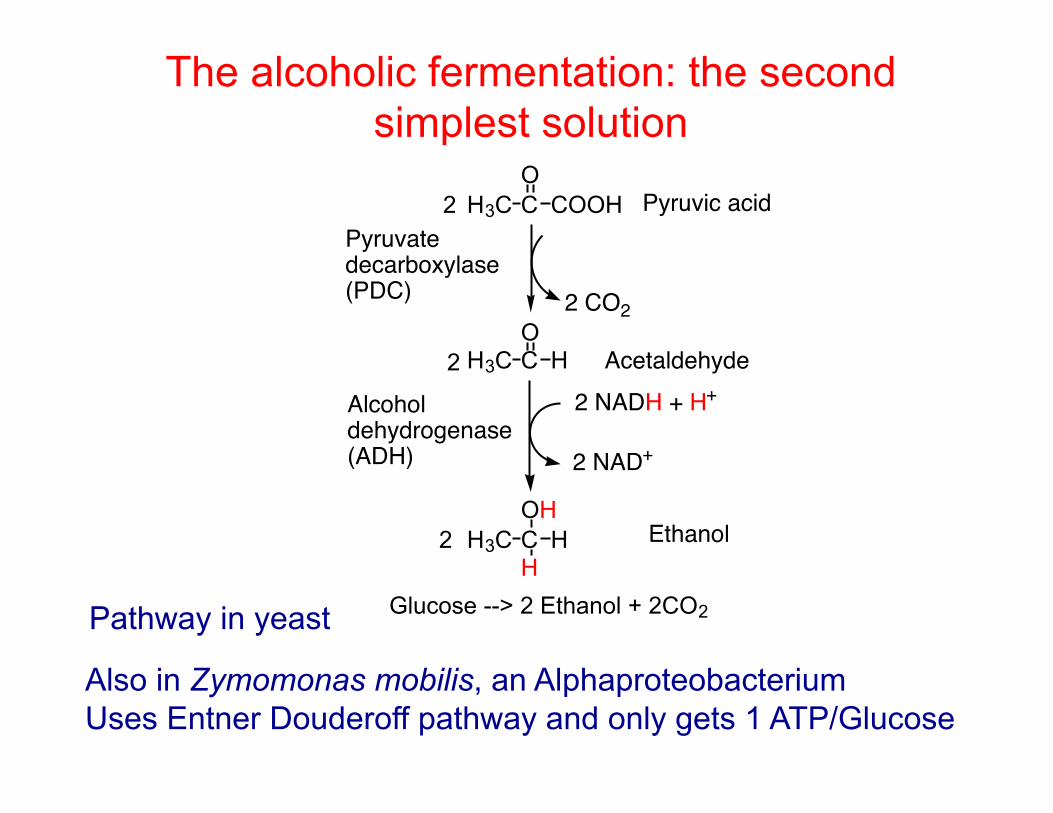
The alcoholic fermentation: the second simplest solution
H3C C COOHO
H3C C HO
2 CO2
2 NADH + H+
2 NAD+
Pyruvic acid
Acetaldehyde
Alcohol dehydrogenase(ADH)
Pyruvate decarboxylase(PDC)
H3C C HOH
HEthanol
Glucose
2 NADH + 2 H+2 ATP
2 ADP + 2Pi 2 NAD+
2
2
2
Glucose --> 2 Ethanol + 2CO2Pathway in yeast
Also in Zymomonas mobilis, an Alphaproteobacterium Uses Entner Douderoff pathway and only gets 1 ATP/Glucose
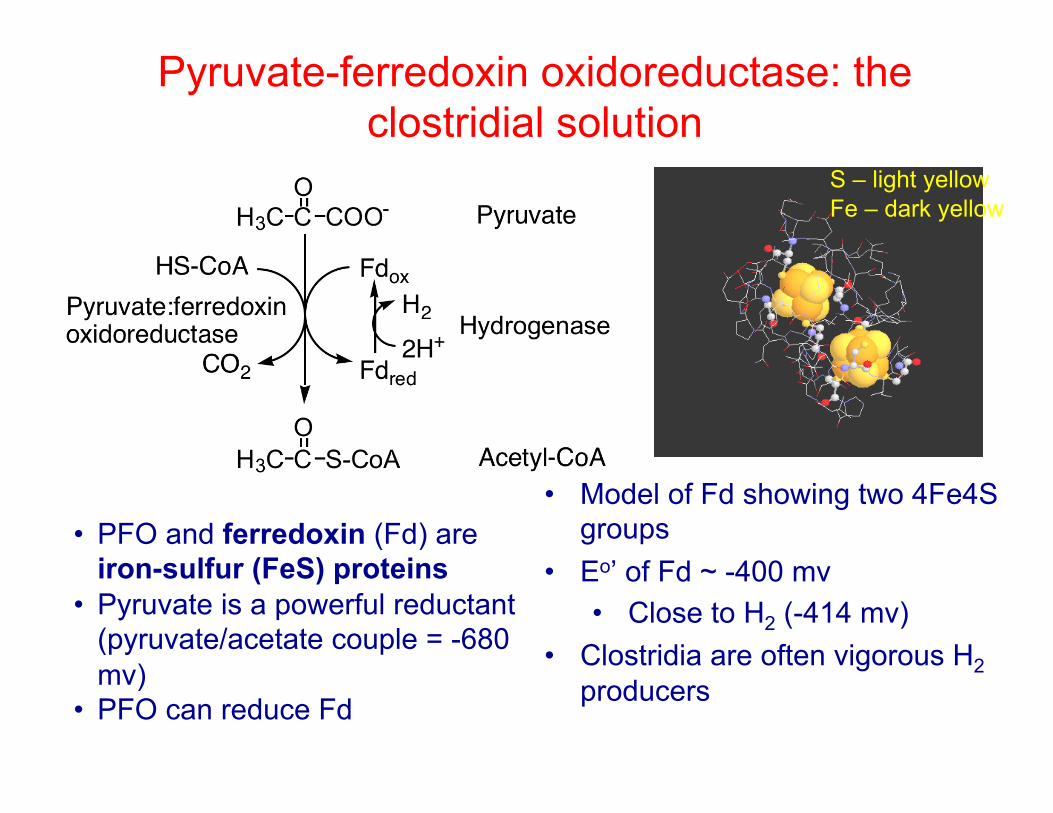
Pyruvate-ferredoxin oxidoreductase: the clostridial solution
• PFO and ferredoxin (Fd) are iron-sulfur (FeS) proteins
• Pyruvate is a powerful reductant (pyruvate/acetate couple = -680 mv)
• PFO can reduce Fd
• Model of Fd showing two 4Fe4S groups
• Eo’ of Fd ~ -400 mv • Close to H2 (-414 mv)
• Clostridia are often vigorous H2 producers
S – light yellow Fe – dark yellow

Hydrogenases • Carry out the seemingly simple reaction: H2 2e– + 2H+
Ribbon model of [FeFe] hydrogenase showing FeS centers leading to active site
H2ase active sites
From: Science 321:572, 2008
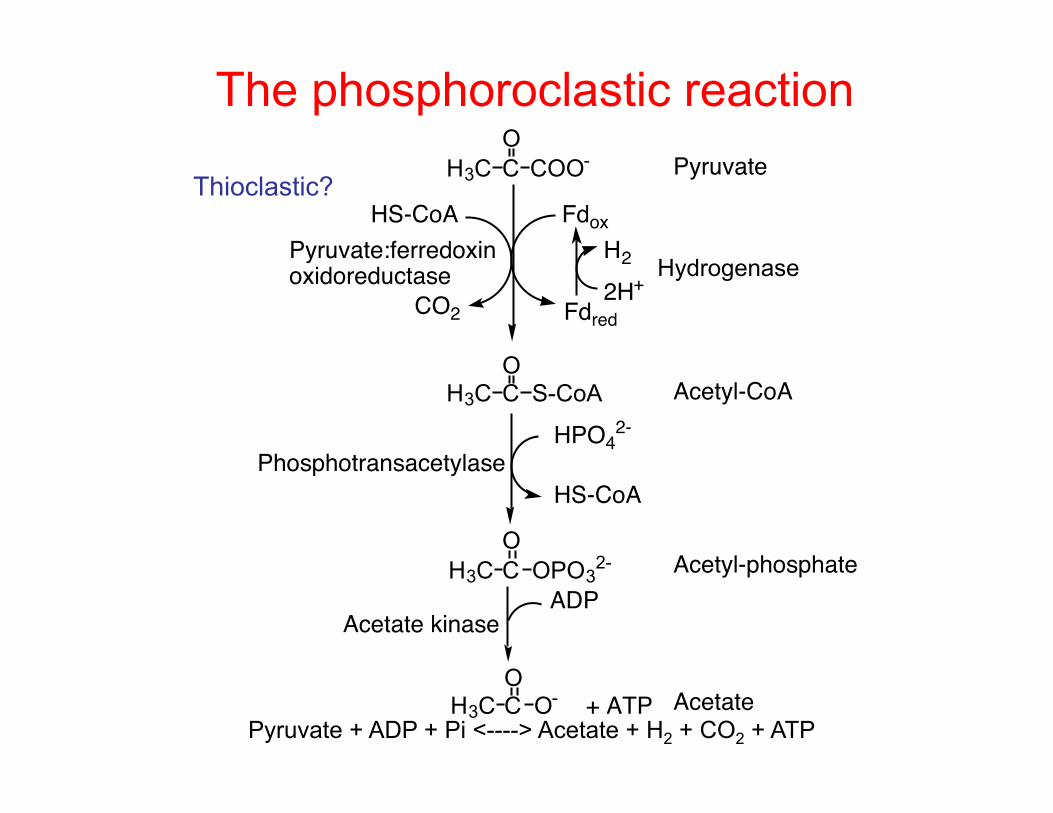
The phosphoroclastic reaction H3C C COO-
O
H3C C S-CoAO
H3C C OPO32-
CO2
HS-CoA
O
Fdox
Fdred2H+
H3C C O-
HPO42-
H2
O
HS-CoA
ADP
+ ATP
Pyruvate
Acetyl-CoA
Acetyl-phosphate
Acetate
Acetate kinase
Phosphotransacetylase
Pyruvate:ferredoxinoxidoreductase Hydrogenase
Pyruvate + ADP + Pi <----> Acetate + H2 + CO2 + ATP
Thioclastic?
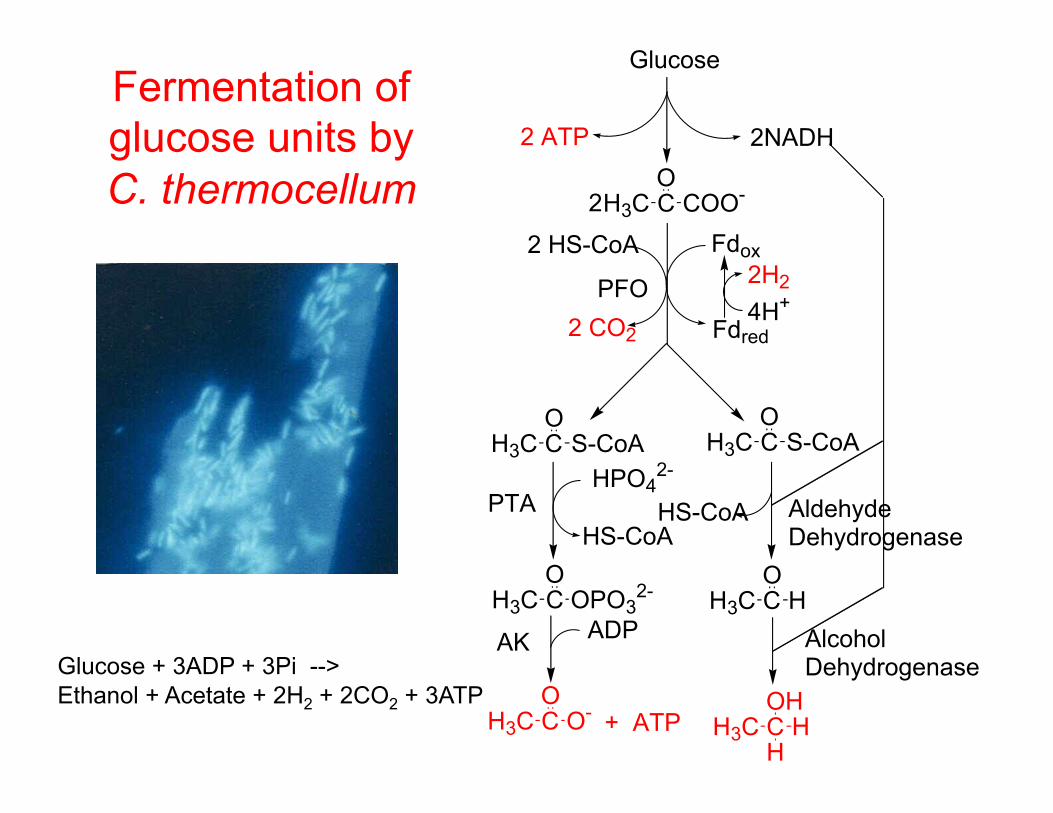
Fermentation of glucose units by C. thermocellum
Glucose + 3ADP + 3Pi --> Ethanol + Acetate + 2H2 + 2CO2 + 3ATP
H3C C COO-O
H3C C S-CoAO
H3C C OPO32-
O
H3C C O-O
2 HS-CoA
2 CO2
Fdox
Fdred4H+
H3C C S-CoAO
2H2PFO
Glucose
2NADH
H3C C HO
HPO42-
HS-CoA
2
H3C C HOH
+ ATPH
ADP
2 ATP
HS-CoAPTA
AK
AldehydeDehydrogenase
AlcoholDehydrogenase
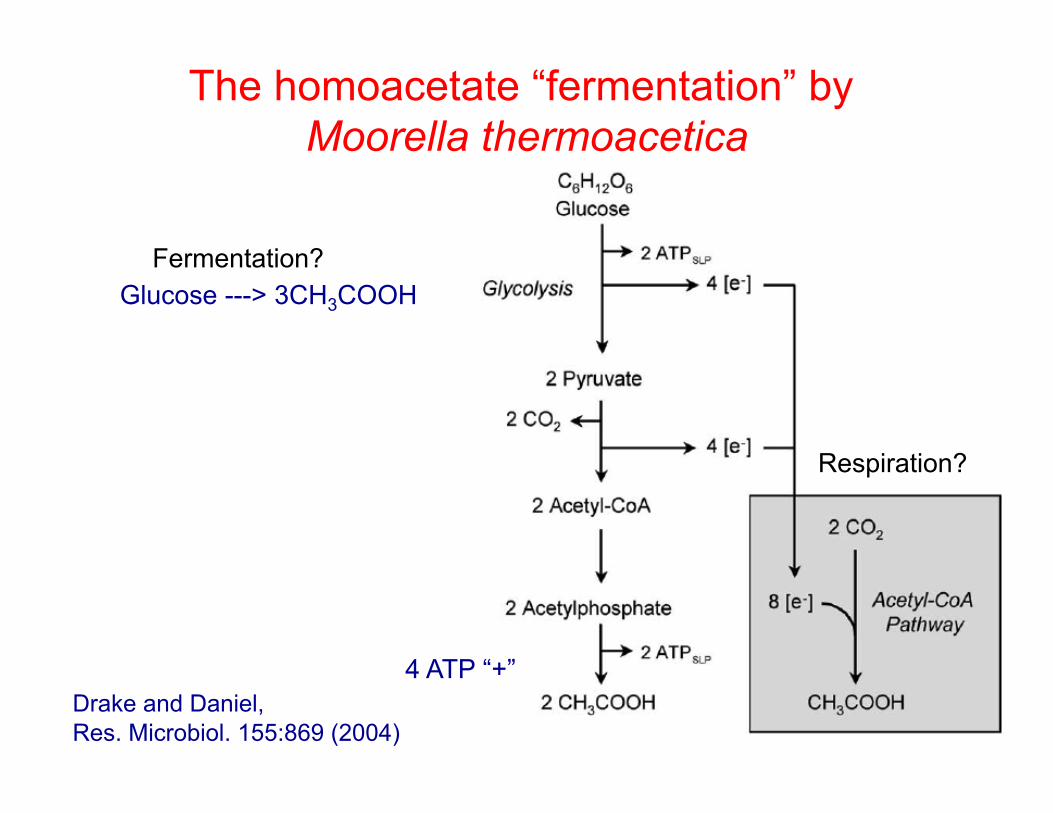
The homoacetate “fermentation” by Moorella thermoacetica
Glucose ---> 3CH3COOH
Respiration?
Fermentation?
4 ATP “+” Drake and Daniel, Res. Microbiol. 155:869 (2004)

Butyrate fermentation in clostridia
From Gottschalk
Some Cl. ferment AAs Products include: putrescine, cadaverine, branched chain FAs, H2S, methyl mercaptan
Also butanol and acetone
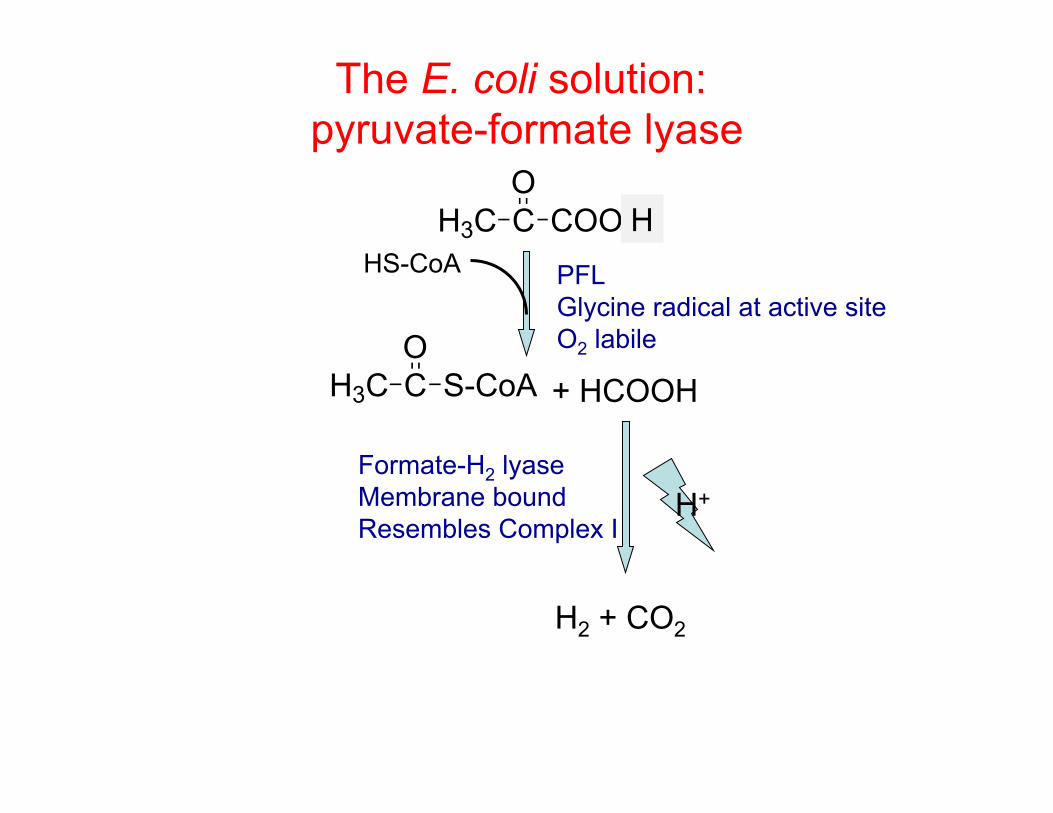
The E. coli solution: pyruvate-formate lyase
H3C C S-CoAO
HS-CoA H3C C COO-
O
+ HCOOH
Formate-H2 lyase Membrane bound Resembles Complex I
H
H2 + CO2
H+
PFL Glycine radical at active site O2 labile

The E. coli mixed acid fermentation - a little bit of everything glucose
(H) ADP ATP
PEP ADP ATP
pyruvate
CO2 oxalo- acetate
TCA cycle
fumarate
succinate
Fumarate reductase
H+
formate
(100)
CO2 H2
Acetyl-CoA
CoA
Acetaldehyde
(H)
ethanol Acetyl-Pi
acetate
lactate
ADP ATP
Pi CoA
(H)
(H)
H+
(49.8)
(35.5)
(79.5)
(88.0) (75.0)
(2.4)
(10.7)
After Gottschalk, Bacterial Metabolism, 1985
= 600 mol C
Products = 531 mol C

Fermentations: summary • O2 is limiting in many environments and organisms
need to dispose their electrons • One solution is fermentation, using the organic
substrate as the electron acceptor • Some facultative and aerotolerant anaerobes use
simple fermentations producing lactate or ethanol as products
• Most true anaerobes increase their energetic yield by making acyl-CoA intermediates which can be cashed in as ATP
• These pathways usually involve disposing of electrons as H2 in the phosphoroclastic reaction or PFL

A controversial proposal

Proton motive force
+++----+
Pump
A
B
H+H+
H+H+
H+
H+
H+
H+
H+H+H+
H+H+
H+
H+
H+ H+H+H+
H+ H+H+
H+H+
H+H+
H+H+
H+H+
H+
H+
H+
H+H+H+
H+
H+H+H+
H+H+
H+
H+
H+
H+
H+
Protons are pumped from compartment A to B. Two forces can drive them back into A 1) the concentration difference
(∆pH) 2) electrostatic attraction (∆Ψ).
The H+ concentration gradient component of the force can be expressed in volts as: RT/nF ln (H+
out)/(H+in)
= 0.059 log (H+out)/(H+
in) = -0.059 ∆pH The electrostatic force can be expressed in volts as: RT/nF ln (ionsout)/(ionsin) = 0.059 log (ionsout)/(ionsin) = ∆Ψ The total proton motive force (∆p) is: ∆p = ∆Ψ - 0.059 ∆pH
In an actively metabolizing cell, ∆p is typically 150-200 mv (0.15-0.2 v)
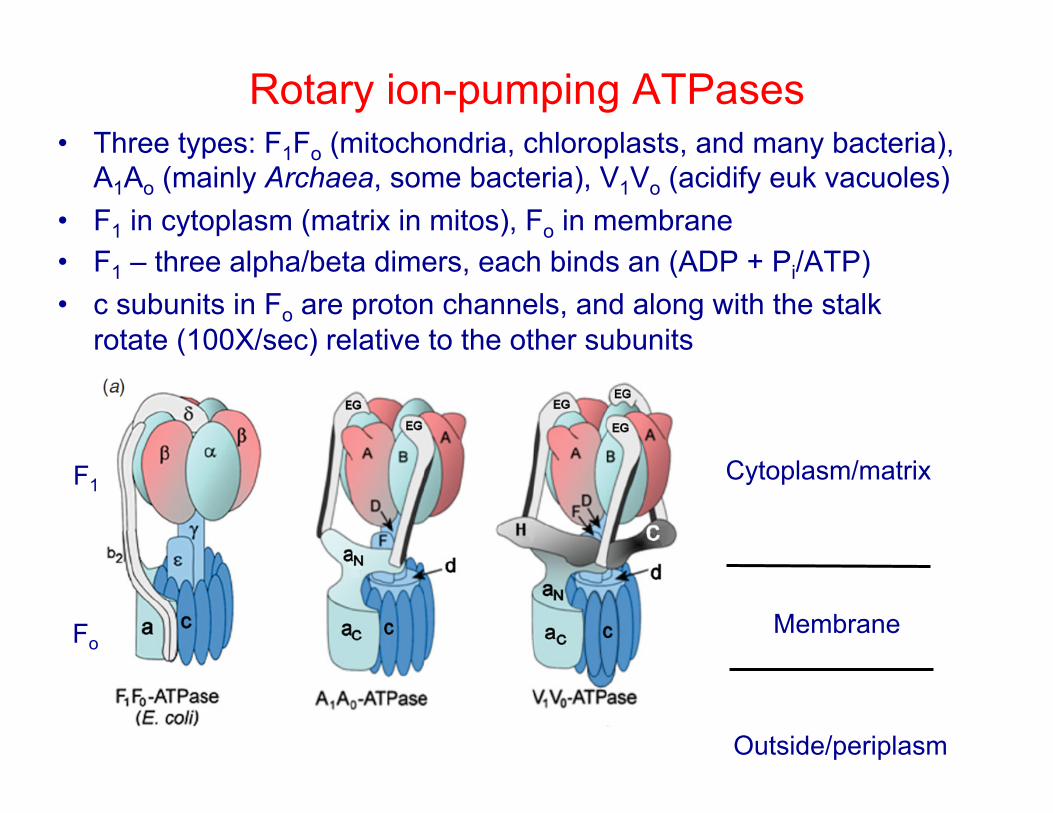
Rotary ion-pumping ATPases • Three types: F1Fo (mitochondria, chloroplasts, and many bacteria),
A1Ao (mainly Archaea, some bacteria), V1Vo (acidify euk vacuoles) • F1 in cytoplasm (matrix in mitos), Fo in membrane • F1 – three alpha/beta dimers, each binds an (ADP + Pi/ATP) • c subunits in Fo are proton channels, and along with the stalk
rotate (100X/sec) relative to the other subunits
Cytoplasm/matrix
Membrane
Outside/periplasm
F1
Fo

F1Fo in action Higher H+ outside (PMF)

Rotary ion-pumping ATPases • ATPases are reversible – may help you to think of as an ATP-
powered proton pump (fan vs windmill) • Each alpha/beta dimer in F1 converts 1 ADP + Pi to ATP per
360o rotation so there are 3 ATP altogether • The question of how many H+/ATP (2,3,4?) was only settled
by a crystal structure of a yeast mitochondrion ATPase • It had 10 c subunits so that per rotation there are 10/3 or 3.33
H+/ATP Science 286:1700 (1999)

ATPase predicted stoichiometries
Organism c subunits cation/ATP Beef heart mitochondria 8 2.67 Yeast mitochondria 10 3.33 Escherichia coli 10 3.33 Acetobacterium woodii 11 3.67 Propionigenium modestum 11 3.67 Thermus thermophilus 12 4 Spinach chloroplast 14 4.67 Various cyanobacteria 13-15 4.33-5 Methanopyrus kandleri 13 4.33
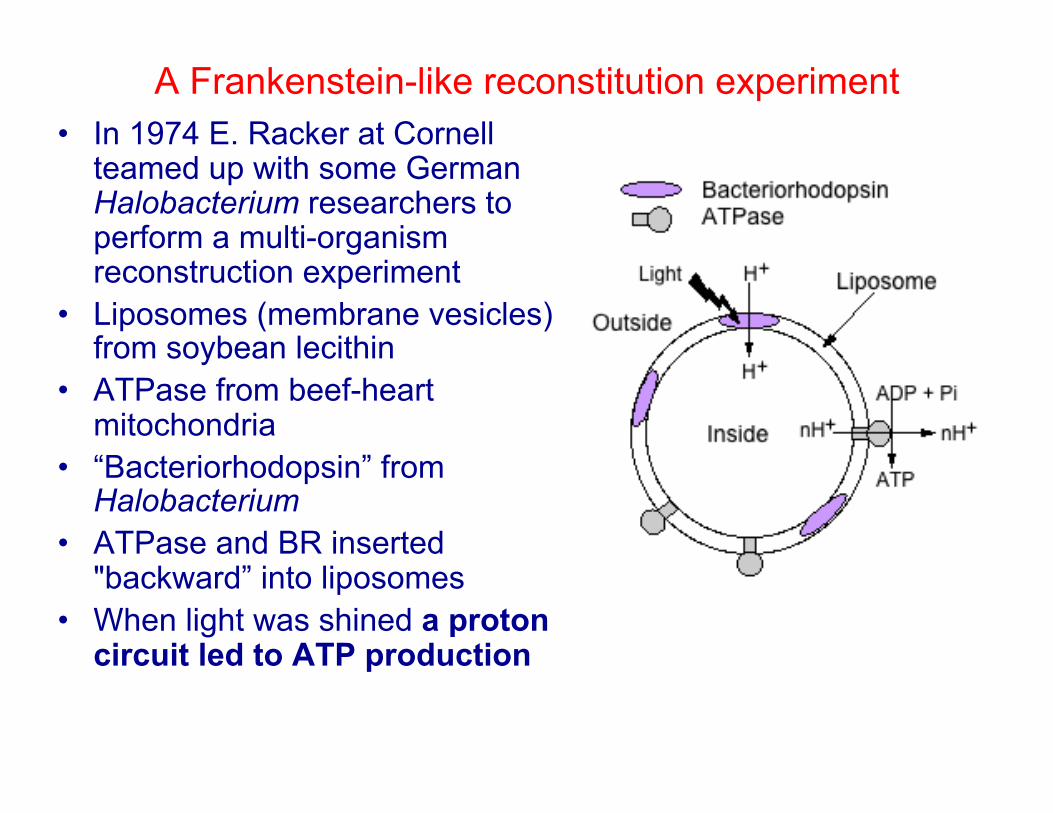
A Frankenstein-like reconstitution experiment • In 1974 E. Racker at Cornell
teamed up with some German Halobacterium researchers to perform a multi-organism reconstruction experiment
• Liposomes (membrane vesicles) from soybean lecithin
• ATPase from beef-heart mitochondria
• “Bacteriorhodopsin” from Halobacterium
• ATPase and BR inserted "backward” into liposomes
• When light was shined a proton circuit led to ATP production

E. coli expressing proteorhodopsin gene
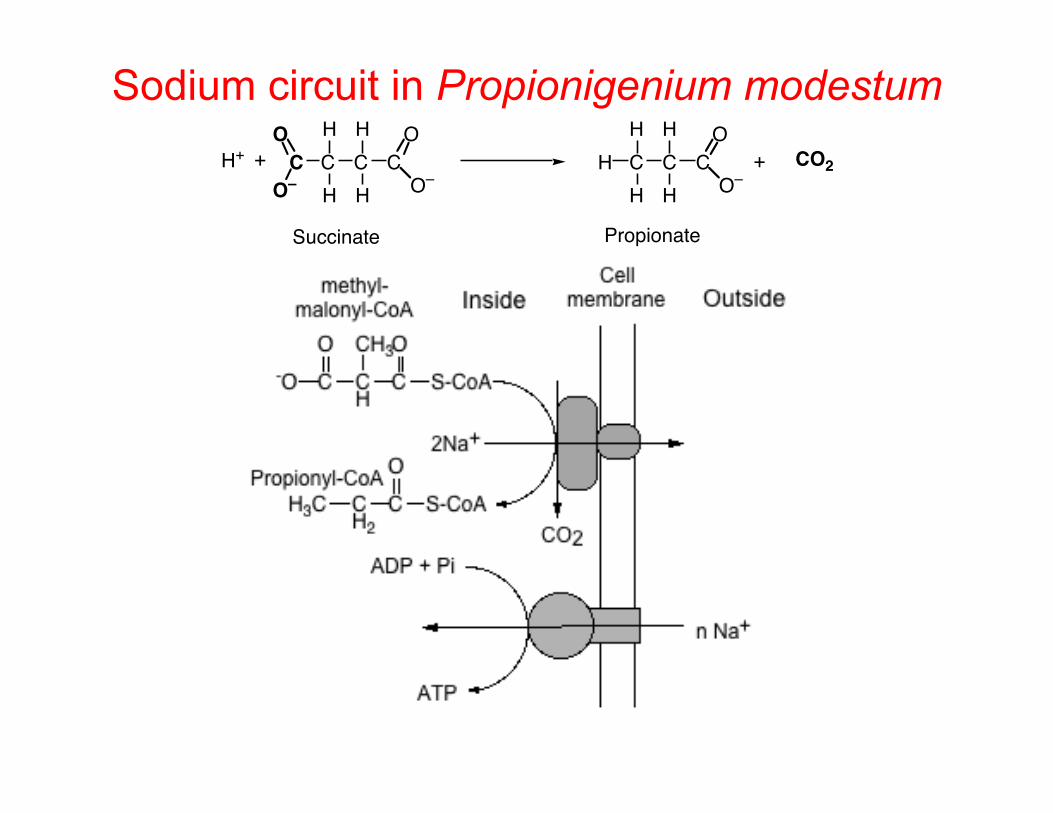
Sodium circuit in Propionigenium modestum C C C C
H
H
H
H O–
OO
O–H+ + C C C
H
H
H
H O–
OH + CO2
Succinate Propionate

Proton motive force and rotary ATPases • ∆p values in respiring organisms are typically 0.15 to 0.2 v, • The production of ATP using 3.3 H+ is energetically
feasible – ∆G= -nFE = 3.3 x 96.4 x (0.15-0.20) – = –48-63 kJ/3.3 H+
• ATPases with higher ratios don’t need as high ∆p – for 15 subunits and 100 mv = 5 x 96.4 (0.10) = 48 kJ/ATP – Like gears on a bike – but organisms can’t change
• ATPases are reversible • In fermentative heterotroph making ATP from SLP (e.g.
Streptococcus) – Low ∆p and high ATP levels – Needs ∆p for transport and motility – ATPase hydrolyzes ATP to pump protons and provide a ∆p (or ∆Na+)

Electron transport-carriers
Couple E°' (v)
H+/H2 -0.41Ferredoxin (Clostridium) -0.41NAD(P)/NAD(P)H -0.32FMN/FMNH2 in NADH dehydrogenase -0.30FeS centers in NADH dehydrogenase -0.35 - 0.0Free FAD/FADH2 -0.22Free FMN/FMNH2 -0.19Menaquinone/menaquinol -0.07Ubiquinone/ubiquinol +0.13cytochrome b +0.06 - +0.26cytochrome c +0.25 - + 0.36Rieske iron sulfur protein +0.28cytochrome a +0.29cytochrome a3 +0.55O2/H2O +0.81
Properties of e- carriers at the end.

Moving protons via electron transport
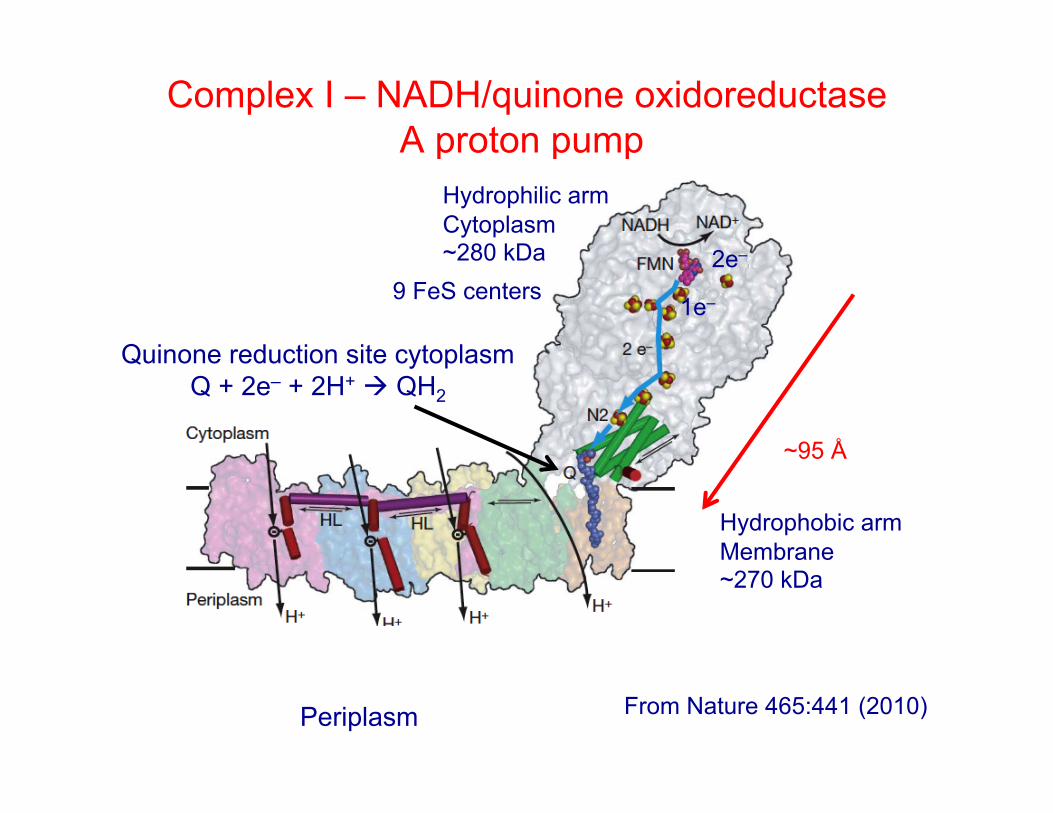
Complex I – NADH/quinone oxidoreductase A proton pump
~95 Å
Quinone reduction site cytoplasm Q + 2e– + 2H+ QH2
Hydrophilic arm Cytoplasm ~280 kDa
Hydrophobic arm Membrane ~270 kDa
From Nature 465:441 (2010)
9 FeS centers 2e–
1e–
Periplasm

Electron transport in aerobically grown E. coli
NADH + H+ NAD+
QH2
QH2 QH2 Q
2H+
2e–
Q QH2
2H+ 4H+
4H+
b
CuB/o3
2e–
2H+ + 0.5O2 H2O
2H+
2H+ 8H+
8H+ 2.4 ADP + Pi 2.4 ATP
Out (periplasm)
In (cytoplasm)
Complex I Complex IV – quinol oxidase b and o3 are hemes CuB is a copper site
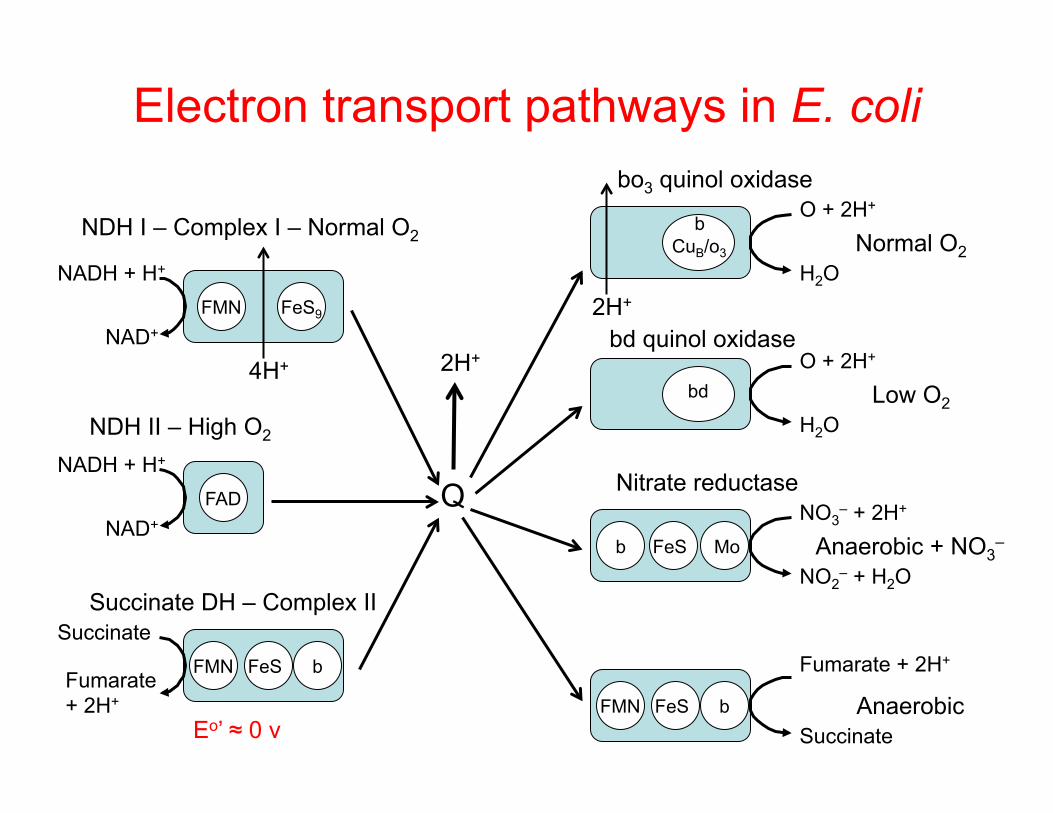
Electron transport pathways in E. coli
b CuB/o3
O + 2H+
H2O FMN FeS9
NADH + H+
NAD+
Q
4H+
2H+
NDH I – Complex I – Normal O2 Normal O2
bo3 quinol oxidase
NADH + H+
NAD+
FAD
NDH II – High O2
FMN FeS b
Fumarate + 2H+
Succinate Anaerobic
bd O + 2H+
H2O Low O2
bd quinol oxidase
Nitrate reductase
b FeS Mo
NO3– + 2H+
NO2– + H2O
Anaerobic + NO3–
2H+
FMN FeS b
Succinate
Fumarate + 2H+
Succinate DH – Complex II
Eo’ ≈ 0 v
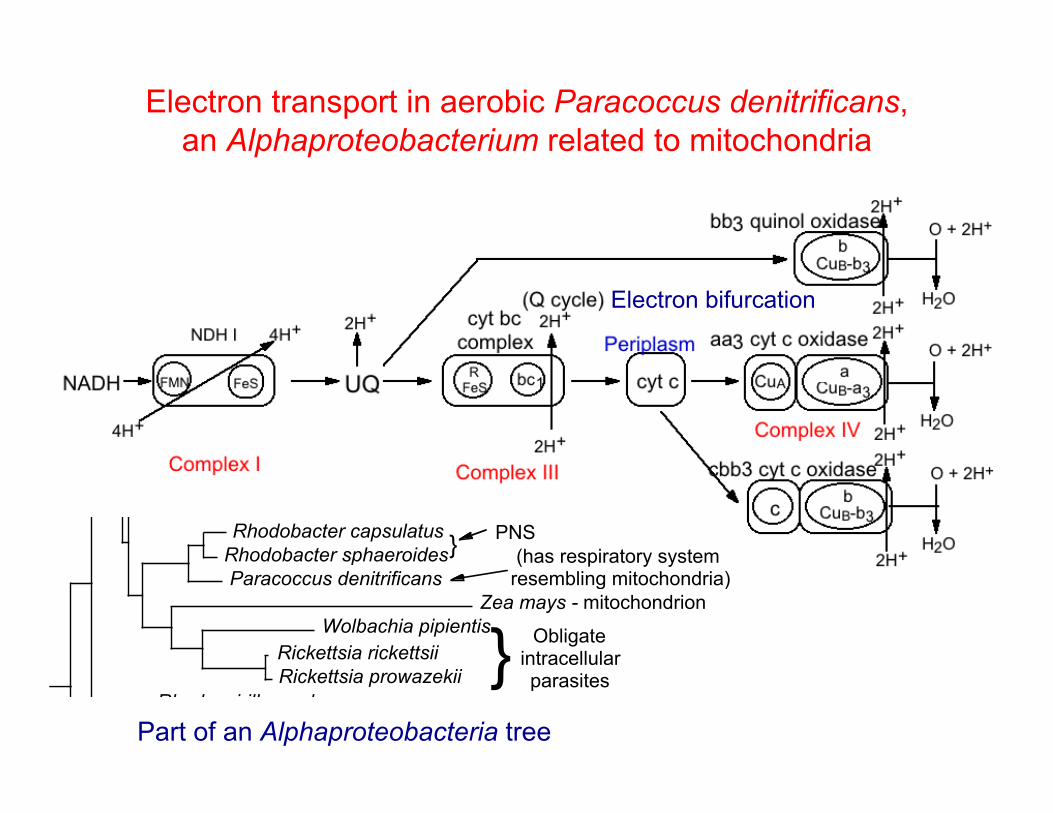
Electron transport in aerobic Paracoccus denitrificans, an Alphaproteobacterium related to mitochondria
Rhodopseudomonas palustrisBradyrhizobium japonicum"Photorhizobium thompsonianum"
Nitrobacter winogradskyiMethylobacterium extorquens
Rhodopseudomonas viridisMethylosinus trichosporium
Rhodomicrobium vannieliiHyphomicrobium vulgare
Rhizobium lotiBrucella abortusBartonella bacilliformis
Rhizobium melilotiAgrobacterium tumefaciensRhizobium leguminosarumAgrobacterium rhizogenes
Caulobacter crescentusSphingomonas paucimobilis
Zymomonas mobilisErythrobacter longus
Rhodobacter capsulatusRhodobacter sphaeroidesParacoccus denitrificans
Zea mays - mitochondrionWolbachia pipientis
Rickettsia rickettsiiRickettsia prowazekii
Rhodospirillum rubrumMagnetospirillum magnetotacticumAzospirillum brasilense
Rhodopila globiformisAcetobacter aceti
Obligate intracellularparasites
(has respiratory system resembling mitochondria)
}
} PNS
Part of an Alphaproteobacteria tree
Electron bifurcation
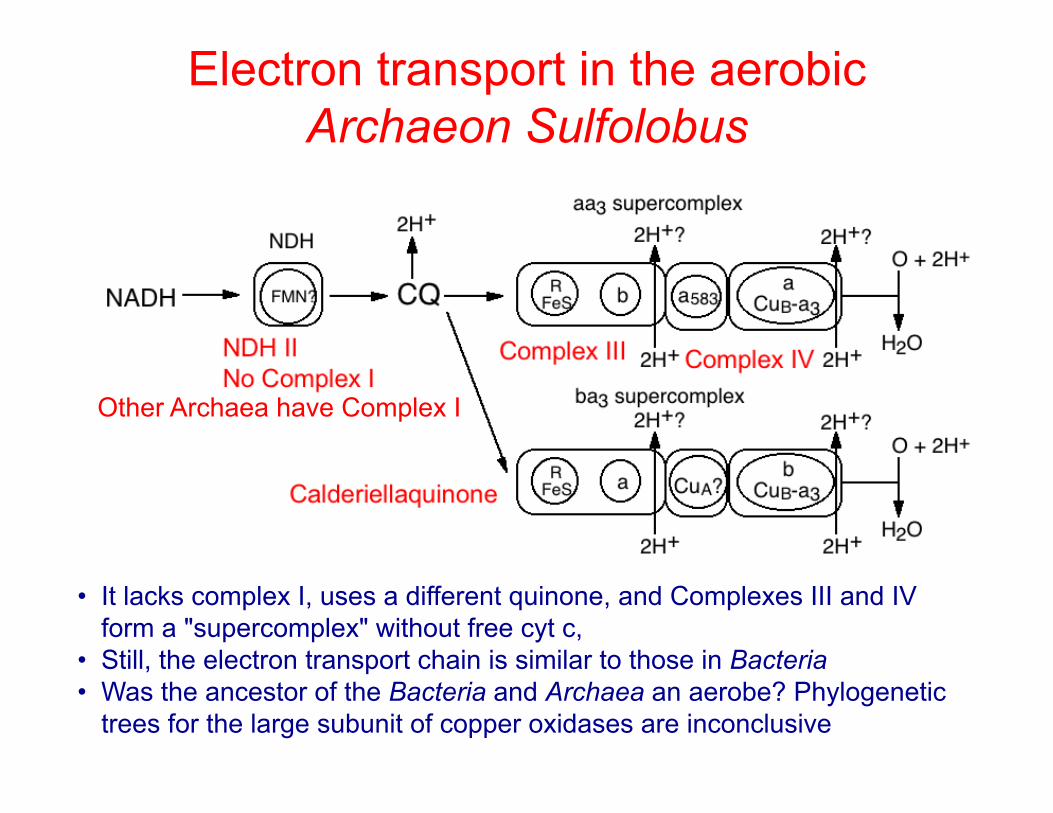
Electron transport in the aerobic Archaeon Sulfolobus
• It lacks complex I, uses a different quinone, and Complexes III and IV form a "supercomplex" without free cyt c,
• Still, the electron transport chain is similar to those in Bacteria • Was the ancestor of the Bacteria and Archaea an aerobe? Phylogenetic
trees for the large subunit of copper oxidases are inconclusive
Other Archaea have Complex I

A truncated electron transport chain in Acidithiobacillus ferrooxidans
• The lithotroph A. ferrooxidans (formerly Thiobacillus) grows aerobically at pH 2 by oxidizing Fe2+ to Fe3+
• Fe2+ is not a strong enough reductant (+0.65 v at pH 2) to reduce NAD+ or quinones
• The electrons feed into the terminal oxidase through two high potential carriers (rusticyanin and cytc533) in the periplasm
Fe2+ Fe3+
cyt aa3
O + 2H+ H2O
2H+
Cyc2? Outer Membrane
Inner Membrane
Periplasm RC cytc553
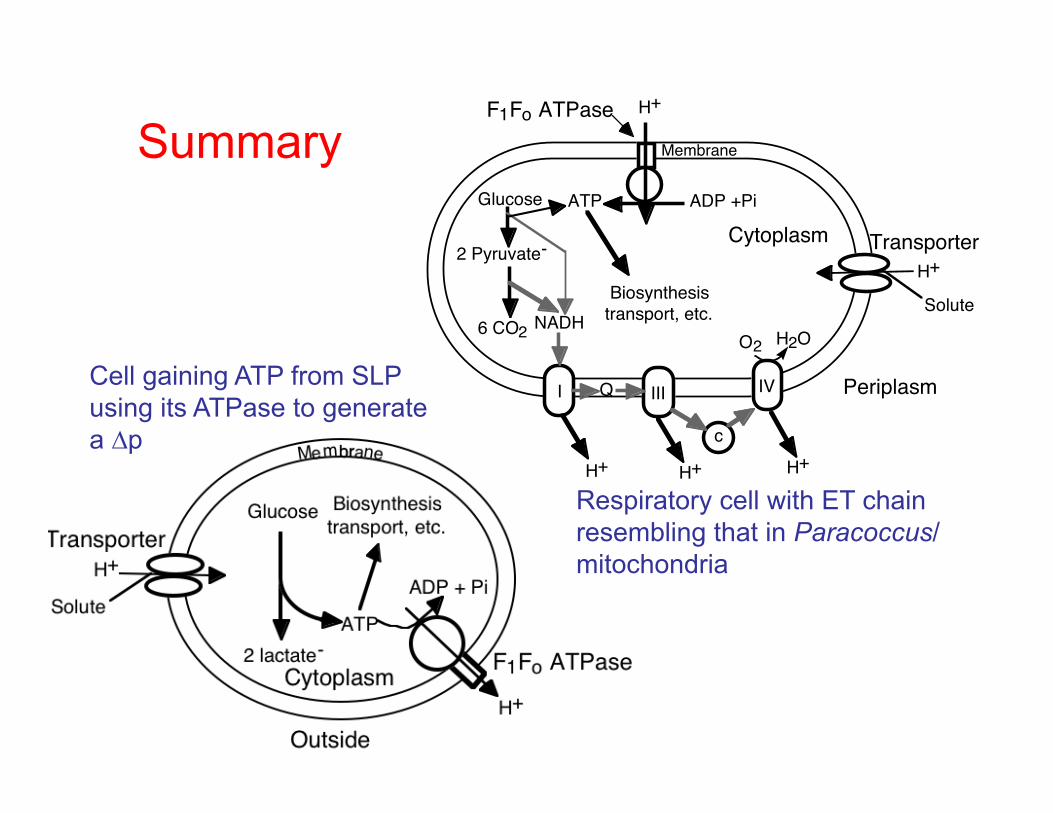
Cell gaining ATP from SLP using its ATPase to generate a ∆p
Summary H+
NADH
Glucose
2 Pyruvate-
6 CO2
I IIIQ
c
H+ H+ H+
O2 H2O
IV
H+
ADP +PiATP
Periplasm
Membrane
SoluteBiosynthesistransport, etc.
Cytoplasm
F1Fo ATPase
Transporter
Respiratory cell with ET chain resembling that in Paracoccus/mitochondria

Volta 2013
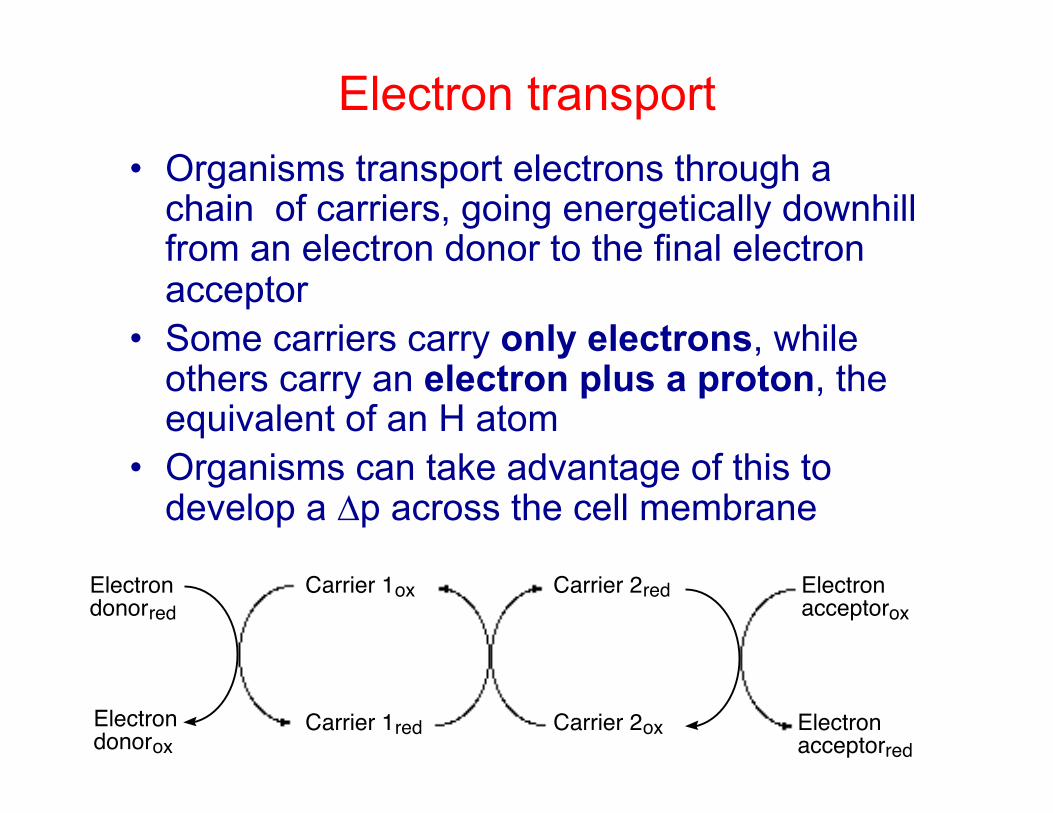
Electron transport • Organisms transport electrons through a
chain of carriers, going energetically downhill from an electron donor to the final electron acceptor
• Some carriers carry only electrons, while others carry an electron plus a proton, the equivalent of an H atom
• Organisms can take advantage of this to develop a ∆p across the cell membrane
Electron donorox
Electron donorred
Carrier 1red
Carrier 1ox
Carrier 2ox
Carrier 2red Electron acceptorox
Electron acceptorred
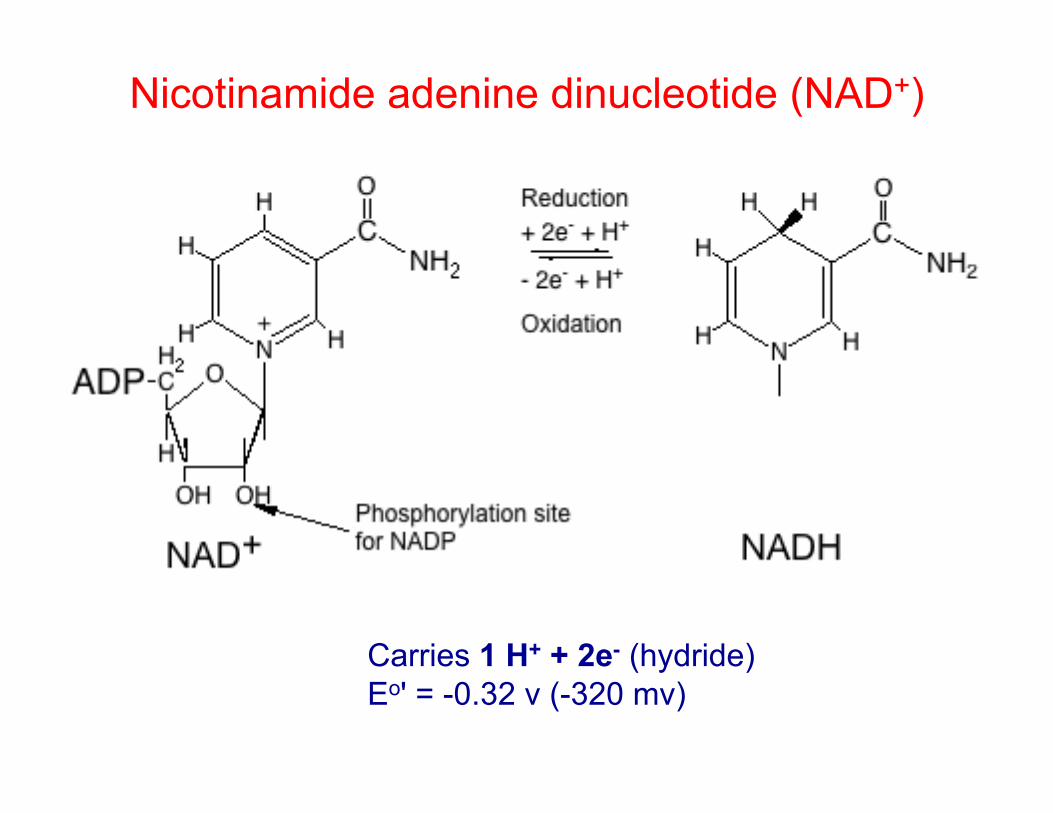
Nicotinamide adenine dinucleotide (NAD+)
Carries 1 H+ + 2e- (hydride)
Eo' = -0.32 v (-320 mv)
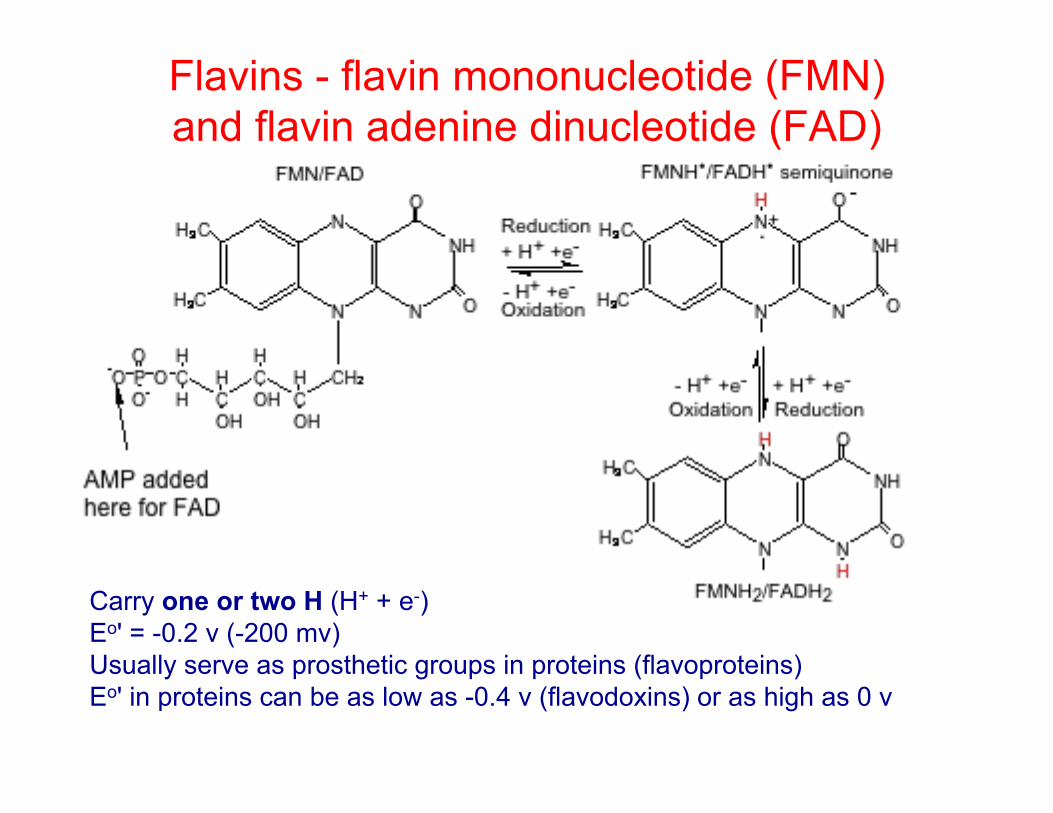
Flavins - flavin mononucleotide (FMN) and flavin adenine dinucleotide (FAD)
Carry one or two H (H+ + e-) Eo' = -0.2 v (-200 mv) Usually serve as prosthetic groups in proteins (flavoproteins) Eo' in proteins can be as low as -0.4 v (flavodoxins) or as high as 0 v

Quinones O
O
CH3H3CO
H3CO
OH
OR
CH3H3CO
H3CO
OH
OHR
CH3H3CO
H3CO
O
OR
CH3
O
OR'
SCH3
S
O
O
.
H+ + e-
R
HH3C
H3C
H+ + e-
H+ + e- H+ + e-Ubiquinone
4-8
Semi-quinone radical
Ubiquinol Plastiquinone
Menaquinone
"Calderiellaquinone"
• Have long hydrocarbon chain that anchors them to the membrane
• Carry one or two H (H+ + e-)
• Ubiquinone commonly found in aerobes, Eo' = +0.13 v
• Menaquinone more common in anaerobes, Eo' = -0.07 v
• Plastiquinone found in chloroplasts and cyanobacteria, Eo' = 0 v
• Calderiellaquinone is found in Sulfolobus, a sulfur-oxidizing member of the Crenarchaeota, Eo' = +0.1 v
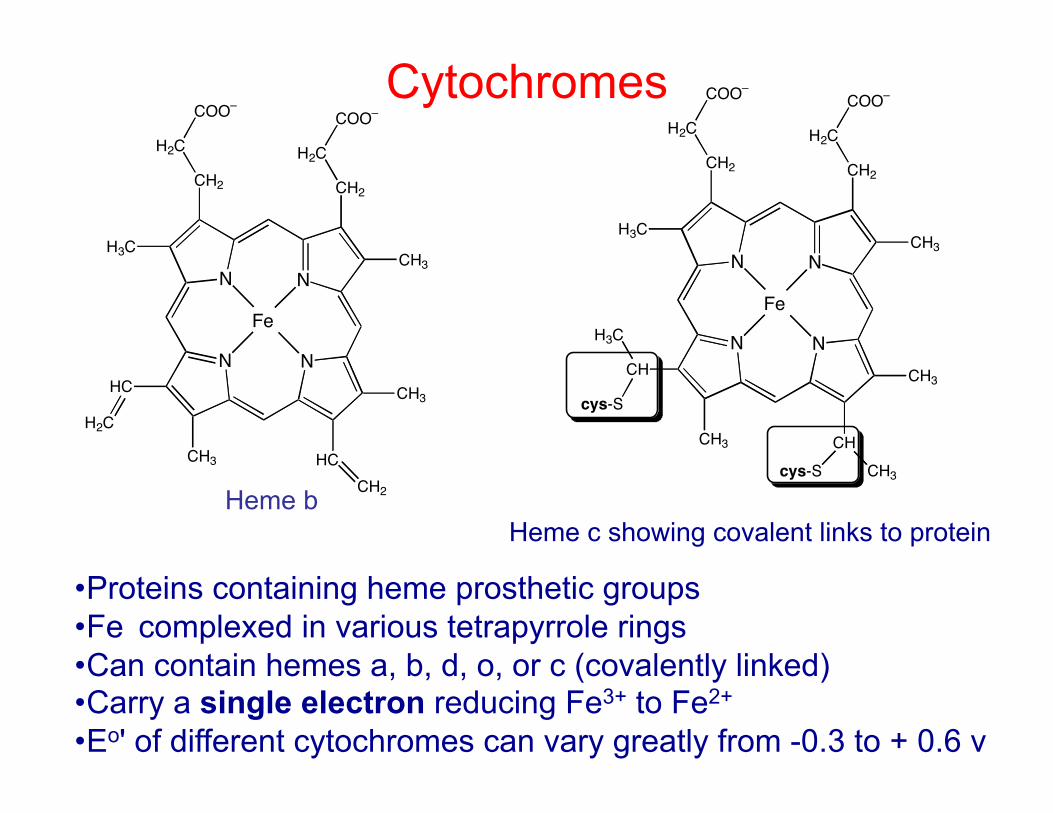
Cytochromes
N N
N N
CH2 CH2
CH3H3C
HC
CH3 HC
CH3
H2C
COO–
H2C
COO–
CH2
H2C
Fe
N N
N N
CH2 CH2
CH3H3C
CH
CH3 CH
CH3
H2C
COO–
H2C
COO–
CH3
cys-S
H3C
cys-S
Fe
Heme b Heme c showing covalent links to protein
• Proteins containing heme prosthetic groups • Fe complexed in various tetrapyrrole rings • Can contain hemes a, b, d, o, or c (covalently linked) • Carry a single electron reducing Fe3+ to Fe2+ • Eo' of different cytochromes can vary greatly from -0.3 to + 0.6 v

Iron-sulfur proteins
2Fe/2S cluster
Rieske 2Fe/2S cluster
4Fe/4S cluster
• Iron sulfur (FeS) proteins have FeS clusters as electron carrying prosthetic groups
• Each cluster can carry a single electron (reducing an Fe3+ to Fe2+) and FeS proteins can have more than one FeS group
• FeS clusters are usually liganded by sulfur groups of cysteines except in the "Rieske" proteins, in which two of the ligands are Ns in histidine
• The Eo' for FeS proteins is typically reducing (-0.53 to 0) except for the Rieske type with a potential of + 0.28
• The importance of FeS proteins was not appreciated because their light/UV spectrum doesn't change significantly on reduction. Can use EPR to detect.
• FeS proteins are probably ancient, derived from naturally forming FeS precipitates