biotechnology and veterinary science: production of veterinary - oie
TRANSCRIPT

Rev. sci. tech. Off. int. Epiz., 1990, 9 (3), 779-794
Biotechnology and veterinary science: production of veterinary vaccines
C. WRAY and M.J. WOODWARD *
Summary: The fundamental principles of genetic manipulation are explained, as are the methods ofproduction of vaccines of veterinary importance. Specific attenuation of micro-organisms may result from genetically-engineered mutations which produce metabolic blocks, from the development of effective bacterial toxoids and from viral gene segment reassortment. Recombinant DNA techniques have been used to produce multi-component vaccines by the insertion of genes into carrier organisms, such as pox viruses and attenuated salmonella, and also to produce subunit vaccines. The use of synthetic peptide vaccines and anti-idiotype vaccines, together with problems of parasite vaccines, are described. Biotechnology would appear to have a major role to play in the future prevention of many animal diseases.
KEYWORDS: Attenuation - Bacteria - Biotechnology - Genetic manipulation -Parasites - Recombinant DNA - Subunit vaccines - Virus.
I N T R O D U C T I O N
Biotechnology may be defined as the application of scientific and engineering principles to the processing of materials by biological agents to produce goods and services. There is nothing new about biotechnology: man has used these techniques for thousands of years to manufacture products such as beer, wine and bread. What is new, however, is the increased capabilities of biotechnology since the development of genetic engineering, more correctly termed recombinant D N A technology, and monoclonal antibody techniques.
Although vaccines against many animal diseases have been available for many years, their efficacy has varied. Some of the available bacterial vaccines, such as clostridial toxoids, have been very successful; others, such as those prepared against Gram-negative organisms, have been less successful because of their poor immunogenicity and the occurrence of fatal adverse reactions. Likewise, some viral vaccines, such as that against rinderpest, have been very successful, whereas others have shown poor attenuation and stability. Some viral vaccines have been inadvertently contaminated with other agents, and many of the recent European outbreaks of foot and mouth disease resulted from inadequately inactivated vaccine.
* Ministry of Agriculture, Fisheries and Food, Central Veterinary Laboratory, New Haw, Weybridge, Surrey KT 15 3NB, United Kingdom.

780
The purpose of this article is to show briefly how the techniques of biotechnology may be used to produce vaccines. It is not our intention to compile an exhaustive list of all known veterinary vaccines. Biotechnology also provides methods that can lead to a greater understanding of microbial virulence and the nature of protective immunogens, bo th of which are important for vaccine development.
G E N E T I C M A N I P U L A T I O N
The information needed for cell growth and metabolism is encoded by genes which are made from a simple chemical alphabet within long sequences of deoxyribonucleic acid (DNA). The D N A consists of two strands each with a backbone of alternate sugar (deoxyribose) and phosphate groups. Linking the two strands in the form of a double helix are pairs of the bases adenine (A), guanine (G), cytosine (C) and thymine (T). Each base is attached to a sugar molecule in one strand, and the two strands of the double helix are linked together by complementary base pairing (that is, A with T and C with G). The sequence of these bases along the D N A strand is the code from which specific proteins and enzymes are synthesised, each of the numerous amino acids that make up a protein molecule being encoded by a sequence of three bases.
During reproduction, genetic information from both parents may be reassorted. Bacteria, however, reproduce by binary fission resulting in clonal populat ions. Gene transfer in bacteria may also occur by the following mechanisms:
a) Transformation: the entry of naked D N A into bacteria. This usually requires some pretreatment of bacterial cells, for example, exposure to high concentrations of calcium ions.
b) Transduction: the transfer of genetic information by bacterial viruses (bacteriophages) which pick up host D N A by accident and transfer it to the host cell.
c) Conjugation: a sexual process involving union between two cells, during which the donor passes D N A to the recipient. Plasmid transfer occurs in this manner.
These mechanisms usually operate only between bacteria of the same or related species, but following a series of fundamental discoveries in the 1960's it has become possible to transfer genetic material across these boundaries by the technique known as "genetic engineering".
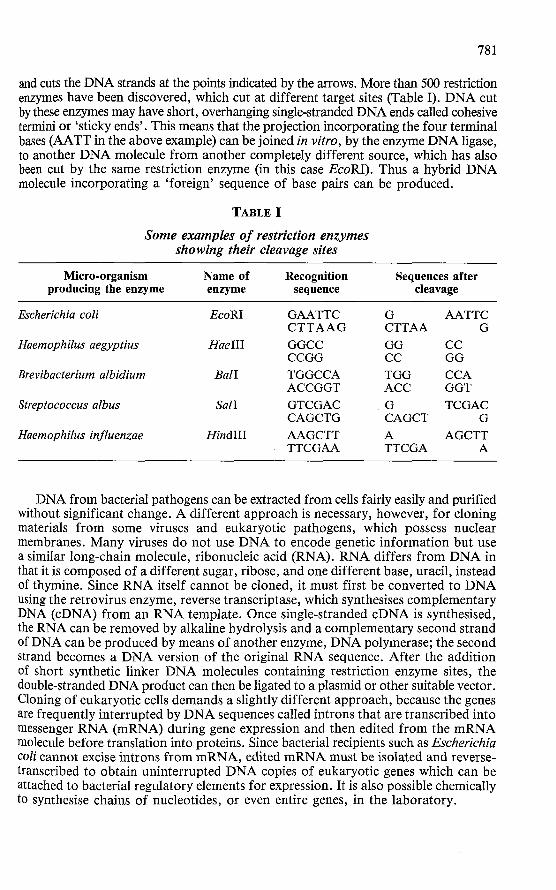
Several stages are involved. D N A is first carefully extracted from the source tissue or organism, then cleaved by one of a group of bacterial enzymes called restriction endonucleases. These enzymes, the natural role of which is to destroy foreign DNA entering a bacterial cell, cut D N A in a predictable manner by recognising a specific sequence of bases in the double helix. D N A fragments are thereby produced. For example, the enzyme EcoRI recognises the sequence:
5' - G A A TT C - 3 '
3 ' - C T T A A G - 5' i
781
and cuts the DNA strands at the points indicated by the arrows. More than 500 restriction enzymes have been discovered, which cut at different target sites (Table I). D N A cut by these enzymes may have short, overhanging single-stranded DNA ends called cohesive termini or 'sticky ends ' . This means that the projection incorporating the four terminal bases (AATT in the above example) can be joined in vitro, by the enzyme D N A ligase, to another D N A molecule from another completely different source, which has also been cut by the same restriction enzyme (in this case EcoRI). Thus a hybrid D N A molecule incorporating a 'foreign' sequence of base pairs can be produced.
T A B L E I
Some examples of restriction enzymes showing their cleavage sites
Micro-organism Name of Recognition Sequences after producing the enzyme enzyme sequence cleavage
Escherichia coli EcoRI GAATTC G AATTC C T T A A G CTTAA G
Haemophilus aegyptius Haelll GGCC GG CC CCGG CC GG
Brevibacterium albidium Ball TGGCCA TGG CCA ACCGGT ACC GGT
Streptococcus albus Sal I GTCGAC G TCGAC CAGCTG CAGCT G
Haemophilus influenzae Hind III AAGCTT A AGCTT TTCGAA TTCGA A
DNA from bacterial pathogens can be extracted from cells fairly easily and purified without significant change. A different approach is necessary, however, for cloning materials from some viruses and eukaryotic pathogens, which possess nuclear membranes. Many viruses do not use D N A to encode genetic information but use a similar long-chain molecule, ribonucleic acid (RNA). R N A differs from D N A in that it is composed of a different sugar, ribose, and one different base, uracil, instead of thymine. Since RNA itself cannot be cloned, it must first be converted to D N A using the retrovirus enzyme, reverse transcriptase, which synthesises complementary DNA (cDNA) from an R N A template. Once single-stranded cDNA is synthesised, the RNA can be removed by alkaline hydrolysis and a complementary second strand of DNA can be produced by means of another enzyme, D N A polymerase; the second strand becomes a D N A version of the original RNA sequence. After the addition of short synthetic linker D N A molecules containing restriction enzyme sites, the double-stranded D N A product can then be ligated to a plasmid or other suitable vector. Cloning of eukaryotic cells demands a slightly different approach, because the genes are frequently interrupted by D N A sequences called introns that are transcribed into messenger R N A (mRNA) during gene expression and then edited from the m R N A molecule before translation into proteins. Since bacterial recipients such as Escherichia coli cannot excise introns from m R N A , edited m R N A must be isolated and reverse-transcribed to obtain uninterrupted D N A copies of eukaryotic genes which can be attached to bacterial regulatory elements for expression. It is also possible chemically to synthesise chains of nucleotides, or even entire genes, in the laboratory.

782
Having obtained a hybrid D N A molecule in vitro, the next step is to transfer the D N A into a bacterium or other suitable host cell and persuade it to produce the gene product . In bacteria, this can be achieved by the use of bacterial viruses or plasmids (Table II) . The simplest self-replicating unit is the plasmid, which is a small circle of D N A encoding very few genes. The circle can be opened by restriction enzyme digestion and a new fragment inserted by ligation to complete the circle once more. It is possible to tailor the insert so that it is placed under the control of a promoter, a gene switch, which can turn on the gene, resulting in its expression and in the product ion of the protein for which it codes.
T A B L E I I
Characterisation of vectors for cloning recombinant DNA
Class of vector Size of vector DNA (kb) Maximum size of foreign DNA insert (kb)
Plasmid 3-15 15 Phage about 50 25 Cosmid 25 50
An alternative vector is a bacterial virus (phage), whose DNA is larger than plasmid DNA; consequently, it is more difficult to manipulate. However, phages will accommodate a larger D N A insert and the recombinant D N A will replicate many hundred-fold in a single bacterial cell, emerging with a complete virus particle which will reinfect surrounding bacteria and so repeat the cycle of amplification.
A third class of vector is a synthetic hybrid of plasmid and phage known as a cosmid. It comprises the D N A insert to be cloned plus two plasmid elements, an antibiotic resistance gene and a short sequence required to start the process of DNA copying in the bacterial host. The cohesive end sequences are phage-derived and they are necessary for circularising the D N A in the bacterial host, itself a prerequisite for replication, and for packaging the D N A in a viral capsid. These vectors are ideal for preparing gene libraries because they may carry inserts as large as 50 kb (that is 50,000 base pairs of double helix DNA) . Other viruses and D N A viruses such as adenovirus and SV40 which have been modified, e.g. small RNA retroviruses, are used to infect animal cells and viral tissue cultures.
Efficient expression of cloned genes requires both genetic and biochemical compatibility within the recipient cell. While the genetic code is the same for all organisms, the DNA-encoded signals that control gene transcription and translation vary from one organism to the next and these signals must be recognised by the host cell if protein synthesis is to proceed. Consequently, much ingenuity has gone into the development of suitable cloning vectors, which now include multiple restriction endonuclease sites located adjacent to promoters , and genes for antibiotic resistance which can be used to select for the presence of recombinant regulatory sequences which influence the expression of the inserted D N A fragment within the host cell.

783
Invertebrate virus expression vectors have been developed for the expression of foreign genes in insect cells (18). Autographa californica nuclear polyhedrosis virus (AcNPV), the prototype baculovirus, has a wide host range and infects more than thirty species of Lepidoptera. The baculovirus utilises the polyhedrin promoter, which has been modified so that the insertion of prokaryotic and eukaryotic genes produces recombinant proteins. One of the major advantages of this vector over bacteria and yeast expression systems is an abundant expression of recombinant proteins, which are in many cases antigenically and functionally similar to their authentic counterpart. Many proteins are naturally glycosylated. This system is particularly useful because the expression of foreign genes in the insect cells often results in near-native glycosylation.
Host cells for gene expression must be found which are suitable for expression of the recombinant protein. While E. coli is probably the most commonly used host, other prokaryotes such as Bacillus subtilis, and simple eukaryotes such as the yeast Saccharomyces cerevisiae, have also been used. The gene encoding the hepatitis B surface antigen was cloned and expressed in E. coli, but expression of the highly immunogenic 22 nm particulate form of this antigen was obtained only when the gene had been inserted into recipient yeast cells (30).
V A C C I N E P R O D U C T I O N
A number of techniques can be used for the development of new vaccines. In some cases recombinant D N A technology provides greater specificity than the empirical approaches of years past, and in other cases the concept is completely novel (Table III) .
T A B L E I I I
Biotechnology for vaccine production
Specific attenuation Bacterial vaccines
a) Mutations - metabolic blocks b) Toxoid vaccines
Viral vaccines a) Recombinations and gene segment reassortments b) Mutations
Recombinant DNA techniques a) Subunit vaccines b) Subunit vaccines in heterologous carriers
Viral carriers Bacterial carriers
Synthetic peptides
Anti-idiotype vaccines

784
SPECIFIC ATTENUATION
It has been known for many years that immunisation with live attenuated organisms can be very effective for producing immunity. Attenuat ion has been achieved by selecting strains of reduced virulence (in the case of bacteria, for example, rough or semi-rough variants), by repeated passage in the laboratory or by growing the organisms under growth-limiting conditions. Live vaccine strains selected in such a manner have always had the disadvantage that they tend to revert to virulence.
BACTERIAL VACCINES
Mutations - metabolic blocks
Cell wall deficient mutants of Salmonella dublin and Salmonella gallinarum which are partially rough strains, have been shown to be effective vaccines in calves (24) and poultry (25) for preventing salmonellosis. However, the S. dublin vaccine causes swelling at the site of inoculation and the occasional fatal allergic reaction. More recently, galE mutants of salmonella with genetic defects created by chemical mutagenesis have been evaluated for their utility as vaccines. These organisms have a lesion which affects galactose metabolism, such that they grow normally in the absence of galactose, but the presence of galactose in the animal leads to their lysis (11). However, they survive long enough to produce strong immunity.
Auxotrophic mutants , also obtained by chemical mutagenesis, have defects which render them dependent on metabolites not present in the vaccinated animal. Examples of such mutants are the purine-requiring (pur) salmonella strains which are commercially available in the German Democratic Republic (19). These strains also possess a second at tenuation marker , usually histidine auxotrophy (his), and a detection marker to distinguish them from wild-type strains.
An alternative approach to the production of attenuated auxotrophs has been the construction of aroA mutants (14). The aroA gene is one of several genes encoding enzymes which form the pre-chorismate biosynthetic pathway, the only route by which aromatic compounds can be synthesised in bacteria. This pathway is absent in animal cells. The use of aro A mutants as vaccines for farm animals has given good results (26).
Another approach to selecting attenuated strains with previously characterised mutations is to introduce random mutations into the genome by means of transposons and to screen the resulting bacteria for attenuation. The transposon TnphoA contains a coding region for bacterial alkaline phosphatase. This t ransposon is constructed in such a way that if it inserts into a gene for a secreted protein, active alkaline phosphatase is secreted; a simple chromagenic substrate in the plating medium differentiates between strains producing active and inactive alkaline phosphatase (28). Thus , TnphoA can be used to identify genes encoding secreted proteins. Deletion mutat ions, usually induced by UV-irradiation, are very stable and may be used preferentially.
Toxoid vaccines
In bacteria, small deletions within toxin genes may be used to produce toxoid vaccines which react immunologically the same way as toxin, but lack the active site of toxicity. Enterotoxigenic E. coli (ETEC) are important causes of diarrhoea and

785
possess two main virulence determinants - an adhesin, which allows the organism to adhere to the mucosa, and enterotoxins, which are responsible for fluid loss. The enterotoxins may be of two types, a heat-stable toxin (ST), which is a small peptide and poorly immunogenic, and a heat-labile toxin (LT), which is highly immunogenic. LT consists of two subunits, the enzymically active A subunit, which is responsible for the toxic activity, and an immunogenic B subunit, which is responsible for binding the A subunit to the epithelial cells of the intestine. The toxin is not readily purified by conventional means and cannot be converted to a toxoid. The LT genes were cloned, the coding sequences within the A subunit gene deleted and a powerful E. coli promoter introduced in front of the B subunit gene. This recombinant D N A was introduced by transfection into a non-pathogenic laboratory strain of E. coli by using a high copy number vector. High levels of B subunit were produced in the form of a toxoid which, when used as a vaccine, conferred protective immunity against LT-producing E. coli (7).
Likewise, tetanus toxoid has been produced by cloning portions of the tetanus toxin gene into E. coli and determining the entire D N A sequence of the gene. Using E. coli expression vectors, E. coli strains were produced which expressed port ions of the tetanus polypeptide containing the non-toxic but highly immunogenic C-fragment. These portions protected mice against challenge with tetanus toxin (8).
VIRAL VACCINES
Recombinations and gene segment reassortments
Viruses with segmented genomes, such as influenza and rotavirus, have the natural capability to undergo reassortment of segments following viral co-infection. Highly attenuated temperature-sensitive mutants of influenza virus or avian strains of the virus, which do not replicate efficiently in mammals , have been used with virulent virus strains to co-infect cell cultures. Some reassortments can be isolated which contain the six internal genes from the avirulent parent and the two genes for the surface proteins H A and NA of the wild-type virus. Such reassortments are also being developed for equine influenza virus (21).
The genes of rotaviruses can also undergo reassortment during mixed infections. The reassortment of the major surface antigen (VP7) of human rotavirus into bovine or rhesus rotavirus has been used to produce a strain having reduced growth characteristics, which is being evaluated as a vaccine (12).
Mutations
Deletion has also been used to produce attenuated strains of the herpes group of viruses. Missense mutants of influenza and respiratory syncytial viruses have been produced; temperature-sensitive strains of these viruses, although attenuated for growth at high temperatures, were found to be genetically unstable (5).
Another approach has been to select attenuated virus with monoclonal antibodies. Rotavirus mutants selected in this way contain single amino-acid substitutions although other viral mutants (e.g. rabies), selected in a similar manner , were unstable and reverted to virulence (9).

786
RECOMBINANT D N A TECHNIQUES
Subunit vaccines
Genes from many infectious agents have been cloned and expressed in prokaryotic and eukaryotic cells by using recombinant D N A techniques. Hepatitis B virus (HBV) vaccine is perhaps the most successful vaccine produced so far with these techniques. The current licensed HBV vaccine which contains purified 22 run HB surface antigen, is made from plasma of human carriers and is costly to prepare. Although hepatitis B surface antigen (HBsAG) could be synthesised in E. coli, the yields in this host were poor . However, when the gene was transferred into yeast (Saccharomyces cerevisiae), immunogenic particulate forms of HBsAG, which closely resembled those found naturally, were produced and readily extracted (30). Evaluation of the utility of this preparat ion as a vaccine is underway.
Other viral genes which have been cloned and expressed include those encoding the VP1 protein of foot and mouth disease virus (FMDV), influenza A virus haemagglutinin (HA), rabies virus glycoprotein and herpes simplex glycoprotein. In the case of FMDV, the sequence coding for V P 1 , one of the viral proteins against which neutralising antibody is directed, was cloned in E. coli and expressed as a fused polypeptide. However, the polypeptide prepared in this way had low efficacy as a vaccine, apparently because the VP1 polypeptide obtained by cloning is folded differently from the VP1 polypeptide in its native configuration in the viral coat, and the immunogenicity is thereby altered (16). Likewise, although a number of different viral proteins have been expressed successfully, the level of immunity they induce has been low and much more work is necessary to produce stable immunogenic products .
Enterotoxigenic E. coli possess somatic pili composed of 14 to 22 kDa protein molecules that enable the organism to adhere to the host 's mucosa. Immunologically distinct pili have been isolated from E T E C , such as K88 and 987P from swine and K99 from cattle, sheep and swine. Vaccines based on purified preparations of these fimbriae have been used in veterinary medicine for a number of years. In producing the first commercial recombinant D N A (rDNA) vaccine, a high copy number plasmid was utilised to increase the yield of the antigen (K88). However, the use of DNA technology to construct pilus vaccines is complicated by the fact that several genes are necessary for the t ransport and processing of the pilus subunit to its final site. Bacteroides nodosus, the cause of ovine footrot, possesses fimbriae which consist of a single repeated structural subunit of about 18 kDa. When these genes were cloned into E. coli, the subunits were not assembled into mature fimbriae but remained embedded in the cell membrane. However, by cloning into Pseudomonas aeruginosa, which possesses compatible type 4 fimbriae, high levels of fimbriae were produced.
The haemagglutinin of fowl plague virus was expressed in larvae of Heliothis viruscens by using recombinant baculovirus, Autographa californica nuclear polyhedrosis virus (AcNPV), as a vector. The larvae were infected with the recombinant virus either by parenteral injection or by feeding. The amount of haemagglutinin synthesised in larvae after parenteral infection was 0 . 3 % of the total protein as compared with 5 % obtained in cultured insect cells. Likewise, the haemagglutinin from larvae was found to be only half as active as that from vertebrate or insect cell culture. However, chickens immunised with larval tissues containing haemagglutinin, when challenged with the virus, were protected from infection (17).

787
Subunit vaccines in heterologous carriers
Virus carriers
Vaccinia virus, adenovirus and herpes virus have been used as carriers, but in general many viruses are unsuitable because of a restricted capacity for foreign D N A and limited growth capacity. Vaccinia virus and related pox viruses, however, retain infectivity even after accommodat ion of up to 25,000 base pairs of foreign D N A (equivalent to 20 average genes) in the viral genome. The principle of construction is that foreign D N A flanked by vaccinia D N A sequences within a plasmid vector can be introduced by transfection into virus-infected cells. Recombination between plasmid and virus creates recombinant vaccinia virus which express antigens from other viruses. Also, these recombinant viruses tend to be less virulent than the parental vaccinia virus in experimental animals and have the potential to produce both cellular and humoral immunity. Vaccinia virus recombinants carrying hepatitis B surface antigen gene have been produced which express high levels of surface antigen in the immunologically active, 22 nm particle form (23).
A recombinant vaccinia virus expressing rabies G glycoprotein has been developed (15). This virus produced high levels of virus neutralising antibodies when fed to foxes, raccoons and skunks, and conferred long-term protection (4). A hybrid vaccinia virus expressing the antigens of influenza virus, hepatitis B virus and herpes hominis virus type 1 elicited antibodies against all three when inoculated into rabbits. Whilst vaccinia-based vaccines have given good results in experimental animals, the use of such vaccines in humans is controversial because vaccinia itself may produce adverse reactions and complications at a frequency of 1:1000. Likewise, repeated inoculation with vaccinia virus cannot be used because this will induce immunity to the vaccinia vector. Consequently, other carriers are being examined.
At present fowl-pox virus appears to be the candidate of choice because its host range is limited and consequently it is unable to multiply in non-avian hosts . Rabies glycoprotein genes and avian influenza genes incorporated into the fowl pox genome are correctly expressed in avian and non-avian cells and, in the latter, poxviral particles are not produced (27).
Bacterial carriers
The attenuated salmonella strains mentioned previously are also being investigated as to their suitability as carriers of heterologous genes. Depending on the strain of salmonella, a variety of immune responses in the host, ranging from local to cell-mediated, can be produced. Because salmonella and E. coli belong to the same bacterial family, foreign genes expressed in E. coli can be propagated in salmonella vaccine strains, usually by means of bacteriophage. Antigens from bacteria (3, 6), viruses (29) and parasites (22) have all been expressed in salmonella. Thus , a plasmid from Shigella sonnei has been introduced into the galE typhoid vaccine (Ty21a) and shown to be safe in volunteers. The gene coding for the E. coli heat labile B subunit toxin has been introduced in salmonella aroA mutants and shown to induce an anti-LT-B secretory immune response and also the product ion of serum antibodies. The mice immunised with these constructs are also immune to subsequent challenge with virulent salmonella. Likewise, the nucleoprotein of influenza virus has been introduced into S. typhimurium, where it is expressed at a high level.

788
SYNTHETIC PEPTIDES
Only a small port ion of an antigen is necessary to stimulate an immune response as long as it is immunogenic or has been made immunogenic by being linked to a carrier. Individual antibodies recognise the small regions of the antigen known as epitopes, and if the structure of the epitope is known, it is possible to synthesise it biochemically. Synthetic peptides whose sequences correspond to epitopes recognised by antibodies, or sequences which are essential for the function of a particular protein, have been manufactured and used to immunise animals.
The first peptide immunogen, corresponding to the VP1 polypeptide of FMDV, stimulated the development of neutralising antibodies in guinea pigs. It was, however, necessary to couple the peptide to a suitable carrier (keyhole limpet haemocyanin) and to use an adjuvant (2). Even so, the immunogenicity was only 1-10% of that of the whole inactivated virus.
However, recent studies have found that the immunological non-responsiveness to FMDV peptide can be overcome by addition of 'foreign' helper T-cell determinants from ovalbumin or sperm-whale myoglobin (10). It was found, however, that the virus-neutralising activity of the antibody raised against peptide was dependent on the specific peptide sequence used. Thus, FMDV peptides with the added sequence 132-148 from sperm-whale myoglobin elicited a high degree of virus neutralising activity in mice, whereas sera from mice which received the peptides with the added sequence 105-121 from sperm-whale myoglobin did not neutralise the virus although they had high levels of anti-peptide activity. The results indicated that the T-cell help given by foreign epitopes is B-cell clone specific.
Synthetic peptides from other viruses have been less successful. Peptides representing various regions of the influenza A virus haemagglutinin elicited antibodies, but these were neither protective nor capable of neutralising virus (13). Another synthetic peptide corresponding to an influenza A antigen was conjugated to tetanus toxoid and, when administered with Freund's complete adjuvant, stimulated the product ion of antibodies and gave partial protection to mice against challenge (20). Chemical synthesis of peptide vaccines for bacterial diseases has been reported. A synthetic peptide, corresponding to an amino-acid sequence of the A chain of the diphtheria toxin, when conjugated to a protein carrier elicited antibodies in guinea pigs that neutralised the dermonecrotic and lethal effects of the toxin (1).
Two antibodies are known to protect against cholera, one being directed against Vibrio cholerae lipopolysaccharide and the other against cholera toxin. Antibodies to the B subunit of the toxin prevent the binding of toxin molecules to mucosal receptors. Consequently, several peptides corresponding to regions of the amino-acid chain of the B subunit have been synthesised and linked to either tetanus toxoid or multi-chain poly D-L alanine; these are currently being evaluated (1).
Although synthetic peptides are a promising development and have a number of important potential applications, their use for vaccines remains uncertain for a number of reasons. To produce a good immune reponse, Freund's adjuvant is often necessary. Furthermore, many synthetic peptides lack T-cell recognition sites, which leads to insufficient co-operation between T and B cells during the early immune response. Finally, many important epitopes are discontinuous and their antigenicity is therefore dependent on the conformation of the viral protein.

789
ANTI - IDIOTYPE VACCINES
The region of an antibody that interacts with an antigen is called the paratope. This consists of six hypervariable loops, each of which is made up of a number of determinants called idiotypes. A portion of the idiotype is the antigen-binding site, and thus antibodies directed against this site, anti-idiotypes, may mimic the conformation of the antigen and serve as surrogate antigens. The advent of monoclonal antibodies and rDNA techniques has made the development of anti-idiotype vaccines a possibility. So far a number of anti-idiotype have been prepared that stimulate immunity, e.g. injection of mice with HBsAg anti-idiotype primes an anamnestic response to subsequent injection with HBsAG. The theoretical possibilities of antiidiotype vaccines are numerous. Such vaccines could be used to prime the immune system, since their immunity is generally specific and of short duration. They could also be used to overcome the immune restriction to complex carbohydrates that exists during the neonatal period. Also, they may be able to eliminate the problems associated with defective configuration of antigens produced by gene cloning of synthetic peptides.
P A R A S I T E S
Parasite vaccines have been considered separately because of the multicellular nature of parasites and their wide biological spectrum, which ranges from intracellular organisms to complex endo- and ectoparasites. During parasite infections, numerous antigens may be processed by the host immune system and the presentation of multiple stages has rendered the isolation of the protective antigen(s) difficult.
A number of existing parasite vaccines are available commercially and these range from the classic lungworm vaccine, which consists of irradiated larvae, to at tenuated Coccidia, Anaplasma and Babesia. It has been shown that non-viable coccidial sporozoite antigens could induce protective immunity in young chickens and that this immunity was cross-species. Published evidence suggests that recombinant molecules have also been successful in conferring immunity. A major surface protein of Anaplasma marginale, M S P - 1 , has been used to immunise cattle and reduce post-challenge parasitaemia and clinical disease. D N A from A. marginale was cloned and expressed in E. coli and MSP-1 was obtained.
Protective immunity against malaria can be induced by vaccination with irradiation-attenuated sporozoites. The immunity is mediated by neutralising antibodies and T-cell effector mechanisms which recognise epitopes in the circumsporozoite. Thus, passive transfer of cytotoxic T-cell clones into mice provided protection against challenge. Likewise, salmonella vaccine strains have been engineered to express the circumsporozoite protein; vaccination with these bacteria provides protection against murine malaria in the absence of antibodies.
Trichinella spiralis is an important public health problem in many countries. Infection of an animal host results in solid immunity. It has been shown that vaccination with a combination of adult and larval antigens confers nearly complete immunity. A number of studies have identified protective antigens, but the yields of antigen from the intact parasite have been low; consequently, in vitro translation of T. spiralis mRNA has been used to prepare a number of polypeptides for protection studies.

790
In the case of ticks and Haemonchus it has been found that parasite gut antigens can induce protection in the host animal, and it may be possible to produce the antigen by molecular techniques.
A D J U V A N T S
Adjuvants are necessary to augment the immunogenicity of many vaccines. Since many synthetic antigens are water-soluble, they tend to be less immunogenic than particulate antigens and adjuvants are often necessary to confer protection. The adjuvants currently in use are aluminium gels and Freund 's complete adjuvant (FA), which is a water-in-oil emulsion with killed mycobacteria. Whilst F A has been used in laboratory animals, it may cause severe local reactions and as a consequence safer adjuvants are being sought.
One candidate is muramyl dipeptide (MDP), which is a synthetic analogue of monosaccharide tripeptide, the active mycobacterial component of FA. M D P in an aqueous solution is non-pyrogenic and active even when administered orally. Synthetic peptide antigens from diphtheria toxin have been covalently linked to M D P and a carrier; when injected into mice, these induced an effective anti-toxin response. MDP has also been added to surfactant polymer, squalene-pluronic block emulsion, which has the ability to bind proteins to the surface of oil drops in the emulsion and possibly produces good antigen conformation.
Another promising adjuvant is immunostimulating complex (ISCOM), which is made from the adjuvant Quil A (active component of saponin). ISCOM forms a complex with amphipathic peptides which are presented on the surface of the matrix. Not all antigens can be incorporated in ISCOMs but they seem to be particularly suited to detergent-soluble membrane proteins. ISCOMs are very immunogenic and stimulate both T- and B-cell response. An equine influenza ISCOM is commercially available and membrane proteins from parainfluenza-3 (PI3), rabies and measles have been combined with ISCOMs.
Liposomes, which are spheres of phospholipid bilayers separated by aqueous compartments , have also been used to entrap antigens; under certain conditions, the l iposome/antigen complex produces higher titres than when the antigen is used alone.
C O N C L U S I O N
Biotechnology is being used increasingly to clarify the pathogenic mechanisms and immune response to microbial disease. This will lead to the production of more effective vaccines in the future. During the last decade, many candidate vaccines have been produced by these techniques, but few are yet in commercial production. Problems of strain stability and poor immunogenicity have occurred and production costs may be higher for synthetic vaccines than for conventional vaccines. Licensing of live genetically-manipulated vaccines may also present additional problems because

791
the use of such vaccines may be construed as the release of genetically-manipulated organisms into the environment. However, given the progress of recent years, biotechnology has a major role to play in the prevention of many animal diseases.
* * *
BIOTECHNOLOGIE ET SCIENCE VÉTÉRINAIRE : PRODUCTION DE VACCINS VÉTÉRINAIRES. - C. Wray et M.J. Woodward.
Résumé: Les auteurs expliquent les principes fondamentaux des manipulations génétiques ainsi que les méthodes de production des vaccins vétérinaires les plus importants. L'atténuation spécifique des microorganismes peut résulter soit des mutations introduites par génie génétique et qui produisent des blocages métaboliques, soit du développement de toxoïdes bactériens efficaces, mais également du réarrangement d'un fragment de gène viral. Les techniques d'ADN recombinant ont été utilisées pour produire des vaccins sous-unitaires et pour concevoir des vaccins multivalents grâce à l'insertion de gènes dans des organismes porteurs tels que poxvirus et salmonelles atténuées. Cet article fait état de l'utilisation de vaccins à base de peptides synthétiques et de vaccins antiidiotypes, ainsi que des problèmes posés par les vaccins antiparasitaires. La biotechnologie devrait apparaître dans le futur comme jouant un rôle majeur dans la prévention de nombreuses maladies animales.
MOTS-CLÉS : ADN Recombinant - Atténuation - Bactéries - Biotechnologie -Manipulation génétique - Parasites - Vaccins sous-unitaires - Virus.
* * *
BIOTECNOLOGÍA Y CIENCIA VETERINARIA: PRODUCCIÓN DE VACUNAS VETERINARIAS. - C. Wray y M.J. Woodward.
Resumen: Los autores explican los principios fundamentales de las manipulaciones genéticas, así como las técnicas de producción de las vacunas veterinarias más importantes. La atenuación especifica de los microorganismos puede resultar de las mutaciones producidas por ingeniería genética, que producen bloqueos metabólicos, del desarrollo de toxoides bacterianos eficaces, o bien del reordenamiento de un segmento de gene vírico. Se han utilizado técnicas de ADN recombinante para producir vacunas de subunidades y también vacunas multivalentes mediante la inserción de genes en organismos portadores, como poxvirus y salmonelas atenuadas. Este artículo comenta el uso de vacunas a base de péptidos sintéticos y vacunas antiidiotipos, así como los problemas planteados por las vacunas antiparasitarias. En el futuro, la biotecnología desempeñará sin duda un papel esencial en la prevención de numerosas enfermedades animales.
PALABRAS CLAVE: ADN recombinante - Atenuación - Bacterias -Biotecnología - Manipulación genética - Parásitos - Vacunas de subunidades -Virus.
* * *

792
REFERENCES
1. AUDIBERT F . , JOLIVET M., CHEDID L., ARNON R . & SELA M. (1982). - Successful immunisation with a totally synthetic diphtheria vaccine. Proc. Natl Acad. Sci. USA, 79, 5042-5046.
2. BITTLE J .L. , HOUGHTON R.A. , ALEXANDER H., SHINNICK T.M., SUTCLIFF J . G . , LERNER R.A. , ROWLANDS D . J . & BROWN F . (1982). - Protection against foot and mouth disease by immunisation with a chemically synthesized peptide predicted from the viral nucleotide sequence. Nature, 1 9 8 , 30-34.
3. BLACK W . J . & FALKOW S. (1987). - Construction and characterisation of Bordetella pertussis toxin mutants. Infect. & Immunity, 5 5 , 2465-2470.
4. BLANCOU J . , KIENY M.P., LATHE R . , LECUCQ J.P., PASTORET P.-P., SOULEBOT J.P. & DESMOTTRE P. (1986). - Oral vaccination of the fox against rabies using a live recombinant vaccinia virus. Nature, 3 3 2 , 373-375.
5. CHANOCK R.M. & MURPHY B . R . (1980). - Use of a temperature-sensitive and cold-adapted mutant virus in immunoprophylaxis of acute respiratory tract disease. Rev. infect. Dis., 2 , 421-432.
6. CLEMENTS J . D . & EL-MORSHIDY S. (1984). - Construction of a potential live oral bivalent vaccine for typhoid fever and cholera-Escherichia coli related diarrhoeas. Infect. & Immunity, 4 6 , 564-569.
7. DALLAS W . S . , GILL D . M . & FALKOW S. (1979). - Cistrons encoding Escherichia coli heat-labile toxin. J. Bact., 139 , 850-858.
8. FAIRWEATHER N.F. , LYNESS V.A. & MASKELL D . J . (1987). - Immunisation of mice against tetanus with fragments of tetanus toxin synthesized in Escherichia coli. Infect. & Immunity, 5 5 , 2541-2545.
9. FLAMAND A., COULON P., PEPIN M., BLANCOU J., ROLLIN P. & PORTNOI D . (1984). -Immunogenic and protective power of avirulent mutants of rabies virus selected with neutralizing monoclonal antibodies. In Modern approaches to vaccines (R.M. Chanock and R .A. Lerner, eds.). Cold Spring Harbour Laboratory, New York, 289-294.
10. FRANCIS M.J., HASTINGS G . Z . , SYRED A.D. , MCGINN B . , BROWN F . & ROWLANDS D.J . (1987). - Non-responsiveness to a foot and mouth disease virus peptide overcome by the addition of foreign helper T-cell determinants. Nature, 3 3 0 , 168-170.
11. GERMANIER R . & FURER E. (1975). - Isolation and characterisation of galE mutant Ty 21a of Salmonella typhi: a candidate strain for a live, oral typhoid vaccine. J. infect. Dis., 1 3 1 , 553-558.
12. GREENBERG H., MIDTHUN K . , WYATT R . , FLORES J., HOSHINO Y., CHANOCK R.M. & KAPIKIAN A. (1984). - Use of reassortant rotaviruses and monoclonal antibodies to make gene-coding assignments and construct rotavirus vaccine candidates. In Modern approaches to vaccines (R.M. Chanock and R.A. Lerner, eds.). Cold Spring Harbor Laboratory, New York, 319-327.
13. GREEN N., ALEXANDER H., OLSON A., ALEXANDER S., SHINNICK T.M., SUTCLIFFE J.G., LERNER R.A. (1982). - Immunogenic structure of the influenza virus haemagglutinin. Cell, 2 8 , 477-487.
14. HOISETH S.K. & STOCKER B . A . D . (1981). - Aromatic dependent Salmonella typhimurium are non-virulent and effective as live vaccines. Nature (London), 2 9 1 , 238-239.
15. KIENY M.P., LATHE R . , DRILLIEN R . , SPEHNER D . , SKORY S., SCHMITT D . , WIKTOR T., KOPROWSKI H. & LECOCQ J.P. (1984). - Expression of rabies virus glycoprotein from a recombinant vaccinia virus. Nature, 3 1 2 , 163-166.
16. KLEID D . G . , YANSURA D . , SMALL B. , DOWBENKO D . , MOORE D .M. , GRUBMAN M.J., MCKERCHER P . D . , MORGAN D . O . , ROBERTSON B.H. & BACHRACH H.L. (1981). -Cloned viral protein vaccine for foot-and-mouth disease: responses in cattle and swine. Science, 2 1 4 , 1125-1129.

793
17. KURODA K . , GRONER A., FRESE K . , DRENCKHAHN D., HAUSER C., ROTT R., DOERFLER W . , KLENK H . O . (1989). - Synthesis of biologically active influenza virus haemagglutinin in insect larva. J. Virol., 63, 1677-1685 .
18. LUCKOW V.A. & SUMMERS M.D. (1988) . - Trends in the development of Baculovirus expression vectors. Biotechnology, 6, 4 7 - 5 5 .
19. MEYER H., DEJA O . , FACH U., GROB W., GRUNERT G . , HARTMANN H., HEINRICH B., HUBRIG T., KINTSCHER M., KOCH H., LINDE K . , LUMM K . , MICHAEL M., OTTA J . , PICK B., PLOTNER J . , SCHIPPEL R., SCHOLL W., SCHULZ W., SCHEIEGER H., STEINBACH G . , UKBANECK D. & ZEPEZAUER V. (1980) . - Bovisaloral "Dessau" orally applicable live vaccine against S. dublin infection of calves. Mh. Vet.Med., 35, 405 -410 .
20. MULLER G . M . , SHAPIRA M. & ARNON R. (1982) . - Anti-influenza response achieved by immunisation with a synthetic conjugate. Proc. Natl Acad. Sci. USA, 79, 5 6 9 - 5 7 3 .
2 1 . MURPHY B.R., SLY D.L., TIERNEY E.L., HOSIER N.T., MASSICOT J .D. , LONDON W.T., CHANOCK R.M., WEBSTER R .G . & HINSHAW V.S. (1982) . - Reassortant virus derived from avian and human influenza A viruses is attenuated and immunogenic in monkeys. Science, 218, 1330-1332 .
22. SADOFF J .C . , BALLOU R . W . , BARON L.S., MAJARÍAN W . R . , BREY R.N., HOCKMEYER W . T . , YOUNG J . F . , CRYZ J . J . , Ou J . , LOWELL G . H . & CHULAY J .D . (1988) . - Oral Salmonella typhimurium vaccine expressing circumsporozoite protein protects against malaria. Science, 240, 2 3 6 - 2 4 0 .
23 . SMITH G.L. , MACKETT M., Moss B. (1983). - Infectious vaccinia virus recombinants that express hepatitis B virus surface antigen. Nature, 302, 4 9 0 - 4 9 5 .
24. SMITH H . W . (1956) . — The use of live vaccines in experimental Salmonella gallinarum infection in chickens with observations on their interference effect. J. Hyg., Camb., 54, 4 1 9 - 4 2 9 .
25. SMITH H . W . (1955) . - The immunization of mice, calves & pigs against Salmonella dublin and Salmonella choleraesuis infections. J. Hyg., Camb., 63, 117-135 .
26. STOCKER B.A.D., HOISETH S.K. & SMITH B.P. (1983) . - Aromatic-dependent Salmonella spp. as live vaccines in mice and calves. Dev. Biol. Standards, 53, 4 7 - 5 4 .
27. TAYLOR J . & PAOLETTI E. (1988) . - Fowlpox virus as a vector in non-avian species. Vaccine, 6, 4 6 6 - 4 6 7 .
28. TAYLOR R.K. , MILLER V.L., FURLONG D.G. & MEKALANOS J . J . (1987) . - Use of phoA gene fusions to identify a Apilus colonization factor coordinately regulated with cholera toxin. Proc. Natl Acad. Sci. USA, 84, 2 8 3 3 - 2 8 3 7 .
29. TITE J . P . , RUSSELL S.M., DOUGAN G . , O'CALLAGHAN D., JONES I., BROWNLEE G . & LIEW F .Y. (1988) . - Antiviral immunity induced by recombinant nucleoprotein of influenza A virus. I. Characteristics and cross reactivity of T-cell responses. J. Immun., 141, 3 9 8 0 - 3 9 8 7 .
30. VALENZUELA P., MEDINA A., RUTTER W . J . , AMMERER G . & HALL B.D. (1982) . -Synthesis and assembly of hepatitis B surface antigen particles in yeast. Nature, 298, 347-350.
* * *
For further reading:
Books ARNON R. (1987) . - Synthetic Vaccines, Vols. 1 and 2 . CRC Press, Inc., Boca Raton, Florida, USA. CHANOCK R.M. & LERNER R.A. (1984) . - Modern approaches to vaccines: molecular and chemical basis of virus virulence and immunogenicity. Cold Spring Harbor Laboratory, USA.

794
GERMANIER R. (ed.) (1984). - Bacterial Vaccines. Academic Press, London. KOHLER H. & LOVERDE P.T. (1988). - Vaccines: new concepts and developments. In Proceedings of the 10th International Convention on Immunology, Buffalo, New York, July 1986. Harlow Longmans Scientific & Technical, New York, 14-17. PEARSON T.W. (ed.) (1986). - Parasite antigens: toward new strategies for vaccines. Marcel Dekker Inc., New York.
Reviews ARNON R., SHAPIRA M. & JACOB C.O. (1983). - Synthetic Vaccines. J. Immunol. Meth., 6 1 , 261-273. BACHRACH H.L. (1985). - New approaches to vaccines. Academic Press Inc., New York. Adv. vet. Sci. comp. Med., 3 0 , 1-38. FIELD B.N. & CHANOCK R.M. (1989). - What biotechnology has to offer vaccine development. Rev. infect. Dis., II. Supplement 3, S519-S523. GAMBLE H.R. & ZARLENGA D.S. (1986). - Biotechnology in the development of vaccines for animal parasites. Vet. Parasit., 2 0 , 237-250. LIEW F . J . (1985). — New aspects of vaccine development. Clin. exp. Immunol., 6 2 , 225-241. LUCKOW V.A. & SUMMERS M.D. (1988). - Trends in the development of Baculovirus expression vectors. Biotechnology, 6 , 47-55. MURRAY P .K. (1989). - Molecular vaccines against animal parasites. Vaccine, 7, 291-299.