blackwell publishing ltd the impact of european occupation...
TRANSCRIPT

Journal of Ecology
2006
94
, 987–1002
© 2006 The AuthorsJournal compilation© 2006 British Ecological Society
Blackwell Publishing Ltd
The impact of European occupation on terrestrial and aquatic ecosystem dynamics in an Australian tropical rain forest
SIMON G. HABERLE, JOHN TIBBY*, SOPHIA DIMITRIADIS† and HENK HEIJNIS‡
Resource Management in Asia-Pacific Program, Research School of Pacific and Asian Studies, Australian National University, Canberra, ACT 0200, Australia,
*
Geographical and Environmental Studies, University of Adelaide, Adelaide, SA 5005, Australia,
†
School of Botany and Zoology, The Australian National University, Canberra, ACT 0200, Australia, and
‡
Australian Nuclear Science and Technology Organization, PMB 1, Menai, NSW 2234, Australia
Summary
1
The long-term impact of changes in land use, fire and climate on species diversity inAustralia are only just beginning to be understood. We combined fine-resolutionpalaeoecological proxies for terrestrial and aquatic ecosystems to investigate theresponses of a tropical rain forest catchment over the last 700 years.
2
Sediment cores were sampled at 1-cm intervals to a depth of 100 cm from LakeEuramoo on the Atherton Tableland of north-east Queensland, dated and analysed forpollen, charcoal, diatom, chironomid and inorganic content.
3
The pollen and charcoal record shows a rapid loss of forest diversity (particularly the long-lived taxa
Agathis
and
Podocarpus
) and increased burning coinciding with the arrival ofEuropean settlers. The aquatic environment is also subject to rapid changes at this time, witha possible increase in pH and subsequent shifts in local algal and insect communities
.
Thisevent was outside the historic range of variability in both rain forest and aquatic communities.
4
The present mosaic of vegetation types is a complex function of environmental changesoperating across a range of spatial and temporal scales: millennial climate change, short-term climatic variations associated with El Niño events and, most significantly, a shift fromindigenous to ‘European’ land-use practices, including clearance and burning activitiesassociated with the timber and farming industry between about
1880 and 1920.
5
The establishment of a World Heritage reserve around the lake catchment and thesuppression of fire over the last 50 years have not yet restored the terrestrial or aquaticecosystem to its pre-European state and are unlikely to, given the current predictions offuture climate change. This supports the notion that ecological and climate thresholdsare not necessarily the same, and that the effects of crossing them are not necessarilyreversible. Retrospective studies of the historic range of variability within small catchmentscan provide an understanding of the limits of natural and human-induced variabilitythat can inform management decisions and resource planning.
Key-words
: alternate states, Atherton Tableland, Australia, charcoal, chironomids,diatoms, disturbance, historic range of variability, pollen, tropical rain forest
Journal of Ecology
(2006)
94
, 987–1002doi: 10.1111/j.1365-2745.2006.01140.x
Introduction
Australia, with its diversity of ecosystems, late colonialsettlement and absence of previous agricultural activity,
offers advantages over other developed nations forstudying the effects of industrial agriculture on thephysical environment. Description of the ecologicalchanges during the period of European settlement ofAustralia has generally fallen to those deriving theirprimary data from historical documents or ‘long-term’ecological data (Kirkpatrick 1999; Bowman
et al
. 2001;
Correspondence: Dr Simon Haberle (tel. + 61 26125 3373;fax + 6126125 4896; e-mail [email protected]).

988
S. G. Haberle
et al.
© 2006 The AuthorsJournal compilation © 2006 British Ecological Society,
Journal of Ecology
,
94
, 987–1002
Griffiths 2001, 2002; Fensham & Fairfax 2002; Lunt 2002).These studies show that Australian ecosystems arehighly dynamic and have undergone significant changethrough most of the last century due to land-use andclimate change. However, there is a rapid loss of qualityand quantity of primary data with increasing antiquityand, with this, a diminishing capacity to understand therate, magnitude and root causes of ecological changeduring the early stages of settlement. The need to under-stand these deeper temporal dynamics of ecosystemchange, which can occur at decadal to millennial scales,is crucial if realistic models of ecosystem response tofuture change are to be developed (Whittaker
et al
. 2005).Although vegetation dynamics on millennial time-
scales are well understood in the Australian wet tropicalrain forest, our understanding of vegetation dynamicsduring the last thousand years is limited to insights fromfragmentary pollen records from the Atherton Tableland(Kershaw 1970, 1971, 1975, 1983). The original sedimentsequences from Lake Euramoo (Kershaw 1970), QuincanCrater (Kershaw 1971), Bromfield Swamp (Kershaw 1975)and Lynch’s Crater (Kershaw 1983) are discontinuous ordisturbed near the surface, with the youngest age beingaround 800
for the upper samples. The contrastbetween recent (European period) and pre-European(> 800
) depends on inclusion of pollen assemblagesfrom surface samples (floating root mats) into thesepollen records. In general, this comparison shows thatan increase in disturbance-related pollen taxa, suchas
Cyclosorus
and
Trema
, and a loss of some long-livedforest pollen taxa, including
Elaeocarpus
and
Agathis
,occurred around the time of European settlement. A1000-year laminated sediment record from Lake Barrineshows little identifiable change in vegetation character-istics over this time period beyond the appearanceof pollen of introduced species of
Solanum
(Walker &Owen 1999; Walker
et al
. 2000), although the large size(
c
. 1 km
2
) and rain forest covered catchment of the lakemay have limited its sensitivity to changes in land useover this time.
European settlement of the rain forest ecosystems ofnorth-eastern Australia was a relatively late event anddid not occur until the end of the 19th century becauseof the remoteness from the then administrative centreof Port Jackson (Sydney) and a perception of the regionas impenetrable and economically barren (Birtles 1982).In addition, early encounters between European explorersand Aboriginal peoples created a perception of theindigenous population as dangerous, although resistanceto encroachment appears to have been brief, possiblybecause disease led to rapidly diminished populations.Little is known about occupation of the rain forest priorto European settlement, although recent archaeologicalresearch indicates that Aboriginal peoples have had thetechnological capacity to subsist in these forests for atleast 7500 years, and to occupy them intensively for thelast 1800 years (Cosgrove 2005). Such people relied onseveral species of fruit trees (
Beilschmeidia
spp.,
Elaeocarpus
spp. and
Aleurites moluccana
), as well as rootstocks of
wild yam (
Dioscorea
spp.), and had mastered the complexprocessing required to obtain substantial quantities ofprotein and carbohydrates from highly toxic cycads (
Cycas
spp.) (Hill & Baird 2003). Fire was used to maintainopen tracks and grassland pockets throughout the rainforest up until the arrival of Europeans in the 1880s(Birtles 1982; Hill
et al
. 1999).Recent, and future, climate change may be a potentially
more significant influence on the extent and survival ofrain forest species and ecosystems (Williams
et al
. 2003).Bioclimatic models of rain forest habitats suggest that,with only 1
°
C warming and 10% decrease in rainfall,the spatial extent of highland rain forest environmentscould decrease by 50%, posing a threat to the survivalof several key vertebrate species (Hilbert
et al
. 2004).However, the widespread cessation of aboriginal burningpractices in the 19th Century and increases in atmos-pheric CO
2
in the late 20th Century favour expansion ofsuch habitat (Hopkins
et al
. 1996; Crowley & Garnett2000; Russell-Smith
et al
. 2004). How these complexinteractions between climate and land-use change willplay out in the next century is critical for the long-termsurvival and management of this ecosystem.
We explore the potential of using high-resolutionmultiproxy palaeoecological data spanning the last 700years to investigate the nature and direction of ecologicalchange in the rain forest environment of the AthertonTableland. We use fossil records of pollen, charcoal,invertebrates and aquatic algae to provide a window intopast ecosystem change, concentrating on the period oftransition marked by the arrival of European settlementin 1881, to address specific questions:
1.
How did European land-use activity influence terres-trial and aquatic ecosystems and lead to the modernrain forest environment?
2.
Did European land-use activity represent an un-precedented event that lay outside the historic range ofvariability?
3.
How has climate change influenced terrestrial andaquatic ecosystems and what impact might it have onthe future of the rain forest environment?
Lake Euramoo is located near the western edge of theWorld Heritage Wet Tropics Bioregion of north-eastQueensland (718 m above sea level, 17
°
10
′
S, 146
°
38
′
E)on the Tertiary uplifted highlands of the AthertonTableland (Fig. 1) and within an area that containsAustralia’s most significant expanse of tropical rainforest. Lake Euramoo is a maar, described as an ovatedouble explosion crater that formed during the LatePleistocene period (Kershaw 1970), with a relativelysmall catchment area of around 4.4 ha and no inflow oroutflow channels. The lake is warm monomictic, with awater depth averaging around 20 m in the northernbasin and 16 m in the southern basin, though there areseasonal fluctuations in water depth of between 2 and3 m (Timms 1976).
989
European impact on Australian rain forest
© 2006 The AuthorsJournal compilation © 2006 British Ecological Society,
Journal of Ecology
,
94
, 987–1002
More than 60% of the estimated annual averagerainfall of 1500 mm falls between January and March(Kershaw 1970), so that the winter is distinctly dry. Thelake lies at the drier end of a steep, east to west rainfallgradient. The dominant source of precipitation is thesouth-east Trades, although occasional north-westerlymonsoonal flows, and associated tropical cyclones, alsobring high but infrequent rainfall events during the australsummer months when the intertropical convergencezone (ITCZ) is at its most southerly extent (Godfred-Spenning & Reason 2002). During El Niño episodes, anorthward movement of the ITCZ and a north-eastwardmigration of the South Pacific convergence zone resultin a significant decrease in summer precipitation(typically 150–300 mm below seasonal average) overthe region (Dai & Wigley 2000). Mean daily maximumand minimum temperatures are around 25.9
°
C and14.4
°
C, respectively, and frosts occur infrequentlyduring the austral winter months at times of weak tradewinds and low cloud cover.
The first European incursions over the escarpmentand into the Atherton Tableland region were associatedwith timber taking (mainly of
Toona australis
‘Red Cedar’and
Agathis palmerstonii
‘Kauri’), beginning in 1881.Pastoral interests, timber and a series of gold rusheshad lured colonists northwards in the 1870s and 1880s,increasing efforts to settle the rain forest region. Estab-lishment of dairy farms to supply the goldfields towardsthe coast, as well as the developing upland settlements ofHerberton and Atherton, exacerbated the rate of forestclearance. Aboriginal populations were dispersed anddecimated during these early decades and soldier settle-ment schemes, from 1905 to 1915 and during the mid20th century, increased land clearance and populationpressure in the region (Birtles 1988). The designation ofremnant patches of rain forest, such as Lake Barrine andLake Eacham, as National Parks in the 1930s provided
some protection from logging. While it is estimated thatonly around 14% of the rain forest had been clearedby 1985 (Winter
et al
. 1987), the selective extraction ofeconomically valuable species had a far wider impact.
The terrestrial vegetation surrounding Lake Euramoois a remnant of moist submontane rain forest surroundedby previously cleared land that, within the last 50 years, haseither been planted with endemic Hoop Pine (
Araucariacunninghamii
) and exotic conifers or has undergonesecondary colonization towards re-establishing rainforest. Over 100 species of rain forest tree and shrub havebeen recorded within 100 m of Lake Euramoo (A. P.Kershaw personal communication). Typical moist sub-montane rain forest species found near Lake Euramooinclude members of the Araliaceae (e.g.
Polysciasaustraliana
,
Schefflera actinophylla
), Araucariaceae (e.g.
Agathis robusta
), Moraceae (e.g.
Ficus
sp.), Elaeocar-paceae (e.g.
Elaeocarpus grandis
), Euphorbiaceae (e.g.
Aleurites moluccana
,
Macaranga
spp.), Myrtaceae (e.g.
Austromyrtus
spp.,
Eugenia cormiflora
) and Rubiaceae(e.g.
Flindersia brayleyana
,
Euodia bonwickii
). The nearby(< 1 km) drier vegetation communities include tallopen forest (sclerophyll woodland) dominated by
Casuarina
spp. and
Eucalyptus
spp. At higher altitude,cool lower montane rain forest is increasingly dominatedby members of families such as the Cunoniaceae (e.g.
Caldcluvia australiensis
,
Schizomeria whitei
), Podo-carpaceae (e.g.
Podocarpus
spp.,
Prumnopitys
spp.) andRhamnaceae (e.g.
Alphitonia
spp.).Weedy species, of which the shrub
Lantana camara
isthe most common, frequently invade roadsides anddisturbed areas. The native vegetation around the lakemargin consists of conspicuous zones of aquatic plantcommunities governed by water depth and seasonalfluctuations in water level (Kershaw 1978). Rain forestlianas (e.g.
Parsonsia
spp.) intertwine with the tall swampgrass
Phragmites australis
around the shallow lake edge.
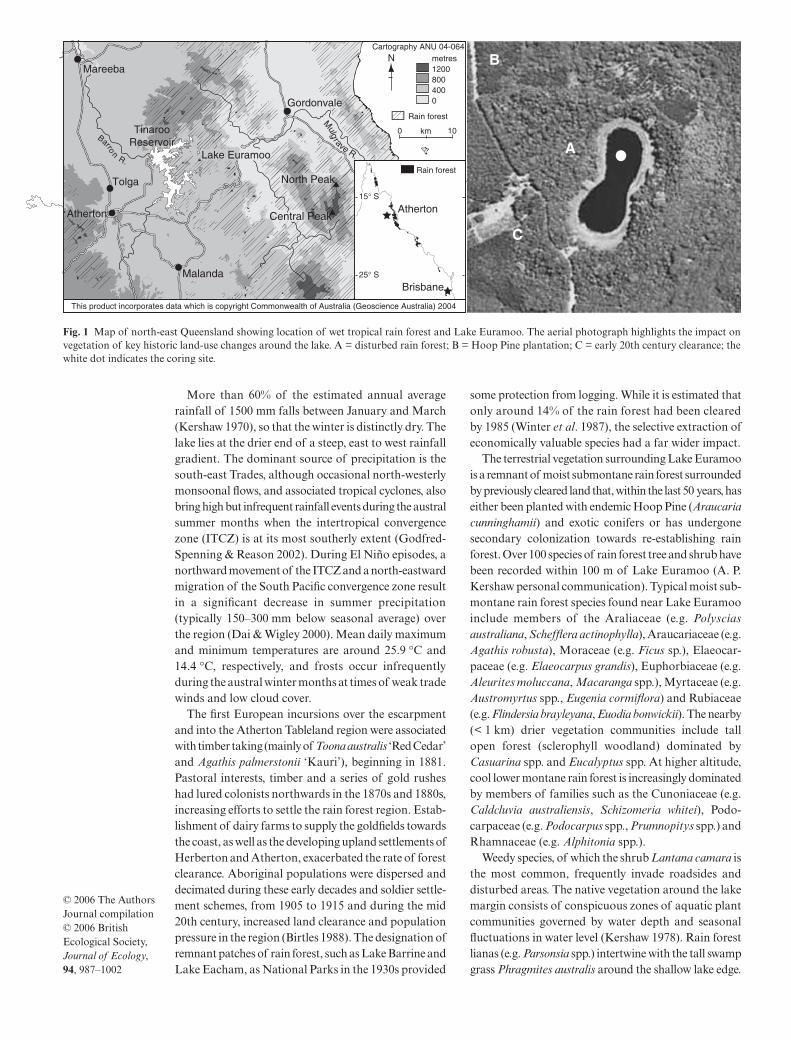
Fig. 1 Map of north-east Queensland showing location of wet tropical rain forest and Lake Euramoo. The aerial photograph highlights the impact onvegetation of key historic land-use changes around the lake. A = disturbed rain forest; B = Hoop Pine plantation; C = early 20th century clearance; thewhite dot indicates the coring site.

990
S. G. Haberle
et al.
© 2006 The AuthorsJournal compilation © 2006 British Ecological Society,
Journal of Ecology
,
94
, 987–1002
Hibiscus
spp. and
Ludwigia
spp. become more commonin the deeper (but still less than 1 m water depth) swampmargin (i.e. outside the influence of canopy shade andliana growth). A zone of rooted emergent aquatics andfloating vegetation mats is characterized by
Cyclosorusgongylodes
and
Oenanthe
spp., with
Blechnum
spp. and
Eleocharis
spp. becoming common on less stable floatingmats. As water depth increases to greater than 1 m, thereis a transition from rooted emergent aquatics to floatingaquatics, mainly
Nymphoides
spp., which can occur asmuch as 30 m from the lake shore.
Methods
Fieldwork was carried out in April–May 1999. Theuppermost 100 cm of sediment was obtained, using aclear plastic piston corer, from the centre of the northernbasin of Lake Euramoo, where the water depth measured1600 cm. This method allows for undisturbed recovery ofthe mud–water interface. The two sections of core, whichmeasured from 0 to 68 cm depth and 69 to 100 cm depth,were divided into contiguous 1-cm intervals on-site.
Samples were analysed for loss-on-ignition (%LOI)by combusting dried sediment at 500
°
C for 5 hours.Chronological control of the record was providedusing a combination of
14
C,
210
Pb and first appearanceof exotic pollen types (Tables 1 and 2). Conventional
14
C dates were obtained on two bulk organic sedimentsamples at 95 cm and 190 cm depth (Table 1), the latterfrom a longer core (Haberle 2005). Fourteen sedimentsamples from the upper 25 cm of the core were driedovernight at 75
°
C, finely ground, and analysed for
210
Pbby ANSTO (Australian Nuclear Science and Technology
Organisation, Sydney, Australia). Sedimentation rateswere derived from the analysis of
210
Pb activity usingthe constant initial concentration (CIC) model (Appleby& Oldfield 1983). Constant rate of supply (CRS) modelcalculations were not derived, because a full inventoryof excess
210
Pb could not be calculated as excess
210
Pbwas still present in the bottom sample.
,
The terrestrial and aquatic flora is represented by thepollen and spore assemblage preserved in the lakesediments. Pollen and spore analysis followed standardprocedures (Bennett & Willis 2001). Pollen counts areexpressed as percentages of the total pollen sum (excludingpollen of aquatic vascular plants and spores), whichreaches a minimum of 200 in all samples. The timingof European settlement in the core was assessed fromchanges in pollen representing introduced weeds,including
Lantana
,
Mimosa
,
Plantago
,
Pinus
, Solanaceaeand
Rumex
(based on Queensland Herbarium records).Concentrations of pollen, spores and microscopic charcoal(
c
. 10–125
µ
m) were calculated by relative abundancewith respect to a known concentration of
Lycopodium
.Concentrations of larger particles of charcoal (> 125
µ
m)were determined as an indicator of fires occurringrelatively close to the site (Clark & Royall 1995;Millspaugh & Whitlock 1995).
Diatoms are unicellular organisms characterized bytheir siliceous cell walls and are particularly sensitiveto physical, chemical or biological changes in aquatichabitats (Battarbee
et al
. 2001). Samples for diatomanalysis were taken at 2–4 cm intervals down the core
Table 1 Lake Euramoo: radiocarbon dates and calibrated ages (cal year = years before 1950) determined using the Calib 4.0software (Stuiver et al. 1998)
Sample material
Depth from sediment surface (cm)
Laboratory code
δ13C (‰)
Percentage modern carbon
Conventional 14C age Calibrated age
range cal year (% prob. 1σ)pMC 1σ (year ) 1σ
Organic detritus 95–96 OZE682 92.90 0.54 590 40 642–592 (71.9%)563–549 (28.1%)
Organic detritus 190–191 OZE683 84.36 0.36 1370 40 1303–1272 (100%)
Table 2 Time markers used to construct the age-depth model for sedimentation and sedimentation accumulation rates at LakeEuramoo
Depth (cm)Age estimate (year ) Age estimate based upon
Sediment accumulation rate (cm year−1)
0–1 1999 Assumed to represent present 0.3212–13 c. 1960 Appearance of weed pollen: Pinus and Plantago 0.2624–25 c. 1912 Appearance of weed pollen: Mimosa 0.4840–41 c. 1880s Appearance of weed pollen: Lantana 0.1345–46 1850−1920 210Pb age determination 0.1095–96 1340 Calibrated AMS 14C age determination 0.13190–191 660 Calibrated AMS 14C age determination 0.15

991European impact on Australian rain forest
© 2006 The AuthorsJournal compilation © 2006 British Ecological Society, Journal of Ecology, 94, 987–1002
from the same depth as the samples for pollen analysis(with the exception of 1 and 3 cm, which were not analysedfor diatoms) and prepared using a modified versionof the technique outlined in Renberg (1990). Diatomswere identified at 1000× magnification, using a ZeissAxioscope with differential interference contrast optics,with reference to published floras (in particular, Krammer& Lange-Bertalot 1986; 1988; 1991; Vyverman 1991;and references outlined in Tibby 2001).
Diatom data are expressed as relative abundances ofthe diatom sum where all taxa, with the exception ofthe weakly silicified taxon Rhizoselenia sp. 1, abundantonly in the uppermost sample, are included in the sum.Modified terms from Tibby (2001) were used to describethe relative abundance of diatom valves: trace (< 1%relative abundance), minor (1–5%), important (5–20%),subdominant (20–50%), and dominant (> 50%).
Chironomidae are abundant in fossil records and havebeen used to trace the palaeoproductivity of lakes, toassess anthropogenic eutrophication and acidification,to monitor the impact of salinity fluctuations on benthicassemblages and, due to their sensitivity to temperature,in palaeoclimate reconstruction (Rossaro 1991; Walkeret al. 1995; Dimitriadis & Cranston 2001). Sedimentsamples of 2 cm3 were extracted at c. 4 cm intervals downthe core; given the limited sediment available for analysis,these do not correspond to the samples used for pollenand diatom analysis. Sample treatment followedDimitriadis & Cranston (2001).
To assess whether ordination techniques, which implicitlyassume linear or Gaussian responses, should be applied,detrended correspondence analysis was used to determinethe gradient length of the biological data sets (Birks1998). All data sets had gradient lengths < 2.0, with thepollen and chironomid data sets < 1.0. Hence lineartechniques (principal components analysis, PCA, andredundancy analysis, RDA) are appropriate. PCA is usedto reduce the fossil biotic data to a two-dimensionalplot and the resulting data set is displayed as a biplotfor samples and taxa (Birks & Gordon 1985). PCA wasperformed using only those taxa whose percentagevalues exceeded 5% at least once. The data for each fossilgroup were square-root transformed to reduce theinfluence of very abundant taxa. Redundancy analysiswas used to assess the effect of landscape disturbanceand biomass burning (inferred from the inorganic andcharcoal fractions, respectively) on all data sets. Theinfluence of vegetation change on the lake ecosystem wasassessed by determining the explanatory power of axes1 and 2 of the pollen data on the diatom and chironomiddata sets. The statistical significance of those variableswas determined using 9999 Monte Carlo permutationsof the ordination solution. RDA was conducted where
both pollen and diatom data were determined for eachdepth. However, for analysis of the chironomid dataset, where adjacent data were not available, pollen datafrom the next available down core sample were utilized.All numerical analyses, with the exception of RDA,have been implemented within PSIMPOLL, a programfor plotting pollen data, developed by Bennett (1994).Numerical zonation employed optimal splitting by sum-of-squares analysis to partition the stratigraphicallyconstrained fossil assemblage data into significantlydifferent zones (Bennett 1996). RDA was implemented in for Windows 4.5 (ter Braak & Smilauer 2002).
Results
Figure 2(a) illustrates the two lines of best fit throughthe excess 210Pb activity down core. Application of theCIC model to these alternatives provides a range ofpotential ages down core, estimating the lowest dated210Pb sample (46 cm) to be between 1850 and 1920. The sequential first appearance of exotic pollen types(Table 2) provides an additional age control on thesedimentation rate and these data have been includedin construction of the age-depth model (Fig. 2b). The
Fig. 2 Lake Euramoo Core B age-depth model. (a) Plot ofexcess Pb-210 activity vs. depth with two possible lines of bestfit (CIC model). (b) Age-depth model based on first appearanceof exotic pollen, Pb-210 and C-14 ages for the top 100 cm ofsediment (year ).
992S. G. Haberle et al.
© 2006 The AuthorsJournal compilation © 2006 British Ecological Society, Journal of Ecology, 94, 987–1002
ages derived for each sample are likely to have greateruncertainties than represented by the envelope drawnaround the preferred age-depth model due to the lowresolution of age analysis points (Telford et al. 2004). Thefinal age-depth model adopted for this study (Fig. 2b,solid line) is derived by linear interpolation between thekey markers outlined in Table 2.
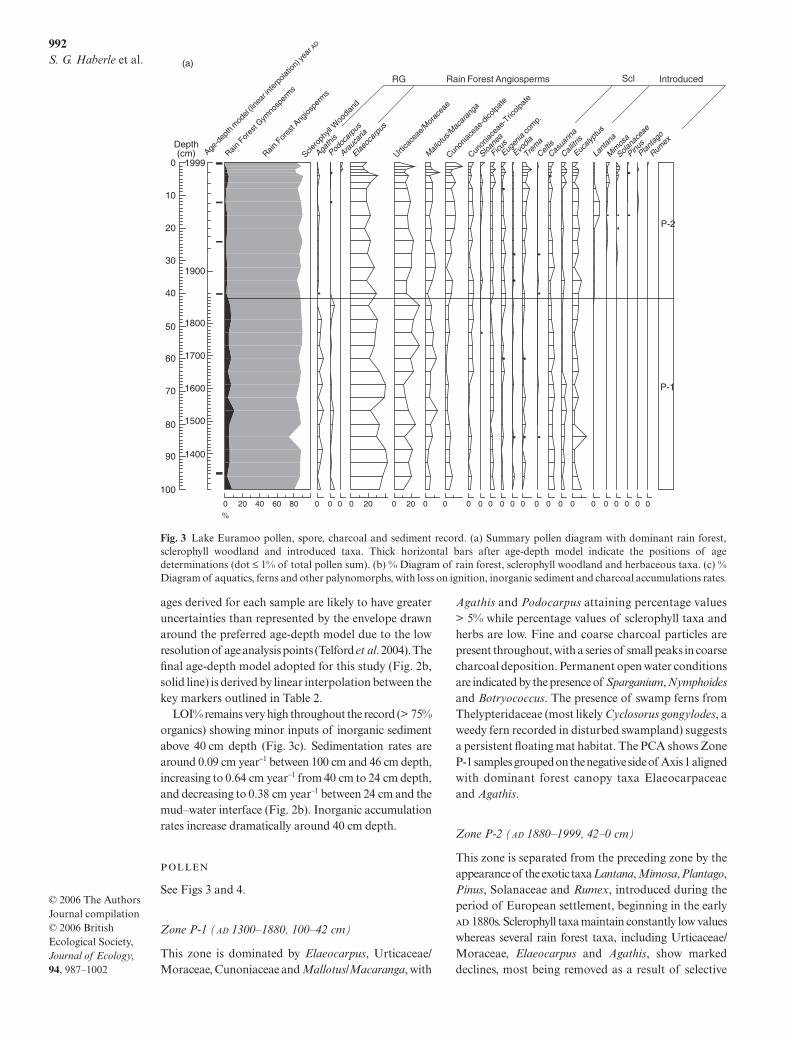
LOI% remains very high throughout the record (> 75%organics) showing minor inputs of inorganic sedimentabove 40 cm depth (Fig. 3c). Sedimentation rates arearound 0.09 cm year−1 between 100 cm and 46 cm depth,increasing to 0.64 cm year−1 from 40 cm to 24 cm depth,and decreasing to 0.38 cm year−1 between 24 cm and themud–water interface (Fig. 2b). Inorganic accumulationrates increase dramatically around 40 cm depth.
See Figs 3 and 4.
Zone P-1 (AD 1300–1880, 100–42 cm)
This zone is dominated by Elaeocarpus, Urticaceae/Moraceae, Cunoniaceae and Mallotus/Macaranga, with
Agathis and Podocarpus attaining percentage values> 5% while percentage values of sclerophyll taxa andherbs are low. Fine and coarse charcoal particles arepresent throughout, with a series of small peaks in coarsecharcoal deposition. Permanent open water conditionsare indicated by the presence of Sparganium, Nymphoidesand Botryococcus. The presence of swamp ferns fromThelypteridaceae (most likely Cyclosorus gongylodes, aweedy fern recorded in disturbed swampland) suggestsa persistent floating mat habitat. The PCA shows ZoneP-1 samples grouped on the negative side of Axis 1 alignedwith dominant forest canopy taxa Elaeocarpaceaeand Agathis.
Zone P-2 (AD 1880–1999, 42–0 cm)
This zone is separated from the preceding zone by theappearance of the exotic taxa Lantana, Mimosa, Plantago,Pinus, Solanaceae and Rumex, introduced during theperiod of European settlement, beginning in the early 1880s. Sclerophyll taxa maintain constantly low valueswhereas several rain forest taxa, including Urticaceae/Moraceae, Elaeocarpus and Agathis, show markeddeclines, most being removed as a result of selective
Fig. 3 Lake Euramoo pollen, spore, charcoal and sediment record. (a) Summary pollen diagram with dominant rain forest,sclerophyll woodland and introduced taxa. Thick horizontal bars after age-depth model indicate the positions of agedeterminations (dot ≤ 1% of total pollen sum). (b) % Diagram of rain forest, sclerophyll woodland and herbaceous taxa. (c) %Diagram of aquatics, ferns and other palynomorphs, with loss on ignition, inorganic sediment and charcoal accumulations rates.

993European impact on Australian rain forest
© 2006 The AuthorsJournal compilation © 2006 British Ecological Society, Journal of Ecology, 94, 987–1002
logging during the late 19th and early 20th centuries(Birtles 1988). Increases are recorded in Rutaceae/Araliaceae, Trema, Cunoniaceae, Sloanea and Eugenia.Charcoal particle accumulation rates attain high values,although there is little apparent impact on the total rainforest taxa percentage of the pollen sum. The PCAshows Zone P-2 samples grouped on the positive sideof Axis 1, aligned with disturbance taxa such as Trema,Lantana and grasses.
See Figs 5 and 6.
Zone D-1 (AD 1300–1720, 100–62 cm)
Zone D-1 is characterized by high relative abundances ofEunotia pectinalis. Aulacoseira sp. 1 is important lowerin this zone but, after peaking to dominance at 80 cm,it is replaced by Eunotia aff. implicata and Gomphonemagracile, while Cyclotella aff. glomerata has a notablepeak at 76–72 cm. The separation of Zone D-1 samplesto the positive side of axis 1 on the PCA is driven by thedominance of Eunotia pectinalis.
Zone D-2 (AD 1720–1850, 62–46 cm)
In D-2, Aulacoseira sp. 1 replaces Eunotia pectinalis as thedominant diatom taxon in the Euramoo record. Gomph-onema gracile is more abundant in this zone, though itdoes not reach subdominant proportions. The combinedpresence of Staurosira construens f. venter, Staurosirellaelliptica and Staurosirella pinnata at minor to importantrepresentation in the 56-cm sample is notable. In theupper samples in this zone, Aulacoseira ambigua occursabove 2% for the first time in the record. The shift in samplegrouping to the negative side of axis 1 in the PCA isdriven by the shift to dominance of Aulacoseira sp. Thestrong negative values on axis 2 reflect peak values beingreached in Gomphonema gracile and Eunotia aff. implicata.
Zone D-3 (AD 1850–1950, 46–12 cm)
Zone D-3 is characterized by continued reduction ofEunotia pectinalis, although, unlike D-1 and D-2, thereare no corresponding changes in Aulacoseira sp. 1.Cyclotella aff. glomerata exhibits a variable, gradualincrease through the zone. Gomphonema gracile and G.parvulum are important taxa, which reach their greatest
Fig. 3 continued

994S. G. Haberle et al.
© 2006 The AuthorsJournal compilation © 2006 British Ecological Society, Journal of Ecology, 94, 987–1002
abundance in this zone. There is a minor shift in sampledistribution in the PCA towards the negative side ofaxis 1, where a range of minor taxa, including Encyonemagracilis and Gomphonema parvulum, reach maximumpercentage values.
Zone D-4 (AD 1950–1999, 12–0 cm)
Aulacoseira sp. 1 has a higher mean relative abun-dance in zone D-4 than in other zones. Aulacoseiraambigua and Cyclotella aff. glomerata are also at
Fig. 3 continued
Fig. 4 Principal components analysis of the pollen assemblage with eigenvalue axis 1 = 36.1% and axis 2 = 12.6%. (a) Pollen taxa(includes only taxa with > 5% representation: data have undergone square-root transformation). (b) Individual samples, withsymbols distinguishing those belonging to pollen zones P-1 and P-2.

995European impact on Australian rain forest
© 2006 The AuthorsJournal compilation © 2006 British Ecological Society, Journal of Ecology, 94, 987–1002
their most abundant in this zone, with the latterpeaking in the youngest two samples in the record.In this zone, Eunotia pectinalis is all but eliminatedfrom the record, as are Gomphonema gracile and G.
parvulum. Peak values in Cyclotella aff. glomerata,Aulacoseira ambigua and Fragilaria sp. shift thesample grouping in the PCA towards the positiveside of axis 1.
Fig. 5 Lake Euramoo summary diatom record. Conventions as in Fig. 3.
Fig. 6 Principal components analysis of the diatom assemblage with eigenvalue axis 1 = 56.7% and axis 2 = 18.5%. (a) Diatom taxa(includes only taxa with > 5% representation: data have undergone square-root transformation). (b) Samples with symbolsdistinguishing those belonging to diatom zones D-1 to D-4.

996S. G. Haberle et al.
© 2006 The AuthorsJournal compilation © 2006 British Ecological Society, Journal of Ecology, 94, 987–1002
See Figs 7 and 8.
Zone C-1 (c. AD 1300–1955, 104–14 cm)
Zone C-1 is characterized by consistent dominanceof Monopelopia sp., Procladius paludicola, Chironomussp. 1 and Dicrotendipes jobetus. Djamabatista sp., Chi-ronomus sp. 2, Microtendipes umbrosus, Polypedilumleei, P. nr serosum and Zavreliella marmorata are foundonly in this zone, and only at low levels. Parakiefferiellasp. percentage values begin to increase towards the topof this zone (after 1920). The separation of ZoneC-1 samples to the negative side of axis 1 in the PCAis driven by the dominant taxa, particularly Dicroten-dipes jobetus.
Zone C-2 (AD 1955–1999, 14–0 cm)
Parakiefferiella sp. percentage values continue to increasein this zone, becoming dominant by the surface sample.Monopelopia sp., Procladius paludicola and Chironomussp. 1 remain common taxa, whereas Dicrotendipes jobetusdecreases. Xenochironomus sp. also increases to peakvalues. Polypedilum K1 is present only in this zone. Theshift of sample grouping to the positive side of axis 1 inthe PCA is due to the increased dominance of Paraki-efferiella sp. and Xenochironomus sp. at the expense ofother common chironomid taxa.
Interpretation and discussion
Historical and palaeoecological analysis of ecosystemchange attempts to extend the range of spatial and
Fig. 7 Lake Euramoo summary chironomid record.

997European impact on Australian rain forest
© 2006 The AuthorsJournal compilation © 2006 British Ecological Society, Journal of Ecology, 94, 987–1002
temporal perspectives, beyond those attainable with onlyecological data, on the environmental drivers and dis-turbance processes that interact to control patterns andchange in biotic assemblages. A single coring locality(Lake Euramoo) provides a window into both theterrestrial environment, through pollen and charcoal, andthe aquatic environment, through pollen, chironomidsand diatoms. Each assemblage reflects environmentalchanges at different spatial scales and can provide agreater understanding of the pre-European and Europeanhistory of natural and human disturbance, environmentalchange and catchment ecosystem dynamics.
Lake Euramoo has a surface area of approximately 4.4 ha,which should receive pollen from within a < 100 m to10 km radius (assuming that the general pollen dispersalmodel of Jacobson & Bradshaw 1981 is applicable in thetropics). The dominant contribution will be from thelocal (< 100 m) to extra-local (100–1000 m) taxa and, asthese will be mainly those contained within the crater,the data will provide a detailed history of vegetationdynamics within the catchment. The pollen recordshows that the composition of rain forest in this regionhas not been stable over the last 700 years (Figs 3 and4). The occurrence between the 13th and 19th century,of only minor fluctuations in the dominant rain foresttaxa Elaeocarpus, Mallotus/Macaranga and Urticaceae/Moraceae appears to reflect a stable rain forest environ-ment. Periodic fire events during the early part of therecord (13th to 16th century) may reflect a potentiallyhigher level of disturbance in the rain forest or at therain forest-sclerophyll margin, but fire events seem tohave been suppressed at both regional and local levelbetween the 17th and 19th century. The most significantchange in the pollen record occurs at 1880, andincludes the reduction of key taxa in the pollen record,such as Agathis and Podocarpus (both exploited for thetimber industry during the late 19th and early 20thcentury), the sequential appearance of exotic taxaLantana, Mimosa, Plantago, Pinus, Solanaceae and
Rumex, and increased charcoal accumulation. Theserapid changes coincide with the onset of Europeanoccupation and are most likely the result of clearanceand burning activities associated with the timber andfarming industry between about 1880 and 1920. Theirimpact appears to have persisted through to the present,even after the cessation of logging and clearance activityin the 1950s. While there are no significant zonal changesidentified in the pollen record of the post-Europeansettlement period, invasions of exotic taxa continue tothe present.
The ability to reconstruct precisely the aquatic historyof Lake Euramoo is hampered by the absence ofdedicated biotic data sets from northern Australian lakes,and a lack of species-level identifications. Despite this,much information can still be gleaned for key taxa. Inparticular, we infer general water quality shifts fromsouthern Australian diatom and chironomid data sets(e.g. Dimitriadis & Cranston 2001; Tibby et al. 2003)and changes in the nature of the physical lacustrineenvironment from habitat preferences of key taxa (e.g.plant associated vs. planktonic).
The diatom record exhibits the greatest variability ofthe proxy data sets, with the major change being theswitch in dominance between Eunotia pectinalis andAulacoseira sp. 1 that occurs between 64 and 60 cm, clearlypredating European arrival (see Fig. 2b and below).However, a further, longer record from Lake Euramooindicates that levels of these two taxa were variable overat least the thousand years before European arrival(J. Tibby, unpublished data). Hence, although loweredvalues of E. pectinalis predate European influence, theearly fall might merely reflect natural variability, whichis compounded by suppression as a result of Europeanarrival.
The next substantial change in the diatom recordoccurs between 42 and 46 cm (i.e. the zone D-2 and D-3boundary). Given that the pollen of introduced Lantanafirst appears at 40 cm and pollen of introduced taxa may
Fig. 8 Principal components analysis of the chironomid assemblage with eigenvalue axis 1 = 49.8% and axis 2 = 16.9%.(a) Chironomid taxa. (b) Samples belonging to pollen zones C-1 and C-2.

998S. G. Haberle et al.
© 2006 The AuthorsJournal compilation © 2006 British Ecological Society, Journal of Ecology, 94, 987–1002
post-date the first evidence of European-induced waterquality change (Leahy et al. 2005), it seems likely thatthis change is due to European impact. Indeed, the firstdefinitive evidence of European arrival appears to beregistered in the sample immediately below the zoneboundary (46 cm). Staurosira construens f. venter, Stauro-sirella elliptica and Staurosirella pinnata exhibit relativelyminor but distinctive peaks at this depth. These taxamake up 17% of the diatom sum at this point, whereasthey are always less than 0.5% of the count for all othersamples.
These taxa are associated predominantly with alkalinewaters (Bennion 1995; Sayer 2001), often with highturbidity (Gell et al. 2002). In south-eastern Australiathey have pH optima of 7.7 (Staurosira construens f.venter), 8.2 (Staurosirella pinnata) and 8.6 (Staurosirellaelliptica) (Tibby et al. 2003 and unpublished data). Thepreference for alkaline waters contrasts with theoptimum for the dominant pre-European diatom Eunotiapectinalis (6.9) and suggests an increase in pH.
In south-eastern Australia, increased pH has beenobserved in several riverine aquatic systems shortly afterEuropean arrival (Reid et al. 2002; Tibby et al. 2003;Gell et al. 2005). This increase is attributed in part to anincreasing load of base cations due to catchment erosion(Tibby et al. 2003), a process that may in itself be causedby fires (Korhola et al. 1996). A similar phenomenonmay have occurred at Lake Euramoo. Certainly there isa general association between increased macrocharcoalin the post-European phase and the reductions in Eunotiapectinalis. However, as the redundancy analysis suggests(Table 3), the effects of burning on lake pH are likely tobe indirect. The sustained reduction of E. pectinalis fromthe record suggests that elevated pH may have persistedas a result of deforestation and its tendency to raise soilpH in the region (Rasiah et al. 2004).
Associated with the decline of Eunotia pectinalis arezone D-2 increases in Aulacoseira ambigua and Gom-phonema parvulum, which are competitive in elevatednutrient environments. The abundance of these taxaindicates a notable enrichment of the lake waters in thepost-European phase. The rise in the aquatic micro-phyte Sparganium during this phase might also relate tonutrient enrichment in the lake margin habitats. Themost likely source of these nutrients would be localizedcatchment erosion resulting from the activities of timbercutters and, certainly, increases in the delivery ofinorganic material (Fig. 3c).
The diatom and chironomid communities show anapparently strong response to the cessation of loggingand clearance activities in the region, with a number ofsignificant taxa changes at the top of zone D-1 (Fig. 5)and zone C-1 (Fig. 7). The elimination of the diatomsEunotia aff. implicata, Fragilaria tenera and Gomphonemagracile and the nutrient rich indicator Gomphonemaparvulum, result largely from increases in Aulacoseira sp. 1and Cyclotella aff. glomerata. The rise in the chironomidParakiefferiella sp. and Xenochironomus sp. and thereduction in Dicrotendipes jobetus and Tanytarsus nr
fuscithorax at the same time represent the most signifi-cant shift in chironomid assemblages over the preceding600 years. Xenochironomus sp. is known to be closelyassociated with decaying wood and these general trendsmay indicate a shift towards expanding littoral habitats.Unfortunately the ambiguous taxonomy and lack ofecological data for these taxa makes the precise reasonfor these changes elusive.
,
Did European land-use activity represent an un-precedented event that lay outside the historic rangeof variability? The pre-contact pollen, diatom andchironomid records indicate a degree of variability oncentennial time-scales, with, for example, periods ofdominance by the acidic periphytic diatom taxon Eunotiapectinalis in the 13th, 14th and 17th centuries interspersedwith periods of dominance by Aulacoseira sp. 1 in the 15thand 18th centuries, and similar amplitude variation inthe pollen and chironomid records. The rain forest anddiatom communities appear to rapidly register the firsteffect of European arrival, with the boundary betweenzones D-2 and D-3 and P-1 and P-2 located very closeto the known historical first European occupation event.
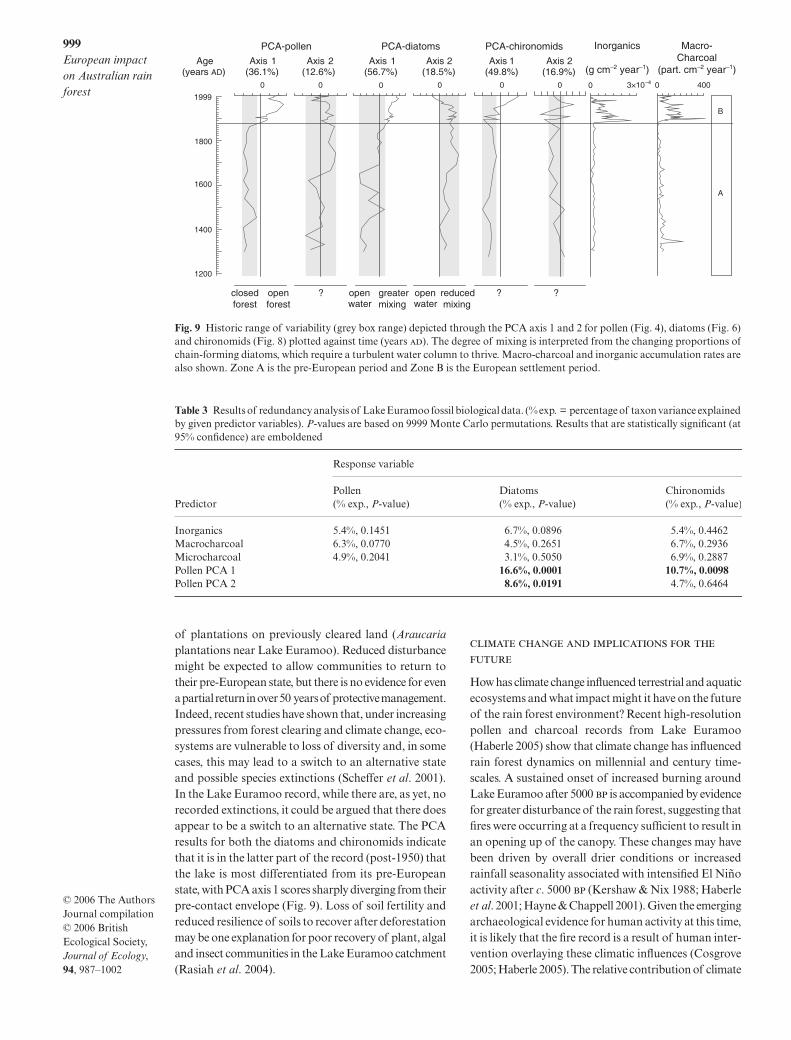
An historic range of variability perspective (Morganet al. 1994) is one way of examining the propositionthat the impact of European land-use on rain forestecosystems lies outside the known range of variabilityin terms of extent and severity. In Fig. 9 we use the firstand second axis of the PCA as a proxy for biotic vari-ability through time. All 1st axis PCA curves indicate thatunprecedented changes do occur during the Europeanperiod although these are neither unidirectional norsynchronous. The strong and sustained shift in rain forestand algae community composition beginning at the timeof European settlement implies that the communityresponse was rapid and related, at least in part, to burningand sediment mobilization in and around the catchment.However, not all aspects of the lake catchment ecosystemwere as sensitive: the first axis of the chironomid recordand all second axes show a much delayed or benignresponse to European settlement. Redundancy analysis ofthese data sets shows that catchment–lake interactions areimplicated in aquatic community perturbation (Table 3).Major shifts in vegetation (as summarized on PCA axis1 of the pollen data) explain a significant (P < 0.05) andsubstantial amount of variation in both the diatom (16.6%)and chironomid (10.7%) data. A stronger influence ofvegetation change on the algal community is implicatedas axis 2 of the pollen PCA also explains a significantamount of diatom variance (8.6%, P = 0.02). This may bedue to the more direct relationship between diatoms andwater quality (Brooks et al. 2001), although the sampleoffset between the two indicators may play a role.
The Lake Euramoo catchment has been protectedfrom clearance and burning activity since the 1950s dueto designation of National Parks and the establishment
999European impact on Australian rain forest
© 2006 The AuthorsJournal compilation © 2006 British Ecological Society, Journal of Ecology, 94, 987–1002
of plantations on previously cleared land (Araucariaplantations near Lake Euramoo). Reduced disturbancemight be expected to allow communities to return totheir pre-European state, but there is no evidence for evena partial return in over 50 years of protective management.Indeed, recent studies have shown that, under increasingpressures from forest clearing and climate change, eco-systems are vulnerable to loss of diversity and, in somecases, this may lead to a switch to an alternative stateand possible species extinctions (Scheffer et al. 2001).In the Lake Euramoo record, while there are, as yet, norecorded extinctions, it could be argued that there doesappear to be a switch to an alternative state. The PCAresults for both the diatoms and chironomids indicatethat it is in the latter part of the record (post-1950) thatthe lake is most differentiated from its pre-Europeanstate, with PCA axis 1 scores sharply diverging from theirpre-contact envelope (Fig. 9). Loss of soil fertility andreduced resilience of soils to recover after deforestationmay be one explanation for poor recovery of plant, algaland insect communities in the Lake Euramoo catchment(Rasiah et al. 2004).
How has climate change influenced terrestrial and aquaticecosystems and what impact might it have on the futureof the rain forest environment? Recent high-resolutionpollen and charcoal records from Lake Euramoo(Haberle 2005) show that climate change has influencedrain forest dynamics on millennial and century time-scales. A sustained onset of increased burning aroundLake Euramoo after 5000 is accompanied by evidencefor greater disturbance of the rain forest, suggesting thatfires were occurring at a frequency sufficient to result inan opening up of the canopy. These changes may havebeen driven by overall drier conditions or increasedrainfall seasonality associated with intensified El Niñoactivity after c. 5000 (Kershaw & Nix 1988; Haberleet al. 2001; Hayne & Chappell 2001). Given the emergingarchaeological evidence for human activity at this time,it is likely that the fire record is a result of human inter-vention overlaying these climatic influences (Cosgrove2005; Haberle 2005). The relative contribution of climate
Fig. 9 Historic range of variability (grey box range) depicted through the PCA axis 1 and 2 for pollen (Fig. 4), diatoms (Fig. 6)and chironomids (Fig. 8) plotted against time (years ). The degree of mixing is interpreted from the changing proportions ofchain-forming diatoms, which require a turbulent water column to thrive. Macro-charcoal and inorganic accumulation rates arealso shown. Zone A is the pre-European period and Zone B is the European settlement period.
Table 3 Results of redundancy analysis of Lake Euramoo fossil biological data. (% exp. = percentage of taxon variance explainedby given predictor variables). P-values are based on 9999 Monte Carlo permutations. Results that are statistically significant (at95% confidence) are emboldened
Predictor
Response variable
Pollen (% exp., P-value)
Diatoms (% exp., P-value)
Chironomids (% exp., P-value)
Inorganics 5.4%, 0.1451 6.7%, 0.0896 5.4%, 0.4462Macrocharcoal 6.3%, 0.0770 4.5%, 0.2651 6.7%, 0.2936Microcharcoal 4.9%, 0.2041 3.1%, 0.5050 6.9%, 0.2887Pollen PCA 1 16.6%, 0.0001 10.7%, 0.0098Pollen PCA 2 8.6%, 0.0191 4.7%, 0.6464

1000S. G. Haberle et al.
© 2006 The AuthorsJournal compilation © 2006 British Ecological Society, Journal of Ecology, 94, 987–1002
change and human activity to these changes remains acomplex question that ultimately may not be resolved ifwe assume an interdependence of human response toenvironmental conditions (Haberle & David 2004).
On shorter time-scales, and during the last millennium,our understanding of north-east Australian climatechange is limited to high resolution data from coralgrowth records from the Great Barrier Reef, whereHendy et al. (2002) infer persistent elevated sea surfacetemperatures in the SW Pacific throughout the LittleIce Age (17th–19th centuries) compared with the 20thcentury. However, a conflicting sea surface temperaturerecord has been derived from coral in New Caledonia,suggesting a 1.4 °C cooling around 1730 (Crowleyet al. 1999). It has been postulated that the apparentregional differences in the temperature records forthe Little Ice Age are the result of significant shifts inthe ocean-atmosphere system during this period, withtemperature gradients between tropical low latitudesand mid-to-high latitudes being greater during theLittle Ice Age (Hendy et al. 2002). For the AthertonTableland this may have produced higher rainfall in alow amplitude El Niño period (D’Arrigo et al. 2005). Theevidence from Lake Euramoo seems to confirm thisLIA climate scenario, with the combination of lowcharcoal accumulation and low open forest-disturbancetaxa in the rain forest pointing to wetter conditions andpossibly muted interannual climate variability (El Niño,Southern Oscillation) during the 17th to 19th century.
Clearly temperatures, rainfall and the amplitude andfrequency of El Niño-related climate variability havefluctuated through time and space in the region. At thetime of European arrival on the Atherton Tableland,the region appears to have been entering a period ofheightened climate variability, with severe droughts(e.g. Australia’s Federation Drought of 1895–1902)coinciding with the early settlement period and thusexacerbating the impact of human activity on a previouslyvery wet and relatively stable environment. The combinedimpact of these climate and land-use changes led to anunprecedented shift in rain forest community compo-sition that still persists, despite 50 years of protection.It therefore seems unlikely that ongoing protection andmanagement will allow rain forest habitats to return toa pre-European state, particularly given that futureclimate change is expected to bring about further shiftsin rain forest habitat distribution and loss of keyvertebrate species (Williams et al. 2003). This supportsthe notion that ecological and climate thresholds arenot necessarily the same, and the effects of crossingthem are not necessarily reversible (Maslin 2004).
Conclusions
Pre-European rain forests on the Atherton Tablelandwere highly dynamic, responding to climate change at arange of time-scales (millennium to decades), as well asto fire-related disturbance and Aboriginal activity. Theresponse of both terrestrial and aquatic ecosystems
to European settlement in the region was rapid andprofound, and was most likely driven by changes in fireregime and by forest clearance. The loss of diversity mayhave led to an irreversible switch to an alternative state.
Future research should include studies of local environ-mental change in similar small catchments in order tocompare species and community responses to the Europeansettlement event on the Atherton Tableland. Retrospec-tive studies of the historic range of variability of forestand aquatic ecosystems and fire may provide at leasttentative answers to the three key questions that can theninform management decisions and resource planning.
Acknowledgements
The authors wish to thank Peter Kershaw for encourag-ing this work. Nick Porch and Ainsley Noakes helpedwith fieldwork. Andy Moores prepared the diatomsamples. Roger Flower and Patrick Riaoul provideduseful diatom taxonomic advice. Permission to core atLake Euramoo was granted through the QueenslandParks and Wildlife Service, Queensland Government.Radiocarbon and Lead-210 analysis was completed atthe Australian Nuclear Science and Technology Organ-ization funded through an AINSE Grant 00/60. S.H.was supported by an Australian Research Council QEIIFellowship at Monash University and the AustralianNational University. J.T. was supported by an AustralianResearch Council grant. S.D. was supported by anAustralian Postgraduate Award.
References
Appleby, P.G. & Oldfield, F. (1983) Assessment of 210Pb fromsites with varying sediment accumulation rates. Hydrobio-logia, 103, 29–35.
Battarbee, R.W., Jones, V.J., Flower, R.J., Cameron, N.G.,Bennion, H., Carvalho, L. et al. (2001) Diatoms. TrackingEnvironmental Change Using Lake Sediments. Volume 3:Terrestrial, Algal and Siliceous Indicators (eds J.P. Smol,H.J.B. Birks & W.M. Last), pp. 155–202. Kluwer Academic,Dordrecht, The Netherlands.
Bennett, K.D. (1994) ‘psimpoll’, Version 2.23: a C Programfor analysing pollen data and plotting pollen diagrams.INQUA Commission for the Study of the Holocene. WorkingGroup on Data-handling Methods, Newsletter, 11, 4–6.
Bennett, K.D. (1996) Determination of the number of zones in abiostratigraphical sequence. New Phytologist, 132, 155–170.
Bennett, K.D. & Willis, K.J. (2001) Pollen. Tracking Environ-mental Change Using Lake Sediements. Volume 3: Terrestrial,Algal, and Siliceous Indicators (eds J.P. Smol, H.J.B. Birks &W.M. Last), pp. 355–361. Kluwer, Dordrecht, The Netherlands.
Bennion, H. (1995) Surface-sediment diatom assemblages inshallow, artificial, enriched ponds, and implications forreconstructing trophic status. Diatom Research, 10, 1–19.
Birks, H.J.B. (1998) Numerical tools in palaeolimnology:progress, potentialities, and problems. Journal of Paleo-limnology, 20, 307–332.
Birks, H.J.B. & Gordon, A.D. (1985) Numerical Methods inQuaternary Pollen Analysis. Academic Press, London.
Birtles, T.G. (1982) Trees to burn: settlement in the Atherton-Evelyn rainforest, 1880–1900. North Australia ResearchBulletin, 8, 31–86.

1001European impact on Australian rain forest
© 2006 The AuthorsJournal compilation © 2006 British Ecological Society, Journal of Ecology, 94, 987–1002
Birtles, T.G. (1988) European interpretation of the Atherton-Evelyn ‘vine scrub’ of tropical north Queensland, 1880–1920. Changing Tropical Forests: Historical Perspectives onToday’s Challenges in Asia, Australasia and Oceania (eds J.Dargavel, K. Dixon & N. Semple), pp. 197–216. WorkshopMeeting, Canberra, 16–18 May 1988. Centre for Resourceand Environmental Studies, ANU, Canberra.
Bowman, D.M.J.S., Walsh, A. & Milne, D.J. (2001) Forestexpansion and grassland contraction within a Eucalyptussavanna matrix between 1941 and 1994 at Litchfield NationalPark in the Australian monsoon tropics. Global Ecologyand Biogeography, 10, 535–548.
ter Braak, C.J.F. & Smilauer, P. (2002) CANOCO Reference Manualand CanoDraw for Windows User’s Guide: Software for Canon-ical Community Ordination, Version 4.5. MicrocomputerPower, Ithaca.
Brooks, S.J., Bennion, H. & Birks, H.J.B. (2001) Tracing laketrophic history with a chironomid–total phosphorusinference model. Freshwater Biology, 46, 513–533.
Clark, J.S. & Royall, P.D. (1995) Transformation of a northernhardwood forest by aboriginal (Iroquois) fire: charcoalevidence from Crawford Lake, Ontario, Canada. Holocene,5, 1–9.
Cosgrove, R. (2005) Coping with noxious nuts. Nature Australia,28 (6), 47–53.
Crowley, G.M. & Garnett, S.T. (2000) Changing fire managementin the pastoral lands of Cape York Peninsula of northeastAustralia, 1623–1996. Australian Geographical Studies, 38,10–26.
Crowley, T.J., Quinn, T.M. & Hyde, W.T. (1999) Validation of coraltemperature calibrations. Paleoceanography, 14, 605–615.
D’Arrigo, R., Cook, E.R., Wilson, R.J., Allan, R. & Mann, M.E.(2005) On the variability of ENSO over the past six centuries.Geophysical Research Letters, 32, L03711.
Dai, A. & Wigley, T.M.L. (2000) Global patterns of ENSO-induced precipitation. Geophysical Research Letters, 27,1283–1286.
Dimitriadis, S. & Cranston, P. (2001) An Australian Holoceneclimate reconstruction using Chironomidae from a tropicalvolcanic maar lake. Mammology, Palaeoclimatology, Palae-oecology, 176, 109–131.
Fensham, R.J. & Fairfax, R.J. (2002) Aerial photography forassessing vegetation change: a review of applications andthe relevance of findings for Australian vegetation history.Australian Journal of Botany, 50, 415–429.
Gell, P.A., Sluiter, I.R. & Fluin, J. (2002) Seasonal andinter-annual variations in diatom assemblages in MurrayRiver-connected wetlands in northwest Victoria, Australia.Marine and Freshwater Research, 53, 981–992.
Gell, P., Tibby, J., Fluin, J., Leahy, P., Reid, M., Adamson, K.et al. (2005) Accessing limnological change and variabilityusing fossil diatom assemblages, south-east Australia. RiverResearch and Applications, 21, 257–269.
Godfred-Spenning, C.R. & Reason, C.J.C. (2002) Interannualvariability of lower-tropospheric moisture transport duringthe Australian Monsoon. International Journal of Climatology,22, 509–532.
Griffiths, T. (2001) One hundred years of environmental crisis.Rangeland Journal, 23, 5–14.
Griffiths, T. (2002) How many trees make a forest? Culturaldebates about vegetation change in Australia. AustralianJournal of Botany, 50, 375–389.
Haberle, S.G. (2005) A 23 000-yr pollen record from LakeEuramoo, Wet Tropics of NE Queensland, Australia.Quaternary Research, 64, 343–356.
Haberle, S.G. & David, B. (2004) Climates of change: humandimensions of Holocene environmental change in lowlatitudes of the PEPII transect. Quaternary International,118–119, 165–179.
Haberle, S.G., Hope, G.S. & van der Kaars, W.A. (2001) Biomassburning in Indonesia and Papua New Guinea: natural and
human induced fire events in the fossil record. Mammology,Palaeoclimatology, Palaeoecology, 171, 259–268.
Hayne, M. & Chappell, J. (2001) Cyclone frequency duringthe last 5000 years at Curacoa Island, north Queensland,Australia. Mammology, Palaeoclimatology, Palaeoecology,168, 207–219.
Hendy, E.J., Gagan, M.K., Alibert, C.A., McCulloch, M.T.,Lough, J.M. & Isdale, P.J. (2002) Abrupt decrease intropical Pacific sea surface salinity at end of Little Ice Age.Science, 295, 1511–1514.
Hilbert, D.W., Bradford, M., Parker, T. & Westcott, D.A.(2004) Golden bowerbird (Prionodura newtonia) habitat inpast, present and future climates: predicted extinction of avertebrate in tropical highlands due to global warming.Biological Conservation, 116, 367–377.
Hill, R. & Baird, A. (2003) Kuku-Ya;anji rainforest aboriginalpeople and carbohydrate resource management in theWet Tropics of Queensland, Australia. Human Ecology, 31,27–52.
Hill, R., Baird, A. & Buchanan, D. (1999) Aborigines and firein the Wet Tropics of Queensland, Australia: ecosystemmanagement across cultures. Society and Natural Resources,12, 205–223.
Hopkins, M.S., Head, J., Ash, J., Hewett, R.K. & Graham, A.W.(1996) Evidence for Holocene and continuing recentexpansion of lowland rain forest in humid, tropical NorthQueensland. Journal of Biogeography, 23, 737–745.
Jacobson, G.L. Jr & Bradshaw, R.H.W. (1981) The selectionof sites for paleovegetational studies. Quaternary Research,16, 80–96.
Kershaw, A.P. (1970) A pollen diagram from Lake Euramoo,North-East Queensland, Australia. New Phytologist, 69,785–805.
Kershaw, A.P. (1971) A pollen diagram from Quincan Crater,north-east Queensland, Australia. New Phytologist, 70,669–681.
Kershaw, A.P. (1975) Stratigraphy and pollen analysis ofBromfield Swamp, north-east Queensland, Australia. NewPhytologist, 75, 173–191.
Kershaw, A.P. (1978) The analysis of aquatic vegetation onthe Atherton Tableland, north-east Queensland, Australia.Australian Journal of Ecology, 3, 23–42.
Kershaw, A.P. (1983) A Holocene pollen diagram from Lynch’sCrater, north-eastern Queensland, Australia. New Phytologist,94, 669–682.
Kershaw, A.P. & Nix, H.A. (1988) Quantitative palaeo-climatic estimates from pollen data using bioclimatic profilesof extant taxa. Journal of Biogeography, 15, 589–602.
Kirkpatrick, J.B. (1999) A Contintent Transformed. OxfordUniversity Press, Melbourne, Australia.
Korhola, A., Virkanen, J., Tikkanen, M. & Blom, T. (1996)Fire-induced pH Rise in a naturally acid hill-top lake, SouthernFinland: a palaeoecological survey. Journal of Ecology, 84,257–265.
Krammer, K. & Lange-Bertalot, H. (1986) Bacillariophyceae.1: Teil: Naviculaceae. Gustav Fischer Verlag, Jena.
Krammer, K. & Lange-Bertalot, H. (1988) Bacillariophyceae.2: Teil: Bacillariaceae, Epthimiaceae, Surirellaceae. GustavFischer Verlag, Jena.
Krammer, K. & Lange-Bertalot, H. (1991) Bacillariophyceae.3: Centrales, Fragilariaceae, Eunotiaceae. Gustav FischerVerlag, Stuttgart.
Leahy, P.J., Kershaw, A.P., Tibby, J. & Heijnis, H. (2005) Apalaeoecological reconstruction of the impact of Europeansettlement on Bolin Billabong, Yarra River floodplain.Rivers Research and Applications, 21, 131–149.
Lunt, I.D. (2002) Grazed, burnt and cleared: how ecologistshave studied century-scale vegetation changes in Australia.Australian Journal of Botany, 50, 391–407.
Maslin, M. (2004) Ecological versus climate thresholds. Science,306, 2197–2198.

1002S. G. Haberle et al.
© 2006 The AuthorsJournal compilation © 2006 British Ecological Society, Journal of Ecology, 94, 987–1002
Millspaugh, S.H. & Whitlock, C. (1995) A 750-year fire historybased on lake sediment records in central YellowstoneNational Park, USA. Holocene, 5, 283–292.
Morgan, P., Aplet, G.H., HauOer, J.B., Hum, H.C., Moore, M.M.& Wilson, W.D. (1994) Historical range of variability: auseful tool for evaluating ecosystem change. Journal ofSustainable Forestry, 2, 87–111.
Rasiah, V., Florentine, S.K., Williams, B.L. & Westbrooke, M.E.(2004) The impact of deforestation and pasture abandonmenton soil properties in the wet tropics of Australia. Geoderma,120, 35–45.
Reid, M.A., Fluin, J., Ogden, R.W., Tibby, J.C. & Kershaw, A.P.(2002) Long-term perspectives on human impacts onfloodplain-river ecosystems, Murray-Darling Basin,Australia. Internationale Vereinigung für Theoretische undAngewandte Limnologie: Verhandlungen, 28, 710–716.
Renberg, I. (1990) A procedure for preparing large sets of diatomslides from sediment cores. Journal of Paleolimnology, 19,399–416.
Rossaro, B. (1991) Chironomids and water temperature.Aquatic Insects, 13, 87–98.
Russell-Smith, J., Stanton, P.J., Edwards, A. & Whitehead, P.J.(2004) Rain forest invasion of eucalypt-dominated woodlandsavanna, Iron Rangee, north-eastern Australia. II. Rates oflandscape change. Journal of Biogeography, 31, 1305–1316.
Sayer, C.D. (2001) Problems with the application of diatom-totalphosphorus transfer functions: examples from a shallowEnglish lake. Freshwater Biology, 46, 743–757.
Scheffer, M., Carpenter, S., Foley, J.A., Folke, C. & Walker, B.(2001) Catastrophic shifts in ecosystems. Nature, 413, 591–596.
Stuiver, M., Reimer, P.J., Bard, E., Beck, J.W., Burr, G.S.,Hughen, K.A. et al. (1998) INTCAL98 Radiocarbon agecalibration 24 000–0 cal . Radiocarbon, 40, 1041–1083.
Telford, R.J., Heegaard, E. & Birks, H.J.B. (2004) All age-depthmodels are wrong: but how badly? Quaternary ScienceReviews, 23, 1–5.
Tibby, J. (2001) Diatoms as indicators of sedimentary
processes in Burrinjuck reservoir, New South Wales,Australia. Quaternary International, 83–85, 245–256.
Tibby, J., Reid, M., Fluin, J., Hart, B.T. & Kershaw, A.P.(2003) Assessing long-term pH change in an Australian rivercatchment using monitoring and palaeolimnological data.Environmental Science and Technology, 37, 3250–3255.
Timms, B.V. (1976) Morphology of Lakes Barrine, Eachamand Euramoo, Atherton Tableland, North Queensland.Proceedings of the Royal Society of Queensland, 87, 81–84.
Vyverman, W. (1991) Diatoms from Papua New Guinea.Cramer, Berlin.
Walker, D., Head, M.J., Hancock, G.J. & Murray, A.S. (2000)Establishing a chronology for the last 1000 years of laminatedsediment accumulation at Lake Barrine, a tropical uplandmaar lake, northeastern Australia. Holocene, 10, 415–427.
Walker, D. & Owen, J.A.K. (1999) The characteristics andsource of laminated mud at Lake Barrine, Northeast Australia.Quaternary Science Reviews, 18, 1597–1624.
Walker, I.R., Wilson, S.E. & Smol, J.P. (1995) Chironomidae(Diptera) – quantitative palaeosalinity indicators for lakes ofWestern Canada. Canadian Journal of Fisheries and AquaticSciences, 52, 950–960.
Whittaker, R.J., Araújo, M.B., Jepson, P., Ladle, R.J.,Watson, J.E.M. & Willis, K.J. (2005) Conservation biogeo-graphy: assessment and prospect. Diversity and Distributions,11, 3–23.
Williams, S.E., Bolitho, E.E. & Fox, S. (2003) Climate changein Australian tropical rainforests: an impending environ-mental catastrophe. Proceedings of the Royal Society ofLondon B, 270, 1887–1892.
Winter, J.W., Bell, F.C., Pahl, L.I. & Atherton, R.G. (1987)Rainforest clearance in northeastern Australia. Proceed-ings of the Royal Society of Queensland, 98, 41–57.
Received 4 October 2005 revision accepted 2 February 2006 Handling Editor: Peter Moore