boletin 39 del invemar
TRANSCRIPT

BOLETÍN DE INVESTIGACIONES MARINAS Y COSTERAS(Bulletin of Marine and Coastal Research)
BOLE
TÍN D
E INV
ESTIG
ACIO
NES M
ARIN
AS Y
COST
ERAS
39 ( 1
)
(Bulletin of Marine and Coastal Research)(Bulletin of Marine and Coastal Research)
Instituto de Investigaciones Marinas y Costeras “José Benito Vives De Andréis”Vinculado al Ministerio de Ambiente,Vivienda y Desarrollo Territorial
Año 2010 - Volumen 39 (1)
VOL. 39 (1)
Santa Marta - Colombia
Santa Marta, Colombia, 2010
ISSN: 0122-9761
Serie de Publicaciones Periódicas
BOLETÍN DE INVESTIGACIONES MARINAS Y COSTERAS(Bulletin of Marine and Coastal Research)
(ISSN 0122-9761)
La revista científica BOLETÍN DE INVESTIGACIONES MARINAS Y COSTERAS, premiada por Colciencias como la mejor publicación en su género en Colombia en 1997, y clasificada dentro de Publindex (Colciencias) como revista indexada en categoría A2, considera para su publicación trabajos inéditos sobre cualquier tópico de investigación en el mar o en ambientes acuáticos de la zona costera, realizados en las áreas tropicales y subtropicales de América. Es editada desde 1967 por el Instituto de Investigaciones Marinas y Costeras “José Benito Vives De Andréis”-INVEMAR. Hasta el volumen 8 llevó el nombre de “Mittelungen aus dem Instituto Colombo-Alemán de Investigaciones Científicas Punta de Betín” y entre los volúmenes 9 y 24 se denominó “Anales del Instituto de Investigaciones Marinas y Costeras”. A partir de 2008 la revista se publica en dos números por volumen anual. El Boletín se distribuye en 37 países alrededor del mundo a través del sistema de canje. Las instrucciones a los autores para la preparación y envío de manuscritos se encuentran en las últimas páginas de este número y en el portal de Internet www.invemar.org.co/boletin
Director INVEMARFranciso Arias Isaza INVEMAR, Colombia Director de la RevistaJesús A. Garay, M. Sc. INVEMAR, Colombia
Editor Responsable, Vol. 39 (1)Arturo Acero Pizarro, Ph. D. Univ.Nacional de Colombia sede Caribe-CECIMAR,Colombia
Comité EditorialArturo Acero Pizarro, Ph. D. Univ. Nacional de Colombia sede Caribe-CECIMAR, ColombiaCarlos Andrade, Ph. D. Escuela Naval de Cadetes, ColombiaAdriana Bermúdez, Dr. rer. nat. Universidad de Cartagena, ColombiaGladys Bernal, Ph. D. Universidad Nacional de Colombia sede Medellín, ColombiaJacobo Blanco, M. Sc. Universidad del Magdalena, ColombiaMaribeb Castro-González, Dr. en Oceanografía Universidad del Tolima, ColombiaJuan Manuel Díaz, Dr. rer. nat. Centro de Investigaciones Biológicas, ColombiaGuillermo Díaz-Pulido, Ph. D. Griffith University, AustraliaGuillermo Duque, Ph. D. Universidad Nacional de Colombia sede Palmira, ColombiaLuisa Fernanda Espinosa, Ph. D. INVEMAR, ColombiaAndrés Franco, Ph. D. Universidad de Bogotá Jorge Tadeo Lozano, ColombiaDiego L. Gil-Agudelo, Ph. D. INVEMAR, ColombiaJavier Gómez-León, Ph. D. INVEMAR, ColombiaGeorgina Guzmán, Ph. D. INVEMAR, ColombiaJuan Carlos Narváez, M. Sc. Universidad del Magdalena, Colombia Gabriel Navas, M. Sc. INVEMAR, ColombiaCarlos Palacio, Ph. D. Universidad de Antioquia, ColombiaJorge Páramo, M. Sc. Universidad del Magdalena, ColombiaMónica Puyana, Ph. D. Universidad de Bogotá Jorge Tadeo Lozano, ColombiaEfraín Rodríguez, Ph. D. Centro Control Contaminación del Pacífico-CCCP, ColombiaMario Rueda, Ph. D. INVEMAR, ColombiaAdolfo Sanjuán, M. Sc. Universidad de Bogotá Jorge Tadeo Lozano, ColombiaMarie-Luise Schnetter, Dr. rer. nat. Justus Liebig Universität Giessen, AlemaniaYves-François Thomas, Dr. en Geografía Université Paris, FranciaLuisa Villamil, Ph. D. Universidad de Bogotá Jorge Tadeo Lozano, Colombia
Asistente EditorialAmalia María Cano-Castaño, Bióloga INVEMAR, Colombia
CONTENIDO - CONTENTS
L. Gónima, L. E. Ruiz y M. E. González
Desarrollo de una metodología sencilla para la georreferenciación y medición de distancias
a partir de imágenes de satélite sistemáticamente georreferenciadas
[Development of a simple methodology for georeferencing and distance measuring from
systematic georeferenced satellite images] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .7
A. M. Galeano Chavarría y E. Arteaga Sogamoso
Distribución y abundancia de diatomeas del género Pseudo-nitzschia en condición de El
Niño 2007 sobre la cuenca pacífica colombiana
[Distribution and abundance of diatoms of the genus Pseudo-nitzschia in El Niño conditions
in 2007 in the Colombian Pacific Basin] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .25
L. Flórez-Leiva, A. Rangel-Campo, M. Díaz-Ruiz, D. E. Venera-Pontón y G. Díaz-Pulido
Efecto de la sedimentación en el reclutamiento de las macroalgas Dictyota spp. y Lobophora
variegata: un estudio experimental en el Parque Nacional Natural Tayrona, Caribe colombiano
[Effects of sedimentation on the recruitment of the macroalgae Dictytota spp. and Lobophora
variegata: an experimental study in the Tayrona National Natural Park, Colombian Caribbean] . . . . . . . . . . . . . . . . . .41
N. Novoa-Bustos, A. C. Hernández-Zanuy y R. Viquez-Portuguez
Distribución y abundancia de las ascidias de los fondos rocosos de la bahía de Cuajiniquil,
Costa Rica
[Distribution and abundance of the rocky bottom ascidians of Cuajiniquil Bay, Costa Rica] . . . . . . . . . . . . . . . . . .57
T. Posada P., Á. L. Peña C. y G. R. Navas S.
Hidrozoos de la familia Aglaopheniidae de la plataforma continental y talud superior del
Caribe colombiano
[Hydroids of the family Aglaopheniidae from the Colombian Caribbean continental shelf and
upper slope] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .67
Y. Lozano-Duque, L. A. Vidal y G. R. Navas S.
Listado de diatomeas (Bacillariophyta) registradas para el mar Caribe colombiano
[Check-list of diatoms (Bacillariophyta) reported from the Colombian Caribbean Sea] . . . . . . . . . . . . . . . . . .83
BOLETÍN DE INVESTIGACIONESMARINAS Y COSTERASBOLETÍN DE INVESTIGACIONESMARINAS Y COSTERAS
J. G. Domínguez, J. C. Gómez, C. Ricaurte, G. Mayo, J. Orejarena, J. M. Díaz y
C. A. Andrade
Cobertura de los fondos y paisajes bentónicos asociados a formaciones diapíricas en los bancos
de Salmedina, plataforma continental del Caribe colombiano
[Seafloor coverage and benthic landscapes associated to diapiric (mud volcanism) formations
in Salmedina Banks, Colombian Caribbean continental shelf] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 117
K. Gómez-Campo, M. Rueda y C. García-Valencia
Distribución espacial, abundancia y relación con características del hábitat del caracol pala
Eustrombus gigas (Linnaeus) (Mollusca: Strombidae) en el archipiélago Nuestra Señora del Rosario,
Caribe colombiano
[Spatial distribution, abundance and relationship to habitat features of the queen conch
Eustrombus gigas (Linnaeus) (Mollusca: Strombidae) in Nuestra Señora del Rosario archipelago,
Caribbean coast of Colombia] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .137
S. Espinosa G., M. F. Delgado Hernández, B. Orobio Riofrio, L. M. Mejía-Ladino y D. L.
Gil-Agudelo
Estado de la población y valoración de algunas estrategias de conservación del recurso piangua
Anadara tuberculosa (Sowerby) en sectores de Bazán y Nerete, costa pacífica nariñense de
Colombia
[Population status and valuation of some conservation strategies of piangua Anadara tuberculosa
(Sowerby) in areas of Bazán and Nerete, Nariño, Pacífic coast of Colombia] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .161
J. López-Navarro, J. Gómez-León, C. Castellanos Romero y E. Acosta Ortiz
Efecto de la densidad de siembra sobre el crecimiento y supervivencia de Nodipecten nodosus
(Bivalvia: Pectinidae) en un nuevo arte de cultivo
[Effect of the density on growth and survival of the scallop Nodipecten nodosus (Bivalvia: Pectinidae)
in a suspended culture] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .177
NOTAS – NOTES
S. A. Balaguera-Reina, J. F. González-Maya y A. Acero P.
Fauna nocturna asociada a los manglares y otros humedales en la Vía Parque Isla de Salamanca,
departamento del Magdalena, Caribe colombiano
[Nocturnal fauna associated to mangroves and other wetlands in the Vía Parque Isla de
Salamanca, Departamento Magdalena, Colombian Caribbean] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .191
Continúa en la contraportada interior
Viene de la contraportada

BOLETÍN DE INVESTIGACIONES MARINAS Y COSTERAS(Bulletin of Marine and Coastal Research)
BOLE
TÍN D
E INV
ESTIG
ACIO
NES M
ARIN
AS Y
COST
ERAS
39 ( 1
)
(Bulletin of Marine and Coastal Research)(Bulletin of Marine and Coastal Research)
Instituto de Investigaciones Marinas y Costeras “José Benito Vives De Andréis”Vinculado al Ministerio de Ambiente,Vivienda y Desarrollo Territorial
Año 2010 - Volumen 39 (1)
VOL. 39 (1)
Santa Marta - Colombia
Santa Marta, Colombia, 2010
ISSN: 0122-9761
Serie de Publicaciones Periódicas
BOLETÍN DE INVESTIGACIONES MARINAS Y COSTERAS(Bulletin of Marine and Coastal Research)
(ISSN 0122-9761)
La revista científica BOLETÍN DE INVESTIGACIONES MARINAS Y COSTERAS, premiada por Colciencias como la mejor publicación en su género en Colombia en 1997, y clasificada dentro de Publindex (Colciencias) como revista indexada en categoría A2, considera para su publicación trabajos inéditos sobre cualquier tópico de investigación en el mar o en ambientes acuáticos de la zona costera, realizados en las áreas tropicales y subtropicales de América. Es editada desde 1967 por el Instituto de Investigaciones Marinas y Costeras “José Benito Vives De Andréis”-INVEMAR. Hasta el volumen 8 llevó el nombre de “Mittelungen aus dem Instituto Colombo-Alemán de Investigaciones Científicas Punta de Betín” y entre los volúmenes 9 y 24 se denominó “Anales del Instituto de Investigaciones Marinas y Costeras”. A partir de 2008 la revista se publica en dos números por volumen anual. El Boletín se distribuye en 37 países alrededor del mundo a través del sistema de canje. Las instrucciones a los autores para la preparación y envío de manuscritos se encuentran en las últimas páginas de este número y en el portal de Internet www.invemar.org.co/boletin
Director INVEMARFranciso Arias Isaza INVEMAR, Colombia Director de la RevistaJesús A. Garay, M. Sc. INVEMAR, Colombia
Editor Responsable, Vol. 39 (1)Arturo Acero Pizarro, Ph. D. Univ.Nacional de Colombia sede Caribe-CECIMAR,Colombia
Comité EditorialArturo Acero Pizarro, Ph. D. Univ. Nacional de Colombia sede Caribe-CECIMAR, ColombiaCarlos Andrade, Ph. D. Escuela Naval de Cadetes, ColombiaAdriana Bermúdez, Dr. rer. nat. Universidad de Cartagena, ColombiaGladys Bernal, Ph. D. Universidad Nacional de Colombia sede Medellín, ColombiaJacobo Blanco, M. Sc. Universidad del Magdalena, ColombiaMaribeb Castro-González, Dr. en Oceanografía Universidad del Tolima, ColombiaJuan Manuel Díaz, Dr. rer. nat. Centro de Investigaciones Biológicas, ColombiaGuillermo Díaz-Pulido, Ph. D. Griffith University, AustraliaGuillermo Duque, Ph. D. Universidad Nacional de Colombia sede Palmira, ColombiaLuisa Fernanda Espinosa, Ph. D. INVEMAR, ColombiaAndrés Franco, Ph. D. Universidad de Bogotá Jorge Tadeo Lozano, ColombiaDiego L. Gil-Agudelo, Ph. D. INVEMAR, ColombiaJavier Gómez-León, Ph. D. INVEMAR, ColombiaGeorgina Guzmán, Ph. D. INVEMAR, ColombiaJuan Carlos Narváez, M. Sc. Universidad del Magdalena, Colombia Gabriel Navas, M. Sc. INVEMAR, ColombiaCarlos Palacio, Ph. D. Universidad de Antioquia, ColombiaJorge Páramo, M. Sc. Universidad del Magdalena, ColombiaMónica Puyana, Ph. D. Universidad de Bogotá Jorge Tadeo Lozano, ColombiaEfraín Rodríguez, Ph. D. Centro Control Contaminación del Pacífico-CCCP, ColombiaMario Rueda, Ph. D. INVEMAR, ColombiaAdolfo Sanjuán, M. Sc. Universidad de Bogotá Jorge Tadeo Lozano, ColombiaMarie-Luise Schnetter, Dr. rer. nat. Justus Liebig Universität Giessen, AlemaniaYves-François Thomas, Dr. en Geografía Université Paris, FranciaLuisa Villamil, Ph. D. Universidad de Bogotá Jorge Tadeo Lozano, Colombia
Asistente EditorialAmalia María Cano-Castaño, Bióloga INVEMAR, Colombia
CONTENIDO - CONTENTS
L. Gónima, L. E. Ruiz y M. E. González
Desarrollo de una metodología sencilla para la georreferenciación y medición de distancias
a partir de imágenes de satélite sistemáticamente georreferenciadas
[Development of a simple methodology for georeferencing and distance measuring from
systematic georeferenced satellite images] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .7
A. M. Galeano Chavarría y E. Arteaga Sogamoso
Distribución y abundancia de diatomeas del género Pseudo-nitzschia en condición de El
Niño 2007 sobre la cuenca pacífica colombiana
[Distribution and abundance of diatoms of the genus Pseudo-nitzschia in El Niño conditions
in 2007 in the Colombian Pacific Basin] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .25
L. Flórez-Leiva, A. Rangel-Campo, M. Díaz-Ruiz, D. E. Venera-Pontón y G. Díaz-Pulido
Efecto de la sedimentación en el reclutamiento de las macroalgas Dictyota spp. y Lobophora
variegata: un estudio experimental en el Parque Nacional Natural Tayrona, Caribe colombiano
[Effects of sedimentation on the recruitment of the macroalgae Dictytota spp. and Lobophora
variegata: an experimental study in the Tayrona National Natural Park, Colombian Caribbean] . . . . . . . . . . . . . . . . . .41
N. Novoa-Bustos, A. C. Hernández-Zanuy y R. Viquez-Portuguez
Distribución y abundancia de las ascidias de los fondos rocosos de la bahía de Cuajiniquil,
Costa Rica
[Distribution and abundance of the rocky bottom ascidians of Cuajiniquil Bay, Costa Rica] . . . . . . . . . . . . . . . . . .57
T. Posada P., Á. L. Peña C. y G. R. Navas S.
Hidrozoos de la familia Aglaopheniidae de la plataforma continental y talud superior del
Caribe colombiano
[Hydroids of the family Aglaopheniidae from the Colombian Caribbean continental shelf and
upper slope] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .67
Y. Lozano-Duque, L. A. Vidal y G. R. Navas S.
Listado de diatomeas (Bacillariophyta) registradas para el mar Caribe colombiano
[Check-list of diatoms (Bacillariophyta) reported from the Colombian Caribbean Sea] . . . . . . . . . . . . . . . . . .83
BOLETÍN DE INVESTIGACIONESMARINAS Y COSTERASBOLETÍN DE INVESTIGACIONESMARINAS Y COSTERAS
J. G. Domínguez, J. C. Gómez, C. Ricaurte, G. Mayo, J. Orejarena, J. M. Díaz y
C. A. Andrade
Cobertura de los fondos y paisajes bentónicos asociados a formaciones diapíricas en los bancos
de Salmedina, plataforma continental del Caribe colombiano
[Seafloor coverage and benthic landscapes associated to diapiric (mud volcanism) formations
in Salmedina Banks, Colombian Caribbean continental shelf] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 117
K. Gómez-Campo, M. Rueda y C. García-Valencia
Distribución espacial, abundancia y relación con características del hábitat del caracol pala
Eustrombus gigas (Linnaeus) (Mollusca: Strombidae) en el archipiélago Nuestra Señora del Rosario,
Caribe colombiano
[Spatial distribution, abundance and relationship to habitat features of the queen conch
Eustrombus gigas (Linnaeus) (Mollusca: Strombidae) in Nuestra Señora del Rosario archipelago,
Caribbean coast of Colombia] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .137
S. Espinosa G., M. F. Delgado Hernández, B. Orobio Riofrio, L. M. Mejía-Ladino y D. L.
Gil-Agudelo
Estado de la población y valoración de algunas estrategias de conservación del recurso piangua
Anadara tuberculosa (Sowerby) en sectores de Bazán y Nerete, costa pacífica nariñense de
Colombia
[Population status and valuation of some conservation strategies of piangua Anadara tuberculosa
(Sowerby) in areas of Bazán and Nerete, Nariño, Pacífic coast of Colombia] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .161
J. López-Navarro, J. Gómez-León, C. Castellanos Romero y E. Acosta Ortiz
Efecto de la densidad de siembra sobre el crecimiento y supervivencia de Nodipecten nodosus
(Bivalvia: Pectinidae) en un nuevo arte de cultivo
[Effect of the density on growth and survival of the scallop Nodipecten nodosus (Bivalvia: Pectinidae)
in a suspended culture] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .177
NOTAS – NOTES
S. A. Balaguera-Reina, J. F. González-Maya y A. Acero P.
Fauna nocturna asociada a los manglares y otros humedales en la Vía Parque Isla de Salamanca,
departamento del Magdalena, Caribe colombiano
[Nocturnal fauna associated to mangroves and other wetlands in the Vía Parque Isla de
Salamanca, Departamento Magdalena, Colombian Caribbean] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .191
Continúa en la contraportada interior
Viene de la contraportada

BOLETÍN DE INVESTIGACIONES MARINAS Y COSTERAS(Bulletin of Marine and Coastal Research)
BOLE
TÍN D
E INV
ESTIG
ACIO
NES M
ARIN
AS Y
COST
ERAS
39 ( 1
)
(Bulletin of Marine and Coastal Research)(Bulletin of Marine and Coastal Research)
Instituto de Investigaciones Marinas y Costeras “José Benito Vives De Andréis”Vinculado al Ministerio de Ambiente,Vivienda y Desarrollo Territorial
Año 2010 - Volumen 39 (1)
VOL. 39 (1)
Santa Marta - Colombia
Santa Marta, Colombia, 2010
ISSN: 0122-9761
Serie de Publicaciones Periódicas
BOLETÍN DE INVESTIGACIONES MARINAS Y COSTERAS(Bulletin of Marine and Coastal Research)
(ISSN 0122-9761)
La revista científica BOLETÍN DE INVESTIGACIONES MARINAS Y COSTERAS, premiada por Colciencias como la mejor publicación en su género en Colombia en 1997, y clasificada dentro de Publindex (Colciencias) como revista indexada en categoría A2, considera para su publicación trabajos inéditos sobre cualquier tópico de investigación en el mar o en ambientes acuáticos de la zona costera, realizados en las áreas tropicales y subtropicales de América. Es editada desde 1967 por el Instituto de Investigaciones Marinas y Costeras “José Benito Vives De Andréis”-INVEMAR. Hasta el volumen 8 llevó el nombre de “Mittelungen aus dem Instituto Colombo-Alemán de Investigaciones Científicas Punta de Betín” y entre los volúmenes 9 y 24 se denominó “Anales del Instituto de Investigaciones Marinas y Costeras”. A partir de 2008 la revista se publica en dos números por volumen anual. El Boletín se distribuye en 37 países alrededor del mundo a través del sistema de canje. Las instrucciones a los autores para la preparación y envío de manuscritos se encuentran en las últimas páginas de este número y en el portal de Internet www.invemar.org.co/boletin
Director INVEMARFranciso Arias Isaza INVEMAR, Colombia Director de la RevistaJesús A. Garay, M. Sc. INVEMAR, Colombia
Editor Responsable, Vol. 39 (1)Arturo Acero Pizarro, Ph. D. Univ.Nacional de Colombia sede Caribe-CECIMAR,Colombia
Comité EditorialArturo Acero Pizarro, Ph. D. Univ. Nacional de Colombia sede Caribe-CECIMAR, ColombiaCarlos Andrade, Ph. D. Escuela Naval de Cadetes, ColombiaAdriana Bermúdez, Dr. rer. nat. Universidad de Cartagena, ColombiaGladys Bernal, Ph. D. Universidad Nacional de Colombia sede Medellín, ColombiaJacobo Blanco, M. Sc. Universidad del Magdalena, ColombiaMaribeb Castro-González, Dr. en Oceanografía Universidad del Tolima, ColombiaJuan Manuel Díaz, Dr. rer. nat. Centro de Investigaciones Biológicas, ColombiaGuillermo Díaz-Pulido, Ph. D. Griffith University, AustraliaGuillermo Duque, Ph. D. Universidad Nacional de Colombia sede Palmira, ColombiaLuisa Fernanda Espinosa, Ph. D. INVEMAR, ColombiaAndrés Franco, Ph. D. Universidad de Bogotá Jorge Tadeo Lozano, ColombiaDiego L. Gil-Agudelo, Ph. D. INVEMAR, ColombiaJavier Gómez-León, Ph. D. INVEMAR, ColombiaGeorgina Guzmán, Ph. D. INVEMAR, ColombiaJuan Carlos Narváez, M. Sc. Universidad del Magdalena, Colombia Gabriel Navas, M. Sc. INVEMAR, ColombiaCarlos Palacio, Ph. D. Universidad de Antioquia, ColombiaJorge Páramo, M. Sc. Universidad del Magdalena, ColombiaMónica Puyana, Ph. D. Universidad de Bogotá Jorge Tadeo Lozano, ColombiaEfraín Rodríguez, Ph. D. Centro Control Contaminación del Pacífico-CCCP, ColombiaMario Rueda, Ph. D. INVEMAR, ColombiaAdolfo Sanjuán, M. Sc. Universidad de Bogotá Jorge Tadeo Lozano, ColombiaMarie-Luise Schnetter, Dr. rer. nat. Justus Liebig Universität Giessen, AlemaniaYves-François Thomas, Dr. en Geografía Université Paris, FranciaLuisa Villamil, Ph. D. Universidad de Bogotá Jorge Tadeo Lozano, Colombia
Asistente EditorialAmalia María Cano-Castaño, Bióloga INVEMAR, Colombia
CONTENIDO - CONTENTS
L. Gónima, L. E. Ruiz y M. E. González
Desarrollo de una metodología sencilla para la georreferenciación y medición de distancias
a partir de imágenes de satélite sistemáticamente georreferenciadas
[Development of a simple methodology for georeferencing and distance measuring from
systematic georeferenced satellite images] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .7
A. M. Galeano Chavarría y E. Arteaga Sogamoso
Distribución y abundancia de diatomeas del género Pseudo-nitzschia en condición de El
Niño 2007 sobre la cuenca pacífica colombiana
[Distribution and abundance of diatoms of the genus Pseudo-nitzschia in El Niño conditions
in 2007 in the Colombian Pacific Basin] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .25
L. Flórez-Leiva, A. Rangel-Campo, M. Díaz-Ruiz, D. E. Venera-Pontón y G. Díaz-Pulido
Efecto de la sedimentación en el reclutamiento de las macroalgas Dictyota spp. y Lobophora
variegata: un estudio experimental en el Parque Nacional Natural Tayrona, Caribe colombiano
[Effects of sedimentation on the recruitment of the macroalgae Dictytota spp. and Lobophora
variegata: an experimental study in the Tayrona National Natural Park, Colombian Caribbean] . . . . . . . . . . . . . . . . . .41
N. Novoa-Bustos, A. C. Hernández-Zanuy y R. Viquez-Portuguez
Distribución y abundancia de las ascidias de los fondos rocosos de la bahía de Cuajiniquil,
Costa Rica
[Distribution and abundance of the rocky bottom ascidians of Cuajiniquil Bay, Costa Rica] . . . . . . . . . . . . . . . . . .57
T. Posada P., Á. L. Peña C. y G. R. Navas S.
Hidrozoos de la familia Aglaopheniidae de la plataforma continental y talud superior del
Caribe colombiano
[Hydroids of the family Aglaopheniidae from the Colombian Caribbean continental shelf and
upper slope] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .67
Y. Lozano-Duque, L. A. Vidal y G. R. Navas S.
Listado de diatomeas (Bacillariophyta) registradas para el mar Caribe colombiano
[Check-list of diatoms (Bacillariophyta) reported from the Colombian Caribbean Sea] . . . . . . . . . . . . . . . . . .83
BOLETÍN DE INVESTIGACIONESMARINAS Y COSTERASBOLETÍN DE INVESTIGACIONESMARINAS Y COSTERAS
J. G. Domínguez, J. C. Gómez, C. Ricaurte, G. Mayo, J. Orejarena, J. M. Díaz y
C. A. Andrade
Cobertura de los fondos y paisajes bentónicos asociados a formaciones diapíricas en los bancos
de Salmedina, plataforma continental del Caribe colombiano
[Seafloor coverage and benthic landscapes associated to diapiric (mud volcanism) formations
in Salmedina Banks, Colombian Caribbean continental shelf] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 117
K. Gómez-Campo, M. Rueda y C. García-Valencia
Distribución espacial, abundancia y relación con características del hábitat del caracol pala
Eustrombus gigas (Linnaeus) (Mollusca: Strombidae) en el archipiélago Nuestra Señora del Rosario,
Caribe colombiano
[Spatial distribution, abundance and relationship to habitat features of the queen conch
Eustrombus gigas (Linnaeus) (Mollusca: Strombidae) in Nuestra Señora del Rosario archipelago,
Caribbean coast of Colombia] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .137
S. Espinosa G., M. F. Delgado Hernández, B. Orobio Riofrio, L. M. Mejía-Ladino y D. L.
Gil-Agudelo
Estado de la población y valoración de algunas estrategias de conservación del recurso piangua
Anadara tuberculosa (Sowerby) en sectores de Bazán y Nerete, costa pacífica nariñense de
Colombia
[Population status and valuation of some conservation strategies of piangua Anadara tuberculosa
(Sowerby) in areas of Bazán and Nerete, Nariño, Pacífic coast of Colombia] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .161
J. López-Navarro, J. Gómez-León, C. Castellanos Romero y E. Acosta Ortiz
Efecto de la densidad de siembra sobre el crecimiento y supervivencia de Nodipecten nodosus
(Bivalvia: Pectinidae) en un nuevo arte de cultivo
[Effect of the density on growth and survival of the scallop Nodipecten nodosus (Bivalvia: Pectinidae)
in a suspended culture] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .177
NOTAS – NOTES
S. A. Balaguera-Reina, J. F. González-Maya y A. Acero P.
Fauna nocturna asociada a los manglares y otros humedales en la Vía Parque Isla de Salamanca,
departamento del Magdalena, Caribe colombiano
[Nocturnal fauna associated to mangroves and other wetlands in the Vía Parque Isla de
Salamanca, Departamento Magdalena, Colombian Caribbean] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .191
Continúa en la contraportada interior
Viene de la contraportada
Comité CientíficoDalila Aldana, Ph. D. CINVESTAV, MéxicoJuan José Carballo, Ph. D. Universidad Nacional Autónoma de México, MéxicoStella de la Torre, Ph. D. Universidad San Francisco de Quito, EcuadorGabriel Delgado, Ph. D. Florida Fish and Wildlife Conservation Comission, EE. UU.Luis Freites, Ph. D. Universidad de Oriente, VenezuelaRocío García, Ph. D. Universidad del Magdalena, ColombiaGabriel Genzano, Ph. D. Universidad Nacional de Mar del Plata, ArgentinaJosep Gili, Ph. D. Instituto de Ciencias del Mar de Barcelona, EspañaDavid Hernández, Ph. D. Universidad Nacional Autónoma de México, MéxicoDiana Maya, M. Sc. PontificiaUniversidadJaveriana,ColombiaYaneth Muñoz, M. Sc. Universidad Nacional de Colombia, ColombiaÁlvaro Navarrete, Ph. D. Colegio Frontera Sur, MéxicoAlicia Ojeda, Ph. D. Instituto Canario de Ciencias Marinas, EspañaEnrique Peña, Ph. D. Universidad del Valle, ColombiaJaime Polanía, Ph. D. Universidad Nacional de Colombia, ColombiaBlanca Posada, M. Sc. INVEMAR, ColombiaRosana Rocha, Ph. D. Universidade Federal do Paraná, BrasilAna Silva, Ph. D. Universidad de Costa Rica, Costa RicaGeorge Vernette, Ph. D. Université Bordeaux, Francia
Foto portada: Francisco A. Arias IsazaCanje: Centro de Documentación, INVEMAR, Colombia (E-mail: [email protected])Distribución y sucripciones: Comité Editorial, Boletín de Investigaciones Marinas y Costeras, INVEMAR. A.A. 1016. Cerro Punta de Betín, Santa Marta, Colombia. E-mail: [email protected] web: www.invemar.org.coBoletín en línea: www.invemar.org.co/boletinDiagramación y montaje electrónico: Alejandro Ríos, Marquillas S.A.Impresión: Marquillas S.A., Sabaneta (Antioquia), Colombia
Derechos Reservados conforme a la ley, los textos pueden ser reproducidos total o parcialmente citando la fuente.
Citar la obra completa INVEMAR. 2010. Boletín de Investigaciones Marinas y Costeras. Vol. 39 (1). Serie de Publicaciones Periódicas. Instituto de Investigaciones Marinas y Costeras, Santa Marta. 207 p.


BOLETÍN DE INVESTIGACIONES MARINAS Y COSTERAS(Bulletin of Marine and Coastal Research)
39 (1)
Instituto de Investigaciones Marinas y Costeras“José Benito Vives De Andréis”
Vinculado al Ministerio de Ambiente,Vivienda y Desarrollo Territorial
Santa Marta, Colombia, 2010
Registrado en ASFA, Biological Abstracts, BIOSIS PREVIEWS, Geobase,Periodica, Zoological Record, Latindex, Publindex y Scielo-Colombia


CONTENIDO – CONTENTS
L. Gónima, L. E. Ruiz y M. E. GonzálezDesarrollo de una metodología sencilla para la georreferenciación y medición de distancias a partir de imágenes de satélite sistemáticamente georreferenciadas[Development of a simple methodology for georeferencing and distance measuring from systematic georeferenced satellite images] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7
A. M. Galeano Chavarría y E. Arteaga SogamosoDistribución y abundancia de diatomeas del género Pseudo-nitzschia en condición de El Niño 2007 sobre la cuenca pacífica colombiana[Distribution and abundance of diatoms of the genus Pseudo-nitzschia in El Niño conditions in 2007 in the Colombian Pacific Basin] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 25
L. Flórez-Leiva, A. Rangel-Campo, M. Díaz-Ruiz, D. E. Venera-Pontón
y G. Díaz-PulidoEfecto de la sedimentación en el reclutamiento de las macroalgas Dictyota spp . y Lobophora variegata: un estudio experimental en el Parque Nacional Natural Tayrona, Caribe colombiano[Effects of sedimentation on the recruitment of the macroalgae Dictytota spp . and Lobophora variegate: an experimental study in the Tayrona National Natural Park, Colombian Caribbean] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41
N. Novoa-Bustos, A. C. Hernández-Zanuy y R. Viquez-PortuguezDistribución y abundancia de las ascidias de los fondos rocosos de la bahía de Cuajiniquil, Costa Rica[Distribution and abundance of the rocky bottom ascidians of Cuajiniquil Bay, Costa Rica] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 57
T. Posada P., Á. L. Peña C. y G. R. Navas S.Hidrozoos de la familia Aglaopheniidae de la plataforma continental y talud superior del Caribe colombiano[Hydroids of the family Aglaopheniidae from the Colombian Caribbean continental shelf and upper slope] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 67
Y. Lozano-Duque, L. A. Vidal y G. R. Navas S.Listado de diatomeas (Bacillariophyta) registradas para el mar Caribe colombiano[Check-list of diatoms (Bacillariophyta) reported from the Colombian Caribbean Sea] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 83
J. G. Domínguez, J. C. Gómez, C. Ricaurte, G. Mayo, J. Orejarena,
J. M. Díaz y C. A. AndradeCobertura de los fondos y paisajes bentónicos asociados a formaciones diapíricas en los bancos de Salmedina, plataforma continental del Caribe colombiano[Seafloor coverage and benthic landscapes associated to diapiric (mud volcanism) formations in Salmedina Banks, Colombian Caribbean continental shelf] . . . . . . . . . . . . . . . . . . . . . . . . 117

K. Gómez-Campo, M. Rueda y C. García-ValenciaDistribución espacial, abundancia y relación con características del hábitat del caracol pala Eustrombus gigas (Linnaeus) (Mollusca: Strombidae) en el archipiélago Nuestra Señora del Rosario, Caribe colombiano[Spatial distribution, abundance and relationship to habitat features of the queen conch Eustrombus gigas (Linnaeus) (Mollusca: Strombidae) in Nuestra Señora del Rosario archipelago, Caribbean coast of Colombia] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 137
S. Espinosa G., M. F. Delgado Hernández, B. Orobio Riofrio, L. M.
Mejía-Ladino y D. L. Gil-AgudeloEstado de la población y valoración de algunas estrategias de conservación del recurso piangua Anadara tuberculosa (Sowerby) en sectores de Bazán y Nerete, costa pacífica nariñense de Colombia[Population status and valuation of some conservation strategies of piangua Anadara tuberculosa (Sowerby) in areas of Bazán and Nerete, Nariño, Pacífic coast of Colombia] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 161
J. López-Navarro, J. Gómez-León, C. Castellanos Romero y E. Acosta OrtizEfecto de la densidad de siembra sobre el crecimiento y supervivencia de Nodipecten nodosus (Bivalvia: Pectinidae) en un nuevo arte de cultivo[Effect of the density on growth and survival of the scallop Nodipecten nodosus (Bivalvia: Pectinidae) in a suspended culture] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 177
NOTAS – NOTES
S. A. Balaguera-Reina, J. F. González-Maya y A. Acero P.Fauna nocturna asociada a los manglares y otros humedales en la Vía Parque Isla de Salamanca, departamento del Magdalena, Caribe colombiano[Nocturnal fauna associated to mangroves and other wetlands in the Vía Parque Isla de Salamanca, Departamento Magdalena, Colombian Caribbean] . . . . . . . . . . . . . . . . . . 191

Instituto de Investigaciones Marinas y Costeras - INVEMAR 7
Bol . Invest . Mar . Cost . 39 (1) ISSN 0122-9761 Santa Marta, Colombia, 2010
DESARROLLO DE UNA METODOLOGÍA SENCILLA PARA LA GEORREFERENCIACIÓN Y MEDICIÓN DE DISTANCIAS A PARTIR DE IMÁGENES DE SATÉLITE
SISTEMÁTICAMENTE GEORREFERENCIADAS
Leonardo Gónima, Libardo E. Ruiz y Marcos E. González
Universidad de Córdoba, Departamento de Física y Electrónica, Grupo de Materiales y Física Aplicada. Carrera 6 No. 76-103, Montería, Córdoba, Colombia. [email protected] (L.G.); [email protected] (L.R.); [email protected] (M.G.)
RESUMEN
Uno de los principales problemas para una precisa georreferenciación y medición de distancias a partir de imágenes de satélite, especialmente en zonas geográficas con una gran dinámica morfológica y ambiental, reside no solamente en la dificultad para la identificación de puntos de control (GCPs, por su sigla en inglés), sino también en las limitaciones reales para el acceso a dichos lugares . En el presente trabajo se propone una metodología relativamente simple para la georreferenciación y la medición de distancias a partir de imágenes de satélite, basada en la utilización de imágenes de la reflectancia de la superficie previamente calculadas y luego orientadas hacia el norte (georreferenciación sistemática) . A partir de estas imágenes y fijando un punto (píxel) de control básico P, medido con GPS, los demás GCPs se obtuvieron midiendo distancias definidas desde dicho punto P hasta puntos (píxeles) representativos sobre la imagen, seleccionados para la georreferenciación de ella . La validación estadística de los resultados obtenidos, utilizando una muestra diferente de GCPs medidos con GPS, muestra que la precisión de la georreferenciación y la medición de distancias utilizando la metodología desarrollada son similares a la obtenida por medios convencionales como la georreferenciación de imágenes por medio de datos de GPS .
PALABRAS CLAVE: Imágenes de satélite, Radiometría, Georreferenciación, Medición de distancias, Errores estadísticos .
ABSTRACT
Development of a simple methodology for georeferencing and distance measuring from systematic georeferenced satellite images. One of the main problems for a precise georeferencing and distance measurements from satellite images, especially in geographical zones with strong morphologic and environmental dynamics, lies not only in the difficulty for identifying ground control points (GCPs), but also in real limitations for accessing such places . In this work a relatively simple methodology is proposed for georeferencing and distance measuring from satellite images, based on the utilization of previously calculated reflectance images from the surface and then oriented toward the north (systematic georeferencing) . From these images and setting a basic control point (pixel) P, measured with GPS, the
7-23

8 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
other GCPs were obtained by measurements of distances defined from the P point to representative points (pixels) on the image, selected for its georeferencing . The statistical validation of the obtained results, using a different sample of GCPs measured with GPS, shows that the precision of the georeferencing and distance measurement utilizing the developed methodology is similar to that obtained by conventional procedures, such as image georeferencing from GPS data .
KEY WORDS: Satellite images, Radiometry, Georeferencing, Distance measurements, Statistical errors .
INTRODUCCIÓN
La utilización de imágenes de satélite es una herramienta muy poderosa
para el estudio y el monitoreo de los fenómenos naturales y antrópicos ocurridos
en el sistema superficie terrestre-atmósfera . En este sentido, para la localización
geográfica de dichos fenómenos se requiere una gran precisión cartográfica de las
imágenes, proceso digital que se conoce con el nombre de georreferenciación .
Para georreferenciar una imagen de satélite es indispensable disponer de
puntos de control (GCPs, por su sigla en inglés), adquiridos a través de mapas, de
otras imágenes georreferenciadas o de mediciones en terreno con GPS . De esta
manera la precisión de la georreferenciación dependerá de la calidad de los GCPs
adquiridos, estando ésta representada por la capacidad que se tenga en localizar
acertadamente, tanto en la imagen como en el terreno o lugar deseado, los puntos
de control a utilizarse en el proceso . En la literatura se encuentran un sinnúmero
de referencias relacionadas con la georreferenciación de imágenes de los satélites
Landsat y SPOT de zonas planas (cultivos, áreas urbanas, planicies y llanuras,
desiertos, entre otros), basada en la utilización de GCPs provenientes de mapas
cartográficos o de imágenes de satélite previamente georreferenciadas (Pratt, 1974;
Lucas y Kanade, 1981; Brown, 1992)
En la búsqueda continua para facilitar y mejorar la precisión de la
georreferenciación de imágenes de satélite, en las últimas décadas se han
desarrollado nuevos métodos para la adquisición de los GCPs . Wisetphanichkij et al . (1998) muestran los resultados obtenidos con la aplicación de algoritmos de
detección secuencial similar (SSDAs) para la determinación precisa de los GCPs
utilizando imágenes ópticas del satélite JERS-1 de la ciudad de Bangkok (Tailandia) .
Por otra parte, Huseby y Solberg (1998) presentan un algoritmo para la detección
automática de GCPs basado en una técnica de “matching”, utilizando imágenes
NOAA-AVHRR, EOS MODIS y Landsat TM . Otro procedimiento desarrollado es la
extracción automática de GCPs comparando radiométrica y visualmente imágenes
IKONOS (Ambrosio et al ., 1996) .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 9
Con la puesta en funcionamiento operacional de los GPS, a partir de 1990,
el uso de este instrumento para la medición de GCPs con el fin de georreferenciar
imágenes de satélite ha tenido una vasta difusión, registrada profusamente en la
bibliografía especializada . Por ejemplo, Wilkie (1990) obtuvo GCPs necesarios para
la georreferenciación de imágenes Landsat TM en zonas con cartografía inexistente o
deficiente (bosque húmedo tropical del noreste de Zaire) utilizando un GPS Magnavox
4400 . Con un Magellan NAV 1000 PRO, Clavet et al. (1993) georreferenciaron
imágenes SPOT con la precisión requerida para la producción de cartografía digital a
escala 1:50000 . Más aún, imágenes digitales georreferenciadas de mayor precisión,
de Landsat TM, SPOT multiespectral y SPOT pancromática fueron obtenidas a partir
de mediciones con GPS, que aquellas con GCPs provenientes de tabletas digitales
(mapas impresos), según lo registrado por Cook y Pinder (1996) y Kardoulas et al. (1996) . Esto último fue posteriormente comprobado por Smith y Atkinson (2001) al
establecer la importancia que tiene la recolección de GCPs con GPS diferenciales
en la calidad de la georreferenciación de imágenes de satélite SPOT . Finalmente,
Gao (2001) mostró que utilizar GPS no diferenciales para la medición de GCPs es
un procedimiento suficientemente preciso para la georreferenciación de imágenes
de satélite de resolución espacial media como Landsat TM y SPOT multiespectral .
A pesar de los avances expuestos anteriormente para lograr una más precisa
georreferenciación de imágenes de satélite, todavía subsisten dificultades en la
ubicación de los respectivos GCPs, aún en zonas planas y de acceso relativamente
fácil, pero especialmente en aquellas aisladas de acceso difícil o prácticamente nulo .
Como una contribución a la solución de este problema, en el presente trabajo se
discute y se propone una metodología relativamente simple y de fácil aplicación para
la georreferenciación satisfactoria de imágenes de satélite de resolución espacial
media (por ejemplo, SPOT 4) y la medición de distancias a partir de ellas, cuya
aplicabilidad también es válida para zonas de difícil o restringida accesibilidad . Esta
metodología está basada en la precisa identificación, en una imagen calculada de la
reflectancia de la superficie georreferenciada sistemáticamente, de un único punto
(píxel) de control básico P, simultáneamente medido con un GPS no diferencial . A
partir de dicho P se obtienen, por medición de distancias (conteo de píxeles), los
demás GCPs indispensables para la adecuada georreferenciación de la imagen de
satélite seleccionada .
ÁREA DE ESTUDIO
La zona de estudio está localizada al norte del departamento de Córdoba como se muestra en la Figura 1 . En ésta se incluye parte de los municipios de San

10 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Bernardo del Viento, Lorica y San Antero, el delta de Tinajones y la bahía de Cispatá . Está caracterizada por ser una zona con una gran dinámica morfológica, debido a los continuos cambios de curso del río Sinú y las consecuentes variaciones de la línea de costa (Sánchez et al., 2005) . Por otra parte, la zona presenta algunas dificultades para el acceso a diversos lugares, tanto por tierra (vías en mal estado) como por vía acuática, por ejemplo: el delta de Tinajones, el sector de las ciénagas de la bahía de Cispatá y aquel denominado Cuchilla de Cispatá (cerros de baja altura) . Además de lo anterior, como consecuencia de la uniformidad en la cobertura vegetal del área de estudio, representada principalmente por vegetación de manglar y pastos, en algunos lugares se presentan inconvenientes en la ubicación de puntos de control, con la suficiente precisión, lo cual se traduce en una menor calidad de los resultados obtenidos con el proceso de georreferenciación .
Figura 1 . Localización geográfica de la zona de estudio (imagen SPOT 4) .
MATERIALES Y MÉTODOS
Está ampliamente difundido en la literatura especializada que una de las
dificultades en la identificación adecuada de los GCPs en una imagen de satélite
óptica es la alteración que produce la atmósfera sobre la radiación solar reflejada por
la superficie terrestre y que posteriormente es captada por los sensores satelitales
(Jensen, 2005) . Esto es aún más evidente en regiones tropicales, en las cuales
la atmósfera, debido a su alto contenido de vapor de agua, modifica aún más la

Instituto de Investigaciones Marinas y Costeras - INVEMAR 11
radiación electromagnética reflejada proveniente de la superficie terrestre y que es
medida por los sensores satelitales . Por otra parte, el que no todas las imágenes de
satélite sean captadas en forma vertical, sino con un ángulo de visión determinado,
también contribuye a alterar radiométricamente los valores de los píxeles: entre
mayor sea la inclinación del sensor respecto a la vertical a la superficie, mayor
será la probabilidad de equivocación en la interpretación de ellos . Para el caso de
la imagen seleccionada en este estudio, SPOT 4, la inclinación del sensor respecto
a la superficie correspondió a un ángulo de visión bastante grande (26 .8O) . Por
consiguiente, para una mejor y más precisa identificación e interpretación de las
imágenes de satélite, se calcularon a partir de éstas imágenes de la reflectancia de
la superficie con correcciones atmosféricas y del ángulo de visión para una banda
espectral i, ri, de acuerdo con la siguiente ecuación (Gónima y Alados, 2002):
ri = (Li ↑-Ai) / [Bi + (Li ↑Ai)*Ci] (1)
Donde, Li↑es la radiación solar espectral reflejada por la superficie en
dirección del sensor y Ai, B
i y C
i son constantes radiativas espectrales .Acorde con lo
expuesto anteriormente, la metodología utilizada fue la siguiente:
1 . Procesamiento digital de una imagen de satélite SPOT 4 del 12 .04 .2007
sin georreferenciar, representado por el cálculo de la reflectancia de la superficie
para cada banda espectral (cuatro en total), de acuerdo con la ecuación 1 .
2 . Agrupamiento de las cuatro imágenes ópticas de reflectancia espectral de
la superficie calculadas en un solo archivo imagen, con el fin de georreferenciarlas
posteriormente en bloque .
3 . Otro de los efectos que no solamente afectan la radiometría de una
imagen, sino que también la distorsionan espacialmente, es que la órbita del sensor
no está muchas veces orientada exactamente hacia el norte geográfico . Este es el
caso particular de la imagen seleccionada, en la cual debido al ángulo de visión del
sensor y a la órbita desfasada respecto al norte, las superficies en la imagen original
aparecen de dimensiones diferentes a las que realmente tienen . Esta situación se
corrige en gran medida georreferenciando la imagen en forma sistemática . Entonces,
la imagen del numeral anterior se georreferenció sistemáticamente utilizando los
cinco GCPs (uno por cada una de las esquinas y uno del centro de la imagen SPOT
de dimensiones originales) suministrados en el archivo VOL_LIST existente en el
“header” de la imagen SPOT 4 mencionada. Este procedimiento, a pesar de que la
precisión de localización absoluta de puntos en las imágenes (zonas planas) registrada
por SPOTimage (2008) es de aproximadamente 350 m, permite orientarlas hacia el
norte geográfico, lo cual es muy importante para el desarrollo de la metodología
propuesta .

12 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
4 . Ubicación en la imagen georreferenciada sistemáticamente del píxel
correspondiente al punto de control básico P, a partir del cual se determinaron los
GCPs indispensables para la georreferenciación de ella . P fue seleccionado de
manera que correspondiera a un píxel lo más precisamente identificable (radiométrica
y visualmente) en la imagen y que representara al mismo tiempo un punto fácilmente
reconocible visualmente en el terreno, en este caso dentro del casco urbano de la
población de San Antero (Figura 2: círculo color rojo) .
5 . Medición en campo de las coordenadas del píxel P con GPS no diferencial .
6 . Georreferenciación de la imagen del numeral 3 utilizando 20 GCPs
medidos en campo con GPS para posterior validación de la metodología propuesta a
continuación (Figura 2: círculos amarillos) .
7 . Determinación de los GCPs necesarios para la georreferenciación de la
imagen orientada hacia el norte . El procedimiento desarrollado para la obtención de
cada uno de los nuevos GCPs fue el siguiente (Figura 3):
a) Selección de un píxel cualquiera en otro lugar sobre la imagen M .
b) A partir del píxel P se traza una línea recta horizontal (en dirección E-W
ó W-E, según M se encuentre al oeste de P o viceversa) que intersecte la línea recta
proyectada verticalmente desde el píxel M (en dirección N-S ó S-N, según M se
encuentre al norte de P o viceversa) . El píxel de intersección entre las dos líneas
rectas se denomina c en la Figura 3 .
c) A continuación se cuenta el número de píxeles existentes entre los píxeles
P y c, multiplicándose dicho número por el tamaño lineal del píxel (resolución
espacial) . Para la imagen seleccionada la resolución espacial es de 20 m .
d) De acuerdo con el literal anterior, la distancia horizontal en metros así
calculada, sumada o restada al valor de la componente x del píxel P, dependiendo
de si c se encuentra al este o al oeste de dicho punto respectivamente, representa la
coordenada x del nuevo GCP .
e) Para la definición de la coordenada y del nuevo GCP se procede de
manera similar a partir del píxel c, midiendo la distancia (línea recta vertical) en
metros desde dicho píxel hasta el píxel M y sumándola o restándola a la coordenada
y del píxel P, de acuerdo a que M se encuentre al norte o al sur de dicho píxel,
respectivamente . El resultado de la operación matemática anterior corresponde con
la coordenada y del nuevo GCP .
Es de anotar que las coordenadas de P y las correspondientes a los nuevos
GCPs, medidas y calculadas respectivamente, fueron asignadas al centro de cada
uno de los píxeles respectivos, como representativo de cada punto de control en

Instituto de Investigaciones Marinas y Costeras - INVEMAR 13
Figura 2. Imagen de la reflectancia de la superficie georreferenciada a través de 20 puntos de GPS y utilizada para la validación estadística con 21 nuevos GCPs medidos con GPS .
Figura 3. Representación gráfica para la definición de los GCPs de acuerdo con la metodología desarrollada .
particular . En la Figura 4 se muestra la ubicación de 26 píxeles M (círculos azul claro)
seleccionados en la imagen corregida (reflectancia de la superficie y orientación
hacia el norte) y que corresponden a los GCPs utilizados para la georreferenciación
de ella de acuerdo con la metodología desarrollada (numeral 7) . Es de anotar que
aún en las zonas marinas y terrestres de acceso restringido, se establecieron también
puntos de control .

14 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Figura 4. Ubicación espacial de los GCPs utilizados para la georreferenciación de la imagen de la reflectancia de la superficie por medio de la metodología desarrollada (imagen SPOT 4) .
RESULTADOS Y DISCUSIÓN
A pesar de que las imágenes calculadas de la reflectancia de la superficie con
correcciones atmosféricas y del ángulo de visión posteriormente fueron orientadas
hacia el norte, permitieron obtener una mayor precisión y facilidad en la interpretación
y la ubicación de los GCPs requeridos para la posterior georreferenciación y
medición de distancias más precisas a partir de ellas . Sólo es posible establecer la
validez de la metodología desarrollada por medio de comparaciones estadísticas
8 . Georreferenciación de la imagen utilizando los GCPs calculados de
acuerdo con el numeral anterior . Comparación de los resultados obtenidos con la
imagen georreferenciada por medio de la metodología desarrollada con los de las
imágenes georreferenciadas sistemáticamente (numeral 3) y con GPS (numeral 6) .
9 . Validación de las tres imágenes georreferenciadas mencionadas
anteriormente con nuevos GCPs medidos con GPS (21 en total) .
10 . Validación de distancias calculadas en las tres imágenes georreferenciadas
por medio de los métodos expuestos anteriormente .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 15
Imagen georreferenciada sistemáticamente
(5 GCPs: “header” imagen SPOT 4)
Imagen georreferenciada con GPS
(20 GCPs: círculos amarillos en Figura 2)
Imagen georreferenciada metodología desarrollada
(26 GCPs: Figura 4)
emc (x) emc (y) emc (x) emc (y) emc (x) emc (y)
388 m 367 m 27 m 27 m 28 m 29 m
Como la comparación de los resultados a partir de los mismos GCPs
utilizados para la georreferenciación de las tres imágenes mencionadas anteriormente
no puede ser interpretada estadísticamente como suficientemente significativa, se
seleccionó una nueva muestra de datos constituida por 21 GCPs diferentes, medidos
en campo con GPS y representados por los círculos verde claro de las Figuras 2, 5 y
6 . La Tabla 2 muestra la aplicación de la metodología desarrollada en la definición
de las coordenadas de los nuevos GCPs por medio del cálculo de las distancias entre
el punto P y cada uno de dichos puntos de control .
Así mismo, en la Tabla 3 se muestran las coordenadas de los 21 GCPs
utilizados para la validación definitiva de la metodología desarrollada, tanto para las
Tabla 1 . Error medio cuadrático emc calculado para las tres imágenes georreferenciadas a partir de sus respectivos GCPs .
entre los resultados obtenidos, inicialmente a partir de los mismos GCPs utilizados
para la georreferenciación de la imagen corregida por los tres métodos expuestos
anteriormente, a saber:
a)Los cinco GCPs suministrados en el “header” de la imagen SPOT 4 adquirida,
procedimiento denominado comúnmente georreferenciación sistemática .
b) 20 GCPs medidos en campo con GPS (Figura 2: círculos amarillos) .
c) La metodología desarrollada (Figura 4) .
La comparación estadística mencionada se muestra en la Tabla 1 . Los
resultados de la Tabla 1 demuestran que la imagen corregida (georreferenciación
sistemática) es la que menor precisión ofrece, tanto por lo pequeño de la muestra
de GCPs seleccionados (cinco en total) como por la calidad de éstos, lo que
corrobora lo informado por SPOTimage (2008) . Si se comparan los resultados
de la georreferenciación realizada con datos de GPS con los obtenidos mediante
la metodología desarrollada, se observa que los errores medios cuadráticos emc
calculados por ambos métodos son bastante similares, lo que significa que la
precisión de ambos procedimientos es de aproximadamente ±1 píxel para las
muestras de GCPs con que ellas mismas fueron georreferenciadas .

16 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Figura 5 . Imagen de la reflectancia de la superficie georreferenciada sistemáticamente y utilizada para la validación estadística con 21 nuevos GCPs medidos con GPS .
Figura 6. Imagen de la reflectancia de la superficie georreferenciada a través de la metodología desarrollada y utilizada para la validación estadística con 21 nuevos GCPs medidos con GPS .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 17
mediciones con GPS como para cada una de las tres imágenes georreferenciadas por
los tres métodos expuestos anteriormente . A partir de los datos de la Tabla 3 se hizo
una comparación estadística entre las coordenadas de los nuevos GCPs medidas con
GPS y las coordenadas extraídas de cada una de las tres imágenes georreferenciadas,
basada en el cálculo de las diferencias entre ellas (Tabla 4) .
Observando las diferencias individuales de la Tabla 4, nuevamente
las imágenes georreferenciadas con datos provenientes de GPS y con aquellos
provenientes de la metodología desarrollada, son las que muestran resultados bastante
similares respecto a las mediciones en campo con GPS de los 21 GCPs . Como era de
esperarse, la imagen georreferenciada sistemáticamente es la que menor precisión
ofrece, mucho menor que cualquiera de las otras dos . Analizando el emc calculado
para dicha muestra de datos, se comprueba que con la imagen georreferenciada
mediante la metodología desarrollada se obtienen prácticamente los mismos
resultados que con la imagen georreferenciada usando datos de GPS: la precisión
alcanzada en la georreferenciación de ambas imágenes es aproximadamente de ±2
píxeles para cada par de coordenadas x y y, lo cual representa el valor aceptado
internacionalmente para considerar dicho proceso como suficientemente preciso .
La precisión obtenida depende en gran medida de la asignación que se hizo a priori
de que la coordenada de un GCP es representativa del centro del píxel en la imagen,
lo cual muchas veces no representa la realidad del sitio o punto seleccionado . Por
otra parte, en imágenes de resolución espacial media es imposible hacer coincidir
exactamente el punto donde se miden las coordenadas con GPS con un píxel en
particular, debido a que mientras la precisión de las medidas con GPS no diferencial
varía generalmente entre 4 y 10 m, la resolución espacial de dichos píxeles es de
algunas decenas de metros, como en el caso del SPOT 4 (20 m) . Esta situación
mejorará en la medida en que se disponga de imágenes de satélite con más altas
resoluciones espaciales como IKONOS, QuickBird, IRS, entre otros .
Respecto a la calidad de la medición de distancias, al estar la metodología
desarrollada basada exclusivamente en dicha medición para el cálculo de las
coordenadas de cualquier punto o píxel de una imagen y ya que los resultados
de la validación de la georreferenciación por medio de esta misma metodología
son altamente satisfactorios, entonces también la medición de distancias es
suficientemente precisa, como lo demuestran los datos de la Tabla 4 . Es decir, por
medio de esta metodología es posible obtener resultados suficientemente precisos
tanto en la georreferenciación de imágenes de satélite como en la medición de
distancias a partir de ellas .

18 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Núm
ero
de p
íxel
es a
par
tir
de P
(con
teo)
Dis
tanc
ia c
alcu
lada
(m)
Coo
rden
adas
imag
en g
eorr
efer
enci
ada
met
odol
ogía
des
arro
llada
(m)
GC
PsL
ugar
xy
xy
xy
1E
rosi
ón r
ío S
inú
972
(Oes
te)
409
(Sur
)19
440
8180
3968
2210
3015
0
2Pu
ente
La
Bal
sa11
83 (
Oes
te)
182
(Sur
)23
660
3640
3926
2210
3459
0
3Pu
ente
La
Doc
trin
a72
6 (O
este
)59
6 (S
ur)
1452
011
920
4017
4210
2641
0
4Pu
nta
Bel
lo97
(E
ste)
157
(Nor
te)
1940
3140
4181
8210
4148
0
5Pu
nta
Reb
ujin
a I
76 (
Oes
te)
69 (
Nor
te)
1520
1380
4147
4210
3971
0
6Pu
nta
Bol
ívar
170
(Est
e)15
8 (N
orte
)34
0031
6041
9662
1041
450
7A
rroy
o Pu
nta
Bel
lo13
5 (E
ste)
146
(Nor
te)
2700
2920
4189
6210
4121
0
8Pl
aya
Bla
nca
I32
(E
ste)
106
(Nor
te)
640
2120
4169
0210
4045
0
9Pl
aya
Bla
nca
II16
(E
ste)
83 (
Nor
te)
320
1660
4165
8210
3999
0
10Pu
nta
Reb
ujin
a II
58 (
Oes
te)
50 (
Nor
te)
1160
1000
4151
0210
3929
0
11C
año
Cab
añas
83 (
Oes
te)
51(N
orte
)16
6010
2041
4602
1039
350
12C
año
Lob
o18
1 (O
este
)84
(Su
r)36
2016
8041
2642
1036
650
13V
olcá
n de
lodo
76 (
Oes
te)
178
(Sur
)15
2035
6041
4742
1034
770
14V
ía a
Cov
eñas
I18
6 (E
ste)
1 (S
ur)
3720
2041
9982
1038
310
15V
ía a
Cov
eñas
II
141
(Est
e)39
(Su
r)28
2078
041
9082
1037
550
16V
ìa S
an A
nter
o45
(E
ste)
118
(Sur
)90
023
6041
7162
1035
970
17A
rroy
o M
amón
75 (
Est
e)95
(Su
r)15
0019
0041
7762
1036
430
18C
año
Sala
do I
444
(Oes
te)
2331
(N
orte
)88
8029
0040
7382
1041
230
19C
año
Sala
do I
I67
9 (O
este
)16
1 (N
orte
)13
580
3220
4027
0210
4155
0
20C
año
Rem
edia
pobr
e41
5 (O
este
)15
8 (S
ur)
8300
3160
4079
6210
3517
0
21D
elta
de
Tin
ajon
es90
1 (O
este
)13
8 (N
orte
)18
020
2760
3982
4210
4109
0
Tab
la 2
. Det
erm
inac
ión
de la
s di
stan
cias
a p
artir
del
pun
to d
e co
ntro
l bás
ico
P (x
= 4
1626
2 m
, y=
103
8330
m)
y la
s co
orde
nada
s de
los
píxe
les
(GC
Ps)
utili
zado
s pa
ra
la v
alid
ació
n de
la m
etod
olog
ía d
esar
rolla
da (
Figu
ra 6
) .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 19
Med
icio
nes
hech
as c
on G
PS
(m)
Imag
en g
eorr
efer
enci
ada
sist
emát
icam
ente
(m)
Imag
en g
eorr
efer
enci
ada
con
GP
S (m
)Im
agen
geo
rref
eren
ciad
a m
etod
olog
ía d
esar
rolla
da (m
)
GC
PsL
ugar
xy
xy
xy
xy
1E
rosi
ón r
ío S
inú
3968
7110
3011
839
7336
1030
408
3968
2210
3015
039
6822
1030
150
2Pu
ente
La
Bal
sa39
2675
1034
615
3931
3310
3487
739
2622
1034
590
3926
2210
3459
0
3Pu
ente
La
Doc
trin
a40
1757
1026
384
4022
5110
2666
940
1742
1026
408
4017
4210
2641
0
4Pu
nta
Bel
lo41
8230
1041
434
4187
1410
4171
541
8182
1041
450
4181
8210
4148
0
5Pu
nta
Reb
ujin
a I
4147
7810
3973
141
5253
1039
955
4147
4210
3971
141
4742
1039
710
6Pu
nta
Bol
ívar
4196
2410
4148
342
0173
1041
735
4196
6010
4145
041
9662
1041
450
7A
rroy
o P .
Bel
lo41
8987
1041
248
4194
7410
4149
541
8963
1041
211
4189
6210
4121
0
8Pl
aya
Bla
nca
I41
6930
1040
403
4174
1310
4069
541
6902
1040
450
4169
0210
4045
0
9Pl
aya
Bla
nca
II41
6617
1039
939
4170
9110
4023
441
6582
1039
990
4165
8210
3999
0
10Pu
nta
Reb
ujin
a II
4150
7810
3929
041
5614
1039
574
4151
0310
3929
141
5102
1039
290
11C
año
Cab
añas
4146
4310
3930
141
5123
1039
594
4146
0310
3934
841
4602
1039
350
12C
año
Lob
o41
2687
1036
645
4131
3510
3694
541
2642
1036
651
4126
4210
3665
0
13V
olcá
n41
4762
1034
735
4152
5210
3501
241
4743
1034
768
4147
4210
3477
0
14V
ía a
Cov
eñas
I42
0034
1038
278
4205
1310
3855
441
9983
1038
310
4199
8210
3831
0
15V
ía a
Cov
eñas
II
4191
4010
3753
541
9594
1037
795
4190
8010
3755
141
9082
1037
550
16V
ìa S
an A
nter
o41
7200
1035
947
4176
7410
3621
341
7160
1035
970
4171
6210
3597
0
17A
rroy
o M
amón
4177
9410
3643
241
8274
1036
674
4177
6210
3643
341
7762
1036
430
18C
año
Sala
do I
4073
1010
4116
440
7894
1041
406
4073
8210
4123
040
7382
1041
230
19C
año
Sala
do I
I40
2636
1041
483
4031
9210
4179
240
2702
1041
551
4027
0210
4155
0
20C
año
Rem
edia
pobr
e40
7930
1035
072
4084
9310
3541
040
7962
1035
170
4079
6210
3517
0
21D
elta
de
Tin
ajon
es39
8282
1041
049
3987
5510
4133
039
8235
1041
049
3982
4210
4109
0
Tab
la 3
. Coo
rden
adas
de
los
píxe
les
(GC
Ps)
sele
ccio
nado
s pa
ra la
val
idac
ión
de la
met
odol
ogía
des
arro
llada
(Fi
gura
s 2,
5 y
6: c
írcu
los
colo
r ve
rde)
.

20 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Imag
en g
eorr
efer
enci
ada
sist
emát
icam
ente
Imag
en g
eorr
efer
enci
ada
con
GP
SIm
agen
geo
rref
eren
ciad
a m
etod
olog
ía
desa
rrol
lada
GC
PsL
ugar
Δx (
m)
Δy (
m)
Δx (
m)
Δy (
m)
Δx (
m)
Δy (
m)
1E
rosi
ón r
ío S
inú
-465
-290
49-3
249
-32
2Pu
ente
La
Bal
sa-4
58-2
6253
2553
253
Puen
te L
a D
octr
ina
-494
-285
15-2
415
-26
4Pu
nta
Bel
lo-4
84-2
8148
-16
48-4
65
Punt
a R
ebuj
ina
I-4
75-2
2436
2036
216
Punt
a B
olív
ar-5
49-2
52-3
633
-38
337
Arr
oyo
Punt
a B
ello
-487
-247
2437
2538
8Pl
aya
Bla
nca
I-4
83-2
9228
-47
28-4
79
Play
a B
lanc
a II
-474
-295
35-5
135
-51
10Pu
nta
Reb
ujin
a II
-536
-284
-25
-1-2
40
11C
año
Cab
añas
-480
-293
40-4
741
-49
12C
año
Lob
o-4
48-3
0045
-645
-5
13V
olcá
n de
lodo
-490
-277
19-3
320
-35
14V
ía a
Cov
eñas
I-4
79-2
7651
-32
52-3
215
Vía
a C
oveñ
as I
I-4
54-2
6060
-16
58-1
516
Vìa
San
Ant
ero
-474
-266
40-2
338
-23
17A
rroy
o M
amón
-480
-242
32-1
322
18C
año
Sala
do I
-584
-242
-72
-66
-72
-66
19C
año
Sala
do I
I-5
56-3
09-6
6-6
8-6
6-6
720
Cañ
o R
emed
iapo
bre
-563
-338
-32
-98
-32
-98
21D
elta
de
Tin
ajon
es-4
73-2
8147
040
-41
emc
(m)
508.
228
4.0
44.2
41.1
43.9
43.5
Tab
la 4
. Eva
luac
ión
esta
díst
ica
de l
a pr
ecis
ión
de l
a ge
orre
fere
ncia
ción
y d
e la
med
ició
n de
dis
tanc
ias
real
izad
as p
or l
os t
res
mét
odos
util
izad
os (
mue
stra
de
21
GC
Ps m
edid
os c
on G
PS s
elec
cion
ados
par
a la
val
idac
ión)
. Δx:
dif
eren
cia
de la
coo
rden
ada
x en
tre
los
GC
Ps m
edid
os c
on G
PS y
los
obte
nido
s a
part
ir d
e la
res
pect
iva
geor
refe
renc
iaci
ón, Δ
y: d
ifer
enci
a de
la c
oord
enad
a y
entr
e lo
s G
CPs
med
idos
con
GPS
y lo
s ob
teni
dos
a pa
rtir
de
la r
espe
ctiv
a ge
orre
fere
ncia
ción
, em
c: e
rror
med
io
cuad
rátic
o de
las
resp
ectiv
as d
ifer
enci
as .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 21
Esta determinación precisa de distancias por medio de imágenes de satélite
es muy importante para la estimación cuantitativa de variaciones espaciales de zonas
urbanas, corrientes de agua (ríos, caños y canales), línea costera y en la delimitación
de zonas con cobertura vegetal, suelos expuestos y superficies de agua (lagos,
ciénagas y embalses, entre otros) . Por ejemplo, se muestra claramente en el estudio
de Pérez et al. (en prensa), en el cual se utilizaron imágenes multiespectrales Landsat
(TM y ETM+) y SPOT 4 para la cuantificación de los procesos de erosión y acreción
en los sectores La Balsa, el delta de Tinajones y la bahía de Cispatá (departamento
de Córdoba) utilizando la metodología desarrollada .
En conclusión, la metodología desarrollada es una manera relativamente
fácil, sencilla y suficientemente precisa para georreferenciar imágenes de satélite
de zonas planas o relativamente planas, con la ventaja de que solamente requiere
de la identificación lo más precisa posible de un punto de control básico en una
imagen de la reflectancia de la superficie y orientada hacia el norte, a partir del
cual es posible obtener los demás GCPs necesarios para la georreferenciación, aún
los de aquellos puntos o lugares inaccesibles como áreas marinas o terrenas . Es de
esperarse, como también está registrado en la bibliografía especializada, que para
otras condiciones orográficas la precisión de la georreferenciación dependerá de
la disponibilidad y calidad de un modelo digital de terreno, con el cual se puedan
corregir adecuadamente las variaciones radiométricas y espaciales causadas por la
pendiente y orientación del terreno .
CONCLUSIONES
La metodología desarrollada permite obtener resultados suficientemente
precisos de la georreferenciación y de la medición de distancias a partir de imágenes
de satélite previamente corregidas radiométrica (reflectancia con correcciones
atmosféricas y del ángulo de visón) y espacialmente al orientarlas hacia el norte por
medio de una georreferenciación sistemática . Es de esperarse que, con la aplicación
de la metodología desarrollada a imágenes de satélite solamente georreferenciadas
sistemáticamente sin estar previamente corregidas radiométricamente, sea posible
obtener resultados relativamente satisfactorios, siempre y cuando la atmósfera del
área de estudio sea bastante clara (bajo contenido de vapor de agua y aerosoles) y
que el sensor satelital haya adquirido la imagen en posición vertical o cuasi-vertical .
La aplicación de la metodología desarrollada es relativamente fácil y
sencilla, ya que está basada en la adquisición de un solo punto de control GCP
identificado lo más precisamente posible en la imagen de satélite y medido en terreno
con GPS, a partir del cual es posible obtener los demás GCPs, independientemente

22 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
de la accesibilidad física a dichos puntos, por medición de distancias . Estas últimas
tienen especial importancia en la cuantificación de variaciones espaciales de zonas
urbanas, corrientes de agua en general, línea costera y en la delimitación de zonas
con cobertura vegetal, suelos expuestos y superficies de agua .
Aunque la precisión alcanzada por medio de la aplicación de la
metodología desarrollada está dentro de los cánones internacionales, existen errores
debidos al tamaño del píxel de las imágenes de satélite de resolución espacial media
comparado con la precisión de los datos medidos con GPS no diferencial, lo cual
genera incertidumbres al momento de hacer coincidir un mismo punto en el terreno
por ambos medios (imagen y GPS) . Por otra parte, las variaciones del relieve del
terreno son otra de las causas de las imprecisiones en la georreferenciación de
imágenes de satélite y en la medición de distancias a partir de ellas . Esto significa
que la metodología desarrollada da los mejores resultados para zonas planas y/o
relativamente planas, como es mayoritariamente el área de estudio seleccionada .
La metodología desarrollada ha sido aplicada a varias imágenes de satélite
Landsat y SPOT con resultados satisfactorios para zonas planas y relativamente
planas . Sin embargo, una validación definitiva de ésta sólo será posible a medida
que tenga una mayor aplicación en diferentes imágenes de satélite . Una versión
modificada que incluye el efecto de la pendiente del terreno está en vías de desarrollo .
BIBLIOGRAFÍA
Ambrosio, G ., J . González y V . Arévalo .1996 . Comparación de imágenes de satélite para la detección
de cambios temporales . http://www .ceafac .es/actividades/jornadas/XXIV/documentos/viar/96 .pdf .
17/02/2009 .
Brown, L . G . 1992 . A survey of image registration techniques . ACM Comp . Surv ., 24 (4): 325-376 .
Clavet, D ., M . Lasserre y J . Pouliot . 1993 . GPS control for 1:50000-scale topographic mapping from
satellite images . Photogram . Engin . Rem . Sens ., 59 (1): 107-111 .
Cook, A . E y J . E . Pinder . 1996 . Relative accuracy of rectifications using coordinates determined from
maps and the Global Positioning System . Photogram . Engin . Rem . Sens ., 62 (1): 73-77 .
Gao, J . 2001 . Non differential GPS an alternate source of planimetric control for rectifying satellite
imagery . Photogram . Engin . Rem . Sens ., 67 (1): 49-55 .
Gónima, L . y L . Alados . 2002 . Cálculo de la reflectancia a partir de datos de satélite, versión alternativa .
Proceedings Tercera Asamblea Hispano – Portuguesa Geodesia y Geofísica (Valencia, España), 3:
408-413 .
Huseby, R . B . y R . Solberg . 1998 . A model-based approach for geometric correction of optical satellite
images . Reporte 932, Norwegian Computing Center, Oslo . 24 p .
Jensen, J . R . 2005 . Introductory digital image processing, a remote sensing perspective . Tercera Edición .
Prentice Hall Series, Upper Saddle River, EE . UU . 526 p .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 23
Kardoulas, N . G ., A . C . Bird y A . I . Lawan . 1996 . Geometric correction of SPOT and Landsat imagery:
A comparison of map -and GPS- derived control points . Photogram . Engin . Rem . Sens ., 62 (10):
73-77 .
Lucas, B . D . y T . Kanade . 1981 . An iterative image registration technique with an application to stereo
vision . Proceedings 7th International Joint Conference on Artificial Intelligence (Vancouver), 2: 674-
679 .
Pérez, M ., L . Gónima y A . Pastrana . (en prensa) . Dinámica de las variaciones morfológicas de la franja
costera del DMI . En: Teledetección aplicada al ordenamiento ambiental del Distrito de Manejo
Integrado de Cispatá . Serie de Publicaciones Especiales No . 19, Instituto de Investigaciones Marinas
y Costeras-INVEMAR, Santa Marta .
Pratt, W . K . 1974 . Correlation techniques of image registration . IEEE Transact . Aerosp . Electr . Syst .-
AES, 10: 353-358 .
Sánchez, H ., G . Ulloa, H . Tavera y W . Gil . 2005 . Plan de manejo integral de los manglares de la zona de
uso sostenible del sector estuarino de la bahía de Cispatá . MVDAT, CONIF, OIMT y Corporación
Autónoma Regional de los Valles del Sinú y del San Jorge (CVS), Montería . 202 p .
Smith, D . P . y S . F . Atkinson . 2001 . Accuracy of rectification using topographic maps versus GPS ground
control points . Photogram . Engin . Rem . Sens ., 67 (5): 585-587 .
SPOTimage . 2008 . www .spotimage .com/web/es/316-satelites-spot .php . 15/01/2009 .
Wilkie, D . S . 1990 . GPS location data: an aid to satellite image analyses of poorly mapped regions . Intern .
J . Rem . Sens . 11 (4): 653-658 .
Wisetphanichkij, S ., K . Dejhan, F . Cheevasuvit, S . Mitatha, S . Hanpipatpongsa, C . Pienvijarnpong y C .
Soonyeekan . 1998 . An improvement of geometric correlation of satellite image .
www .gisdevelopment .net/aars/acrs/1998/ps3/ps3014 .asp . 25/03/2009 .
FECHA DE RECEPCIÓN: 16/06/2008 FECHA DE ACEPTACIÓN:16/12/2009


Instituto de Investigaciones Marinas y Costeras - INVEMAR 25
Bol . Invest . Mar . Cost . 39 (1) ISSN 0122-9761 Santa Marta, Colombia, 2010
DISTRIBUCIÓN Y ABUNDANCIA DE DIATOMEAS DEL GÉNERO PSEUDO-NITZSCHIA EN CONDICIÓN
DE EL NIÑO 2007 SOBRE LA CUENCA PACÍFICA COLOMBIANA*
Ana María Galeano Chavarría y Edgar Arteaga Sogamoso
Instituto de Investigaciones Marinas y Costeras-INVEMAR, sede Pacífico, A.A. 6713 N 36, Parque Científico Agronatura CIAT, Cali, Colombia. [email protected] (A.M.G.); [email protected] (E.A.S.)
RESUMEN
Se describe la distribución espacial y abundancia de diatomeas pertenecientes al género
Pseudo-nitzschia en la cuenca pacífica colombiana (CPC) durante la campaña oceanográfica PACÍFICO
XLIV-ERFEN XLII, evaluando su comportamiento frente a las variables de temperatura y salinidad . Se
realizaron muestreos de fitoplancton (con botellas Niskin de 5 L) a dos profundidades (superficial y 50
m) en 25 estaciones . Se observó que en superficie se registraron las mayores abundancias del género
distribuyéndose especialmente en la zona intermedia de la CPC . Este patrón de distribución coincidió
con valores bajos de temperatura y altos en salinidad . Debido a lo anterior, se encontraron diferencias
significativas en la abundancia de Pseudo-nitzschia entre las dos profundidades de estudio (Mann-
Whitney test, p= 0 .001) . Se estableció una relación directa entre la salinidad y la abundancia del género
(r= 0 .50; p= 0 .01), indicando la afinidad de estas algas por ambientes con altas salinidades, mientras que
la temperatura mostró un comportamiento inverso con la abundancia (r= -0 .42; p= 0 .043) . Aunque los
valores de correlación no son muy altos, la distribución espacial de variables bióticas y abióticas, sugieren
la influencia de la salinidad y temperatura en la abundancia y distribución de Pseudo-nitzschia .
PALABRAS CLAVE: Pacífico colombiano, Pseudo-nitzschia, Distribución, Abundancia, Evento El Niño .
ABSTRACT
Distribution and abundance of diatoms of the genus Pseudo-nitzschia in El Niño conditions in 2007 in the Colombian Pacific Basin. The spatial distribution and abundance of diatoms
belonging to the genus Pseudo-nitzschia in the Colombian Pacific Basin (CPB) during PACÍFICO
XLIV- ERFEN XLII oceanographic survey is described, whereas its relation to temperature and salinity
was tested . Samplings of phytoplankton (Niskin bottles 5 L) at two depths (surface and 50 m deep)
were carried out in 25 stations . The highest abundances of Pseudo-nitzschia were observed at surface,
especially clustered in the intermediate CPB zone . This spatial pattern agreed with low temperature and
25-39
*Contribución No. 1046 del Instituto de Investigaciones Marinas y Costeras – INVEMAR.

26 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
high salinity values . Consistent with previous data, there were significant differences in the abundance of
Pseudo-nitzschia between the two depths sampled (Mann-Whitney test, p= 0 .001) . A direct relationship
between salinity and the genus abundance was set (r= 0 .50; p= 0 .01), indicating the affinity of these
algae to environments with high salinities, whereas an inverse trend was showed with the temperature
(r= -0 .42; p= 0 .043) . Although the correlation values are not high, the spatial distribution of biotic and
abiotic variables, suggest the influence of salinity and temperature on the abundance and distribution of
Pseudo-nitzschia.
KEY WORDS: Colombian Pacific, Pseudo-nitzschia, Distribution, Abundance, El Niño .
INTRODUCCIÓN
El género Pseudo-nitzschia (Peragallo, H .) se halla representado
por diatomeas pennadas cosmopolitas de hábitos planctónicos marinos que se
encuentran distribuidas desde los trópicos hasta las regiones polares, así como
en ambientes oceánicos y costeros (Hasle, 1995, 2002) . Las características
morfológicas más representativas de este género son: células alargadas,
fusiformes, presentando el eje apical considerablemente mayor que los ejes
perivalvar y transapical; formación de colonias con forma escalariforme resultante
de la superposición de los extremos de las células contiguas; localización del rafe
a nivel de la superficie valvar y ausencia de poroides en las paredes del canal
(Ferrario et al ., 2002) .
En los últimos años, este género ha adquirido gran interés desde que se
ha observado que varios de sus representantes pueden ser causantes de floraciones
nocivas o mareas rojas siendo tóxicos al producir ácido domoico (AD) . Esta
neurotoxina de tipo aminoácido origina el Síndrome Amnésico por Marisco o
Amnesic Shellfish Poisoning (ASP) en organismos superiores como vertebrados
y el hombre, debido al consumo de organismos filtradores como bivalvos y
ciertos peces que tienden a acumular la toxina cuando se alimentan de estas algas
(Reguera, 2002) . El primer episodio de intoxicación asociado a este género se
presentó en 1987 en la isla Príncipe Eduardo, Canadá, ocasionando intoxicación
y muerte a varias personas (Bates et al ., 1989), así mismo, se destaca la presencia
de AD producido por Pseudo-nitzschia spp . en países como España, Japón y
Nueva Zelanda (Yasumoto et al ., 1996) . Igualmente, en el cono suramericano y
en Estados Unidos se han registrado proliferaciones tóxicas por parte de algunas
especies de este género (Negri e Inza, 1998; Trainer et al ., 1998) . En el Pacífico
colombiano, hasta el momento no hay registros de un episodio de intoxicación o
marea roja asociada a Pseudo-nitzschia .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 27
En el presente trabajo se describe la distribución y abundancia del género
Pseudo-nitzschia evaluando su relación con la temperatura y salinidad . Esto
permitirá tener los primeros registros del comportamiento del género durante
una condición El Niño en el Pacífico colombiano, sirviendo como base para la
predicción de futuros eventos de floraciones . El seguimiento de estas algas y
en general de todas aquellas productoras de toxinas, se constituye en una labor
fundamental para el manejo de los recursos pesqueros y biológicos de la región .
De acuerdo con Gómez-Aguirre et al. (2004), la importancia de Pseudo-nitzschia
se ha resaltado en diversas conferencias mundiales sobre proliferaciones algales
nocivas y es tema obligado en publicaciones realizadas sobre esta temática
(Hallegraeff et al ., 1995; Reguera et al ., 1998; Sar et al ., 2002) .
ÁREA DE ESTUDIO
La distribución espacial de Pseudo-nitzschia fue estudiada sobre la cuenca
del Pacífico colombiano (CPC), entre los paralelos 01º30’ hasta los 07º10’ de latitud
norte y entre los meridianos 77º40’ y 82º00’ de longitud oeste, donde se encuentra
preestablecida una grilla de 25 estaciones para estudios biológicos (Figura 1) y de
44 para variables oceanográficas . Esto según el diseño de cruceros oceanográficos
PACÍFICO-ERFEN realizados por el Centro Control Contaminación del Pacífico
(CCCP) . Se delimitaron tres zonas de muestreo: zona costera (estaciones a menos
de 35 millas de la costa), zona intermedia (estaciones entre 35 y 210 millas de la
costa) y zona oceánica (estaciones a más de 210 millas de la costa), ubicándose el
área de estudio en parte del área conocida como Panamá Bigth (Camacho y Pineda,
1999) . Según CCCP (2002), la CPC presenta características especiales dentro del
océano Pacífico por estar ubicada dentro de la región de bajas presiones atmosféricas
conocida como Concavidad Ecuatorial, en la cual convergen los vientos Alisios
de cada hemisferio para formar la Zona de Convergencia Intertropical (ZCIT) .
El choque de masas de aire ascendentes con diferencias térmicas y gradientes de
humedad genera la formación de una banda nubosa por proceso de convección, lo
que ocasiona que el clima se caracterice por vientos variables y débiles y por su
alta pluviosidad (CCCP, 2002) . Las variaciones en las precipitaciones dependen del
desplazamiento de las lluvias de norte a sur de la ZCIT, a lo largo del ciclo anual
sobre la costa pacífica (Prahl et al ., 1990) .

28 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
La temperatura media anual no excede 30 °C y la humedad relativa oscila entre 80 y 95 % (Garcés y Medina, 1997) . En general se presenta un clima superhúmedo y, debido a los altos niveles de pluviosidad, existe una compleja red fluvial que causa fuertes erosiones y brinda importantes aportes al mar (Forsbergh, 1969) . El patrón de corrientes superficiales en el Pacífico es complejo y responde normalmente al sistema dominante de los vientos planetarios, determinados por el desplazamiento del cinturón de convergencia intertropical . Contrario a lo que se cree, las corriente del Perú y de Humboldt, que son corrientes frías subantárticas, no tienen efecto sobre la ensenada de Panamá (Cantera, 1993) .
MATERIALES Y MÉTODOS
Diseño y procesamiento de muestrasEn cada estación se realizaron muestreos de fitoplancton mediante colectas
de agua con botellas Niskin de 5 L, enviadas a nivel superficial (0 m) y a 50 m de profundidad . Una vez en el buque, se obtuvieron 500 mL de la muestra original y se fijaron con formalina hasta obtener una concentración final aproximada de 5 % . Las
Figura 1. Estaciones del crucero oceanográfico PACÍFICO XLIV-ERFEN XLII sobre la cuenca del Pacífico colombiano, efectuado entre enero 24 y febrero 19 de 2007 . Se muestra la delimitación por zonas: A) costera, B) intermedia y C) oceánica .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 29
muestras obtenidas fueron trasladadas al laboratorio del Instituto de Investigaciones Marinas y Costeras–INVEMAR (Pacífico), para su procesamiento y análisis. Simultáneamente, en cada estación, se registraron mediciones de temperatura (oC) y salinidad en las profundidades de muestreo usando un perfilador CTD, Seabird 19; estos datos fueron colectados y suministrados a este estudio por el CCCP .
Para la determinación cuantitativa se empleó la metodología de Uthermöhl, extrayendo de cada muestra previamente homogenizada, alícuotas de 50 mL que se depositaron en cámaras de sedimentación por un tiempo de 72 horas . Posteriormente, fueron observadas bajo microscopio óptico invertido realizando el conteo de los organismos de interés mediante barrido total o parcial del fondo de la cámara . Dependiendo de la densidad de los mismos, se determinó el número de organismos presentes en el volumen de muestra analizado y se extrapoló al volumen total para establecer la concentración de células por litro (cel L-1), teniendo en cuenta la metodología para la cuantificación del fitoplancton expuesta por Villafañe y Reid (1995) .
Análisis de datosSe realizó un análisis descriptivo y exploratorio de las abundancias obtenidas
en cada estación mediante pruebas de normalidad y ensayos de transformación con el fin de determinar el tipo de estadística a emplear . De acuerdo con lo anterior, se utilizó el test no paramétrico U de Mann-Whitney para comparar los valores de abundancia entre las profundidades estudiadas (superficial y 50 m), y entre los muestreos realizados durante el día y la noche . Asimismo, se determinó el grado de asociación entre la abundancia de Pseudo-nitzschia y las variables abióticas mediante el coeficiente Spearman (r
s) . Para los análisis antes mencionados se tuvo
en cuenta un nivel de significancia estadística de 0 .05 .
RESULTADOS
De acuerdo con los resultados obtenidos por el CCCP durante la campaña oceanográfica PACÍFICO XLIV-ERFEN XLII sobre la CPC, entre los meses de enero y febrero de 2007 a nivel superficial, los valores de temperatura tuvieron un incremento hacia el costado noroccidental de la cuenca, siendo más notorio en las estaciones cercanas a la costa, ubicadas frente a Chocó . Este calentamiento también se hizo evidente en la columna de agua, provocando termoclinas más profundas, característica típica de la acción de ondas Kelvin, sugiriendo que para esta época aún se presentaron condiciones influenciadas por el evento El Niño 2006-2007 . Por otro lado, también se observó una disminución de la temperatura hacia la zona centro ocasionada por la presencia de un núcleo de temperaturas por debajo de 26 .5 °oC localizado sobre 4oN y 80oW, provocado por la aparición del jet del viento de

30 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Panamá, característico para esta época . Este proceso influyó en la dinámica de las corrientes en esa zona y redujo el calentamiento originado por El Niño sobre la temperatura superficial del mar . La temperatura mínima encontrada para el área fue 25 .3 °oC y la máxima 30 °C, con un ámbito de variación de 4 .7 oC . El promedio para la zona fue 27.7°oC .
La salinidad presentó un ámbito de variación alto de 7 .11 (mínimo: 26 .60, máximo: 33 .71), con un promedio de 31 .76 para toda la cuenca . Los mayores valores se registraron hacia el costado suroccidental de la CPC, mientras que los menores se localizaron hacia el noroccidente y hacia la zona costera, especialmente en la bahía de Tumaco, Buenaventura y en la región norte de Chocó .
Durante el presente estudio, Pseudo-nitzschia fue uno de los géneros que se caracterizaron por presentar mayores valores de abundancia entre las diatomeas y dinoflagelados registrados tanto en superficie (Figura 2A) como a 50 m de profundidad (Figura 2B), siendo superado sólo por el género Chaetoceros . Pseudo-nitzschia también se destacó por estar presente en 92 % de las estaciones en superficie y en 79 % de las estaciones a 50 m de profundidad . A nivel superficial, la abundancia de Pseudo-nitzschia fluctuó entre 38 y 265246 cel·L-1, con promedio de 21964 cel·L-1
y mediana de 4606 cel·L-1 (Tabla 1) . La distribución de las abundancias fue agregada en la zona intermedia de la cuenca y en dos estaciones oceánicas (Figura 3) . Por otra parte, los representantes del género se encontraron dentro del ámbito de temperatura entre 25 .39 y 29.83 °C y entre 28 .68 y 33 .2 de salinidad (Figura 4) . A 50 m de profundidad la abundancia del género varió entre 38 y 6867 cel·L-1, con promedio de 748 y mediana de 247 cel·L-1 (Tabla 1) . La distribución de la abundancia fue más homogénea, con una tendencia mayor hacia la zona intermedia de la CPC; sin embargo, el valor más alto de abundancia se presentó en la estación oceánica 107 (Figura 5) . La escala de temperatura donde se halló Pseudo-nitzschia fue entre 15 .97 y 21.19 °C y la salinidad estuvo comprendida entre 34 .3 y 35 .0 a esta profundidad .
Figura 2. Porcentaje de abundancia de Pseudo-nitzschia respecto a otros grupos dominantes del fitoplancton (diatomeas y dinoflagelados) para: A) superficie y B) 50 m de profundidad .
45 %
18 %
9 %
7 %
7 %
6 % 4 % 2 % 2 %
Chaetoceros
Hemiaulus
Pseudo-nitzschia
Dactyliosolem
Otros
Cylindrotheca
Leptocylindrus
Detonula
Guinardia
A B
59 %21 %
6 %4 %
3 % 2 % 2 % 2 % 1 %
Otros
Thalassionema
Chaetoceros
Leptocylindrus
Pseudo-nitzschia
Rhizosolenia
Cylindrotheca
Guinardia
Hemiaulus

Instituto de Investigaciones Marinas y Costeras - INVEMAR 31
Est
ació
nU
bica
ción
geo
gráfi
caA
bund
anci
a su
perfi
cial
(c
el•L
-1)
Abu
ndan
cia
a 5
0 m
(cel
•L-1)
Tem
pera
tura
su
perfi
cial
(ºC
)
Salin
idad
su
perfi
cial
Tem
pera
tura
a
50 m
(º
C)
Sal
inid
ad
a 50
m
E00
106
°30’
N77
°45’
W38
027
.81
30 .5
017
.53
34 .7
9
E00
305
°30’
N77
°45’
W63
0091
127
.37
29 .9
818
.47
34 .7
3
E00
504
°30’
N77
°45’
W65
3611
427
.72
30 .6
518
.04
34 .7
5
E00
703
°30’
N77
°45’
W42
8711
428
.03
28 .6
818
.43
34 .8
4
E01
006
°00’
N78
°00’
W49
250
28 .0
529
.96
17 .7
934
.75
E01
205
°00’
N78
°00’
W17
0615
127
.41
30 .8
418
.99
34 .6
8
E01
404
°00’
N78
°00’
W68
20
27 .0
031
.77
17 .8
934
.80
E01
603
°00’
N78
°00’
W41
070
26 .9
330
.48
18 .5
134
.79
E02
506
°00’
N79
°00’
W28
043
1229
27 .0
831
.78
17 .0
534
.84
E02
705
°00’
N79
°00’
W19
203
9526
.85
31 .3
817
.40
34 .7
7
E02
904
°00’
N79
°00’
W23
929
3826
.44
32 .0
117
.34
34 .9
2
E03
103
°00’
N79
°00’
W30
341
626
.28
31 .7
218
.78
34 .8
2
E03
302
°00’
N79
°00’
W11
312
1026
.67
31 .6
918
.83
34 .9
6
E04
305
°00’
N80
°00’
W54
2634
226
.80
31 .9
516
.38
34 .9
2
E04
504
°00’
N80
°00’
W26
5246
1934
25 .3
932
.88
16 .9
934
.89
E04
703
°00’
N80
°00’
W42
243
2001
26 .7
032
.91
17 .9
435
.00
E07
505
°00’
N82
°00’
W24
638
26 .8
532
.06
18 .4
334
.51
Tab
la 1
. Val
ores
de
abun
danc
ia (
cel•
L-1),
tem
pera
tura
(o C
) y
salin
idad
reg
istr
ados
a n
ivel
sup
erfi
cial
y a
50
m d
e pr
ofun
dida
d du
rant
e el
cru
cero
PA
CÍF
ICO
XL
IV-
ER
FEN
XL
II r
ealiz
ado
entr
e en
ero
24 y
feb
rero
19
de 2
007
en la
cue
nca
del P
acíf
ico
colo
mbi
ano .
Los
dat
os d
e te
mpe
ratu
ra y
sal
inid
ad f
uero
n su
min
istr
ados
por
el
CC
CP
(200
7) .

32 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Con
tinua
ción
Tab
la 1
.
Est
ació
nU
bica
ción
geo
gráfi
caA
bund
anci
a su
perfi
cial
(c
el•L
-1)
Abu
ndan
cia
a 5
0 m
(cel
•L-1)
Tem
pera
tura
su
perfi
cial
(ºC
)
Salin
idad
su
perfi
cial
Tem
pera
tura
a
50 m
(º
C)
Sal
inid
ad
a 50
m
E07
704
°00’
N82
°00’
W85
30
27 .7
632
.67
16 .3
934
.87
E07
903
°00’
N82
°00’
W67
1857
26 .5
833
.06
15 .9
734
.90
E08
102
°00’
N82
°00’
W62
088
624
27 .0
433
.20
16 .5
234
.92
E10
705
°00’
N84
°00’
W0
6867
29 .8
330
.60
21 .1
934
.30
E10
904
°00’
N84
°00’
W0
456
29 .4
330
.22
17 .4
834
.72
E11
103
°00’
N84
°00’
W92
460
627
.91
32 .6
316
.37
34 .8
7
E11
302
°00’
N84
°00’
W43
230
739
27 .
32 3
3 .13
16 .
36 3
4 .98
Tot
al
52
7146
179
42
Pro
med
io21
964
748
Med
iana
4606
247
Máx
imo
2652
4668
6729
.83
33 .2
021
.19
35 .0
0
Mín
imo
0 0
25 .
39 2
8 .68
15 .
97 3
4 .30

Instituto de Investigaciones Marinas y Costeras - INVEMAR 33
Figura 3. Distribución espacial de las abundancias (cel•L-1) de Pseudo-nitzschia encontrada a nivel superficial en la cuenca del Pacífico colombiano. Crucero PACÍFICO XLIV-ERFEN XLII realizado entre enero 24 y febrero 19 de 2007 .
Figura 4. Dispersión de las abundancias de Pseudo-nitzschia respecto a la temperatura (oC) y la salinidad encontradas en la cuenca del Pacífico colombiano entre enero y febrero de 2007 . El tamaño de las circunferencias corresponde a los valores de abundancia (cel•L-1) en orden de magnitud . Las líneas indican el límite mínimo de salinidad y máximo de temperatura (oC) más allá del cual no se encontraron individuos pertenecientes a Pseudo-nitzschia en este estudio .

34 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
No se encontraron diferencias significativas en la abundancia de las estaciones
entre los muestreos diurnos y nocturnos (Mann-Whitney test, p= 0 .578); sin embargo a
nivel de profundidad (superficial y 50 m) sí se establecieron diferencias significativas
(Mann-Whitney test, p= 0 .001) . En superficie, la correlación de la abundancia con
la temperatura fue negativa (rs=-0 .42; p= 0 .043), mientras que con la salinidad fue
positiva (rs= 0 .50; p= 0 .01) . A la profundidad de 50 m, no se determinó correlación
significativa para estas dos variables (Spearman, p> 0 .05) .
DISCUSIÓN
El presente trabajo es el primero en realizar un análisis de la abundancia y
distribución del género Pseudo-nitzschia a lo largo de la CPC a principio de año y en
un periodo considerado aún como de influencia de El Niño, por lo que las condiciones
ambientales y por ende el comportamiento de éstos podrían ser considerados como
especiales para el periodo estudiado . Como característica particular en los resultados
fue el predominio en abundancia de Pseudo-nitzschia y, en general, de las diatomeas
Figura 5. Distribución espacial de las abundancias (cel•L-1) de Pseudo-nitzschia encontradas a 50 m de profundidad en la cuenca del Pacífico colombiano. Crucero PACÍFICO XLIV-ERFEN XLII realizado entre enero 24 y febrero 19 de 2007 .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 35
respecto a los dinoflagelados, a pesar de presentarse aún una condición El Niño . En
esta condición se esperaría, de acuerdo con los resultados de otros autores (Avaria,
1984; Constain y Delgado, 1985; Ochoa et al ., 1985; Castillo y Vizcaíno, 1992; Osorio
y Castillo, 1993; Vizcaíno, 1993; Mendoza, 1996; García, 2001; CCCP, 2002), que
estos últimos fueran predominantes o, por lo menos, más notorios . Lo anterior podría
deberse, para el caso del Pacífico colombiano, a la diferencia en la época en que se
ejecutó el presente muestreo (a principios de año) con respecto al realizado por dichos
autores (final de año), en donde para el primer trimestre del año el incremento de
la corriente del golfo de Panamá por la acción de los vientos Alisios del norte y del
chorro de vientos de Panamá, así como la presencia de un gran giro ciclónico, son
responsables de la presencia de aguas frías y ricas en nutrientes (Rodríguez-Rubio,
2007) y por lo tanto de mayor productividad primaria en la CPC, favoreciendo el
desarrollo de diatomeas, dentro de las que se destacó Pseudo-nitzschia.La distribución horizontal de las mayores abundancias de Pseudo-nitzschia
encontradas a nivel superficial en la zona intermedia de la cuenca (estaciones 45 y
47) coincidió con los menores registros de temperatura y los mayores de salinidad
para el área y época evaluada, así como con la ubicación del giro ciclónico . Este
comportamiento pondría de manifiesto la preferencia de estas algas hacia aguas frías
y de altas salinidades, confirmado por las tendencias en las correlaciones entre la
abundancia de Pseudo-nitzschia y estos dos factores . Conductas similares han sido
observadas por varios autores en diferentes regiones: Trainer et al . (2000) y Thessen
(2007) para las costas de Estados Unidos; Caroppo et al . (2005) para el mar Adriático .
Para el caso específico de la temperatura, comportamientos afines fueron registrados
por Dortch et al. (1997) para el golfo de México y por Buric et al . (2008) para el
mar Adriático . Respecto a la salinidad, Thessen et al . (2005) sugieren que este es
un factor que puede controlar el crecimiento, desarrollo y distribución de Pseudo-
nitzschia, desarrollándose mejor en aguas de salinidades altas . Sin embargo, ciertas
especies del género pueden tolerar también bajas salinidades en el medio marino
(Thessen et al ., 2005) . Las escalas de temperatura y salinidad en las que se encontraron
estos organismos fueron amplias a nivel superficial (Tabla 1), ámbito que se extiende
aún más si se tienen en cuenta los registros de Pseudo-nitzschia encontrados a 50
m de profundidad; esto demuestra la capacidad eurihalina y euriterma de Pseudo-
nitzschia, descrita por Thessen et al. (2005) . No obstante, es importante anotar que
para este trabajo se desconoce el número de especies o ecotipos involucrados en este
comportamiento, los cuales estarían adaptados a las diversas condiciones ambientales
encontradas para la época analizada .

36 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
A pesar de las diferencias en abundancia encontradas entre las
profundidades analizadas, es probable que existan otros factores ambientales,
diferentes a temperatura y salinidad, que pudieron haber favorecido la presencia
de una mayor densidad de Pseudo-nitzschia en superficie . Por otro lado, el no
encontrar diferencias significativas entre las abundancias de las estaciones, cuyas
muestras fueron tomadas durante el día con las tomadas durante la noche, podría
indicar que el comportamiento arriba observado no estaría influenciado por algún
tipo de migración nictimeral dentro de la columna de agua .
La gran densidad registrada en algunas de las estaciones evaluadas,
especialmente en la zona intermedia de la cuenca, fue superior a lo encontrado
por Vargas y Freer (2004) para el golfo de Nicoya, Costa Rica, quienes registraron
un caso de floración por Pseudo-nitzschia pungens, una especie potencialmente
tóxica con densidades aproximadas de 105 cel L-1, considerando estos valores como
significativos y advirtiendo sobre la atención que se le debe dar a este tipo de eventos .
A pesar de las altas densidades encontradas, no se presentaron registros de algún tipo
de floración durante el desarrollo de este estudio, lo que no permite afirmar o negar
que tal proliferación haya sido nociva, bien porque no tuvieron características de
toxicidad o porque se presentaron en áreas lejanas de la costa, que no permitieron
conocer sus efectos . Aunque la identificación taxonómica realizada sólo fue posible
hasta género, no se debe descartar la posible presencia de especies potencialmente
tóxicas en la CPC, si se tiene en cuenta que aproximadamente una de cada tres
especies del género tienen esta capacidad (Bailey, 2008) .
Es importante mencionar que, para futuros estudios con especies de
Pseudo-nitzschia, es necesario contar con herramientas que permitan una correcta
identificación, como el uso de microscopía electrónica de barrido o de transmisión,
ya que muchas de las especies comparten características que son difíciles de
observar con microscopio óptico (MO) convencional . Sin embargo, no se puede
dejar de lado el uso de MO como primer paso para una identificación aproximada
de los organismos (Ferrario et al ., 2002) . Recientemente se está utilizando la
genética molecular como herramienta para determinar las diferencias o similitudes
en cuanto a su conformación genética (Bailey, 2008) . Finalmente, este estudio se
constituye en el primer antecedente sobre la ecología de Pseudo-nitzschia en el
Pacífico colombiano, para la época y el área estudiada, lo que permitió concluir
que los valores de abundancia en algunas de las estaciones superaron densidades de
105 cel L-1, y que la distribución se dio hacia la zona intermedia de la cuenca, lugar
en donde se presentó un núcleo de bajas temperaturas y altas salinidades, en este

Instituto de Investigaciones Marinas y Costeras - INVEMAR 37
sentido se pudo comprobar la afinidad de Pseudo-nitzschia por este tipo ambientes,
corroborando otros estudios realizados en diferentes partes del mundo . Considerando
esto, es importante seguir avanzando en estudios con Pseudo-nitzschia, así como
de otras especies fitoplanctónicas consideradas como potencialmente nocivas, para
determinar de una manera más clara y precisa los efectos que puedan estar causando
estos organismos al resto de especies marinas en el Pacífico colombiano .
AGRADECIMIENTOS
Los autores expresan su agradecimiento al Instituto de Investigaciones
Marinas y Costera “José Benito Vives de Andréis”-INVEMAR por el apoyo
brindado durante la realización de este trabajo . Este artículo hace parte de la tesis
titulada “Caracterización espacial de la abundancia, composición y diversidad del
fitoplancton de la Zona Oriental del Pacífico colombiano durante el evento El Niño
2006-2007” presentada por A. Galeano a la Universidad del Valle sede Pacífico
como requisito para optar al título de Biólogo . De igual manera, se extiende este
agradecimiento a la Armada Nacional de Colombia, la Dirección General Marítima-
DIMAR y al Centro Control Contaminación del Pacífico-CCCP, por la invitación
extendida al INVEMAR para participar de la campaña oceanográfica PACÍFICO
XLIV-ERFEN XLII, así como por la información suministrada . Finalmente, a
Beatriz Herrera por la realización de los mapas que aquí se presentan
BIBLIOGRAFÍA
Avaria, S . 1984 . Cambios en la composición y biomasa del fitoplancton marino del norte de Chile durante
el fenómeno El Niño 1982-1983 . Rev . Com . Per . Pac . Sur, 15: 303-309 .
Bailey, A . 2008 . Pseudo-nitzschia species distributions in Glacier Bay, AK as measured by automated
ribosomal intergenic spacer analysis (ARISA) . http://hdl .handle .net/1773/3745 . 20/07/2008 .
Bates, S . S ., C . J . Bird, A . Freitas, R . Foxall, M . Gilgan, L . A . Hanic, G . R . Johnson, A . W . McCulloch,
P . Odense, R . Pocklington, M . Quilliam, J . Sim, J . C . Smith, D . V . Subba Rao, E . C . D . Todd, J . A .
Walter y J . L . C . Wright . 1989 . Pennate diatom Nitzschia pungens as the primary source of domoic
acid, a toxin in shellfish from eastern Prince Edward Island, Canada . Can . J . Fish . Aquat . Sci ., 46:
1203-1215 .
Buric, Z ., D . Vilicic, K . C . Mihalic, M . Caric, K . Kralj y N . Ljubesic . 2008 . Pseudo-nitzschia blooms in
the Zrmanja River estuary (Eastern Adriatic Sea) . Diatom Res ., 23: 51-63 .
Camacho, G. y R. Pineda. 1999. Datos oceanográficos del crucero PACÍFICO XXI-ERFEN XXVII.
Mayo/98, Buque ARC Malpelo . Armada Nacional, Dirección General Marítima . Informe técnico,
Centro Control Contaminación del Pacífico, Tumaco, Colombia . 38 p .

38 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Cantera, J . R . 1993 . Oceanografía . 13-23 . En: Leyva, P . (Ed .) . Colombia Pacífico . Tomo I . Fondo de
Protección del Medio Ambiente “José Celestino Mutis”-FEN, Bogotá. 396 p.
Caroppo, C ., R . Congestri, L . Bracchini y P . Albertano . 2005 . On the presence of Pseudo-nitzschia
calliantha Lundholm, Moestrup et Hasle and Pseudo-nitzschia delicatissima (Cleve) Heiden in the
Southern Adriatic Sea (Mediterranean Sea, Italy) . J . Plank . Res ., 27: 763-774 .
Castillo, L . y A . Vizcaíno . 1992 . Los indicadores biológicos del fitoplancton y su relación con el fenómeno
El Niño 1991-1992 en el Pacífico colombiano . Bol . Cient . CIOH, 12: 13-22 .
CCCP . 2002 . Compilación oceanográfica de la cuenca pacífica colombiana . Serie de Publicaciones
Especiales Vol . 1, Centro Control Contaminación del Pacífico-CCCP, DIMAR, Tumaco, Colombia .
168 p .
Constain, L . y L . Delgado . 1985 . Dinoflagelados del Pacífico colombiano como indicadores del fenómeno
El Niño, Crucero PACÍFICO-VIII-ERFEN V, Nov-Dic.1982. Tesis Biol. Mar., Univ. Jorge Tadeo
Lozano, Bogotá . 44 p .
Dortch, Q ., R . Robichaux, S . Pool, D . Milsted, G . Mire, N . N . Rabalais, T . M . Soniat, G . A . Fryxell, R .
E . Turner y M . L . Parsons . 1997 . Abundance and vertical flux of Pseudo-nitzschia in the northern
Gulf of Mexico . Mar . Ecol . Prog . Ser ., 146: 249-264 .
Ferrario, M . E ., E . A . Sar y S . E . Sala . 2002 . Diatomeas potencialmente toxígenas del cono Sur Americano .
167-194 . En: Sar, E . A ., M . E . Ferrario y B . Reguera (Eds .) . Floraciones algales nocivas en el cono
Sur Americano . Instituto Español de Oceanografía, Madrid . 311 p . Forsbergh, E . D . 1969 . On the climatology, oceanography and fisheries of the Panama Bight . Bull . Inter .
Am . Trop . Tuna Comm ., 14 (2): 49-365 .
Garcés, L . y L . Medina . 1997 . Evaluación de las especies fitoplanctónicas reportadas como indicadores
biológicos para el fenómeno de El Niño en el Pacífico colombiano . Cruceros ERFEN 1978-1994 .
Tesis Biol . Mar ., Univ . Jorge Tadeo Lozano, Bogotá . 63 p .
García, I . 2001 .Efecto de los eventos El Niño y La Niña sobre las comunidades de fitoplancton al interior
de la ensenada de Tumaco (Colombia) . Bol . Cient . CCCP, 8: 12-25 .
Gómez-Aguirre, S ., S . Licea y S . Gómez . 2004 . Proliferaciones de Pseudo-nitzschia y otras especies del
microplancton en la bahía de Mazatlán, México . Rev . Biol . Trop ., 52 (Suppl . 1): 69-76 .
Hallegraeff, G ., D . Anderson y A . Cembella (Eds .) . 1995 . Manual on harmful marine microalgae . IOC
Manuals and Guides No . 33, UNESCO, París . 551 p .
Hasle, G . R . 1995 . Pseudo-nitzschia pungens and P. multiseries (Bacillariophyceae) - nomenclatural
history, morphology, and distribution . J . Phycol ., 31: 428-435 .
Hasle, G . R . 2002 . Are most of the domoic acid-producing species of the diatom genus Pseudo-nitzschia
cosmopolites? Harmful Algae, 1: 137-146 .
Mendoza, A . 1996 . Fitoplancton del área comprendida entre Caleta Caracas y La Muerte, Bahía Málaga,
Pacífico colombiano . Tesis Biol ., Univ . del Valle, Cali . 37 p .
Negri, R . y D . Inza . 1998 . Some potentially toxic species of Pseudo-nitzschia in the Argentine sea (35º-39º
S) . 84-85 . En: Reguera, B ., J . Blanco, M . Fernández y T . Wyatt (Eds .) . Harmful algae . Xunta de
Galicia and Intergovernmental Oceanographic Commission of UNESCO, Vigo, España . 635 p .
Ochoa, N ., B . Rojas y O . Gómez . 1985 . Identificación del fenómeno El Niño a través de los organismos
fitoplanctónicos. 23-31. En: Arntz, W. E., A. Landa y J. Tarazona (Eds.). El Fenómeno “El Niño” y
su impacto en la fauna marina . Bol . Inst . Mar Perú-Callao, volumen extraordinario: 23-31 .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 39
Osorio, D ., y F . Castillo . 1993 . Relación climatológica de la temperatura superficial del Pacífico
colombiano sobre el fitoplancton marino durante El Niño 1991-92 . Bol . Cient . CIOH, 14: 131-144 .
Prahl, H .v ., J . Cantera y R . Contreras .1990 . Manglares y hombres del Pacífico colombiano . Fondo FEN-
Colciencias, Bogotá . 139 p .
Reguera, B . 2002 . Establecimiento de un programa de seguimiento de microalgas tóxicas . 19-54 . En: Sar,
E . A ., M . E . Ferrario y B . Reguera (Eds .) . Floraciones algales nocivas en el Cono Sur Americano .
Instituto Español Oceanografía, Madrid . 311 p .
Reguera, B ., J . Blanco, M . Fernádez y T . Wyatt (Eds .) . 1998 . Harmful algae . Xunta de Galicia and
Intergovernmental Oceanographic Commission of UNESCO, Vigo, España . 635 p .
Rodríguez-Rubio, E . 2007 . El Niño y La Niña en la cuenca pacífica colombiana, situación actual . http://
www .cccp .org .co/modules .php?name=News&file=article&sid=269 . 20/08/2007 .
Sar, A ., M . Ferrairo y B . Reguera (Eds .) . 2002 . Floraciones algales nocivas en el Cono Sur Americano .
Instituto Español de Oceanografía, Madrid . 311 p .
Thessen, A . E . 2007 . Taxonomy and ecophysiology of Pseudo-nitzschia in the Chesapeake Bay . Tesis
doctoral, University of Maryland, College Park, EE . UU . 231 p .
Thessen, E ., Q . Dortch, L . Parsons y W . Morrison . 2005 . Effect of salinity on Pseudo-nitzschia species
(Bacillariophyceae) growth and distribution . J . Phycol ., 41: 21-29 .
Trainer, V ., J . Wekell, R . Horner, C . Hatfield y J . Stein . 1998 . Domoic acid production by Pseudo-
nitzschia pungens . 337-339 . En: Reguera, B ., J . Blanco, M . Fernández y T . Wyatt (Eds .) . Harmful
algae . Xunta de Galicia and Intergovernmental Oceanographic Commission of UNESCO, Vigo,
España . 635 p .
Trainer, V ., N . Adams, B . Bill, C . Stehr, J . Wekell, P . Moeller, M . Busman y D . Woodruff . 2000 . Domoic
acid production near California coastal upwelling zones, June 1998 . Limnol . Oceanog ., 45: 1818-
1833 .
Vargas, M . y E . Freer . 2004 . Proliferaciones algales de la diatomea toxigénica Pseudo-nitzschia
(Bacillariophyceae) en el Golfo de Nicoya, Costa Rica . Rev . Biol . Trop ., 52 (1): 127-132 .
Villafañe, V . E . y F . M . Reid . 1995 . Métodos de microscopía para la cuantificación del fitoplancton . 170-
185 . En: Alveal, K ., M . Ferrario, E . de Oliveira y E . Sar (Eds .) . Manual de métodos ficológicos .
Univ . Concepción, Concepción, Chile . 863 p .
Vizcaíno, Z . 1993 . Fitoplancton del Pacífico colombiano como indicador del fenómeno El Niño . Tesis .
Biol ., Univ . Valle, Cali . 64 p .
Yasumoto, T ., Y . Oshima y Y . Fukuyo . 1996 . Proceedings of the seventh international conference on toxic
phytoplankton, harmful and toxic algal blooms . Intergovernmental Oceanographic Commission,
UNESCO, Sendai, Japón . 586 p .
FECHA DE RECEPCIÓN: 02/09/2008 FECHA DE ACEPTACIÓN: 15/03/2010


Instituto de Investigaciones Marinas y Costeras - INVEMAR 41
Bol . Invest . Mar . Cost . 39 (1) ISSN 0122-9761 Santa Marta, Colombia, 2010
EFECTO DE LA SEDIMENTACIÓN EN EL RECLUTAMIENTO DE LAS MACROALGAS DICTYOTA
SPP. Y LOBOPHORA VARIEGATA: UN ESTUDIO EXPERIMENTAL EN EL PARQUE NACIONAL NATURAL
TAYRONA, CARIBE COLOMBIANO*
Lennin Flórez-Leiva1, Alejandro Rangel-Campo2, Martha Díaz-Ruiz3, Dagoberto E. Venera-Pontón2 y
Guillermo Díaz-Pulido4
1 Universidad de Concepción, Departamento de Oceanografía, Chile. [email protected] Universidad del Magdalena, Instituto de Investigaciones Tropicales-INTROPIC, Carrera 32 No 22-08, Santa Marta, Colombia. [email protected] (A.R.C.); [email protected] (D.E.V.P.)3 Instituto de Investigaciones Marinas y Costeras-INVEMAR, A.A. 1016, Santa Marta, Colombia. [email protected] Griffith University, Griffith School of Environment y ARC Centre of Excellence for Coral Reef Studies, Brisbane, Nathan 4111, QLD, Australia. [email protected]
RESUMEN
La sedimentación está frecuentemente implicada como un factor clave en la degradación arrecifal . Sin embargo son pocos los estudios experimentales que evalúan este importante proceso en la dinámica del reclutamiento de las algas en los arrecifes coralinos . En un experimento de campo se manipularon los niveles de sedimentación y se examinaron los impactos en el reclutamiento y crecimiento de las macroalgas pardas Dictyota spp . y Lobophora variegata en dos bahías (Chengue y Granate) del Parque Nacional Natural Tayrona, Caribe colombiano . Se encontró una gran variabilidad de respuestas de las algas a la sedimentación, la cual dependió del nivel de sedimento empleado, tipo de alga estudiada y parámetro poblacional considerado (reclutamiento o crecimiento) . La adición de sedimentos tuvo generalmente un efecto negativo en el reclutamiento y crecimiento de Dictyota spp . Contrariamente, la adición de sedimentos no afectó el reclutamiento de L. variegata, aunque la remoción de sedimentos sí lo hizo . La bahía con mayor tasa de sedimentación (Granate) presentó en general menores valores de reclutamiento y crecimiento de ambas algas que la otra (Chengue) . Estos resultados indican que existe una gran complejidad y variabilidad en las respuestas de las algas arrecifales a los efectos de sedimentación . Sugieren también que la degradación arrecifal tiene el potencial de modificar la dinámica poblacional de los estadios tempranos de vida de las algas . Esto último puede tener implicaciones importantes para el entendimiento ecológico de la degradación arrecifal .
PALABRAS CLAVE: Caribe colombiano, Arrecifes coralinos, Macroalgas, Parque Nacional Natural Tayrona, Sedimentación .
41-56
*Contribución No. 1047 del Instituto de Investigaciones Marinas y Costeras – INVEMAR.

42 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
ABSTRACT
Effects of sedimentation on the recruitment of the macroalgae Dictyota spp. and Lobophora variegata: an experimental study in the Tayrona National Natural Park, Colombian Caribbean. Sedimentation is frequently associated with coral reef degradation . However, there are few experimental studies evaluating the impacts of this process on the dynamics of algal recruitment on coral reefs . In a field experiment, we manipulated the levels of sedimentation and examined the impacts on the recruitment and growth of the brown macroalgae Dictyota spp . and Lobophora variegata, in two localities (Chengue and Granate) on the Tayrona National Natural Park, Colombian Caribbean . We found considerable variability in the responses of algae to sedimentation . This variability depended on the levels of sediments used, the type of algae employed and the population parameter considered (recruitment or growth) . Sediment addition generally had a negative effect on the recruitment and early growth of the alga Dictyota spp . In contrast, sediment addition did not affect the recruitment of L. variegata, while sediment removal had a negative effect on recruitment . The locality with higher sedimentation rates (Granate) generally had lower recruitment and growth of both algae than that of the other locality with lower sedimentation levels (Chengue) . Our study indicates that there is substantial variability and complexity in the responses of reef algae to the effects of sedimentation . The results also suggest that the process of reef degradation can modify early population dynamics of benthic algae, with important implications for understanding the ecology of reef degradation .
KEY WORDS: Colombian Caribbean, Coral reefs, Macroalgae, Tayrona National Natural Park, Sedimentation .
INTRODUCCIÓN
La degradación de los arrecifes coralinos se ha incrementado
considerablemente en los últimos años debido al deterioro de la calidad del agua
(eutroficación e incrementos de sedimentación), sobrepesca, enfermedades de
herbívoros y corales, blanqueamiento coralino producido por el calentamiento
global y acidificación de las aguas oceánicas (Jackson et al ., 2001; Hoegh-Guldberg
et al ., 2007) . Estos procesos de degradación pueden conducir a la pérdida de corales
duros y al incremento de las macroalgas, trayendo como consecuencia un cambio
de estado ecológico en los arrecifes (Done, 1992; Hughes, 1994; McCook, 1999;
Hughes et al ., 2003) .
Estos cambios de estado pueden ser negativos para el ecosistema,
ya que reducen las tasas de acreción arrecifal y empobrecen la biodiversidad
marina, además de traer consigo pérdidas económicas importantes (Hughes,
1994; Grimsditch y Salm, 2006) . Los cambios de estado pueden además alterar
la estructura de los ensamblajes algales, modificando la dominancia y diversidad
algal . Al presentarse estas alteraciones se pueden afectar procesos importantes en
los que participan las macroalgas como, por ejemplo, la productividad primaria,
interacciones ecológicas con los corales y la acreción de los arrecifes coralinos
(debido a la reducción de la abundancia de algas calcáreas) . Además, pueden

Instituto de Investigaciones Marinas y Costeras - INVEMAR 43
reducirse los hábitats para diversas especies de importancia ecológica y comercial
(Jensen, 1993; Adey, 1998; Bula-Meyer, 2004) .
La dinámica del reclutamiento de algas es un proceso crítico durante su
colonización e invasión en los arrecifes coralinos . Para que ocurra la colonización
algal se requiere que esporas y propágulos se asienten y recluten en el sustrato
duro y que muchos juveniles sobrevivan hasta alcanzar los estadios adultos para
sostener la dominancia . Las macroalgas pueden responder de manera diferente a
las perturbaciones antropogénicas dependiendo de los estadios de vida . Algunos
trabajos han evaluado los efectos de la sedimentación en las algas adultas
(Eriksson y Johansson, 2003, 2005); sin embargo, los efectos de la sedimentación
sobre los estadios tempranos de las macroalgas han recibido muy poca atención .
A pesar de la importancia de las algas en la ecología arrecifal y del reclutamiento
como proceso poblacional y ecológico, es muy poco lo que se conoce acerca de la
dinámica del reclutamiento de algas en arrecifes de coral y los procesos y factores
que la regulan (Díaz-Pulido y McCook, 2004) .
Entre los procesos de gran importancia en la degradación arrecifal, a
nivel local, se encuentra el incremento de la tasa de sedimentación marina, debida
principalmente a las actividades humanas tales como agricultura, desarrollo costero,
construcción, minería, dragado y deforestación . Estas actividades, especialmente
la deforestación de los bosques para generar zonas ganaderas o agrícolas,
colectar madera, realizar construcciones y producir carbón vegetal, modifican las
propiedades físicas y químicas del suelo, posibilitando los procesos de erosión
y un consecuente incremento de la sedimentación costera . La sedimentación
constituye una preocupación importante porque afecta a un gran número de
arrecifes coralinos y ha sido relacionada con su salud (Bruijnzeel, 1990; Rogers,
1990; Richardson, 1998) . De esta forma, el objetivo de este estudio fue investigar
los efectos de los incrementos de la sedimentación sobre el desarrollo temprano de
las poblaciones de las algas Dictyota spp . y Lobophora variegata en dos bahías del
Parque Nacional Natural Tayrona .
ÁREA DE ESTUDIO
Bahías de Chengue y GranateChengue es una pequeña bahía (3 .3 km2 de superficie) localizada dentro del
Parque Nacional Natural Tayrona (PNNT, Figura 1) a unos 14 km al nororiente de la ciudad de Santa Marta, en la costa Caribe de Colombia (11º20’N-74º08’W). La topografía costera en el área es muy heterogénea y pendiente, debido a su cercanía con la Sierra Nevada de Santa Marta (Rodríguez-Ramírez y Garzón-Ferreira, 2003) .

44 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Figura 1 . Localidades de estudio en el Parque Nacional Natural Tayrona: bahía de Chengue y bahía de Granate (modificado de Rodríguez-Ramírez y Garzón-Ferreira, 2003) .
El clima y las condiciones oceanográficas son similares a la bahía de Santa
Marta y están determinados por la influencia de los vientos Alisios del nororiente . La
precipitación anual promedio para el área es alrededor de los 350 mm y el promedio
anual de temperatura del aire es de 28 ºC (Salzwedel y Müller, 1983); su clima puede
calificarse como seco y cálido . La oceanografía del área está determinada por la presencia de aguas de surgencia costera, con temperaturas de hasta 21 ºC durante la surgencia (diciembre-enero) y de hasta 31 ºC durante el periodo de no surgencia, o lluvias (Zea, 1993; Díaz-Pulido y Garzón-Ferreira, 2002) .
Por su parte, la bahía de Granate está localizada al noroeste de la ciudad de
Santa Marta (Figura 1) a 11º18’ N -74º12’ W. Esta bahía es arenosa en algunas partes
de su periferia y hasta los 25 m de profundidad presenta fondos coralinos (Erhardt y
Werding, 1975) . Esta bahía, aunque abierta al mar, está bien protegida de los fuertes
vientos Alisios del nororiente y de la corriente norte por la isla de la Aguja; la cual
ocasiona un oleaje tenue durante la mayor parte del año, a excepción del costado
occidental que se encuentra más expuesto al impacto directo de los vientos . Granate
exhibe una profundidad que no sobrepasa los 30 m y, al igual que en otras bahías
del PNNT, su parte central está constituida por fondos sedimentarios recientes
(Acosta, 1989) . Esta bahía se diferencia de Chengue por presentar una elevada tasa

Instituto de Investigaciones Marinas y Costeras - INVEMAR 45
de sedimentación natural . En general existe un gradiente de sedimentación natural
a lo largo del PNNT, siendo mayor en áreas cercanas a las fuentes de descarga (e .g .
Santa Marta, debido a los ríos Manzanares y Magdalena) y decreciendo hacia el
nororiente (Zea, 1994) .
MATERIALES Y MÉTODOS
Macroalgas experimentalesLas macroalgas pardas Dictyota spp . y Lobophora variegata fueron
utilizadas por las siguientes razones: 1) el arrecife de estudio provee abundante
sustrato para un buen desarrollo de algas Dictyota spp ., 2) se encuentra una alta
cobertura de Dictyota spp . de 3 a 10 m de profundidad, 3) son macroalgas de fácil
identificación taxonómica a nivel de género, caso de Dictyota spp . (De Clerck,
2003) y 4) ambas algas están implicadas en procesos de degradación arrecifal y
existe poca información sobre las causas de su dominancia en el PNNT (Loya y
Rinkevich, 1980; Díaz-Pulido y Garzón-Ferreira, 2002; Jompa y McCook, 2002) .
El género Dictyota es bastante diverso en el PNNT, con al menos 14 especies
registradas para el área (Díaz-Pulido y Díaz-Ruiz, 2003) . Si bien sería ideal
comparar el reclutamiento de Dictyota a nivel de especies, esto es bastante difícil,
dados los tamaños microscópicos de los propágulos algales y la falta de estructuras
morfológicas y reproductivas únicas entre ellas .
Experimentos en campoSe analizaron los efectos de la sedimentación en el desarrollo de reclutas de
Dictyota spp . y L. variegata utilizando sustratos artificiales expuestos a diferentes
niveles de sedimentos, a una profundidad entre los 6 y 9 m . Las algas fueron
sembradas en sustratos artificiales, los cuales consistían en placas de cerámica (11 x
11 cm de lado, atornilladas a una estructura de hierro en el fondo) . Se trabajaron tres
niveles de sedimentación: 1) alta, 2) natural (control) y 3) remoción de sedimentos .
Las manipulaciones experimentales se realizaron siguiendo metodologías descritas
previamente (Devinny y Volse, 1978; Umar et al ., 1998; Chapman y Fletcher,
2002; Eriksson y Johansson, 2003, 2005) . El experimento comprendió dos fases:
incubación y manipulación experimental .
IncubaciónLa incubación incluyó el asentamiento natural y crecimiento in situ de
las esporas de las algas estudiadas para así disponer de un número de individuos
que permitiera desarrollar el experimento . La incubación se desarrolló entre abril y

46 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
junio de 2005, cuando los picos de crecimiento y desarrollo algal fueron altos (e .g .
Díaz-Pulido y Garzón-Ferreira, 2002), permitiendo capturar y obtener suficientes
propágulos algales . La incubación se desarrolló en Chengue porque Dictyota spp .
es abundante en esta bahía (Bula-Meyer, 1985; Díaz-Pulido y Garzón-Ferreira,
2002) . Con el fin de determinar los efectos de la localidad en el impacto de la
sedimentación sobre las macroalgas, luego del proceso de incubación, la mitad de
las placas (15) fue trasladada a Granate y la otra mitad (15) permaneció en Chengue .
Manipulación experimentalPara determinar los efectos de la sedimentación se realizaron manipulaciones
experimentales de sedimentos marinos, los cuales fueron adicionados o removidos de las placas . La cantidad de sedimentos adicionados se estandarizó teniendo en cuenta las tasas de sedimentación natural para el Parque Nacional Natural Tayrona . Para evitar que la composición del sedimento adicionado fuera diferente entre localidades, los sedimentos adicionados en ambas bahías fueron obtenidos del plano arenoso adyacente al talud arrecifal de Chengue, entre los 10 y 11 m de profundidad . Las adiciones se hicieron semanalmente durante 12 semanas . Las tasas de sedimentación en los arrecifes del Parque Tayrona son bastante variables, por ejemplo en Chengue se han registrado valores máximos promedio de 1 .5 (Garzón-Ferreira, 1998), 4 .5 (Garzón-Ferreira et al., 2002) y 14 .4 mg/cm2/día (Solano, 1987) y en isla Aguja de 1 .5 mg/cm2/día (Zea, 1994) . Con el objeto de maximizar los efectos de la adición de sedimentos en nuestro experimento, se usó una tasa de sedimentación registrada para el Parque Tayrona . Para determinar las tasas experimentales de sedimentación a ser empleadas por placa se multiplicaron las tasas de sedimentación natural (14 .4 mg/cm2/día) por el área de la placa (100 cm2), por el número de días transcurridos entre una adición y otra (aproximadamente una semana) . De esta manera se determinó que una tasa de sedimentación de 10 g de sedimento/semana/placa sería adecuada para el experimento . Esta tasa de sedimentación junto con la tasa natural de sedimentación, constituyen el tratamiento de sedimentación alta .
Los tratamientos de remoción de sedimentos consistieron en retirar las placas de las estructuras metálicas, invertirlas in situ y sacudirlas suavemente por un período estandarizado de 10 segundos; las placas fueron colocadas nuevamente en las estructuras metálicas . Se tuvo cuidado de no dañar los reclutas fijados en las placas . Los tratamientos naturales no fueron manipulados y se constituyeron en controles . Cada tratamiento consistió en cinco placas (réplicas) . Al final del experimento, las placas con las algas fueron transportadas al laboratorio de la Universidad del Magdalena utilizando tanques plásticos de 60 L de capacidad, protegidos de la luz y en agua de mar permanente .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 47
Variables respuestaLas variables respuesta analizadas fueron altura, ancho, puntos de
bifurcación y reclutamiento algal . Estas mediciones se hicieron en el laboratorio
con cubreobjetos milimetrados . La variable reclutamiento para las dos macroalgas
se registró como el número de reclutas que aparecieron en cada placa, exceptuando
el borde de la misma (aproximadamente 1 cm) . Esto último, para evitar el efecto
potencial que puedan generar las características particulares del borde de la placa
(efecto borde) . La altura de Dictyota spp . se obtuvo como la máxima longitud
alcanzada por el talo desde el disco de fijación hasta su parte más alta . El ancho se
determinó como la distancia entre un extremo y otro del borde del talo en sentido
perpendicular al largo; los puntos de bifurcación (PB, en el caso de Dictyota spp .)
se definieron como aquellos donde la célula apical separa dos ramas contiguas entre
sí . Los puntos de bifurcación dan una idea del número de dicotomías de la planta
y se estudian como una medida de crecimiento . La morfometría realizada se basó
en De Clerck (2003) . Para L. variegata la medición del ancho fue definida como el
diámetro del crecimiento lateral del talo (Díaz-Pulido y McCook, 2004) .
Análisis de datosLos datos fueron analizados mediante Análisis de Varianza (ANOVA)
bifactoriales con niveles de sedimentación (adición, control y remoción) y áreas de
estudio (Chengue y Granate) como factores fijos, y placas como réplicas (n= 5) . La
altura, ancho y puntos de bifurcación fueron medidos en 20 reclutas algales que fueron
seleccionados aleatoriamente para cada placa y luego se calculó el valor promedio
por placa para cada variable . Estos valores constituyeron los datos usados (N= 30: 5
réplicas x 3 tratamientos x 2 localidades) en los análisis estadísticos . El reclutamiento
y las otras variables de crecimiento se utilizaron como respuesta en los ANOVA . Los
datos fueron inspeccionados para revisar los supuestos del ANOVA, incluyendo la
homoscedasticidad (prueba de Cochran) y la normalidad de residuales (Underwood,
1997) .
RESULTADOS
Efectos de la sedimentación sobre el reclutamiento y crecimiento de Dictyota spp.Se observó una gran variabilidad de respuestas del alga Dictyota spp .
ante la sedimentación, la cual dependió del parámetro poblacional considerado
(altura, ancho o reclutamiento) y del nivel de sedimentación (adición o remoción) .
La adición de sedimentos en Granate redujo a cero el número de reclutas de
Dictyota spp ., mientras que en Chengue el reclutamiento fue cuatro veces menor,

48 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
aproximadamente, con respecto al tratamiento control . A pesar de estas claras
diferencias, el ANOVA no detectó diferencias significativas . Por su parte, la localidad
afectó significativamente el reclutamiento y crecimiento del alga . El crecimiento de
Dictyota fue mayor en Chengue que en Granate (p= 0 .002; Figuras 2B y C), así
como los puntos de bifurcación (probabilidad marginal, p= 0 .057); el ancho se vio
levemente reducido en Granate, aunque la diferencia no fue significativa (p= 0 .110;
Tabla 1; Figura 2B) .
Figura. 2. Reclutamiento (A), ancho (B), altura (C) y puntos de bifurcación (D) de Dictyota spp . para cada tratamiento en cada localidad. Los datos representan la media ± 1 error estándar, n= 5 placas.
A) Reclutamiento
Granate Chengue
Recl
uta
s/pla
ca (
#)
7
5
2
0
C) Crecimiento
Granate Chengue
Altu
ra (
mm
)
11
10
9
8
7
6
5
4
3
2
1
D) Crecimiento
Granate Chengue
Pu
nto
s d
e b
ifurc
aci
ón
(#)
6
5
4
3
2
1
0
B) Crecimiento
Granate Chengue
Anch
o (
mm
)
3.5
3.0
2.5
2.0
1.5
1.0
0.5
0.0
Sedimentación Adición Control Remoción
La remoción de sedimentos incrementó ligeramente la altura de Dictyota
spp . en ambas bahías (p= 0 .065, Tabla 1, Figura 2C) . También se observó que
el efecto de la remoción de sedimentos fue mayor en Granate (Figura 2B) que
en Chengue . Por su parte, Chengue presentó mayores valores de crecimiento en
altura (p= 0 .002, Tabla 1) . Por otra parte, los puntos de bifurcación no presentaron
diferencias significativas entre los tratamientos de sedimentación, aunque esta
variable presentó mayores valores en Chengue (p= 0 .057; Tabla 1, Figura 2D) .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 49
Variable Fuente de variación Suma de cuadrados gl Cuadrado
medio F p
Altura de Dictyota spp . Localidad 34 .504 1 34 .504 29 .798 0.002
Sedimentación 10 .374 2 5 .187 4 .479 0 .065
Localidad*Sedimentación 5 .590 2 2 .795 2 .414 0 .170Error 6 .948 6 1 .158
Ancho de Dictyota spp . Localidad 2 .817 1 2 .817 3 .517 0 .110
Sedimentación 0 .151 2 0 .076 0 .094 0 .911
Localidad*Sedimentación 0 .161 2 0 .081 0 .101 0 .906
Error 4 .805 6 0 .801
PB Dictyota spp . Localidad 6 .801 1 6 .801 5 .499 0.057Sedimentación 0 .832 2 0 .416 0 .336 0 .727Localidad*Sedimentación 1 .232 2 0 .616 0 .498 0 .631Error 7 .420 6 1 .237
Densidad Dictyota spp . Localidad 32 .033 1 32 .033 4 .319 0.049Sedimentación 18 .067 2 9 .033 1 .218 0 .313Localidad*Sedimentación 11 .267 2 5 .633 0 .760 0 .479Error 178 .000 24 7 .417
Ancho L.variegata Localidad 53 .844 1 53 .844 9 .977 0.005Sedimentación 14 .090 2 7 .045 1 .305 0 .293Localidad*Sedimentación 25 .855 2 12 .928 2 .395 0 .117Error 107 .933 20 5 .397
Densidad L.variegata Localidad 24 .300 1 24 .300 0 .404 0 .531
Sedimentación 36 .867 2 18 .433 0 .306 0 .739Localidad*Sedimentación 79 .800 2 39 .900 0 .663 0 .524Error 1444 .000 24 60 .167
Tabla 1. Resultados de los Análisis de Varianza (ANOVA) . Los valores de p que aparecen en negrillas, indican diferencias significativas .
La localidad afectó significativamente el reclutamiento de Dictyota spp ., presentándose mayores valores en Chengue (p= 0 .049, Tabla 1, Figura 2) . Como
se mencionó anteriormente, se presentaron menores valores de reclutamiento entre
tratamientos de adición que en los de remoción y controles, aunque la diferencia no
fue significativa, sugiriendo que pudo haber un efecto negativo de la sedimentación
en el reclutamiento de Dictyota .
Efectos de la sedimentación sobre el crecimiento y reclutamiento de Lobophora variegata
Lobophora variegata tuvo una respuesta leve a los tratamientos de
sedimentación . En Chengue, la remoción de sedimentos redujo el reclutamiento
a la mitad, con respecto a los tratamientos control o de adición de sedimentos
(Figura 3B) . Sin embargo, esta diferencia no fue significativa . En Granate no se

50 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
observaron diferencias en el reclutamiento de L. variegata entre los tratamientos de
sedimentación . Por otra parte, sí se observaron diferencias significativas entre las
dos bahías con respecto al ancho de los talos de L. variegata, siendo éstos mayores
en Chengue que en Granate (p= 0 .005; Tabla 1, Figura 3A)
DISCUSIÓN
La manipulación de los niveles de sedimentación tuvo una respuesta
variada en las macroalgas Dictyota spp . y L. variegata . La intensidad del efecto
de la sedimentación dependió de la localidad, cantidad de sedimentos adicionados
(o removidos), además del tipo de alga estudiada y del parámetro poblacional
considerado . Esto ilustra la complejidad y variabilidad que existe entre los
parámetros poblacionales de Dictyota spp . y L. variegata y su relación con los
procesos que afectan su ecología . Por lo tanto, es importante considerar la medición
de distintas variables para lograr un mejor entendimiento de los efectos de los
procesos ecológicos en los organismos .
Efectos de la sedimentación y la localidad en el reclutamiento macroalgalLos dos grupos de algas respondieron diferente a los efectos de los
niveles de sedimentación empleados . A pesar de la baja significancia estadística, los resultados sugieren que existe una tendencia a la disminución del crecimiento (altura) y reclutamiento de Dictyota spp . en respuesta al incremento de niveles de sedimentación . Es posible que la corta duración de los experimentos (tres meses) o la limitada replicación (n= 5), hayan generado un error estadístico Tipo II (Underwood, 1997), en donde los análisis estadísticos muestran una baja significancia a pesar
Figura 3. Reclutamiento (3A) y ancho (3B) de Lobophora variegata para cada tratamiento en cada localidad. Los datos representan la media ±1 error estándar, n= 5 placas.
Sedimentación Adición Control Remoción
B) Crecimiento
Granate Chengue
Re
clu
tas/
pla
ca (
#) 15
10
5
0
A) Crecimiento
Granate Chengue
An
cho
(m
m)
10
5
0

Instituto de Investigaciones Marinas y Costeras - INVEMAR 51
de que el efecto existe . Los resultados de Dictyota son, sin embargo, consistentes con estudios previos, que han demostrado que los incrementos en las cargas de sedimentos en los arrecifes de coral son perjudiciales para las algas adultas (Umar et al ., 1998) . Estudios experimentales en zonas templadas han demostrado también que altos niveles de sedimentos disminuyen el reclutamiento de Macrocystis (Neushul et al ., 1976), impiden la fijación de propágulos en M. pyrifera (Devinny y Volse, 1978) y la fijación de estados microscópicos de varios kelpos como Undaria pinnatifida y Ecklonia cava (Arakawa y Matsuike, 1992) . Espinoza y Rodríguez (1987) también encontraron una reducción en las tallas del talo y la capacidad reproductiva de Sargassum sinicola sujeta a incrementos en los niveles de sedimentos en el golfo de California . Resultados similares se han observado en corales, pastos marinos y manglares, reduciendo su reclutamiento, cobertura, establecimiento, supervivencia y tasas de crecimiento (Pringle, 1989; Hodgson, 1990; Rogers, 1990; Umar et al ., 1998; Airoldi, 2003; Schiel et al ., 2006) . Este estudio demuestra por primera vez efectos negativos de la sedimentación sobre los estadios tempranos de macroalgas de arrecifes coralinos .
Se han postulado cuatro mecanismos principales mediante los cuales, los incrementos de sedimentos afectan las algas marinas (Airoldi, 2003) . 1) Asfixia: implica una reducción de la disponibilidad y calidad de la luz y oxígeno generando potencialmente la acumulación de sulfuro de hidrógeno y desechos de productos metabólicos, cuyo efecto produce cambios en las características químicas del microambiente . 2) Reducción del acceso a nutrientes esenciales: posiblemente debido a alteraciones en las capas de intercambio (“boundary layers”, Larkum et al ., 2003) que afectan la difusión e intercambio de nutrientes . 3) Abrasión: causada por el movimiento de sedimentos que puede dañar y remover el alga o sus partes . 4) Cambios en las características físicas de la superficie del fondo: ocurre como consecuencia del reemplazo de sustratos duros estables por partículas inestables y puede resultar en una pérdida de hábitat para el establecimiento algal (Devinny y Volse, 1978; D´Antonio, 1986; Airoldi et al ., 1996; Airoldi, 1998) . Este trabajo no pretendió examinar cómo estos mecanismos afectan las algas, sin embargo las observaciones sugieren que es posible que todos estos mecanismos operen en conjunto para reducir el reclutamiento y crecimiento del alga Dictyota . Es importante resaltar que se requieren más estudios que profundicen sobre los mecanismos mediante los cuales los sedimentos afectan las algas marinas .
La adición de sedimentos no afectó el reclutamiento de L. variegata,
contrario a lo observado en Dictyota spp . Este resultado es sorprendente porque
generalmente se tiende a asociar el incremento de sedimentación con el deterioro o
daño de las poblaciones algales (como se detalló en el caso de Dictyota) . Por otro
lado, existe evidencia de que algunas algas pueden ser resistentes a la sedimentación

52 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
(D`Antonio, 1986; Kendrick, 1991) . Trabajos en algas rojas muestran que existe
una resistencia a la abrasión causada por arena y muestran que estas algas pueden
prosperar en áreas donde pocas especies sobreviven (D`Antonio, 1986) . Otros
trabajos en campo registran que la presencia de limo podría ser benéfica al suministrar
nutrientes y proteger las algas del movimiento del agua (Kennelly, 1983; Schaffelke,
1999) . Sin embargo, la adición de sedimentos redujo el crecimiento de L. variegata
(Figura 3B) y por tanto es posible que el mecanismo de aumento de disponibilidad
de nutrientes no opere en estas circunstancias particulares .
Estos resultados enfatizan la variabilidad de respuestas encontradas en las
poblaciones algales ante los efectos de la sedimentación . La razón de la falta de
respuestas en el reclutamiento de L. variegata a la adición de sedimentos no es clara,
pero es posible que los sedimentos puedan proteger de una u otra manera al alga
del pastoreo de herbívoros, por ejemplo creando una barrera física especialmente
contra mesoherbívoros (Kennelly, 1983) . Esto puede ser particularmente cierto si se
considera que en el tratamiento de remoción de sedimentos, el reclutamiento de L. variegata se redujo en más de la mitad . Sin embargo, la hipótesis de la protección
de sedimentos contra el pastoreo necesita verificación experimental . Finalmente, la
tolerancia de L. variegata a niveles de sedimentación puede conferir cierto grado
de ventaja en hábitats o situaciones (e .g . cambios de estado de corales a algas) con
niveles de sedimentación moderados o altos .
Los impactos de la sedimentación sobre las algas variaron entre localidades
(arrecifes) expuestas a diferentes grados de sedimentación natural, con valores
menores de reclutamiento y crecimiento algal en el arrecife con mayores tasas
de sedimentación (Granate). Áreas como Granate y la bahía de Santa Marta, que
presentan un mayor grado de impacto antropogénico (al encontrarse cercanas a áreas
urbanas, turísticas y por estar cerca a fuentes de descarga continental como ríos y
desagües de aguas residuales), son aparentemente menos aptas para el crecimiento
de algas pardas rastreras, como es el caso de Dictyota y L. variegata, que áreas
con menores tasas de sedimentación (e .g . Chengue) . Algas pardas de talos erectos
considerables, como por ejemplo Sargassum, sin embargo proliferan en Granate
pero son escasas en los arrecifes de Chengue . Diversos factores pueden también
jugar papeles claves en determinar las distribuciones espaciales de algas en la
región del PNNT, e .g . tasas de pastoreo (Díaz-Pulido et al ., 2007), disponibilidad
de nutrientes, grado de deterioro del arrecife, intensidad de la surgencia (Bula-
Meyer, 1985) . Nuestro experimento demuestra que el tipo y magnitud del impacto
de los sedimentos es altamente heterogéneo en escalas espaciales, contribuyendo
a la complejidad espacial de las comunidades algales de la región del PNNT . Este
estudio indica que existe una gran complejidad y variabilidad en las respuestas de

Instituto de Investigaciones Marinas y Costeras - INVEMAR 53
las algas arrecifales a los efectos de la escorrentía de sedimentos desde la Sierra
Nevada de Santa Marta . Sugiere también que los procesos de degradación arrecifal
pueden modificar la dinámica poblacional y estadios tempranos de vida de las algas
bentónicas, lo cual puede tener implicaciones importantes en el entendimiento de la
ecología de la degradación arrecifal .
AGRADECIMIENTOS
Este estudio fue financiado por el Instituto Colombiano para el Desarrollo
de la Ciencia y la Tecnología-Colciencias, el Fondo Patrimonial para la Investigación
de la Universidad del Magdalena-Fonciencias, y el Instituto de Investigaciones
Marinas y Costeras-INVEMAR . Agradecemos a Olga Camacho, Katrín Perilla,
Sonia Bejarano, Érika Ortíz, Isabel Chacón, Oscar Martínez y Adriana Santacruz
por su ayuda en campo . Igualmente a Laura Airoldi, Bernabé Santelices, Sven Zea,
Adolfo Sanjuán y a dos revisores anónimos por sus comentarios constructivos .
BIBLIOGRAFÍA
Acosta, L . 1989 . Composición y estructura de las comunidades de corales hermatípicos en tres zonas
representativas de la región de Santa Marta, Punta Betín, isla Morro Grande y ensenada de Granate,
mar Caribe colombiano . Tesis de Biol . Univ . Nacional de Colombia, Bogotá . 242 p .
Adey, W . H . 1998 . Coral reefs: algal structured and mediated ecosystems in shallow, turbulent, alkaline
waters . J . Phycol ., 34: 393-406 .
Airoldi, L . 1998 . Roles of disturbance, sediment stress, and substratum retention on spatial dominance in
algal turf . Ecology, 79: 2759-2770 .
Airoldi, L . 2003 . The effects of sedimentation on rocky coast assemblages . Oceanogr . Mar . Biol . Ann .
Rev ., 41: 161-236 .
Airoldi, L ., M . Fabiano y F . Cinelli . 1996 . Sediment deposition and movement over a turf assemblage in a
shallow rocky coastal area on the Ligurian Sea . Mar . Ecol . Prog . Ser ., 133: 241-251 .
Arakawa, H . y K . Matsuike . 1992 . Influence on insertion of zoospores, germination, survival, and
maturation of gametophytes of brown algae exerted by sediments . Nippon Suisan Gakkaishi, 58:
619-625 .
Bruijnzeel, L . 1990 . Hydrology of moist tropical forest and effects of conversion: a state of the knowledge .
UNESCO, París . 224 p .
Bula-Meyer, G . 1985 . Un núcleo nuevo de surgencia en el Caribe colombiano detectado en correlación
con las macroalgas . Bol . Ecotrópica, 12: 2-26 .
Bula-Meyer, G . 2004 . Las macroalgas marinas en la agronomía y el uso potencial de Sargassum flotante
en la producción de fertilizantes en el archipiélago de San Andrés y Providencia, Colombia . Rev .
Intrópica, 1: 91-103 .
Chapman, A . S y R . L . Fletcher . 2002 . Differential effects of sediments on survival and growth of Fucus
serratus embryos (Fucales, Phaeophyceae) . J . Phycology, 38: 894-903 .

54 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
D’Antonio, C. M. 1986. Role of sand in the domination of hard substrata by the intertidal alga Rhodomela
larix. Mar . Ecol . Prog . Ser ., 27: 263-275 .
De Clerck, O . 2003 . The genus Dictyota (Dictyotales, Phaeophyta) in the Indian Ocean . Opera Bot .,
Bélgica, 13: 1-205 .
Devinny, J . S . y L . A . Volse . 1978 . Effects of sediments on the development of Macrocystis pyrifera
gametophytes . Mar . Biol ., 48: 343-348 .
Díaz-Pulido, G . y M . Díaz-Ruiz . 2003 . Diversity of benthic marine algae of the Colombian Atlantic . Biota
Colomb . 4: 203-246 .
Díaz-Pulido, G . y J . Garzón-Ferreira . 2002 . Seasonality in algal assemblages on upwelling-influenced
coral reef in the Colombian Caribbean . Bot . Mar ., 45: 284-292 .
Díaz-Pulido, G . y L . McCook . 2004 . Effects of live coral, epilithic algae communities and substrate type
on algal recruitment . Coral Reefs, 23: 225-233 .
Díaz-Pulido, G ., L . Villamil y V . Almanza . 2007 . Herbivory effects on the morphology of the brown alga
Padina boergesenii (Phaeophyta) . Phycologia, 46: 131-136 .
Done, T . J . 1992 . Phase shifts in coral reef communities and their ecological significance . Hydrobiología,
247: 121-132 .
Erhardt, H . y B . Werding . 1975 . Los corales (Antozoa e Hydrozoa) de la ensenada de Granate, pequeña
bahía al este de Santa Marta, Colombia . Caldasia, 11 (53): 107-138 .
Eriksson, B . K . y G . Johansson . 2003 . Sedimentation reduces recruitment success of Fucus vesiculosus
(Phaeophyceae) in the Baltic Sea . Eur . J . Phycol ., 38: 217-222 .
Eriksson, B . K . y G . Johansson . 2005 . Effects of sedimentation on macroalgae: species-specific responses
are related to reproductive traits . Oecologia, 143: 438-448 .
Espinoza, J . y H . Rodríguez . 1987 . Seasonal phenology and reciprocal transplantation of Sargassum
sinicola Setchell et Gardner in the southern Gulf of California . J . Exp . Mar . Biol . Ecol ., 110: 183-
195 .
Garzón-Ferreira, J . 1998 . Bahía de Chengue, Parque Nacional Natural Tayrona, Colombia . 115-125 . En:
CARICOMP (Ed .) . Caribbean coral reef, seagrass and mangrove sites . UNESCO, París . 345 p .
Garzón-Ferreira, J ., A . Rodríguez-Ramírez, S . Bejarano-Chavarro, R . Navas-Camacho y C . Reyes-
Nivia . 2002 . Estado de los arrecifes coralinos . 29-40 . En: Ospina-Salazar, G . H . y A . Acero P .
(Eds .) . Informe del estado de los ambientes marinos y costeros en Colombia: año 2001 . Serie de
Publicaciones Periódicas No . 8, Instituto de Investigaciones Marinas y Costeras-INVEMAR, Santa
Marta . 292 p .
Grimsditch, G . y R . V . Salm . 2006 . Coral reef resilience and resistance to bleaching . IUCN, Gland, Suiza .
53 p .
Hodgson, G . 1990 . Tetracycline reduces sedimentation damage to corals . Mar . Biol ., 104: 493-496 .
Hoegh-Guldberg, O ., P . J . Mumby, A . J . Hooten, R . S . Steneck, P . Greenfield, E . Gómez, C . D . Harvell,
P . F . Sale, A . J . Edwards, K . Caldeira, N . Knowlton, C . M . Eakin, R . Iglesias-Prieto, N . Muthiga, R .
H . Bradbury, A . Dubi y M . E . Hatziolos . 2007 . Coral reefs under rapid climate change and ocean
acidification . Science, 318: 1737-1742 .
Hughes, T . P . 1994 . Catastrophes, phase shifts and large-scale degradation of a Caribbean coral reef .
Science, 265 (5178): 1547-1551 .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 55
Hughes, T . P ., A . H . Baird, D . R . Bellwood, M . Card, S . R . Connolly, C . Folke, R . K . Grosberg, O . Hoegh-
Guldberg, J . B . C . Jackson, J . Kleypas, J . M . Lough, P . A . Marshall, M . Nystrom, S . R . Palumbi,
J . M . Pandolfi, B . R . Rosen y J . Roughgarden . 2003 . Climate change, human impacts, and the
resilence of coral reefs . Science, 301: 929-933 .
Jackson, J . B . C ., M . X . Kirby, W . H . Berger, K . A . Bjorndal, L . W . Botsford, B . J . Bourque, R . H .
Bradbury, R . Cooke, J . Erlandson, J . A . Estes, T . P . Hughes, S . Kidwell, C . B . Lange, H . S . Lenihan,
J . M . Pandolfi, C . H . Peterson, R . S . Steneck, M . J . Tegner y R . R . Warner . 2001 . Historical
overfishing and the recent collapse of coastal ecosystems . Science, 293: 629-638 .
Jensen, A . 1993 . Present and future needs for algae and algal products . Hydrobiología, 260-261: 15-23 .
Jompa, J . y L . J . McCook . 2002 . The effects of nutrients and herbivory on competition between a hard
coral (Porites cylindica) and a brown algae (Lobophora variegata) . Limnol . Oceanogr ., 47: 527-
534 .
Kendrick, G . A . 1991 . Recruitment of coralline crusts and filamentous turf algae in the Galapagos
archipelago: effect of simulated scour, erosion and accretion . J . Exp . Mar . Biol . Ecol ., 147: 47-63 .
Kennelly, S . J . 1983 . An experimental approach to the study of factors affecting algal colonization in a
sublittoral kelp forest . J . Exp . Mar . Biol . Ecol ., 68: 257-276 .
Larkum, A . W . D ., E . C . Koch y M . Kuehl 2003 . Diffusive boundary layers and photosynthesis of the
epilithic algal community of coral reefs . Mar . Biol ., 142: 1073-1082 .
Loya, Y . y B . Rinkevich . 1980 . Effect of oil pollution on coral reef communities . Mar . Ecol . Prog . Ser .,
3: 176-180 .
McCook, L . J . 1999 . Macroalgae, nutrient and phase shifts on coral reefs: scientific issues and management
consequences for the Great Barrier Reef . Coral Reefs, 18: 357-367 .
Neushul, M ., M . S . Foster, D . A . Coon, J . W . Woessner y B . W . Harquer . 1976 . An in situ study of
recruitment, growth and survival of subtidal marine algae: techniques and preliminary result . J .
Phycol ., 12: 397-408 .
Pringle, A . W . 1989 . The history of dredging in Cleveland Bay, Queensland and its effect on sediment
movement and on growth of mangrove, coral and seagrass . Great Barrier Reef Marine Park Autority,
Townsville, Queensland, Australia . 177 p .
Richardson, L . L . 1998 . Coral diseases: what is really known? Trends Ecol . Evol ., 13: 438-443 .
Rodríguez-Ramírez, A . y J . Garzón-Ferreira . 2003 . Monitoreo de arrecifes coralinos, pastos marinos
y manglares en la bahía de Chengue (Caribe colombiano) 1993-1999 . Series de Publicaciones
Especiales No 8, Instituto de Investigaciones Marinas y Costeras-INVEMAR, Santa Marta . 170 p .
Rogers, C . S . 1990 . Responses of coral reefs and reefs organisms to sedimentation . Mar . Ecol . Prog . Ser .,
62: 185-202 .
Salzwedel, H. y K. Müller. 1983. A summary of meteorological data for the bay Santa Marta, Colombian
Caribbean . An . Inst . Invest . Mar . Punta de Betín, 13: 67-83 .
Schaffelke, B . 1999 . Particulate organic matter as an alternative nutrient source for tropical Sargassum
species (Fucales, Pheophyceae) . J . Phycol ., 35: 1150-1157 .
Schiel, D . R ., S . A . Wood, R . A . Dunmore y D . Taylor . 2006 . Sediment on rocky intertidal reefs: effects
on early post-settlement stages of habitat-forming seaweeds . J . Exp . Mar . Biol . Ecol ., 331: 158-172 .

56 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Solano, O . 1987 . Estructura y diversidad de las comunidades de corales hermatípicos en la bahía de
Chengue (PNNT) . Tesis M . Sc ., Univ . Nacional de Colombia, Bogotá . 111 p .
Umar, M . J ., L . J . McCooke e I . R . Price . 1998 . Effects of sediment deposition on the seaweed Sargassum
on a fringing reef. Coral Reefs, 17: 169-177 .
Underwood, A . J . 1997 . Experiments in ecology: their logical design and interpretation using analysis of
variance . Cambridge University Press, Cambridge, Reino Unido . 503 p .
Zea, S . 1993 . Cover of sponges and other sessile organisms in rocky and coral reef habitats of Santa
Marta, Colombian Caribbean Sea . Caribb . J . Sci ., 29 (1-2): 75-88 .
Zea, S . 1994 . Patterns of coral and sponge abundance in stressed coral reefs at Santa Marta, Colombian
Caribbean . 257-264 . En: Van Soest, R . W . M ., T . M . G . Van Kempen y J . C . Braekman (Eds .) . Sponges
in time and space. Proceedings of the 4th International Porifera Congress. Ámsterdam. 515 p.
FECHA DE RECEPCIÓN: 16/02/2007 FECHA DE ACEPTACIÓN: 19/03/2010.

Instituto de Investigaciones Marinas y Costeras - INVEMAR 57
Bol . Invest . Mar . Cost . 39 (1) ISSN 0122-9761 Santa Marta, Colombia, 2010
DISTRIBUCIÓN Y ABUNDANCIA DE LAS ASCIDIAS DE LOS FONDOS ROCOSOS DE LA BAHÍA DE CUAJINIQUIL,
COSTA RICA
Nidya Nova-Bustos1, Aida C. Hernández-Zanuy2 y Rigoberto Viquez-Portuguez1
1 Universidad Nacional, Heredia 86-3000, Costa Rica. [email protected] Instituto de Oceanología, Avenida Primera Número 18406 entre 184 y 186, Playa, La Habana, Cuba
RESUMEN
Las ascidias de la costa del Pacífico centroamericano están muy poco estudiadas a pesar de que este grupo se destaca entre los miembros de la fauna bentónica de los fondos duros de la región . Teniendo en cuenta lo anterior, el objetivo del presente trabajo es conocer la distribución y abundancia de las ascidias en los fondos rocosos de la bahía de Cuajiniquil . Se ofrecen datos de la densidad (individuos por metro cuadrado de fondo) de las ascidias solitarias Rhopalaea birkelandi (Tokioka) y Ascidia ceratodes (Huntsman) a 5, 10 y 15 m de profundidad en tres transectos perpendiculares a la costa que se ubicaron en Isla David, Bajo Rojo y Bajo Viejón, sitios costeros de la bahía de Cuajiniquil, en el noroeste de Costa Rica . Otras tres especies de ascidias coloniales Didemnum moseleyi (Herdman), Lissoclinum caulleryi (Ritter y Forsyth) y Polyandrocarpa tincta (Van Name) forman parte de los fondos rocosos de esta bahía a profundidades mayores de 10 m . El dendrograma de similitud (coeficiente de similitud de Jaccard) muestra elevada semejanza entre los transectos ubicados en Bajo Viejón e Isla David . La diferencia entre la profundidad de 5 m y las de 10 y 15 m es significativa (H= 6 .24, gl= 2 .51, p= 0 .04) . En Bajo Rojo sólo se encontraron ascidias solitarias con una densidad promedio 10 veces menor que en los otros dos sitios . Las diferencias pudieran estar relacionadas con la menor disponibilidad de sustrato provocada por las algas pardas que cubren la mayor parte del fondo en Bajo Rojo y/o con la mayor estabilidad ambiental de las comunidades de fondos duros que se encuentran en los bordes de la boca de la bahía, más expuestos al intercambio con el océano y alejados del impacto que los sedimentos provenientes del río Cuajiniquil pudiera estar provocando en el centro de la bahía .
PALABRAS CLAVE: Distribución, Abundancia, Ascidias, Densidad, Bahía Cuajiniquil .
ABSTRACT
Distribution and abundance of the rocky bottom ascidians of Cuajiniquil Bay, Costa Rica.The ascidians of theCentral-American Pacific coast are poorly studied, although this group isone of the most important members of the benthic fauna of this region. The present study shows data of density (individuals per square meter) of the solitary ascidians Rhopalaea birkelandi (Tokioka) and Ascidia ceratodes (Huntsman), at 5, 10 and 15 m deep in three transects perpendicular to the coast. These transects were located in Isla David, Bajo Rojo and Bajo Viejón, coastal areas of Cuajiniquil Bay, northeast coast Costa Rica. Other three species of colonial ascidians, Didemnum moseleyi (Herdman), Lissoclinum caulleryi (Ritter y Forsyth) and Polyandrocarpa tincta (Van Name), are part of the benthic
57-66

58 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
communities of the rocky bottoms of this bay at depths greater than 10 m. The cluster analysis (using Jaccardsimilaritycoefficient)showsahighsimilaritybetweentransectslocatedatBajoViejónandIslaDavid.Thedifferencebetween5,10and15mdeepissignificant(H=6.24,gl=2.51,p=0.04).InBajoRojo only solitary ascidians were found with an average density ten times lower than other sites. These differences could be related to lower substratum availability due to the stronger competition with brown algae which covers most of the sea bottom at Bajo Rojo and/or the higher environmental stability of the hard bottom communities that are usually found on the edges of the higher part of the bay, greatly exposed to oceanic waters and away from impacts that sediments from the Cuajiniquil river could be producing in the middle part of the bay.
KEY WORDS: Distribution, Abundance, Ascidians, Density, Cuajiniquil Bay .
INTRODUCCIÓN
Las ascidias de la costa del Pacífico centroamericano están muy poco
estudiadas a diferencia de la fauna cercana correspondiente al Pacífico de
Norteamérica, la que en los últimos años ha recibido una atención especial como
parte del estudio de organismos bioinvasores (Abbott y Newberry, 1980; Lambert,
1993; Lambert y Lambert, 1998, 2003) . Sólo algunos trabajos (Van Name, 1945;
Tokioka, 1971, 1972; Abbott y Newberry, 1980) mencionan la presencia de este
grupo en la costa occidental de América Central .
En Costa Rica las ascidias han recibido poca atención, resultando muy
escasos los estudios de este grupo desde el punto de vista taxonómico y nulos los
estudios ecológicos, a pesar de que este grupo resalta entre los miembros de la
fauna bentónica de los fondos duros de la región (Tokioka, 1972) . En la costa del
Pacífico costarricense sólo Tokioka (1972) señala 13 especies de ascidias que fueron
colectadas en playa del Coco en 1970 . La falta de conocimientos del bentos marino
en la costa del Pacífico, la ausencia de estudios previos sobre ascidias en Costa Rica,
y la abundancia y longevidad de las ascidias solitarias en estas costas, motivó la
presente investigación cuyo objetivo es evaluar la abundancia y distribución de estos
organismos en la bahía de Cuajiniquil, como contribución al conocimiento de la
diversidad de especies y a la línea base ambiental marina de Costa Rica .
ÁREA DE ESTUDIO
El estudio se realizó en la bahía de Cuajiniquil, área marino-costera del
Pacífico noroeste de Costa Rica (Figura 1), en tres transectos perpendiculares a la
costa que fueron ubicados en sitios conocidos como Bajo Rojo [Sitio 1: (10°57’45’’ N y 85°44’03’’W)], Isla David [Sitio 2: (10°57’46’’N y 85°85’43’’ W)] y Bajo Viejón [Sitio 3: (10°57’32’’N y 85°43’75’’W)]. Los muestreos se realizaron de julio
a octubre del 2006, incluyendo parte del llamado “veranillo” (períodos secos de

Instituto de Investigaciones Marinas y Costeras - INVEMAR 59
más de tres días que ocurren en los meses de julio y agosto), y septiembre cuando
se producen las mayores lluvias (Manso et al., 2005) .
Figura 1. Ubicación geográfica de los tres sitios de muestreo (Sitio 1: Bajo Rojo, Sitio 2: Bajo Viejón y Sitio 3: Isla David) estudiados en la bahía de Cuajiniquil, en la costa noroeste de Costa Rica .
MATERIALES Y MÉTODOS
A través de buceo autónomo, en cada transecto y a cada profundidad se
colocaron tres cuadrantes al azar sobre el fondo (marcos de aluminio de 1 x 1 m),
en cada sitio de muestreo, en los cuales se contó el número de ascidias solitarias por
especie . De cada una de las especies (solitarias y coloniales) se colectaron entre 10
y 20 ejemplares (siempre que fue posible) para su identificación en el laboratorio .
Los muestreos fueron realizados una sola vez en cada transecto entre julio y octubre
de 2006 .
Las características del biotopo señalando las peculiaridades físicas
del sustrato de cada sitio (grietas, fisuras o pequeñas ondulaciones) y de la biota
asociada, se anotaron en una tablilla junto a los datos de las especies . La densidad de
ascidias como indicador de abundancia se ofrece en número de individuos por metro
cuadrado (ind/m2) .

60 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Para comparar los tres sitios estudiados y las tres zonas (profundidades de
5, 10 y 15 m) se utilizó la prueba de Kruskal-Wallis para datos no paramétricos
(Statgraphics 1994-1997) . Se aplicó el coeficiente de similitud de Jaccard con un
α= 0 .05, empleando el programa InfoStat (2002) y se construyó un dendrograma de
similitud con el método de encadenamiento completo utilizado para medir el grado
de semejanza entre dos muestras según las especies compartidas entre sitios .
Para la colecta, anestesiado y fijación de las ascidias se utilizó el método
propuesto por Monniot y Monniot (1972) . La identificación se realizó consultando
los trabajos de Van Name (1945), Tokioka (1971, 1972), Abbott y Newberry (1980)
y Vásquez y Young (1996) .
RESULTADOS
Caracterización de las zonas de estudio y abundancia de las ascidiasLas características físico-biológicas de los tres sitios estudiados se muestran
en la Tabla 1 . El fondo es duro en los tres sitios y a las tres profundidades estudiadas,
fundamentalmente rocoso cubierto por algas pardas y verdes así como por el coral
Tubastraea coccinea (Lesson).
Sitio Tipo de fondo Biota
Sitio 1 Bajo Rojo Fondo rocoso, muy regular, con ondulaciones leves y con escasas fisuras, bastante homogéneo desde 5 hasta 15 m, perfil que se ubica en la pared interna de un promontorio rocoso que se eleva desde los 32 m hasta la superficie, frente a la costa central de la bahía de Cuajiniquil . Las ascidias, se ubican directamente sobre la roca tanto en posición horizontal como diagonal y vertical .
Algas pardas y verdes (muy abundantes); cubren la mayor parte del fondo .
Sitio 2 Isla David Fondo rocoso con muy leves ondulaciones, liso pero con grietas también lisas en su interior y muy angostas, lo que dificulta la extracción de los ejemplares de ascidias . Se encuentra de la costa del extremo norte de la bahía de Cuajiniquil con una inclinación de 45o . Las ascidias, se ubican ya sea en el suelo rocoso horizontal, diagonal o vertical .
Algas pardas y verdes no muy abundantes . Presencia de Tubastraea coccinea.
Sitio 3 Bajo Viejón Fondo rocoso muy homogéneo de la costa hacia mayores profundidades con una inclinación menor de 25o y ondulaciones leves, grietas numerosas lisas y muy angostas . Las ascidias, se ubican ya sea en el suelo rocoso horizontal, diagonal o vertical .
Algas pardas y verdes no muy abundantes . Tubastraea coccinea muy abundante .
Tabla 1. Características físico-biológicas de los tres sitios estudiados .
Instituto de Investigaciones Marinas y Costeras - INVEMAR 61
En Bajo Rojo se destaca la abundancia de algas verdes y pardas, siendo
estas últimas los organismos que cubren la mayor parte del fondo . En el Bajo Viejón
e Isla David el coral T. coccinea se destaca por su colorido y abundancia, mayor en
el primer sitio que en el segundo .
Las ascidias estuvieron presentes en los tres sitios y en las tres profundidades
estudiadas (Tabla 2) . Sólo se identificaron cinco especies, dos solitarias y tres
coloniales, estas últimas no aparecieron en el sitio 1 (Bajo Rojo) .
Tabla 2. Especies de ascidias identificadas en los tres sitios de la bahía de Cuajiniquil a tres diferentes profundidades (5, 10 y 15 m) .
Sitio 1 Sitio 2 Sitio 3
Bajo Rojo Isla David Bajo Viejón
5 m 10 m 15 m 5 m 10 m 15 m 5 m 10 m 15 m
Especie
Ascidia ceratodes X X X X X X
Didemnum mosseleyi X X X X
Lissoclinum caulleryi X X
Polyandrocarpa tincta X X X X
Rhopalaea birkelandi X X X X X X X X
Las mayores densidades de ascidias solitarias (sumando el número de
ejemplares de las dos especies en los tres cuadrantes) en las tres profundidades en
cada sitio se presentaron en Isla David (41 ind/m2) seguidas por Bajo Viejón (35
ind/m2), ambos sitios con una densidad casi 10 veces mayor que la encontrada en
Bajo Rojo . La ascidia Rhopalaea birkelandi es la más abundante en los tres sitios,
alcanzando valores de (36 ind/m2) en Isla David, seguidas por las encontradas en
Bajo Viejón (32 ind/m2), particularmente a profundidades de 10 y 15 m (Tabla 3) .
Como se observa en la Tabla 3, las densidades totales de ascidias solitarias
(sumando el número de ejemplares de las dos especies en cada cuadrante) aumentan
con la profundidad (6 ind/m2 a 5 m; 42 ind/m2 a 10 m y 43 ind/m2 a 15 m) . Las
diferencias de densidades entre la profundidad de 5 m y las de 10 y 15 m son
significativas (H= 6 .24, gl= 2 .51, p= 0 .04) . El dendrograma obtenido a partir del
coeficiente de similitud de Jaccard (Figura 2) mostró que entre las zonas 2 y 3 hay
60 % de similitud con relación al número de especies, en cambio entre la zona 1 y 3
sólo se encontró un 29 % de similitud .

62 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
5 m
10 m
15 m
TO
TA
L
Rép
licas
12
3N
Dm
DE
12
3N
Dm
DE
12
3N
Dm
DE
ND
m
(ind
/m
2 )D
E
Sitio
de
m
uest
reo
Esp
ecie
Baj
o R
ojo
Rho
pala
ea
birk
elan
di12
00
124
6 .93
90
09
35 .
27
00
72 .
334 .
0428
3 .11
10 .2
1
Asc
idia
ce
rato
des
90
09
35 .
20
00
00
00
00
00
09
15 .
2
Tota
l de
indi
vidu
os21
712
.19
35 .
27
2 .33
4 .04
374 .
1115
.41
Isla
Dav
idR
hopa
laea
bi
rkel
andi
1717
034
11 .3
39 .
8132
3036
9833
3 .06
5470
7119
565
9 .54
327
36 .3
154 .
05
Asc
idia
ce
rato
des
00
44
1 .33
2 .31
87
621
71
46
1222
7 .33
4 .16
475 .
2221
.66
Tota
l de
indi
vidu
os38
12 .6
78 .
4111
940
14 .2
217
72 .3
32 .2
637
441
.674
.28
Baj
o V
iejó
nR
hopa
laea
bi
rkel
andi
00
00
00
4047
3612
341
5 .57
1572
8417
157
36 .8
629
432
.714
7 .65
Asc
idia
ce
rato
des
00
00
00
1512
027
97 .
940
00
00
027
315
.59
Tota
l de
indi
vidu
os0
00
150
5019
171
5739
321
35 .7
67
TO
TAL
Rho
pala
ea b
irke
land
i46
230
373
Dm
5 .1
25 .5
641
.44
DE
1759
.92
102 .
32
Asc
idia
cer
atod
es
1348
22D
m
1 .4
5 .33
2 .44
DE
4 .5
14 .2
12 .7
Tab
la 3
. C
onte
os d
e la
s do
s es
peci
es d
e as
cidi
as s
olita
rias
en
bahí
a C
uajin
iqui
l (R
hopa
laea
bir
kela
ndi y
Asc
idia
cer
atod
es).
N: n
úmer
o de
indi
vidu
os s
umad
os e
n tr
es
cuad
rant
es, D
m: d
ensi
dad
prom
edio
(in
d/m
2 ), D
E: D
esvi
ació
n es
tánd
ar .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 63
DISCUSIÓN
En la bahía de Cuajiniquil la abundancia de ascidias solitarias que alcanza
valores superiores a 60 ind/m2 y hasta 84 ind/m2 como valor máximo medido en un
cuadrante en Isla David, pudiera estar relacionada con varios factores . En primer lugar,
las ascidias, al igual que la mayoría de los organismos filtradores, son abundantes
en localidades donde hay elevada disponibilidad de alimento disuelto en la columna
de agua y condiciones hidrológicas favorables, según los requerimientos ecológicos
de cada especie . Estas condiciones son comunes en las bahías (Monniot y Monniot,
1985; Hernández-Zanuy, 1999) .
La abundancia de ascidias solitarias encontrada en la bahía de Cuajiniquil
(área marginal del afloramiento de Papagayo) pudiera estar relacionada con el aporte
de nutrientes tanto provenientes de la corriente de California y los afloramientos que
en la zona ocurren (Acuña et al ., 1997; Jiménez, 2001), como de los arrastres del río
Cuajiniquil y de los manglares que bordean la costa . La abundancia de la ascidia
solitaria R. birkelandi en la costa pacífica de Costa Rica fue señalada anteriormente
por Tokioka (1971) a partir de las colectas realizadas por el Dr . Birkeland en playa del
Coco en 1970 . Aunque en el mencionado estudio no se ofrecen datos cuantitativos y
no es posible realizar una comparación detallada, nuestros resultados indican que 36
años después, esta ascidia continúa siendo uno de los miembros más sobresalientes
de la comunidad bentónica del Pacífico de Costa Rica .
El género Rhopalaea en general es considerado por Monniot y Monniot
(1985) como un relicto de la fauna cuaternaria, lo que condiciona su presencia en
Figura 2. Coeficiente de similitud de Jaccard para las especies presentes en las tres áreas de muestreo (Sitio 1: Bajo Rojo, Sitio 2: Bajo Viejón y Sitio 3: Isla David) .
Coeficiente de similitud de Jaccard

64 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
fondos por debajo de la termoclina estival (a partir de 20 m) (Ramos y Ros, 1990) .
La mayor abundancia de R. birkelandi se encontró en Isla David donde la pendiente
del fondo es muy abrupta (>45°) y en las profundidades mayores (10-15 m), donde
generalmente la temperatura del agua es menor .
En Bajo Rojo las poblaciones de ascidias son diferentes a las encontradas
en los otros dos sitios estudiados, en éste sólo se encontraron ascidias solitarias y con
muy poca densidad . En este sitio la biota acompañante también difiere de la observada
en los otros dos sitios . Los ejemplares del coral T. coccinea, muy abundantes en los
otros dos sitios, no aparecieron y las algas pardas cubrían la mayor parte del fondo,
aspecto también diferente en los otros dos sitios donde, a pesar de encontrar algas,
éstas no ocupaban la mayor parte del fondo marino . La elevada cobertura de algas
sobre un fondo rocoso coralino es un indicador de estrés ambiental producido por
contaminación orgánica en la zona .
Con relación a la comparación entre sitios, las medidas de similaridad
(Figura 2) muestran una notoria correspondencia entre los sitios 2 y 3, probablemente
por la distribución agregada de las poblaciones de ascidias a lo largo de los sitios de
muestreo en general o por la disponibilidad, tipo y distribución de sustrato duro apto
para la colonización de estos individuos . Además, según Bassey (2010), Isla David
y Bajo Viejón son comunidades coralinas sobre arena, a diferencia de Bajo Rojo que
es un arrecife de coral masivo .
El éxito de las ascidias R. birkelandi y A. ceratodes también pudiera estar
vinculado a las características de la túnica de estas dos especies . En el caso de A. ceratodes numerosos estudios demuestran particularidades químicas de la túnica
que evitan la depredación de los organismos (Frank y Hodgson, 2000; Frank, 2003),
lo que ha sido muy documentado para otras especies de ascidias (Parry, 1984) .
Sin embargo, R. birkelandi presenta una túnica lisa, totalmente desprovista de
organismos epibiontes e incrustaciones, debido a sus características bioquímicas
como alta acidez y altos contenidos de vanadio que la protegen contra la colonización
y depredación (Stoecker 1978, 1980a, 1980b) .
AGRADECIMIENTOS
Para la realización de este trabajo se contó con el financiamiento del Proyecto
FIDA (Vicerrectora Investigación) denominado “Determinación de actividades
farmacológicas y elucidación de estructuras químicas en los macroinvertebrados
(esponjas, corales blandos y ascidias) en algunas zonas del Pacífico y Caribe de Costa
Rica”, de la Universidad Nacional de Costa Rica, y con la colaboración del Instituto
de Oceanología del CITMA, Cuba . Agradecemos el apoyo de las dos instituciones .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 65
BIBLIOGRAFİA
Abbott, D . P . y A . T . Newberry . 1980 . Urochordata: The tunicates . 177-226 . En: Morris R . H ., D . P . Abbott
y E . C . Haderlie (Eds .) . Intertidal invertebrates of California . Stanford University Press, Stanford,
EE . UU . 690 p .
Acuña, J . A ., J . Cortés y M . M . Murillo . 1997 . Mapa de sensibilidad ambiental para derrames de petróleo
en las costas de Costa Rica . Rev . Biol . Trop ., 44 (3)/45 (1): 463-470 .
Bassey, G . 2010 . Evaluación ecológica de los arrecifes y comunidades coralinas de las islas Murciélago
y sección norte de la península de Santa Elena en el Pacífico de Costa Rica . Tesis M . Sc ., Univ .
Nacional de Costa Rica, Heredia . 132 p .
Frank, P . 2003 . The vanadium environment in blood cells of Ascidia ceratodes is divergent at all
organismal levels: an XAS and EPR spectroscopic study . J . Inorg . Biochem ., 94: 59-71 .
Frank, P . y K . O . Hodgson . 2000 . Defining chemical species in complex environments using K-edge
X-ray absorption spectroscopy: vanadium in intact blood cells and Henze solution from the tunicate
Ascidia ceratodes. Inorg . Chem ., 39: 6018-6027 .
Hernández-Zanuy, A . 1999 . Ascidias de las bahías Vita y Jururú, Costa NE de Cuba . Ciencias del Mar
UAS, 2: 1-14 .
Infostat . 2002 . Infostat Version 1 .1 . Grupo Infostat, FCA, Universidad Nacional de Córdoba, Argentina .
Jiménez, C . 2001 . Arrecifes y ambientes coralinos de bahía Culebra, Pacífico de Costa Rica: aspectos
biológicos, económico-recreativos y de manejo . Rev . Biol . Trop ., 49 (2): 215-231 .
Lambert, G . 1993 . Three new species of stolidobranch ascidians (Chordata: Ascidiacea) from the
California continental shelf . Proc . Calif . Acad . Sci ., 48 (4): 109-118 .
Lambert, C . C . y G . Lambert . 1998 . Non-indigenous ascidians in southern California harbors and marinas .
Mar . Biol ., 130: 675-688 .
Lambert, C . C . y G . Lambert . 2003 . Persistence and differential distribution of non-indigenous ascidians
in harbors of the Southern California Bight . Mar . Ecol . Prog . Ser ., 259: 145-161 .
Manso, P ., W . Stolz y J . Fallas . 2005 . El régimen de la precipitación en Costa Rica . Rev . Ambientico,
144: 7-8 .
Monniot, C . y F. Monniot. 1972. Clé mondiale des genres d’ascidies. Arch. Zool. Exp. Gén., 113: 311-
367 .
Monniot, F . y C . Monniot . 1985 . Ascidies littorals de Guadalupe . IX . Caracteristiques des populations .
Ecologie, Rapports avec la faune mondiale . Tethys, 11 (3-4): 203-213 .
Parry, D . 1984 . Chemical properties of the test of ascidians in relation to depredation . Mar . Ecol . Prog .
Ser ., 17: 279-282 .
Ramos, A . A . y J . D . Ros . 1990 . Tipos biológicos de ascidias litorales de sustrato duro . Bentos, 6: 283-
299 .
Statgraphics . 1994-1997 . Statgraphics Plus for Windows 3 .0 . Statistical Graphics Corp . Warrenton, EE .
UU .
Stoecker, D . 1978 . Resistance of a tunicate to fouling . Biol . Bull ., 155: 615-626 .
Stoecker, D . 1980a . Distribution of acid and vanadium in Rhopalaea birkelandi Tokioka . J . Exp . Mar .
Biol . Ecol ., 48: 277-281 .

66 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Stoecker, D . 1980b . Relationships between chemical defense and ecology in benthic ascidians . Mar .
Biol . Prog . Ser ., 3: 257-265 .
Tokioka, T . 1971 . A new species of Rhopalaea from the Pacific coast of Costa Rica (Tunicate, Ascidiacea) .
Publ . Seto Mar . Biol . Lab ., 19 (2/3): 119-122 .
Tokioka, T . 1972 . On small collection of ascidians from the Pacific coast of Costa Rica . Publ . Seto Mar .
Biol . Lab ., 19 (2/3): 119-122 .
Van Name, W . G . 1945 . The North and South American ascidias . Bull . Am . Mus . Nat . Hist ., 81:1-476 .
Vásquez, E . y C . M . Young . 1996 . Rhopalea clonery (Ascidiacea, Cionidae), a new deep-water ascidian
from the coastal waters of British Columbia . J . Zool . Lond ., 238: 599-610 .
FECHA DE RECEPCIÓN: 12/08/2008 FECHA DE ACEPTACIÓN: 06/04/2010

Instituto de Investigaciones Marinas y Costeras - INVEMAR 67
Bol . Invest . Mar . Cost . 39 (1) ISSN 0122-9761 Santa Marta, Colombia, 2010
HIDROZOOS DE LA FAMILIA AGLAOPHENIIDAE DE LA PLATAFORMA CONTINENTAL Y TALUD SUPERIOR DEL
CARIBE COLOMBIANO*
Tibisay Posada P.1, Álvaro L. Peña C.2 y Gabriel R. Navas S.3
1 Instituto de Investigaciones Marinas y Costeras-INVEMAR, Cerro Punta Betín, Santa Marta, Colombia. [email protected] Universidad de Valencia, Instituto Cavanilles de Biodiversidad y Biología Evolutiva, Valencia, España. [email protected] Universidad de Cartagena, Facultad de Ciencias Exactas y Naturales, Departamento de Biología, Cartagena, Colombia. [email protected]
RESUMEN
Se realizó un estudio taxonómico de los hidrozoos de la familia Aglaopheniidae colectados sobre fondos blandos del Caribe colombiano entre 20 y 500 m de profundidad en marzo de 2001 por la expedición INVEMAR MACROFAUNA II . Fueron encontrados dos géneros y seis especies destacándose Aglaophenia elongata, A. lophocarpa y A. tridentata como nuevos registros para aguas colombianas y además se amplió la distribución batimétrica de Aglaophenia latecarinata y A. tridentata . En general los caracteres morfológicos de las especies estudiadas no presentaron diferencias significativas con respecto a las descripciones taxonómicas realizadas por otros autores . En esta publicación se consignan las fichas taxonómicas e imágenes de las especies respectivas y se realizan anotaciones generales sobre su distribución y sustrato .
PALABRAS CLAVE: Hidrozoos, Aglaopheniidae, Caribe colombiano, Plataforma continental, Talud superior .
ABSTRACT
Hydroids of the familiy Aglaopheniidae from the Colombian Caribbean continental shelf and upper slope. A taxonomic study of hydroids of the Aglaopheniidae family collected on soft bottoms of Colombian Caribbean Sea between 20 and 500 m depth in March 2001 was made . Two genera and six species were found, highlighting Aglaophenia elongata, A. lophocarpa and A. tridentata as new records for Colombian waters . Additionally, the bathymetric distribution for Aglaophenia latecarinata and A. tridentata was expanded . In general, morphological characters of the species studied did not show significant differences regarding the taxonomic descriptions made by other authors . This report contains taxonomic files and images of the species described and annotations about general composition, distribution and substrate .
KEY WORDS: Hydroids, Aglaopheniidae, Colombian Caribbean, Continental shelf, Upper slope .
67-81
*Contribución No. 1048 del Instituto de Investigaciones Marinas y Costeras – INVEMAR.

68 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
INTRODUCCIÓN
Los hidrozoos habitan casi todos los ambientes marinos y algunas especies
también viven en aguas dulces o salobres . Pese a su abundancia e importancia
ecológica son el grupo de cnidarios menos conocido y poco estudiado desde el
punto de vista taxonómico, lo cual hace posible que en las colectas que se realizan
generalmente aparezcan novedades (Grohmann, 2009) .
Entre los principales estudios sobre hidrozoos realizados en el Caribe se
encuentran el de Fraser (1937) quien describió diez nuevas especies para la región
de Puerto Rico . Van Gemerden-Hoogeveen (1965) trabajó las familias Sertulariidae, Plumulariidae y Aglaopheniidae de Curazao y otras islas del Caribe . Kelmo y Vargas
(2002) elaboraron las primeras fichas taxonómicas de hidrozoos de los órdenes
Anthoathecatae y Leptothecatae del Caribe y Pacífico de Costa Rica .
En Colombia se destacan los trabajos de Wedler (1973, 1975) donde
describió algunos aspectos ecológicos de hidrozoos del área de Santa Marta . Bandel
y Wedler (1987) realizaron una caracterización de hidrozoos y gasterópodos en la
zona litoral del Caribe colombiano, donde resaltan la importancia de algunas especies
como indicadoras de las zonas supra, meso y sublitoral . El trabajo taxonómico más
representativo para el país es el de Flórez (1983) que registró 15 familias, 31 géneros
y 51 especies en aguas cercanas a Cartagena . En el presente estudio se adicionan tres
primeros registros de hidrozoos al inventario de especies del Caribe colombiano y
se aporta nueva información morfológica y de ámbito batimétrico para seis especies
presentes en el Caribe sur .
ÁREA DE ESTUDIO
Los hidrozoos estudiados fueron recolectados en las ecorregiones Guajira
(GUA), Palomino (PAL), Tayrona (TAY), Magdalena (MAG), Archipiélagos
Coralinos (ARCO) y Darién (DAR), abarcando el área comprendida entre las
coordenadas 12o°34’36.999”-9o0’78.760”N y 72o18’19.000”-77o53’26.999”
O (Figura 1) . Estas ecorregiones fueron definidas con base en geomorfología de
la costa y de la plataforma continental, hidrografía, sedimentología, productividad
biológica de la columna de agua y ecosistemas costeros presentes (Díaz y Gómez,
2000) . Guajira y Palomino en el extremo norte se caracterizan por poseer una
plataforma ancha con fondos arenosos y presentar aguas turbias debido a los
procesos de surgencia . Tayrona presenta características geomorfológicas y
ecológicas determinadas por las estribaciones noroccidentales de la Sierra Nevada

Instituto de Investigaciones Marinas y Costeras - INVEMAR 69
de Santa Marta; su plataforma continental está prácticamente ausente y presenta
baja influencia de las descargas continentales . Magdalena, Darién y Morrosquillo se
caracterizan por la marcada influencia que ejercen las descargas de ciénagas y ríos .
Archipiélagos Coralinos se extiende paralelamente a la ecorregión Morrosquillo, a
partir de los 40 m de profundidad hasta el límite externo de la plataforma continental
(Díaz y Gómez, 2000) .
MATERIALES Y MÉTODOS
Las campañas de recolección del material se realizaron a bordo del buque
de investigaciones Ancón perteneciente al Instituto de Investigaciones Marinas y
Costeras-INVEMAR (Santa Marta) . Las colectas se hicieron por medio de arrastres
por popa con una red de arrastre demersal de tipo semibalón, con dos puertas
metálicas tipo “V” (91 x 63 cm) a 20, 70 y 150 m de profundidad en todas las
ecorregiones y a 300, 400 y 500 m frente a ellas .
Figura 1. Caribe colombiano mostrando cada ecorregión: (GUA) Guajira, (PAL) Palomino, (COC) Caribe oceánico, (TAY) Tayrona, (MAG) Magdalena, (ARCO) Archipiélagos coralinos, (MOR) Morrosquillo, (DAR) Darién .
vv

70 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Las muestras fueron lavadas con agua en la cubierta del buque y preservadas
en alcohol etílico al 96 %; en el laboratorio fueron separadas por morfotipos y
almacenadas . Para la identificación se emplearon las claves y descripciones realizadas
por Allman (1883), Vervoort (1941, 1959, 1968), Van Gemerden-Hoogeveen (1965)
y Bouillon et al . (2004) . Las imágenes fueron obtenidas con una cámara Nikon
D-200 adaptada al microscopio para la observación de cada una de las estructuras
de la hidroteca e hidrocladio . El material fue almacenado en el Museo de Historia
Natural Marina de Colombia (MHNMC) del INVEMAR y la información referente
a la especie y al material examinado como sinonimias, descripción, distribución y
notas del estado hace parte del Sistema de Información sobre Biodiversidad Marina
(SIBM) . Al material de cada especie se le realizaron medidas de los caracteres
morfológicos importantes para la determinación taxonómica (Figura 2) .
Figura 2. Caracteres morfológicos importantes para la determinación de hidrozoos de la familia Aglaopheniidae . (LI): Longitud del internudo . (PH): Profundidad de la hidroteca . (LNL): Longitud de la nematoteca lateral . (LNM): Longitud de nematoteca media . (DMH): Diámetro al margen de la hidroteca . (LDM): Longitud diente medial . (DM): Número de dientes marginales .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 71
RESULTADOS Y DISCUSIÓN
En total fueron estudiadas trescientas colonias de hidrozoos de la familia
Aglaopheniidae pertenecientes a dos géneros y seis especies: Aglaophenia elongata (Meneghini), A. latecarinata Allman, A. lophocarpa Allman, A. tridentata Versluys,
A. rhynchocarpa Allman y Macrorhynchia allmani (Nutting) . Se realizó una clave
para la identificación de las especies de la familia Aglaopheniidae presentes en
el Caribe colombiano basada en la observación y morfometría de los caracteres
importantes para su determinación taxonómica .
Clave taxonómica para los hidrozoos de la familia Aglaopheniidae conocidos en el Caribe colombiano
1a. Colonias con margen hidrotecal sinuoso………………...…..……………......2
1b. Colonias con margen hidrotecal dentado……………………………………....3
2a. Hidrocladios delgados con dos nematotecas en su base……… . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Macrorhynchia allmani (Figura 3)
Figura 3. Macrorhynchia allmani . Hidroteca . En la parte izquierda de la imagen obsérvese la nematoteca mesial unida a toda la pared abcaulinar de la hidroteca. Escala: 200 μm.

72 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
2b. Hidrocladios delgados sin nematotecas en su base………… . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ....………....Aglaophenia tridentata (Figura 4)
Figura 4 . Aglaophenia tridentata. Nematoteca lateral. Escala: 130 μm.
3a. Hidrocaulo con apófisis en su parte basal………………..………..….…………4
3b. Hidrocaulo sin apófisis en su parte basal…...………….…………………….…5
4a . Nematoteca media unida a dos tercios de la pared abcaulinar de la hidroteca; internudos
tecados con longitudes entre 410 y 413 μm.………Aglaophenia elongata (Figura 5)
4b . Nematoteca media corta, hidroteca grande y alargada; internudos tecados con
595 y 600 μm de longitud......…………Aglaophenia rhynchocarpa (Figura 6)
5a. Hidroteca profunda (600-610 μm) sin diafragma interno, nematoteca mesial
corta y nematotecas laterales en forma de copas anchas……………………
……………………………………………Aglaophenia lophocarpa (Figura 7)
5b. Hidrotecas regularmente grandes (304-310 μm) con diafragma interno que
atraviesa la hidroteca, nematoteca medial larga y nematotecas laterales tubulares y
dobladas en su extremo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .Aglaophenia latecarinata (Figura 8)

Instituto de Investigaciones Marinas y Costeras - INVEMAR 73
Figura 5. Aglaophenia elongata. Nematoteca lateral en forma de copa. Escala: 50 μm.
Figura 6. Aglaophenia rhynchocarpa. Hidrocaulo e hidrotecas . (1) Septo en la base de la nematoteca lateral . (2) Septo entre la nematoteca lateral y el diafragma . (3) Septo donde termina el diafragma hidrotecal. (4) Septo en la base del internudo. Escala: 150 μm.

74 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Figura 8. Aglaophenia latecarinata. Margen hidrotecal con dientes marginales curvados hacia el interior . Escala: 120 μm.
Figura 7 . Aglaophenia lophocarpa. Unión del hidrocladio al hidrocaulo . En la parte superior de la imagen la flecha señala el pliegue intratecal que forma el internudo en el hidrocladio. Escala: 380 μm.

Instituto de Investigaciones Marinas y Costeras - INVEMAR 75
Especies estudiadas
Aglaophenia elongata (Meneghini, 1845)(Figura 5)
Material examinado: INVCNI 3224, colectado frente a Manaure (GUA),
22 m de profundidad, tres colonias sin hidrorriza ni córbula, sin sustrato consolidado .
INVCNI 3225, frente a Manaure (GUA), 70 m de profundidad, seis colonias sin
córbulas, sin sustrato consolidado .
Descripción: Colonia con crecimiento monopodial, con apófisis en la parte
basal del hidrocladio . Las hidrotecas presentan un margen dentado con ocho dientes
prominentes con uno de ellos cubierto por las nematotecas laterales y diente medial
agudo de aproximadamente 45 μm. Las nematotecas laterales de esta especie son
pequeñas y en forma de copas con aproximadamente 100 μm de longitud dirigidas
hacia el hidrocladio (Tabla 1) .Distribución: Registrada por primera vez para el Atlántico por Marktanner-
Turneretscher (1890) . Según Vervoort (1941), su distribución es probablemente sobre áreas tropicales y subtropicales de los océanos Atlántico, Índico y Pacífico. En este estudio esta especie se encontró en la ecorregión Guajira en el sector de Manaure .
Observaciones: En general la mayoría de caracteres morfológicos medidos en los ejemplares coincidieron con lo descrito por Allman (1877), Vervoort (1941, 1966), Van Gemerden-Hoogeveen (1965) y Svoboda (1979) . Sin embargo, la profundidad de la hidroteca y la longitud del internudo del hidrocladio mostraron variaciones en la morfometría, considerándose éstas como variabilidades intraespecíficas por diferencias ecológicas y/o poblacionales . Esta especie fue registrada por Ritchie (1909) para las costas de Brasil, siendo éste el primer registro para el Caribe .
Aglaophenia latecarinata Allman, 1877(Figura 8)
Material examinado: INVCNI 3233, frente a Dibulla (PAL), 20 m de profundidad, una colonia sin sustrato consolidado . INVCNI 3228, frente a Dibulla (PAL), 70 m de profundidad, 32 colonias asociadas a hidrozoos de la familia Syntheciidae, conchas de moluscos, fragmentos de esponja, arena y piedras . INVCNI 3235, frente a Dibulla (PAL), 70 m de profundidad, siete colonias sobre tubos de poliquetos y fragmentos de esponjas . INVCNI 3231, frente a Nenguange (TAY), 20 m de profundidad, seis colonias sin sustrato consolidado . INVCNI 3226, frente a Nenguange (TAY), 74 m de profundidad, seis colonias sin córbulas, sin sustrato

76 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
CARACTERES MORFOLÓGICOS Mínimo – Máximo (μm)
Aglaophenia elongata (tomadas en 45 hidrocladios de nueve especímenes)Longitud del internudo del hidrocladio 410-413Profundidad de la hidroteca 430-450Longitud de la nematoteca mesial o media 170-185Longitud de las nematotecas laterales 110-120Longitud del diente medial 43-45
Aglaophenia latecarinata (tomadas en 1340 hidrocladios de 268 especímenes)Longitud del internudo del hidrocladio 290-310Diámetro al margen de la hidroteca 120-126Profundidad de la hidroteca 304-310Longitud de las nematotecas laterales 109-116Número de dientes al margen de la hidroteca 8
Aglaophenia rhynchocarpa (tomadas en 45 hidrocladios de nueve especímenes)Longitud del internudo del hidrocladio 320-350Diámetro al margen de la hidroteca 125-130Profundidad de la hidroteca 315-320Longitud de las nematotecas laterales 100-110Número de dientes al margen de la hidroteca 9
Aglaophenia lophocarpa (tomadas en 18 hidrocladios de un especímen)Longitud del internudo del hidrocladio 760-765Diámetro al margen de la hidroteca 200-210Profundidad de la hidroteca 600-610Longitud de las nematotecas laterales 110-120Número de dientes al margen de la hidroteca 9
Aglaophenia tridentata (tomadas en nueve hidrocladios de un espécimen)Longitud del internudo del hidrocladio 300-305Profundidad de la hidroteca 275-283Longitud de la nematoteca mesial o media 270-280Longitud de las nematotecas laterales 98-100
Macrorhynchia allmani (tomadas en 60 hidrocladios de nueve especímenes)Longitud del internudo del hidrocladio 450-505Profundidad de la hidroteca 166-170Longitud de las nematotecas laterales 120-130
Tabla 1 . Intervalo de medidas de caracteres morfológicos para la determinación de las especies de hidrozoos estudiados .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 77
consolidado . INVCNI 3230, frente a Bahía Concha (TAY), 150 m de profundidad, fragmento de colonia adherido a fragmentos de coral . INVCNI 3227, frente a la isla de Salamanca (MAG), 20 m de profundidad, una colonia sin córbula . INVCNI 3234, frente a la isla de Salamanca, (MAG), 20 m de profundidad, 22 colonias creciendo sobre fragmentos de esponjas y piedras . INVCNI 3238, frente a Morro Hermoso (MAG), 20 m de profundidad, 110 colonias sobre fragmentos de esponjas . INVCNI 3229, frente a Morro Hermoso (MAG), 20 m de profundidad, 35 colonias asociadas a briozoos, sobre fragmentos de algas coralinas y arena . INVCNI 3236, frente a Morro Hermoso (MAG), 70 m de profundidad diez colonias sin presencia de sustrato consolidado . INVCNI 3237, frente a Morro Hermoso (MAG), 75 m de profundidad, 34 colonias sobre fragmentos de coral y esponjas . INVCNI 3232, frente a Morro Hermoso (MAG), 145 m de profundidad, siete colonias sin sustrato consolidado .
Descripción: Las colonias presentan hidrocladios con pequeñas invaginaciones que dan lugar a internudos tecados . Las córbulas tienen diez pares de hojas que cubren la gonoteca . Las hidrotecas poseen un diafragma oblicuo que surge de la pared adcaulinar y atraviesa la hidroteca . El margen hidrotecal tiene dos dientes más agudos y curvados hacia el interior sobre el margen hidrotecal . Las nematotecas laterales son tubulares, dirigidas basalmente hacia el margen de la hidroteca y en su parte distal se doblan en dirección al hidrocladio, poseen longitudes entre 109 y 116 μm aproximadamente como se referencia en la Tabla 1.
Distribución: Según Vervoort (1959), esta especie presenta una amplia distribución en el Atlántico occidental y ha sido registrada en Bahamas, el Golfo de México y Brasil . En el Caribe colombiano A. latecarinata ha sido recolectada en el área de Cartagena (Flórez, 1983) y acá se registra para las ecorregiones Palomino, Tayrona y Magdalena .
Observaciones: Los caracteres morfológicos presentaron las mismas características descritas por Broch (1913), Van Gemerden-Hoogeveen (1965), García-Corrales et al. (1978), Izquierdo et al. (1986), Kelmo y Vargas (2002) y Varela et al. (2005) . En algunos individuos se observaron variaciones en el grado de desarrollo de los dientes marginales ocasionados posiblemente por variabilidad intraespecífica . Se registra por primera vez entre 20 y 150 m siendo ampliada así su distribución batimétrica .

78 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Aglaophenia rhynchocarpa Allman, 1877
(Figura 6)
Material examinado: INVCNI 3239, frente a Dibulla (PAL), 70 m de
profundidad, siete colonias, sin presencia de sustrato consolidado .
Descripción: Colonia con hidrocaulo dividido en internudos formados por
septos oblicuos, siendo estos más visibles en las partes jóvenes de la colonia . En
el internudo tecado se presentan cuatro septos internodales: uno en la base de la
nematoteca lateral, uno entre la nematoteca lateral y el diafragma hidrotecal, uno
en el sitio donde termina el diafragma hidrotecal y uno en la base del internudo . Se
presentan longitudes del internudo entre 320 y 350 μm (Tabla 1).
Distribución: Se encuentra restringida al Atlántico occidental tropical,
presentándose principalmente en la región Caribe . En el Caribe colombiano fue
encontrada en sectores de Cartagena (Flórez, 1983) y en la ecorregión Palomino .
Observaciones: La profundidad de la hidroteca, el diámetro del margen
hidrotecal y la nematoteca lateral son de menor tamaño que los conocidos para
ejemplares provenientes del área de Cartagena . Es posible que esta variación esté
influenciada por las diferencias ecológicas entre los sitios de colecta .
Aglaophenia lophocarpa Allman, 1877
(Figura 7)
Material examinado: INVCNI 3240, frente a Puerto Escondido (DAR), 70
m de profundidad, una colonia, sin presencia de sustrato consolidado .
Descripción: Hidrocaulo desprovisto en su parte basal de hidrocladios y
nematotecas . En la unión del hidrocladio al hidrocaulo presenta una nematoteca
redondeada . La córbula presenta diez pares de hojas separadas entre sí con sus
bordes armados con una fila de nematotecas a lado y lado de cada hoja . La hidroteca
es estrecha y tiene una profundidad entre 600 y 610 μm (Tabla 1).
Distribución: Registrada en África en sectores de Guinea (Gili et al ., 1989) . Fraser (1947) menciona su presencia en el océano Pacífico . El material fue
recolectado en la ecorregión Darién en el sector de Puerto Escondido .
Observaciones: Las características morfológicas de esta especie descritas
por Nutting (1900), Gili et al . (1989), Svoboda (1979, 1992) y Bouillon et al . (2004)
concuerdan con la morfometría de la única colonia encontrada en la muestra . Los
ejemplares capturados constituyen el primer registro de la especie para el Caribe
colombiano .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 79
Aglaophenia tridentata Versluys, 1899
(Figura 4)
Material examinado: INVCNI 3241, frente al golfo de Salamanca (MAG),
70 m de profundidad una colonia, sin sustrato consolidado .
Descripción: Colonias con hidrocaulo grueso . En los internudos de los
hidrocladios se presenta una hidroteca con una invaginación de su pared abcaulinar
lo cual hace que la nematoteca medial sea alargada (entre 270 y 280 μm). Las
nematotecas laterales son cilíndricas y alargadas con longitudes entre 98 y 100 μm
(Tabla 1) .
Distribución: Fue registrada por Vervoort (1968) para el Atlántico
occidental . Kelmo y Vargas (2002) la registran por primera vez para aguas de Costa
Rica . El material fue recolectado en la ecorregión Magdalena en el sector del golfo
de Salamanca .
Observaciones: En esta colonia se observó la presencia de un hidrozoo
lafoeideo, los cuales suelen encontrarse como epibiontes reptando en el hidrocaulo
de Aglaophenia. Constituye el primer registro de Aglaophenia tridentata para aguas
del Caribe colombiano . Hasta el momento la literatura no registra ámbito batimétrico
para esta especie, en el área de estudio se encontró a 70 m de profundidad en el golfo
de Salamanca .
Macrorhynchia allmani (Nutting, 1900)
(Figura 3)
Material examinado: INVCNI 3245, frente al golfo de Salamanca (MAG),
70 m de profundidad, un fragmento de colonia sin sustrato consolidado . INVCNI
3243, frente a Tolú (ARCO), 70 m de profundidad, cuatro colonias con diminutas
conchas de bivalvos y piedras pequeñas en la hidrorriza . INVCNI 3242, frente a Tolú
(ARCO), 270 m de profundidad, seis colonias sin sustrato . INVCNI 3244, frente a
Puerto Escondido (DAR), 70 m de profundidad, una colonia con diminutas piedras
en su hidrorriza .
Descripción: Las colonias presentan ramificaciones secundarias de los
hidrocladios . En la base de los hidrocladios poseen dos nematotecas grandes de forma
triangular . La nematoteca media está unida en su totalidad a la pared abcaulinar de
la hidroteca y alcanza a sobrepasar el margen hidrotecal . Las hidrotecas tienen una
profundidad entre 166 y 170 μm y poseen un septo intratecal muy pronunciado (Tabla 1).

80 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Distribución: Se ha registrado únicamente en la parte occidental del océano
Atlántico . En el Caribe colombiano fue registrada en Santa Marta por Wedler (2004) .
En la zona de estudio fue encontrada en las ecorregiones Magdalena, Archipiélagos
Coralinos y Darién .
Observaciones: La morfometría del material recolectado en el Caribe
colombiano muestra gran similitud con la descrita por Nutting (1900), Van
Gemerden-Hoogeveen (1965) y Wedler (2004), aunque presenta algunas diferencias
con las longitudes descritas por Van Gemerden-Hoogeveen (1965), lo cual puede ser
atribuido a la edad de la colonia, siendo más jóvenes las colonias descritas por este
autor con respecto a las recolectadas en el Caribe colombiano .
AGRADECIMIENTOS
Los autores expresan sus sinceros agradecimientos a los doctores Adriana
Bermúdez de la Universidad de Cartagena y Arturo Acero de la Universidad Nacional
de Colombia sede Caribe por su revisión en la redacción del documento . Al Instituto
de Investigaciones Marinas y Costeras “José Benito Vives de Andreis”-INVEMAR
y a la Agencia Nacional de Hidrocarburos por el apoyo prestado para la realización
de esta publicación .
BIBLIOGRAFÍA
Allman, G . 1877 . Report on the Hydroida collected during the exploration of the Gulf Stream by L . F . de
Pourtalés . Museum of Comparative Zoology Harvard, 5 (2): 1-66 .
Allman, G. J. 1883. Report on the Hydroida dredged by “H. M. S. Challenger” during the years 1873-
1876, Part 1 - Plumularidae . Report of the scientific results of the voyage of H .M .S . Challenger
during the years 1873-76, Zoology, 20 (7), 55 p .
Bandel, K . y E . Wedler . 1987 . Hydroid, amphineuran and gastropod zonation in the littoral of the
Caribbean Sea, Colombia . Senckenbergiana Marit ., 19 (1/2):1-129 .
Bouillon, J ., M . D . Medel, F . Pagès, J . M . Gili, F . Boero y C . Gravili . 2004 . Fauna of the Mediterranean
Hydrozoa . Sci . Mar ., 68 (suppl .2): 1-454 .
Broch, H. 1913. Hydroida from the “Michael Sars” North Atlantic deep-sea expedition 1910. Report on
the scientific results of the “Michael Sars” North Atlantic Deep-Sea Expedition 1910, 3 (1): 1-18.
Díaz, J . M . y D . I . Gómez . 2000 . Programa nacional de investigaciones en biodiversidad marina . Instituto
de Investigaciones Marinas y Costeras-INVEMAR y Ministerio de Medio Ambiente, Bogotá . 83 p .
Flórez, L . 1983 . Inventario preliminar de la fauna hydroide de la Bahía de Cartagena y áreas adyacentes .
Bol . Mus . Mar, (11):112-140 .
Fraser, C . M . 1937 . New species of hydroids from the Puerto Rico Region . Smith . Misc . Coll ., 91 (28):
1-7 .
Fraser, M . 1947 . Hydroids of the 1939 Allan Hancock Caribbean Sea Expedition . Allan Hancock Atl .
Exped . Rep ., (4): 1-24 .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 81
García-Corrales, P ., A . Aguirre y D . González . 1978 . Contribución al conocimiento de los hidrozoos de
las costas españolas: Parte 1 . Halécidos, campanuláridos y plumuláridos . Bol . Inst . Esp . Oceanogr .,
4 (253): 5-73 .
Gili, J . M ., W . Vervoort y F . Pagès . 1989 . Hydroids from the West African coast: Guinea Bissau, Namibia
and South Africa . Scient . Mar ., 53 (1): 67-112 .
Grohmann, P . 2009 . Hydroids (Cnidaria, Hydrozoa) of the intertidal zone of Governador and Paquetá
islands, Guanabara Bay, Rio de Janeiro, Brazil . Iheringia Série Zoologia, 99 (3): 291-294 .
Izquierdo, M . S ., P . García-Corrales y J . J . Bacallado . 1986 . Contribución al conocimiento de los hidrozoos
caliptoblástidos del Archipiélago Canario, parte I: Haleciidae, Lafoeidae, Campanulariidae y
Syntheciidae . Bol . Inst . Esp . Oceanogr ., 3 (1): 81-94 .
Kelmo, F . y R . Vargas . 2002 . Anthoathecatae and Leptothecatae hydroids from Costa Rica (Cnidaria:
Hydrozoa) . Rev . Biol . Trop ., 50 (2): 599-627 .
Marktanner-Turneretscher, G . 1890 . Die Hydroiden des K .K .naturhistorischen Hofmuseums . Ann . Nat .
Hist . Hofmus . Wien ., 5: 195-286 .
Nutting, C .C . 1900 . American hydroids, part I, the Plumularidae . Smithsonian Inst ., US Nat . Mus . Spec .
Bull ., 4 (1): 1-285 .
Ritchie, J . 1909 . Supplementary report on the hydroids of the Scottish National Antarctic Expeditions .
Transact . Royal Soc . Edinb ., 47: 65-101 .
Svoboda, A . 1979 . Beitrag zur Ökologie, biometrie und Systematic der Mediterranen Aglaophenia Arten
(Hydroidea) . Zool . Verh. Leiden, 167: 114 p .
Svoboda, A . 1992 . Sexual dimorphism in Aglaophenia tubulifera and Aglaophenia lophocarpa (Cnidaria,
Hydrozoa) . Sci . Mar., 56 (2-3): 177-183 .
Van Gemerden-Hoogeveen, G . C . H . 1965 . Hydroids of the Caribbean: Sertulariidae, Plumulariidae and
Aglaopheniidae . Stud . Fauna Curaçao Carib . Isl ., 22 (84): 1-87 .
Varela, C ., M . Ortiz y R . Lalana . 2005 . Nuevas consignaciones de hidrozoos marinos y confirmación de
la presencia de Aglaophenia latecarinata (Cnidaria:Hydrozoa), para aguas cubanas . Rev . Invest .
Mar ., 26 (2): 177-179 .
Vervoort, W . 1941 . Biological results of the Snellius Expedition XI . The hydroida of the Snellius
Expedition (Milleporidae and Stylasteridae excluded) . Temminckia, 6: 186-224 .
Vervoort, W . 1959 . The hydroida of the tropical West coast of Africa . Atlantide Report . Scientific Result
of the Danish Expedition to the Coasts of Tropical West Africa 1945-1946, 5: 211-325 .
Vervoort, W . 1966 . Bathyal and abissal hydroids . Galathea Report, Scientific Results of the Danish Deep
Sea Expedition, 1950-1952, 8: 97-173 .
Vervoort, W . 1968 . Report on a collection of Hydroida from the Caribbean region, including an annotated
checklist of Caribbean hydroids . Zool . Verh . Leiden, 92: 1-124 .
Wedler, E . 1973 . Die Hydroiden der Ciénaga Grande de Santa Marta (Kolumbien) und einiges zu ihrer
Ökologie. Mitt . Inst . Colombo-Alemán Invest . Cient ., 7: 31-39 .
Wedler, E . 1975 . Ökologische Untersuchungen an Hydroiden des Felslitorals von Santa Marta
(Kolumbien) . Helgoländ . wiss . Meeresun ., 27 (3): 324-363 .
Wedler, E . 2004 . Inducción de formación de gonóforos de hidrozoos en el medio natural para fines
taxonómicos . Intrópica, 1: 85-90 .
FECHA DE RECEPCIÓN: 21/04/2008 FECHA DE ACEPTACIÓN: 06/04/2010


Instituto de Investigaciones Marinas y Costeras - INVEMAR 83
Bol . Invest . Mar . Cost . 39 (1) ISSN 0122-9761 Santa Marta, Colombia, 2010
LISTADO DE DIATOMEAS (BACILLARIOPHYTA) REGISTRADAS PARA
EL MAR CARIBE COLOMBIANO*
Yesid Lozano-Duque1, Luis Alfonso Vidal2 y Gabriel R. Navas S.3
1 Universidad del Valle, Facultad de Ciencias, Departamento de Biología, Grupo de Investigación en Estuarios y Manglares. Calle 13 No 100-00. Cali, Colombia. [email protected], [email protected] Santa Marta, Colombia. [email protected] 3 Universidad de Cartagena, Facultad de Ciencias Exactas y Naturales, Departamento de Biología. Cartagena, Colombia. [email protected]
RESUMEN
Se realizó una revisión bibliográfica de la información disponible sobre las especies registradas de diatomeas (Bacillariophyta) para las zonas costera y oceánica del mar Caribe colombiano . Se presenta el listado de 337 taxones correspondientes a 312 especies, 19 variedades y siete formas, agrupadas en 54 familias y 106 géneros . La clasificación taxonómica se basó en el sistema propuesto por Round et al . (1990) .
PALABRAS CLAVE: Fitoplancton, Diatomophyceae, Bacillariophyta, Mar Caribe, Plancton .
ABSTRACT
Check-list of diatoms (Bacillariophyta) reported from the Colombian Caribbean Sea . We conducted a literature review of available information on the species of marine (Bacillariophyta) diatoms reported for coastal and ocean Colombian Caribbean Sea . We present a checklist of 337 taxa for 312 species, 19 varieties and seven forms, grouped into 54 families and 106 genera . The taxonomic classification was based on the system proposed by Round et al . (1990) .
KEY WORDS: Phytoplankton, Diatomophyceae, Bacillariophyta, Caribbean Sea, Plankton .
83-116
*Contribución No. 1049 del Instituto de Investigaciones Marinas y Costeras – INVEMAR.

84 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
INTRODUCCIÓN
Las diatomeas se caracterizan por la presencia de una pared celular o
frústula compuesta de sílice; la cual se divide en una parte superior (epiteca) y una
parte inferior (hipoteca) . La estructura y ornamentación de la frústula son la base de
la clasificación de las diatomeas (Hasle y Fryxell, 1995) . Las diatomeas se dividen
en dos grandes grupos por su morfología: las formas céntricas con valvas simétricas
radialmente y las formas pennadas con valvas simétricas bilateralmente . Según la
clasificación de diatomeas propuesta por Round et al . (1990), éstas pertenecen a una
división con tres clases, la clase Coscinodiscophyceae, donde se clasifican todas las
diatomeas céntricas, y las clases Fragilariophyceae y Bacillariophyceae dentro de las
cuales se agrupan las diatomeas pennadas .
Con más de 150 años de observación microscópica, el conocimiento de la
historia de vida en diatomeas sigue siendo escaso en comparación con otros grupos
de algas y plantas superiores . Las recientes comunicaciones entre investigadores
sugieren que actualmente se reconocen aproximadamente 20000 especies de
diatomeas, aunque este número podría alcanzar en el futuro 105-106 especies
(Edlund y Stoermer, 1997) . Una mayor comprensión de la historia de vida de las
diatomeas ha dado lugar a la aplicación de un nuevo enfoque de su clasificación .
Para la mayor parte del siglo XX, la taxonomía y sistemática de diatomeas se
ha basado en la morfología de la frústula . Sin embargo, dos acontecimientos
han cambiado radicalmente la visión de la sistemática en diatomeas (Edlund y
Stoermer, 1997) . En primer lugar, el advenimiento de la microscopía electrónica
de transmisión y de barrido dio una visión más detallada de la morfología frustular,
mejorando considerablemente nuestra comprensión de la taxonomía y la estructura
de diatomeas . En segundo lugar, el redescubrimiento de algunos trabajos que se
basaron en características citológicas .
El número existente de especies y géneros de fitoplancton marino aún es
desconocido, a pesar de su importante interés en estudios de ecología, ficología y
evolución . Sournia et al . (1991) publicaron una sinopsis de los géneros oceánicos
a nivel mundial indicando el número de especies de cada género . Estos autores
registraron 77 géneros de diatomeas céntricas con 865-999 especies y 87 géneros
para diatomeas pennadas con 500-784 especies . Las diatomeas marinas han
sido estudiadas en el mar Caribe desde 1857 con varios trabajos realizados por
investigadores muy reconocidos en la taxonomía de este grupo de microalgas
(Navarro y Hernández-Becerril, 1997) . Margalef (1969), en su estudio del ecosistema
pelágico del NE de Venezuela, registra 300 especies fitoplanctónicas; sin embargo
con el aporte de otros autores y la información publicada de otras áreas del Caribe

Instituto de Investigaciones Marinas y Costeras - INVEMAR 85
no menos de 450 especies podrían ser mencionadas . Navarro y Hernández-Becerril
(1997) realizaron una revisión de literatura disponible sobre diatomeas marinas y un
listado de las especies citadas hasta ese momento para el área total del mar Caribe .
Estos autores registraron 1083 taxa de diatomeas marinas, 305 taxa pertenecientes a
diatomeas céntricas y 778 taxa pertenecientes a diatomeas pennadas . En la Tabla 1 se
presentan los resultados de dicho trabajo de Navarro y Hernández-Becerril (1997) .
El estudio del fitoplancton marino en el mar Caribe colombiano se registra desde
los trabajos de Arosemena et al . (1973), Caycedo (1975, 1977) y Vidal y Carbonell
(1977), los cuales fueron realizados en aguas costeras .
Familias No. Géneros
No. Especies y variedades Familias No.
GénerosNo. Especies y variedades
Thalassiosiraceae 2 13 Psammodiscaceae 1 1Skeletonemataceae 2 4 Ardissoneaceae 1 5Stephanosdiscaceae 1 5 Toxariaceae 1 2Lauderiaceae 1 2 Thalassionemataceae 2 6Chrysanthemodiscaceae 1 1 Rhabdonemataceae 1 3Melosiraceae 1 6 Striatellaceae 4 29Stephanopyxidaceae 1 3 Cyclophoraceae 1 1Endictyaceae 3 7 Climacospheniaceae 2 4Paraliaceae 1 2 Protoraphidaceae 2 2Aulacoseiraceae 1 2 Lyrellaceae 1 17Coscinodiscaceae 2 28 Mastogloiaceae 3 91Aulacodiscaceae 1 2 Dictyoneidaceae 1 2Gossleriellaceae 1 1 Rhoicospheniaceae 1 1Hemidiscaceae 4 17 Achnanthaceae 1 15Heliopeltaceae 1 4 Cocconeidaceae 3 29Ethmodiscaceae 1 1 Berkeleyaceae 3 8Asterolampraceae 2 10 Diadesmidaceae 1 1Triceratiaceae 8 37 Amphipleuraceae 1 2Plagiogrammaceae 3 22 Scoliotropidaceae 4 4Biddulphiaceae 4 18 Sellaphorineae 2 3Hemiaulaceae 5 10 Pinnulariaceae 3 33Bellerocheaceae 1 3 Diploneidaceae 1 49Streptothecaceae 1 2 Naviculaceae 5 88Anaulaceae 2 4 Pleurosigmataceae 4 60Lithodesmiaceae 2 3 Plagiotropidaceae 2 11Corethraceae 1 1 Stauroneidaceae 2 8Cymatosiraceae 1 1 Catenulaceae 2 61Rhizosoleniaceae 5 21 Bacillariaceae 8 115Chaetocerotaceae 2 71 Rhopalodiaceae 1 3Leptocylindraceae 1 3 Entomoneidaceae 1 2Fragilariaceae 14 30 Auriculaceae 1 3Licmophoraceae 1 28 Surirellaceae 4 38Rhaphoneidaceae 3 7 Totales 150 c .a 1067
Tabla 1. Listado de familias y su número de géneros y taxones para el mar Caribe según Navarro y Hernández-Becerril (1997) .

86 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Actualmente para el mar Caribe colombiano no se encuentra una recopilación
de la información disponible sobre las especies fitoplanctónicas presentes, ni guías
publicadas para la identificación de especies y la información que existe en gran parte
está sin publicar . En este trabajo se realizó la recopilación de información y revisión
nomenclatural de las diatomeas marinas citadas para la zona costera y oceánica de
Colombia incluyendo tesis de pregrado e informes de salidas de campo que aunque
no se consideran publicaciones como tal, constituyen una fuente muy importante de
información del grupo a nivel local . El propósito es proporcionar una lista que sirva
como material de base y referencia .
MATERIALES Y MÉTODOS
Se llevó a cabo la revisión bibliográfica de estudios realizados en la
identificación de especies de diatomeas fitoplanctónicas en aguas costeras y oceánicas
en el mar Caribe colombiano . Se consultaron los informes, tesis y publicaciones
disponibles en centros de documentación como el del Instituto de Investigaciones
Marinas y Costeras–INVEMAR y en bibliotecas de las universidades Jorge Tadeo
Lozano, del Magdalena y del Atlántico . Además, se consultaron bases de datos,
revistas e información disponible en Internet .
La presentación del listado de especies especifica la distribución espacial
en la zona utilizando la división política por departamentos (dado el tipo y la poca
información adquirida de las fuentes consultadas) . Esta lista estará disponible a
través del Sistema de Información sobre Biodiversidad Marina del INVEMAR .
En esta lista se consideraron sólo los trabajos que describieron hasta el nivel
de especie . Sólo en tres casos se presentaron a nivel de género por considerarse
importantes por su escaso registro para el área . Las especies fueron corregidas en
su nomenclatura y autores y se confirmaron los nombres aceptados actualmente .
Cuando el nombre de una especie citado por un autor fue corregido por su nombre
aceptado actualmente, seguido de este último se colocó un (=) para indicar que era
una sinonimia, y cuando se presentó un error de identificación o taxón mal escrito,
se colocó (como) . En estos casos se escribió el nombre de la especie citado por el
autor y que no es aceptado actualmente; y con los símbolos de asteriscos (* y **)
se relaciona el autor que citó el nombre no aceptado . Es decir, cuando se actualizó
el nombre de una especie, con un asterisco se relacionó un autor y el nombre de la
especie que cita este autor en su trabajo pero que ya no es aceptado; dos asteriscos
relacionan lo mismo pero con otro autor que cita otro nombre de la especie que ya no
es aceptado . En la columna donde se indica la distribución en Colombia se escribió
el departamento donde se realizó el trabajo con las siguientes siglas bl (Bolívar),

Instituto de Investigaciones Marinas y Costeras - INVEMAR 87
lg (La Guajira), ma (Magdalena) y sp (San Andrés y Providencia) . La clasificación
taxonómica se basó en Round et al . (1990) . Para la revisión de los nombres
taxonómicos y las sinonimias se consultó la base de datos www .marinespecies .org,
y como apoyo se consultaron www .catalogueoflife .org y los trabajos de Schmidt
(1874-1959), Stosch y Simonsen (1984), Round et al . (1990), Snoeijs (1993), Snoeijs
y Vilbaste (1994), Hasle y Fryxell (1995), Snoeijs y Potapova (1995), Hartley et al. (1996), Snoeijs y Kasperoviciene (1996), Tomas (1996), Snoeijs y Balashova (1998)
y Sunesen y Sar (2007) .
RESULTADOS
Este listado de especies de diatomeas citadas en estudios realizados en la
zona oceánica y costera del mar Caribe colombiano presenta 337 taxones con 312
especies, 19 variedades y siete formas . De éstas, 151 especies, tres variedades y seis
formas pertenecen a formas céntricas (clase Coscinodiscophyceae) y 160 especies,
16 variedades y una forma pertenecen a formas pennadas (Clases Fragilariophyceae
y Bacillariophyceae) . Estos taxones son representantes de 54 familias de las cuales
26 pertenecen a céntricas y 28 a formas pennadas, y 106 géneros con 50 céntricas
y 56 pennadas . A continuación se presentan en la Tabla 2 las especies citadas en el
mar Caribe colombiano según la bibliografía consultada .
Tabla 2. Listado de especies de diatomeas registradas para el mar Caribe colombiano según la bibliografía consultada . Bolívar (bl), La Guajira (lg), Magdalena (ma) y San Andrés y Providencia (sp), sinonimia (=), error de identificación o de escritura (como), en estos casos se escribió el nombre de la especie citado por el autor y que no es aceptado actualmente (* y **) se relaciona el autor que citó el nombre no aceptado .
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
División Bacillariophyta (Haeckel 1878)Clase Coscinodiscophyceae Round et Crawford 1990Subclase Thalassiosirophycidae Round et Crawford 1990Orden Thalassiosirales Glezer et Makarova 1986
Familia Thalassiosiraceae Lebour 1930
Género Planktoniella Schütt 1892
Planktoniella sol (Wallich) Schutt 1983 bl lg ma Duarte (1996), Ramos (2005), Suárez (2007)
Género Thalassiosira Cleve 1873

88 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
Thalassiosira allenii Takano 1965 ma Fernández et al . (1999)
Thalassiosira decipiens (Grunow) Jørgensen 1905 sensu Hasle, 1979
ma Ramos (2005)
Thalassiosira eccentrica (Ehrenberg) Cleve 1904
= Coscinodiscus eccentricus Ehrenberg 1841*
bl ma Vidal y Carbonell (1977), Fernández et al. (1999), *Ramos (2005)
Thalassiosira lineata Jousé 1968
= Coscinodiscus lineatus Ehrenberg 1839* ma *Caycedo (1975, *1977), Ramos (2005)
Thalassiosira punctigera (Castracane) Hasle 1983
ma Ramos (2005)
Thalassiosira symmetrica Fryxell et Hasle 1972
ma Fernández et al . (1999)
Familia Skeletonemataceae Lebour 1930 emend . Round 1990
Género Detonula Schütt 1894
Detonula pumila (Castracane) Schütt 1896 bl Vidal y Carbonell (1977), Carbonell (1979)
Detonula moseleyana (Castracane) Gran 1900
bl Carbonell (1979)
Género Skeletonema Greville 1865
Skeletonema costatum (Greville) Cleve 1878 bl ma sp Caycedo (1975, 1977), Vidal y Carbonell (1977), Carbonell (1979), Márquez y Herrera (1986), Arias y Durán (1982, 1984), Cañón et al. (2005), Gavilán et al . (2005), Ramos (2005), Franco-Herrera y Torres-Sierra (2007), Suárez (2007)
Skeletonema tropicum Cleve 1873 ma Caycedo (1975, 1977)
Familia Stephanodiscaceae Glezer et Makarova 1986
Género Cyclotella (Kützing) Brébisson 1838
Cyclotella glomerata Bachmann 1911 ma Ramos (2005)
Cyclotella meneghiniana Kützing 1844 bl Suárez (2007)
Cyclotella planctonica Brunnthaler 1901 ma Ramos (2005)
Familia Lauderiaceae (Schütt) Lemmermann 1899, emend . Round et Crawford 1990
Género Lauderia Cleve 1873
Lauderia annulata Cleve 1873
= Lauderia borealis Gran 1900* bl ma sp *Márquez y Herrera (1986), *Ramos (2005), Suárez (2007)
Subclase Coscinodiscophycidae Round et Crawford 1990
Continuación Tabla 2.

Instituto de Investigaciones Marinas y Costeras - INVEMAR 89
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
Orden Melosirales Crawford 1990Familia Melosiraceae Kützing 1844 emend. Crawford 1990
Género Melosira Agardh 1824
Melosira moniliformis (Müller) Agardh 1783 bl Vidal y Carbonell (1977), Gavilán et al. (2005), Suárez (2007)
Melosira nummuloides Agardh, 1824 emend . Crawford 1975
bl Suárez (2007)
Melosira pseudogranulata Cleve-Euler 1948 bl Vidal y Carbonell (1977), Arias y Durán (1982, 1984), Gavilán et al . (2005)
Familia Stephanopyxidaceae Nikolaev 1988
Género Stephanopyxis Ehrenberg 1844
Stephanopyxis palmeriana (Greville) Grunow 1884
bl Carbonell (1979)
Stephanopyxis turris (Greville) Ralfs in Pritchard 1861
lg ma Duarte (1996), Franco-Herrera y Torres-Sierra (2007)
Familia Endictyaceae Crawford 1990
Género Endictya Ehrenberg 1845
Endyctia oceanica Ehrenberg 1845 bl Vidal y Carbonell (1977), Suárez (2007)
Orden Paraliales Crawford 1990Familia Paraliaceae Crawford 1988
Género Paralia Heiberg 1863
Paralia sulcata (Ehrenberg) Cleve 1873 ma Fernández et al . (1999), Ramos (2005)
Orden Aulacoseirales Crawford 1990Familia Aulacoseiraceae Crawford 1990
Género Aulacoseira Thwaites 1848
Aulacoseira granulata (Ehrenberg) Simonsen 1979
= Melosira granulata (Ehrenberg) Ralfs 1861*
bl *Cañón et al . (2005), *Gavilán et al . (2005)
Orden Coscinodiscales Round et Crawford 1990Familia Coscinodiscaceae Kützing 1844 emend . Round et Crawford 1990
Género Coscinodiscus Ehrenberg emend . Rattray 1890
Coscinodiscus argus Ehrenberg 1838 bl Carbonell (1979)
Coscinodiscus asteromphalus Ehrenberg 1811
bl Vidal y Carbonell (1977), Carbonell (1979), Arias y Durán (1982, 1984), Cañón et al. (2005), Gavilán et al. (2005), Suárez (2007)
Continuación Tabla 2.

90 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
Coscinodiscus centralis Ehrenberg 1844 bl Arias y Durán (1982, 1984), Cañón et al . (2005), Gavilán et al . (2005)
Coscinodiscus gigas Ehrenberg 1841 bl lg ma Vidal y Carbonell (1977), Carbonell (1979), Arias y Durán (1982, 1984), Duarte (1996), Gavilán et al . (2005), Ramos (2005), Franco-Herrera y Torres-Sierra (2007), Suárez (2007)
Coscinodiscus granii Gough 1905 bl ma Cañón et al . (2005), Gavilán et al . (2005), Ramos (2005), Franco-Herrera y Torres-Sierra (2007), Suárez (2007)
Coscinodiscus jonesianus (Greville) Ostenfeld 1915
ma Ramos (2005)
Coscinodiscus marginatus Ehrenberg 1841 ma Franco-Herrera y Torres-Sierra (2007)
Coscinodiscus perforatus Ehrenberg 1844 bl Arosemena et al . (1973)
Coscinodiscus radiatus Ehrenberg 1841 bl ma sp Arosemena et al . (1973), Carbonell (1979), Arias y Durán (1982, 1984), Márquez y Herrera (1986), Cañón et al . (2005), Gavilán et al . (2005), Ramos (2005), Franco-Herrera y Torres-Sierra (2007), Suárez (2007)
Coscinodiscus reniformis Castracane 1886 ma Fernández et al . (1999)
Coscinodiscus thorii Pavillard 1925 bl Vidal y Carbonell (1977), Gavilán et al . (2005)
Coscinodiscus wailesii Gran et Angst 1931 bl Suárez (2007)
Familia Gossleriellaceae Round 1990
Género Gossleriella Schütt 1893
Gossleriella tropica Schütt 1893 ma Ramos (2005)
Familia Hemidiscaceae Hendey 1937 emend . Simonsen 1975
Género Actinocyclus Ehrenberg 1837
Actinocyclus normanii (W . Gregory) Hustedt 1957
ma Ramos (2005)
Actinocyclus octonarius Ehrenberg 1838 bl ma Vidal y Carbonell (1977), Ramos (2005)
Actinocyclus subtilis (Gregory) Ralfs in Pritchard 1861
bl Vidal y Carbonell (1977), Carbonell (1979)
Género Azpeitia Peragallo in Tempére et Peragallo 1912
Azpeitia africana (Janisch in A . Schmidt) Fryxell et Watkins 1986
ma Fernández et al . (1999), Ramos (2005)
Azpeitia nodulifera (Schmidt) Fryxell et Sims in Fryxell et al . 1986
= Coscinodiscus nodulifer A . Schmidt 1878* bl ma *Carbonell (1979), Ramos (2005)
Continuación Tabla 2.
Continuación Tabla 2.

Instituto de Investigaciones Marinas y Costeras - INVEMAR 91
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
Género Palmeria Greville 1865
Palmeria hardmaniana Greville 1865
= Hemidiscus hardmanianus (Greville) Mann 1907
bl *Vidal y Carbonell (1977), Carbonell (1979), *Ramos (2005), Suárez (2007)
Familia Heliopeltaceae H .L . Smith 1872
Género Actinoptychus Ehrenberg 1839
Actinoptychus splendens (Shadbolt) Ralfs in Pritchard 1861
ma Ramos (2005)
Orden Asterolamprales Round et Crawford 1990Familia Asterolampraceae H .L . Smith 1872 emend . Gombos 1980
Género Asterolampra Ehrenberg 1844
Asterolampra marylandica Ehrenberg 1844 bl sp Vidal y Carbonell (1977), Márquez y Herrera (1986)
Género Asteromphalus Ehrenberg 1844
Asteromphalus cleveanus Grunow 1876 ma Ramos (2005)
Asteromphalus flabellatus (Brébisson) Greville 1859
bl ma Carbonell (1979), Ramos (2005)
Asteromphalus heptactis (Brébisson) Ralfs 1861
Como Asteromphalus flabellatus (Brébisson) Greville 1859*
ma *Vidal y Carbonell (1977), Ramos (2005)
Subclase Biddulphiophycidae Round et Crawford 1990Orden Triceratiales Round et Crawford 1990Familia Triceratiaceae (Schütt) Lemmermann 1899
Género Cerataulus Ehrenberg 1843
Cerataulus smithii Ralfs 1861 ma Fernández et al . (1999), Ramos (2005)
Cerataulus turgidus (Ehrenberg) Ehrenberg 1843
ma Ramos (2005)
Género Eupodiscus Bailey 1851
Eupodiscus radiatus Bailey 1851 ma Fernández et al . (1999), Ramos (2005)
Género Lampriscus A . Schmidt 1882
Lampriscus shadboltianum (Greville) Peragallo et Peragallo 1897-1908
= Triceratium shadboltianum Greville 1862* bl *Vidal y Carbonell (1977), *Carbonell (1979, 1980)
Género Odontella Agardh 1832
Odontella aurita (Lyngbye) Agardh 1832 bl Suárez (2007)
Continuación Tabla 2.

92 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
Odontella longicruris (Greville) Hoban 1983
= Biddulphia longicruris Greville 1859* bl *Vidal y Carbonell (1977), *Carbonell (1979)
Odontella mobiliensis (Bailey) Grunow 1884
= Biddulphia mobiliensis (Bailey) Grunow in van Heurck 1882*
bl ma *Caycedo (1975, 1977), *Vidal y Carbonell (1977), *Carbonell (1979), Gavilán et al. (2005), Ramos (2005), Suárez (2007)
Odontella rhombus (Ehrenberg) Kützing 1849
ma Ramos (2005)
Odontella regia (Schultze) Simonsen 1974
= Biddulphia regia (Schultze) Ostenfeld 1908*
ma bl *Caycedo (1975, 1977), *Vidal y Carbonell (1977), *Arias y Durán (1982, 1984), *Gavilán et al. (2005), Ramos (2005), Suárez (2007)
Odontella sinensis (Greville) Grunow 1884 bl Suárez (2007)
Género Pleurosira (Meneghini) Trevisan emend . Compére 1982
Pleurosira laevis (Ehrenberg) Compère 1982
= Biddulphia laevis Ehrenberg 1843* bl ma *Vidal y Carbonell (1977), Ramos (2005)
Género Triceratium Ehrenberg 1839
Triceratium antediluvianum (Ehrenberg) Grunow 1870
bl Vidal y Carbonell (1977)
Triceratium dubium Brightwell 1859 bl Vidal y Carbonell (1977), Carbonell (1979)
Triceratium formosum f . formosum Brightwell 1856
bl Carbonell (1980)
Triceratium formosum f . quadrangularis (Greville) Hustedt 1930
bl Carbonell (1980)
Triceratium formosum f . quinquelobulata (Greville) Hustedt 1930
bl Carbonell (1980)
Triceratium gibbosum Harvey et Bailey 1854 bl Carbonell (1979)
Triceratium pelagicum (Schroder) Sournia 1968
bl Vidal y Carbonell (1977), Carbonell (1979, 1980)
Triceratium pentacrinus (Ehrenberg) Wallich 1858
bl Vidal y Carbonell (1977), Carbonell (1979), Suárez (2007)
Triceratium robertsianum Greville 1863 ma Fernández et al . (1999)
Triceratium shadboltianum f . elongata (Grunow) Hustedt 1930
bl Carbonell (1979, 1980)
Familia Plagiogrammaceae De Toni 1890
Género Dimeregramma Ralfs in Pritchard 1861
Dimeregramma sp . bl Vidal y Carbonell (1977)
Continuación Tabla 2.

Instituto de Investigaciones Marinas y Costeras - INVEMAR 93
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
Género Plagiogramma Greville 1859
Plagiogramma obesum Greville 1865 bl Vidal y Carbonell (1977)
Plagiogramma decussatum Greville 1866 ma Ramos (2005)
Orden Biddulphiales Krieger 1954Familia Biddulphiaceae Kützing 1844
Género Biddulphia Gray 1821
Biddulphia alternans (Bailey) van Heurck 1880-1885
= Odontella alternans Schutt 1899* bl *Suárez (2007)
Biddulphia azorica Pavillard 1931 bl Carbonell (1979, 1980)
Biddulphia biddulphiana (Smith) Boyer 1901
= Biddulphia pulchella Gray 1821* bl ma *Vidal y Carbonell (1977), *Carbonell (1979), Ramos (2005)
Biddulphia obtusa (Kützing) Ralfs in Pritchard 1861
= Odontella obtusa Kützing 1844* bl ma Vidal y Carbonell (1977), Arias y Durán (1982, 1984), *Fernández et al. (1999), *Suárez (2007)
Biddulphia pellucida Castracane, Forma Schroder 1908
bl Carbonell (1980)
Biddulphia schroederiana Schussnig 1915 bl Vidal y Carbonell (1977), Carbonell (1979, 1980)
Biddulphia tuomeyi (Bailey) Roper 1859 ma Fernández et al . (1999)
Género Biddulphiopsis von Stosch et Simonsen 1984
Biddulphiopsis membranacea (Cleve) von Stosch et Simonsen 1984= Biddulphia membranacea Cleve 1878* bl *Carbonell (1979, 1980)
Biddulphiopsis titiana (Grunow) von Stosch et Simonsen 1984
= Biddulphia titiana (Grunow) Hustedt 1930*
Como Biddulphia membranacea Cleve 1878**
bl **Vidal y Carbonell (1977), *Carbonell (1980)
Género Isthmia Agardh 1832
Isthmia enervis Ehrenberg 1838 bl ma Caycedo (1975, 1977), Carbonell (1979), Fernández et al . (1999), Ramos (2005)
Isthmia minima Harvey et Bailey 1854 bl Vidal y Carbonell (1977), Carbonell (1979)
Género Trigonium Cleve 1868
Trigonium diaphanum Mann 1925 bl Carbonell (1979)
Trigonium formosum (Brightwell) Cleve bl Carbonell (1979)
Trigonium gibbosum Harvey et Bailey 1854 bl Carbonell (1979)
Orden Hemiaulales Round et Crawford 1990
Continuación Tabla 2.

94 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
Familia Hemiaulaceae Heiberg 1863
Género Cerataulina Péragallo in Schütt 1896
Cerataulina pelagica (Cleve) Hendey 1937
= Cerataulina bergonii (H . Peragallo) H . Peragallo in Schütt 1896*
bl ma Arosemena et al . (1973), *Caycedo (1975, 1977), Vidal y Carbonell (1977), Carbonell (1979), Ramos (2005)
Género Climacodium Grunow 1868
Climacodium biconcavum Cleve 1897 ma Caycedo (1975, 1977), Ramos (2005)
Género Eucampia Ehrenberg 1839
Eucampia cornuta (Cleve) Grunow in van Heurck 1885
bl Vidal y Carbonell (1977), Carbonell (1979)
Eucampia zodiacus Ehrenberg 1840 ma Ramos (2005)
Género Hemiaulus Heiberg 1863
Hemiaulus hauckii Grunow in van Heurck 1882
bl lg ma sp Caycedo (1975, 1977), Vidal y Carbonell (1977), Carbonell (1979), Márquez y Herrera (1986), Duarte (1996), Gavilán et al .( 2005), Ramos (2005), Franco-Herrera y Torres-Sierra (2007)
Hemiaulus sinensis Greville 1865 bl lg ma sp Caycedo (1975, 1977), Vidal y Carbonell (1977), Carbonell (1979), Márquez y Herrera (1986), Arias y Durán (1982, 1984), Duarte (1996), Gavilán et al . (2005), Ramos (2005), Franco-Herrera y Torres-Sierra (2007)
Hemiaulus membranaceus Cleve 1873 bl lg ma sp Vidal y Carbonell (1977), Carbonell (1979), Márquez y Herrera (1986), Duarte (1996), Gavilán et al . (2005), Ramos (2005)
Familia Bellerocheaceae Crawford 1990
Género Bellerochea van Heurck 1885
Bellerochea malleus (Brightwell) van Heurck 1885
bl Vidal y Carbonell (1977), Suárez (2007)
Familia Streptothecaceae Crawford 1990
Género Helicotheca Ricard 1987
Helicotheca tamesis (Shrubsole) Ricard 1987
= Streptotheca tamesis Shrubsole 1890* bl ma *Vidal y Carbonell (1977), Ramos (2005)
Orden Anaulales Round et Crawford 1990Familia Anaulaceae (Schütt) Lemmermann 1899
Continuación Tabla 2.

Instituto de Investigaciones Marinas y Costeras - INVEMAR 95
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
Género Eunotogramma Weisse 1854
Eunotogramma marinum (Smith) Peragallo 1897-1908
bl Vidal y Carbonell (1977)
Subclase Lithodesmiophycidae Round et Crawford 1990Orden Lithodesmiales Round et Crawford 1990Familia Lithodesmiaceae H . et M . Peragallo 1897-1908 emend . Simonsen 1979
Género Ditylum Bailey 1861
Ditylum brightwellii (West) Grunow in van Heurck 1883
ma Caycedo (1975, 1977), Ramos (2005)
Ditylum sol (van Heurck) De Toni 1894 bl Vidal y Carbonell (1977), Carbonell (1979)
Género Lithodesmium Ehrenberg 1839
Lithodesmium undulatum Ehrenberg, 1839 bl ma Caycedo (1975, 1977), Vidal y Carbonell (1977), Carbonell (1979), Ramos (2005), Suárez (2007)
Subclase Corethrophycidae Round et Crawford 1990Orden Corethrales Round et Crawford 1990Familia Corethraceae Lebour 1930
Género Corethron Castracane 1886
Corethron criophilum Castracane 1886
= Corethron hystrix Hensen 1887* ma bl sp lg Caycedo (1975, 1977), Vidal y Carbonell (1977), Márquez y Herrera (1986), Duarte (1996), Ramos (2005), *Franco-Herrera y Torres-Sierra (2007)
Subclase Rhizosoleniophycidae Round et Crawford 1990Orden Rhizosoleniales Silva 1962 emend . Round et Crawford 1990Familia Rhizosoleniaceae De Toni 1890
Género Dactyliosolen Castracane 1886
Dactyliosolen antarcticus Castracane 1886 bl Vidal y Carbonell (1977), Carbonell (1979), Arias y Durán (1982, 1984)
Dactyliosolen fragilissimus (Bergon) Hasle 1996
= Rhizosolenia fragilissima Bergon 1903* bl lg ma sp *Arosemena et al . (1973), *Caycedo (1975, 1977), *Vidal y Carbonell (1977), *Carbonell (1979), *Márquez y Herrera (1986), *Duarte (1996), *Gavilán et al . (2005), Ramos (2005), *Franco-Herrera y Torres-Sierra (2007)
Continuación Tabla 2.

96 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
Género Guinardia Peragallo 1892
Guinardia cylindrus (Cleve) Hasle 1996 ma bl Ramos (2005), Suárez (2007)
Guinardia flaccida (Castracane) Peragallo 1892
bl ma sp Arosemena et al. (1973), Vidal y Carbonell (1977), Carbonell (1979), Márquez y Herrera (1986), Arias y Durán (1982, 1984), Gavilán et al. (2005), Ramos (2005)
Guinardia striata (Stolterfoth) Hasle 1996
= Rhizosolenia stolterfothii H . Péragallo 1888*
bl lg ma sp *Caycedo (1975, 1977), *Vidal y Carbonell (1977), *Carbonell (1979), *Márquez y Herrera (1986), *Duarte (1996), Ramos (2005)
Género Neocalyptrella Hernández-Becerril et Meave del Castillo 1997
Neocalyptrella robusta (Norman) Hernández-Becerril et Meave 1997
= Calyptrella robusta (Norman in Ralfs) Hernández-Becerril et Meave del Castillo 1996*
= Rhizosolenia robusta Norman in Pritchard 1861**
bl ma sp **Vidal y Carbonell (1977), **Carbonell (1979), **Arias y Durán (1982), **(1984), **Márquez y Herrera (1986), *Gavilán et al . (2005), *Ramos (2005), **Franco-Herrera y Torres-Sierra (2007), *Suárez (2007)
Género Proboscia Subdström 1986
Proboscia alata (Brightwell) Sundström 1986
= Rhizosolenia alata (Brightwell) Sundström 1986*
bl lg ma sp *Caycedo (1975, 1977), *Vidal y Carbonell (1977), *Carbonell (1979), *Márquez y Herrera (1986), *Arias y Durán (1982, 1984), *Duarte (1996), *Gavilán et al . (2005), *Franco-Herrera y Torres-Sierra (2007), Suárez (2007)
Proboscia alata f . gracillima (Cleve) Gran 1986
= Rhizosolenia alata var . gracillima (Cleve) Gran 1905*
bl ma *Vidal y Carbonell (1977), *Carbonell (1979), Ramos (2005)
Proboscia indica (Peragallo) Hernández-Becerril emend . Jordan et Ligowski 2004
Continuación Tabla 2.

Instituto de Investigaciones Marinas y Costeras - INVEMAR 97
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
= Rhizosolenia alata f . indica (Péragallo) Gran 1905*
= Proboscia alata f . indica (Peragallo) Ostenfeld**
bl ma *Vidal y Carbonell (1977), *Carbonell (1979), **Ramos (2005)
Género Pseudosolenia Subdström 1986
Pseudosolenia calcar-avis (Schultze) Sundström 1986
= Rhizosolenia calcar-avis Schultze 1858* bl lg ma sp *Vidal y Carbonell (1977), *Carbonell (1979), *Arias y Durán (1982, 1984), *Márquez y Herrera (1986), *Duarte (1996), Ramos (2005), *Franco-Herrera y Torres-Sierra (2007), Suárez (2007)
Género Rhizosolenia Ehrenberg emend . Brightwell 1858
Rhizosolenia acuminata (Peragallo) Gran 1905
bl ma Arosemena et al . (1973), Ramos (2005)
Rhizosolenia castracanei Peragallo 1888 Franco-Herrera y Torres-Sierra (2007)
Rhizosolenia clevei Ostenfeld 1902 ma Ramos (2005)
Rhizosolenia bergonii Peragallo 1892 ma Franco-Herrera y Torres-Sierra (2007)
Rhizosolenia formosa Peragallo 1888 ma Ramos (2005)
Rhizosolenia hebetata Bailey 1856 lg ma sp Duarte (1996), Márquez y Herrera (1986), Franco-Herrera y Torres-Sierra (2007)
Rhizosolenia hebetata f . semispina (Hensen) Gran 1905
bl Vidal y Carbonell (1977), Carbonel (1979)
Rhizosolenia hyalina Ostenfeld 1901 sp Márquez y Herrera (1986)
Rhizosolenia imbricata Brightwell 1858
= Rhizosolenia imbricata var . schrubsolei (Cleve) Schröder 1906*
= Rhizosolenia cf . shrubsolei Cleve 1881** bl ma *Caycedo (1975, 1977), **Vidal y Carbonell (1977), Carbonell (1979), Ramos (2005), Franco-Herrera y Torres-Sierra (2007), Suárez (2007)
Rhizosolenia setigera Brightwell 1858 bl lg ma sp Arosemena et al . (1973), Caycedo (1975, 1977), Vidal y Carbonell (1977), Carbonell (1979), Márquez y Herrera (1986), Duarte (1996), Cañón et al . (2005), Gavilán et al. (2005), Ramos (2005), Franco-Herrera y Torres-Sierra (2007), Suárez (2007)
Continuación Tabla 2.

98 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
Rhizosolenia styliformis Brightwell 1858 bl lg ma sp Arosemena et al . (1973), Márquez y Herrera (1986), Duarte (1996), Cañón et al . (2005), Gavilán et al. (2005), Ramos (2005)
Rhizosolenia styliformis var . longispina Hustedt 1914
bl Carbonell (1979)
Subclase Chaetocerotophycidae Round et Crawford 1990Orden Chaetocerotales Round et Crawford 1990Familia Chaetoceraceae Ralfs in Pritchard 1861
Género Bacteriastrum Shadbolt 1854
Bacteriastrum comosum Pavillard 1916 bl ma sp Vidal y Carbonell (1977), Arias y Durán (1982, 1984), Márquez y Herrera (1986), Cañón et al. (2005), Gavilán et al . (2005), Ramos (2005), Franco-Herrera y Torres-Sierra (2007)
Bacteriastrum delicatulum Cleve 1897
= Bacteriastrum furcatum Shadbolt 1854* bl ma sp Caycedo (1975, 1977), Vidal y Carbonell (1977), Carbonell (1979), Arias y Durán (1982, 1984), Márquez y Herrera (1986), Gavilán et al . (2005), Ramos (2005), Franco-Herrera y Torres-Sierra (2007), Suárez (2007), * Suárez (2007)
Bacteriastrum elegans Pavillard 1916 bl ma Arosemena et al . (1973), Ramos (2005)
Bacteriastrum elongatum Cleve 1897 ma sp Márquez y Herrera (1986), Ramos (2005)
Bacteriastrum hyalinum Lauder 1864 bl lg ma sp Caycedo (1975, 1977), Márquez y Herrera (1986), Duarte (1996), Gavilán et al.( 2005), Ramos (2005), Suárez (2007)
Género Chaetoceros Ehrenberg 1844
Chaetoceros affinis Lauder 1864 bl lg ma sp Vidal y Carbonell (1977), Carbonell (1979), Márquez y Herrera (1986), Duarte (1996), Gavilán et al . (2005), Ramos (2005), Franco-Herrera y Torres-Sierra (2007), Suárez (2007)
Chaetoceros atlanticus Cleve 1873 bl lg ma Duarte (1996), Gavilán et al . (2005), Ramos (2005)
Chaetoceros atlanticus var . neopolitana (Schröder) Hustedt 1930
bl Vidal y Carbonell (1977), Carbonell (1979)
Continuación Tabla 2.

Instituto de Investigaciones Marinas y Costeras - INVEMAR 99
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
Chaetoceros borealis Bailey 1854 bl ma Ramos (2005), Suárez (2007)
Chaetoceros brevis Schütt 1895 bl Vidal y Carbonell (1977), Carbonell (1979), Cañón et al . (2005), Gavilán et al. (2005), Ramos (2005)
Chaetoceros coarctatus Lauder 1864 bl ma sp Vidal y Carbonell (1977), Carbonell (1979), Márquez y Herrera (1986), Ramos (2005)
Chaetoceros compressus Lauder 1864 ma sp Márquez y Herrera (1986), Ramos (2005)
Chaetoceros constrictus Gran 1897 ma Franco-Herrera y Torres-Sierra (2007)
Chaetoceros curvisetus Cleve 1889 bl lg ma sp Vidal y Carbonell (1977), Carbonell (1979), Arias y Durán (1982, 1984), Márquez y Herrera (1986), Duarte (1996), Ramos (2005), Franco-Herrera y Torres-Sierra (2007), Suárez (2007)
Chaetoceros danicus Cleve 1889 bl ma Gavilán et al . (2005), Ramos (2005)
Chaetoceros decipiens Cleve 1873 bl Carbonell (1979), Suárez (2007)
Chaetoceros debilis Cleve 1894 bl Carbonell (1979)
Chaetoceros dichaeta Ehrenberg 1844 bl sp Carbonell (1979), Márquez y Herrera (1986)
Chaetoceros didymus Ehrenberg 1845 bl lg ma sp Caycedo (1975, 1977), Vidal y Carbonell (1977), Carbonell (1979), Márquez y Herrera (1986), Duarte (1996), Cañón et al. (2005), Gavilán et al. (2005), Ramos (2005), Franco-Herrera y Torres-Sierra (2007), Suárez (2007)
Chaetoceros didymus var . anglicum (Grunow) Gran 1905
bl ma Caycedo (1975, 1977), Vidal y Carbonell (1977)
Chaetoceros diversum Cleve 1873
= Chaetoceros laevis Leuduger-Fortmorel 1892*
bl ma lg sp Caycedo (1975, 1977), Vidal y Carbonell (1977), Carbonell (1979), Arias y Durán (1982, 1984), Márquez y Herrera (1987), *Duarte (1996), Ramos (2005), Franco-Herrera y Torres-Sierra (2007), *Franco-Herrera y Torres-Sierra (2007), Suárez (2007), *Suárez (2007)
Chaetoceros eibenii Grunow in van Heurck 1882
bl Suárez (2007)
Chaetoceros distans Cleve 1894
Continuación Tabla 2.

100 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
= Chaetoceros laciniosus Schütt 1895* bl ma sp *Caycedo (1975, 1977), *Márquez y Herrera (1986), *Gavilán et al. (2005), *Ramos (2005), *Franco-Herrera y Torres-Sierra (2007)
Chaetoceros lauderi Ralfs in Lauder 1864 ma Franco-Herrera y Torres-Sierra (2007)
Chaetoceros lorenzianus Grunow 1863 bl lg ma sp Vidal y Carbonell (1977), Carbonell (1979), Márquez y Herrera (1986), Duarte (1996), Ramos (2005), Franco-Herrera y Torres-Sierra (2007), Suárez (2007)
Chaetoceros messanensis Castracane 1875 ma Ramos (2005), Franco-Herrera y Torres-Sierra (2007)
Chaetoceros neogracilis van Landingham 1968
= Chaetoceros gracilis Schütt 1895* bl sp *Márquez y Herrera (1986), *Gavilán et al . (2005)
Chaetoceros pelagicus Cleve 1873 ma Franco-Herrera y Torres-Sierra (2007)
Chaetoceros pendulus Karsten 1905 bl Vidal y Carbonell (1977), Carbonell (1979)
Chaetoceros peruvianus Brightwell 1856 bl lg ma sp Caycedo (1975, 1977), Vidal y Carbonell (1977), Carbonell (1979), Márquez y Herrera (1986), Duarte (1996), Ramos (2005), Franco-Herrera y Torres-Sierra (2007), Suárez (2007)
Chaetoceros protuberans Lauder 1864
= Chaetoceros didymus var . protuberans (Lauder) Gran et Yendo 1914*
bl *Suárez (2007)
Chaetoceros pseudocurvisetus Mangin 1910 bl Gavilán et al . (2005)
Chaetoceros radicans Schütt 1895 lg ma sp Márquez y Herrera (1986), Duarte (1996), Suárez (2007)
Chaetoceros subtilis Cleve 1896 bl Cañón et al . (2005), Gavilán et al. (2005)
Chaetoceros socialis Lauder 1864 bl Cañón et al. (2005), Gavilán et al. (2005)
Chaetoceros tortissimus Gran 1900 ma Ramos (2005)
Chaetoceros willei Gran 1897 bl sp Vidal y Carbonell (1977), Carbonell (1979), Márquez y Herrera (1986)
Orden Leptocylindrales Round et Crawford 1990Familia Leptocylindraceae Lebour 1930
Género Leptocylindrus Cleve 1889
Continuación Tabla 2.

Instituto de Investigaciones Marinas y Costeras - INVEMAR 101
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
Leptocylindrus danicus Cleve 1889 bl lg ma sp Caycedo (1975, 1977), Vidal y Carbonell (1977), Carbonell (1979), Márquez y Herrera (1986), Duarte (1996), Ramos (2005), Suárez (2007)
Leptocylindrus mediterraneus (Peragallo) Hasle 1975
= Dactyliosolen mediterraneus Peragallo 1892*
ma *Caycedo (1975, 1977)
Clase Fragilariophyceae Round 1990Subclase Fragilariophycidae Round 1990Orden Fragilariales Silva 1962 emend . Round 1990Familia Fragilariaceae Greville 1833
Género Asterionella Hassall 1855
Asterionella formosa Hassall 1850 ma Ramos (2005)
Género Asterionellopsis Round in Round et al . 1990
Asterionellopsis glacialis (Castracane) Round in Round et al. 1990
= Asterionella japonica Cleve in Cleve et Möller 1878*
bl ma sp *Caycedo (1975, 1977), *Vidal y Carbonell (1977), Márquez y Herrera (1986), Gavilán et al . (2005), Ramos (2005), Franco-Herrera y Torres-Sierra (2007), Suárez (2007)
Género Bleakeleya Round in Round et al. 1990
Bleakeleya notata (Grunow) Round in Round et al . 1990
= Asterionella notata Grunow in van Heurck 1885*
bl ma *Vidal (1981), Ramos (2005)
Género Fragilaria Lyngbye 1819
Fragilaria longissima Hustedt 1913 bl Ramos (2005)
Género Opephora Petit 1888
Opephora schwartzii (Grunow) Petit in Pelletan 1889
ma Fernández et al . (1999)
Género Podocystis Kützing 1844
Podocystis adriatica (Kützing) Ralfs in Pritchard 1861
bl Vidal y Carbonell (1977), Vidal (1981)
Género Synedra Ehrenberg 1830
Synedra goulardii Brehicon Cleve et Grunow 1887
ma Ramos (2005)
Continuación Tabla 2.

102 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
Synedra hantzschiana Sournia 1963 bl Vidal y Carbonell (1977)
Synedra ulna (Nitzsch) Ehrenberg 1832 bl ma Gavilán et al. (2005), Ramos (2005)
Synedra ulna var . amphirhynchus (Ehrenberg) Grunow 1862
bl ma Vidal y Carbonell (1977), Fernández et al (1999), Ramos (2005)
Synedra ulna var . contracta Øestrup bl Vidal y Carbonell (1977)
Synedra ulna var. oxyrhynchus f . mediocontracta (Forti) Hustedt 1932
bl Vidal y Carbonell (1977)
Orden Lichmophorales Round 1990Familia Lichmophoraceae Kützing 1844
Género Licmophora C . Agardh 1827
Licmophora ehrenbergii (Kützing) Grunow 1867
ma Ramos (2005)
Licmophora flabellata (Carmichael) Agardh 1831
bl Vidal y Carbonell (1977)
Lichmophora grandis (Kutzing) Grunow 1880
ma Ramos (2005)
Licmophora hyalina (Kützing) Grunow 1867 ma Ramos (2005)
Licmophora Lyngbyei (Kützing) Grunow in van Heurck 1885
bl Vidal y Carbonell (1977)
Licmophora paradoxa (Lyngbye) Agardh 1836
ma Ramos (2005)
Licmophora remulus Grunow 1867 ma Ramos (2005)
Género Licmosphenia Mereschkowsky 1902
Licmosphenia clevei Mereschkowsky 1902 bl Vidal (1981)
Licmosphenia grunowii Mereschkowsky 1902
bl Vidal (1981)
Orden Rhaphoneidales Round 1990Familia Rhaphoneidaceae Forti 1912
Género Delphineis Andrews 1977
Delphineis surirella (Ehrenberg) Andrews 1982
bl Vidal y Carbonell (1977)
Género Neodelphineis Takano 1982
Neodelphineis pelagica Takano 1982 ma Ramos (2005)
Orden Ardissoneales Round 1990Familia Ardissoneaceae Round 1990
Género Ardissonea de Notaris 1842
Ardissonea crystallina (Agardh) Grunow in Cleve et Grunow 1880
= Synedra crystallina (Ag.) Kützing 1844* bl *Vidal y Carbonell (1977)
Orden Toxariales Round 1990Familia Toxariaceae Round 1990
Género Toxarium Bailey 1854
Toxarium hennedyanum (Gregory) Pelletan 1889
Continuación Tabla 2.

Instituto de Investigaciones Marinas y Costeras - INVEMAR 103
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
= Synedra hennedyana Gregory 1857* bl ma *Vidal y Carbonell (1977), Fernández et al . (1999), Ramos (2005)
Toxarium undulatum Bailey 1854
= Synedra undulata (Bailey) Gregory 1857* bl ma *Vidal y Carbonell (1977), Fernández et al. (1999), *Ramos (2005), Suárez (2007)
Orden Thalassionematales Round 1990Familia Thalassionemataceae Round 1990
Género Lioloma Hasle in Hasle et Syvertsen 1996
Lioloma elongatum (Grunow) Hasle 1996 ma Ramos (2005)
Lioloma pacificum (Cupp) Hasle 1996 ma Ramos (2005)
Género Thalassionema Grunow ex Hustedt 1932
Thalassionema capitulata (Castracane) Hustedt 1958
ma Ramos (2005)
Thalassionema elegans Hustedt 1958 ma Ramos (2005)
Thalassionema frauenfeldii (Grunow) Hallegraeff 1986
= Thalassiothrix frauenfeldii Grunow 1810* bl ma sp *Caycedo (1975, 1977), *Vidal y Carbonell (1977), Arias y Durán (1982, 1984), Márquez y Herrera (1986), Ramos (2005), *Suárez (2007)
Thalassionema nitzschioides Grunow in Hustedt 1932
bl lg ma sp Arosemena et al (1983) Arias y Durán (1982, 1984)
Género Thalassiotrix Cleve et Grunow 1880
Thalassiotrix heteromorpha var . mediterránea (Pavillard) Hallegraeff 1986
= Thalassiothrix mediterranea Pavillard 1916*
= Asterionella mediterránea** bl ma *Arosemena et al . (1973), **Caycedo (1975, 1977), *Vidal y Carbonell (1977), *Arias y Durán (1982, 1984), Gavilán et al . (2005)
Thalassiothrix longissima Cleve et Grunow 1880
ma Franco-Herrera y Torres-Sierra (2007)
Orden Rhabdonematales Round et Crawford 1990Familia Rhabdonemataceae Round et Crawford 1990
Género Rhabdonema Kützing 1844
Continuación Tabla 2.

104 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
Rhabdonema adriaticum Kützing 1811 bl ma Caycedo (1975, 1977), Vidal y Carbonell (1977), Vidal (1981), Fernández et al. (1999), Ramos (2005)
Orden Striatellales Round 1990Familia Striatellaceae Kützing 1844
Género Grammatophora Ehrenberg 1840
Grammatophora marina (Lyngbye) Kützing 1844
bl ma Caycedo (1975, 1977), Vidal y Carbonell (1977), Franco-Herrera y Torres-Sierra (2007), Suárez (2007)
Género Microtabella Round 1990
Microtabella interrupta (Ehrenberg) Round in Round et al . 1990
= Striatella interrupta (Ehrenberg) Heiberg* bl *Vidal (1981)
Hyalosira interrupta (Ehrenberg) Navarro in Navarro et Williams, 1991
= Striatella interrupta (Ehr .) Heiberg* bl *Vidal (1981)
Género Striatella Agardh 1832
Striatella unipunctata (Lyngbye) Agardh 1830
bl ma Vidal y Carbonell (1977), Ramos (2005), Suárez (2007)
Orden Climacospheniales Round 1990Familia Climacospheniaceae Round 1990
Género Climacosphenia Ehrenberg 1841
Climacosphenia moniligera Ehrenberg 1843 bl ma Vidal y Carbonell (1977), Fernández et al (1999), Ramos (2005), Franco-Herrera y Torres-Sierra (2007), Suárez (2007)
Clase Bacillariophyceae Haeckel 1878 emend . D .G . MannSubclase Eunotiophycidae D .G . Mann 1990Orden Eunotiales Silva 1962Familia Eunotiaceae Kützing 1844
Género Eunotia Ehrenberg 1837
Eunotia sp . bl Vidal y Carbonell (1977), Cañón et al. (2005), Gavilán et al . (2005)
Subclase Bacillariophycidae D .G . MannOrden Lyrellales D .G . Mann 1990Familia Lyrellaceae D .G . Mann 1990Género Lyrella Karayeva 1978
Lyrella clavata (Gregory) Mann 1990 ma Ramos (2005)
Lyrella genifera (A . Schmidt) Mann in Round et al . 1990
Continuación Tabla 2.

Instituto de Investigaciones Marinas y Costeras - INVEMAR 105
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
Como Navicula hennedyi Smith 1856* bl ma *Vidal y Carbonell (1977), Fernández et al . (1999)
Lyrella lyra (Ehrenberg) Karayeva 1978
= Navicula lyra Ehrenberg 1843* bl *Vidal y Carbonell (1977), *Vidal (1981)
Género Petroneis Stickle et Mann in Round et al. 1990
Petroneis granulata (Bailey) Mann in Round et al. 1990
= Navicula granulata Bailey 1854* bl *Vidal y Carbonell (1977)
Petroneis marina (Ralfs in Pritchard) Mann in Round et al . 1990
= Navicula marina Ralfs in Pritchard 1861* bl *Vidal y Carbonell (1977)
Petroneis plagiostoma (Grunow in Cleve et Möller) Mann in Round et al. 1990
= Navicula plagiostoma Grunow 1879* bl *Vidal (1981)
Orden Mastogloiales D .G . Mann 1990Familia Mastogloiaceae Mereschkowsky 1903
Género Mastogloia Thwaites in . Smith 1856
Mastogloia angulata Lewis 1861 bl Vidal y Carbonell (1977), Vidal (1981)
Mastogloia apiculata Smith 1856 bl Vidal (1981)
Mastogloia baldjickiana Grunow 1893
Como Mastogloia sp . B* *Vidal (1981)
Mastogloia cribrosa Grunow 1860 bl Vidal (1981)
Mastogloia cruciata Cleve 1891
Como Mastogloia sp . C* *Vidal (1981)
Mastogloia erythraea Grunow 1860 bl Vidal y Carbonell (1977)
Mastogloia fimbriata (Brightwell) Cleve 1895
ma Fernández et al. (1999)
Mastogloia mauritiana Brun 1893
Como Mastogloia sp . A* *Vidal (1981)
Mastogloia punctatissima (Greville) Ricard 1975
bl Vidal (1981)
Mastogloia pusilla var . linearis Østrup 1910 ma Fernández et al . (1999)
Mastogloia seychellensis Grunow 1878
Como Mastogloia sp . D* ma *Vidal (1981), Ramos (2005)
Mastogloia smithii Thwaites in Smith 1856 bl Vidal y Carbonell (1977)
Mastogloia smithii var . amphicephala Grunow 1880
bl Vidal y Carbonell (1977)
Mastogloia tropica Hustedt 1927 ma Ramos (2005)
Orden Achnanthales Silva 1962
Continuación Tabla 2.

106 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
Familia Achnanthaceae Kützing 1844 emend. D .G . Mann 1990
Género Achnanthes Bory 1822
Achnanthes brevipes Agardh 1824 bl Vidal y Carbonell (1977)
Achnanthes controversa Hustedt 1937 ma Ramos (2005)
Achnanthes manifera Brun 1895 bl ma Vidal y Carbonell (1977), Fernández et al. (1999), Ramos (2005)
Familia Cocconeidaceae Kützing 1844
Género Cocconeis Ehrenberg 1838
Cocconeis placentula Ehrenberg 1838 ma Ramos (2005)
Cocconeis scutellum Ehrenberg 1833 bl Vidal y Carbonell (1977)
Orden Naviculales Bessey 1907 emend . D .G . Mann 1990Suborden Neidiineae D .G . Mann 1990Familia Berkeleyaceae D .G . Mann 1990
Género Berkeleya Greville emend . Cox 1975
Berkeleya scopulorum (Brébisson in Kützing) Cox 1979
= Navicula scopulorum Brébisson in Kützing 1849
ma *Ramos (2005)
Género Climaconeis Grunow emend . Cox 1982
Climaconeis lorenzii Grunow 1862 ma Ramos (2005)
Suborden Sellaphorineae D .G . Mann 1990Familia Sellaphoraceae Mereschkowsky 1902
Género Fallacia Stickle et D .G . Mann in Round et al. 1990
Fallacia spp . ma Fernández et al . (1999)Familia Pinnulariaceae D .G . Mann 1990
Género Caloneis Cleve 1894
Caloneis liber (Smith) Cleve 1894 ma Fernández et al . (1999)
Género Öestrupia Heiden 1906
Öestrupia powellii (Lewis) Heiden ex Hustedt
bl Vidal y Carbonell (1977)
Género Pinnularia Ehrenberg 1843
Pinnularia nobilis Ehrenberg 1840 bl Vidal y Carbonell (1977)
Pinnularia gentilis (Donkin) Cleve 1891 bl Vidal y Carbonell (1977)
Suborden Diploneidineae D .G . Mann 1990Familia Diploneidaceae D .G . Mann 1990
Género Diploneis Ehrenberg ex Cleve 1894
Diploneis bombus Ehrenberg 1844
Continuación Tabla 2.

Instituto de Investigaciones Marinas y Costeras - INVEMAR 107
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
Como Diploneis bombus (Ehrenberg) Cleve 1894*
Como Diploneis bombus (Ehr .) Ehrenberg ex Cleve**
bl ma *Vidal y Carbonell (1977), **Ramos (2005)
Diploneis chersonensis (Grunow) Cleve 1894 bl Vidal (1981)
Diploneis crabro Ehrenberg 1844 bl ma Vidal y Carbonell (1977), Vidal (1981), Fernández et al . (1999), Ramos (2005), Franco-Herrera y Torres-Sierra (2007)
Diploneis fusca (Gregory) Cleve 1894 ma Ramos (2005)
Diploneis incurvata (Gregory) Cleve 1894
Como Diploneis crabro Ehrenberg 1844 partim*
bl *Vidal y Carbonell (1977)
Diploneis notabilis (Greville) Cleve 1894 ma Franco-Herrera y Torres-Sierra (2007)
Diploneis smithii (Brébisson) Cleve 1894 bl Vidal y Carbonell (1977)
Diploneis subadvena Hustedt, 1937 ma Ramos (2005)
Diploneis suborbicularis (Gregory) Cleve 1894
Como Diploneis sp .* bl *Vidal y Carbonell (1977)
Suborden Naviculineae Hendey 1937Familia Naviculaceae Kützing 1844 emend. D .G . Mann 1990
Género Haslea Simonsen 1974
Haslea wawrikae (Hustedt) Simonsen 1974
= Navicula wawrikae Hustedt 1961* bl ma *Arias y Durán (1982, 1984), Ramos (2005)
Género Navicula Bory 1822
Navicula abunda Hustedt 1955 ma Fernández et al . (1999)
Navicula cancellata Donkin 1870 bl lg sp Arias y Durán (1982, 1984), Márquez y Herrera (1986), Duarte (1996)
Navicula directa (Smith) Ralfs in Pritchard 1861
bl sp Vidal y Carbonell (1977), Márquez y Herrera (1986)
Navicula longa (Gregory) Ralfs 1861 ma Fernández et al . (1999)
Navicula radiosa Kützing 1844 ma Ramos (2005)
Navicula tersa Hustedt 1956 ma Ramos (2005)
Navicula uniseriata Østrup 1913 ma Fernández et al. (1999)
Navicula yarrensis var . americana Cleve 1895
Como Navicula cancellata Donkin 1870* bl *Vidal y Carbonell (1977)
Género Trachyneis Cleve 1894
Continuación Tabla 2.

108 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
Trachyneis aspera (Ehrenberg) Cleve 1894 bl ma Vidal y Carbonell (1977), Fernández et al (1999), Ramos (2005), Franco-Herrera y Torres-Sierra (2007)
Familia Pleurosigmataceae Mereschkowsky 1903
Género Gyrosigma Hassall 1845
Gyrosigma balticum (Ehrenberg) Rabenhorst 1894
bl ma Vidal y Carbonell (1977), Arias y Durán (1982, 1984), Fernández et al . (1999), Ramos (2005), Suárez (2007)
Gyrosigma macrum (Smith) Griffith et Henfrey 1856
Como Cylindrotheca fusiformis Reimann et Lewin var . fusiformis 1964*
bl *Vidal y Carbonell (1977)
Gyrosigma rectum (Donkin) Cleve 1894
= Donkinia recta (Donkin) Grunow ex van Heurck 1880*
bl *Vidal y Carbonell (1977)
Gyrosigma simile (Grunow) Boyer 1916 bl Vidal (1981)
Gyrosigma spathulatum Ricard 1975 ma Ramos (2005)
Gyrosigma tenuissimum (Smith) Griffith et Henfrey 1856
ma Fernández et al . (1999), Ramos (2005)
Género Pleurosigma Smith 1852
Pleurosigma decorum Smith 1853 ma Ramos (2005)
Pleurosigma delicatulum Smith 1853 ma Ramos (2005)
Pleurosigma elongatum Smith 1852 ma Franco-Herrera y Torres-Sierra (2007)
Pleurosigma formosum Smith 1852 bl ma Vidal (1981), Arias y Durán (1982, 1984), Fernández et al . (1999)
Pleurosigma hamuliferum Brun 1889 ma Ramos (2005), Franco-Herrera y Torres-Sierra (2007)
Pleurosigma nicobarium (Grunow) Grunow 1878
ma Franco-Herrera y Torres-Sierra (2007)
Pleurosigma normanii Ralfs 1861 ma Fernández et al. (1999)
Pleurosigma portoricense Hagelstein 1938 ma Fernández et al . (1999)
Pleurosigma rigidum Smith 1853 ma Fernández et al . (1999), Ramos (2005)
Pleurosigma rhombeum Grunow 1880 ma Ramos (2005)
Pleurosigma speciosum Smith 1852 ma Fernández et al . (1999)
Pleurosigma strigosum Smith 1852 ma Ramos (2005), Franco-Herrera y Torres-Sierra (2007)
Familia Plagiotropidaceae D .G . Mann 1990
Género Plagiotropis Pfitzer 1871
Plagiotropis lepidoptera (Gregory) Kuntze 1898
Continuación Tabla 2.

Instituto de Investigaciones Marinas y Costeras - INVEMAR 109
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
= Tropidoneis lepidoptera (Gregory) Cleve 1894*
bl ma *Vidal y Carbonell (1977), Ramos (2005), *Franco-Herrera y Torres-Sierra (2007), Suárez (2007)
Plagiotropis longa (Cleve) Kuntze 1898 ma Fernández et al . (1999)
Plagiotropis maxima (Gregory) Kuntze 1898 ma Fernández et al. (1999)
Plagiotropis seriata (Cleve) Kuntze 1898 ma Fernández et al. (1999)Familia Stauroneidaceae D .G . Mann 1990
Género Meuniera P .C . Silva
Meuniera membranacea (Cleve) P .C . Silva
= Stauroneis membranacea (Cleve) Hustedt 1931*
bl ma *Vidal y Carbonell (1977), *Ramos (2005)
Orden Thalassiophysales D .G . Mann 1990Familia Catenulaceae Mereschkowsky 1902
Género Amphora Ehrenberg 1840
Amphora clevei Grunow in Schmit 1875 ma Ramos (2005)
Amphora coffeaeformis (Agardh) Kützing 1844
bl ma Vidal y Carbonell (1977), Ramos (2005)
Amphora crassa Gregory 1857 ma Ramos (2005)
Amphora decussata Grunow 1857 bl Vidal (1981), Fernández et al. (1999), Ramos (2005)
Amphora exigua Gregory 1857 bl Vidal y Carbonell (1977)
Amphora lineolata Ehrenberg 1838 bl Vidal y Carbonell (1977)
Amphora marina van Heurck 1896 bl Vidal y Carbonell (1977)
Amphora obtusa Gregory 1857 bl ma Vidal y Carbonell (1977), Vidal (1981), Fernández et al . (1999), Ramos (2005)
Amphora ostrearia de Brebisson in Kützing var . ostrearia Hendey 1964
bl Vidal y Carbonell (1977), Vidal (1981)
Amphora ostrearia var . lineata Cleve 1895 bl Vidal y Carbonell (1977)
Amphora ostrearia var . vitrea Cleve 1895 bl Vidal y Carbonell (1977)
Amphora rhombica Kitton 1876 ma Fernández et al . (1999)
Amphora terroris Ehrenberg 1853 bl Vidal (1981)
Amphora ventricosa Gregory 1857 bl ma Vidal y Carbonell (1977), Ramos (2005)
Familia Thalassiophysaceae D .G . Mann 1990
Género Thalassiophysa Conger 1954
Thalassiophysa hyalina (Greville) Paddock et Sims 1981
ma Ramos (2005)
Orden Bacillariales Hendey 1937 emend . Round et al . 1990Familia Bacillariaceae Ehrenberg 1831
Género Bacillaria Gmelin 1788
Bacillaria paxillifer (Müller) Hendey 1951
Continuación Tabla 2.

110 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
= Bacillaria paradoxa Gmelin in L . 1788* bl ma Vidal y Carbonell (1977), Vidal (1981), Arias y Durán (1982, *1984), Ramos (2005), Suárez (2007)
Género Cylindrotheca Rabenhorst emend . Reimann et Lewin 1964
Cylindrotheca closterium (Ehrenberg) Reiman et Lewin 1964
= Nitzschia closterium (Ehrenberg) Smith 1853*
bl lg ma sp *Caycedo (1975, 1977), *Vidal y Carbonell (1977), Márquez y Herrera (1986), Duarte (1996), Gavilán et al . (2005), Ramos (2005), *Franco-Herrera y Torres-Sierra (2007), Suárez (2007)
Género Fragilariopsis Hustedt in Schmidt 1913 emend . Hasle 1993
Fragilariopsis doliolus (Wallich) Medlin et Sims 1993
ma Ramos (2005)
Género Hantzschia Grunow 1877
Hantzschia amphioxys (Ehrenberg) Grunow 1880
ma Ramos (2005)
Genero Nitzschia Hassall 1845
Nitzschia bicapitata Cleve1900 sp Márquez y Herrera (1986)
Nitzschia braarudii Hasle 1960 ma Ramos (2005)
Nitzschia distans var . tumescens Grunow in Cleve et Grunow 1880
ma Ramos (2005)
Nitzschia fluminensis Grunow 1862 ma Fernández et al. (1999)
Nitzchia habirshawii (Grunow) Boyer
Como Nitzchia habirshawii Febiger in Cleve et Möller 1877*
bl *Vidal y Carbonell (1977), *Arias y Durán (1982, 1984)
Nitzschia interruptestriata (Heiden) Simonsen 1974
ma Ramos (2005)
Nitzschia longissima (Bréb .) Grunow
Como Nitzschia longissima (Brébisson in Kützing) Ralfs in Pritchard 1861*
bl ma sp *Caycedo (1975, 1977), *Vidal y Carbonell (1977), *Arias y Durán (1982, 1984), Márquez y Herrera (1986), Fernández et al . (1999), *Ramos (2005), Franco-Herrera y Torres-Sierra (2007), Suárez (2007)
Nitzschia longissima var . chinensis Grunow in Cleve et Grunow 1880
ma Ramos (2005)
Nitzschia martiana (Agardh) van Heurck 1896
ma Fernández et al . (1999)
Nitzschia panduriformis var . minor Grunow in Cleve et Grunow 1880
ma Ramos (2005)
Continuación Tabla 2.

Instituto de Investigaciones Marinas y Costeras - INVEMAR 111
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
Nitzschia pulcherrima (Grunow) Grunow in Cleve et Möller 1878
ma Fernández et al. (1999)
Nitzschia scalaris (Ehrenberg) Smith 1853
Como Nitzschia sigmoidea (Nitzsch) Smith* bl *Vidal y Carbonell (1977)
Nitzschia scalpelliformis (Grunow) Grunow in Cleve et Grunow 1880
= Nitzschia obtusa var . scalpelliformis Grunow in Cleve et Möller*
ma *Ramos (2005)
Nitzschia sicula var . sicula (Castracane) Hustedt 1958
ma Ramos (2005)
Nitzschia sigma (Kützing) Smith 1853 bl ma sp Vidal y Carbonell (1977), Márquez y Herrera (1986), Fernández et al. (1999), Franco-Herrera y Torres-Sierra (2007), Suárez (2007)
Nitzschia sigmoidea (Nitzsch) Smith 1853
Como Nitzschia sp .* bl ma *Vidal y Carbonell (1977), Ramos (2005)
Nitzschia socialis Gregory 1857 ma Fernández et al . (1999)
Nitzschia ventricosa Kitton 1869
= Nitzchia longissima f . costata Schmidt 1873*
ma *Vidal y Carbonell (1977), Fernández et al. (1999), *Gavilán et al .( 2005), Ramos (2005)
Género Psammodictyon D .G . Mann 1990
Psammodictyon constrictum (Gregory) D .G . Mann in Round et al . 1990
= Nitzschia constricta (Gregory) Grunow in Cleve et Grunow 1880*
bl sp *Vidal (1981), *Márquez y Herrera (1986), *Gavilán et al. (2005)
Psammodictyon panduriforme (Gregory) D .G . Mann in Round et al . 1990
= Nitzschia panduriformis Gregory 1857* ma *Fernández et al . (1999), *Ramos (2005)
Género Pseudo-nitzschia Peragallo in H . et M . Peragallo 1900
Pseudo-nitzschia delicatissima (Cleve) Heiden in Heiden et Kolbe 1928
= Nitzschia delicatissima Cleve 1897* ma sp *Márquez y Herrera (1986), Ramos (2005)
Pseudo-nitzschia pungens (Grunow) Hasle 1993
= Nitzschia pungens Grunow in Cleve 1897* bl ma sp *Caycedo (1975, 1977), *Vidal y Carbonell (1977), Arias y Durán (1982, 1984), Márquez y Herrera (1986), Ramos (2005), *Franco-Herrera y Torres-Sierra (2007), Suárez (2007)
Continuación Tabla 2.

112 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
Género Tryblionella Smith 1853
Tryblionella acuminata Smith 1853
= Nitzschia acuminata (Smith) Grunow 1878*
ma *Ramos (2005)
Tryblionella coarctata (Grunow in Cleve et Grunow) D .G . Mann 1990
Como Nitzschia aërophila Hustedt 1942* bl *Vidal (1981)
Tryblionella levidensis Smith 1856
= Nitzschia levidensis (Smith) Grunow in van Heurck 1881*
ma *Ramos (2005)
Tryblionella littoralis (Grunow in Cleve et Grunow) D .G . Mann in Round et al . 1990
Como Nitzchia aërophila Hustedt 1942* bl *Vidal y Carbonell (1977), *Vidal (1981)
Tryblionella plana (Smith) Pelletan 1889
= Nitzschia plana Smith 1853* bl *Vidal (1981)
Orden Surirellales D .G . Mann 1990Familia Entomoneidaceae Reimer in Patrick et Reimer 1975
Género Entomoneis Ehrenberg 1845
Entomoneis alata Ehrenberg 1845
= Amphiprora alata (Ehrenberg) Kützing 1844*
bl ma *Vidal y Carbonell (1977), Ramos (2005), *Franco-Herrera y Torres-Sierra (2007)
Familia Auriculaceae Hendey 1964
Género Auricula Castracane emend . Paddock et Sims 1980
Auricula complexa (Gregory) Cleve 1894 ma Fernández et al . (1999)Familia Surirellaceae Kützing 1844
Género Campylodiscus Ehrenberg 1840
Campylodiscus bicostatus Smith 1854 bl Vidal y Carbonell (1977)
Campylodiscus fastuosus Ehrenberg 1845 bl Vidal y Carbonell (1977)
Campylodiscus imperialis Greville 1860 ma Fernández et al. (1999)
Campylodiscus incertus Schmidt 1875 ma Fernández et al . (1999)
Campylodiscus latus var . pertinax A . Schmidt 1893
Como Campylodiscus biangulatus Greville 1862*
bl *Vidal y Carbonell (1977)
Campylodiscus limbatus Brébisson 1854 ma Fernández et al. (1999), Ramos (2005)
Campylodiscus undulatus Greville 1863 bl Vidal (1981)
Género Petrodictyon D .G . Mann in Round et al. 1990
Petrodictyon gemma (Ehrenberg) D .G . Mann in Round et al. 1990
Continuación Tabla 2.

Instituto de Investigaciones Marinas y Costeras - INVEMAR 113
Taxón/Taxon
Distribución en Colombia/
Distribution in Colombia
Referencias/References
Como Surirella gemma (Ehrenberg) Kützing 1844*
bl ma *Vidal y Carbonell (1977), *Ramos (2005)
Género Surirella Turpin 1828
Surirella fastuosa (Ehrenberg) Kützing 1844 bl ma Vidal y Carbonell (1977), Vidal (1981), Fernández et al. (1999), Ramos (2005), Suárez (2007)
Surirella fastuosa var . lata Ehrenberg 1854 bl Vidal y Carbonell (1977)
Surirella inducta Schmidt 1875
Como Surirella robusta Ehrenberg 1841* bl *Vidal y Carbonell (1977)
Surirella kattrayi Schmidt 1875 ma Ramos (2005)
Surirella linearis Smith 1853 ma Ramos (2005)
Surirella smithii Ralfs in Pritchard 1861
Como Surirella biseriata Brebisson 1838* bl ma *Vidal y Carbonell (1977), Ramos (2005)
DISCUSIÓN
Muchos de los trabajos desarrollados en el mar Caribe colombiano se han
realizado en zonas costeras, especialmente en las bahías de Cartagena, Nenguange y
Santa Marta y en el golfo de Salamanca . En zonas insulares se encuentran trabajos en
el archipiélago de San Andrés y Providencia y en el Parque Nacional Natural Corales
del Rosario . En la zona noroeste de La Guajira se destaca el trabajo realizado durante
el crucero Caribe 1-93 .
Existen muchas áreas en el Caribe colombiano que aún no han sido
estudiadas y la información obtenida en este trabajo se basa en estudios realizados en
lugares muy puntuales . Los trabajos hasta el momento desarrollados y publicados en
el mar Caribe colombiano son muy pocos . La mayoría de la información se encuentra
en informes institucionales o en trabajos de grado de las universidades de la región .
Este listado de especies es un inicio ya que aún faltan muchas áreas por
estudiar, las cuales podrían aumentar este elenco con nuevos registros y nuevas
especies para el área . Además, debido al constante cambio de la sistemática, y a
la descripción de nuevos taxones es difícil tener un listado definitivo . Esta clase de
listados exige que sea constantemente actualizado . También es importante resaltar
que la información que no ha sido divulgada y que no es de fácil acceso se logre
publicar .
En comparación con el trabajo de Navarro y Hernández-Becerril (1997),
el cual presenta 66 familias, 150 géneros, y alrededor de 1067 taxones, el presente
Continuación Tabla 2.

114 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
listado cuenta con un menor número en cuanto a familias, géneros y taxones; es decir
que existen muchas especies que pueden estar presentes en el área pero que no han
sido registradas por la falta de estudios en el mar Caribe colombiano . Se debe tener
en cuenta que el trabajo de Navarro y Hernández-Becerril (1997) también se basa en
la clasificación propuesta por Round et al . (1990) .
Vale la pena anotar que en la región no se encuentran colecciones de
referencia . Es importante la creación de una colección de referencia con el montaje
de placas permanentes y/o de registros fotográficos de buena calidad de las especies
fitoplanctónicas del mar Caribe colombiano, lo que daría más validez a los registros .
AGRADECIMIENTOS
Al Instituto de Investigaciones Marinas y Costeras-INVEMAR y a la
Agencia Nacional de Hidrocarburos-ANH por el apoyo y financiación del proyecto
“Especies, ensamblajes y paisajes de los bloques marinos sujetos a exploración de
hidrocarburos” dentro del cual se realizó el presente trabajo.
BIBLIOGRAFÍA
Arias, F . A . y J . C . Durán . 1982 . Variación anual del fitoplancton en la bahía de Cartagena para 1980 . Tesis
Biol . Mar ., Univ . Jorge Tadeo Lozano, Bogotá . 80 p .
Arias, F . A . y J . C . Durán . 1984 . Variación anual del fitoplancton en la bahía de Cartagena . Bol . Cient .
CIOH, 5: 61-116 .
Arosemena, D ., H . Cárdenas, F . Garzón, F . Ibáñez, C . Moreno y J . Sierra . 1973 . Algunos dinoflagelados
y diatomeas de la bahía de Cartagena y sus alrededores . Inf . Mus . Mar, 8, 11 p .
Cañón, M . L ., T . Vanegas, M . Gavilán, L . F . Morris y G . Tous . 2005 . Dinámica planctónica, microbiológica
y fisicoquímica en cuatro muelles de la bahía de Cartagena y buques de tráfico internacional . Bol .
Cient . CIOH, 23: 46-59 .
Carbonell, M . C . 1979 . Fitoplancton de las islas del Rosario y alrededores, Colombia (mar Caribe) .
Diatomeas centrales y algunos dinoflagelados . Informe, Centro de Investigaciones Oceanográficas
e Hidrográficas–CIOH, Cartagena. 82 p.
Carbonell, M . C . 1980 . Discusión taxonómica de algunas especies interesantes de los géneros Biddulphia
Gray y Triceratium Ehrenberg (Bacillariophyceae) . Bol . Inst . Ocean . Mem .V Simp . Latinoam .
Oceanogr . Biol ., 29 (2): 75-78 .
Caycedo, I . E . 1975 . Productividad primaria y fitoplancton en la bahía de Neguanje, Parque Nacional
Tayrona . Tesis Biol . Mar ., Univ . Jorge Tadeo Lozano, Bogotá . 49 p .
Caycedo, I . E . 1977 . Fitoplancton de la bahía de Nenguange (Parque Nacional Tayrona), mar Caribe,
Colombia . An . Inst . Invest . Mar . Punta Betín, 9: 17-44 .
Duarte, G . 1996 . Estructura del fitoplancton al noroeste de La Guajira durante el crucero Caribe 1-93 .
Tesis Biol . Mar ., Univ . Jorge Tadeo Lozano, Bogotá . 98 p .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 115
Edlund, M . B . y E . F . Stoermer . 1997 . Review: Ecological, evolutionary, and systematic significance of
diatom life histories . J . Phycol ., 33: 897-918 .
Fernández, C . E ., L . A Vidal y C . B . García . 1999 . Guide for the identification of diatoms found in
the Gulf of Salamanca, Central Colombian Caribbean: Estudio ecológico-pesquero de los recursos
demersales del Golfo de Salamanca, Caribe colombiano, estimación de la variabilidad de los
componentes biológicos del sistema . Informe final, Instituto de Investigaciones Marinas y Costeras-
INVEMAR, Santa Marta . 245 p .
Franco-Herrera, A . y E . A . Torres-Sierra . 2007 . La comunidad fitoplanctónica en el evento de surgencia
frente al mar Caribe Centro Colombiano . Revista . Univ . Cien . Apl . y Amb ., 10 (1): 159-172 .
Gavilán, M ., M . L . Cañón y G . Tous . 2005 . Comunidad fitoplanctónica en la bahía de Cartagena y en
aguas de lastre de buques de tráfico internacional . Bol . Cient . CIOH, 23: 60-73 .
Hartley, B ., H . G . Barber, J . R . Carter y P . A Sims .1996 . An atlas of British diatoms . Biopress Limited,
Dorchester, Reino Unido . 601 p .
Hasle, G . R . y G . A . Fryxell . 1995 . Taxonomy of diatoms . 339-364 . En: Hallegraeff, G . M ., D . M .
Anderson y A . D . Cembella (Eds .) . Manual of harmful marine microalgae . Intergovernmental
Oceanographic Commission of UNESCO, París . 793 p .
Margalef, R . 1969 . El ecosistema pelágico del mar Caribe . Mem . Soc . Cienc . Nat . La Salle, 29 (32), 36 p .
Márquez, G . y M . Herrera . 1986 . Estudios en la biomasa del fitoplancton y su distribución geográfica
durante el crucero Océano área I: levantamiento Providencia en el Caribe colombiano . Informe
final, Anexo II, Univ . Nacional de Colombia, Bogotá . 153 p .
Navarro, J . N . y D . U . Hernández-Becerril . 1997 . Listados florísticos de México . XV . Check-list of marine
diatoms from the Caribbean Sea . Univ . Autónoma de México, México D .F . 48 p .
Ramos, L . M . 2005 . Caracterización de la comunidad fitoplanctónica de la bahía de Santa Marta (Caribe
colombiano) . Tesis Biol . Univ . del Magdalena, Santa Marta . 86 p .
Round, F . E ., R . M . Crawford y D . G . Mann . 1990 . The diatoms, biology and morphology of the genera .
Cambridge University Press, Cambridge . 747 p .
Schmidt, A . 1874-1959 . Atlas der diatomaceen-Kunde . Reprint 1984 . Koeltz Scientific Books, Leipzig,
Alemania Occidental . Heft 1-120, Tafem 1-460 .Snoeijs, P . 1993 . Intercalibration and distribution of diatom species in the Baltic Sea . Vol . 1 . The Baltic
marine biologists . Publ . No . 16 a . Opulus Press, Uppsala, Suecia .129 p .
Snoeijs, P . y N . Balashova . 1998 . Intercalibration and distribution of diatom species in the Baltic Sea . Vol .
5 . The Baltic marine biologists . Publ . No . 16 e . Opulus Press, Uppsala, Suecia . 144 p .
Snoeijs, P . y J . Kasperoviciene . 1996 . Intercalibration and distribution of diatom species in the Baltic Sea .
Vol . 4 . The Baltic marine biologists . Publ . No 16 d . Opulus Press, Uppsala, Suecia . 126 p .
Snoeijs, P . y M . Potapova . 1995 . Intercalibration and distribution of diatom species in the Baltic Sea . Vol .
3 . The Baltic marine biologists . Publ . No 16 c . Opulus Press, Uppsala, Suecia . 126 p .
Snoeijs, P . y S . Vilbaste . 1994 . Intercalibration and distribution of diatom species in the Baltic Sea . Vol . 2 .
The Baltic marine biologists . Publ . No 16 b . Opulus Press, Uppsala, Suecia . 125 p .
Sournia, A ., M . J . Chrdtiennot-Dinet y M . Ricard . 1991 . Marine phytoplankton: how many species in the

116 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
world ocean? J . Plankton Res ., 13 (5): 1093-1099 .
Stosch, S . A von y R . Simonsen 1984 . Biddulphiospsis, a new genus of the Biddulphiaceae . Bacillaria,
7: 9-36 .
Suárez, V . C . 2007 . Biodiversidad del fitoplancton en aguas de lastre de buques de tráfico internacional y
en los principales muelles cargueros de la bahía de Cartagena – Colombia. Tesis Biol. Mar., Univ.
Jorge Tadeo Lozano, Bogotá . 130 p .
Sunesen, I . y E . Sar . 2007 . Marine diatoms from Buenos Aires coastal waters (Argentina) . IV . Rhizosolenia
s . str ., Neocalyptrella, Pseudosolenia, Proboscia . Phycologia, 46 (6): 628-643 .
Tomas, C . 1996 . Identifying marine phytoplankton . Academic Press, San Diego, EE . UU . 598 p .
Vidal, L . A . 1981 . Diatomeas y dinoflagelados en las islas del Rosario (arrecife coralino) y alrededores .
Bol . Cient . CIOH, 3: 75-133 .
Vidal, L .A . y M . C . Carbonell . 1977 . Diatomeas y dinoflagelados en la bahía de Cartagena . Tesis Biol .
Mar ., Univ . Jorge Tadeo Lozano, Bogotá . 360 p .
FECHA DE RECEPCIÓN: 05/08/2008 FECHA DE ACEPTACIÓN: 06/04/2010

Instituto de Investigaciones Marinas y Costeras - INVEMAR 117
Bol . Invest . Mar . Cost . 39 (1) ISSN 0122-9761 Santa Marta, Colombia, 2010
COBERTURA DE LOS FONDOS Y PAISAJES BENTÓNICOS ASOCIADOS A FORMACIONES DIAPÍRICAS EN LOS
BANCOS DE SALMEDINA, PLATAFORMA CONTINENTAL DEL CARIBE COLOMBIANO
Juan Gabriel Domínguez1, Juan Carlos Gómez1, Constanza Ricaurte1, Gisela Mayo1, Jaime Orejarena1, Juan Manuel Díaz2 y Carlos Alberto Andrade1
1 Escuela Naval de Cadetes “Almirante Padilla”, Grupo de Investigación en Oceanología (GIO), Cartagena, Colombia. [email protected] Corporación para Investigaciones Biológicas, Unidad de Biodiversidad, Medellín, Colombia. [email protected]
RESUMEN
Se determinaron las unidades ecológicas submarinas alrededor de un volcán activo de lodo en los bancos de Salmedina, Caribe colombiano, y se cartografiaron mediante video-transectos submarinos georreferenciados, análisis de la batimetría e interpretación de imágenes satelitales . Se describen la distribución, la cobertura relativa y los organismos bentónicos sésiles más representativos de las diferentes unidades ecológicas . La zona somera (0-10 m) de los bancos presenta 60 % de cobertura algal y 10 % de coral vivo; en las zonas más profundas (<10 m) la cobertura de coral vivo varía entre 33 % en los cantiles semiverticales y 55 % en las cimas de los bancos menos elevados; las especies de coral dominantes fueron Siderastrea siderea y Montastraea annularis en todos los sectores . Los bancos de Salmedina reciben la influencia cada vez mayor de plumas turbias que probablemente han aumentado la sedimentación, y las comunidades biológicas se ven posiblemente afectadas por el empleo de explosivos para la pesca . Aunque la zona más somera del banco Salmedina es la que evidencia mayor afectación, las comunidades coralinas más profundas aún mantienen una cobertura viva significativa en comparación con otras zonas similares en la región . La actividad diapírica reciente con emanaciones constantes de lodo es determinante en la estructura de la comunidad coralina en el sector occidental .
PALABRAS CLAVE: Coberturas fondo marino, Procesos kársticos, Formaciones diapíricas, Caribe colombiano .
ABSTRACT
Seafloor coverage and benthic landscapes associated to diapiric (mud volcanism) formations in Salmedina Banks, Colombian Caribbean continental shelf. The submarine ecological units around an active mud volcano in the offshore banks of Salmedina, Colombian Caribbean, were determined and mapped employing geo-referenced submarine video-transects, bathymetry analysis, and interpretation of satellite imagery . Distribution, relative cover, and the most representative benthic, sessile organisms of the different ecological units are described . In the shallow zone of the banks, 60 % of the
117-135

118 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
substrate was covered by algae and only 10 % by living corals . The relative cover of living corals in deeper zones increased to 33 % in semi vertical slopes and up to 55 % on the upper flats of deeper located banks . The dominant coral species overall were Siderastrea siderea and Montastraea annularis . A determining natural factor in the reef structure in the zone seems to be recent diapiric activity, with a constant mud emanation in the western sector . The Banks of Salmedina are under the influence of turbid plumes that seemingly increase sedimentation and are impacted by blast fishing as well . Although the shallowest zone of the bank shows the highest affectation, the coral community in the deeper zones still exhibit a noticeable cover of living coral, even higher than in other areas in the region with similar features . The recent diapiric activity of constant mud ejection determines the coral community structure in the western sector .
KEYWORDS: Seafloor coverage, Karstic processes, Diapiric (mud volcanic) formations, Colombian
Caribbean .
INTRODUCCIÓN
La importancia ecológica de los arrecifes coralinos es reconocida a nivel
mundial debido a su biodiversidad y elevada producción biológica . El progresivo
y avanzado deterioro generalizado de estos ecosistemas, especialmente en el Caribe,
debido a múltiples factores, entre los que se destacan la sobrepesca, la sedimentación,
el enriquecimiento por nutrientes (eutrofización) y el calentamiento global (Burke
y Maidens, 2004), ha convertido en una prioridad los estudios sobre el estado y
distribución de los arrecifes de coral, principalmente para los países caribeños .
La generación de mapas ecológicos o de coberturas a partir de muestreos in situ y con ayuda de imágenes satelitales y fotografías aéreas, utilizados anteriormente
para valorar ecosistemas terrestres, se ha implementado con gran éxito para el mapeo
de áreas coralinas (Díaz, 1998) . En Colombia se han realizado varios trabajos
utilizando este tipo de metodología (Monsalve y Restrepo, 1990; Sánchez et al ., 1992; Díaz et al ., 1996a, 1996b, 1997, 2000), particularmente en áreas conocidas y
de gran extensión, dejando de lado formaciones menos visitadas pero no de menor
importancia ecológica .
Localizados en la parte central de la plataforma continental del Caribe
colombiano, los bancos de Salmedina, al igual que los demás bajos e islas característicos
del área, son producto de una progresiva colonización coralina sobre bajos de origen
diapírico (Vernette, 1985), y constituyen el límite norte de un cordón arrecifal que se extiende
por unos 200 km en sentido NNE-SSW hasta Isla Fuerte (Geister, 1983; Díaz et al., 1996a) .
Los bancos de Salmedina ocupan una extensión aproximada de 10 km2, de los cuales 4
km2 corresponden a áreas propiamente coralinas (Ricaurte et al., 2004), prácticamente
desconocidas pero de gran importancia para las comunidades de pescadores de la región .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 119
La presencia de actividad diapírica de lodo en estos bancos los hace particularmente
interesantes, ya que esta condición les confiere características especiales y los constituye
en un escenario privilegiado para el estudio de este fenómeno y su influencia sobre las
comunidades biológicas, especialmente las coralinas . Hasta el momento, el interés por los
bancos de Salmedina se ha centrado en el riesgo que representan para la navegación, ya
que se localizan en la trayectoria de las rutas de acceso al puerto de Cartagena de Indias,
uno de los de mayor tráfico en el Caribe colombiano (Ricaurte et al., 2004) . No obstante,
aparte de una descripción somera por Díaz et al. (2000), no han sido objeto de estudios
detallados . El objetivo de este trabajo es describir la composición, la distribución y el
estado de las formaciones coralinas de los bancos de Salmedina como un aporte a su
conocimiento y un insumo para la implementación de medidas de manejo tendientes
a la conservación de este sector poco conocido del Caribe colombiano .
MATERIALES Y MÉTODOS
Los bancos de Salmedina se localizan entre las latitudes 10º21’15’’ y 10º
23,10’’N y las longitudes 75º38’05’’ y 75º41,45’’ W, aproximadamente a 7 km al
W de la punta norte de la isla de Tierra Bomba, frente a la ciudad de Cartagena .
(Figura 1) . Están conformados por cinco elevaciones, a saber: banco Salmedina,
banco Burbujas, banco Ukura, banco Kubina y banco Ygio, que suman una
extensión de 10 .5 km2, de los cuales 4 .45 km2 corresponden a las plataformas más
altas (Ricaurte et al., 2004) .
Figura 1. Área de estudio: Bancos de Salmedina frente a la bahía de Cartagena.

120 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Durante 2004 se llevaron a cabo 17 transectos de video sobre los bancos
de Salmedina, cuya longitud varió entre 240 y 960 m, trazados intentando cubrir
homogéneamente el área de estudio; trece de ellos fueron dispuestos en sentido
sur-norte siguiendo la corriente predominante en la zona, con el fin de economizar
tiempo y energía, y los cuatro restantes se trazaron en sentido este-oeste para cubrir
las zonas intermedias . Los videos se realizaron mediante buceo autónomo empleando
una cámara digital (Panasonic 3CCD, PV-GS70®) dentro de un recipiente estanco
(Equinos PROPAC 6-GS70®) . Durante las filmaciones, la cámara se mantuvo siempre
aproximadamente a una distancia de 2 m del fondo, con la lente perpendicular a éste; para
mantener la dirección constante a lo largo del transecto, un buzo acompañante guiaba
al camarógrafo unos metros adelante de éste empleando una brújula . Los transectos
fueron georeferenciados usando un boyarín en superficie, equipado con un GPS
(Garmín MAP76®), que registraba la posición aproximadamente cada 30 segundos; un
tercer buzo, en superficie, se encargaba de mantener el boyarín perpendicular sobre el
camarógrafo para registrar datos precisos de posición (Figura 2) .
Los datos obtenidos en los transectos se complementaron con 42 inmersiones en apnea . En cada punto de inmersión se anotó el tipo de sustrato y la posición geográfica; los puntos georreferenciados, tanto de los video-transectos como en las observaciones complementarias, fueron superpuestos sobre un mapa batimétrico de la zona a escala 1:12500, utilizando el programa computacional MapInfo Professional 5® .
Figura 2. Diagrama de la metodología empleada en los video-transectos .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 121
Figura 3 . Localización en los bancos de los video-transectos y los puntos de observación (profundidades en metros) .
Las filmaciones fueron examinadas imagen por imagen cada 30 segundos, hasta obtener toda la secuencia a lo largo del video-transecto y cada una fue proyectada sobre el mapa batimétrico, incorporando los datos de posición registrados con ayuda del programa MapInfo Professional 7 .0® (Figura 3) . Se elaboró un mapa de coberturas detallado del área con base en los cambios topográficos y su relación con las variaciones en el tipo de cobertura detectado en las imágenes y en las observaciones hechas in situ .
El mapa de coberturas fue sobrepuesto a una imagen del satélite SPOT de diciembre 20 de 1990, utilizando el programa Surfer 5® . Con ello, al detectarse variaciones en la coloración del fondo atribuibles a cambios en la cobertura, inferidos por los registros de las filmaciones y distintos a los producidos por los cambios de profundidad, se complementó la información obtenida mediante interpolación de los video-transectos y los muestreos puntuales .
Los tipos de cobertura se determinaron con base en las unidades ecológicas de áreas coralinas propuestas por Díaz et al. (2000) para permitir la comparación de la información con la existente para otras áreas del Caribe colombiano . No obstante, dicha clasificación fue complementada adicionando nuevas unidades ecológicas y subdividiendo otras según características morfológicas propias del área de estudio . Para la identificación de los organismos más representativos de cada unidad ecológica, se examinaron las filmaciones y se ubicaron cinco puntos al azar sobre la pantalla; la filmación fue detenida cada 30 s y se anotó el tipo de sustrato u organismo marcado en cada uno de los puntos; en promedio se obtuvieron unos 200 puntos por transecto; según lo permitió la resolución del video, se anotó hasta el menor nivel taxonómico posible y se calcularon las abundancias relativas .

122 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
El procesamiento de la imagen satelital se efectuó mediante el programa ENVI (Environment for Visualizing Images) versión 3 .5® y fue georreferenciada a partir de una imagen ya corregida, usando la técnica de “registro imagen-imagen” y empleando el algoritmo bilinear para el ajuste de los puntos de control mediante translación y rotación de la imagen . Las principales características, bandas espectrales y resolución de la imagen se presentan en la Tabla 1 . La clasificación multiespectral se realizó en forma no supervisada por “clases”. El sistema agrupó los píxeles con características espectrales similares, permitiendo clasificar posteriormente los tipos de cobertura, y se agruparon las propiedades espectrales similares de los píxeles .
Tabla 1. Características generales de la imagen del satélite SPOT 2 de diciembre 20 de 1990 y bandas espectrales y resoluciones de los sensores (según http://www .spotimage .fr/html/_253_311_554_320_ .php, consulta de diciembre de 2004) .
Radiómetro ARV-2KJ 641 .329Fecha 20/12/1990Hora 15h 37m 47sElevación del sol 50.8ºAzimut del sol 150 .0Orientación 8.8ºIncidencia 2 .0(D)Modo espectral multibanda Pancromático: 0.5-0.73 μm
B1: verde (0.5-0.59 μm)B2: rojo (0.61-0.68 μm)B3: infrarrojo cercano (0.78-0.89 μm)
Resolución 15-20 m
Con base en la batimetría de alta resolución levantada para la zona
(Andrade et al., 2005) se elaboró una imagen “batimétrica”, que fue transformada
hasta una resolución espacial de 15 m, coincidente con la de la imagen satelital .
Posteriormente se extrajo una ventana correspondiente al área de los bancos de
Salmedina y se utilizó la imagen batimétrica para construir una máscara que
descartó los valores de píxeles superiores a 40 m de la imagen satelital, con lo
cual se resaltaron los píxeles correspondientes a los bancos y se eliminó el ruido
introducido por las aguas turbias circundantes que pudiera alterar la eficiencia
de los algoritmos de clasificación . Para esta clasificación no supervisada se
utilizó el algoritmo de agrupación que aplica la técnica de análisis de datos auto-
organizados “ISODATA” (Iterative Self-Organizing Data Analysis Technique).
Se calcularon los valores promedio de cada clase que estaban distribuidos dentro
de todos los datos e iterativamente se agruparon los píxeles sobrantes mediante

Instituto de Investigaciones Marinas y Costeras - INVEMAR 123
la técnica de la mínima distancia . Cada iteración recalcula el valor medio y
reclasifica los píxeles con respecto al nuevo valor medio (Jensen, 1996) .
Con la clasificación obtenida se identificaron las zonas donde las
clases son similares y se hizo una comparación espacial con los sitios donde
se identificaron determinado tipo de cobertura respetando los valores de
profundidad similar . ISODATA representa un completo y equilibrado grupo de
procesos heurísticos que han sido incorporados en un algoritmo de clasificación
iterativa . Muchos de esos procesos son el resultado de experiencia ganada a través
de la experimentación y es auto-organizativo porque requiere relativamente poca
información previa dada por el usuario .
RESULTADOS
Las observaciones realizadas y la interpretación de las imágenes
satelitales revelaron que los bancos de Salmedina poseen una inusitada diversidad
de paisajes; se obtuvo un mapa de coberturas del área que discrimina 12 unidades
ecológicas diferentes (Figura 4) que difieren entre sí en el tipo de cobertura
dominante o en características morfológicas relevantes que permiten adoptarlas
como unidades independientes; los rasgos predominantes de cada unidad se
describen en la Tabla 2 . Las zonas menos profundas se encuentran sobre la meseta
del banco Salmedina, el mayor de los cinco que componen el área de estudio,
que alcanza profundidades entre 4 y 8 m en su parte más somera; la meseta está
cubierta en su mayoría por sustrato calcáreo colonizado por algas verdes, algas
calcáreas y macroalgas verdes y pardas, principalmente Dictyota sp . con un 60 %
del área total . Sobreviven colonias muy aisladas de corales masivos (Montastraea
annularis, Siderastrea siderea) que conforman menos del 10 % de la cobertura
(Figura 5A) . En algunos sectores de la meseta el sustrato se eleva 2 m sobre el
fondo, formando terrazas surcadas por canales en los que se acumula arena y
escombros pequeños de coral; la cobertura en las crestas de las terrazas es similar
al tapete de algas del resto de la meseta, con una dominancia clara de las algas (70
%) sobre el coral (Figura 5B), aunque en los surcos sobreviven algunas colonias
grandes de corales masivos y octocorales .

124 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
El cantil norte de Salmedina y Burbujas y el cantil alrededor de Ukura,
Kubina e Ygio poseen pendientes muy pronunciadas, con paredes semi-verticales que
caen varios metros y cuya cobertura coralina corresponde principalmente a colonias
costrosas de Montastraea sp . que cubren en promedio un 33 % de la totalidad del
sustrato; algunos antipatarios, frondosas macroalgas verdes y grandes esponjas
barril (Xetospongia muta) complementan el paisaje (Figura 5C) . En contraste con
el sector norte, los sectores sur y occidental del cantil de los bancos Burbujas y
Salmedina carecen de cobertura viva . Se trata de un sustrato arenoso donde el
elemento predominante, casi en un 100 %, es arena fina con algunos parches de
coral que desaparecen rápidamente en los primeros metros de profundidad (Figura
5D) . Al sur del banco Salmedina la arena da paso a un sustrato totalmente cubierto
por escombros de corales ramificados, posiblemente Porites porites, que cubren el
cantil hasta varios metros de profundidad (Figura 5E) .El banco Salmedina presenta dos formaciones morfológicas muy especiales
y puntuales . La primera se encuentra en el sector suroccidental del cantil donde el sustrato está conformado por rocas de diversos tamaños, dando la apariencia de un “derrumbe” de grandes proporciones (unidad Roc); algunas rocas han sido colonizadas por algas, hidrocorales (Millepora sp .) y esponjas, pero en general el desarrollo coralino es muy bajo en este sector . La otra formación consiste en dos depresiones que se ubican en el sector nororiental de la meseta del Banco; su contorno es circular, de unos 150 m de diámetro y 10 m de profundidad, con paredes muy inclinadas que poseen una cobertura similar a la encontrada en el cantil norte (unidad Cmix);
Figura 4. Unidades ecológicas presentes en los bancos de Salmedina .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 125
Convención Biotopo Descripción
T-Oct-Cmix Terrazas de octocorales-corales mixtos
Estructuras coralinas a manera de espolones que se elevan 1-2 m sobre el fondo arenoso, surcados por canales en dirección norte, están cubiertas por varias especies de corales y octocorales .
Oct-Cmix Octocorales-corales mixtos
Tapete coralino conformado principalmente por corales masivos y octocorales, algas calcáreas, esponjas y macroalgas .
P-Cmix Paredes de corales mixtos
Cantil muy vertical, colonizado por diferentes especies de corales costrosos, esponjas y antipatarios .
T-Algesc Terrazas de algas sobre escombros
Formaciones coralinas que se elevan 1 ó 2 m del sustrato, cubiertas por macroalgas verdes e incrustantes y gorgonáceos en algunas zonas, surcadas por valles de fondo arenoso .
Algesc Algas sobre escombros
Sustrato calcáreo colonizado por macroalgas e hidrocorales, algunas zonas cubiertas por arena y escombros de coral .
Are Arena Sustrato dominado por arenas finas .
Esc Escombros coralinos
Sustrato formado por escombros de Porites porites colonizados por algas incrustantes y esponjas y octocorales dispersos .
Apal-m Acropora muerta
Setos muertos de Acropora palmata con colonias dispersas de corales masivos . Corresponde a la versión degradada de la unidad palmata-strigosa de Díaz et al. (2000) .
Algesc-Litos Algas sobre coral y lodolitos
Sustrato dominado por rocas de lodo consolidado, colonizadas parcialmente por algas, y escombros de coral también colonizados por algas .
Are-lodo Arena y lodo Arena mezclada con lodo .
L-Litos Lodolitos y lodo fino
Planicie de lodolitos pequeños y lodos finos, ausencia de macroalgas, octocorales y otras formas de vida sésil .
Roc Rocas Sustrato rocoso, rocas sueltas de diversos tamaños .
Algcal Algas calcáreas Fondo dominado por algas calcáreas que crecen sobre extensas terrazas coralinas que se elevan 2 m sobre el fondo arenoso, corales masivos dispersos y octocorales .
Sed Sedimentos finos
Sustrato conformado por una capa profunda de sedimentos finos y parches de octocorales muertos .
Tabla 2. Unidades ecológicas y sus rasgos morfológicos y de cobertura más relevantes .
el fondo plano de las depresiones está cubierto por una gruesa capa de sedimentos finos y lodosos, y acumulaciones de octocorales muertos agrupados hacia el centro; los únicos organismos vivos evidentes en esta zona se hallan en un tapete irregular de algas filamentosas que crecen sobre el fango (Figura 5F) .

126 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Figura 5. A.Tapete algal predominante sobre la meseta del banco Salmedina (unidad Algesc) . B. Surco entre dos terrazas al norte del banco (unidad T-Algesc) . C. Cobertura predominante en el cantil norte del banco Ygio (unidad Cmix) . D. Fondo del cantil arenoso al occidente del banco Burbujas (unidad Are) . E. Escombros de coral en el cantil sur del banco Salmedina (unidad Esc) . F. Fondo del cráter en la meseta del banco Salmedina (unidad Sed) .
A
C
E
B
D
F

Instituto de Investigaciones Marinas y Costeras - INVEMAR 127
El área con comunidades coralinas vivas cubre un total de 1 .5 km2 de las plataformas superiores de los bancos; los bancos del sur, Kubina y Ukura, están dominados por un paisaje de octocorales, principalmente del género Plexaura, que crecen entre formas de corales masivos y costrosos (Diploria sp., Montastraea sp., S. siderea), cubriendo en promedio 55 % del sustrato; también se encuentran representadas varias especies de algas calcáreas, dominando Halimeda sp ., esponjas y algas incrustantes; las macroalgas frondosas ocupan menos de 20 % del sustrato (Figura 6A) . Las zonas más elevadas en estos dos bajos están conformadas por setos muertos de Acropora palmata que cubren 55 % del sustrato; en algunos sectores se desarrollan parches de colonias costrosas de Montastraea spp ., colonias masivas de S. siderea y coral de fuego Millepora sp . sobre los esqueletos de A. palmata; la cobertura de algas en esta zona es muy baja, inferior a 5 % (Figura 6B) .
La meseta del bajo Ygio está dividida en dos sectores: el sector sur, un poco más profundo, está dominado por corales y octocorales, un ambiente similar al de los bajos Ukura y Kubina (unidad Oct-Cmix), pero con la característica especial de presentar una densidad considerable de esponjas barril (X. muta) . El sector norte de la meseta está dominado totalmente por terrazas de origen coralino cubiertas casi en su totalidad por un tapete continuo de algas calcáreas como Halimeda sp . y Amphiroa sp ., que cubren 50 % del total del sustrato; el resto de la cobertura es compartida por algas incrustantes, Millepora sp . y esponjas incrustantes (Figura 6C) . Este tapete sólo es interrumpido por canales profundos entre terrazas donde se acumulan restos de algas calcáreas que cubren el fondo y en sus paredes prosperan corales de formas foliáceas, masivas y costrosas (Agaricia sp ., Montastraea spp . y Siderastrea sp .) y diversas especies de octocorales .
La meseta superior del banco Burbujas presenta una clara división en dos paisajes definidos y muy diferentes . El sector norte corresponde a una unidad ecológica muy similar, en composición y cobertura de coral vivo, a los bancos Kubina y Ukura, pero se diferencia de éstos porque el sustrato se eleva hasta 2 m por encima del fondo formando terrazas que están surcadas por valles arenosos, casi siempre orientados en sentido norte-sur. Sobre las mesetas se desarrollan “jardines” de octocorales de varios géneros que crecen sobre tapetes coralinos de formas costrosas, foliáceas y masivas, Halimeda sp ., esponjas de varios tipos, algas incrustantes, algas pardas (Dyctiota sp .) y Millepora sp .; la cobertura de coral vivo es de 40 %, contra 20 % de coral muerto cubierto por algas (Figura 6D) .
Hacia el sur del banco Burbujas la zona de terrazas termina abruptamente dando paso a un valle que domina todo el sector sur del banco; el sustrato en el fondo del valle está totalmente cubierto por una capa de lodo fino y lodolitos pequeños, con una aparente ausencia de organismos sésiles (Figura 6E) . Hacia la periferia del valle se encuentra una franja de transición, entre la zona coralina y la zona de lodos, donde el sustrato es de arena blanca dispuesta en rizos (ripple marks) paralelos al oleaje en cuyos surcos se acumula lodo y algunos lodolitos; octocorales muy dispersos crecen fijados sobre lodolitos grandes o fragmentos coralinos desprendidos de las terrazas (Figura 6F) .

128 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Figura 6. A. Formaciones coralinas predominantes en el banco Ukura (unidad Oct-Cmix) . B. Cementerio de Acropora sp . (unidad Apal-m) . C. Tapete de algas en la meseta del banco Ygio (unidad Algcal) . D. Paisaje característico de las terrazas coralinas del banco Burbujas (unidad T-Oct-Cmix) . E. Vista parcial del fondo del valle de lodo (unidad L-Litos) . F. Vista parcial de la zona de transición en la periferia del valle (unidad Are-Lodo), las piedras grises en las líneas de sedimentos más oscuros son lodolitos . G. Lodolitos parcialmente incrustados por algas en la falda sur de la cima en el banco Burbujas (unidad Alges-Litos) .
A
C
E
G
B
D
F

Instituto de Investigaciones Marinas y Costeras - INVEMAR 129
En el centro del valle, en medio de la planicie de lodo, se eleva un domo de 8
m de altura, cuyas laderas presentan dos sectores claramente definidos, uno al norte,
dominado por terrazas de coral similares a las que dominan todo el sector norte del
banco, aunque un poco más dispersas, y el otro en las secciones occidental, oriental
y sur del domo, que están cubiertas por lodolitos de diferentes tamaños, la mayoría
colonizados por algas calcáreas . Hacia el norte del área se encuentran escombros
de coral entre los lodolitos (Figura 6G) . En la porción más elevada de la cresta la
actividad diapírica es evidente y se manifiesta con un flujo constante de lodo; los
lodos finos en suspensión enturbian el agua dificultando la observación clara del
paisaje . Sin embargo, a pesar de esta limitación, se puede afirmar que el sustrato no
cambia sustancialmente, siguiendo dominado por rocas de lodo; se logró observar
dos octocorales, posiblemente Plexaura sp ., sobreviviendo en estas condiciones .
El análisis de la imagen satelital reveló la presencia de cinco diferentes
coloraciones agrupando los píxeles por su grado de reflectancia (Figura 7); los
cambios de coloración correspondieron primordialmente a variaciones en la
profundidad (desde amarillo en las zonas más someras entre 4 y 5 m de profundidad
hasta rojo, por debajo de 30 m .) . La imagen fue usada para aumentar la resolución
de la batimetría en los primeros 10 m de profundidad, ya que permitió ver detalles
mucho más finos de la morfológica de la zona y corregir los contornos generados por
la interpolación de los datos batimétricos (Figura 8) . La imagen también resultó útil
para diferenciar cambios de cobertura en ambientes someros y muy contrastantes,
siempre que la profundidad se mantenía con escasa variación y que observaciones
de campo permitían reconocer el tipo de sustrato mostrado por la imagen (Figura
9); gracias a ello se logró delimitar el parche de arena sobre el banco Salmedina,
el cual, debido a su gran extensión, presentaba dificultades para ello a partir de las
observaciones in situ .
Por otro lado, la panorámica general que ofrece la imagen satelital permitió
identificar y ubicar accidentes importantes en el área de estudio, como el volcán
de lodo del banco Burbujas o su pluma de lodo (Figura 10), haciendo posible
comprender la magnitud del fenómeno diapírico observado in situ . En general, la
imagen satelital permitió dar mayor detalle al mapa de coberturas incrementando
la resolución de la batimetría y brindando la posibilidad de definir contornos a gran
escala de algunos rasgos submarinos a poca profundidad . Debido a que el área de
estudio está rodeada por fondos profundos, la caracterización con mayor detalle de
éstos a partir de la imagen se hace muy difícil; en estudios realizados en ambientes
más someros, la definición de las coberturas desde imágenes satelitales es mucho
más detallada (Genet, 1992) .

130 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Figura 7. Imagen satelital SPOT tratada de los bancos de Salmedina .
Figura 8. Batimetría sobrepuesta a la imagen satelital . Se aprecian las diferencias en la zona de terrazas coralinas (a) y en la zona nororiental de Salmedina (b) .
Figura 9. Detalle de la imagen satelital que muestra el cambio de cobertura sobre el banco Salmedina .
a bContornos reales de las terrazas coralinas (en amarillo)
Contornos generados por la batimetría
Dirección realmostrada por la imagen de satélite
Dirección de ladepresión según labatimetría

Instituto de Investigaciones Marinas y Costeras - INVEMAR 131
Figura 10. Detalle de la imagen satelital mostrando el volcán de lodo sobre el banco Burbujas .
DISCUSIÓN
Estudios recientes en los bancos de Salmedina (Orejarena et al., 2004; Ricaurte
et al., 2004) muestran que éstos presentan diferencias muy marcadas entre sí en morfología,
profundidad, nivel de la actividad diapírica y grado de exposición al tren de olas y a plumas
de agua continental . Esta diversidad de factores favorece el desarrollo de múltiples biotopos
en un área relativamente pequeña .
El aporte de aguas continentales superficiales, cargadas de nutrientes y
sedimentos, favorecen el crecimiento algal y afectan negativamente el desarrollo
coralino (Alvarado y Corchuelo, 1992); aportes de este tipo, provenientes
principalmente del Canal del Dique, un tributario artificial del río Magdalena, llegan
a los bancos de Salmedina en forma de parches y plumas turbias (Orejarena et al., 2004) . Estas masas de agua turbia son muy superficiales y pueden explicar la
dominancia de macroalgas en la cobertura en las áreas menos profundas de los bancos,
entre 4 y 8 m, donde la cobertura algal alcanza valores de hasta 70 % y las coberturas de
coral vivo no sobrepasan 10 % . Por otro lado, la presencia de grandes extensiones de
escombros de P. porites y setos colapsados de A. palmata muestran que hasta tiempos
recientes la cobertura de coral vivo era considerable pero que resultó severamente afectada
aparentemente por la pesca con explosivos, una práctica frecuente en la región (Díaz et al ., 2000) .
El arrastre continuo de sedimentos y escombros coralinos causado
por las fuertes corrientes superficiales, que predominantemente van hacia el sur,
determinan en buena parte el paisaje submarino al sur de Salmedina . Éste consiste
principalmente de acumulaciones de escombros coralinos y arena desprovistos de

132 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
cobertura coralina . En contraste, la meseta de los bancos Ukura y Kubina, ubicados
a menos de 100 m del banco Salmedina, presentan una cobertura de coral vivo
considerable (55 %), superior a la registrada en áreas cercanas similares con mayor
desarrollo arrecifal, tales como bajo Tortuga (40 %) y bajo Bushell (31 %) (Díaz
et al., 2000) . El principal factor que puede explicar esta diferencia es un profundo
cañón que alcanza 38 m de profundidad y que separa estos bajos del Salmedina . Este
último podría estar actuando a barlovento como una barrera que mantiene protegido
el sector de sotavento; sin embargo, algunas especies hermatípicas importantes ya
han desaparecido, a juzgar por la abundancia de escombros y setos muertos de A. palmata sobre las crestas superiores en esta área .
La influencia de las plumas de agua turbia disminuye notablemente en los
primeros 7 a 8 m de la columna de agua; por debajo de 9 m el agua permanece clara la
mayor parte del año, lo que favorece el desarrollo coralino en este sector . Las mesetas
de los bajos Ygio y Burbujas exhiben una cobertura de coral vivo de 40 %, aunque la
dominancia de especies como S. siderea y M. annularis, particularmente resistentes
a la sedimentación y exitosas en la competencia con otras especies coralinas (Díaz
et al., 1996b; Ferriz-Domínguez y Horta-Puga, 2001) y la presencia de amplias
praderas de algas calcáreas, que suelen desarrollarse sobre el sustrato duro dejado
por los escombros de coral (Frydl, 1982), muestran que estas formaciones están
sometidas a variaciones fuertes de ciertas condiciones ambientales, especialmente
de las tasas de sedimentación .
El alto grado de exposición al oleaje de fondo (swell) proveniente
predominantemente del nor-noreste, propicia un mayor crecimiento coralino vertical
a barlovento, creando una mayor elevación del andamiaje arrecifal y pendientes
pronunciadas (Ricaurte et al ., 2004); este patrón se repite en cada uno de los bajos de
Salmedina, al igual que en otras estructuras arrecifales de la plataforma continental
del Caribe colombiano (Díaz, 1998) . La cobertura del talud norte de los bancos,
cubierta por formas costrosas de coral, esponjas tubulares y antipatarios, es típica
de este tipo de paisaje con alto grado de pendiente y escasa iluminación (Sánchez y
Ortiz, 1992; Díaz, 1998) .
De acuerdo con Ricaurte et al. (2004), el origen del valle que domina el
sur del bajo Burbujas podría estar relacionado con el desplazamiento de material
diapírico al formarse un domo descrito por Vernette (1985) . La estructura prominente
que se observa en medio del valle es muy probablemente de origen diapírico, pues
actualmente se encuentra en actividad; en la cima de la prominencia existe una
emanación constante de lodo en un área de unos 50 m2 acompañada de una copiosa
salida de burbujas de gas en varios sectores y la formación de una “nube” turbia de
varias decenas de metros . El flujo continuo de lodo que se solidificó sobre los flancos

Instituto de Investigaciones Marinas y Costeras - INVEMAR 133
oriental, occidental y sur del domo evitan el desarrollo de formaciones coralinas; la
dominancia de algas costrosas en este ambiente sugiere una alta resistencia de estos
organismos a perturbaciones ambientales y a la turbidez . En contraste, el flanco
norte está menos expuesto al flujo de lodo, y la fuerte corriente predominante del
norte la excluye de la influencia de los sedimentos finos, propiciando el desarrollo
coralino .
El resto del valle consiste en sustrato cubierto por una capa de lodos finos
desprovista de organismos epibentónicos, debido posiblemente al aporte continuo
de lodo proveniente de las estructuras diapíricas vecinas (Ricaurte et al ., 2004) .
Hacia los límites del valle, donde la influencia del lodo disminuye, el paisaje cambia
abruptamente a terrazas de jardines de octocorales sobre terrazas de coral, donde se
concentra gran cantidad de peces; ello sugiere la existencia de un efecto secundario
de la actividad diapírica en la comunidad arrecifal, pero hace falta mayor información
que sustente esta apreciación .
Las dos conspicuas depresiones circulares que se encuentran en el bajo
Salmedina son evidentemente de origen kárstico . Al igual que en los complejos
arrecifales de las islas del Rosario, el bajo Tortuga y el archipiélago de San Bernardo,
estas estructuras debieron originarse durante una de las glaciaciones del Pleistoceno,
como resultado de la disolución de las formaciones calcáreas pre-existentes, al quedar
éstas expuestas a la acción de las aguas de lluvia, creándose un intrincado sistema
de cavernas; posteriormente, al elevarse nuevamente el nivel de mar, inundando las
estructuras, los techos de las cavernas colapsaron en algunas zonas produciendo
hondas depresiones que moldean el actual paisaje de los arrecifes (Díaz et al ., 2000;
López-Victoria y Díaz, 2000) .
Para otros bajos arrecifales de la plataforma continental del Caribe
colombiano se han descrito características similares a las halladas en el bajo
Burbujas . Por ejemplo, Díaz et al . (1996b) registraron en el bajo Bushnell una notoria
disminución de la cobertura coralina hacia la mitad sur del bajo, cambiando de un
ambiente coralino a uno dominado por arenas, esponjas y octocorales . Asimismo,
Díaz et al. (2000) mencionaron la presencia de aguas de aspecto lechoso en las
hondonadas del banco Tortuga, atribuidas a la resuspensión de sedimentos muy
finos . La nueva evidencia sobre la influencia que ejercen las emanaciones de lodo
sobre las coberturas y comunidades epibentónicas aledañas, sumada a las analogías
registradas en otras áreas arrecifales de origen y condiciones similares, permiten
sugerir que el diapirismo submarino de lodo es un factor generalizado importante
en la configuración de la composición y la estructura de las comunidades coralinas
a lo largo del cordón de formaciones coralinas y complejos arrecifales de la región
central del Caribe colombiano .

134 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
AGRADECIMIENTOS
Los autores desean agradecer a la Dra . Gladys Bernal, Universidad Nacional
de Colombia Sede Medellín, por su interés, sus comentarios para el manuscrito así
como por su colaboración y la de su grupo en la toma de imágenes submarinas . Al
Sr . José Ladeut por su gran pericia en el timón de las embarcaciones . Al Sr . Capitán
de Navío Mauricio Ospina, director del CIOH, por su disposición para apoyar todos
los esfuerzos necesarios para ir al mar . Este proyecto se llevó a cabo con el apoyo
logístico de la Armada Nacional a través de la Escuela Naval “Almirante Padilla”,
la Dirección General Marítima y sus unidades en Cartagena, y de COLCIENCIAS
en la financiación del proyecto “Estudio oceanográfico integral de los bancos de
Salmedina, Caribe colombiano”. Código: 112609-12399.
BIBLIOGRAFÍA
Alvarado, E . M . y M . C . Corchuelo . 1992 . Los nutrientes, la temperatura y la salinidad provenientes del
Canal del Dique como factores de deterioro en el Parque Nacional Natural Corales del Rosario
(Cartagena, Colombia) . 277-287 . En: CCO (Ed .) . Mem . VIII Sem . Nal . Cienc . Tecn . Mar, Santa
Marta . 608 p .
Andrade, C . A ., G . Bernal, C . Ricaurte, G . Mayo, J . Domínguez, J . Orejarena, A . Gutiérrez y W . Castro .
2005 . Estudio oceanográfico de los bancos de Salmedina, Caribe colombiano . Escuela Naval
Almirante Padilla, Cartagena . 113 p .
Burke, L . y J . Maidens . 2004 . Reefs at risk in the Caribbean . World Resources Institute, Washington . 81 p .
Díaz, J . M . 1998 . Maps for Colombian reefs . Reef Encounter, 12: 11-12 .
Díaz, J . M ., J . A . Sánchez, S . Zea y J . Garzón-Ferreira . 1996a . Morphology and marine habitats of two
southwestern Caribbean atolls: Albuquerque and Courtown . Atoll Res . Bull ., 435: 1-33 .
Díaz, J . M ., J . A . Sánchez y G . Díaz-Pulido . 1996b . Geomorfología y formaciones arrecifales recientes
de Isla Fuerte y Bajo Bushnell, plataforma continental del Caribe colombiano . Bol . Invest . Mar .
Cost ., 25: 87-105 .
Díaz, J . M ., J . A . Sánchez y J . Geister . 1997 . Development of lagoonal reefs in oceanic reef complexes
of the southwestern Caribbean: geomorphology, structure and distribution . Proc . 8th Internat . Coral
Reef Symp ., Panamá, 1: 779-784 .
Díaz, J . M ., L . M . Barrios, M . H . Cendales, J . Garzón-Ferreira, J . Geister, M . López-Victoria, G . H .
Ospina, F. Parra-Velandia, J. Pinzón, B. Vargas-Angel, F. A. Zapata y S. Zea. 2000. Áreas coralinas
de Colombia . Serie Publicaciones Especiales No . 5, Instituto de Investigaciones Marinas y Costeras-
INVEMAR, Santa Marta . 175 p .
Ferriz-Domínguez, N . y G . Horta-Puga . 2001 . Short-term aggressive behavior in scleractinian corals from
La Blanquilla reef, Veracruz Reef System . Rev . Biol . Trop ., 49 (1): 67-75 .
Frydl, P . M . 1982 . Holocene ostracods in the southern Boso Peninsula . Mobil Oil Canada, Calgary,
Canada . Bull . Univ . Tokio, 20, 10 p .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 135
Geister, J . 1983 . Holozäne westindische Korallenriffe: Geomorphologie, Ökologie und Fazies, Facies,
9: 173-284 .
Genet, P . Y . 1992 . Informe final proyectos cartografía fondos submarinos islas del Rosario (Colombia) y
apreciación de sus áreas, a partir de imágenes de satélite SPOT . Bol . Cient . CIOH, 11: 15-26 .
Jensen, J . R . 1996 . Introductory digital image processing . Segunda edición, Prentice Hall, New Jersey .
450 p .
López-Victoria, M . y J . M . Díaz . 2000 . Morfología y estructura de las formaciones coralinas del
archipiélago de San Bernardo, Caribe colombiano . Rev . Acad . Col . Cienc ., 24 (91): 219-230 .
Monsalve, C . B . y J . D . Restrepo . 1990 . Aproximación a la fotointerpretación y cartografía de los
ecosistemas arrecifales, Isla Caribarú, Caribe colombiano . 325-336 . En: CCO (Ed .) . Mem . VII Sem .
Nal . Cienc . Tecn . Mar, Cali . 792 p .
Orejarena, J . A ., J . G . Domínguez, C . Ricaurte, G . Mayo, C . A . Andrade, H . M . Ospina y W . Castro . 2004 .
Variaciones de la concentración de la clorofila-a y su relación con los parámetros físicos medidos
en los bancos de Salmedina durante 2003 - 2004, Caribe colombiano . Bol . Cient . CIOH, 22: 56-63 .
Ricaurte, C ., J . G . Domínguez, G . Mayo, C . A . Andrade, H . M . Ospina y A . de J . Gutiérrez . 2004 . Nota
sobre algunos rasgos geomorfológicos de los bancos de Salmedina, Bol . Cient . CIOH, 22: 64-76 .
Sánchez, J . A . y V . Ortiz . 1992 . Distribución de los corales negros (Antipatharia) de arrecifes entre
Cartagena y el golfo de Urabá, Caribe colombiano . 362-369 . En: CCO (Ed .) . Mem . VIII Sem . Nal .
Cienc . Tecn . Mar, Santa Marta . 608 p .
Sánchez, J . A ., A . Ramírez y R . Quintero . 1992 . Estudio de reconocimiento de la morfología y las
comunidades bénticas de los arrecifes de isla Tesoro, Parque Nacional Natural Corales del Rosario
(PNNCR), Caribe colombiano . 263-276 . En: CCO (Ed .) . Mem . VIII Sem . Nal . Cienc . Tecn . Mar,
Santa Marta . 608 p .
Vernette, G . 1985 . La plateforme continentale Caraïbe de Colombie (du Débouché du Magdalena au
Golfe de Morrosquillo), importance du diapirisme argileux sur la morphologie et la sédimentation .
Tesis Doctor de Estado en Ciencias, Univ . de Bordeaux I, Bordeaux, Francia . 381 p .
FECHA DE RECEPCIÓN: 07/10/2009 FECHA DE ACEPTACIÓN: 16/04/2010


Instituto de Investigaciones Marinas y Costeras - INVEMAR 137
Bol . Invest . Mar . Cost . 39 (1) ISSN 0122-9761 Santa Marta, Colombia, 2010
DISTRIBUCIÓN ESPACIAL, ABUNDANCIA Y RELACIÓN CON CARACTERÍSTICAS DEL HÁBITAT DEL CARACOL
PALA EUSTROMBUS GIGAS (LINNAEUS) (MOLLUSCA: STROMBIDAE) EN EL ARCHIPIÉLAGO NUESTRA SEÑORA DEL ROSARIO, CARIBE COLOMBIANO*
Kelly Gómez-Campo, Mario Rueda y Carolina García-Valencia
Instituto de Investigaciones Marinas y Costeras-INVEMAR, A.A. 1016, Cerro Punta Betín, Santa Marta, Colombia. [email protected] (K.G.C.), [email protected] (M.R.); [email protected] (C.G.V.)
RESUMEN
Se evaluó la densidad poblacional y distribución espacial de Eustrombus gigas (Linnaeus) en el archipiélago Nuestra Señora del Rosario, así como su asociación a diferentes hábitats, a partir del muestreo sistemático de 184 estaciones en dos épocas climáticas, húmeda (mayo-noviembre) y seca (diciembre-abril) que coinciden con el período reproductivo de la especie . En cada estación se contaron y midieron los individuos presentes en un área circular de 1256 .6 m2 mediante buceo autónomo, a la vez que se hicieron observaciones in situ sobre los tipos de hábitats presentes . La densidad promedio (± DE) fue baja comparada con otras regiones del Caribe, 4.0 ind/ha ±10.8 y 3.7 ind/ha ±9.3, para las épocas húmeda y seca, respectivamente . Los individuos adultos, representaron 89 y 64 % de la población muestreada en cada época climática, especialmente agrupados en bajo Tortugas e isla Tesoro . Aunque se observó actividad reproductiva durante la época húmeda, la densidad de individuos estuvo por debajo de niveles críticos que garanticen el éxito reproductivo de la población . El análisis de distribución mostró que la abundancia de adultos estuvo espacialmente estructurada en ambas épocas climáticas, con agregaciones de individuos principalmente en bajo Tortugas e isla Tesoro . Los juveniles no mostraron estructura espacial debido a la baja abundancia observada . Los modelos de regresión múltiple explicaron 48 y 14 % de la varianza total en la abundancia de adultos para la época húmeda y seca, respectivamente, con la profundidad, el tamaño medio de grano y porcentaje de coral mixto como variables predictoras . Se recomienda declarar zonas de protección especial a isla Tesoro y bajo Tortugas para la protección de población desovante; mientras que la misma medida de conservación se sugiere implementar en isla Arena para la protección de juveniles en praderas de pastos marinos .
PALABRAS CLAVE: Archipiélago Nuestra Señora del Rosario, Eustrombus gigas, Abundancia, Distribución espacial, Caracol pala .
137-159
*Contribución No. 1050 del Instituto de Investigaciones Marinas y Costeras-INVEMAR.

138 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
ABSTRACT
Spatial distribution, abundance and relationship to habitat features of the queen conch Eustrombus gigas (Linnaeus) (Mollusca: Strombidae) in Nuestra Señora del Rosario archipelago, Caribbean coast of Colombia. Population density and spatial distribution of the Caribbean conch, Eustrombus gigas (Linnaeus), as well as its association to different habitats were evaluated at 184 stations sampled systematically in two seasons (wet and dry) in Nuestra Señora del Rosario archipelago . Each individual was counted and measured in a circular area of 1256 .6 m2 (sampling unit) by SCUBA diving and in situ observations on the types of habitats were made. The average density (±SD) of the species was low compared with other regions in the Caribbean, 4.0 ind/ha ±10.8 and 3.7 ind/ha ±9.3, for the wet and dry seasons, respectively . Adults represented 89 and 64 % of the population surveyed for each season, and were specifically located at Bajo Tortugas and Isla Tesoro . Although reproductive activity was observed during the wet season, population density was below critical levels to guarantee the population’s reproductive success . The spatial distribution analysis showed that the abundance of adults was spatially structured in both seasons, with important aggregations at Bajo Tortugas and Isla Tesoro . Juveniles did not show spatial structure for neither season due to the low observed abundance . Multiple regression models explained 48 and 14 % of the total variance in the abundance of adults during the wet and dry seasons, respectively; with depth, mean grain size, and percentage of mixed coral as predictor variables . It is recommended to establish Bajo Tortugas and Isla Tesoro as special protected areas in order to maintain the adult spawning stock, while the same conservation measure should be implemented at Isla Arena in order to protect juveniles on seagrass beds .
KEY WORDS: Nuestra Señora del Rosario archipelago, Eustrombus gigas, Abundance, Spatial distribution, Queen conch .
INTRODUCCIÓN
El gasterópodo Eustrombus gigas (Linnaeus), anteriormente llamado
Strombus gigas (Petuch y Roberts, 2007), es conocido comúnmente en Colombia
como caracol pala y debe su gran importancia al valor económico de su carne y al
papel ecológico en su hábitat (Randall, 1964) . Es una especie herbívora que habita
generalmente entre 2 y 20 m de profundidad sobre fondos de praderas de pastos
marinos, parches de arena, cascajo y praderas de macroalgas a lo largo de todo
el Caribe, desde Bermuda y el sur de Florida hasta la costa norte de Sudamérica
(Randall, 1964; Ray y Stoner, 1994; Glazer y Kidney, 2004) .
En Colombia se ha trabajado considerablemente sobre los aspectos
biológicos de E. gigas (Duque, 1974; García, 1991; Lagos, 1994; Chiquillo et al.,
1997), destacando una talla media de madurez sexual de 240 mm y 221 mm para
hembras y machos, respectivamente; y evidenciando una época de reproducción
entre los meses de junio y enero por la presencia de masas de huevos en el hábitat y
la migración de individuos adultos a aguas más someras, que coincide con el periodo

Instituto de Investigaciones Marinas y Costeras - INVEMAR 139
de lluvias o época húmeda . Adicionalmente, Márquez (1993) observó una clara
estratificación de la estructura poblacional en donde los juveniles se agruparon en
aguas someras y sobre praderas de pastos marinos, mientras los adultos en sustratos
arenosos en aguas más profundas, infiriendo una correlación entre la densidad de
individuos, la talla y las características del hábitat .
En las últimas décadas se ha desarrollado a gran escala la pesca comercial de
E. gigas como respuesta a la alta demanda internacional, hasta el punto de llevarlo a la
Lista Roja de Animales Amenazados de la Unión Internacional para la Conservación
de la Naturaleza -UICN, 1994- como especie ‘comercialmente amenazada’, la
especie está además listada en el Apéndice II de la Convención sobre el Comercio
Internacional de Especies Amenazadas de Fauna y Flora Silvestres CITES (CITES,
2003) . Además de la presión pesquera, la destrucción de hábitats de crianza a causa
de diferentes métodos de pesca y contaminación constituye un factor importante
que influye potencialmente en la amenaza de la especie (Mora, 1994) . Por todo lo
anterior, la regulación por parte de CITES para el manejo pesquero y conservación
en los países afectados, pretende propiciar la recuperación de las poblaciones y los
hábitats esenciales para evitar la potencial extinción de la especie . En este sentido
la literatura ha documentado que la creación de áreas marinas protegidas (AMPs)
(Stoner, 1997) y las estrategias de repoblación y trasplante (Stoner y Glazer, 1998),
son medidas de conservación prometedoras para la recuperación de la especie . No
obstante, el diseño de AMPs para la protección de agregaciones reproductivas o
mecanismos de recuperación de poblaciones a través de estrategias de repoblación y
trasplante, requiere el conocimiento de la distribución espacial de la especie objeto
de estudio y su asociación con el hábitat que ocupa (Stoner, 1997, 2003) .
En la última reunión realizada para el manejo del recurso “Taller Regional
para el Desarrollo de Estrategias Colaborativas en el Manejo de Caracol Pala en el
Caribe Sur Occidental, Islas de San Andrés, 2008”, se reconoció que el archipiélago
Nuestra Señora del Rosario (NSR), junto con San Bernardo, son lugares del Caribe
continental colombiano donde aún se registra E. gigas, con algunas observaciones
aisladas en La Guajira y los bajos de Isla Fuerte . Aunque el archipiélago NSR
está declarado como Parque Nacional Natural, es un lugar sometido a intensas
actividades antrópicas, como turismo y pesca, actividades que han conducido a que
la población de E. gigas presente signos de sobreexplotación con una disminución
en las capturas y en su talla media en la pesca artesanal . Medidas de conservación,
como la declaración de áreas intangibles (cerradas a la intervención humana), no

140 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
han rendido los frutos esperados, probablemente por la falta de control y vigilancia
o por la falta de conservación dirigida exclusivamente a E. gigas con base en el
conocimiento del estado actual de la población y su contexto espacial asociado a la
heterogeneidad de hábitats existentes en el área de estudio (Díaz et al ., 2000) .
Es fundamental reconocer que en la naturaleza los organismos se encuentran
formando agregaciones o gradientes en respuesta a características estructurales
controladas por variables bióticas y abióticas que presentan los hábitats (Legendre,
1993), para contribuir a un mayor entendimiento de la dinámica poblacional de los
organismos y su relación con el ambiente . Lo anterior ha redundado en importantes
aplicaciones para el manejo y conservación de especies usando nuevos enfoques de
la ecología, basados en la incorporación de la modelación espacial de poblaciones en
estudios ecológicos (Legendre, 1993) . Esta investigación, basándose en muestreos
intensivos de la población y el hábitat que ocupa, buscó identificar patrones de
distribución espacial y temporal teniendo en cuenta períodos de actividad reproductiva
(época húmeda) y no reproductiva (época seca) de E. gigas en el archipiélago NSR,
además de estimar su abundancia y determinar su relación con variables descriptoras
del hábitat . Esta información permitirá generar un conocimiento fundamental para
apoyar y contribuir a la formulación de planes de repoblación, trasplante y/o diseño
de áreas marinas protegidas dirigidas a conservar la especie .
ÁREA DE ESTUDIO
El archipiélago NSR hace parte del Parque Nacional Natural Corales del
Rosario y San Bernardo (PNNCRSB), se encuentra en el mar Caribe colombiano,
dentro de la jurisdicción del Distrito Turístico, Cultural e Histórico de Cartagena
de Indias en el departamento de Bolívar . Presenta una localización geográfica entre
10°06’ y 10°15’ N y 75°36’ y 75°50’ W (Alvarado et al ., 1986; Díaz et al ., 2000) . De
este Parque hacen parte 28 islas de distinto tamaño (Genet, 1992) . El archipiélago
fue declarado Parque Nacional Natural en 1977 (Figura 1), dada la necesidad de
conservar y proteger áreas naturales poco intervenidas para la realización de
investigaciones científicas, actividades recreativas y de educación ambiental . El
área del PNNCRSB posee un clima tropical costero, con dos épocas climáticas en
el año: época húmeda (mayo-noviembre) con lluvias frecuentes, temperatura del
agua alrededor de 29 °C y oleaje suave; y época seca (diciembre-abril) sin lluvias,
temperatura de 27 °C y oleaje fuerte (Alvarado et al ., 1986; Bohórquez, 2002) .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 141
MATERIALES Y MÉTODOS
Diseño de muestreoTomando como base la cartografía preexistente de fondos potencialmente
favorables para E. gigas en el archipiélago NSR (Díaz et al ., 1996), se diseñó
un muestreo sistemático . Teniendo en cuenta que la época de reproducción del
caracol pala -junio a diciembre- en el área de estudio (Lagos, 1994) coincide con
el aumento de la temperatura del agua, la baja influencia de los vientos Alisios y
el período de mayores lluvias, se organizaron 87 estaciones separadas cada 500 m
durante una época húmeda (agosto 2004) y 97 en una época seca (febrero 2005), que
correspondieron a las mismas estaciones de la época húmeda y 10 complementarias .
Por razones prácticas, las estaciones se agruparon en diferentes sectores segregados
espacialmente (Figura 1): Bajo Tortugas (BT), Isla Tesoro (TROS), Isla Grande
norte (IGN), Isla Grande sur (IGS), Isla Grande oriental (IGO), Isla Rosario (IR) e
Isla Arena (IA) . En cada estación (unidad de muestreo) se hizo una inmersión con
equipo de buceo autónomo (SCUBA), cubriendo un área circular de 1256.64 m²
descrita por un radio de 20 m, en la que se hicieron recorridos concéntricos a 10 y 20
m, alrededor de un lastre de 30 kg atado con un cabo a una boya . Todos los caracoles
Figura 1 . Ubicación y delimitación del archipiélago Nuestra Señora del Rosario (modificado de INVEMAR et al., 2003) .

142 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
encontrados en el área circular se contaron y se midieron, teniendo en cuenta su
longitud total o sifonal LT (medida que se toma del ápice de la espira al final del
canal sifonal) y grosor de labio GL (medida que se toma a 40 mm de profundidad
del borde de la concha en su parte central) . Simultáneamente se consideraron
como variables descriptoras del hábitat los siguientes parámetros: profundidad,
temperatura, salinidad, densidad de vástagos de Thalassia testudinum y tipo de
fondo determinado por medio de la cuantificación de la cobertura del sustrato en
porcentaje a partir de la observación de las proporciones de los diferentes fondos
(porcentaje de esponjas, porcentaje de T. testudinum, porcentaje de arena, porcentaje
de cascajo, porcentaje de algas, porcentaje de coral mixto, porcentaje de octocorales,
porcentaje de Syringodium filiforme) . Adicionalmente, se extrajo una muestra de
sedimento por estación de muestreo para determinar contenido de materia orgánica
y tipo de sedimento de acuerdo con el tamaño de grano .
Análisis de informaciónDensidad. La densidad de individuos de cada estación se estandarizó a
individuos por hectárea (ind/ha) . Para efectos de evaluar la estructura de la población,
los individuos se clasificaron en dos grupos de edades: juveniles observables en
campo (<12 cm de LT) y adultos . Appeldoorn (1988) menciona que un aspecto
inusual en la biología del caracol pala es que el crecimiento en longitud total de la
concha cesa en el momento de la maduración sexual, incrementándose sólo el grosor
del labio de la concha, por lo cual se ha enfatizado que el grosor de labio es una
medida para identificar juveniles y adultos de E. gigas . El ámbito de variación de la
medida de grosor de labio que determina la edad de primera madurez y la fase adulta
varia de >1 mm, utilizado por Pérez y Aldana (2000), hasta 14 .2 mm, registrado por
Appeldoorn (1988), como promedio para el primer grupo de edad de individuos
adultos . Para este estudio se tuvo en cuenta la presencia de un labio extendido y se
consideraron individuos juveniles cuando su grosor de labio no superó 8 mm, de
acuerdo con Lagos (1994) quien determinó el ciclo reproductivo y la talla de madurez
sexual de E. gigas en el área de estudio . La relación grosor de labio vs . longitud total
(Figura 2) se ajustó a una función potencial que corresponde al ámbito de variación de
grosores de labio de individuos maduros registrados en la bibliografía (Appeldoorn,
1988; Pérez y Aldana, 2000) . Se realizó un análisis exploratorio de datos con fines
de conocer las medidas de tendencia central y variabilidad, las cuales condujeron
a aplicar la prueba de Mann-Whitney (U; Zar, 1996) para evaluar diferencias en la
abundancia de juveniles y adultos entre épocas climáticas, períodos que coinciden
con el ciclo reproductivo de E. gigas en el archipiélago NSR (Lagos, 1994) .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 143
Distribución espacial. Se usaron técnicas geoestadísticas (análisis
semivariográfico y kriging en bloques) para identificar patrones de distribución
espacial de la densidad de E. gigas y para estimar la abundancia total efectiva en el
área de estudio por medio del programa . El contexto espacial abordado asumió que
la densidad de E. gigas (variable Z), medida en un sitio (x), representó un proceso
estocástico Z(x), con una variable regionalizada . Lo anterior, unido a un gran número
de estaciones cubiertas en un período de tiempo corto, posibilitó la aplicación de
métodos estadísticos (Cressie, 1993; Legendre, 1993; Burrough y McDonell, 1998) .
Debido a que el propósito del análisis geoestadístico es modelar datos espacialmente
correlacionados, la función de estructura espacial o semivariograma experimental
γ (h)fue calculada como (Matheron, 1971):
y = 6E-08x3.4622
0
5
10
15
20
25
30
35
50 75 100 125 150 175 200 225 250 275 300 325 350
GL
(mm
)
LT (mm)
donde Z (xi) es el valor de la variable (e .g . densidad) en la estación xi, Z
(xi+h) es otro valor de la variable separado de xi por una distancia discreta h y N(h) y es el número de pares de observaciones separadas por h. Para determinar el
tipo de distribución espacial, los semivariogramas experimentales se ajustaron a los
modelos teóricos: esférico, exponencial, lineal y Gaussiano (Burrough y McDonell,
1998) . La selección del mejor modelo se hizo con base en los siguientes criterios:
suma de cuadrados reducida (SCR), coeficiente de determinación (r2) y componente
espacialmente estructurado o varianza espacial explicada (C/(Co+C)) . Los modelos
Figura 2. Relación de la longitud total (LT) y el grosor de labio (GL) de la población de Eustrombus gigas en el Parque Nacional Natural Corales del Rosario y San Bernardo . Se traza la línea de tendencia y su ecuación .

144 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
ajustados permitieron estimar los siguientes parámetros que definieron la función
de dependencia espacial: variabilidad de microescala (Co: efecto pepita); valor
asintótico de la semivarianza que indica la distancia más allá de la cual no existe
estructura espacial (Co+C: meseta) y distancia de influencia espacial o tamaño
relativo del parche (Ao: rango) . La predicción espacial de la variable en los sitios
no muestreados se hizo usando los parámetros estimados del semivariograma,
mediante la técnica de kriging en bloques, la cual posibilitó la elaboración de mapas
de densidad . La abundancia total se estimó mediante la sumatoria de las medias de
densidad de los bloques Z (xi) a través del área irregular muestreada (A) (Burrough
y McDonell, 1998):
εββββ ++++= kk XXXY ...22110
Para evaluar los resultados de kriging se utilizó el análisis de validación
cruzada, en la que los valores observados y estimados por el modelo espacial son
relacionados evaluando el ajuste con una regresión lineal . Una relación 1:1 indica
que el modelo predice exactamente los valores observados, entonces la pendiente (ß)
es 1 .0 y el intercepto (α) es 0 .0 . Todos los análisis geoestadísticos se realizaron en el
programa GS+ Versión7 .0, Gamma Design Software, Geoestadística para Ciencias
Ambientales .
Relación entre la densidad y variables descriptoras del hábitat. Para
evaluar la relación entre variables predictoras del hábitat (profundidad, salinidad,
temperatura, densidad de vástagos, tamaño medio de grano, tipo de sedimento,
porcentaje de materia orgánica en el sedimento, porcentaje de esponjas, porcentaje
de T. testudinum, porcentaje de arena, porcentaje de cascajo, porcentaje de algas,
porcentaje de coral mixto, porcentaje de octocorales, porcentaje de S. filiforme)
y la variable dependiente (densidad de juveniles y adultos) se usó análisis de
regresión múltiple . Inicialmente se verificó la relación lineal entre variables
mediante correlación de Pearson; aquellas variables predictoras correlacionadas
significativamente (p< 0 .05), fueron tomadas en cuenta para el análisis de regresión
múltiple (Zar, 1996):
donde ß0
,ß1
,ß2
,...ßk
son los coeficientes de regresión múltiple y X1, X2,... Xk son las variables descriptoras del hábitat . El modelo se construyó
mediante la técnica de regresión por pasos hacia atrás, es decir, que las variables
potencialmente predictoras fueron removidas una a una hasta obtener el mejor

Instituto de Investigaciones Marinas y Costeras - INVEMAR 145
modelo . La remoción secuencial de variables descriptoras del hábitat se determinó
por una restricción realizada a priori sobre el nivel de significancia que debe tener
cada variable para integrar el modelo (valores del estadístico F>3) . La contribución
independiente de cada variable descriptora del hábitat a la predicción de la densidad
de caracol, fue evaluada por medio de la correlación parcial y la redundancia . Todos
los análisis de correlación y regresión múltiple se realizaron con el programa
STATISTICA 6 .1 Data Analysis Software System .
RESULTADOS
DensidadLa abundancia de individuos no presentó diferencias entre épocas
climáticas ni en adultos ni en juveniles (U87, 97
= 4170 .5; p= 0 .82 y U87, 97
= 3893;
p= 0 .06; respectivamente); sin embargo, es claro el carácter marginal de la segunda
comparación . En época húmeda se registraron 44 individuos en total, de los cuales
11 % fueron juveniles y 89 % adultos . Los individuos juveniles sólo se encontraron
en los sectores isla Grande sur, isla Tesoro e isla Rosario, con densidades medias
en cada sector que no sobrepasaron 1 .1 ind/ha (Tabla 1) . Debido a la baja presencia
de juveniles, los individuos adultos representaron la población de E. gigas en época
húmeda, con una densidad media de 3.6±10.77 ind/ha. En el sector bajo Tortugas,
todos los individuos encontrados fueron adultos con un ámbito de 0 .0-55 .70 ind/ha
y un valor medio de 23.9±23.5 ind/ha, así como en el sector isla Rosario donde se
encontró también una densidad relevante de 9.7±8.70 ind/ha (Tabla 1).
Tabla 1. Medidas de tendencia central (media: ; mediana: M) y dispersión (DE: desviación estándar) de la densidad de individuos (ind/ha) en diferentes sectores del archipiélago NSR . IA: Isla Arena, IGO: Isla Grande (sector oriente), TROS: Isla Tesoro, IGN: Isla Grande (sector norte), BT: Bajo Tortugas, IR: Isla Rosario, IGS: Isla Grande (sector sur) durante dos épocas climáticas, húmeda y seca . --No se encontró ningún individuo en el sector .
Época húmeda (mayo – noviembre) Época seca (diciembre – abril)
Juveniles Adultos Juveniles Adultos
Sector M DE M DE M DE M DE
IA -- -- -- -- -- -- 2 .39 0 .00 3 .84 -- -- --
IGO -- -- -- -- -- -- -- -- -- 0 .72 0 .00 2 .40
TROS 0 .88 0 .00 2 .65 9 .73 7 .96 8 .70 1 .77 0 .00 3 .51 9 .73 0 .00 15 .80
IGN -- -- -- -- 2 .89 0 .00 9 .60 0 .72 0 .00 2 .40
BT -- -- -- 23 .87 15 .92 23 .54 -- -- -- 11 .49 7 .96 17 .84
IR 0 .42 0 .00 1 .83 -- -- -- 1 .45 0 .00 3 .14 0 .36 0 .00 1 .70
IGS 1 .09 0 .00 3 .72 0 .36 0 .00 1 .70 0 .95 0 .00 3 .50 0 .95 0 .00 3 .50
Total 0 .46 0 .00 2 .22 3 .57 0 .00 10 .77 1 .31 0 .00 4 .25 2 .38 0 .00 8 .14
X~
X~ X~ X~X~

146 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
En época seca se encontraron 45 individuos vivos en total, de los cuales 36 % fueron juveniles y 64 % adultos. La densidad media de juveniles fue 1.3±4.25 ind/ha (Tabla 1) . Los valores más altos se registraron en los sectores isla Grande norte (2.9±9.6 ind/ha) e isla Arena (2.4±3.8 ind/ha), seguidos de isla Tesoro, isla Rosario e isla Grande sur, mientras que los sectores bajo Tortugas e isla Grande oriental no presentaron individuos juveniles . Con relación a la población adulta, los sectores bajo Tortugas e isla Tesoro mostraron las densidades medias más altas en el archipiélago (11.5±17.84 ind/ha y 9.7±15.8 ind/ha). Las densidades medias más bajas se registraron en isla Grande sur, isla Grande norte, isla Grande oriental e isla Rosario (<1 .00 ind/ha) . En el sector bajo Tortugas, todos los individuos encontrados fueron adultos, con un ámbito de 0 .0-55 .70 ind/ ha, mientras que en el sector isla Arena no se encontraron individuos adultos .
Distribución espacialPor la presencia de ceros informativos y distribución asimétrica de las
variables analizadas, se transformaron los datos con Log(x+1). Los semivariogramas experimentales de la abundancia de adultos fueron explicados para ambas épocas climáticas por el modelo Gaussiano (λ(h)= Co + C[1-exp(-h2/Ao2)]) en términos de los valores más bajos de SCR y más altos de r2 (Figura 3) . Los semivariogramas alcanzaron asíntotas que definieron distancias de dependencia espacial de 11310 m y 9220 m para las épocas húmeda y seca, respectivamente, con un valor muy alto de varianzas espaciales explicadas (C/(Co+C)= 0 .99) y un valor muy bajo de variabilidad de microescala (Co= 0 .001) (Tabla 2) . La validación cruzada de las interpolaciones de kriging arrojaron valores en los parámetros de la regresión que indicaron mayor calidad de las interpolaciones durante la época húmeda (α= 0 .205; β= 0 .629; r2= 0 .44) que durante la seca (α= 0 .205; β= 0 .379; r2= 0 .07) (Tabla 2) . Dicha calidad se constató con los mapas de desviación estándar (Gómez-Campo, 2005) que mostraron baja variabilidad de la densidad en el área interpolada .
Los mapas de kriging generados a partir de la densidad de individuos mostraron concentraciones importantes de caracoles en el norte y al suroeste del área de estudio (isla Tesoro y bajo Tortugas, respectivamente) (Figura 4) . De esta información, la abundancia total efectiva de E. gigas se estimó en 936±11 individuos para la época húmeda y 422±7 individuos para la seca. La densidad de individuos juveniles no se ajustó a algún modelo, por tanto una idea de su distribución fue determinada usando mapa de cuartiles . Estos mapas indicaron que el 75 % del área interpolada mostró valores de densidad de cero, mientras que el 25 % restante del área mostró densidades ≤15.92 ind/ha (2 ind/1256.64 m2) durante la época húmeda y ≤31.83 ind/ha (4 ind/1256.64 m2) durante la época seca en los sectores isla Grande, isla Tesoro, isla Rosario e isla Arena (Figura 4) .
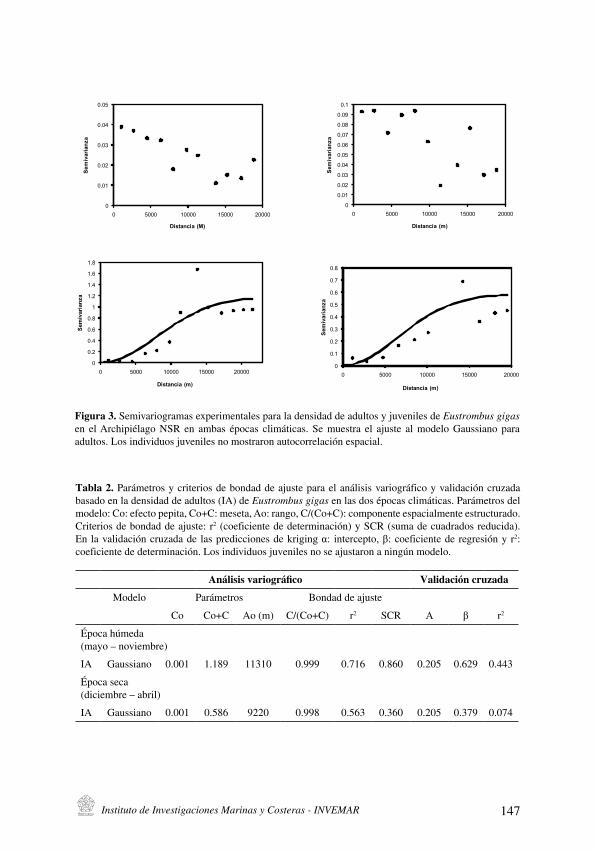
Instituto de Investigaciones Marinas y Costeras - INVEMAR 147
0
0.01
0.02
0.03
0.04
0.05
0 5000 10000 15000 20000
Sem
ivar
ianz
a
Distancia (M)
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
0 5000 10000 15000 20000
Sem
ivar
ianz
a
Distancia (m)
0
0.01
0.02
0.03
0.04
0.05
0.06
0,07
0.08
0.09
0.1
0 5000 10000 15000 20000
Sem
ivar
ianz
a
Distancia (m)
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0 5000 10000 15000 20000
Sem
ivar
ianz
a
Distancia (m)
Tabla 2. Parámetros y criterios de bondad de ajuste para el análisis variográfico y validación cruzada basado en la densidad de adultos (IA) de Eustrombus gigas en las dos épocas climáticas . Parámetros del modelo: Co: efecto pepita, Co+C: meseta, Ao: rango, C/(Co+C): componente espacialmente estructurado . Criterios de bondad de ajuste: r2 (coeficiente de determinación) y SCR (suma de cuadrados reducida) . En la validación cruzada de las predicciones de krigingα: intercepto,β:coeficiente de regresión y r2: coeficiente de determinación . Los individuos juveniles no se ajustaron a ningún modelo .
Análisis variográfico Validación cruzada
Modelo Parámetros Bondad de ajuste
Co Co+C Ao (m) C/(Co+C) r2 SCR Α β r2
Época húmeda (mayo – noviembre)
IA Gaussiano 0 .001 1 .189 11310 0 .999 0 .716 0 .860 0 .205 0 .629 0 .443
Época seca (diciembre – abril)
IA Gaussiano 0 .001 0 .586 9220 0 .998 0 .563 0 .360 0 .205 0 .379 0 .074
Figura 3. Semivariogramas experimentales para la densidad de adultos y juveniles de Eustrombus gigas en el Archipiélago NSR en ambas épocas climáticas . Se muestra el ajuste al modelo Gaussiano para adultos . Los individuos juveniles no mostraron autocorrelación espacial .

148 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Relación entre la abundancia y variables que describen el hábitatDe las variables abióticas descriptoras del hábitat (Tabla 3), se encontró que
la temperatura difirió entre épocas climáticas (H1,183
= 137 .62; p<< 0 .05), siendo más
baja en la época seca (U: p< 0 .05) . De igual manera la salinidad presentó diferencias
estacionales (H1,184
= 146 .94; p<< 0 .05), con valores más bajos durante la época
húmeda (U: p< 0 .05); datos que coinciden con las condiciones físico-químicas de cada
época climática . En lo referente a las variables que describen el fondo, se observaron
diferencias en la densidad de vástagos, 56 .51 vást/m2 (DE= 125 .27) en la época
húmeda y 90 .89 vást/m2 (DE= 162 .27) en la seca, con altas desviaciones en los datos
observados en campo (Tabla 3) . El análisis de sedimentos, detectó una clasificación
que exhibe predominio de ‘arenas medias’ en los sectores isla Arena, isla Grande
oriental, isla Grande norte, isla Rosario e isla Grande sur, con valores de tamaño medio
de grano que van desde 1.11±0.71 hasta 1.34±0.48 Ø en isla Arena e isla Grande norte,
respectivamente, y un valor máximo de 3.53 Ø en isla Grande sur. En los sectores bajo
Tortuga y Tesoro se registraron ‘arenas gruesas y gravas’ con un tamaño medio de
grano de 0.53±0.72 Ø y 0.08±0.48 Ø, respectivamente, y un valor máximo de -0.62
Ø. Por otra parte, la materia orgánica presente en el sedimento registró valores medios
entre 3 y 4 %, el máximo valor se encontró en el sector isla Grande Sur con 8 .61 %,
mientras que el valor más bajo se presentó en Tesoro con 2 .30 % .
Figura 4. Mapas de cuartiles y kriging para la densidad de juveniles y adultos . respectivamente (ind/ 1256 .64 m2) en el archipiélago NSR durante ambas épocas climáticas .
Isla Tesoro
Isla TesoroIsla Tesoro
Isla Tesoro
Isla Grande
Isla GrandeIsla Grande
Isla Grande
Isla Rosario
Isla RosarioIsla Rosario
Isla Arena
Isla ArenaIsla Arena
Isla Arena Bajo Tortugas
Bajo TortugasBajo Tortugas
Bajo Tortugas
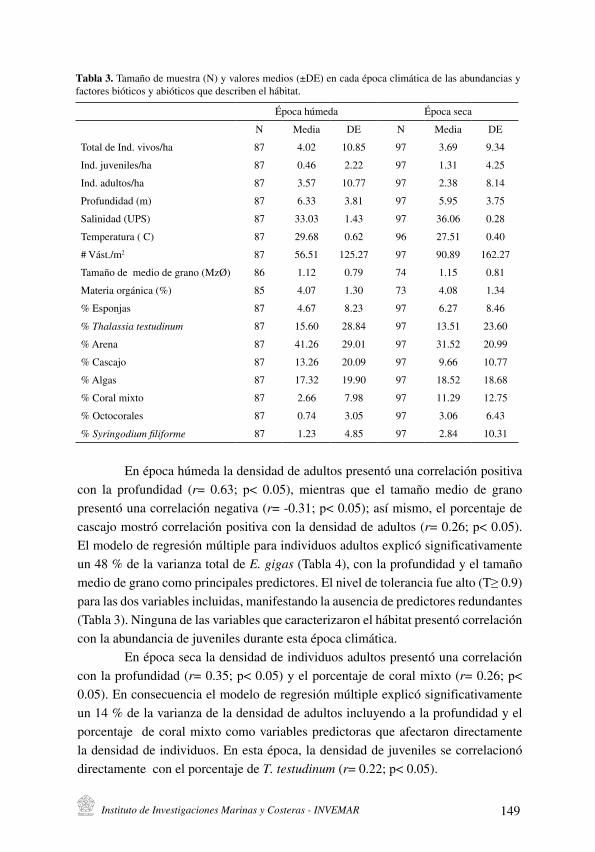
Instituto de Investigaciones Marinas y Costeras - INVEMAR 149
En época húmeda la densidad de adultos presentó una correlación positiva
con la profundidad (r= 0 .63; p< 0 .05), mientras que el tamaño medio de grano
presentó una correlación negativa (r= -0 .31; p< 0 .05); así mismo, el porcentaje de
cascajo mostró correlación positiva con la densidad de adultos (r= 0 .26; p< 0 .05) .
El modelo de regresión múltiple para individuos adultos explicó significativamente
un 48 % de la varianza total de E. gigas (Tabla 4), con la profundidad y el tamaño
medio de grano como principales predictores . El nivel de tolerancia fue alto (T≥ 0 .9)
para las dos variables incluidas, manifestando la ausencia de predictores redundantes
(Tabla 3) . Ninguna de las variables que caracterizaron el hábitat presentó correlación
con la abundancia de juveniles durante esta época climática .
En época seca la densidad de individuos adultos presentó una correlación
con la profundidad (r= 0 .35; p< 0 .05) y el porcentaje de coral mixto (r= 0 .26; p<
0 .05) . En consecuencia el modelo de regresión múltiple explicó significativamente
un 14 % de la varianza de la densidad de adultos incluyendo a la profundidad y el
porcentaje de coral mixto como variables predictoras que afectaron directamente
la densidad de individuos . En esta época, la densidad de juveniles se correlacionó
directamente con el porcentaje de T. testudinum (r= 0 .22; p< 0 .05) .
Época húmeda Época seca
N Media DE N Media DE
Total de Ind . vivos/ha 87 4 .02 10 .85 97 3 .69 9 .34
Ind . juveniles/ha 87 0 .46 2 .22 97 1 .31 4 .25
Ind . adultos/ha 87 3 .57 10 .77 97 2 .38 8 .14
Profundidad (m) 87 6 .33 3 .81 97 5 .95 3 .75
Salinidad (UPS) 87 33 .03 1 .43 97 36 .06 0 .28
Temperatura (°C) 87 29 .68 0 .62 96 27 .51 0 .40
# Vást ./m2 87 56 .51 125 .27 97 90 .89 162 .27
Tamaño de medio de grano (MzØ) 86 1 .12 0 .79 74 1 .15 0 .81
Materia orgánica (%) 85 4 .07 1 .30 73 4 .08 1 .34
% Esponjas 87 4 .67 8 .23 97 6 .27 8 .46
% Thalassia testudinum 87 15 .60 28 .84 97 13 .51 23 .60
% Arena 87 41 .26 29 .01 97 31 .52 20 .99
% Cascajo 87 13 .26 20 .09 97 9 .66 10 .77
% Algas 87 17 .32 19 .90 97 18 .52 18 .68
% Coral mixto 87 2 .66 7 .98 97 11 .29 12 .75
% Octocorales 87 0 .74 3 .05 97 3 .06 6 .43
% Syringodium filiforme 87 1 .23 4 .85 97 2 .84 10 .31
Tabla 3. Tamaño de muestra (N) y valores medios (±DE) en cada época climática de las abundancias y factores bióticos y abióticos que describen el hábitat .

150 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
DISCUSIÓN
DensidadEs difícil realizar comparaciones directas de registros de densidad con
otros estudios dadas las diferencias metodológicas en muestreo, ausencia parcial de
información y falencias en la clasificación de la estructura de la población (juveniles-
adultos) . Una evaluación discreta de algunos estudios realizados sugirió que existe
una gran variabilidad regional en las densidades de E. gigas (Tabla 5) . Valores medios
en el archipiélago NSR fueron siempre <5 ind/ha, muy cercanos y comparables a
los registrados en Bermuda, Florida, Panamá y Colombia (Quitasueño) . Es claro
cómo, a través de los años, las densidades medias de la población del caracol
disminuyen drásticamente . Es el caso de República Dominicana, donde tan sólo en
tres años las densidades pasaron de 188 a 15 ind/ha . La densidad más alta registrada
en este estudio -Bajo Tortugas, 24 ind/ha- representó 3 y 1 % de la densidad más
alta registrada en México (830 ind/ha; Martínez-Vásquez, 1995) y Venezuela (1886
ind/ha; Weil y Laughlin, 1984) . Sin embargo, un área protegida en Venezuela (Los
Roques) fue abruptamente afectada por la pesca ilegal de E. gigas, como ha ocurrido
con otras poblaciones de muchas regiones del Caribe (Appeldoorn, 1994) .
Tabla 4. Resultados del análisis de regresión múltiple para predecir la abundancia de adultos (ind/ha) de Eustrombus gigas en el archipiélago NSR durante las dos épocas climáticas . Se detalla la media del intercepto (±EE) y los coeficientes de regresión para las variables descriptoras del hábitat. CP: correlación parcial; T: tolerancia; r2aj: coeficiente de determinación ajustado; ns: variable removida (no significativa); N: tamaño de muestra . *p< 0 .1 . **p< 0 .05 . (--) variable no incluida en el análisis de regresión .
Época húmeda Época seca
(mayo – noviembre) (diciembre – abril)
Intercepto -5 .715 (2 .220) -2 .992 (1 .508)Profundidad 0 .650 (0 .082)** 0 .308 (0 .097)** CP 0 .661 0 .312 T 0 .932 0 .958Tamaño medio de grano -0 .141 (0 .082)* -- CP -0 .188 T 0 .932% Cascajo ns -- CP T% Coral mixto -- 0 .195 (0 .097)** CP 0 .204
T 0 .958r2 ajustado 0 .478 0 .139p << 0 .01 << 0 .01EE estimado 7 .263 7 .547N 81 94

Instituto de Investigaciones Marinas y Costeras - INVEMAR 151
Ubicación Año de muestreo ind/ha Referencia
Bahamas
Great Bahamas Bank
Área no protegida 1983 20.8 Smith y Neirop (1984)
Área protegida 1991-1994 53.6 Stoner y Ray-Culp (1996)
Bermuda 1988 0.5 Berg et al. (1992)
República Dominicana
del este 1996 187.5 Delgado et al. (1998)
del este 1999 15.0 Torres y Sullivan-Sealy (2000)
Florida Keys 1987-1988 2.4 Berg y Glazer (1995)
1990 1.5 Berg y Glazer (1995)
Honduras
Cayos Cochinos 1996 14.6 Tewfik et al. (1998)
Jamaica
Pedro Bank 1994 88.6 Tewfik(1996)
Pedro Bank 1997 314.0 TewfikyAppeldoorn(1998)
Panamá
Bocas del Toro 2000 1.4 TewfikyGuzmán(2003)
México
Cozumel
Área no protegida 1989 89.0 Martínez-Vásquez (1995)
Área protegida 1995 830.0 Martínez-Vásquez (1995)
Arrecife Alacranes
Parque Nacional 84.0 Pérez y Aldana (2000)
Banco Chinchorro 1992 26510.0 De Jesús-Navarrette et al. (2003)
1997 60.0 De Jesús-Navarrette et al. (2003)
Venezuela
Los Roques
Área no protegida 160.0 Weil y Laughlin (1984)
Área protegida 1886.0 Weil y Laughlin (1984)
Colombia
Quitasueño 1999 2.4 Valderrama y Hernández (1999)
Serrana 1999 285.0 Valderrama y Hernández (1999)
Roncador 1999 37.6 Valderrama y Hernández (1999)
Archipiélago NSR
Parque Nacional 2004 4.0 Este estudio
2005 3.7 Este estudio
Tabla 5 . Comparación de valores de densidad media de Eustrombus gigas en el Caribe, determinados por conteo visual en diferentes estudios .

152 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
El componente estructural dominante de la población de E. gigas en el
archipiélago NSR son los adultos . La baja densidad de juveniles en el archipiélago puede atribuirse a la intensa actividad pesquera en el área de hace unos años . La condición de sitio turístico estimuló hace varios años la intensa extracción de caracol para suplir la demanda que, sin tener en cuenta volúmenes de captura, tallas, ni áreas específicas de pesca, llevó a disminuir de forma importante el recurso (Mora, 1994) . Otro factor que pudo haber influenciado la baja densidad de juveniles es la urbanización del conjunto de islas que componen el archipiélago, lo que ha implicado a través de los años la destrucción de grandes extensiones de fondos compuestos principalmente por praderas de pastos (Alvarado, 1988), hábitat más importante para el desarrollo de los primeros años de vida de la especie . Por esta razón, es claro que los sectores menos intervenidos por actividades antrópicas, como bajo Tortugas e isla Tesoro (declaradas Áreas Intangibles, Decreto 622/77), puedan presentar las abundancias más altas de E. gigas .
La población de adultos en isla Tesoro y bajo Tortugas (individuos ≥de cuatro años de edad) podrían representar un grupo de donadores o parentales para el sistema y el futuro de las poblaciones en el área de estudio . No obstante, estudios sobre actividad reproductiva de adultos realizados en Bahamas (Stoner y Ray-Culp, 2000) evidenciaron el efecto ‘Allee’ en las poblaciones de E. gigas, es decir, la ocurrencia de tasas de crecimiento poblacional negativas promovidas por la baja probabilidad de encuentro para la cópula entre los especímenes . Estos autores mostraron que nunca ocurrieron encuentros reproductivos cuando las densidades poblacionales fueron <56 ind/ha . En el archipiélago NSR, durante el período comprendido entre julio y agosto (época reproductiva según Lagos et al ., 1996), se observó actividad reproductiva evidenciada en la presencia de masas de huevos, en dos áreas con una densidad poblacional de 24 ind/ha (bajo Tortugas) y 11 ind/ha (isla Tesoro) . Aunque en este estudio no se evaluó un comportamiento reproductivo denso-dependiente, es posible que si bien existen adultos disponibles para una recuperación de la población, la actividad reproductiva podría estar siendo afectada por la baja probabilidad de encuentros . Stoner y Ray-Culp (2000) proponen que a densidades tan bajas como las registradas en el archipiélago NSR la población podría no tener progenie y el reclutamiento dependería sólo de fuentes externas, como la retención de larvas y la sobrevivencia de juveniles en los hábitats de crianza apropiados .
Distribución espacialLa estimación de la abundancia total efectiva de E. gigas generó una visión
del estado de la población en el archipiélago NSR . Las estimaciones registradas en la literatura están basadas en una extrapolación de datos que no tiene en cuenta la distribución de la población ni el contexto espacial en el que habitan (Petitgas, 1996) .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 153
Tewfik et al . (1998) estimaron una abundancia total aproximada de 14 .6 ind/ha en Honduras (223818 individuos en un área total de 15330 ha), mientras que en este estudio la abundancia total estimada fue de 0.27 ind/ha en la época húmeda (936±11 individuos en 3041 ha) y 0.14 ind/ha durante la época seca (422±7 individuos en 3041 ha) . Estas abundancias tan bajas reflejan un indicio del severo decaimiento de la población en el área de estudio que enfatiza su inminente riesgo de extinción . El análisis variográfico reveló que la abundancia se estructuró espacialmente dentro de un ámbito de 11000 a 9000 m en las épocas húmeda y seca, respectivamente . La baja contribución del efecto pepita (<10 %) indicó que la escala de muestreo fue adecuada para medir el proceso espacial de la abundancia de E. gigas . Los tamaños de parche estimados en el archipiélago NSR son de gran magnitud considerando el área de estudio, lo cual sugiere que los individuos están muy alejados, generando además baja probabilidad de encuentros reproductivos .
Los mapas de distribución espacial de la densidad, reflejaron un patrón muy parecido entre épocas climáticas, con sectores específicos que representan altas densidades de individuos adultos (bajo Tortugas e isla Tesoro) . Entre los aspectos que pudieron ocasionar la concentración de individuos adultos en dichos sectores está su condición como áreas exclusivas por la lejanía con las islas de interés turístico y el acceso . Bajo Tortugas es un sector que promueve alta sobrevivencia de E. gigas, por su difícil accesibilidad tanto para pescadores como para el sector turístico y por sus mayores profundidades (>10 m). En el sector isla Tesoro ‘área intangible’, la vigilancia y control son estrictos y se aplica la restricción completa de ‘no pesca’, a diferencia de isla Rosario que a pesar de ser también un ‘área intangible’ no recibe la atención y vigilancia requerida de acuerdo a lo que reflejan los resultados y las actividades observadas durante el trabajo de campo de este estudio .
El grupo de juveniles son individuos que se presentan en el ambiente natural en agregaciones ocupando sectores específicos del hábitat . Según Stoner y Ray-Culp (1993), las agregaciones de juveniles en Bahamas, forman una elipse con un eje longitudinal paralelo al eje principal de la dirección de la corriente . A primera vista, la ausencia de autocorrelación espacial de juveniles en ambas épocas climáticas en este estudio debida a la baja abundancia imposibilitó identificar patrones de distribución para juveniles . Lo anterior fue corroborado al registrarse en ambas épocas climáticas el 75 % del área muestreada con densidades de cero . El 25 % restante se encontró en los sectores isla Grande, isla Rosario, isla Tesoro e isla Arena .
Relación entre la abundancia y variables que describen el hábitatEn el archipiélago NSR la profundidad es una variable que contribuye a
predecir la abundancia de individuos durante ambas épocas climáticas, con un modelo de regresión que explicó significativamente el 48 % de la varianza de densidad de

154 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
individuos adultos, valores que sugieren un buen modelo para explicar la variabilidad de la abundancia de E. gigas . Estos resultados coinciden con lo registrado por Alcolado (1976), en el sentido que la distribución de la especie está ligada con la profundidad . Este autor encontró que los caracoles adultos tienen una distribución batimétrica más fuerte, que se encuentran formando grupos hasta los 40 m de profundidad . Sin embargo, esta distribución de caracoles en aguas más profundas del archipiélago NSR puede ser atribuida a la presión pesquera (Bené y Tewfik, 2003) . En el mismo sentido, Stoner (1997) afirma que en lugares donde se pesca con equipo autónomo SCUBA (Puerto Rico) los caracoles se encuentran distribuidos a mayores profundidades (18-25 m), mientras que en lugares donde está prohibida la pesca con este tipo de equipo (Exuma Park, Bahamas), las mayores abundancias se registran a menores profundidades (10-15 m) . Por esta razón, teniendo en cuenta la pesca indiscriminada en el archipiélago NSR, la mayor cantidad de individuos se encontró en las mayores profundidades de difícil acceso .
Otra variable predictora de la densidad de E. gigas fue el tamaño medio de grano MzØ, cuyos valores más bajos corresponden a arenas muy gruesas y los más altos a arenas muy finas . Esta variable presentó una correlación con la abundancia de la especie, por lo tanto, donde se registraron arenas más gruesas (menores MzØ) se encontraron a su vez una mayor cantidad de individuos adultos . Según Randall (1964), el caracol pala hace parte del grupo de organismos herbívoros que habitan en los fondos marinos basando su alimentación en epífitos y afectando directamente la estructura de la comunidad bentónica . Así, es lógico pensar que los individuos prefieran arenas gruesas sobre las que pueden moverse para un pastoreo sin mucha dificultad, de esta manera funcionan ecológicamente como reguladores de la abundancia de detrito en su hábitat (Stoner et al ., 1995) . E. gigas se caracteriza por presentar su actividad reproductiva asociada a fondos de arenas gruesas y cascajo (Weil y Laughlin, 1984; Lagos et al ., 1996; Glazer y Kidney, 2004), como se observó en este estudio durante la época húmeda en bajo Tortugas e isla Tesoro . Es probable que los fondos semidesérticos sean los lugares óptimos para las actividades reproductivas de E. gigas, ya que presentan mayor facilidad de desplazamiento sumado a la necesidad de formar depresiones en la arena (D’Asaro, 1965) para la depositación de las masas de huevos y su posterior camuflaje mediante la adherencia de granos de arena .
La única variable que presentó correlación con la abundancia de caracoles juveniles, fue el porcentaje de cobertura de T. testudinum en el sustrato marino . Durante la época seca, se observó una mayor cantidad de juveniles asociados directamente a este tipo de hábitat . Estos organismos se caracterizan por alimentarse de macrodetritos, incluyendo partes muertas de hojas de pastos marinos (Stoner, 1989), lo que hace que dichos ambientes sean propicios para el ciclo de vida de

Instituto de Investigaciones Marinas y Costeras - INVEMAR 155
caracoles juveniles . Estos resultados concuerdan con lo registrado por varios autores en las islas Vírgenes y Bahamas, Gran Caribe (Randall, 1964; Stoner y Ray-Culp, 1993; Stoner y Schwarte, 1994), donde los juveniles de E. gigas están directamente asociados a parches de pastos marinos . Según Stoner et al . (1995), los juveniles se agregan durante cierta época del año en sectores específicos de praderas de pastos, donde la protección contra depredadores y la disponibilidad de alimento hace del hábitat un lugar seguro y propicio para el crecimiento . Es claro que la tasa de depredación disminuye con el aumento de la complejidad de la pradera de pastos (Ray y Stoner, 1994) . Sin embargo, parece ser que la escogencia de praderas densas por parte de individuos juveniles es un comportamiento adaptativo y la distribución puede ser consecuencia de efectos como la preferencia por detritos, presencia de ciertas algas y componentes del sedimento . El comportamiento de juveniles coincide con lo registrado por Stoner y Ray-Culp (1993) en Shark Rock (Bahamas), donde la distribución de caracoles juveniles en praderas de pastos marinos fue independiente a la densidad de vástagos, lo que sugiere que la escogencia de la pradera puede llegar a ser específica de un sitio .
Los resultados aquí expuestos permiten contribuir y apoyar la generación de medidas de manejo y conservación para E. gigas en el archipiélago NSR, por lo tanto se sugiere declarar sectores de protección especial: I) las áreas de desove de isla Tesoro y bajo Tortugas, y II) el sector de isla Arena, considerando que es un área potencial de crianza con predominancia de praderas de pastos, aislada, sin mayor intervención humana y donde se encontraron individuos juveniles; y/o usarla para programas de repoblación o trasplante . Cualquier medida debe estar basada en un sistema de manejo con control efectivo y que garantice la activa participación de la comunidad que reside en el área: nativos, pescadores, ocupantes y sector turístico .
AGRADECIMIENTOS
Este documento es parte del trabajo de grado de KGC . Los autores desean agradecer al Instituto Colombiano para el Desarrollo de la Ciencia y la Tecnología (COLCIENCIAS) por el apoyo financiero (proyecto código 2105-09-13528), al Instituto de Investigaciones Marinas y Costeras “José Benito Vives de Andréis” (INVEMAR), el Centro de Investigaciones, Educación y Recreación de las Islas del Rosario (CEINER) y la Unidad Administrativa Especial del Sistema de Parques Nacionales Naturales (UAESPNN) por el apoyo técnico y logístico . Al grupo de trabajo del proyecto por la asistencia, colaboración y apoyo incondicional en el trabajo de campo, en el desarrollo de la investigación y elaboración del documento .

156 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
BIBLIOGRAFÍA
Alcolado, P . 1976 . Crecimiento, variaciones morfológicas de la concha y algunos datos biológicos del
cobo Strombus gigas L . (Mollusca, Mesogastropoda) . Acad . Cien . Ser . Oceanol ., 34: 1-36 .
Alvarado, E . 1988 . El Parque Nacional Natural Corales del Rosario: situación actual . VI Seminario
Nacional de las Ciencias y Tecnologías del Mar . Comisión Colombiana del Oceano, Bogotá . 48 p .
Alvarado, E ., F . Duque, L . Flórez y R . Ramírez . 1986 . Evaluación cualitativa de los arrecifes coralinos
de las islas del Rosario (Cartagena–Colombia). Bol. Ecotrópica: Ecosistemas Tropicales, 15: 1-30.
Appeldoorn, R . 1988 . Age determination, growth, mortality and age of first reproduction in adult queen
conch, Strombus gigas L ., off Puerto Rico . Fish . Res ., 6: 363-378 .
Appeldoorn, R . 1994 . Spatial variability in the morphology of queen conch and its implications for
management regulations . 145-157 . En: Appeldoorn, R . y B . Rodríguez (Eds .) . Biología, pesquería y
cultivo del caracol Strombus gigas . Fundación Científica Los Roques, Caracas . 376 p .
Bené, C . y A . Tewfik . 2003 . Biological evaluation of marine protected area: evidence of crowding effect
on a protected population of queen conch in the Caribbean . Mar . Ecol ., 24 (1): 45-58 .
Berg, C . J . y R . Glazer . 1995 . Stock assessment of a large marine gastropod (Strombus gigas) using
randomized and stratified towed-diver censusing . ICES Mar . Sci . Symp ., 199: 247-258 .
Berg, C . J ., F . Couper . K . Nisbet y J . Ward . 1992 . Stock assessment of queen conch, Strombus gigas, and
harbour conch, S. costatus, in Bermuda . Proc . Gulf Carib . Fish . Inst ., 41: 433-438 .
Bohórquez, C . 2002 . Guía Parque Nacional Natural los Corales del Rosario y San Bernardo . Convenio
CEINER-UEASPNN-MINAMBIENTE, Cartagena . 44 p .
Burrough, P . y R . McDonnell . 1998 . Principles of geographical information systems . Oxford University
Press, Nueva York . 333 p .
Chiquillo, E ., J . Gallo y J . Ospina . 1997 . Aspectos biológicos del caracol pala, Strombus gigas Linnaeus,
1758 (Mollusca: Gastropoda: Strombidae), en el departamento archipiélago de San Andrés,
Providencia y Santa Catalina (Caribe colombiano) . Bol . Cient . INPA, 5: 159-179 .
CITES, 2003 . Convención sobre el comercio internacional de especies amenazadas de fauna y flora
silvestres . Examen del comercio significativo de especies del Apéndice II (resolución Conf . 12 .8 y
Decisión 12 .75) . Progresos realizados en la aplicación del examen de comercio significativo (fases
IV y V) . Decimonovena reunión del comité de fauna, Ginebra, Suiza . 73 p .
Cressie, N . 1993 . Statistics for spatial data . John Wiley & Sons, Nueva York . 900 p .
D´Asaro, C . 1965 . Organogenesis, development and metamorphosis in the queen conch Strombus gigas
with notes on breeding habits . Bull . Mar . Sci ., 15 (2): 359-416 .
De Jesús-Navarrette, A ., A . Medina-Quej y J . J . Oliva-Rivera . 2003 . Changes in the queen conch
(Strombus gigas l.) population structure at Banco Chinchorro, Quintana Roo, Mexico, 1990–1997.
Bull . Mar . Sci ., 73 (1): 219-229 .
Delgado, G . A ., M . Chiappone, F . X . Geraldes, E . Pugibet, K . M . Sullivan, R . E . Torres y M . Vega . 1998 .
Abundance and size frequency of queen conch in relation to benthic community structure in Parque
Nacional del Este, Dominican Republic . Proc . Gulf Carib . Fish . Inst ., 50: 1-31 .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 157
Díaz, J . M ., G . Díaz, J . Garzón, J . Geister, J . A . Sánchez y S . Zea . 1996 . Atlas de los arrecifes coralinos
del Caribe colombiano, I: complejos arrecifales oceánicos . Publicación Especial No . 2, Instituto de
Investigaciones Marinas y Costeras-INVEMAR, Santa Marta . 83 p .
Díaz, J . M ., L . Barrios, M . Cendales, J . Garzón, J . Geister, M . López, G . H . Ospina, F . Parra, J . Pinzón, B .
Vargas, F. A. Zapata y S. Zea. 2000. Áreas coralinas de Colombia. Serie de Publicaciones Especiales
No . 5, Instituto de Investigaciones Marinas y Costeras-INVEMAR, Santa Marta . 176 p .
Duque, F . 1974 . Estudio biológico-pesquero de Strombus (Tricornis) gigas L . (Mollusca, Gastropoda) en
el archipiélago de San Bernardo (Bolívar) . Trabajo de grado . Univ . Jorge Tadeo Lozano, Bogotá .
74 p .
García, M . I . 1991 . Biología y dinámica poblacional del caracol de pala Strombus gigas L . (Mollusca,
Mesogastropoda) en las diferentes áreas del archipiélago de San Andrés y Providencia . Trabajo de
grado . Univ . Valle, Cali . 183 p .
Genet, P . 1992 . Informe final proyecto: cartografía y fondos submarinos islas del Rosario (Colombia) y
apreciación de sus áreas a partir de imágenes de satélite Spot . Bol . Cient . CIOH, 11: 15-26 .
Glazer, R . y J . Kidney . 2004 . Habitat associations of adult queen conch (Strombus gigas L .) in an unfished
Florida Keys back reef: applications to essential fish habitat . Bull . Mar . Sci ., 75 (2): 205-224 .
Gómez-Campo, K . 2005 . Distribución espacial de la población de Strombus gigas Linnaeus, 1758
(Mollusca: Strombidae) y características del hábitat que la explican en el archipiélago Nuestra
Señora del Rosario, Caribe colombiano . Trabajo de grado, Univ . Jorge Tadeo Lozano, Santa Marta .
103 p .
INVEMAR, UAESPNN, CARDIQUE, EPA-CARTAGENA y DIMAR . 2003 . Elaboración de un modelo
de desarrollo sostenible para los archipiélagos de Nuestra Señora del Rosario y San Bernardo .
Informe técnico . Resolución del MAVDT número 456, Bogotá . 256 p .
Lagos, A . L . 1994 . Algunos aspectos biológicos y pesqueros del caracol pala Strombus gigas Linnaeus,
1758 . Trabajo de grado, Univ . Jorge Tadeo Lozano, Bogotá . 154 p .
Lagos, A ., S . Hernández, H . Rodríguez y P . Victoria . 1996 . Algunos aspectos bioecológicos y reproductivos
del caracol de pala Strombus gigas Linnaeus, 1758 en el archipiélago de San Bernardo, Caribe
colombiano . Bol . Cient . INPA, (4): 141-160 .
Legendre, P . 1993 . Spatial autocorrelation: trouble or paradigm? Ecology, 74: 1659-1673 .
Márquez, E . 1993 . Biología poblacional del caracol pala Strombus gigas Linnaeus, 1758 en las islas de
Providencia y Santa Catalina . Trabajo de grado, Univ . Jorge Tadeo Lozano, Bogotá . 154 p .
Martínez-Vásquez, D. 1995. ¿Qué pasa con las poblaciones de caracol en Cozumel? Informe CRIP,
Puerto Morelos, 2 (2), 15 p .
Matheron, G . 1971 . The theory of regionalized variables and its applications . Cahiers du CMM, facs . 5,
Ecole Nat . Sup . Mines, París . 211 p .
Mora, O . 1994 . Análisis de la pesquería del caracol pala (Strombus gigas L .) en Colombia . 137-144 . En:
Appeldoorn, R . y B . Rodríguez (Eds .) . Biología, pesquería y cultivo del caracol Strombus gigas .
Fundación Científica Los Roques, Caracas . 376 p .
Pérez, M . y D . Aldana . 2000 . Distribución, abundancia, densidad y mofometría de Strombus gigas
(Mesogasteropoda: Strombidae) en el arrecife Alacranes, Yucatán, México . Rev . Biol . Trop ., 48
(1): 51-57 .

158 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Petitgas, P . 1996 . Geostatistics and their applications to fisheries survey data . 113-142 . En: Megrey, B . y
E . Moksness (Eds .) . Computers in fishery research . Chapman y Hall, Londres . 142 p .
Petuch, E . J . y C . E . Roberts . 2007 . The geology of the Everglades and adjacent areas . CRC Press, Boca
Ratón, EE . UU . 212 p .
Randall, J . 1964 . Contributions to the biology of the queen conch Strombus gigas . Bull . Mar . Sci ., 14:
246-295 .
Ray, M . y A . Stoner . 1994 . Experimental analysis of growth and survivorship in a marine gastropod
aggregation: balancing growth with safety in numbers . Mar . Ecol . Prog . Ser ., 105: 47-59 .
Smith, G . B . y M . Neirop . 1984 . Distribution, abundance, and potential yield of shallow-water
fishery resource of the Little and Great Bahamas Banks: UNDP/FAO Fisheries Division Project
BHA/82/002 . Food and Agriculture Organization of the United Nations-FAO, Roma . 78 p .
Stoner, A . 1989 . Density-dependent growth and grazing effects of juvenile queen conch Strombus gigas
L . in a tropical seagrass meadow . J . Exp . Mar . Biol . Ecol ., 130: 119-133 .
Stoner, A . 1997 . The status of queen conch, Strombus gigas, research in the Caribbean . Mar . Fish . Rev .,
59 (13): 14-22 .
Stoner, A . 2003 . What constitutes essential nursery habitat for a marine species? A case of study of habitat
form and function for queen conch . Mar . Ecol . Prog . Ser ., 257: 275-289 .
Stoner, A . y R . Glazer . 1998 . Variation in natural mortality: implications for queen conch stock
enhancement . Bull . Mar . Sci ., 62 (2): 427-442 .
Stoner, A . y M . Ray-Culp . 1993 . Aggregation dynamics in juvenile queen conch (Strombus gigas):
population structure, mortality, growth and migration . Mar . Biol ., 116: 571-582 .
Stoner, A . y M . Ray-Culp . 1996 . Queen conch, Strombus gigas, in fished and unfished locations of the
Bahamas: effects of a marine fishery reserve on adults, juveniles, and larval production . Fish . Bull .,
94: 551-565 .
Stoner, A. y M. Ray-Culp. 2000. Evidence for ‘Allee Effects’ in an over-harvested marine gastropod:
density-dependent mating and egg production . Mar . Ecol . Prog . Ser ., 202: 297-302 .
Stoner, A . y K . Schwarte . 1994 . Queen conch, Strombus gigas, reproductive stocks in the central Bahamas:
distribution and probable sources . Fish . Bull ., 92: 171-179 .
Stoner, A ., M . Ray-Culp y M . Waite . 1995 . Effects of a large gastropod on macrofaunal communities in
tropical sea grass meadows . Mar . Ecol . Prog . Ser ., 121: 125-137 .
Tewfik, A . 1996 . An assessment of the biological characteristics, abundance and potential yield of the
queen conch (Strombus gigas) fishery on the Pedro Bank off Jamaica . Tesis M .Sc . Acadia University,
Wolfville, Canadá . 139 p .
Tewfik, A . y R . S . Appeldoorn . 1998 . Queen conch (Strombus gigas) abundance survey and potential
yield estimates for Pedro Bank, Jamaica . Fisheries Division, Jamaica . 31 p .
Tewfik, A . y H . Guzmán . 2003 . Shallow-water distribution and population characteristics of Strombus
gigas and S. costatus in Bocas del Toro, Panamá . J . Shelf . Res ., 22 (3): 789-794 .
Tewfik, A ., H . Guzmán y G . Jácome . 1998 . Assessment of the queen conch Strombus gigas (Gastropoda)
population in Cayos Cochinos, Honduras . Rev . Biol . Trop ., 46: 137-150 .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 159
Torres, R . E y K . M . Sullivan-Sealey . 2002 . Abundance, size frequency and spatial distribution of queen
conch (S. gigas) in south-eastern Dominican Republic: a four-year population study in Parque
Nacional del Este . Proceedings of the Gulf and Caribbean Fishery Institute, 53: 120-128 .
Valderrama, M . y S . Hernández . 1999 . Determinación de la abundancia del recurso caracol de pala
Strombus gigas en los bajos Serrana, Serranilla, Roncador y Quitasueño como base para la
definición de cuotas de pesca en el archipiélago de San Andrés y Providencia, Caribe colombiano .
INPA Subdirección de Investigaciones, Bogotá . 16 p .
Weil, E . y R . Laughlin . 1984 . Ecología, cultivo y repoblación del botuto Strombus gigas L . en el Parque
Nacional Archipiélago de los Roques . Fundación Científica Los Roques, Informe final, Caracas .
481 p .
Zar, J . 1996 . Biostatistical analysis, Tercera Edicion . Prentice-Hall, Inc ., New Jersey, 662 p .
FECHA DE RECEPCIÓN:14/10/2005 FECHA DE ACEPTACIÓN: 21/04/2010


Instituto de Investigaciones Marinas y Costeras - INVEMAR 161
Bol . Invest . Mar . Cost . 39 (1) ISSN 0122-9761 Santa Marta, Colombia, 2010
ESTADO DE LA POBLACIÓN Y VALORACIÓN DE ALGUNAS ESTRATEGIAS DE CONSERVACIÓN DEL RECURSO PIANGUA
ANADARA TUBERCULOSA (SOWERBY) EN SECTORES DE BAZÁN Y NERETE, COSTA PACÍFICA NARIÑENSE DE
COLOMBIA*
Silvana Espinosa G., María Fernanda Delgado Hernández, Bernardo Orobio Riofrío, Luz Marina Mejía-Ladino y Diego L. Gil-Agudelo
Instituto de Investigaciones Marinas y Costeras "José Benito Vives de Andreis"-INVEMAR. Cerro Punta de Betín, Santa Marta, Colombia. [email protected] (S.E.G.); [email protected] (L.M.M.L.); [email protected] (D.L.G.A.)
RESUMEN
Esta investigación tuvo como objetivo el conocer el estado de la población de piangua en dos lugares del departamento de Nariño (costa pacífica colombiana) y valorar estrategias para su conservación como el establecimiento de épocas de descanso de la actividad de extracción, rotación de áreas de concheo y control de la captura por debajo de la talla mínima de 5 cm reglamentada por el INPA (resolución 539 de 2000) . Para esto se establecieron parcelas permanentes donde se colectaron todas las pianguas utilizando la técnica manual tradicional, registrando su abundancia y datos morfométricos de cada individuo . Muestreos posteriores se realizaron a diferentes intervalos de tiempo según la estrategia a evaluar . Se colectaron 10441 individuos de A. tuberculosa, 61 % en Bazán y 39 % en Nerete . Las estrategias de conservación evaluadas mostraron efectos positivos en la conservación y aprovechamiento sostenible del recurso, siendo la época de descanso con un tiempo de dos meses entre colectas la práctica de conservación que dio mejores resultados (p= 0 .027 para Bazán y p= 8 .57E-06 para Nerete; Tukey: p< 0 .05); en Bazán únicamente esta estrategia arrojó resultados positivos mientras que en Nerete las tres estrategias de conservación mostraron ser eficientes .
PALABRAS CLAVE: Piangua, Anadara tuberculosa, Especie amenazada, Nariño, Pacífico colombiano .
ABSTRACT
Population status and valuation of some conservation strategies of piangua Anadara tuberculosa (Sowerby) in areas of Bazán and Nerete, Nariño, Pacific coast of Colombia. The objective of this investigation was to evaluate the status of piangua (Anadara tuberculosa, Sowerby) populations in two areas of the province of Nariño (Pacific coast of Colombia), and to assess the effect on natural communities of conservation strategies such as closed periods, rotation on extracting areas, and enforcement of the 5 cm minimum size declared by INPA (resolution 539, 2000) . Permanent plots were set up and all piangua were collected using the traditional manual techniques . Morphometric measurements of gathered organisms were recorded . Subsequent samplings were performed at different
161-176
*Contribución No. 1051 del Instituto de Investigaciones Marinas y Costeras – INVEMAR.

162 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
times depending on the conservation strategy evaluated . A total of 10441 individuals of A. tuberculosa were collected, 61 % corresponding to Bazán and 39 % to Nerete . All conservation strategies assessed showed positive effects on the conservation and sustainable use of the resource, being closed periods of two months the one with better results (p= 0 .027 for Bazán and p= 8 .57E-06 for Nerete; Tukey: p< 0 .05); in Bazán, this strategy was the only one showing positive results, while in Nerete the three strategies showed to be effective .
KEY WORDS: Piangua, Anadara tuberculosa, Threatened species, Nariño, Colombian Pacific .
INTRODUCCIÓN
Anadara tuberculosa (Sowerby) es un molusco bivalvo que se distribuye
en el Pacífico americano, desde Baja California hasta Perú . Posee varios nombres
comunes de acuerdo al lugar de procedencia tales como piangua, concha negra,
curil, chucheca, concha prieta y piangua hembra (Candelo, 2005) . Habita la zona
intermareal asociada a los sustratos fangosos, arcillosos o limo-arcillosos que
soportan las raíces de mangle (Camacho, 1999) . La piangua sostiene una de las
principales pesquerías artesanales de la zona costera en la región constituyendo
una importante fuente de proteína para las comunidades humanas del litoral, y sus
volúmenes de extracción han dependido tanto de las necesidades de abastecimiento
familiar como del comercio local . La deforestación del manglar y las altas tasas
de explotación han llevado a las poblaciones naturales de piangua a situaciones
críticas en países como El Salvador, Honduras y Ecuador . La colecta no regulada
constituye la principal amenaza a la especie en todos los países de la región donde es
fuertemente explotada (MacKenzie, 2001) .
En el Pacífico colombiano, A. tuberculosa está ligada al consumo tradicional
de las comunidades afrodescendientes que se asientan en las márgenes de los bosques
de manglar y es el principal recurso de la pesquería de moluscos, una actividad
de extracción exclusivamente artesanal efectuada principalmente por mujeres y
niños, aunque en los últimos años se ha presentado una gradual incorporación de
hombres que ven en la extracción de ese recurso una oportunidad para aumentar los
ingresos familiares . Aproximadamente 30000 familias de estratos económicos 1 y 2
se dedicaban a esta actividad económica en 2004 (WWF, 2005) .
Si bien ha existido un aprovechamiento tradicional de la piangua en la
región basado principalmente en el autoconsumo, la demanda comercial del recurso
ha aumentado en las últimas décadas, especialmente por parte del Ecuador . Esto ha

Instituto de Investigaciones Marinas y Costeras - INVEMAR 163
provocado una mayor e incontrolada captura de A. tuberculosa, especialmente en la
parte sur del país, estimándose que anualmente se extraen más de 300 millones de
unidades solamente en los siete municipios costeros de Nariño (WWF, 2005) .
Estadísticas de pesca puntuales indican que en la década de los años 70
del siglo pasado, las capturas eran de aproximadamente 100 ton/año (Artunduaga y
Mora, 1975), alcanzando para el 2004, 3283 ton/año en el área de Tumaco (Borda y
Cruz, 2004a) . Este importante aumento en la explotación plantea una problemática
asociada a la oferta natural del recurso, a la pesquería y a las comunidades que
obtienen su sustento diario de esa actividad (Espinosa et al., 2007) .
González (2006) mostró una reducción en la captura de 800 individuos/
persona/faena entre 1960 y 1970, a sólo 140 individuos/persona/faena en 2005 . Por
éstos y otros indicios de disminución en sus poblaciones, A. tuberculosa fue incluida
en el Libro Rojo de Invertebrados Marinos de Colombia como especie amenazada
en la categoría Vulnerable (Ardila et al ., 2002) .
Las principales iniciativas planteadas para aportar soluciones al problema
de la disminución del recurso piangua en la zona costera del departamento de Nariño
han partido de las comunidades de piangüeras directamente afectadas, quienes
han diseñado tres estrategias de conservación: 1) establecer épocas de descanso
de la actividad de extracción, 2) rotar áreas de concheo, y 3) evitar la captura por
debajo de la talla mínima de 5 cm propuesta por el Instituto Nacional para la Pesca
y la Acuicultura (INPA), en la resolución 539 de 2000 . El objetivo de la presente
investigación fue conocer el estado de las poblaciones de piangua, en cuanto a su
abundancia y tallas, en los lugares de estudio, y valorar estas estrategias locales para
la conservación del recurso a partir de su influencia en la abundancia .
ÁREA DE ESTUDIO
La costa pacífica nariñense de Colombia posee aproximadamente 50 % de
la extensión de manglares del país, equivalente a 117576 hectáreas, de las cuales
80000 están declaradas como área protegida (Parque Nacional Natural Sanquianga)
(INVEMAR et al., 2006; MAVDT et al., 2007) . En algunas zonas los manglares
ocupan franjas que alcanzan hasta 20 km de ancho en su máximo desarrollo y biomasa
forestal, alcanzando alturas de 30 m y raíces fúlcreas de 5 m . Se seleccionaron dos
áreas de muestreo: la vereda Bazán, municipio de El Charco, en jurisdicción del
PNN Sanquianga; y la vereda Nerete, en el municipio de Tumaco (Figura 1) .

164 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
MATERIALES Y MÉTODOS
En cada área se realizó un muestreo exploratorio para definir los esteros en los cuales se ubicarían las parcelas a evaluar . Se establecieron ocho parcelas, tres para la estrategia Época de descanso, cuatro para la estrategia Rotación de áreas, y una para Talla mínima . En cada una de ellas se demarcaron mediante cinta reflectiva (sin cerramiento de ningún tipo), tres unidades muestreales de 10 x 10 m paralelas a la línea de costa, distanciadas 10 m una de otra .
En el muestreo inicial se extrajeron las pianguas presentes en las unidades muestreales . Luego, en cada uno de los tiempos designados para evaluar cada estrategia de conservación, se regresó a las unidades muestreales y se extrajeron los ejemplares presentes . Las estrategias de conservación fueron evaluadas a diferentes tiempos así:
Figura 1. Área de estudio en la costa nariñense del Pacífico colombiano. La cobertura del manglar aparece sombreada .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 165
Época de descanso a los meses 2, 4 y 6; Rotación de áreas a las semanas 1, 2, 3 y 4; y Talla mínima a los meses 2 y 4; en este último caso, se devolvieron los ejemplares con talla por debajo de los 5 cm . Todos los tratamientos fueron evaluados entre octubre de 2006 y abril de 2007 . Como referencia al comportamiento del recurso sin aplicar ninguna medida de conservación (control), se evaluaron tres unidades muestreales en una parcela en una misma localidad, tanto en el área de Bazán como de Nerete, en las cuales se hacía extracción semanal de todos los individuos . Las colectas fueron realizadas por dos piangüeras mediante la técnica manual tradicional durante marea baja. Las pianguas se contaron y midieron con un calibrador (precisión 0 .01 mm) para determinar su longitud (distancia antero posterior de la concha), altura (máxima distancia entre las superficies externas de las dos valvas) y ancho (distancia dorso ventral del ápice) .
Se empleó estadística descriptiva (promedio, desviación estándar, valores mínimos y máximos, entre otros) para presentar las generalidades de las poblaciones de piangua evaluadas en cada lugar, resaltando la abundancia relativa (expresada en porcentaje), densidad (número de pianguas por metro cuadrado) y clases de tallas de las poblaciones . El contraste entre el control, los tratamientos (estrategias de conservación) y niveles (intervalos de tiempo), se hizo restando la abundancia final de la abundancia inicial, interpretándose un mayor valor negativo como una mayor eficiencia de la estrategia de conservación . La normalidad de las diferencias fue probada con la corrección de Lilliefors (Conover, 1980) y la homogeneidad de varianzas con la prueba de Levene .
Para determinar el efecto de los tratamientos se hizo un análisis de varianza (ANOVA a una vía) y a través de la Prueba Tukey (Zar, 1996) se establecieron los niveles en los cuales se da esa diferencia . Se usó el paquete estadístico R versión 2 .5 .0 y el complementario NORTEST, de libre distribución (http://cran .r-project .org, descargado en 2007) .
RESULTADOS
Abundancia y tallasEn total se evaluaron 10441 individuos de A. tuberculosa, la mayor parte
(61 %) en Bazán . Las tallas promedio encontradas fueron de 4 .36 cm para Bazán y
4 .23 cm para Nerete, mientras que la mayor parte de la población se encontró en el
ámbito de 3 .5 a 4 .9 cm (65 % para Bazán y 72 % para Nerete) . El ámbito de tallas
con mayor abundancia se encontró entre 4 .0 y 4 .4 cm de longitud: 24 % en Bazán y
30 % en Nerete (Figura 2) .
La longitud máxima encontrada en Nerete fue de 6 .8 cm y la mínima de
0 .9 cm . De los especímenes colectados en Bazán, uno de ellos tuvo la mayor talla

166 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
del presente estudio, 7 .8 cm . La mayoría de los individuos colectados tanto en
Bazán (76 %) como en Nerete (85 %) se encontraron por debajo de la talla mínima
reglamentaria de 5 .0 cm, según la Resolución 0539 de 2000, expedida por el INPA
(Figura 2) .
DensidadLos datos de densidad obtenidos fueron variables tanto entre localidades
como entre los esteros donde se ubicaron las parcelas . El estero La Cagada (1 .02 ind
m-2 ± 0.50 DE) en Bazán y el conocido como Nerete (0.93 ± 0.10 ind m-2) en Nerete,
fueron los que presentaron las mayores densidades registradas (Tabla 1) . En general,
los valores promedio de densidad no superaron 1 .00 ind m-2 .
Estrategias de conservaciónLa extracción de ejemplares en las parcelas control de cada área evaluada,
ocasionó una disminución rápida del recurso en las primeras tres semanas sin un
agotamiento total, pero manteniendo un número bajo de individuos frente a la
extracción permanente [en promedio 28 pianguas por 100 m2 para Bazán (Figura 3a)
y 22 pianguas por 100 m2 para Nerete (Figura 3b)] .
Comparando el comportamiento de los controles con las diferentes
estrategias de conservación, se encontraron diferencias significativas en el efecto de
las estrategias de conservación en la abundancia entre las dos áreas de estudio (p=
0 .0017) (Tabla 2a, Figura 4) . Igualmente todas las interacciones entre las localidades
y las estrategias tienen influencia en la diferencia de abundancias: época de descanso
Figura 2. Abundancia relativa (expresada en porcentaje) de Anadara tuberculosa diferenciada en ocho clases de tallas entre 2 .0 y 5 .9 cm en las veredas Bazán (n= 6249) y Nerete (n= 4192) del Pacífico nariñense . La línea indica la talla mínima de captura, reglamentada en 5 cm para Colombia . No se incluyen los valores extremos de talla .
2
5
12
2124
20
11
4
02
11
20
30
3
22
12
0
5
10
15
20
25
30
2.0 – 2.4 2.5 – 2.9 3.0 – 3.4 3.5 – 3.9 4.0 – 4.4 4.5 – 4.9 5.0 – 5.4 5.5 – 5.9
Clases de talla (cm)
Abu
ndan
cia
rela
tiva
(exp
resa
da en
por
cent
aje)
Bazán Nerete

Instituto de Investigaciones Marinas y Costeras - INVEMAR 167
Tabla 1. Densidad de Anadara tuberculosaenlosesterosdelasveredasBazányNeretedelPacíficonariñense (n: número de unidades muestreales, DE: desviación estándar).
Localidad Estero Longitud W
Latitud N
Densidad ( ind m-2) n DE
Bazán La Cagada 72º08’34” 02º40’25” 1 .02 3 0 .50
El Muertero (oriental)
78º09’43” 02º40’32” 0 .85 9 0 .44
El Muertero (occidental)
Sin Datos Sin Datos 1 .01 12 0 .50
La Correntosa 78º08’41” 02º40’32” 0 .33 3 0 .10
Nerete Nerete 78º40’13” 01º42’50” 0 .93 9 0 .41
Los Cabezas Sin Datos Sin Datos 0 .47 12 0 .32
Tulicio 78º40’25” 01º43’0” 0 .49 3 0 .09
X~
(p= 0 .010), rotación de áreas (p= 0 .017) y talla mínima (p= 0 .024) (Tabla 2a) . Para
las localidades se observó que en Bazán la única estrategia que parece tener un
efecto positivo en la diferencia de abundancias fue la época de descanso (p= 0 .027)
mientras que en Nerete las tres estrategias de conservación tuvieron influencia en la
diferencia de las abundancias respecto al control (p< 0 .005) (Tabla 2b y c) .
Asimismo, la época de descanso con un tiempo de dos meses entre colectas
fue la estrategia de conservación que mostró un mayor incremento de abundancia
de piangua, al final de los tratamientos en las dos localidades estudiadas (Tukey:
p< 0 .05) (Figura 4b) . Al cuarto mes de descanso se presentaron diferencias entre
las localidades: en Bazán hubo aumento del número de individuos (Figura 5a) pero
en Nerete disminuyeron (Figura 5b); mientras que en el descanso de seis meses se
presentó un menor número en ambas localidades .
En la estrategia de conservación con control de captura de la talla mínima
hubo una mayor abundancia de individuos en colectas realizadas en ambas
localidades para el tiempo de descanso de cuatro meses (Figura 4d) . Sin embargo,
esta estrategia no pareció ser un factor influyente en la diferencia de abundancia de
individuos de piangua en Bazán (p= 0 .061) pero sí en Nerete (p= 0 .0006); en esta
última localidad, la mayor diferencia en las abundancias relativas se obtuvo para el
tiempo de descanso de cuatro meses (Tukey: p< 0 .05) . La estrategia de rotación de
áreas es influyente en la diferencia de abundancias en Nerete (p= 0 .004) cuando se
tenía un tiempo de descanso de cuatro semanas .

168 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Tabla 2. Análisis de varianza para las diferencias de abundancias con: a) datos totales; b) diferenciando por localidad: Bazán y Nerete .
Grados de libertad
Suma de cuadrados
Cuadrados medios Valor F p
Locación 1 11102 11102 10 .712 0 .0016648
Época de descanso 3 12549 4183 4 .0359 0 .0105228
Rotación de áreas 4 13368 3342 3 .2246 0 .017408
Talla mínima 2 8107 4053 3 .9111 0 .0246045
Grados de libertad
Suma de cuadrados
Cuadrados medios Valor F p
Época de descanso 3 14264 4755 3 .3931 0 .02756
Rotación de áreas 4 9102 2276 1 .624 0 .18818
Talla mínima 2 8398 4199 2 .9967 0 .06188
Grados de libertad
Suma de cuadrados
Cuadrados medios Valor F p
Época de descanso 3 23769 .7 7923 .2 13 .4474 8 .57 E-06
Rotación de áreas 4 11283 .9 2821 4 .7878 0 .0039975
Talla mínima 2 11152 .3 5576 .1 9 .4639 0 .0006192
Tabla 2a.
Tabla 2b. Bazán
Tabla 2b. Nerete
Abu
ndan
cia
(núm
ero
de in
divi
duos
)
0
20
40
60
80
100
120
140
160
180
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
Semanas
Abu
ndan
cia
(núm
ero
de in
divi
duos
)
0
20
40
60
80
100
120
140
160
180
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
Semanas
a
b
Figura 3. Abundancias promedio de Anadara tuberculosa en la parcela control: a) Bazán (n= 1390) y b) Nerete (n= 791) .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 169
Figura 4. Diagramas de caja (mediana, cuartiles del 25 y 75 %, valores máximo y mínimo, y valores extremos) que muestran las diferencias de abundancia de Anadara tuberculosa al aplicar las estrategias de conservación estudiadas (los valores negativos indican que la abundancia final fue mayor que la inicial) . Se compara con la parcela control .

170 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
DISCUSIÓN
En el presente estudio los valores de densidad obtenidos variaron entre 0 .33 y 1 .02 ind m-2; trabajos de las décadas de los años 70 y 80 registran densidades entre 1 .0 y 3 .0 ind m-2 en distintos sitios de la costa pacífica colombiana (Tabla 3), mientras que investigaciones más recientes en Colombia presentan cifras similares a las encontradas en el presente estudio . A pesar de que las densidades de piangua son variables entre localidades (Borda y Cruz, 2004b), los resultados muestran que la condición de amenaza sigue presentándose en el departamento de Nariño debido a la disminución histórica de sus poblaciones . En otros países, como Costa Rica y Ecuador donde también se presentan importantes capturas, los ámbitos de abundancia registrados son más amplios (Tabla 3) . Así se confirma el estado de amenaza de la especie A. tuberculosa en algunos sectores de Bazán y Nerete en el departamento de Nariño .
0
50
100
150
200
250
300
350
400
450
Mes 2 Mes 4 Mes 6
Abu
ndac
ia(n
úmer
o de
indi
vidu
os)
Abundancia Inicial Abundancia Final
0
50
100
150
200
250
300
350
400
450
Mes 2 Mes 4 Mes 6
Abu
ndac
ia(n
úmer
o de
indi
vidu
os)
Abundancia Inicial Abundancia Final
Figura 5. Abundancias relativas totales de Anadara tuberculosa en la parcela Época de descanso: a) Bazán (n= 1810) y b) Nerete (n= 1422) .
a
b

Instituto de Investigaciones Marinas y Costeras - INVEMAR 171
Departamento y/o País Localidad Valor medido Autor
Densidad relativa (ind m-2)
Chocó, ColombiaEnsenada de Utría, Jurubidá y Chorí
0 .4-3 .8 Franco (1995)
Valle del Cauca, Colombia
Bahía Málaga 0 .5-0 .8 Lucero (2007)
Buenaventura 1 .0-4 .0 Squires et al . (1975)
Punta Soldado 1 .0-3 .0 Bolívar (1984)
Nariño, Colombia Salahonda 1 .0-3 .0 Haskoning (1986)
Bazán 0 .3-1 .0 Presente estudio
Nerete 0 .5-0 .9 Presente estudio
Costa Rica Sierpe-Térraba 1 .0 Campos et al . (1990)
Costa Rica Golfo Dulce 0 .2-0 .9 Silva y Bonilla (2001)
Ecuador 0 .5-2 .0 Mora y Moreno (2007)
Longitud (cm)
Nariño, Colombia Bazán Mínima: 1 .6 Promedio: 4 .4 Máxima: 7 .8
Presente estudio
Nerete Mínima: 0 .9 Promedio: 4 .2 Máxima: 6 .8
Presente estudio
PNN Sanquianga Promedio: 5 .0-5 .4 Puentes y Celis (1997)
Bazán Máxima: 9 .0 Gutiérrez et al. (1998)
Tumaco Promedio: 4 .8-5 .6 Ardila (1989)
Tumaco Máxima: 7 .5 González (2006)
Valle del Cauca, Colombia
Bahía MálagaMínima: 1 .9 Promedio: 4 .6 Máxima: 10 .0
Lucero (2007)
Costa Rica Golfo Dulce Promedio: 4 .3 Silva y Bonilla (2001)
Pacífico Oriental Tropical Máxima: 7 .5 MacKenzie (2001)
Tabla 3. Valores de densidad relativa (ind m-2) y longitud (cm) de Anadara tuberculosa en diferentes localidades del Pacífico colombiano y otros países (valor preciso disminuyendo a una cifra decimal) .
En cuanto a la talla media de captura, Borda (2001) encuentra un valor de 4 .76 cm y porcentajes de captura por talla de 20 .1 % (4 .0 y 4 .5 cm), 8 .9 % (entre los 2 .5 y 3 .0 cm) y 0 .5 % (6 .5 y 7 .0 cm) . En el presente trabajo, el porcentaje de individuos de la población natural que se obtuvo por debajo de la talla reglamentaria de 5 cm, correspondiente a 85 %, es mayor al indicado por Borda y Cruz (2004a) en los sitios de acopio, donde aproximadamente el 55 % de la piangua comercializada está por debajo de esta cifra . La mayoría de los estudios que registran tallas de A. tuberculosa se ha realizado a partir de individuos obtenidos en los centros de

172 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
acopio, favoreciendo el registro de pianguas de tallas grandes mientras que las tallas medianas o pequeñas se ven subrepresentadas debido a que no son colectadas o son separadas para autoconsumo y no llegan al centro de acopio . El presente estudio muestra resultados in situ, reflejando una mejor estimación de la longitud promedio y la abundancia de los diferentes ámbitos de talla (Tabla 3) .
Gutiérrez et al. (1998), en un estudio sobre el impacto del canal Naranjo en la zona de Bazán, registraron pianguas de hasta 9 cm de longitud en el período 1997-1998, valores claramente más altos que los encontrados en este estudio (7 .8 cm) . Lo anterior evidencia una posible disminución en los individuos de tallas grandes en las poblaciones . Los valores de talla máxima encontrados en este estudio en la localidad de Bazán son más cercanos al de 7 .5 cm que registra MacKenzie (2001) para el Pacífico Oriental Tropical, que los encontrados en Nerete (Tabla 3) .
El comportamiento de la parcela control muestra la dinámica del recurso cuando la extracción se hace en una misma área de manera permanente; así, se observó una disminución progresiva del número de individuos como se encontró en este estudio (Figura 3) . Por el contrario, al valorar las estrategias de conservación propuestas, se puede observar un efecto positivo sobre la abundancia del recurso, especialmente en su dinámica frente a tiempos de descanso mensual de las áreas de extracción y control de talla mínima de captura . Al dejar de extraer el recurso aparecen otros individuos con el tiempo y aumenta su abundancia en los lugares donde en un principio se habían extraído . La manera distinta como inciden las estrategias de conservación en la población de piangua entre Bazán y Nerete (en Nerete las tres estrategias influyen, en Bazán sólo la época de descanso) puede ser explicada en parte, por la distribución del recurso y la influencia de las condiciones ambientales . El área de cobertura del manglar, las especies de manglar dominantes y ciertos parámetros físico-químicos como el pH, son algunas de las variables no controladas que pueden estar influyendo en el comportamiento de las estrategias de conservación en cada una de las localidades estudiadas . Por ejemplo, la mayor disponibilidad de manglar en Bazán, localidad que hace parte del Parque Nacional Natural Sanquianga que posee la mayor extensión de manglar de la costa pacífica colombiana, calculada en 80000 hectáreas, puede favorecer la recuperación de la abundancia de A. tuberculosa al ofrecer más sitios disponibles para colonizar . Sin embargo, la información es insuficiente ya que otros factores pueden afectar esa dinámica . Al respecto, Naranjo (1982) explica que la distribución de A. tuberculosa parece estar determinada principalmente por el tipo de sustrato y el nivel de las mareas .
Si bien la época de descanso se evaluó en tres tiempos, los mayores valores de abundancia se obtuvieron, contrario a lo esperado, a los dos meses de descanso en ambas localidades . Se presumía que al dejar el área más tiempo sin colectar, se obtendría una mayor abundancia . Al respecto, Caddy y Defeo (2003) muestran

Instituto de Investigaciones Marinas y Costeras - INVEMAR 173
evidencias de que cierres extendidos de explotación de moluscos pueden inhibir el reclutamiento de nuevos individuos y así el crecimiento de la población en lugares con altas densidades de adultos, sin embargo se considera que en este caso el tiempo de descanso de la colecta fue corto para obtener este tipo de conclusiones . Así, los picos de reproducción de la especie, la tasa de mortalidad y los movimientos de las larvas pueden ser factores influyentes en un estudio temporal, por lo tanto se sugiere realizar posteriores trabajos que permitan corroborar esas hipótesis y su relación con el tiempo de descanso .
Con relación a las medidas de manejo, algunos países han reglamentado vedas y tallas mínimas de captura . Actualmente las regulaciones en Costa Rica establecieron una talla mínima de captura (TMC) de 4 .7 cm (Decreto 13371-A, expedido por el Instituto Costarricense de Pesca y Acuacultura) (Silva y Bonilla, 2001) y 15 años atrás, la TMC era de 4 .4 cm para ese mismo país (Campos et al ., 1990) . En el Perú y El Salvador se estableció la TMC en 4 .5 cm (Borda y Cruz, 2004a) . Con el presente estudio, la combinación del descanso bimensual de la colecta y el control de la talla mínima de 5 cm establecida en Colombia, incidió de manera positiva en la población de A. tuberculosa para las dos localidades, y pueden ser las estrategias de conservación más viables para implementar y alcanzar un manejo adecuado de ese recurso . Sin embargo, es necesario complementar la información con otros estudios poblacionales de A. tuberculosa, además de la dinámica espacial de las poblaciones, sus tasas de natalidad, reclutamiento y mortalidad, que pueden explicar otras dimensiones de las poblaciones de piangua . Igualmente, hay que avanzar en el entendimiento de su comportamiento frente a las estrategias de conservación, así como de las particularidades del entorno, para sustentar las diferencias y contribuir con información base para el manejo de este recurso biológico y la pesquería que sustenta .
La identificación de estrategias de conservación para la piangua es de gran importancia debido a los altos volúmenes de explotación actuales y a la gran dependencia del recurso que poseen comunidades afrodescendientes del Pacífico colombiano . Los resultados obtenidos en el presente estudio pueden servir de guía para la aplicación de estas estrategias en otros lugares de la costa pacífica colombiana, pero es necesaria la determinación de su viabilidad en cada uno de los sitios en que se deseen ensayar . Igualmente, sumarse a las iniciativas que desde la dinámica social y de los acuerdos colectivos de manejo se llevan a cabo, por parte de las piangueras mismas, con su disposición de cumplir reglas y normas, y realizar acciones para su conservación (Cárdenas et al., 2002, 2003; Cárdenas, 2004, 2005; Espinosa et al., 2009) y las promovidas por entidades como la WWF (Candelo, 2005), Ministerio del Ambiente, Vivienda y Desarrollo Territorial, Unidad Administrativa Especial del Sistema de Parques Nacionales y Fundación Chonapi, entre otras .

174 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
AGRADECIMIENTOS
Este trabajo fue realizado en el marco de los proyectos “Algunas estrategias
de conservación de la piangua Anadara tuberculosa (especie amenazada según
categoría de la UICN) en la costa pacífica nariñense de Colombia, mediante la
articulación de los saberes tradicional y científico”, financiado por COLCIENCIAS,
Código 2105-10-17933, Contrato 246-2005 y “Potencial productivo de las
poblaciones naturales de la piangua Anadara tuberculosa y Anadara similis dentro
de una perspectiva espacio-temporal en la costa pacífica colombiana”, financiado
por Ministerio de Agricultura y Desarrollo Rural, contrato: 161-2008T6344-3459 y
con el apoyo del BPIN PNIBM (años 2008 y 2009) y BEM (año 2010) . Los autores
agradecen a la Asociación de Concheras de Nariño, Asconar y la Fundación Chonapi,
por el apoyo en la coordinación de las actividades con las piangüeras. También a la
Alcaldía del municipio de El Charco y la oficina de Incoder de Tumaco, las cuales
colaboraron con la logística de las salidas de campo . A X . Zorrilla (UAESPNN), G .
Arteaga (Corponariño), J . Cantera (Univalle), J . A . López-Cerezo (Universidad de
Oviedo, España) y G . Navas (INVEMAR) por sus valiosos aportes en el desarrollo
del proyecto y mejoras al manuscrito .
BIBLIOGRAFÍA
Ardila, C . 1989 . Diagnóstico de la pesca artesanal de moluscos en algunas regiones del Pacífico
colombiano . Tesis de grado, Univ . Valle, Cali . 140 p .
Ardila, N . E ., G . R . Navas y J . O . Reyes (Eds .) . 2002 . Libro rojo de invertebrados marinos de Colombia .
Serie Libros Rojos de Especies Amenazadas de Colombia . Instituto de Investigación de Recursos
Biológicos Alexander Von Humboldt y Ministerio del Medio Ambiente, Bogotá . 136 p .
Artunduaga, E . y O . Mora . 1975 . Diagnóstico sobre la pesca industrial y artesanal en el Pacífico colombiano .
Divulg . Pesq ., 5: 37-60 .
Bolívar, G . 1984 . Evaluación de la oferta de semilla de Anadara spp . en el estero Veneno, bahía de
Buenaventura, Pacífico colombiano . Informe técnico, Univ . Valle, Cali . 8 p .
Borda, C . 2001 . Evaluación pesquera de la piangua (Anadara tuberculosa) en el Pacífico colombiano . Tesis
M . Sc ., Univ . La Habana, La Habana . 74 p .
Borda, C . y R . Cruz . 2004a . Pesca artesanal de bivalvos (Anadara tuberculosa y A. similis) y su relación con
eventos ambientales del Pacífico colombiano . Rev . Invest . Mar ., 25 (3): 197-208 .
Borda, C . y R . Cruz . 2004b . Reproducción y reclutamiento del molusco Anadara tuberculosa (Sowerby,
1833) en el Pacífico colombiano . Rev . Invest . Mar ., 25 (3): 185-195 .
Caddy, J . F . y O . Defeo . Enhancing or restoring the productivity of natural populations of shellfish and other
marine invertebrate resources . FAO Fish . Tech . Paper, 448, Roma . 159 p .
Camacho, Y . 1999 . Especies de Costa Rica . Anadara tuberculosa . Instituto Nacional para la Biodiversidad,
San José . 3 p .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 175
Campos, J ., M . Fournier, R . Soto y R . Arrion . 1990 . Estimación de la población de Anadara tuberculosa
(Bivalvia: Arcidae) en Sierpe-Térraba, Costa Rica . Rev . Biol . Trop ., 38 (1): 477-480 .
Candelo, C . 2005 . Fortalecimiento de capacidades: oportunidad para el cambio social y la experiencia de
las piangüeras en Nariño (Colombia). Sem. Internal. “Enfoques y Perspectivas de la Enseñanza del
Desarrollo Rural”. Pont. Univ. Javeriana, Bogotá. 16 p.
Cárdenas, J . C . 2004 . Norms from outside and from inside: An experimental analysis on the governance of
local ecosystems . Forest Pol . Econ ., 6: 229-241 .
Cárdenas, J . C . 2005 . Social preferences among the people of Sanquianga in Colombia . Univ . Los Andes,
Bogotá . 35 p .
Cárdenas, J ., D . Maya y M . López . 2002 . Manglares, comunidad y cooperación . Pont . Univ . Javeriana,
Fondo Mundial para la Naturaleza, WWF, Cali . 28 p .
Cárdenas, J ., D . Maya y M . López . 2003 . Métodos experimentales y participativos para el análisis de la
acción colectiva y la cooperación en el uso de recursos naturales por parte de comunidades rurales .
Cuad . Des . Rur . Pont . Univ . Javeriana, Bogotá . 200 p .
Conover, W . J . 1980, Practical nonparametric statistics . Segunda edición, John Wiley & Sons, Nueva York .
297 p .
Espinosa, S ., L . A . Zapata, C . Candelo, M . X . Zorrilla, G . Arteaga, M . F . Delgado, B . Orobio y D . L . Gil-
Agudelo . 2007 . Diagnóstico de especies amenazadas: Estado actual de las poblaciones de piangua
Anadara tuberculosa en la costa pacífica colombiana . 261-268 . En: INVEMAR (Ed .) . Informe
del Estado de los Ambientes Marinos y Costeros en Colombia: Año 2006 . Serie de Publicaciones
Periódicas No . 8, Instituto de Investigaciones Marinas y Costeras-INVEMAR, Santa Marta . 378 p .
Espinosa, S ., D . L . Gil-Agudelo, C . Candelo y L . A . Zapata . 2009 . Las piangueras en la costa pacífica
colombiana: investigación participativa para la conservación de la piangua y la actividad económica
de este recurso biológico . 159-167 . En: Vicepresidencia de la República, Comisión Colombiana del
Océano y Observatorio del Pacífico Colombiano (Eds .) . Los pueblos del mar . Bogotá . 167 p .
Franco, C . 1995 . Distribución, aspectos ecológicos y estado actual de la piangua, Anadara similis y Anadara
tuberculosa y observaciones ecológicas sobre la sángara, Anadara grandis en la ensenada de Utría y
Parque Nacional Natural Utría . Informe final, Fundación para la Promoción de la Investigación y la
Tecnología, Bogotá . 42 p .
González, J . 2006 . Reglas de manejo, percepciones y estado del recurso piangua (Anadara tuberculosa):
caso Asconar . Informe técnico ASCONAR, Tumaco, Colombia . 89 p .
Gutiérrez, C ., X . Zorrilla, A . Villa y V . Puentes . 1998 . Aspectos biológico pesqueros de la piangua (Anadara
tuberculosa) en el Parque Nacional Natural Sanquianga . Informe técnico, UAESPNN, Cali . 39 p .
Haskoning, C . 1986 . Study on the environmental impact of dredging and excavation on mangroves and
the associated fishery on the mangrove cockle (Anadara tuberculosa) . Informe técnico, Corporación
Autónoma Regional del Valle del Cauca-CVC, Cali . 65 p .
INVEMAR, CRC y CORPONARIÑO . 2006 . Unidad ambiental costera de la llanura aluvial del sur:
caracterización, diagnóstico integrado y zonificación ambiental . INVEMAR-CRC-CORPONARIÑO,
Santa Marta . 383 p .
Lucero, C . 2007 . Aportes de las bases científicas para el acompañamiento de procesos de conservación de
la piangua en bahía Málaga, Pacífico colombiano . Informe técnico, Univ . Valle, Cali . 26 p .
MacKenzie, C . Jr . 2001 . The fisheries for mangrove cockles, Anadara spp ., from Mexico to Peru, with

176 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
descriptions of their habitats and biology, the fishermen’s lives, and the effects of shrimp farming.
Mar . Fish . Rev ., 1: 1-39 .
Ministerio de Ambiente, Vivienda y Desarrollo Territorial-MAVDT, Fondo de Compensación Ambiental-
FCA, Corporación Autónoma Regional de Nariño-Corponariño y Consejo Comunitario Río Tablón
Salado . 2007 . Caracterización, diagnóstico y zonificación de los manglares en el departamento de
Nariño . Informe . Versión 1 .2 . Bogotá . 405 p .
Mora, E . y J . Moreno . 2007 . Estado de la pesquería del recurso concha (Anadara tuberculosa y A . similis)
en la costa ecuatoriana . Instituto Nacional de Pesca . Guayaquil . 15 p .
Naranjo, L . 1982 . Crecimiento y observaciones generales del bivalvo Anadara tuberculosa (Sowerby, 1833)
en Punta Soldado (Bahía de Buenaventura) . Tesis de pregrado, Univ . Valle, Cali . 79 p .
Puentes, V . y J . Celis . 1997 . Aspectos biológico-pesqueros de la piangua Anadara spp . en el Parque Natural
Sanquianga . Ministerio del Medio Ambiente, Bogotá . 41 p .
Silva, A . y R . Bonilla . 2001 . Abundancia y morfometría de Anadara tuberculosa y A. similis (Mollusca:
Bivalvia) en el manglar de Purrujá, Golfo Dulce, Costa Rica . Rev . Biol . Trop ., 40 (2): 315-320 .
Squires M ., M . Estevéz, O . Barona y O . Mora . 1975 . Mangrove cockles, Anadara spp . (Mollusca: Bivalvia)
of the Pacific coast of Colombia . The Veliger, 18: 57-68 .
WWF. 2005. Diagnóstico socioeconómico del sector piangüero y estado actual del recurso hidrobiológico
piangua Anadara tuberculosa en la costa pacífica de Nariño . Fondo Mundial para la Naturaleza-
WWF, Cali . 35 p .
Zar, J.H. 1996. Biostatistical analisys. Tercera Edición, Prentice Hall, Englewood Cliffs, EE. UU. 662 p.
FECHA DE RECEPCIÓN:19/08/2008 FECHA DE ACEPTACIÓN: 22/04/2010

Instituto de Investigaciones Marinas y Costeras - INVEMAR 177
Bol . Invest . Mar . Cost . 39 (1) ISSN 0122-9761 Santa Marta, Colombia, 2010
EFECTO DE LA DENSIDAD DE SIEMBRA SOBRE EL CRECIMIENTO Y SUPERVIVENCIA DE NODIPECTEN NODOSUS (BIVALVIA: PECTINIDAE) EN UN NUEVO
ARTE DE CULTIVO*
Johann López-Navarro1 , Javier Gómez-León2, Claudia Castellanos Romero2 y Ernesto Acosta Ortiz2
1 Universidad del Magdalena, Programa de Ingeniería Pesquera, Santa Marta,Colombia [email protected] 2 Instituto de Investigaciones Marinas y Costera-INVEMAR. A.A. Cerro Punta Betín, Santa Marta, Colombia. [email protected] (J.G.L.); [email protected](C.C.R.); [email protected] (E.A.O.)
RESUMEN
Como un aporte para determinar la factibilidad biológica y tecnológica del cultivo de Nodipecten nodosus en el Caribe colombiano, entre abril y noviembre de 2007 se evaluó el efecto de dos densidades de cobertura (30 y 50 %) sobre su crecimiento y supervivencia . En jaulas suspendidas en el mar a 5 m de profundidad se colocaron juveniles entre 60 y 65 mm de longitud . Se realizaron mediciones quincenales de la longitud de la concha y de la supervivencia . De igual manera se registró la temperatura, salinidad, seston total y materia orgánica particulada para determinar la relación presentada con estos factores; N. nodosus alcanzó una longitud de 84 .7 y 87 .9 mm a 30 y 50 % de densidad, respectivamente . Los tratamientos (30 y 50 %) no difirieron en la supervivencia, pero sí en el crecimiento . Se observó una relación directa de la tasa de crecimiento con la materia orgánica y el seston total . La supervivencia mantuvo una tendencia similar hasta julio en las dos densidades, presentando a su vez una relación directa con la materia orgánica particulada . Los resultados observados demuestran que N. nodosus puede soportar altas densidades, lo que permite tener un mayor aprovechamiento del arte de cultivo, aumentando así la productividad del mismo .
PALABRAS CLAVE: Crecimiento, Supervivencia, Nodipecten nodosus, Densidad, Caribe colombiano
ABSTRACT
Effect of the density on growth and survival of the scallop Nodipecten nodosus (Bivalvia: Pectinidae) in a suspended culture . As a contribution to determine the biological and technological feasibility of the culture of N. nodosus in the Colombian Caribbean, the effect of two cover densities (30 and 50 %) upon growth and survival was evaluated between April and November 2007 . In cages suspended in the sea at 5 m depth juveniles between 60 and 65 mm length were placed . Shell lengths and survival were measured each 15 days . In the same manner, temperature, salinity, seston and particulate
177-189
*Contribución No. 1052 del Instituto de Investigaciones Marinas y Costeras – INVEMAR.

178 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
organic matter were measured in order to determine relationships with these factors; N. nodosus reached a length of 84 .7 and 87 .9 mm at densities of 30 and 50 %, respectively . Treatments affected growth but did not affect significantly survival . A direct relation between growth and organic matter and between growth and total seston were detected . Survival maintained a similar trend until July at both densities, presenting in turn a direct relation to particulate organic matter . Results demonstrate that N. nodosus presents a good development at high densities, which allow to have a greater utilization of the culture art, hence increasing its productivity .
KEY WORDS: Growth, Survival, Nodipecten nodosus, Density, Colombian Caribbean
INTRODUCCIÓN
Entre los moluscos, los bivalvos son considerados un grupo de gran importancia para la acuicultura marina porque muchas especies presentan un amplio mercado y un alto valor comercial (Maeda et al., 1996; Valero et al ., 2000; Pereira y Ferreira, 2006) . Una de estas especies es Nodipecten nodosus, que alcanza un tamaño entre 120 y 140 mm de longitud de la concha (Lodeiros et al., 1998; Acosta et al., 2000) y una coloración externa anaranjado vivo, rojo marrón o vino tinto (Díaz y Puyana, 1994) . Este bivalvo hermafrodita (Román et al., 2001) se distribuye por todo el Atlántico occidental desde Carolina del Norte hasta Brasil (Lodeiros et al., 1998) .
El crecimiento y la supervivencia son procesos poblacionales clave para determinar la viabilidad del cultivo de estos organismos . Estos procesos en los bivalvos se ven afectados por variables ambientales como la temperatura y la salinidad (Román et al., 2001), la oferta y calidad del alimento, que es el principal factor que afecta la fisiología alimentaria de los organismos filtradores (Navarro, 2001) . Todos estos factores se encuentran estrechamente relacionados con otros como la densidad, la profundidad, la localidad y el arte de cultivo (Freites et al ., 1995; Lodeiros et al ., 1998) . Además con la presencia de depredadores (principalmente caracoles y cangrejos), así como la de bioincrustantes (algas y animales), que aumentan drásticamente su mortalidad (Freites et al ., 2000) .
El cultivo de N. nodosus se ha llevado a nivel piloto en Venezuela, Brasil y Colombia (Gómez et al., 1998; Lodeiros et al., 1998; Acosta et al., 2000; Pereira y Ferreira, 2006; Reiser y Manzini, 2007; Rupp et al., 2007; Silva et al., 2007), basado en el modelo japonés, el cual fue desarrollado en la década de los años 60 del siglo pasado (Ventilla, 1982; Ito, 1991) . Así mismo, se han basado en el modelo desarrollado en Chile y Perú, donde se cultiva Argopecten purpuratus a nivel comercial (Illanes, 1995; IMARPE, 2006) .
A pesar de lo anterior, la aplicación de estos procesos de cultivo ha generado inconvenientes tanto de carácter biológico, como económico, ya que en

Instituto de Investigaciones Marinas y Costeras - INVEMAR 179
muchos casos se han visto afectados por las altas inversiones y la baja rentabilidad . Esto se debe a la falta de experiencia y a la mala aplicación de ciertos procesos biológicos y técnicos, haciendo necesaria la búsqueda de mejorar las técnicas de producción, y de esta forma convertirlo en un sistema de producción diferente y atractivo para las comunidades de pescadores e inversionistas del país, ofreciéndoles una oportunidad económica y social encaminadas a mejorar la calidad de vida mediante un aprovechamiento racional de los recursos .
En este estudio se investigó la factibilidad biológica y tecnológica del cultivo de N. nodosus en el Caribe colombiano, evaluando el crecimiento y la supervivencia en dos densidades (30 y 50 %) en cultivo suspendido, utilizando un nuevo arte que consistió en un diseño en cajas suspendidas a 5 m de profundidad . Así mismo, se evaluó la relación entre las variables biológicas y la disponibilidad de alimento, y fisicoquímicas, como temperatura y salinidad . Además de lo anterior, se determinó la presencia de depredadores durante el estudio .
ÁREA DE ESTUDIO
Este estudio se llevó a cabo en la bahía de Neguanje, Parque Nacional Natural Tayrona (PNNT; 11o 20,N, 74o10, W), la cual está ubicada sobre la costa al nororiente de la ciudad de Santa Marta en el Caribe colombiano (Figura 1) . Sus costas son de arenas blancas y gruesas; su característica corresponde a la de una bahía bastante abierta, presentado algunas zonas protegidas (Wedler y Álvarez, 1989; Díaz, 1990) . Además, presenta un régimen global de precipitaciones concentradas en dos períodos de lluvia, siendo la máxima entre septiembre y noviembre (Galvis, 1994) . Estas precipitaciones se alternan con períodos secos, donde predominan los vientos Alisios (diciembre-abril) provenientes del nororiente, que generan una escasez de lluvias y un ambiente drásticamente seco (Márquez, 1982), donde predomina una surgencia local intensa de aguas de la masa subtropical sumergida (Bula, 1985), generada debido al hundimiento progresivo de la plataforma en el sentido este-oeste . Esto origina una menor homogenización de la columna de agua (Blanco, 1993) que, en algunos casos, producen surgencia de agua “fría” proveniente de 100-175 m de profundidad que hacen descender la temperatura marina superficial (a veces hasta 22 oC), a la vez que incrementa la salinidad hasta 38 y los niveles de nutrientes (Garzón y Cano, 1991) . Durante los meses de abril a noviembre los Alisios son usualmente débiles y las lluvias pueden presentarse con mayor frecuencia, caracterizándose a su vez por el aumento de las temperaturas marinas superficiales (hasta 30 °C) y cargada de sedimento (Bula, 1985) . Las lluvias locales pueden originar también importantes descargas fluviales a las bahías, contribuyendo con el incremento de la turbidez y disminución de la salinidad hasta menos de 34 en esta época (Garzón y Cano, 1991) .

180 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
MATERIALES Y MÉTODOS
El experimento se llevó acabo entre abril y noviembre de 2007 . Inicialmente,
se capturaron juveniles con una longitud total entre 55 y 65 mm, provenientes del
medio natural; se tomó la longitud anteroposterior de la concha de al menos 50
individuos al azar y de esta forma se ajustaron las siembras en el arte de cultivo a
las densidades estudiadas . Los individuos seleccionados fueron distribuidos en cajas
que tenían forma rectangular, compuestas por tres compartimientos, construidas en
tubería de polivinilcloruro (PVC) de 2 .3 cm y forradas con malla de nylon de 1
cm de ojo de malla (Figura 2) . Cada compartimiento presentó una dimensión de
35 x 35 x 20 cm, y un área del fondo del arte de 1225 cm2 . Las cajas de cultivo
fueron suspendidas a 5 m de profundidad, en un “long-line” de 100 m de longitud.
Se establecieron dos tratamientos (densidades de 30 y 50 %) con dos réplicas y una
de sustitución, en función de utilizar los ejemplares de ésta última para mantener
la densidad a través del período de estudio, haciendo de cada una de las cajas una
unidad muestral .
El crecimiento se determinó cada quince días, tomando la longitud
anteroposterior de la concha a 10 ejemplares por unidad muestral con la ayuda de
un calibrador de 0 .1 mm de precisión . Se determinó la tasa de crecimiento absoluto
mensual asumiendo que ifif TTAATCA −−= , donde A corresponde a
la longitud en (mm), T al tiempo (días) e i y f a los tiempos iniciales y finales,
respectivamente (Reiser y Manzini, 2007) . La supervivencia se estimó mediante el
Figura 1. Área de estudio y ubicación de la estación de cultivo experimental de bivalvos.

Instituto de Investigaciones Marinas y Costeras - INVEMAR 181
recuento quincenal del número de ejemplares vivos por unidad muestral en cada
densidad estudiada . Los organismos que se encontraron muertos en cada uno de
los recuentos realizados fueron reemplazados por los de la caja de sustitución para
mantener la densidad constante y estimación del crecimiento .
Figura 2. Diseño del arte de cultivo empleado en el experimento .
Cada 30 días se realizó la limpieza de las cajas y de las conchas de los
organismos, donde se logró retirar los organismos bioincrustantes (algas y
animales) . La temperatura se registró una vez al día a la misma hora, con la ayuda
de un termómetro manual de mercurio con precisión de 1 oC; la salinidad se tomó
tres veces por semana mediante un refractómetro; y la disponibilidad de alimento
fue medida como seston y materia orgánica particulada quincenalmente a través de
muestras de agua (5000 mL), siguiendo la técnica descrita por Strickland y Parsons
(1972) . Además, se llevó a cabo el registro de la presencia de depredadores, para
determinar el efecto que éstos causaban a la supervivencia de los organismos en
cultivo .
Para determinar el efecto de la densidad sobre el crecimiento y la
supervivencia de N. nodosus se utilizó la prueba no paramétrica de Mann-Whitney,

182 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
dado que los datos no cumplieron con los supuestos de homogeneidad de varianzas
y normalidad de los residuos . Se hizo un análisis para cada variable, usando como
factor la densidad . Finalmente, se realizaron análisis de correlaciones de Pearson
para comprobar la influencia de las variables ambientales sobre el crecimiento y la
supervivencia de N. nodosus (Zar, 1984) .
RESULTADOS
Variables ambientalesDurante el período de estudio, la temperatura del agua en la estación de
cultivo fluctuó entre 28 .2 °C en abril y 29 .6 °C en julio (Figura 3A); mientras
que la salinidad osciló entre 34 .5 en junio y 33 .2 en noviembre (Figura 3A) . La
concentración del seston total presentó un valor máximo de 1 .7 mg L-1 en noviembre
y un valor mínimo de 0 .9 mg L-1 en julio (Figura 3B); entre tanto la materia orgánica
particulada registró valores de 0 .96 mg L-1 en noviembre y 0 .42 mg L-1 en julio .
Figura 3. Variación de la temperatura y salinidad (A), seston total y materia orgánica particulada (B) en bahía de Neguanje entre abril y noviembre de 2008. La barra indica el error estándar.
A
B

Instituto de Investigaciones Marinas y Costeras - INVEMAR 183
Figura 4. Crecimiento promedio mensual de la longitud de la concha de Nodipecten nodosus en cajas a densidad de 30 y 50 % de cobertura en bahía de Neguanje, entre abril y noviembre de 2008. La barra indica el error estándar.
Tabla 1. Prueba no paramétrica de Mann-Whitney aplicada para evaluar las diferencias de longitud, tasa de crecimiento y supervivencia entre las dos densidades. gl: grados de libertad; w: estadístico; p: nivel designificancia.
CrecimientoAl inicio del estudio, N. nodosus presentó una longitud media de 64 .4
mm (±3.9) y al final alcanzó 84.7 (±0.86) y 87.9 mm (±1.29) de longitud a
30 y 50 % de densidad de cobertura, respectivamente . Las tendencias en ambos
tratamientos fueron similares, demostrando que la talla de los animales aumentó
proporcionalmente durante el período experimental (Figura 4); sin embargo, las
diferencias de longitud entre las densidades fueron significativas (Tabla 1), dado que
los animales mantenidos a la densidad del 50 % de cobertura presentaron un mayor
crecimiento . Por otro lado, se determinó una relación directa entre la longitud de
ambas densidades con la materia orgánica particulada (Tabla 2) .
50
60
70
80
90
ABR MAY JUN JUL AGO SEP OCT NOV
Long
itud
(mm
)
Tiempo (meses)
30 % 50 %
Variable Densidad % n Medias Desviación
estándar gl W p
Longitud30 280 75 .84 7 .04
1 69577 .5 0 .000150 280 75 .46 8 .11
Tasa de crecimiento
30 8 3 .01 0 .811 69 .00 0 .9591
50 8 3 .08 1 .44
Supervivencia30 28 1 .28 0 .16
1 860 .0 0 .308250 28 1 .23 0 .16

184 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Con respecto a la tasa de crecimiento, no se observaron diferencias entre
las densidades (Tabla 1), dado que al final del período se observó una tasa de
crecimiento promedio muy similar con 3.0 (±0.81) y 3.1 mm mes-1 (±1.43) para las
densidades del 30 y 50 %, respectivamente . La tasa de crecimiento observada por la
densidad del 50 % se relacionó indirectamente con el seston total (Tabla 2); mientras
que con el resto de variables ambientales no se presentó relación alguna .
SupervivenciaLa supervivencia mantuvo una tendencia muy similar hasta julio, siendo al
final del estudio mayor para los individuos mantenidos en el tratamiento con densidad
del 30 % . La mayor mortalidad se presentó entre agosto y octubre para la densidad
del 50 % (Figura 5); sin embargo, no se observaron diferencias en los porcentajes de
supervivencia entre los tratamientos (Tabla 2) . De las variables ambientales, sólo se
observaron relaciones indirectas entre la supervivencia mensual de los tratamientos
con la materia orgánica particulada (Tabla 2) .
Tabla 2. Correlación de Pearson. Relación del alimento (materia orgánica particulada: MOP y seston total)entrelongitud,supervivenciaytasadecrecimiento.n=8;p:niveldesignificancia;r=coeficientede correlación.
Alimento Variable Resultado estadístico
Longitud 30%r = 0 .800
p = 0 .016
Longitud 50%r = 0 .780
p = 0 .022
MOPSupervivencia 30%
r = -0 .850
p = 0 .007
Supervivencia 50%r = 0 .780
p = 0 .003
Seston total Tasa de crecimiento 50%r = 0 .825
p = 0 .006

Instituto de Investigaciones Marinas y Costeras - INVEMAR 185
Figura 5. Supervivencia de Nodipecten nodosus cultivada a dos densidades (30 y 50 %) en bahía de Neguanje, entre abril y noviembre de 2008. La barra indica el error estándar.
50
60
70
80
90
100
ABR MAY JUN JUL AGO SEP OCT NOV
Tiempo (meses)
Sup
ervi
venc
ia (%
)
30 % 50 %
DISCUSIÓN
Los valores registrados de temperatura no influyeron en el crecimiento y la
supervivencia de N. nodosus durante el experimento . Estos resultados se asimilan a
los documentados por Lodeiros et al. (1998) quienes evaluaron el crecimiento de la
misma especie . Mientras que Freites et al . (1999) observaron que los altos niveles de
temperatura afectaron la susceptibilidad de N. nodosus, convirtiéndose en un factor
adverso que disminuyó su crecimiento y causó mortalidades . Así mismo, Pereira
y Ferreira (2006) observaron que temperaturas entre 13 .7 y 22 .7 °C inhibieron el
crecimiento de esta especie en Brasil . En cuanto a la salinidad, los valores registrados
durante el estudio presentaron fluctuaciones que no afectaron el crecimiento o la
supervivencia .
Otro factor de gran importancia que, desde un punto de vista ecológico, es
fundamental para considerar la conducta alimentaria de estos organismos (Navarro,
2001) es el seston, el cual durante este estudio presentó un comportamiento opuesto
con la tasa de crecimiento y la supervivencia en ambos tratamientos . Estos resultados
son contrarios a los registrados por otros trabajos con N. nodosus, donde documentan
un comportamiento similar de la concentración del seston con la tasa de crecimiento
(Lodeiros et al., 1998; Acosta et al ., 2000; Freites y Núñez, 2001; Rupp y Pearson,
2004; Pereira y Ferreira, 2006) .

186 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
La densidad de cobertura es un factor determinante para establecer la
rentabilidad de un cultivo; una densidad no adecuada a la cual es sometida una
especie en cultivo se verá reflejada en una baja tasa de crecimiento y, en el menor de
los casos, en una alta mortalidad (Freites et al ., 1995; Acosta et al., 2000; Villarroel
et al ., 2005) . El mayor crecimiento observado en este trabajo a la mayor densidad
probada (50 %), indica que no ocurrió una competencia intraespecífica por alimento
y espacio, resultados que son similares a los documentados por Reiser y Manzini
(2007) al comparar densidades del 50 y 75 % y no encontrar diferencias entre éstas,
lo que pone de manifiesto que N. nodosus presenta un buen crecimiento a altas
densidades, contrastando a su vez lo argumentado por Acosta et al. (2000) y Pereira
y Ferreira (2006) que experimentaron densidades de cobertura del 50 y 100 % sobre
la misma especie y no encontraron buenos resultados .
Desde un punto de vista biológico, las cajas de cultivo empleadas resultaron
un método apropiado para el crecimiento de N. nodosus, al compararlas con las
tradicionales redes linternas, utilizadas por Lodeiros et al. (1998) y Freites et al. (1999) . Sin embargo, el poco número de compartimientos lo convierte en un arte
ineficaz desde un punto de vista de producción con fines comerciales, ya que al
aumentar el número de compartimientos, se convierte en un arte de difícil manejo
(limpieza, extracción y manipulación), más pesado y de mayor ocupación de área en
la línea madre, generando altos costos en cuanto a su producción y mantenimiento .
Varios estudios en pectínidos han evaluado diferentes artes de cultivo suspendidos:
redes perleras, conos, linternas y bolsas, registrando mayor eficiencia en cuanto
al crecimiento y supervivencia en los artes diferentes a las tradicionales linternas
(Freites et al., 1993; Mendoza et al., 2003) .
En cuanto a las mortalidades, éstas fueron atribuidas posiblemente al
estrés generado por la manipulación de los animales durante los muestreos y entre
ellos al introducirse uno dentro del otro, originando que se cortasen el músculo
aductor (Avendaño et al ., 2001), ya que durante este trabajo no hubo presencia de
depredadores del grupo de los gasterópodos, principalmente del género Cymatium,
ni de los decápodos de las familias Xanthidae, Majidae y Portunidae, considerados
como los principales causantes de las mortalidades en el cultivo de estos organismos
(Freites et al ., 2000) . Esto posiblemente fue debido al monitoreo exhaustivo (cada
15 días), lo que impidió probablemente el desarrollo desde su estado larvario en
las artes de cultivo, convirtiéndose así en depredadores potenciales (Lodeiros y
Hilmmelman, 2000) .
La elevada productividad generada por N. nodosus hace de su cultivo
una actividad atractiva con fines comerciales, por presentar altos porcentajes de
supervivencia, gran tamaño y rápido crecimiento, generando así buena rentabilidad
(INVEMAR, 2003) . La talla de 80 mm obtenida por esta especie en tan sólo 11

Instituto de Investigaciones Marinas y Costeras - INVEMAR 187
meses la hace altamente competitiva al compararla con especies de importancia
comercial . Por ejemplo, si se tiene en cuenta el cultivo de Argopecten purpuratus
en Perú y Chile, su talla comercial de 65 mm se logra en un año o año y medio en
condiciones normales (IMARPE, 2006) . Lo anterior sugiere que N. nodosus tiene un
alto potencial biológico y tecnológico en el Caribe colombiano, ya que actualmente
existen alternativas de producción de semillas en hatchery (Velasco y Barros, 2008),
así como los estudios de cultivo en campo que han generado buenos resultados .
AGRADECIMIENTOS
Los autores expresan sus agradecimientos al Departamento Administrativo
de Ciencia, Tecnología e Innovación-Colciencias por la financiación de este estudio,
que hizo parte del trabajo de grado del primer autor, y que estuvo enmarcado en
el proyecto “Optimización de la producción de post-larvas del ostión Nodipecten nodosus y la conchuela Argopecten nucleus en el Caribe colombiano”, código 2105-
09-17982 . De igual manera a los directivos del Instituto de Investigaciones Marinas
y Costera “José Benito Vives De Andréis”-INVEMAR, por el apoyo prestado a lo
largo de esta investigación. Igualmente a María Fernanda Virgüez y Edgard Arias,
propietarios de Hidrocultivos de la Costa Ltda . y a Fabián Cortés, Osman Aragón,
Nelson Bolaño y Katherine Carreño .
BIBLIOGRAFÍA
Acosta, V ., L . Freites y C . Lodeiros . 2000 . Densidad, crecimiento y supervivencia de juveniles de
Lyropecten (Nodipecten) nodosus (Pteroida: Pectinidae) en cultivo suspendido en el golfo de
Cariaco, Venezuela . Rev . Biol . Trop ., 48 (4): 799-806 .
Avendaño, M ., M . Cantillanez, M . Le Pennec, C . Lodeiros y L . Freites . 2001 . Cultivo de pectínidos
iberoamericanos en suspensión . 193-211 . En: Maeda-Martínez, A . N . (Ed .) . Los moluscos pectínidos
de Iberoamérica: ciencia y acuicultura . Editorial Limusa, México D .F . 501 p .
Blanco, J . 1993 . Reconocimiento piloto de fondos, ambiente, fauna asociadas y recursos pesqueros en
aguas costeras del departamento del Magdalena . Informe final, Instituto de Investigaciones Marinas
y Costeras-INVEMAR, Santa Marta . 102 p .
Bula, M . G . 1985 . Un núcleo nuevo de surgencia en el Caribe colombiano detectado en correlación con
las macroalgas . Bol . Ecotrópica, 12: 3-25 .
Díaz, J . M . 1990 . Estudio ecológico integrado de la zona costera de Santa Marta y Parque Nacional
Natural Tayrona . Informe final, Instituto de Investigacioneas Marinas y Costeras-INVEMAR, Santa
Marta . 439 p .
Díaz, J . y M . Puyana . 1994 . Moluscos del Caribe colombiano: un catalogo ilustrado . Colciencias,
Fundación Natura, INVEMAR, Bogotá . 291 p .

188 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
Freites, L . y M . Núñez . 2001 . Cultivo suspendido de Lyropecten (Nodipecten) nodosus (L ., 1758)
mediante los métodos de bolsa y aurícula “ear hanging”. Bol. Inst. Oceanog., 4 (1 y 2): 21-29.
Freites, L ., A . Vélez y C . Lodeiros . 1993 . Crecimiento y productividad de la vieira Euvola (Pecten)
ziczac bajo varios sistemas de cultivo suspendido . Mem . 4th Congr . Latinoam . Cienc . Mar, Serie
Ocasional, 2: 259-269 . Freites, L ., B . Vera, C . Lodeiro y A . Vélez . 1995 . Efecto de la densidad sobre el crecimiento y la producción
secundaria de juveniles de Euvola (Pecten) ziczac, bajo condiciones de cultivo suspendido . Cienc .
Mar ., 21 (4): 361-372 .
Freites, L ., J . Cote, J . Hilmmelman y C . Lodeiros . 1999 . Effects of wave action on the growth and survival
of the scallop Euvola ziczac and Lyropecten nodosus in suspended culture . J . Exp . Mar . Biol . Ecol .,
239: 47-59 .
Freites, L ., J . Hilmmelman y C . Lodeiros . 2000 . Impact of predation and crabs recruiting onto culture
enclosure on the survival of the scallop Euvola ziczac (L .) in suspended culture . J . Exp . Mar . Biol .
Ecol ., 244: 47-59 .
Galvis, R . 1994 . Comportamiento generalizado de las condiciones oceanográficas (temperatura, salinidad,
corrientes y vientos) en la plataforma continental del Caribe colombiano . Informe interno, INPA-
VECEP-UE, Santa Marta . 124 p
Garzón, J . y M . Cano . 1991 . Tipos, distribución, extensión y estado de conservación de los ecosistemas
marinos costeros del Parque Nacional Natural Tayrona . Versión presentada al Séptimo Concurso
Nacional de Ecología, “Enrique Pérez Arbelaez” Fondo para la Protección del Medio Ambiente –
FEN Colombia, Bogotá . 82 p .
Gómez, J ., A . Córdoba y H . Urban . 1998 . Aspectos del crecimiento de Pteria colymbus, Nodipecten
nodosus y Argopecten nucleus (Bivalvia: Pteriidae, Pectinidae) en cultivo suspendido en el Parque
Nacional Natural Tayrona, Caribe colombiano . Resúmenes XI Seminario Nacional de Políticas,
Ciencia y Tecnología del Mar, Bogotá . 80 p .
Illanes, J . E . 1995 . Cultivo de larvas y postlarvas . Mem . 8th Curso Internal . Cultivo Moluscos . UCN-
JICA, Coquimbo, Chile . 384 p .
IMARPE . 2006 . Investigación de invertebrados marinos . http://www .imarpe .gob .pe/imarpe/invertebrados .
php . 12/12/2007 .
INVEMAR . 2003 . Validación y desarrollo de un cultivo piloto de bivalvos en la región de Santa Marta,
Caribe colombiano . Informe final, ECOFONDO-INVEMAR, Santa Marta . 259 p .
Ito, H . 1991 . Fisheries and aquaculture: Japan . 1017-1056 . En: Shumway . S . E . (Ed .) . Scallop: Biology,
ecology and aquaculture. Elsevier Publi. Co., Ámsterdam. 1095 p.
Lodeiros, C . y J . Hilmmelman . 2000 . Identification of factors affecting growth and survival of the tropical
scallop Euvola (Pecten) ziczac in the Golfo de Cariaco, Venezuela . Aquaculture, 182: 91-114 .
Lodeiros, C ., J . Rengel, L . Freites, F . Morales y J . Hilmmelman . 1998 . Growth and survival of the
tropical scallop Lyropecten (Nodipecten) nodosus maintained in suspended culture at three depths .
Aquaculture, 165: 41-50 .
Maeda, A ., P . Omar, T . Moctezuma y V . Osorio . 1996 . Cultivo de scallops en Ecuador . Una alternativa de
desarrollo . Acuicult . Ecuador, 13: 29-36 .
Márquez, G . 1982 . Ecosistemas marinos del sector adyacente a Santa Marta Caribe colombiano . Ecol .
Trop, 2 (1): 5-24 .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 189
Mendoza, Y ., L . Freites, C . Lodeiros, J . López y J . Hilmmelman. 2003 . Evaluation of biological and
economical aspects of the culture of scallop Lyropecten (Nodipecten) nodosus in suspended and
bottom culture . Aquaculture, 221: 207-219 .
Navarro, J . 2001 . Fisiología energética de los pectínidos iberoamericanos . 61-76 . En: Maeda-Martínez,
A . N . (Ed .) . Los moluscos pectínidos de Iberoamérica: ciencia y acuicultura . Editorial Limusa,
México D .F . 501 p .
Pereira, M . y J . Ferreira . 2006 . Eficiencia comparada do cultivo da vieira Nodipecten nodosus (Linnaeus,
1758) (Bivalvia: Pectinidae) em diferentes densidades e profundidades . Biotemas, 19 (2): 37-45 .
Reiser, A . y C . Manzini. 2007 . Efeitos da densidade de estocagem no deselvolvimento da viera Nodipecten
nodosus (Linnaeus, 1758) . XII Congreso COLACMAR, Florianópolis, Brasil . 642 p .
Román, G ., G . Martínez, O . García y L . Freites . 2001 . Reproducción . 27-59 . En: Maeda-Martinez, A . N .
(Ed .) . Los moluscos pectínidos de Iberoamérica: ciencia y acuicultura . Editorial Limusa, Balderas,
México . 501 p .
Rupp, S . y J . Pearson . 2004 . Effects of salinity and temperature on the survival and byssal attachment of
the lion’s paw scallop Nodipecten nodosus at its southern distribution limit . J . Exp . Mar . Bio ., 309:
173-198 .
Rupp, S ., L . Iwersen, G . Manzoni, C . Buglione, C . Herrera, G . Maes y M . Bem . 2007 . Influencia da
profundidade no cultivo da viera Nodipecten nodosus (Linneus, 1758) na enseada da armacao do
Itapocoroy, Santa Catarina . XII Congreso COLACMAR, Florianópolis, Brasil . 642 p .
Silva, R ., M . Bastos, V . Moschen, L . Zaganelli y F . Castro. 2007 . Cultivo experimental da viera Nodipecten
nodosus (Linnaeus, 1758) na Baia da Ilha Grande – Angra dos Reis – Rio de Janeiro. XII Congreso
COLACMAR, Florianópolis, Brasil . 642 p .
Strickland, J . y T . Parsons . 1972 . A practical handbook of seawater análysis . Bull . Fish . Res . Board Can .,
167: 167-315 .
Valero, A ., S . Sánchez, C . García, A . Córdoba, J . Roche, J . Gómez, C . Castellanos, C . Torres, M . Rueda,
K . Hernández, R . Bautista, J . Assmun y Y . Caballero . 2000 . Cultivo de bivalvos marinos del Caribe
colombiano . Colombia Ciencia y Tecnología, COLCIENCIAS, 18 (3): 10-13 .
Velasco, L . y J . Barros . 2008 . Experimental larval culture of the Caribbean scallops Argopecten nucleus
and Nodipecten nodosus . Aqua Res ., 39: 603-618 .
Ventilla R . F . 1982 . The scallop industry in Japan . Adv . Mar . Biol ., 20: 309-382 .
Villarroel, E ., E . Buitrago y C . Lodeiros . 2005 . Identificación de factores ambientales que afectan al
crecimiento y la supervivencia de Crassostrea rhizophorae (Mollusca: Bivalvia) bajo condiciones
de cultivo suspendido en el Golfo de Cariaco, Venezuela . Veterinaria, 14 (1): 1-12 .
Wedler, E. y R. Álvarez. 1989. Estructura de una sección del arrecife coralino en la Playa del Muerto
(bahía de Neguanje, Caribe colombiano) . Carib . J . Sci ., 25 (3-4): 203-217 .
Zar, J . H . 1984 . Biostadistical análysis . Prentice-Hall, Nueva Jersey . 718 p .
FECHA DE RECEPCIÓN:16/09/2008 FECHA DE ACEPTACIÓN: 22/04/2010


Instituto de Investigaciones Marinas y Costeras - INVEMAR 191
Bol . Invest . Mar . Cost . 39 (1) ISSN 0122-9761 Santa Marta, Colombia, 2010
NOTA:
FAUNA NOCTURNA ASOCIADA A LOS MANGLARES Y OTROS HUMEDALES EN LA VÍA PARQUE ISLA DE SALAMANCA, DEPARTAMENTO DEL MAGDALENA,
CARIBE COLOMBIANO*
Sergio A. Balaguera-Reina1, José F. González-Maya1 y Arturo Acero P.2
1 Proyecto de Conservación de Aguas y Tierras, ProCAT Colombia/Internacional, Calle 100 No. 11A-07 Of. 501, Bogotá, Colombia. [email protected] (S.A.B.R.); [email protected] (J.F.G.M.)2 Universidad Nacional de Colombia sede Caribe, CECIMAR/INVEMAR, Cerro Punta Betín, Santa Marta, Colombia. [email protected]
ABSTRACT
Nocturnal fauna associated to mangroves and other wetlands in the Vía Parque Isla de Salamanca, Departamento Magdalena, Colombian Caribbean. This note reports the faunal species associated with mangroves and other wetlands in nocturnal periods in the Vía Parque Isla de Salamanca in order to support management and conservation planning for this natural park . The study was carried out from September through December 2006 using nocturnal surveys . We report 19 species including invertebrates such as arachnids, crustaceans, and mollusks, and vertebrates such as amphibians, reptiles, birds, and mammals . We focused on those species that are classified as threatened either nationally or internationally, and those for which VIPIS represents an information need and an important area for ensuring their persistence .
KEY WORDS: Nocturnal activity, Fauna, Species richness, Mangroves, Vía Parque Isla de Salamanca .
Los ecosistemas de manglar y estuarinos del Caribe colombiano ostentan
una alta relevancia de conservación ya que albergan relictos de fauna únicos de los
antiguos ecosistemas prístinos, siendo así considerados como refugios únicos y
valiosos (Sánchez-Páez et al., 2004) . El estudio de la historia natural de estos relictos,
como sus patrones y períodos de actividad, representa una herramienta fundamental
para el entendimiento de las comunidades y ecosistemas naturales dependientes de
conservación, siendo ésta una base importante para la planificación (González-Maya
191-199
*Contribución No. 331 del Centro de Estudios en Ciencias del Mar, CECIMAR de la Facultad de Ciencias, Universidad Nacional de Colombia.

192 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
et al., 2009a) . A nivel nocturno, existe una notable mayoría de especies de fauna
que presentan una elevada actividad debido, entre otros aspectos, a características
climáticas favorables para la búsqueda de refugio y alimento (Dolsan y Albarran,
1998), lo que a su vez representa un elemento de importancia para el manejo de
sistemas complejos .
A nivel nacional, aunque se han efectuado estudios sobre la fauna presente
en algunos ecosistemas de manglar y estuarios (Sánchez-Páez et al., 1997a, 1997b,
2004), aún no se han realizado trabajos periódicos ni a largo plazo sobre este tema,
existiendo por lo tanto vacíos de información que afectan fuertemente la toma de
decisiones de manejo y conservación . Es por ello que los registros de especies
en ecosistemas protegidos aportan conocimiento base sobre la riqueza presente,
volviéndose un punto de referencia importante para detectar patrones, cambios u
alteraciones en la composición y estructura de los ensamblajes; también ayudan
en gran medida a vislumbrar problemáticas presentes, siendo una base esencial
para el desarrollo de planes y acciones de conservación . Con base en lo anterior,
el presente trabajo busca brindar información básica sobre la fauna nocturna que
habita en los sistemas de estuario y manglar de la Vía Parque Isla de Salamanca
(VIPIS), necesaria para la formulación de planes de manejo, en este caso de áreas
protegidas, pero aplicables a regiones similares bajo diferentes estrategias de
manejo . Sin embargo, es importante desarrollar mayores esfuerzos investigativos
para entender la composición y estructura de estos ecosistemas que deriven en
ordenamientos efectivos, y que a la vez aclaren el panorama sobre la biodiversidad
allí presente .
La VIPIS está ubicada entre 10o53’0.7”-11o0.7’19”N y 74o20’34”-
74o51’0.0”O y cuenta con una extensión de 56200 ha. Limita al norte con el mar
Caribe, al sur con el complejo Pajarales, caño Clarín Nuevo y el municipio de Sitio
Nuevo, al este con el municipio de Pueblo Viejo y la Ciénaga Grande de Santa
Marta (CGSM) y al oeste con el río Magdalena (Figura 1) . Está catalogada como
sitio RAMSAR (González-Maya et al., 2009b) y reconocida como Reserva de la
Biósfera por la UNESCO (Moreno-Bejarano y Álvarez-León, 2003). Presenta una
temperatura promedio anual de 28 °C, una precipitación que oscila entre 400 .1 y
760 .2 mm/año y una humedad relativa media anual de 75 % (Tavera y Gamba,
2001) . Muestra además un mosaico dinámico espacio-temporal de interacción
entre aguas marinas y continentales, generando una alta variedad de hábitats y
zonas de transición para los organismos existentes (Balaguera-Reina, 2007) . La

Instituto de Investigaciones Marinas y Costeras - INVEMAR 193
vegetación característica del área incluye bosques de manglar alrededor de los
caños (asociación R. mangle-A. germinans), ciénagas estuarinas (bordeadas por
Rhizophora mangle con alternancia de Avicennia germinans y vegetación acuática)
y pantanos de aguas continentales, dominando el 21 % de la superficie total de la
VIPIS (12126 .45 ha; Balaguera-Reina, 2007) . Entre otras formaciones vegetales
relevantes en la zona están los bosques sub-xerofíticos y xerofíticos, situados en el
sector nororiental de la isla y bosques riparios y en las orillas de los caños que se
Figura 1. Distribución de las especies que presentan amenazas o relevancia en los procesos de conservación que se están llevando a cabo en VIPIS .
desprenden del río Magdalena (Tavera y Gamba, 2001) .
Los registros se llevaron a cabo dentro de recorridos realizados entre
septiembre y diciembre de 2006 como parte del proyecto “Distribución espacial
y descripción de hábitat de los Crocodylia de la Vía Parque Isla de Salamanca”,
abarcando la época de lluvias y el comienzo de la época seca (Balaguera-Reina,
2007) . Se realizaron 20 recorridos nocturnos en una embarcación de fibra de vidrio
impulsada por un motor fuera de borda (40 hp) a velocidad constante (7 km/h), por
cinco rutas de muestreo durante los cuatro meses de campo, abarcando de norte a sur

194 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
todo el sector centro-occidental del Parque . Los recorridos se efectuaron entre 19:00
y 02:00 h, con un esfuerzo muestreal de 42 h y 110 km por semana a lo largo de los
cuatro meses (promedio por noche 22.0 km ± 14.8), para un total de 168 h y 436.8
km mensuales . Para el muestreo se utilizaron lámparas halógenas de dos millones
de candelas, usando una batería de automotor de 12 V, con las cuales se realizaban
barridos verticales desde la proa a lo largo de las dos márgenes del río (en el caso de
los caños) o el costado visible desde el cuerpo de agua (en el caso de las ciénagas),
detectando la fauna presente en la estructura vegetal adyacente (bosques de manglar
o ribereños) o sobre el espejo de agua (hidrófitia y vegetación acuática) . Al avistar
el animal se acercarba al máximo a este para así determinar la especie y tamaño del
individuo, y en caso de ser posible se tomaron registros fotográficos o se procedió
a realizar captura . La metodología siguió el procedimiento descrito por Balaguera-
Reina y González-Maya (2008, 2009) . Debido a la variedad taxonómica registrada
en el presente estudio, y al ecotono existente en la VIPIS por la interacción entre
sistemas continentales y marinos, se agruparon las especies observadas teniendo en
cuenta sus hábitos (acuáticos o terrestres) y exigencias de hábitat (generalistas o
especialistas), con el fin de obtener una visión más integral de las relaciones de estas
especies con su ecosistema .
Se registraron un total de 19 especies con hábitos nocturnos dentro de
los bosques de manglar de la VIPIS . A nivel de invertebrados se avistaron cinco
especies, dos pertenecientes al filo Mollusca, dos al subfilo Crustacea y uno a la
clase Arachnida (Tabla 1) . En el caso de vertebrados, se observaron anfibios, reptiles,
aves y mamíferos tanto voladores como terrestres, de los cuales algunos presentan
estados de amenaza tanto nacional como internacional . Se encontraron un total de 13
especies con hábitos acuáticos y seis terrestres (Tabla 1), además de un alto número
de especies generalistas como Callinectes bocourti, Nephila clavipes, Rhinella marina (Savage, 2002; Solís et al., 2008), Iguana iguana y Caiman crocodilus fuscus, las cuales presentan amplios ámbitos de distribución y pocas restricciones de
hábitat (Medem, 1981; Balaguera-Reina y González-Maya, 2009) . Aratus pisonii y Polymesoda solida, por su parte, aunque presentan amplios ámbitos de distribución,
muestran mayores restricciones de hábitat (Rueda y Urban, 1998) .
En el caso de la estructura del paisaje, las áreas en donde fueron avistadas la
mayoría de especies de mamíferos y reptiles (Figura 1), presentaron características
notables como una alta cobertura boscosa con una estratificación definida con pocas
alteraciones de origen humano (sector central, ciénagas del Medío y Marchena y
caños El Dedo y Las Lanchas). Áreas de alto impacto, como caño Clarín Nuevo,
Limón, Almendros y la ciénaga La Dársena, mostraron un reducido número de

Instituto de Investigaciones Marinas y Costeras - INVEMAR 195
Grupo taxonómico
Orden Familia EspecieNombre común
Sitios de observación
Subfilo Crustacea
Decapoda Grapsidae Aratus pisonii*Cangrejo del mangle
1,2
PortunidaeCallinectes bocourti*
Jaiba 1
Araneae TetragnathidaeNephila clavipes
Araña hilo dorado
3, 5, 4
Clase Bivalvia
Veneroida CorbiculidaePolymesoda solida*
Ostra 6,7
Clase Gasteropoda
Mesogastropoda Pilidae Pomacea sp.* 3,4
Clase Amphibia
Anura BufonidaeRhinella marina*
Sapo 8,9,10,11
Clase Reptilia
Crocodylia AlligatoridaeCaiman crocodilus fuscus*
Babilla
1, 4, 5, 6, 7, 8, 9, 12, 13, 14, 15, 16, 17, 18
CrocodylidaeCrocodylus acutus*
Caimán aguja
3, 5
Sauria Iguanidae Iguana iguana Iguana 3
Clase Aves
Ciconiformes ArdeidaeButorides striata*
Baco común 4
Nycticorax nycticorax *
Garza nocturna
1, 3, 4, 5, 14
Charadriiformes JacanidaeJacana jacana*
Gallito de ciénaga
3
Coraciiformes AlcedinidaeMegaceryle torquata*
Martín pescador
2, 4, 10, 21
Gruiformes RallidaeAramides cajanea*
Piscingo patas rojas
22
Podicipediformes PodicipedidaeTachybaptus dominicus*
Raspa culo 3
Strigiformes StrigidaePseudoscops clamator
Búho cachón
4
Clase Mammalia
Carnivora Procyonidae Procyon lotorMapache manglero
3, 4, 14, 15, 18, 19, 20, 22
Chiroptera NoctilionidaeNoctilio cf. leporinus*
Murciélago pescador
Todos
Xenarthra MyrmecophagidaeTamandua mexicana
Oso hormiguero
20
Tabla 1. Fauna nocturna registrada entre agosto y diciembre de 2006 en el Vía Parque Isla de Salamanca, departamento de Magdalena, Caribe colombiano . Clave sitios: (1) ciénaga Manatíes, (2) caño de Las Lanchas, (3) caño Clarín Viejo, (4) caño El Dedo, (5) estación Los Cocos, (6) ciénaga El Torno, (7) ciénaga Poza Verde, (8) caño Clarín Nuevo, (9) caño Limón, (10) caño Almendros, (11) ciénaga La Dársena, (12) caño Loro, (13) ciénaga Baco, (14) ciénaga Playa del Perro, (15) ciénaga Las Moras, (16) ciénaga del Medio, (17) ciénaga Cuatro Bocas, (18) ciénaga Marchena, (19) ciénaga Atascosa, (20) caños de Marchena, (21) caño Calentura, (22) ciénaga Lirial . * Especies acuáticas .

196 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
especies avistadas, siendo R. marina y C. c. fuscus los más representativos .Fue notoria una segregación en la distribución de las especies, mostrando
núcleos de acumulación de los avistamientos, encontrándose el 87 % de éstos en bosques inundables (45 %), manglares (14 %) y cuerpos de agua (28 %) . A su vez, se observó que las distancias promedio entre los puntos de observación de todas las especies fueron bajas con una variación amplia (3.08 ± 7.4 km), respecto al tamaño del área total muestreada . Especies como R. marina y C. c. fuscus mostraron una mayor dispersión (18 .0 y 12 .65 km, respectivamente) que las demás especies registradas, siendo la primera de éstas considerada generalista (Savage, 2002) y la segunda catalogada como de amplia distribución y alta abundancia en la zona (Balaguera-Reina, 2007) . Se observó una distribución sectorizada en el caso de C. acutus al igual que un reducido número de individuos de T. mexicana (una única observación), mostrando esto las difíciles condiciones en las cuales se encuentran las poblaciones de estas especies (Figura 1), debido, probablemente, a la reducción de sus hábitats y la presión de caza existente (Balaguera-Reina y González-Maya, en prensa), resaltándose los continuos atropellamientos de T. mexicana en la vía que divide dicho Parque (obs . pers) .
A pesar que la especie aquí registrada es T. mexicana, basado en la identificación directa del individuo y en previos registros para la zona (Moreno-Bejarano y Álvarez-León, 2003), es importante recalcar la posible simpatría con T. tetradactyla en el área, planteada en la Lista Roja de Especies Amenazadas de UICN 2008 (Abba et al., 2008; Meritt et al., 2008; Schipper et al., 2008) . Caso parecido sucede con P. lotor, el cual, según los planteamientos de Alberico et al. (2000), se distribuye en la región Caribe colombiana . Sin embargo, Timm et al. (2008) registran esta especie desde Norteamérica hasta el centro de Panamá, siendo P. cancrivorus la que habita el Caribe colombiano (Reid y Helgen, 2008) . Por su parte, Guzmán-Lenis (2004) plantea que estas últimas especies son simpátricas en la región Caribe, mostrándose la necesidad de realizar aclaraciones concernientes a su distribución, con el fin de verificar qué especie o especies se encuentran presentes en el área . Es importante resaltar que algunas de las especies avistadas se encuentran catalogadas con algún grado de amenaza tanto a nivel nacional como internacional, siendo su registro un importante aporte para los análisis de distribución de estas especies; C. c. fuscus y C. acutus están incluidos en los apéndices II y I de CITES (CITES, 2009), respectivamente, y éste último es considerado como Vulnerable según la UICN (Crocodile Specialist Group, 1996; Ross, 1998) y en “Peligro Crítico” según el Libro Rojo de Reptiles de Colombia (Rodríguez, 2002) . En términos generales, el número de especies detectadas en el área es relativamente bajo comparado con registros anteriores de la misma región (40 especies de reptiles, 194 de aves y 46 de mamíferos; Moreno-Bejarano y Álvarez-León, 2003), sin embargo, de los totales anteriormente registrados, la mayoría se refieren a especies con hábitos principalmente diurnos .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 197
AGRADECIMIENTOS
A la Unidad Administrativa Especial del Sistema de Parques Nacionales
Naturales Seccional Caribe, en especial al personal de la VIPIS . A Giovanni Ulloa,
Bernardo Maya (Reptibol Ltda .), Daniel Rozo, Diego Zárrate, Amancay Cepeda y
Mauricio González por su apoyo en todo el proceso hasta la publicación de este
trabajo .
BIBLIOGRAFÍA
Abba, A ., P . Lara-Ruiz y Members of the IUCN SSC Edentate Specialist Group . 2008 . Tamandua
tetradactyla . http://www .iucnredlist .org . 06/10/2009 .
Alberico, M ., A . Cadena, J . Hernández-Camacho y Y . Muñoz-Saba . 2000 . Mamíferos (Synapsida: Theria)
de Colombia . Biota Colomb ., 1 (1): 43-75 .
Balaguera-Reina, S . A . 2007 . Distribución espacial y descripción de hábitat de los Crocodylia en el
Parque Nacional Natural Vía Isla de Salamanca, Caribe colombiano . Trabajo de grado, Biol . Mar .,
Univ . Jorge Tadeo Lozano, Bogotá . 105 p .
Balaguera-Reina, S . A . y J . F . González-Maya . 2008 . Population structure, density and habitat of
Crocodylus acutus Cuvier 1807 in the Vía Parque Isla de Salamanca, Magdalena Department,
Colombia . Herpetotropicos, 4 (2): 59-63 .
Balaguera-Reina, S . A . y J . F . González-Maya . 2009 . Estructura poblacional y distribución de Caiman
crocodilus fuscus (Cope 1868) en el Vía Parque Isla de Salamanca, Caribe colombiano . Rev . Biol .
Mar . Oceanogr ., 44 (1): 145-152 .
Balaguera-Reina, S . A . y J . F . González-Maya . En prensa . Percepciones, conocimiento y relaciones
entre los Crocodylia y dos poblaciones humanas cercanas al Vía Parque Isla de Salamanca (Caribe
colombiano) . Rev . Latinoam . Conserv .
CITES . 2009 . Apéndices I, II y III en vigor a partir del 22 de mayo de 2009 . CITES-UNEP, Ginebra . 41 p .
Crocodile Specialist Group . 1996 . Crocodylus acutus . http://www .iucnredlist .org . 06/10/2009 .
Dolsan, A . y T . Albarran . 1998 . La problemática de la contaminación lumínica en la conservación de
la biodiversidad . I Sesión de Trabajo sobre la Contaminación Lumínica . Departamento de Medio
Ambiente, Universidad de Cataluña, Barcelona . 8 p .
González-Maya, J . F ., A . Cepeda, S . A . Balaguera-Reina, D . A . Zárrate-Charry y C . Castaño-Uribe .
2009a . Análisis de integralidad ecológica, definición de objetos de conservación y delimitación del
Sitio Ramsar Sistema Delta Estuarino del Río Magdalena Ciénaga Grande de Santa Marta . Informe
técnico, Conservación Internacional-Colombia, Ministerio de Ambiente Vivienda y Desarrollo
Territorial, Bogotá . 95 p .
González-Maya, J . F ., J . Schipper y A . Benítez . 2009b . Activity patterns and community ecology of small
carnivores in the Cordillera Talamanca, Costa Rica . Small Carnivore Conserv ., 41: 9-14 .
Guzmán-Lenis, A . 2004 . Revisión preliminar de la familia Procyonidae en Colombia . Acta Biol . Colomb .,

198 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
9 (1): 69-76 .
Medem, F . 1981 . Los Crocodylia de Sur América, Volumen 1 . Los Crocodylia de Colombia . Colciencias,
Bogotá . 354 p .
Meritt, D ., R . Samudio y Members of the IUCN SSC Edentate Specialist Group . 2008 . Tamandua
mexicana . http://www .iucnredlist .org . 06/10/2009 .
Moreno-Bejarano, M. y R. Álvarez- León. 2003. Fauna asociada a los manglares y otros humedales en
el delta-estuarino del río Magdalena, Colombia . Rev . Acad . Colomb . Cienc ., 27 (105): 517-534 .
Reid, F . y K . Helgen . 2008 . Procyon cancrivorus . http://www .iucnredlist .org . 06/10/2009 .
Rodríguez, M . 2002 . Crocodylus acutus. 41-44 . En: Castaño-Mora, O . V . (Ed .) . Libro rojo de reptiles
de Colombia . Libros rojos de especies amenazadas de Colombia . Instituto de Ciencias Naturales-
Universidad Nacional de Colombia, Ministerio del Medio Ambiente, Conservación Internacional,
Bogotá . 160 p .
Ross, J . P . 1998 . Crocodiles, status survey and conservation action plan . IUCN/SSC Crocodiles Specialist
Group . IUCN, Gand, Suiza, y Cambridge, Reino Unido . 96 p .
Rueda, M . y H . J . Urban . 1998 . Population dynamics and fishery of the fresh-water clam Polymesoda
solida (Corbiculidae) in Ciénaga Poza Verde, Salamanca Island, Colombian Caribbean . Fish . Res .,
39: 75-86 .
Sánchez-Páez, H., R. Álvarez-León, O. A. Guevara-Mancera, A. Zamora-Guzmán, H. Rodríguez-Cruz, y
H . Bravo-Pazmiño . 1997a . Diagnóstico y zonificación preliminar de los manglares del Pacífico de
Colombia . Proyecto de conservación y manejo para el uso múltiple y el desarrollo de los manglares
de Colombia . Ministerio del Medio Ambiente (MMA), Organización Internacional de Maderas
Tropicales-OMIT, Dirección de Proyectos de Repoblación y Ordenación Forestal, Bogotá . 343 p .
Sánchez-Páez, H., R. Álvarez-León, F. Pinto-Nolla, A. S. Sánchez-Alférez, J. C. Pino-Rengifo, I. García-
Hansen y M . T . Acosta-Peñalosa . 1997b . Diagnóstico y zonificación preliminar de los manglares
del Caribe de Colombia . Proyecto de conservación y manejo para el uso múltiple y el desarrollo de
los manglares de Colombia . Ministerio del Medio Ambiente (MMA), Organización Internacional de
Maderas Tropicales-OMIT, Dirección de Proyectos de Repoblación y Ordenación Forestal, Bogotá .
511 p .
Sánchez-Páez, H ., G . Ulloa-Delgado y H . Tavera-Escobar . 2004 . Manejo integral de los mangles por
comunidades locales Caribe de Colombia . Ministerio de Ambiente Vivienda y Desarrollo Territorial,
Coorporación Nacional de Investigación y Fomento Forestal-CONIF, Organización Internacional
de Maderas Tropicales-OIMT . Proyecto Manejo Sostenible y Restauración por las Comunidades
Locales del Caribe de Colombia, Bogotá D . C . 335 p .
Savage, J . M . 2002 . The amphibians and reptiles of Costa Rica . The University of Chicago Press, Miami .
934 p .
Schipper, J ., J . Chanson, F . Chiozza, N . Cox, M . Hoffmann, V . Katariya, J . Lamoreux, A . Rodrigues,
S . Stuart, H . Temple, J . Baillie, L . Boitani, T . Lacher, R . Mittermeier, A . Smith, D . Absolon, J .
Aguiar, G . Amori, N . Bakkour, R . Baldi, R . Berridge, J . Bielby, P . A . Black, J . Blanc, T . Brooks,
J . Burton, T . Butynski, G . Catullo, R . Chapman, Z . Cokeliss, B . Collen, J . Conroy, J . Cooke, G . da
Fonseca, A . Derocher, H . Dublin, J . Duckworth, L . Emmons, R . Emslie, M . Bianchet, M . Foster, S .
Foster, D . Garshelis, C . Gates, M . Giménez-Dixon, S . González, J . F . González-Maya, T . Good, G .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 199
Hammerson, P . Hammond, D . Happold, M . Happold, J . Hare, R . Harris, C . Hawkins, M . Haywood,
L . Heaney, S . Hedges, K . Helgen, C . Hilton-Taylor, S . Hussain, N . Ishii, T . Jefferson, R . Jenkins,
C . Johnston, M . Keith, J . Kingdon, D . Knox, K . Kovacs, P . Langhammer, K . Leus, R . Lewison, G .
Lichtenstein, L . Lowry, Z . Macavoy, G . Mace, D . Mallon, M . Masi, M . McKnight, R . Medellín, P .
Medici, G . Mills, P . Moehlman, S . Molur, A . Mora, K . Nowell, J . Oates, W . Olech, W . Oliver, M .
Oprea, B . Patterson, W . Perrin, B . Polidoro, C . Pollock, A . Powel, Y . Protas, P . Racey, J . Ragle, P .
Ramani, G . Rathbun, R . Reeves, S . Reilly, J . Reynolds, C . Rondinini, R . Rosell-Ambal, M . Rulli,
A . Rylands, S . Savini, C . Schank, W . Sechrest, C . Self-Sullivan, A . Shoemaker, C . Sillero-Zubiri,
N . De Silva, S . Smith, C . Srinivasulu, P . Stephenson, N . van Strien, B . Kumar Talukdar, B . Taylor,
R . Timmins, D . Tirira, M . Tognelli, K . Tsytsulina, L . Veiga, J . C . Vié, E . Williamson, S . Wyatt, Y .
Xie y B. Young. 2008. The status of the world’s land and marine mammals: Diversity, threat, and
knowledge . Science, 322 (5899): 225-230 .
Solís, F ., R . Ibáñez, G . Hammerson, B . Hedges, A . Diesmos, M . Matsui, J . Hero, S . Richards, L . Coloma,
S . Ron, E . La Marca, J . Hardy, R . Powell, F . Bolaños y G . Chaves . 2008 . Rhinella marina . http://
www .iucnredlist .org . 06/10/2009 .
Tavera, H . y N . Gamba . 2001 . Caracterización de la vegetación de la Vía Parque Isla de Salamanca,
Magdalena, Colombia . Trabajo de grado, Ing . Forestal, Univ . Distrital Francisco José de Caldas,
Bogotá D .C . 208 p .
Timm, R ., A . D . Cuarón, F . Reid y K . Helgen . 2008 . Procyon lotor . http://www .iucnredlist .org .
06/10/2009 .
FECHA DE RECEPCIÓN:19/04/2008 FECHA DE ACEPTACIÓN:15/04/2010


Instituto de Investigaciones Marinas y Costeras - INVEMAR 201
FE DE ERRATAS
BOLETÍN DE INVESTIGACIONES MARINAS Y COSTERASVOL. 38 No. 1 (2009)
• En el artículo “La variación anual de los ángulos solares en la latitud de
Santa Marta y su importancia local” (páginas 151-169), la Figura 4, de la página 161,
es incorrecta . La Figura 4 correcta es la siguiente:
-30
-20
-10
0
10
20
30
1 10 19 28 37 46 55 64 73 82 91 100 109 118 127 136 145 154 163 172 181 190 199 208 217 226 235 244 253 262 271 280 289 298 307 316 325 334 343 352 361
días
Dec
linac
ión
(º)
670
680
690
700
710
720
730
740
750
760
770
Dur
ació
n dí
a (m
in)
EpSv Eo Si
Figura 4. Variación anual de la declinación solar (trazo fino) y de la duración del día en minutos (trazo grueso) a mediodía . Puntos críticos cuando para la latitud de Santa Marta se presentan las condiciones de equinoccios (Ep, Eo) y de solsticios (Sv, Si) .


Instituto de Investigaciones Marinas y Costeras - INVEMAR 203
INSTRUCCIONES A LOS AUTORES
Se consideran para publicación en el Boletín de Investigaciones Marinas y Costeras únicamente artículos originales, es decir, aquellos que incluyan resultados significativos que no han sido publicados ni están siendo considerados para publicación en otra revista . Se reciben también artículos de revisión crítica, que compilen y resuman adecuadamente el trabajo en un campo particular y señalen líneas de investigación a seguir . Se tendrán en cuenta los manuscritos preparados según las normas siguientes . Como complemento, se recomienda a los autores siempre examinar cuidadosamente el estilo de presentación de artículos en el último número de la revista .
Idioma: Español o inglés, con resumen en ambos idiomas .
Presentación: Una copia impresa o anexando el archivo en formato digital, como archivo adjunto al correo electrónico: boletin@invemar .org .co o someterlo a la página del boletín: www .invemar .org .co/boletin . Tipo de letra: preferiblemente Times New Roman, tamaño 11 o su equivalente, configuración de página tamaño carta y a doble espacio . Máximo 40 páginas incluyendo figuras y tablas; sólo en casos excepcionales y justificados se aceptarán manuscritos más extensos . Márgenes: izquierdo y superior de 3 cm y derecho e inferior de 2 cm como mínimo . Todos los títulos van en negrilla: título del trabajo y títulos principales en situación centrada y mayúsculas (incluidos los nombres científicos), dejando un espacio para comenzar el párrafo; los títulos de segundo rango centrados pero en minúsculas y sin dejar espacio para comenzar el párrafo; los de tercer rango al lado izquierdo y en minúsculas, también sin espacio . Todas las hojas numeradas consecutivamente, incluyendo las de figuras, tablas y leyendas para las figuras . Una vez aceptado el trabajo, el texto final deberá ser entregado nuevamente en archivo electrónico (textos, figuras y tablas) y/o enviado como archivo adjunto a: boletin@invemar .org .co . El contenido del archivo electrónico debe corresponder exactamente a la copia final impresa .
Texto: Conformado por las siguientes secciones en su orden: Título, Autor(es), Filiación y dirección del (de los) Autor(es), Resumen, Abstract, Introducción, Área de Estudio, Materiales y Métodos, Resultados, Discusión, Conclusiones, Agradecimientos y Bibliografía . Este formato puede ser modificado si la naturaleza del trabajo lo exige . Los artículos en forma de “Notas” deben ser idealmente de máximo 5 páginas de texto y sin división en secciones como los artículos extensos (excepto Abstract cuando el texto sea en español, Resumen cuando el texto sea en inglés), Agradecimientos y Bibliografía . Resumen de máximo 30 renglones en un sólo párrafo . El Abstract debe ser una traducción fiel de lo escrito en el resumen . Tanto el Resumen como el Abstract deben llevar al final y en un renglón aparte, máximo cinco Palabras Clave (Key Words), las cuales deben comenzar en mayúscula . El título del trabajo debe incluir una traducción fiel al inglés cuando el manuscrito sea en español, la cual se incluye en negrita al comienzo del Abstract, y viceversa cuando el trabajo sea en inglés .
Tablas: Elaboradas en procesador de texto (con la opción “tabla”), a doble espacio, cada una comenzando en página aparte y evitando trazar líneas verticales . La leyenda debe ir encima, en minúsculas, y ser suficientemente explicativa . Deben ser numeradas consecutivamente, citadas (en orden) en el texto y presentadas juntas al final del texto . La información incluida en las tablas no puede aparecer repetida en las figuras .
Figuras: Incluir bajo esta denominación todas las ilustraciones, como mapas, esquemas, fotografías y láminas . Deben ser excluidas del texto y el número correspondiente se indicará en cada figura, sobre el margen inferior . Las leyendas deben ir en minúsculas, presentarse juntas en hoja aparte y ser suficientemente explicativas . Dibujos: Letras y símbolos en tamaño apropiado, de modo que al

204 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
reducirlos no pierdan su nitidez . Se aceptan gráficas de computador pero de alta calidad (impresión láser o equivalente) . Fotografías: La revista asume los costos de publicar fotografías en blanco y negro . Los costos de fotografías y figuras a color deben ser asumidos por el autor . Los archivos de las fotografías o imágenes a color deben ser guardados en formato TIF, con una resolución de al menos 360 dpi y los colores deben corresponder a la escala CMYK . Se recomienda que la primera versión del manuscrito a ser sometido utilice una resolución de 72 dpi, en escala RGB, para facilitar el envío de estos archivos por correo electrónico .
Abreviaturas: Pueden usarse abreviaturas o símbolos comunes, tales como m, cm, km, g, kg, mg, mL, L, %, o °C, siempre y cuando vayan acompañando números. Evitar los puntos en las abreviaturas. Todas las cifras decimales deberán separarse con punto y no con coma .
Referencias: Dentro del texto citarse como en los siguientes ejemplos, evitando el uso de op . cit.: “La especie vive sobre sustratos rocosos y coralinos (Pérez, 1980; Darwin y Scott, 1991), aunque, según García (1992), prefiere fondos de cascajo”. Cuando la referencia involucra a más de dos autores debe citarse como: Castro et al . (1965) o (Castro et al., 1965), donde “et al.” debe aparecer siempre en letra cursiva . Las comunicaciones personales deben evitarse en lo posible pero serán aceptadas a juicio del editor a cargo de cada artículo, según sea el caso y deberán seguir el siguiente formato: Nombre completo (punto) Institución (punto) Ciudad (coma) País (punto) Año (punto) Com . Pers . No se aceptan las referencias indirectas, p . ej . Pérez 2007 (En: Fernández, 2008) .
Bibliografía: Incluir todas y únicamente las referencias citadas en el texto, en orden alfabético de apellidos y en orden cronológico cuando haya varios trabajos del mismo autor . El símbolo “&”, comúnmente usado, deberá reemplazarse por la letra “y” o la palabra “and” según el idioma del manuscrito . El estilo de elaboración de las citas bibliográficas debe verse en los trabajos publicados en este volumen para seguir obligatoriamente el mismo formato y su exactitud es responsabilidad únicamente de los autores . No se admiten citas de trabajos que no hayan sido aceptados para publicación y las citas de artículos en prensa deben incluir el volumen, el año, y el nombre de la revista en que saldrán publicados . Los artículos publicados en revistas periódicas se deben citar de la siguiente manera: Autor (punto) Año (punto) Título (punto) Nombre de la revista (coma) Volumen (dos puntos) Ámbito de páginas (punto). Ejemplo: Blanco, J .A ., J .C . Nárvaez y E .A . Viloria . 2007 . ENSO and the rise and fall of a tilapia fishery in northern Colombia . Fish . Res ., 88: 100-108 . En los casos en que la referencia incluya la totalidad de las páginas de la revista periódica, se debe citar así: Autor (punto) Año (punto) Título (punto) Nombre de la revista (coma) Volumen (coma) páginas totales (punto) . Ejemplo: Pang, R .K . 1973 . The systematics of some Jamaican excavating sponges . Postilla, 161, 75 p . Debe abreviarse el nombre de la revista cuando esté formado por más de una palabra, siguiendo la información presentada en www .abbreviations .com . Para revistas no listadas debe presentarse el nombre completo . Los libros se deben citar de la siguiente manera: Autor (punto) Año (punto) Título (punto) Editorial (coma) Ciudad (punto) número total de páginas (punto) . Ejemplo: Dahl, G . 1971 . Los peces del norte de Colombia . Inderena, Bogotá . 391 p . Los capítulos de libros se deben citar de la siguiente manera: Autor (punto) Año (punto) Nombre del capítulo (punto) Ámbito de páginas (punto). Editores (punto) Título del libro (punto) Editorial (coma) Ciudad (punto) número total de páginas (punto). Ejemplo: Álvarez-León, R. y J. Blanco. 1985. Composición de las comunidades ictiofaunísticas de los complejos lagunares estuarinos de la bahía de Cartagena, ciénaga de Tesca y Ciénaga Grande de Santa Marta, Caribe colombiano . 535-555 . En: Yáñez-Arancibia, A . (Ed .) . Fish community ecology in estuaries and coastal lagoons . UNAM, México D .F . 653 p . Las memorias de reuniones y documentos inéditos y de escasa divulgación (tesis y literatura gris) sólo se citarán a criterio del editor del manuscrito . Las tesis se deben citar como los libros . Ejemplo (tesis): Arboleda, E . 2002 . Estado actual del conocimiento y riqueza de peces, crustáceos decápodos, moluscos, equinodermos

Instituto de Investigaciones Marinas y Costeras - INVEMAR 205
y corales escleractíneos del océano Pacifico colombiano . Tesis Biol . Mar ., Univ . Jorge Tadeo Lozano, Bogotá . 125 p .; (informe): Pedraza, R .A ., S . Suárez y R . Julio . 1979 . Evaluación de captura en la Ciénaga Grande de Santa Marta . Informe final, Inderena, Bogotá . 39 p .
Recursos electrónicos: Los documentos electrónicos varían constantemente dentro de las páginas web, por lo tanto se prefiere que los autores citen documentos de este tipo únicamente en casos en que sea indispensable hacerlo y serán aceptadas a juicio del editor a cargo de cada artículo, según sea el caso; deberán seguir el siguiente formato: Autor (punto) Año (punto) Título (punto) URL completa (punto) Fecha de consulta (punto). Ejemplo: Hammer, O., D.A.T. Harper y P.D. Ryan. 2008. PAST – Palaentological Statistics, ver . 1 .81 . http://folk .uio .no/ohammer/past/past .pdf . 08/06/2008 .
Otros: No se admitirán notas de pie de página . Los nombres científicos de géneros y especies deben escribirse en letra cursiva o en su defecto subrayarse . Los registros taxonómicos deben hacerse siguiendo el código internacional de nomenclatura para lo cual el Boletín tiene un modelo con ejemplos, disponible para quien lo solicite . Las medidas se deben expresar en unidades del sistema métrico decimal . Recordar que los valores de salinidad deben presentarse sin unidades o símbolos . Se deben sugerir los nombres de al menos tres evaluadores para el documento sometido, incluyendo la información necesaria para contactarlos (dirección postal y correo electrónico) .
Trámite: Los manuscritos en archivo electrónico deben ser ingresados a la página del boletín www .invemar .org .co/boletin, anexando una carta remisoria dirigida al: Comité Editorial, Boletín de Investigaciones Marinas y Costeras, INVEMAR, Cerro Punta de Betín, Santa Marta, Colombia (e-mail: boletin@invemar .org .co) . Los trámites posteriores de evaluación y comunicación de resultados se harán preferentemente a través de la página o a través de correo electrónico, para mayor agilidad del proceso . Todo trabajo será evaluado por tres especialistas y en un término no mayor de tres meses el Comité Editorial decidirá e informará si el artículo se acepta para publicación, si es aceptable pero debe ser corregido siguiendo las recomendaciones de los evaluadores o si se rechaza . En caso de correcciones los autores deberán entregar una nueva versión en un plazo no mayor a dos meses .

206 Boletín de Investigaciones Marinas y Costeras - Vol. 39 (1) - 2010
INSTRUCTIONS TO AUTHORS
Potential contributors to the Bulletin of Marine and Coastal Research should follow the subsequent instructions: authors must submit original articles; that means those that have significant unpublished results, nor being considered for another journal . The Bulletin also accepts manuscripts on critical revisions that compile and analyze studies in particular fields and propose accurate research lines . We suggest that authors, in any case, check the presentation style in the last Bulletin’s issue to follow the exact structure .
Languages: Spanish or English, abstract in both languages .
Presentation: Attach digital complete text file and figures to an email and send it to boletin@invemar .org .co . Type of font: preferably times New Roman, size 11 or its equivalent, double- spaced . Maximum 40 pages, including figures and tables . Only in exceptional cases we will accept papers longer than that . Margins: Left and upper, 3 cm; right and lower, 2 cm . All titles must be written in boldface type . Manuscript title and main titles must be written in capital letters, centered, and there must be a space line before the first line of the text; subtitles must be also centered but as lower-case letters and there must be no space before the first line of the text; and third titles must be written on the left side of the paper, as lower-case letter, and also with no space . All pages must be numbered consecutively, including figures, tables, and legend for figures pages . Once the paper is accepted, the complete final file must be sent again as attachment to: boletin@invemar .org .co . Final file must include exactly the same information that was accepted as the last version .
Text: Text must be structured as follows: Title, Author (s), Author’s address, Abstract, Abstract in Spanish (Resumen), Introduction, Study area, Materials and Methods, Results, Discussion, Conclusion (optional), Acknowledgements, and Bibliography . This format can be modified depending on the type of work. Articles in “Note” format must be preferably five pages long or shorter, without divisions and subtitles like the long manuscripts (except a Spanish Resumen), Acknowledgements, and Bibliography) . Abstract must be one paragraph and should not exceed 30 lines . Spanish Resumen should be an exact translation of the abstract, both must include at the end a maximum of five key words, which have to start with capital letters . The title of the manuscript should be given in both languages .
Tables: Must be processed using the “table” tool as text file, double-spaced, vertical lines should be avoided, one table per page . Tables legend must be placed on top, in lower-case letters, and present clearly the content . Tables must be numerated consecutively, cited so in the text, and presented together at the end of the manuscript . Information included in tables should not be repeated in figures .
Figures: Figures include illustrations such as maps, pictures, plates, and schemes . Theymust be numbered and presented, excluded from the text, on separated pages . Legends for
figures must be presented in a separated page . Drawings: letters and symbols on drawings should have a suitable size, so that if they get reduced they will not loose their shape and details . We can accept high resolution digital drawings . Photographs: the journal covers the costs for publishing black and white photographs . If there are color images in the manuscript, the authors will have to afford color page fees . Photographs and image files should be saved as TIF files, with at least 360 dpi resolution and colors should correspond to CMYK scale . We suggest sending images for first version in 72 dpi resolution, in RGB scale, so that files are not too heavy to be sent by email .

Instituto de Investigaciones Marinas y Costeras - INVEMAR 207
Abbreviations: Abbreviations may be used and also common symbols, such as m, km, g, mg, %, or ºC, as long as they are next to numbers. Do not use period marks after symbols or abbreviations. All decimal numbers have to be separated by periods . References: In the text please reference the bibliography as follows, avoiding use of op.cit: “The species lives in rocky and coral substrates (Pérez, 1980; Darwin and Scott, 1991), although following García (1992), it prefers gravel substrate.” When the reference includes more than two authors, it has to be quoted as: Castro et al. (1965) or (Castro et al., 1965), with “et al.” always written in italics. Personal communications should not be used but they may be accepted by the editor depending on the specific case . If they are included they should be presented as: Complete name . Institution . City, Country . Year . Pers . Comm .
Bibliography: The bibliography must include all and just the references cited in the text, in alphabetical order and in chronological order when there is more than one article by the same author (s) . The symbol “&”, frequently used, has to be changed for the word “and”. The bibliographic style can be checked in any article in the last issue of the Bulletin and its accuracy is the author’s responsibility. Unpublished data or manuscripts are not supposed to be cited . If a manuscript in press has to be cited, it should include information about issue, year, and title of the journal of the future publication .
Electronic resources: Web documents vary constantly . Therefore, they should be avoided as references in the manuscripts if it is not really necessary. In special cases, it will be the editor’s decision to accept it or not . These citations should follow the next format: Author . Date . Title . Complete URL .
Others: Use of footnotes in the text is strongly discouraged . Scientific names of genera and species must be written in italics type or underlined . Taxonomic reports have to be written following the international nomenclature code . Bulletin office have some examples that may asked for if needed . Measurements have to be marked using metric system . Salinity values have to be presented without symbols or units . You should suggest the names of at least two potential reviewers for the manuscript, including their contact information .
Procedure: Manuscripts, digital files, and communications should be sent to: Comité Editorial, Boletín de Investigaciones Marinas y Costeras, INVEMAR, Cerro Punta de Betín, Santa Marta, Colombia (e-mail: boletin@invemar .org .co) . Communications about results and correction suggestions should be preferably sent by email, in order to abbreviate the complete process . Manuscripts are first reviewed by three experts in the relevant discipline . No longer than three months later, the editorial board will decide and notify if the article is acceptable for publication, if it is acceptable but need some corrections, or if it is rejected . If some corrections are needed, authors have to write a new version and return it no longer than two months later .


Nombre completo:Full name:
Fecha:Date:
Filiación:Affiliation:
Grado académico:Degree:
Dirección:Address:
Ciudad:City:
Departamento:State:
Código postal:ZIP:
País:Country:
Teléfono:Telephone:
Fax:
Correo electrónico:E-mail:
PRECIOSPRICES
COLOMBIA OTROS PAÍSES/OTHER COUNTRIES
$ 20 .000 pesosEnvío: $5 .500 (kilo adicional $1 .200) más IVA
US$ 10más envío / plus shipment (USD$10)
Forma de suscripción (seleccione marcando con una X)Subscription details (select with X)
Canje (especificar la revista)Exchange (specify journal)
Pago Payment
Para pago nacional:BANCO DAVIVIENDACuenta de ahorros No . 1160 6999994-8INVEMAR
Para pagos internacionales:For International payment:ABA Routing Number 0660011392OCEAN BANK780 NW 42nd Avenue, Miami, Florida 33126Account 0082-7657-20INSTITUTO DE INVESTIGACIONES MARINAS Y COSTERAS - INVEMAR
BOLETÍN DE INVESTIGACIONES MARINAS Y COSTERAS(BULLETIN OF MARINE AND COASTAL RESEARCH)
FORMATO DE SUSCRIPCIÓNSUSCRIPTION FORM


BOLETÍN DE INVESTIGACIONES MARINAS Y COSTERAS(Bulletin of Marine and Coastal Research)
BOLE
TÍN D
E INV
ESTIG
ACIO
NES M
ARIN
AS Y
COST
ERAS
39 ( 1
)
(Bulletin of Marine and Coastal Research)(Bulletin of Marine and Coastal Research)
Instituto de Investigaciones Marinas y Costeras “José Benito Vives De Andréis”Vinculado al Ministerio de Ambiente,Vivienda y Desarrollo Territorial
Año 2010 - Volumen 39 (1)
VOL. 39 (1)
Santa Marta - Colombia
Santa Marta, Colombia, 2010
ISSN: 0122-9761
Serie de Publicaciones Periódicas
BOLETÍN DE INVESTIGACIONES MARINAS Y COSTERAS(Bulletin of Marine and Coastal Research)
(ISSN 0122-9761)
La revista científica BOLETÍN DE INVESTIGACIONES MARINAS Y COSTERAS, premiada por Colciencias como la mejor publicación en su género en Colombia en 1997, y clasificada dentro de Publindex (Colciencias) como revista indexada en categoría A2, considera para su publicación trabajos inéditos sobre cualquier tópico de investigación en el mar o en ambientes acuáticos de la zona costera, realizados en las áreas tropicales y subtropicales de América. Es editada desde 1967 por el Instituto de Investigaciones Marinas y Costeras “José Benito Vives De Andréis”-INVEMAR. Hasta el volumen 8 llevó el nombre de “Mittelungen aus dem Instituto Colombo-Alemán de Investigaciones Científicas Punta de Betín” y entre los volúmenes 9 y 24 se denominó “Anales del Instituto de Investigaciones Marinas y Costeras”. A partir de 2008 la revista se publica en dos números por volumen anual. El Boletín se distribuye en 37 países alrededor del mundo a través del sistema de canje. Las instrucciones a los autores para la preparación y envío de manuscritos se encuentran en las últimas páginas de este número y en el portal de Internet www.invemar.org.co/boletin
Director INVEMARFranciso Arias Isaza INVEMAR, Colombia Director de la RevistaJesús A. Garay, M. Sc. INVEMAR, Colombia
Editor Responsable, Vol. 39 (1)Arturo Acero Pizarro, Ph. D. Univ.Nacional de Colombia sede Caribe-CECIMAR,Colombia
Comité EditorialArturo Acero Pizarro, Ph. D. Univ. Nacional de Colombia sede Caribe-CECIMAR, ColombiaCarlos Andrade, Ph. D. Escuela Naval de Cadetes, ColombiaAdriana Bermúdez, Dr. rer. nat. Universidad de Cartagena, ColombiaGladys Bernal, Ph. D. Universidad Nacional de Colombia sede Medellín, ColombiaJacobo Blanco, M. Sc. Universidad del Magdalena, ColombiaMaribeb Castro-González, Dr. en Oceanografía Universidad del Tolima, ColombiaJuan Manuel Díaz, Dr. rer. nat. Centro de Investigaciones Biológicas, ColombiaGuillermo Díaz-Pulido, Ph. D. Griffith University, AustraliaGuillermo Duque, Ph. D. Universidad Nacional de Colombia sede Palmira, ColombiaLuisa Fernanda Espinosa, Ph. D. INVEMAR, ColombiaAndrés Franco, Ph. D. Universidad de Bogotá Jorge Tadeo Lozano, ColombiaDiego L. Gil-Agudelo, Ph. D. INVEMAR, ColombiaJavier Gómez-León, Ph. D. INVEMAR, ColombiaGeorgina Guzmán, Ph. D. INVEMAR, ColombiaJuan Carlos Narváez, M. Sc. Universidad del Magdalena, Colombia Gabriel Navas, M. Sc. INVEMAR, ColombiaCarlos Palacio, Ph. D. Universidad de Antioquia, ColombiaJorge Páramo, M. Sc. Universidad del Magdalena, ColombiaMónica Puyana, Ph. D. Universidad de Bogotá Jorge Tadeo Lozano, ColombiaEfraín Rodríguez, Ph. D. Centro Control Contaminación del Pacífico-CCCP, ColombiaMario Rueda, Ph. D. INVEMAR, ColombiaAdolfo Sanjuán, M. Sc. Universidad de Bogotá Jorge Tadeo Lozano, ColombiaMarie-Luise Schnetter, Dr. rer. nat. Justus Liebig Universität Giessen, AlemaniaYves-François Thomas, Dr. en Geografía Université Paris, FranciaLuisa Villamil, Ph. D. Universidad de Bogotá Jorge Tadeo Lozano, Colombia
Asistente EditorialAmalia María Cano-Castaño, Bióloga INVEMAR, Colombia
CONTENIDO - CONTENTS
L. Gónima, L. E. Ruiz y M. E. González
Desarrollo de una metodología sencilla para la georreferenciación y medición de distancias
a partir de imágenes de satélite sistemáticamente georreferenciadas
[Development of a simple methodology for georeferencing and distance measuring from
systematic georeferenced satellite images] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .7
A. M. Galeano Chavarría y E. Arteaga Sogamoso
Distribución y abundancia de diatomeas del género Pseudo-nitzschia en condición de El
Niño 2007 sobre la cuenca pacífica colombiana
[Distribution and abundance of diatoms of the genus Pseudo-nitzschia in El Niño conditions
in 2007 in the Colombian Pacific Basin] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .25
L. Flórez-Leiva, A. Rangel-Campo, M. Díaz-Ruiz, D. E. Venera-Pontón y G. Díaz-Pulido
Efecto de la sedimentación en el reclutamiento de las macroalgas Dictyota spp. y Lobophora
variegata: un estudio experimental en el Parque Nacional Natural Tayrona, Caribe colombiano
[Effects of sedimentation on the recruitment of the macroalgae Dictytota spp. and Lobophora
variegata: an experimental study in the Tayrona National Natural Park, Colombian Caribbean] . . . . . . . . . . . . . . . . . .41
N. Novoa-Bustos, A. C. Hernández-Zanuy y R. Viquez-Portuguez
Distribución y abundancia de las ascidias de los fondos rocosos de la bahía de Cuajiniquil,
Costa Rica
[Distribution and abundance of the rocky bottom ascidians of Cuajiniquil Bay, Costa Rica] . . . . . . . . . . . . . . . . . .57
T. Posada P., Á. L. Peña C. y G. R. Navas S.
Hidrozoos de la familia Aglaopheniidae de la plataforma continental y talud superior del
Caribe colombiano
[Hydroids of the family Aglaopheniidae from the Colombian Caribbean continental shelf and
upper slope] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .67
Y. Lozano-Duque, L. A. Vidal y G. R. Navas S.
Listado de diatomeas (Bacillariophyta) registradas para el mar Caribe colombiano
[Check-list of diatoms (Bacillariophyta) reported from the Colombian Caribbean Sea] . . . . . . . . . . . . . . . . . .83
BOLETÍN DE INVESTIGACIONESMARINAS Y COSTERASBOLETÍN DE INVESTIGACIONESMARINAS Y COSTERAS
J. G. Domínguez, J. C. Gómez, C. Ricaurte, G. Mayo, J. Orejarena, J. M. Díaz y
C. A. Andrade
Cobertura de los fondos y paisajes bentónicos asociados a formaciones diapíricas en los bancos
de Salmedina, plataforma continental del Caribe colombiano
[Seafloor coverage and benthic landscapes associated to diapiric (mud volcanism) formations
in Salmedina Banks, Colombian Caribbean continental shelf] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 117
K. Gómez-Campo, M. Rueda y C. García-Valencia
Distribución espacial, abundancia y relación con características del hábitat del caracol pala
Eustrombus gigas (Linnaeus) (Mollusca: Strombidae) en el archipiélago Nuestra Señora del Rosario,
Caribe colombiano
[Spatial distribution, abundance and relationship to habitat features of the queen conch
Eustrombus gigas (Linnaeus) (Mollusca: Strombidae) in Nuestra Señora del Rosario archipelago,
Caribbean coast of Colombia] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .137
S. Espinosa G., M. F. Delgado Hernández, B. Orobio Riofrio, L. M. Mejía-Ladino y D. L.
Gil-Agudelo
Estado de la población y valoración de algunas estrategias de conservación del recurso piangua
Anadara tuberculosa (Sowerby) en sectores de Bazán y Nerete, costa pacífica nariñense de
Colombia
[Population status and valuation of some conservation strategies of piangua Anadara tuberculosa
(Sowerby) in areas of Bazán and Nerete, Nariño, Pacífic coast of Colombia] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .161
J. López-Navarro, J. Gómez-León, C. Castellanos Romero y E. Acosta Ortiz
Efecto de la densidad de siembra sobre el crecimiento y supervivencia de Nodipecten nodosus
(Bivalvia: Pectinidae) en un nuevo arte de cultivo
[Effect of the density on growth and survival of the scallop Nodipecten nodosus (Bivalvia: Pectinidae)
in a suspended culture] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .177
NOTAS – NOTES
S. A. Balaguera-Reina, J. F. González-Maya y A. Acero P.
Fauna nocturna asociada a los manglares y otros humedales en la Vía Parque Isla de Salamanca,
departamento del Magdalena, Caribe colombiano
[Nocturnal fauna associated to mangroves and other wetlands in the Vía Parque Isla de
Salamanca, Departamento Magdalena, Colombian Caribbean] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .191
Continúa en la contraportada interior
Viene de la contraportada

BOLETÍN DE INVESTIGACIONES MARINAS Y COSTERAS(Bulletin of Marine and Coastal Research)
BOLE
TÍN D
E INV
ESTIG
ACIO
NES M
ARIN
AS Y
COST
ERAS
39 ( 1
)
(Bulletin of Marine and Coastal Research)(Bulletin of Marine and Coastal Research)
Instituto de Investigaciones Marinas y Costeras “José Benito Vives De Andréis”Vinculado al Ministerio de Ambiente,Vivienda y Desarrollo Territorial
Año 2010 - Volumen 39 (1)
VOL. 39 (1)
Santa Marta - Colombia
Santa Marta, Colombia, 2010
ISSN: 0122-9761
Serie de Publicaciones Periódicas
BOLETÍN DE INVESTIGACIONES MARINAS Y COSTERAS(Bulletin of Marine and Coastal Research)
(ISSN 0122-9761)
La revista científica BOLETÍN DE INVESTIGACIONES MARINAS Y COSTERAS, premiada por Colciencias como la mejor publicación en su género en Colombia en 1997, y clasificada dentro de Publindex (Colciencias) como revista indexada en categoría A2, considera para su publicación trabajos inéditos sobre cualquier tópico de investigación en el mar o en ambientes acuáticos de la zona costera, realizados en las áreas tropicales y subtropicales de América. Es editada desde 1967 por el Instituto de Investigaciones Marinas y Costeras “José Benito Vives De Andréis”-INVEMAR. Hasta el volumen 8 llevó el nombre de “Mittelungen aus dem Instituto Colombo-Alemán de Investigaciones Científicas Punta de Betín” y entre los volúmenes 9 y 24 se denominó “Anales del Instituto de Investigaciones Marinas y Costeras”. A partir de 2008 la revista se publica en dos números por volumen anual. El Boletín se distribuye en 37 países alrededor del mundo a través del sistema de canje. Las instrucciones a los autores para la preparación y envío de manuscritos se encuentran en las últimas páginas de este número y en el portal de Internet www.invemar.org.co/boletin
Director INVEMARFranciso Arias Isaza INVEMAR, Colombia Director de la RevistaJesús A. Garay, M. Sc. INVEMAR, Colombia
Editor Responsable, Vol. 39 (1)Arturo Acero Pizarro, Ph. D. Univ.Nacional de Colombia sede Caribe-CECIMAR,Colombia
Comité EditorialArturo Acero Pizarro, Ph. D. Univ. Nacional de Colombia sede Caribe-CECIMAR, ColombiaCarlos Andrade, Ph. D. Escuela Naval de Cadetes, ColombiaAdriana Bermúdez, Dr. rer. nat. Universidad de Cartagena, ColombiaGladys Bernal, Ph. D. Universidad Nacional de Colombia sede Medellín, ColombiaJacobo Blanco, M. Sc. Universidad del Magdalena, ColombiaMaribeb Castro-González, Dr. en Oceanografía Universidad del Tolima, ColombiaJuan Manuel Díaz, Dr. rer. nat. Centro de Investigaciones Biológicas, ColombiaGuillermo Díaz-Pulido, Ph. D. Griffith University, AustraliaGuillermo Duque, Ph. D. Universidad Nacional de Colombia sede Palmira, ColombiaLuisa Fernanda Espinosa, Ph. D. INVEMAR, ColombiaAndrés Franco, Ph. D. Universidad de Bogotá Jorge Tadeo Lozano, ColombiaDiego L. Gil-Agudelo, Ph. D. INVEMAR, ColombiaJavier Gómez-León, Ph. D. INVEMAR, ColombiaGeorgina Guzmán, Ph. D. INVEMAR, ColombiaJuan Carlos Narváez, M. Sc. Universidad del Magdalena, Colombia Gabriel Navas, M. Sc. INVEMAR, ColombiaCarlos Palacio, Ph. D. Universidad de Antioquia, ColombiaJorge Páramo, M. Sc. Universidad del Magdalena, ColombiaMónica Puyana, Ph. D. Universidad de Bogotá Jorge Tadeo Lozano, ColombiaEfraín Rodríguez, Ph. D. Centro Control Contaminación del Pacífico-CCCP, ColombiaMario Rueda, Ph. D. INVEMAR, ColombiaAdolfo Sanjuán, M. Sc. Universidad de Bogotá Jorge Tadeo Lozano, ColombiaMarie-Luise Schnetter, Dr. rer. nat. Justus Liebig Universität Giessen, AlemaniaYves-François Thomas, Dr. en Geografía Université Paris, FranciaLuisa Villamil, Ph. D. Universidad de Bogotá Jorge Tadeo Lozano, Colombia
Asistente EditorialAmalia María Cano-Castaño, Bióloga INVEMAR, Colombia
CONTENIDO - CONTENTS
L. Gónima, L. E. Ruiz y M. E. González
Desarrollo de una metodología sencilla para la georreferenciación y medición de distancias
a partir de imágenes de satélite sistemáticamente georreferenciadas
[Development of a simple methodology for georeferencing and distance measuring from
systematic georeferenced satellite images] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .7
A. M. Galeano Chavarría y E. Arteaga Sogamoso
Distribución y abundancia de diatomeas del género Pseudo-nitzschia en condición de El
Niño 2007 sobre la cuenca pacífica colombiana
[Distribution and abundance of diatoms of the genus Pseudo-nitzschia in El Niño conditions
in 2007 in the Colombian Pacific Basin] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .25
L. Flórez-Leiva, A. Rangel-Campo, M. Díaz-Ruiz, D. E. Venera-Pontón y G. Díaz-Pulido
Efecto de la sedimentación en el reclutamiento de las macroalgas Dictyota spp. y Lobophora
variegata: un estudio experimental en el Parque Nacional Natural Tayrona, Caribe colombiano
[Effects of sedimentation on the recruitment of the macroalgae Dictytota spp. and Lobophora
variegata: an experimental study in the Tayrona National Natural Park, Colombian Caribbean] . . . . . . . . . . . . . . . . . .41
N. Novoa-Bustos, A. C. Hernández-Zanuy y R. Viquez-Portuguez
Distribución y abundancia de las ascidias de los fondos rocosos de la bahía de Cuajiniquil,
Costa Rica
[Distribution and abundance of the rocky bottom ascidians of Cuajiniquil Bay, Costa Rica] . . . . . . . . . . . . . . . . . .57
T. Posada P., Á. L. Peña C. y G. R. Navas S.
Hidrozoos de la familia Aglaopheniidae de la plataforma continental y talud superior del
Caribe colombiano
[Hydroids of the family Aglaopheniidae from the Colombian Caribbean continental shelf and
upper slope] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .67
Y. Lozano-Duque, L. A. Vidal y G. R. Navas S.
Listado de diatomeas (Bacillariophyta) registradas para el mar Caribe colombiano
[Check-list of diatoms (Bacillariophyta) reported from the Colombian Caribbean Sea] . . . . . . . . . . . . . . . . . .83
BOLETÍN DE INVESTIGACIONESMARINAS Y COSTERASBOLETÍN DE INVESTIGACIONESMARINAS Y COSTERAS
J. G. Domínguez, J. C. Gómez, C. Ricaurte, G. Mayo, J. Orejarena, J. M. Díaz y
C. A. Andrade
Cobertura de los fondos y paisajes bentónicos asociados a formaciones diapíricas en los bancos
de Salmedina, plataforma continental del Caribe colombiano
[Seafloor coverage and benthic landscapes associated to diapiric (mud volcanism) formations
in Salmedina Banks, Colombian Caribbean continental shelf] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 117
K. Gómez-Campo, M. Rueda y C. García-Valencia
Distribución espacial, abundancia y relación con características del hábitat del caracol pala
Eustrombus gigas (Linnaeus) (Mollusca: Strombidae) en el archipiélago Nuestra Señora del Rosario,
Caribe colombiano
[Spatial distribution, abundance and relationship to habitat features of the queen conch
Eustrombus gigas (Linnaeus) (Mollusca: Strombidae) in Nuestra Señora del Rosario archipelago,
Caribbean coast of Colombia] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .137
S. Espinosa G., M. F. Delgado Hernández, B. Orobio Riofrio, L. M. Mejía-Ladino y D. L.
Gil-Agudelo
Estado de la población y valoración de algunas estrategias de conservación del recurso piangua
Anadara tuberculosa (Sowerby) en sectores de Bazán y Nerete, costa pacífica nariñense de
Colombia
[Population status and valuation of some conservation strategies of piangua Anadara tuberculosa
(Sowerby) in areas of Bazán and Nerete, Nariño, Pacífic coast of Colombia] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .161
J. López-Navarro, J. Gómez-León, C. Castellanos Romero y E. Acosta Ortiz
Efecto de la densidad de siembra sobre el crecimiento y supervivencia de Nodipecten nodosus
(Bivalvia: Pectinidae) en un nuevo arte de cultivo
[Effect of the density on growth and survival of the scallop Nodipecten nodosus (Bivalvia: Pectinidae)
in a suspended culture] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .177
NOTAS – NOTES
S. A. Balaguera-Reina, J. F. González-Maya y A. Acero P.
Fauna nocturna asociada a los manglares y otros humedales en la Vía Parque Isla de Salamanca,
departamento del Magdalena, Caribe colombiano
[Nocturnal fauna associated to mangroves and other wetlands in the Vía Parque Isla de
Salamanca, Departamento Magdalena, Colombian Caribbean] . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .191
Continúa en la contraportada interior
Viene de la contraportada