bordetella bronchiseptica pagp is a bvg-regulated lipid a palmitoyl transferase that is required for...
TRANSCRIPT

Molecular Microbiology (2003)
48
(3), 725–736
© 2003 Blackwell Publishing Ltd
Blackwell Science, LtdOxford, UKMMIMolecular Microbiology1365-2958Blackwell Publishing Ltd, 200348
3
Original Article
B. bronchiseptica pagPA. Preston et al.
Accepted 27 January, 2003. *For correspondence. [email protected]; Tel. (
+
44) 122 333 7688; Fax (
+
44) 122333 7610.
Bordetella bronchiseptica
PagP is a Bvg-regulated lipid A palmitoyl transferase that is required for persistent colonization of the mouse respiratory tract
Andrew Preston,
1
* Elizabeth Maxim,
1
Elinor Toland,
1
E. Jane Pishko,
2
Eric T. Harvill,
2
Martine Caroff
3
and Duncan J. Maskell
1
1
Centre for Veterinary Science, Department of Clinical Veterinary Medicine, University of Cambridge, Madingley Road, Cambridge CB3 0ES, UK.
2
The Pennsylvania State University, Department of Veterinary Science, University Park, PA 16802, USA.
3
Equipe Endotoxines, UMR 8619 du Centre National de la Recherche Scientifique, Biochimie, Universite de Paris-Sud, Orsay, France.
Summary
Bordetella bronchiseptica
lipopolysaccharide (LPS)expression varies depending on growth conditions,regulated by the Bvg system. A
B. bronchisepticapagP
homologue was identified that is required forBvg-mediated modification of the lipid A core regionof LPS that occurs on switching from the Bvg
–
to theBvg++++
phase. Structural analysis demonstrated that thelipid A of a
B. bronchiseptica pagP
mutant differedfrom wild-type lipid A by the absence of a palmitategroup in secondary acylation at the C3
¢¢¢¢
position. Theputative
pagP
promoter drove the expression of agreen fluorescent protein (GFP) reporter gene in aBvg-regulated fashion. These data suggest that
B.bronchiseptica pagP
encodes a Bvg-regulated lipid Apalmitoyl transferase that mediates modification ofthe lipid A as part of the overall Bvg-mediated adap-tation of this organism to changing environmentalconditions. We also show that
pagP
is not requiredfor the initial colonization of the mouse respiratorytract by
B. bronchiseptica
, but is required for persis-tence of the organism within this organ.
Introduction
The genus
Bordetella
currently contains eight species,some of which are respiratory tract pathogens. Three of
these species have been studied in greater detail, namely
B. pertussis
,
B. parapertussis
and
B. bronchiseptica
.
B.bronchiseptica
infects many mammalian species and iscommonly found associated with atrophic rhinitis in pigs,snuffles in rabbits and kennel cough in dogs (Rutter, 1981;Thrusfield
et al
., 1991a,b; Burns
et al
., 1993). It has alsobeen described occasionally as a respiratory tract patho-gen in humans, but only in the very old or immunosup-pressed (Reina
et al
., 1991; Gueirard
et al
., 1995; Tamion
et al
., 1996).Most of the putative virulence determinants of the bor-
detellae that have been described are proteins whoseexpression is regulated by the Bvg two-component sys-tem in response to environmental stimuli (reviewed byCotter and Miller, 2001).
Bvg
is regarded as a globalcontrol system that mediates modulation between differ-ent phases. In each phase, different genes areexpressed or repressed. In the Bvg-plus (Bvg
+
) phase,BvgA protein is phosphorylated by the sensor proteinBvgS in response to environmental stimuli, and BvgA isthen able to upregulate the expression of several of theproteins that are considered to be important for coloniza-tion of the host respiratory tract. These include putativeadhesins such as pertactin and filamentous haemaggluti-nin, toxins such as pertussis toxin and adenylate cyclase/haemolysin and other systems including the type IIIsecretory system. Other genes are repressed in the Bvg
+
phase (the so-called
vrg
genes). In the Bvg-minus (Bvg
–
)phase, the expression of the virulence-associated genesis downregulated, whereas the expression of the
vrg
genes is upregulated. Thus the Bvg
+
phase may beregarded as the state that the bacterium adopts on enter-ing the host respiratory tract, whereas the Bvg
–
phasemay be regarded as the state that the bacterium adoptsoutside the host. More recently, distinct phases interme-diate between plus and minus (Bvg-i) have been recog-nized (Cotter and Miller, 1997). The stimuli to which Bvgresponds
in vivo
are unknown, and thus the precise con-ditions that cause switching between phases areunknown.
The role of lipopolysaccharide (LPS) in
Bordetella
pathogenicity and immunity has been largely overlookedpartly because the genetic basis for LPS biosynthesis inthe bordetellae was completely unknown until recently.

726
A. Preston
et al.
© 2003 Blackwell Publishing Ltd,
Molecular Microbiology
,
48
, 725–736
The small amount of research addressing the issue of
Bordetella
LPS and its role in infection has shown that itis highly immunogenic, is an immunological adjuvant anddisplays the properties expected of an endotoxin (Chabyand Caroff, 1988; Amano
et al
., 1990; Watanabe
et al
.,1990). Furthermore,
Bordetella
LPS mutants are attenu-ated in a mouse model of infection, suggesting thatexpression of a full-length LPS molecule is required forvirulence (Harvill
et al
., 2000; West
et al
., 2000; Burns
et al
., 2003).
Bordetella bronchiseptica
LPS comprises a lipid Adomain, a branched-chain core oligosaccharide and an Oantigen-like repeat (Caroff
et al
., 2001) (Fig. 1). The coretogether with lipid A forms the structure called band B(Peppler, 1984; Caroff
et al
., 1990). This may be substi-tuted further by a complex trisaccharide, the fully substi-tuted LPS (band B
+
trisaccharide) being called band A(Peppler, 1984; Caroff
et al
., 1990).
B. bronchiseptica
lipidA structures appear to be variable between strains, withunusual fatty acyl substitutions on the diglucosaminebackbone of the molecule (Zarrouk
et al
., 1997). In addi-
tion,
B. bronchiseptica
and
Bordetella hinzii
are the onlybordetellae in which hexa-acylated lipid A has beenobserved as a major LPS species (Zarrouk
et al
., 1997;Aussel
et al
., 2000).The LPS phenotype of a bacterium can vary in
response to environmental parameters. It is now clear thatprecise regulation of LPS phenotype in response to par-ticular environmental cues is a widespread phenomenonamong Gram-negative bacteria and that this regulationcan be an important component of the responses ofpathogens to different econiches within the host duringthe course of infection (reviewed by Preston and Maskell,2002).
For example,
Salmonella typhimurium
lipid A under-goes a number of covalent modifications mediated bythe PhoPQ and PmrAB two-component systems inresponse to environmental stimuli (Raetz, 2001). Someof these modifications render the bacterium more resis-tant to the killing action of certain cationic antimicrobialpeptides (CAMPs) (Gunn, 2001) and involve the addi-tion of 4-amino-4-deoxy-
L
-arabinose (
L
-ara4N) to the C-
Fig. 1.
Schematic representation of
B. bronchiseptica
LPS. The molecule comprises lipid A (green), a branched-chain core (red), a distal trisaccharide (blue) and a homopolymeric O antigen (magenta) that is attached somewhere on the core/trisaccharide region. The SDS-PAGE profile (see
Experimental procedures
) of
B. bronchiseptica
RB50 is shown in the inset and comprises three LPS species. Band B is composed of lipid A and core. Band A is composed of band B further substituted with the terminal trisaccharide. O antigen-containing LPS resolves as a diffuse band. The core structure shown is that of
B. pertussis
strain 1414, and is the one complete
Bordetella
LPS core structure known (Caroff
et al
., 2000).
B. bronchiseptica
core is very similar if not identical to this structure (Caroff
et al
., 2001).

B. bronchiseptica pagP 727
© 2003 Blackwell Publishing Ltd,
Molecular Microbiology
,
48
, 725–736
4
¢
and/or C-1 phosphates and/or the addition of palmi-tate in acyloxyacyl linkage to one of the primary hydrox-ylated acyl chains. This addition requires the enzymePagP (Guo
et al
., 1998; Bishop
et al
., 2000). PagP isunusual as an enzyme for LPS biosynthesis in that it islocated in the outer membrane, whereas most otherLPS biosynthesis functions are located at the cytoplas-mic–inner membrane interface (Bishop
et al
., 2000).PagP transfers palmitate from phospholipids to matureLPS molecules in the outer membrane under conditionsof PhoPQ activation (Bishop
et al
., 2000), thus increas-ing the acylation of the LPS molecules. In addition to arole in resistance to CAMP-mediated killing, the result-ant hepta-acylated, palmitoylated
S. typhimurium
lipid Amay act as an antagonist towards the cytokine inductionactivity of
Escherichia coli
LPS on human cell lines(Tanamoto and Azumi, 2000), indicating that the palmi-tate modification modulates the activity of lipid A.Together, these lipid A modifications might be expectedto lead to salmonellae less likely to induce host cytokineresponses and better able to survive innate host resis-tance mechanisms.
We report here the identification of a PagP homo-logue in
B
.
bronchiseptica
. Mutation of
B. bronchisepticapagP
abrogated Bvg-mediated LPS core modification.Structural analyses confirmed that
B. bronchisepticapagP
LPS lacked palmitate in acyl-oxyacyl linkage at theC3
¢
position present in wild-type LPS. Thus, we hypoth-esize that
B. bronchiseptica
LPS undergoes palmitoyla-tion by PagP as part of the Bvg-mediated response tohost stimuli that is necessary to adapt to the changingenvironments in the host during infection. Furthermore,although
pagP
was not required for the initial coloniza-tion of the mouse respiratory tract by
B. bronchiseptica
,it was required for persistence of the bacteria within thisorgan, confirming a role for
pagP
in the infectiousprocess.
Results
Bordetella bronchiseptica
LPS is subject to Bvg-mediated modifications
We observed that the LPS phenotype of
B. bronchiseptica
varied depending on growth conditions (Fig. 2). This is inagreement with a previous report in which the tempera-ture- and growth condition-dependent variation of the LPSof several
Bordetella
species was noted (van den Akker,1998).
B. bronchiseptica
grown at 37
∞
C in the presenceof 50 mM MgSO
4
adopt the Bvg
–
phase. Under theseconditions, the SDS-PAGE LPS profile is that classicallydescribed for these bacteria, comprising band B LPS,band A LPS and LPS containing O antigen. There was amarked change in the LPS profile of bacteria grown in theabsence of MgSO
4
at 37
∞
C, conditions in which they adoptthe Bvg
+
phase. In these conditions, O-antigen expressionwas markedly decreased and the core lipid A region wasmodified to produce a doublet band that migrates toapproximately the same position as band A LPS (Fig. 2).Both the bands in this doublet react with an anti-band Amonoclonal antibody (data not shown) and thus containthe band A epitope recognized by this antibody, suggest-ing that the LPS in this doublet contains a modification ofthe Bvg
–
phase LPS rather than a completely novel LPS.Thus, the LPS profile is modified by switching betweengrowth conditions that also modulate Bvg activity. Toinvestigate the role of Bvg in this LPS modification, theLPS profiles of several Bvg phase-locked mutants wereanalysed (Fig. 2). A Bvg
+
phase-locked mutant had thesame profile as the wild-type (WT) strain grown in Bvg
+
phase conditions. A Bvg
–
phase-locked mutant had thesame profile as the WT strain grown in Bvg
–
phase con-ditions. Neither of these phase-locked mutants respondedto the presence or absence of MgSO
4
with a change inLPS profile. This strongly suggests that the LPS modifica-tion that occurs in the WT strain on switching from the
Fig. 2.
SDS-PAGE LPS profiles (see
Experimental procedures
) of
B. bronchiseptica
RB50 and Bvg mutants of this strain. Bacteria were grown at either 37
∞
C in the Bvg
+
phase or 37
∞
C in the pres-ence of 50 mM MgSO
4
in the Bvg
–
phase. Lanes 1–5, Bvg
–
phase. Lanes 6–10, Bvg
+
phase. Lanes 1 and 6, RB50 wild type; lanes 2 and 7, RB53, a Bvg
+
phase-locked mutant; lanes 3 and 8, RB53I, a Bvg intermediate phase mutant; lanes 4 and 9, RB50I, a Bvg intermediate phase mutant; lanes 5 and 10, RB54, a Bvg
–
phase-locked mutant. In the WT, but not in the Bvg phase-locked mutants, the LPS phenotype is modified on switching from the Bvg
– to the Bvg+ phase, involving a decrease in expression of O antigen-containing LPS and mod-ification of the core/lipid A region.

728 A. Preston et al.
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 48, 725–736
Bvg– to the Bvg+ phase requires a functional Bvg systemand that B. bronchiseptica LPS is subject to Bvg-mediatedmodification. Consistent with this hypothesis, two Bvg-intermediate phase mutants had LPS profiles intermedi-ate between the Bvg+ and Bvg– phases (Fig. 2).
Identification of a PagP homologue in B. bronchiseptica genome sequence data
The existence of a B. pertussis PagP homologue hadbeen noted previously (Bishop et al., 2000). The aminoacid sequence of S. typhimurium PagP was used in aBLASTP search of the B. bronchiseptica genome sequenceidentifying an open reading frame (ORF) that is similar tothe S. typhimurium sequence (1.5e-30). Upstream of thisORF is an ORF that is very similar (e-111) to members ofthe MgtE family of putative magnesium transporters(pfam01769). This ORF is arranged in the same directionbut separated from pagP by an intergenic region of104 bp. Downstream of pagP is an ORF that is similar(e-31) to a number of putative cation transporters fromdifferent bacteria, including Mesorhizobium loti, Ralstoniasolanacearum and Brucella melitensis. This ORF andpagP are arranged in opposite directions and have over-lapping stop codons.
PCR amplification, cloning and mutagenesis of the B. bronchiseptica PagP locus
Using genome sequence information, primers weredesigned, and the pagP locus (extending 500 bpupstream of the pagP ATG and 65 bp downstream of thepagP stop codon) was amplified by polymerase chainreaction (PCR) and cloned. An erythromycin resistancecassette was inserted into a BsrGI site within the pagPcoding sequence. This mutation was moved on to the B.bronchiseptica chromosome via allelic exchangemutagenesis to produce a pagP mutant. The expectedchromosomal rearrangement was confirmed by Southernhybridization (data not shown).
Analysis of B. bronchiseptica pagP LPS phenotype
The LPS profile of B. bronchiseptica pagP was analysedby SDS-PAGE and silver staining (Fig. 3). In the Bvg–
phase, both WT and pagP mutant LPS profiles were thesame, comprising band B LPS, band A LPS and O anti-gen-substituted LPS. However, in the Bvg+ phase,whereas the WT profile displays the doublet core lipid Acharacteristic of Bvg-mediated modification, the pagPmutant did not express this modified structure, but insteadexpressed a single band A LPS structure as observed inthe Bvg– phase. This result suggested that pagP wasrequired for Bvg-mediated modification of the core lipid A
region of B. bronchiseptica LPS and that this modificationis added in the Bvg+ phase. Interestingly, the LPS of thepagP mutant in the Bvg+ phase migrated to a slightlyhigher position in the gel than did the LPS of the pagPmutant in the Bvg– phase. This suggests that modificationof the LPS occurs on switching from the Bvg– to the Bvg+
phase in addition to that mediated by PagP. The nature ofthis modification is at present unknown.
To confirm that the loss of LPS modification observedresulted from mutation of the pagP gene, and not from asecond site mutation, the mutation was complemented.pagP, including its putative promoter region, was ampli-fied by PCR, cloned into pCR2.1 Topo and then movedinto pBBR1kan. This construct was introduced into B.bronchiseptica pagP by conjugation. Clones were veri-fied by plasmid extraction and restriction analysis. TheLPS profiles of a representative clone grown under Bvg+
and Bvg– conditions are shown in Fig. 3. The introduc-tion of the plasmid-borne copy of pagP restored modifi-cation of the LPS, suggesting that the loss ofmodification in B. bronchiseptica pagP resulted frommutation of pagP and not from a second site mutation. Inthe complemented clone, some modification wasobserved in the Bvg– phase. This is probably caused bythe complementing gene being carried on a multicopyplasmid leading to a low level of transcription from thepagP promoter occurring in the Bvg– phase. In this sce-nario, in the wild type, the level of PagP expression inthe Bvg– phase is insufficient to give rise to observable
Fig. 3. SDS-PAGE LPS profiles (see Experimental procedures) of B. bronchiseptica RB50, B. bronchiseptica pagP and B. bronchiseptica pagP complemented with a plasmid-borne copy of the pagP gene. Lane 1, RB50, Bvg+ phase; lane 2, RB50, Bvg– phase; lane 3, B. bronchiseptica pagP, Bvg+ phase; lane 4, B. bronchiseptica pagP, Bvg– phase; lane 5, the complemented strain, Bvg+ phase; and lane 6, the complemented strain, Bvg– phase. The pagP mutant does not affect modification of the core/lipid A that occurs in WT on switching from the Bvg– to the Bvg+ phase. This modification is restored by complementing the pagP mutation with a wild-type copy of the gene.

B. bronchiseptica pagP 729
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 48, 725–736
LPS modification. However, in the complementing clone,the pagP copy number is high enough to give sufficientPagP expression to cause the LPS modification seen inFig. 3. In agreement with this, low-level transcriptionfrom the pagP promoter is observed in the Bvg– phasewhen it is carried on a reporter gene plasmid (seebelow).
LPS purification and structural analysis
The structures of the lipid As from LPS purified from Bvg+
phase WT and mutant bacteria were determined. Previ-ously, four different lipid A structures have been describedfor five Bordetella species examined (Caroff et al., 2001).The lipid A acylation patterns of B. bronchiseptica werefound to be different from those of other members of thegenus. In three B. bronchiseptica strains examined, eitherdodecanoic acid (C12) or 3-hydroxydodecanoic acid (C12-OH) was present at the C-3 position of glucosamine I ofthe lipid A, and only one strain had a major hexa-acylatedcomponent (Zarrouk et al., 1997), the other two havingmajor tetra- and/or penta-acylated components asobserved in other Bordetella species. Determinationof the total fatty acid composition of the WT lipid Arevealed that C12, C12-OH, tetradecanoic acid (C14), 3-hydroxytetradecanoic acid (C14-OH) and hexadecanoicacid (C16) occurred in the proportions 0.3:0.5:1:2.8:1. Themutant lipid A also contained these fatty acids but in theproportions 0.3:0.45:1:3:0.1, the main difference from WTbeing the much reduced presence of C16.
The esterified substituents at the C-3 and C-3¢ positionsof the diglucosamine were liberated by mild alkali treat-ment and analysed. From WT lipid A, C12/C12-OH, C14-OHand C16 as well as C14OC16 were liberated but, from themutant lipid A, only C12/C12-OH and C14-OH were liber-ated. The free C14-OH and C16 fatty acids originated fromthe C14OC16 in the WT lipid A.
The residual lipid obtained after mild alkali treatmentwas analysed by matrix-assisted laser desorption ioniza-tion (MALDI) mass spectrometry in the negative- andpositive-ion modes (not shown). Molecular species andfragments were observed. A major peak in the negative-ion mode was obtained at m/z 1162, corresponding to twoglucosamines, two phosphates, two C14-OH in amide link-age and one C14 in acyl-oxyacyl position on glucosamine-II. All are characteristic of bordetellae lipid As.
The lipid A isolated from the WT strain gave a negative-ion MALDI spectrum displaying signals for major penta-and hexa-acylated molecular species at m/z 1571 and1809 respectively (data not shown). They were doubledby smaller peaks at m/z 1587 and 1825 corresponding tomolecular species in which C12 was replaced by C12-OH.The spectrum of the mutant lipid A did not have the C16-containing hexa-acylated molecular ion peak.
Spectra obtained in the positive-ion mode (Fig. 4) gaveinformation about the location of fatty acids. In the WT lipidA spectrum, peaks were observed in the lower massregion at m/z 1143, 916, 904, 677 and 662. The peak atm/z 1143 (Fig. 4A, peak B1) corresponded to an ion con-taining one glucosamine, one phosphate, two C14-OH, oneC14 and one C16 [Mr (–H2O) = 161.16, 80, 226.36 ¥ 2,210.36 and 238.41 respectively]. According to the frag-mentation pattern already described in plasma desorptionmass spectrometry studies (Karibian et al., 1999) andobserved with reference lipids A in MALDI, the C16 couldbe localized on glucosamine-II. The peak at m/z 1143 wasabsent from the mutant lipid A spectrum in which the B1
fragment appeared at m/z 904 (1143 minus C16). In thehigher mass region of the WT lipid A spectrum, smallpeaks at m/z 1491, 1508, 1747 and 1769 were observed.They corresponded to monophosphoryl forms (Fig. 4,peak B2) of the penta- and hexa-acylated molecular spe-cies as well as a Na+ adduct of the latter. In the mutantlipid A spectrum, no hexa-acylated molecular species (m/z 1747) containing C16 was observed (Fig. 4B).
Thus, mass spectrometry determined that the C16 wason glucosamine-II, and the liberation of the C14OC16 bymild alkali treatment further localized it at position C-3¢(Fig. 4, insets). Both techniques showed the absence ofesterified C16 in the lipid A from the pagP mutant.
Bvg-dependent B. bronchiseptica pagP promoter activity
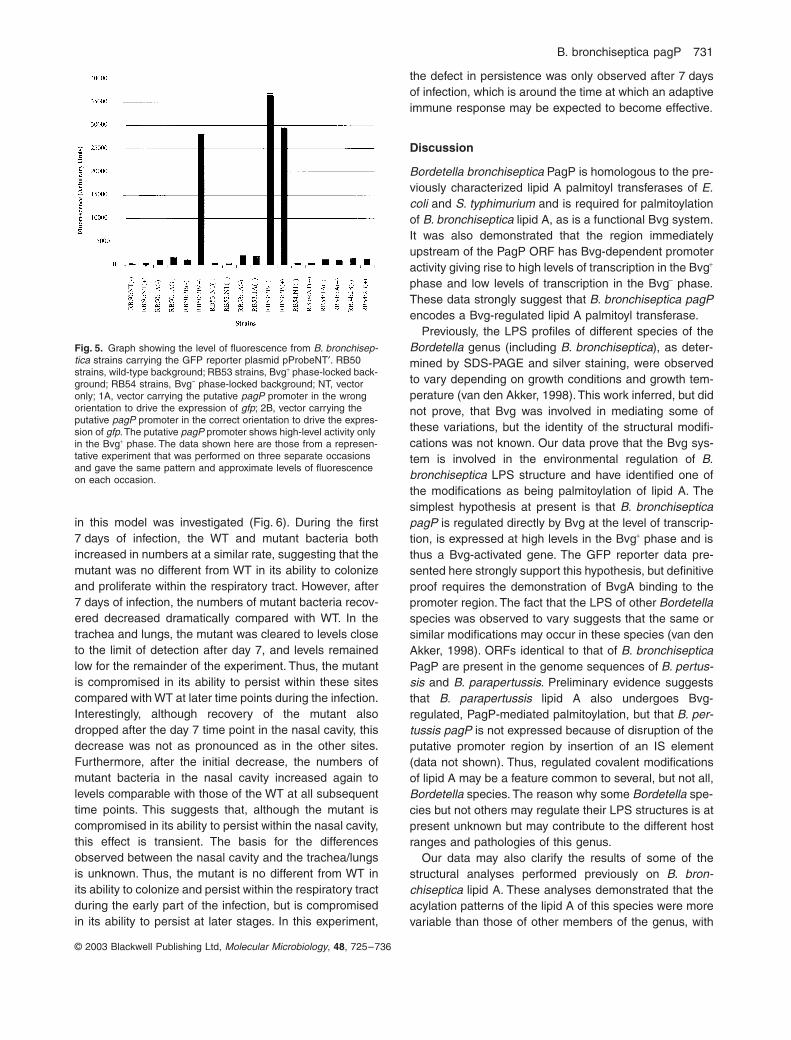
The PagP ORF is separated from the putative MgtEupstream ORF by 104 bp. Thus, it is possible that thisregion contains a promoter for PagP expression. To testthis hypothesis, the region was amplified by PCR andcloned into the green fluorescent protein (GFP) reporterplasmid, pProbeNT¢, to generate a transcriptional fusionbetween the putative promoter and gfp. pProbeNT¢ isderived from the B. bronchiseptica plasmid pBBR1MCSand thus replicates in B. bronchiseptica (Kovach et al.,1994). The putative promoter region was cloned in bothorientations within the vector. The reporter constructswere introduced into wild-type B. bronchiseptica and theBvg phase-locked mutants RB53 and RB54. Thesestrains were grown under both Bvg+ and Bvg– phase con-ditions, and the fluorescence from each strain was mea-sured (Fig. 5). Strains carrying vector alone gave very lowlevels of fluorescence that resulted from either naturalfluorescence of the bacteria or very low levels of GFPexpression from promoter sequences unrelated to thepagP gene on the vector. Strains carrying the putativepromoter region in the wrong orientation for driving tran-scription of gfp gave higher levels of fluorescence thanvector alone, regardless of the Bvg phase, which sug-gests that the cloned Bordetella DNA had some ability torecruit RNA polymerase and initiate transcription of gfp to

730 A. Preston et al.
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 48, 725–736
very low levels. Wild-type strains carrying the putativepromoter in the correct orientation for driving transcriptionof gfp demonstrated low levels of fluorescence whengrown in the Bvg– phase. Growth in the Bvg+ phaseresulted in an approximately 10-fold increase in fluores-cence over that observed for the Bvg– phase. In the Bvgphase-locked mutants, the RB53 clone gave high-levelfluorescence and the RB54 clone low-level fluorescenceregardless of the growth conditions. These data demon-strate that the putative promoter region is able to drivetranscription of the downstream gfp gene but only whenin the correct orientation. Furthermore, this promoteractivity is Bvg dependent, with high-level activity in the
Bvg+ phase. These data are in agreement with PagP-mediated lipid A palmitoylation occurring in the Bvg+
phase and suggest that regulation occurs through Bvgregulation of pagP transcription.
Bordetella bronchiseptica pagP is attenuated in a mouse model of infection
Many Bvg-regulated genes that are expressed in the Bvg+
phase have been implicated in the pathogenesis of B.bronchiseptica. A murine model of infection has beenused extensively to study B. bronchiseptica pathogenesis.Thus, the ability of the pagP mutant to colonize and persist
Fig. 4. Positive ion mode mass spectra of the Bvg+ phase lipid A from (A) B. bronchiseptica RB50 and (B) the pagP mutant of this strain. The WT lipid A contains a palmitate group that is absent from the mutant lipid A.
B. bronchiseptica pagP 731
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 48, 725–736
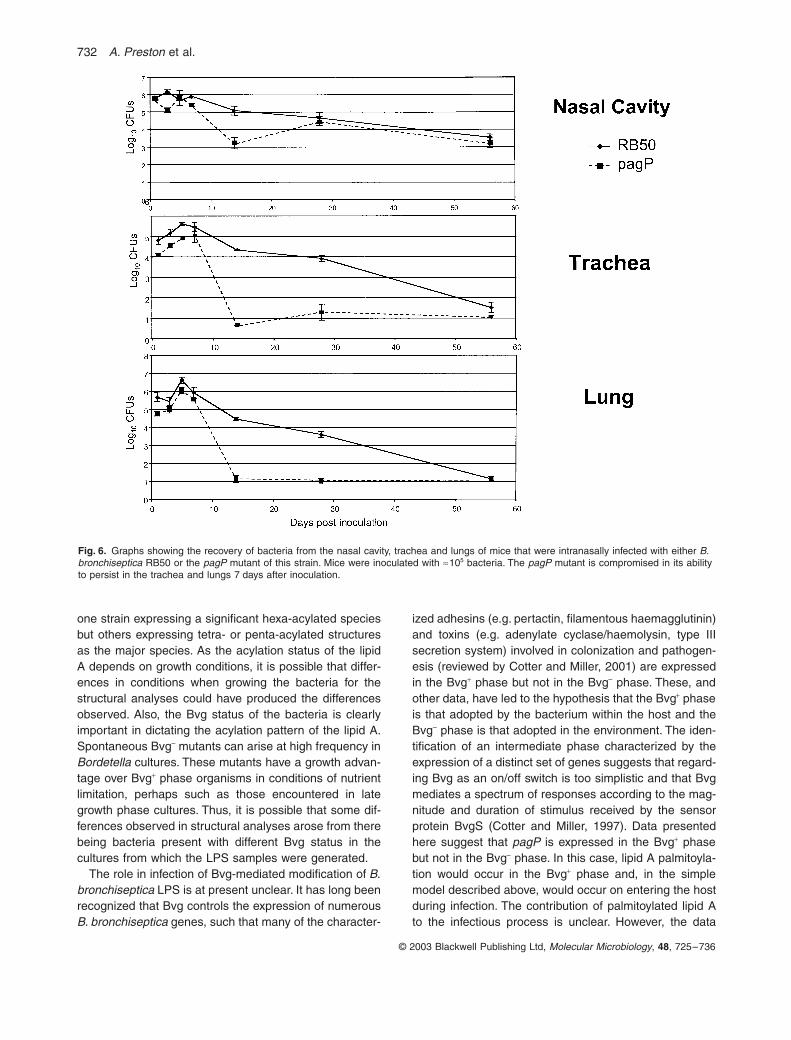
in this model was investigated (Fig. 6). During the first7 days of infection, the WT and mutant bacteria bothincreased in numbers at a similar rate, suggesting that themutant was no different from WT in its ability to colonizeand proliferate within the respiratory tract. However, after7 days of infection, the numbers of mutant bacteria recov-ered decreased dramatically compared with WT. In thetrachea and lungs, the mutant was cleared to levels closeto the limit of detection after day 7, and levels remainedlow for the remainder of the experiment. Thus, the mutantis compromised in its ability to persist within these sitescompared with WT at later time points during the infection.Interestingly, although recovery of the mutant alsodropped after the day 7 time point in the nasal cavity, thisdecrease was not as pronounced as in the other sites.Furthermore, after the initial decrease, the numbers ofmutant bacteria in the nasal cavity increased again tolevels comparable with those of the WT at all subsequenttime points. This suggests that, although the mutant iscompromised in its ability to persist within the nasal cavity,this effect is transient. The basis for the differencesobserved between the nasal cavity and the trachea/lungsis unknown. Thus, the mutant is no different from WT inits ability to colonize and persist within the respiratory tractduring the early part of the infection, but is compromisedin its ability to persist at later stages. In this experiment,
the defect in persistence was only observed after 7 daysof infection, which is around the time at which an adaptiveimmune response may be expected to become effective.
Discussion
Bordetella bronchiseptica PagP is homologous to the pre-viously characterized lipid A palmitoyl transferases of E.coli and S. typhimurium and is required for palmitoylationof B. bronchiseptica lipid A, as is a functional Bvg system.It was also demonstrated that the region immediatelyupstream of the PagP ORF has Bvg-dependent promoteractivity giving rise to high levels of transcription in the Bvg+
phase and low levels of transcription in the Bvg– phase.These data strongly suggest that B. bronchiseptica pagPencodes a Bvg-regulated lipid A palmitoyl transferase.
Previously, the LPS profiles of different species of theBordetella genus (including B. bronchiseptica), as deter-mined by SDS-PAGE and silver staining, were observedto vary depending on growth conditions and growth tem-perature (van den Akker, 1998). This work inferred, but didnot prove, that Bvg was involved in mediating some ofthese variations, but the identity of the structural modifi-cations was not known. Our data prove that the Bvg sys-tem is involved in the environmental regulation of B.bronchiseptica LPS structure and have identified one ofthe modifications as being palmitoylation of lipid A. Thesimplest hypothesis at present is that B. bronchisepticapagP is regulated directly by Bvg at the level of transcrip-tion, is expressed at high levels in the Bvg+ phase and isthus a Bvg-activated gene. The GFP reporter data pre-sented here strongly support this hypothesis, but definitiveproof requires the demonstration of BvgA binding to thepromoter region. The fact that the LPS of other Bordetellaspecies was observed to vary suggests that the same orsimilar modifications may occur in these species (van denAkker, 1998). ORFs identical to that of B. bronchisepticaPagP are present in the genome sequences of B. pertus-sis and B. parapertussis. Preliminary evidence suggeststhat B. parapertussis lipid A also undergoes Bvg-regulated, PagP-mediated palmitoylation, but that B. per-tussis pagP is not expressed because of disruption of theputative promoter region by insertion of an IS element(data not shown). Thus, regulated covalent modificationsof lipid A may be a feature common to several, but not all,Bordetella species. The reason why some Bordetella spe-cies but not others may regulate their LPS structures is atpresent unknown but may contribute to the different hostranges and pathologies of this genus.
Our data may also clarify the results of some of thestructural analyses performed previously on B. bron-chiseptica lipid A. These analyses demonstrated that theacylation patterns of the lipid A of this species were morevariable than those of other members of the genus, with
Fig. 5. Graph showing the level of fluorescence from B. bronchisep-tica strains carrying the GFP reporter plasmid pProbeNT¢. RB50 strains, wild-type background; RB53 strains, Bvg+ phase-locked back-ground; RB54 strains, Bvg– phase-locked background; NT, vector only; 1A, vector carrying the putative pagP promoter in the wrong orientation to drive the expression of gfp; 2B, vector carrying the putative pagP promoter in the correct orientation to drive the expres-sion of gfp. The putative pagP promoter shows high-level activity only in the Bvg+ phase. The data shown here are those from a represen-tative experiment that was performed on three separate occasions and gave the same pattern and approximate levels of fluorescence on each occasion.
732 A. Preston et al.
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 48, 725–736
one strain expressing a significant hexa-acylated speciesbut others expressing tetra- or penta-acylated structuresas the major species. As the acylation status of the lipidA depends on growth conditions, it is possible that differ-ences in conditions when growing the bacteria for thestructural analyses could have produced the differencesobserved. Also, the Bvg status of the bacteria is clearlyimportant in dictating the acylation pattern of the lipid A.Spontaneous Bvg– mutants can arise at high frequency inBordetella cultures. These mutants have a growth advan-tage over Bvg+ phase organisms in conditions of nutrientlimitation, perhaps such as those encountered in lategrowth phase cultures. Thus, it is possible that some dif-ferences observed in structural analyses arose from therebeing bacteria present with different Bvg status in thecultures from which the LPS samples were generated.
The role in infection of Bvg-mediated modification of B.bronchiseptica LPS is at present unclear. It has long beenrecognized that Bvg controls the expression of numerousB. bronchiseptica genes, such that many of the character-
ized adhesins (e.g. pertactin, filamentous haemagglutinin)and toxins (e.g. adenylate cyclase/haemolysin, type IIIsecretion system) involved in colonization and pathogen-esis (reviewed by Cotter and Miller, 2001) are expressedin the Bvg+ phase but not in the Bvg– phase. These, andother data, have led to the hypothesis that the Bvg+ phaseis that adopted by the bacterium within the host and theBvg– phase is that adopted in the environment. The iden-tification of an intermediate phase characterized by theexpression of a distinct set of genes suggests that regard-ing Bvg as an on/off switch is too simplistic and that Bvgmediates a spectrum of responses according to the mag-nitude and duration of stimulus received by the sensorprotein BvgS (Cotter and Miller, 1997). Data presentedhere suggest that pagP is expressed in the Bvg+ phasebut not in the Bvg– phase. In this case, lipid A palmitoyla-tion would occur in the Bvg+ phase and, in the simplemodel described above, would occur on entering the hostduring infection. The contribution of palmitoylated lipid Ato the infectious process is unclear. However, the data
Fig. 6. Graphs showing the recovery of bacteria from the nasal cavity, trachea and lungs of mice that were intranasally infected with either B. bronchiseptica RB50 or the pagP mutant of this strain. Mice were inoculated with ª105 bacteria. The pagP mutant is compromised in its ability to persist in the trachea and lungs 7 days after inoculation.

B. bronchiseptica pagP 733
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 48, 725–736
presented here suggest that palmitoylation is not requiredfor initial colonization of the respiratory tract or persistencewithin it during the early stages of infection. This is incontrast to the small amount of data regarding the role oflipid A palmitoylation in the biology of other bacteria. PagPmutants in S. typhimurium and Legionella pneumophiladisplayed increased sensitivity to cationic antimicrobialpeptide-mediated killing, suggesting that, in these spe-cies, lipid A palmitoylation may increase resistance tothese peptides (Guo et al., 1998; Robey et al., 2001).These peptides are part of the innate immune system thatis considered to be particularly important during the earlystages of infection. The B. bronchiseptica pagP mutant isnot compromised in colonization or persistence early onin infection, suggesting that lipid A palmitoylation in thisspecies is not required for resistance to early innatedefences. In fact, B. bronchiseptica pagP is more resistantto killing by the peptide cecropin P1 than WT (data notshown), indicating that lipid A palmitoylation plays adifferent role in B. bronchiseptica than it does in otherbacteria.
That B. bronchiseptica pagP is attenuated around thetime at which an adaptive immune response mightbecome effective suggests that either the mutant is moresusceptible than WT to killing by effector mechanisms ofthe adaptive immune response or the mutant triggers amore effective adaptive immune response than the WT.Which (if either) of these scenarios is correct is at presentunclear. However, LPS is critical for stimulating signaltransduction in mammalian cells that is an integral part ofthe host response to infection. The nature of this signaltransduction is important to the interaction between hostand pathogen that dictates the outcome of infection. Theacyl groups of lipid A are particularly involved in elicitingthese signal transduction responses. Thus, it is possiblethat modification of lipid A acylation will alter the interac-tion between host and pathogen through alteration of thehost signal transduction response. Several reports haveindicated that mammalian cells do produce differentresponses to palmitoylated lipid A compared with non-palmitoylated lipid A. Chemically synthesized, palmitoy-lated, hepta-acylated lipid A acted as an antagonisttowards the activity of hexa-acylated lipid A with regard tothe induction of tumour necrosis factor (TNF)a inductionand NFkB activation in human-derived THP-1 cells, sug-gesting that palmitoylation may decrease mammalian cellresponses (Tanamoto and Azumi, 2000). On the otherhand, palmitoylated Pseudomonas aeruginosa LPSinduced significantly more interleukin (IL)-8 from humanumbilical cord endothelial cells (HUVECs) than did non-palmitoylated LPS, suggesting that, in this species, palm-itoylation may stimulate increased responses (Ernst et al.,1999). It is thus possible that lipid A palmitoylation damp-ens the adaptive immune response that is triggered by B.
bronchiseptica, decreasing the effectiveness of theresponse. How this may occur is unclear, but the role oflipid A in the stimulation of signal transduction in host cellsmakes alteration of this process an attractive hypothesis.In this case, B. bronchiseptica pagP might be unable tosuppress the immune response, resulting in its earlierclearance from the respiratory tract. This would representa novel immune modulation mechanism.
The work presented here is the first characterization atthe molecular level of regulated modification of BordetellaLPS. The involvement of Bvg in this process stronglysuggests that LPS modification forms part of the adapta-tion of this organism to changing environmental conditionsincluding those encountered upon entry into the host dur-ing the course of infection. This work also demonstratesthat Bvg-mediated modification of LPS is required for fullvirulence of B. bronchiseptica.
Experimental procedures
Bacterial strains and plasmids
Bordetella bronchiseptica strain RB50 is a wild-type strain.RB53 and RB54 are Bvg phase-locked mutants, and RB50iand RB53i are Bvg-intermediate phase mutants that havebeen described previously (Cotter and Miller, 1994; 1997).These strains were generously provided by J. F. Miller, UCLA.Whole-genome sequencing of RB50 has been completedrecently (http://www.sanger.ac.uk). Escherichia coli XL1-Blue(Stratagene) was used for cloning and maintenance of plas-mids. E. coli SM10 (Simon et al., 1983) or CC118lpir wasused as donor strains in conjugations. pUC18 Ready-To-Go™ (Pharmacia) or pCR2.1 topo (Invitrogen) was used toclone PCR products. pBluescript-II SK+ (Stratagene) wasused as a general cloning vector. pEX100T (Schweizer andHoang, 1995) was used as a suicide vector for allelicexchange mutagenesis in B. bronchiseptica. pBBR1kan is aderivative of pBBR1MCS in which the chloramphenicol resis-tance cassette has been replaced with a kanamycin resis-tance cassette. pBBR1MCS is a broad-host-range vector thatis capable of replication in Bordetella (Kovach et al., 1994).pBBR1kan was used to introduce a complementing copy ofthe pagP gene into the B. bronchiseptica pagP mutant.pProbeNT¢ (Miller et al., 2000) is a promoter trap vector thatcreates transcriptional fusions between cloned promoter frag-ments and a gfp reporter gene.
Growth media and conditions
Bordetella bronchiseptica was grown on Bordet–Gengouagar (Difco) supplemented with 15% defibrinated horse blood(TCS Biologicals) at 37∞C. E. coli was grown on LB agar at37∞C. For liquid culture, B. bronchiseptica and E. coli weregrown in LB broth at 37∞C with aeration. For growth of B.bronchiseptica in the Bvg– phase, 50 mM MgSO4 was addedto growth medium. Streptomycin (200 mg ml-1), ampicillin(100 mg ml-1), erythromycin (10 mg ml-1) and kanamycin(50 mg ml-1) were used where appropriate.

734 A. Preston et al.
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 48, 725–736
Chemicals and reagents
Chemicals and reagents were from Difco or Sigma.
DNA purification
Plasmid DNA was purified using a Qiagen plasmid DNApurification kit according to the manufacturer’s instructions.Genomic DNA was purified using the agarose plug method(Sambrook et al., 1989).
DNA manipulation
DNA manipulations were performed according to standardmethods. DNA restriction and modifying enzymes were fromNew England Biolabs, Roche or Gibco Life Technologies.
PCR and primers
Genomic DNA template was made by resuspending plate-grown bacteria in 0.5 ml of water, boiling in a water bath for5 min, spinning at top speed in a bench-top microcentrifugefor 2 min and taking 0.2 ml of supernatant. A sample of 1 mlof supernatant was used per PCR. Each 50 ml PCR com-prised genomic DNA template, buffer as directed by the man-ufacturer, dNTPs (25 mM each), 20 ng of each primer, 5%(v/v) DMSO, 5 mM MgCl2 and 2.5 units of Taq DNA poly-merase (Promega). Primers used to amplify the B. bron-chiseptica pagP locus were PagPF (5¢-GCGTTTCTCATGTCCATC-3¢) and PagPR (5¢-TCAACGAGCATGGCATTG-3¢).Primers used to amplify the putative pagP promoter wereProm1 (5¢-ACGATCTTCCTACTGTAG-3¢) and Prom 2 (5¢-ATGCTGCGCTAACGGGAA-3¢). Primers used to amplifypagP with its putative promoter for construction of the com-plementation construct were Prom1 and PagPR. PCRs wereincubated at 94∞C for 5 min followed by 30 cycles of 94∞C for75 s, 60∞C for 75 s and 72∞C for 90 s, followed by a final stepof 72∞C for 7 min.
Complementation of the pagP mutation
The region encompassing the putative pagP promoter andthe pagP coding sequence was amplified by PCR (seeabove), and the PCR product was cloned into pCR2.1 topo.The insert was excised by digestion with BamHI and XbaI,and the fragment was subcloned into BamHI–XbaI-digestedpBBR1kan. This construct was introduced into B. bronchisep-tica pagP by conjugation.
SDS-PAGE/silver stain analysis of LPS
Whole-cell lysate LPS samples were prepared and analysedby SDS-PAGE/silver staining as described previously(Preston et al., 1996).
LPS purification
LPS was purified according to a protocol based on thatdescribed previously (Johnson and Perry, 1976). Briefly, 4 l
of overnight bacterial cultures were pelleted at 18 500 g. Thepellets were washed three times with distilled water (dH2O).The entire bacterial pellet was resuspended in a small vol-ume of dH2O to produce a thick slurry. This was then soni-cated with three pulses of 30 s each before the addition oflysozyme to a concentration of 0.2% (w/v) and incubationovernight on a tumbling rack at room temperature. The slurrywas then incubated for a further 24 h at 4∞C without agitation,sonicated for 30 s and then 40 U of micrococcal nucleaseand RNase A (100 mg ml-1) were added. The slurry was thenincubated for 48 h at 4∞C. A volume of phenol equivalent tothe slurry volume was melted, and both slurry and phenolwere equilibrated to 65∞C, mixed, sonicated for 15 s and thenincubated at 65∞C for 30 min. The mixture was cooled on icefor 30 min to enhance phase separation before centrifugingat 20 000 g for 30 min. The aqueous phase was collected andthen dialysed extensively against dH2O to remove all tracesof phenol. The resultant LPS preparation was precipitated bythe addition of 1/10th volume 3 M NaAc, pH 5.4, and twovolumes of ethanol with incubation at -20∞C overnight. TheLPS was pelleted by centrifugation at 15 000 g, and the pelletwas air dried overnight. Finally, the pellet was resuspendedin a small volume of dH2O and lyophilized to give the finalLPS product.
Structural analysis of LPS
Preparation of lipid As. The LPS samples were resus-pended to a concentration of 10 mg ml-1 and then purified byextraction with chloroform–methanol (1:2) to remove phos-pholipids and free fatty acids. The dried, extracted LPSs weresuspended in water and recovered by centrifugation at90 000 g for 45 min, and this was repeated until thin-layerchromatography (TLC) and UV spectra showed no detectablecontaminants. Lipid A was prepared by hydrolysing LPS in20 mM NaAc, pH 4.5–1% SDS at 100∞C for 1 h followed bylyophilization. Detergent was removed by repeated extractionwith acidified ethanol, the LPS/lipid A was recovered by cen-trifugation, dried under a stream of N2, and the lipid A in thedried pellet was extracted with chloroform–methanol–water(12:6:1) (Caroff et al., 1988).
Fatty acid analyses. To determine the total lipid composition,fatty acids were released as described previously (Haeffneret al., 1977). To identify fatty acids substituting glucosaminepositions C-3 and C-3¢, 1 mg of lipid A was treated with 0.2 MNaOH (0.5 ml) at 37∞C for 5 min (Karibian et al., 1999). Thereleased fatty acids were extracted with ethyl acetate, meth-ylated with diazomethane and identified by gas chromatog-raphy (GC) on an HP5 column (30 m ¥ 0.32 mm) with aprogramme of 150∞C to 300∞C at 6∞ min-1. GC/mass spec-trometry (MS) was performed on a DB5ms capillary column(30 m) coupled to a Finnigan MAT 95.S electron-impact massspectrometer to characterize the fatty acids.
Mass spectrometry. MALDI/MS was carried out on a Persep-tive Voyager STR model (PE Biosystem) time-of-flight massspectrometer (IBBMC). Gentisic acid (2,5-dihydroxybenzoicacid) was used as a matrix: a suspension of lipid A in amixture of chloroform–methanol–water (1 mg ml-1) was

B. bronchiseptica pagP 735
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 48, 725–736
desalted with a few grains of Dowex 50 W-X8(H+), 0.5 ml wasdeposited on the target, mixed with 0.5 ml of the matrix sus-pended at 10 mg ml-1 in either water or 0.1 M aqueous citricacid (Therisod et al., 2001) and dried. Analyte ions weredesorbed from the matrix with pulses from a 337 nm nitrogenlaser. Spectra were obtained in the negative- and positive-ion modes at 20 kV with an average of 128 pulses.
Assay of pagP promoter activity
The putative promoter region was amplified by PCR andcloned into pProbeNT¢. The construct was introduced into B.bronchiseptica by conjugation. GFP activity was assayed byresuspending bacteria from plates in PBS to an OD600 = 1.0.Aliquots of 200 ml were transferred to the wells of a 96-wellflat-bottomed microtitre plate, and fluorescence from tripli-cate samples was measured on a CytoFluor series 400 flu-orescent plate reader (Perseptive Biosystems) usingexcitation at 480 nm and measuring emission at 530 nm. Thefluorescence of PBS alone was subtracted from each samplevalue. The average fluorescence and standard deviation ofthe triplicate samples is shown in Fig. 5.
Animal experiments
Female BALB/c mice, 4–6 weeks old, were obtained fromCharles River Laboratories. Bacteria were grown to mid-logphase in Stainer–Scholte medium (Stainer and Scholte,1970), pelleted and resuspended in PBS. Mice lightlysedated with isofluorane were inoculated with ª 5 ¥ 105 bac-teria by pipetting 50 ml of inoculum onto the tip of the externalnares. Groups of four animals were sacrificed on each ofdays 0, 3, 5, 7, 14, 21, 28 or 50 after inoculation. Colonizationof various organs was quantified by homogenizing each tis-sue in PBS, plating aliquots onto BG-blood agar and countingcolonies after 2 days of incubation at 37∞C. Animals werehandled in accordance with institutional guidelines. Statisticalsignificance was determined using an unpaired t-test.
Acknowledgements
This work was supported by The Wellcome Trust, programmegrant number 054588 (A.P., E.M., E.T. and D.J.M.) and grantsfrom Neose Corporation, Pennsylvania Department of Agri-culture grant ME44067 and US Department of Agriculturegrant 2002-35204-11684 (J.P. and E.T.H.).
References
van den Akker, W.M. (1998) Lipopolysaccharide expressionwithin the genus Bordetella: influence of temperature andphase variation. Microbiology 144: 1527–1535.
Amano, K., Fukushi, K., and Watanabe, M. (1990) Biochem-ical and immunological comparison of lipopolysaccharidesfrom Bordetella species. J Gen Microbiol 136: 481–487.
Aussel, L., Brisson, J.R., Perry, M.B., and Caroff, M. (2000)Structure of the lipid A of Bordetella hinzii ATCC 51730.Rapid Commun Mass Spectrom 14: 595–599.
Bishop, R.E., Gibbons, H.S., Guina, T., Trent, M.S., Miller,
S.I., and Raetz, C.R. (2000) Transfer of palmitate fromphospholipids to lipid A in outer membranes of gram-negative bacteria. EMBO J 19: 5071–5080.
Burns, E.H., Jr, Norman, J.M., Hatcher, M.D., and Bemis,D.A. (1993) Fimbriae and determination of host speciesspecificity of Bordetella bronchiseptica. J Clin Microbiol 31:1838–1844.
Burns, V.C., Pishko, E.J., Preston, A., Maskell, D.J., andHarvill, E.T. (2003) Role of Bordetella O antigen in respi-ratory tract infection. Infect Immun 71: 86–94.
Caroff, M., Tacken, A., and Szabo, L. (1988) Detergent-accelerated hydrolysis of bacterial endotoxins and deter-mination of the anomeric configuration of the glycosylphosphate present in the ‘isolated lipid A’ fragment of theBordetella pertussis endotoxin. Carbohyd Res 175: 273–282.
Caroff, M., Chaby, R., Karibian, D., Perry, J., Deprun, C., andSzabo, L. (1990) Variations in the carbohydrate regions ofBordetella pertussis lipopolysaccharides: electrophoretic,serological, and structural features. J Bacteriol 172: 1121–1128.
Caroff, M., Brisson, J., Martin, A., and Karibian, D. (2000)Structure of the Bordetella pertussis 1414 endotoxin.FEBS Lett 477: 8–14.
Caroff, M., Aussel, L., Zarrouk, H., Martin, A., Richards, J.C.,Therisod, H., et al. (2001) Structural variability and origi-nality of the Bordetella endotoxins. J Endotoxin Res 7: 63–68.
Chaby, R., and Caroff, M. (1988) Lipopolysaccharide of Bor-detella pertussis endotoxin. In Pathogenesis and Immunityin Pertussis. Wardlaw, A.C., and Parton, R. (eds). NewYork: John Wiley, pp. 247–271.
Cotter, P.A., and Miller, J.F. (1994) BvgAS-mediated signal-transduction – analysis of phase-locked regulatory mutantsof Bordetella bronchiseptica in a rabbit model. InfectImmun 62: 3381–3390.
Cotter, P.A., and Miller, J.F. (1997) A mutation in the Borde-tella bronchiseptica bvgS gene results in reduced virulenceand increased resistance to starvation, and identifies a newclass of Bvg-regulated antigens. Mol Microbiol 24: 671–685.
Cotter, P.A., and Miller, J.F. (2001) Bordetella. In Principlesof Bacterial Pathogenesis. Groisman, E. (ed.). San Diego,CA: Academic Press, pp. 619–674.
Ernst, R.K., Yi, E.C., Guo, L., Lim, K.B., Burns, J.L., Hackett,M., and Miller, S.I. (1999) Specific lipopolysaccharidefound in cystic fibrosis airway Pseudomonas aeruginosa.Science 286: 1561–1565.
Gueirard, P., Weber, C., Lecoustumier, A., and Guiso, N.(1995) Human Bordetella bronchiseptica infection relatedto contact with infected animals – persistence of bacteriain host. J Clin Microbiol 33: 2002–2006.
Gunn, J.S. (2001) Bacterial modification of LPS and resis-tance to antimicrobial peptides. J Endotoxin Res 7: 57–62.
Guo, L., Lim, K.B., Poduje, C.M., Daniel, M., Gunn, J.S.,Hackett, M., and Miller, S.I. (1998) Lipid A acylation andbacterial resistance against vertebrate antimicrobial pep-tides. Cell 95: 189–198.
Haeffner, N., Chaby, R., and Szabo, L. (1977) Identificationof 2-methyl-3-hydroxydecanoic and 2-methyl-3-hydroxytetradecanoic acids in the ‘lipid X’ fraction of the

736 A. Preston et al.
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 48, 725–736
Bordetella pertussis endotoxin. Eur J Biochem 77: 535–544.
Harvill, E.T., Preston, A., Cotter, P.A., Allen, A.G., Maskell,D.J., and Miller, J.F. (2000) Multiple roles for Bordetellalipopolysaccharide molecules during respiratory tract infec-tion. Infect Immun 68: 6720–6728.
Johnson, K.G., and Perry, M.B. (1976) Improved techniquesfor the preparation of bacterial lipopolysaccharides. Can JMicrobiol 22: 29–34.
Karibian, D., Brunelle, A., Aussel, L., and Caroff, M. (1999)252Cf-plasma desorption mass spectrometry of unmodi-fied lipid A: fragmentation patterns and localization of fattyacids. Rapid Commun Mass Spectrom 13: 2252–2259.
Kovach, M.E., Phillips, R.W., Elzer, P.H., Roop, R.M., II, andPeterson, K.M. (1994) pBBR1MCS: a broad-host-rangecloning vector. Biotechniques 16: 800–801.
Miller, W.G., Leveau, J.H., and Lindow, S.E. (2000) Improvedgfp and inaZ broad-host-range promoter-probe vectors.Mol Plant–Microbe Interact 13: 1243–1250.
Peppler, M.S. (1984) Two physically and serologically distinctlipopolysaccharide profiles in strains of Bordetella pertus-sis and their phenotype variants. Infect Immun 43: 224–232.
Preston, A., and Maskell, D.J. (2002) Molecular genetics androle in infection of environmentally regulated lipopolysac-charide expression. Int J Med Microbiol 292: 1–9.
Preston, A., Maskell, D., Johnson, A., and Moxon, E.R.(1996) Altered lipopolysaccharide characteristic of the I69phenotype in Haemophilus influenzae results from muta-tions in a novel gene, isn. J Bacteriol 178: 396–402.
Raetz, C.R.H. (2001) Regulated covalent modifications oflipid A. J Endotoxin Res 7: 73–78.
Reina, J., Bassa, A., Llompart, I., Borrell, N., Gomez, J., andSerra, A. (1991) Pneumonia caused by Bordetella bron-chiseptica in a patient with a thoracic trauma. Infection 19:46–48.
Robey, M., O’Connell, W., and Cianciotto, N.P. (2001) Iden-tification of Legionella pneumophila rcp, a pagP-like genethat confers resistance to cationic antimicrobial peptidesand promotes intracellular infection. Infect Immun 69:4276–4286.
Rutter, J.M. (1981) Quantitative observations on Bordetellabronchiseptica infection in atrophic rhinitis of pigs. VetRecord 108: 451–454.
Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989) Molecu-lar Cloning: A Laboratory Manual. Cold Spring Harbor, NY:Cold Spring Harbor Laboratory Press.
Schweizer, H.P., and Hoang, T.T. (1995) An improved sys-tem for gene replacement and XylE fusion analysis inPseudomonas aeruginosa. Gene 158: 15–22.
Simon, R., Priefer, U., and Pühler, A. (1983) A broad hostrange mobilisation system for in vivo genetic engineering:transposon mutagenesis in Gram negative bacteria. Bio-techniques 1: 784–791.
Stainer, D.W., and Scholte, M.J. (1970) A simple chemicallydefined medium for the production of phase I Bordetellapertussis. J Gen Microbiol 63: 211–220.
Tamion, F., Girault, C., Chevron, V., Pestel, M., and Bon-marchand, G. (1996) Bordetella bronchiseptica pneumoniawith shock in an immunocompetent patient. Scand J InfectDis 28: 197–198.
Tanamoto, K., and Azumi, S. (2000) Salmonella-type heptaa-cylated lipid A is inactive and acts as an antagonist oflipopolysaccharide action on human line cells. J Immunol164: 3149–3156.
Therisod, H., Labas, V., and Caroff, M. (2001) Direct microex-traction and analysis of rough-type lipopolysaccharides bycombined thin-layer chromatography and MALDI massspectrometry. Anal Chem 73: 3804–3807.
Thrusfield, M.V., Aitken, C.G.G., and Muirhead, R.H. (1991a)A field investigation of kennel cough – efficacy of differenttreatments. J Small Anim Pract 32: 455–459.
Thrusfield, M.V., Aitken, C.G.G., and Muirhead, R.H.(1991b) A field investigation of kennel cough – incubationperiod and clinical signs. J Small Anim Pract 32: 215–220.
Watanabe, M., Takimoto, H., Kumazawa, Y., and Amano, K.(1990) Biological properties of lipopolysaccharides fromBordetella species. J Gen Microbiol 136: 489–493.
West, N.P., Jungnitz, H., Fitter, J.T., McArthur, J.D., Guzman,C.A., and Walker, M.J. (2000) Role of phosphoglucomu-tase of Bordetella bronchiseptica in lipopolysaccharide bio-synthesis and virulence. Infect Immun 68: 4673–4680.
Zarrouk, H., Karibian, D., Bodie, S., Perry, M.B., Richards,J.C., and Caroff, M. (1997) Structural characterization ofthe lipids A of three Bordetella bronchiseptica strains: vari-ability of fatty acid substitution. J Bacteriol 179: 3756–3760.