bradykinin receptors in isolated intestinal smooth muscle
TRANSCRIPT

Drug Development Research 2:l-16 (1982)
Current Trends Review
Bradykinin Receptors in Isolated Intestinal Smooth Muscle Arleen Chase and Ladislav Volicer
Department of Pharmacology, Boston University School of Medicine, and E, N. Rogers Memorial Veterans Hospital, Bedford, Massachusetts
Introduction Bradykinin Receptor Studies
Structure-Activity Studies Direct Binding Studies Characterization of Drug Receptor Response
Differential Specificity and Sensitivity of Various Tissues Enzymatic Degradation-the Use of Bradykinin Potentiating Peptides Sex of the Animal Receptor-Effector Coupling
Discussion Conclusion Acknowledgments References
ABSTRACT
Chase, A., and Vollcer, L.: Bradykinin receptors in isolated intestinal smooth muscle. Drug Dev. Res. 2:OOl-016, 1982.
Although bradykinin has been long studied, its biological relevance is still unclear. The enzymes that form and degrade bradykinin appear to be numerous and ubiquitous, and its “receptors” are both species and tissue specific. Therefore, despite the fact that a multitude of compounds have been synthesized and screened as receptor antagonists, to date only one specific antagonist has been identified and it is only active in the rabbit aorta. This review is designed to present what is known about bradykinin receptors in intestinal smooth muscle. Relevant background material and a discussion of evidence for bradykinin’s mech- anism of action are also presented. Key words: Bradykininlsmooth muscle receptors, receptor studies
Received final version July 14, 1980; accepted December 10, 1980.
Address reprint requests to Arleen Chase, Instrumentation Laboratory, Inc., 113 Hartwell Avenue, Lexington, MA 02173.
0272-4391/82/0201-01$04.50 0 1982 Alan R. Liss, Inc.

2 Chase and Volicer
INTRODUCTION
This review is divided into three major sections: 1. Definition of some relevant nomenclature and outlines of the major synthetic and degradative pathways for bradykinin; 2. characterization of the bradykinin receptors in intestinal smooth muscle; and 3. biochemical evidence for the mechanisms of action of bradykinin intestinal smooth muscle.
Bradykinin is a nonapeptide with the following structure:
H-Arg'-Pro2-Pro3-Gly4-Phe5-Ser6-Pro7-Phes-Argg-OH.
It belongs to the group of polypeptides called kinins. Kinin is a descriptive term for polypeptides with either common structural features, regardless of their biological activity, or for peptides with a specific profile of biological activities. Kinins induce hypotension, increase blood flow, increase capillary permeability, and produce pain. They can be identified by their ability to contract or relax specific intestinal smooth muscle preparations. Bradykinin is one of three kinins that have been identified in man. The other two kinins are metlys-bradykinin and lys-bradykinin (kallidin). The
TABLE 1. Relative Activities of Kinins in Man*
Guinea pig ileum Rat uterus Rat duodenum Agonist effect" activity effect activity effecta activity
Bradykinin C 1 C 1 R 1 Ly s-bradykinin C 0.3 C 0.6 R 0.5 Met-lys-bradykinin C 0.1-0.3 C 0.3 R 0.25 Threshold for bradykinin (not 1 .O ngiml 0.03 ng/ml 0.10 ngiml defined)
T, contraction; R, relaxation. *From Erdos [ 19661,
INACTIVE OR PREKALLIKREIN
PANCREAS
COLON, ETC.
\
ACTIVE KALLIKREINS
SEX GLANDS KIDNEY
PREKALLIKREIN KALLIKREIN
INHIBITORS ACT I VATORS \ i -=b" KININOGENS
KlN lNS
Fig. 1. A simplified diagram of the kallikrein-kininogen-kinin System (modified from Schachter [ 19791). a) Prekallikrein activators include: Hageman factor, trypsin. b) Kallikrein inhibitors include: Trasylol, CI esterase inhibitor, a2-macroglobulin, al-antitrypsin, antithrombin 111.
Bradykinin Receptors in Intestinal Muscle 3
PRE-HAGEMAN FACTOR
c * - - - - - - - - - - - - - - - - - I I I I
ACTIVE H.F.
PRE-PTA PLASMINOGEN
PRE- ACTIVATOR I
I I
* I
PTA (XI) I PLASMINOGEN ACTIVATOR
PRE- KA LLI K RE I NS I I
INTRINSIC CLOTTING
SYSTEM KALLIKREINS
PLASMINOGEN
PCASMIN (FAST)
(PLASMA)
t METLYS - BK
L Y S ~ K I
(T ISSUES)
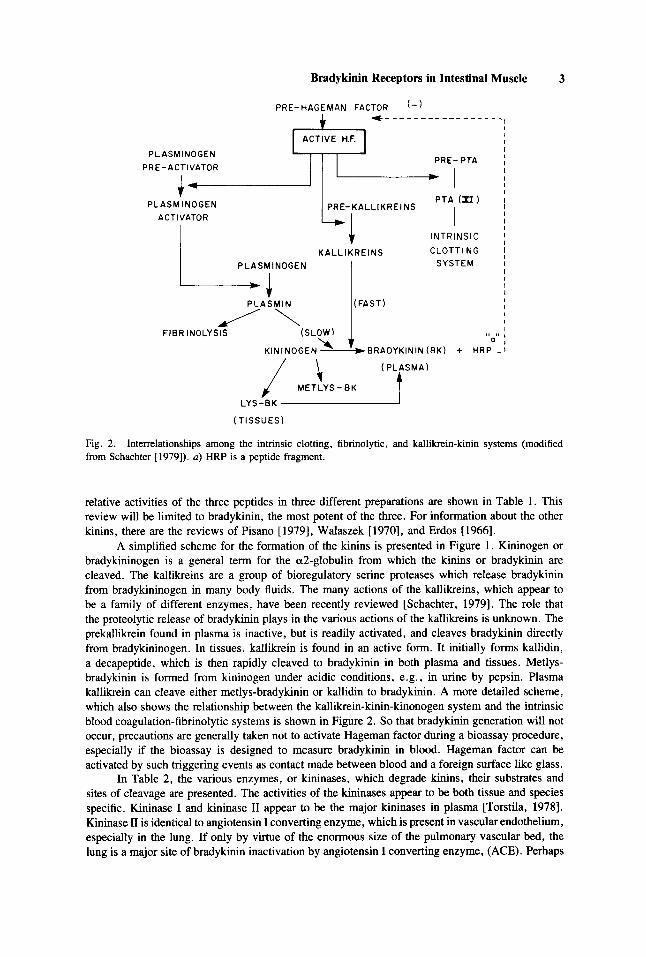
Fig. 2. from Schachter [1979]). a) HRP is a peptide fragment.
Interrelationships among the intrinsic clotting, fibrinolytic, and kallikrein-kinin systems (modified
relative activities of the three peptides in three different preparations are shown in Table 1. This review will be limited to bradykinin, the most potent of the three. For information about the other kinins, there are the reviews of Pisano [1979], Walaszek [1970], and Erdos [1966].
A simplified scheme for the formation of the kinins is presented in Figure 1. Kininogen or bradykininogen is a general term for the a2-globulin from which the kinins or bradykinin are cleaved. The kallikreins are a group of bioregulatory serine proteases which release bradykinin from bradykininogen in many body fluids. The many actions of the kallikreins, which appear to be a family of different enzymes, have been recently reviewed [Schachter, 19791. The role that the proteolytic release of bradykinin plays in the various actions of the kallikreins is unknown. The prekallikrein found in plasma is inactive, but is readily activated, and cleaves bradykinin directly from bradykininogen. In tissues, kallikrein is found in an active form. It initially forms kallidin, a decapeptide, which is then rapidly cleaved to bradykinin in both plasma and tissues. Metlys- bradykinin is formed from kininogen under acidic conditions, e.g., in urine by pepsin. Plasma kallikrein can cleave either metlys-bradykinin or kallidin to bradykinin. A more detailed scheme, which also shows the relationship between the kallikrein-kinin-kinonogen system and the intrinsic blood coagulation-fibrinolytic systems is shown in Figure 2. So that bradykinin generation will not occur, precautions are generally taken not to activate Hageman factor during a bioassay procedure, especially if the bioassay is designed to measure bradykinin in blood. Hageman factor can be activated by such triggering events as contact made between blood and a foreign surface like glass.
In Table 2, the various enzymes, or kininases, which degrade kinins, their substrates and sites of cleavage are presented. The activities of the kininases appear to be both tissue and species specific. Kininase I and kininase I1 appear to be the major kininases in plasma [Torstila, 19781. Kininase I1 is identical to angiotensin I converting enzyme, which is present in vascular endothelium, especially in the lung. If only by virtue of the enormous size of the pulmonary vascular bed, the lung is a major site of bradykinin inactivation by angiotensin I converting enzyme, (ACE). Perhaps

4 Chase and Volicer
TABLE 2. Kiniiases*
Bradykinin (BK) : Arg'-F'roz-Pro3-Gly4-Phe5-Se~-Pro7-Phes-Arg9-OH Lys-bradykinin :
Metlys-bradykinin: H-Met'-Lys2-Arg3-Pro4-Pro5-Gly6-Phe7-Se$-Prog-e'o-Arg' '-OH
H-Lys I -ArgZ-Pro3-Pro4-Gly5-Phe6-Ser7-Pro8-Phe9-Arg'o-OH (Kallidin)
Enzyme Substrate Site(s) of cleavage
Trypsin
Chymotrypsin
Carboxypeptidase A Carboxypeptidase B
Kininase I" Carboxypeptidase
(Kidney) Catheptic carboxypeptidase (spleen) Carboxycathepsinb Aminopeptidase
(purified and plasma)
(purified and erythrocyte)
(identical to angiotensin I)
converting enzyme (vascular endothe.
(pancreas)
Prolidase
Kininase 11'
Lys-BK and Metlys-BK Metlys-BK and BK BK BK
BK
BK BK only Lys-BK and Metlys-BK
only BK
BK
angiotensin I
LysVArgZ LysZiArg3 Phe7/SeP, Phe"/Arg" Phe'ISefi, Phe8/Arg9 Pro7/Phe8, after removal of Arg' Phe8/Arg9
Phe*/ Arg'
Pro7/Phe8, after removal of Arg' Pro7/Phe8 Lys '/ArgZ Met'/Lys2, and Lys2/Arg3
Arg'/Pro2
sequential: Pro'iPhe', then Phe5/Ser6
Asp- Arg-Val-Tyr-Ile-His-Pro-Phe/His-Leu
lium, esp, lung)
"Also called: carboxypeptidase N (plasma), or arginine carboxypeptidase. bAlso called: "enzyme in kidney cortex", or carboxydipeptidylpeptide hydrolase. "Also called: peptidyl dipeptide hydrolase, or cathepsin A. *Modified from Erdos [ 19661.
with the development of specific antagonists for this and the number of other catabolic enzymes which act on bradykinin, the relative importance of each in particular tissues will be discovered. The multiple pathways for the release and degradation of bradykinin have greatly confounded the progress of bradykinin research. Still unclear are its physiological role and biochemical mechanisms of action. Even the identification of specific bradykinin receptor antagonists has been difficult. For example, in in vivo studies, it has been found that bradykinin exhibits a different spectrum of activity, or much less activity, when injected IV as opposed to IA, primarily because of the effects of the kininases .
Ferreira [1965] discovered that venom from the Bothrops jururucu, a South American pit viper, contained a mixture of peptides that potentiated the effects of bradykinin by inhibiting kininases activity. A number of the peptides have been isolated, their structures determined, and their angiotensin I converting enzyme (ACE) inhibitory activities quantified [Ferreira et al., 1970; Soffer, 1976; Stewart, 19791. The names and structures of a few of these peptides are presented in Table 3. They are frequently used in bioassays and receptor studies to inhibit the kininases [Ferreira and Vane, 19671.

Bradykinin Receptors in Intestinal Muscle 5
TABLE 3. Bradykinin Potentiating Peptides
Source Names Structure
Snake venom BPP5,, or V-3A or Pyr-Lys-Trp- Ala-Pro
Snake venom BPPg,, or V-6-1, or Pyr-Trp-Pro- Arg-Pro-Gln-Ile-Pro-Pro SQ 20.475 (a pentapeptide)
SQ 20.881 or (a nonapeptide) Teprotide
Tryptic A-VIJ Val-Glu-Ser-Set-Ly s hydrolysate (a pentapeptide) of plasma
Another class of bradykinin potentiating compound includes those which contain a sulfhydryl group. A number of sulfhydryl-containing compounds inhibit bradykinin degradation [Iso et al., 19791. One very potent angiotensin converting enzyme inhibitor which contains a thiol group is SQ 14.225. In this type of compound, the thiol group is thought to bind to the zinc residue in the active site of ACE to inhibit the enzyme, and thereby potentiate bradykinin [Iso et al., 1979; Murphy et al., 1978; Stewart, 19791.
Two other kininase inhibitors have been evaluated with respect to their ability to inhibit kininase I or kininase 11. Trasylol has been found to only partially inhibit kininase I, and does not inhibit kininase 11; whereas 1, 10-phenanthroline inhibits both kininase I and kininase II (Trautsc- hold, 19701. These compounds have also been used in assays of bradykinin to inhibit kininase activity.
Kinins have been implicated in pathological conditions including: carcinoid syndrome [Oates and Melmon, 19663, various shock states (hemorrhagic, anaphylactic, endotoxin), acute pancrea- titis, “dumping syndrome,” inflammation of the joints, and migraine headaches [Roberts and Mashford, 19721. Recently, the role of the kinins in acute myocardial infarction was investigated [Torstila, 19781. Its role in hypertension via its actions on the cardiovascular and renal systems, if any, is currently under study [Stewart, 1979; Torstila, 19781. The significance of the role of bradykinin in these and other pathological conditions is yet unclear as is its role in normal phys- iological processes. The main sources of difficulty are that the enzymes which release and degrade bradykinin appear to be numerous and ubiquitous, and that its “receptors” are both species and tissue specific. Despite the fact that a multitude of compounds have been synthesized and screened as receptor antagonists, to date only one specific antagonist has been identified, and it is active only in the rabbit aorta [Regoli et al., 19771. New drug development in the area of bradykinin research is required to address the need for specific antagonists at the bradykinin receptors in specific tissues and species. There is also a need to decipher the relative importance of each of the enzymes which form and degrade bradykinin in various tissues and under various conditions. For this purpose it is desirable to have specific antagonists for each of the enzymes. Perhaps with these new pharmacologic probes, the physiological role of bradykinin and the significance of its presence in diverse pathological conditions will be clarified.
Of the multitude of effects attributed to bradykinin, a number of them have been used as the basis of bioassays. One review of assay methods using kinins is that of Trautschold [1970]. Intestinal smooth muscle has been frequently used to bioassay bradykinin. It is the study of the bradykinin-receptor interaction in intestinal smooth muscle that will be the focus of this review. Other reviews on bradykinin receptors include those of Walaszek [ 19701 and Odya and Goodfriend (19791.
BRADYKlNlN RECEPTOR STUDIES Experimental approaches to receptor studies generally include: 1. Studies of structure-activity
relationships to define the structural requirements for receptor binding; 2. direct binding studies;
6 Chase and Volicer
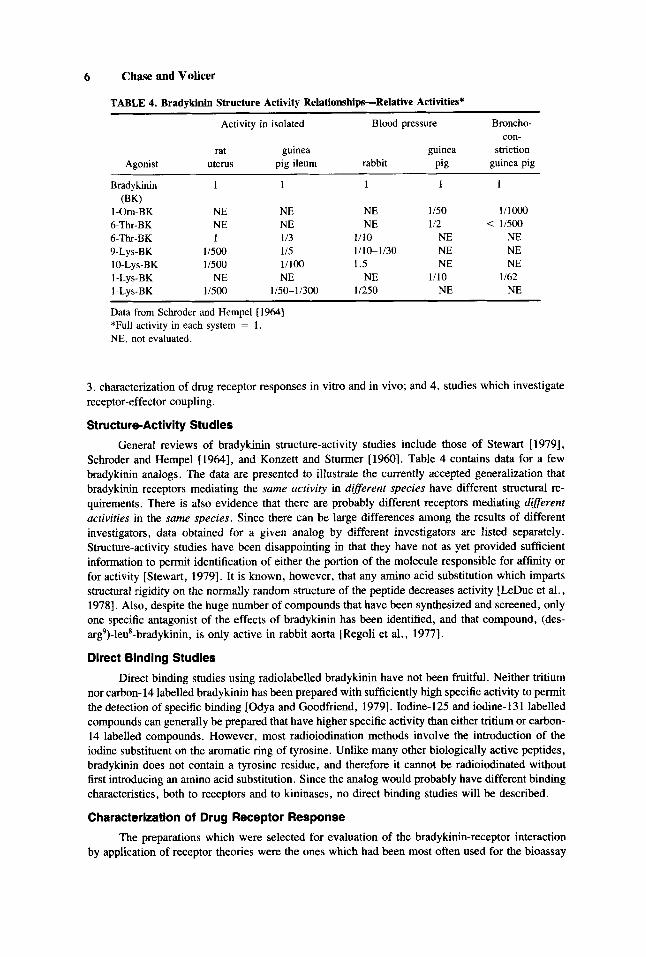
TABLE 4. Bradykinin Structure Activity Relationships-Relative Activities*
Activity in isolated Blood pressure Broncho-
rat guinea guinea striction con-
guinea pig Agonist uterus pig ileum rabbit Pig
Bradykinin (BK)
I-Om-BK 6-Thr-BK 6-Tk-BK 9-Lys-BK 10-Lys-BK I-Lys-BK 1 -Lvs-BK
1 1 1 1 1
NE NE 1
1/500 11500
NE 1/500
NE NE 1 /3 I15 11100 NE
1150-1 1300
NE NE
1/10 1110-1130 1.5
NE 11250
1150 1/2 <
NE NE NE
NE 1/10
l/lOOo 11500
NE NE NE
NE 1/62
Data from Schroder and Hempel [ 19641 *Full activity in each system = 1 . NE, not evaluated.
3. characterization of drug receptor responses in vitro and in vivo; and 4. studies which investigate receptor-effector coupling.
Structure-Activity Studies
General reviews of bradykinin structure-activity studies include those of Stewart [ 19791, Schroder and Hempel [1964], and Konzett and Stunner [1960]. Table 4 contains data for a few bradykinin analogs. The data are presented to illustrate the currently accepted generalization that bradykinin receptors mediating the same activity in difSerent species have different structural re- quirements. There is also evidence that there are probably different receptors mediating difSerent activities in the same species. Since there can be large differences among the results of different investigators, data obtained for a given analog by different investigators are listed separately. Structure-activity studies have been disappointing in that they have not as yet provided sufficient information to permit identification of either the portion of the molecule responsible for affinity or for activity [Stewart, 19791. It is known, however, that any amino acid substitution which imparts structural rigidity on the normally random structure of the peptide decreases activity [LeDuc et al., 19781. Also, despite the huge number of compounds that have been synthesized and screened, only one specific antagonist of the effects of bradykinin has been identified, and that compound, (des- arg’)-leu*-bradykinin, is only active in rabbit aorta [Regoli et al., 19771.
Direct Binding Studies
Direct binding studies using radiolabelled bradykinin have not been fruitful. Neither tritium nor carbon- 14 labelled bradykinin has been prepared with sufficiently high specific activity to permit the detection of specific binding [Odya and Goodfriend, 19791. Iodine-125 and iodine-1 3 1 labelled compounds can generally be prepared that have higher specific activity than either tritium or carbon- 14 labelled compounds. However, most radioiodination methods involve the introduction of the iodine substituent on the aromatic ring of tyrosine. Unlike many other biologically active peptides, bradykinin does not contain a tyrosine residue, and therefore it cannot be radioiodinated without first introducing an amino acid substitution. Since the analog would probably have different binding characteristics, both to receptors and to kininases, no direct binding studies will be described.
Characterization of Drug Receptor Response
The preparations which were selected for evaluation of the bradykinin-receptor interaction by application of receptor theories were the ones which had been most often used for the bioassay

Bradykinin Receptors in Intestinal Muscle 7
TABLE 5. Analysis of Bradykinin-Receptor Interactions ~ ~~~
Type Minimum effective to of maximum effective PD2
Tissue response concentration (moledliter) (M) 16%/84% Reference
Cat terminal C 0.8-800” 7.77 1/28 Barabe et al. ileum [I9751 Cat jejunum C 1 .6-800’ 7.68 1/23 Barabe et al.
[1975] Rat R 0.06- lo0 9.27 not Paegelow et al. duodenum evaluated [I9771
“Cascade superfusion in Krebs buffer was used. C, contraction; R, relaxation.
of bradykinin. In fact, the analyses were performed to validate the use of the tissues for bioassay purposes since a bioassay should employ a system wherein the drug-receptor interaction is well- characterized.
Barabe et al. [ 19751 have analyzed the effects of bradykinin on two segments of cat intestine, the cat terminal ileum and cat jejunum. Paegelow et al. [1977] evaluated the response of the rat duodenum to bradykinin by inducing a prolonged relaxation (2 minutes) and then used planimetry to quantify the response as area. In Table 5, the data are summarized. Barabe et al. [ 19751 reported that the dose-response curves for bradykinin in both segments of cat intestine were “classified.” The effects of bradykinin were direct; they were not antagonized by specific anticholinergic, antiardrenergic, antihistaminic, or antiserotonergic drugs. When possible, agonists were included as positive and negative controls for the specificity of the antagonism. Neither indomethacin nor polyphloretine phosphate inhibited the effect of bradykinin. For both cat tissues, Barabe et al. [1975] determined the pD2, the negative log of the concentration which produced 50% of the maximal response. Using the pD2, theoretical dose-response curves were constructed, as described by Clark [ 19371 and were compared to the experimental curves. For both the cat ileum and jejunum, the theoretical and experimental curves were superimposable. Clark’s ratio, or the ratio of the dose producing 16% to that producing 84% of a maximal response for both the cat ileum and jejunum was in satisfactory agreement with the theoretical value of 1/27, proposed by Clark [I9371 for a bimolecular interaction. The relationship between stimulus (dose) and effect (size of contraction) was also investigated. Stephenson [ 19561 formulated and experimentally evaluated the hypothesis that the effect, as the percent of maximal effect, versus the stimulus, as percent of maximal stimulus, is linear with a slope of 1 .O if the response is directly proportional to receptor occupancy. Barabe et al. [1975] plotted the data from both cat ileum and cat jejunum in this way and, in both cases, the relationship was linear and the slope was 1.0. Since bradykinin is a full agonist in both preparations and since the response appears to be determined by receptor occupancy, the pD2 may be assumed to reflect the dissociation constant of the bradykinin-receptor complex. These results suggest that in the cat ileum and cat jejunum there are receptors which are specific for bradykinin, which behave in accordance with occupation theory, and which have similar affinities for bradykinin. However, because of the lack of a specific antagonist for bradykinin at its receptor, and the lack of specific antagonists for its degradative enzymes, these results must still be considered preliminary.
The results must also be considered preliminary until the in vitro conditions that were used to collect the data are evaluated. Only under reasonably physiological conditions can one expect that, for example, any modulatory systems which operate normally will be intact and that the electrical and mechanical activities of the muscle will sufficiently well resemble conditions in vivo to permit the hope of biological relevance. Bioassay data for bradykinin will now be evaluated, not from a statistically rigorous sense of bioassay, e.g., as an assay, but, rather, the bio aspect of bioassay will be considered. One of two kinds of data were generally collected: 1 ) “Threshold data”
TABLE
6. C
ompa
riso
n of
Effe
ctiv
e* C
once
ntra
tions
? (nm
olef
iiter
) of
Var
ious
Ago
- in I
ntes
tinel
Pre
para
tions
Prep
arat
ions
5-
HT
Ach
A
Tn
BK
Nor
epi
EP1
Ref
eren
ce
Rat c
olon
(as
c.)
Rat c
olon
(des
c.)
Rat s
tom
. shi
p
Rat
duo
denu
m
Rat
ileu
m (
I 8 cm
be
fore
cae
cum
) Ra
t ile
um (j
ust
befo
re c
aecu
m)
Gui
nea
pig
ileum
Gui
nea p
ig il
eum
Gui
nea
pig
ileum
(term. 4
cm.)
C
at je
junu
m
Cat
jeju
num
Gui
nea
pig
taen
ia c
oli
Pige
on re
ctum
Chi
ck re
ctum
Ill-
I111
c
11-1
11
c
0.11
-5.5
C
11-1
11
c
1,11
1 c
111-
1111
c
11-1
11
c
-
1m
20
00
c
23,0
00
C
230
C
> 1
,111
c
11-1
11
c -
9.5-
95
c
9.5-
95
c
9.5-
95
c
9.5-
95 c
95-9
50
C
95-9
50
C
9.5-
95
c -
11
c
340
C
4.8-
14
C
-
9.5-
95
c
9.5-
95
c
0.2-
2.0
c
0.2-
2.0
c
0.2-
2.0
c
200
c
20-2
00
c
20-2
00
c
2-20
c
0.05
-0.1
0 C
-
-
up to
100
0
2-20
c
2-20
c
inse
ns. t
o 20
00
18-1
80
R
1.8-
18
R
1.8-
18
C
R
0.18
-1.8
R
18-1
80
.C
1.8-
18
C
> 1
80
C
1.0
c 7.
5 c
20
c
0.4-
1.8
C
> 1
80
C
or
R or
-
-
120-
1200
R
> 1
200
R
>I2
00
R
> 1
200
R
>I20
0 R
> 1
200
R
> 1
200
R
-
-
-
120
R
> I
200
R
120-
1200
R
120-
1200
R
111-
1111
R
3.
1111
R
> 1
111
R
> 1
111
R
> 1
111
R
> 1
111
R
> 1
111
R
-
-
-
110
R
> 1
111
R
111-
1111
R
11-1
11
R
Reg
oli a
nd
Reg
oli a
nd
Reg
oli a
nd
Van
e [ 1
9641
Van
e I1
9641
Van
e [I
9641
Reg
oli a
nd
Van
e [I9
641
Reg
oli a
nd
Van
e [I
9641
R
egol
i and
V
ane
1196
41
Reg
oli a
nd
Van
e [I
%]
Kha
iralla
h an
d Pa
ge [I
9631
BU
Y a
nd M
ash-
fo
rd (
1977
1 Barabe e
t al.
1197
51
Ferr
eira
and
V
ane
[1%
7]
Reg
oli a
nd
vane
[I9
641
Reg
oli a
nd
Reg
oli a
nd
Van
e 11
9641
Van
e [1
964]
*Eff
ectiv
e is
vario
usly
def
ined
. Reg
oli a
nd V
ane
[196
41: C
ontra
ctio
ns o
f gr
eate
r tha
n or
equ
al to
3 c
m o
r rel
axat
ions
of
grea
ter t
han
or e
qual
to
0.5
cm. are
calle
d “t
hres
hold
dos
es.”
Bury a
nd M
ashf
ord
[197
7]: E
C~S
-JJ.
Kha
iralla
h an
d Pa
ge [1
%3]
: Rep
orte
d “m
inim
um e
ffec
tive”
con
cent
ratio
ns, b
ut d
id n
ot d
efin
e the
bio
logi
cal
end
poin
t. Th
ey a
lso
did
not s
peci
fy if
con
cent
ratio
n In
glm
l) w
as in
term
s of
base
or s
alt.
Dat
a w
ere
trans
form
ed in
term
s of
the
base
. Bar
abe
et a
l. [1
975]
: Rep
orte
d co
rres
pond
ing
mm
of
resp
onse
. For
5-H
T, 1
2.7 2 1
.3; F
or A
ch,
30.2
2
1.2;
For
BK
, 20
nm
olel
liter
pro
duce
d 32
.0 2 1
.0, w
here
as 1
.6 n
mol
ellit
er w
as “
min
. ef
fect
ive
conc
entra
tion.
” (n
ot d
efin
ed).
Ferr
eira
and
Van
e [1
%7]
: C
ontra
ctio
ns (C
) are
thos
e gr
eate
r th
an o
r equ
al to
3.0
cm. R
elax
atio
ns (
R) a
re th
ose
grea
ter t
han
or e
qual
to 0
.5 cm
. sm. =
sm
all,
or le
ss th
an numbers d
escr
ibed
. C
= co
ntra
ctio
n, R
= re
laxa
tion,
0 =
no r
espo
nse.
5-H
T,
sero
toni
n; A
ch,
acet
ylch
olin
e; A
Tn,
angi
oten
sin
11;
BK, b
rady
kini
n; N
orep
i, no
repi
neph
rine;
EPi
, epi
neph
rine.
tC
once
ntra
tions
wer
e ca
lcul
ated
from
orig
inal
dat
a w
hen
nece
ssar
y. U
nits
of w
eigh
thit
vol
ume
wer
e co
nver
ted
to n
mol
e/lit
er in
ter
ms
of t
he c
orre
spon
ding
bas
e.
Syst
ems u
sed
by th
e va
rious
inve
stig
ator
s: R
egol
i and
Van
e [I%
]: 5
ml o
rgan
bat
hiK
rebs
. Bur
y an
d M
ashf
ord
[197
7]: 2 m
l org
an b
awy
md
e. K
haira
llah and
Page
[1
%3]
: Onl
y sp
ecifi
ed o
rgan
bat
h. B
arab
e et
al.
[197
5]: E
ither
10
ml o
rgan
bat
h or
sup
erfu
sion-
not
clea
rly d
efin
ed/K
rebs
. Fer
reira
and
Van
e [1
%7]
: Su
perf
usio
d V
-A.”
00 6 B LD s a
c a L. n
Bradykinin Receptors in Intestinal Muscle 9
TISSUE
RAT STOMACH STRlF
CHICK RECTUM
RAT COLON
RAEBIT RECTUM
CAT JEJUNUM STRIP
NE I A T n 1 BK I PG’s I 5-HT I ADH I
I I I 1 1 1
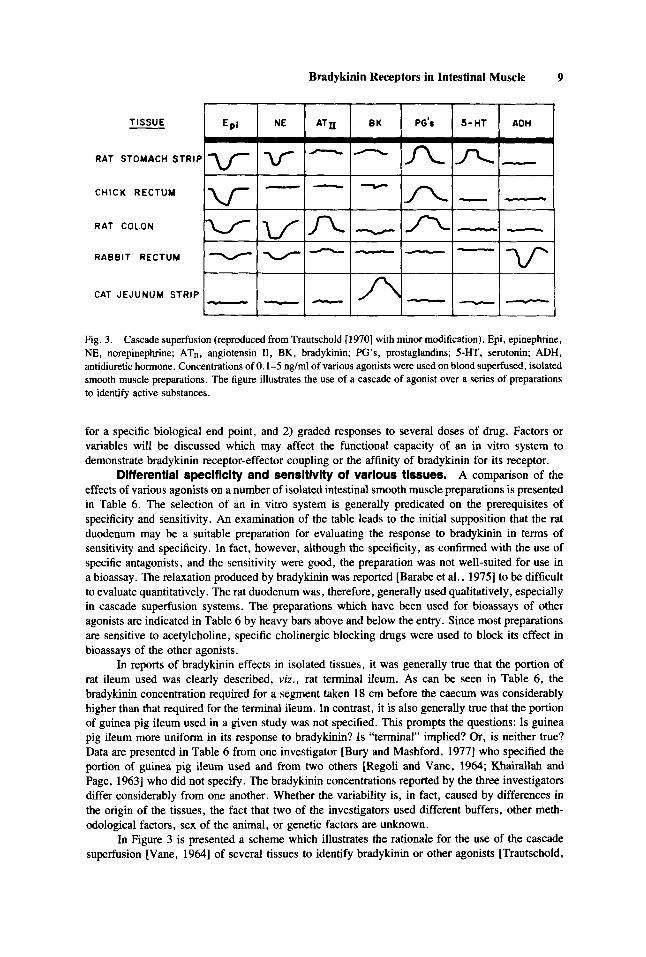
Fig. 3. Cascade superfusion (reproduced from Trautschold [ 19701 with minor modification). Epi, epinephrine, NE, norepinephrine; ATn, angiotensin 11, BK, bradykinin; PG’s, prostaglandins; 5-HT, serotonin; ADH, antidiuretic hormone. Concentrations of 0.1-5 ng/ml of various agonists were used on blood superfused, isolated smooth muscle preparations. The figure illustrates the use of a cascade of agonist over a series of preparations to identify active substances.
for a specific biological end point, and 2) graded responses to several doses of drug. Factors or variables will be discussed which may affect the functional capacity of an in vitro system to demonstrate bradykinin receptor-effector coupling or the affinity of bradykinin for its receptor.
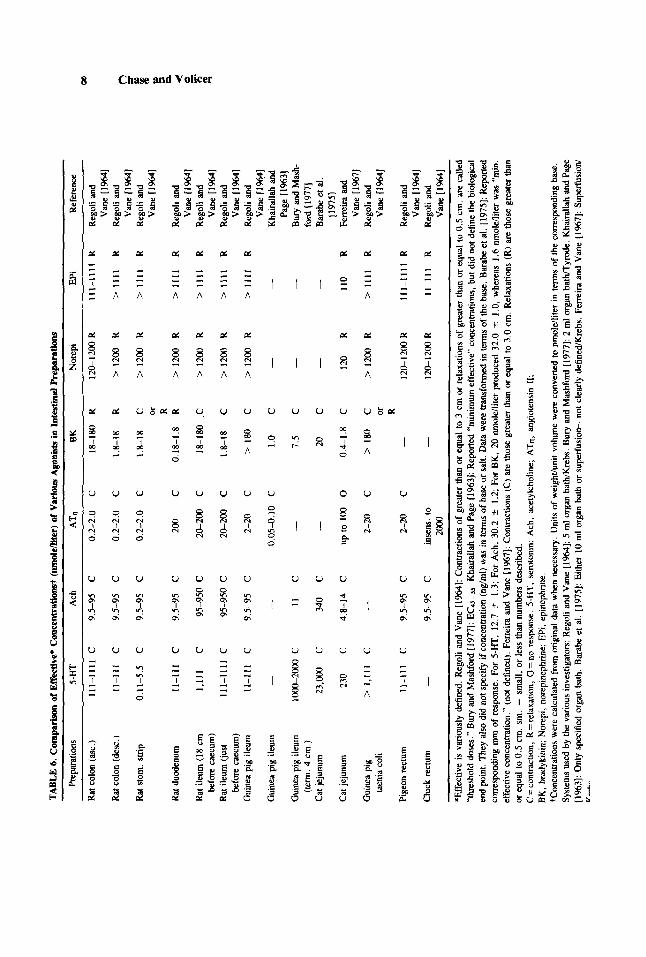
Differential specificity and sensitivity of various tissues. A comparison of the effects of various agonists on a number of isolated intestinal smooth muscle preparations is presented in Table 6. The selection of an in vitro system is generally predicated on the prerequisites of specificity and sensitivity. An examination of the table leads to the initial supposition that the rat duodenum may be a suitable preparation for evaluating the response to bradykinin in terms of sensitivity and specificity. In fact, however, although the specificity, as confirmed with the use of specific antagonists, and the sensitivity were good, the preparation was not well-suited for use in a bioassay. The relaxation produced by bradykinin was reported [Barabe et al., 19751 to be difficult to evaluate quantitatively. The rat duodenum was, therefore, generally used qualitatively, especially in cascade superfusion systems. The preparations which have been used for bioassays of other agonists are indicated in Table 6 by heavy bars above and below the entry. Since most preparations are sensitive to acetylcholine, specific cholinergic blocking drugs were used to block its effect in bioassays of the other agonists.
In reports of bradykinin effects in isolated tissues, it was generally true that the portion of rat ileum used was clearly described, viz., rat terminal ileum. As can be seen in Table 6, the bradykinin concentration required for a segment taken 18 cm before the caecum was considerably higher than that required for the terminal ileum. In contrast, it is also generally true that the portion of guinea pig ileum used in a given study was not specified. This prompts the questions: Is guinea pig ileum more uniform in its response to bradykinin? Is “terminal” implied? Or, is neither true? Data are presented in Table 6 from one investigator [Bury and Mashford, 19771 who specified the portion of guinea pig ileum used and from two others [Regoli and Vane, 1964; Khairallah and Page, 19631 who did not specify. The bradykinin concentrations reported by the three investigators differ considerably from one another. Whether the variability is, in fact, caused by differences in the origin of the tissues, the fact that two of the investigators used different buffers, other meth- odological factors, sex of the animal, or genetic factors are unknown.
In Figure 3 is presented a scheme which illustrates the rationale for the use of the cascade superfusion [Vane, 19641 of several tissues to identify bradykinin or other agonists [Trautschold,

10 Chase and Volicer
19701. The cat jejunum strip has frequently been used as the primary bioassay preparation for bradykinin in the presence of other active substances. Ferreira and Vane [1967] described the following criteria for identifying bradykinin in circulating blood using simultaneous parallel bioas- says: 1. Contraction of the cat jejunum, but not the rat stomach strip, rat colon or chick rectum; 2. that the concentration should be reduced by including a circuit in which the kininases in the blood can continue to degrade bradykinin; and 3. that the contraction be increased by the infusion of bradykinin potentiating peptides.
The data for the cat jejunum from Barabe et al. [1975] and Ferreira and Vane [1967] are also presented in Table 6. Although similar responses were measured by the two investigators for some agonists (see footnotes to Table 6), the concentrations required to produce those responses were very dissimilar. Both investigators used cats of either sex and neither defined the exact origin of the tissue. Barabe et al. I19751 reported the use of both organ bath and superfusion systems, yet did not clearly specify on which system the data were obtained. Therefore, it is possible that the disparate sensitivities were merely reflecting the fact that Ferreira and Vane [1967] used superfusion, a more sensitive system, and that Barabe et al. [ 19751 may have used an organ bath, but this is only speculation. Barabe et al. (19751 reported a “minimum effective” concentration for bradykinin in his report which is similar to the concentration reported by Ferreira and Vane [1967] but, unfortunately, Barabe et al. [1975] did not define “minimum effective.”
The inability to compare results quantitatively from investigator to investigator may be caused by unavoidable biological variability. Regardless of the cause, one can only, therefore, evaluate the apparent internal consistency of a given report. This unfortunately leads to the possibility that different research groups are evaluating receptor-effector coupling, etc., at concentrations of agonist that can vary as much as 100-fold. That the effect of a drug may be different at high than at low dose is not an uncommon occurrence.
Enzymatic degradation-the use of bradykinin potentlating peptldes. The bra- dykinin potentiating peptides have been frequently used in in vitro binding studies. In one study, Ufkes et al. [1976] compared the effects of two bradykinin potentiating peptides in several tissues. The peptides were BPP,, (from snake venom), and A-Vl-5 (from tryptic plasma hydrolysates). Although there was no obvious structural similarity between the two peptides, they potentiated the response to bradykinin to the same maximal degree or showed no potentiating effect in the same tissues. Also, once a maximal stimulation was achieved with one peptide, addition of the other caused no further increase, although the maximal response to bradykinin was not achieved.
There also appeared to be species differences in the effectiveness of the peptides. Ufkes et al. [1976] reported that both BPP5, and A-V1-5 did not potentiate the response to bradykinin in rat tissues (duodenum, ileum or uterus); they did potentiate bradykinin in guinea pig ileum and rabbit ileum. However, it is also known that BPP5, is a potentiating peptide with a half recovery time in vitro of only 4 minutes [Soffer, 19761 because it acts as a substrate for ACE; BPP,, does not act as a substrate. Therefore, BPP,, has been found to completely block the pulmonary de- struction of bradykinin in rats; BPPsa was not as effective. It is, therefore, important to consider which peptide is used in a given study. All peptides with two prolyl residues at the C-terminus are resistant to pulmonary ACE; BPP5, has only one C-terminal prolyl group and, therefore, can act as substrate for ACE. Consequently, its lack of effect in rat tissues, but activity in rabbit and guinea pig tissues, may be interpreted as species differences in kininase specificity and/or activity.
Although the mechanism of action of the potentiating peptides, is, at least, in part kininase inhibition, there is some evidence that kininase inhibition may not be their sole mechanism of action. Stewart [ 19791 provided some supporting evidence for a mechanism of action which is not consistent with inhibition of kininase activity. He reported that similar potentiating effects were observed in ileum by pretreatment with potentiator as by its addition when the bradykinin response was maximal. If the mechanism were kininase inhibition, pretreatment would have been expected to have produced a more marked effect.
LeDuc et al. [ 19781 designed a study to attempt to distinguish between kininase and receptor actions of the potentiating peptides using bradykinin analogs. They concluded that BPP9, acts on

Bradykinin Receptors in Intestinal Muscle 11
either another kininase in addition to ACE, or on a bradykinin-linked receptor. The data did not permit them to distinguish between these two possibilities.
Edery and Shemesh [1978] have presented data which suggest that the potentiating peptides act to increase response to bradykinin by affecting prostaglandin release. These data will be discussed in more detail in the section on “bradykinin and prostaglandins.”
It has also been proposed that some BPPs act by increasing intracellular calcium content. BPPI1, has been demonstrated to increase intracellular calcium. By analogy this has been proposed also for BPP,, [Odya and Goodfriend, 19791.
Is0 et al. [1979] examined the effects of YS-980, a sulfhydryl compound, in guinea pig ileum. They found that its actions were specific for bradykinin potentiation in that it had no significant effect on the dose response curves of agonists other than angiotensin I and bradykinin. YS-980 shifted the dose-response curve of bradykinin to the left about tenfold and that of angiotensin I to the right about tenfold, while not changing either the slopes or maximal response of either peptide. The data for the sulfhydryl compounds as bradykinin potentiators are consistent with a mechanism of action of kininase inhibition.
Specific antagonists, which are known to not affect bradykinin receptor interactions, are needed for each of the kininases. There is also the necessity to clarify the mechanism(s) of action of each of these bradykinin potentiators so that results of the many studies using these agents can be properly interpreted.
Sex of the animal. One aspect of the in vitro systems to which virtually no attention was paid was the sex of the animal from which the tissues were taken. Weinberg et al. [1976] found that although there were no differences between acetylcholine or histamine receptors in male and female guinea pig ileal preparations for their respective agonists, the affinity of the bradykinin receptor for bradykinin was about four times higher in ileal strips from female than from male guinea pigs. Also, longitudinal ileal muscle strips from castrated male guinea pigs exhibited receptor affinities for bradykinin which were closer to those obtained for strips from female animals. However, when castrated males were treated with estradiol, prior to sacrifice of the animal, the affinities were nearly identical to those of strips from female animals. Conversely, ileal muscle strips from females that were treated with testosterone prior to sacrifice, had in vitro affinities which were comparable to those of strips from males. It is important to note that addition of the sex hormones to the organ bath during the in vitro study had no effect on subsequently measured affinities. For example, the affinities of strips from females were not detectably altered by the addition of testosterone to the organ bath. Both the physiological reason for the sex difference in guinea pig ileum and the mechanism for the modification of the bradykinin receptor interaction are unknown. Since species differences and tissue differences are possible, whether this phenomenon occurs in other tissues and species is also unknown.
Receptor-effector coupllng. There is little that is yet known of the cellular events that the bradykinin receptors mediate or modulate. We will review the available information linking response to bradykinin with prostaglandins, calcium, adenylate cyclase, and guanylate cyclase.
Bradykinin and prostaglandins: Liebman and Arnold [1978] have determined the effect of bradykinin on prostaglandin formation in microsomal fractions in rat duodenum and uterus. Bra- dykinin enhanced the formation of both PGE, and PGF, in duodenum; PGF, was enhanced more than PGE,. The specificity of the effect was supported by the fact that o-phenanthroline, which inhibits the degradation of bradykinin, increased the effect of bradykinin. In contrast, bradykinin inhibited the synthesis of PGE, in the rat uterus without significantly affecting the synthesis of PGF,,. Control levels of prostaglandin E2 are about threefold higher in rat uterus than in duodenum. Therefore, in the tissue where the control level of PGE, was higher, bradykinin inhibited the synthesis of prostaglandins, whereas in the tissue where control prostaglandin levels were lower, bradykinin enhanced prostaglandin formation. Whether the effect was due to an interaction with the synthetase or activation of lipase activity was not examined.
Edery and Shemesh [1978] reported on an apparent relationship between the tone, prostag- landin accumulation, and response to bradykinin in rat terminal ileum. It was noted that soon after

12 Chase and Volicer
setting up the tissue in a tissue bath, PGE2, PGFI,, and PGF2, began to accumulate and the tissue relaxed. If bradykinin were added in the presence of prostaglandins, a strong contraction ensued which was preceeded by a short relaxation. Thereafter, prostaglandins declined, and response to bradykinin declined in a corresponding fashion. When the bradykinin potentiating peptide, SQ 20.881, was added to the bath, about three times as much prostaglandin accumulated. Subsequent addition of bradykinin in the presence of SQ 20.881 led to a larger response to bradykinin. These data suggest that the bradykinin potentiating peptide may have potentiated the response to bradyk- inin, not by inhibiting kininases alone, but rather by causing the release of prostaglandins.
There are data which suggest the existence of species differences in the relationship between bradykinin actions, prostaglandins, and the tone of the preparation. Edery and Shemesh [1978] reported that, although in the rat terminal ileum prostaglandin accumulation accompanied a relax- ation of the tissue, in the guinea pig ileum, prostaglandin accumulation accompanied an increase in tone of the preparation. Crocker and Willavoys [1976] reported that aspirin, indomethacin, and polyphloretin phosphate antagonized the response to bradykinin in the rat terminal ileum. Barabe et al. [ 19751, however, found that neither indomethacin, nor polyphloretin phosphate antagonized the action of bradykinin in the cat terminal ileum.
Prostaglandin synthesis has been reported to be influenced by bradykinin in a number of non-intestinal smooth muscle preparations as well [Alexander and Gimbrone, 1976; Odya and Goodfriend, 1979; Hong and Levine, 1976; Terragno et al., 19751. A number of these studies indicated that bradykinin could activate a phospholipase and, thus, increase substrate (arachidonic acid) availability. There have also been reports that bradykinin could increase conversion of PGE2 to PGF2 by stimulating prostaglandin E 9-ketoreductase in canine mesenteric veins [Limas, 19771. One study suggested that prostaglandins attenuated the response to bradykinin [Goldberg et al., 19761. When the response to bradykinin was maximal (6 hours after setting up the preparation), prostaglandin inhibitors had no effect; whereas, when the strips were the least responsive to bradykinin (within the first two hours after setting up the tissue), addition of prostaglandin inhibitors increased the response to bradykinin. These studies suggest that bradykinin has the ability to alter the net effect of prostaglandins by altering the PGE/PGF ratio and the possibility that prostaglandin levels may modulate the response of a tissue to bradykinin.
Bradykinin and calcium: Ufkes et al. [I9761 examined the response to bradykinin in the presence of medium containing normal and low calcium concentrations in rat duodenum. The response in the presence of a normal level of calcium was a dose-dependent relaxation. In a low calcium medium, the rat duodenum contracted to bradykinin in a dose-dependent manner, but over a higher dose range. This is unlike responses to epinephrine, for example, which are not calcium dependent.
Aarsen [ 19771 determined both the electrical and mechanical aspects of the responses to both bradykinin and BPP9, in guinea pig taenia coli using the double sucrose gap technique. The author concluded that bradykinin enhanced sodium ion permeability and that the observed effect may be determined by concentrations of chloride, calcium, and other ions in the system. It was also found that bradykinin and BPP9, appeared to have different mechanisms of action. The results obtained in this study, however, were not in agreement with those obtained using the single sucrose gap technique. Aarsen [I9771 reported that in previous studies using the single sucrose gap technique, the taenia coli showed " . . . greater spontaneous activity and usually responded bi-phasically to bradykinin." In this study, bradykinin produced " . . . a small contraction accompanied by an increase in spike frequency and a decrease in membrane potential, with a change in membrane resistence." Aarsen [ 19771 suggested that the different results obtained when the double sucrose gap was used may have been due to hyperpolarization of the membrane in the center of the preparation, the area used to apply drug, because the adjoining portions of tissues were immersed in sucrose solution.
The effects of bradykinin have been linked to calcium ion concentration in several systems unrelated to intestinal smooth muscle as well [Clyman et al., 1975; Odya and Goodfriend, 1979; Perris and Whitfield, 1969; Zelck et al., 19741.

Bradykinin Receptors in Intestinal Muscle 13
Roemer et al. 119781 reported on the effect of bradykinin in rat duodenal and uterine plasma membrane fractions on guanylate cyclase, both in the presence and absence of calcium ion. Although calcium itself stimulated guanylate cyclase up to 20% over basal in both preparations, bradykinin in the presence of calcium significantly decreased guanylate cyclase activity in the duodenum only. In the absence of calcium ion, bradykinin had no significant effect on guanylate cyclase in either tissue. This study indicated a tissue-specific, calcium-dependent capability of bradykinin to alter guanylate cyclase activity.
Bradykinin and adenylate cyclase: Paegelow et al. [1977] have provided data which suggest coupling of the effect of bradykinin with adenylate cyclase in the rat duodenum. Some of the results are unusual, however. It was reported that pre-treatment with ACTH or sodium fluoride significantly increased the relaxation response to bradykinin by stimulating adenylate cyclase. Sodium fluoride was reported to have had a stronger effect than ACTH. However, sodium fluoride is not thought to stimulate membrane-bound adenylate cyclase in the intact cell [Robison et al., 19711. That sodium fluoride produced such an effect may suggest the speculation that there was excessive tissue damage, or that the sodium fluoride may have altered the electricaUmechanica1 properties of the preparation and thereby the response to bradykinin, or perhaps even that bradykinin permitted the sodium fluoride to enter the cell by means of a receptor-operated channel. The authors did not discuss the fact that sodium fluoride appeared to have stimulated adenylate cyclase in an intact cell preparation.
Paegelow et al. [1977] further reported that bradykinin had no effect in an in vitro phos- phodiesterase assay. In the presence of imidazole, which stimulates phosphodiesterase, the bra- dykinin effect was diminished. In the presence of theophylline, which inhibits phosphodiesterase, the response to bradykinin was enhanced. The response of the tissue to isoproterenol, but not to bradykinin, was antagonized by propranolol. Therefore, Paegelow et al. [ 19771 concluded that the bradykinin-induced relaxation was not caused by interaction of bradykinin at the beta-adrenergic receptor. Rather, the effect of bradykinin was mediated by some linkage between bradykinin and adenylate cyclase.
DISCUSSION
Clearly, the data herein assembled indicate marked differences in the effective range of concentrations of bradykinin for given biological response in particular segments of intestinal smooth muscle. The reasons for these differences are unknown as is their impact on elucidation of bradykinin-receptor effector coupling. What is known, however, is that there are marked dif- ferences in how individual investigators dealt with the fact that intestinal smooth muscle is char- acterized by varying degrees of spontaneous electrical and mechanical activity. Since in any in vitro system wherein the response to a drug is to be quantitated a stable baseline is desirable, steps are generally taken in order to attain this end. However, it is possible that in their choice of buffer, time interval between sacrifice of the animal and setting up the tissue, temperature, or in choosing to add drugs to suppress spontaneous activity, investigators may be sufficiently perturbing the system as to alter the tissue response to a given drug. Drug effects, or the lack of them, and/or receptor-contraction coupling may, in effect, be determined by the excitability of the tissue [Daniel and Sama, 19781 and the excitability of the tissue by the conditions used. In the results of studies that were presented, buffer, time between setting up the tissue and measuring the response, tem- perature, and the exact description of the site of origin of the tissue were all factors which were not held constant. The effects of these variables and others, including the extent of physical trauma to the tissue, on the results that were obtained are all unknown and, therefore, evaluation of data using these systems, with respect to the characteristics of the bradykinin receptor must necessarily await further studies which specifically examine these factors.
One point that is troublesome throughout the bradykinin literature is that the standard bra- dykinin preparation (Sandoz BRS 640), to which even synthetic bradykinin activities are compared, contained chlorobutanol as a preservative. Yet vehicle controls were never described as containing chlorobutanol. There is, for example, some controversy concerning the possibility that chlorobutanol

14 Chase and Volicer
may interfere with the response of isolated hen rectal caecum to bradykinin [Walaszek, 19701. Hemsmeyer and Aprigliano [ 19761 reported that the negative inotropic effect of the Sandoz prep- aration was due to chlorobutanol. Since chlorobutanol has local anesthetic as we11 as antiseptic activities and can, at some concentrations, exhibit similar effects to bradykinin, I would have appreciated having seen the use of specific vehicle controls. A limited amount of data comparing isolated, purified bradykinin to the synthetic material appeared in the 1961 “Proceedings of the First International Pharmacological Meeting on the Mode of Actions of Drugs.” Rocha e Silva [1963] presented data comparing his standard, isolated bradykinin to the newly available, synthetic Sandoz BRS 640 bradykinin. The experiments used a 2 X 2 design to obtain discrimination coef- ficients as described by Gaddum [1955]. Identity between the two preparations would yield a discrimination coefficient of 1 .00. The discrimination coefficients for rat duodenum, cat blood pressure, and rat uterus were compared to that of guinea pig ileum, with guinea pig ileum defined as 1.00. The coefficients obtained were 1.3, 0.7, and 1.0, respectively. The difference between 1 .O and 1.3 in the rat duodenum was not considered significant because of the difficulty in obtaining reliable results with the tissue. The discrimination coefficient of 0.7 obtained for cat blood pressure, however, was considered significantly different from 1 .OO “. . . since it was rather persistent and obtained under a variety of conditions [in which the bradykinin standard prepared by Rocha e Silva was] consistently more active than the corresponding number of units of the synthetic material” [Rocha e Silva, 19631. These experiments, however, appear to have been designed assuming the greater stability and purity of the synthetic material would permit evaluation of the chemical purity of the isolated bradykinin.
Similarly, although the data on the relationship between bradykinin and prostaglandins have been intensively studied, there is one experimental detail which is generally not specified, but which may have a significant effect on the results. Frequently, in studies which consider response of the tissue to bradykinin in the presence of indomethacin, the solvent for the indomethacin is not specified. Yet, in at least one preparation, the rat uterus [Barabe et a]., 19771 indomethacin in buffer was not antagonistic to the effects of bradykinin, whereas indomethacin in alcohol was antagonistic, despite the fact that the alcohol itself had no effect.
Another difficulty in the literature is that some reviewers reported “threshold doses” for bradykinin in particular tissues, but did not define “threshold.” When the original reference is examined for the definition, it is sometimes discovered that there is little resemblance between the cited information and the experimental framework, focus, or conclusions of the source.
CONCLUSION Although there is a huge literature on bradykinin describing its relationship to and meas-
urement in any systems, there is still very little that is known about its mechanism of action, even in such a limited sphere as in intestinal smooth muscle. There do appear to be bradykinin receptors which are species and tissue specific. However, the effect of following factors on bradykinin receptors is not clear: 1) The numerous kininases which are species and tissue specific, 2) numerous kallikreins, 3) at Ieast three kininogens in plasma alone, 4 ) the unclear relationship in smooth muscle among prostaglandins, muscle tone, tissue trauma, and the observed response to bradykinin receptor stimulation. Therefore, the completion of the characterization of the nature of the bradykinin receptor, and of receptor-effector coupling requires further investigations.
ACKNOWLEDGMENTS This work was supported by the Pharmaceutical Manufacturers Association Foundation Inc.
and by the Veterans Administration. The expert assistance of B. Farrell, J. Costa, and B. Landrigan in the manuscript preparation is greatly appreciated.
REFERENCES Aarsen, P.N.: The effects of bradykinin and the bradykinin potentiating peptide, BPP5,, on the electrical and
mechanical responses of the guinea-pig taenia coli. Br. J . Pharmacol. 61: 523-532, 1977.

Bradykinin Receptors in Intestinal Muscle 15
Alexander, R.W. and Gimbrone, M.A.: Stimulation of prostaglandin E synthesis in cultured human umbilical
Barabe, J., Park, W.K. and Regoli, D.: Application of drug receptor theories to the analysis of the myotropic
Barabe, J. , Drouin, J.N., Regoli, D. and Park, W.: Receptors for bradykinin in intestinal and uterine smooth
Bury, R.W. and Mashford, M.L.: A pharmacological investigation of synthetic substance P on the isolated
Clark, A.J.: “General Pharmacology.” Berlin: Verlag Springer, 1937. Clyman, R.L., Blacksin, A.S., Sandler, J.A., Manganiello, V.C. and Vaughan, M.: The role of calcium in
regulation of cyclic nucleotide content in human umbilical artery. J. Biol. Chem. 250: 47184721, 1975.
Crocker, A.D. and Willavoys, S.P.: Possible involvement of prostaglandins in the contractile action of bra- dykinin on rat terminal ileum. Commun. J. Pharm. Pharmacol. 28: 78, 1976.
Daniel, E.E. and Sama, S.: The generation and conduction of activity in smooth muscle. AM. Rev. Pharmacol. Toxicol. 18: 145-166, 1978.
Edery, H. and Shemesh, M.: Release of prostaglandins mediating the potentiation of bradykinin by BPF and chymotrypsin in rat isolated ileum. Agents and Actions 8(1-2): 159-160, 1978.
Erdos, E.: Hypotensive peptides: bradykinin, kallidin, and eledoisin. Adv. Pharmacol. 4 1-90, 1966. Ferreira, S.H.: A bradykinin-potentiating factor (BPF) present in the venom of Borhrops jararaca. Br. J.
Ferreira, S.H. and Vane, J.R.: The detection and estimation of bradykinin in the circulating blood. Br. J .
Ferreira, S.H., Baxtelt, D.C. and Greene, L.J.: Isolation of bradykinin-potentiating peptides from Bofhrups
Gaddum, J.H.: In “Polypeptides Which Stimulate Plain Muscle.” Edinburgh and London: Livingstone, 1955. Goldberg, M.R., Chapnick, B.M., Joiner, P.D., Hyman, A.L. and Kadowitz, P.J.: Influence of inhibitors of
prostaglandin synthesis on venoconstrictor responses to bradykinin. J. Pharmacol. Exp. Ther. 198:
Hermsmeyer, K. and Aprigliano, 0.: Effects of chlorobutanol and bradykinin on myocardial excitation. Am. J. Physiol. 230(2): 306-310, 1976.
Hong, S.-C.L., Levine, L.: Stimulation of prostaglandin synthesis by bradykinin and thrombin and their mechanisms of action on MC 5-5 fibroblasts. J. Biol. Chem. 251: 5814-5816, 1976.
Iso, T., Nishimura, K., Oya, M. and Iwao, J.-I.: Potentiating mechanism of bradykinin action on smooth muscle by sulfhydryl compounds. Eur. J. Pharmacol. 54: 303-305, 1979.
Khairallah, P.A. and Page, I.H.: Effects of bradykinin and angiotensin on smooth muscle. Ann. N.Y. Acad. Sci. 104: 212-221, 1963.
Konzett, H. and Stunner, E.: Biological activity of synthetic polypeptides with bradykinin-like properties. Br. J. Pharmacol. 15: 544, 1960.
LeDuc, L.E., Marshall, G.R. and Needleman, P.: Differentiation of bradykinin receptors and of kininases with conformational analogues of bradykinin. Mol. Pharmacol. 14 413421, 1978.
Liebmann, C. and Arold, H.: Effect of bradykinin on prostaglandin synthetase activity in microsomal fractions from rat duodenum and uterus. Acta Biol. Med. Germ. 37: 901-903, 1978.
Limas, C.J.: Selective stimulation of venous prostaglandin E 9-keto-reductase by bradykinin. Biochim. Biophys. Acta 498: 306-315, 1977.
Murthy, V., Waldron, T. and Goldberg, M.: The mechanism of bradykinin potentiation after inhibition of angiotensin converting enzyme by SQ 14.225 in conscious rabbits. Circ. Res. (Suppl. 1) 43(1): 140-145, 1978.
Oates, J.A. and Melmon, K.L.: Biochemical and physiological studies of the kinins in carcinoid syndrome. In “Hypotensive Peptides.” New York Springer-Verlag, 1966, pp. 563.
Odya, C.E. and Goodfriend, T.L.: Bradykinin receptors. In “Handbook of Experimental Pharmacology.” Vol. XXV(Suppl.), New York: Springer-Verlag, 1979, pp. 287-300.
Paegelow, I., Reissmann, S., Vietinghoff, G., Romer, W and Arnold, H.: Bradykinin action in the rat duodenum through the cyclic AMP system. Agents and Actions 7(4): 447451, 1977.
Pems, A.D. and Whitfield, J.F.: The mitogenic action of bradykinin on thymic lymphocytes and its dependence on calcium. Proc. Soc. Exp. Med. Biol. 130: 1198-1201, 1969.
Pisano, J.J.: Kinins in nature. In “Handbook of Experimental Pharmacology.” Vol. XXV(Suppl.), New York: Springer-Verlag, 1979, pp. 273-285.
vein smooth muscle cells. Proc. Natl. Acad. Sci. USA 7 3 1617-1620, 1976.
effects of bradykinin. Can. J. Physiol. Pharmacol. 53: 345-353, 1975.
muscle. Can. J. Physiol. Pharmacol. 55: 1270-1285, 1977.
guinea-pig ileum. Clin. Exp. Pharmacol. Physiol. 4 453461, 1977.
Pharmacol. 24: 163-169, 1965.
Pharmacol. Chemother. 29: 367-377, 1967.
jaruraca venom. Biochemistry 9(13): 2583-2592, 1970.
357-365, 1976.

16 Chase and Volicer
Regoli, D. and Vane, J.R.: A sensitive method for the assay of angiotensin. Br. J. Pharmacol. 23: 351-359,
Regoli, D., Barabe, 1. and Park, W.K.: Receptors for bradykinin in rabbit aortae. Can. J. Physiol. Pharmacol.
Roberts, M.L. and Mashford, M.L. : Possible physiological and pathological roles of the kallikrein-kinin system.
Robison, G.A., Butcher, R.W. and Sutherland, E.W.: “Cyclic AMP.” New York: Academic Press. Rocha e Silva, M.: Definition of bradykinin and other kinins. In “Proceedings of the First Intemat. Pharmacol.
Meeting.” vol. 9(1), New York: Pergamon Press, MacMillan Co., 1963, vol 9(1), pp. 3-21. Roemer, W., Liebmann, C., Reissmann, S. and Arold, H.: Effects of prostaglandins, calcium, and bradykinin
on guanylate cyclase in different organs. Acta biol. Med. Germ. 37: 905-906, 1978. Schachter, M.: Kallikreins (kinin0genases)-a group of serine proteases with bioregulatory actions. Pharmacol.
Rev. 31(1): 1-17, 1979. Schroder, E. and Hempel, R.: Bradykinin, kallidin and their synthetic analogues. Experientia, 20(10): 529-592,
1964. Soffer, R.L.: Angiotensin-converting enzyme and the regulation of vasoactive peptides. Ann. Rev. Biochem.
Stephenson, R.P.: A modification of receptor theory. Br. J. Pharmacol. Chemother. 11: 379-393, 1956. Stewart, J.M.: Chemistry and biologic activity of peptides related to bradykinin. In “Handbook of Experimental
Pharmacology.” Vol. XXV (Suppl.), New York: Springer-Verlag, 1979, Vol. 25, pp. 227-272. Terragno, D.A., Crowshaw, K., Tenagno, N.A. and McGiff, J.C.: Prostaglandin synthesis by bovine mesenteric
arteries and veins. Circ. Res. (Suppl. 1) 36: 176-180, 1975. Torstila, I.: The plasma kinin system in acute myocardial infarction. Acta Med. Scand. (Suppl.) 620: 162,
1978. Trautschold, I.: Assay methods in the kinin system. In “Handbook of Experimental Pharmacology.” Vol. XXV,
New York: Springer-Verlag, 1970, vol 25, pp. 52-81. Ufkes, J.G.R., Aarsen, P.N. and Van der Meer, C.: The bradykinin potentiating activities of two pentapeptides
on various isolated smooth muscle preparations. Eur. J. Pharmacol. 40(1): 137-144, 1976. Vane, J.R.: The use of isolated organs for detecting active substances in the circulating blood. Br. J. Pharmacol.
Walaszek, E.J.: The effect of bradykinin and kallidin on smooth muscle. In “Handbook of Experimental Pharmacology.” New York Springer-Verlag, 1970, vol 25, pp. 421429.
Weinberg, J . , Diniz, C.R. and Mares-Guia, M.: Influence of sex and sexual hormones in the bradykinin- receptor interaction in the guinea pig ileum. Biochem. Pharmacol. 25(4): 433437, 1976.
Y-K Wong, P., Terragno, D.A., Tenagno, N.A. and McGiff, J.C.: Dual effects of bradykinin on prostaglandin metabolism: relationship to the dissimilar vascular actions of kinins. Prostaglandins 13(6): 1 1 13-1 125, 1977.
Zelck, U., Konya, L. and Albracht, E.: ATP-dependent calcium uptake by microsomal fractions of pig coronary artery and its dependence on bradykinin and angiotensin. Acta Biol. Med. Ger. 32: Kl-K5, 1974.
1964.
55: 855-867, 1977.
Med. J. Aust. 2: 887-891, 1972.
45: 73-94, 1976.
23: 360-373, 1964.