branching 81-6n-acetylglucosaminetransferases and
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1993 by The American Soeiety for Bioehemistry and Molecular Biology, Inc.
Vol. 268, No. 2, Issue of January 15, pp. 1242-1251,1993 Printed in U.S.A.
Branching 81-6N-Acetylglucosaminetransferases and Polylactosamine Expression in Mouse F9 Teratocarcinoma Cells and Differentiated Counterparts*
(Received for publication, April 3, 1992)
Michael HeffernanS, Reuben Lotan$, Brad Amos$, Monica Palcicll, Ryo Takano$, and James W. Dennis4 )I From the $Samuel Lunenfeld Research Institute, Mt. Sinai Hospital, Toronto, Ontario M5G 1x5 and the Department of Medical Genetics, University of Toronto, Toronto, Ontario M5G 1x5, Canada, the §Department of Tumor Biology, University of Texas, M. D. Anderson Cancer Center. Houston. Texas 77030. and the qDepartment of Food Science, University of Alberta, Edmonton, Alberta T6G 2P5, Canada
8-All-trans-retinoic acid (RA)-induced endodermal differentiation of mouse F9 teratocarcinoma cells is accompanied by changes in glycoprotein glycosylation, including expression of i antigen (Le. polylactosamine) and leukophytohemagglutinin-reactive oligosaccha- rides (Le. -GlcNAcj31-6Manal-6-branched N-linked). We have used the F9 teratocarcinoma cells as a model to study developmental regulation of glycosyltransfer- ase activities which are responsible for the biosyn- thesis of 81-6GlcNAc-branched N- and 0-linked oli- gosaccharides and polylactosamine. Growth of F9 cells in the presence of lo-‘ M RA for 4 days increased core 2 GlcNAc transferase and GlcNAc transferase V ac- tivities by 13- and g-fold, respectively, whereas the activities of GlcNAc transferase I, 81-3GlcNAc trans- ferase (i), 81-4Gal transferase, and 81-3Gal transfer- ase increased 2-4-fold. Induction of glycosyltransfer- ase activities by RA was dose-dependent and showed a biphasic response with approximately half of the in- crease observed 3 days after RA treatment and the remainder occurred by day 4. PYS-2, a parietal endo- derm cell line, showed levels of glycosyltransferase activities similar to those of RA-treated F9 cells. Gly- cosyltransferase activities in the RA-resistant F9 cell line (RA-3-10) were low and showed only a small induction by RA. These observations suggest that dif- ferentiation of F9 cells is closely associated with in- duction of multiple glycosyltransferase activities, with most pronounced increases in GlcNAc transferase V and 2’, 5”tetradenylate (core 2) GlcNAc transferase. The increase in GlcNAc transferase V was also re- flected by the 4-6-fold increase in the binding of ”‘1- leukophytohemagglutinin to several cellular glycopro- teins, which occurred after 3 days of RA treatment.
The endo-8-galactosidase-sensitive polylactosamine content of membrane glycoproteins and, in particular, the LAMP-1 glycoprotein was markedly increased after RA treatment of F9 cells. Consistent with these observations, fucosylated polylactosamine (Le. dimeric
* This work was supported by research grants from the National Cancer Institute of Canada and the Medical Research Council of Canada (to J. W. D.) and by the M. D. Anderson Annual Campaign Fund (to R. L.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
11 Senior Research Scientist of the National Cancer Institute of Canada. To whom correspondence should be addressed Samuel Lu- nenfeld Research Institute, Mt. Sinai Hospital, 600 University Ave., Toronto, Ontario M5G 1x5, Canada.
Lex) was also increased in RA-treated cells. Analysis of the aryl oligosaccharides produced by F9 cells cul- tured in the presence of aryl a-D-GalNAc showed that RA treatment enhanced the synthesis of disialyl core 2 0-linked oligosaccharides and increased the polylac- tosamine content of the aryl oligosaccharides by >20- fold. The results suggest that differentiation of FB cells into endoderm is closely associated with increased GlcNAc transferase V and core 2 GlcNAc transferase activities, enzymes which control the level of 81- 6GlcNAc-branched N- and 0-linked oligosaccharides, the preferred substrates for polylactosamine addition.
The mouse teratocarcinoma cell line F9 exhibits character- istics of embryonic cells from inner cell mass of the blastula stage, and following RA’ treatment, the cells differentiate into primitive endoderm-like cells (1-3). F9 differentiation occur- ring over 4-5 days in the presence of RA is associated with changes in gene expression typified by sequential expression of homeotic (i.e. Hox) genes, and the expression of collagen type IV and laminin (4-6). In addition, cell surface carbohy- drates change following RA-induced differentiation as deter- mined using anti-carbohydrate antibodies (7,8). In particular, loss of SSEA-1 (i.e. Le’, Galp1-4(Fucl-3)GlcNAc~) (9) and acquisition of SLe” (10) and Galal-3Gal (11) occurs in RA- induced F9 cells. These antigenic carbohydrate sequences are commonly found at the nonreducing termini of polylactosa- mine ( i e . repeating Galpl-4GlcNAcpl-3 which also increases following differentiation of embryonal carcinoma cells (7,121. Polylactosamine or i antigen expressed on differentiated EC cells was found almost exclusively on glycoproteins (8), pre- sumably associated with N- and/or 0-linked oligosaccharides. Expression of SSEA-1 at the 8-16-cell stage has been impli- cated in compaction of the mouse embryo (13,14), suggesting that developmentally regulated changes in oligosaccharide biosynthesis and processing may be required for embryo de- velopment.
Following RA-induced differentiation of F9 cells, the LAMP-1 glycoprotein, a major cellular carrier of N-linked
’ The abbreviations used are: RA, 6-all-trans-retinoic acid; L-PHA, leukoagglutinin; pNP, para-nitrophenyl; MES, 4-morpholineethane- sulfonic acid; ELISA, enzyme-linked immunosorbent assay; BSA, bovine serum albumin; TES, N-tris(hydroxymethyl)methyl-2-ami- noethanesulfonic acid PBS, phosphate-buffered saline; Me2S0, di- methyl sulfoxide; PAGE, polyacrylamide gel electrophoresis; HPLC, high performance liquid chromatography; SA, sialic acid; BtZcAMP, N6,02-dibutyryl CAMP.
1242

P1-6GlcNAc Transferases and Polylactosamine in F9 Cells 1243
oligosaccharides (15-18), shows greatly increased binding to leukophytohemagglutinin (L-PHA) (19). This lectin binds specifically to the more branched tri- and tetraantennary complex-type N-linked oligosaccharides (20). L-PHA reactiv- ity with LAMP-1 also increases in oncogene-transformed rat2 fibroblasts and in metastatic sublines of the SP1 murine mammary carcinoma 1 cells as a result of a 3-10-fold increase in UDP-GlcNAc:aGMan @6-N-acetylglucosaminyltransferase V (i .e. GlcNAc transferase V) activity (21, 22). The polylac- tosamine content in N-linked oligosaccharides of MDAY-D2 lymphoma cells and SP1 cell lines correlated with changes in GlcNAc transferase V activity, whereas UDP-G1cNAc:Gal- P4GlcNAc-R P3-N-acetylglucosaminyltransferase (GlcNAc to Gal) (i .e. GlcNAc transferase (i)) activity remained un- changed (22). This suggests that GlcNAc transferase V cata- lyzes a rate-limiting reaction in expression of polylactosamine in N-linked oligosaccharides and is consistent with earlier substrate specificity studies on partially purified GlcNAc transferase (i) which showed the enzyme preferentially sub- stituted acceptors with the Gal@l-4GlcNAc~l-6Mancu anten- nas (23).
Polylactosamine is also found in 0-linked oligosaccharides (24), and its expression in SP1 tumor cells appeared to be dependent upon UDP-GlcNAc:Gal@3GalNAc-R P6-N-acetyl- glucosaminyltransferase (GlcNAc to GalNAc) (i.e. core 2 GlcNAc transferase) activity which substitutes Gal@1- 3GalNAc producing Gal~l-3(GlcNAc~l-6)GalNAc (22). Sub- stitution of the GlcNAc by UDP-Ga1:GlcNAc-R P4-galacto- syltransferase ( i e . @1-4Gal transferase) provides a lactosa- mine substrate which can be extended by GlcNAc transferase (i) into polylactosamine. Core 2 GlcNAc transferase activities in human B lymphocytes have recently been shown to regulate polylactosamine levels in the 0-linked oligosaccharides of CD43 (i.e. leukosialin) (25).
These observations suggest that during embryogenesis, expression of polylactosamine, as well as the extended chain antigens ( i e . poly-Le' and -LeY) based on this linear sequence, may be regulated by the Pl-6 branching GlcNAc transferases. T o address this question, we have compared glycosyltransfer- ase activities and polylactosamine levels in N- and 0-linked oligosaccharides of F9 cells and RA-treated F9 cells. The results show that differentiation of F9 cells into primitive or parietal endoderm is associated with distinct patterns of gly- cosyltransferase activity. Furthermore, enhanced GlcNAc transferase V and core 2 GlcNAc transferase activities in differentiated F9 cells were associated with expression of endo-/?-galactosidase-sensitive polylactosamine sequences in both N- and 0-linked oligosaccharides.
MATERIALS AND METHODS
Chemicals and Glycosyltransferase Substrates
UDP-6-[3H]N-acetylglucosamine (26.8 Ci/mmol, Du Pont-New England Nuclear) and UDP-6-[3H]galacto~e (20 Ci/mmol, Amer- sham, United Kingdom) were diluted with the respective unlabeled sugar nucleotides purchased from Sigma. GalNAca-pNP and GalP1- 3GalNAca-pNP were purchased from Toronto Research Chemicals (Toronto). GlcNAcpl-3Galp-OCH3 was purchased from Sigma. GlcNAc~l-2Mannl-6Man/3-O(CHz)aCOOCH3; Manal-3(Man al-6)Man~1-O(CH2)aCOOCH3; Ga1(31-4GlcNAc(3-2Manal- 6Manp-O-(CH2)&OOCH3 were kindly providedby Dr. 0. Hindsgaul, University of Alberta. RA was a gift from BASF Aktiengeselschaft (Ludwigshafen, Germany). L-PHA was purchased from Vector Lab- oratories (Burlingame, CA). SH2 monoclonal antibody was kindly provided by Drs. A. K. Singhal and S. Hakomori, Biomembrane Institute, Seattle (26).
Cell Lines
Mouse F9 teratocarcinoma cells were cultured in 1:l (v/v) Ham's F-12 and Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum (Hazelton, Lenexa, KS) and antibiotics (100 pg/ml of both penicillin and streptomycin). Cells were induced to differentiate in monolayer cultures by adding RA to the cultures from a 10 mM solution in MezSO to a final concentration of M (19). Alternatively, F9 cells were cultured in the presence of 2 mM sodium butyrate, 1 mM N6,0Z-dibutyryl CAMP (Bt2cAMP) or combinations of these compounds with M RA. Control cultures received MezSO at the same concentration as the RA-treated cultures (0.01%). All cultures were refed with fresh medium every 48 h. The cells were cultured in the presence of differentiating agents for 4 days and then harvested for analysis. The origin and analysis of oligosaccharides in the highly metastatic DBA/2 strain lymphoid tumor called MDAY- D2 have been described previously (27).
To label oligosaccharides metabolically, F9 cells were grown for 3 days in medium supplemented with 0.01% MezS04 or M RA. The cultures were then fed with medium containing 10 pCi/ml D-[6-%] glucosamine (25 Ci/mmol, ICN Radiochemicals, Irvine CA) and in- cubated for an additional 48 h. Cells were harvested, and LAMP-1 was immunoprecipitated and digested with endo-0-galactosidase for 16 h at 37 "C as described elsewhere (19).
Glycosyltransferase Assays
Cells were washed in phosphate-buffered saline and lysed in 0.9% NaCl, 1% Triton X-100 at 0 "C for all assays. The reactions contained 25 pl of cell lysate (8-12 mg/ml of protein), 0.5 pCi of 3H-sugar nucleotide donor (approximately 4,400 cpm/nmol) in a total volume of 50 p1 and were incubated for 1 or 2 h a t 37 "C followed by processing as indicated for each substrate. For all transfer assays, endogenous activity was measured in the absence of acceptor and subtracted from values determined in the presence of added acceptor. For assays using pNP and O(CH,)8COOCH3-coupled substrates, the reactions were diluted to 5 ml in H,O and applied to a C18 Sep-Pak column (Millipore Waters) in H,O, washed with 20 ml of HzO. The products were then eluted with 5 ml of methanol, and radioactivity was counted in a p- liquid scintillation counter.
Core 2 GlcNAc Transferase-The reaction contained 50 mM MES, pH 7.0, 1 mM UDP-GlcNAc, 0.5 pCi of UDP-[3H]GlcNAc, 0.1 M GlcNAc, 1 mM Galpl-3GalNAca-pNP as substrate in a final volume of 33 11.
fll-2GlcNAc Transferase I (UDP-GlcNAc:cu3Man p2-N-Acetylglu- cosaminyltransferase I)-The reaction contained 50 mM MES, pH 7.0, 1 mM UDP-GlcNAc, 0.5 pCi of UDP-[3H]GlcNAc, 0.1 M GlcNAc, 25 mM MnCI,, and 1 mM Manal-3(Manal-6)Man(?l- O(CHz)8COOCH3 as substrate.
pH 7.0, 1 mM UDP-GlcNAc, 2 pCi of UDP-[3H]GlcNAc, 0.1 M /3-6GlcNAc Transferase V-The reactions contained 50 mM MES,
GlcNAc, 10 mM Na2EDTA, 1 mM GlcNAc@1-2Manal-6Manpl-
fi13GlcNAc Transferase (+The reactions contained 50 mM MES, pH 7.0, 1 mM UDP-GlcNAc, 0.5 pCi of UDP-[3H]GlcNAc, 0.1 M GlcNAc, 25 mM MnClZ, and 1 mM Gal~l-4GlcNAcpl-2Mancul- 6ManB-O-(CH2)a-COOCH3.
p1-3Gal Transferase (UDP-Ga1:GalNAca-R 03-Galactosyltransfer- me)-The reactions contained 50 mM MES, pH 7.0, 1 mM UDP-Gal, 0.5 pCi of UDP-[3H]Gal, 25 mM MnC12, and 1 mM GalNAca-phenyl.
p1-4Gal Transferase-The reactions contained 50 mM MES, pH 7.0, 1 mM UDP-GlcNAc, 0.5 pCi of UDP-[3H]Gal, 25 mM MnCIZ, and 20 mM GlcNAc as substrate. The reactions were passed over Dowex AG 1-X8 in H,O and excluded radioactivity counted.
O(CHz)aCOOCH3.
ELISA for GlcNAc Transferase V Activity
Immulon 96-well microtiter plates were coated with 2 pg/well of the acceptor neoglycoprotein GlcNAc(/31-2)Man(al-6)Manpl- O(CHz)aCO-BSA as described previously (28). The GlcNAc transfer- ase V reactions contained 20 mM EDTA, pH 8.0, 100 mM TES, pH 7.1, 10 mM AMP, 0.2 mM GlcNAc, 4 mM UDP-GlcNAc in a total volume of 100 pllwell. Following a 2-h incubation a t 37 "C, the microtiter plates were washed with water and phosphate-buffered saline containing 0.05% Tween 20 (PBST), followed by a 2-h incu- bation a t room temperature with a 1:8,000 dilution of a refined rabbit antiserum in PBST + 1% BSA which specifically recognizes the GlcNAc~l-2GlcNAc/31-6Manal-6Man@l-O(CHz)aCO-BSA GlcNAc transferase V reaction product. Following PBST washes the

1244 @l-GGlcNAc Transferases and Polylactosamine in F9 Cells
plates were incubated with a 1:1,000 dilution in PBST + 1% BSA of alkaline phosphatase-conjugated goat anti-rabbit IgG (Bio-Rad) at room temperature for 2 h. Plates were washed with PBST and water prior to a 1-h color development using a p-nitrophenyl phosphate substrate solution (Bio-Rad). Absorbance at 410 nm was determined using a Bio-Tek EL-309 enzyme immunoassay plate reader. All assays were performed in duplicate at six to eight lysate protein concentra- tions. Activities (i.e. A410 units/mg/h) in different cell lysates were compargd in experiments preformed on the same day.
Measurement of Endo-@-Galactosidase-sensitiue Polylactosamine Endo-@-galactosidase-sensitive polylactosamine was quantitated as
described previously (22). Briefly, microsomal membranes in 1 mM HEPES, pH 7.0, were heated to 65 "C for 15 min to inactivate 8- galactosidases. The samples were diluted with 0.15 M NaCl, 10 mM HEPES, pH 7.0,0.5% Triton X-100 to 3 mg/ml, and terminal GlcNAc residues, particularly 0-linked GlcNAc residues (29), were blocked with cold galactose by bovine p4-Gal transferase. The reaction buffer contained 100 mM galactose, 100 mM HEPES, pH 7.3, 0.15 M NaC1, 50 mM MnC12, 1.2 mM AMP, 1 mM UDP-Gal, 0.5% Triton X-100, 300 milliunits/ml bovine Gal transferase (Sigma) and 1.0 mg/ml microsomal membrane protein. The reaction was incubated for 2 h at 37 'C, and then the samples were exhaustively dialyzed against 0.15 M NaCl, 50 mM McClZ, and 25 mM sodium acetate, pH 5, to remove the cold UDP-Gal. Half of each sample was treated with 5 milliunits of endo-@-galactosidase (Miles) per 100 pg of protein for 16 h at 50 "C. Samples (50 pg), both endo-@-galactosidase-treated and untreated, were diluted 1:l with the reaction buffer, this time with 2 pCi of UDP-[3H]Gal (20 Ci/mmol, Amersham) instead of cold UDP- Gal. After 2 h the reaction was terminated by the addition of ice-cold 10% trichloroacetic acid, and the pellet was washed three times with 10% trichloroacetic acid and counted in a liquid scintillation counter.
To release N-linked oligosaccharides, samples were labeled with [3H]Gal as described above and then heated to 65 "C for 15 min, made 0.4% sodium deoxycholate, and incubated with and without 1.5 units of glycopeptidase F from Flavobacterium meningosepticurn (Boehrin- per) for 16 h at 37 "C. The samples were then precipitated and washed three times with 10% trichloroacetic acid and counted.
Binding of lZ51-L-PHA to Cellular Glycoproteins Cells were solubilized by suspending 2.5 X lo7 cells/ml in 0.5%
Nonidet P-40 in 10 mM Tris-HC1, pH 7.4, and incubating them for 20 min on ice. The cell suspension was centrifuged at 12,000 X g, and the supernatant fraction (cell extract) was collected. The cellular proteins were subjected to polyacrylamide gel electrophoresis (PAGE), in the presence of sodium dodecyl sulfate (SDS), and then the proteins were transferred electrophoretically to a nitrocellulose membrane. Following an overnight blocking of nonspecific binding sites on the nitrocellulose with a solution containing 15% skim milk, 0.02% sodium azide, and 0.15 M NaCl in 10 mM Tris-HC1, pH 7.4 (blocking buffer), the membrane was incubated for 2 h at 23 "C with lZ6I-labeled L-PHA (4-8 X lo5 cpm/mg protein; 1 X lo6 cpm/ml in the blocking solution) with agitation. Following four 15-min washes with a solution containing 0.05% Polysorbate 20 and 0.15 M NaCl in 10 mM Tris-HC1, pH 7.4, the membrane was air-dried and placed on an x-ray film (XAR-5, Kodak) between two intensifying screens for autoradiography. Quantitation of specific bands on the autoradi- ograms was achieved by scanning the autoradiogram using a Beckman DU-8 scanning spectrophotometer.
Western Blotting Proteins separated by 12% SDS-PAGE under reducing conditions
were transferred electrophoretically onto nitrocellulose sheets in buffer consisting of 25 mM Tris, 0.2 M glycine which contained 20% methanol. Blots were blocked for 1 h at 37 "C in 50 mM Tris-HC1, pH 8.0, 0.15 M NaCl (TBS), 5% BSA, then incubated with a 1:50 dilution of culture supernatant containing SH2 monoclonal antibody in PBS, 0.1% BSA. Following three 5-min washes in PBS, 0.1% BSA, lectin blots were incubated for 1 h with horseradish peroxidase- coupled anti-mouse antibody diluted 1:1,000 in TBS, 0.1% BSA. Blots were washed four times (5 min each) followed by one wash in TBS, 0.1% Tween 20, and one wash in TBS. The blots were developed using the ECL chemiluminescence Western blotting kit as per man- ufacturer's instructions (Amersham).
Separation of Aryl Oligosaccharides from Cell Culture Medium F9 cells, either untreated or 5 days in M RA, were cultured at
2 X 106/ml in serum-free minimal essential medium with 2 mM GalNAccu-pNP for 6 b as described previously (30). The accumulation of aryl oligosaccharides in the medium is linear during this incubation. Lyophilized cell culture medium was reconstituted in 0.5 ml of H20, and aryl oligosaccharides were desalted on a Bio-Gel P2 column (1 X 45 cm) developed in HzO. The oligosaccharides were pooled, lyophi- lized, and separated on a Protein Pak DEAE 5PW HPLC column (Millipore Waters) equilibrated in 2.5 mM KH2P04, pH 5.2. The column was eluted with the starting buffer for 15 min followed by a gradient to a final concentration of 250 mM KH2P04, pH 5.2, for a further 60 min. The disialyl fraction was pooled and desalted on a Bio-Gel P2 column (1 X 45 cm) developed in H20. Aliquots of aryl oligosaccharides were treated with 0.1 M HCl for 1 h at 80 "C and then separated on a 1 X 25-cm Ultrahydrogel HPLC column (Milli- pore Waters) developed isocratically in 80% acetonitrile, 20% 5 mM KH2P04, pH 5.2. The HPLC column effluent was monitored at 303 nm with a Waters LC spectrometer model 481, and the data were analyzed with a Waters 740 data module. A standard curves using pl-3GalNAccu-pNP produced an extinction coefficient of 600,000 AaO3 units/nmol.
Endo-@-galactosidase-sensitive polylactosamine in the disialyl aryl oligosaccharides was measured as described above using bovine Gal transferase and UDP-[3H]Gal. The products of the reaction were retained on Sep-Pak CIS and eluted with 100% methanol. A standard curve was generated using GlcNAc@1-2Manal-GManP- O(CH2)~COOCH3 as acceptor. Product formed over a range of 1-750 pmol of substrate was linear and provided a measure of P1-4Gal transferase efficiency of approximately 10% conversion of substrate to product.
RESULTS
Glycosyltransferase Activity in F9 and RA-induced F9 Cells-RA-induced differentiation of F9 cells was shown to increase L-PHA reactivity of cellular glycoproteins greatly (19), particularly LAMP-1, which has 18 N-linked glycosyla- tion sites bearing "complex-type" oligosaccharides (17). Since L-PHA binds to the Gal~l-4GlcNAc~l-6(Gal~l-4Glc- NAcpl-2)Manpl-6 portion of tri- and tetraantennary N- linked glycans (20), increased reactivity of the lectin with RA- treated F9 cells may be caused by an increase in GlcNAc transferase V activity, similar to that observed previously in transformed cell lines (21, 31) and human carcinomas (32, 33). Initial experiments measuring GlcNAc transferase V activity in F9 cells with radiolabeled UDP-GlcNAc and GlcNAc~l-2Manal-6Man~1-O(CH2)~C00CH~ showed 30 pmol/mg/h in F9 cell lysates and a 3-fold increase in enzyme activity in cells treated with loe6 M RA for 4 days (Fig. 1). The acceptor K , was 0.075 mM, similar for RA-treated and untreated F9 cells, as well as that reported previously for GlcNAc transferase V activity in baby hamster kidney cell lysates (34).
Since GlcNAc transferase V activity is often low compared with most glycosyltransferases, a more sensitive assay using the synthetic substrate GlcNAc~l-2Manal-GMan~l- O(CH2)&O-BSA and an ELISA for detection of GlcNAc transferase V product was used in subsequent experiments (28). The specificity of the ELISA was tested using Chinese hamster ovary cells and the GlcNAc transferase V-deficient Lec4 cell lines (Fig. 2). Titration of the Chinese hamster ovary cell lysate produced a typical second order curve, at protein concentrations approximately 100-fold less than that used in assays using radiolabeled UDP-GlcNAc, whereas Lec4 cell lysates showed essentially no GlcNAc transferase V activity. Linear curves were created by expressing activity (i.e. A410) as a log function of protein concentration and used to calculate reaction rates (Fig. 3). For comparison, the specific activity of GlcNAc transferase V in Chinese hamster ovary and F9 cell lysates was 0.20 and 0.02 Ak1O/pg/h, compared with 250

@l-GGlcNAc Transferases and Polylactosamine in F9 Cells 1245
65 - 60 - 55 - 50 - 45 - 40 - 35 - 30 - 25 -
I 1 1 1
-20 -15 -10 -5 0 5 10 15 20 25
1/[Sl (mu)
FIG. 1. GlcNAc transferase activity in F9 (A) and F9 + RA (m) was determined as a function of acceptor concentration and is shown as a Lineweaver-Burk plot. The assay contained 1 mM UDP-[3H]GlcNAc and was incubated a t 37 “C for 2 h as described under “Materials and Methods.”
0.7 I I
0.6 I / CHo I
0 10 20 30 40
Protein (up)
FIG. 2. GlcNAc transferase V activity in Chinese hamster ovary (O), Lec4 (V), F9 (Y), and RA-treated F9 (0) cell lysates was measured with the ELISA as described under “Materials and Methods.”
and approximately 30 pmol/mg/h using radiolabeled UDP- GlcNAc.
Increased expression of LAMP-1- and L-PHA-reactive ol- igosaccharides in F9 cells was detected previously at lo-’ M and optimal a t loT6 M RA (19). GlcNAc transferase V activity showed a similar dose-dependent increase, with maximal ac- tivity at IOw7 M RA ( i e . a 5.5-fold increase) (Fig. 3). The 0- linked glycosyltransferases core 2 GlcNAc transferase and P1-3Gal transferase also showed a dose-dependent increase in activity with a 12- and 3-fold increase a t M RA, respectively (Fig. 3B). The RA dose response for core 2 GlcNAc transferase (Fig. 3C) was similar to that of GlcNAc transferase V, whereas P l J G a l transferase required higher concentrations of RA > lo-’ M (Fig. 3 0 ) similar to that reported for induction of the LAMP-2 in F9 cells (19).
Time courses for RA-stimulated induction of GlcNAc trans- ferase V, core 2 GlcNAc transferase, P1-3Gal transferase, and P1-4Gal transferase activities showed biphasic responses with a n increase occurring over days 1-3 followed by a further more precipitous increase on 4th day after RA treatment (Fig. 4). A similar time course was observed for induction of L-
PHA-reactive glycoconjugates (i.e. LAMP-1, LAMP-2, and laminin), with a small increase on day 3 and the major change 4 days after RA treatment (Fig. 5). Under the same experi- mental conditions, induction of laminin B1, LAMP-1, and LAMP-2 also began 2-3 days after RA treatment (19). In human and mouse embryonal carcinoma cells, RA treatment results in the sequential and ordered induction of Hox 2 genes begins within 3 h and extending over 5 days (5, 6). The biphasic induction of glycosyltransferases may also be caused by sequential regulatory events in this complex process.
The data in Fig. 6 summarize the fold increase in GlcNAc transferase V, core 2 GlcNAc transferase, P1-4Gal transfer- ase, and P1-3Gal transferase as well as GlcNAc transferase I and GlcNAc transferase (i) activities after 4 days in the presence of M RA. GlcNAc transferase V and core 2 GlcNAc transferase showed the largest induction, 6- and 12- fold, respectively, whereas increases in the other glycosyl- transferases were 2-3.6-fold. GlcNAc transferase V assay using UDP-[3H]GlcNAc showed only a 3-fold increase com- pared with 6-fold with the ELISA. The difference may be because of the concentration of UDP-GlcNAc used in the two assays. The ELISA contained 4 mM UDP-GlcNAc. The ra- dioactive assay was performed at 1 mM, approximately the K,,, for UDP-GlcNAc (34), and may therefore have underes- timated the difference between enzyme activity in untreated and RA-treated cells.
To examine further the relationship between F9 cell differ- entiation and glycosyltransferase induction, cells were treated with agents previously shown to have distinct effects on F9 differentiation pathways. RA induces F9 cells to differentiate into primitive endoderm, Bt2cAMP plus RA induces parietal endoderm, and Bt2cAMP alone does not induce differentia- tion (35). In contrast, butyrate has been shown to inhibit RA- dependent differentiation of F9 cells (36). Consistent with this relationship, induction of GlcNAc transferase V and core 2 GlcNAc transferase activities was RA > RA + butyrate > BtzcAMP (Fig. 7A). RA + Bt,cAMP stimulated GlcNAc transferase V activity, but less than RA alone. Butyrate alone also showed intermediate levels of GlcNAc transferase V and core 2 GlcNAc transferase stimulation, which suggests that it may affect F9 cell differentiation in a manner independent of RA. The patterns of induction of GlcNAc transferase V and core 2 GlcNAc transferase activities in F9 cells treated with combinations of RA, butyrate, Bt2cAMP were similar for these two enzymes and differed from that of the Gal transfer- ases. P1-4Gal transferase and P1-3Gal transferase activities showed a 2-3-fold induction with all treatments except RA + BtZcAMP, in which induction was 3-4-fold (Fig. 7). L-PHA reactivity of glycoproteins from F9 cells treated with these agents correlated with GlcNAc transferase V activity in cells treatment with RA or with RA + Bt2cAMP as shown in Fig. 5. However, there was no increase in L-PHA binding in cells treated with butyrate alone.
PYS-2 and PSA-5E are murine embryonal lines with pari- etal and visceral phenotypes, respectively (37), and both show L-PHA reactivity comparable to RA-treated F9 cells (19). Consistent with this phenotype, both PYS-2 and PSA-5E cells showed elevated levels of GlcNAc transferase V activity similar to that of RA-induced F9 cells (Table I). PYS-2 cells also had core 2 GlcNAc transferase activity comparable to that of RA-treated F9 cells, but the visceral endoderm line PSA-5E showed a distinctive phenotype with very low levels of core 2 GlcNAc transferase activity, below that of untreated F9 cells. Differentiation into parietal endoderm, represented by both the PYS-2 cell line and RA + BtzcAMP-treated F9 cells, was distinguished by elevated P1-4Gal transferase and

1246 pl-6GlcNAc Transferases and Polylactosamine i n F9 Cells
FIG. 3. Dose-response curve for RA-induced increase in GlcNAc transferasev (panelsA and B ) , core 2 GlcNAc transferase (panel C), and 01-3Gal transferase (panel D ) ac- tivities in F9 teratocarcinoma cells. F9 cells were treated with various con- centrations of RA (0) or 0.01% Me2S0 (0) for 4 days. GlcNAc transferase V activity was measured at seven lysate protein concentrations using the ELISA. Panel A shows each GlcNAc transferase V activity curve as a function of log[protein], and in panel B the data are expressed as specific activity versus RA concentration. 0, V, V, 0, W, and A in panel A are in order of decreasing RA concentration.
0.10
Y s 0.05
$ 0.04 -
2 0.03 - 0.02
-
-
> -
O.O1 0.00 0 L 1 2 3 4 5
1 .o 0.9
A 0.125
B
< a 0.4 Y
._
0.2
0.1
0.0 1 10 100
Protein (up)
5 C 1
V
01 I 5 6 7 8 9 1 0 1 1
RA Concentrotion (-IogM)
0.100 -
s 5 0.075
-
d .- * 0.050 - n
.- I U <
0.025 -
0.000 1 5 6 7 8 9 1 0 1 1
V
RA Concentration (-IoqM)
5
01 5 6 7 8 9 1 0 1
RA Concentration (-IoqM)
6 I
C 5
FIG. 4. Time course for RA-de-
ferase V (panel A ) , 81-4Gal trans- ferase (panel B ) , core 2 GlcNAc
transferase (panel D ) in F9 cells. F9
:: 1 - pendent induction of GlcNAc trans-
0 1 2 3 4 5 transferase (panel C), and 81-3Gal 0 .
40 cells were treated with M RA (0) or 5 .
B vested, and glycosyltransferases activi- 0.01% Me2S0 (V). The cells were har-
ties were measured as described under "Materials and Methods."
I J : v ; D / J I 30
20
+ P .
v , o 1 1 " 10 - z -
:" 1 -
0 0 1 2 3 4 5 0
Tim. (day.) Time (Days)
1 2 3 4 5 0
P1-3Gal transferase activities compared with primitive en- Glycosyltransferase activities were measured in RA-3-10, a doderm (RA-treated F9) (Table I and Fig. 7). The results RA-resistant F9 cell line which does not differentiate into suggest that embryonal cell differentiation is closely associ- parietal endoderm in the presence of RA (38). Partial induc- ated with changes in glycosyltransferase activity, which are tion of GlcNAc transferase V activity (ie. 2-fold) was observed characteristic of the differentiated cell lineage. following treatment of RA-3-10 cells for 4 days with M
/31-6GlcNAc Transferases and Polylactosamine in F9 Cells
L 1RO
e116
Core
/ t I / I
" 0 1 2 3 4 5
Tlnle (Days)
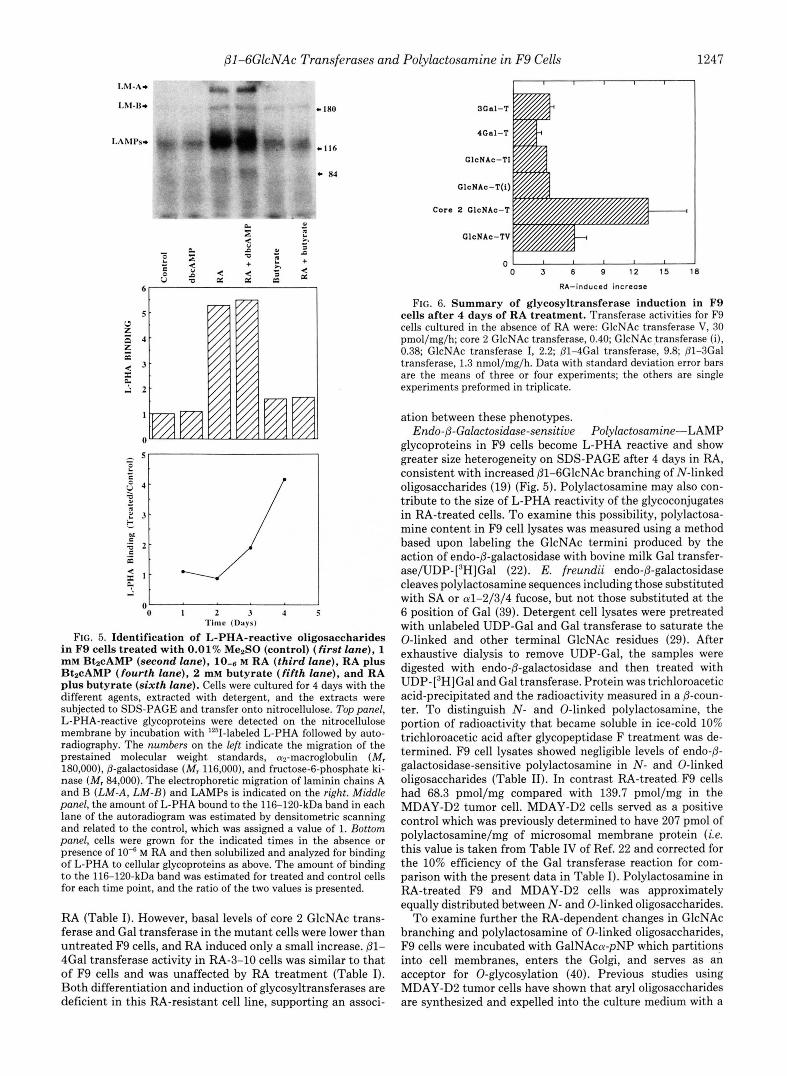
FIG. 5. Identification of L-PHA-reactive oligosaccharides in F9 cells treated with 0.01% Me2S0 (control) (first lane), 1 m M BtzcAMP (second lane), lods M RA (third lane), RA plus Bt2cAMP (fourth lune), 2 mM butyrate ( f i f th lune), and RA plus butyrate (sixth lane). Cells were cultured for 4 days with the different agents, extracted with detergent, and the extracts were subjected to SDS-PAGE and transfer onto nitrocellulose. Top panel, L-PHA-reactive glycoproteins were detected on the nitrocellulose membrane by incubation with 12sII-labeled L-PHA followed by auto- radiography. The numbers on the left indicate the migration of the prestained molecular weight standards, cu2-macroglobulin (M, 180,000), 8-galactosidase (M, 116,000), and fructose-6-phosphate ki- nase (M, 84,000). The electrophoretic migration of laminin chains A and B (LM-A, LM-B) and LAMPS is indicated on the right. Middle panel, the amount of L-PHA bound to the 116-120-kDa band in each lane of the autoradiogram was estimated by densitometric scanning and related to the control, which was assigned a value of 1. Bottom panel, cells were grown for the indicated times in the absence or presence of M RA and then solubilized and analyzed for binding of L-PHA to cellular glycoproteins as above. The amount of binding to the 116-120-kDa band was estimated for treated and control cells for each time point, and the ratio of the two values is presented.
RA (Table I). However, basal levels of core 2 GlcNAc trans- ferase and Gal transferase in the mutant cells were lower than untreated F9 cells, and RA induced only a small increase. 61- 4Gal transferase activity in RA-3-10 cells was similar to that of F9 cells and was unaffected by RA treatment (Table I). Both differentiation and induction of glycosyltransferases are deficient in this RA-resistant cell line, supporting an associ-
JGal-T
4Gal-T
GlcNAc-TI
GlcNAc-T( i )
2 GIcNAc-T
ClcNAc-TV
-I
1247
RA-induced increase
FIG. 6. Summary of glycosyltransferase induction in F9 cells after 4 days of RA treatment. Transferase activities for F9 cells cultured in the absence of RA were: GlcNAc transferase V, 30 pmol/mg/h; core 2 GlcNAc transferase, 0.40; GlcNAc transferase (i), 0.38; GlcNAc transferase I, 2.2; 81-4Gal transferase, 9.8; 81-3Gal transferase, 1.3 nmol/mg/h. Data with standard deviation error bars are the means of three or four experiments; the others are single experiments preformed in triplicate.
ation between these phenotypes. Endo-6-Galactosidase-sensitive Polylactosamine-LAMP
glycoproteins in F9 cells become L-PHA reactive and show greater size heterogeneity on SDS-PAGE after 4 days in RA, consistent with increased 61-6GlcNAc branching of N-linked oligosaccharides (19) (Fig. 5). Polylactosamine may also con- tribute to the size of L-PHA reactivity of the glycoconjugates in RA-treated cells. To examine this possibility, polylactosa- mine content in F9 cell lysates was measured using a method based upon labeling the GlcNAc termini produced by the action of endo-6-galactosidase with bovine milk Gal transfer- a~e /uDP-[~H]Gal (22). E. f r eud i i endo-&galactosidase cleaves polylactosamine sequences including those substituted with SA or a1-2/3/4 fucose, but not those substituted at the 6 position of Gal (39). Detergent cell lysates were pretreated with unlabeled UDP-Gal and Gal transferase to saturate the 0-linked and other terminal GlcNAc residues (29). After exhaustive dialysis to remove UDP-Gal, the samples were digested with endo-@-galactosidase and then treated with UDP-[3H]Gal and Gal transferase. Protein was trichloroacetic acid-precipitated and the radioactivity measured in a 6-coun- ter. To distinguish N- and 0-linked polylactosamine, the portion of radioactivity that became soluble in ice-cold 10% trichloroacetic acid after glycopeptidase F treatment was de- termined. F9 cell lysates showed negligible levels of endo-6- galactosidase-sensitive polylactosamine in N- and 0-linked oligosaccharides (Table 11). In contrast RA-treated. F9 cells had 68.3 pmol/mg compared with 139.7 pmol/mg in the MDAY-D2 tumor cell. MDAY-D2 cells served as a positive control which was previously determined to have 207 pmol of polylactosamine/mg of microsomal membrane protein (i.e. this value is taken from Table IV of Ref. 22 and corrected for the 10% efficiency of the Gal transferase reaction for com- parison with the present data in Table I). Polylactosamine in RA-treated F9 and MDAY-D2 cells was approximately equally distributed between N- and 0-linked oligosaccharides.
To examine further the RA-dependent changes in GlcNAc branching and polylactosamine of 0-linked oligosaccharides, F9 cells were incubated with GalNAca-pNP which partitions into cell membranes, enters the Golgi, and serves as an acceptor for 0-glycosylation (40). Previous studies using MDAY-D2 tumor cells have shown that aryl oligosaccharides are synthesized and expelled into the culture medium with a
1248 (?l-GGkNAc Transferases and Polylactosamine in F9 Cells
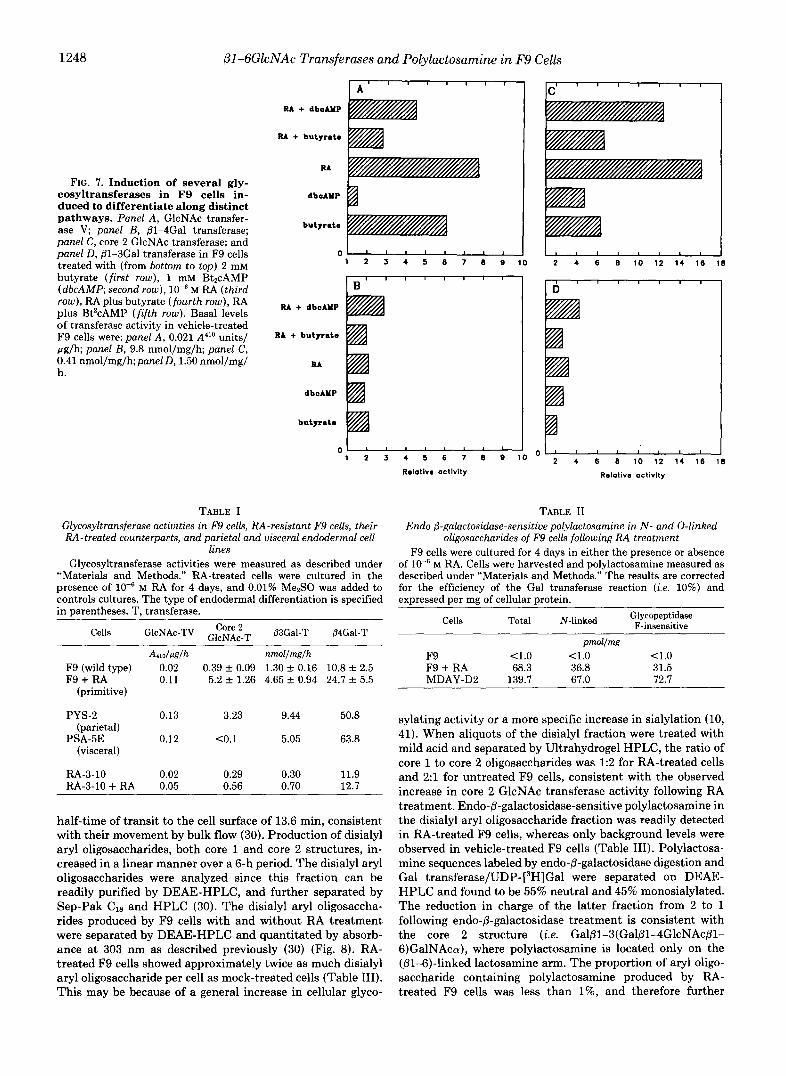
FIG. 7. Induction of several gly- cosyltransferases in F9 cells in- duced to differentiate along distinct pathways. Panel A, GlcNAc transfer- ase V; panel B, (3-4Gal transferase; panel C, core 2 GlcNAc transferase; and panel D, B1-3Gal transferase in F9 cells treated with (from bottom to top) 2 mM butyrate (f irst row), 1 mM Bt2cAMP (dbcAMP; second row), M RA (third row), RA plus butyrate (fourth row), RA plus Bt’cAMP (f i f th row). Basal levels of transferase activity in vehicle-treated F9 cells were: panel A , 0.021 A”” units/ pg/h; panel B, 9.8 nmol/mg/h; panel C, 0.41 nmol/mg/h;panel D, 1.50 nmol/mg/ h.
RA + dboAMP
RA + butyrate
RA
dbcAYP
butyrate
0 1 2 3 4 5 8 7 8 9 1 0
RA + dbcAYP
RA + butyrate
RA
dbaAYP
butyrata
u 2 4 6 E 10 12 14 16 18
1 2 3
TABLE I Glycosyltransferase activities i n F9 cells, RA-resistant F9 cells, their RA-treated counterparts, and parietal and visceral endodermal cell
lines Glycosyltransferase activities were measured as described under
“Materials and Methods.” RA-treated cells were cultured in the presence of M RA for 4 days, and 0.01% Me2S0 was added to controls cultures. The type of endodermal differentiation is specified in parentheses. T, transferase.
Cells GlcNAc-TV Ggi:”T P3Gal-T P4Gal-T
A m l d h nmollmglh F9 (wild type) 0.02 0.39 f 0.09 1.30 f 0.16 10.8 f 2.5 F9 + RA 0.11 5.2 f 1.26 4.65 k 0.94 24.7 k 5.5
(primitive)
PYS-2 0.13 3.23 9.44 50.8
PSA-5E 0.12 <0.1 5.05 63.8 (parietal)
(visceral)
RA-3-10 0.02 0.29 0.30 11.9 RA-3-10 + RA 0.05 0.56 0.70 12.7
half-time of transit to the cell surface of 13.6 min, consistent with their movement by bulk flow (30). Production of disialyl aryl oligosaccharides, both core 1 and core 2 structures, in- creased in a linear manner over a 6-h period. The disialyl aryl oligosaccharides were analyzed since this fraction can be readily purified by DEAE-HPLC, and further separated by Sep-Pak CIS and HPLC (30). The disialyl aryl oligosaccha- rides produced by F9 cells with and without RA treatment were separated by DEAE-HPLC and quantitated by absorb- ance at 303 nm as described previously (30) (Fig. 8). RA- treated F9 cells showed approximately twice as much disialyl aryl oligosaccharide per cell as mock-treated cells (Table 111). This may be because of a general increase in cellular glyco-
Relative activity Relative activity
TABLE I1 Endo 0-galactosidase-sensitive polylactosamine i n N - and 0-linked
oligosaccharides of F9 cells following R A treatment F9 cells were cultured for 4 days in either the presence or absence
of M RA. Cells were harvested and polylactosamine measured as described under “Materials and Methods.” The results are corrected for the efficiency of the Gal transferase reaction (i.e. 10%) and expressed per mg of cellular protein.
Cells Total N-linked Glwpeptidase F-insensitive
pmollmg F9 4 . 0 4 . 0 4 . 0 F9 + RA 68.3 36.8 31.5 MDAY-D2 139.7 67.0 72.7
sylating activity or a more specific increase in sialylation (10, 41). When aliquots of the disialyl fraction were treated with mild acid and separated by Ultrahydrogel HPLC, the ratio of core 1 to core 2 oligosaccharides was 1:2 for RA-treated cells and 2:l for untreated F9 cells, consistent with the observed increase in core 2 GlcNAc transferase activity following RA treatment. Endo-@-galactosidase-sensitive polylactosamine in the disialyl aryl oligosaccharide fraction was readily detected in RA-treated F9 cells, whereas only background levels were observed in vehicle-treated F9 cells (Table 111). Polylactosa- mine sequences labeled by endo-@-galactosidase digestion and Gal tran~ferase/UDP-[~H]Gal were separated on DEAE- HPLC and found to be 55% neutral and 45% monosialylated. The reduction in charge of the latter fraction from 2 to 1 following endo-@-galactosidase treatment is consistent with the core 2 structure (i.e. Gal@l-3(Gal@l-4GlcNAc@1- 6)GalNAca), where polylactosamine is located only on the (@1-6)-linked lactosamine arm. The proportion of aryl oligo- saccharide containing polylactosamine produced by RA- treated F9 cells was less than 1%, and therefore further
fi1-6GlcNAc Transferases and Polylactosamine in F9 Cells 1249
2 N 1 2 + + + 1
0.0 22.5 45.0 0.0 22.5 45.0
Time (min.) nme (mln.)
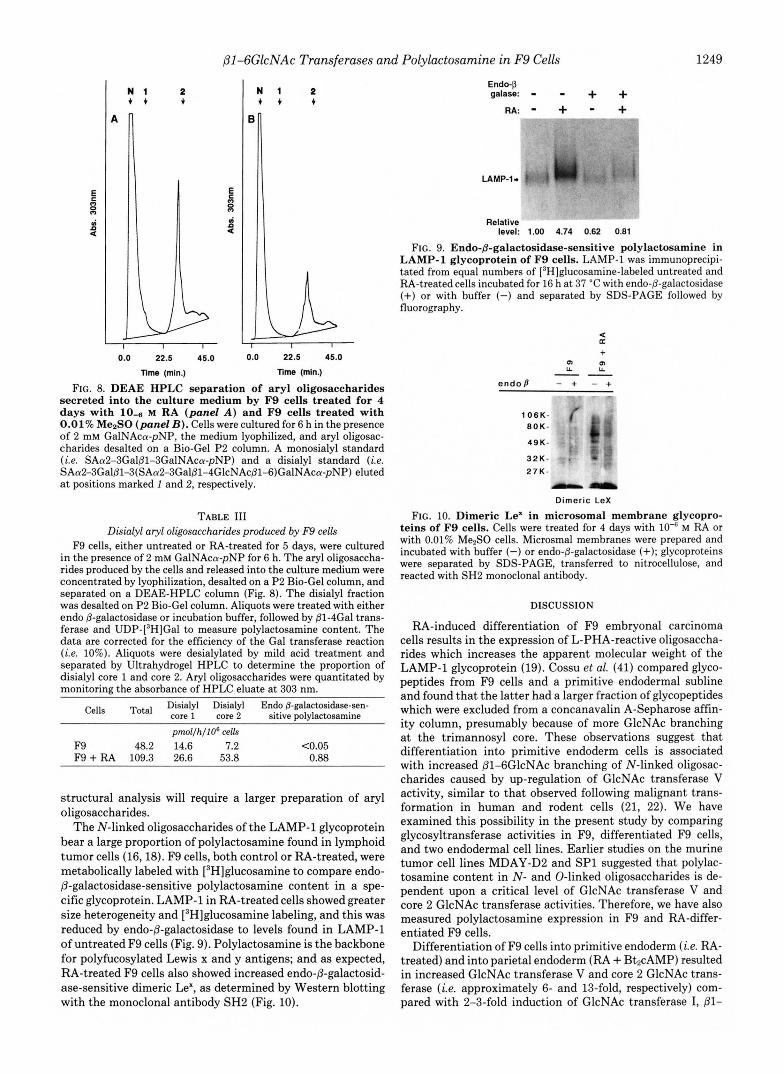
FIG. 8. DEAE HPLC separation of aryl oligosaccharides secreted into the culture medium by F9 cells treated for 4 days with 10-6 M RA (panel A ) and F9 cells treated with 0.01% Me2S0 (panel 23). Cells were cultured for 6 h in the presence of 2 mM GalNAca-pNP, the medium lyophilized, and aryl oligosac- charides desalted on a Bio-Gel P2 column. A monosialyl standard (i.e. SAa2-3Galj31-3GalNAca-pNP) and a disialyl standard (i.e. SAa2-3Galp1-3(SAa2-3Gal@l-4GlcNAc@1-6)GalNAca-pNP) eluted at positions marked 1 and 2, respectively.
TABLE 111 Disialyl aryl oligosaccharides produced by F9 cells
F9 cells, either untreated or RA-treated for 5 days, were cultured in the presence of 2 mM GalNAca-pNP for 6 h. The aryl oligosaccha- rides produced by the cells and released into the culture medium were concentrated by lyophilization, desalted on a P2 Bio-Gel column, and separated on a DEAE-HPLC column (Fig. 8). The disialyl fraction was desalted on P2 Bio-Gel column. Aliquots were treated with either endo @-galactosidase or incubation buffer, followed by @1-4Gal trans- ferase and UDP-[3H]Gal to measure polylactosamine content. The data are corrected for the efficiency of the Gal transferase reaction (i.e. 10%). Aliquots were desialylated by mild acid treatment and separated by Ultrahydrogel HPLC to determine the proportion of disialyl core 1 and core 2. Aryl oligosaccharides were quantitated by monitoring the absorbance of HPLC eluate a t 303 nm.
Cells Total Disialyl Disialyl Endo p-galactosidase-sen- core 1 core 2 sitive polylactosamine pmlfhflOs cells
F9 48.2 14.6 7.2 C0.05 F9 + RA 109.3 26.6 53.8 0.88
structural analysis will require a larger preparation of aryl oligosaccharides.
The N-linked oligosaccharides of the LAMP-1 glycoprotein bear a large proportion of polylactosamine found in lymphoid tumor cells (16,18). F9 cells, both control or RA-treated, were metabolically labeled with [3H]glucosamine to compare endo- @-galactosidase-sensitive polylactosamine content in a spe- cific glycoprotein. LAMP-1 in RA-treated cells showed greater size heterogeneity and [3H]glucosamine labeling, and this was reduced by endo-@-galactosidase to levels found in LAMP-1 of untreated F9 cells (Fig. 9). Polylactosamine is the backbone for polyfucosylated Lewis x and y antigens; and as expected, RA-treated F9 cells also showed increased endo-@-galactosid- ase-sensitive dimeric Le', as determined by Western blotting with the monoclonal antibody SH2 (Fig. 10).
Endo-P galase: - - + +
RA: * + - +
LAMP-1- !. i w .
Relative level: 1.00 4.74 0.62 0.81
FIG. 9. Endo-B-galactosidase-sensitive polylactosamine in LAMP-1 glycoprotein of F9 cells. LAMP-1 was immunoprecipi- tated from equal numbers of [3H]glucosamine-labeled untreated and RA-treated cells incubated for 16 h a t 37 "C with endo-@-galactosidase (+) or with buffer (-) and separated by SDS-PAGE followed by fluorography.
K +
m m L L L L "
endop - + - + - 7 - IC(
106K- 80K-
49K-
32K- 27K-
" Dimeric Lex
FIG. 10. Dimeric Le' in microsomal membrane glycopro- teins of F9 cells. Cells were treated for 4 days with 10"' M RA or with 0.01% Me2S0 cells. Microsmal membranes were prepared and incubated with buffer (-) or endo-p-galactosidase (+); glycoproteins were separated by SDS-PAGE, transferred to nitrocellulose, and reacted with SH2 monoclonal antibody.
DISCUSSION
RA-induced differentiation of F9 embryonal carcinoma cells results in the expression of L-PHA-reactive oligosaccha- rides which increases the apparent molecular weight of the LAMP-1 glycoprotein (19). Cossu et al. (41) compared glyco- peptides from F9 cells and a primitive endodermal subline and found that the latter had a larger fraction of glycopeptides which were excluded from a concanavalin A-Sepharose affin- ity column, presumably because of more GlcNAc branching at the trimannosyl core. These observations suggest that differentiation into primitive endoderm cells is associated with increased Dl-6GlcNAc branching of N-linked oligosac- charides caused by up-regulation of GlcNAc transferase V activity, similar to that observed following malignant trans- formation in human and rodent cells (21, 22). We have examined this possibility in the present study by comparing glycosyltransferase activities in F9, differentiated F9 cells, and two endodermal cell lines. Earlier studies on the murine tumor cell lines MDAY-D2 and SP1 suggested that polylac- tosamine content in N- and 0-linked oligosaccharides is de- pendent upon a critical level of GlcNAc transferase V and core 2 GlcNAc transferase activities. Therefore, we have also measured polylactosamine expression in F9 and RA-differ- entiated F9 cells.
Differentiation of F9 cells into primitive endoderm ( i e . RA- treated) and into parietal endoderm (RA + Bt2cAMP) resulted in increased GlcNAc transferase V and core 2 GlcNAc trans- ferase ( i e . approximately 6- and 13-fold, respectively) com- pared with 2-3-fold induction of GlcNAc transferase I, @l-
i i\ .i "

1250 pl-GGlcNAc Transferases and Polylactosamine in F9 Cells
3Gal transferase, @1-4Gal transferase, and GlcNAc transfer- ase (i). GlcNAc transferase I, @1-3Gal transferase are widely expressed and involved in the biosynthesis of core N- and 0- linked oligosaccharide sequences, respectively. The differen- tiation-associated increase in their activities suggests a gen- eral increase in cellular glycosylation capacity. This may account in part for the observed increase in disialyl aryl oligosaccharides made by RA-treated F9 cells compared with untreated F9 cells. However, elevated glycosyltransferase ac- tivity is not caused by a general increase in catabolism, as [’Hlleucine incorporation into proteins is similar in untreated and RA-treated F9 cells (42).
01-4Gal transferase and GlcNAc transferase (i) are re- quired for polylactosamine biosynthesis, and although the change in their activities paralleled that of the ubiquitous GlcNAc transferase I and p1-3Gal transferase enzymes, the larger increase in GlcNAc transferase V and core 2 GlcNAc transferase may be responsible for the observed up-regulation of polylactosamine content of 0- and N-linked oligosaccha- rides. The increase in GlcNAc transferase V when compared with the increase in other enzymes examined was only 2-fold greater. However, as little as a 2-3-fold increase in GlcNAc transferase V activity in the SP1 tumor cells and T24H-ras- transfected rat2 fibroblasts is associated with a dramatic increase in L-PHA reactivity of membrane glycoproteins in- cluding LAMP-1 (21, 31) and greatly increased polylactosa- mine levels, whereas @1-4Gal transferase and GlcNAc trans- ferase (i) were not altered (22). Conversely, a 3-4-fold reduc- tion in GlcNAc transferase V activity in KBL-1 cells, a somatic mutant of the MDAY-D2 lymphoma, results in al- most complete loss of polylactosamine in N-linked, whereas 0-linked polylactosamine remained unchanged (22). Endo-@-galactosidase-sensitive polylactosamine in mem-
brane glycoconjugates and LAMP-1 increased dramatically following RA-dependent differentiation of F9 cells, an obser- vation consistent with those made earlier on whole mouse embryos using antibodies reactive with “i” antigen (43). Endo- &galactosidase is inhibited by six substitutions to Gal, such as “I” antigen (-GlcNAc@l-3(GlcNAc@l-6)Gal@l-4Glc- NAc-), a structure which has been detected in cultures of untreated F9 cells (7, 8). If “I” structures saturate the poly- lactosamine in our untreated F9 cells, endo-@-galactosidase would not be useful in detection of polylactosamine. However, this is unlikely, since Childs et al. (8) found that branching at Gal does not saturate polylactosamine chains present in un- treated F9 cells as the majority of I antigen found in the cells was sensitive to endo-@-galactosidase digestion (8). Therefore, our failure to detect endo-@-galactosidase-sensitive sequences in untreated F9 cell line used in the present experiments suggests that the cells have very little polylactosamine and therefore are phenotypically similar to the 16-cell or early embryonic stage which F9 is thought to resemble (44). How- ever, embryonal carcinoma cell lines do not show patterns of carbohydrate antigens identical to any specific cell type in the mouse embryo. The low expression of i antigen detected in F9 cells has been suggested to correspond to the subpopulation of spontaneously differentiating cells known to occur in em- bryonic carcinoma cell cultures (8).
RA-induced F9 cells and several endodermal cell lines strongly express endo-6-galactosidase-sensitive i antigen sim- ilar to that observed in 5-8-day embryonic and extraem- bryonic endoderm in uiuo. Pennington et al. (43) performed immunohistochemistry with anti-carbohydrate antibodies and found that i antigen (Le. polylactosamine) was first expressed in endoderm, ectoderm, and extraembryonic endo- derm but not mesoderm cells of the 5-day mouse embryo,
increased on day 6, and persisted to day 8. Le” and Ley antigens were observed in the same cells at approximately the same time, and their detection was enhanced by removal of terminal Galal-3 and sialic acid residues. This may be be- cause of a combination of increases in glycosyltransferase activities, which synthesize the terminal antigenic sequences, and increased Dl-6-branching GlcNAc transferase activities. Regarding the former enzymes, F9 cells treated with M RA for 3 days showed a %fold increase in al-3Gal transferase activity (11). Human embryonal carcinoma cells treated with RA for 4 days showed a 3-fold increase in a2-3 sialyltransfer- ase and a 2-fold increase al-3 fucosyltransferase, which may account for the conversion from Le“ to SLe” following RA treatment (10,45). Similarly, RA induced a 3-fold increase in fucosyltransferase in F9 cells (42) and a 2-3-fold increase in sialyltransferase activity in melanoma cells (46). P1-4Gal transferase mRNA levels have been shown to increase 2-3- fold in parallel with enzyme activity in RA-treated F9 cells, suggesting a differentiation-dependent increase in gene tran- scription or mRNA stability (47). The magnitude of change in these transferase activities is similar to that observed in the present study for the ubiquitous GlcNAc transferase I and @1-3Gal transferase. Therefore, the relatively greater stimu- lation of the branching GlcNAc transferases may be an im- portant element regulating the levels of the terminal antigenic sequences associated with polylactosamine. In this regard, the majority of SLe”, LeY, and Galal-3Gal sequences were found on endo-@-galactosidase-susceptible polylactosamine se- quences in the glycoprotein fraction of developing mouse embryos (43) and embryonal carcinoma cells (8).
Glycosyltransferase activity in the parietal endoderm cell line PYS-2 was similar to that of RA-treated F9 cells, sug- gesting that changes are not strictly RA-dependent but are associated with cell differentiation. The visceral endoderm cell line PAS-5E showed a pattern of glycosyltransferase activity similar to that of RA + BtpcAMP-treated F9 cells, with the exception that core 2 GlcNAc transferase was very low in PAS-5E. This may reflect a further phase of differen- tiation occurring after that of 4-day RA + BtzcAMP-treated F9 cells. Based on expression of laminin, collagen, and Hox genes in RA-treated F9 cells, differentiation appears to require ordered and interdependent changes in gene expression (4-6) and phenotypic changes, particularly in glycosyltransferase activities which extend up to and beyond 4 days (42, 47).
In conclusion, the results of this study demonstrate that differentiation of F9 cells into distinct types of endodermal cells is accompanied by marked and characteristic changes in specific glycosyltransferases and in polylactosaminoglycans. Developmental regulation of polylactosamine and associated Lewis antigens may have functional consequences for cellular interactions during embryogenesis (13,141. Transgenic exper- iments will be required to test this hypothesis, as has recently been done by Varki et al. (48) to explore the role of 9-0- acetylneuraminic acid in murine embryogenesis. The mecha- nism(s) by which RA alone or in combination with BtzcAMP increases the activity of several glycosyltransferases is not known. It is likely that RA regulates the transcription of their genes via nuclear retinoic acid receptors (49-51). The eluci- dation of such a mechanism must await the cloning of these enzymes and the isolation and analysis of their promoters.
Acknowledgments-We thank Dr. Lorraine J. Gudas for the F-9 (wild type) and RA-resistant mutant RAS-3-10 cells, Dr. Angie Rizzino for the PYS-2 and PSA-5E cells, and Zofia Krzyzek for secretarial assistance.

P1-6GlcNAc Transferases and Polylactosamine in F9 Cells 1251
1. 2. 3.
4.
5.
6.
7.
8.
9.
10.
11. 12.
13. 14.
15.
16.
18. 17.
19. 20. 21.
22.
23.
24.
25.
REFERENCES Martin, G. R. (1980) Science 209, 768-776 Strickland, S., and Mahdavi, V. (1978) Cell 16,393-403 Hogan, B. L. M., Barlow, D. P., and Tilly, R. (1983) Cancer Suru. 2, 115-
Rogers, M. B., Watkins, S. C., and Gudas, L. J. (1990) J. Cell BWL 110,
Simeone, A,, Acampora, D., Arcioni, L., Andrews, P. W., Boncinelli, E.,
Papalopulu, N., Lovell-Badge, R., and Krumlauf, R. (1991) Nucleic Acids
Kapadia, A., Feizi, T., and Evans, M. J. (1981) Exp. Cell Res. 131, 185-
140
1767-1777
and Mavilio, F. (1990) Nature 346, 763-766
Res. 19,54975506
Childs, R. A., Pennington, J., Uemura, K., Scudder, P., Goodfellow, P. N.,
Solter. D.. and Knowles. B. B. (1978) Proc. Natl. Acad. Sci. U. S. A. 75.
195
Evans, M. J., and Feizi, T. (1983) Biochem. J. 216,491-503
5565-5569
Res. 180,326-340
. ,
Rosenman, S. J., Fenderson, B. A,, and Hakomori, S.-I. (1989) Exp. Cell
Cummings, R. D., and Mattox, S. A. (1988) J. Biol. Chem. 263,511-519 Fenderson, B. A., Holmes, E. H., Fukushi, Y., and Hakomori, %-I. (1986)
Bird, J. M., and Kimber, S. J. (1984) Deu. Biol. 104,449-460 Fenderson, B. A,, Zehavi, U., and Hakomori, S. (1984) J. Exp. Med. 160,
Deu. Biol. 114,lZ-21
Chen, J. W., Pan, W., D'Souza, M. P., and August, J. T. (1985) Arch.
Laferte, S., and Dennis, J. W. (1989) Biochem. J. 269,569-576 Heffernan, M., and Dennis, J. (1989) Cancer Res. 49,6077-6084 Carlsson, S. R., and Fukuda, M. (1990) J. BWL Chem. 266,20488-20495 Amos, B., and Lotan, R. (1990) J. Biol. Chem. 266, 19192-19198
Dennis, J. W., Laferte, S., Waghorne, C., Breitman, M. L., and Kerbel, R. Cumrnings, R. D., and Kornfeld, S. (1982) J. Biol. Chem. 267,11230-11234
Yousefi, S., Higgins, E., Doaling, Z., Hindsgaul, O., Pollex-Kruger, A,, and
van den Eiinden. D. H.. Koenderman. A. H. L.. and Schiuhorst. W. E. C.
1591-1596
Biochem. Biophys. 239,547-586
S. (1987) Science 236,582-585
Dennis, J. W. (1991) J. BWL Chem. 266,1772-1783
M. (1988j J. Eiol. Chim. 263,12461-12465 '
261, 12796-12806
- .
Fukuda, M., Carlsson, S. R., Koek, J. C., and Dell, A. (1986) J. Biol. Chem.
26. Sin hal, A. K., Orntoft, T. F., Nudelman, E., Nance, S., Schibig, L., Stroud, J. W. (1991) J. Biol. Chem. 266,6280-6290
27. Kerbel, R. S., Florian, M. S., Man, S., Dennis, J. W., and McKenzie, I. d. R., Clausen, H., and Hakomori, S. (1990) Cancer Res. 60,1375-1380
28. Crawley, S. C., Hindsgaul, O., Alton, G., Pierce, M., and Palcic, M. M. (1980) J. Natl. Cancer Inst. 64,1221-1230
29. Torres, C. R., and Hart, G. W. (1984) J. Biol. Chem. 269,3308-3317 (1990) A d . Biochem. 186, 1-6
30. Zhuang, D., Grey, A., Harris-Brandts, M., Higgins, E., Kashem, M. A., and
31. Dennis, J. W., Kosh, K., Bryce, D.-M., and Breitman, M. (1989) Oncogene
33. Fernandes, B., Sagman, U.. Auger, M., Demetrio, M., and Dennis, J. W. 32. Dennis, J. W., and Laferte, S. (1989) Cancer Res. 49,945-950
34. Palcic, M. M., Ripka, J., Kaur, K. J., Shoreibah, M., Hindsgaul, O., and
35. Strickland, S., Smith, K. K., and Marotti, K. R (1980) Cell 21, 393-403 36. Grippo, J. F., and Gudas, L. J. (1987) J. Biol. Chem. 262,4492-4500 37. Lehman, J. M., Speers, W. C., Swartzendruber, D. E., and Pierce, G. B.
38. Wang, S.-Y., and Gudas, L. J. (1984) J. Biol. Chem. 269,5899-5906 39. Fukuda, M., and Fukuda, M. M. (1984) in The Biology of Glycoproteins
(Ivatt, R. J., ed) p. 183-234, Plenum Publishing Cop., New York 40. Kuan, S.-F., Byrd, j. C., Basbaum, C., and Kim, Y. S. (1989) J. Biol. Chem.
41. Cossu, G., Cortesi, E., and Warren, L. (1985) Differentiation 29.63-67 42. Amos, B., Lotan, D., and Lotan, R. (1990) Int. J. Cancer 46,86-94 43. Pennington, J. E., Rastan, S., Roelcke, D., and Feizi, T. (1985) J. Embryol.
44. Warren, L., Buck, C. A,, and Tuszynski, G. P. (1978) Biochim. Biophys.
45. Chen, C., Fenderson, B. A., Andrews, P. W., and Hakomori, S. (1989)
46. Deutsch, V., and Lotan, R. (1983) Exp. Cell Res. 149,237-245 47. Lo ez, L. C., Maillet, C. M., Oleszkowicz, K., and Shur, B. D. (1989) Mol.
48. Varki, A., Hooshmand, F., Diaz, S., Varki, N. M., and Hedrick, S. M. (1991)
49. Lotan, R., and Clifford, J. L. (1991) Eiomd. Phurmacother. 46, 145-156 50. Glass, C. K., DiRenzo, J., Kurokawa, R., and Han, Z. (1991) Cell Biol. 10,
Dennis, J. W. (1991) Glycobiology 1,425-433
4,853-860
(1991) Cancer Res. 61,718-723
Pierce, M. (1990) J. Biol. Chem. 266,6759-6769
(1992) J. Cell. Physiol. 84, 13-28
264 , 19271-19277
90,335-361
Acta 616,97-127
Biochemistry 28, 2229-2238
Cell. Biol. 9,2370-2377
Cell 66,65-74
623-638 Higgins, L. A., Siminovitch, K. A., Zhuang, D., Brockhausen, I., and Dennis, 51. DeLuca, L. M. (1991) FASEB J. 6,2924-2933
~~. ~~.