ca binding proteins a0001347-001-000.pdf
TRANSCRIPT

7/27/2019 Ca binding proteins A0001347-001-000.pdf
http://slidepdf.com/reader/full/ca-binding-proteins-a0001347-001-000pdf 1/8
Calcium-binding ProteinsBeat Schwaller, University of Fribourg, Fribourg, Switzerland
Calcium ions act as important second messengers for many intracellular processes and the
information contained in this calcium signal is modulated by specific calcium-binding
proteins. According to well-conserved structural elements these proteins can be groupedinto different families including annexins, C2 domain proteins and EF-hand proteins.
Physiological Calcium Concentrationsand the Calcium Ion as UbiquitousCytosolic Second Messenger
The extracellular concentration of Ca21 ions is approxi-mately 2.5 mmol L2 1 and is tightly regulated by the twohormones calcitonin and parathyroid hormone (PTH) inan antagonistic way. Calcitonin produced in the thyroid
gland lowers the Ca21
concentration by favouringformation of hydroxyapatite, while PTH stimulates theactivity of osteoclasts resulting in an increased concentra-tion of Ca21 . Secretion of PTH is regulated by levels of extracellular Ca21 sensed by calcium receptor protein(CaSR, a G protein-coupled receptor) which is expressedon the surface of parathyroid cells (Brown et al ., 1993). Asimilar yet unidentified receptor on osteoblasts is suggestedto transmit the extracellular Ca21 signal to stimulateosteoblastic proliferation resulting in increased boneformation. In addition, 1,25 dihydroxyvitamin D3 affectstheserumCa21 concentrationby increasing the resorptionof Ca21 from the intestine and the kidney.
The intracellular concentration of Ca21
in a resting cellis approximately four orders of magnitude lower (50– 100 nmolL21) than in the serum and its regulation isextremely sophisticated. All cells have in their plasmamembranes extrusion systems to remove intracellularCa21 , the most important being the Ca21 -ATPase andthe Na1 /Ca21 exchanger which either use ATP or thesodium gradient to drive Ca21 out of the cell, respectively.On the other hand, influx of Ca21 via opening of Ca21
channels in the plasma membrane or from activation of intracellular Ca21 -release channels such as inositol-1,4,5-trisphosphate (IP3) and ryanodine receptors of intracel-lular stores (e.g. endoplasmic reticulum) leads to an abrupt
increase in [Ca21
]i which is used in the cell as a cytosolicsecond messenger. The information contained in this signalis modulated by specific Ca21 -binding proteins (CaBP).The kinetics and geometry of Ca21 influx or release frominternal stores, the characteristics of soluble Ca21 -bindingproteins and the efficiency of Ca21 extrusion and/orreuptake into organelles determine the amplitude andshape of the Ca21 transients. This in turn determineswhich systems are activated by the second messengerCa21 .
Overview of Calcium-binding Proteins
One of the advantages of Ca21 as a second messenger is iability to tightly bind to proteins. Oxygen ligands, as founin carboxylic side-chain groups of aspartate or glutamator carbonyl oxygen atoms of the main-chain (or side-chaioxygen from the carbonyl group from asparagine anglutamine), bind well to Ca21 ions. A special ligand igamma-carboxyglutamate (g-Gla, Figure 5) with twcarboxylic side-chains present in some specific serumproteins (see below). Ca21 in a complex with oxygeatoms has a coordination of 6–8 ligands. This allows thformation of bonds with oxygen groups from differenamino acids of a protein, thus generatinga particularthreedimensional complex structure. The Ca21 -induced conformational change of a protein can lead to the activatioor inhibition of enzymatic activities or the binding tspecific ligands either in the presence or absence of bounCa21 .
According to their location (extracellular, cytosoli
inside of organelles), proteins capable of binding Ca
2
ions face a variety of completely different environmentFor example, the free Ca21 concentration is high in thextracellular space and in organelles, but extremely low ithe cytosol, and the pH and ionic composition of the milievary. Ca21 -binding proteins therefore have affinities foCa21 that are finely tuned to exert their precise biologicafunctions. Lately it has become clear that not only affinitiefor Ca21 (thermodynamic properties), but also thkinetics of binding and releasing of Ca21 ions arimportant parameters and it is necessary to understanthese before we can explain how a Ca21 signal can controevents of such extremely different time scales, such a
neurotransmitter release (10
2 5
s), muscle contractio(1023 s), metabolic reactions (102 1 s) and cell cyclerelated reactions (up to hours). To date the metal-bindinproperties of only a minority of the known CaBPs havbeen explored, and even fewer kinetic studies have beeperformed.
There is no one way in which proteins specifically binCa21 , but based on sequence and structure homologyCa21 -binding proteins can be grouped into differenfamilies. The ones discussed in detail in this chapte
Article Contents
Secondary article
. PhysiologicalCalcium Concentrationsand the Calcium
Ion as Ubiquitous Cytosolic Second Messenger
. Overview of Calcium-binding Proteins
. Families of Calcium-binding Proteins
. Transport of Calcium
ENCYCLOPEDIA OF LIFE SCIENCES / & 2001 Nature Publishing Group / www.els.net

7/27/2019 Ca binding proteins A0001347-001-000.pdf
http://slidepdf.com/reader/full/ca-binding-proteins-a0001347-001-000pdf 2/8
include the annexins, the C2 domain proteins, the EF-handfamily of CaBPs, the pentraxins and the vitamin K-dependent proteins. Furthermore, some low-affinity, high-capacity CaBPs present in intracellular organelles andserum will be described.
Families of Calcium-binding Proteins
Annexins
The annexins are a family of widely distributed Ca21 - andphospholipid-binding proteins, all of which possess aconserved repeated domain called the annexin fold(Burgoyne and Geisow, 1989). Family members containfour domains of 70–75 amino acids making up the proteincore, with the exception of annexin VI in which eight suchdomains are present (Figure 1). The core domain contains
the binding sites for Ca21 and negatively chargedphospholipids (phosphatidylserine). Each annexin foldconsists of five a-helical stretches joined by small linkerregions. Most of the initial structural information of annexins was obtained from the determination of thecrystal structure of annexin V (Huber et al ., 1990). MostrecentdataontheCa21 -binding stochiometrysuggestthatannexin V binds 10 Ca21 ions as determined from the X-ray structure of this protein co-crystallized with phospho-lipid head groups (Swairjo et al ., 1995). Different annexinsare distinguishedfrom one anotherby N-terminal domainsof variable length that may confer specific functions. Todate 10 mammalian annexins have been fully sequenced (I,
II, III, IV, V, VI, VII, VIII, XI and XIII). Of over 20members known, at least one can be detected in nearlyevery eukaryotic cell. The Ca21 -binding affinities of theannexins are greatly increased in the presence of phospho-lipids and vary widely between family members.
Annexins were initially considered to be intracellulacytosolic proteins involved in intracellular vesicular trafficexocytosis and endosome–endosome fusion. The higaffinity of annexin II for Ca21 (in the presence ophospholipids) made it an interesting candidate foinvolvement in exocytosis, but annexins I and VII alshave strong vesicle-aggregating activities. The ability o
annexin II to associate with cytoskeletal proteins hasuggested that annexins may regulate cytoskeletal interactions with membranes. Several annexins undergo transientranslocation to membranes (plasma membrane, secretorgranules) when cells are activated and [Ca21 ]i is elevatedPurified annexins (e.g. V and VII) can form pores isynthetic bilayers. Most annexins (with the exception oannexin V) are also substrates for protein kinases C (PKCfamily members of the C2 family of Ca21 -bindinproteins. The activity of cytosolic PKCs and of phospholipase A2 seems to be modulated by annexin V, suggestinthat this protein could play a role in Ca21 anphospholipid signalling pathways.
Although annexins have no apparent signal sequencefor secretion and the mechanism by which annexins artransported out of cells is as yet unknown, some anneximembers (I and II) have been detected in the serum and othe external surface of plasma membranes. This indicative of specific extracellular functions. The annexiII heterotetramer, consisting of two molecules of annexiII and two molecules of S100A10 (a member of the EFhand family), binds tissue-type plasminogen activato(tPA), plasminogen and plasmin. It accelerates thactivation of the clot-dissolving protease plasmin bcomplexing with plasminogen and tPA and the formatioof this complex suggests that the annexin II heterotetrame
is the key physiological receptor for plasminogen on thextracellular surface of endothelial cells.
Both up- and downregulation of annexins has beeobserved under pathological conditions. These proteincan have a suppressive action on some tumours whil
Figure 1 Structure of the annexins. (a) Annexins consist of an N-terminal domain of variable length and a protein core composed of four annexin folwhich contain thebindingsitesfor Ca21 and phospholipids.Boxes (a – e ) representahelices as observedin thecrystal structureof annexin V. Annexin VImade up from eight annexin folds. Shown here for annexin II, in theN-terminal region, phosphorylation sites andbinding sites for targets (S100A10) a
present.(b) Structureof annexinV, whichin thepresenceof phospholipidsbinds 10 moleculesof Ca21 . Modified fromHuber et al .(1990)andSwairjo et a
(1995). (Brookhaven Data Base, PDB entry code: 1A8A.)
Calcium-binding Proteins
2 ENCYCLOPEDIA OF LIFE SCIENCES / & 2001 Nature Publishing Group / www.els.net
7/27/2019 Ca binding proteins A0001347-001-000.pdf
http://slidepdf.com/reader/full/ca-binding-proteins-a0001347-001-000pdf 3/8
favouring tumour progression in others. The differenteffects may be linked to the transient expression patternsduring development and differentiation and are similar tothe pro- or anti-tumorigenic effects of proteins belongingto the EF-hand family of CaBPs.
C2 domain calcium-binding proteins
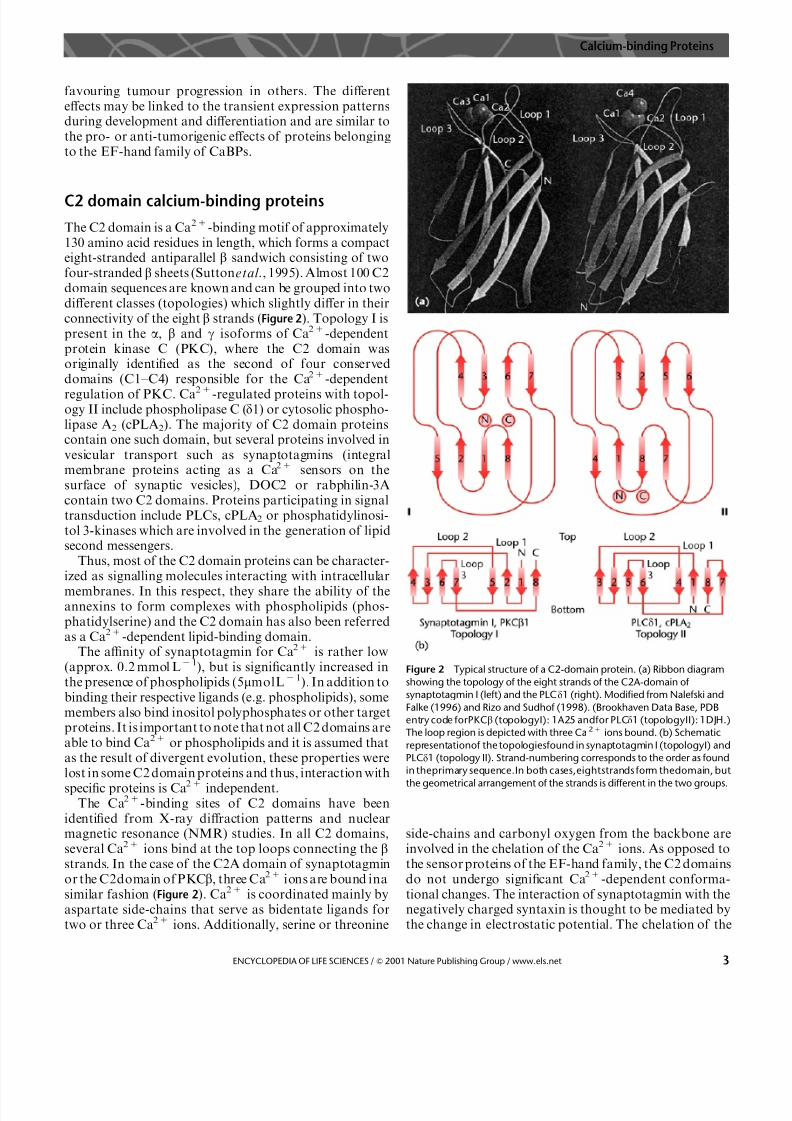
The C2 domain is a Ca21 -binding motif of approximately130 amino acid residues in length, which forms a compacteight-stranded antiparallel b sandwich consisting of twofour-stranded b sheets (Sutton etal ., 1995). Almost 100 C2domain sequences are known and can be grouped into twodifferent classes (topologies) which slightly differ in theirconnectivity of the eight b strands (Figure 2). Topology I ispresent in the a, b and g isoforms of Ca21 -dependentprotein kinase C (PKC), where the C2 domain wasoriginally identified as the second of four conserveddomains (C1–C4) responsible for the Ca21 -dependent
regulation of PKC. Ca21
-regulated proteins with topol-ogy II include phospholipase C (d1) or cytosolic phospho-lipase A2 (cPLA2). The majority of C2 domain proteinscontain one such domain, but several proteins involved invesicular transport such as synaptotagmins (integralmembrane proteins acting as a Ca21 sensors on thesurface of synaptic vesicles), DOC2 or rabphilin-3Acontain two C2 domains. Proteins participating in signaltransduction include PLCs, cPLA2 or phosphatidylinosi-tol 3-kinases which are involved in the generation of lipidsecond messengers.
Thus, most of the C2 domain proteins can be character-ized as signalling molecules interacting with intracellular
membranes. In this respect, they share the ability of theannexins to form complexes with phospholipids (phos-phatidylserine) and the C2 domain has also been referredas a Ca21 -dependent lipid-binding domain.
The affinity of synaptotagmin for Ca21 is rather low(approx. 0.2 mmol L2 1), but is significantly increased inthe presence of phospholipids (5mmolL2 1). In addition tobinding their respective ligands (e.g. phospholipids), somemembers also bind inositol polyphosphates or other targetproteins. It is important to note that not all C2 domains areable to bind Ca21 or phospholipids and it is assumed thatas the result of divergent evolution, these properties werelost in some C2 domain proteins and thus, interaction with
specific proteins is Ca21 independent.The Ca21 -binding sites of C2 domains have been
identified from X-ray diffraction patterns and nuclearmagnetic resonance (NMR) studies. In all C2 domains,several Ca21 ions bind at the top loops connecting the bstrands. In the case of the C2A domain of synaptotagminor the C2domain of PKCb, three Ca21 ions are bound inasimilar fashion (Figure 2). Ca21 is coordinated mainly byaspartate side-chains that serve as bidentate ligands fortwo or three Ca21 ions. Additionally, serine or threonine
side-chains and carbonyl oxygen from the backbone arinvolved in the chelation of the Ca21 ions. As opposed tthe sensor proteins of the EF-hand family, the C2 domaindo not undergo significant Ca21 -dependent conformational changes. The interaction of synaptotagmin with thnegatively charged syntaxin is thought to be mediated bthe change in electrostatic potential. The chelation of th
Figure 2 Typical structure of a C2-domain protein. (a) Ribbon diagram
showing the topology of the eight strands of the C2A-domain of synaptotagmin I (left) and the PLCd1 (right). Modified from Nalefski and
Falke (1996) and Rizo and Sudhof (1998). (Brookhaven Data Base, PDBentry code forPKCb (topologyI): 1A25 andfor PLCd1 (topologyII): 1DJHThe loop region is depicted with three Ca21 ions bound. (b) Schematic
representationof the topologiesfound in synaptotagmin I (topologyI) anPLCd1 (topology II). Strand-numbering corresponds to the order as founin theprimary sequence.In both cases,eightstrands form thedomain, b
the geometrical arrangement of the strands is different in the two group
Calcium-binding Proteins
ENCYCLOPEDIA OF LIFE SCIENCES / & 2001 Nature Publishing Group / www.els.net

7/27/2019 Ca binding proteins A0001347-001-000.pdf
http://slidepdf.com/reader/full/ca-binding-proteins-a0001347-001-000pdf 4/8
aspartates in the Ca21 -binding sites of synaptotagminthen allow the interaction of basic residues surroundingthis Ca21 -binding site with syntaxin.
A similar mechanism has been proposed for theinteraction with negatively charged phospholipids, sincereplacing the same acidic and basic residues in the C2Adomain of synaptotagmin inhibited binding of phospho-
lipids. It is proposed that the Ca21
-binding sites of C2domainproteins act as electrostatic switches. This might beimportant for very fast Ca21 -triggered reactions such asexocytosis (e.g. neurotransmitter release) because it doesnot necessitate large conformational changes of the C2domain proteins.
EF-hand proteins
The typical canonical EF-hand Ca21 -binding site consistsof a stretch of 29 amino acids consisting of an a helix(residues 1– 10), a loop around the Ca21 ion (residues 10–
21), and a seconda helix (residues 19–29).The second helixis oriented almost perpendicularto the first one (Kretsingerand Nockolds, 1973). The Ca21 ion is coordinated byseven oxygen ligands, derived from side-chains of aspar-tateand glutamate residues, carbonyl groups of the peptidebackbone and a bridging water molecule (Figure 3). Thegeometrical arrangement of the ligands can be described asa pentagonal bipyramid with the Ca21 ion occupying thecentre of the pyramid.
The name EF-hand stems from the protein structure of parvalbumin, in which six a helices named A–F form atotal of three EF-hand domains (AB, CD, EF) and the lastdomain (EF) gave the entire family its name. Although
most proteins within this family contain the typical EF-hand motif, a variation is present, for example, in the firstCa21 -binding site of the S100 proteins (two EF-handproteins) where two additional residues occur in the firstCa21 -binding loop.
In addition, the Ca21 -binding loop contains a b-pleatedsheet, which is important for the pairing of two EF-handdomains. This double EF-hand motif appears to be a basicstructural feature common to all EF-hand Ca21 -bindingproteins, and almost all of the family members have aneven number of EF-hand domains (2, 4, 6 or 8). In the caseof parvalbumin, which consists of three EF-hand domains,only two (CD and EF) are capable of metal binding. This
organization into tandem domains seems to be importantfor correct structural folding during protein synthesis andalso strongly affects the functional properties of individualCa21 -binding sites.
Based on differences in selectivity and affinity of EF-hand proteins for Ca21 and Mg21 binding, two types of Ca21 -binding sites have been characterized: mixed Ca21 /Mg21 sites and Ca21 -specific sites. The mixed Ca21 /Mg21 sites bind Ca21 with high affinity and Mg21 withmoderate affinity in a competitive manner (dissociation
constants: K Ca51027 –1029molL21; K Mg51023 –102
molL21). Under physiological conditions, i.e. at loresting levels of Ca21 , they are occupied by Mg21 ; wheintracellular Ca21 concentration is elevated Mg21
replaced by Ca21 , a reaction that is often characterized brelatively slow kinetics. In comparison with mixed Ca21
Mg21 sites, the affinity range spanned for Ca21 -specifisitesis two orders of magnitude lower with respectto Ca2
binding and still further reduced in the case of Mg2
(K Ca51025 –1027mol L21; K Mg51021 –1022molL21
Mg21 ions thushave onlya minor effect on Ca21 binding
In the resting cell, Ca21 -specific sites are considered to bin a metal-free state, and hence in readiness to bind Ca2
ions directly, with fast kinetics, when intracellular levels othis cation are raised. These sites are believed to bresponsible for Ca21 -dependent regulatory functioninvolved in protein activation.
Important functions of EF-hand Ca21 -binding proteininclude the modulation of Ca21 signals, direct Ca21
dependent enzymatic activities, and the buffering ocytosolic Ca21 concentration. Members of the first grou
Figure 3 EF-hand motif. (a) The three-dimensional arrangement of theEF-hand motif can be simulated by the right hand, with the index finger
representing the E-helix (residues 1 – 10), the bent middle finger symbolizing the Ca21 -bindingloop (10 – 21), andthethumbdepictingth
F-helix 19 – 29). Thegeometricalarrangement of thesevenoxygen ligancoordinating the Ca21 ion can best be described as a pentagonalbipyramid. Modified from Celio et al . (1996). (b) Crystal structurefrom E
domain of parvalbumin. Modified from Kretsinger and Nockolds (1973)(c) Coordination of the Ca21 ion in calmodulin with the seven oxygenligands (five from side-chains, one from a carbonyl group of the backbon
andone from a watermolecule). (d)Consensussequence forthe canonicEF-hand domain. The symbol n denotes nonpolar side-chains and the
positionsX, Y,Z, 2 Y, 2Xand 2Z provide theoxygenligand forthe Ca2
binding. At position 2 Y a carbonyl oxygen bonds to the Ca21 ion. The2X ligand (usually glutamate) binds Ca21 with both oxygen atoms fro
the carboxylate group (Kawasaki et al ., 1998). * any amino acid; I,isoleucine.
Calcium-binding Proteins
4 ENCYCLOPEDIA OF LIFE SCIENCES / & 2001 Nature Publishing Group / www.els.net

7/27/2019 Ca binding proteins A0001347-001-000.pdf
http://slidepdf.com/reader/full/ca-binding-proteins-a0001347-001-000pdf 5/8
interact with target proteins in a Ca21 -dependent manner,thereby modulating protein activation. For example, thebinding of Ca21 to calmodulin induces marked conforma-tional changes, which enable it to interact with, andregulate the activity of, a large number of different targetproteins. In contrast, troponin C, which has a very similaroverall structure to calmodulin is involved solely in the
Ca21
-dependent regulation of skeletal and heart musclecontraction.
Proteins within the second group manifest direct Ca21 -dependent enzymatic activity. Currently, eight subfamilieshave been described, including aequorin, Ca21 -dependentprotein kinase, calcineurin, diacylglycerol-kinase, proteinphosphatase, calpain, glycerol phosphate dehydrogenaseand calmodulin domain protein kinase.
Members of the third group do not interact with otherproteins but act as intracellular Ca21 buffers, therebymodulating the temporal and spatial aspects of Ca21
transients. Proteins expressed in specific subpopulation of neurons include calretinin (CR), calbindin D-28k (CB) and
parvalbumin. While the former two have five and fourCa21 -specific sites, respectively with presumably fastCa21 -binding kinetics, the latter one with two Ca21 /Mg21 mixed sites is a slow-onset Ca21 buffer. This meansthat CR and CB are fast enough to immediately bufferCa21 entering via channels from the outside or releasedfrom internal stores and thus to limit the amplitude of Ca21 transients. PVon the other handis too slowto play arole in the rising phase of [Ca21 ]i, but helps to shortentransients. The hypothesis on the protective role of theseproteins against Ca21 overload has been put forward, butseveral recent findings in transgenic mice lacking one ormore of these proteins do not generally support this
function in vivo. Overexpression of Ca21
-binding proteinsin culture systems can modulate Ca21 transients andprotect cells from apoptosis due to elevated levels of Ca21 .The downregulation of some CaBPs before and during thedevelopment of several neurodegenerative diseases hasbeen linked with a failure of these neurons to withstandexcitotoxic insults. Therefore, the involvement of theseproteins in such a protective mechanism remains to befurther investigated.
Calmodulin
Calmodulin (CaM) is a ubiquitously expressed four EF-
hand Ca21 -binding protein (17 kDa) that appears to bethe primary Ca21 sensor in eukaryotic cells. It acts as aCa21 signal-transmitting subunit for many proteins inmammalian cells. It consists of two pairs of two EF-handslinked by a relatively flexible tether and is dumbbell-shapedin the metal-free form (Figure 4a; Babu et al ., 1988). Whenbound with three or four molecules of Ca21 , CaMbecomes globular and is involved in the regulation of multiple cellular functions including intermediary meta-bolism, secretion, motility, signal transduction, cell growth
and division. CaM is essential for the survival of all cellsbut expression levels can vary considerably. In vertebratea multigene family of three to four different, maximalldivergent genes encode an identical 148-amino-aciprotein. CaM is the best conserved protein detected sfar and no naturally occurring mutations have as yet beeidentified in multicellular organisms.
Upon Ca21 binding CaM undergoes a conformationachange that exposes hydrophobic pockets on the undersidof each pair of EF-hands. In the presence of target peptid
a further conformational change results in the flexiblinker between the two heads being dramatically benpartially unwound and wrapped around the target peptid(Figure 4b; Ikura et al ., 1992). Calmodulin has been showto interact with well over 100 different target proteins anaffinities for bona fide CaM targets are usually in thnanomolar range. CaM is found in the cytosol, associatewith membranes and also in the nucleus. The regulateredistribution of CaM between the cytosol and the nucleuappears to play an important role in the proper regulatio
Figure4 Structureof calmodulin (CaM).(a)In theCa21 -freeformCaMdumbbell-shaped and the two pairs of EF-hand domains are linked by a
flexible tether (Babu et al , 1988). (b) After Ca21 binding, CaM becomemore globular and can wrap around target proteins (e.g. myosin light-chain kinase; Ikura et al ., 1992). (Brookhaven Data Base, PDB entry code1CLL and 3CLN.)
Calcium-binding Proteins
ENCYCLOPEDIA OF LIFE SCIENCES / & 2001 Nature Publishing Group / www.els.net

7/27/2019 Ca binding proteins A0001347-001-000.pdf
http://slidepdf.com/reader/full/ca-binding-proteins-a0001347-001-000pdf 6/8
of CaM function. Of the most important targets for CaMare the calmodulin-dependent protein kinases II and IV(CaMKII and IV). CaMKII phosphorylates target pro-teins implicated in many important cellular processes,including neurotransmitter synthesis and release, modula-tion of ion channels and energy metabolism.
Troponin C
Skeletal muscle troponin C (TnC) is the Ca21 -sensingcomponent of the troponin complex and contains a pair of high-affinity, Ca21 /Mg21 mixed sites in the C-terminalhalf and a pair of low-affinity Ca21 -specific sites in the N-terminal half. The structure of TnC is very similar to that of calmodulin (Figure 4a). The high-affinity metal-bindingsites are probably saturated with Ca21 or Mg21 duringmuscle relaxation and contraction, respectively and allowconstitutive association of TnCwith the troponin complex.Ca21 -binding to the low-affinity site(s) with fast kineticsinduces a conformational change of the N-terminal half
which is transmitted to the other components and is thetriggering event of muscular contraction.
Parvalbumin
Parvalbumin (PV) possesses two high-affinity Ca21 /Mg21 mixed sites and is highly expressed in fast-twitchmuscles. Under resting [Ca21 ]i, the metal-binding sites areoccupied by Mg21 ions. The kinetics of Ca21 binding isdetermined by the relatively slow off-rate for Mg21 .During muscle contraction PV does not compete with TnCfor the binding of Ca21 , but helps to increase the initialrate of [Ca21 ]i decay. Thus, it shortens the relaxationphase after very brief contractions. In this example, it is
evident that Mg21
is nearly as important as Ca21
; it notonly lowers the Ca21 affinity to the physiological range,but significantly affects the kinetics of Ca21 binding andrelease. Structural data is available fromBrookhaven DataBase (PDB: entry codes: 1BU3, 1PVA and 3PAT).
Pentraxins
Like annexins, the pentraxins belong to the family of peripheral membrane-binding proteins (Schwalbe et al .,1992). Members are divided into two subfamilies: serumamyloid P (SAP) and C-reactive proteins (CRP). As forannexins or some C2-domain proteins, human SAP has a
high affinity for negatively charged phospholipids. Theprotein consists of a dimer of pentamers and eachpentamer can bind 10 Ca21 ions (Brookhaven Data Base,PDB entry code: 1SAC). The mechanism of binding tomembranes is similar to that of annexins, whereby theCa21 ions form a complex with the protein and the acidicphospholipids. The exact biological functions of pentrax-ins are not known, but recently an interaction of SAP withcalumenin, a secreted Ca21 -binding protein, has beendemonstrated.
Vitamin K-dependent proteins
The most characteristic feature of proteins in this family ithe presence of 9–13 gamma-carboxyglutamate (g-GlaFigure 5a) in the N-terminal Gla domain (residues 1–45This carboxylation reaction is carried out by a vitamin-Kdependent enzyme system. g-Gla is a much stronge
chelator for Ca
21
ions than glutamate. Gla domains arpresent in serum proteins involved in blood coagulatioand include prothrombin and factors VIIa, IX and X. Afor the annexins, it was postulated that the negativelcharged phospholipids exposed on the surface of blooplatelets after injury anchor prothrombin to the plasmmembrane by the bridging function of Ca21 ions. Thattachment would allow the interaction with factor Xa (serine protease) and factor V (a stimulatory protein) whicthen leads to the cleavage of prothrombin and the newlformed thrombin is released from the membrane.
Although the order of reactions taking place during thprocess is clear, the mode of attachment of prothrombi
via electrostatic interactions is less certain. The structure othe prothrombin fragment 1 revealed that most Ca21 ionand Gla residues were buried inside the Gla domai(Soriano-Garcia et al ., 1992). A similar structure was alsfound in the Gla domains of factors VIIa and IX. Morrecent models suggest ‘specific-site’ binding of these factorto membranes. The interaction may involve specifibinding to one or more phospholipid head groups, bualso a cluster of hydrophobic residues in the Gla domainseems to be important for membrane binding (Figure 5). Athe moment it is not known whether these residues ardirectly involved in the actual membrane contact.
Figure 5 Vitamin K-dependent proteins. (a) Structure of the modifiedamino acidg-glutamate (Gla). (b) Picture of prothrombin fragment 1. Thpeptide is in wire-frame form except for Ala1, Phe5, Leu6 and Val9.
Modified from Nelsestuen and Ostrowski (1999). (Brookhaven Data BasePDB entry code: 2PF2.)
Calcium-binding Proteins
6 ENCYCLOPEDIA OF LIFE SCIENCES / & 2001 Nature Publishing Group / www.els.net

7/27/2019 Ca binding proteins A0001347-001-000.pdf
http://slidepdf.com/reader/full/ca-binding-proteins-a0001347-001-000pdf 7/8
Low-affinity, high-capacity CaBPs
The Ca21 concentration in the lumen of organelles(endoplasmic reticulum, mitochondria) is in the millimolarrange, like the extracellular milieu, and thus the affinities of CaBPs in these organelles are adapted to these conditions.Proteins belonging to this group are calnexin, calmegin,
calreticulin and calsequestrin. They are all associated withendo(sarco)plasmic reticulum membranes and are thoughtto play a role in a variety of cellular functions (e.g. Ca21
homeostasis, protein folding, adhesion, gene expression,etc.). The proteins can be divided into distinct structuraland functional domains which sometimes show stronghomologies. A proline-rich domain (P domain) is presentin calreticulin, calnexin and calmegin. The C domainsimplicated in Ca21 -binding are veryacidic andbind Ca21 ,Mg21 , Zn21 with low affinity and high capacity (forCa21 : K d50.5–2 mmol L2 1; Bmax4 25–50 moles of Ca21 per mol). In some cases they can also bind Fe31 orK1 . Calnexin is one of the major Ca21 -binding proteins
of the endoplasmic reticulum membrane and bindstransiently to newly synthesized membrane and solubleproteins that are destined for the secretory pathway. Thismembrane-bound chaperone is thought to be directlyinvolved in the quality-control mechanism of the endo-plasmic reticulum (ER). Stable, if not permanent, complexformation has been reported with unassembled multi-subunit and misfolded proteins. Calreticulin binds toblood clotting factors (factor IX, factor X and prothrom-bin) and has antithrombotic activity. A chaperone func-tion for the protein has also been suggested. Calsequestrin(CS) is the major Ca21 -binding protein in the sarcoplas-mic reticulum (SR) of skeletal and cardiac muscle. It is
localized within the terminal cisternae of the SR, close tothe luminal site of the junctional membrane and inpotential contact with the Ca21 -release channels. Itsproposed functions include Ca21 storage and an involve-ment in Ca21 release. About 30% of the total amino acidsof CS are acidic residues, mainly concentrated within theC-terminus.
Transport of Calcium
Human serum albumin is by far the most abundant Ca21 -
binding protein in serum, but a2-macroglobulin, immu-noglobulin M (IgM) and haptoglobins bind approximatelythe same amount of calcium per gram as albumin. Besidesthese proteins, small molecules (e.g. bicarbonate, phos-phate, citrate) and the pH determine the free serumconcentration of Ca21 . Ca21 ions in the serum have beenshown to act as a first messenger by binding to a calcium-sensing receptor (CaSR) which was first detected on thesurface of parathyroid cells, but is also present in epithelialand kidney cells. Another protein, a putative calcium-
sensing protein (CAS, also named gp330 or megalin) habeen implicated in ‘sensing’ the extracellular concentratioof Ca21 .
1,25-Dihydroxyvitamin D3 induces absorption of Ca2
from the intestinal lumen and also Ca21 resorption in thkidney. It upregulates the expression of two EF-hanCaBPs, calbindin D-28k and calbindin D-9k by
mechanism similar to that of other steroid hormoneCalbindin D-9k is present in absorptive epithelial cells omammalian duodenum and its tissue concentration correlated with active transcellular, vitamin D-dependenCa21 absorption. Furthermore, it is assumed to play a roin Ca21 reabsorption in the kidney distal tubules. In thtissue also calbindin D-28k is abundantly expressed and thought to participate in Ca21 resorption. Calbindin D-9does not exist in amphibians and birds and insteadcalbindin D-28k is expressed in the enterocytes in thesspecies. In both cases, the proteins act as Ca21 transporters, shuttling these potentially cytotoxic ions from thluminal to the apical side (blood vessels).
References
Babu YS,Bugg CE andCook WJ (1988)Structure of calmodulin refin
at 2.2 A resolution. Journal of Molecular Biology 204: 191–204.
Brown EM, Gamba G, Riccardi D et al . (1993) Cloning an
characterization of an extracellular Ca21 -sensing receptor fro
bovine parathyroid. Nature 366: 575–580.
Burgoyne RD and Geisow MJ (1989) The annexin family of calcium
binding proteins. Cell Calcium 10: 1–10.
Celio M, Pauls T and Schwaller B (1996) Guidebook to the Calcium
Binding Proteins, pp. 1–238. Oxford: Oxford University Press.
Huber R, Romisch J and Paques EP (1990) The crystal and molecul
structure of human annexin V, an anticoagulant protein that binds
calcium and membranes. EMBO Journal 9: 3867–3874.
Ikura M, Clore GM, Gronenborn AM, Zhu G, Klee CB and Bax
(1992) Solution structure of a calmodulin-target peptide complex
multidimensional NMR. Science 256: 632–638.
Kawasaki H, Nakayama S and Kretsinger RH (1998) Classification an
evolution of EF-hand proteins.Biometals 11: 277–295.
Kretsinger RH and Nockolds CE (1973) Carp muscle calcium-bindin
protein. II. Structure determination and general description.Journ
of Biological Chemistry 248: 3313–3326.
Nalefski EA and Falke JJ (1996) TheC2 domaincalcium-binding mot
structural and functional diversity. Protein Science 5: 2375–2390.
Nelsestuen GL and Ostrowski BG (1999) Membrane association wi
multiple calcium ions: vitamin-K-dependent proteins, annexins an
pentraxins. Current Opinions in Structural Biology 9: 433–437.
Rizo J and Sudhof TC (1998) C2-domains, structure and function of
universal Ca21 -binding domain. Journal of Biological Chemistry 27
15879–15882.
Schwalbe RA,DahlbackB, CoeJE andNelsestuen GL (1992)Pentrax
family of proteins interact specifically with phosphorylcholine and/
phosphorylethanolamine.Biochemistry 31: 4907–4915.
Soriano-Garcia M,PadmanabhanK, de VosAM andTulinskyA (199
The Ca21 ion and membrane binding structure of the Gla domain
Ca-prothrombin fragment 1. Biochemistry 31: 2554–2566.
Sutton RB, Davletov BA, Berghuis AM, Sudhof TC and Sprang S
(1995) Structure of the first C2 domain of synaptotagmin I: a nov
Ca21 phospholipid-binding fold.Cell 80: 929–938.
Calcium-binding Proteins
ENCYCLOPEDIA OF LIFE SCIENCES / & 2001 Nature Publishing Group / www.els.net

7/27/2019 Ca binding proteins A0001347-001-000.pdf
http://slidepdf.com/reader/full/ca-binding-proteins-a0001347-001-000pdf 8/8
Swairjo MA, Concha NO, Kaetzel MA, Dedman JR and Seaton BA
(1995) Ca21 -bridging mechanism and phospholipid head group
recognition in the membrane-binding protein annexin V. Nature
Structural Biology 2: 968–974.
Further Reading
Brown EM (1999) Physiology and pathophysiology of the extracellularcalcium-sensing receptor.American Journal of Medicine106: 238–253.
Gewurz H, Zhang XH and Lint TF (1995) Structure and function of the
pentraxins. Current Opinions in Immunology 7: 54–64.
McDonald JF, Evans TC, Jr, Emeagwali DB et al . (1997) Ion
propertiesof membrane association by vitamin K-dependentprotein
the case for univalency. Biochemistry 36: 15589–15598.
Persson E and Petersen LC (1995) Structurally and functionally distin
Ca21 binding sites in the gamma-carboxyglutamic acid-containin
domain of factor VIIa. European Journal of Biochemistry 234: 293
300.
Stenflo J (1999) Contributions of Gla and EGF-like domains to t
function of vitamin K-dependent coagulation factors.Critical Revieon Eukaryotic Gene Expression 9: 59–88.
Calcium-binding Proteins
8 ENCYCLOPEDIA OF LIFE SCIENCES / & 2001 Nature Publishing Group / www.els.net