calcareous nannofossil bioevents and … in the mediterranean sea (castradori, 1993; sprovieri et...
TRANSCRIPT

www.elsevier.com/locate/marmicro
Marine Micropaleontology
Calcareous nannofossil bioevents and environmental control on
temporal and spatial patterns at the early–middle Pleistocene
Patrizia Maiorano*, Maria Marino
Dipartimento di Geologia e Geofisica-Universita di Bari, Via E. Orabona, 4-70125 Bari, Italy
Received 13 April 2004; received in revised form 9 August 2004; accepted 10 August 2004
Abstract
Quantitative distributions of calcareous nannofossils are analysed in the early–middle Pleistocene at the small Gephyrocapsa
and Pseudoemiliania lacunosa zone transition in deep-sea cores from the Mediterranean Sea and North Atlantic Ocean (Ocean
Drilling Program [ODP] Sites 977, 964 and 967, Deep Sea Drilling Project [DSDP] Site 607). The temporal and spatial mode of
occurrence of medium-sized gephyrocapsids and reticulofenestrids has been examined to refine biostratigraphic constraints and
evaluate possible relationships of stratigraphic patterns to environmental changes during a period of global climatic
deterioration. The timing of bioevents has been calibrated using high-resolution sampling and correlation to the y18O record in
chronologically well-constrained sections. Newly identified events and ecostratigraphical signals enhance the stratigraphic
resolution at the early–middle Pleistocene. The first occurrence (FO) of intermediate morphotypes between Pseudoemiliania
and Reticulofenestra (Reticulofenestra sp.) is proposed as a reliable event within marine isotope stage (MIS) 35 or at the MIS
35/34 transition. The distribution of Reticulofenestra asanoi is characterized by rare and scattered occurrences in its lowest
range, but the first common occurrence (FCO) is consistently identified at MIS 32 or 32/31; the last common occurrence (LCO)
of the species is a distinctive event at MIS 23. In the studied interval, Gephyrocapsa omega dominates among medium-sized
Gephyrocapsa. The FO of G. omega and contemporaneous re-entry of medium-sized gephyrocapsids at the lower–middle
Pleistocene transition are diachronous between the Atlantic Ocean and Mediterranean Sea and from the western to eastern
Mediterranean. In the Mediterranean, the LO of G. omega falls at MIS 15, insolation cycle 54 and is isochronous among the
sites. Abundance fluctuations of G. omega show notable relations to early–middle Pleistocene climate changes; they
considerably increase in abundance at the interglacial stages, suggesting warm water preferences. Gephyrocapsa omega
temporarily disappears during the glacial MIS 22 and MIS 20. Above MIS 20, an impoverishment in G. omega and in the total
abundance of medium-sized gephyrocapsids occurs. A decrease in abundance of G. omega is observed between the western Site
977 and the easternmost Site 967 in the Mediterranean Sea, as a possible response to high salinity and/or low nutrient content.
Possible environmental influences on the distribution of R. asanoi and of Reticulofenestra sp. are discussed.
D 2004 Elsevier B.V. All rights reserved.
Keywords: Pleistocene; calcareous nannofossils; biochronology; temporal distribution; Mediterranean Sea; Atlantic Ocean
0377-8398/$ - s
doi:10.1016/j.m
* Correspon
E-mail addr
53 (2004) 405–422
ee front matter D 2004 Elsevier B.V. All rights reserved.
armicro.2004.08.003
ding author. Tel.: +39 80 544 3455; fax: +39 80 544 2625.
ess: [email protected] (P. Maiorano).

P. Maiorano, M. Marino / Marine Micropaleontology 53 (2004) 405–422406
1. Introduction
The Pleistocene was characterized by significant
orbitally forced climate fluctuations, which are
well recorded in the sedimentary record through
changes in sediment properties, fossil commun-
ities, chemical and isotopic characteristics. In
more detail, during the early–middle Pleistocene
transition, the Earth climatic system experienced a
major change related to intensification of the
Northern Hemisphere glaciation, with a significant
shift toward more intense glacial conditions,
occurred between 0.9 and 0.6 Ma, together with
the development of 100 ky glacial–interglacial
oscillations (Ruddiman et al., 1989). Quantitative
biostratigraphic techniques, high-resolution sam-
pling and calibration of bioevents through the
astronomical tuning of the cyclic patterns in the
sedimentary record, have led to a high-quality
biochronological framework of the Pleistocene
record (Shackleton et al., 1990; Hilgen, 1991;
Hilgen et al., 1993; Lourens et al., 1996).
However, the early–middle Pleistocene transition
falls within the Pseudoemiliania lacunosa zone,
which has the lowest biostratigraphic resolution of
the entire Pleistocene. Previous studies suggested
that calcareous nannofossil biostratigraphy could
be improved by collecting further data on poorly
known taxa and on ecostratigraphical signals. The
Fig. 1. Location map of
availability of precise Pleistocene chronology
provides a powerful reference frame for this.
Moreover, the increasing documentation of the
ecological preferences of modern coccolithophorids
can be a key tool for interpreting distinctive
distribution patterns of calcareous nannofossils,
during the past, in terms of response to palae-
oenvironmental changes.
The aim of this study is to understand charac-
ters in the stratigraphic and spatial patterns of
selected calcareous nannofossils through the small
Gephyrocapsa and Pseudoemiliania lacunosa zones
in high-quality Mediterranean and extra-Mediterra-
nean deep-sea cores, within a well-constrained
temporal framework. In particular, the study focu-
ses on the distribution of medium-sized gephy-
rocapsids and selected reticulofenestrids, on their
correlation to oxygen isotopes fluctuations and on
geographical differences in the abundance of taxa,
in order to discuss the potential stratigraphical
value of new or contradictory bioevents, their
patterns of diachrony and their possible relation
to environmental parameters.
2. Material and methods
Three Mediterranean deep-sea sections located
along an east–west transect (ODP Sites 977, 964,
the studied sites.

Table 1
Summary of the sites investigated in this study and related references
Studied sites Reference
Leg/Site Location, latitude,
longitude
Previous calcareous
nannofossil
studies
Oxygen isotope
stratigraphy
Sapropel stratigraphy,
insolation cycle
Age model
DSDP 94/
Site 607
Eastern Atlantic,
418 N, 338 WTakayama and Sato (1987),
Raffi et al. (1993),
Wei (1993), Raffi (2002)
Ruddiman et al.
(1986)
/ Mix et al.
(1995)
ODP 161/
Site 977
Western Mediterranean,
368 N, 18 Wde Kaenel et al. (1999) von Grafenstein
et al. (1999)
Murat (1999), de
Kaenel et al. (1999)
von Grafenstein
et al. (1999)
ODP 160/
Site 964
Eastern Mediterranean,
368 N, 17 E
Sprovieri et al. (1998),
Maiorano et al. (2004)
Howell et al.
(1998),
Sprovieri et al.
(1998)
Sakamoto et al.
(1998), Emeis
et al. (2000)
Howell et al.
(1998)
ODP 160/
Site 967
Eastern Mediterranean,
348 N, 328 ERaffi (2002) Kroon et al.
(1998)
Kroon et al. (1998) Kroon et al.
(1998)
P. Maiorano, M. Marino / Marine Micropaleontology 53 (2004) 405–422 407
967) and a reference mid-latitude extra-Mediterranean
section (DSDP Site 607) located in the North Atlantic
Ocean (Fig. 1) were selected for this study. Age
control, oxygen isotope stratigraphy and sapropel
stratigraphy when possible, are available in the
sections (Table 1). A total of 664 samples were
analysed in the interval across marine oxygen isotope
stages 35 to 15 in all the sections. Sample spacing
varied from mean values of 10 and 20 cm at Sites 607,
964, 967 to 40 cm at Site 977 to obtain approximately
one sample per 3–5 ky at all sites, according to the
different sedimentation rates, ranging from mean
values of about 150 m/Ma at Site 977 to 25–40 m/
Ma at the other sites. Frequently, samples used for
oxygen isotopes have been investigated. Simple smear
slides were prepared from unprocessed samples
according to standard technique (Bown and Young,
1998) and analyzed under a polarized light micro-
scope at 1000� magnification. Quantitative data were
collected by counting the number of specimens per
unit area (Backman and Shackleton, 1983). About 150
fields of view having approximately the same density
of nannofossils (nearly 50–60 specimens N4 Am) were
investigated, and abundances were plotted as number
of specimens/mm2.
3. Taxonomic remarks
The taxonomic criteria for unambiguous recog-
nition of Reticulofenestra asanoi and intermediate
morphotypes between Reticulofenestra and Pseudoe-
miliania are discussed in recent papers (Marino,
1996; Maiorano et al., 2004). Circular to subcircular
reticulofenestrids larger than 6 Am are referred to R.
asanoi (Plate 1, Figs. 1–4); the number of slits (up to
four) on the distal shield as taxonomic definition of
the species (de Kaenel et al., 1999) is not followed
because we found that it cannot be consistently
applied in sediments with varying preservation state.
Intermediate morphotypes between Reticulofenestra
and Pseudoemiliania are distinguished from R.
asanoi and are indicated here as Reticulofenestra
sp. (Plate 1, Figs. 5–8); they are larger than 5 Am,
exhibit a subcircular outline and a prominent collar.
The size of the central opening is wider than in R.
asanoi and smaller than in Pseudoemiliania, repre-
senting about 30% of the total length of the
reticulofenestrid; only a few slits are visible on the
distal shield. The absence of slits in R. asanoi is an
important taxonomic criterion if compared to Retic-
ulofenestra sp.
As far as Gephyrocapsa is concerned, it is
worthwhile mentioning that taxonomic concepts vary
between different authors. Syntheses on the equiv-
alence among gephyrocapsid taxonomic units are
given by Flores et al. (1999) and Raffi (2002). In
particular, gephyrocapsids are generally subdivided
by using biometric criteria, mainly among biostratig-
raphers and/or grouped in morphological associations
(probable genotypes) having different environmental
preferences (Bollmann, 1997), as frequently adopted

Plate 1. Light microscope photographs of selected calcareous nannofossil species. XP—crossed polarized light, PL—parallel light. Scale bar
represents 5 Am.
1–2. Reticulofenestra asanoi Sato and Takayama, 1 XP, 2 PL, ODP Hole 967A, 4–4, 90 cm.
3–4. Reticulofenestra asanoi Sato and Takayama, 3XP, 4 PL, DSDP 607, 5–6, 88 cm.
5–6. Reticulofenestra sp., 5 XP, 6 PL, ODP Hole 967A, 4–3, 110 cm.
6–7. Reticulofenestra sp., 6 XP, 7 PL, ODP Hole 967A, 4–4, 90 cm.
8–9. Pseudoemiliania lacunosa (Kamptner) Gartner, 8 XP, 9 PL, ODP Hole 967A, 4–2, 90 cm.
10. Gephyrocapsa omega Bukry, XP, ODP Hole 967A, 3–3, 50 cm.
11. Gephyrocapsa oceanica s.l. sensu Rio, 1982, XP, ODP Hole 967A, 4–2, 30 cm.
P. Maiorano, M. Marino / Marine Micropaleontology 53 (2004) 405–422408
in late Pleistocene to recent. In this study, the
classification of gephyrocapsids follows the morpho-
metric criteria of Raffi et al. (1993), which is a
powerful subdivision for stratigraphic purposes;
medium-sized Gephyrocapsa includes all early and
middle Pleistocene gephyrocapsids 4–5.5 Am in size.
However, Gephyrocapsa omega (gephyrocapsids N4
Am in size with a high angle bridge) is also
distinguished within medium-sized Gephyrocapsa to
enhance the data set on stratigraphic meaning of
Pleistocene temporary disappearances of the species
(Marino et al., 2003; Maiorano et al., 2004) and of its

Fig. 2. Quantitative patterns of selected calcareous nannofossils and oxygen isotope stratigraphy at Site 607. Position of re-entrance of medium-
sized Gephyrocapsa (reemG) is indicated according to Raffi (2002). Oxygen isotopes and interglacial stage numbers are from Ruddiman et al.
(1989). FO—first occurrence; FCO—first common occurrence; LO—last occurrence; LCO—last common occurrence; td—temporary
disappearance.
P. Maiorano, M. Marino / Marine Micropaleontology 53 (2004) 405–422 409
LO in the Mediterranean Sea (Castradori, 1993;
Sprovieri et al., 1998).
4. Results
The present results mainly focus on quantitative
distributions of Reticulofenestra sp., Reticulofenestra
asanoi , total medium-sized Gephyrocapsa and
Gephyrocapsa omega, which are major components
of the investigated Pleistocene calcareous nannofossil
assemblage. The studied interval corresponds to the
transitional record through the small Gephyrocapsa
and Pseudoemiliania lacunosa zones. The recognized
calcareous nannofossil events, their correlation with
oxygen isotope stages and estimated biochronology are
listed in Table 2. The discussion that follows outlines
the mode of occurrence of the recognized bioevents,
from older to younger, between the mid-latitude
Atlantic site and the Mediterranean sections (Figs. 2–
5), as well as from western to eastern Mediterranean,
and evaluates correlation of bioevents among different
areas (Fig. 6). Comparison of abundance patterns to
oxygen isotope stratigraphy is examined (Figs. 7 and 8)
in an attempt to understand possible relations of the
evolutionary and/or ecostratigraphical events to envi-
ronmental parameters.
4.1. Stratigraphical patterns
4.1.1. FO of Reticulofenestra sp.
Reticulofenestra sp. is very rare in its lowermost
distribution, and this is a consistent pattern among the
Mediterranean sites (Figs. 3–5). The first occurrence
(FO) of the species can be identified and correlated
among the sections (Fig. 6); it always predates the
lowest occurrences of Reticulofenestra asanoi and
falls at marine isotope stage (MIS) 35 at Sites 607,
977, 967 or at MIS 35/34 at Site 964. At Sites 977 and
967, where sapropel stratigraphy is available and

Fig. 3. Quantitative patterns of selected calcareous nannofossils and oxygen isotope stratigraphy at Site 977. Sapropel layers are according to
Murat (1999) and the corresponding insolation cycles are from de Kaenel et al. (1999). Oxygen isotope stratigraphy and stage assignments are
from von Grafenstein et al. (1999). For abbreviations, refer to Fig. 2.
P. Maiorano, M. Marino / Marine Micropaleontology 53 (2004) 405–422410
correlated to the insolation cycles (i-cycle), the event
is located between i-cycles 110 and 112. The
calibrations of the event are well comparable among
the Sites 607, 977 and 967 (Table 2); at Site 964, the
age of the FO of Reticulofenestra sp. is about 29 ky
younger than at the other sites. This difference is
unlikely related to the slightly higher error in the age
estimate at Site 964, which is on the order of 6 ky
(Table 2), and can be the result of comparing different
age models. The uppermost distribution of Reticulo-
fenestra sp. is variable between the studied sections
(Figs. 2–5), and therefore, neither the last common
occurrence (LCO) nor the LO can be identified with
certainty.
Close morphological affinities between Reticulofe-
nestra sp. and Reticulofenestra asanoi as well as
remarkable similarity in abundance patterns of the
taxa suggest that they may be phylogenetically
relationed.
4.1.2. FCO of Reticulofenestra asanoi
The distribution of Reticulofenestra asanoi is
restricted to the lower–middle Pleistocene transition,
as documented by several authors (Matsuoka and
Okada, 1989; Sato et al., 1991; Wei, 1993; Marino,
1996; de Kaenel et al., 1999; Fornaciari, 2000; Flores
et al., 2000; Raffi, 2002; Maiorano et al., 2004).
However, wide disagreement exists in previous
correlations of FO and LO of the species to oxygen
isotope data. In our opinion, these discrepancies can
be related to indistinct concepts between R. asanoi
and Reticulofenestra sp. In all the sections, R. asanoi
is rare and discontinuous at the beginning of the
distribution (Figs. 2–5); the first common occurrence
(FCO) of the species is more distinctive than the
absolute FO, and it is recorded at MIS 31/32 at Sites
607, 977 and 964 (Figs. 2–4) or at MIS 32 at Site 967.
The event falls slightly above i-cycle 102 at Sites 964
and 967 (Fig. 6). It is noteworthy that a younger

Fig. 4. Quantitative patterns of selected calcareous nannofossils at Site 964. Position of sapropel layers is from Sakamoto et al. (1998); i-cycles
are from Emeis et al. (2000). Oxygen isotope data are from Howell et al. (1998); interglacial stage numbers are indicated according to Sprovieri
et al. (1998) with slight revision after Maiorano et al. (2004). For abbreviations refer to Fig. 2.
P. Maiorano, M. Marino / Marine Micropaleontology 53 (2004) 405–422 411
correlation of FCO of R. asanoi at Site 964 to MIS 30
(Maiorano et al., 2004) is revised here as a conse-
quence of the higher sampling resolution adopted in
the present study, which has considerably improved
the recognition of the event. The age of FCO of R.
asanoi is significantly comparable among the sections
(Table 2). In our opinion, the previous older calibra-
tions of the FO of R. asanoi at MIS 33/34 at Site 977
(mbsf 183.53–182.62, de Kaenel et al., 1999) and at
MIS 35 at Site 607 (mcd 47.85–48.66, Wei, 1993) are
probably caused by different taxonomic criteria
adopted by different authors.
4.1.3. LCO of Reticulofenestra asanoi
The significant decrease in abundance (LCO) of
Reticulofenestra asanoi is recorded in all the inves-
tigated sites and appears as a more reliable event with
respect to the absolute LO; above the LCO, the
distribution of R. asanoi is characterized by discon-
tinuous occurrences of the species (Figs. 2–5), and the
abundances are generally lower than 2 specimens/
mm2. The event is recorded at MIS 23 at all the
investigated sites (Fig. 6), in agreement with Maior-
ano et al. (2004), and slightly predates the severe MIS
22 glaciation. Biochronologic data on the LCO of R.
asanoi are comparable among the sections (Table 2)
inasmuch as differences in the age assignments range
between 1 and 19 ky. The age of the LCO of R. asanoi
is correlatable to data on the LO of the species at MIS
23 or 22/23 from Raffi (2002). On the other hand, the
event is over 30 ky older if compared with the age of
the LO (not LCO), from Wei (1993) at Site 607, where
the event has been correlated to MIS 22. Previous
correlations of the LO of R. asanoi to MIS 19 (de
Kaenel et al., 1999) are most probably caused by
different taxonomic concepts.
4.1.4. Re-entry of medium-sized Gephyrocapsa and
FO of Gephyrocapsa omega
The re-entry of medium-sized Gephyrocapsa and
the contemporaneous FO of Gephyrocapsa omega are
well recognizable events in the studied sites (Figs.

Fig. 5. Quantitative patterns of selected calcareous nannofossils and oxygen isotope stratigraphy at Site 967. Sapropel layers and corresponding
insolation cycles, oxygen isotope stratigraphy and glacial stage numbers are from Kroon et al. (1998). For abbreviations refer to Fig. 2.
P. Maiorano, M. Marino / Marine Micropaleontology 53 (2004) 405–422412
2–5) and define the small Gephyrocapsa and Pseu-
doemiliania lacunosa zonal boundary (Rio et al.,
1990). The abundance patterns of total medium
Gephyrocapsa essentially match that of G. omega in
all the sections, confirming that the reappearance of the
medium-sized gephyrocapsids is largely composed of
G. omega and subordinately of Gephyrocapsa oce-
anica s.l. (sensu Rio, 1982); Gephyrocapsa carib-
beanica represents a minor component in the studied
sections. With regard to the available biochronolog-
ical data set, the middle Pleistocene reappearance of
medium-sized Gephyrocapsa is known to be consis-
tently diachronous at different latitudes, between MIS
29 and 25, and is considered as a possible migratory
event from low to mid-high latitudes (Wei, 1993;
Raffi et al., 1993; Flores et al., 1999; Raffi, 2002). In
the Mediterranean, the event is substantially recorded
in the interval between MIS 26 and MIS 25
(Castradori, 1993; de Kaenel et al., 1999; Raffi,
2002; Maiorano et al., 2004). At Site 607, the event
falls at MIS 27 (Raffi, 2002). In the Mediterranean,
our results indicate that at the westernmost Site 977,
the re-entrance of medium-sized Gephyrocapsa and
the FO of G. omega are characterized by very rare
occurrences correlated to MIS 26/27 transition below
i-cycles 92 (Fig. 3), before the abrupt increase
recorded at MIS 25 within i-cycle 88. At the eastern
sites, the FO of G. omega is sudden and occurs at
MIS 25 between i-cycles 90–92 at Site 964 and
within sapropel layer correlated to i-cycle 90 at Site
967 (Fig. 6). Correlation of bioevents to oxygen
isotopes (Fig. 6) and biochronological data (Table 2)
suggests a diachrony from Atlantic to Mediterranean,
as well as from western to eastern Mediterranean, on
the order of 30–60 ky between Atlantic and Medi-
terranean and of about 30 ky within the Mediterra-
nean. It is worth noting that various sampling
resolution in the sections may affect evaluation of
slight diachrony; in this interval, temporal resolution
in the Mediterranean sites is slightly lower at Site 964

Fig. 6. Position of bioevents relative to age models adopted at the investigated sites. Rsp—Reticulofenestra sp.; Ra—Reticulofenestra asanoi; reemG—re-entry of medium
Gephyrocapsa; Gom—Gephyrocapsa omega; td—temporary disappearance. Dashed lines indicate selected i-cycles. Ages of sapropel layers are relative to their midpoints.
P.Maiorano,M.Marin
o/Marin
eMicro
paleo
ntology53(2004)405–422
413

P. Maiorano, M. Marino / Marine Micropaleontology 53 (2004) 405–422414
(4.5 ky) but is comparable between Sites 977 and 967
(about 3 ky), supporting the value of the estimated
diachrony of the event (Table 2). However, it cannot
be excluded that uncertainty in the evaluation of small
age constraints in diachrony may be also affected by
comparing different age models. The abrupt increase
(FCO) of G. omega at MIS 25 appears to be a more
distinctive pattern rather than the FO in the Medi-
terranean; however, it is variously correlated between
the sites with respect to sapropel stratigraphy, result-
ing younger in age in the western Mediterranean than
in the eastern basin, and therefore, the FCO is not
suggested in replacement of the standard and widely
adopted FO.
4.1.5. Temporary disappearance of Gephyrocapsa
omega
An interval of temporary disappearance of
medium-sized Gephyrocapsa is well known in the
small Gephyrocapsa zone (Gartner, 1977) in the
lower Pleistocene. More recently, Marino (1996),
Marino et al. (2003), Maiorano et al. (2004) discussed
the potential biostratigraphic value of an additional
interval of temporary disappearance of Gephyrocapsa
omega in the Pseudoemiliania lacunosa zone.
Although intervals of temporary disappearance can
be considered as unconventional and unreliable
biostratigraphic markers, inasmuch as they may be
strongly environmentally controlled, they are often
well documented in the calcareous nannofossil assem-
blage in different time intervals. They are widely used
as biostratigraphic proxies although their causes are
not always explained.
Our results allow the data set on the occurrence of
temporary disappearances of Gephyrocapsa omega in
different geographic areas to be extended. The taxon
clearly shows a consistent pattern among the inves-
tigated sites; two intervals of remarkable rarity or
absence of the species can be observed independently
among sections indicative of diverse water masses
(Figs. 2–5), suggesting that they can be considered as
a primary variation in the nannofossil assemblage. In
fact, both the older interval of temporary disappear-
ance (td 1) and the younger one (td 2) appear as
essentially correlated to MIS 22 and 20, respectively
(Fig. 6), that represent two severe oxygen isotope
glacial signals of the middle Pleistocene. In more
details, td 1 is a short-term ecostratigraphic signal
(about 15 ky) and occurs from MIS 23/22 or MIS 22
to MIS 22/21 transition or MIS 21 in the Mediterra-
nean sites, with the ages of bottom and top of
temporary disappearance in good agreement among
all the investigated sections (Table 2) and with an
error in the age estimates not exceeding 2 ky. The
beginning of td 2 is well comparable among the sites
(Fig. 6, Table 2) occurring at MIS 21/20 or MIS 20 at
Site 964. This result is consistent with data from a
southern Apennine foredeep section (Maiorano et al.,
2004). The end of td 2 has a slightly variable
extension in the sites (Fig. 6, Table 2), ranging from
MIS 19/20 to MIS 19/18; however, difference in the
age assignments is not higher than 29 ky and is
unlikely related to sampling resolution inasmuch as
the estimated error is not higher than 7 ky (Table 2).
4.1.6. LO of Gephyrocapsa omega
Gephyrocapsa omega is still present in the open
oceans today; the LO of the species is known only in
the Mediterranean Sea, where it was first identified by
Castradori (1993) at 0.58 Ma and correlated to MIS 15
at Site 964 (Sprovieri et al., 1998). In the present
study, the LO of G. omega is also recognized at Sites
977 and 967 for the first time and correlated to MIS
15 in both sections, in agreement with previous
results. Biochronological data obtained in this study
indicate a high synchroneity of the event with an
estimated age ranging from 570 to 577 ky (Table 2)
and correlated to i-cycle 54 at Sites 977 and 964 (Fig.
6). The slight differences with previous age assign-
ments can be ascribed to different references used for
age evaluation; however, the best biochronologic
datum with the lower age uncertainty (F2 ky) among
the investigated records is that obtained at Site 977.
4.2. Environmental control on stratigraphic and
geographic patterns
High-resolution sampling, correlation to oxygen
isotope stratigraphy and comparison of the quantita-
tive distribution of gephyrocapsids and reticulofenes-
trids among different sites suggest a possible
environmental control on abundance patterns.
As shown in Fig. 7, Gephyrocapsa omega, which
represents almost the total medium-sized Gephyro-
capsa, shows significant variations in abundance in
the open ocean Atlantic site and in the Mediterranean

Table 2
List of calcareous nannofossil events: depth, correlation to marine oxygen isotope (MIS) and age evaluation
Calcareous
nannofossil event
Site 607 Site 977 Site 964 Site 967
Depth
(mcd)
MIS Age (Ma) Depth
(mbsf)
MIS Age (Ma) Depth
(rmcd)
MIS Age
(Ma)
Depth
(rmcd)
MIS Age (Ma)
LO G. omega 90.1 15 0.577F0.002 24.07 15 0.575F0.008 22.26 15 0.57F0.004
End td 2 31.64 19 0.778F0.004 127.77 20/19 0.79F0.001 29.88 19 0.786F0.0002 27.46 19/18 0.761F0.007
Beginning td 2 33.31 21/20 0.816F0.002 131.37 21/20 0.809F0.001 30.63 20 0.799F0.001 29.04 21/20 0.817F0.0004
End td 1 34.51 22/21 0.862F0.001 140.9 21 0.86F0.001 31.99 22/21 0.863F0.002 30.24 21 0.864F0.0003
Beginning td 1 35.09 23/22 0.88F0.002 142.6 22 0.874F0.002 32.36 22 0.88F0.001 31.2 22 0.887F0.001
LCO R. asanoi 36.23 23 0.912F0.003 147.65 23 0.913F0.002 32.93 23 0.906F0.004 31.43 23 0.894F0.004
FO G. omega 41.32* 27* 1.005F0.005* 160.67 27/26 0.985F0.001 34.31 25 0.965F0.004 33.19 25 0.956F0.002
FCO R. asanoi 44.78 32/31 1.087F0.003 178.3 32/31 1.079F0.002 37.36 32/31 1.089F0.0003 36.39 32 1.089F0.002
FO Reticulofenestra sp. 47.41 35 1.17F0.003 192.05 35 1.178F0.003 38.56 35/34 1.148F0.006 38.75 35 1.168F0.003
mcd =meters composite depth; mbsf =meters below sea floor; rmcd = revised meters composite depth.
* Data from Raffi (2002) and refer to re-entry of medium Gephyrocapsa.
P.Maiorano,M.Marin
o/Marin
eMicro
paleo
ntology53(2004)405–422
415

Fig. 7. Comparison of abundance patterns of medium-sized gephyrocapsids and y18O stratigraphy among the investigated sites. Shaded areas indicate intervals of temporary
disappearance of medium-sized Gephyrocapsa.
P.Maiorano,M.Marin
o/Marin
eMicro
paleo
ntology53(2004)405–422
416
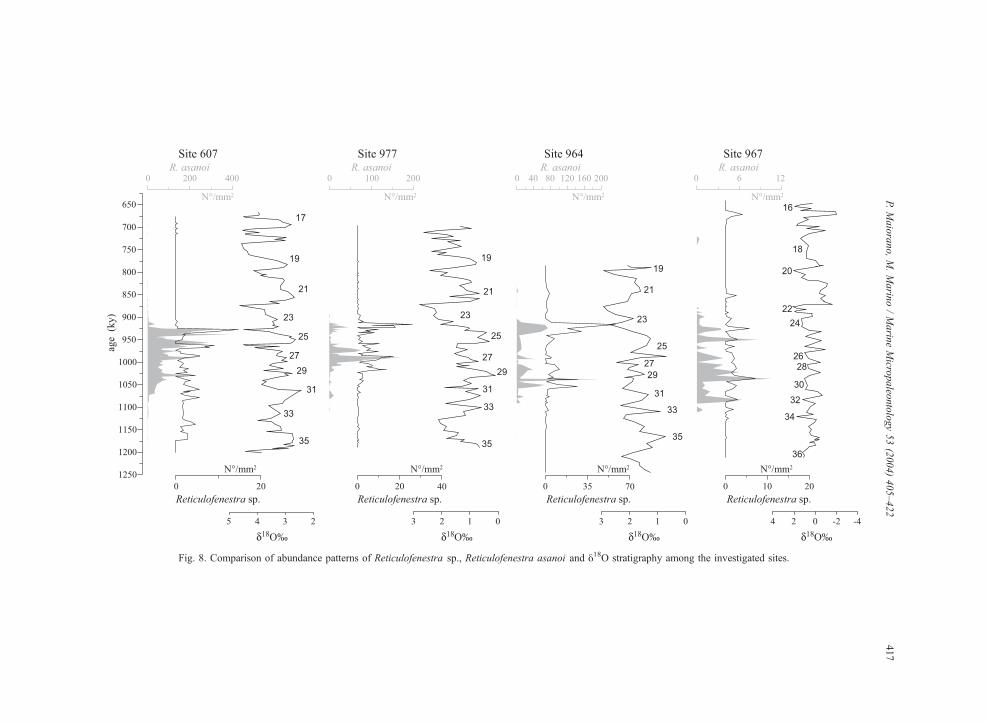
Fig. 8. Comparison of abundance patterns of Reticulofenestra sp., Reticulofenestra asanoi and y18O stratigraphy among the investigated sites.
P.Maiorano,M.Marin
o/Marin
eMicro
paleo
ntology53(2004)405–422
417

P. Maiorano, M. Marino / Marine Micropaleontology 53 (2004) 405–422418
sections. In particular, a positive correlation is noted
between increase in abundance of gephyrocapsids and
the interglacial stages (Fig. 7), suggesting warm water
preferences of the taxon. In this context, the two
intervals of temporary disappearance of G. omega at
MIS 22 and MIS 20 appear to be a response to glacial
conditions (Fig. 7). In fact, the glacial MIS 22 and 20
represent intense cold stages during the middle
Pleistocene climate deterioration, when the perennial
ice condition replaced the seasonal ice condition in the
Arctic Ocean (Herman and Hopkins, 1980; Worsley
and Herman, 1980) from 0.9 to 0.7 Ma (Pisias and
Moore, 1981; Prell, 1982; Ruddiman et al., 1986). It is
worth noting that gephyrocapsids are considered
dissolution-resistant coccoliths (Roth and Berger,
1975; Roth and Coulbourn, 1982; Roth, 1994;
Bollmann et al., 1998) so it is unlikely that the
temporary disappearances of G. omega are related to
dissolution. A similar argument was made by McIn-
tyre and McIntyre (1971) and Gartner (1988) for the
small Gephyrocapsa zone. An environmental influ-
ence on the distribution of gephyrocapsids is also
observed in changes in the abundances of gephyr-
ocapsids within the Mediterranean; an impoverish-
ment of G. omega and of all medium-sized
Gephyrocapsa occurs above MIS 20 (Fig. 7). Mean
abundances of G. omega change from about 500
specimens/mm2 (Sites 977 and 964) or 120 (Site 967)
during interglacial stages 25–21 to about 70 speci-
mens/mm2 (Sites 977 and 964) or 8 (Site 967) during
interglacial 19–15, suggesting that its distribution was
affected by the climate cooling of the glacial
Pleistocene. Further considerations arise from com-
parative analysis of the abundance of G. omega
between the cores. A noticeable decrease in abun-
dance is observed between the western and the eastern
Mediterranean (Fig. 7); mean values in the total
abundance of G. omega range from about 280
specimens/mm2 at Site 977 and Site 964 to about 65
at Site 967 during interglacial stages (MIS 25–15).
This pattern is probably a response to increase in
salinity and/or decrease in nutrient content in the
eastern basin (Rohling et al., 2000; Bethoux et al.,
1999).
Our results suggest that during the early–middle
Pleistocene, Gephyrocapsa omega was a warm water
taxon having higher abundances in low salinity and
high nutrient waters. The temperature dependence of
G. omega is not known in the fossil record; however,
our data seem to be in agreement with studies from the
recent, suggesting warm water preferences for gephyr-
ocapsids having high angle bridge (McIntyre et al.,
1970; Bollmann, 1997; Takahashi and Okada, 2000).
Moreover, comparable environmental preferences are
widely known for Gephyrocapsa oceanica Kamptner
(N3 Am). In fact, in many studies, G. oceanica is
proved to be a warm water taxon both in the
Pleistocene record (Thierstein et al., 1977; Gartner,
1988; Flores et al., 1999; Sprovieri et al., 2003) and in
the recent (Okada and Honjo, 1973; Honjo and Okada,
1974; Geitzenauer et al., 1977; Okada and McIntyre,
1979; Kleijne et al., 1989; Giraudeau, 1992; Jordan
and Chamberlain, 1997; Jordan and Winter, 2000;
Hagino et al., 2000; Findlay and Giraudeau, 2000) and
to prefer high nutrient content (Winter, 1982; Mitchell-
Innes and Winter, 1987; Gartner, 1988; Houghton and
Guptha, 1991; Kinkel et al., 2000; Andruleit and
Rogalla, 2002; Cortes et al., 2001) and low salinity
(Kleijne, 1993; Jordan and Winter, 2000). In the
Mediterranean, it is absent in the column water and
surface sediments from the eastern basin and is
considered a tracer for Atlantic surface waters (Knap-
pertsbusch, 1993).
Unlike gephyrocapsids, abundance of reticulofe-
nestrids, although highly variable through the sec-
tions, is not clearly related to oxygen isotope
fluctuations and shows increase both in glacial and
interglacial stages (Fig. 8). The abundances of
Reticulofenestra asanoi and of Reticulofenestra sp.
are higher in the Atlantic than in the Mediterranean
and decrease eastward within the Mediterranean
basin. Although no ecological preferences are known
for these taxa, this trend may be a response to increase
in salinity, suggesting a preference for normal salinity
waters. However, further data need to be collected on
this topic.
5. Conclusion
The quantitative data collected from the Mediter-
ranean and North Atlantic deep-sea sections allowed
to outline characters in the distributions of selected
calcareous nannofossils at the early–middle Pleisto-
cene transition. Correlation between abundance pat-
terns of calcareous nannofossils, oxygen isotopes and

P. Maiorano, M. Marino / Marine Micropaleontology 53 (2004) 405–422 419
sapropel stratigraphy enabled testing of the strati-
graphical value of bioevents to provide biochrono-
logic data and to evaluate patterns of diachrony.
Possible relations of evolutionary and/or ecostrati-
graphical events to environmental parameters have
also been inferred.
The results on the distribution of Reticulofenestra
sp. and Reticulofenestra asanoi allow few consid-
erations on the previous inconsistent calibrations of
both the lowest and highest occurrences of R.
asanoi to marine isotope stages. Unambiguous
taxonomy is necessary to differentiate Reticulofe-
nestra sp. from R. asanoi; the two taxa have
comparable abundance patterns and similar mor-
phology, and their phylogenetic relationship cannot
be excluded. However, the occurrences of specimens
of Reticulofenestra sp. slightly below the lowest
occurrence of R. asanoi may be responsible of the
controversial results in previous findings. Clear
taxonomic criteria allow us to propose the FO of
Reticulofenestra sp. as reliable event occurring at
MIS 35 or MIS 35/34 transition. The distribution of
R. asanoi is characterized by very rare and scattered
occurrences in the lower part of the range and
suggests a low reproducibility of the FO of the
species; on the other hand, the increase in abun-
dance of the species (FCO) can be identified at MIS
32 or 31/32. The uppermost distribution of R.
asanoi indicates a high reliability of the LCO of
the species at MIS 23, well comparable to previous
data.
In the investigated interval, the medium-sized
Gephyrocapsa are almost exclusively represented by
Gephyrocapsa omega, and their distribution appears
significantly environmentally controlled. The reap-
pearance of medium-sized gephyrocapsids (contem-
poraneous to the FO of G. omega) in the middle
Pleistocene, which is known to follow a latitudinal
trend, is probably also affected by a migratory trend
from the Atlantic to the Mediterranean Sea and by a
regional environmental gradient (salinity and/or
nutrient) within the Mediterranean. This is sug-
gested by a pattern in diachrony of the event
between Atlantic and western Mediterranean, as
well as between western and eastern Mediterranean,
on the order of 30–60 ky between Atlantic and
Mediterranean and of about 30 ky within the
Mediterranean.
The two intervals of temporary disappearance of
Gephyrocapsa omega correlated to the glacial MIS
22 and MIS 20 remarkably improve the stratigraphic
constraints at the lower–middle Pleistocene transition
if high resolution data (3–5 ky) are available and are
interpreted as an ecological response to significant
glacial conditions. The positive correlation between
increases in the abundance of G. omega and
interglacial fluctuations suggests that the species
may provide valuable signals of paleoclimatic
amelioration in the middle Pleistocene record
through MIS 25–15. In this framework, the impov-
erishment of G. omega and of all medium-sized
Gephyrocapsa from MIS 22/20 transition upwards
can be interpreted as a response to the global
climatic deterioration. Moreover, the eastward
decrease of G. omega within the Mediterranean
seems to be related to increased salinity and
decreased nutrients.
Acknowledgements
We wish to thank the Ocean Drilling Program for
providing samples of all the investigated sites. We are
deeply indebted to J. Henderiks and J. Young for the
careful review of the manuscript and their valuable
suggestions, which greatly improved the early ver-
sion of the paper. This research was financially
supported by MIUR Grant 60% (2002) to A.
D’Alessandro.
Appendix A. Taxonomic list
All taxa cited in the paper are listed below
Gephyrocapsa Kamptner, 1943
G. oceanica Kamptner, 1943
G. oceanica s.l >4 Am sensu Rio, 1982
G. caribbeanica Boudreaux and Hay, 1967
G. omega Bukry, 1973
Pseudoemiliania, Gartner, 1969
P. lacunosa (Kamptner, 1963) Gartner, 1969
Reticulofenestra Hay et al., 1966
R. asanoi Sato and Takayama, 1992

P. Maiorano, M. Marino / Marine Micropaleontology 53 (2004) 405–422420
References
Andruleit, H.A., Rogalla, U., 2002. Coccolithophores in surface
sediments of the Arabian Sea in relation to environmental
gradients in surface waters. Mar. Geol. 186, 505–526.
Backman, J., Shackleton, N.J., 1983. Quantitative biochronoly of
Pliocene and early Pleistocene calcareous nannofossils from the
Atlantic, Indian and Pacific oceans. Mar. Micropaleontol. 8,
141–170.
Bethoux, J.P, Gentili, B., Morin, P., Nicolas, E., Pierre, C.,
Ruiz-Pino, D., 1999. The Mediterranean Sea: a miniature
ocean for climatic and environmental studies and a key for
the climatic functioning of the North Atlantic. Prog. Oceanogr.
44, 131–146.
Bollmann, J., 1997. Morphology and biogeography of the genus
Gephyrocapsa coccoliths, in Holocene sediments. Mar. Micro-
paleontol. 29, 319–350.
Bollmann, J., Baumann, K.H., Thierstein, R.H., 1998. Global
dominance of Gephyrocapsa coccoliths in the late Pleistocene:
selective dissolution, evolution or global environmental change?
Paleoceanography 13, 517–529.
Bown, P.R., Young, J.R., 1998. Chapter 2: techniques. In: Bown,
P.R. (Ed.), Calcareous Nannofossil Biostratigraphy. Kluwer
Academic Publishing, Dordrecht, pp. 16–28.
Castradori, D., 1993. Calcareous nannofossil biostratigraphy and
biochronology in eastern Mediterranean deep-sea cores. Riv.
Ital. Paleontol. Stratigr. 99, 107–126.
Cortes, M., Bollmann, J., Thierstein, H.R., 2001. Coccolithophore
ecology at the HOT station ALOHA Hawai. Deep-Sea Res., II
48, 1957–1981.
de Kaenel, E., Siesser, W.G., Murat, A., 1999. Pleistocene
calcareous nannofossil biostratigraphy and the western Medi-
terranean sapropels, Sites 974 to 977 and 979. In: Zhan, R.,
Comas, M.C., Klaus, A. (Eds.), Proc. ODP Sci. Results 161.
College Station, Texas, pp. 15–183.
Emeis, K.C., Sakamoto, T., Wehausen, R., Brumsack, H.J., 2000.
The sapropel record of the eastern Mediterranean sea-results of
Ocean Drilling Program Leg 160. Palaeogeogr. Palaeoclimatol.
Palaeoecol. 158, 371–395.
Findlay, C.S., Giraudeau, J., 2000. Extant calcareous nannoplaknton
in the Australia sector of the southern ocean (Austral summer
1994 and 1995). Mar. Micropaleontol. 40, 417–439.
Flores, J.A., Gersonde, R., Sierro, F.J., 1999. Pleistocene fluctua-
tions in the Agulhas current retroflection based on the
calcareous plankton record. Mar. Micropaleontol. 37, 1–22.
Flores, J.A., Gersonde, R., Sierro, F.J., Niebler, H.S., 2000.
Southern ocean Pleistocene calcareous nannofossil events:
calibration with isotope and geomagnetic stratigraphies. Mar.
Micropaleontol. 40, 377–402.
Fornaciari, E., 2000. Calcareous nannofossil biostratigraphy of
the California margin. In: Lyle, M., Koizumi, I., Richter, C.,
Jr., T.C. (Eds.), Proc. ODP Sci. Res. 167. College Station,
Texas, pp. 3–40.
Gartner, S., 1977. Calcareous nannofossil biostratigraphy and
revised zonation of the Pleistocene. Mar. Micropaleontol. 2,
1–25.
Gartner, S., 1988. Paleoceanography of the mid-Pleistocene. Mar.
Micropaleontol. 13, 23–46.
Geitzenauer, K.R., Roche, M.B., McIntyre, A., 1977. Coccolith
biogeography from North Atlantic and Pacific surface sedi-
ments; a comparison of species distribution and abundances. In:
Ramsay, A.T.S. (Ed.), Oceanic Micropaleontology. Academic
Press, London, pp. 973–1008.
Giraudeau, J., 1992. Distribution of recent nannofossils beneath the
Benguela system: southwest African continental margin. Mar.
Geol. 108, 219–237.
Hagino, K., Okada, H., Matsuoka, H., 2000. Spatial dynamics of
coccolithophore assemblages in the Equatorial western–central
Pacific Ocean. Mar. Micropaleontol. 39, 53–72.
Herman, Y., Hopkins, D.M., 1980. Arctic oceanic climate in late
Cenozoic time. Science 209, 557–562.
Hilgen, F.J., 1991. Astronomical calibration of Gauss to
Matuyama sapropels in the Mediterranean and implication
for the geomagnetic polarity time scale. Earth Planet. Sci. Lett.
104, 226–244.
Hilgen, F.J., Lourens, L.J., Berger, A., Loutre, M.F., 1993.
Evaluation of the astronomically calibrated time scale for the
late Pliocene and earliest Pleistocene. Paleoceanography 8,
549–565.
Honjo, S., Okada, H., 1974. Community structure of coccolitho-
phores in the photic layer of the mid-Pacific. Micropaleontology
20, 209–230.
Houghton, S.D., Guptha, M.V.S., 1991. Monsoonal and fertility
control on recent marginal sea and continental shelf coccolith
assemblages from the western Pacific and northern Indian
oceans. Mar. Geol. 97a, 251–259.
Howell, M.W., Thunell, R.C., Di Stefano, E., Tappa, E.J.,
Sakamoto, T., 1998. Stable isotope chronology and paleocea-
nography history of Sites 963 and 964, eastern Mediterranean
Sea. In: Robertson, H.F., Emeis, K., Richter, C., et al. (Eds.),
Proc. ODP Sci. Res. 160. College Station, Texas, pp. 67–180.
Jordan, R.W., Chamberlain, A.H-L., 1997. Biodiversity among
haptophyte algae. Biodivers. Conserv. 6, 131–152.
Jordan, R.W., Winter, A., 2000. Assemblages of coccolithophorids
and other living microplankton off the coast of Puerto Rico
during January–May 1995. Mar. Micropaleontol. 39, 113–130.
Kinkel, H., Baumann, K.-H., Cepek, M., 2000. Coccolithophores in
the equatorial Atlantic Ocean: response to seasonal and late
Pleistocene surface water variability. Mar. Micropaleontol. 39,
87–112.
Kleijne, A., 1993. Morphology, Taxonomy and Distribution of
Extant Coccolithophorids (Calcareous Nannoplankton). PhD
thesis, Vrije Universiteit, Amsterdam, 321 pp.
Kleijne, A., Kroon, D., Zevemboom, W., 1989. Phytoplankton and
foraminiferal frequencies in northern Indian Ocean and Red Sea
surface waters. Neth. J. Sea Res. 24, 531–539.
Knappertsbusch, M., 1993. Geographic distribution of living and
Holocene coccolithophores in the Mediterranean Sea. Mar.
Micropaleontol. 21, 219–247.
Kroon, D., Alexander, I., Little, M., Lourens, L.J., Mattehewson,
A., Robertson, A.H.F., Sakamoto, T., 1998. Oxygen isotope and
sapropel stratigraphy in the eastern Mediterranean during the
last 3.2 million years. In: Robertson, H.F., Emeis, K.C., Richter,

P. Maiorano, M. Marino / Marine Micropaleontology 53 (2004) 405–422 421
C., et al. (Eds.), Proc. ODP Sci. Res. 160. College Station,
Texas, pp. 181–190.
Lourens, L.J., Antonarakou, A., Hilgen, F.J., Van Hoof, A.A.M.,
Vergnaud-Grazzini, C., Zachariasse, W.J., 1996. Evaluation of
the Plio–Pleistocene astronomical timescale. Paleoceanography
11, 391–413.
Maiorano, P., Marino, M., Di Stefano, E., Ciaranfi, N., 2004.
Calcareous nannofossil events in the lower–middle Pleistocene
transition at the Montalbano Jonico section and ODP Site 964:
calibration with isotope and sapropel stratigraphy. Riv. Ital.
Paleontol. Stratigr. 110 (2), 547–556.
Marino, M., 1996. Quantitative calcareous nannofossil biostratig-
raphy of lower–middle Pleistocene Montalbano Ionico section
(southern Italy). Palaeopelagos 6, 347–360.
Marino, M., Maiorano, M., Monechi, S., 2003. Quantitative
Pleistocene calcareous nannofossil biostratigraphy of Leg 86
Site 577 (Shatsky Rise northwestern Pacific Ocean). J.
Nannoplankton Res. 25 (1), 25–37.
Matsuoka, H., Okada, H., 1989. Quantitative analysis of Pleistocene
nannoplankton in the subtropical nothwestern Pacific Ocean.
Mar. Micropaleontol. 14, 97–118.
McIntyre, A., McIntyre, R., 1971. Coccolith concentrations and
differential solution in oceanic sediments. In: Funnell, B.M.,
Riedel, W.R. (Eds.), The Micropaleontology of Oceans. Cam-
bridge Univ. Press, London, pp. 253–261.
McIntyre, A., Be, A.W.H., Roche, M.B., 1970. Modern Pacific
coccolithophorida: a paleontological thermometer. N. Y. Acad.
Sci. Trans. Ser. II 32, 720–731.
Mitchell-Innes, B.A., Winter, A., 1987. Coccolithophores: a major
phytoplankton component in mature upwelled water off the
Cape Peninsula, South Africa in March, 1983. Mar. Biol. 95,
25–30.
Mix, A.C., Le, J., Shackleton, N.J., 1995. Benthic foraminiferal
stable isotope stratigraphy of Site 846: 0–1.8 Ma.. In: Pisias,
N.G., Mayer, L., Janecek, T., Palmer-Julson, A., (Eds.), Proc.
ODP, Sci. Res. 138. College Station, Texas, pp. 839–856.
Murat, A., 1999. Pliocene–Pleistocene occurrence of sapropels
in the western Mediterranean Sea and their relation to
eastern Mediterranean sapropels. In: Zahn, R., Comas, M.C.,
Klaus, A. (Eds.), Proc. ODP, Sci. Res. 161. College Station,
Texas, pp. 519–528.
Okada, H., Honjo, S., 1973. The distribution of oceanic coccoli-
thophorids in the Pacific. Deep-Sea Res. 20, 355–374.
Okada, H., McYntyre, A., 1979. Seasonal distribution of modern
coccolithophores in the western North Atlantic Ocean. Mar.
Biol. 54, 319–328.
Pisias, N.G., Moore, T.C., 1981. The evolution of Pleistocene
climate: a time series approach. Earth Planet. Sci. Lett. 52,
450–458.
Prell, W.L., 1982. Oxygen and carbon isotope stratigraphy for the
Pleistocene of Hole 502B: evidence for two modes of isotopic
variability. In: Prell, W.L., Gardner, J.V., et al. (Eds.), DSDP
Init. Reports 68. Government Printing Office, Washington, DC,
U.S., pp. 455–466.
Raffi, I., 2002. Revision of the early–middle Pleistocene calcareous
nannofossil biochronology (1.75–0.85 Ma). Mar. Micropaleon-
tol. 45, 25–55.
Raffi, I., Backman, J., Rio, D., Shackleton, N.J., 1993. Early
Pleistocene and late Pliocene nannofossil biostratigraphy and
calibration to oxygen isotope stratigraphies from DSDP Site 607
and ODP Site 677. Paleoceanography 8, 387–404.
Rio, D., 1982. The fossil distribution of coccolithophore genus
Gephyrocapsa Kamptner and related Plio–Pleistocene chrono-
stratigraphic problems. In: Prell, W.L., Gardner, J.V., et al.
(Eds.), DSDP Init. Reports 68. Government Printing Office,
Washington, DC, U.S., pp. 325–343.
Rio, D., Raffi, I., Villa, G., 1990. Pliocene–Pleistocene calcareous
nannofossil distribution patterns in the western Mediterranean.
In: Kastens, K.A., Mascle, J., et al. (Eds.), Proc. ODP, Sci. Res.
107. College Station, Texas, pp. 513–533.
Rohling, E.J., De Rijk, S., Myers, P.G., Haines, K., 2000.
Palaeoceanography and numerical modelling: the Mediterranean
Sea at times of sapropel formation. In: Hart, M.B. (Ed.),
Climates: Past and Present, Geol. Soc. 181. Spec. Publ.,
London, pp. 135–149.
Roth, P.H., 1994. Distribution of coccoliths in oceanic sediments.
In: Winter, A., Siesser, W.G. (Eds.), Coccolithophores. Uni-
versity Press, Cambridge, pp. 199–218.
Roth, P.H., Berger, W.H., 1975. Distribution and dissolution of
coccoliths in the south and central Pacific. In: Sliter, W.V.,
Be, A.W., Berger, W.H. (Eds.), Dissolution of Deep Sea
Carbonates. Cushman Found, Foraminifera Res. Spec. Publ.
13, pp. 87–113.
Roth, P.H., Coulbourn, W.T., 1982. Floral and solution patterns of
coccoliths in surface sediments of the north Pacific. Mar.
Micropaleontol. 7, 1–52.
Ruddiman, W.F., Raymo, M.E., McIntyre, A., 1986. Matuyama
41,000-year cycles: North Atlantic Ocean and Northern Hemi-
sphere ice sheets. Earth Planet. Sci. Lett. 80, 117–129.
Ruddiman, W.F., Raymo, M.E., Martinson, D.G., Clement, B.M.,
Backman, J., 1989. Pleistocene evolution of Northern Hemi-
sphere climate. Paleoceanography 4, 353–412.
Sakamoto, T., Janecek, T., Emeis, K.C., 1998. Continuous
sedimentary sequences from the eastern Mediterranean Sea:
composite depth sections. In: Robertson, A.H.F., Emeis, K.C.,
Richter, C., Camerlenghi, A. (Eds.), Proc. ODP Sci. Res. 160.
College Station, Texas, pp. 29–36.
Sato, T., Kameo, K., Takayama, T., 1991. Coccolith biostratigraphy
of Arabian Sea. In: Prell, W.L., et al. (Eds.), Proc. ODP Sci. Res.
117. College Station, Texas, pp. 37–54.
Shackleton, N.J., Berger, A., Peltier, W.R., 1990. An alternative
astronomical calibration of the lower Pleistocene time scale
based on ODP site 677. Trans. R. Soc. Edinb. Earth Sci. 81,
251–261.
Sprovieri, R., Di Stefano, E., Howell, M., Sakamoto, T., Di Stefano,
A., Marino, M., 1998. Integrated calcareous plankton biostra-
tigraphy and cyclostratigraphy at Site 964. In: Robertson,
A.H.F., Emeis, K.C., Richter, C., Camerlenghi, A. (Eds.), Proc.
ODP, Sci. Res. 160. College Station, Texas, pp. 155–165.
Sprovieri, R., Di Stefano, E., Incarbona, A., Gargano, M.E., 2003.
A high-resolution record of the last deglaciation in the Sicily
channel based on foraminifera and calcareous nannofossil
quantitative distribution. Palaeogeogr. Palaeoclimatol. Palae-
oecol. 202, 119–142.

P. Maiorano, M. Marino / Marine Micropaleontology 53 (2004) 405–422422
Takahashi, K., Okada, H., 2000. The paleoceanography for the
last 30,000 years in the southeastern Indian Ocean by
means of calcareous nannofossils. Mar. Micropaleontol. 40,
83–103.
Takayama, T., Sato, T., 1987. Coccolith biostratigraphy of the North
Atlantic Ocean, DSDP Leg 94. In: Ruddiman, W.F., Kidd, R.B.,
Thomas, E., et al. (Eds.), DSDP Init. Reports 94. Government
Printing Office, Washington, DC, U.S., pp. 651–702.
Thierstein, H.R., Geitzenauer, K., Molfino, B., Shackleton, N.J.,
1977. Global synchroneity of late Pleistocene coccolith
datum levels; validation by oxygen isotopes. Geology 5,
400–404.
von Grafenstein, R., Zahn, R., Tiedemann, R., Murat, A., 1999.
Planktonic y18O records at Sites 976 and 977, Alboran Sea:
stratigraphy, forcing, and paleoceanographic implications. In:
Zahn, R., Comas, M.C., Klaus, A. (Eds.), Proc. ODP, Sci. Res.
161. College Station, Texas, pp. 469–480.
Wei, W., 1993. Calibration of upper Pliocene–lower Pleistocene
nannofossil events with oxygen isotope stratigraphy. Paleocea-
nography 8, 85–89.
Winter, A., 1982. Paleoenvironmental interpretation of Pleistocene
coccolith assemblages from the gulf of Aqaba (Elat) Red Sea.
Rev. Esp. Micropaleontol. 14, 291–314.
Worsley, T.R., Herman, Y., 1980. Periodic ice-free Arctic Ocean in
Pliocene and Pleistocene time: calcareous nannofossil evidence.
Science 210, 323–325.